المعادن الحيوية، وهي المعادن غير العضوية للكائنات الحية، معروفة بشكل رئيسي بوظائفها المتعلقة بالخصائص الفيزيائية في الكائنات الحية الحديثة. اكتشافنا الأخير للأنشطة الشبيهة بالإنزيمات للمواد النانوية، التي أطلق عليها اسم نانوإنزيم، يلهم الفرضية بأن المعادن الحيوية النانوية قد تعمل كعوامل تحفيز شبيهة بالإنزيمات في الخلايا. هنا نبلغ أن النوى الحديدية للفريتينات البيولوجية تعمل كنانوزيمات طبيعية لالتقاط الجذور الحرة من سوبر أكسيد. من خلال تحليل ثمانية عشر فريتينا تمثل ثلاث ممالك حية، نجد أن النواة الحديدية لفريتينا بدائية النواة تمتلك نشاطًا أعلى في تقليل سوبر أكسيد مقارنة بتلك الخاصة بالنواة الحقيقية. تكشف التحقيقات الإضافية أن الاختلافات في القدرة التحفيزية ناتجة عن تغييرات في نسبة الحديد/الفوسفات في النواة الحديدية، والتي تحددها بشكل رئيسي هياكل الفريتينات. يقوم الفوسفات في النواة الحديدية بتحويل النواة الحديدية من هيكل بلوري مفرد إلى هيكل شبيه بالفوسفات الحديدي غير المتبلور، مما يؤدي إلى تقليل الألفة لبروتون الهيدروجين في النواة الشبيهة بالفيريهايدرايت، مما يسهل تفاعلها مع سوبر أكسيد بطريقة تختلف عن تلك الخاصة بأيونات الحديد الثلاثي. علاوة على ذلك، فإن التعبير المفرط عن الفريتينات ذات الأنشطة العالية في تقليل سوبر أكسيد فيزيادة مقاومة الإشريكية القولونية للأكسجين الفائق، بينما يؤدي حذف بكتيريوفيريتين أو إدخال فيريتين بشري إلى تقليل تحمل الجذور الحرة، مما يبرز الدور الفسيولوجي لمضادات الأكسدة لهذا النوع من النانوإنزيمات.
المعادن غير العضوية التي تحدث بشكل طبيعي كمواد صلبة متجانسة ذات تركيبة كيميائية محددة، موزعة على نطاق واسع على الأرض. حتى الآن، تم تسجيل ما يقرب من 5800 نوع مختلف من المعادن من قبل الجمعية الدولية للمعادن (IMA).وفقًا لروبرت م. هازن، شهدت تطور المعادن 10 مراحل متداخلة جزئيًا، بدءًا من ظهور المعادن الكوندريتية الأولية إلى التمعدن الحيوي في العصر الظاهر.من الجدير بالذكر أن ما يقرب من ثلثي جميع المعادن الطبيعية نتجت عن تحول الأرض بواسطة الكائنات الحية، خاصة في الفترة التي تلت “حدث الأكسدة العظيم”، مما يشير إلى العلاقة الوثيقة بين توليد المعادن وتطور الحياة. في الواقع، يُعتقد أن المعادن لا تشارك فقط في نشأة الحياة، بل أصبحت أيضًا مكونات متكاملة للكائنات الحية في معظم، إن لم يكن جميع، الممالك الحية.في زمن ما قبل الحياة، كان يُعتبر هيكل الإنزيمات معقدًا جدًا ليكون متاحًا. وكان يُفترض أن المعادن غير العضوية تحفز تخليق وتعدد الجزيئات الأولية.
الجزيئات الحيويةيمكن أن تحفز المعادن التخليق العضوي الأساسي، مثل تثبيت الكربون في البيئة البدائية.وإدراك اختيار الجزيئات الكيرالية على السطح الكيرالي للمعادن الطبيعية، مثل الكالسيتبالإضافة إلى تخليق وتركيز الجزيئات الصغيرة، يمكن أن يتم تحفيز بلمرة الببتيدات وجزيئات RNA أيضًا بواسطة المعادن الطينية مثل معادن الهيدروكسيد المزدوجة الطبقات.في الكائنات الحية الحديثة، توجد كميات كبيرة من المعادن الحيوية في معظم الأنواع، مثل الكربونات والمغنيتيت في البكتيريا.أكسالات الكالسيوم في النباتات والعظام في الحيوانات. ومع ذلك، على عكس أدوارها التحفيزية في الفترة ما قبل الحيوية، تلعب المعادن الحيوية بشكل رئيسي أدوار الدعم الفيزيائي والحماية، مثل دور الدعم الذي تقدمه السيليكات في الدياتومات والعظام في الحيوانات..
كان الدور التحفيزي للمواد البيومعدنية قليل النقاش حتى اكتشاف النشاط الشبيه بالإنزيمات للمواد النانوية غير العضوية.. لقد تم دراسة هذه النانوإنزيمات على نطاق واسع منذ ذلك الحين في مجالات متنوعة بما في ذلك علوم البيئة، وصناعة الأدوية، وما إلى ذلك. بالإضافة إلى خصائصها الفيزيائية والكيميائية الجوهرية مثل المغناطيسية والتألقتمتلك النانو زيمات أنشطة تحفيزية وحركيات تفاعل مشابهة للإنزيمات الطبيعية.حالياً، تم تطوير مجموعة متنوعة من النانوإنزيمات من الجسيمات المغناطيسية الأصلية إلى آلاف المواد النانوية المختلفة وحتى النانوإنزيمات ذات الذرة الواحدة التي يمكن أن تنافس البيروكسيداز الطبيعي من الفجل.تجعل هذه الخصائص الفريدة من النانو زيم محاكياً مفضلاً للإنزيمات في التطبيقات المتعلقة بحماية البيئة والزراعة وتشخيص الأمراض وعلاجها..
من المثير للاهتمام أنه تم نشر عدد قليل من التقارير حول النانوإنزيمات الطبيعية من المعادن الحيوية، بما في ذلك المغناطيسوم في البكتيريا المغناطيسية، والنانوإنزيم المعدني في الفطريات، والفيريتين من طحال الحصان.التي تظهر أنشطة شبيهة بالبيروكسيداز بالإضافة إلى كونها مكونات طبيعية. علاوة على ذلك، تم إثبات أن المغناطيسوم والمعادن النانوية للواجهة الفطرية لعبت دورًا مضادًا للأكسدة في حماية مضيفها من الجذور الحرة السوبر أكسيد أو الهيدروكسيل، مما يشير إلى أن هذه الأنزيمات النانوية قد تكون لها وظيفة فسيولوجية مهمة. نظرًا للتنوع والتوزيع الواسع للمعادن الحيوية، فمن المحتمل أن تكون الأنزيمات النانوية الطبيعية موجودة وفعالة في أنواع متعددة من الكائنات الحية. ومن الجدير بالذكر أن بروتين تخزين الحديد الفيريتين يوجد كهيكل نانوي طبيعي في جميع الكائنات الحية تقريبًا بما في ذلك البكتيريا والعتائق واليوكاريوت.على الرغم من الاختلافات الكبيرة في تسلسلاتها، تخزن الفيريتينات من جميع الأنواع الحديد في شكل نواة الحديد غير العضوي، وهو نوع كلاسيكي من المواد النانوية غير العضوية في الكائنات الحية.
في الدراسة الحالية، نقوم بفحص 18 فيريتين من البكتيريا والعتائق واليوكاريوتيات من حيث أنشطتها كنانوزيم ونفكك الآليات الأساسية. وقد وُجد أنه، بالمقارنة مع فيريتينات اليوكاريوتي، تظهر فيريتينات البروكاريوتي نشاطًا أعلى يشبه نشاط سوبر أكسيد ديسموتاز (SOD). تُظهر بياناتنا أن الاختلافات في نشاط SOD الشبيه لهذه الفيريتينات ناتجة عن تغييرات في نسبة الحديد/الفوسفات في النوى الحديدية. تمتلك فيريتينات العتائق والبكتيريا محتوى فوسفات أعلى مقارنةً بفيريتيين الإنسان، وهو ما تحدده تسلسلات الأحماض الأمينية والهياكل الخاصة بها. علاوة على ذلك، تُظهر تحليلات الامتصاص بالأشعة السينية الممتدة (EXAFS) ونظرية الكثافة الوظيفية (DFT) أن الفوسفات داخل النواة الحديدية، وليس الموجود على السطح، يغير قدرة ارتباط بروتون الهيدروجين بالنواة الحديدية، مما يؤثر على القدرة التفاعلية مع السوبر أكسيد. والأهم من ذلك، أن الفيريتينات ذات الأنشطة العالية الشبيهة بـ SOD يمكن أن تلعب دورًا مضادًا للأكسدة في الحالة النفسية. تُظهر هذه النتائج أن النوى الحديدية للفيريتيين البيوجيني من أنواع مختلفة لديها قدرات مميزة على القضاء على الجذور الحرة للسوبر أكسيد. تحدد هياكل الفيريتينات تركيبة النوى الحديدية التي بدورها تقرر نشاط SOD الشبيه، مما يوضح أن النشاط الشبيه بالإنزيم للنانومعادن الحيوية الطبيعية يعتمد على الهيكل ومحفوظ تطوريًا، وقد يلعب دورًا مهمًا في الحفاظ على التوازن الداخلي للكائنات الحية.
النتائج والمناقشة
نوى الحديد البيوجيني للفيريتينات من كائنات مختلفة تظهر أنشطة مشابهة لإنزيم سوبر أكسيد ديسموتاز
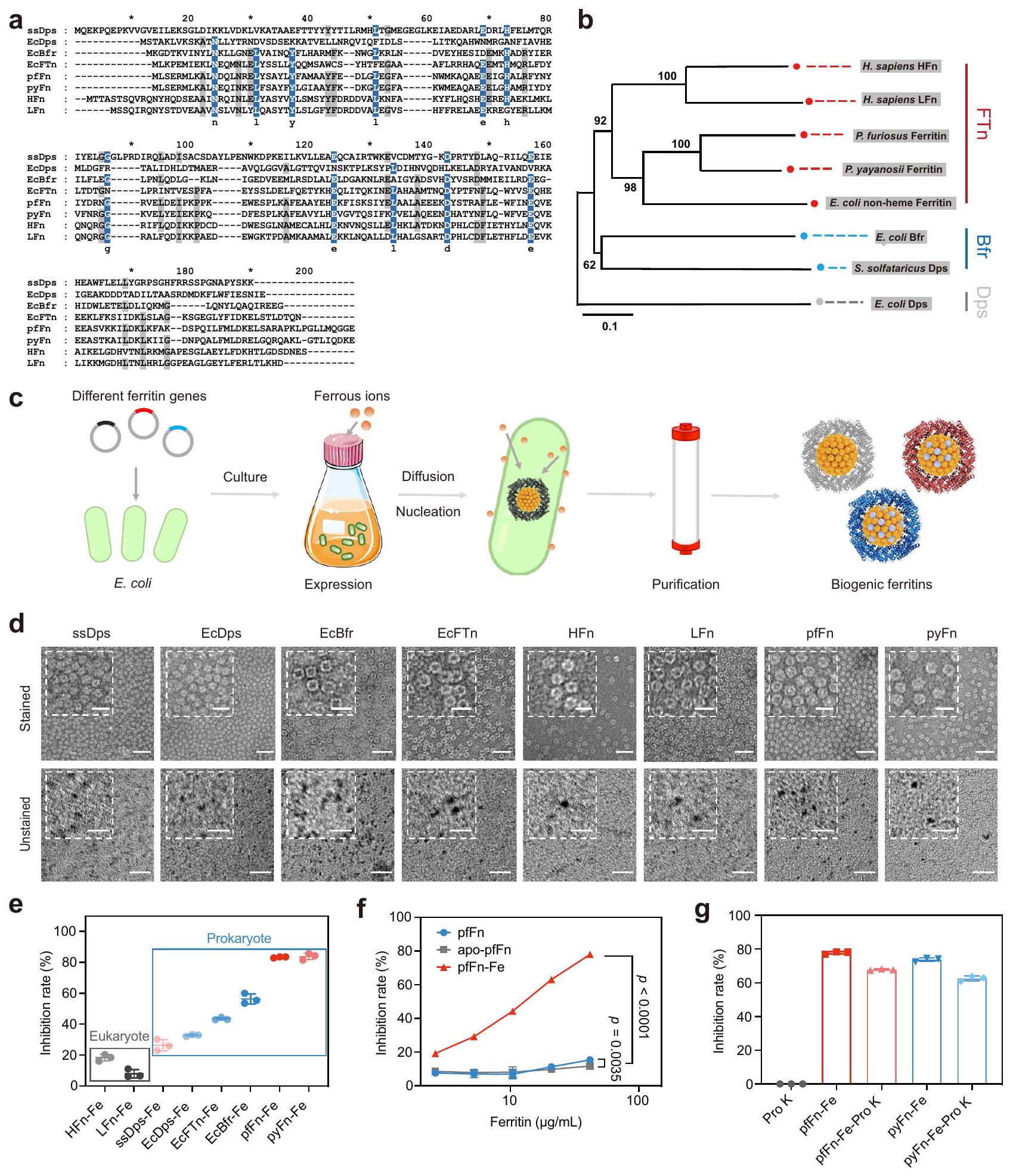
استنادًا إلى هياكلها، يتم تصنيف الفيريتينات عادةً إلى ثلاث عائلات فرعية: بروتين ربط الحمض النووي من الخلايا الجائعة (Dps)، بكتيريوفيريتين (Bfr)، والفيريتين الكلاسيكي (FTn).. من بينها، تم توزيع Dps بهيكل نواة-غلاف مكون من 12 وحدة (بقطر خارجي 9 نانومتر) في العتائق والبكتيريا، بينما توجد الفيريتينات من مجموعتي Bfr وFTn كهيكل أكبر مكون من 24 وحدة بقطر خارجي 12 نانومتر. الفيريتين FTn كفيريتين نموذجي موجود في جميع المجالات الثلاثة للحياة، بينما توجد الفيريتينات من عائلة Bfr بشكل رئيسي في البكتيريا وتحتوي على هيكل هيم خاص في محورها الثنائي، مما يسهل نقل الإلكترون للحديد.من أجل مقارنة الفيريتينات ذات الهياكل المختلفة من المجالات الثلاثة للحياة بشكل منهجي، تم اختيار ثمانية فيريتينات شائعة، بما في ذلك Dps من البكتيريا (E. coli، EcDps) والعتائق (S. solfataricus، ssDps)، FTn من البكتيريا (E. coli، EcFTn)، والعتائق (P. furiosus و P. yayanosii، pfFn و pyFn) واليوكاريوت.. سابينس، سلسلة ثقيلة من الفيريتين (HFn) وسلسلة خفيفة من الفيريتين (LFn)، Bfr من البكتيريا (E. coli، EcBfr). أولاً، تم إجراء محاذاة تسلسلية متعددة لتحديد تشابه التسلسل لهذه الفيريتينات. كما هو موضح في الشكل 1a، فإن الفيريتينات الثمانية لديها تشابه منخفض بشكل عام في الأحماض الأمينية. ومع ذلك، فإن التحليل النشوي (الشكل 1b) بواسطة طريقة الجوار القريب كشف أن معظم هذه الفيريتينات يمكن أن تتجمع في عائلات Dps وBfr وFTn، مما يوضح أن الفيريتينات من نفس المجموعة أظهرت تسلسلات أحماض أمينية متشابهة نسبياً. ومع ذلك، تم تجميع Dps من S. solfataricus مع Bfr من E. coli، وهو ما يتماشى مع التقرير السابق الذي أشار إلى أنه يمتلك مركز أكسدة الحديد مشابه لـ. بعد ذلك، لتقييم الأنشطة الشبيهة بالإنزيمات لنواة الحديد في الفيريتين، قمنا أولاً بتخليق نواة الفيريتين حيوياً عن طريق إضافة ملح الحديد الثنائي إلى وسط الثقافة أثناء تعبيرها (الشكل 1c) بحيث يمكن أن يتم تمعدن الحديد الزائد في الفيريتينات على شكل نواة الحديد غير العضوي. ثم تم تنقية هذه الفيريتينات المتمعدنة حيوياً كما هو موصوف سابقاً.كما هو موضح في SDS-PAGE (الشكل التكميلي 1a)، كان لعملية التخليق الحيوي تأثير ضئيل على وحدات الفيريتين مقارنة بتلك المنقاة من الوسائط الأساسية. أشارت صور المجهر الإلكتروني الناقل (TEM) الملونة وطيف تشتت الضوء الديناميكي (DLS) إلى أن الفيريتينات المعدلة حيوياً حافظت على هياكلها الأصلية من البروتينات. والأهم من ذلك، أن اللون الأحمر البني المميز وصور TEM غير الملونة للفيريتينات المعدلة حيوياً أشارت إلى توليد النواة الحديدية الطبيعية في الفيريتينات. ومع ذلك، فإن الفيريتين ذو السلسلة الخفيفة البشرية وأعضاء عائلة Dps الفرعية تظهر أحجام نوى أصغر وقليل من تغير اللون، على الأرجح بسبب عدم وجود مركز الفيروكسي داز في الأول وقطر Dps الداخلي الأصغر. بعد تنقيتها، تم تقييم هذه الفيريتينات لأنشطة النانو زيم الشائعة بما في ذلك أنشطة SOD و الأكسيداز و البيروكسيداز و الكاتالاز (الشكل التكميلي 2). ومن المثير للاهتمام، أن نشاط SOD الشبيه بهذه الفيريتينات كان مرتبطًا بأصلها. كما هو موضح في الشكل 1e والشكل التكميلي 3a، أظهرت كل من الفيريتينات الأصلية والمصنعة حيوياً الميل المتسق بأن قوة أنشطة SOD الشبيهة بها كانت مرتبة بالترتيب: بدائيات النوى > حقيقيات النوى.
لتحديد مصدر الأنشطة الشبيهة بالـ SOD، تم استخدام حمض الثيوغليكوليك و2،2′-بيبيريديل لتقليل وإزالة النواة الحديدية في الفيريتينات. كما هو موضح في الشكل If والشكل التكميلي 3b-d، كانت الأنشطة الشبيهة بالـ SOD قد انخفضت بشكل كبير مع إزالة النواة، في حين أن النشاط الشبيه بالإنزيم للفيريتينات لم يتأثر بشكل كبير بعد هضم الغلاف البروتيني باستخدام البروتياز K (الشكل 1g والشكل التكميلي 3e)، مما يثبت أن النواة الحديدية كانت مسؤولة عن الأنشطة. بالإضافة إلى ذلك، من خلال إضافة كميات متدرجة من ملح الحديد الثنائي إلى وسط التخليق الحيوي، وُجد أن الأنشطة الشبيهة بالـ SOD لهذه الفيريتينات كانت مرتبطة إيجابياً بمحتواها من الحديد (الشكل التكميلي 3f-h). علاوة على ذلك، لاستبعاد أن النشاط المقلل للأكسيد الفائق لم يكن ناتجاً عن تثبيط أكسيداز الزانثين، قمنا أيضاً بالكشف عن آخر
الشكل 1 | تنقية، توصيف، ونشاط شبيه SOD للفيريتينات من تنظيم تسلسلات متعددة من تسلسلات الأحماض الأمينية لمختلف الفيريتينات؛ الألوان تشير إلى تشابه الأحماض الأمينية، الأزرق:رمادي: . شجرة النشوء والتطور لتسلسلات الأحماض الأمينية للفيريتين؛ الأرقام على الفروع تمثل قيمة البوتستراب. ج. مخطط لعملية التخليق الحيوي لنوى الحديد المختلفة للفيريتين.صور TEM سلبية ممثلة وغير ملونة للفيريتينات المعدلة حيوياً من ثلاث تجارب مستقلة مع نتائج مشابهة؛ تمثل الأشكال المرفقة صورًا مكبرة جزئيًا؛ تشير شريط القياس إلى 50 نانومتر وشريط القياس المرفق يشير إلى 20 نانومتر. e نشاط مشابه لنشاط SOD لـ
الفيريتينات المعدنية الحيوية مع التركيز النهائي للبروتين عند; تُعرض البيانات كمتوسط SD ( تجارب مستقلة).نشاط مشابه لـ SOD لـ apo-pfFn (الفيريتين الذي أُزيل منه الحديد)، pfFn، وpfFn المعدني الحيوي (pfFn-Fe)؛ البيانات مقدمة كمتوسط SD ( تجارب مستقلة)؛ تم تقييم الفرق الكبير بواسطة تحليل التباين ثنائي الاتجاه مع اختبار توكي HSD بعد الاختبار. ج النشاط الشبيه بـ SOD للفيريتينات الأثرية بعد هضم البروتياز K (ProK)؛ البيانات مقدمة كمتوسط SD ( تجارب مستقلة). يتم توفير بيانات المصدر كملف بيانات المصدر.
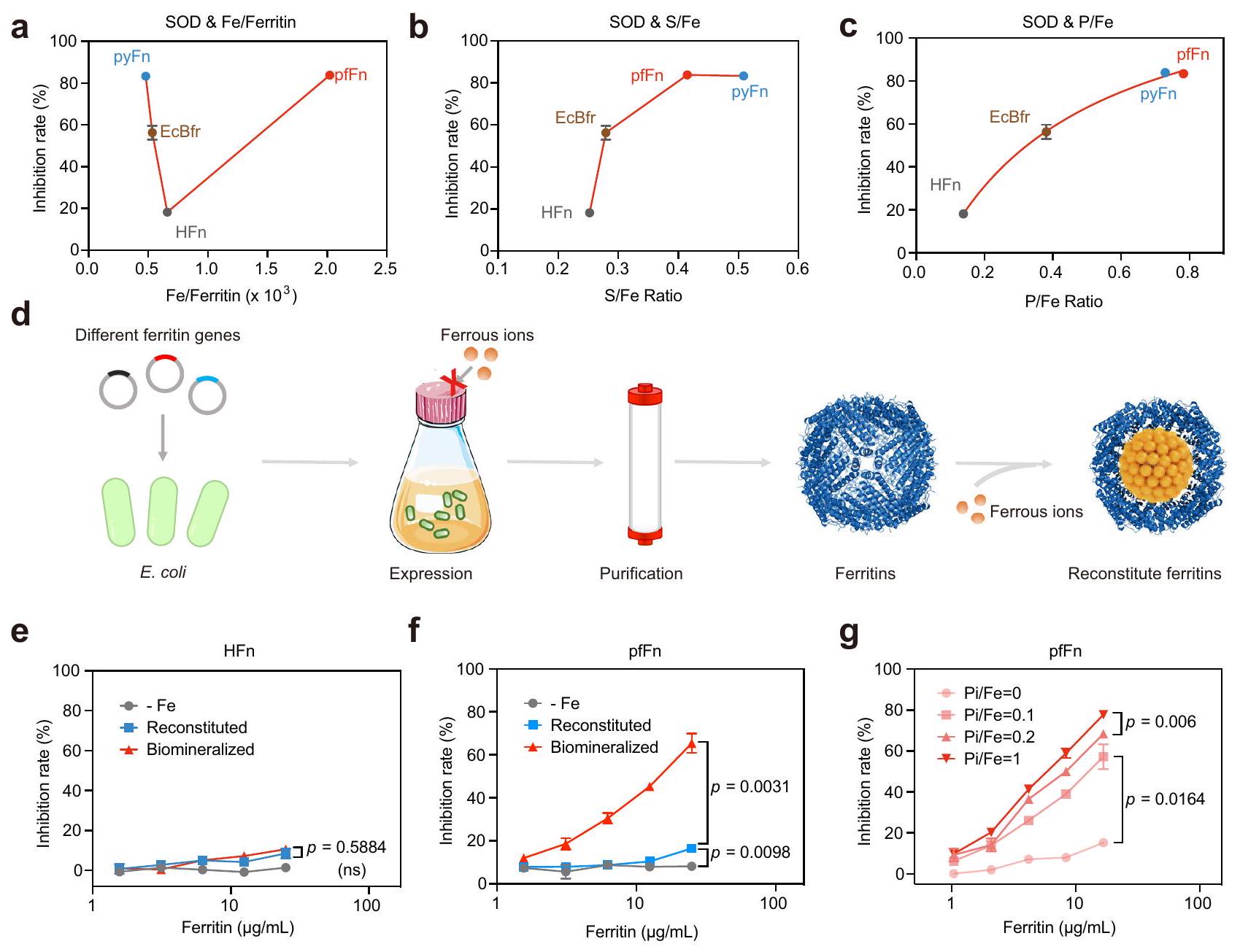
الشكل 2 | تأثير العناصر المعدنية الرئيسية في نواة الفيريتين على نشاط شبيه SOD. نشاط شبيه SOD و (أ) محتوى الحديد. (ب) نسبة S/Fe و (ج) نسبة P/Fe للفيريتينات من كائنات مختلفة؛ البيانات مقدمة كمتوسط SD ( تجارب مستقلة). د مخطط عملية تخليق الفيريتين مع نواة الحديد المعاد تكوينها.نشاط شبيه بـ SOD للمعاد تشكيله، المعدني الحيوي
الفيريتينات والفيريتينات المنقاة من الوسط الأساسي (-Fe). ج النشاط الشبيه بـ SOD لـ pfFn المعاد تكوينه مع نسب الفوسفات/الحديد المتدرجة ( ); البيانات في تُعرض كمتوسط ( تجارب مستقلة)؛ تم تقييم الفرق الكبير بواسطة تحليل التباين ثنائي الاتجاه مع اختبار توكي بعد الاختبار؛ ns: غير دال،تُقدم بيانات المصدر كملف بيانات مصدر. توليد اليورات من نظام توليد السوبر أوكسيد. كما هو موضح في الشكل التكميلي 3i و j، على عكس السوبر أوكسيد، لم يتأثر إنتاج اليورات بشكل واضح بالفيريتينات المعدلة حيوياً. مجتمعة، أظهرت هذه النتائج أن الفيريتينات تظهر قدرات تقليل السوبر أوكسيد المرتبطة بالأنواع وأن النوى الحديدية هي مصدر النشاط. من المهم أن تثبت دراستنا للآلية أن هذا النشاط ليس ناتجًا ببساطة عن أيون المعدن الذي يتم إطلاقه من الفيريتينات.
علاوة على ذلك، بما أن الفيريتينات المختارة هنا كانت محدودة بعدد قليل من الفصائل، للتحقق بشكل أكبر من العلاقة بين النشاط والمصادر، قمنا بعد ذلك بمقارنة تسلسلات الفيريتين من فصائل مختلفة في الممالك النسبية. كما هو موضح في الشكل التوضيحي 4، كانت تسلسلات الفيريتين في الحيوانات من فصائل مختلفة متشابهة. ومع ذلك، كان هناك القليل من التشابه في الأحماض الأمينية في تسلسلات الفيريتين للبكتيريا والعتائق في فصائل مختلفة. بعد ذلك، اخترنا 8 فيريتينات أخرى من Caenorhabditis elegans وCrassostrea gigas وPseudomonas aeruginosa وThermus scotoductus وMycobacterium tuberculosis وThermosphaera aggregans وThermococcus barophilus، والتي تضمنت فصائل مختلفة مقارنةً بالفيريتينات الثمانية السابقة. ومن المثير للاهتمام، أنه بعد التنقية والتوصيف، كان النشاط الشبيه بـ SOD لهذه الفيريتينات لا يزال مرتبطًا بالمصادر (الشكل التوضيحي 5)، مما يؤكد بشكل أكبر القواعد النشطة التي وجدناها.
التغير في نشاط شبيه SOD لنوى الحديد في الفيريتين الناتج عن اختلافات في نسبة الحديد/الفوسفور
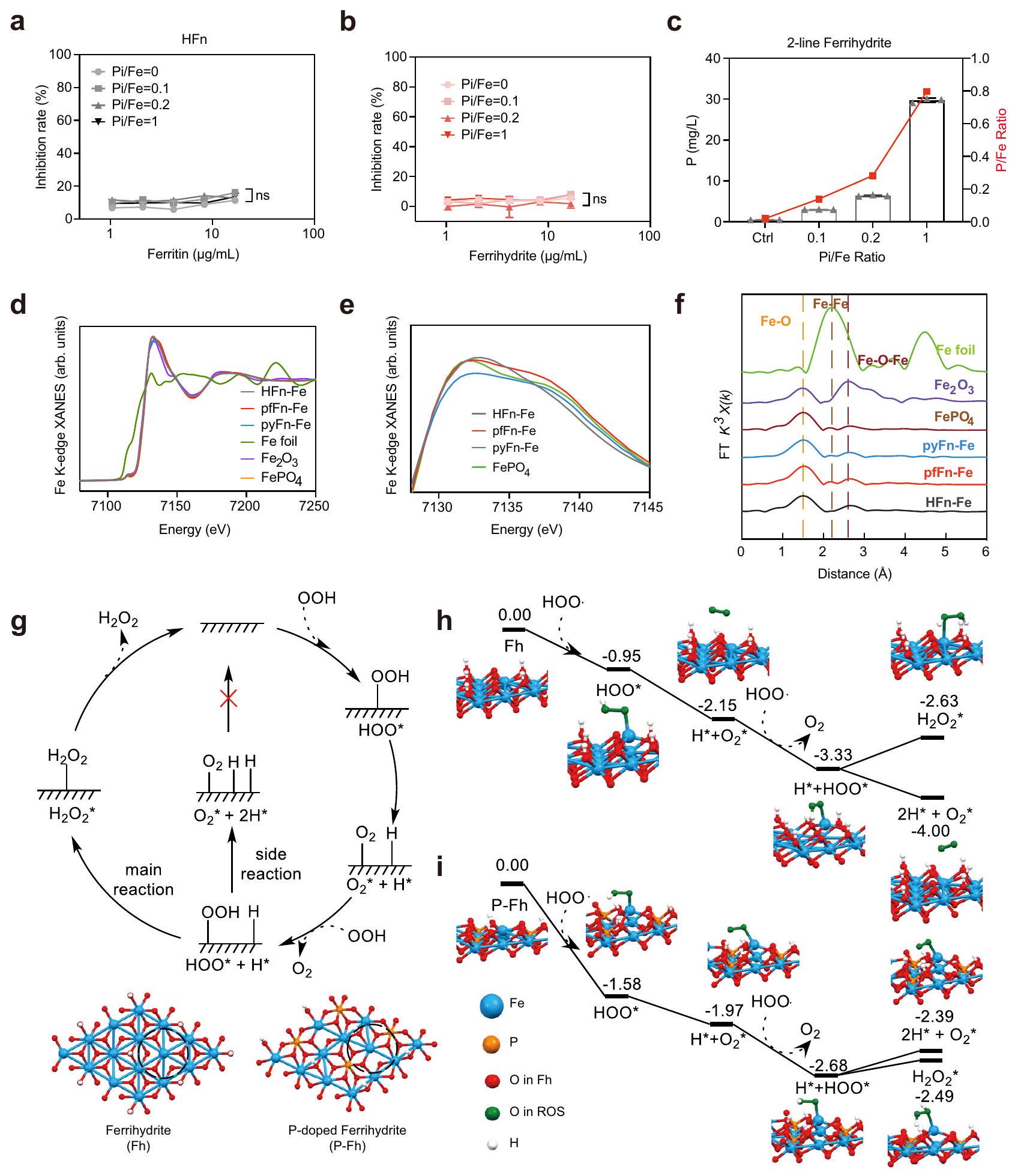
نظرًا لأن النشاط الشبيه بـ SOD جاء من النواة الحديدية وكان مرتبطًا إيجابيًا بمحتوى الحديد في نفس الفيريتينات، تم استخدام مطيافية الكتلة مع الاقتران بالتحليل الطيفي بالتحفيز الكهربائي (ICP-MS) لفحص العناصر المعدنية الرئيسية في أربعة فيريتينات نموذجية لتحديد ما إذا كانت الفروقات في نشاط الفيريتينات المختلفة ناتجة عن محتويات الحديد المختلفة. بشكل غير متوقع، لم تتوافق محتويات الحديد في الفيريتينات المختلفة مع أنشطتها الشبيهة بـ SOD (الشكل 2أ)، كما لم يتوافق محتوى الكبريت، وهو عنصر موجود عادة في البلورات الفيريتينية (الشكل 2ب). ومن المثير للاهتمام أن نسب P/Fe للنوى الحديدية كانت مرتبطة ارتباطًا وثيقًا بأنشطة SOD الشبيهة في الفيريتينات المختلفة (الشكل 2ج)، وهو ما يتماشى مع التقارير السابقة التي أفادت بأن الفيريتينات من البكتيريا أظهرت محتوى فوسفات أعلى من الفيريتينات الثديية.. بعد ذلك، للتحقق بشكل أكبر من دور محتوى الحديد، تم إعادة تكوين نواة الفيريتين في المختبر بعد تنقيتها لإنتاج فيريتينات تحتوي فقط على عنصر الحديد (الشكل 2d). كما هو متوقع، لم يظهر الفيريتين المعاد تكوينه HFn أي اختلاف كبير عن الفيريتين المعدني الحيوي الذي يمتلك نسبة P/Fe منخفضة (الشكل 2e)، بينما أظهرت الفيريتينات الأثرية المعاد تكوينها نشاطًا أقل بكثير من تلك المعدنية الحيوية (الشكل 2f)، مما يشير إلى أن الحديد لم يكن العامل الوحيد الذي يحدد نشاط SOD-like. بعد ذلك، للتحقق من تأثير نسبة P/Fe في الأنشطة، تم إجراء تجربة إعادة التكوين مرة أخرى باستخدام ملح الحديد والفوسفات غير العضوي المتدرج (Pi). كما هو موضح في الشكل التوضيحي 6a، فإن إضافة الفوسفات خففت بشكل واضح من لون الفيريتينات، مما يشير بشكل أساسي إلى أنه تم دمجه في النواة الحديدية. بالإضافة إلى ذلك، تم اكتشاف باستخدام ICP-MS أن الحديد المدمج زاد في البداية مع إضافة الفوسفات، على الأرجح بسبب أن أكسدة الحديد الثنائي في مركز الفيروكسي داز قد تسهلت في وجود الفوسفات.، ثم تم إسقاطه عندما كانت تركيزات الفوسفات مرتفعة بما يكفي لتشكيل ترسب فوسفات الحديد الثلاثي خارج الفيريتين (الشكل التوضيحي 6ب). من المهم أن الفيريتين المعاد تكوينه مع الحديد الثنائي والفوسفات أظهر نشاطًا يشبه نشاط إنزيم سوبر أكسيد ديسموتاز أعلى بكثير من تلك التي لا تحتوي على فوسفات، وكان النشاط يظهر ارتباطًا إيجابيًا معنسبة في إما HFn أو الفيريتينات الأثرية (الشكل 2g والشكل التكميلية 6c-g)، مما يثبت الدور الحاسم لـالنسب. تشير النتائج أعلاه إلى أن الفرق في نشاط SOD-like للفيريتينات ناتج عن تباين الـنسبة، ونواة الحديد ذات نسبة أعلىالنسبة تظهر نشاطًا شبيهًا بـ SOD أعلى.
الجهد السطحي لمحور الثلاثي للفيريتين يؤثر على نسبة الحديد/الفوسفات في النواة
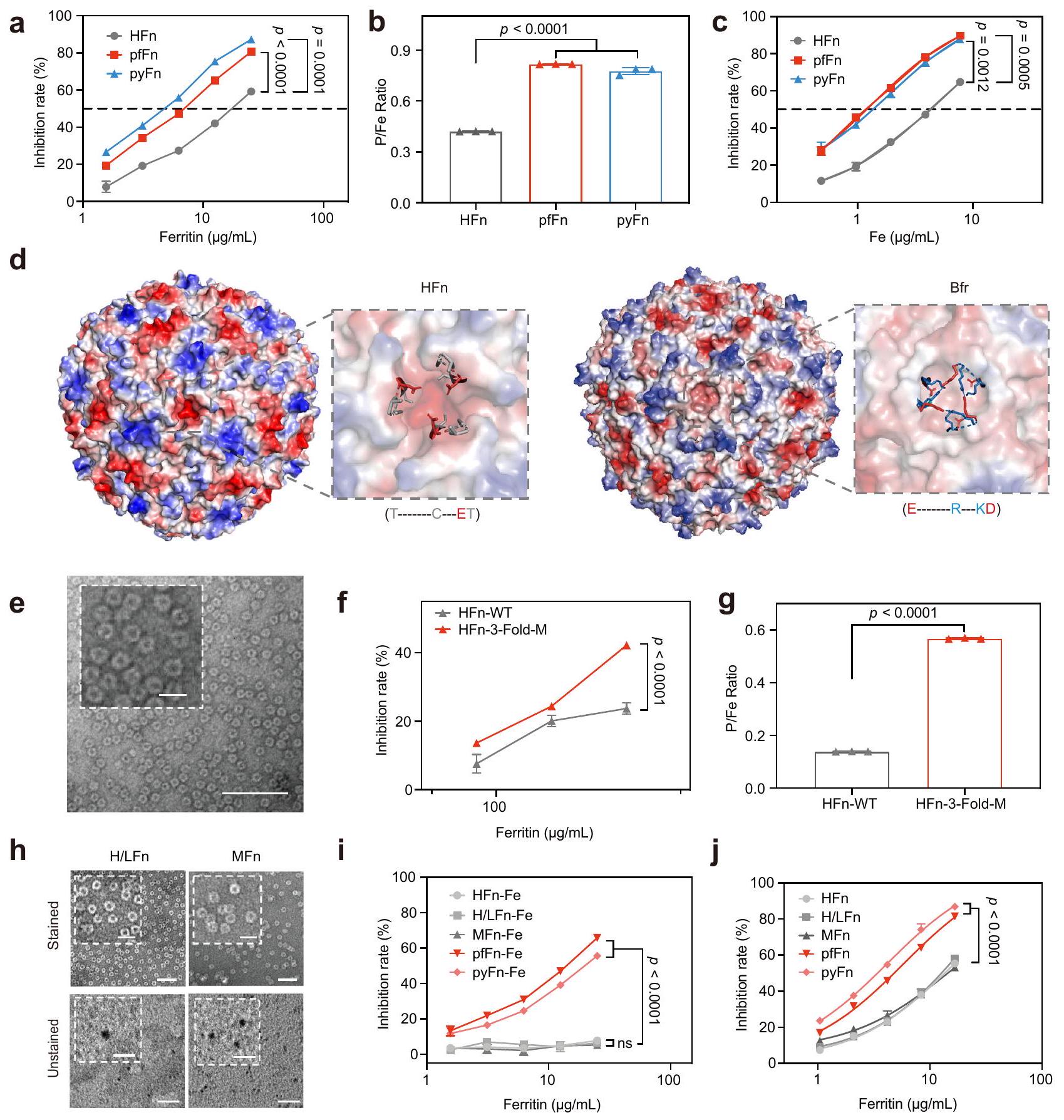
على الرغم من أنه تم الإبلاغ عن أن تشكيل نواة الفيريتين تأثر بالبيئة في السيتوبلازمكانت الفيريتينات التي تم التعبير عنها في دراساتنا منتجة تحت نفس الظروف واحتوت على نوى حديدية مختلفة، مما يشير إلى أن هيكل الفيريتين يشكل نواة الحديد. لاستكشاف التفاعلات بشكل أكبر، تم إعادة تركيب ثلاثة فيريتينات ذات نشاط مختلف بشكل ملحوظ يشبه نشاط SOD، وهي HFn، بالإضافة إلى pfFn الأركي والـ pyFn، في نفس المحلول (نسبة مولارية من الفوسفات/الحديد = 1:1). من الجدير بالذكر أن نشاط HFn المعاد تركيبه كان أقل بشكل واضح من نشاط pfFn وpyFn (الشكل 3أ). وبما يتماشى مع تغيير النشاط، كانت نسبة P/Fe لـ HFn المعاد تركيبه أيضًا أقل من تلك الخاصة بـ pfFn وpyFn (الشكل 3ب). بالإضافة إلى ذلك، نظرًا لأن HFn استوعبت كمية أقل من الحديد مقارنة بـ pfFn وpyFn، تمت مقارنة أنشطة الفيريتينات المعاد تركيبها والفيريتينات المعدنية الحيوية بنفس كمية الحديد لاستبعاد التأثير المحتمل لتغير الحديد. كما هو موضح في الشكل 3ج والشكل التكميلية 7أ، أظهرت أنشطتهم نفس الاتجاه، مما يدل على أن الاختلافات بين هذه الفيريتينات في نشاط SOD-like ناتجة عن اختلافها في هياكل البروتين. ومن المثير للاهتمام، أن الأعمال السابقة حول هياكل البكتيريوفيريتين وجدت أن الأنيون الكبريتي موجود في مدخل المحور الثلاثي لـ البكتيريوفيريتين المغمور بالفوسفات من E. coli.أو في منتصف المحور الثلاثي المغمور بالحديدبكتيريوفيريتين أيروجينوزا. والأهم من ذلك، عندما تم نقع Feتم نقع بكتيريوفيريتين Pseudomonas aeruginosa مرة أخرى في محلول الحديد الثنائي، وستظهر هذه الأنيون الكبريتات في التجويف الداخلي للبكتيريوفيريتين، مما يشير إلى أن المحور الثلاثي قد يكون الموقع لعملية إدخال الأنيون أثناء ترسيب نواة الحديد.هيكليًا، كانت هناك أحماض أمينية كارهة للماء وأخرى قطبية وفتحة ثلاثية أوسع نسبيًا في الفيريتين البكتيري وFTn، مما قد يؤدي إلى نسبة عالية من الفوسفات/الحديد في نوى الفيريتين البكتيري.. دعمًا للفكرة، فإن الجهد الكهربائي السطحي . كان محور Bfr ثلاثي الطي أقل سلبية من محور HFn (الشكل 3d)، مما يسهل على ما يبدو عملية إدخال الأنيونات. لفهم تأثير قشرة الفيريتين على النواة الحديدية بشكل أفضل، قمنا بتحوير الأحماض الأمينية النسبية في محور HFn ثلاثي الطي لتكون مشابهة لتلك الخاصة بـ. بفر Pseudomonas aeruginosa لزيادة تقاربها للفوسفات. أظهرت التحليلات باستخدام TEM، وطيف DLS، وSDS- وPAGE الأصلية (الشكل 3e، الشكل التكميلية 7b-d) أن الطفرات لم يكن لها تأثير على الهيكل الكامل لـ HFn. ومع ذلك، كانت نشاط SOD الشبيه بالطفراتفولد-مكان أعلى من نوع HFn البري (HFn-WT)، وزادت نسبة P/Fe في نواته الحديدية بشكل ملحوظ (الشكل 3f، g والشكل التوضيحي 7e). في تجارب إعادة التركيب، أظهر الطافرة HFn أيضًا نشاطًا أعلى يشبه SOD وقدرة على امتصاص الفوسفات مقارنةً بنوع HFn البري (الشكل التوضيحي 7f)، مما يوضح تأثير بنية البروتين على الأنشطة والمكونات الأساسية.
بالإضافة إلى ذلك، حيث أن الفيريتينات الثديية عادة ما توجد كأشكال هيتروبوليمرية، للتحقق بشكل أكبر من العلاقة بين هياكل الفيريتين وأنشطة SOD-like، تم تحضير وتنقية الفيريتين الطبيعي كأشكال هوموبوليمرية في الفيريتين الميتوكوندري للإنسان (MFn) والفيريتينات الهتروبوليمرية H/L (H/LFn) (الشكل التكميلي 7g، h). وبالمثل، لم يؤثر عملية التمعدن الحيوي على الهيكل وكان هناك تكوين واضح للنواة بعد عملية التخليق الحيوي (الشكل 3h والشكل التكميلي 7i-k). والأهم من ذلك، أن MFn و H/LFn أظهرا أيضًا نشاطًا منخفضًا مثل HFn سواء في التجارب المتمعدنة حيويًا (الشكل 3i) أو التجارب المعاد تكوينها (الشكل 3j والشكل التكميلي 7I). لتوضيح الفرق في النشاط المحدد، قمنا أيضًا بحساب وحدات SOD بناءً على CuZnSOD البشري التجاري أو الطريقة المقدمة من مجموعة اختبار SOD. كما هو موضح في الشكل التكميلي 7m، n، كانت أنشطة SOD للفيريتين البشري المعاد تكوينه قريبة منومع ذلك، يمكن أن تصل الفيريتينات الأركائية إلى.
من النتائج أعلاه، يُستنتج أن هيكل البروتين، وخاصةً إمكانيات المحور الثلاثي، لعب دورًا رئيسيًا في النشاط التحفيزي لنواة الحديد في الفيريتين. للتحقيق بشكل أعمق فيما إذا كانت الفجوة المحتملة بين HFn وPseudomonas aeruginosa Bfr عالمية، بحثنا في قاعدة بيانات PDB وحسبنا 20 فيريتينا بكتيرية و20 فيريتينا حقيقية النواة. من المRemarkable، أظهرت جميع الفيريتينات الحقيقية النواة العشرين إمكانيات أكثر سلبية في المحور الثلاثي مقارنةً بتلك الخاصة بجميع الفيريتينات البكتيرية العشرين (الأشكال التكميلية 8 و9)، مما يكشف عن قاعدة الهيكل بين الفيريتينات البكتيرية والحقيقية النواة.
نسبة الحديد/الفوسفات المختلفة تؤدي إلى تغيير في هيكل النواة الحديدية
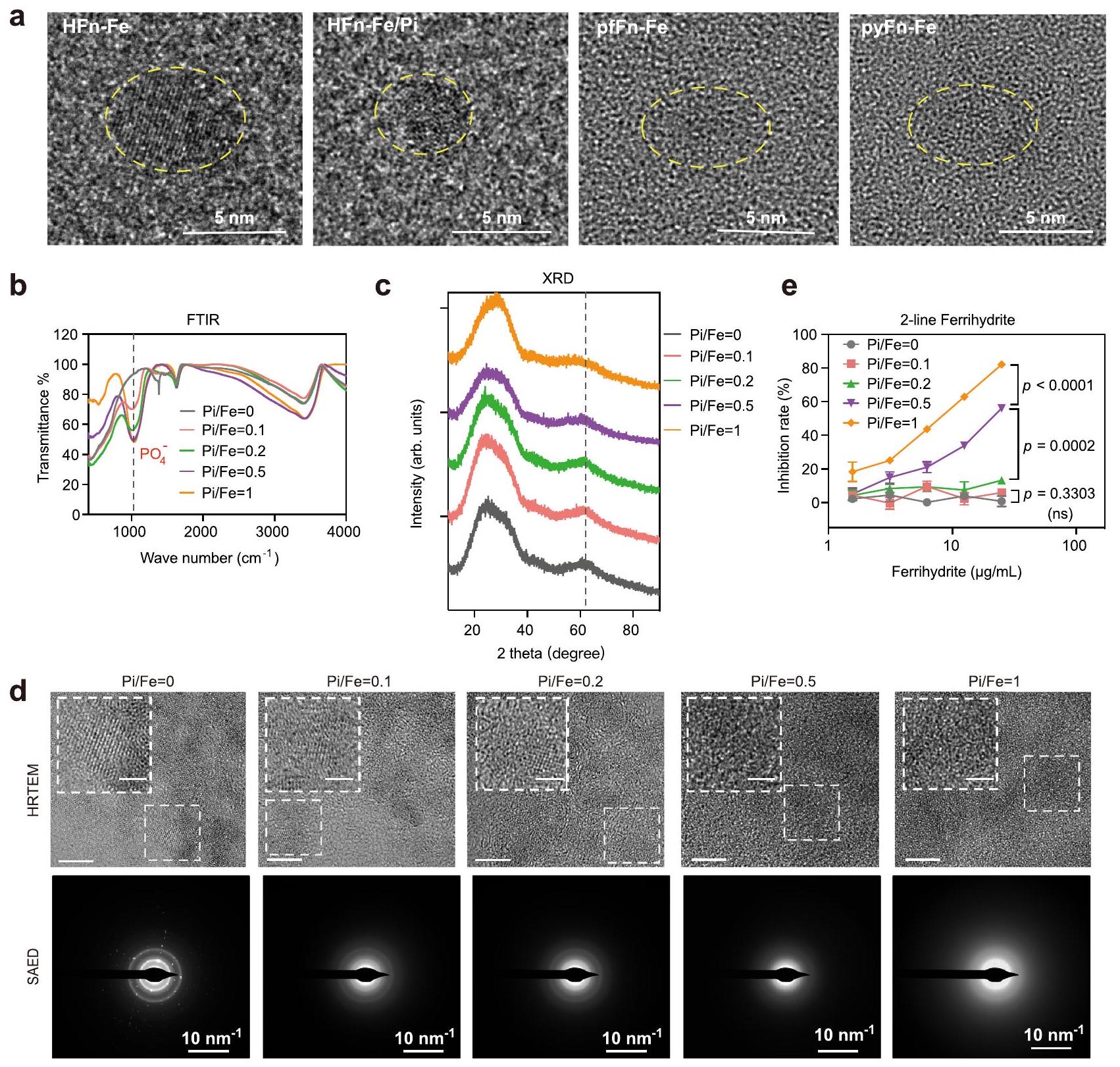
نظرًا لأن الفوسفات أثر بشكل كبير على نشاط النواة الحديدية، فقد استكشفنا الفرق بين النواة الحديدية مع أو بدون فوسفات. ومن المثير للاهتمام أنه تم العثور على أن النواة الحديدية من الفيريتين في الطحال البشري شكلت هيكلًا أكثر ترتيبًا من تلك الموجودة في الليمبت و. أيروجينوس ، مما دفعنا إلى فحص بنية نوى الحديد في الفيريتين ذات النشاط الشبيه بـ SOD المختلف. تحت المجهر الإلكتروني الناقل عالي الدقة (HRTEM)، ظهر نواة الحديد في HFn بشكل أساسي كبلورة ذات مجال واحد، بينما يبدو أنها تحولت إلى بنية متعددة البلورات عندما تم إعادة تكوين الفوسفات في النواة. نواة الحديد في الفيريتين الأركي التي تمتلك مستوى أعلى من ظهر النسبة كمجال غير متبلور تمامًا (الشكل 4 أ والشكل التكميلي 10 أ). تشير هذه البيانات إلى أن دمج الفوسفات في النواة الحديدية يغير هيكلها وفي نفس الوقت يعزز نشاطها التحفيزي.
تم الإبلاغ عن أن الحديد المودع في الفيريتين يكون على شكل أكسيد الحديد الثلاثي أو فيريهايدرايت.لتأكيد تأثير الفوسفات على الشكل والنشاط للنواة الحديدية، تم أولاً تخليق فيريهايدرايت مزدوج الخطوط المدعوم بالفوسفات لمحاكاة نواة الفيريتين التي تحتوي على كميات مختلفة من الفوسفات. كما هو موضح في صور TEM وطيف DLS، عرض الفيريهايدرايت مع أو بدون فوسفات كلاهما هيكل غير متبلور (الأشكال التكميلية 10b و 11a). التغير الملحوظ في اللون وظهور قمة الفوسفات عندفي طيف مطيافية الأشعة تحت الحمراء بواسطة تحويل فورييه (FTIR) أظهر الدمج الناجح للفوسفات (الشكل 4ب والشكل التكميلي 10ب). كما تم استخدام مطيافية الإلكترونات الضوئية بالأشعة السينية (XPS) للكشف عن الحالة الكيميائية والبنية الإلكترونية العامة للعناصر. كما هو موضح في الشكل التكميلي 11ب-ز، ظلت حالة أكسدة الحديد والفوسفور و على التوالي في جميع مواد الفيريهيدرايت. تم إجراء تحليل حيود الأشعة السينية (XRD) لتوصيف الهيكل البلوري لهذه الفيريهيدرايت. كما هو موضح في الشكل 4c، مع إضافة الفوسفات، تضاءل الذروة الثانية في طيف XRD تدريجياً، مما يشير بشكل أساسي إلى فقدان البلورية. تم العثور على نفس الاتجاه من خلال التحليلات باستخدام HRTEM وSAED. تسبب الفوسفات المدعوم في انتقال هيكل الشبكة من بلورة أحادية المجال إلى غير متبلور واختفاء بقع الحيود (الشكل 4d)، والتي كانت متوافقة للغاية مع النتائج في الفيريتينات. مجتمعة، تشير هذه البيانات إلى
الشكل 3 | تأثير بنية البروتين على نشاط شبيه SOD ونسبة الحديد/الفوسفور في الفيريتين. أ نشاط شبيه SOD و (ب)نسبة HFn و pfFn و pyFn المعاد تكوينها في نفس المحلول (نسبة مولارية من ). أنشطة شبيهة بـ SOD لـ HFn المعاد تكوينه و pfFn و pyFn بنفس محتوى الحديد الكلي. د الجهد السطحي للهيكل الكامل والمحور الثلاثي لـ HFn و Bfr (P. aeruginosa) مع الأحماض الأمينية المحيطة بالمحور الثلاثي موضحة في الأسفل؛ الألوان تشير إلى الجهد الكهروستاتيكي، الأحمر: سالب، الأبيض: محايد، الأزرق: موجب. هـ صورة TEM ممثلة ملونة سلبية للثقب الثلاثي المتحور لـ HFn من ثلاث تجارب مستقلة مع نتائج مشابهة؛ الأشكال المضمنة تمثل صورًا مكبرة جزئيًا؛ شريط القياس يشير إلى 50 نانومتر، شريط القياس المضمن يشير إلى 20 نانومتر. و نشاط شبيه بـ SOD و (ز) نسبة P/Fe لـ WT المعدني حيويًا (HFn-WT) و HFn المتحور (HFn-3-fold-M). المجهر الإلكتروني الناقل الملون سلبياً وغير الملون
صور لـ و MFn لثلاث تجارب مستقلة مع نتائج مشابهة؛ تمثل الأشكال المرفقة صورًا مكبرة جزئيًا؛ تشير شريط القياس إلى 50 نانومتر، بينما تشير شريط القياس المرفق إلى 20 نانومتر. i نشاط يشبه SOD للـ HFn البشري المعدني الحيوي، MFn، H/LFn و pfFn، pyFn الأركيائية.صمام بين HFn-Fe وكان 0.8465، وبين HFn-Fe و MFn-Fe كان 0.1187. نشاط يشبه SOD للبشر المعاد تكوينهم HFn و MFn و H/LFn و pfFn الأركيائي و pyFn. جميع البيانات مقدمة كمتوسط ( التجارب المستقلة). تم تقييم الفروق المهمة بواسطة تحليل التباين ثنائي الاتجاه مع اختبار توكي HSD بعد الاختبار لـ (أ، ج، ي، ج)، بواسطة تحليل التباين أحادي الاتجاه مع اختبار توكي HSD بعد الاختبار لـ (ب)، وبواسطة اختبار الطالب غير المقترن ذو الجانبين.-اختبار لـ( ); غير مهم، تُقدم بيانات المصدر كملف بيانات مصدر. أن الفوسفات المدمج في الفيريتين يغير بنية نواة الحديد بطريقة تعتمد على التركيز.
تم فحص نشاط شبيه SOD لهذه الفيريهيدرايتات إما بنفس الجودة أو بنفس محتوى الحديد. كما هو موضح في الشكل 4e. و الشكل التوضيحي الإضافي 11h، كانت النشاط مرتبطًا إيجابيًا معنسبة في هذه المواد. علاوة على ذلك، حيث أن الفيريهيدرايت في الفيريتينات يوجد في تحول من خطين إلى ستة خطوط.تم أيضًا تخليق الفيريهيدرايت ذو الخطوط الستة المدعوم بالفوسفات (المكملات
الشكل 4 | توصيف نواة الحديد في الفيريتين والفيريهايدرايت المدعوم بالفوسفات. أ صور HRTEM لنواة الحديد في الفيريتين مع اختلافاتنسب ثلاث تجارب مستقلة ذات نتائج مشابهة.طيف FTIR لفيريهيدرايت ذو الخطين المدعوم بالفوسفات المتدرج.طيف XRD لفيريهيدرايت ذو الخطين المدعوم بالفوسفات المتدرج. صور HRTEM وSAED لفيريهيدرايت ذو الخطين المدعوم بالفوسفات المتدرج من ثلاث تجارب مستقلة مع نتائج مشابهة؛ الصورة المرفقة.
الأشكال تمثل صورًا مكبرة جزئيًا؛ شريط المقياس في صور HRTEM يشير إلى 5 نانومتر؛ شريط المقياس في الزاوية يشير إلى 2 نانومتر. النشاط الشبيه بـ SOD للفيريهايدريت ذو الخطين المدعوم بالفوسفات؛ البيانات هي متوسط. SD ( تجارب مستقلة)؛ تم تقييم الفرق الكبير بواسطة تحليل التباين ثنائي الاتجاه مع اختبار توكي بعد الاختبار؛ ns: غير دال،تُقدم بيانات المصدر كملف بيانات مصدر.
الشكل 12أ). مشابهًا لتوصيف الفيريهيدرايت ذو الخطين، أظهرت تحليلات FTIR وXRD الدمج الناجح للفوسفات (الشكل التكميلي 12ب، ج). والأهم من ذلك، زادت نشاطات SOD-like أيضًا مع إضافة الفوسفات (الشكل التكميلي 12د)، مما يوضح الدور المهم للفوسفات على هيكل الفيريهيدرايت ونشاطه.
فوسفات مرتبط بذرة الحديد مما يؤدي إلى نشاط شبيه بـ SOD مرتفع
لقد تم إظهار سابقًا أن الفوسفات يوجد إما على سطح النواة الحديدية في الفيريتين الموجود في طحال الحصان أو في داخل الفيريتين البكتيري.، مما يشير إلى المواقع المختلفة للفوسفات في نواة الفيريتين. للتحقيق في التأثير المحدد للفوسفات السطحي، تم إعادة تكوين الفيريتينات بالحديد و”خط 2 تم تعريض الفيريهيدرايت للفوسفات المرتبط بالسطح. على عكس تلك المعاد تكوينها بالفوسفات، لم يتغير لون الفيريتينات المرتبطة بالسطح (الشكل التكميلي 13أ)، مما يوحي بأن الفوسفات السطحي لم يغير بنية نواة الحديد. بالإضافة إلى ذلك، لم يظهر الفيريتين المُعد من محلول الفوسفات المتدرج زيادة في نشاطه الشبيه بنشاط إنزيم سوبر أكسيد ديسموتاز (SOD) (الشكل 5أ والشكل التكميلي 13ب-د). علاوة على ذلك، أظهرت الفيريهيدرايت التي امتصت الفوسفات المتدرج أيضًا تغييرات ضئيلة في أنشطتها الشبيهة بنشاط SOD، على الرغم من أن نسبة P/Fe قد زادت لتقترب من 1 (الشكل 5ب، ج). تشير هذه النتائج إلى أن الفوسفات الممتص على السطح لا يساهم في نشاط الفيريتين الشبيه بنشاط نواة الحديد.
لتحديد موقع الفوسفات في النواة الحديدية بشكل أكبر، تم استخدام مطيافية امتصاص الأشعة السينية عند حافة الحديد (XAFS) لتحليل الحالة الكيميائية وبيئة التنسيق لـ
الشكل 5 | التركيب الذري وتحليل DFT لنوى الحديد في الفيريتين. أ شبيه SOD نشاط HFn المعاد تكوينه مع ربط السطح المتدرج للفوسفات.قيمة لـ مقارنة مع كان، على التوالي. ب نشاط شبيه بـ SOD للفيريهايدريت ذو الخطين مع ربط الفوسفات على السطح المتدرج. الـقيمة لـمقارنة معكان0.4216 على التوالي. البيانات في (أ، ب) هي المتوسط ( تجارب مستقلة)؛ تم تقييم الفرق الكبير بواسطة تحليل التباين ثنائي الاتجاه مع اختبار توكي بعد المقارنة؛ ns: عدم الدلالة،. تحليل ICP-MS لمحتوى الفوسفور ونسب في الفيريهيدرايت ذو الخطين مع ربط الفوسفات على السطح المتدرج؛ البيانات مقدمة كمتوسط SD ( تجارب مستقلة). د، هـ طيف XANES لـ المواد المرجعية النسبية HFn المعدلة حيوياً، pfFn، pyFn.أحجام التحويل فورييه لإشارات EXAFS عند حافة الحديد K التجريبية للـ HFn المعدني الحيوي، pfFn، pyFn، والمواد المرجعية النسبية. g اقترح مسارين تفاعليين محتملين للجذور الحرة مع المنتجات من و (رد الفعل الرئيسي) أو و (تفاعل جانبي) ومراكز التحفيز في الفيريهيدرايت والفيريهيدرايت المدعوم بالفوسفور.ملفات تفاعل مع الهياكل الوسيطة الرئيسية وطاقة التفاعل (eV) للفيريهايدريت. i ملفات تفاعل مع الهياكل الوسيطة الرئيسية وطاقة التفاعل (eV) للفيريهايدريت المدعوم بالفسفور. تم عرض فقط الأجزاء الهيكلية المهمة من أجل الوضوح. تم توفير بيانات المصدر كملف بيانات مصدر. الحديد المعدني الحيوي HFn و pfFn و pyFn. كما هو موضح من خلال ملفات هيكل الامتصاص القريب من حافة الأشعة السينية (XANES)، كانت حواف الامتصاص لجميع الفيريتينات مشابهة لـ و ، مما يشير إلى أن الحديد في الفيريتينات أظهر هيكلًا إلكترونيًا مشابهًا لهذه المواد المشار إليها (الشكل 5d). من ناحية أخرى، اقترح ذروة ما قبل الحافة لجميع الفيريتينات الهيكل الثماني السطوح لذرة الحديد.. علاوة على ذلك، بالمقارنة مع HFn، كان هناك كتف عندفي ملف XANES للفيريتينات الأثرية التي تم تقديمها أيضًا فيالمعيار (الشكل 5e)، مما يشير بشكل أساسي إلى وجود الفوسفور في البيئة المحيطة بذرة الحديد. تم استخدام المنطقة الممتدة من طيف XAFS للحصول على معلومات هيكلية محددة حول الغلاف الذري المحيط بالحديد.كما هو موضح في الشكل 5f، أظهرت جميع الفيريتينات قمة رئيسية عندالذي يُنسب إلىالتنسيق في القشرة الأساسية الأولى، والذي يتوافق بشكل جيد مع وجود و . علاوة على ذلك، كان لدى الثلاثة فيريتينات نفس ذروة عندالذي يُعزى إلى وجودالتنسيق في القشرة الثانية. ومع ذلك، بالمقارنة مع HFn، كان هناك قمة صغيرة عندفي طيفي pfFn و pyFn، الذي اتفق مع الهيكل واقترح أن بعض الفوسفور قد حل محل ذرة الحديد في القشرة الثانية. ثم تم إجراء تحليل ملاءمة منحنى EXAFS الكمي للتحقيق في تكوين التنسيق (الشكل التكميلي 14 أ والجدول التكميلي 1). كان نموذج الملاءمة المناسب لجميع الفيريتينات هو قشرتين تنسيقيتين حيث كانت القشرة الأولى مع كان القطر مشغولاً بست ذرات أكسجين ونحو ذرتين من الحديد تمثلان القشرة الثانية عندالمسافة. بالإضافة إلى ذلك، كان هناك غلاف تنسيق الحديد والفوسفور فيقطر الفيريتينات الأثرية ولكن ليس HFn، مما يشير إلى أن بعض الفوسفور استبدل الحديد في القشرة الثانية. مجتمعة، توضح هذه النتائج أن الاختلاف الرئيسي بين نواة HFn الحيوية ونواة الفيريتين الأثري pfFn وpyFn هو تنسيق الفوسفور في القشرة الثانية من ذرة الحديد، مما يؤثر على القدرة التفاعلية مع السوبر أكسيد.
من أجل معرفة كيف أن الفوسفات المنسق يعزز النشاط الشبيه بـ SOD، قمنا بإجراء حسابات DFT لكشف عمليات التفاعل على الفيرهيت والفيرهيت المدعوم بالفوسفات. لأن أيون السوبرأكسيديتأين بسهولة لتكوينفي حالة مائية، الـاستخدمت كلمة راديكالي لتمثيلفي حسابات DFT. كما هو موضح في الشكل 5g، يتضمن المسار المقترح امتصاصوانحلاله اللاحق لتشكيل و دراستنا السابقة لقد أظهرت أن هذه التفاعلات يمكن أن تحدث بحواجز منخفضة. وبالتالي، اعتبرنا فقط امتصاص الوسائط المهمة على سطح المحفز. تم اعتماد نموذج شريحة لسطح الفيريهيدرايت (001) في هذه الحسابات، حيث تم تحديد الموقع النشط بخط متقطع (الشكل 5g والشكل التكميلي 14b). أظهرت الأشكال 5h و i ملفات الطاقة المقابلة للتفكك على الفيريهيدرايت والفيريهيدرايت المدعوم بالفوسفور، على التوالي. ومن الجدير بالذكر أن امتصاص الأولوما يليهاالانقسام للإفراجتكون طاردة للحرارة، مما يشير إلى أنها يمكن أن تحدث بسهولة على كل من الفيريهيدرايت والفيريهيدرايت المدعوم بالفوسفور. ومع ذلك، فإن تشكيلبعد امتصاص الثانيعلى الفيروهيدرايت هو تفاعلي ماص للحرارة مع زيادة في الطاقة بمقدار 0.7 إلكترون فولت (الشكل 5h). بالمقابل، فإن توليد هو طارد للحرارة مع انخفاض الطاقة بمقدار 0.67 إلكترون فولت (الشكل 5i). من المحتمل أن يكون ذلك بسبب أن السطح (001) من الفيريهيدرايت لديه ميل قوي جداً تجاه الهيدروجين، مما جعل الهيكل الممتص للهيدروجين مستقراً جداً. تشير نتيجة الشكل 5h إلى أن الفيريهيدرايت يميل إلى الأكسدة.لتشكيلبدلاً من تحفيز تفككهامثل SOD. من ناحية أخرى، تشكيلكان أكثر ملاءمة من الناحية الطاقية منبمقدار 0.29 إلكترون فولت في الفيريهيدرايت المدعوم بالفوسفور (الشكل 5i)، مما يشير إلى أن الفيريهيدرايت المدعوم بالفوسفات يمكنه بسهولة تحفيز تفكك السوبرأكسيد. تتوافق هذه النتائج من نظرية الكثافة الوظيفية مع التجربة التي تظهر أن الفيريهيدرايت لديه نشاط تحفيزي شبيه بنشاط إنزيم سوبر أكسيد ديسموتاز بشكل متفوق من خلال إضافة الفوسفات.
الميزة الحفازة لنشاط شبيه SOD للفيريتينات
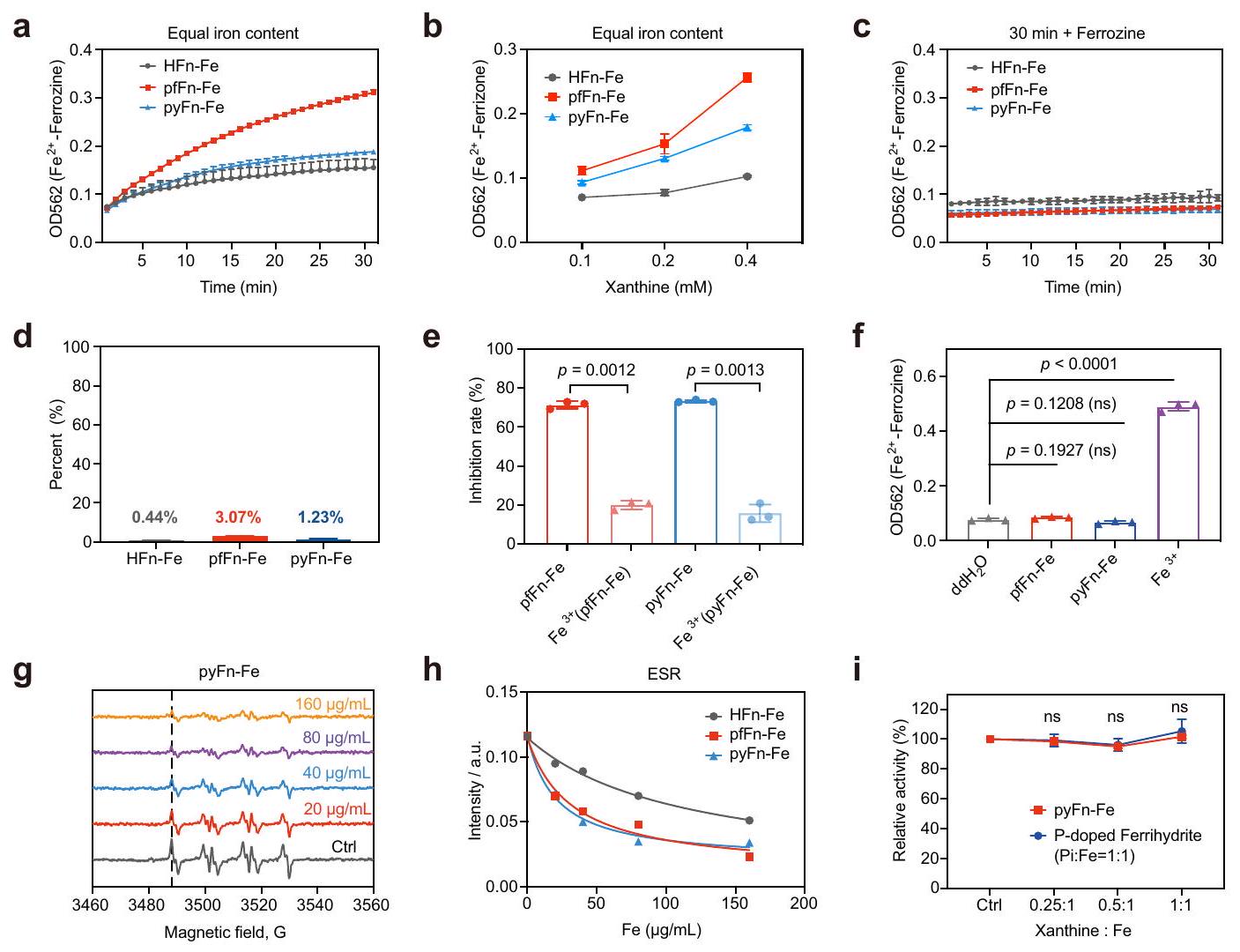
من أجل التحقق من الدور التحفيزي لنواة الحديد في الفيريتين، تم فحص عمليات التفاعل مع السوبرأكسيد لاحقًا. أولاً، تم تتبع المنتج الوسيط أيون الحديد الثنائي بواسطة الخالب المحدد فيروزين. كما هو موضح في الشكل 6أ، تم الكشف عن أيون الحديد الثنائي خلال تفاعل الفيريتينات مع زانثين أوكسيداز (XOD)/زانثين (Xan) وأظهر ارتباطًا إيجابيًا مع مستويات الزانثين (الشكل 6ب). علاوة على ذلك، لم يتم إنتاج أيون الحديد الثنائي بدون XOD أو زانثين، مما يشير إلى أن أيون الحديد الثنائي جاء بشكل رئيسي من الاختزال بواسطة السوبرأكسيد (الشكل التكميلي 15أ-ج). ومع ذلك، أصبح أيون الحديد الثنائي غير قابل للكشف عندما أضيف الفيروزين بعد التفاعل، على الأرجح بسبب إعادة أكسدة أيونات الحديد الثنائي (الشكل 6ج). كما دعمت هذه النتائج تقارير سابقة تفيد بأن نواة الحديد تبقى في الغالب داخل الفيريتين بعد اختزال الحديد الثلاثي ويمكن أن تعاد أكسدتها لاحقًا.. تم الحصول على نفس النتائج في التجارب مع الفيريهيدرايت المدعوم بالفوسفات، حيث عزز الفوسفات المدعوم قدرة الفيريهيدرايت على التفاعل مع السوبر أوكسيد، ولم يتم الكشف عن أي أيون حديد ثنائي بعد التفاعل (الشكل التكميلي 15د، هـ). لتمييز النشاط الشبيه بـ SOD للفيريتين مع تفاعل هابر-وايس المحفز بأيون الحديد الثلاثي، قمنا بتقدير بناءً على منحنى تركيز الحديد الثنائي القياسي أن الأيونات الحديدية الناتجة تمثل من الحديد في HFn و pfFn و pyFn على التوالي في مسار اختبار نشاط SOD (الشكل 6d). بالمقابل، كانت القدرة على تقليل السوبر أوكسيد لهذه الأيونات الحديدية أقل بكثير من الفيريتينات المقابلة لها (الشكل 6e). علاوة على ذلك، تم التحقيق أيضًا في كفاءة تقليل السوبر أوكسيد لملح الحديد الثلاثي عند نفس مستوى الحديد الكلي. كما هو موضح في الشكل التكميلي 15f، كان نشاط SOD لملح الحديد الثلاثي عند نفس مستوى الحديد الكلي أقل بكثير من الأركيا ولكنه أعلى من مجموعة HFn، مما يشير إلى أن نشاط SOD الشبيه لا ينتج عن التفاعل الأيوني. بالإضافة إلى ذلك، تم تقييم توليد أيونات الحديد الثنائي من الفيريتينات الأركية وملح الحديد الثلاثي عند نفس محتوى الحديد الكلي. كما هو موضح في الشكل التكميلي 15g، مقارنةً بالفيريتينات، أنتج ملح الحديد الثلاثي كمية أكبر بكثير من الحديد الثنائي بينما أظهر في المقابل قدرة منخفضة على تقليل السوبر أوكسيد. علاوة على ذلك، على عكس التفاعل مع الفيريتين، ظل هناك مستوى عالٍ من الحديد الثنائي بعد التفاعل (الشكل 6f)، مما يشير إلى أن الفيريتينات تستنفد السوبر أوكسيد بطريقة مختلفة عن الحديد الثلاثي. ثم تم استخدام الرنين المغناطيسي الإلكتروني (ESR) لتقييم قدرة الفيريتينات على استنفاد السوبر أوكسيد. كما هو متوقع، قللت الفيريتينات بشكل كبير من كمية السوبر أوكسيد بطريقة تعتمد على الجرعة (الشكل 6g والشكل التكميلي 15h، i)، وأظهرت الفيريتينات الأركية قدرة أقوى من HFn في تقليل السوبر أوكسيد (الشكل 6h). من الجدير بالذكر أن تدهور السوبر أوكسيد المحفز بواسطة الفيريتين كان مختلفًا تمامًا عن التفاعل الناتج عن أيونات الحديد الثلاثي، حيث أنتج الأخير جذور هيدروكسيل واضحة (الشكل التكميلي 15j). قمنا أيضًا بتقييم نشاط SOD الشبيه لـ pyFn-Fe و ferrihydrite المدعوم بالفوسفات بعد تفاعلاتها مع كميات متدرجة من الزانثين. كلاهما أظهر نشاط SOD شبيه متسق (الشكل 6i)، ولم تنخفض محتويات الحديد الخاصة بهما مع التفاعل (الشكل التكميلي 15k، l)، مما يثبت مرة أخرى الدور التحفيزي لنواة الحديد. باختصار، أظهرت هذه النتائج القدرة الأقوى على استنفاد السوبر أوكسيد للفيريتين الأركي مقارنةً بـ HFn وأظهرت أن نواة الحديد في الفيريتين تحفز استنفاد السوبر أوكسيد بطريقة مختلفة عن أيونات الحديد الثلاثي.
القدرة المضادة للأكسدة للفيريتين في الإشريكية القولونية المعدلة وراثياً والمتحورة
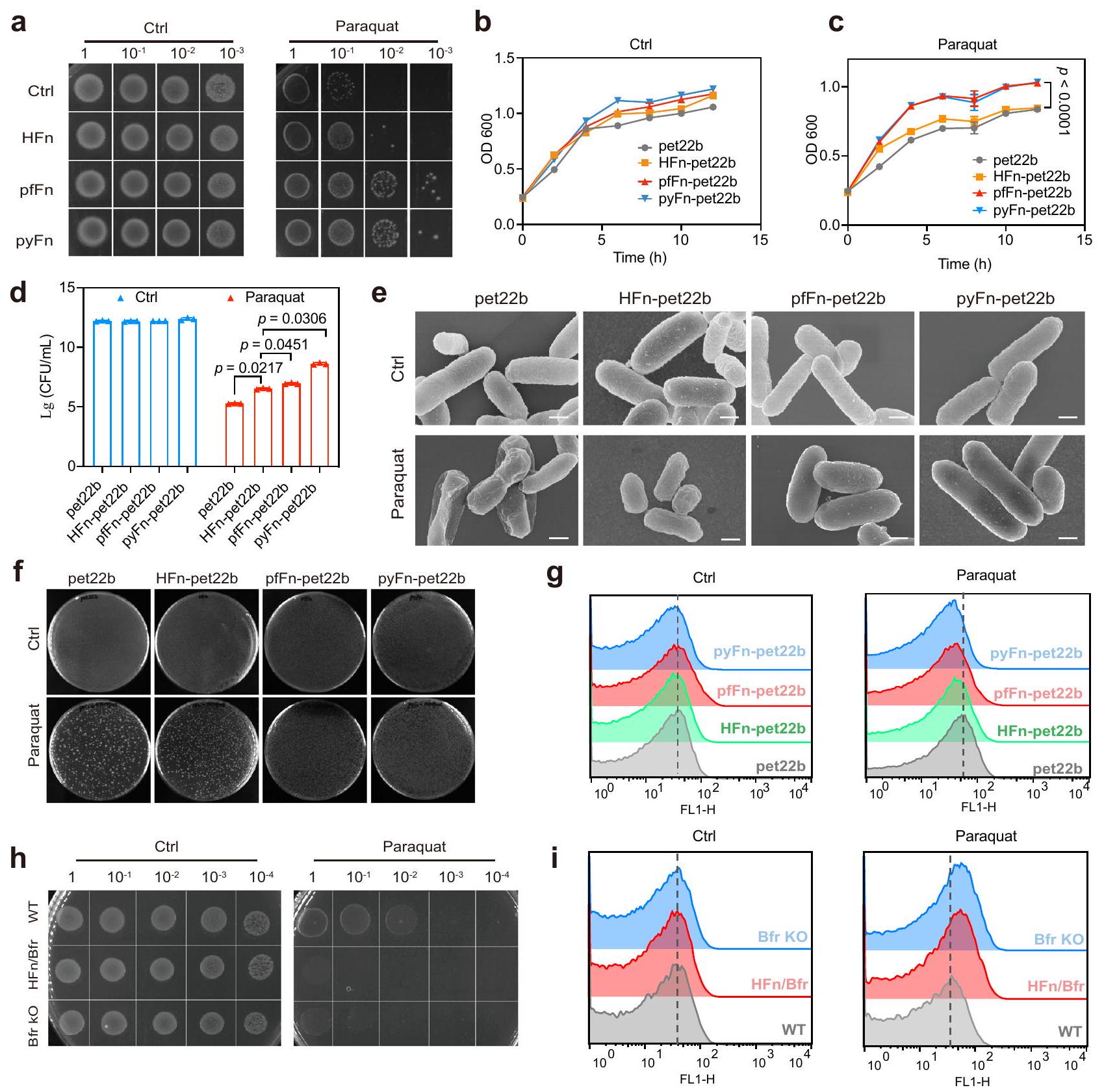
استلهمنا من نشاط الفيريتينات البكتيرية الشبيه بنشاط SOD، وقمنا بالتحقيق في قدرتها المضادة للأكسدة في بكتيريا الإشريكية القولونية المعدلة وراثيًا المعرضة للباراكوين، الذي يمكن أن يحفز إنتاج السوبر أكسيد.كما هو موضح في الشكل التكميلي 16a، فإن خلايا BL21(DE3) المؤهلة التي تم تحويلها مع البلازميدات التي تشفر HFn وpfFn وpyFn أظهرت مستويات مشابهة من الفيريتينات. بعد ذلك، للكشف عن حساسية هذه السلالات للأكسجين الفائق، تم إجراء اختبار النقاط عن طريق إسقاط تخفيفات متسلسلة من الكائنات المعدلة وراثيًا.
الشكل 6 | توصيف عملية التفاعل لنشاط شبيه SOD للفيريتينات. أ منحنى إنتاج الحديدوز-فيروزين في تفاعل الفيريتينات مع XOD-Xan.محتوى إنتاج الحديد الثنائي-فيروزين في تفاعل الفيريتين مع XOD وزانثين التدرج. ج إنتاج الحديد الثنائي-فيروزين في تفاعل الفيريتين وXOD-زانثين عندما تم إضافة الفيروزين بعد 30 دقيقة من أكسيداز الزانثين.نسبة أيونات الحديد الثنائي المتوسطة إلى إجمالي محتوى الحديد في الفيريتينات المختلفة بعد تفاعلها مع XOD-Xan. النشاط الشبيه بـ SOD للفيريتينات الأثرية وأيون الحديد الثلاثي النسبي، الذي تم توليده في تفاعل الفيريتينات الأثرية مع XOD-Xan خلال 30 دقيقة.الإنتاج الحديدي للفيريتين الأركي والملح الحديدي بعد التفاعل مع XOD-Xan في تركيز الحديد في. طيف ESR لـ DMPO/HOO• في وجود
XOD/Xan و pyFn المتدرج؛ التركيز المحدد المشار إليه بمحتوى الحديد. h شدة ESR لـ DMPO/HOO• لمجموعات HFn وpfFn وpyFn. تم استخدام الخط عند 3488.125 G من طيف DMPO/HOO• لتمثيل الشدة. i نشاط شبيه SOD لـ pyFn والفيريهايدريت المدعوم بالفوسفات بعد تفاعلها مع زانثين متدرج. جميع البيانات في (a-f, i) هي متوسط SD ( التجارب المستقلة). تم تقييم الفرق الكبير بواسطة اختبار الطالب غير المقترن ذو الذيلين-اختبار (e)، ومن خلال تحليل التباين الأحادي مع اختبار توكي بعد الاختبار لـ (i). الـالقيمة في (i) مقارنةً مع Ctrl لمجموعة pyFn من اليسار إلى اليمين كانت 0.7532، 0.0742، 0.1398؛ وللفيريهيدرايت المدعوم بالفوسفات كانت 0.9967، 0.7771، 0.5676؛ ns: غير دالة إحصائياً،تُقدم بيانات المصدر كملف بيانات مصدر. البكتيريا على اللوحة التي تحتوي على باراكوين. كما هو موضح في الشكل 7أ، أظهرت البكتيريا المعدلة وراثيًا التي تعبر عن الفيريتينات الأثرية مقاومة أعلى بشكل ملحوظ لباراكوين مقارنة بتلك التي تم تحويلها مع المتجه الفارغ أو المتجه الذي يشفر HFn. بينما نمت جميع البكتيريا بمعدل مشابه بدون باراكوين، نمت البكتيريا المحولة مع البلازميدات التي تشفر pyFn أو pfFn بشكل أسرع بشكل ملحوظ من تلك المحولة مع المتجه الفارغ أو البلازميد الذي يشفر HFn في وجود باراكوين (الشكل 7ب، ج)، مما يوضح تحملًا أعلى للأكسجين الفائق لمجموعات pyFn أو pfFn. تم تقييم حيوية البكتيريا من خلال تجربة تشكيل المستعمرات. كما هو موضح في الشكل 7د، تبقى المزيد من المستعمرات الفردية لمجموعات pyFn أو pfFn مقارنة بالمجموعة التي تحتوي على المتجه الفارغ ومجموعة HFn. وبالمثل، تم عكس حيوية البكتيريا أيضًا من خلال نشاط الديهيدروجيناز للبكتيريا، والذي يمكن أن يقلل من مؤشر WST-8 ويظهر قمة امتصاص محددة عند 450 نانومتر. كما هو موضح في الشكل التكميلي 16ب، أظهرت البكتيريا المحولة مع البلازميدات التي تعبر عن pfFn و pyFn نشاطًا أعلى بشكل ملحوظ للديهيدروجيناز في وجود باراكوين. تم الحصول على نتيجة مماثلة من خلال صورة المجهر الضوئي الماسح بالليزر (CLSM) بعد التلوين المشترك مع SYTO-9 (بكتيريا حية) و بروبيديوم (PI، بكتيريا ميتة) (الشكل التكميلي 16ج). تم تحليل شكل البكتيريا بشكل أكبر بواسطة المجهر الإلكتروني الماسح (SEM). أظهرت البكتيريا في جميع المجموعات المختلفة الشكل النموذجي على شكل قضيب مع جدران خلوية سليمة وناعمة. بينما عانت البكتيريا في مجموعات التحكم من تلف كبير في الغشاء وانكماش الخلايا بعد باراكوين. العلاج، أولئك الذين يعبرون عن HFn قد خففوا من ضرر الباراقوات ولكن لا يزالوا يظهرون انكماشًا معتدلاً. بالمقابل، فإن التعبير عن pfFn و pyFn عكس إلى حد كبير آثار الباراقوات (الشكل 7e). بالإضافة إلى ذلك، فإن إعادة زراعة البكتيريا المعالجة بالباراقوات على أطباق LB قد أكدت أيضًا على ارتفاع نسبة بقاء الخلايا في مجموعات المحولة بـ pfFn و pyFn. بعد ذلك، استخدمنا المجس الفلوري 2′-7′-dichlorofluorescein diacetate (DCFH-DA) للكشف عن مستويات ROS في البكتيريا المعرضة للباراقوات. كما هو متوقع، أظهرت البكتيريا التي تحتوي على pfFn و pyFn مستويات أقل بكثير من ROS مقارنة بتلك المحولة مع المتجه الفارغ أو المتجه الذي يعبر عن HFn (الشكل 7g). هذه النتائج أظهرت أنه يتماشى مع نشاطها الشبيه بـ SOD، فإن الفيريتينات البكتيرية تمارس وظيفة قوية مضادة للأكسدة في البكتيريا وقد يكون لها وظائف فسيولوجية مهمة.
بعد ذلك، للتحقق بشكل أكبر من القدرات المختلفة لتقليل فوق أكسيد الهيدروجين للفيريتينات في البشر والبكتيريا، قمنا باستبدال كل منجين مع الـالجين أو تم إزالتهالجين في سلالة BL21(DE3) بواسطة نظام CRISPR/Cas9 (الشكل التوضيحي التكميلي 16d). كما هو موضح في الشكل التوضيحي التكميلي 16e، بعد قطع المتبرع مع تم تحويل الجين والبلازميدات التي تحتوي على gRNA و Cas9 إلى سلالة BL21، وتم اختيار المستعمرات ذات المقاومة المزدوجة. بعد ذلك، باستخدام البرايمر الأمامي الجينومي والبرايمر العكسي الداخليتم اختيار المستعمرات التي حققت تحرير الجينات بنجاح لعلاج البلازميدات، وأشارت نتائج الرحلان الكهربائي إلى وجود استبدال جيني واضح. وبالمثل، تم إلغاء
الشكل 7 | القدرة المضادة للأكسدة للفيريتين في الإشريكية القولونية المعدلة وراثياً. أ اختبار النقاط للفيريتين المعدل وراثياً المختلف. القولون تحت ضغط الباراكوات من ثلاث تجارب مستقلة مع نتائج مشابهة. Ctrl تشير إلى المجموعات التي لم تتلق علاج الباراكوات. ب، ج منحنى النمو لجراثيم E. coli المتحولة الفيريتين المختلفة مع أو بدون تحفيز الباراكوات؛ البيانات هي متوسط ( تم تقييم الفرق الكبير بواسطة تحليل التباين الثنائي (Two-way ANOVA) مع اختبار توكي (Tukey HSD) بعد الاختبار.كمية تشكيل المستعمرات لبكتيريا الإشريكية القولونية المعدلة وراثياً مع أو بدون تحفيز الباراكوات؛ البيانات هي متوسط SD ( تم تقييم الفرق الكبير بواسطة تحليل التباين الأحادي (One-way ANOVA) في التجارب المستقلة.
مع اختبار توكي بعد التحليل. هـ صور SEM تمثيلية من الفيريتين المعدل وراثيًا. القولونية بعد معالجة الباراقوات لثلاث تجارب مستقلة مع نتائج مشابهة؛ الشريط القياسي المشار إليهصور رقمية للبروتين الفيريتين المعاد تشكيله في بكتيريا الإشريكية القولونية المعدلة وراثياً بعد معالجة الباراقوات لثلاث تجارب مستقلة ذات نتائج مشابهة.مستويات ROS من مختلف سلالات الإشريكية القولونية المحورة جينياً بعد معالجة الباراكوات. اختبار النقاط للسلالة البرية (WT) وسلالة نقص Bfr (Bfr KO) وسلالة استبدال Bfrمن ثلاثة تجارب مستقلة ذات نتائج مشابهة. مستويات ROS من السلالة البرية، وسلالة نقص Bfr، وسلالة الاستبدال بعد معالجة الباراكوات. تم توفير بيانات المصدر كملف بيانات مصدر. تم الحصول على السلالة بواسطة الجزء المتماثل دون وتم التحقق من كفاءة الإزالة بواسطة البرايمرات العلوية والسفلية في الجينوم لـ الجين (الشكل التوضيحي 16f). بعد البناء الناجح لهذين السلالتين، تم مقارنة قدراتهما المضادة للأكسدة ضد الباراقوات. كما هو موضح في اختبار البقع (الشكل 7h)، زاد حذف Bfr بشكل واضح من حساسية السلالة BL21 للأكسجين الفائق، ولم يساهم استبدال HFn في القدرات المضادة للأكسدة. علاوة على ذلك، من خلال قياس مستويات ROS، وجدنا أيضًا مستويات أعلى من ROS في كل منالإقصاء والاستبدال السلالات، مما يدل بشكل أكبر على وظيفة مضادات الأكسدة للفيريتينات ذات النشاط الشبيه بسوبر أكسيد ديسموتاز (الشكل 7i).
نقاش
توزع المعادن الحيوية على نطاق واسع وتلعب دورًا فسيولوجيًا مهمًا في الكائنات الحية المختلفة. ومع ذلك، فإنها تستغل بشكل رئيسي الخصائص المتعلقة بالفيزياء مثل الدعم الميكانيكي والاستشعار البصري أو المغناطيسي في الكائنات الحية. إن ظهور النانوإنزيمات يوحي بأن المعادن الحيوية النانوية قد تعمل كعوامل تحفيز تشبه الإنزيمات. في هذه الدراسة، نحن اخترنا الفيريتينات كنموذج بحث. من خلال مقارنة الأنشطة الشبيهة بـ SOD للفيريتينات من ثلاث ممالك حية، وجدنا لأول مرة العلاقة بين قدرة الفيريتين على التخلص من السوبر أكسيد ونوعياته، والتي تأتي بالترتيب: بدائيات النوى > حقيقيات النوى. من المفهوم أن الاختلاف يعكس الحاجة الملحة لخلايا الكائنات وحيدة الخلية للبقاء في بيئة مؤكسدة، والتي لا تتعرض لها معظم خلايا الكائنات العليا بشكل مباشر.
من خلال تحديد التركيب الأساسي، وجدنا أن الفرق في النشاط كان ناتجًا عن الفوسفات المرتبط بذرة الحديد داخل الفيريتين، الذي تم تشكيله في البداية بواسطة التركيب. في الواقع، وجدت الدراسات السابقة أن محتوى الفوسفات غير العضوي كان أعلى في الفيريتينات الخاصة بالبكتيريا ودموع الرخويات مقارنة بتلك الموجودة في الطحال البشري. و كان لدى الفيريتين من فينلاندii فوسفات مجاور لذرة الحديد في النواة، مقارنةً بفيريتينات الطحال من الخيول، مما يتوافق جيدًا مع دراستنا.من الجدير بالذكر أنه تم التكهن في الأدبيات السابقة بأن هذا التباين في بكتيريا وفيريتين الثدييات كان ناتجًا عن اختلاف محتوى الفوسفات في السيتوبلازم. أشارت تجارب إعادة التركيب والطفرات لدينا إلى أن بنية البروتين كانت أيضًا العامل الحاسم في تباين نواة الحديد.
من المثير للاهتمام معرفة كيف يؤثر الفوسفات في النواة على النشاط الشبيه بالإنزيم. وقد تم الإبلاغ عن أن الفوسفات في نواة الفيريتين يؤثر على الأكسدة وترسيب الحديد الثنائي في طحال الحصان وفيريتين البكتيريا من خلال تعزيز إزاحة الحديد (III) بواسطةفي مركز الفيروكسي داز وتسريع النشاط الأكسدة والاختزال الحديدي على سطح النواة الحديدية. بالإضافة إلى ذلك، وجد وات وآخرون أن الفيريتينات من طحال الحصان وكانت الفينيلاندية تمتلك إمكانيات اختزال مختلفة. بالمقارنة مع الفيريتين من طحال الحصان، و -416 مللي فولت عند ، و 9.0 ، على التوالي)، تظهر الفيريتين من فينلاندii إمكانيات اختزال أكثر سلبية وثباتًا (-420 مللي فولت) عند مستويات pH أعلى، مما يدل على قدرة أقوى على الاحتفاظ بالحديد بواسطةالفيريتين من فينلاندii. بالإضافة إلى ذلك، خلال اختزال الفيريتين من طحال الحصان، هناك تقريباًنُقل إلى النواة لكلمخفض إلىالذي كان غائبًا في الفيريتين البكتيري. يتوافق هذا الظاهرة بشكل جيد مع نتيجة DFT لدينا التي تشير إلى أن النواة الحديدية في HFn تمتلك قدرة أقوى على ربط بروتون الهيدروجين مقارنةً بالنوى الأثرية.أظهرت هذه النتائج أن الفوسفات يؤثر على عملية التكوين وكذلك على قدرة التفاعل للنواة الحديدية.
الأهم من ذلك، فإن الكشف عن النانوإنزيم الطبيعي للفيريتين SOD وآلية التحفيز سيوفر منصة جديدة للتطبيقات الطبية الحيوية. لا يمكن أن يتسبب السوبرأكسيد فقط في إلحاق الضرر المباشر بالإنزيم الذي يحتوي على مركز الحديد-كبريت، مثل الأكونيتاز وسوكينيت ديهيدروجيناز، بل يمكن أن يتحول أيضًا إلى أنواع أخرى من الجذور الحرة السامة للغاية مثل الجذور الهيدروكسيلية والبروكسينيتريت، والتي تضر بمعظم الخلايا والأنسجة. لذا يلعب السوبرأكسيد دورًا مهمًا في العديد من الأمراض، بما في ذلك أضرار الإشعاع، والسكتة الدماغية، والأمراض التنكسية العصبية، والسرطان، وما إلى ذلك.لقد استخدمت الأبحاث السابقة إنزيم السوبر أوكسيد ديسموتاز الطبيعي لعلاج التهاب المفاصل الروماتويدي.التهاب المفاصل العظميوتخفيف أضرار العلاج الإشعاعي والعلاج الكيميائيومع ذلك، بسبب انخفاض امتصاص الخلايا، والمناعة، ونصف العمر القصير، كانت ترجمتها السريرية محدودة للغاية.مع الاستقرار العالي وسهولة تعديل هياكل الفيريتين، فإن اكتشاف النشاط الشبيه بسوبر أكسيد ديسموتاز للنانوزيم الطبيعي للفيريتين لن يسهم فقط في فهم المعادن الحيوية، بل سيقدم أيضًا استراتيجية جديدة للأمراض المرتبطة بالسوبر أكسيد. على سبيل المثال، أثبتت أعمالنا السابقة أن HFn نفسه يمكن أن يستهدف خلايا الورم بدقة وحساسية عالية بناءً على قدرته على الارتباط بمستقبل TfR1.بالإضافة إلى ذلك، تم العثور على أن الإفراط في التعبير عن إنزيم سوبر أوكسيد ديسموتاز يثبط نمو أو غزو خلايا الورم.بالإضافة إلى هاتين الخاصيتين، كان يُعتقد أن النانوإنزيم الفيريتين يمكن استخدامه لتعزيز علاج الأورام. علاوة على ذلك، يمكن تعديل أقفاص الفيريتين بسهولة لتزويدها بخصائص استهداف مختلفة، كما يمكن أن تعمل كحامل جيد للأدوية.، مما وسع بشكل أكبر سيناريوهات تطبيقها. بالإضافة إلى علاج الأورام، فإن نانوإنزيم الفيريتين قد يُستخدم أيضًا كمضاد للأكسدة لكل من التطبيقات في المختبر وفي الجسم (مثل، مضاد للالتهابات ومضاد للشيخوخة). ومع ذلك، فإن اكتشافنا هنا قد حل بشكل أولي آلية التحفيز للنانوزيم الفيريتين، ولا يزال هناك الكثير من العمل اللازم لتحسين هياكلها وأنشطتها قبل تطبيقاتها.
علاوة على ذلك، من الجدير بالذكر أننا قمنا فقط بدراسة الفرق وآلية نشاط إنزيمات مثل SOD للفيريتينات. من المحتمل أن تكون هناك أنشطة إنزيمية أخرى تشبه الفيريتين الطبيعي قد تكون لها أيضًا وظائف فسيولوجية وتطورية مهمة تستحق الاستكشاف. بالإضافة إلى ذلك، تشكل الفيريتين جزءًا صغيرًا فقط من النانوإنزيمات الطبيعية. هناك حاجة إلى مزيد من العمل لاكتشاف وجود أنواع مختلفة من النانوإنزيمات الطبيعية واستكشاف وظائفها الفسيولوجية. نأمل أن تساهم دراساتنا حول النشاط الإنزيمي للفيريتينات في تمهيد الطريق لمزيد من التحقيقات المنهجية.
طرق
المواد
جميع المواد الكيميائية كانت من الدرجة التحليلية. NaCl، مستخلص الخميرة، البيبتون،بيروكسيد الهيدروجين ( ), 3، 3’، 5، 5′-تترا ميثيل بنزيدين (TMB)، 2’، ديكلوروفلورسئين (DCFH-DA)، أمبيسيلين، كاناميسين، سبيكتينوميسين وإيزوبروبيل-تم شراء -D-thiogalactoside (IPTG) من SigmaAldrich (الولايات المتحدة الأمريكية). تم شراء ورق الحديد من Alfa Aesar. تم شراء CuZnSOD البشري من Absin (الصين)، ومجموعة اختبار SOD، و5,5-Dimethyl-1-pyrroline-N-oxide (DMPO)، ومجموعة اختبار صلاحية الميكروبات من DOJINDO (اليابان). تم شراء مجموعة اختبار صلاحية البكتيريا LIVE/DEAD BacLight من Thermo Fisher (الولايات المتحدة الأمريكية).
محاذاة التسلسلات المتعددة وبناء شجرة النشوء
تم الحصول على تسلسلات الأحماض الأمينية لبروتين ربط الحمض النووي من الإشريكية القولونية المنحدرة من سلالة K-12 الفرعية MG1655 (EcDps، NP_415333.1)، بروتين حماية الحمض النووي من سولفولوبوس سولفاتيكي (ssDps، NP_343470.1)، باكتيريوفيريتين من الإشريكية القولونية المنحدرة من سلالة K-12 الفرعية MG1655 (EcBfr، NP_417795.1)، فيريتين غير الهيم من عائلة الإشريكية القولونية (EcFTn، WP_000917208.1)، فيريتين السلسلة الثقيلة من الإنسان (HFn، NP_002023.2)، فيريتين السلسلة الخفيفة من الإنسان (LFn، NP_000137.2)، فيريتين من Pyrococcusfuriosus (pfFn، WP_011011871.1)، فيريتين من Pyrococcus yayanosii (pyFn، WP_013905435.1)، فيريتين من Caenorhabditis elegans (CeFn، NP_504944.2)، فيريتين من Crassostrea gigas (CgFn، NP_001292267.1)، فيريتين من Siniperca chuatsi (XP_044062311)، فيريتين السلسلة الثقيلة من Xenopus laevis (NP_001084057.1)، فيريتين السلسلة الثقيلة من Anolis carolinensis (XP_003221882)، فيريتين السلسلة الثقيلة من Gopherus eugoodei (XP_030416668)، فيريتين السلسلة الثقيلة من Coturnix japonica (XP_015719168)، باكتيريوفيريتين من Parabacteroides goldsteinii سلالة BFG-241 (WP_007659302)، باكتيريوفيريتين من Alistipes onderdonkii السلالة الفرعية vulgaris سلالة 3BBH6 (WP_022332956.1)، باكتيريوفيريتين من Thermus scotoductus (WP_038069286.1)، باكتيريوفيريتين من Streptococcus equinus (WP_074533576.1)، باكتيريوفيريتين من Leptospira weilii (WP_002621798)، باكتيريوفيريتين من Pseudomonas aeruginosa (PaBfr، WP_003092078.1)، فيريتين من Pseudomonas aeruginosa (PaFTn، WP_003093668.1)، فيريتين من Thermus scotoductus (TsFn، WP_038069286.1)، فيريتين من Mycobacterium tuberculosis (MtFn، NP_218358.1)، فيريتين من Candidatus Korarchaeum (WP_012309233.1)، فيريتين من Staphylothermus marinus (WP_011839058)، فيريتين من Thermosphaera aggregans (TaFn، WP_013129153) وفيريتين من Thermococcus barophilus (TbFn، WP_013467695) من قاعدة بيانات NCBI. تم إجراء محاذاة التسلسل المتعددة بواسطة Cluster W وتم تحديد تشابه التسلسل بواسطة برنامج Genedoc (اللون يشير إلى تشابه الأحماض الأمينية، الأحمر: أزرق: رمادي: )، وتم استخدام Mega5 لبناء الشجرة التطورية بطريقة الانضمام الجار، مع إعادة التكرار 1000 مرة.
تحضير البلازميد، التعبير عن الفيريتين البري والمعادن الحيوية وتنقيته
تم اختيار 18 نوعًا مختلفًا من الفيريتين للتنقية واختبار النشاط الشبيه بالإنزيم، بما في ذلك EcDps وssDps وEcFTn وEcBfr وHFn وLFn وpfFn وpyFn وCeFn وCgFn وPaBfr وPaFTn وTsFn وMtFn وTaFn وTbFn وفيريتين الميتوكوندريا البشري (MFn) والبشر.الفيريتين الهتروبوليمر (H/LFn). باستثناء H/LFn، تم استنساخ 17 تسلسل ترميز للفيريتين الآخر إلى بلازميد pet22b مع موقع تقييد Ndel و BamHI. تم بناء متجه التعبير H/LFn عن طريق استنساخ التسلسلين الترميزين في بلازميد petDuet-1 من خلال Ncol/BamHI و Ndel/Xhol. تم تحويل البلازميدات المبنية إلى خلايا BL21(DE3) القابلة للتحول للتعبير عن البروتينات. تم زراعة خلايا التحول القابلة للتحول طوال الليل في وسط LB معأمبيسيلين. تم تحفيز تعبير البروتين عن طريق إضافة 1 مللي مول من الإيزوبروبيل--D-thيogalactoside (IPTG، سيغما) عندما وصلت OD600 إلى 0.6-0.8، ثم تم زراعة البكتيريا المحفزة لمدة 8 ساعات فيتم تصنيع الفيريتينات البيومعدنية عن طريق إضافة ملح الحديد الثنائي المحضر حديثًا إلى وسط ثقافة البكتيريا قبل إضافة IPTG مباشرةً، بحيث يمكن أن يتم تمعدن الحديد الزائد في تجويف الفيريتين أثناء تعبير الفيريتين. تم جمع البكتيريا المستحثة بواسطةتمت عملية الطرد المركزي لمدة 20 دقيقة وتم إعادة تعليق الرواسب لتنقية البروتين. تم الإشارة إلى عمليات التنقية في المنشورات السابقة.باستثناء أنه تم استبدال جميع محاليل الفوسفات المالحة بمحلول تريس أو محلول هيبس.
توصيف الفيريتين البري والفيريتين الحيوي من أنواع مختلفة
TEM: عينات الفيريتين ( ) تم تضمينها في شبكة نحاسية معالجة بواسطة جهاز تنظيف البلازما HPDC32G وتم صبغها بأسيتات اليورانيوم 1% لمدة دقيقة واحدة، ثم تم تصويرها باستخدام مجهر إلكتروني نافذ JEM-1400 80 kV (JEOL، اليابان). DLS: متوسط قطر عينات الفيريتين ( ، تم تحليل ) بواسطة جهاز DynaPro Titan مع عينة ميكروية ذات تحكم في درجة الحرارة من (وايات تكنولوجي، الولايات المتحدة الأمريكية). مطيافية الكتلة بالتحليل الطيفي للبلازما المقترنة بالحث (ICP-MS): تم تحليل محتويات العناصر في عينات الفيريتين من أنواع مختلفة بواسطة جهاز Agilent ICPMS7800.
إزالة نواة الحديد من الفيريتين وهضم الغلاف البروتيني
تم الإشارة إلى طريقة إزالة النواة الحديدية في المنشور السابقتم إضافة الفيريتينات الحيوية الاصطناعية إلى محلول 0.1 م من أسيتات الصوديوم يحتوي على 1% من حمض الثيوغليكوليك، pH 5.5 وتم وضعه فيلمدة 18 ساعة، ثم أضيفت كمية زائدة من 2-2′-بيبيريديل لتخليط الحديد الثنائي. ثم تم الحصول على الأبو-فيريتين بواسطة عمود إزالة الملح G75 (GE Healthcare). تم هضم غلاف البروتين بواسطة البروتياز K: تم إضافة البروتياز K إلى محلول الفيريتين في محلول عازل تريس بتركيز 20 مللي مولار ودرجة حموضة 8.0 وتم وضعه فيلهضم كامل لقشرة الفيريتين.
تركيب الفيريتين المعاد تكوينه بنسب مختلفة من الحديد/الفوسفات
تم تعديل تجربة إعادة التكوين من المنشور السابقتم وضع الفيريتينات في محلول 50 مليمول من MES، pH 6.5، ثم أضيفت محلول كبريتات الأمونيوم الحديدي الطازج إلى محلول الفيريتين بتردد 500/فيريتين كل 10 دقائق عند درجة حرارة الغرفة للأكسدة الحديدية والتبلور. تم تصنيع الفيريتينات بنسب مختلفة من الحديد/الفوسفات عن طريق إضافة فوسفات الصوديوم الهيدروجيني النسبي قبل ملح الحديد. بعد التفاعل، تم طرد الخليط في جهاز الطرد المركزي عندلمدة 10 دقائق لإزالة الرواسب الزائدة. ثم تمت إزالة أيونات الحديد الثلاثي الحرة بواسطة عمود إزالة الملح G75.
اختبار النشاط الشبيه بالإنزيم
اختبار نشاط يشبه سوبر أكسيد ديسموتاز: تم تقييم الأنشطة الشبيهة بسوبر أكسيد ديسموتاز للفيريتيين والفيريهايدرايت باستخدام مجموعة اختبار سوبر أكسيد ديسموتاز (Dojindo، اليابان)، والتي كانت تعتمد على نظام زانثين-زانثين أوكسيداز. يمكن لزانثين أوكسيداز أكسدة الركيزة زانثين. يؤدي ذلك إلى تقليل زانثين أوكسيداز، الذي يقوم بدوره بأكسدة الأكسجين لتوليد السوبر أوكسيد. يمكن أن يتفاعل السوبر أوكسيد مع ملح التترازوليوم القابل للذوبان في الماء (WST-1) في مجموعة الاختبار لتوليد صبغة الفورمازان WST-1، التي تظهر قمة امتصاص محددة عند 450 نانومتر، لذا فإن مستوى السوبر أوكسيد يرتبط بامتصاص OD 450. يمكن لوكيل SOD تقليل السوبر أوكسيد، مما يؤدي إلى انخفاض الامتصاص عند 450 نانومتر. لذلك، تم تعريف نشاط SOD على أنه “معدل التثبيط (%)” لتفاعل السوبر أوكسيد مع WST-1، وتم حسابه كالتالي: [(Ablank 1 – Ablank 3) – (Asample – Ablank 2)] / (Ablank 1 – Ablank 3) × 100، حيث تشير Ablank 1 إلى OD450 لنظام التفاعل أعلاه بدون وكلاء SOD، وتشير Ablank 3 إلى نظام التفاعل بدون وكيل SOD وزانثين أوكسيداز، وتشير Asample إلى نظام التفاعل مع وكلاء SOD، وتشير Ablank 2 إلى نظام التفاعل مع وكلاء SOD ولكن بدون زانثين أوكسيداز. خلال الاختبار الفعلي،تم خلط عينات بتركيزات بروتين محددة تم تحديدها بواسطة اختبار BCA أو محتوى الحديد الذي تم تحديده بواسطة ICP-MS معمحلول العمل WST-1، pH 7.4، ثمتم إضافة زانثين أوكسيداز لبدء التفاعل وتم وضع المزيج فيلمدة 30 دقيقة، ثم تم مراقبة OD450 بواسطة قارئ الميكرو بلايت. تم تعريف وحدة واحدة من SOD على أنها كمية الإنزيم فيمن محلول العينة الذي أعاق تفاعل الاختزال لـ WST-1 مع أيون السوبر أوكسيد بواسطة. وبالمثل، تم حساب وحدات SOD المستندة إلى CuZnSOD التجارية بناءً على IC50 لـ SOD التجارية ووحدات SOD النسبية لها.
اختبار نشاط يشبه البيروكسيداز ويشبه الأكسيداز: تم تقييم النشاط الذي يشبه البيروكسيداز والنشاط الذي يشبه الأكسيداز باستخدام TMB (مذاب في DMSO، ) كمادة أساسية. باختصار، خليط يحتوي على TMB (التركيز النهائي ) و (التركيز النهائي 0.5 م) في محلول 0.2 م من NaAc، pH 4.5 تم إضافته إلى عينات الفيريتين أو الفيريهيدرايت (التركيز النهائي ). تم تقييم النشاط من خلال مراقبة امتصاص الأوكسيداز TMB عند 652 نانومتر على مدى 30 دقيقة. تم تنفيذ النشاط الشبيه بالأكسيداز بنفس الطريقة باستثناء أن كان غائبًا في الخليط.
اختبار نشاط يشبه الكاتالاز: تم تحديد نشاط يشبه الكاتالاز بواسطة إلكترود الأكسجين المحدد على محلل متعدد المعلمات (JPSJ606L، ليشي الصين). تم خلط الفيريتين أو الفيريهيدرايت مع الـمحلول مائي بتركيزات مختلفة في محلول هيفيس 20 مللي مول، pH 7.4، كانت التركيز النهائي للفيريتين أو الفيريهيدرايتوتم تسجيل معدل توليد الأكسجين لمدة دقيقتين.
اختبار توليد اليورات لنظام زانثين أوكسيداز
تم إجراء اختبار توليد اليورات بالإشارة إلى الأدبيات السابقة.باختصار، أكسيداز الزانثين (منتم إضافة أكسيداز الزانثين إلى محلول يحتوي على 20 مللي مول من عازلة هيبس، pH 7.4، تحتوي على زانثين ) عند حجم نهائي قدره 3.0 مل عند تم مراقبة إنتاج اليورات من خلال تغيير الامتصاص عند 290 نانومتر. بالنسبة لمجموعات الفيريتين، تم إضافة الفيريتينات قبل زانثين أوكسيداز بتركيز نهائي من.
تعديل وتنقية الفيريتين الثقيل البشري المتحور ذو المحور الثلاثي
تم استبدال الأحماض الأمينية عند المحور الثلاثي لـ HFn بالأحماض الأمينية النسبية في Bfr من Pseudomonas aeruginosa (T123E، C131R، E135K، T136D). تم استنساخ تسلسل الطافرة المعدلة لـ HFn إلى بلازميد pet22b، وتم تنقية الطافرة HFn كما هو الحال في النوع البري.
تركيب الفيريهيدرايت المخلوط بالفوسفات
تم تحضير الفيريهيدريت المخدر بالفوسفات ذو الخطين أو الستة خطوط بطريقة معدلة من منشور سابق.. تم إعداد جميع الحلول بواسطة مع مقاومة كهربائية قدرهافيريهيدرايت ذو خطين و فيريهيدرايت ذو خطين مشبع بالفوسفات:تم إضافتها إلى 500 مل من الماء المقطر أو تلك التي تحتوي على نسبةتحت التحريك القوي، ثم أضيف 1 م NaOH إلى المحلول مع التحريك المستمر لضبط الرقم الهيدروجيني إلى 7-8. ثم تم طرد الفيريهيدرايتات في جهاز الطرد المركزي عندلـ
20 دقيقة لترسيب الفيريهيدرايت وغسلها بالماء المقطر 5 مرات لإزالة الإلكتروليت. ثم تم تجميد الترسيبات للتخزين أو الاستخدام لاحقًا. فيريهيدرايت ذو 6 خطوط وفيريهدرايت مخدر بالفوسفات ذو 6 خطوط: الماء المقطر الذي لا يحتوي على أو 0.215 جرام،تم وضعها فيفرن للتسخين المسبق، ثمتمت إضافة إلى المحلول المسخن مسبقًا تحت التحريك وتمت إعادة الخليط إلى الفرن لـبعد التفاعل، تم نقل المحلول إلى الثلج للتبريد السريع. ثم تم غسيل المحاليل المبردة ضد الماء المقطر لمدة 7 أيام، وتم استبدال الماء المغسول 3 مرات يوميًا. تم تجميد المواد المغسولة بالتجفيف بالتجميد للتخزين أو للاستخدام لاحقًا.
توصيف نواة الحديد في الفيريتين والفيريهايدرايت المدعوم بالفوسفات
تم استخدام مجهر الإلكترون الناقل عالي الدقة (HRTEM، FEI Tecnai G2 F30، الولايات المتحدة الأمريكية) مع حيود الإلكترونات في المنطقة المختارة (SAED) لوصف شكل وهيكل الشبكة البلورية لنواة الحديد في الفيريتين والفيريهايدريت المدعوم بالفوسفات. تم استخدام مطياف الأشعة السينية للأشعة السطحية (XPS، Thermo escalab 250Xi، ألومنيوم أحادي اللون)جهد تسريع الإشعاع 14.8 كيلوفولت، القدرة علىتم تطبيقه لتوصيف تركيبة العنصر وحالة التكافؤ. تم استخدام طيف FTIR (Nicolet IS10، الولايات المتحدة الأمريكية) لتحديد المجموعة الوظيفية المحددة عن طريق المسح في النطاق منفي خطوةتم استخدام جهاز حيود الأشعة السينية (XRD، Brucker D8 advance، ألمانيا) لتوصيف التركيب البلوري للفيريتيين والفيريهايدرايت المدعوم بالفوسفات معالإشعاع في نطاقمع خطوة.
امتصاص الفوسفات السطحي لنواة الحديد في الفيريتين والفيريهايدرايت
تم تعديل تجربة امتصاص سطح الفيريهيدرايت من المنشور السابقتم تعليق الفيريهيدرايت المجفف بالتجميد ذو الخطين في ماء منزوع الأيونات يحتوي على 0.1 م من NaCl عن طريق الموجات فوق الصوتية. تم تحديد محتوى الحديد في محلول الفيريهيدرايت المحضر بواسطة ICP-MS، ثم تم تحضير ملح الفوسفات ذو الكتلة المولية النسبية.نسبة المولات ) تم إضافته إلى محلول الفيريهيدرايت. تم تحريك الخليط لمدة 24 ساعة لضمان الامتصاص الكافي وتجنب الترسيب المحتمل. ثم تم طرد العينات مركزياً عند لمدة 10 دقائق لفصل الفيريهيدرايت الممتص للفوسفات وغسله ثلاث مرات بـ. تم تجميد الترسيب لتحديد محتوى الحديد والفوسفات واختباره لنشاطات شبيهة بـ SOD. تم إجراء تجربة امتصاص سطح نواة الحديد في الفيريتين بنفس الطريقة. أولاً، تم تصنيع الفيريتين المعاد تكوينه بالحديد عن طريق إضافة كبريتات الحديدوز الأمونيوم إلى الفيريتين المختلفة في محلول 50 مليمول من MES، pH 6.5، وتم تنقيته لاحقًا بواسطة عمود إزالة الملح G75. ثم تم إضافة محلول الفوسفات بتركيز مولي نسبي إلى الفيريتين المعاد تكوينه وتمت حضانة الخلائط بشكل إضافي في لمدة 24 ساعة، ثم تم طرد العينات في جهاز الطرد المركزي عندلمدة 10 دقائق وتم تنقيته بواسطة عمود إزالة الملح G75. تم الكشف عن تركيز البروتين بواسطة مجموعة اختبار بروتين BCA وتم اختبار نشاط شبيه SOD.
توصيف إشعاع السنكروترون لنواة الحديد في الفيريتين تم تقييم الحالة الكيميائية للحديد وبيئة التنسيق من خلال هيكل الامتصاص الدقيق للأشعة السينية (XAFS) باستخدام رقائق الحديد.، و كمواد مرجعية. خضعت عينات الفيريتين المعدني الحيوي لعملية غسيل ضد الماء المقطر لإزالة الأملاح. بعد ذلك، تم تجفيف المحاليل التي تم غسلها بالتجميد، وطحنها بدقة، وضغطها إلى كريات متجانسة ملصقة بشريط 3M. تم الحصول على طيف XAFS عند حافة Fe-K في خط الشعاع 1W1B من منشأة الإشعاع المتزامن في بكين (BSRF) في الصين، باستخدام وضع النقل لقياسات العينة والمرجع. تم تحليل بيانات XANES المعيارية عبر ملاءمة المربعات الصغرى باستخدام برنامج IFEFFIT Athena من CARS (التحالف للإشعاع المتقدم).
مصادر من جامعة شيكاغو لتحديد نسبة أنواع الحديد. لتحليل بيانات EXAFS، تم استخدام برنامج IFEFFIT Artemis (أيضًا من CARS)، معتمدين على طور Fe (Im-3m) لأغراض ملاءمة البيانات.
دراسات DFT لنشاط شبيه SOD لفيريهيدريت الفوسفات
تم استخدام نموذج الشريحة لسطح (001) من الفيريهيدرايت للتحقيق في أنشطته التحفيزية لتفكك السوبرأكسيد. تم بناء الفيريهيدرايت المدعوم بالفوسفور الأكثر استقرارًا عن طريق استبدال ذرة حديد واحدة بذرة فوسفور. باستخدام هذه النماذج الشريطية، تم إجراء حسابات DFT من خلال حزمة المحاكاة الأولية فيينا (VASP 5.4.4).تم استخدام حسابات دالة بيردو-بورك-إرنزرهوف، معززة بمصطلح التشتت شبه التجريبي “D3BJ” من غريمي (PBE-D3BJ).للوصف الإلكترونات التكافؤية، تم استخدام مجموعات أساس الموجة المسطحة مع حد طاقة يبلغ 450 إلكترون فولت، بينما وصفت البسودو إمكانيات الموجة المعززة بالمشاريع الإلكترونات الأساسية. تم تعيين معايير التقارب عندللطاقة وللقوة الذرية. تم تحقيق أخذ عينات من منطقة بريلوان باستخداممونكهورست-باكشبكات نقاط k في جميع الحسابات.
الكشف عن الحديد الوسيط واختبار نشاط إعادة التدوير
تم إجراء الكشف عن أيونات الحديد الثنائي الوسيطة وفقًا للطريقة المعدلة من الطريقة السابقة.باختصار، تم خلط الفيريتينات أو الفيريهيدرايت المدعوم بالفوسفات مع الزانثين والفيروزين بنفس محتوى الحديد الكلي.في محلول هيفيس 20 مللي مول، pH 7.4. ثم تمت إضافة زانثين أوكسيداز لبدء التفاعل وتمت مراقبة توليد الحديد الثنائي بواسطةتم عرض الفيروزين الذي أظهر امتصاصًا محددًا عند 562 نانومتر. تم الحصول على منحنى القياسية للحديد الثنائي عن طريق إضافة كبريتات الأمونيوم الحديدية المتدرجة إلى الفيروزين. تم حساب منحنى القياسية بواسطة طريقة الانحدار الخطي. تم إجراء تجربة نشاط إعادة التدوير عن طريق تفاعل الفيريتينات أو الفيريهيدرايت المدعوم بالفوسفات مع زانثين متدرج لمدة 24 ساعة في وجود XOD. ثم تم تسخين مجموعات الفيريتين الأركي.لمدة 15 دقيقة لتغيير طبيعة إنزيم زانثين أوكسيداز، وتم فصل الفيريتينات عن المزيج بواسطة الطرد المركزي وعمود إزالة الملح. وتم فصل الفيريهيدرايت المدعوم بالفوسفات بواسطة الطرد المركزي والغسيل. ثم تم استخدام الفيريتينات المنفصلة والفيريهايدريت المدعوم بالفوسفات لاختبار نشاط مشابه لـ SOD.
رنين دوران الإلكترون
تم استخدام مطيافية رنين دوران الإلكترون (ESR) لفحص قدرة الفيريتينات على التخلص من السوبر أكسيد: تم استخدام أكسيد الزانثين (XOD) والزانثين لتوليد السوبر أكسيد في محلول هيفس 20 مللي مول.تم استخدام DMPO لاحتجاز السوبر أكسيد عن طريق تشكيل DMPO/OOH. تم بدء التفاعل بإضافة XOD.
اختبار البقعة: تم تقييم حساسية البكتيريا المعدلة وراثيًا لمولد السوبر أكسيد باراكوين من خلال اختبار البقعة. تم زراعة السلالات المحولة مع بلازميد فارغ أو ناقلات تعبير معاد تكوينها في وسط LB جديد. عندما وصلت OD600 إلى 0.8، تم تحفيز العينة بواسطة 1 مللي مول من IPTG عندلمدة 8 ساعات، ثم تم تخفيف هذه السلالات بتركيز 10 مرات ورشها على وسط LB الصلب المحتوي على 0.2 مللي مول من الباراكوات لتحليل حساسيتها.
مراقب منحنى النمو: تم زراعة بكتيريا BL21(DE3) المحولة مع البلازميد الفارغ أو المتجهات التعبيرية المعاد تكوينها في وسط LB الطازج الذي يحتوي علىأمبيسيلين حتى وصل OD600 إلى، ثم تم تخفيف اللقاح إلى نفس بواسطة وسط LB الطازج. بعد ذلك، تم إضافة 1 مللي مول من IPTG و 1 مللي مول من باراكوين في نفس الوقت إلى الوسط عندما وصل OD600 إلى 0.2 وتمت زراعة الملقحات بشكل إضافي في 200 دورة في الدقيقة لمدة 12 ساعة. تم تسجيل امتصاص OD600 كل ساعتين.
اختبار أنواع الأكسجين التفاعلية (ROS): تم تحديد مستوى ROS داخل الخلايا بواسطة-ديكلوروفلورسئين داي أستات (DCFH-DA). يمكن أن يتم امتصاص DCFH-DA بواسطة الخلايا حيث يقوم الإستراز الخلوي بقطع مجموعات الأسيتيل، مما يؤدي إلى إنتاج DCFH. يمكن أن تؤكسد الجذور الحرة الناتجة عن الخلايا DCFH إلى DCF، الذي ينبعث منه فلوريسنس أخضر مع طول موجي للإثارة يبلغ 488 نانومتر وطول موجي للإصدار يبلغ 525 نانومتر. تم تقييم مستويات الفلورسنس بواسطة تقنية قياس التدفق الخلوي. يمكن لتقنية قياس التدفق الخلوي تخصيص قناة لخلية الفلورسنس وفقًا لشدة الفلورسنس الخاصة بها. مع زيادة شدة الفلورسنس الخلوية، سيتم توزيع الخلايا في قنوات أعلى (مما يتوافق مع انتقال إلى نطاق أعلى على المحور السيني). خلال الاختبار الفعلي، تم معالجة البكتيريا المتحولة كما في السابق. بعد المعالجة، تم جمع البكتيريا وغسلها بمحلول PBS، pH 7.4. ثم تم إضافة DCFH-DA إلى البكتيريا المعاد تعليقها بتركيز نهائي من وتم حضنه في لمدة 30 دقيقة، ثم تم غسل البكتيريا مرتين بمحلول PBS، pH 7.4 وتم الكشف عنها بواسطة قياس التدفق الخلوي.
اختبار صلاحية البكتيريا: تم التحقق من صلاحية البكتيريا من خلال تجربة تشكيل المستعمرات. باختصار، تم معالجة البكتيريا التي تحتوي على بلازميد مختلف كما هو مذكور أعلاه. بعد معالجة الباراكوات، تم زراعة البكتيريا المعدلة وراثيًا بالتخفيف المناسب وزراعتها فيبين عشية وضحاها، ثم تم حساب أعداد البكتيريا بوحدات تشكيل المستعمرات (CFU).
اختبار نشاط ديهيدروجيناز البكتيريا: تم استخدام مجموعة اختبار قابلية الحياة الميكروبية (Dojindo) للكشف عن قابلية الحياة للبكتيريا من خلال نشاطها في الديهيدروجيناز. تم معالجة البكتيريا التي تحتوي على بلازميد مختلف كما هو مذكور أعلاه. بعد المعالجة،تم نقل inoculum البكتيريا إلى ألواح 96 بئرًا وتم إضافة محلول العمل (الذي يحتوي على WST-8 والوسيط) إلى كل بئر. ثم تم وضع اللوحة فيلـتم تسجيل OD450 بواسطة SpectraMax. بالإضافة إلى ذلك، تم استخدام مجموعة قياس حيوية البكتيريا Living/Dead Baclight لتحديد البكتيريا الحية (SYTO-9، FITC) والميتة (PI، Texas Red) بواسطة المجهر الضوئي الماسح بالليزر (CLSM) وفقًا لمخطوطة المورد.
ملاحظة الشكل: تم استخدام المجهر الإلكتروني الماسح لمراقبة شكل البكتيريا المتحولة. تم جمع البكتيريا المعالجة بالباراكوين وغسلها بمحلول PBS، وتم تثبيت الراسب بواسطةجلوتارالدهيد فيلمدة 8 ساعات. ثم تم غسل العينة الثابتة ثلاث مرات بالمحلول الملحي وتم تجفيفها بشكل إضافي بواسطة سلسلة من خليط الإيثانول/الماء. بعد ذلك، تم تجفيف العينة بواسطة جهاز التجفيف عند النقطة الحرجة الأوتوماتيكي (CPD300) وتمت ملاحظتها بواسطة المجهر الإلكتروني الماسح (SU8010).
طريقة حذف الجين والاستبدال بواسطة CRISPR/Cas9 في BL21(DE3)
التم إجراء حذف الجين والاستبدال وفقًا لـ يانغ وآخرون.، وgRNA لـ تم تصميمه بواسطة موقع CRISPOR. البلازميد pecCas9 الذي يحتوي على-تم تحويل جين RED و Cas9 إلى سلالة BL21(DE3)، وتم اختيار المستعمرات الإيجابية لتحفيز التعبير عن-RED وصنع خلايا كفؤة. بعد ذلك، بلازميد pecgRNA وقطعة متجانسة من Bfrupstream و downstream مع أو بدونتم إدخال الإدخالات عن طريق التحفيز الكهربائي في الخلايا القابلة. تم التحقق من المستعمرات الفردية بواسطة تفاعل البوليميراز المتسلسل للمستعمرات وتسلسل الحمض النووي. أخيرًا، تم استخدام المستعمرات التي تم التحقق منها لعلاج بلازميدات pecgRNA وpecCas عن طريق إضافة الراهاموز والسكر على التوالي. تم إدراج جميع البلازميدات والبرايمرات المستخدمة في هذه الدراسة في الجدول التكميلي 2.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
البيانات التي تدعم نتائج هذه الدراسة متاحة من المؤلفين المراسلين عند الطلب. جميع هياكل PDB هي متاح على قاعدة بيانات PDB (https://www.rcsb.org/). يتم توفير بيانات المصدر للأرقام والأشكال التكميلية كملف بيانات مصدر. يتم توفير بيانات المصدر مع هذه الورقة.
References
Warr, L. N. IMA-CNMNC approved mineral symbols. Mineral. Mag. 85, 291-320 (2021).
Hazen, R. M. et al. Mineral evolution. Am. Mineral. 93, 1693-1720 (2008).
Hazen, R. M. & Ferry, J. M. Mineral evolution: mineralogy in the fourth dimension. Elements 6, 9-12 (2010).
Perez-Aguilar, C. D. & Cuellar-Cruz, M. The formation of crystalline minerals and their role in the origin of life on Earth. Prog. Cryst. Growth Charact. Mater. 68, 100558 (2022).
Ferris, J. P., Hill, A. R., Liu, R. H. & Orgel, L. E. Synthesis of long prebiotic oligomers on mineral surfaces. Nature 381, 59-61 (1996).
Huber, C. & Wachtershauser, G. Activated acetic acid by carbon fixation on (Fe,Ni)S under primordial conditions. Science 276, 245-247 (1997).
Roldan, A. et al. Bio-inspired conversion by iron sulfide catalysts under sustainable conditions. Chem. Commun. 51, 7501-7504 (2015).
Hazen, R. M. & Sholl, D. S. Chiral selection on inorganic crystalline surfaces. Nat. Mater. 2, 367-374 (2003).
Ferris, J. P. Mineral catalysis and prebiotic synthesis: Montmorillonite-catalyzed formation of RNA. Elements 1, 145-149 (2005).
Joshi, P. C., Aldersley, M. F., Delano, J. W. & Ferris, J. P. Mechanism of montmorillonite catalysis in the formation of RNA Oligomers. J. Am. Chem. Soc. 131, 13369-13374 (2009).
Konhauser, K. O. Diversity of bacterial iron mineralization. Earth-Sci. Rev. 43, 91-121 (1998).
He, H., Veneklaas, E. J., Kuo, J. & Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 19, 166-174 (2014).
Nudelman, F. & Sommerdijk, N. A. J. M. Biomineralization as an Inspiration for Materials Chemistry. Angew. Chem.-Int. Ed. 51, 6582-6596 (2012).
Gao, L. et al. Intrinsic peroxidase-like activity of ferromagnetic nanoparticles. Nat. Nanotechnol. 2, 577-583 (2007).
Fan, K. et al. In vivo guiding nitrogen-doped carbon nanozyme for tumor catalytic therapy. Nat. Commun. 9, 1440 (2018).
Yang, Y. et al. Platinum-carbon-integrated nanozymes for enhanced tumor photodynamic and photothermal therapy. Nanoscale 12, 13548-13557 (2020).
Wu, J., Li, S. & Wei, H. Multifunctional nanozymes: enzyme-like catalytic activity combined with magnetism and surface plasmon resonance. Nanoscale Horiz. 3, 367-382 (2018).
Chen, J. et al. Glucose-oxidase like catalytic mechanism of noble metal nanozymes. Nat. Commun. 12, 3375 (2021).
Singh, N., NaveenKumar, S. K., Geethika, M. & Mugesh, G. A Cerium Vanadate Nanozyme with specific superoxide dismutase activity regulates mitochondrial function and ATP synthesis in neuronal cells. Angew. Chem.-Int. Ed. 60, 3121-3130 (2021).
Ji, S. et al. Matching the kinetics of natural enzymes with a singleatom iron nanozyme. Nat. Catal. 4, 407-417 (2021).
Wu, J. et al. Nanomaterials with enzyme-like characteristics (nanozymes): next-generation artificial enzymes (II). Chem. Soc. Rev. 48, 1004-1076 (2019).
Li, K. et al. Magnetosomes extracted from Magnetospirillum magneticum strain AMB-1 showed enhanced peroxidase-like activity under visible-light irradiation. Enzym. Microb. Technol. 72, 72-78 (2015).
Tang, Z., Wu, H., Zhang, Y., Li, Z. & Lin, Y. Enzyme-mimic activity of ferric nano-core residing in ferritin and its biosensing applications. Anal. Chem. 83, 8611-8616 (2011).
Chi, Z.-L., Yu, G.-H., Kappler, A., Liu, C.-Q. & Gadd, G. M. Fungalmineral interactions modulating intrinsic peroxidase-like activity of iron nanoparticles: implications for the biogeochemical cycles of nutrient elements and attenuation of contaminants. Environ. Sci. Technol. 56, 672-680 (2021).
Andrews, S. C. The Ferritin-like superfamily: Evolution of the biological iron storeman from a rubrerythrin-like ancestor. Biochim. Et. Biophys. Acta-Gen. Subj. 1800, 691-705 (2010).
Yasmin, S., Andrews, S. C., Moore, G. R. & Le Brun, N. E. A new role for heme, facilitating release of iron from the bacterioferritin iron biomineral. J. Biol. Chem. 286, 3473-3483 (2011).
Gauss, G. H. et al. Structure of the DPS-like protein from Sulfolobus solfataricus reveals a bacterioferritin-like dimetal binding site within a DPS-like dodecameric assembly. Biochemistry 45, 10815-10827 (2006).
Wiedenheft, B. et al. An archaeal antioxidant: Characterization of a Dps-like protein from Sulfolobus solfataricus. Proc. Natl. Acad. Sci. USA 102, 10551-10556 (2005).
Uchida, M. et al. The archaeal Dps nanocage targets kidney proximal tubules via glomerular filtration. J. Clin. Investig. 129, 3941-3951 (2019).
Andrews, S. C. et al. Overproduction, purification and characterization of the bacterioferritin of Escherichia-coli and a c-terminally extended variant. Eur. J. Biochem. 213, 329-338 (1993).
Karas, V. O., Westerlaken, I. & Meyer, A. S. Application of an In vitro DNA protection assay to visualize stress mediation properties of the Dps Protein. Jove-J. Visual. Exp. 75, 50390 (2013).
Hudson, A. J. et al. Overproduction, purification and characterization of the Escherichia-coli ferritin. Eur. J. Biochem. 218, 985-995 (1993).
Yu, J. et al. Thermostable iron oxide nanoparticle synthesis within recombinant ferritins from the hyperthermophile Pyrococcus yayanosii CH1. Rsc Adv. 9, 39381-39393 (2019).
Watt, G. D., Frankel, R. B., Papaefthymiou, G. C., Spartalian, K. & Stiefel, E. I. Redox properties and mossbauer-spectroscopy of Azotobacter-vinelandii bacterioferritin. Biochemistry 25, 4330-4336 (1986).
Rohrer, J. S., Islam, Q. T., Watt, G. D., Sayers, D. E. & Theil, E. C. Iron environment in ferritin with large amounts of phosphate, from Azotobacter-vinelandii and horse spleen, analyzed using extended X-ray absorption fine-structure (EXAFS). Biochemistry 29, 259-264 (1990).
Ebrahimi, K. H., Hagedoorn, P.-L. & Hagen, W. R. Phosphate accelerates displacement of by in the ferroxidase center of Pyrococcus furiosus ferritin. Febs Lett. 587, 220-225 (2013).
Garcia-Prieto, A. et al. On the mineral core of ferritin-like proteins: structural and magnetic characterization. Nanoscale 8, 1088-1099 (2016).
Crow, A., Lawson, T. L., Lewin, A., Moore, G. R. & Brun, N. E. L. Structural basis for iron mineralization by bacterioferritin. J. Am. Chem. Soc. 131, 6808-6813 (2009).
Rivera, M. Bacterioferritin: structure, dynamics, and protein-protein interactions at play in iron storage and mobilization. Acc. Chem. Res. 50, 331-340 (2017).
Weeratunga, S. K. et al. Structural studies of Bacterioferritin B from Pseudomonas aeruginosa suggest a gating mechanism for iron uptake via the ferroxidase center. Biochemistry 49, 1160-1175 (2010).
Aronovitz, N., Neeman, M. & Zarivach, R. In Iron Oxides: From Nature to Applications. (eds. Damien, F.) 117-142 (Wiley, 2016).
Mann, S., Bannister, J. V. & Williams, R. J. P. Structure and composition of ferritin cores isolated from human spleen, limpet (patella-
vulgata) hemolymph and bacterial (pseudomonas-aeruginosa) cells. J. Mol. Biol. 188, 225-232 (1986).
Harrison, P. M., Fischbac, F. A., Hoy, T. G. & Haggis, G. H. Ferric oxyhydroxide core of ferritin. Nature 216, 1188 (1967).
Kauko, A. et al. Iron incorporation in Streptococcus suis Dps-like peroxide resistance protein Dpr requires mobility in the ferroxidase center and leads to the formation of a ferrihydrite-like core. J. Mol. Biol. 364, 97-109 (2006).
Towe, K. M. & Bradley, W. F. Mineralogical constitution of colloidal “hydrous ferric oxides”. J. Coll. interface Sci. 24, 384-392 (1967).
Snow, C. L. et al. Ferritin iron mineralization proceeds by different mechanisms in MOPS and imidazole buffers. J. Inorg. Biochem. 105, 972-977 (2011).
Westre, T. E. et al. A Multiplet Analysis of Fe K-Edge 1s 3d Pre-Edge Features of Iron Complexes. J. Am. Chem. Soc. 119, 6297-6314 (1997).
Orikasa, Y. et al. Noncrystalline nanocomposites as a remedy for the low diffusivity of multivalent ions in battery cathodes. Chem. Mater. 32, 1011-1021 (2020).
Shen, X. et al. Mechanisms of oxidase and superoxide dismutationlike activities of gold, silver, platinum, and palladium, and their alloys: a general way to the activation of molecular oxygen. J. Am. Chem. Soc. 137, 15882-15891 (2015).
Wang, Z. et al. Accelerated discovery of superoxide-dismutase nanozymes via high-throughput computational screening. Nat. Commun. 12, 6866 (2021).
Bolann, B. J. & Ulvik, R. J. Decay of superoxide catalyzed by ferritin. Febs Lett. 318, 149-152 (1993).
Watt, G. D., Frankel, R. B. & Papaefthymiou, G. C. Reduction of mammalian ferritin. Proc. Natl. Acad. Sci. USA 82, 3640-3643 (1985).
Hassan, H. M. & Fridovich, I. Superoxide radical and oxygen enhancement of toxicity of paraquat in Escherichia-coli. J. Biol. Chem. 253, 8143-8148 (1978).
Jiang, Y. et al. Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol 81, 2506-2514 (2015).
Cheng, Y. G. & Chasteen, N. D. Role of phosphate in initial iron deposition in apoferritin. Biochemistry 30, 2947-2953 (1991).
Huang, H. Q., Watt, R. K., Frankel, R. B. & Watt, G. D. Role of phosphate in binding to horse spleen holoferritin. Biochemistry 32, 1681-1687 (1993).
Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Heath Sci. 12, 88 (2018).
Goebel, K.-M., Storck, U. & Neurath, F. J. T. L. Intrasynovial orgotein therapy in rheumatoid arthritis. Lancet 317, 1015-1017 (1981).
Lund-Olesen, K. & Menander-Huber, K.B. Intra-articular orgotein therapy in osteoarthritis of the knee. A double-blind, placebocontrolled trial. Arzneimittelforschung 33, 1199-1203 (1983).
Menander-Huber, K., Edsmyr, F. & Huber, W. J. U. R. Orgotein (superoxide dismutase): A drug for the amelioration of radiationinduced side effects: A double-blind, placebo-controlled study in patients with bladder tumours. Urol. Res. 6, 255-257 (1978).
Fan, K. et al. Magnetoferritin nanoparticles for targeting and visualizing tumour tissues. Nat. Nanotechnol. 7, 459-464 (2012).
Teoh, M. L. T., Fitzgerald, M. P., Oberley, L. W. & Domann, F. E. Overexpression of extracellular superoxide dismutase attenuates heparanase expression and inhibits breast carcinoma cell growth and invasion. Cancer Res. 69, 6355-6363 (2009).
Kinnula, V. L. & Crapo, J. D. Superoxide dismutases in malignant cells and human tumors. Free Radic. Biol. Med. 36, 718-744 (2004).
He, J., Fan, K. & Yan, X. Ferritin drug carrier (FDC) for tumor targeting therapy. J. Controlled Release 311-312, 288-300 (2019).
Levi, S. et al. Mechanism of ferritin iron uptake – activity of the H-chain and deletion mapping of the ferro-oxidase site – a study of
iron uptake and ferro-oxidase activity of human-liver, recombinant H-chain ferritins, and of 2 H -chain deletion mutants. J. Biol. Chem. 263, 18086-18092 (1988).
Liu, G. et al. Characterization and surface reactivity of ferrihydrite nanoparticles assembled in ferritin. Langmuir 22, 9313-9321 (2006).
Weiss, R. H. et al. Evaluation of activity of putative superoxide dismutase mimics. Direct analysis by stopped-flow kinetics. J. Biol. Chem. 268, 23049-23054 (1993).
Faulkner, K. M., Liochev, S. I. & Fridovich, I. Stable Mn(III) porphyrins mimic superoxide dismutase in vitro and substitute for it in vivo. J. Biol. Chem. 269, 23471-23476 (1994).
Durot, S. et al. Series of Mn Complexes Based on N-Centered Ligands and Superoxide – Reactivity in an Anhydrous Medium and SOD-Like Activity in an Aqueous Medium Correlated to Mnll/Mnlll Redox Potentials. Eur. J. Inorg. Chem. 2005, 3513-3523 (2005).
Policar, C. In Redox-Active Therapeutics (eds. Batinić-Haberle, I., Rebouças, J. S. & Spasojević, I.) 125-164 (Springer Publishing, 2016).
Kim, J., Li, W., Philips, B. L. & Grey, C. P. Phosphate adsorption on the iron oxyhydroxides goethite (alpha-FeOOH), akaganeite (beta, and lepidocrocite (gamma-FeOOH): a P-31 NMR Study. Energy Environ. Sci. 4, 4298-4305 (2011).
Kresse, G. & Furthmüller, J. Efficiency of ab-initio total energy calculations for metals and semiconductors using a plane-wave basis set. Comput. Mater. Sci. 6, 15-50 (1996).
Grimme, S. et al. A consistent and accurateab initioparametrization of density functional dispersion correction (DFT-D) for the 94 elements H-Pu. J. Chem. Phys. 132, 154104 (2010).
Monkhorst, H.J. & Pack, J.D. Special points for Brillouin-zone integrations. Phys. Rev. B 13, 5188-5192 (1976).
Biemond, P., Swaak, A. J. G., Beindorff, C. M. & Koster, J. F. Superoxide-dependent and superoxide-independent mechanisms of iron mobilization from ferritin by xanthine-oxidase – implications for oxygen-free-radical-induced tissue destruction during ischemia and inflammation. Biochem. J. 239, 169-173 (1986).
Li, Q. et al. A modified pCas/pTargetF system for CRISPR-Cas9assisted genome editing in Escherichia coli. Acta Biochim. Biophys. Sin. 53, 620-627 (2021).
شكر وتقدير
نشكر البروفيسور يالينغ وانغ ومرفق الإشعاع السنكروتروني في بكين (BSRF)، الصين على الدعم الفني لهيكل الامتصاص الدقيق للأشعة السينية. نشكر الدكتور يوجينغ رين على مساعدته الفنية في تحليل بيانات XAFS. مدعوم من المؤسسة الوطنية للعلوم الطبيعية في الصين (82122037، ك.ف.، 81930050، إكس.واي.، 22121003، إكس.واي.)، البرنامج الوطني الرئيسي للبحث والتطوير في الصين (رقم 2021YFC2102900، ك.ف.)، مشروع الأكاديمية الصينية للعلوم للعلماء الشباب في البحث الأساسي (YSBR-089، ك.ف.)، فريق الابتكار بين التخصصات في الأكاديمية الصينية للعلوم (JCTD-2020-08، ك.ف.)، و
مختبر البيومكرومولكولات، الأكاديمية الصينية للعلوم (O3CCSDZ301، ك.ف.).
مساهمات المؤلفين
قام ك.ف. بتصور وتصميم المشروع بالكامل. صمم ل.م. وأجرى التجارب. صمم وأجرى ج.ج.ز.، إكس.ف.ج. حسابات DFT. صمم ونقى ل.م.، ن.ز. الطفرة البروتينية HFn. ساهم ر.ز.، ل.ف. في تنقية البروتين وتحليل البيانات. ساهم ل.ي. في كتابة المخطوطة. أشرف ي.س. على XAFS. كتب ك.ف.، ل.م.، وإكس.ي. المخطوطة بمشاركة جميع المؤلفين.
المصالح المتنافسة
يعلن المؤلفون عدم وجود مصالح متنافسة.
معلومات إضافية
معلومات إضافية النسخة الإلكترونية تحتوي على المواد التكميلية متاحة على https://doi.org/10.1038/s41467-023-44463-w. يجب توجيه المراسلات والطلبات للحصول على المواد إلى شييون يان أو كيلونغ فان.
معلومات مراجعة الأقران تشكر مجلة Nature Communications المراجعين المجهولين على مساهمتهم في مراجعة هذا العمل. يتوفر ملف مراجعة الأقران.
معلومات إعادة الطباعة والتصاريح متاحة على http://www.nature.com/reprints ملاحظة الناشر: تظل شركة سبرينجر ناتشر محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
مختبر الهندسة في الأكاديمية الصينية للعلوم للنانزيم، المختبر الرئيسي للجزيئات الحيوية الكبيرة (CAS)، مركز التميز في الجزيئات الحيوية الكبيرة، معهد البيوفيزياء، الأكاديمية الصينية للعلوم، بكين 100101، الصين.جامعة الأكاديمية الصينية للعلوم، الأكاديمية الصينية للعلوم، بكين 100408، الصين.مختبر العلوم النانوية النظرية والحاسوبية، المركز الوطني للعلوم النانوية والتكنولوجيا في الصين، بكين 100190، الصين.مدرسة سافيد الطبية، جامعة الأكاديمية الصينية للعلوم، بكين 100049، الصين.مركز الأبحاث الإقليمي في الصين، المركز الدولي للهندسة الوراثية والتكنولوجيا الحيوية، تايتشو، جيانغسو 225316، الصين.مختبر مفتاح الأكاديمية الصينية للعلوم لتأثيرات المواد النانوية الحيوية وسلامة النانو ومركز الأكاديمية الصينية للتميز في علوم النانو، المركز الوطني لعلوم النانو والتكنولوجيا، بكين 100190، الصين.مركز نانو زيم الطبي، كلية العلوم الطبية الأساسية، جامعة تشنغتشو، تشنغتشو، هنان 450052، الصين.مختبر نانو زيم في تشونغيوان، تشنغتشو، خنان 451163، الصين. -البريد الإلكتروني: yanxy@ibp.ac.cn; fankelong@ibp.ac.cn
Biominerals, the inorganic minerals of organisms, are known mainly for their physical property-related functions in modern living organisms. Our recent discovery of the enzyme-like activities of nanomaterials, coined as nanozyme, inspires the hypothesis that nano-biominerals might function as enzyme-like catalyzers in cells. Here we report that the iron cores of biogenic ferritins act as natural nanozymes to scavenge superoxide radicals. Through analyzing eighteen representative ferritins from three living kingdoms, we find that the iron core of prokaryote ferritin possesses higher superoxide-diminishing activity than that of eukaryotes. Further investigation reveals that the differences in catalytic capability result from the iron/phosphate ratio changes in the iron core, which is mainly determined by the structures of ferritins. The phosphate in the iron core switches the iron core from single crystalline to amorphous iron phosphate-like structure, resulting in decreased affinity to the hydrogen proton of the ferrihydrite-like core that facilitates its reaction with superoxide in a manner different from that of ferric ions. Furthermore, overexpression of ferritins with high superoxide-diminishing activities in . coli increases the resistance to superoxide, whereas bacterioferritin knockout or human ferritin knock-in diminishes free radical tolerance, highlighting the physiological antioxidant role of this type of nanozymes.
Inorganic minerals naturally occurring as homogeneous solids with a definite chemical composition, are widely distributed on the earth. Until now, there are almost 5800 different kinds of minerals recorded by the International Mineralogical Association (IMA) . According to Robert M. Hazen, the mineral evolution has experienced 10 partially overlapping stages, from the occurrence of primary chondritic minerals to the phanerozoic biomineralization . Notably, nearly twothirds of all the natural minerals resulted from the transformation of
the earth by living organisms, especially in the period after the “Great Oxidant Event”3, indicating the close relationship between mineral generation and life evolution. Indeed, the minerals not only are believed to participate in the origin of life but also have become integrated components of organisms in most, if not all the living kingdoms . In the prebiotic time, the enzyme architecture was considered too complicated to be available. Inorganic minerals were supposed to catalyze the synthesis and polymerization of the initial
biomolecules . The minerals could catalyze the primary organic synthesis, like the carbon fixation under the primordial environment , and realize the chiral molecule selection on the chiral surface of natural minerals, like the calcite . In addition to the small molecule synthesis and concentration, polymerization of the peptides and RNA molecules could also be catalyzed by the clay minerals such as the layered double hydroxide mineral . In modern living organisms, there are considerable amounts of biominerals in almost all the species, such as the carbonate and magnetite in bacteria , calcium oxalate in plants and bones in animals. However, unlike their catalytic roles in the prebiotic period, the biominerals play mainly physical support and protective roles, such as the supporting role of silicate in diatom and bones in animals .
The catalytic role of the biomineralized materials was little discussed until the discovery of the enzyme-like activity of the inorganic nanomaterials . These nanozymes have since been widely studied in various areas including environmental science, the pharmaceutical industry, and so on . In addition to their intrinsic physical and chemical properties like magnetism and fluorescence , the nanozymes have catalytic activities and reaction kinetics similar to the natural enzymes . Currently, a variety of nanozymes have been developed from the original magnetic particles to thousands of different nanomaterials and even the single-atom nanozyme that could match the natural horseradish peroxidase . These unique properties make nanozyme a preferred enzyme mimic for applications in environmental protection, agriculture, disease diagnosis and treatment .
Interestingly, there have been a few published reports on natural nanozymes from biominerals, including the magnetosome in magnetic bacteria, bio-interface mineral nanozyme in the fungus and horse spleen ferritin , which exhibit peroxidase-like activities in addition to being natural constituents. Furthermore, it was shown that the magnetosome and fungal bio-interface mineral nanozyme played an antioxidant role in protecting their host from the superoxide or hydroxyl radical, indicating that these nanozymes might have an important physiological function. Given the diverse and wide distribution of biominerals, it is conceivable that natural nanozymes are present and functional in multiple species of living organisms. Of note, iron storage protein ferritin exists as a natural nano-structure in almost all living organisms including bacteria, archaea, and eukaryotes . Despite significant differences in their sequences, ferritins from all species store iron in the form of the inorganic iron core, a classical type of inorganic nanomaterials in vivo.
In the present study, we examine 18 ferritins from bacteria, archaea, and eukaryotes for their nanozyme activities and decipher the underlying mechanisms. It is found that, compared to eukaryote ferritins, the prokaryote ferritins exhibit higher superoxide dismutase (SOD)-like activity. Our data demonstrate that the differences in SODlike activity of these ferritins result from the changes of the iron/ phosphate ratio in the iron cores. The archaea and bacteria ferritins possess higher phosphate content in relative to human ferritin, which is determined by their amino acid sequences and structures. Furthermore, extended X-ray absorption fine structure (EXAFS) and density functional theory (DFT) analyses show that the phosphate inner the iron core, not the surface one, changes the hydrogen proton binding ability of the iron core, which affects the reaction ability with the superoxide. More importantly, the ferritins with high SOD-like activities can play an antioxidant role in the psychological condition. These results demonstrate the biogenic ferritin iron cores from different species have distinct abilities to scavenge superoxide radicals. The structures of the ferritins determine the composition of the iron cores that in turn decide the SOD-like activity, illustrating that the enzymelike activity of the natural nano-biomineral is structure-dependent and evolutionary conserved, and may play an important role in maintaining the homeostasis of living organisms.
Results and discussion
Biogenic iron cores of ferritins from distinct organisms exhibiting different SOD-like activities
Based on their structures, ferritins are typically classified into three sub-families, DNA binding protein from starved cells (Dps), bacterioferritin (Bfr), and classical ferritin (FTn) . Among them, Dps with a 12-mer core-shell structure ( 9 nm outer diameter) was distributed in the archaea and bacteria, whereas both Bfr and FTn groups ferritins exist as a larger 24 -mer structure with a 12 nm outer diameter. The FTn as an archetypical ferritin exists in all three domains of life, while the Bfr family ferritins exist mainly in the bacteria and contain a special heme structure in its two-fold axis, which facilitates the electron transfer of the iron corre . In order to systematically compare ferritins with different structures from the three domains of life, eight common ferritins were selected, including Dps from bacteria (E. coli, EcDps) and archaea (S. solfataricus, ssDps), FTn from bacteria (E. coli, EcFTn), archaea (P. furiosus and P. yayanosii, pfFn & pyFn) and eukaryote ( . sapiens, heavy chain ferritin (HFn) & light chain ferritin (LFn)), Bfr from bacteria (E. coli, EcBfr). Firstly, multiple sequence alignments were performed to determine the sequence similarity of these ferritins. As shown in Fig. 1a, the 8 ferritins have overall low amino acid similarities. However, the phylogenetic analysis (Fig. 1b) by the neighbor-joining method revealed that most of these ferritins could be clustered into Dps, Bfr, and FTn families, which illustrated that the ferritins from the same group revealed relatively similar amino acid sequences. Yet, the S. solfataricus Dps was clustered with E. coli Bfr, which was consistent with the previous report that it owned a similar ferroxidase center to . Subsequently, to evaluate the enzyme-like activities of the ferritin iron core, we first biosynthesized the ferritin core by adding the ferrous salt into the culture medium during their expression (Fig. 1c) so that the excess iron could be biomineralized in the ferritins in the form of the inorganic iron core. Then these biomineralized ferritins were purified as previously described . As shown by the SDS-PAGE (Supplementary Fig. 1a), the biosynthetic process had a negligible effect on the ferritin monomers compared with those purified from the basic mediums. The stained transmission electron microscope (TEM) images and dynamic light scattering (DLS) spectra indicated that the biomineralized ferritins maintained their original protein-cage structures (Fig. 1d and Supplementary Fig. 1b, c). More importantly, the distinct red-brown color and the unstained TEM images of the biomineralized ferritins indicated the generation of the natural iron core in the ferritins (Fig. 1d and Supplementary Fig. 1d). However, the human light chain ferritin and the Dps sub-family members exhibit smaller core sizes and little color change, likely due to the lack of the ferroxidase center in the former and the smaller inner diameter of Dps. After their purifications, these ferritins were assessed for common nanozyme activities including SOD-, oxidase-, peroxidase- , and catalase-like activities (Supplementary Fig. 2). Interestingly, the SOD-like activity of these ferritins correlated with their origination. As shown in Fig. 1e and Supplementary Fig. 3a, both native and biosynthetic ferritins exhibited the consistent tendency that the strength of their SOD-like activities ranked in the order prokaryote > eukaryote.
To figure out the source of the SOD-like activities, thioglycolic acid and 2,2′-bipyridyl were then utilized to reduce and remove the iron core in the ferritins. As shown in Fig. If and Supplementary Fig. 3b-d, the SOD-like activities were largely diminished with their core removal, whereas the enzyme-like activity of the ferritins was largely unaffected after digestion of the protein shell with protease K (Fig. 1g and Supplementary Fig. 3e), proving that the iron core was responsible for the activities. Additionally, by adding gradient amounts of ferrous salt to the biosynthesis medium, it was found that the SOD-like activities of these ferritins positively correlated with their iron content (Supplementary Fig. 3f-h). Moreover, to exclude that the superoxide diminishing activity was not caused by the inhibition of the xanthine oxidase, we also detected another
Fig. 1 | Purification, characterization, and SOD-like activity of ferritins from
different organisms. a Multiple sequence alignment of amino acid sequences of different ferritins; the colors referred to the amino acid similarity, blue: , gray: . b Phylogenetic tree of the ferritin amino acid sequences; the numbers on the branch represented the bootstrap value. c Scheme of biosynthesis process of different ferritin iron cores. Representative negative stained and unstained TEM images of biomineralized ferritins of three independent experiments with similar results; inset figures represented partially enlarged images; the scale bar referred to 50 nm and inset scale bar referred to 20 nm . e The SOD-like activity of
biomineralized ferritins with the final protein concentration at ; data are presented as mean SD ( independent experiments). The SOD-like activity of apo-pfFn (iron-removed ferritin), pfFn, and biomineralized pfFn (pfFn-Fe); data are presented as mean SD ( independent experiments); the significant difference was evaluated by the Two-way ANOVA with post-hoc Tukey HSD test. g The SODlike activity of archaeal ferritins after the protease K (ProK) digestion; data are presented as mean SD ( independent experiments). Source data are provided as a Source Data file.
Fig. 2 | The effect of main mineral elements in the ferritin core on the SOD-like activity. SOD-like activity and (a) iron content. (b) S/Fe ratio and (c) P/Fe ratio of ferritins from different organisms; data are presented as mean SD ( independent experiments). d Scheme of the synthesis process of ferritin with reconstituted iron core. The SOD-like activity of the reconstituted, biomineralized
ferritins and ferritins purified from the basic medium (-Fe). g The SOD-like activity of the pfFn reconstituted with gradient phosphate/iron ratios ( ); data in are presented as mean ( independent experiments); the significant difference was evaluated by the Two-way ANOVA with post-hoc Tukey HSD test; ns: not significant, . Source data are provided as a Source Data file.
product-urate generation of the superoxide generation system. As shown in Supplementary Fig. 3i, j, unlike the superoxide, the urate production was not affected apparently by the biomineralized ferritins. Taken together, these results demonstrated that ferritins exhibit species-related superoxide diminishing abilities and that the iron cores are the source of the activity. Importantly, our mechanism study will prove that this activity is not simply caused by the metal ion released from ferritins.
Moreover, as the ferritins chosen here were limited to a few phyla, to further verify the relationship between the activity and sources, we then compared ferritin sequences of different phyla in relative kingdoms. As shown in Supplementary Fig. 4, the ferritin sequences in the animals of different phyla were similar. Nevertheless, there was little amino acid similarity in the ferritin sequences of bacteria and archaea ferritins in different phyla. Subsequently, we chose the other 8 ferritins from Caenorhabditis elegans, Crassostrea gigas, Pseudomonas aeruginosa, Thermus scotoductus, Mycobacterium tuberculosis, Thermosphaera aggregans, and Thermococcus barophilus, which involved different phyla compared with the previous 8 ferritins. Interestingly, after the purification and characterization, the SOD-like activity of these ferritins still correlated with the sources (Supplementary Fig. 5), further confirming the activity rules we found.
The variation in SOD-like activity of ferritin iron cores resulting from differences in the iron/phosphorous ratio
As the SOD-like activity came from the iron core and positively correlated with the iron content in the same ferritins, to determine whether the discrepancies in the activity of various ferritins were due to different iron contents, inductively coupled plasma mass spectrometry (ICP-MS) was utilized to examine the major mineral elements in typical four ferritins. Unexpectedly, the iron contents of different ferritins did not correlate with their SOD-like activities (Fig. 2a), nor did the content of sulfur, an element usually present in the ferritin crystalline (Fig. 2b). Interestingly, the P/Fe ratios of iron cores were closely associated with the SOD-like activities in different ferritins (Fig. 2c), which was consistent with previous reports that ferritins from bacteria exhibited higher phosphate content than mammalian ferritins . Next, to further verify the role of iron content, the ferritin core was reconstituted in vitro after their purification to generate ferritins with the only iron element (Fig. 2d). As except, the reconstituted HFn exhibited no significant difference with the biomineralized one that owned low P/Fe ratio (Fig. 2e), while the reconstituted archaeal ferritins exhibited markedly lower activity than the biomineralized ones (Fig. 2f), indicating that the iron was not the only factor determining the SOD-like activity. Subsequently, to verify the effect of the P/Fe ratio
on the activities, the reconstitution experiment was conducted again with ferrous salt and gradient inorganic phosphate (Pi). As shown by Supplementary Fig. 6a, the addition of phosphate apparently lightened the color of ferritins, primarily indicating that it was incorporated into the iron core. Additionally, it was found by using the ICP-MS that the incorporated iron increased initially with the addition of phosphate, presumably due to the ferrous oxidation in the ferroxidase center could be facilitated in the presence of phosphate , and then dropped when the phosphate concentration was high enough to form the ferric phosphate precipitation outside the ferritin (Supplementary Fig. 6b). Importantly, the ferritin reconstituted with ferrous and phosphate exhibited markedly higher SOD-like activity than those without phosphate, and the activity showed a positive correlation with the ratio in either HFn or archaeal ferritins (Fig. 2g and Supplementary Fig. 6c-g), proving the determining role of the ratios. These results above indicated that the difference in the SOD-like activity of ferritins results from the variance of the ratio, and the iron core with a higher ratio exhibits higher SOD-like activity.
Surface potential of the 3-fold axis of ferritin affecting the iron/ phosphate ratio in the core
Although it has been reported that the formation of ferritin core was affected by the environment in the cytoplasm , ferritins expressed in our studies were produced under the same conditions and contained different iron cores, suggesting that the structure of ferritin shapes up the iron core. To further explore the interactions, three ferritins with markedly different SOD-like activity-namely, HFn, as well as archaeal pfFn and pyFn-were reconstituted in the same buffer (molar ratio of phosphate/Fe=1:1). Noteworthily, the activity of reconstituted HFn was apparently lower than that of pfFn and pyFn (Fig. 3a). Consistent with the alteration of activity, the P/Fe ratio of reconstituted HFn was also lower than that of pfFn and pyFn (Fig. 3b). In addition, as the HFn internalized less iron compared with pfFn and pyFn, the activities of reconstituted ferritins and biomineralized ferritins were compared in the same amount of iron to exclude the potential influence of iron variation. As shown in Fig. 3c and Supplementary Fig. 7a, their activities exhibited the same trend, demonstrating that the differences of these ferritins in SOD-like activity resulted from their discrepancy in protein structures. Interestingly, previous works about the bacterioferritin structures have found that the sulfate anion existed in the entrance of the three-fold axis of the phosphate-soaked E.coli bacterioferritin or in the middle of the three-fold axis of the Fe-soaked . aeruginosa bacterioferritin . More importantly, when Fe-soaked . aeruginosa bacterioferritin was soaked in the ferrous solution again, this anion sulfate would appear in the interior cavity of bacterioferritin, indicating that the 3 -fold axis might be the site for the anion internalization during its iron core deposition . Structurally, there existed hydrophobic and polar amino acids and relatively wider threefold pore in the bacterial Bfr and FTn, which might lead to the high phosphate/iron ratio in bacterial ferritin cores . In support of the notion, the surface electrostatic potential of the . aeruginosa Bfr 3 -fold axis was less negative than that of the HFn (Fig. 3d), which apparently facilitates the anion internalization. To further understand the effect of the ferritin shell on the iron core, we mutated relative amino acids in the 3 -fold axis of HFn to those of . aeruginosa Bfr to increase its affinity for phosphate. Analyses with TEM, DLS spectra, SDS- and native PAGE (Fig. 3e, Supplementary Fig. 7b-d) revealed that the mutations had no effect on the whole HFn structure. However, the SOD-like activity of the mutated Fold-M was higher than that of wild-type HFn (HFn-WT), and the P/Fe ratio of its iron core increased markedly (Fig. 3f, g and Supplementary Fig. 7e). In the reconstitution experiments, the HFn mutant also exhibited higher SOD-like activity and phosphate intake ability than the wild-type HFn (Supplementary Fig. 7f), which illustrated the effect of the protein structure on the core activities and compositions.
In addition, as mammalian ferritins usually exist as heteropolymers, to further verify the correlation of the ferritin structures and SOD-like activities, the natural homopolymers ferritin in humanmitochondrial ferritin (MFn) and H/L heteropolymers ferritins (H/LFn) were biomineralized and purified (Supplementary Fig. 7g, h). Likewise, the biomineralization process did not affect the structure and there was obvious core formation after the biosynthesis process (Fig. 3h and Supplementary Fig. 7i-k). More importantly, the MFn and H/LFn also exhibited low SOD-like activity as HFn either in the biomineralized (Fig. 3i) or the reconstituted experiments (Fig. 3j and Supplementary Fig. 7I). To illustrate the specific activity discrepancy, we also calculate the SOD units based on commercial human CuZnSOD or the method offered by the SOD assay kit. As seen in Supplementary Fig. 7m, n, the SOD activities of reconstituted human ferritin were near Fe, however, the archaea ferritins could arrive at .
From the above results, it is reasoned that protein structure, especially the three-fold axis potential, played a key role in the catalytic activity of the ferritin iron core. To further investigate whether the potential discrepancy of HFn and Pseudomonas aeruginosa Bfr was universal, we searched the PDB database and calculated 20 bacteria ferritins and 20 eukaryote ferritins. Remarkably, all 20 eukaryote ferritins revealed more negative potential in the three-fold axis than those of all the 20 bacteria ferritins (Supplementary Figs. 8 and 9), disclosing the structure rule between the bacteria and eukaryote ferritins.
Different iron/phosphate ratio leading to the structure change of the iron core
As phosphate significantly affected the activity of the iron core, we thus explored the difference of the iron core with or without phosphate. Interestingly, it has been found that the iron core of human spleen ferritin formed a more ordered structure than those from limpet and . aeruginos , which prompted us to examine the structure of the ferritin iron cores with different SOD-like activity. Under highresolution transmission electron microscopy (HRTEM), the iron core in HFn exhibited mostly as single-domain crystallite, whereas, it apparently switched to the polycrystalline structure when phosphate was reconstituted into the core. The iron core of archaeal ferritin that possesses a higher ratio appeared as the completely amorphous domain (Fig. 4a and Supplementary Fig. 10a). These data indicated that the incorporation of phosphate into the iron core changes its structure and at the same time, augments its catalytic activity.
Iron deposited in ferritin was reported to be in the form of ferric oxyhydroxide or ferrihydrite . To further confirm the effect of the phosphate on the morphology and activity of the iron core, phosphate-doped 2-line ferrihydrite was first synthesized to mimic the ferritin iron core containing various amounts of phosphate. As shown in the TEM images and DLS spectra, the ferrihydrite with or without phosphate both displayed the amorphous structure (Supplementary Figs. 10b and 11a). The marked color change and the appearance of phosphate peak at in the Fourier transform infrared spectroscopy (FTIR) spectrum demonstrated the successful incorporation of the phosphate (Fig. 4b and Supplementary Fig. 10b). X-ray photoelectron spectroscopy (XPS) was also deployed to detect the chemical state and the overall electronic structure of elements. As shown in Supplementary Fig. 11b-g, the valence of iron and phosphorus remained and respectively in all the ferrihydrite materials. X-ray diffraction analysis (XRD) was performed to characterize the crystallographic structure of these ferrihydrites. As shown in Fig. 4c, with the addition of the phosphate, the second peak in the XRD spectra diminished gradually, primarily suggesting the loss of crystallinity. The same tendency was found by analyses with HRTEM and SAED. The doped phosphate caused the lattice structure to transit from the single domain crystallite to amorphous and the disappearance of the diffraction spots (Fig. 4d), which were highly consistent with results in ferritins. Taken together, these data indicated
Fig. 3 | Effect of protein structure on SOD-like activity and iron/phosphorous ratio of ferritin. a SOD-like activity and (b) ratio of HFn , pfFn and pyFn reconstituted in the same buffer (molar ratio of ). c SOD-like activities of reconstituted HFn , pfFn and pyFn in the same total iron content. d The surface potential of the whole structure and three-fold axis of the HFn and Bfr (P. aeruginosa) with the amino acids around the three-fold axis marked in the bottom; the colors referred to electrostatic potential, red: negative, white: neutral, blue: positive. e Representative negative stained TEM image of the three-fold pore mutated HFn of three independent experiments with similar results; inset figures represented partially enlarged images; the scale bar referred to 50 nm , inset scale bar referred to 20 nm . f SOD-like activity and (g) P/Fe ratio of the biomineralized WT (HFn-WT) and mutated HFn (HFn-3-fold-M). Negative stained and unstained TEM
images of the and MFn of three independent experiments with similar results; inset figures represented partially enlarged images; the scale bar referred to 50 nm , inset scale bar referred to 20 nm . i SOD-like activity of biomineralized human HFn, MFn, H/LFn and the archaeal pfFn, pyFn. valve between HFn-Fe and was 0.8465, and between HFn-Fe and MFn-Fe was 0.1187. j SOD-like activity of reconstituted human HFn, MFn, H/LFn, and the archaeal pfFn, pyFn. All the data are presented as mean ( independent experiments). The significant differences were evaluated by the Two-way ANOVA with post-hoc Tukey HSD test for (a, c, i, j), by the One-way ANOVA with post-hoc Tukey HSD test for (b), and by Two-sided unpaired Student’s -test for( ); ns: not significant, . Source data are provided as a Source Data file.
that the phosphate incorporated in the ferritin changes the structure of the iron core in a concentration-dependent manner.
The SOD-like activity of these ferrihydrites was then examined in either the same quality or the same iron content. As shown in Fig. 4e
and Supplementary Fig. 11h, the activity positively correlated with the ratio in these materials. Moreover, as the ferrihydrite in the ferritins exists transformation from 2 -line to 6 -line , the phosphatedoped 6 -line ferrihydrite was also synthesized (Supplementary
Fig. 4 | Characterization of the ferritin iron core and phosphate-doped ferrihydrite. a HRTEM images of ferritin iron core with different ratios of three independent experiments with similar results. FTIR spectra of 2-line ferrihydrite doped with gradient phosphate. XRD spectra of 2-line ferrihydrite doped with gradient phosphate. d HRTEM and SAED images of 2-line ferrihydrite doped with gradient phosphate of three independent experiments with similar results; inset
figures represented partially enlarged images; the scale bar in HRTEM images referred to 5 nm ; the inset scale bar referred to 2 nm . e SOD-like activity of the phosphate-doped 2-line ferrihydrite; the data are mean SD ( independent experiments); the significant difference was evaluated by the Two-way ANOVA with post-hoc Tukey HSD test; ns: not significant, . Source data are provided as a Source Data file.
Fig. 12a). Similar to the 2 -line ferrihydrite characterization, the FTIR and XRD analyses illustrated the successful incorporation of the phosphate (Supplementary Fig. 12b, c). More importantly, the SOD-like activity also increased with the addition of the phosphate (Supplementary Fig. 12d), further demonstrating the important role of the phosphate on the ferrihydrite structure and activity.
Phosphate coordinated with the iron atom leading to high SODlike activity
It has been previously shown that phosphate exists on either the surface of the iron core in horse spleen ferritin or in the interior of the bacteria ferritin , which suggests the different locations of the phosphate in the ferritin core. To investigate the specific effect of the surface phosphate, the iron-reconstituted ferritins and the 2 -line
ferrihydrite were exposed to surface-binding phosphate. Unlike those reconstituted with phosphate, the color of the surface binding ferritins did not change (Supplementary Fig. 13a), implying that the surface phosphate has not altered the iron core structure. In addition, the ferritin prepared from the gradient phosphate buffer did not exhibit increased SOD-like activity (Fig. 5a and Supplementary Fig. 13b-d). Additionally, ferrihydrites absorbed gradient phosphate also showed negligible changes in their SOD-like activities, even though the P/Fe ratio has increased close to 1 (Fig. 5b, c). These results indicated that surface-absorbed phosphate does not contribute to the SOD-like activity of the ferritin iron core.
To further define the location of phosphate in the iron core, the Fe K-edge X-ray absorption fine structure (XAFS) spectroscopy was used to analyze the chemical state and coordination environment of the
Fig. 5 | Atomic structure and DFT analysis of ferritin iron cores. a SOD-like
activity of reconstituted HFn with gradient surface binding phosphate. The value of compared with was , respectively. b SOD-like activity of the 2-line ferrihydrite with gradient surface binding phosphate. The value of compared with was , 0.4216 , respectively. The data in (a,b) are mean ( independent experiments); the significant difference was evaluated by the Two-way ANOVA with posthoc Tukey HSD test; ns: non-significance, . c ICP-MS of P content and ratios in the 2 -line ferrihydrite with gradient surface binding phosphate; the data are presented as mean SD ( independent experiments). d, e XANES spectra of
biomineralized HFn , pfFn, pyFn and relative reference materials. Fourier-transformed magnitudes of the experimental Fe K-edge EXAFS signals of biomineralized HFn , pfFn, pyFn, and relative reference materials. g Proposed two possible reaction pathways for superoxide with products of and (main reaction) or and (side reaction) and catalytic centers in Ferrihydrite and P -doped Ferrihydrite. Reaction profiles with key intermediate structures and reaction energies ( eV ) for ferrihydrite. i Reaction profiles with key intermediate structures and reaction energies (eV) for P -doped ferrihydrite. Only important structural fragments were shown for clarity. Source data are provided as a Source Data file.
biomineralized HFn , pfFn and pyFn. As shown by the X-ray absorption near edge structure (XANES) profiles, the absorption edges of all the ferritins were similar to the and , indicating that the Fe in the ferritins exhibited a similar electronic structure as these two referred materials (Fig. 5d). On the other hand, the pre-edge peak of all the ferritins suggested the octahedral structure of the iron atom . Furthermore, compared with that of HFn , there was a shoulder at in the XANES profile of archaeal ferritins that was also presented in the standard (Fig. 5e), primarily suggesting the presence of phosphorus in the surrounding environment of the iron atom. The extended region of the XAFS spectra was used to gain the specific structure information on the atomic shell around the iron . As shown in Fig. 5f, all the ferritins exhibited a major peak at that is attributed to the coordination in the first core shell, which agrees nicely with the presence of and . Moreover, all three ferritins had the same peak at which is attributable to the presence of the coordination in the second shell. However, compared with HFn , there was a minor peak at in the spectrums of pfFn and pyFn , which agreed with the structure and suggested that some phosphorus had replaced the iron atom in the second shell. Quantitative EXAFS curve-fitting analysis was then performed to investigate the coordination configuration (Supplementary Fig. 14a and Supplementary Table 1). The proper fit model for all the ferritins was two coordination shells in which the first shell with diameter was occupied by six oxygen atoms and nearly 2 iron atoms accounted for the second shell at a distance. In addition, there existed the Fe-P coordination shell at diameter for the archaeal ferritins but not the HFn , indicating that some phosphorus replaced the iron in the second shell. Taken together, these results illustrated that the major difference of biosynthetic HFn core and archaeal ferritin pfFn and pyFn core is the phosphorus coordination in the second shell of the iron atom, which likely affects the reaction ability with superoxide.
In order to figure out how the coordinated phosphate promoted the SOD-like activity, we carried out DFT calculations to uncover the reaction processes on ferrihydrite and phosphatedoped ferrihydrite. Because superoxide anion readily protonates to form in aqueous condition , the radical was used to represent in the DFT calculations. As shown in Fig. 5g, the proposed pathway involves the adsorption of and its subsequent disproportionation to form and . Our previous study has shown that these reactions can occur with low barriers. Thus, we considered only the adsorption of important intermediates on the catalyst surface. A slab model of the ferrihydrite (001) surface was adopted in these calculations, where the active site was circled with a dashed line (Fig. 5g and Supplementary Fig. 14b). Figures 5h, i showed the corresponding energy profiles for the disproportionation of on ferrihydrite and P -doped ferrihydrite, respectively. It is worth noting that the adsorption of the first and subsequent splitting to release are exothermic, indicating that they can easily occur on both ferrihydrite and P -doped ferrihydrite. However, the formation of after adsorption of the second on ferrihydrite is endothermic with an increase in energy by 0.7 eV (Fig. 5h). In contrast, the generation of is exothermic with energy decreased by 0.67 eV (Fig. 5i). This is probably due to the (001) surface of ferrihydrite having a very strong affinity to H , which made the H -adsorbed structure very stable. The result of Fig. 5h suggested that ferrihydrite tended to oxidize to form rather than catalyzed its disproportionation of like a SOD. On the other hand, the formation of was energetically more favorable than that of by 0.29 eV in P -doped ferrihydrite (Fig. 5i), indicating that the phosphate-doped ferrihydrite can readily catalyze the disproportionation of superoxide. These DFT results agree with the experiment that ferrihydrite has a superior SOD-like catalytic activity by phosphate doping.
The catalytic feature of the SOD-like activity of ferritins
In order to verify the catalytic role of the ferritin iron core, the reaction processes with superoxide were subsequently examined. Firstly, the intermediate product ferrous ion was tracked by the specific chelator ferrozine. As shown in Fig. 6a, the ferrous ion was detected during the reaction of ferritins with the xanthine oxidase (XOD)/xanthine (Xan) and exhibited a positive correlation with the xanthine levels (Fig. 6b). Moreover, there was no ferrous ion generated without the XOD or xanthine, indicating the ferrous ion mainly came from the reduction by the superoxide (Supplementary Fig. 15a-c). However, the ferrous ion became undetectable when ferrozine was added after the reaction, likely due to the re-oxidation of the ferrous ions (Fig. 6c). These results were also supported by previous reports that the iron core remains mostly inside ferritin after ferric reduction and could be re-oxidated subsequently . The same results were obtained in experiments with phosphate-doped ferrihydrite, in which the doped phosphate enhanced the reaction ability of ferrihydrite with the superoxide, and there was also no ferrous ion detected after the reaction (Supplementary Fig. 15d, e). To distinguish the SOD-like activity of ferritin with the ferric ion catalyzed Haber-Weiss reaction, we estimated based on the standard ferrous concentration curve that the generated ferrous ions accounted for of the iron in the HFn , pfFn and pyFn respectively in the time course of SOD activity test (Fig. 6d). In contrast, the superoxide diminishing ability of these ferric ions was much lower than their corresponding ferritins (Fig. 6e). Furthermore, superoxide diminishing efficiency of ferric salt in the equivalent total iron level was also investigated. As shown in Supplementary Fig. 15f, the SOD activity of the ferric salt in the same total iron level was much lower than the archaea but higher than the HFn group, suggesting that the SOD-like activity dose not result from the ionic reaction. Additionally, the generation of ferrous ions from archaeal ferritins and ferric salt in the same total iron content was also evaluated. As shown in Supplementary Fig. 15g, compared with the ferritins, the ferric salt produced much more ferrous while on the contrary exhibited low superoxide diminishing ability. Furthermore, unlike the reaction with ferritin, there remained a high level of ferrous after the reaction (Fig. 6f), suggesting that the ferritins deplete the superoxide in a way different from the ferric. Electron spin resonance (ESR) was then utilized to evaluate the ability of ferritins to deplete superoxide. As expected, the ferritins significantly decreased the amount of superoxide in a dose-dependent manner (Fig. 6g and Supplementary Fig. 15h, i), and the archaea ferritins exhibited a stronger ability than HFn in reducing superoxide (Fig. 6h). Of note, the ferritin-catalyzed superoxide decay was completely different from the ferric ion-induced reaction, as the latter produced obvious hydroxyl radicals (Supplementary Fig. 15j). We further assessed the SOD-like activity of pyFn-Fe and phosphate-doped ferrihydrite after their reactions with gradient amounts of xanthine. Both of them exhibited consistent SOD-like activity (Fig. 6i), and their iron contents did not decrease with the reaction (Supplementary Fig. 15k, l), further validating the catalytic role of the iron core. In summary, these results demonstrated the stronger superoxide-depleting ability of archaeal ferritin when compared with that of HFn and showed that the ferritin iron core catalyzes the superoxide depletion in a way different from ferric ions.
Antioxidant ability of the ferritin in transgenic and mutated E. coli
Inspired by the SOD-like activity of the prokaryote ferritins, we investigated their antioxidant ability in transgenic E. coli exposed to paraquat, which could stimulate superoxide production . As shown in Supplementary Fig. 16a, BL21(DE3) competent cells transformed with plasmids encoding HFn, pfFn, pyFn expressed similar levels of the ferritins. Next, to detect the superoxide sensitivity of these strains, the spot assay was conducted by dropping serially diluted transgenic
Fig. 6 | Characterization of the reaction process of the SOD-like activity of ferritins. a Ferrous-ferrozine production curve in the reaction of ferritins with XOD-Xan. Content of the ferrous-ferrozine production in the reaction of ferritin with XOD and gradient xanthine. c The ferrous-ferrozine production in the reaction of ferritin and XOD-Xan when the ferrozine was added 30 min later than xanthine oxidase. The proportion of intermediate ferrous ions to the whole iron content in different ferritins after their reaction with XOD-Xan. e SOD-like activity of the archaeal ferritins and relative ferric ion, which was generated in the reaction of archaeal ferritins with XOD-Xan in the 30 min course. The ferrous production of archaeal ferritin and ferric salt after the reaction with the XOD-Xan in the iron concentration at . g ESR spectra of the DMPO/HOO• in the presence of
XOD/Xan and gradient pyFn; the marked concentration referred to iron content. h The ESR intensity of DMPO/HOO• of HFn, pfFn and pyFn groups. The line at 3488.125 G of DMPO/ HOO• spectra was used to represent the intensity. i SOD-like activity of the pyFn and phosphate-doped ferrihydrite after their reaction with gradient xanthine. All data in (a-f, i) are mean SD ( independent experiments). The significant difference was evaluated by Two-tailed unpaired Student’s -test for (e), and by One-way ANOVA with post-hoc Tukey HSD test for (i). The value in (i) compared with Ctrl for pyFn group from left to right were 0.7532, 0.0742, 0.1398; and for phosphate-doped ferrihydrite were 0.9967, 0.7771, 0.5676; ns: not significant, . Source data are provided as a Source Data file.
bacteria onto the plate containing paraquat. As shown in Fig. 7a, transgenic bacteria expressing the archaeal ferritins exhibited markedly higher resistance to paraquat than those transformed with the empty vector or the HFn-encoding vector. While all the bacteria grew at a similar rate without paraquat, bacteria transformed with plasmids encoding pyFn or pfFn grew significantly faster than those transformed with empty vector or plasmid encoding HFn in the presence of paraquat (Fig. 7b, c), illustrating higher superoxide tolerance of the pyFn or pfFn groups. The bacteria viability was assessed by the colony formation experiment. As shown in Fig. 7d, there remained more single colonies of the pyFn or pfFn groups compared with the empty vector and HFn group. Likewise, the viabilities of bacteria were also reflected by the dehydrogenase activity of bacteria, which could reduce the WST-8 indicator and exhibit a specific absorption peak at 450 nm . As shown in Supplementary Fig. 16b, the bacteria transformed with plasmids expressing pfFn and pyFn exhibited markedly higher dehydrogenase activities in the presence of paraquat. A similar result was obtained by the confocal laser scanning microscopy (CLSM) image after co-staining with SYTO-9 (living bacteria) and propidium (PI, dead bacteria) (Supplementary Fig. 16c). The morphology of bacteria was further analyzed by scanning electron microscope (SEM). The bacteria in all the different groups exhibited the typical rod-like shape with intact and smooth cell walls. While the bacteria in the control groups suffered significant membrane damage and cell shrink after paraquat
treatment, those expressing HFn had relieved paraquat damage but still showed moderate shrinkage. In contrast, the expression of pfFn and pyFn largely reversed the effects of paraquat (Fig. 7e). Additionally, replating the paraquat-treated bacteria on the LB plates also verified the high cell viabilities in the pfFn and pyFn transformed groups. Next, we used the fluorescent probe 2′-7′-dichlorofluorescein diacetate (DCFH-DA) to detect ROS levels in paraquat-exposed bacteria. As expected, the bacteria containing pfFn and pyFn showed significantly lower levels of ROS in comparison with those transformed with the empty vector or vector expressing HFn (Fig. 7g). These results demonstrated that consistent with their SOD-like activity, the prokaryote ferritins exert strong antioxidant function in bacteria and may have important physiological functions.
Subsequently, to further verify the different superoxidediminishing abilities of ferritins in human and bacteria, we both substituted the gene with the gene or knocked out the gene in the BL21(DE3) strain by the CRISPR/Cas9 system (Supplementary Fig. 16d). As shown in Supplementary Fig. 16e, after the donor fragments with the gene and the plasmids containing the gRNA and Cas9 were transformed into the BL21 strain, colonies with double resistance were selected. Next, by using the genomic forward primer and reverse primer inner the , the colonies with successful gene editing were selected for the plasmids curing and the electrophoresis result indicated the obvious gene substitution. Likewise, the knockout
Fig. 7 | The antioxidant ability of ferritin in transgenic E. coli. a Spot assay of the different transgenic . coli under the pressure of paraquat of three independent experiments with similar results. Ctrl referred to groups without paraquat treatment. b, c The growth curve of different ferritin transgenic E. coli with or without the paraquat stimulation; the data are mean ( independent experiments); the significant difference was evaluated by the Two-way ANOVA with post-hoc Tukey HSD test. The colony formation amount of the ferritin transgenic E. coli with or without the paraquat stimulation; the data are mean SD ( independent experiments); the significant difference was evaluated by the One-way ANOVA
with post-hoc Tukey HSD test. e Representative SEM images of ferritin transgenic . coli after the paraquat treatment of three independent experiments with similar results; the scale bar referred to . f Digital images of replated ferritin transgenic E. coli after the paraquat treatment of three independent experiments with similar results. ROS levels of the different ferritin transgenic E. coli after the paraquat treatment. h Spot assay of the WT, Bfr knockout (Bfr KO) and Bfr substitution strain of three independent experiments with similar results. i ROS levels of WT, Bfr knockout and substitution strain after the paraquat treatment. Source data are provided as a Source Data file.
strain was acquired by the homologous fragment without and the knockout efficiency was verified by the upstream and downstream primers in the genome of the gene (Supplementary Fig. 16f). After the successful construction of these two strains, their antioxidant abilities against paraquat were compared. As shown by the spot assay (Fig. 7h), the Bfr knockout apparently increased the superoxide sensitivity of the BL21 strain and the HFn substitution did not contribute to the antioxidant abilities. Moreover, by detecting the ROS levels, we also found higher ROS levels in both the knockout and substitution
strains, further indicating the antioxidant function of the ferritins with SOD-like activity (Fig. 7i).
Discussion
The biominerals are widely distributed and play an important physiological role in different organisms. However, they mainly exploited physical-related properties like mechanical support and optical or magnetic sensing in vivo. The rise of nanozymes implied that nanobiominerals might function as enzyme-like catalyzers. In this study, we
chose ferritins as the research model. By comparing the SOD-like activities of ferritins from three living kingdoms, we found for the first time the relationship between the superoxide scavenging ability of ferritin iron cores and their sources of species, which is in the order of prokaryote > eukaryote. It is conceivable that the difference reflects the imminent requirement for cells of unicellular organisms to survive the oxidative environment, to which most of the cells in higher organisms are not exposed directly.
By identifying the core constitution, we found that the difference in the activity was caused by the phosphate coordinated with the iron atom inner the ferritin, which was initially shaped by the structure. In fact, previous studies have found higher inorganic phosphate content existed in the ferritins of bacteria and limpet hemolymph than those of human spleen and . vinelandii ferritin had phosphate neighbored to the iron atom in the core, compared with horse spleen ferritins, which agreed well with our study . Of note, it was speculated in previous literature that this variance in the bacteria and mammalian ferritin core was caused by the different phosphate content in the cytoplasm. Our reconstitution and mutant experiments indicated that the protein structure was also the decisive factor for the variance of the iron core.
It is interesting to know how phosphate in the core affects enzyme-like activity. It was reported that the phosphate in the ferritin core affected the oxidant and deposition of the ferrous in the horse spleen and bacteria ferritin by promoting the displacement of Fe (III) by in the ferroxidase center and accelerating the ferrous redox activity on the surface of the iron core . In addition, Watt et al. have found that ferritins from horse spleen and . vinelandii possessed different redox potentials. Compared with the horse spleen ferritin ( , and -416 mV at , and 9.0 , respectively), the . vinelandii ferritin exhibits a more negative and stable reduction potential ( -420 mV ) at above pHs , indicative of stronger iron retaining ability by the . vinelandii ferritin. In addition, during the reduction of horse spleen ferritin, there are almost transferred to the core for each reduced to the , which was absent in the bacteria ferritin . This phenomenon corresponds well to our DFT result that the iron core in HFn owned stronger hydron proton binding ability compared with the archaeal ones . These results demonstrated that phosphate affects the formation process as well as the reaction ability of the iron core.
More importantly, our disclosing of the natural ferritin SOD nanozyme and catalytic mechanism would provide a new platform for biomedical applications. Superoxide could not only directly damage the enzyme with Fe-S center, like the aconitase and succinate dehydrogenase, but also transform into other highly toxic reactive oxygen radicals such as hydroxyl radicals and peroxynitrite, which are harmful to most cells and tissues. So superoxide plays an important role in many diseases, including radiation damage, stroke, neurodegenerative diseases, cancer, and so on . Previous research has applied the natural superoxide dismutase to treat rheumatoid arthritis , osteoarthritis and mitigate the radiotherapy and chemotherapy damage . However, owing to the low cellular uptake, immunogenicity, and short half-life, their clinical translation was greatly restricted . With the high stability and easy modification of ferritin structures, the finding of the SOD-like activity of natural ferritin nanozyme would not only contribute to the understanding of the biominerals but also offer a new strategy for the superoxide-related disease. For example, our previous works have proved that HFn itself could target the tumor cells with high specificity and sensitivity based on their TfR1 binding ability . In addition, the overexpression of superoxide dismutase has been found to inhibit the growth or invasion of the tumor cells . Combined with these two characteristics, it was believed the ferritin nanozyme could be used to enhance tumor therapy. Moreover, the ferritin cages could not only be easily modified to endow them with different targeting characteristics but also act as a good drug carrier , which further broadened their application scenarios. Besides tumor therapy, the ferritin nanozyme
might be also used as an antioxidant for both in vitro and in vivo applications (e.g., anti-inflammation, and anti-aging). Nevertheless, our finding here just preliminarily resolved the catalytic mechanism of the ferritin nanozyme, much work was needed to further optimize their structures and activities before their applications.
Furthermore, it is worth noting that we only examined the difference and mechanism of SOD-like activity of the ferritins. It is likely that other enzyme-like activities of natural ferritin iron core may also have important physiological and evolutionary functions that are interesting and remain to be further explored. In addition, ferritin only constituted a few parts of the natural nanozymes. Much more work is needed to discover the existence of various natural nanozymes and explore their physiological functions. Hopefully, our studies on the enzyme-like activity of ferritins may pave the way for more systematic investigations.
Methods
Materials
All chemicals were of analytical grade. NaCl, Yeast extract, peptone, , hydrogen peroxide ( ), 3, 3′, 5, 5′-Tetramethylbenzidine (TMB), 2′, dichlorofluorescein (DCFH-DA), ampicillin, kanamycin, spectinomycin and isopropyl- -D-thiogalactoside (IPTG) were purchased from SigmaAldrich (USA). Fe foil was purchased from the Alfa Aesar. Human CuZnSOD was purchased from Absin (China), SOD assay kit, 5,5-Dimethyl-1-pyrroline-N-oxide (DMPO), and Microbial viability assay kit were purchased from DOJINDO (Japan). LIVE/DEAD BacLight Bacterial Viability Kit was purchased from Thermo Fisher (USA).
Multiple sequence alignment and phylogenetic tree construction
The amino acid sequences of the DNA binding protein from starved Escherichia coli str. K-12 substr. MG1655 (EcDps, NP_415333.1), DNA protection protein from Sulfolobus solfataricus (ssDps, NP_343470.1), Bacterioferritin from Escherichia coli str. K-12 substr. MG1655 (EcBfr, NP_417795.1), Non-heme ferritin from Enterobacteriaceae (EcFTn, WP_000917208.1), heavy chain ferritin from Homo sapiens (HFn, NP_002023.2), light chain ferritin from Homo sapiens (LFn, NP_000137.2), ferritin from Pyrococcusfuriosus (pfFn, WP_011011871.1), ferritin from Pyrococcus yayanosii (pyFn, WP_013905435.1), ferritin from Caenorhabditis elegans (CeFn, NP_504944.2), ferritin from Crassostrea gigas (CgFn, NP_001292267.1), ferritin from Siniperca chuatsi (XP_044062311), heavy chain ferritin from Xenopus laevis (NP_001084057.1), heavy chain ferritin from Anolis carolinensis (XP_003221882), heavy chain ferritin from Gopherus eugoodei (XP_030416668), heavy chain ferritin from Coturnix japonica (XP_015719168), bacterioferritin from Parabacteroides goldsteinii strain BFG-241 (WP_007659302), bacterioferritin from Alistipes onderdonkii subsp. vulgaris strain 3BBH6 (WP_022332956.1), bacterioferritin from Thermus scotoductus (WP_038069286.1), bacterioferritin from Streptococcus equinus (WP_074533576.1), bacterioferritin from Leptospira weilii (WP_002621798), bacterioferritin from Pseudomonas aeruginosa (PaBfr, WP_003092078.1), ferritin from Pseudomonas aeruginosa (PaFTn, WP_003093668.1), ferritin from Thermus scotoductus (TsFn, WP_038069286.1), ferritin from Mycobacterium tuberculosis (MtFn, NP_218358.1), ferritin from Candidatus Korarchaeum (WP_012309233.1), ferritin from Staphylothermus marinus (WP_011839058), ferritin from Thermosphaera aggregans (TaFn, WP_013129153) and ferritin from Thermococcus barophilus (TbFn, WP_013467695) were obtained from the NCBI. The multiple sequence alignment was conducted by Cluster W and the sequence similarity was marked by Genedoc software (the color referred to amino acid similarity, red: , blue: , gray: ), and the Mega5 was applied to construct the phylogenetic tree by the neighbor-joining method, bootstrapped with 1000 replicates.
Plasmid preparation, wild type and biomineralized ferritin expression and purification
Total 18 different ferritins were selected for the purification and enzyme-like activity test, including EcDps, ssDps, EcFTn, EcBfr, HFn, LFn, pfFn, pyFn, CeFn, CgFn, PaBfr, PaFTn, TsFn, MtFn, TaFn, TbFn, human mitochondrial ferritin (MFn) and human heteropolymer ferritin (H/LFn). Except for the H/LFn, other 17 ferritin coding sequences were cloned to the pet22b plasmid with the Ndel and BamHI restriction site. The H/LFn expression vector was constructed by cloning the two coding sequences into petDuet-1 plasmid through Ncol/BamHI and Ndel/Xhol. The constructed plasmids were transformed to the BL21(DE3) competent cells to express the proteins. The transformed competent cells were cultured overnight in LB medium with ampicillin. The protein expression was induced by adding 1 mM isopropyl- -D-thiogalactoside (IPTG, Sigma) when the OD600 reached 0.6-0.8, then the induced bacteria were further cultured for 8 h at . The biomineralized ferritins were synthesized by adding freshly prepared ferrous salt into the bacteria culture medium just before the IPTG so that the excess iron could be biomineralized in the ferritin cavity during the ferritin expression. The induced bacteria were harvested by centrifugation for 20 min and the pellets were resuspended for protein purification. The purification processes were referred to previous publications , except that all the phosphate salt buffer was substituted for the Tris-base or Hepes buffer.
Characterization of the wild type and biosynthetic ferritin from different species
TEM: the ferritin samples ( ) were embedded in a Plasma Cleaner HPDC32G-treated copper grid and stained with 1% uranyl acetate for 1 min , then imaged with a JEM- 140080 kV TEM (JEOL, Japan). DLS: the average diameter of the ferritin samples ( , ) was analyzed by the DynaPro Titan with a temperaturecontrolled micro-sampler of (Wyatt Technology, U.S.A.). Inductively coupled plasma mass spectrometry (ICP-MS): the element contents of ferritin samples from different species were analyzed by the Agilent ICPMS7800.
Ferritin iron core removal and protein shell digestion
The iron core removal method was referred to previous publication : the biosynthetic ferritins were added to the 0.1 M sodium acetate buffer containing 1% thioglycolic acid, pH 5.5 and placed in for 18 h , then excess 2-2′-bipyridyl was added to chelate the ferrous. The apo-ferritin was then acquired by the G75 desalting column (GE Healthcare). The protein shell was digested by the protease K : protease K was added to the ferritin solution in 20 mM Tris-base buffer pH 8.0 and placed at for complete digestion of the ferritin shell.
Synthesis of ferritin reconstituted with different iron/ phosphate ratio
The reconstitution experiment was modified from previous publication : ferritins were placed in the 50 mM MES buffer, pH 6.5 , then the freshly prepared ammonium ferrous sulfate solution was added to the ferritin solution in the frequency of 500/ferritin every 10 min at the room temperature for the ferrous oxidant and nucleation. Ferritins with different iron/phosphate ratios were synthesized by adding relative sodium hydrogen phosphate before the ferrous salt. After the reaction, the mixture was centrifugated at for 10 min to remove the excess sediment. The free ferric ions were then removed by the G75 desalting column.
Enzyme-like activity test
Superoxide dismutase-like activity test: the SOD-like activities of the ferritins and ferrihydrites were evaluated by the SOD assay kit (Dojindo, Japan), which was based on the Xanthine-Xanthine oxidase system. The xanthine oxidase could oxidase the substrate xanthine
leading to the reduced xanthine oxidase, which subsequently oxidase the oxygen to generate the superoxide. The superoxide could react with the water-soluble tetrazolium salt (WST-1) in the assay kit to generate the WST-1 formazan dye, which exhibited a specific absorption peak at 450 nm , so the superoxide level correlated with OD 450 absorbance. The SOD agent could diminish the superoxide, leading to the decrease of the absorbance at 450 nm . Therefore, SOD activity was defined as the “inhibition rate (%)” of the superoxide’s reaction with WST-1, and calculated as [(Ablank 1 – Ablank 3) – (Asample – Ablank 2)]/ (Ablank 1 – Ablank 3) x 100, where the Ablank 1 referred to OD450 of above reaction system without SOD agents, Ablank 3 referred to reaction system without the SOD agent and xanthine oxidase, Asample referred to reaction system with SOD agents, Ablank 2 referred to reaction system with SOD agents but without the xanthine oxidase. During the actual test, samples with specific protein concentrations determined by the BCA assay or iron content determined by ICP-MS were mixed with WST-1 working solution, pH 7.4 , then xanthine oxidase was added to start the reaction and the mixture was placed at for 30 min , then OD450 was monitored by the microplate reader. One unit of SOD was defined as the amount of the enzyme in of the sample solution that inhibited the reduction reaction of WST-1 with superoxide anion by . Likewise, the SOD units base on the commercial CuZnSOD was calculated based on the IC50 of commercial SOD and its relative SOD units.
Peroxidase-like and oxidase-like activity test: the peroxidase-like and oxidase-like activity was evaluated by using TMB (dissolved in DMSO, ) as the substrate. Briefly, mixture containing TMB (final concentration ) and (final concentration 0.5 M ) in the 0.2 M NaAc buffer, pH 4.5 was added to the ferritin or ferrihydrite samples (final concentration ). The activity was evaluated by monitoring the absorbance of the oxTMB at 652 nm in a 30 min time course. The oxidase-like activity was performed likewise except that the was absent in the mixture.
Catalase-like activity test: catalase-like activity was determined by the specific oxygen electrode on a multi-parameter analyzer (JPSJ606L, Leici China). The ferritin or ferrihydrite was mixed with the aqueous solution of different concentrations in the 20 mM Hepes buffer, pH 7.4 , the final concentration of the ferritin or ferrihydrite was , and the oxygen generation rate was recorded for 2 min .
Urate generation Assay of the Xanthine oxidase system
The urate generation assay was conducted by referring to previous literatures . Briefly, xanthine oxidase ( of xanthine oxidase) was added to a solution of 20 mM Hepes buffer, pH 7.4 , containing xanthine ( ) at a final volume of 3.0 mL at . Urate production was monitored by the change of absorbance at 290 nm . For the ferritin groups, the ferritins were added before xanthin oxidase in the final concentration of .
Modification and purification of the three-fold axis mutated human heavy chain ferritin
Amino acids at the 3 -fold axis of HFn were substituted for the relative amino acids in the Pseudomonas aeruginosa Bfr (T123E, C131R, E135K, T136D). The modified HFn mutant sequence was cloned to the pet22b plasmid, and the HFn mutant was purified as the wild type.
Synthesis of phosphate doped ferrihydrite
Phosphate doped 2-line or 6-line ferrihydrite was synthesized by the method modified from previous publication . All the solutions were prepared by the with a resistivity of . 2-line ferrihydrite and phosphate-doped 2-line ferrihydrite: were added to 500 mL distilled water or those containing relative under vigorously stirring, then 1 M NaOH was added to the solution with continuous stirring to adjust the pH to 7-8. Then the ferrihydrites were centrifuged at for
20 min to precipitate the ferrihydrites and washed with distilled water 5 times to remove the electrolyte. The precipitations were then freezefried for storage or further use. 6 -line ferrihydrite and phosphate doped 6-line ferrihydrite: the distilled water containing no or 0.215 g , were placed in the oven for pre-heating, then was added to the pre-heated solution under stirring and the mixture was placed back to the oven for . After the reaction, the solution was transferred to the ice for rapid cooling. The cooled solutions were then dialyzed against distilled water for 7 days, the dialyzed water was replaced 3 times every day. The dialyzed materials were freeze-fried for storage or further use.
Characterization of the ferritin iron core and the phosphatedoped ferrihydrite
High resolution transmission electron microscope (HRTEM, FEI Tecnai G2 F30, USA) with the selected area electron diffraction (SAED) was used to characterize the morphology and crystal lattice structure of the ferritin iron core and the phosphate-doped ferrihydrites. X-ray photoelectron spectroscopy (XPS, Thermo escalab 250Xi, monochromatic AlK radiation accelerating voltage of 14.8 kV , power of was applied to characterize the element constitution and valence state. FTIR spectrum (Nicolet IS10, USA) was used to determine the specific functional group by scanning in the range of in the step of . X-ray powder diffractometer (XRD, Brucker D8 advance, Germany) was used to characterize the crystal structure of the ferritins and phosphate-dope ferrihydrite with the radiation in the range of with the step of .
Surface phosphate absorption of the ferritin iron core and the ferrihydrite
The ferrihydrite surface absorption experiment was modified from the previous publication . The freeze-dried 2-line ferrihydrite was suspended in deionized water containing 0.1 M NaCl by sonication. The iron content of the prepared ferrihydrite solution was determined by the ICP-MS, and then phosphate salt of relative molar ( molar ratio ) was added to the ferrihydrite solution. The mixture was stirred for 24 h for sufficient absorption and to avoid potential precipitation. Then the samples were centrifuged at for 10 min to separate the phosphate-absorbed ferrihydrite and washed three times with . The precipitation was freeze-fried for determining the iron and phosphate content and tested for SOD-like activities. Ferritin iron core surface absorption experiment was performed likewise. Firstly the iron-reconstituted ferritin was synthesized by adding the ammonium ferrous sulfate to the different ferritin in 50 mM MES buffer, pH 6.5 , and further purified by the G75 desalting column. Then the phosphate solution of relative molar concentration was added to the reconstituted ferritin and the mixtures were further incubated at for 24 h , then the samples were centrifuged at for 10 min and purified by G75 desalting column. The protein concentration was detected by the BCA protein assay kit and SODlike activity was tested.
Synchrotron radiation characterization of the ferritin iron core The iron chemical state and coordination environment were assessed via X-ray absorption fine structure (XAFS) using Fe foil, , and as reference materials. Biomineralized ferritin samples underwent a dialysis process against distilled water to eliminate salts. Subsequently, the dialyzed solutions were freeze-dried, thoroughly ground, and compressed into uniform pellets affixed to 3 M tape. Fe-K edge XAFS spectra were acquired at beamline 1W1B of the Beijing Synchrotron Radiation Facility (BSRF) in China, employing the transmission mode for sample and reference measurements. Normalized XANES data were analyzed via least-squares fitting using IFEFFIT Athena software from CARS (Consortium for Advanced Radiation
Sources at the University of Chicago) to ascertain the ratio of iron species. For EXAFS data analysis, IFEFFIT Artemis software (also from CARS) was utilized, employing the Fe (Im-3m) phase for data fitting purposes.
DFT studies of SOD-like activity of phosphate ferrihydrite
The slab model of the (001) surface of ferrihydrite was employed to investigate its catalytic activities for the disproportionation of superoxide. The most stable P-doped ferrihydrite was constructed by replacing one Fe atom with a P atom. Utilizing these slab models, DFT calculations were conducted through the Vienna Ab initio Simulation Package (VASP 5.4.4) . The calculations employed the Perdew-Burke-Ernzerhof functional, augmented with Grimme’s semi-empirical “D3BJ” dispersion term (PBE-D3BJ) . To describe valence electrons, plane wave basis sets with an energy cutoff of 450 eV were utilized, while projector-augmented-wave pseudopotentials described core electrons. Convergence criteria were set at for energy and for atomic force. Brillouin zone sampling was achieved using Monkhorst-Pack k-point meshes in all calculations.
Intermediate ferrous detection and the recycle activity test
The intermediate ferrous ions detection was performed according to the method modified from the previous method . Briefly, the ferritins or the phosphate-doped ferrihydrite were mixed with xanthine and ferrozine in the same total iron content of in the 20 mM Hepes buffer, pH 7.4 . Then xanthine oxidase was added to start the reaction and the generation of ferrous was monitored by the . Ferrozine that exhibited specific absorbance at 562 nm . The Ferrous standard curve was acquired by adding gradient ammonium ferrous sulfate to ferrozine. The standard curve was calculated by the linear regression method. The recycle activity experiment was performed by reacting ferritins or phosphate-doped ferrihydrite with gradient xanthine for 24 h in the presence of XOD. Then the archaeal ferriting groups were heated at for 15 min to denature the xanthine oxidase, and ferritins were separated from the mixture by the centrifugation and desalting column. The phosphate-doped ferrihydrite was separated by centrifugation and washing. Then the separated ferritins and phosphate-doped ferrihydrites were used for further SOD-like activity test.
Electron spin resonance
The electron spin resonance spectroscopy (ESR) was used to examine the superoxide scavenging ability of the ferritins: xanthine oxidase (XOD) and xanthine was used to generate superoxide in the 20 mM Hepes buffer, DMPO was used to trap the superoxide by forming the DMPO/OOH. The reaction was started by the addition of the XOD.
The antioxidant experiments of ferritin transgenic . coli
Spot assay: The sensitivity to the superoxide generator paraquat of transgenic bacteria was evaluated by the spot assay. Transformed strains with empty plasmid or reconstituted expression vectors were cultured in the fresh LB medium. When the OD600 reached 0.8, the inoculum was induced by 1 mM IPTG at for 8 h , then these strains were 10 times gradient diluted and spotted on the solid LB medium containing 0.2 mM paraquat to analyze their sensitivity.
Growth curve monitor: The transformed BL21(DE3) bacteria with empty plasmid or the reconstituted expression vectors were cultured in the fresh LB medium containing ampicillin until the OD600 reached , then the inoculum was diluted to the same by the fresh LB medium. Subsequently, 1 mM IPTG and 1 mM paraquat were simultaneously added to the medium when the OD600 reached 0.2 and the inoculums were further incubated at , 200 rpm for 12 h . The OD600 absorbance was recorded every 2 h .
Reactive oxygen species (ROS) assay: the intracellular ROS level was determined by the -dichlorofluorescein diacetate (DCFH-DA). DCFH-DA could be taken up by cells where cellular esterase cleaves off the acetyl groups, resulting in the DCFH. The cellular-generated ROS could oxidize the DCFH to DCF, which emits green fluorescence with an excitation wavelength of 488 nm and an emission wavelength of 525 nm . The fluorescence levels were evaluated by Flow Cytometry. Flow Cytometry could assign a channel to the fluorescence cell according to their fluorescence intensity. With the increase of cellular fluorescence intensity, the cells would be distributed in higher channels (corresponding to a shift to a higher X-axis range). During the actual assay, transformed bacteria were treated as before. After the treatment, the bacteria were harvested and washed with PBS buffer, pH 7.4. Then DCFH-DA was added to the resuspend bacteria in the final concentration of and incubated at for 30 min , then the bacteria were washed twice by PBS, pH 7.4 and detected by Flow Cytometry.
Bacteria viability assay: The bacterial viability was checked by colony formation experiment. Briefly, the bacteria containing different plasmid was treated as mentioned above. After the paraquat treatment, the transgenic bacteria were plated with proper dilution and cultured at overnight, then the bacterial numbers were calculated as CFU .
Bacteria dehydrogenase activity assay: The microbial viability assay kit (Dojindo) was used to detect the bacteria viability through their dehydrogenase activity. The bacteria containing different plasmid was treated as mentioned above. After treatment, bacteria inoculum was transferred to the 96 -well plates and working solution (containing WST-8 and mediator) was added to each well. Then the plate was placed at for and OD450 was recorded by SpectraMax. Besides, the living/Dead Baclight bacteria viability kit was used to determine the living (SYTO-9, FITC) and dead (PI, Texas Red) bacteria by the confocal laser scanning microscopy (CLSM) according to the supplier’s manuscript.
Morphology observation: Scanning electron microscopy was applied to observe the transformed bacteria morphology. The paraquat-treated bacteria were harvested and washed with PBS buffer, and the precipitate was fixed by glutaraldehyde at for 8 h . Then the fixed sample was washed three times with the saline and further dehydrated by a series of ethanol/water mixture. The sample was then dried by the Automatic critical point dryer (CPD300) and observed by the scanning electron microscopy (SU8010).
Gene knockout and substitution method by CRISPR/Cas9 in BL21(DE3)
The gene knockout and substitution were conducted according to Yang et al. , and gRNA for was designed by the CRISPOR website. The pecCas9 plasmid containing -RED and Cas9 gene was transformed into the BL21(DE3) strain, and positive colonies were selected to induce the expression of -RED and make competent cells. Next, the pecgRNA plasmid and homologous fragment of Bfrupstream and downstream with or without insertion were electroporated into the competent cells. The individual colonies were verified by colony PCR and DNA sequencing. Finally, the verified colonies were used for pecgRNA and pecCas plasmids curing by adding the rhamnose and sucrose respectively. All the plasmids and primers used in this study were listed in Supplementary Table 2.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data supporting the findings of this study are available from the corresponding authors upon request. All the PDB Structures are
available on the PDB database (https://www.rcsb.org/). Source data for the figures and supplementary figures are provided as a Source Data file. Source data are provided with this paper.
References
Warr, L. N. IMA-CNMNC approved mineral symbols. Mineral. Mag. 85, 291-320 (2021).
Hazen, R. M. et al. Mineral evolution. Am. Mineral. 93, 1693-1720 (2008).
Hazen, R. M. & Ferry, J. M. Mineral evolution: mineralogy in the fourth dimension. Elements 6, 9-12 (2010).
Perez-Aguilar, C. D. & Cuellar-Cruz, M. The formation of crystalline minerals and their role in the origin of life on Earth. Prog. Cryst. Growth Charact. Mater. 68, 100558 (2022).
Ferris, J. P., Hill, A. R., Liu, R. H. & Orgel, L. E. Synthesis of long prebiotic oligomers on mineral surfaces. Nature 381, 59-61 (1996).
Huber, C. & Wachtershauser, G. Activated acetic acid by carbon fixation on (Fe,Ni)S under primordial conditions. Science 276, 245-247 (1997).
Roldan, A. et al. Bio-inspired conversion by iron sulfide catalysts under sustainable conditions. Chem. Commun. 51, 7501-7504 (2015).
Hazen, R. M. & Sholl, D. S. Chiral selection on inorganic crystalline surfaces. Nat. Mater. 2, 367-374 (2003).
Ferris, J. P. Mineral catalysis and prebiotic synthesis: Montmorillonite-catalyzed formation of RNA. Elements 1, 145-149 (2005).
Joshi, P. C., Aldersley, M. F., Delano, J. W. & Ferris, J. P. Mechanism of montmorillonite catalysis in the formation of RNA Oligomers. J. Am. Chem. Soc. 131, 13369-13374 (2009).
Konhauser, K. O. Diversity of bacterial iron mineralization. Earth-Sci. Rev. 43, 91-121 (1998).
He, H., Veneklaas, E. J., Kuo, J. & Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 19, 166-174 (2014).
Nudelman, F. & Sommerdijk, N. A. J. M. Biomineralization as an Inspiration for Materials Chemistry. Angew. Chem.-Int. Ed. 51, 6582-6596 (2012).
Gao, L. et al. Intrinsic peroxidase-like activity of ferromagnetic nanoparticles. Nat. Nanotechnol. 2, 577-583 (2007).
Fan, K. et al. In vivo guiding nitrogen-doped carbon nanozyme for tumor catalytic therapy. Nat. Commun. 9, 1440 (2018).
Yang, Y. et al. Platinum-carbon-integrated nanozymes for enhanced tumor photodynamic and photothermal therapy. Nanoscale 12, 13548-13557 (2020).
Wu, J., Li, S. & Wei, H. Multifunctional nanozymes: enzyme-like catalytic activity combined with magnetism and surface plasmon resonance. Nanoscale Horiz. 3, 367-382 (2018).
Chen, J. et al. Glucose-oxidase like catalytic mechanism of noble metal nanozymes. Nat. Commun. 12, 3375 (2021).
Singh, N., NaveenKumar, S. K., Geethika, M. & Mugesh, G. A Cerium Vanadate Nanozyme with specific superoxide dismutase activity regulates mitochondrial function and ATP synthesis in neuronal cells. Angew. Chem.-Int. Ed. 60, 3121-3130 (2021).
Ji, S. et al. Matching the kinetics of natural enzymes with a singleatom iron nanozyme. Nat. Catal. 4, 407-417 (2021).
Wu, J. et al. Nanomaterials with enzyme-like characteristics (nanozymes): next-generation artificial enzymes (II). Chem. Soc. Rev. 48, 1004-1076 (2019).
Li, K. et al. Magnetosomes extracted from Magnetospirillum magneticum strain AMB-1 showed enhanced peroxidase-like activity under visible-light irradiation. Enzym. Microb. Technol. 72, 72-78 (2015).
Tang, Z., Wu, H., Zhang, Y., Li, Z. & Lin, Y. Enzyme-mimic activity of ferric nano-core residing in ferritin and its biosensing applications. Anal. Chem. 83, 8611-8616 (2011).
Chi, Z.-L., Yu, G.-H., Kappler, A., Liu, C.-Q. & Gadd, G. M. Fungalmineral interactions modulating intrinsic peroxidase-like activity of iron nanoparticles: implications for the biogeochemical cycles of nutrient elements and attenuation of contaminants. Environ. Sci. Technol. 56, 672-680 (2021).
Andrews, S. C. The Ferritin-like superfamily: Evolution of the biological iron storeman from a rubrerythrin-like ancestor. Biochim. Et. Biophys. Acta-Gen. Subj. 1800, 691-705 (2010).
Yasmin, S., Andrews, S. C., Moore, G. R. & Le Brun, N. E. A new role for heme, facilitating release of iron from the bacterioferritin iron biomineral. J. Biol. Chem. 286, 3473-3483 (2011).
Gauss, G. H. et al. Structure of the DPS-like protein from Sulfolobus solfataricus reveals a bacterioferritin-like dimetal binding site within a DPS-like dodecameric assembly. Biochemistry 45, 10815-10827 (2006).
Wiedenheft, B. et al. An archaeal antioxidant: Characterization of a Dps-like protein from Sulfolobus solfataricus. Proc. Natl. Acad. Sci. USA 102, 10551-10556 (2005).
Uchida, M. et al. The archaeal Dps nanocage targets kidney proximal tubules via glomerular filtration. J. Clin. Investig. 129, 3941-3951 (2019).
Andrews, S. C. et al. Overproduction, purification and characterization of the bacterioferritin of Escherichia-coli and a c-terminally extended variant. Eur. J. Biochem. 213, 329-338 (1993).
Karas, V. O., Westerlaken, I. & Meyer, A. S. Application of an In vitro DNA protection assay to visualize stress mediation properties of the Dps Protein. Jove-J. Visual. Exp. 75, 50390 (2013).
Hudson, A. J. et al. Overproduction, purification and characterization of the Escherichia-coli ferritin. Eur. J. Biochem. 218, 985-995 (1993).
Yu, J. et al. Thermostable iron oxide nanoparticle synthesis within recombinant ferritins from the hyperthermophile Pyrococcus yayanosii CH1. Rsc Adv. 9, 39381-39393 (2019).
Watt, G. D., Frankel, R. B., Papaefthymiou, G. C., Spartalian, K. & Stiefel, E. I. Redox properties and mossbauer-spectroscopy of Azotobacter-vinelandii bacterioferritin. Biochemistry 25, 4330-4336 (1986).
Rohrer, J. S., Islam, Q. T., Watt, G. D., Sayers, D. E. & Theil, E. C. Iron environment in ferritin with large amounts of phosphate, from Azotobacter-vinelandii and horse spleen, analyzed using extended X-ray absorption fine-structure (EXAFS). Biochemistry 29, 259-264 (1990).
Ebrahimi, K. H., Hagedoorn, P.-L. & Hagen, W. R. Phosphate accelerates displacement of by in the ferroxidase center of Pyrococcus furiosus ferritin. Febs Lett. 587, 220-225 (2013).
Garcia-Prieto, A. et al. On the mineral core of ferritin-like proteins: structural and magnetic characterization. Nanoscale 8, 1088-1099 (2016).
Crow, A., Lawson, T. L., Lewin, A., Moore, G. R. & Brun, N. E. L. Structural basis for iron mineralization by bacterioferritin. J. Am. Chem. Soc. 131, 6808-6813 (2009).
Rivera, M. Bacterioferritin: structure, dynamics, and protein-protein interactions at play in iron storage and mobilization. Acc. Chem. Res. 50, 331-340 (2017).
Weeratunga, S. K. et al. Structural studies of Bacterioferritin B from Pseudomonas aeruginosa suggest a gating mechanism for iron uptake via the ferroxidase center. Biochemistry 49, 1160-1175 (2010).
Aronovitz, N., Neeman, M. & Zarivach, R. In Iron Oxides: From Nature to Applications. (eds. Damien, F.) 117-142 (Wiley, 2016).
Mann, S., Bannister, J. V. & Williams, R. J. P. Structure and composition of ferritin cores isolated from human spleen, limpet (patella-
vulgata) hemolymph and bacterial (pseudomonas-aeruginosa) cells. J. Mol. Biol. 188, 225-232 (1986).
Harrison, P. M., Fischbac, F. A., Hoy, T. G. & Haggis, G. H. Ferric oxyhydroxide core of ferritin. Nature 216, 1188 (1967).
Kauko, A. et al. Iron incorporation in Streptococcus suis Dps-like peroxide resistance protein Dpr requires mobility in the ferroxidase center and leads to the formation of a ferrihydrite-like core. J. Mol. Biol. 364, 97-109 (2006).
Towe, K. M. & Bradley, W. F. Mineralogical constitution of colloidal “hydrous ferric oxides”. J. Coll. interface Sci. 24, 384-392 (1967).
Snow, C. L. et al. Ferritin iron mineralization proceeds by different mechanisms in MOPS and imidazole buffers. J. Inorg. Biochem. 105, 972-977 (2011).
Westre, T. E. et al. A Multiplet Analysis of Fe K-Edge 1s 3d Pre-Edge Features of Iron Complexes. J. Am. Chem. Soc. 119, 6297-6314 (1997).
Orikasa, Y. et al. Noncrystalline nanocomposites as a remedy for the low diffusivity of multivalent ions in battery cathodes. Chem. Mater. 32, 1011-1021 (2020).
Shen, X. et al. Mechanisms of oxidase and superoxide dismutationlike activities of gold, silver, platinum, and palladium, and their alloys: a general way to the activation of molecular oxygen. J. Am. Chem. Soc. 137, 15882-15891 (2015).
Wang, Z. et al. Accelerated discovery of superoxide-dismutase nanozymes via high-throughput computational screening. Nat. Commun. 12, 6866 (2021).
Bolann, B. J. & Ulvik, R. J. Decay of superoxide catalyzed by ferritin. Febs Lett. 318, 149-152 (1993).
Watt, G. D., Frankel, R. B. & Papaefthymiou, G. C. Reduction of mammalian ferritin. Proc. Natl. Acad. Sci. USA 82, 3640-3643 (1985).
Hassan, H. M. & Fridovich, I. Superoxide radical and oxygen enhancement of toxicity of paraquat in Escherichia-coli. J. Biol. Chem. 253, 8143-8148 (1978).
Jiang, Y. et al. Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol 81, 2506-2514 (2015).
Cheng, Y. G. & Chasteen, N. D. Role of phosphate in initial iron deposition in apoferritin. Biochemistry 30, 2947-2953 (1991).
Huang, H. Q., Watt, R. K., Frankel, R. B. & Watt, G. D. Role of phosphate in binding to horse spleen holoferritin. Biochemistry 32, 1681-1687 (1993).
Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Heath Sci. 12, 88 (2018).
Goebel, K.-M., Storck, U. & Neurath, F. J. T. L. Intrasynovial orgotein therapy in rheumatoid arthritis. Lancet 317, 1015-1017 (1981).
Lund-Olesen, K. & Menander-Huber, K.B. Intra-articular orgotein therapy in osteoarthritis of the knee. A double-blind, placebocontrolled trial. Arzneimittelforschung 33, 1199-1203 (1983).
Menander-Huber, K., Edsmyr, F. & Huber, W. J. U. R. Orgotein (superoxide dismutase): A drug for the amelioration of radiationinduced side effects: A double-blind, placebo-controlled study in patients with bladder tumours. Urol. Res. 6, 255-257 (1978).
Fan, K. et al. Magnetoferritin nanoparticles for targeting and visualizing tumour tissues. Nat. Nanotechnol. 7, 459-464 (2012).
Teoh, M. L. T., Fitzgerald, M. P., Oberley, L. W. & Domann, F. E. Overexpression of extracellular superoxide dismutase attenuates heparanase expression and inhibits breast carcinoma cell growth and invasion. Cancer Res. 69, 6355-6363 (2009).
Kinnula, V. L. & Crapo, J. D. Superoxide dismutases in malignant cells and human tumors. Free Radic. Biol. Med. 36, 718-744 (2004).
He, J., Fan, K. & Yan, X. Ferritin drug carrier (FDC) for tumor targeting therapy. J. Controlled Release 311-312, 288-300 (2019).
Levi, S. et al. Mechanism of ferritin iron uptake – activity of the H-chain and deletion mapping of the ferro-oxidase site – a study of
iron uptake and ferro-oxidase activity of human-liver, recombinant H-chain ferritins, and of 2 H -chain deletion mutants. J. Biol. Chem. 263, 18086-18092 (1988).
Liu, G. et al. Characterization and surface reactivity of ferrihydrite nanoparticles assembled in ferritin. Langmuir 22, 9313-9321 (2006).
Weiss, R. H. et al. Evaluation of activity of putative superoxide dismutase mimics. Direct analysis by stopped-flow kinetics. J. Biol. Chem. 268, 23049-23054 (1993).
Faulkner, K. M., Liochev, S. I. & Fridovich, I. Stable Mn(III) porphyrins mimic superoxide dismutase in vitro and substitute for it in vivo. J. Biol. Chem. 269, 23471-23476 (1994).
Durot, S. et al. Series of Mn Complexes Based on N-Centered Ligands and Superoxide – Reactivity in an Anhydrous Medium and SOD-Like Activity in an Aqueous Medium Correlated to Mnll/Mnlll Redox Potentials. Eur. J. Inorg. Chem. 2005, 3513-3523 (2005).
Policar, C. In Redox-Active Therapeutics (eds. Batinić-Haberle, I., Rebouças, J. S. & Spasojević, I.) 125-164 (Springer Publishing, 2016).
Kim, J., Li, W., Philips, B. L. & Grey, C. P. Phosphate adsorption on the iron oxyhydroxides goethite (alpha-FeOOH), akaganeite (beta, and lepidocrocite (gamma-FeOOH): a P-31 NMR Study. Energy Environ. Sci. 4, 4298-4305 (2011).
Kresse, G. & Furthmüller, J. Efficiency of ab-initio total energy calculations for metals and semiconductors using a plane-wave basis set. Comput. Mater. Sci. 6, 15-50 (1996).
Grimme, S. et al. A consistent and accurateab initioparametrization of density functional dispersion correction (DFT-D) for the 94 elements H-Pu. J. Chem. Phys. 132, 154104 (2010).
Monkhorst, H.J. & Pack, J.D. Special points for Brillouin-zone integrations. Phys. Rev. B 13, 5188-5192 (1976).
Biemond, P., Swaak, A. J. G., Beindorff, C. M. & Koster, J. F. Superoxide-dependent and superoxide-independent mechanisms of iron mobilization from ferritin by xanthine-oxidase – implications for oxygen-free-radical-induced tissue destruction during ischemia and inflammation. Biochem. J. 239, 169-173 (1986).
Li, Q. et al. A modified pCas/pTargetF system for CRISPR-Cas9assisted genome editing in Escherichia coli. Acta Biochim. Biophys. Sin. 53, 620-627 (2021).
Acknowledgements
We thank Prof. Yaling Wang and Beijing Synchrotron Radiation Facility (BSRF), China for the technical support for the X-ray absorption fine structure. We thank Dr. Yujing Ren for his technical help in the XAFS data analysis. Supported by the National Natural Science Foundation of China (82122037, K.F., 81930050, X.Y., 22121003, X.Y.), National Key Research and Development Program of China (No. 2021YFC2102900, K.F.), CAS Project for Young Scientists in Basic Research (YSBR-089, K.F.), CAS Interdisciplinary Innovation Team (JCTD-2020-08, K.F.), and Key
Laboratory of Biomacromolecules, Chinese Academy of Sciences (O3CCSDZ301, K.F.).
Author contributions
K.F. conceived and designed the whole project. L.M. designed and performed the experiments. J.J.Z., X.F.G. designed and performed the DFT calculation. L.M., N.Z. designed and purified the HFn protein mutant. R.Z., L.F. contributed to protein purification and data analysis. L.Y. contributed to the manuscript writing. Y.C. supervised the XAFS. K.F., L.M., and X.Y. wrote the manuscript with input from all authors.
Competing interests
The authors declare no competing interests.
Additional information
Supplementary information The online version contains
supplementary material available at https://doi.org/10.1038/s41467-023-44463-w.
Correspondence and requests for materials should be addressed to Xiyun Yan or Kelong Fan.
Peer review information Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Reprints and permissions information is available at http://www.nature.com/reprints
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
CAS Engineering Laboratory for Nanozyme, Key Laboratory of Biomacromolecules (CAS), CAS Center for Excellence in Biomacromolecules, Institute of Biophysics, Chinese Academy of Sciences, Beijing 100101, China. University of Chinese Academy of Sciences, Chinese Academy of Sciences, Beijing 100408, China. Laboratory of Theoretical and Computational Nanoscience, National Center for Nanoscience and Technology of China, Beijing 100190, China. Savaid Medical School, University of Chinese Academy of Sciences, Beijing 100049, China. China Regional Research Centre, International Centre for Genetic Engineering and Biotechnology, Taizhou, Jiangsu 225316, China. CAS Key Laboratory for Biomedical Effects of Nanomaterials and Nanosafety and CAS Center for Excellence in Nanoscience, National Center for Nanoscience and Technology, Beijing 100190, China. Nanozyme Medical Center, School of Basic Medical Sciences, Zhengzhou University, Zhengzhou, Henan 450052, China. Nanozyme Laboratory in Zhongyuan, Zhengzhou, Henan 451163, China. -e-mail: yanxy@ibp.ac.cn; fankelong@ibp.ac.cn