استجابات مضادات الأكسدة في الذرة المتأثرة بالكروم كما تأثرت بتطبيقات حمض الفولفيك على الأوراق والجذور Antioxidant Responses in Chromium-Stressed Maize as Influenced by Foliar and Root Applications of Fulvic Acid
يواجه الذرة (Zea mays L.) تحديات كبيرة في نموه وإنتاجيته بسبب ضغط المعادن الثقيلة، وخاصة ضغط الكروم (Cr)، الذي يحفز توليد أنواع الأكسجين التفاعلية (ROS) ويؤدي إلى تلف الأنسجة الضوئية. كانت هذه الدراسة تهدف إلى التحقيق في تأثير تطبيق حمض الفولفيك (FA)، من خلال الرش الورقي أو ري الجذور، في التخفيف من ضغط الكروم على الذرة من خلال تقييم تأثيره على نشاط مضادات الأكسدة ومعايير النمو. تم تعريض نوعين من الذرة، P3939 و30Y87، لضغط الكروم. عند تركيزات من و لمدة 5 أسابيع. تم إجراء التجربة في بيت سلكي تحت ظروف بيئية طبيعية في مركز البذور، معهد علم النبات، جامعة البنجاب، لاهور، باكستان. شملت التقييمات الفسيولوجية تسرب الإلكتروليت، محتوى صبغة الكلوروفيل، مستويات المالونديالديهايد (MDA)، وأنشطة إنزيمات مضادات الأكسدة مثل الكاتالاز (CAT)، بيروكسيداز الأسكوربات (APX)، وبيروكسيداز الجوايacol (GPX) في أوراق الذرة. كما تم مراقبة معايير النمو. كشفت النتائج أن إجهاد الكروم قلل بشكل كبير من محتوى الكلوروفيل وزاد من الإجهاد التأكسدي، كما يتضح من ارتفاع مستويات MDA وتسرب الإلكتروليت. ومع ذلك، فإن تطبيق FA خفف بشكل ملحوظ من هذه التأثيرات: تحسن محتوى الكلوروفيل بنسبة، وانخفضت مستويات MDA بشكل ملحوظ. كانت الري باستخدام FA فعالة بشكل خاص، حيث خفضت مستويات MDA بنسبةمقارنة بـعلاج الكروم. علاوة على ذلك، بينما عزز إجهاد الكروم أنشطة إنزيمات مضادات الأكسدة، فإن تطبيق حمض الفوليك زاد من مستويات البروتين القابل للذوبان الكلي وأنشطة إنزيمات مضادات الأكسدة تحت ظروف الإجهاد. في الختام، يظهر تطبيق حمض الفوليك إمكانيته في تحسين تحمل الذرة لإجهاد المعادن الثقيلة من خلال تعزيز نظام الدفاع المضاد للأكسدة والحفاظ على أصباغ التمثيل الضوئي. تسلط هذه النتائج الضوء على وعد حمض الفوليك كاستراتيجية عملية للتخفيف من الآثار السلبية لإجهاد الكروم على الذرة، مما يعزز الممارسات الزراعية المستدامة في البيئات الملوثة.
الكلمات المفتاحية: إنزيمات مضادة للأكسدة، سمية الكروم، نمو الذرة، تلف أكسيدي، أنواع الأكسجين التفاعلية، تخفيف الإجهاد
تلوث المعادن الثقيلة هو مشكلة كبيرة في جميع أنحاء العالم بسبب تأثيراته السلبية على نمو النباتات والإنتاج وجودة الغذاء.تدخل المعادن الثقيلة إلى التربة من خلال الأنشطة الطبيعية (مثل البراكين والتجوية) والأنشطة البشرية، أي النفايات والمواد الكيميائية من الصناعات.في باكستان، يتم دفع تلوث المعادن الثقيلة بشكل أساسي من خلال التصنيع، والتحضر، وري المياه المستعملة، واستخدام المبيدات الحشرية. ومن الجدير بالذكر أن حوالي 600 مصنع دباغة جلود تعمل في كراتشي وكاسور وسيالكوت.تم تحديد الإفراج المفرط عن مياه الصرف الصحي والمخلفات من هذه المدابغ كمصدر رئيسي لتلوث التربة والمياه بالمعادن الثقيلة.الكروم هو معدن ثقيل منتشر يتم إطلاقه من عمليات الدباغة و
صناعات الطلاءالكروم (VI) ، على وجه الخصوص ، شديد السمية بسبب قدرته العالية على الأكسدة وقابليته للذوبان مقارنة بالكروم (III).تؤدي سمية الكروم في النباتات إلى آثار ضارة متنوعة، بما في ذلك تثبيط النمو، وتدهور الصبغات، وكبح نشاط إنزيمات مضادات الأكسدة (SOD، POD، APX، وCAT)، وتوليد أنواع الأكسجين التفاعلية (ROS)، التي تسبب تلفًا خلويًا. استجابةً لذلك، تعمل مضادات الأكسدة مثل حمض الأسكوربيك، والجلوثاثيون،تصبح -توكوفيرول، الفلافونويدات، والكاروتينات (CAR) ضرورية، حيث تلعب دورًا محوريًا في تحييد الجذور الحرة وتقليل الإجهاد التأكسدي.الإجهاد الناتج عن الكروم يعطل نقل الإلكترونات في الغشاء الثايلاكويدي أثناء عملية التمثيل الضوئي، مما يؤدي إلى الإجهاد التأكسدي وانخفاض القدرة على التمثيل الضوئي.إحدى الحلول المحتملة على المدى القصير للتخفيف من الآثار السلبية للكروم هي تطبيق تعديلات التربة أو الرش الورقي الذي يحتوي على مركبات مضادة للأكسدة..
تلعب المواد الهيومية، مثل حمض الفولفيك، دورًا أساسيًا في تحسين نمو النباتات، ونشاط الإنزيمات، وكيمياء التربة. حمض الفولفيك هو الجزء القابل للذوبان في الحمض من المواد الهيومية. تميل هذه المركبات العضوية إلى تكوين روابط مع المعادن الثقيلة، مما يقلل من الآثار الضارة للإجهاد المعدني على نمو النباتات.حمض الفولفيك يُستخدم كتحسين طبيعي للتربة، والذي يتكون من التحلل البيولوجي والكيميائي للنباتات.تشمل المجموعات الوظيفية الرئيسية لحمض الفولفيك الحلقات العطرية، والهيدروكسيلات الفينولية، ومجموعات الكربوكسيل.حمض الفولفيك يعزز نمو الذرة والتمثيل الضوئي بينما يقلل من امتصاص العناصر السامة من خلال تكوين روابط كيميائية مع المعادن الثقيلة. يمكن أن يعزز حمض الفولفيك إنبات البذور ويحفز عملية التمثيل الغذائي للنبات.. يعزز نشاط إنزيمات مضادات الأكسدة، بما في ذلك الكاتالاز (CAT) والبيروكسيداز (POD)، بالإضافة إلى إنزيمات أخرى مثل الفوسفاتاز القلوي (على سبيل المثال، تحسين تحريك الفوسفات في النباتات تحت ظروف الإجهاد)الذرة (Zea mays L.) هي واحدة من أكثر المحاصيل الحبوب المزروعة على نطاق واسع في العالم، وتلعب دورًا حيويًا في الأمن الغذائي، وعلف الحيوانات، والتطبيقات الصناعية.إنه غني بالنشا والبروتينات والألياف، مما يجعله ضروريًا لتغذية الإنسان والحيوان.نظرًا لأهميته الاقتصادية والتغذوية، يُزرع الذرة غالبًا في المناطق التي تواجه تحديات بيئية، بما في ذلك تلوث التربة بالمعادن الثقيلة. الكروم، وهو معدن ثقيل سام ومستمر، يمكن أن يؤثر بشكل كبير على نمو الذرة وتطورها، مما يؤدي إلى تقليل الغلات وتدهور الجودة الغذائية.كمحصول أساسي، يعتبر الذرة عرضة بشكل خاص لضغوط المعادن الثقيلة، التي تعيق وظائفه الفسيولوجية، بما في ذلك عملية التمثيل الضوئي ونشاط إنزيمات مضادات الأكسدة.على الرغم من أن الذرة معروفة بأنها عرضة للإجهاد الناتج عن المعادن الثقيلة، إلا أن الآليات المحددة التي يؤثر بها الكروم على وظائفها الفسيولوجية، بما في ذلك عملية التمثيل الضوئي ونشاط إنزيمات مضادات الأكسدة، ليست مفهومة تمامًا. كانت أهداف هذه الدراسة تقييم الآثار الضارة للكروم على نمو الذرة ونشاط إنزيمات مضادات الأكسدة، وتقييم الدور المحتمل لحمض الفولفيك في مواجهة سمية الكروم. بينما تم استكشاف استراتيجيات تخفيف مختلفة، لا يزال دور حمض الفولفيك في تعزيز الدفاع المضاد للأكسدة تحت ضغط الكروم في الذرة غير مستكشف بشكل كافٍ. تقترح الفرضية العاملة أن تطبيق حمض الفولفيك يعزز نظام الدفاع المضاد للأكسدة في الذرة تحت ضغط الكروم، مما يقدم نهجًا مبتكرًا لتحسين التحمل وتخفيف الضرر التأكسدي الناتج عن سمية الكروم.
المواد والأساليب
إعداد تجريبي وإجهاد الكروم مع تطبيق حمض الفولفيك في أصناف الذرة P3939 و30Y87
لتقييم تأثير حمض الفولفيك على صنفين من الذرة، تم إجراء تجربة في أصص في مركز البذور، معهد علم النبات، جامعة البنجاب، لاهور، باكستان، في نوفمبر 2018 باستخدام تصميم عشوائي كامل (CRD) كما هو موضح في الشكل 1. تم الحصول على بذور صنفي الذرة P3939 و30Y87 من شركة بايونير للبذور في ساهيوال. تم شراء حمض الفولفيك المستخدم في التجربة من سيغما-ألدريتش. تم زراعة الأصص في بيت سلكي تحت ظروف بيئية طبيعية، مع ري منتظم مرتين في الأسبوع ورش ورقي من N وP وK لتلبية المتطلبات الغذائية. شملت التجربة ثلاث تكرارات لكل معالجة. تم تحفيز إجهاد الكروم باستخدام كلوريد الكروم. و عن طريق إذابة الكميات المناسبة في الماء المقطر في مرحلة الورقة الرابعة. بعد أسبوع من التعرض لإجهاد المعادن الثقيلة، تم تطبيق حمض الفولفيك عن طريق الرش الورقي ( ) وتطبيق الجذر (تم تخصيصها عشوائيًا لأصص باستخدام مياه الري لتقليل التحيز المحتمل وتباين البيئة.
تقييم معايير النمو
لتحقيق تأثيرات حمض الفولفيك على تطوير أصناف الذرة تحت ضغط الكروم، تم تقييم عدة معايير نمو (طول الورقة، عرض الورقة، مساحة الورقة، ارتفاع النبات، وعدد الأوراق).
الحصاد
بعد أربعة أسابيع من تطبيق علاج حمض الفولفيك، تم حصاد نباتات الذرة وتقسيمها بعناية إلى سيقان وجذور. قبل الحصاد، تم جمع عينات من كل من الأوراق الشابة والناضجة تمامًا لكل نبات. ثم خضعت هذه العينات لتحليل مفصل لتقييم معايير النمو المختلفة والمؤشرات الكيميائية الحيوية. شمل هذا التحليل قياسات تسرب الإلكتروليت، ومحتوى المالونديالديهايد (MDA) (علامة على أكسدة الدهون)، ومحتوى صبغة الكلوروفيل، والبروتينات القابلة للذوبان الكلية (TSP)، وأنشطة إنزيمات مضادات الأكسدة. قدمت هذه التقييمات رؤى حول الاستجابات الفسيولوجية والتغيرات الكيميائية الحيوية التي أحدثها علاج حمض الفولفيك في نباتات الذرة.
تحديد محتوى الكلوروفيل
تم تحديد محتوى الكلوروفيل وفقًا لطريقة أرنون،مع تعديلات طفيفة. لتحديد محتوى الكلوروفيل، تم جمع 0.1 جرام من كل نسيج نباتي في أنابيب إيبندورف 1.5 مل مع وضع علامة، وتم إضافة 1 مل من DMSO (ثنائي ميثيل سلفوكسيد). بعد ذلك، تم وضع أنابيب الإيبندورف في الظلام لمدة 48 ساعة، ثم تم قياس امتصاص كل عينة عند 663 نانومتر و 645 نانومتر.
محتوى الكلوروفيل الكليالمذيبوزن النبات الطازج بالجرام.
الشكل 1. يتضمن التخطيط التجريبي نوعين من الذرة، P3939 و30Y87، تعرضا لمستويات مختلفة من إجهاد الكروم (Cr) وعلاجات بحمض الفولفيك. يوضح الشكل ترتيب الأواني ويسلط الضوء على الأعراض المرئية في نباتات الذرة المعرضة للكروم، مثل تغير لون الأوراق.
تحديد تسرب الإلكتروليت
تم إعداد عينات أوراق بحجم موحد باستخدام مثقاب الفلين ووضعها في 20 مل من الماء المقطر. ثم تم هز هذه الأنابيب لمدة 24 ساعة على جهاز هزاز مداري لكسر أغشية الخلايا. الموصلية الكهربائية (تم قياسه بعد 24 ساعة ثم تم تعقيمه بالبخار تحت الضغطلمدة ساعة واحدة قبل وضعها على هزاز مداري لـلقياس الموصلية الكهربائيةتم حساب تسرب الإلكتروليت باستخدام الصيغة التالية.
تسرب الإلكتروليت
تقييم محتوى MDA
تم قياس محتوى MDA ونشاط إنزيمات مضادات الأكسدة (CAT وAPX وGPX) باستخدام تقنية (He et al., 2001). تم سحق 0.25 جرام من الأوراق في 150 مللي مول من محلول الفوسفات (4 مل) ذو pH 7.0 وتم الطرد المركزي عندلـلتحديد محتوى MDA، تم خلط 1 مل من مستخلص الإنزيم مع 2 مل من محلول التفاعل الذي يحتوي على TCA و سيتم الإعلان عنه لاحقًا. بعد ذلك، تم تسخين المحلول في حمام مائي.لمدة 30 دقيقة قبل أن يتم الطرد المركزي عندلمدة 15 دقيقة. ثم تم تسجيل الامتصاص عند 532 و
تحديد إنزيمات مضادات الأكسدة (CAT، APX، و GPX)
تم تقييم نشاط الكاتالاز باستخدام نسخة معدلة من التقنية التي وصفها تشانس ومايلي.محلول التفاعل الذي يحتوي على pHبيروكسيد الهيدروجينمستخلص إنزيميتم جمعها. تم استخدام مقياس الطيف الضوئي لقياس التغيرات في الامتصاص لكل عينة عند 240 نانومتر.
نهجتم استخدامه مع بعض التغييرات لتحديد نشاط APX.محلول التفاعل 100 مللي مولار أسيتات الصوديوم (رقم الهيدروجيني 5.8)،حمض الإيثيلين diamine tetraacetic (EDTA)حمض الأسكوربيك، وتم جمع المحلول المستخرج وبدأت التفاعل بإضافة مستخلص الإنزيم. تم استخدام مقياس الطيف الضوئي لقياس التغيرات في الامتصاص عند 290 نانومتر كل 10 ثوانٍ لمدة 60 ثانية.
تم تقييم نشاط GPX باستخدام نهج مع تعديلات طفيفةمحلول التفاعل الذي يتضمنمحلول منظم أسيتات الصوديومجوايكول، و تم جمع مستخلص الإنزيم. تم استخدام مقياس الطيف الضوئي لقياس التغيرات في الامتصاص عند 460 نانومتر كل 10 ثوانٍ لمدة 60 ثانية.
تحديد إجمالي البروتين القابل للذوبان
تم حساب محتوى البروتين القابل للذوبان في الورقة عن طريق إضافة السائل العلويمن مستخلص مادة نباتية ثم تم خلطه مع 3 مل من مادة ملونة. بعد 5 دقائق، تم قياس امتصاص كل عينة عند.
التحليل الإحصائي
باستخدام برنامج SAS (نظام التحليل الإحصائي)، تم تحليل البيانات إحصائيًا باستخدام إجراء المزيج (PROC MIXED) وإجراء النموذج الخطي العام (PROC GLM) (معهد SAS). تم استخدام اختبار نطاق دنكان المتعدد للكشف عن الفروق في المتوسطات. تم إجراء تحليلات الارتباط والمكونات الرئيسية باستخدام RStudio.
النتائج
تأثيرات حمض الفولفيك على معايير نمو أصناف الذرة
الدراسة بحثت في تأثيرات العلاجات المختلفة على معايير النمو لاثنين من أصناف الذرة، P3939 و30Y87. لوحظت اختلافات كبيرة في طول الورقة، عرض الورقة، مساحة الورقة، ارتفاع النبات، وعدد الأوراق عبر العلاجات (الجدول 1). بالنسبة لكلا الصنفين، أظهر مجموعة التحكم قيمًا متوسطة لمعظم المعايير. تطبيقانخفاض ملحوظ في طول الورقة، عرض الورقة، مساحة الورقة، وارتفاع النبات مقارنةً بالتحكم، مما يشير إلى تأثير سلبي على النمو. على العكس، العلاجات التي تجمع بينمع تطبيق حمض الفولفيك كرش ورقي (FFA، ) أو من خلال وسيط الجذر (IFA، ) أظهرت تأثيرات متباينة. تطبيق عموماً خففت بعض الآثار السلبية المرتبطة بالتعرض للكروم، في حين أنأسفرت عن نتائج مختلطة، أحيانًا تزيد من
علاجات
النوع P3939
النوع 30Y87
طول الورقة (سم)
عرض الورقة (سم)
مساحة الورقة ( )
ارتفاع النبات (سم)
عدد الأوراق
طول الورقة (سم)
عرض الورقة (سم)
مساحة الورقة ( )
ارتفاع النبات (سم)
عدد الأوراق
تحكم ( )
أ-ب
أب
أب
FFA
أ
أ
أ
أ
أ
IFA
أب
أ
أب
أ-ب
أ-ب
أ-ب
أ-ب
e
ج-د
ج-د
كر + ففا
ب-ج
ب-ج
ب-ج
ب-ج
كروم + الحديد
ج-د
ج-د
ب-ج
ب-ج
ب-ج
ب-ج
أ-ج
ب-ج
د-هـ
ب-هـ
ب-ج
د-هـ
د-هـ
د-هـ
كر + ففا
ج-د
ج-د
ب-ج
ب-ج
ج-د
ج
ج-د
كروم + الحديد
د-هـ
ج-د
ج-د
الجدول 1. تأثيرات حمض الفولفيك المطبق كرش ورقي (FFA، ) ومن خلال وسيلة الري الجذرية (IFA، ) على معايير النمو لأسلاف الذرة (P3939 و30Y87). تمثل القيم المتوسط ± الخطأ المعياري (SE)، مع الإشارة إلى الاختلافات المعنوية بين المعالجات باستخدام حروف صغيرة.
الآثار الضارة للكروم. بشكل عام، تسلط هذه النتائج الضوء على التفاعلات المعقدة بين التعرض للكروم وطرق تطبيق حمض الفولفيك على مقاييس نمو هذه الأصناف من الذرة.
آثار حمض الفولفيك على محتوى الكلوروفيل في أصناف الذرة
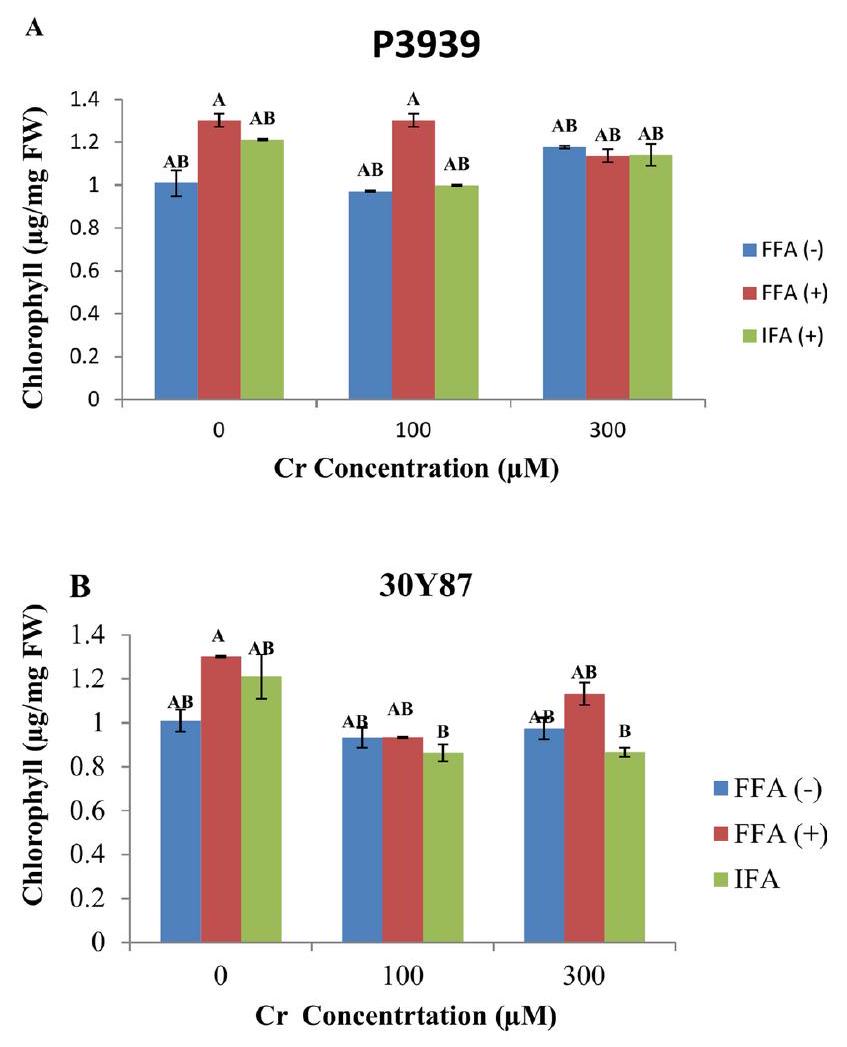
تم قياس تركيز الكلوروفيل في كلا الصنفين من الذرة عبر تركيزات مختلفة من الكروم وعلاجات التطبيق الورقي. في الصنف P3939، تحت الظروف العادية (كانت مستويات الكلوروفيل متسقة نسبيًا عبر العلاجات: FW بدون FFA، إعادة توجيه مع FFA، وإعادة توجيه مع IFA. ومع ذلك، التعرض لـأدى الكروم إلى انخفاض في تركيز الكلوروفيل عبر جميع المعاملات: FW بدون FFA، إعادة توجيه مع FFA، وإعادة توجيه مع IFA. فيتفاوتت تركيزات الكلوروفيل بشكل أكبر: FW بدون FFA، إعادة توجيه مع FFA، و FW مع IFA (الشكل 2A). تحت ظروف السيطرة (في الصنف 30Y87، اختلفت مستويات الكلوروفيل مع العلاجات الورقية: FW بدون FFA، إعادة توجيه مع FFA، وإعادة توجيه مع IFA. التعرض لـأدى إلى انخفاض في تركيز الكلوروفيل عبر جميع المعالجات: FW بدون FFA، إعادة توجيه مع FFA، وإعادة توجيه مع IFA. فياستمرت تركيزات الكلوروفيل في الانخفاض: FW بدون FFA، إعادة توجيه مع FFA، وتمت الموافقة على ذلك مع IFA. تشير هذه النتائج إلى أنه بينما حافظت تطبيقات الفولفيك أسيد (FFA) على مستويات الكلوروفيل أو زادتها قليلاً مقارنةً بالمجموعة الضابطة، فإن التعرض للكروم قلل بشكل مستمر من إنتاج الكلوروفيل (الشكل 2B).
آثار حمض الفولفيك على الإجهاد التأكسدي (تسرب الإلكتروليت ومحتوى MDA) في أصناف الذرة
نسبة تسرب الإلكتروليت (%)
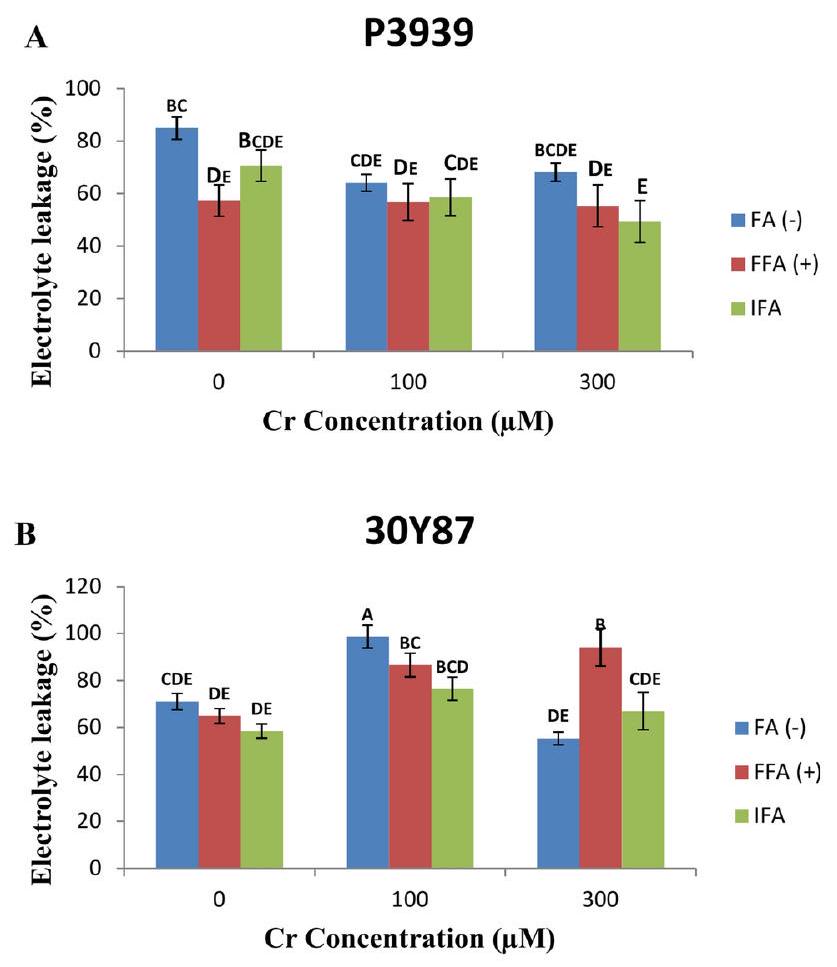
في صنف الذرة P3939، تم فحص تأثير تركيزات الكروم على نسبة تسرب الإلكتروليت عبر علاجات التطبيق الورقي المختلفة. تحت الظروف العادية (نسب تسرب الإلكتروليت كانتبدون حمض الفولفيك، مع FFA، و مع IFA (حمض الفولفيك من خلال وسط الجذور). مع زيادة تركيز الكروم إلىانخفض تسرب الإلكتروليت إلىبدون مع FFA، و مع IFA. عند أعلى مستوى من الكروم تم اختبارهأظهرت تسرب الإلكتروليت اتجاهات مختلطة:بدون FFA، مع FFA، و مع IFA (الشكل 3A).
في صنف الذرة 30Y87، تم فحص نسب تسرب الإلكتروليت عبر تركيزات مختلفة من الكروم وعلاجات التطبيق الورقي. عندزيادة تسرب الإلكتروليت مقارنةً بالتحكم
الشكل 2. تأثيرات حمض الفولفيك المطبق كرش ورقي (FFA، ) ومن خلال ري وسط الجذور (IFA، 300 لتر/هكتار) على محتوى الكلوروفيل في أصناف الذرة P3939 (A) و30 Y87 (B) تحت ضغط الكروم. القيم تمثل المتوسط ± الخطأ المعياري (SE)، مع وجود حروف صغيرة تشير إلى الفروق المعنوية بين المعاملات. -FFA تشير إلى عدم تطبيق حمض الفولفيك (التحكم). +FFA تمثل تطبيق حمض الفولفيك ورقيًا، تم رشه على الأوراق. IFA تشير إلى تطبيق حمض الفولفيك من خلال الري، يتم توصيله عبر وسط الجذور.
الشكل 3. تأثيرات حمض الفولفيك المطبق كرش ورقي (FFA، ) ومن خلال ري وسط الجذور (IFA، 300 لتر/هكتار) على تسرب الإلكتروليت (EL) لاصناف الذرة P3939 (A) و30Y87 (B) تحت ضغط الكروم. القيم تمثل المتوسط ± الخطأ المعياري (SE)، مع وجود حروف صغيرة تشير إلى الفروق المعنوية بين المعاملات. -FFA تشير إلى عدم تطبيق حمض الفولفيك (التحكم). +FFA تمثل تطبيق حمض الفولفيك ورقيًا، تم رشه على الأوراق. IFA تشير إلى تطبيق حمض الفولفيك من خلال الري، يتم توصيله عبر وسط الجذور.
Cr) عبر جميع العلاجات: FFA ( – ) زادت بمقدارFFA ( + ) بواسطة، و IFA ( + ) بواسطة . ومع ذلك، في ، كانت الاستجابات متنوعة: أظهرت FFA ( – ) انخفاضًا في التسرب بنسبةFFA ( + ) أظهر زيادة قدرها، وأظهر IFA (+) زيادة متواضعة قدرها (الشكل 3B).
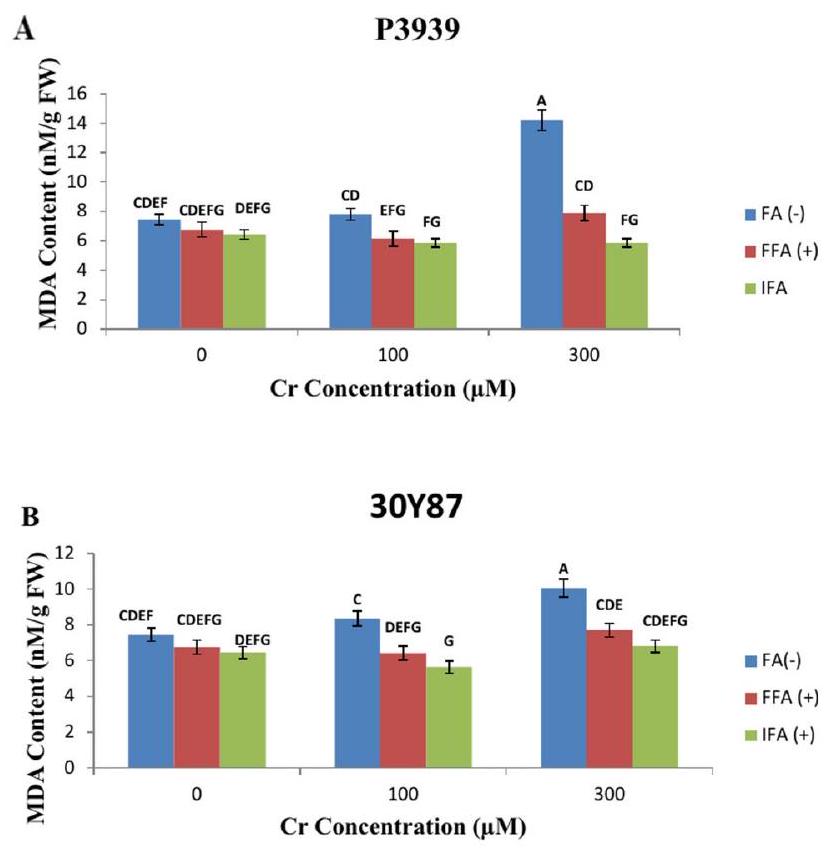
محتوى MDA (نانو مول/غرام وزن رطب) تظهر سلالة الذرة P3939 تغييرات واضحة في محتوى MDA (مالونديالديهايد) (نانو مول/غرام وزن رطب) استجابة لتركيزات مختلفة من الكروم وحمض الفولفيك.، يبدأ محتوى MDA للصنف من تحت ظروف FA(-). مع زيادة تركيز الكروم إلى تظهر مستويات MDA زيادة طفيفة إلىومع ذلك، يحدث زيادة ملحوظة أكثر عند، حيث يرتفع محتوى MDA بشكل ملحوظ إلىتشير هذه الملاحظات إلى أن صنف الذرة P3939 يظهر استجابة تعتمد على التركيز للكروم (Cr) (الشكل 4A). يظهر صنف الذرة 30 Y 87 تغييرات واضحة في محتوى MDA (مالونديالديهايد) (نانو مول/غرام وزن رطب) استجابة لتركيزات مختلفة من الكروم (Cr) وعلاجات ورقية مختلفة مع حمض الفولفيك (FA). تحت ظروف السيطرة ( )، يتم تسجيل محتوى MDA عند عند التعرض لـ بدون أي معالجة ورقية (FA-)، يزيد محتوى MDA إلى إعادة توجيه. من المثير للاهتمام، أن إضافة حمض الفولفيك الورقي (FFA+) بنفس تركيز الكروم (يؤدي إلى انخفاض طفيف في MDA إلىإعادة توجيه، مما يشير إلى تأثير مخفف محتمل للأحماض الدهنية الحرة ضد الإجهاد التأكسدي الناتج عن الكروم. مزيد من التحليل فيمع حمض الفولفيك المروي (IFA+) يظهر زيادة في محتوى MDA إلى FW، مما يدل على زيادة معتدلة مقارنة بالتحكم (الشكل 4B).
آثار حمض الفولفيك على محتوى البروتين القابل للذوبان الكلي (TSP) (ملغ/غ من الوزن الطازج)
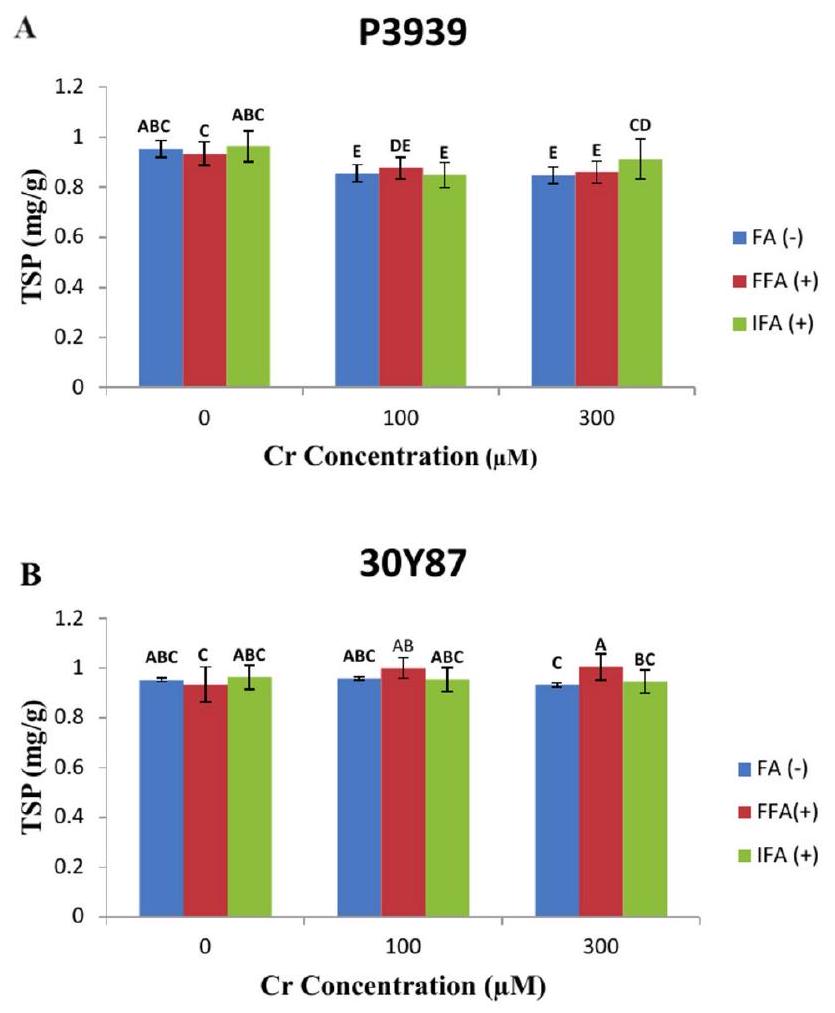
يظهر صنف الذرة P3939 مستويات متفاوتة من البروتين القابل للذوبان الكلي (TSP) ) استجابة لتركيزات مختلفة من الكروم ومعاملات الأوراق مع حمض الفولفيك (FA). تحت ظروف طبيعية دون مستويات TSP هيلـ FA ( – )لـ FFA ( + )، ولـ IFA (+). مع زيادة تركيز الكروم إلىيوجد انخفاض ملحوظ في TSP عبر جميع العلاجات الورقية:لـ FA (-)لـ FFA (+)، ولـ IFA (+). تشير هذه الانخفاضات إلى أن إجهاد الكروم قد يؤثر سلبًا على تخليق البروتين أو استقراره في صنف الذرة P3939. عند أعلى تركيز للكروم تم اختباره (تستمر مستويات TSP في الانخفاض:لـ FA( – )لـ FFA ( + )، ولـ IFA ( + ) (الشكل 5A). تحت الظروف العادية دونتظل مستويات TSP مستقرة نسبيًا عبر جميع العلاجات الورقية:لـ FA( –لـ FFA، و لـ IFA. ومع ذلك، مع زيادة تركيز الكروم إلى و تظهر مستويات TSP تقلبات طفيفة بين العلاجات. عندتزداد مستويات الكروم و TSP إلىلـ FA (-)لـ FFA (+)، ولـ IFA (+). وهذا يشير إلى استجابة محتملة للصنف تجاه الضغط المعتدل الناتج عن الكروم، ربما
الشكل 4. تأثيرات حمض الفولفيك المطبق كرش ورقي (FFA، ) ومن خلال ري وسط الجذور (IFA، 300 لتر/هكتار) على محتوى MDA من أصناف الذرة P3939 (A) و30Y87 (B) تحت ضغط الكروم. القيم تمثل المتوسط ± الخطأ المعياري (SE)، مع وجود حروف صغيرة تشير إلى الفروق المعنوية بين المعاملات. -FFA تشير إلى عدم تطبيق حمض الفولفيك (التحكم). +FFA تمثل تطبيق حمض الفولفيك ورقيًا، تم رشه على الأوراق. IFA تشير إلى تطبيق حمض الفولفيك من خلال الري، يتم توصيله عبر وسط الجذور.
تأثير على استقلاب البروتين أو تخليقه. عندتنخفض مستويات TSP قليلاً إلىلـ FA(-)لـ FFA ( + )، ولـ IFA ( + )، مما يشير إلى تأثير أكثر وضوحًا لتركيزات الكروم الأعلى على مستويات البروتين في نباتات الذرة (الشكل 5B).
تأثيرات حمض الفولفيك على نشاط الإنزيمات المضادة للأكسدة تحت ضغط الكروم
في النتائج، وُجد أن حمض الفولفيك لعب دورًا مهمًا في تعديل نشاط الإنزيمات المضادة للأكسدة في النباتات تحت ظروف إجهاد الكروم. على وجه الخصوص، تم ملاحظة أن الإنزيمات مثل الكاتالاز (CAT) وأكسيداز الأسكوربات (APX) وأكسيداز الجلوتاثيون (GPX) كانت مكونات حاسمة في نظام الدفاع المضاد للأكسدة الذي ساعد في التخفيف من الضرر التأكسدي الناجم عن سمية الكروم.
نشاط الكاتالاز (وحداتبروتين
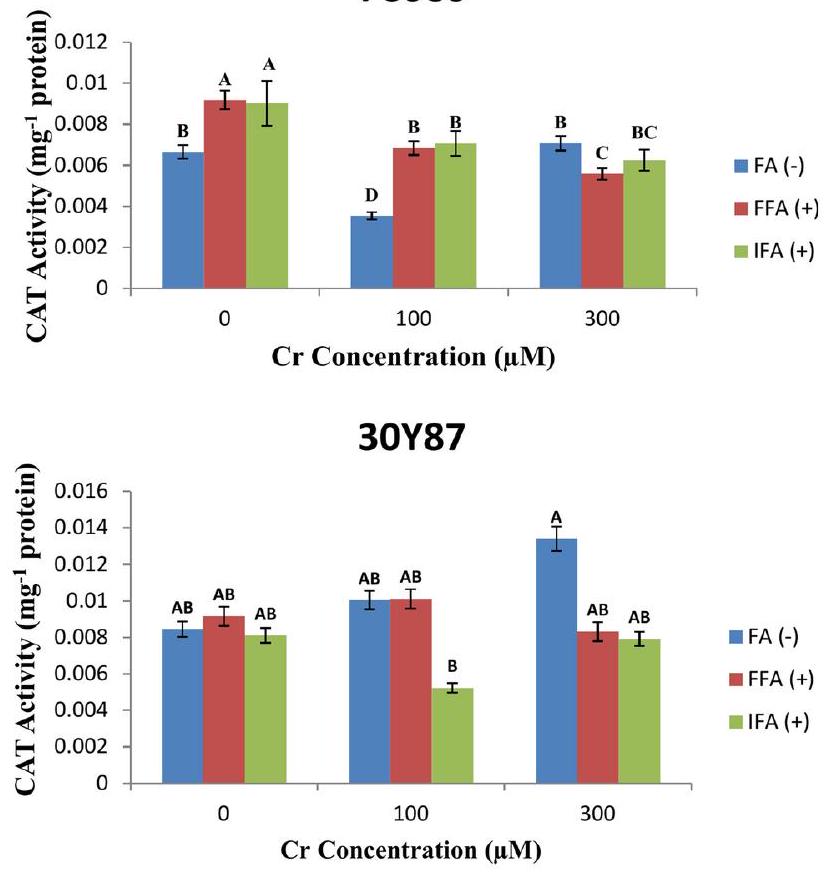
تظهر سلالة الذرة P3939 نشاطًا متغيرًا لإنزيم الكاتالاز (CAT)البروتين) استجابة لتركيزات مختلفة من الكروم ومعاملات الأوراق مع حمض الفولفيك (FA). تحت ظروف السيطرة دونتُقاس مستويات نشاط CAT عندبروتين لـ FA( –بروتين لـ FFA ( + )، وبروتين للاختبار المناعيمع زيادة تركيز الكروم إلى و تُلاحظ التغيرات في نشاط CAT عبر جميع العلاجات الورقية. عندتنخفض نشاط CAT إلىبروتين لـ FA (-)بروتين لـ FFA (+)، وبروتين لـ IFA (+). تشير هذه الانخفاض إلى احتمال كبح وظيفة إنزيم CAT تحت ظروف ضغط الكروم المعتدل. عندتظهر أنشطة CAT مزيدًا من التباينات:بروتين ل FAبروتين لـ FFA ( + )، والبروتين لـ IFA (+) (الشكل 6A). يظهر صنف الذرة 30 Y 87 مستويات متفاوتة من نشاط الكاتالاز (CAT)البروتين) استجابة لتركيزات مختلفة من الكروم (Cr) ومعاملات ورقية مع حمض الفولفيك (FA). تحت ظروف طبيعية بدونتُسجل مستويات نشاط القطط عندبروتين لـ FA( –بروتين للـ FFA، و البروتين لـ IFA ( + ). مع زيادة تركيز الكروم إلىتلاحظ تغييرات في نشاط CAT عبر العلاجات الورقية. يزيد نشاط CAT قليلاً إلىبروتين لـ FA (-)، يبقى مستقرًا عند 0.010112بروتين لـ FFA (+) ، وينخفض إلىبروتين لـ IFA (+). فيتُلاحظ مزيد من التغيرات في نشاط CAT:بروتين لفابروتين للـ FFA، و بروتين لـ IFA ( + ) (الشكل 6B).
نشاط APX (الوحداتبروتين
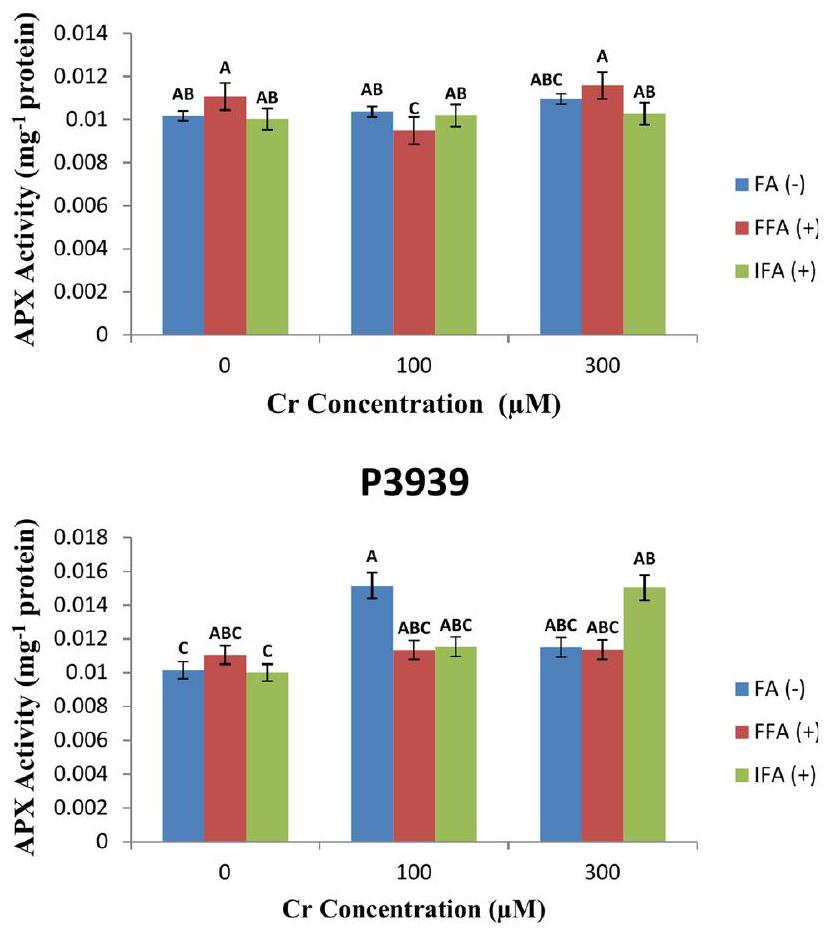
تظهر سلالة الذرة P3939 نشاطًا متغيرًا لإنزيم أسكوربات بيروكسيداز (APX)البروتين) استجابة لتركيزات مختلفة من الكروم ومعاملات الرش بحمض الفولفيك (FA). تحت الظروف العادية دون الكرومتُقاس مستويات نشاط APX عندبروتين لـ FA(-)بروتين للـ FFA، و البروتين لـ IFA ( + ). مع زيادة تركيز الكروم إلىتظهر نشاط APX تقلبات عبر العلاجات الورقية. يزيد نشاط APX إلىبروتين لـ FA( – )، يبقى
الشكل 5. تأثيرات حمض الفولفيك المطبق كرش ورقي (FFA، ) ومن خلال وسيلة الري الجذرية (IFA، ) على إجمالي البروتين القابل للذوبان (TSP) لاصناف الذرة P3939 (A) و 30Y87 (B) تحت ضغط الكروم. القيم تمثل المتوسط ± الخطأ المعياري (SE)، مع وجود حروف صغيرة تشير إلى الفروق المعنوية بين المعاملات. -FFA تشير إلى عدم تطبيق حمض الفولفيك (التحكم). +FFA تمثل تطبيق حمض الفولفيك عن طريق الرش على الأوراق. IFA تشير إلى تطبيق حمض الفولفيك من خلال الري، المقدم عبر وسط الجذور.
مستقر نسبيًا عندبروتين لـ FFA (+) ، ويزداد قليلاً إلىبروتين للاختبار المناعي. في تُلاحظ مزيد من التغيرات في نشاط APX:بروتين لـ FA(-)، 0.011373بروتين لـ FFA(+) والبروتين لـ IFA (+) (الشكل 7A). يظهر صنف الذرة 30 Y 87 مستويات متفاوتة من نشاط أسكوربات بيروكسيداز (APX) (البروتين) استجابة لتركيزات مختلفة من الكروم (Cr) ومعاملات ورقية مع حمض الفولفيك (FA). تحت ظروف السيطرة دونمستويات نشاط APX متسقة نسبيًا:بروتين لـ FA( –بروتين للـ FFA، و البروتين لـ IFA (+). مع زيادة تركيز الكروم إلىتظهر نشاط APX تقلبات طفيفة عبر العلاجات الورقية. ينخفض نشاط APX قليلاً إلىبروتين لـ FA(-)، ينخفض إلىبروتين لـ FFA (+) ، ويزداد بشكل طفيف إلىبروتين لـ IFA( + ).عندتُلاحظ مزيد من التغيرات في نشاط APX:بروتين لـ FA(-)بروتين لـ FFA( + )، وبروتين لـ IFA ( + ) (الشكل 7B).
نشاط GPX (الوحداتبروتين)
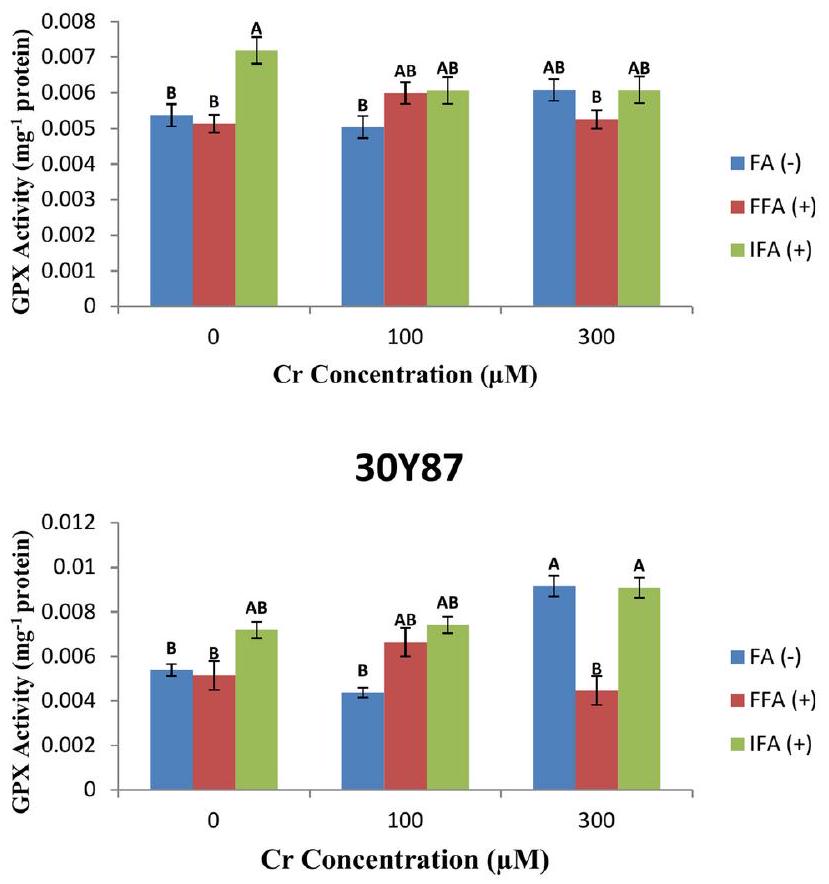
تم تقييم صنف الذرة P3939 لاستجابته لتركيزات مختلفة من الكروم، مع التركيز على النشاط الإنزيمي لبيروكسيداز الجلوتاثيون (GPX). تدرس الدراسة نشاط GPX من حيث الميكرو مولات ( ) من الكروم بتركيزات 0 و 100 و . تشير النتائج إلى أنه مع زيادة تركيزات الكروم، تظل نشاط GPX في صنف الذرة P3939 ضمن نطاق ثابت، مما يشير إلى استجابة مضادة للأكسدة قوية. تظهر مستويات FA (-) و FFA (+) و IFA (+) في الصنف استجابات مختلفة للتعرض للكروم، مع ملاحظات لتقلبات عبر العلاجات. على وجه التحديد، يظهر FA (-) زيادة طفيفة من 0.005377 إلىالبروتين مع ارتفاع مستويات الكروم من 0 إلىبينماوأظهرت FA( – ) و IFA( + ) استجابات أكثر تنوعًا (الشكل 8A). تم دراسة صنف الذرة 30 Y87 لاستجابته لتركيزات متزايدة من الكروم (Cr)، مع التركيز على نشاط إنزيم الجلوتاثيون بيروكسيداز (GPX) ومستويات FA ( – ) و FFA ( + ) و IFA ( + ) في أنسجته. قامت الدراسة بقياس نشاط GPX بوحدات لكل ملليجرام من البروتين عبر تركيزات Cr تبلغ 0 و 100 و. تشير النتائج إلى استجابات متباينة في نشاط GPX مع زيادة مستويات الكروم، مما يوحي بقدرة مضادة للأكسدة معقدة في الصنف 30Y87. على وجه التحديد، يتقلب نشاط GPX، مع انخفاض ملحوظ من 0.007192 إلىالبروتين مع زيادة تركيز الكروم من 0 إلى. هذه التباينات تبرز قدرة الصنف على التكيف مع الإجهاد التأكسدي الناتج عن الكروم. من حيث مستويات FA ( – ) و FFA ( + ) و IFA ( + )، يظهر الصنف 30Y87 استجابات مختلفة عبر علاجات الكروم. مستويات FA (-) تزيد بشكل ملحوظ من 0.005377بروتين فيإلىبروتين فيبينما تظهر FFA (+) و IFA (+) أنماط تغيير أكثر تعقيدًا (الشكل 8B).
الشكل 6. تأثيرات حمض الفولفيك المطبق كرش ورقي (FFA، ) ومن خلال ري وسط الجذور (IFA، 300 لتر/فدان) على إنزيم الكاتالاز (CAT) لاصناف الذرة P3939 (A) و30 Y87 (B) تحت ضغط الكروم. القيم تمثل المتوسط ± الخطأ المعياري (SE)، مع وجود حروف صغيرة تشير إلى الفروق المعنوية بين المعاملات. -FFA تشير إلى عدم تطبيق حمض الفولفيك (التحكم). +FFA تمثل تطبيق حمض الفولفيك ورقيًا، تم رشه على الأوراق. IFA تشير إلى تطبيق حمض الفولفيك من خلال الري، يتم توصيله عبر وسط الجذور.
تحليل المكونات الرئيسية (PCA) وتحليل الارتباط
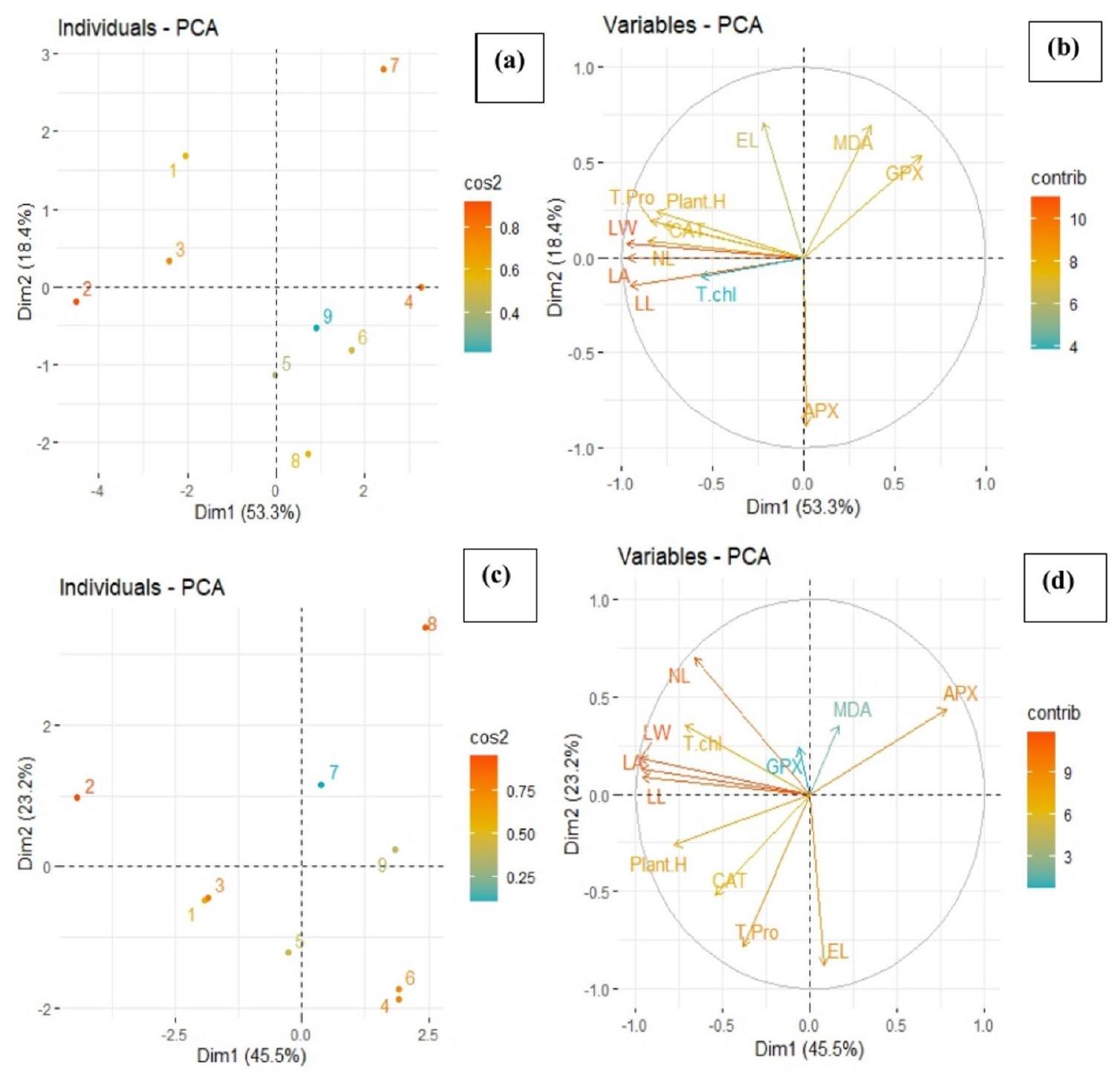
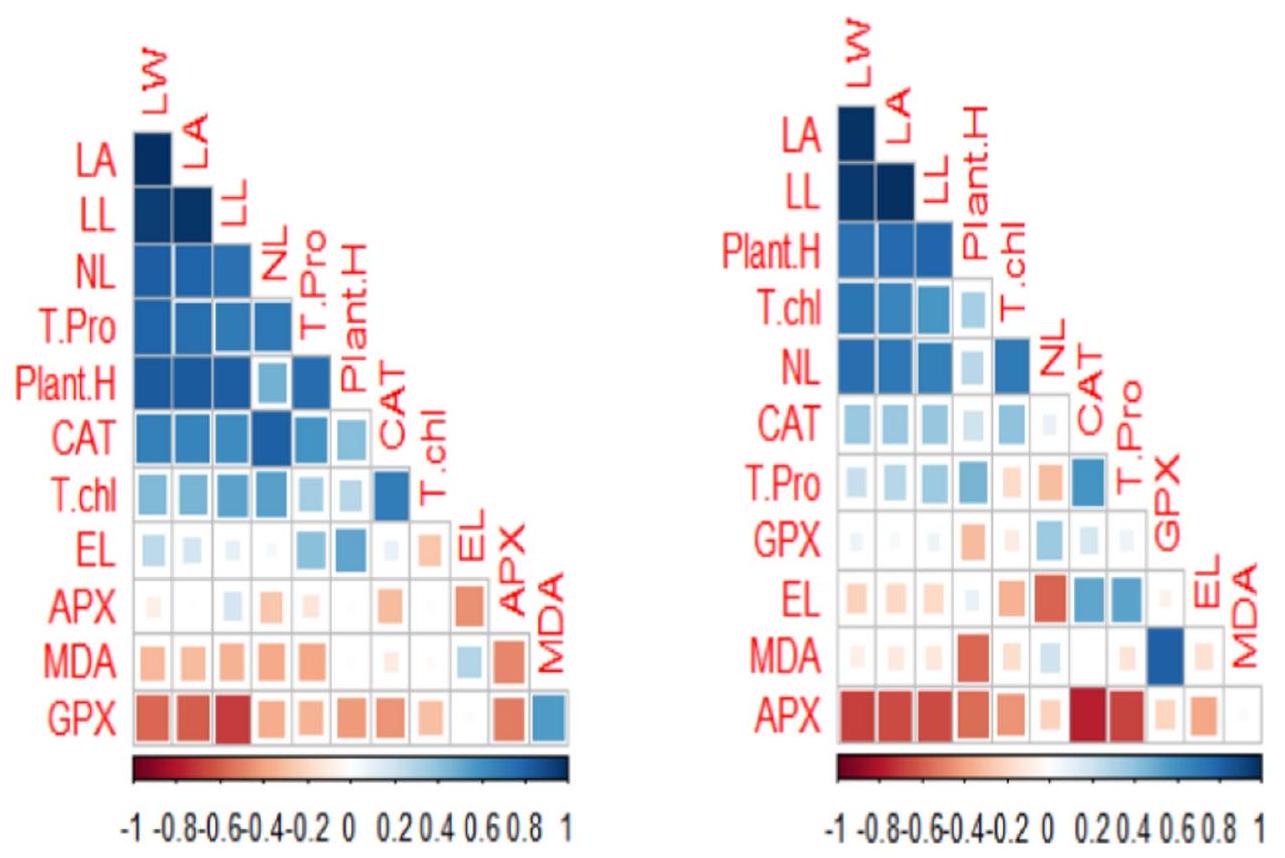
استخدمت هذه الدراسة تحليل المكونات الرئيسية (PCA) وتحليل الارتباط لتقييم المعايير البيوكيميائية والشكلية لأسلاف الذرة تحت مستويات مختلفة من إجهاد الكروم وتطبيق حمض الفولفيك (FFA) و/أو حمض الفولفيك غير العضوي (IFA) من خلال طرق الرش والري (الشكل 9). تم توحيد جميع المعايير لضمان قابلية المقارنة بين المتغيرات من خلال التقييس إلى متوسط صفر وتباين وحدة. تم تحليل درجات المكونات الرئيسية لتحديد الأنماط والعلاقات بين المعايير عبر العلاجات التجريبية المختلفة. ساعد ذلك في فهم كيفية تأثير إجهاد الكروم وتطبيقات حمض الفولفيك على الخصائص البيوكيميائية والشكلية لأسلاف الذرة. تم حساب معاملات ارتباط بيرسون لتحديد قوة واتجاه العلاقات الخطية بين أزواج المعايير داخل كل مجموعة تجريبية (الشكل 10).
نقاش
في هذه الدراسة، بحثنا في دور أوضاع تطبيق حمض الفولفيك المختلفة على النشاط المضاد للأكسدة للذرة (Zea mays L.) المعرضة لإجهاد الكروم الاصطناعي. يعتبر تلوث الكروم في التربة الزراعية مصدر قلق بيئي كبير، حيث يسبب آثارًا ضارة على نمو النباتات وإنتاجيتها.تم تطبيق حمض الفولفيك، المعروف بخصائصه الخلبية وقدرته على تعزيز تحمل النباتات للإجهاد، من خلال طرق مختلفة لتقييم فعاليته في التخفيف من الإجهاد التأكسدي الناتج عن الكروم في نباتات الذرة.تعتبر النشاط المضاد للأكسدة في نباتات الذرة مؤشراً حاسماً على قدرتها على مواجهة تراكم أنواع الأكسجين التفاعلية (ROS) تحت ظروف إجهاد الكروم.من خلال دراسة تأثيرات طرق تطبيق حمض الفولفيك المختلفة، تهدف هذه الدراسة إلى تقديم رؤى حول تحسين الممارسات الزراعية لإنتاج المحاصيل المستدام في التربة الملوثة بالكروم. لوحظت زيادة غير دالة في معايير النمو في النباتات المعالجة بالمعادن والنباتات المعالجة بالكروم مع تطبيق حمض الفولفيك. ومع ذلك، قد تُعزى هذه النتائج غير الدالة إلى التباين الفطري بين العلاجات أو مرحلة تطور النبات في وقت التقييم. أشارت نتائجنا إلى انخفاض في معايير النمو في النباتات المعالجة بالكروم. هذه النتائج تتماشى مع عمل أرشد وآخرون.الذين لاحظوا انخفاضًا في معايير النمو في براسكا جونسيه تحت ضغط الكروم. الشغايهي وآخرون.كما تم الإبلاغ عن انخفاض في معايير النمو في نباتات القمح المعالجة بمعدن النحاس. يمكن أن تُعزى هذه الانخفاضات في معايير النمو إلى التغيرات فوق الهيكلية والفيزيائية الكيميائية في النباتات المعرضة لإجهاد الكروم.يرتبط هذا الانخفاض في مقاييس النمو، مثل طول الساق والجذر والكتلة الحيوية، بتقليل امتصاص العناصر الغذائية.تشير نتائجنا إلى انخفاض في مساحة الورقة تحت ضغط الكروم. يمكن ربط هذا الانخفاض في مساحة الورقة بتقييد توفر المياه الذي يؤثر على توسع الورقة، مشابهًا للملاحظات تحت نقص البوتاسيوم.لقد أظهرت سمية الفلورايد في القمح أيضًا أنها تقلل من مساحة الورقة..
في تجربتنا، أظهر تطبيق حمض الفولفيك الورقي زيادة في معايير النمو المختلفة مثل طول الورقة، وعرضها، ومساحتها، وعدد الأوراق. دراستنا تتماشى مع الدراسات السابقة التي أظهرت
الشكل 7. تأثيرات حمض الفولفيك المطبق كرش ورقي (FFA، ) ومن خلال ري وسط الجذور (IFA، 300 لتر/هكتار) على إنزيم الأسكوربات (APX) لاصناف الذرة P3939 (A) و30Y87 (B) تحت ضغط الكروم. القيم تمثل المتوسط ± الخطأ المعياري (SE)، مع وجود حروف صغيرة تشير إلى الفروق المعنوية بين المعاملات. -FFA تشير إلى عدم تطبيق حمض الفولفيك (التحكم). +FFA تمثل تطبيق حمض الفولفيك ورقيًا، تم رشه على الأوراق. IFA تشير إلى تطبيق حمض الفولفيك من خلال الري، يتم توصيله عبر وسط الجذور.
تأثيرات تطبيق حمض الفولفيك على نمو النباتات ومحتوى العناصر الغذائية. على سبيل المثال، ركزت إحدى الدراسات على تأثير التطبيق الورقي لحمض الفولفيك على نمو النباتات وجودة ثمار الطماطم (Lycopersicon esculentum L.). بينما درست دراسة أخرى تأثيرات تطبيق حمض الفولفيك في التربة على نمو ومحتوى العناصر الغذائية لنباتات الخيار (Cucumis sativus).إن قدرة الأحماض الدهنية على اختراق جذور النباتات وتحفيز التعبير عن الجينات المرتبطة بانقسام الخلايا تدعم دورها في تعزيز النمو الخضري..
تم تقليل محتوى الكلوروفيل في نباتات كلا الصنفين من الذرة بسبب إجهاد الكروم. يتماشى هذا الانخفاض مع النتائج التي تم العثور عليها في القمح (Triticum aestivum) تحت إجهاد المضادات الحيوية.تؤدي نتائج إجهاد الكروم إلى إنتاج أنواع الأكسجين التفاعلية التي تتفاعل مع مجمعات الصبغ والبروتين وتلحق الضرر بأغشية الثايلاكويد للكلوروفيل بسبب استبدال المغنيسيوم بـأيونات. يبرز هذا الاضطراب البيوكيميائي ضعف الآلات الضوئية تحت ضغط المعادن الثقيلة. من ناحية أخرى، أظهرت تطبيقات حمض الفولفيك على الأوراق زيادة في محتوى الكلوروفيل. على سبيل المثال، تمت ملاحظة زيادة في محتوى الكلوروفيل بسبب تطبيق حمض الفولفيك على الأوراق فيفي نباتات الذرة من الصنف P3939. علاوة على ذلك، أدى تطبيق حمض الفولفيك على الأوراق إلىزيادة في محتوى الكلوروفيل في نباتات الذرة المزروعة تحت ظروف طبيعية. هذه النتائج مشابهة لبعض الدراسات السابقة، حيث تم ملاحظة زيادة في محتوى الكلوروفيل من خلال تطبيق حمض الفولفيك في نباتات الذرةتشير هذه الزيادة إلى أن حمض الفولفيك يخفف من الضرر التأكسدي عن طريق تقليل تركيز أيونات الكروم الحرة من خلال الامتصاص.
تسرب الإلكتروليت هو مؤشر على الضرر الخلوي الناتج عن ضغط المعادن الثقيلة وضغوط غير حيوية أخرى. أدى إضافة الكروم () إلى وسط النمو إلى زيادة تسرب الإلكتروليت في أوراق كلا صنفي الذرة. الزيادة في التسرب تتماشى مع الإفراط في إنتاج أنواع الأكسجين التفاعلية تحت ضغط المعادن، مما يساهم في الضرر التأكسدي. ومع ذلك، أظهر تطبيق حمض الفولفيك (تطبيق ورقي + ري) تقليل تسرب الإلكتروليت في النباتات المعالجة بالكروم. تم ملاحظة حواليانخفاض في تسرب الإلكتروليت مع تطبيق حمض الفولفيك عندمقارنة معالعلاج بمفرده. محتوى المالونديالديهايد هو مؤشر موثوق للضغط التأكسدي الناتج عن الضغوط غير الحيوية، وضغط المعادن الثقيلة وهجوم مسببات الأمراض. تم ملاحظة زيادة كبيرة في محتوى المالونديالديهايد () عندتركيزات في كلا الصنفين. يتماشى هذا مع النتائج في عباد الشمس واللفت، حيث أدى الضرر التأكسدي الناتج عن الكروم إلى ارتفاع مستويات المالونديالديهايدحيث تفاعلت أنواع الأكسجين التفاعلية الناتجة عن الكروم مع الدهون، مما تسبب في تدمير الغشاء والضرر الخلوي. أظهر تطبيق حمض الفولفيك تقليل محتوى المالونديالديهايد في نباتات الكروم من كلا الصنفين. ومع ذلك، وُجد أن تطبيق حمض الفولفيك من خلال التربة كان أكثر فعالية في تخفيف الضغط التأكسدي عن طريق إزالة السموم من الجذور الحرة.
تشير نتائجنا إلى انخفاض نسبي في محتوى البروتين القابل للذوبان الكلي عندوالعلاجات مقارنة بالتحكم. كما أفادت دراسة سابقة بانخفاض في محتوى البروتين القابل للذوبان الكلي تحت ضغط الكروم. قد يكون هذا الانخفاض في البروتين القابل للذوبان الكلي ناتجًا أيضًا عن زيادة امتصاص الكروم مما يتداخل مع استقرار البروتين. من ناحية أخرى
الشكل 8. تأثيرات تطبيق حمض الفولفيك كرش ورقي (FFA،) ومن خلال ري وسط الجذور (IFA،) على إنزيم الجلوتاثيون بيروكسيداز (GPX) من أصناف الذرة P3939 (A) و30Y87 (B) تحت ضغط الكروم. تمثل القيم المتوسط ± الخطأ القياسي (SE)، مع الحروف الصغيرة تشير إلى اختلافات معنوية بين العلاجات. -FFA تشير إلى عدم تطبيق حمض الفولفيك (التحكم). +FFA تمثل تطبيق حمض الفولفيك على الأوراق، ورشها على الأوراق. IFA تشير إلى تطبيق حمض الفولفيك من خلال الري، المقدم عبر وسط الجذور.
من ناحية أخرى، أدى تطبيق حمض الفولفيك من خلال وسط الجذور إلى تعزيز محتوى البروتين القابل للذوبان الكلي في النباتات المعالجة بالمعادن. هذه النتائج تتماشى مع عمل أسكاري وآخرون.الذين سجلوا أيضًا زيادة في محتوى البروتين القابل للذوبان الكلي في نباتات القمح المعالجة بالكروم. قد يكون الزيادة في البروتين القابل للذوبان الكلي بسبب التأثيرات الإيجابية لحمض الفولفيك على نمو النبات وأصباغ التمثيل الضوئي. يبرز هذا التحسن دور حمض الفولفيك في دعم تخليق البروتين واستقراره تحت الضغط. في دراستنا التي تحقق في دور أوضاع تطبيق حمض الفولفيك المختلفة على النشاط المضاد للأكسدة للذرة تحت ضغط الكروم الاصطناعي، تم ملاحظة زيادة كبيرة في إنزيمات ومركبات مضادة للأكسدة الرئيسية. على وجه التحديد، أظهرت مستويات الكاتالاز (CAT)، أسكوربات بيروكسيداز (APX)، بيروكسيداز (POD)، وغلوتاثيون ريدوكتاز (GPX) تعزيزًا ملحوظًا. تلعب هذه الإنزيمات أدوارًا حاسمة في تخفيف الضغط التأكسدي عن طريق تحلل أنواع الأكسجين التفاعلية، ومنع الضرر الخلوي، والحفاظ على التوازن الخلوي. الكاتالاز هو مكون مهم من نظام الدفاع المضاد للأكسدة الذي يشارك في تحلل بيروكسيد الهيدروجين الناتج أثناء التنفس الضوئي وأكسدة الأحماض الدهنية. أشارت نتائجنا إلى عدم وجود زيادة معنوية في نشاط الكاتالاز في نباتات الكروم من كلا الصنفين. تم تسجيل زيادة في نشاط الكاتالاز عند تركيز أعلى من الكروم أيمقارنة مع التحكم والكروم. جابين وآخرون.سجلوا أيضًا زيادة في نشاط الكاتالاز في نباتات الفاصوليا تحت ضغط الكروم. قد يكون الانخفاض في نشاط الكاتالاز عندبسبب أن الكاتالاز هو جزيء بروفيرين حديد وقد يتفاعل الكروم مع الحديد مما يؤثر على توفر الشكل النشط من الحديد. أظهر تطبيق حمض الفولفيك (ورقي) زيادة في نشاط الكاتالاز عندتركيز. تم ملاحظة زيادة حواليفي نشاط الكاتالاز مع تطبيق حمض الفولفيك الورقي عندمقارنة معالعلاج بمفرده. البيروكسيداز الذي يحللباستخدام الأسكوربات كمانح للإلكترون يعرف باسم أسكوربات بيروكسيداز. APX هو إنزيم مضاد للأكسدة أساسي موجود في البيروكسيسومات، السيتوسول والكلوروبلاست. أظهرت نتائجنا انخفاضًا في نشاط APX عند تركيز أعلى من. تم الإبلاغ عن انخفاض في APX أيضًا في الخردل الهندي, زيت بذور اللفتوZea mays. قد يكون سبب الانخفاض في نشاط APX هو أن سمية الكروم العالية يمكن أن تثبط نظام الدفاع المضاد للأكسدة المسؤول عن إزالة السموم من أنواع الأكسجين التفاعلية. ومع ذلك، زاد تطبيق حمض الفولفيك على الأوراق من نشاط APX في نباتات الصنف 30 Y 87 تحت ضغط الكروم. تتماشى هذه النتائج مع عمل وانغ وآخرون.الذين سجلوا أيضًا زيادة في نشاط APX مع تطبيق حمض الفولفيك في النباتات المعالجة بالمعادن. تم ملاحظة زيادة حواليفي نشاط APX مع تطبيق حمض الفولفيك الورقي عندالعلاج. GPX هو إنزيم مضاد للأكسدة يقوم بتحليلباستخدام الجوايacol كمانح للإلكترون. يلعب GPX دورًا حيويًا في التصلب؛ الدفاع وشفاء الجروح. لم يتغير نشاط GPX بسبب ضغط الكروم أو التطبيق الخارجي لحمض الفولفيك. قد تكون الزيادة في نشاط إنزيمات مضادة للأكسدة (CAT وGPX) تحت ضغط الكروم استجابة لتوليد الجذور الفائقة من خلال انسداد الكروم لسلسلة نقل الإلكترون في الميتوكوندريا. قد يكون الانخفاض في نشاط إنزيم مضاد للأكسدة (APX) بسبب التأثيرات المثبطة لأيونات الكروم على الإنزيم. في تجربتنا، حسن تطبيق حمض الفولفيك من نشاط إنزيمات مضادة للأكسدة (CAT وAPX)
الشكل 9. مخطط النقاط (أ، ج) ومخططات التحميل (ب، د) لتحليل المكونات الرئيسية (PCA) على مختلف المعلمات المدروسة لأصناف الذرة المزروعة في تربة تحت ضغط الكروم. يمثل مخطط النقاط فصل العلاجات كـ (1) التحكم (بدون تلوث بالكروم)؛ (2) FFA (3) IFA (4)مستوى الكروم، 5)7)8)9)على التوالي. الاختصارات للمعلمات هي كما يلي: ارتفاع النبات: ارتفاع النبات؛ NL: عدد الأوراق؛ LW: عرض الورقة؛ T chl: الكلوروفيل الكلي؛ EL: تسرب الإلكتروليت؛ MDA: أكسدة الدهون؛ T pro: البروتين الكلي؛ CAT: الكاتالاز؛ APX: أسكوربات بيروكسيداز GPX: بيروكسيداز الجوايacol.
تحت ضغط الكروم. قد يكون ذلك بسبب أن حمض الفولفيك يعمل ككاشف للجذور الحرة ومضاد للأكسدة. يمتلك حمض الفولفيك القدرة على التفاعل مع كل من الإلكترونات غير المتزاوجة المشحونة إيجابًا وسلبًا وبالتالي إزالة السموم من الجذور الحرة قد يكون بسبب زيادة امتصاص العناصر الغذائية.
الاستنتاج
تؤثر سمية الكروم سلبًا على نشاط إنزيمات مضادة للأكسدة، وأصباغ التمثيل الضوئي، ونمو النبات بشكل عام. أدى تطبيق حمض الفولفيك (المروي) إلى زيادة في محتوى الكلوروفيل والبروتينات القابلة للذوبان الكلي، على الرغم من أن هذه النتائج لم تكن ذات دلالة إحصائية. بالإضافة إلى ذلك، قلل حمض الفولفيك من محتوى المالونديالديهايد (MDA) وتسرب الإلكتروليت في النباتات المعالجة بالكروم. زاد تطبيق حمض الفولفيك على الأوراق بشكل كبير من نشاط إنزيمات مضادة للأكسدة، وخاصة الكاتالاز (CAT) وأسکوربات بيروكسيداز (APX)، بينما لم يؤثر على نشاط بيروكسيداز الجوايacol في النباتات المعالجة بالمعادن. تشير الزيادة في نشاط CAT وAPX إلى أن تطبيق حمض الفولفيك على الأوراق قد يحسن من مقاومة النبات ضد ضغط الكروم من خلال إزالة أنواع الأكسجين التفاعلية بشكل فعال. تشمل الآلية المحتملة لعمل حمض الفولفيك في هذا السياق تعزيز نشاط إنزيمات مضادة للأكسدة، مما يساعد في تخفيف الضغط التأكسدي الناتج عن التعرض للكروم. على الرغم من أن ضغط الكروم أدى إلى تقليل معلمات النمو، إلا أن تطبيق حمض الفولفيك من خلال الأوراق والري الجذري كان له تأثير إيجابي على مقاييس النمو المختلفة. نوصي بتطبيق حمض الفولفيك
الشكل 10. العلاقة بين النمو والمعايير الفسيولوجية لأسلاف الذرة المزروعة في تربة متأثرة بالكروم.
كأسلوب أكثر فعالية للتخفيف من سمية الكروم، نظرًا لتأثيره الكبير على نظام الدفاع المضاد للأكسدة. يجب أن تستكشف الأبحاث المستقبلية الآثار طويلة الأمد لحمض الفولفيك الورقي مقابل الري، فضلاً عن إمكانيته في تخفيف أنواع أخرى من الضغوط غير الحيوية.
توفر البيانات
جميع البيانات التي تم إنشاؤها أو تحليلها خلال هذه الدراسة مدرجة في هذه المقالة المنشورة.
تاريخ الاستلام: 25 سبتمبر 2024؛ تاريخ القبول: 27 ديسمبر 2024
تم النشر على الإنترنت: 08 يناير 2025
References
Fatima, A., Shabaan, M., Ali, Q., Malik, M., Asghar, H.N., Aslam, M., Zulfiqar, U., Hameed, A., Nazim, M., Mustafa, A.E.Z.M. and Elshikh, M.S. Integrated application of metal tolerant . fluorescens and press mud for conferring heavy metal tolerance to aloe vera (Aloe barbadensis). Plant Stress, 11, 100333 (2024).
Sarwar, M.J., Shabaan, M., Asghar, H.N., Ayyub, M., Ali, Q., Zulfiqar, U., Nazim, M., Alarjani, K.M. and Elshikh, M.S. Interaction of chromium (Cr) resistant plant growth promoting rhizobacteria with compost to phytostabilize Cr in spinach rhizosphere. Plant Stress, 10, 100261 (2023).
Ali, H. Q. et al. Tanneries impact on groundwater quality: A case study of Kasur city in Pakistan. Environ. Monit. Assess. 194(11), 823 (2022).
Mehdiniya Afra, J. et al. Influence of chemical fertilizers and animal manure on morphological traits of medicinal plants in Northern Iran. Res. Crop Ecophysiol. 17(2), 86-101 (2022).
Laxmi, V. & Kaushik, G. Toxicity of hexavalent chromium in environment, health threats, and its bioremediation and detoxification from tannery wastewater for environmental safety. In Bioremediation of industrial waste for environmental safety: volume I: industrial waste and its management (eds Saxena, G. & Bharagava, R. N.) 223-243 (Springer Singapore, 2020).
Zulfiqar, U., Haider, F.U., Ahmad, M., Hussain, S., Maqsood, M.F., Ishfaq, M., Shahzad, B., Waqas, M.M., Ali, B., Tayyab, M.N. and Ahmad, S.A., 2023. Chromium toxicity, speciation, and remediation strategies in soil-plant interface: A critical review. Front Plant Sci. 13, 1081624 (2023).
Arshed, M. Z., Zulfiqar, A. & Saleem, A. Alleviation of chromium stress in Brassica juncea L. and soil remediation by plant growth promoting (PGP) bacteria. Pak. J. Sci. Ind. Res. Ser. Biol. Sci. 67(1), 42-48 (2024).
Sheteiwy, M. S. et al. Elevated differentially attenuates beryllium-induced oxidative stress in oat and alfalfa. Physiol. Plant. 175(5), e14036 (2023).
Hussnain, M., Shabaan, M., Ali, Q., Ashraf, S., Ahmad, M., Ghafoor, U., Akhtar, M.J., Zulfiqar, U., Hussain, S., Al-Ashkar, I. and Elshikh, M.S. Microbial phytoremediation of chromium-contaminated soil with biogas slurry for enhancing the performance of Vigna radiata L. Plant Stress, 10, 100206 (2023).
Younis, U., Danish, S., Datta, R., Alahmadi, T. A. & Ansari, M. J. Sustainable remediation of chromium-contaminated soils: Boosting radish growth with deashed biochar and strigolactone. BMC Plant Biol. 24(1), 115 (2024).
Nardi, S., Schiavon, M. & Francioso, O. Chemical structure and biological activity of humic substances define their role as plant growth promoters. Molecules 26(8), 2256 (2021).
Gong, G. et al. Extraction of fulvic acid from lignite and characterization of its functional groups. ACS Omega 5(43), 27953-27961 (2020).
Shahrajabian, M. H. & Sun, W. The importance of salicylic acid, humic acid and fulvic acid on crop production. Lett. Drug Des. Discov. 21(9), 1465-1480 (2024).
Dawood, M. F., Tahjib-Ul-Arif, M., Sohag, A. A. M. & Abdel Latef, A. A. H. Role of acetic acid and nitric oxide against salinity and lithium stress in Canola (Brassica napus L.). Plants 13(1), 51 (2023).
Jabeen, N. et al. Glycinebetaine mediates chromium tolerance in mung bean through lowering of Cr uptake and improved antioxidant system. Arch. Agron. Soil Sci. 62(5), 648-662 (2016).
Abu-Omar, M. M. High-valent iron and manganese complexes of corrole and porphyrin in atom transfer and dioxygen evolving catalysis. Dalton Trans. 40(14), 3435-3444 (2011).
Sousa, R. H. et al. Impairment of peroxisomal APX and CAT activities increases protection of photosynthesis under oxidative stress. J. Exp. Bot. 70(2), 627-639 (2019).
Nouairi, I. et al. Antioxidant defense system in leaves of Indian mustard (Brassica juncea) and rape (Brassica napus) under cadmium stress. Acta Physiol. Plant. 31, 237-247 (2009).
Wu, Z. et al. Antioxidant enzyme systems and the ascorbate-glutathione cycle as contributing factors to cadmium accumulation and tolerance in two oilseed rape cultivars (Brassica napus L.) under moderate cadmium stress. Chemosphere 138, 526-536 (2015).
Chugh, V., Kaur, N., Gupta, A. K. Evaluation of oxidative stress tolerance in maize (Zea mays L.) seedlings in response to drought (2011).
Dawood, M. F., Tahjib-Ul-Arif, M., Sohag, A. A. M. & Abdel Latef, A. A. H. Fluoride mitigates aluminum-toxicity in barley: Morpho-physiological responses and biochemical mechanisms. BMC Plant Biol. 22(1), 287 (2022).
Bala, S., Asthir, B. & Bains, N. Activities of cell wall peroxidases in relation to lignification in six wheat (Triticum aestivum L.) genotypes under heat and drought stress. J. Environ. Biol. 37(3), 437 (2016).
Yilmaz, S. H., Kaplan, M., Temizgul, R. & Yilmaz, S. Antioxidant enzyme response of sorghum plant upon exposure to aluminum, chromium and lead heavy metals. Turk. J. Biochem. 42(4), 503-512 (2017).
Wang, W., Chen, M., Wang, D., Yan, M. & Liu, Z. Different activation methods in sulfate radical-based oxidation for organic pollutants degradation: Catalytic mechanism and toxicity assessment of degradation intermediates. Sci. Total Environ. 772, 145522 (2021).
الشكر والتقدير
تم دعم الدراسة من قبل وزارة العلوم والتنمية التكنولوجية والابتكار في جمهورية صربيا (المنح رقم 451-03-66/2024-03/200032). يود المؤلفون أن يعبروا عن تقديرهم لمشروع دعم الباحثين رقم (RSP2025R390)، جامعة الملك سعود، الرياض، المملكة العربية السعودية.
مساهمات المؤلفين
التصور، ف.إ، أ.ز، وأ.ك؛ المنهجية، ف.إ؛ البرمجيات، أ.س، وم.ز.أ؛ التحقق والتحليل الرسمي، أ.س، وف.إ؛ الموارد، إ.د، و ب.ف.ب؛ تنسيق البيانات، أ.ز؛ إعداد المسودة الأصلية، ف.إ، أ.ز، أ.س، أ.ز، وم.ز.أ؛ الكتابة-المراجعة والتحرير، إ.د، ب.ف.ب، و و.س؛ الإشراف، أ.ز. جميع المؤلفين قرأوا ووافقوا على النسخة المنشورة من المخطوطة.
التمويل
تم دعم الدراسة من قبل وزارة العلوم والتنمية التكنولوجية والابتكار في جمهورية صربيا (المنح رقم 451-03-66/2024-03/200032). يود المؤلفون أن يعبروا عن تقديرهم لمشروع دعم الباحثين رقم (RSP2025R390)، جامعة الملك سعود، الرياض، المملكة العربية السعودية.
الإعلانات
موافقة الأخلاقيات والموافقة على المشاركة
تم تنفيذ جميع الطرق وفقًا للإرشادات واللوائح ذات الصلة. لقد حصلنا على إذن لجمع المواد النباتية والشتلات.
المصالح المتنافسة
يعلن المؤلفون عدم وجود مصالح متنافسة.
معلومات إضافية
يجب توجيه المراسلات وطلبات المواد إلى أ.ز أو إ.د.
معلومات إعادة الطبع والتصاريح متاحة على www.nature.com/reprints.
ملاحظة الناشر تظل Springer Nature محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
الوصول المفتوح هذه المقالة مرخصة بموجب رخصة المشاع الإبداعي للاستخدام غير التجاري، والتي تسمح بأي استخدام غير تجاري، ومشاركة، وتوزيع، وإعادة إنتاج في أي وسيلة أو صيغة، طالما أنك تعطي الائتمان المناسب للمؤلفين الأصليين والمصدر، وتوفر رابطًا لرخصة المشاع الإبداعي، وتوضح إذا قمت بتعديل المادة المرخصة. ليس لديك إذن بموجب هذه الرخصة لمشاركة المواد المعدلة المشتقة من هذه المقالة أو أجزاء منها. الصور أو المواد الأخرى من طرف ثالث في هذه المقالة مشمولة في رخصة المشاع الإبداعي للمقالة، ما لم يُذكر خلاف ذلك في سطر الائتمان للمادة. إذا لم تكن المادة مشمولة في رخصة المشاع الإبداعي للمقالة واستخدامك المقصود غير مسموح به بموجب اللوائح القانونية أو يتجاوز الاستخدام المسموح به، ستحتاج إلى الحصول على إذن مباشرة من صاحب حقوق الطبع والنشر. لعرض نسخة من هذه الرخصة، قم بزيارة http://creativecommons.org/licenses/by-nc-nd/4.0/.
معهد علم النبات، جامعة البنجاب، لاهور، باكستان.قسم الزراعة، كلية الزراعة والبيئة، الجامعة الإسلامية في بهاولبور، بهاولبور 63100، باكستان.معهد المحاصيل الحقلية والخضروات، المعهد الوطني لجمهورية صربيا، ماكسيم غوركي 30، 21000 نوفي ساد، صربيا.قسم الزراعة، جامعة ولاية كانساس، 108 قاعة ووترز، 1603 مكان كلافلين القديم، مانهاتن، KS 66506، الولايات المتحدة الأمريكية.قسم إنتاج النباتات، كلية علوم الأغذية والزراعة، جامعة الملك سعود، 11451 الرياض، المملكة العربية السعودية.البريد الإلكتروني: usman.zulfiqar@iub.edu.pk; ivica.djalovic@ifvcns.ns.ac.rs
Antioxidant Responses in Chromium-Stressed Maize as Influenced by Foliar and Root Applications of Fulvic Acid
Farwa Iftikhar , Asma Zulfiqar , Atif Kamran , Ammara Saleem , Muhammad Zeeshan Arshed , Usman Zulfiqar , Ivica Djalovic , P. V. Vara Prasad & Walid Soufan
Abstract
Maize (Zea mays L.) faces significant challenges to its growth and productivity from heavy metal stress, particularly Chromium (Cr) stress, which induces reactive oxygen species (ROS) generation and damages photosynthetic tissues. This study aimed to investigate the effects of fulvic acid (FA) application, via foliar spray or root irrigation, on mitigating chromium stress in maize by evaluating its impact on antioxidant activity and growth parameters. Two maize varieties, P3939 and 30Y87, were subjected to chromium stress at concentrations of and for a duration of 5 weeks. The experiment was conducted in a wire house under natural environmental conditions at the Seed Centre, Institute of Botany, University of the Punjab, Lahore, Pakistan. Physiological assessments included electrolyte leakage, chlorophyll pigment content, malondialdehyde (MDA) levels, and activities of antioxidant enzymes such as catalase (CAT), ascorbate peroxidase (APX), and guaiacol peroxidase (GPX) in maize leaves. Growth parameters were also monitored. The results revealed that chromium stress significantly reduced chlorophyll content and increased oxidative stress, as evidenced by elevated MDA levels and electrolyte leakage. However, FA application notably mitigated these effects: chlorophyll content improved by , and MDA levels decreased significantly. Irrigation with FA was particularly effective, reducing MDA levels by compared to the chromium treatment. Furthermore, while chromium stress enhanced antioxidant enzyme activities, FA application further boosted total soluble protein levels and antioxidant enzyme activities under stress conditions. In conclusion, FA application demonstrates potential in improving maize tolerance to heavy metal stress by enhancing the antioxidant defense system and preserving photosynthetic pigments. These findings highlight FA’s promise as a practical strategy for mitigating the negative impacts of chromium stress on maize, promoting sustainable agricultural practices in contaminated environments.
Heavy metal contamination is a major problem across the world owing to its negative impacts on plant development, production, and food quality . Heavy metals are introduced into soil by both natural (volcanism and weathering) and human activities, i.e., wastes and chemicals from industries . In Pakistan, heavy metal contamination is primarily driven by industrialization, urbanization, wastewater irrigation, and the use of pesticides. Notably, approximately 600 leather tanneries are operating in Karachi, Kasur, and Sialkot . The excessive release of wastewater and effluents from these tanneries has been identified as a major source of soil and water contamination with heavy metals . Chromium is a widespread heavy metal released by tanning and
plating industries . Chromium (VI), in particular, is highly toxic due to its high oxidation potential and solubility compared to chromium (III) . Chromium toxicity in plants leads to various harmful effects, including inhibited growth, pigment degradation, suppression of antioxidant enzyme activity (SOD, POD, APX, and CAT), and the generation of reactive oxygen species (ROS), which cause cellular damage. In response, antioxidants like ascorbic acid, glutathione, -tocopherol, flavonoids, and carotenoids (CAR) become essential, as they play a pivotal role in neutralizing ROS and mitigating oxidative stress . Chromium-induced stress disrupts electron transport in the thylakoidal membrane during photosynthesis, leading to oxidative stress and a decrease in photosynthetic capacity . One potential short-term solution to mitigate chromium’s adverse effects is the application of soil amendments or foliar sprays containing antioxidant compounds .
Humic substances, such as fulvic acid, play an essential role in improving plant growth, enzyme activity, and soil chemistry. Fulvic acid is the acid-soluble fraction of humic substances. These organic compounds have a tendency to form bonds with heavy metals, thereby reducing the detrimental impacts of metal stress on plant development . Fulvic acid is used as natural soil amendment which is formed by biological and chemical decomposition of plants . The major functional groups of fulvic acid include aromatic rings, phenolic hydroxyls and carboxyl groups . Fulvic acid promotes maize growth and photosynthesis while reducing toxic element uptake by forming chemical bonds with heavy metals. Fulvic acid can enhance seed germination and stimulate plant metabolism . It enhances antioxidant enzyme activity, including catalase (CAT) and peroxidase (POD), as well as other enzymes like alkaline phosphatase (e.g., improving phosphate mobilization in plants under stress conditions) .Maize (Zea mays L.) is one of the most widely cultivated cereal crops globally, playing a crucial role in food security, animal feed, and industrial applications . It is rich in starch, proteins, and fiber, making it crucial for both human and animal nutrition . Due to its economic and nutritional importance, maize is often grown in regions that face environmental challenges, including soil contamination from heavy metals. Chromium, a persistent and toxic heavy metal, can severely affect maize growth and development, leading to reduced yields and compromised nutritional quality . As a staple crop, maize is particularly vulnerable to heavy metal stress, which inhibits its physiological functions, including photosynthesis and antioxidant enzyme activity . Although maize is known to be vulnerable to heavy metal stress, the specific mechanisms by which chromium affects its physiological functions, including photosynthesis and antioxidant enzyme activity, are not fully understood. The objectives of this study were to evaluate the detrimental effects of chromium on maize growth and antioxidant enzyme activity and to assess the potential mitigating role of fulvic acid in counteracting chromium toxicity. While various mitigation strategies have been explored, the role of fulvic acid in enhancing antioxidant defense under chromium stress in maize remains underexplored. The working hypothesis proposes that fulvic acid application enhances maize’s antioxidant defense system under chromium stress, offering an innovative approach to improve tolerance and alleviate oxidative damage caused by chromium toxicity.
Materials and methods
Experimental setup and chromium stress with fulvic acid application in maize cultivars P3939 and 30Y87
To evaluate the impact of fulvic acid on two maize cultivars, a pot experiment was conducted at the Seed Centre, Institute of Botany, University of the Punjab, Lahore, Pakistan, in November 2018 utilizing a complete randomized design (CRD) as shown in Fig. 1. Seeds of maize cultivars P3939 and 30Y87 were sourced from Pioneer Seed Company in Sahiwal. The fulvic acid used in the experiment was purchased from Sigma-Aldrich. Pots were grown in a wire house under natural environmental conditions, with regular irrigation twice a week and a foliar spray of N, P, K applied to meet nutritional requirements. The experiment included three replicates for each treatment. Chromium stress was induced using chromium chloride and by dissolving the appropriate amounts in distilled water at the 4th leaf stage. After one week of exposure to heavy metal stress, fulvic acid applications via foliar spray ( ) and root application ( ) with irrigated water, were randomly allocated to pots to minimize potential bias and environmental variability.
Evaluation of growth parameters
To investigate the effects of fulvic acid on the development of maize cultivars under to chromium stress, several growth parameters (leaf length, leaf breadth, leaf area, plant height, and number of leaves) were evaluated.
Harvesting
After four weeks of applying fulvic acid treatment, maize plants were harvested and carefully divided into shoots and roots. Prior to harvesting, samples were collected from both young and fully matured leaves of each plant. These samples were then subjected to detailed analysis to assess various growth parameters and biochemical indicators. This analysis included measurements of electrolyte leakage, malondialdehyde (MDA) content (a marker of lipid peroxidation), chlorophyll pigment content, total soluble proteins (TSP), and activities of antioxidant enzymes. These assessments provided insights into the physiological responses and biochemical changes induced by the fulvic acid treatment in maize plants.
Determination of chlorophyll content
Chlorophyll content was determined according to the method of Arnon, with slight modifications. For determination of chlorophyll content, leaf 0.1 g of each plant tissue was collected in labelled Eppendorf tubes 1.5 mL and DMSO (Dimethyl sulfoxide.) 1 mL was added. After the Eppendorf were placed in the dark for 48 h , the absorbance of each sample was then measured at 663 nm and 645 nm .
Total Chlorophyll Content of solvent gram fresh weight of the plant.
Fig. 1. The experimental layout includes two maize cultivars, P3939 and 30Y87, subjected to different chromium ( Cr ) stress levels and treatments with fulvic acid. The figure depicts the arrangement of the pots and highlights the visual symptoms in maize plants exposed to Cr , such as leaf discoloration.
Electrolyte-leakage determination
Leaf samples of uniform size were made using a cork borer and put in 20 mL distilled water. These tubes were then shaken for 24 h on an orbital shaker to break the cell membranes. The electrical conductivity ( ) was measured after 24 h then autoclaved at for 1 h before being put on an orbital shaker for to measure electrical conductivity . Electrolyte leakage was calculated by using following formula.
Electrolyte leakage
Evaluation of MDA content
MDA content and antioxidant enzyme activity (CAT, APX, and GPX) were measured using the technique of (He et al., 2001). 0.25 g of leaves were crushed in 150 mM phosphate buffer ( 4 mL ) having pH 7.0 and centrifuged at for . For MDA content determination, 1 ml of enzyme extract was mixed with 2 ml of a reaction solution containing TCA and TBA. After that, the solution was heated in a water bath for 30 min before being centrifuged at for 15 min . The absorbance then recorded at 532 and
Determination of antioxidant enzymes (CAT, APX, and GPX)
Catalase activity was assessed using a modified version of the technique described by Chance and Maehly . Reaction solution containing pH , hydrogen peroxide , and enzyme extract were collected. A spectrophotometer was used to measure changes in absorbance of each sample at 240 nm .
The approach of with certain changes was used to determine APX activity. of the reaction solution 100 mM sodium acetate ( pH 5.8), , Ethylenediaminetetracetic acid (EDTA) ascorbic acid, and extracted solution) was collected and the reaction was started by adding enzyme extract. A spectrophotometer was used to measure changes in absorbance at 290 nm every 10 s for 60 s .
With minor adjustments, GPX activity was assessed using the approach of reaction solution comprising sodium acetate buffer, guaiacol, , and enzyme extract were collected. Spectrophotometer was used to measure changes in absorbance at 460 nm every 10 s for 60 s .
Determination of total soluble protein
The total soluble protein content of the leaf was calculated by adding supernatant from extract of plant material then mixed with 3 mL of colour reagent. After 5 min , the absorbance of each sample was measured at .
Statistical analysis
Using the SAS (Statistical Analysis System) statistical software programme, the data was statistically analyzed using procedure mixed (PROC MIXED) and procedure generalized linear model (PROC GLM) (SAS Institute). The Duncan Multiple range test was used to detect mean differences. Correlation and principal component analyses were performed using RStudio’.
Results
Effects of fulvic acid on growth parameters of maize cultivars
The study investigated the effects of different treatments on the growth parameters of two maize cultivars, P3939 and 30Y87. Significant variations were observed in leaf length, leaf width, leaf area, plant height, and number of leaves across treatments (Table 1). For both cultivars, the control group exhibited intermediate values for most parameters. The application of notably reduced leaf length, leaf width, leaf area, and plant height compared to the control, indicating a negative impact on growth. Conversely, treatments combining with fulvic acid applied as a foliar spray (FFA, ) or through the rooting medium (IFA, ) showed varying effects. The application of generally mitigated some of the adverse effects associated with chromium exposure, whereas yielded mixed outcomes, occasionally intensifying the
Treatments
Cultivar P3939
Cultivar 30Y87
Leaf length (cm)
Leaf width (cm)
Leaf area ( )
Plant height (cm)
No. of leaves
Leaf length (cm)
Leaf width (cm)
Leaf area ( )
Plant height (cm)
No. of leaves
Control ( )
a-b
ab
ab
FFA
a
a
a
a
a
IFA
ab
a
ab
a-b
a-b
a-b
a-b
e
c-d
c-d
Cr + FFA
b-c
b-c
b-c
b-c
Cr + IFA
c-d
c-d
b-c
b-c
b-c
b-c
a-c
b-c
d-e
b-e
b-c
d-e
d-e
d-e
Cr + FFA
c-d
c-d
b-c
b-c
c-d
c
c-d
Cr + IFA
d-e
c-d
c-d
Table 1. Effects of fulvic acid applied as a foliar spray (FFA, ) and through the rooting medium irrigation (IFA, ) on growth parameters of maize cultivars (P3939 and 30Y87). Values represent mean ± standard error (SE), with lowercase letters indicating significant differences among treatments.
harmful impacts of chromium. Overall, these findings highlight the complex interactions between chromium exposure and fulvic acid application methods on the growth metrics of these maize cultivars.
Effects of fulvic acid on chlorophyll content maize cultivars
In both maize cultivar chlorophyll concentration was measured across varying Cr concentrations and foliar application treatments. In cultivar P3939, under normal conditions ( ), chlorophyll levels were relatively consistent across treatments: FW without FFA, FW with FFA, and FW with IFA. However, exposure to Cr resulted in a decrease in chlorophyll concentration across all treatments: FW without FFA, FW with FFA, and FW with IFA. At , chlorophyll concentrations varied further: FW without FFA, FW with FFA, and FW with IFA (Fig. 2A). Under control conditions ( ), In cultivar 30Y87 chlorophyll levels varied with foliar treatments: FW without FFA, FW with FFA, and FW with IFA. Exposure to resulted in a decline in chlorophyll concentration across all treatments: FW without FFA, FW with FFA, and FW with IFA. At Cr, chlorophyll concentrations continued to decrease: FW without FFA, FW with FFA, and FW with IFA. These findings indicate that while foliar application of fulvic acid (FFA) generally maintained or slightly elevated chlorophyll levels compared to controls, chromium exposure consistently diminished chlorophyll production (Fig. 2B).
Effects of fulvic acid on oxidative stress (electrolyte leakage and MDA) content maize cultivars
Electrolyte leakage (%)
In maize cultivar P3939, the impact of Cr concentrations on electrolyte leakage percentage was examined across different foliar application treatments. Under normal conditions ( ), electrolyte leakage percentages were without FFA (fulvic acid), with FFA, and with IFA (fulvic acid through the rooting medium). As chromium concentration increased to , electrolyte leakage decreased to without with FFA, and with IFA. At the highest Cr level tested , electrolyte leakage showed mixed trends: without FFA, with FFA, and with IFA (Fig. 3A).
In maize cultivar 30Y87, electrolyte leakage percentages were examined across varying Cr concentrations and foliar application treatments. At , electrolyte leakage increased compared to the control (
Fig. 2. Effects of fulvic acid applied as a foliar spray (FFA, ) and through the rooting medium irrigation (IFA, 300 L/A) on Chlorophyll content of maize cultivars P3939 (A) and 30 Y87 (B) under chromium stress. Values represent mean ± standard error (SE), with lowercase letters indicating significant differences among treatments. -FFA indicates no fulvic acid application (control). + FFA represents foliar application of fulvic acid, sprayed on leaves. IFA refers to fulvic acid application through irrigation, delivered via the rooting medium.
Fig. 3. Effects of fulvic acid applied as a foliar spray (FFA, ) and through the rooting medium irrigation (IFA, 300 L/A) on Electrolyte leakage (EL) of maize cultivars P3939 (A) and 30Y87 (B) under chromium stress. Values represent mean ± standard error (SE), with lowercase letters indicating significant differences among treatments. -FFA indicates no fulvic acid application (control). + FFA represents foliar application of fulvic acid, sprayed on leaves. IFA refers to fulvic acid application through irrigation, delivered via the rooting medium.
Cr) across all treatments: FFA ( – ) increased by , FFA ( + ) by , and IFA ( + ) by . However, at , the responses varied: FFA ( – ) showed a decrease in leakage by , FFA ( + ) exhibited an increase by , and IFA ( + ) showed a modest increase by (Fig. 3B).
MDA content (nm/g FW)
Maize cultivar P3939 demonstrates distinct changes in MDA (Malondialdehyde) content (nM/g FW) in response to varying concentrations of Cr and fulvic acid (FA). At , the MDA content for the cultivar starts at under FA(-) conditions. As the Cr concentration increases to , MDA levels show a slight rise to FW. However, a more notable increase occurs at , where MDA content significantly jumps to . These observations indicate that maize cultivar P3939 exhibits a concentration-dependent response to Cr (Fig. 4A). Maize cultivar 30 Y 87 displays distinct changes in MDA (Malondialdehyde) content (nM/g FW) in response to varying concentrations of chromium (Cr) and different foliar treatments with fulvic acid (FA). Under control conditions ( ), the MDA content is recorded at . When exposed to Cr without any foliar treatment (FA-), the MDA content increases to FW. Interestingly, the addition of foliar fulvic acid (FFA+) at the same Cr concentration ( ) results in a slight decrease in MDA to FW, suggesting a potential mitigating effect of FFA against Cr-induced oxidative stress. Further analysis at with irrigated fulvic acid (IFA+) shows a rise in MDA content to FW, indicating a moderate increase compared to the control (Fig. 4B).
Effects of fulvic acid on total soluble protein (TSP) content (mg/g FW)
Maize cultivar P3939 displays varying levels of total soluble protein (TSP) ( ) in response to different concentrations of Cr and foliar treatments with fulvic acid (FA). Under normal conditions without , TSP levels are for FA ( – ), for FFA ( + ), and for IFA ( + ). As the Cr concentration increases to , there is a noticeable decrease in TSP across all foliar treatments: for FA (-), for FFA ( +), and for IFA ( +). This reduction suggests that chromium stress may negatively impact protein synthesis or stability in maize cultivar P3939. At the highest Cr concentration tested ( ), TSP levels continue to decrease: for FA( – ), for FFA ( + ), and for IFA ( + ) (Fig. 5A). Under normal conditions without , the TSP levels are relatively stable across all foliar treatments: for FA( – for FFA , and for IFA . However, as the Cr concentration increases to and , TSP levels show slight fluctuations among the treatments. At Cr, TSP levels increase to for FA (-), for FFA (+), and for IFA ( +). This suggests a potential response of the cultivar to moderate stress induced by chromium, possibly
Fig. 4. Effects of fulvic acid applied as a foliar spray (FFA, ) and through the rooting medium irrigation (IFA, 300 L/A) on MDA content of maize cultivars P3939 (A) and 30Y87 (B) under chromium stress. Values represent mean ± standard error (SE), with lowercase letters indicating significant differences among treatments. -FFA indicates no fulvic acid application (control). + FFA represents foliar application of fulvic acid, sprayed on leaves. IFA refers to fulvic acid application through irrigation, delivered via the rooting medium.
influencing protein metabolism or synthesis. At , TSP levels decrease slightly to for FA(-), for FFA ( + ), and for IFA ( + ), indicating a more pronounced effect of higher chromium concentrations on protein levels in the maize plants (Fig. 5B).
Effects of fulvic acid on antioxidant enzymatic activity under chromium stress
In the results, it was found that fulvic acid played a significant role in modulating antioxidant enzymatic activity in plants under chromium stress conditions. Specifically, enzymes such as catalase (CAT), ascorbate peroxidase (APX), and glutathione peroxidase (GPX) were observed to be crucial components of the antioxidant defense system that helped mitigate oxidative damage caused by chromium toxicity.
Catalase activity (units protein)
Maize cultivar P3939 exhibits varying catalase (CAT) activity ( protein) in response to different concentrations of Cr and foliar treatments with fulvic acid (FA). Under control conditions without , CAT activity levels are measured at protein for FA( – protein for FFA ( + ), and protein for IFA . As the Cr concentration increases to and , changes in CAT activity are observed across all foliar treatments. At , CAT activity decreases to protein for FA (-), protein for FFA (+), and protein for IFA (+). This reduction suggests a potential suppression of CAT enzyme function under moderate chromium stress conditions. At , CAT activity shows further variations: protein for FA protein for FFA ( + ), and protein for IFA ( + ) (Fig. 6A). Maize cultivar 30 Y 87 exhibits varying levels of catalase (CAT) activity ( protein) in response to different concentrations of chromium ( Cr ) and foliar treatments with fulvic acid (FA). Under normal conditions without , CAT activity levels are recorded at protein for FA( – protein for FFA , and protein for IFA ( + ). As the Cr concentration increases to , changes in CAT activity are observed across the foliar treatments. CAT activity slightly increases to protein for FA (-), remains stable at 0.010112 protein for FFA (+), and decreases to protein for IFA (+). At , further variations in CAT activity are noted: protein for FA protein for FFA , and protein for IFA ( + ) (Fig. 6B).
APX activity (units protein)
Maize cultivar P3939 demonstrates varying ascorbate peroxidase (APX) activity ( protein) in response to different concentrations of Cr and foliar treatments with fulvic acid (FA). Under normal conditions without Cr , APX activity levels are measured at protein for FA(-), protein for FFA , and protein for IFA ( + ). As the Cr concentration increases to , APX activity shows fluctuations across the foliar treatments. APX activity increases to protein for FA( – ), remains
Fig. 5. Effects of fulvic acid applied as a foliar spray (FFA, ) and through the rooting medium irrigation (IFA, ) on Total Soluble protein (TSP) of maize cultivars P3939 (A) and 30Y87 (B) under chromium stress. Values represent mean ± standard error (SE), with lowercase letters indicating significant differences among treatments. -FFA indicates no fulvic acid application (control). + FFA represents foliar application of fulvic acid, sprayed on leaves. IFA refers to fulvic acid application through irrigation, delivered via the rooting medium.
relatively stable at protein for FFA ( + ), and slightly increases to protein for IFA . At , further variations in APX activity are observed: protein for FA(-), 0.011373 protein for FFA(+), and protein for IFA (+) (Fig. 7A). Maize cultivar 30 Y 87 displays varying levels of ascorbate peroxidase (APX) activity ( protein) in response to different concentrations of chromium ( Cr ) and foliar treatments with fulvic acid (FA). Under control conditions without , APX activity levels are relatively consistent: protein for FA( – protein for FFA , and protein for IFA ( + ). As the Cr concentration increases to , APX activity shows minor fluctuations across the foliar treatments. APX activity decreases slightly to protein for FA(-), decreases to protein for FFA ( + ), and increases marginally to protein for IFA( + ).At , further variations in APX activity are observed: protein for FA(-), protein for FFA( + ), and protein for IFA ( + ) (Fig. 7B).
GPX activity (units protein)
Maize cultivar P3939 is evaluated for its response to varying concentrations of Cr , focusing on the enzymatic activity of glutathione peroxidase (GPX). The study examines GPX activity in terms of micromoles ( ) of Cr at concentrations of 0,100 , and . Results indicate that at increasing Cr concentrations, GPX activity in P3939 maize cultivar remains within a consistent range, suggesting a robust antioxidative response. The levels of FA (-), FFA (+), and IFA (+) in the cultivar show differential responses to Cr exposure, with fluctuations observed across the treatments. Specifically, FA (-) exhibits a slight increase from 0.005377 to protein as Cr levels rise from 0 to , while and IFA( + ) show more varied responses (Fig. 8A). Maize cultivar 30 Y87 was investigated for its response to increasing concentrations of chromium (Cr), focusing on the activity of glutathione peroxidase (GPX) and the levels of FA (-), FFA (+), and IFA (+) in its tissues. The study measured GPX activity in units per milligram of protein across Cr concentrations of 0,100 , and . Results indicate varying responses in GPX activity as Cr levels increase, suggesting a nuanced antioxidative capacity in cultivar 30Y87. Specifically, GPX activity fluctuates, with a noticeable decrease from 0.007192 to protein as Cr concentration rises from 0 to . This variability underscores the cultivar’s adaptability to oxidative stress induced by chromium. In terms of FA ( – ), FFA ( + ), and IFA ( + ) levels, cultivar 30Y87 displays differential responses across the Cr treatments. FA (-) levels notably increase from 0.005377 protein at to protein at , while FFA ( + ) and IFA ( + ) show more complex patterns of change (Fig. 8B).
Fig. 6. Effects of fulvic acid applied as a foliar spray (FFA, ) and through the rooting medium irrigation (IFA, 300 L/A) on Catalase enzyme (CAT) of maize cultivars P3939 (A) and 30 Y87 (B) under chromium stress. Values represent mean ± standard error (SE), with lowercase letters indicating significant differences among treatments. -FFA indicates no fulvic acid application (control). + FFA represents foliar application of fulvic acid, sprayed on leaves. IFA refers to fulvic acid application through irrigation, delivered via the rooting medium.
Principal component analysis (PCA) and correlation analysis
This study employed Principal Component Analysis (PCA) and correlation analysis to assess the biochemical and morphological parameters of maize cultivars under varying levels of Cr stress and application of fulvic acid (FFA) and/or inorganic fulvic acid (IFA) through foliar and irrigated methods (Fig. 9). All parameters were standardized to ensure variables were comparable by scaling to zero mean and unit variance. PC scores were analyzed to identify patterns and relationships among parameters across different experimental treatments. This helped in understanding how Cr stress and fulvic acid applications influenced the biochemical and morphological characteristics of maize cultivars. Pearson correlation coefficients were calculated to determine the strength and direction of linear relationships between pairs of parameters within each experimental group (Fig. 10).
Discussion
In this study, we investigated the role of different modes of fulvic acid application on the antioxidant activity of maize (Zea mays L.) subjected to artificial chromium stress. Chromium contamination in agricultural soils is a significant environmental concern, posing detrimental effects on plant growth and productivity . Fulvic acid, known for its chelating properties and ability to enhance plant stress tolerance, was applied through various methods to assess its efficacy in mitigating chromium-induced oxidative stress in maize plants . The antioxidant activity of maize plants serves as a critical indicator of their ability to counteract reactive oxygen species (ROS) accumulation under chromium stress conditions . By examining the effects of different fulvic acid application modes, this study aims to provide insights into optimizing agricultural practices for sustainable crop production in chromium-contaminated soils. A non-significant increase in growth parameters was observed in metal treated plants and Cr treated plants with FA application. However, these non-significant findings may be attributed to the inherent variability among treatments or the stage of plant development at the time of assessment. Our results indicated a reduction in growth parameters in chromium treated plants. These findings are in consistent with work of Arshed et al. who observed the reduction in growth parameters in Brassica juncea under chromium stress. Alshegaihi et al. also reported reduction in growth parameters in cooper metal treated plants of wheat. These reductions in growth parameters can be attributed to ultra-structural and physio-chemical alterations in plants exposed to Cr stress . This decrease in growth metrics, such as shoot and root length and biomass, is linked to diminished nutrient uptake . Our results indicated a reduction in leaf area under chromium stress. This reduction in leaf area could be linked to restricted water availability impacting leaf expansion, similar to observations under potassium deficiency . Fluoride toxicity in wheat has similarly been shown to reduce leaf area .
In our experiment, foliar fulvic acid application showed an increase in various growth parameters such as leaf length, width, area, and number of leaves. Our study is consistent with previous studies that demonstrated
Fig. 7. Effects of fulvic acid applied as a foliar spray (FFA, ) and through the rooting medium irrigation (IFA, 300 L/A) on Ascorbate enzyme (APX) of maize cultivars P3939 (A) and 30Y87 (B) under chromium stress. Values represent mean ± standard error (SE), with lowercase letters indicating significant differences among treatments. -FFA indicates no fulvic acid application (control). + FFA represents foliar application of fulvic acid, sprayed on leaves. IFA refers to fulvic acid application through irrigation, delivered via the rooting medium.
the effects of fulvic acid application on plant growth and nutrient content. For instance, one study focused on the effect of foliar application of fulvic acid on plant growth and fruit quality of tomato (Lycopersicon esculentum L.). Another study investigated the effects of soil application of fulvic acid on the growth and nutrient content of cucumber (Cucumis sativus) plants . The capacity of FA to penetrate plant roots and stimulate the expression of genes involved in cell division supports its role in promoting vegetative growth .
Chlorophyll content was reduced in plants of both maize cultivars due to chromium stress. This reduction aligns with findings in Triticum aestivum under antibiotic stress . Chromium stress results in the production of reactive oxygen species that reacts with pigment-protein complexes and damages the thylakoid membranes of chlorophyll due to the substitution of magnesium with ions . This biochemical disruption underscores the vulnerability of photosynthetic machinery under heavy metal stress. On the other hand, foliar application of fulvic acid showed an increase in chlorophyll content. For example, increase in chlorophyll content due to foliar application of fulvic acid was observed at in plants of maize cultivar P3939. Moreover, foliar application of fulvic acid caused a increase in chlorophyll content in maize plants grown under normal conditions. These results are similar to some earlier studies, in which an increase in chlorophyll content by the fulvic acid application has observed in maize plants . This increase suggests that FA mitigates oxidative damage by reducing free Cr ion concentration through adsorption .
Electrolyte leakage is an indicator of cellular damage induced by heavy metal stress and other abiotic stresses. Addition of chromium ( ) to the growth medium increased the electrolyte leakage in the leaves of both maize cultivars. The rise in leakage is consistent with overproduction of ROS under metal stress, contributing to oxidative damage . However, fulvic acid application (foliar + irrigated) showed reduced electrolyte leakage in chromium treated plants. About decrease in electrolyte leakage was observed with IFA application at as compared with treatment alone. Malondialdehyde content is a reliable indicator of oxidative stress induced by abiotic stresses, heavy metal stress and pathogen attack . A significant increase in MDA content ( ) was observed at concentrations in both varieties. This aligns with findings in sunflower and turnip, where Cr-induced oxidative damage elevated MDA levels where Cr-induced ROS reacted with lipids, causing membrane disruption and cellular damage . Fulvic acid application showed reduced MDA content in chromium treated plants of both cultivars. However, the application of FA through the soil was found more effective in alleviating oxidative stress by free radical detoxification.
Our results indicate relatively reduced total soluble protein content at and treatments as compared to control. Previous study also reported a decrease in total soluble protein content under Cr stress . This decline in TSP may also stem from increased Cr uptake interfering with protein stability . On the other
Fig. 8. Effects of fulvic acid applied as a foliar spray (FFA, ) and through the rooting medium irrigation (IFA, ) on glutathione peroxidase (GPX) of maize cultivars P3939 (A) and 30Y87 (B) under chromium stress. Values represent mean ± standard error (SE), with lowercase letters indicating significant differences among treatments. -FFA indicates no fulvic acid application (control). + FFA represents foliar application of fulvic acid, sprayed on leaves. IFA refers to fulvic acid application through irrigation, delivered via the rooting medium.
hand, application of fulvic acid through rooting medium enhanced the total soluble protein content in metal treated plants. These results are in consistent with work of Askari et al. who also recorded an increase in TSP content in Cr treated wheat plants. Increase in total soluble protein might be due to positive effects of fulvic acid on plant growth and photosynthetic pigments. This improvement underscores the role of FA in supporting protein synthesis and stability under stress. In our study investigating the role of different modes of fulvic acid application on the antioxidant activity of maize under artificial chromium stress, a significant increase in key antioxidant enzymes and compounds was observed. Specifically, levels of catalase (CAT), ascorbate peroxidase (APX), peroxidase (POD), and glutathione reductase (GPX) showed marked enhancement. These enzymes play critical roles in mitigating oxidative stress by decomposing ROS, preventing cellular damage, and maintaining cellular homeostasis . Catalase is an important component of antioxidant defense system that is involved in breakdown of hydrogen peroxide generated during photorespiration and fatty acids oxidation . Our results indicated no significant increase in CAT activity in chromium treated plants of both cultivars. An increase in catalase activity was recorded at a higher concentration of Cr i.e. as compared with control and Cr. Jabeen et al. also recorded increase in CAT activity in bean plants under Cr stress. The decrease in CAT activity at might be due to reason that catalase is iron porphyrin molecule and Cr may react with iron affecting the availability of the active form of iron . Fulvic acid (foliar) application showed an increase in catalase activity at concentration. About increase in CAT activity was observed with foliar FA at as compared with alone treatment. Peroxidase that breakdown by using ascorbate as an electron donor is known as ascorbate peroxidase. APX is an essential antioxidant enzyme present in peroxisomes, cytosol and chloroplasts . Our results showed reduced APX activity at a higher concentration of . The decrease in APX was also reported in Indian mustard , oil seed rape and Zea mays . The reason for the decline in APX activity might be that higher Cr toxicity can inhibit the antioxidant defence system responsible for detoxification of ROS. However, foliar application of fulvic acid increased APX activity in chromium stressed plants of cultivar 30 Y 87 . These findings are in consistent with work of Wang et al. also recorded an increase in APX activity with fulvic acid application in metal treated plants. About increase in APX activity was observed with foliar FA application at treatment. GPX is an antioxidant enzyme that breaks down by using guaiacol as an electron donor . GPX plays vital role in lignifications; defense and wound healing . The activity of GPX did not change due to chromium stress or exogenous application of fulvic acid. An increase in antioxidant enzyme activities (CAT and GPX) under Cr stress is might in response to generation of superoxide radicals by Cr induced blockage of electron transport chain in mitochondria. The decrease in antioxidant enzyme activity (APX) might be due to the inhibitory effects of Cr ions on the enzyme . In our experiment, FA application improved the activity of antioxidant enzymes (CAT and APX)
Fig. 9. Score (a, c) and loading plots (b, d) of principal component analysis (PCA) on various studied parameters of maize cultivars grown in Cr stressed soil. Score plot represents separation of treatments as (1) Control (without Cr contamination); (2) FFA (3) IFA (4) Cr level, 5) , 7) , 8) 9) respectively. The abbreviations of parameters are as follows: Plant H: plant height; NL: number of leaves; LW: leaf width; T chl: total chlorophyll; EL: electrolyte leakage; MDA: Lipid peroxidation; T pro: total protein; CAT: catalase; APX: Ascorbate peroxidase GPX: Guaiacol peroxidases.
under chromium stress. It might be due to reason that FA acts as a free radical scavenger and antioxidant. FA has the ability to react with both positive and negatively charges unpaired electrons and hence detoxify the free radicals might be due to enhanced uptake of nutrients .
Conclusion
Chromium toxicity adversely affects antioxidant enzyme activities, photosynthetic pigments, and overall plant growth. Fulvic acid application (irrigated) led to an increase in chlorophyll content and total soluble proteins, although these results were statistically non-significant. Additionally, fulvic acid reduced malondialdehyde (MDA) content and electrolyte leakage in chromium-treated plants. Foliar fulvic acid application significantly enhanced the activities of antioxidant enzymes, particularly catalase (CAT) and ascorbate peroxidase (APX), while it did not affect the activity of guaiacol peroxidase in metal-treated plants. The increased activities of CAT and APX suggest that foliar fulvic acid application may improve plant resilience against chromium stress by effectively scavenging reactive oxygen species (ROS). The potential mechanism of fulvic acid’s action in this context involves enhancing antioxidant enzyme activities, which helps mitigate oxidative stress caused by chromium exposure. Although chromium stress resulted in reduced growth parameters, foliar and root irrigated fulvic acid application had a positive impact on various growth measures. We recommend fulvic acid application
Fig. 10. Correlation between growth and physiological parameters of maize cultivars grown in Cr stressed soil.
as a more effective approach for mitigating chromium toxicity, given its significant impact on the antioxidant defense system. Future research should explore the long-term effects of foliar versus irrigated fulvic acid, as well as its potential to alleviate other types of abiotic stress.
Data availability
All data generated or analyzed during this study are included in this published article.
Received: 25 September 2024; Accepted: 27 December 2024
Published online: 08 January 2025
References
Fatima, A., Shabaan, M., Ali, Q., Malik, M., Asghar, H.N., Aslam, M., Zulfiqar, U., Hameed, A., Nazim, M., Mustafa, A.E.Z.M. and Elshikh, M.S. Integrated application of metal tolerant . fluorescens and press mud for conferring heavy metal tolerance to aloe vera (Aloe barbadensis). Plant Stress, 11, 100333 (2024).
Sarwar, M.J., Shabaan, M., Asghar, H.N., Ayyub, M., Ali, Q., Zulfiqar, U., Nazim, M., Alarjani, K.M. and Elshikh, M.S. Interaction of chromium (Cr) resistant plant growth promoting rhizobacteria with compost to phytostabilize Cr in spinach rhizosphere. Plant Stress, 10, 100261 (2023).
Ali, H. Q. et al. Tanneries impact on groundwater quality: A case study of Kasur city in Pakistan. Environ. Monit. Assess. 194(11), 823 (2022).
Mehdiniya Afra, J. et al. Influence of chemical fertilizers and animal manure on morphological traits of medicinal plants in Northern Iran. Res. Crop Ecophysiol. 17(2), 86-101 (2022).
Laxmi, V. & Kaushik, G. Toxicity of hexavalent chromium in environment, health threats, and its bioremediation and detoxification from tannery wastewater for environmental safety. In Bioremediation of industrial waste for environmental safety: volume I: industrial waste and its management (eds Saxena, G. & Bharagava, R. N.) 223-243 (Springer Singapore, 2020).
Zulfiqar, U., Haider, F.U., Ahmad, M., Hussain, S., Maqsood, M.F., Ishfaq, M., Shahzad, B., Waqas, M.M., Ali, B., Tayyab, M.N. and Ahmad, S.A., 2023. Chromium toxicity, speciation, and remediation strategies in soil-plant interface: A critical review. Front Plant Sci. 13, 1081624 (2023).
Arshed, M. Z., Zulfiqar, A. & Saleem, A. Alleviation of chromium stress in Brassica juncea L. and soil remediation by plant growth promoting (PGP) bacteria. Pak. J. Sci. Ind. Res. Ser. Biol. Sci. 67(1), 42-48 (2024).
Sheteiwy, M. S. et al. Elevated differentially attenuates beryllium-induced oxidative stress in oat and alfalfa. Physiol. Plant. 175(5), e14036 (2023).
Hussnain, M., Shabaan, M., Ali, Q., Ashraf, S., Ahmad, M., Ghafoor, U., Akhtar, M.J., Zulfiqar, U., Hussain, S., Al-Ashkar, I. and Elshikh, M.S. Microbial phytoremediation of chromium-contaminated soil with biogas slurry for enhancing the performance of Vigna radiata L. Plant Stress, 10, 100206 (2023).
Younis, U., Danish, S., Datta, R., Alahmadi, T. A. & Ansari, M. J. Sustainable remediation of chromium-contaminated soils: Boosting radish growth with deashed biochar and strigolactone. BMC Plant Biol. 24(1), 115 (2024).
Nardi, S., Schiavon, M. & Francioso, O. Chemical structure and biological activity of humic substances define their role as plant growth promoters. Molecules 26(8), 2256 (2021).
Gong, G. et al. Extraction of fulvic acid from lignite and characterization of its functional groups. ACS Omega 5(43), 27953-27961 (2020).
Shahrajabian, M. H. & Sun, W. The importance of salicylic acid, humic acid and fulvic acid on crop production. Lett. Drug Des. Discov. 21(9), 1465-1480 (2024).
Dawood, M. F., Tahjib-Ul-Arif, M., Sohag, A. A. M. & Abdel Latef, A. A. H. Role of acetic acid and nitric oxide against salinity and lithium stress in Canola (Brassica napus L.). Plants 13(1), 51 (2023).
Jabeen, N. et al. Glycinebetaine mediates chromium tolerance in mung bean through lowering of Cr uptake and improved antioxidant system. Arch. Agron. Soil Sci. 62(5), 648-662 (2016).
Abu-Omar, M. M. High-valent iron and manganese complexes of corrole and porphyrin in atom transfer and dioxygen evolving catalysis. Dalton Trans. 40(14), 3435-3444 (2011).
Sousa, R. H. et al. Impairment of peroxisomal APX and CAT activities increases protection of photosynthesis under oxidative stress. J. Exp. Bot. 70(2), 627-639 (2019).
Nouairi, I. et al. Antioxidant defense system in leaves of Indian mustard (Brassica juncea) and rape (Brassica napus) under cadmium stress. Acta Physiol. Plant. 31, 237-247 (2009).
Wu, Z. et al. Antioxidant enzyme systems and the ascorbate-glutathione cycle as contributing factors to cadmium accumulation and tolerance in two oilseed rape cultivars (Brassica napus L.) under moderate cadmium stress. Chemosphere 138, 526-536 (2015).
Chugh, V., Kaur, N., Gupta, A. K. Evaluation of oxidative stress tolerance in maize (Zea mays L.) seedlings in response to drought (2011).
Dawood, M. F., Tahjib-Ul-Arif, M., Sohag, A. A. M. & Abdel Latef, A. A. H. Fluoride mitigates aluminum-toxicity in barley: Morpho-physiological responses and biochemical mechanisms. BMC Plant Biol. 22(1), 287 (2022).
Bala, S., Asthir, B. & Bains, N. Activities of cell wall peroxidases in relation to lignification in six wheat (Triticum aestivum L.) genotypes under heat and drought stress. J. Environ. Biol. 37(3), 437 (2016).
Yilmaz, S. H., Kaplan, M., Temizgul, R. & Yilmaz, S. Antioxidant enzyme response of sorghum plant upon exposure to aluminum, chromium and lead heavy metals. Turk. J. Biochem. 42(4), 503-512 (2017).
Wang, W., Chen, M., Wang, D., Yan, M. & Liu, Z. Different activation methods in sulfate radical-based oxidation for organic pollutants degradation: Catalytic mechanism and toxicity assessment of degradation intermediates. Sci. Total Environ. 772, 145522 (2021).
Acknowledgements
The study was supported by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia (grants no. 451-03-66/2024-03/200032). The authors extend their appreciation to Researchers Supporting Project number (RSP2025R390), King Saud University, Riyadh, Saudi Arabia.
Author contributions
Conceptualization, F.I., A.Z., and A.K.; methodology, F.I.; software, A.S., and M.Z.A.; validation and formal analysis, A.S., and F.I.; resources, I.D., and P.V.P.; data curation, U.Z.; writing-original draft preparation, F.I., A.Z., A.S., U.Z., and M.Z.A,; writing-review and editing, I.D., P.V.P., and W.S.; supervision, A.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia (grants no. 451-03-66/2024-03/200032). The authors extend their appreciation to Researchers Supporting Project number (RSP2025R390), King Saud University, Riyadh, Saudi Arabia.
Declarations
Ethics approval and consent to participate
All methods were performed in accordance with the relevant guidelines and regulations. We have obtained permission to collect plant material and seedlings.
Competing interests
The authors declare no competing interests.
Additional information
Correspondence and requests for materials should be addressed to U.Z. or I.D.
Reprints and permissions information is available at www.nature.com/reprints.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
Institute of Botany, University of the Punjab, Lahore, Pakistan. Department of Agronomy, Faculty of Agriculture and Environment, The Islamia University of Bahawalpur, Bahawalpur 63100, Pakistan. Institute of Field and Vegetable Crops, National Institute of the Republic of Serbia, Maxim Gorki 30, 21000 Novi Sad, Serbia. Department of Agronomy, Kansas State University, 108 Waters Hall, 1603 Old Claflin Place, Manhattan, KS 66506, USA. Plant Production Department, College of Food and Agriculture Sciences, King Saud University, 11451 Riyadh, Saudi Arabia. email: usman.zulfiqar@iub.edu.pk; ivica.djalovic@ifvcns.ns.ac.rs