الاستخدام الانتقائي للبوليسكاريدات الطبية بواسطة أنواع بكتيرويدس وبارابكتيرويدس في أمعاء الإنسان Selective utilization of medicinal polysaccharides by human gut Bacteroides and Parabacteroides species
تلعب أنواع بكتيريا بكتيرويدس وبارابكتيرويدس في أمعاء الإنسان أدوارًا حاسمة في صحة الإنسان وتشتهر بقدرتها على استخدام مجموعة متنوعة من البوليسكاريدات. إن فهم كيفية استخدام هذه البكتيريا للبوليسكاريدات الطبية يعد أساسًا لتطوير بروبيوتيك وأدوية قائمة على البوليسكاريدات. هنا، قمنا برسم خرائط منهجية لملفات استخدام 20 نوعًا مختلفًا من البوليسكاريدات الطبية بواسطة 28 نوعًا من بكتيرويدس وبارابكتيرويدس في أمعاء الإنسان. أظهرت ملفات النمو تباينًا كبيرًا عبر الأنواع البكتيرية المختلفة والبوليسكاريدات الطبية. عززت البوليسكاريدات المستخرجة من الجينسنغ نمو عدة أنواع من بكتيرويدس وبارابكتيرويدس؛ في المقابل، عززت البوليسكاريدات المستخرجة من دندروبيوم بشكل انتقائي نمو بكتيرويدس يونيكوميس. كان هذا الملف المميز للاستخدام مرتبطًا بالتنوع الجينومي في الإنزيمات النشطة للكربوهيدرات، بدلاً من تباين تركيب السكريات الأحادية بين البوليسكاريدات الطبية. من خلال التحليل المقارن للتعبير الجيني والتلاعب الجيني، تحققنا من أن موضع استخدام البوليسكاريد PUL34_Bu مكن بكتيرويدس يونيكوميس من استخدام البوليسكاريدات المستخرجة من دندروبيوم (أي الجلوكومانان). بالإضافة إلى ذلك، وجدنا أن إنزيم GH26 في PUL34_Bu سمح لبكتيرويدس يونيكوميس باستخدام مانان مستخرج من النباتات. بشكل عام، كشفت نتائجنا عن الاستخدام الانتقائي للبوليسكاريد الطبي بواسطة أنواع بكتيرويدس وبارابكتيرويدس وقدمت رؤى حول استخدام البوليسكاريدات في هندسة ميكروبيوم أمعاء الإنسان.
تلعب بكتيريا الأمعاء البشرية من نوع باكتيرويدس وباراباكتيرويدس، المستهلكين الرئيسيين للبوليسكاريدات، أدوارًا حيوية في صحة الإنسان والمرض، مثل التطور والعلاج المناعي مقاومة السرطان للاستعمار ضد مسببات الأمراض، إلخ. إنهم يستخدمون أوليغو/بوليسكاريدز ذات المنشأ الداخلي (المستمدة من المضيف) والخارجي (مثل المستمدة من النباتات)، مما يوفر التغذية والفيتامينات للمضيف وميكروبات الأمعاء الأخرى.تُبرز الدراسات الحديثة أن البوليسكاريدات تؤثر على صحة المضيف من خلال تنظيم نمو وملفات الأيض لبكتيرويدس..
فهم كيفية استخدام البكتيرويدس والبارابكتيرويدس لهذه البوليسكاريدات أمر ضروري لتطوير بروبيوتيك وأدوية جديدة قائمة على البوليسكاريدات لتعزيز الصحة.
تحتوي البوليسكاريدات الطبية المستخلصة من الأعشاب والفطر على هياكل معقدة ومتنوعة، تتكون من روابط جليكوسيدية مختلفة (مثل، – أو -روابط جليكوسيدية) في السلاسل الرئيسية أو المتفرعة وتشمل عشرة أو أكثر من السكريات الأحادية، أحيانًا مع تعديلات مثل الميثيلاation و أسيتيلتظهر الأبحاث أن البولي سكريات الطبية وغيرها من البولي سكريات المستمدة من النباتات (مثل تلك الموجودة في الخضروات والفواكه) تمتلك أنشطة بيولوجية متنوعة.بما في ذلك التأثيرات المضادة للأورام، المضادة للفيروسات، المضادة للالتهابات، وتأثيرات تعديل المناعة. يمكن تعزيز أنواع البكتيرويدز والبارابكتيرويدز بواسطة العديد من البوليسكاريدات الطبية. لقد أظهرت بوليسكاريدات الجينسنغ (GPs) أنها تعزز كل من بكتيرويدز فulgatus وبارابكتيرويدز ديستاسونيس، مما يزيد من معدل الاستجابة لعلاج المناعة PD-1/PD-L1.تم العثور على أن البوليسكاريدات من Ophiocordyceps sinensis تعزز من نمو البكتيريا المعوية المفيدة Parabacteroides goldsteinii، مما يؤدي إلى تحسينات في السمنة والاضطرابات الأيضية.يسترجع بوليسكاريد دندروبيوم أوفيكينا التنوع في ميكروبيوتا الأمعاء، ويزيد من وفرة بكتيرويدس، مما له تأثير وقائي ضد التهاب القولون الناتج عن سلفات الدكستران في الفئران..
على الرغم من التقدم في استخدام البوليسكاريدات من قبل البكتيرويدات، إلا أنه ليس من الواضح كيف تستخدم البكتيرويدات البوليسكاريدات الطبية. تمتلك البكتيرويدات والبارابكتيرويدات المئات من الإنزيمات النشطة على الكربوهيدرات (CAZymes) المسؤولة عن تحلل البوليسكاريدات.عادةً ما تكون إنزيمات CAZymes منظمة في مجموعات تعرف بمواقع استخدام البوليسكاريد (PULs)، والتي يتم تنظيمها بشكل إيجابي بواسطة بوليسكاريدات محددة.الهيولاز الجليكوزيدية (GH) هي إنزيمات مسؤولة عن التحلل المائي للروابط الجليكوزيدية. عادةً ما تحتوي جينومات أنواع البكتيرويد على عدة وحدات استخدام البوليسكاريد (PULs) لاستغلال بوليسكاريد مختلفة. تم توضيح بعض الآليات، على سبيل المثال، يستخدم B. thetaiotaomicron VPI-5482 النشا باستخدام نظام Sus.; تقوم أنواع البكتيرويد بتفكيك السكريات المعقدة O الموجودة في المخاط باستخدام السلفاتاز; B. plebeius يستخدم بوليسكاريد الكبريتات البورفيران باستخدام إنزيمات البورفيران، إلخ.
في هذه الدراسة، قمنا بتوصيف أنماط نمو 28 نوعًا من بكتيرويدس وبارابكتيرويدس في أمعاء الإنسان باستخدام 20 بوليسكاريد طبي. لاحظنا اختلافات كبيرة في ملفات النمو بين الأنواع والبوليسكاريدات. ومن الجدير بالذكر أن بوليسكاريدات دندروبيوم (DPs، نوع من الجلوكومانان) حفزت بشكل خاص نمو B. uniformis DA183. من خلال دراسات التعبير الجيني والتلاعبات الجينية، حددنا مجموعة الجينات الرئيسية PUL34_Bu والإنزيم الحاسم GH26_BuDA183 المعنيين باستخدام DPs. كما أوضحت اختبارات نشاط الإنزيم في المختبر والتوصيل الجزيئي الآلية الجزيئية لعمل GH26. توفر هذه النتائج رؤى قيمة حول كيفية استخدام ميكروبات أمعاء الإنسان للبوليسكاريدات الطبية وتقدم معلومات مهمة لتطوير أدوية جديدة قائمة على البوليسكاريد.
النتائج
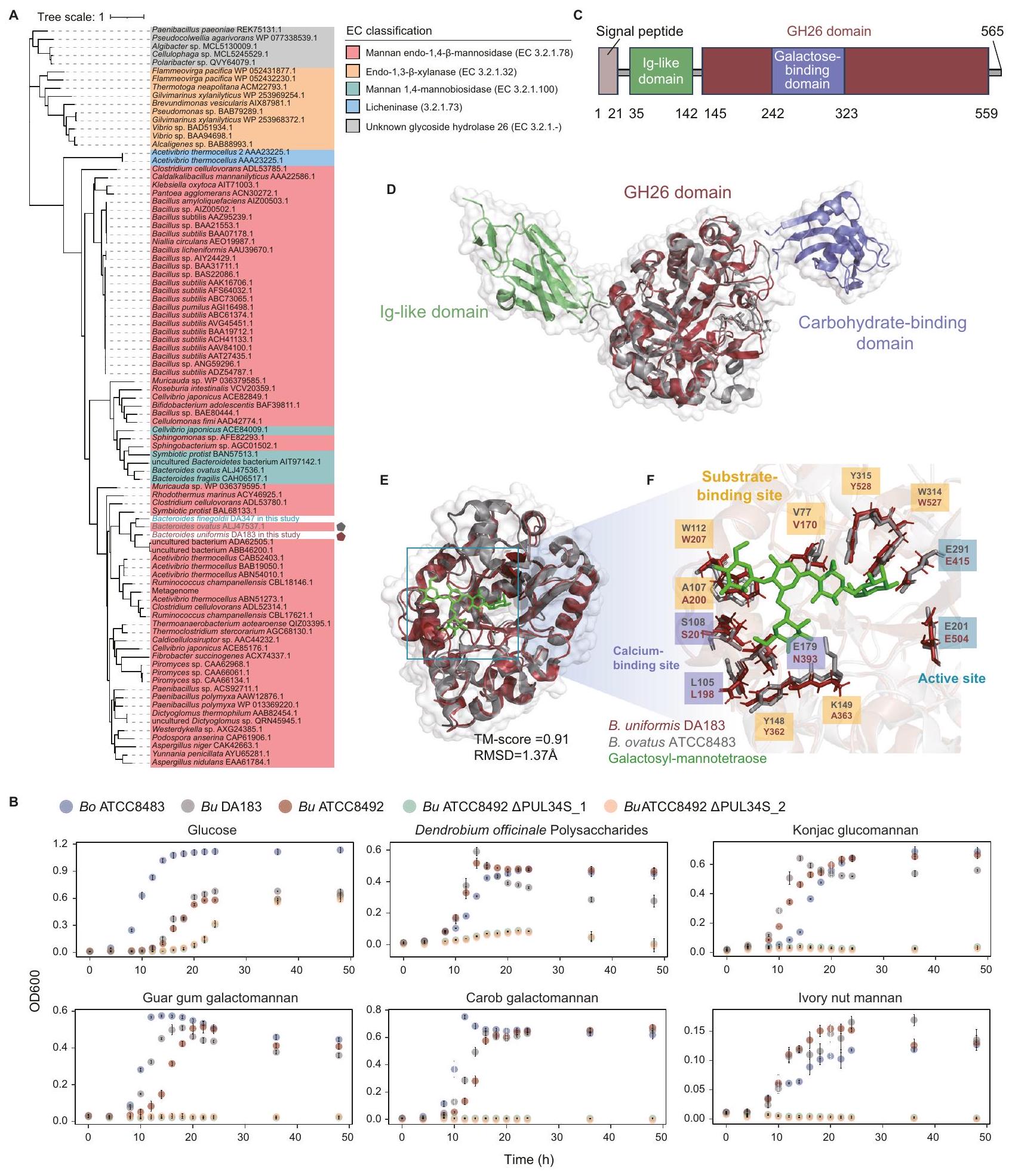
رسم خريطة استخدام البولي سكريات الطبية بواسطة أنواع بكتيرويدس وبارابكتيرويدس في أمعاء الإنسان
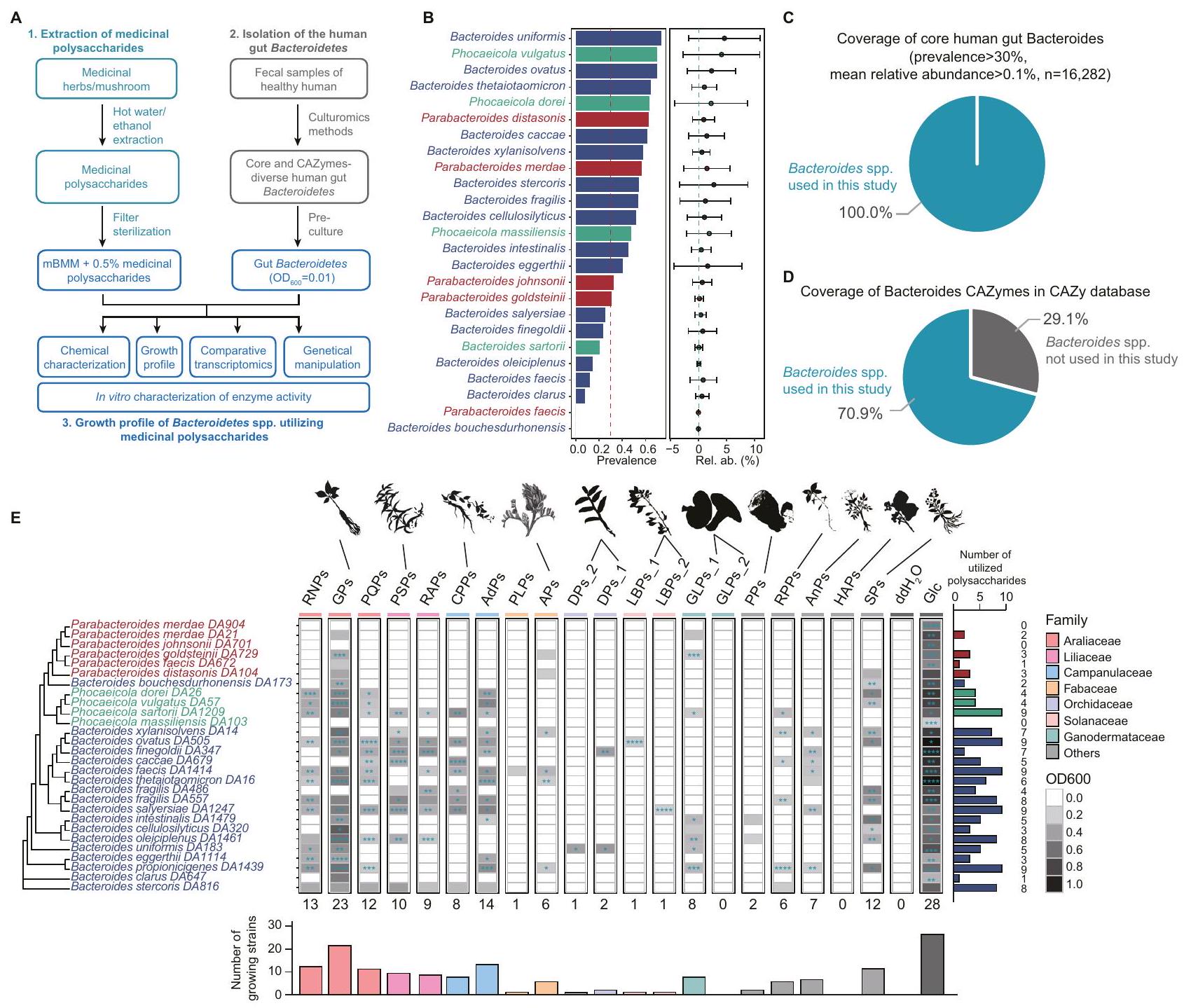
لتحليل شامل لاستخدام البوليسكاريدات الطبية بواسطة بكتيريا الأمعاء البشرية من نوع باكتيرويدس وباراباكتيرويدس، قمنا بتطوير منصة لملف نمو البكتيريا المعوية في المختبر كما هو موضح في الشكل 1A، بدءًا من اختبار ثلاثة وسائط غذائية بسيطة. أظهرت منحنيات النمو أن الوسط الغذائي المعدل لبكتيرويدس (mBMM)، وهو وسط صناعي، يدعم عمومًا نمو أنواع باكتيرويدس وباراباكتيرويدس (الشكل التوضيحي التكميلي 1).
تقوم بكتيريا باكتيرويدس وبارا باكتيرويدس، التي تتواجد بكثرة ووفرة في ميكروبيوم الأمعاء البشرية، بترميز العديد من PULs. تشمل المنصة 18 نوعًا من باكتيرويدس، و6 أنواع من بارا باكتيرويدس، و4 أنواع من فوكايكولا (سابقًا أنواع باكتيرويدس). تم اختيار بعض السلالات بشكل متكرر على مستوى الأنواع (مثل B. fragilis DA486 وDA557) بسبب ملفاتها المختلفة من CAZyme. لتقييم انتشار ومتوسط وفرة هذه الأنواع من باكتيرويدس في ميكروبيوم الأمعاء البشرية، قمنا بتحليل مجموعات بيانات جينات 16S rRNA المتاحة للجمهور. ) من GMrepo . وجدنا أن لأنواع البكتيرويديس كانت لها انتشار أكبر من، و كان لديه وفرة نسبية متوسطة أكبر من 0.1% (الشكل 1B، الجداول التكميلية 1، 2). كانت تغطية الأنواع الأساسية من بكتيرويدس الأمعاء البشرية (انتشارالمتوسط النسبي وفرة ) وإنزيمات CAZymes في البكتيرويدز (قاعدة بيانات CAZy ) كان و على التوالي (الشكل 1C، D). تضمن هذه الأنواع المختارة من البكتيرويد تحليلًا منهجيًا وشاملًا لاستخدام البوليسكاريد.
تم الإبلاغ عن البكتيرويدات والبارابكتيرويدات كأهداف للعديد من البوليسكاريدات الطبية في الأمعاء، لكن آليات استخدامها لا تزال غير مستكشفة بشكل كاف. لتوسيع فهمنا، اخترنا 20 بوليسكاريد طبي مختلف من الناحية التصنيفية مع هيكل مميز (أي ارتباط السكر والتعديل) (الجدول 1). على سبيل المثال، تحتوي البوليسكاريدات من عائلة الأراليات على جلوكانات شبيهة بالنشا، وأرابينوجالاكتانات،-هيكل غالا (هوموغالكتورونان)، بكتين غني بالرمانوغالاكتورونانتتميز بوليسكاريدات الدندروبيوم (DPs_1 و DPs_2، المُعدة باستخدام طرق مختلفة) بهياكل عظمية من )- -D-جلوكوز- – -دي-مانب--2-O-أسيتيل--دي-مانب-، و -3-O-أسيتيل--دي-مانب-تظهر هذه العشرون بوليسكاريد طبي نشاطات بيولوجية متعددة، بما في ذلك مضادة للورم، ومضادة للأكسدة، ومضادة للالتهابات، وتنظيم المناعة، مع الحفاظ على سمية منخفضة وآثار جانبية minimal. يمكن العثور على مزيد من التفاصيل حول بوليسكاريدات طبية أخرى مستخدمة في هذه الدراسة في الجدول التكميلي 3A وB.
وجدنا أن بيانات نمو البكتيريا كانت متسقة عبر ثلاثة تكرارات فنية. ) (الشكل التوضيحي الإضافي ) وبين تجربتين مستقلتين ( ، ) (الشكل التوضيحي التكميلي 2D)، مما يشير إلى أن طريقتنا قابلة للتكرار بشكل كبير. كشف تصنيف أنماط النمو عن تنوع في ملفات النمو بين أنواع البكتيرويد المختلفة والبوليسكاريدات الطبية (الشكل 1E، الشكل التوضيحي التكميلي 3A-C). أظهرت التحليلات الإحصائية أن كل سلالة يمكن أن تستخدم في المتوسط 4.79 نوعًا من البوليسكاريدات الطبية، حيث يمكن لبعض السلالات استخدام ما يصل إلى 9 بوليسكاريدات طبية، بينما لا تستطيع سلالات أخرى استخدام أي من 20 بوليسكاريدًا طبيًا. يمكن لكل نوع من البوليسكاريدات الطبية دعم نمو متوسط 6.8 سلالات. تدعم بعض البوليسكاريدات، مثل GPs، نمو عدة أنواع من البكتيرويد وبارابكتيرويد، بينما تدعم أخرى، مثل DPs_2، نوعًا واحدًا فقط من البكتيرويد. وجدنا أن البوليسكاريدات المستخرجة من النباتات ذات النسب القريبة أدت إلى ملفات نمو بكتيرية مشابهة (الشكل التوضيحي التكميلي 3D). من المحتمل أن تكون هذه الشبه ناتجة عن تركيبات وهياكل بوليسكاريد مشابهة، كما هو موضح في RNPs و PQPs، التي تحتوي كلاهما على Ara و Gal و GalA و Glc و GlcA و Man و Rha، مع الروابط الجليكوسيدية التي تشكل هياكلها الخطية .
قد تكون اختلافات ملفات نمو البوليسكاريدات الطبية ناتجة عن الاختلافات في الخصائص الفيزيائية والكيميائية للبوليسكاريدات، مثل تركيب السكريات الأحادية وبنية البوليسكاريدات. لتحديد ما إذا كانت بنية البوليسكاريد أو تركيب السكريات الأحادية تسبب اختلافات في ملفات النمو، اخترنا ثمانية بوليسكاريدات طبية تظهر ملفات نمو مختلفة. أولاً، قمنا بتوصيف الخصائص الفيزيائية والكيميائية لهذه البوليسكاريدات (الشكل التوضيحي التكميلي 4A-B، الشكل التوضيحي التكميلي 5A-D، والجداول التكميلية 5-7). أظهر تحليل تركيب السكريات الأحادية أن APs و GPs و GLPs_1 و GLPs_2 تتكون بشكل رئيسي من الجلوكوز (Glc) (92.8%،، و ، على التوالي). كانت DPs_1 و DPs_2 تتكون بشكل رئيسي من المانوز (Man) (و78.7%، على التوالي). كانت LBPs_1 تتكون بشكل رئيسي منأرابينوز (Ara)جالاكتوز، وحمض الجالاكتورونيك (GalA)، بينما كانت LBPs_2 تتكون بشكل رئيسي من، و أظهرت السكريات المتعددة الطبية المختارة اختلافات في محتوى السكر الكلي، والوزن الجزيئي والتوزيع، وتركيب السكريات الأحادية، مما يدل على أن السكريات المتعددة الطبية كانت متنوعة في الخصائص الفيزيائية والكيميائية. ومن الجدير بالذكر أننا لم نجد تقريبًا أي ارتباط بين مسافة تركيب السكريات الأحادية وملف النمو، مما يشير إلى أن السكريات المتعددة ذات التركيب المشابه للسكريات الأحادية لم تظهر ملفات نمو مشابهة (الشكل التكميلي 4C). على سبيل المثال، سكر متعدد الفصوص (APs) و GPs،
الشكل 1 | ملف استخدام البوليسكاريدات الطبية بواسطة أنواع بكتيرويدس وبارابكتيرويدس في أمعاء الإنسان. أ مخطط التدفق الذي يوضح العملية التجريبية لتوصيف ملف النمو للبوليسكاريدات الطبية، بما في ذلك i) استخراج البوليسكاريدات الطبية باستخدام الماء الساخن/الإيثانول، ii) عزل أنواع بكتيرويدس من أمعاء الإنسان من خلال الثقافة، iii) ملف نمو أنواع بكتيرويدس باستخدام وسط غذائي بسيط مضاف إليهالبوليسكاريدات الطبية في غرفة لاهوائية، و تم قياس كثافة النمو الضوئي (OD) وتسجيلها كل 12 ساعة باستخدام قارئ الميكرو بلايت. ب. انتشار (مخطط عمودي) ووفرة نسبية (مخطط نقطي) لأنواع بكتيرويد بناءً على بيانات تسلسل الميتاجينوم البشري العامة. ). الـ ‘ يشير إلى عدد عينات البراز المجمعة من الأفراد الأصحاء في GMrepo قاعدة البيانات. تم تلوين أنواع البكتيرويد بواسطة الجنس، حيث تم تلوين البكتيرويد باللون الأزرق، والبارابكتيرويد باللون الأحمر، والفوكايكولا باللون الأخضر. الانتشار0.3 (خط متقطع أحمر)، الوفرة النسبية (خط متقطع أخضر). تُعبر بيانات الوفرة النسبية عن القيم المتوسطة ± الانحراف المعياري (SD). C أظهر الرسم البياني الدائري تغطية بكتيريا باكتيرويدس الأساسية في الأمعاء البشرية (انتشار >متوسط الوفرة النسبية ). د أظهر الرسم البياني الدائري تغطية إنزيمات الكربوهيدرات النشطة لبكتيرويدس الأمعاء البشرية في قاعدة بيانات CAZy. هـ أظهر خريطة الحرارة نمو OD600 لـ 28 نوعًا من بكتيرويدس بعد 48 ساعة تحت 20 البوليسكاريدات الطبيةتم تحديد الدلالة الإحصائية باستخدام اختبار t غير المتزاوج ثنائي الجانب بين كل مجموعة تجريبية ومجموعة التحكم السلبية، مع إجراء تعديلات على المقارنات المتعددة باستخدام طريقة بونفيروني.القيم موضحة كما يلي: *<0.05؛; *** < 0.001; تم إجراء كل تجربة ثلاث مرات لضمان الموثوقية. الرسم البياني الفرعي على اليمين يحصي عدد السكريات المتعددة الطبية المستخدمة من قبل أنواع البكتيرويديت. الرسم البياني الفرعي في الأسفل يحصي عدد أنواع البكتيرويديت التي تستخدم السكريات المتعددة الطبية. RNPs: سكريات راديكس نوتوجينسينغ؛ GPs: سكريات الجينسنغ؛ PQPs: سكريات بانكس كوينكوفوليوم؛ PSPs: سكريات بوليغوناتوم سيبيريكوم؛ RAPs: سكريات ريزوما أنيمارينا؛ CPPs: سكريات كودونوبسيس بيلوسولا؛ AdPs: سكريات أدينوفورا؛ PLPs: سكريات بويراريا لوبياتا؛ APs: سكريات أستراجالوس؛ DPs_1 و DPs_2: سكريات دندروبيوم؛ GLPs_1 و GLPs_2: سكريات غانوديرما لوسيدوم؛ LBPs_1 و LBPs_2: سكريات ليكيم بارباروم؛ PPs: سكريات بوريا؛ RPPs: سكريات راديكس بسيودوكسيلاريا؛ AnPs: سكريات أنجيليكا؛ HAPs: سكريات هوفينيا أسيبا؛ SPs: سكريات عائلة سكروفولاريا؛ PC: التحكم الإيجابي؛ NC: التحكم السلبي. على الرغم من أن لديها تركيبات مونو سكريد مشابهة، إلا أنها أظهرت أنماط نمو مختلفة بشكل ملحوظ لبكتيريا الأمعاء البشرية. ومن الجدير بالذكر أن APs تتكون من غالاكتورونيك ميثيل.وسلاسل جانبية متنوعة، تختلف عن السكريات الأحادية. تشير هذه النتائج إلى أن بنية البوليسكاريد، بدلاً من تركيب السكريات الأحادية، هي التي تؤثر على اختلافات نمط النمو.
يرتبط ملف استخدام البوليسكاريدات الطبية بالتنوع الجيني في الإنزيمات النشطة على الكربوهيدرات
توصيف التركيب الكيميائي للبوليسكاريدات هو تحدٍ معترف به عالميًا. ومع ذلك، أدت البوليسكاريدات المعقدة إلى تطور العديد من عائلات الإنزيمات النشطة على الكربوهيدرات (CAZymes) المتنوعة. بدورها، فإن خصوصية التحلل
الجدول 1 | قائمة البوليسكاريدات الطبية
اختصار
البوليسكاريدات
مصدر المواد المستخرجة
RNPs
بوليسكاريد راديكس نوتوجينس
ف_أرالياسية؛ س_جذر النوتوجينسنج
الأطباء العامون
بوليسكاريد الجينسنغ
فصيلة الأرالية؛ نوع بانكس جينسنغ
PQPs
بوليسكاريد بانكس كوينكويفوليوم
فصيلة الأرالية؛ نوع بانكس كوينكوفوليوم
PSPs
بوليسكاريدات بوليغوناتوم سيبيريكوم
ف_ عائلة الزنبق؛ ص_ بوليغوناتوم سيبيريكوم
راب
بوليسكاريد أنيمارينا أسبوديلوديس
ف_ عائلة الزنبق؛ س_ أنيمارينا أسبوديلوديس
CPPs
بوليسكاريد من كودونوبسيس بيلوسولا
ف_ عائلة الجرسية؛ س_ كودونوبسيس بيلوسولا
إعلانات الدفع لكل نقرة
بوليسكاريدات غلينيا ليتوراليس
ف_ عائلة الجرسية؛ س_ غلينيا الساحلية
PLPs
بوليسكاريد بويراريا لوبياتا
ف_ الفاباسيا؛ س_ بويراريا لوباتا
APs
بوليسكاريد الأستراجالوس
ف_ الفصيلة البقولية؛ س_ الأستراجالوس الغشائي
دي بيز_1
بوليسكاريد دندروبيوم أوفيسينال 1
ف_ الأوركيدية؛ س_ دندروبيوم أوفيسينالي
دي بيز_2
بوليسكاريد دندروبيوم أوفيسينال 2
ف_ الأوركيدية؛ س_ دندروبيوم أوفيسينال
LBPs_1
بوليسكاريدس ليكيم بارباروم 1
ف_ الباذنجانية؛ س_ ليكيم بارباروم
LBPs_2
بوليسكاريدس ليكيوم بارباروم 2
ف_ الباذنجانية؛ س_ ليكيم بارباروم
GLPs_1
بوليسكاريد غانوديرما لوسيدوم 1
ف_ عائلة الغانوديرماتاس; س_ غانودرما لوسيدوم
GLPs_2
بوليسكاريد غانوديرما لوسيدوم 2
ف_ عائلة الغانوديرماتاس; س_ غانوديرما لوسيدوم
بي بي إس
بوليسكاريد البوريا
فصيلة البوليبوراسيا؛ نوع بورتيا كوكوس
RPPs
بوليسكاريدات راديكس بوسيدوكسيلاريا
ف_ القرنفلية؛ ص_ بيسودوستيلاريا هيتيروفيلا
أنبس
بوليسكاريد أنجيليكا
ف_ الخيميات؛ س_ أنجليكا سينيensis
HAPs
بوليسكاريدات هوفينيا أسيباربا
فصيلة الرمانيات؛ نوع هوفنيا الحامضة
مزودو الخدمة
بوليسكاريد من عائلة السكروفولاريا
ف_ عائلة السّكروفولاريا; س_ سكروفولاريا نينغبوينسيس
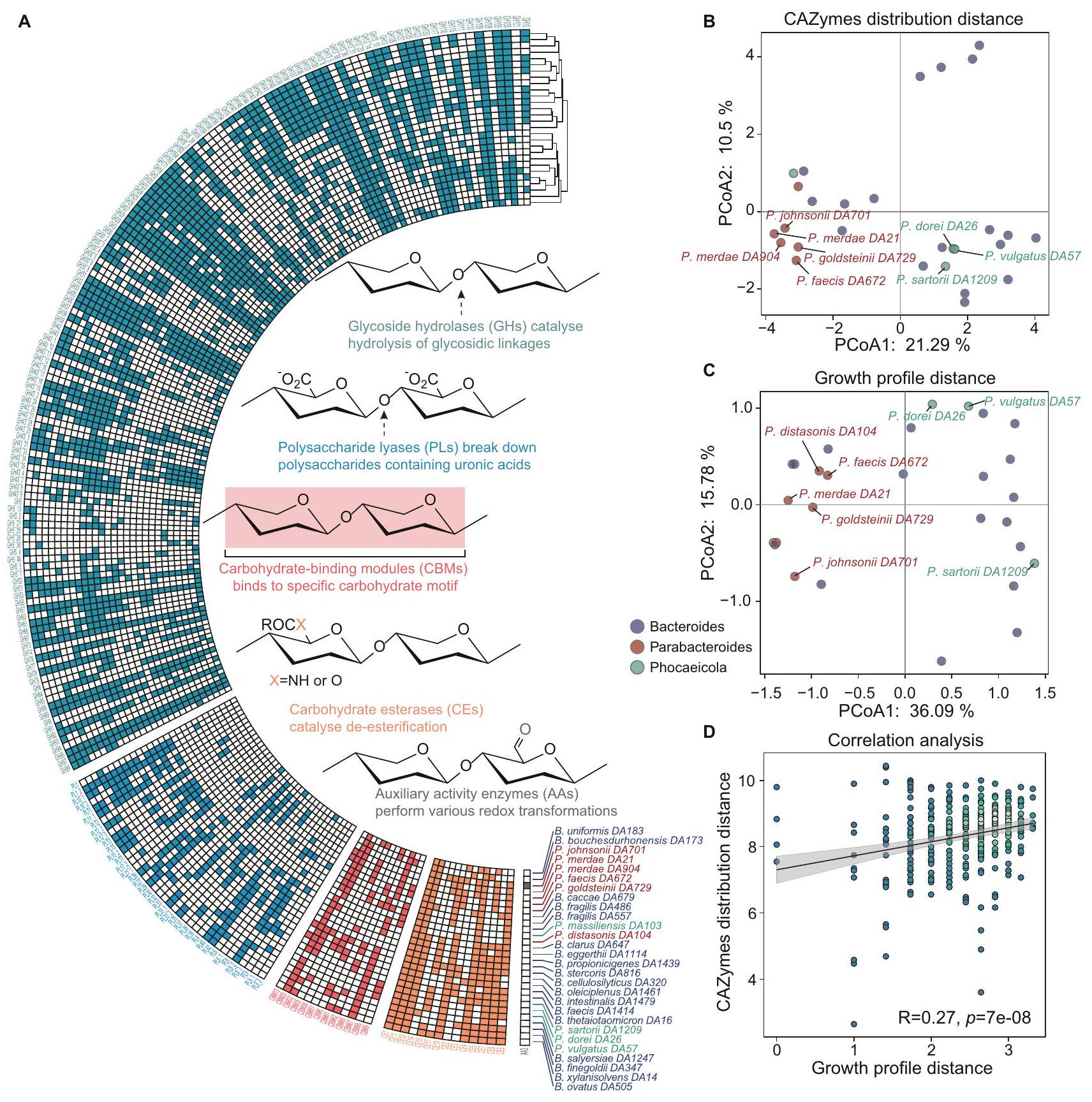
ملاحظات: f_ تعني العائلة؛ s_ تعني النوع يمكن أن توفر الإنزيمات معلومات حول بنية البوليمرات السكرية المتحللة. يمكن أن تساعدنا الجينات المشفرة بواسطة بكتيريا الأمعاء المختلفة في فهم ملف الاستخدام. تشفر بكتيريا باكتيرويدز مئات من CAZymes لاستخدام البوليمرات السكرية المعقدة.الإنزيمات المعتمدة على الكربوهيدرات المشاركة في تحلل الكربوهيدراتتشمل الهيدرازات الجليكوسيدية (GHs) لتحليل الروابط الجليكوسيدية، والليازات متعددة السكاريد (PLs) لتفكيك متعددات السكاريد التي تحتوي على الأحماض الأورونية، ووحدات ربط الكربوهيدرات (CBMs) لربط أنماط الكربوهيدرات، وإسترازات الكربوهيدرات (CEs) لإزالة الاستر، وإنزيمات النشاط المساعد (AAs) لتحولات الأكسدة والاختزال المختلفة. باستخدام dbCAN (الإصدار 4.1.4)لتحليل CAZymes المعلّمة، لاحظنا تباينات في توزيع CAZyme على مستويات الجنس والأنواع والسلالات بين أنواع Bacteroides وParabacteroides (الشكل 2A). أظهر تجميع الـ 28 نوعًا من Bacteroides بناءً على المسافة الإقليدية لـ CAZymes أن Bacteroides وParabacteroides ذوي التصنيف المشابه يتشاركون في أنماط توزيع CAZyme مشابهة (الشكل 2A). كانت GHs الأكثر ترميزًا على نطاق واسع، في حين كانت AAs أقل شيوعًا بسبب البيئة اللاهوائية في الأمعاء التي تحد من AAs المعتمدة على الأكسجين..
لتفسير اختلافات ملف نمو الأنواع المختلفة من بكتيرويدس وبارابكتيرويدس، قمنا بتجميع 28 سلالة بناءً على المسافة الإقليدية للإنزيمات المرتبطة بالسكريات (CAZymes) (الشكل 2B)، وتوزيع ملف النمو (الشكل 2C). وجدنا أن الأنواع من بكتيرويدس التي لديها توزيع مشابه لجينات CAZyme كانت لديها نمط أيضي مشابه للسكريات المتعددة. على سبيل المثال، أظهرت P. dorei DA26 وP. vulgatus DA57 توزيعات مشابهة لجينات CAZyme وأنماط أيضية مشابهة للسكريات المتعددة. لوحظ ظاهرة مشابهة بين الأنواع من بارابكتيرويدس. بالإضافة إلى ذلك، لوحظ ارتباط كبير بين المسافات لتوزيع CAZyme وملفات النمو، مما يشير إلى أن بكتيرويدس وبارابكتيرويدس الذين لديهم توزيعات مشابهة للـ CAZyme يظهرون ملفات نمو مشابهة (الشكل 2D). تشير هذه النتائج إلى أن توزيع CAZyme يدفع اختلافات ملفات النمو.
تحليل النسخ الجيني يكشف أن بعض وحدات الجينات المحددة مرتفعة بشكل كبير بواسطة البوليسكاريدات الطبية
يساعد التعبير الجيني التفاضلي في اكتشاف الإنزيمات والمسارات لاستخدام مصادر الكربون المحددة. أولاً، اخترنا بوليسكاريدات الجينسنغ (GPs) التي دعمت نمو العديد من
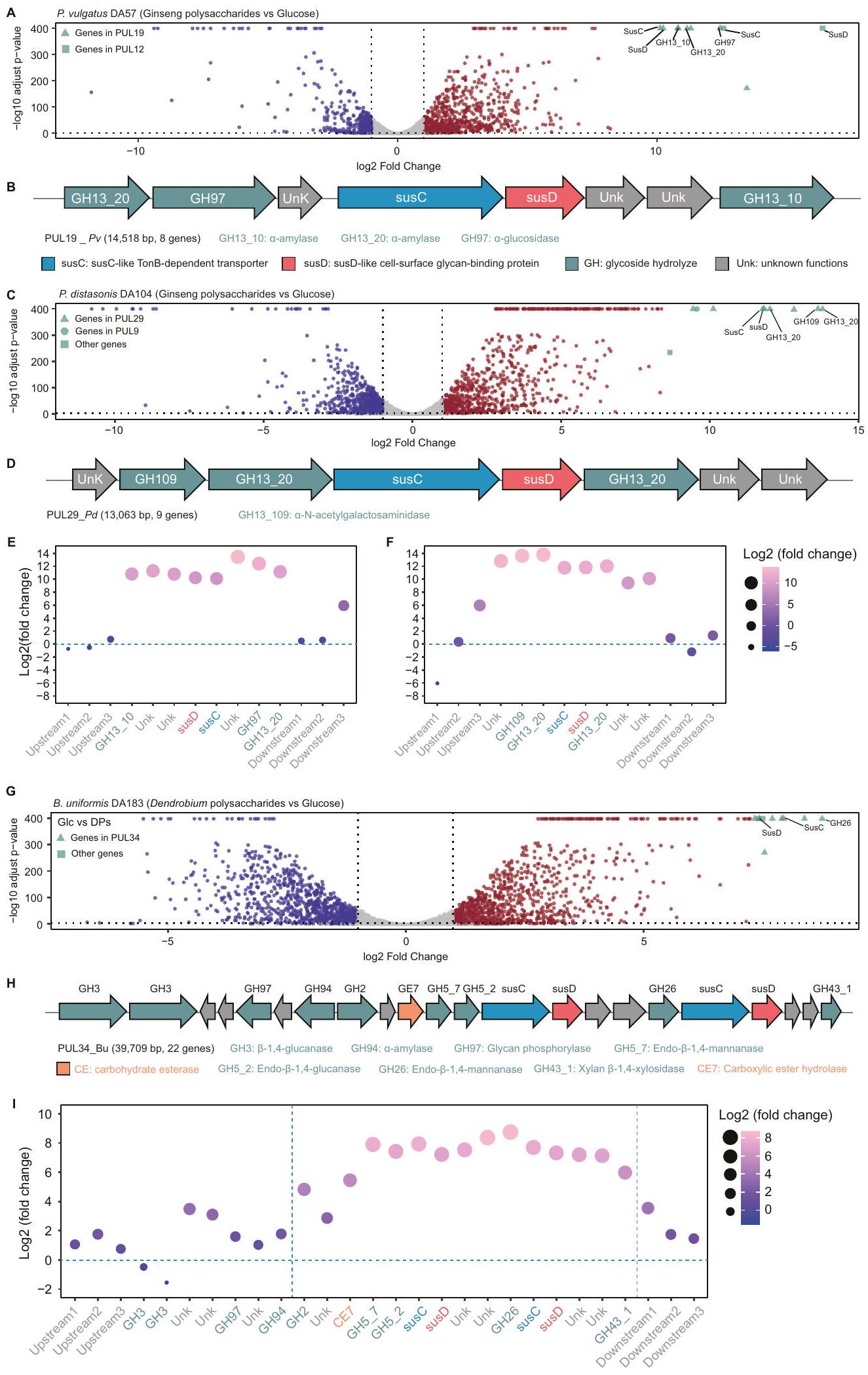
باكتيرويدس. قمنا بتحليل بيانات النسخ الجيني لـ P. vulgatus DA57 و. تم زراعة P. distasonis DA104 باستخدام GPs والجلوكوز كمصدرين للكربون الوحيدين (الشكل 3A-F، الشكل التوضيحي 6A-D، والجداول التكميلية 8 و9). أظهر تحليل النسخ المقارن أن أعلى 10 جينات في عملية التمثيل الغذائي للكربوهيدرات كانت موجودة في PUL19 من P. vulgatus DA57 (PUL19_Pv) (الشكل 3A) وPUL29 من P. distasonis DA104 (PUL29_Pd) (الشكل 3C) كما تم توضيحه بواسطة PULpy.قمنا بتنقيح حدود PUL بناءً على مستويات التعبير للجينات العليا والسفلى (الشكل 3C وE وI، والشكل التوضيحي 7C) (انظر “الطرق” لمزيد من التفاصيل). ومن الجدير بالذكر أن GH13_10 وGH13_20، تم وصفهما كـالأميلازات -أ بواسطة قاعدة بيانات CAZyكانت مرتفعة في كل من PUL19 Pv و PUL29 Pd. نظرًا لأن السكريات المتعادلة، بما في ذلك خلطات الأميلويدتشير هذه النتائج إلى أن GH13_10 و GH13_20 هما عنصران حاسمان في استخدام GPs.
بعد ذلك، قمنا بفحص بيانات التعبير الجيني لبكتيرويدس باستخدام بوليسكاريد غانوديرما لوسيدوم (GLPs_1)، والذي دعم نمو عدد معتدل من بكتيرويدس. كانت أعلى 10 جينات لتمثيل الكربوهيدرات التي تم تنظيمها تقع في PUL16 وPUL25 وPUL39 من B. uniformis DA183 (الشكل التوضيحي التكميلي 6E-F، الشكل التوضيحي التكميلي 7A، والجدول التكميلي 10). أظهر تحليل هياكل تجمعات الجينات أن هذه PULs ترمز إلى GH55 (أي، إكسو/إندو--1,3-غلوكانيز) في PUL16، GH9 (أي، إندو--1,4-غلوكانيز) في PUL16&25، GH3 (أي، إندو--1,6-غلوكوزيداز) في PUL39، و GH16_3 و GH158 (أي، إندو-(-1,3-غلوكانيز) في PUL39 (الشكل التوضيحي 7B، C). وجود عدة PULs مسؤولة عن نفس البوليسكاريد يتماشى مع الدراسات السابقةتتكون GLPs من )- -D-GIcp- –-D-جالكتوز-، و – -D-جلوكوز-تشير هذه النتائج إلى أن PUL16 و PUL25 و PUL39 من المحتمل أن تكون مسؤولة عن استخدام GLP.
قمنا أيضًا بتحليل بيانات التعبير الجيني لبكتيرويدس باستخدام DPs_2، التي تعزز بشكل انتقائي نمو B. uniformis. مع التركيز على B. uniformis DA183، قمنا بمقارنة بيانات النسخ الجيني عند زراعتها باستخدام DPs والجلوكوز كمصدرين للكربون. كانت أعلى 10 جينات في عملية التمثيل الغذائي للكربوهيدرات التي تم تنظيمها بشكل إيجابي تقع في PUL34 من B. uniformis DA183 (PUL34_Bu) (الشكل 3G-I، الشكل التوضيحي 6E-F، الجدول التوضيحي 11). DPs هي أوليغوسكاريد مانوز يتكون هيكله العظمي من-D-جلك-دي مان
الشكل 2 | التباين الجينومي في الإنزيمات النشطة على الكربوهيدرات يحدد الملف المميز لاستخدام البوليسكاريدات الطبية بين أنواع باكتيرويدس وبارا باكتيرويدس. أ تظهر خريطة الحرارة توزيع جينات CAZyme المعنية في تحلل الكربوهيدرات. تم تجميع أنواع باكتيرويديتس بناءً على المسافة الإقليدية لجينات CAZyme. تم تجميع جينات CAZymes بناءً على المسافة الإقليدية للتوزيع في باكتيرويدس وبارا باكتيرويدس. هيدرازات الجليكوسيد (GHs): التحلل المائي و/أو إعادة ترتيب الروابط الجليكوسيدية؛ ليزات البوليسكاريد (PLs): الانقسام غير التحللي للروابط الجليكوسيدية؛ وحدات ربط الكربوهيدرات (CBMs): الالتصاق بالكربوهيدرات؛ استرازات الكربوهيدرات (CEs): التحلل المائي لاسترازات الكربوهيدرات؛ الأنشطة المساعدة (AAs): إنزيمات الأكسدة والاختزال التي تعمل بالتزامن مع CAZymes. ب تحليل المكونات الرئيسية (PCA) المعتمد على براي كورتيس يظهر تحليل تقليل الأبعاد لـ إنزيمات الكربوهيدرات النشطة في بكتيرويدس. النقاط على الرسم البياني ملونة وفقًا لجنس بكتيرويديتس. التحليل الرئيسي للمكونات (PCA) القائم على المسافة الإقليدية يظهر تحليل تقليل الأبعاد لاستخدام البوليسكاريدات بعد 48 ساعة بواسطة بكتيرويدس. النقاط على الرسم البياني ملونة وفقًا لجنس بكتيرويديتس. يعرض مخطط النقاط العلاقة بين مسافة ملف النمو ومسافة توزيع إنزيمات الكربوهيدرات.تشير القيم إلى النموذج الأكثر اقتصادية. تم تحديد الدلالة الإحصائية باستخدام اختبار ارتباط بيرسون ثنائي الجانب (لقيمة R) وتحليل الانحدار العادي (OLS) (للتناسب الخطي). تتضمن الشكل أيضًا “فترة الثقة حول أفضل ملاءمة خطية. كانت النقاط الخضراء الفاتحة تمثل كثافة نقاط عالية والنقاط الخضراء الداكنة تمثل كثافة نقاط منخفضة.
أو-أسيتيل--دي-مانب-، و -3-O-أسيتيل--دي-مانب-. تؤكد التركيبة المقاسة من السكريات الأحادية لـ DPs أن المانوز هو المكون الرئيسي (الشكل التوضيحي 4A، B). تم وصف إنزيمين من الجليكوسيد هيدراز (GHs) تم تنظيمهما بشكل كبير في PUL34_Bu، GH26 و GH5_7، على أنهما إكسو- -1,4-مانوبيوز و إندو--1,4ماناناز بواسطة قاعدة بيانات CAZy.
مجموعة جينات PUL34_Bu في B. uniformis مطلوبة لاستغلال بوليسكاريدات دندروبيوم
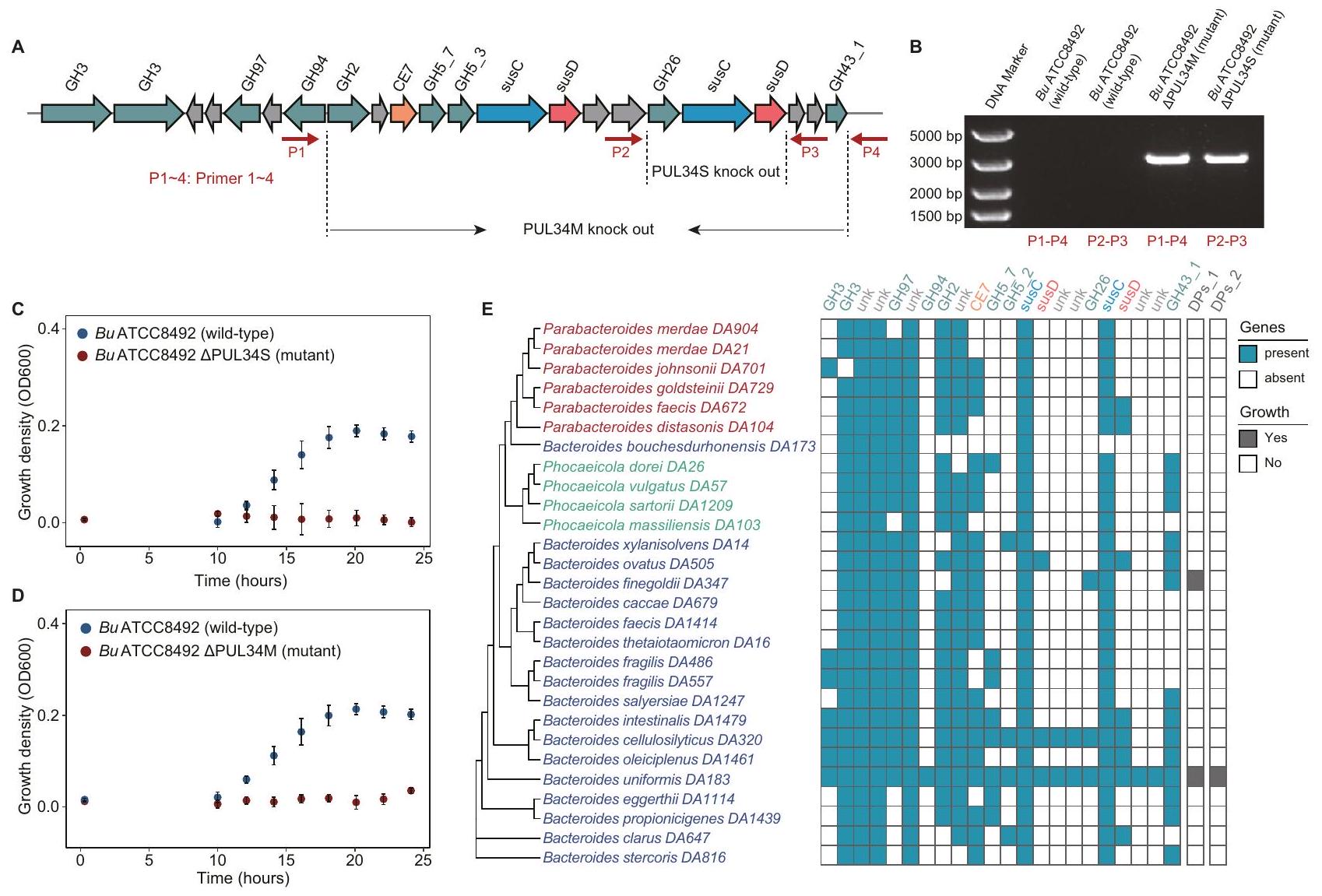
لاستكشاف الأساس الجيني الذي يكمن وراء الاستخدام المحدد للغاية لسكريات دندروبيوم (DPs) بواسطة بكتيريا ب. يونيكوميس، استخدمنا أداة تحرير جينوم بكتيرويديس المعتمدة على تقنية كريسبر/كاس.لبناء طفرات حذف PUL34 (الطرق). لضمان نجاح الجين الضربة القاضية، استهدفنا منطقتين في مجموعة الجينات لإجراء ضربة قاضية منفصلة (الشكل 4A). كانت المنطقة الأولى (PUL34M_Bu) مقطعًا متوسط الحجم مع زيادة ملحوظة في مستوى النسخ وجينات يتم نسخها في نفس الاتجاه. احتوت المنطقة الثانية (PUL34S_Bu) على ثلاثة إنزيمات رئيسية كانت نظائر إنزيمات رئيسية ثلاثة متورطة في استخدام الإينولين.بما في ذلك بروتين مشابه لبروتين SusD لارتباط البوليسكاريد في الغشاء الخارجي، يوجد بروتين شبيه بـ SusC لنقل البوليمرات السكرية، و GH26 لتحلل الروابط الجليكوسيدية. أكدت تقنية الرحلان الكهربائي للهلام وتسلسل الحمض النووي نجاح إلغاء المنطقتين (الشكل 4B، الشكل التوضيحي 9A-D). عند زراعتها في وسط mBMM مع DPs كمصدر الكربون الوحيد، نمت السلالة البرية، بينما لم تنمو السلالة الطافرة (الشكل 4C وD).
الشكل 3 | زيادة تنظيم جينات مواقع استخدام البوليسكاريد المحددة بواسطة البوليسكاريدات الطبية. رسم بياني بركاني لـتعبير جين DA57 في الفولغاتوس مقارنةً بالوسط الأدنى المدعوم بالـ GPs مع الجلوكوز، موضحًا نسبة التغير ( -المحور) مقابل الدلالة التفاضلية (-لوغاريتم 10 المعدل-قيمة،المحور). تم تمييز أعلى 10 جينات مرتفعة التعبير باللون الأخضر. تم إجراء تحليل التعبير التفاضلي باستخدام DESeq2، مع استخدام اختبار والد ذو الجانبين. تم تعديلتم الحصول على القيم بعد تصحيحها لاختبار الفرضيات المتعددة باستخدام طريقة بنجاميني وهوشبرغ للتحكم في معدل الاكتشاف الخاطئ (FDR). الجينات التي تم تعديلها-قيمةتعتبر ذات دلالة إحصائية.-المحور يمثلتغير الطي في تعبير الجين، بينما-المحور يظهر الـ-المعدل-القيمة. تم إجراء كل تجربة ثلاث مرات لضمان الموثوقية. ب يوضح المخطط أن PUL19 من P. vulgatus DA57 (PUL19_Pv) يحتوي على 8 جينات، بحجم مجموعة يبلغ 14,518 قاعدة. susC: ناقل معتمد على TonB يشبه susC؛ susD: سكر سطحي يشبه susD- بروتين الربط؛ GH: هيدروكسي الجليكوسيد؛ Unk: وظائف غير معروفة. C مخططات البركان للتعبير الجيني لـ P. distasonis DA104 مقارنةً بالوسائط الدنيا المدعمة بـ GPs والجلوكوز. التحليل الإحصائي هو نفسه كما في (A). D يوضح المخطط التخطيطي أن PUL29 من P. distasonis DA104 (PUL29 Pd) يحتوي على 8 جينات، بحجم مجموعة يبلغ 13,063 قاعدة. E-F يعرض مخطط الفقاعات التعبير التفاضلي لـ E) جينات PUL19 Pv ، و F) جينات PUL29 Pd وجيناته الثلاثة المجاورة، باستخدام GPs والجلوكوز. G مخططات البركان للتعبير الجيني لـ B. uniformis DA183 مقارنةً بالوسائط الدنيا المدعمة بـ DPs والجلوكوز. التحليل الإحصائي هو نفسه كما في (A). H يوضح المخطط التخطيطي أن PUL34_Bu يحتوي على 22 جينًا، بحجم مجموعة يبلغ 39,709 قاعدة. CE: استراز الكربوهيدرات. I يعرض مخطط الفقاعات التعبير التفاضلي لجين PUL34_Bu وجيناته الثلاثة المجاورة، باستخدام DPs والجلوكوز. المنطقة بين خطين عموديين متقطعين باللون السماوي تشير إلى الجينات التي تم تنظيمها بشكل كبير مع اتجاه نسخ متسق.
الشكل 4 | مجموعة جينات PUL34_Bu في B. uniformis مطلوبة لاستخدام بوليسكاريد دندروبيوم. A يوضح الرسم التخطيطي منطقتين من جينات PUL34_Bu التي تم حذفها. تشير PUL34M_Bu إلى الجينات التي تم تنظيمها بشكل كبير مع اتجاه النسخ المتسق. تشير PUL34S_Bu إلى الجينات الثلاثة الرئيسية المحتملة. B يظهر مخطط الرحلان الكهربائي للأحماض النووية أربعة منتجات DNA: المنتجات التي تم تضخيمها باستخدام بادئات مصممة ضمن منطقة 1.5 كيلوباز تتاخم منطقة PUL34M_Bu (P1 ~ P4)، وDNA الجينومي من النوع البري، والسلالة المتحورة؛ المنتجات التي تم تضخيمها باستخدام بادئات مصممة ضمن منطقة 1.5 كيلوباز تتاخم منطقة PUL34S_Bu (P2 ~ P3)، وDNA الجينومي من النوع البري، والسلالة المتحورة. إذا تم حذف الشريحة بنجاح، سيتم تضخيم منتج بطول 3000 قاعدة. بالمقابل، إذا لم تنجح عملية الحذف، لن يتم تضخيم أي منتج بسبب الطول المفرط لشريط الهدف. هذه الصورة التمثيلية هي من تجربة واحدة. C-D يوضح الرسم البياني للنقاط كثافة النمو لسلالات C) النوع البري وسلالات PUL34S المحذوفة، وD) النوع البري وسلالات PUL34M المحذوفة في وسط mBMM مع DPs_2 كمصدر الكربون الوحيد. يتم تقديم البيانات كقيم متوسطة ± الانحراف المعياري. تم إجراء الاختبارات في ثلاث تكرارات فنية.تظهر خريطة الحرارة العلاقة بين PUL34_Bu وكثافة النمو OD600. تم تحديد جينات PUL34_Bu بواسطة BLASTP (الإصدار 2.6.0) (تغطيةحددتم تلوين الأنواع الحاملة لجينات PUL34_Bu باللون السماوي، بينما تم تلوين الأنواع الأخرى باللون الأبيض. الأنواع التي تستخدم DPs تم تلوينها باللون الرمادي، بينما تم تلوين الأنواع الأخرى باللون الأبيض.
لتحقيق توزيع PUL34_Bu في جينومات 27 نوعًا آخر من بكتيرويدس وبارابكتيرويدس، قمنا بإجراء محاذاة تسلسلية مع عتبة تشابه التسلسل بـوعتبة التغطية لـ (الطرق). وجدنا أن B. uniformis فقط هو الذي يشفر مجموعة جينات PUL34_Bu كاملة (الشكل 4E). باختصار، كشفنا أن مجموعة جينات PUL34_Bu الكاملة سمحت بالاستخدام المحدد للغاية للـ DPs بواسطة B. uniformis.
إن إنزيمات GH26 ضرورية لاستخدام المانان المستخرج من النباتات في B. uniformis
قمنا بإنشاء شجرة تطورية لـ 89 تسلسل بروتين GH26 المميز المودع في قاعدة بيانات CAZy. (طرق). تم تجميع B. uniformis GH26 مع إنزيمات أخرى (بما في ذلك إنزيمات GH26 من B. ovatus ATCC8483) الموصوفة كإنزيمات مانان إندو-1,4-مانوزيداز (EC 3.2.1.78) (الشكل 5A). لاستكشاف توزيع. الجرثومة GH26 الموحدة في بكتيريا الأمعاء البشرية، قمنا بإجراء محاذاة تسلسلية في قاعدة بيانات UHGPباستخدام طرق الماس. وجدنا أن المتجانسات B. uniformis GH26 كانت موجودة بشكل رئيسي في أنواع بكتيرويدس عند عتبة المحاذاة الوظيفية المحفوظة (الهوية تغطية )، بما في ذلك 64/66 ب. يونيكوم، 26/32 ب. سيلولوسيلتيكوس، 3/17 ب. أوفاتوس، 2/2 ب. ستيركوريوروس، و 2/2 ب. دوراي (الشكل التوضيحي التكميلي 10A، B).
للتحقيق في ضرورة PUL34_Bu لاستخدام المانانات المستمدة من النباتات بواسطة بكتيريا الأمعاء Bacteroides، قمنا بإجراء تحليل نمو لـ B. ovatus ATCC8483 وB. uniformis DA183 وB. uniformis ATCC8492 (النوع البري) وB. uniformis ATCC8492. PUL34S_1&_2 (تحور) باستخدام مانان مستخلص من النباتات، مثل بوليسكاريد دندروبيوم أوفيسينالي (DPs)، جلوكومانان كونجاكصمغ الغوار جالاكتومانانغالاكتومانان الخروبوزيت جوز العاجوجدنا أن سلالات B. uniformis و B. ovatus من النوع البري كانت قادرة على استخدام هذه المانان المستمدة من النباتات، بينما لم تتمكن الطفرات الناتجة عن حذف PUL34S من النمو (الشكل 5B). تشير هذه النتائج إلى أن GH26 في B. uniformis يقوم بتحلل مجموعة واسعة من ركائز المانان.
لفهم الخصائص الهيكلية لـ B. uniformis GH26 بشكل أفضل، قمنا بتوقع هيكله باستخدام AlphaFold2. (pLDDT=93.04). وجدنا أن B. uniformis GH26 يحتوي على ببتيد إشارة (البقايا 1 إلى 30)، ونطاق شبيه بالأجسام المضادة البكتيرية (البقايا 31 إلى 131)، ونطاق GH26 (البقايا 135 إلى 564)، ولبتين مرتبط بالغالكتوز (البقايا 239 إلى 354) (الشكل 5C، D). تم تحديد بنية إنزيم GH26 من B. ovatus ATCC8483 بواسطة البلورة (رقم الوصول في GenBank ALJ47537.1؛ رمز PDB 6HF4) . مقارنة مع هيكل . في B. uniformis GH26، وجدنا أن هيكل B. uniformis GH26 يحتوي على وحدتين إضافيتين: 1) مجال مشابه للأجسام المضادة والذي يتعلق باستقرار الإنزيمأو العمل كموصل بين مجالين; 2) وحدة ربط الكربوهيدرات. وجدنا أن المواقع النشطة، ومواقع ارتباط الكالسيوم، ومواقع ارتباط الركيزة في B. uniformis GH26 و B. ovatus GH26 محفوظة (درجة TM 0.91 ، RMSD ) (الشكل 5E، والشكل التكميلي 11).
البقايا المحفوظة من B. uniformis GH26 (W527 و Y528) و B. ovatus GH26 (W314 و Y315) هي المسؤولة عن ربط المانوز بواسطة الروابط الهيدروجينية.، مما يشير إلى أن الركيزة لـ. يحتوي GH26 من بكتيريا B. ovatus على المانوز. ومع ذلك، فإن بقايا K149 المسؤولة عن ربط الجالاكتوز ليست محفوظة بين الإنزيمين، مما يشير إلى خصوصية مختلفة لربط الركيزة. في الواقع، فإن ركيزة B. ovatus GH26 هي جالاكتومانان، الذي يختلف هيكليًا عن DPs (أي الجلوكومانان).
لقد لاحظنا أن DPs_1 قد حفز أيضًا نمو B. finegoldii DA347، الذي يحتوي على جين GH26 في جينومه (الشكل 4E). وقد دفعنا هذا الملاحظة إلى إجراء محاذاة تسلسلية وهيكلية لجين GH26 من B. finegoldii DA347. أظهرت محاذاة التسلسل تشابهًا منخفضًا بين GH26 من B. uniformis DA183 وB. ovatus ATCC8483 وB. finegoldii DA347. هوية التسلسل هيالتغطية) بين B. uniformis DA183 و B. ovatus ATCC8483، 49.1% (تغطية 52.0%) بين B. uniformis DA183 و B. finegoldii DA347، و 48.3% (التغطية) بين B. ovatus ATCC8483 و B. finegoldii DA347 (الشكل التوضيحي 12A). أظهر المحاذاة الهيكلية أن B. finegoldii DA347 ترمز لإنزيم مشابه هيكليًا لـ B. ovatus ATCC8483 GH26 (درجة TMRMSD ) (الشكل التكميلي 12B-C). علاوة على ذلك، فإن المواقع النشطة (E217 و E321)، مواقع ربط الكالسيوم (L109 و S112)، ومواقع ربط الركيزة (W116 و W353 و Y354) من GH26 من B. finegoldii DA347 (نماذج عصا زرقاء)، والمواقع النشطة (E201 و E291)، مواقع ربط الكالسيوم (L105 و S108)، ومواقع ربط الركيزة (W112 و W314 و Y315) من B. ovatus GH26 (نماذج عصا رمادية) محفوظة (الشكل التكميلي 12D). قد تفسر هذه النتائج قدرتها على النمو باستخدام DPs_1.
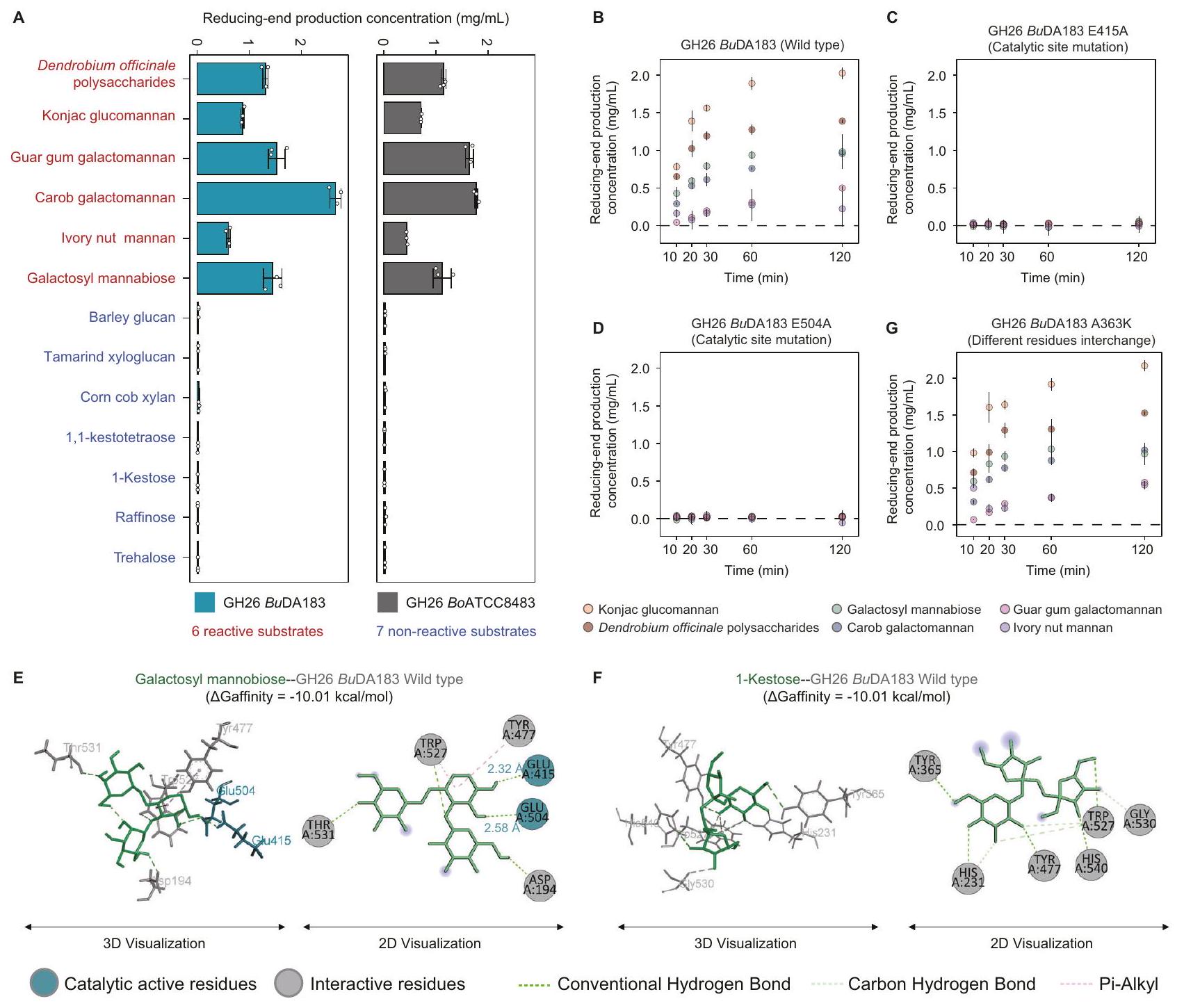
النشاط الإنزيمي والخصوصية لإنزيم Bacteroides GH26
لتوصيف نشاط الإنزيم وخصوصية الركيزة لبكتيرويدس GH26 بشكل أكبر، قمنا بتعبير وتنقية إنزيم GH26 من. قمنا بدراسة الإنزيمات uniformis DA183 و B. ovatus ATCC8483 (الشكل التكميلي 13A، B) واختبرناها مع 13 ركيزة، بما في ذلك 6 مانانات، و3 ركائز محتملة من بروتينات مشابهة هيكليًا أو من نفس عائلة البروتين، و4 ركائز محددة جيدًا كضوابط سلبية محتملة (الجدول التكميلي 3C). تم قياس إنتاج النهاية المخفضة باستخدام اختبار حمض 3,5-داينيتروساليسيليك (DNS) مع المانوز كمعيار (الطرق). وجدنا أن كلا الإنزيمين قام بتحليل جميع المانانات الستة، ولكن ليس الركائز السبع الأخرى (الشكل 6A، والجدول التكميلي 16A، B).
المخلفات الحفازة المفترضة هي اثنان من بقايا الجلوتاميك (E2O1 وE291 في B. ovatus ATCC8483 وE415 وE504 في B. uniformis DA183)، والتي تقوم بتحلل المانانات عبر آلية عامة للتبادل المزدوج للحمض والقاعدة.قمنا بإجراء طفرات نقطية على الموقعين الحفازين لبكتيريا B. uniformis GH26 (E415A و E504A) ووجدنا أن طفرات أي من بقايا الجلوتاميك تؤدي إلى فقدان نشاط الإنزيم، مما يشير إلى أن كلا الموقعين الحفازين ضروريان لنشاط الإنزيم (الشكل 6B، والجدول التكميلي 17).
للتحقيق في الأساس الهيكلي لتخصص الركيزة المختلفة، قمنا بإجراء محاكاة ربط جزيئي لـ 5 ركيزات محددة هيكليًا (1 ركيزة تفاعلية و 4 ركيزات غير تفاعلية) باستخدام AutoDock Vina (الإصدار 1.2.0).توقعنا تفاعلات الربط الخاصة بهم وقمنا بترتيب أوضاع الربط لمجمعات الإنزيم-الركيزة باستخدام CSM-الكربوهيدراتكانت الطاقة الحرة المرتبطة بالتفاعل للركائز غير التفاعلية أعلى قليلاً فقط من تلك الخاصة بالغالكتوزيل مانوبيوز التفاعلي، وكان للترهالوز غير المحفز حتى طاقة ربط حرة أقل. وهذا يشير إلى أن خصوصية الركيزة لا يمكن تفسيرها فقط من خلال مقارنة الطاقات الحرة للربط للركائز. ومن الجدير بالذكر أن بقايا التحفيز وُجدت في بقايا التفاعل لمجمعات الإنزيم-الركيزة التفاعلية، ولكن لم توجد في تلك الخاصة بمجمعات الإنزيم-الركيزة غير التفاعلية، مما يشير إلى أن تفاعل الركيزة يعتمد على قدرتها على التفاعل مع بقايا التحفيز (الشكل 6C، D والشكل التوضيحي 14A-C).
من بين الستة مانان التفاعلي، أظهر GH 26 من B. uniformis DA183 أعلى نشاط تجاه الجلوكو مانان (مثل مانان الكونجاك) (الشكل 6B)، بينما كان GH26 من B. ovatus ATCC8483 لديه أعلى نشاط تجاه الجالاكتومانان (الشكل التوضيحي 15A). للتحقيق في دور مواقع ربط الركيزة المختلفة في نشاط GH26 من B. uniformis DA183 و B. ovatus ATCC8483، حددنا الموقع الفريد المختلف (A363 في B. uniformis DA183 و K149 في B. ovatus ATCC8483) من خلال المحاذاة الهيكلية وقمنا بتبادل البقايا. من المثير للاهتمام أن الطفرة K149A في GH26 BoATCC8483 أظهرت نشاطًا منخفضًا بشكل ملحوظ تجاه جالاكتومانان صمغ الغوار وأصبحت الأكثر نشاطًا تجاه جلوكو مانان الكونجاك (الشكل التوضيحي 15C). بشكل غير متوقع، أظهر النشاط الإنزيمي للطفرة A363K في GH26 BuDA183 زيادة طفيفة ولم ينتج عنها تغيير كبير في خصوصية الركيزة، على عكس الطفرة K149A في GH26 BoATCC8483، مما يشير إلى أن اختلافات غير متوقعة أخرى في مواقع ربط الركيزة تساهم في تفضيلات الركيزة المميزة للإنزيمين (الشكل 6G، والجدول التوضيحي 17).
لتقييم دور مجال CBM في GH26 BuDA183، قمنا أولاً بمقارنة النشاط الإنزيمي لنوع البرية الكامل و GH26 المقطوع من CBM. ومن المثير للاهتمام أنه لم يتم ملاحظة أي فرق كبير في نشاط الإنزيم بين GH26 نوع البرية (Km=3.75 ± 1.44 ملغ/مل، Kcatوتم تقصير CBM GH26ككات ) (الشكل التكميلي 13A-B، الشكل التكميلي 16A-B، والجدول التكميلي 18). من الجدير بالذكر أن بعض الدراسات أظهرت أن مجال CBM يوفر حماية حرارية لبعض الإنزيمات المحللة للكربوهيدرات.من خلال مقارنة أنشطة الإنزيمات لنوع GH26 البري و GH26 المقطوع من CBM عند درجات حرارة مختلفة، وجدنا أن GH26 المقطوع من CBM أصبح أقل استقرارًا حراريًا. على وجه التحديد، كانت قيم Tm هيلنوع البرية GH26 وللنمط المختصر من CBM (الشكل التكميلي 16C، والجدول التكميلي 19). وهذا يشير إلى أن الوظيفة المحتملة لمجال CBM من GH26 مرتبطة بالاستقرار الحراري.
الشكل 5 | استخدام المانان المستمد من النباتات بواسطة إنزيمات Bacteroides GH26. شجرة تطورية لتسلسلات بروتين GH26، بما في ذلك 89 GH26 موصوفة.. ب. يونيورميس GH26 في هذه الدراسة. أسماء الكائنات تشير إلى مستوى الأنواع، بجانب رقم الوصول إلى GenBank لبروتين GH33 الذي تم تحليله. تم إنشاء الشجرة باستخدام طريقة الاحتمال الأقصى. تم تلوين علامة الشجرة حسب تصنيف EC. تم تمييز ب. يونيورميس GH26 في هذه الدراسة بنجمة حمراء ذات خمس نقاط. تم تمييز GH26 المتبلور من ب. أوفاتوس بنجمة رمادية ذات خمس نقاط. ب. تُظهر النقطة المخطط كثافة نمو ب. أوفاتوس ATCC8483 (باللون الأزرق)، ب. يونيورميس DA183 (باللون الرمادي)، ب. يونيورميس ATCC8492 (باللون الأحمر)، سلالة الطفرة من ب. يونيورميس ATCC8492 المحذوف منها PUL34S (باللون الأخضر) وسلالة الطفرة الأخرى من ب. يونيورميس ATCC8492 المحذوف منها PUL34S (باللون البرتقالي) التي تستخدم 6 مصادر كربون، مثل الجلوكوز، بوليسكاريد دندروبيوم أوفيكينايل (DPs)، غلوكومانان كونجاك، صمغ الغوار. الجالاكتومانان، جالاكتومانان الخروب، ومانان جوز العاج. تُعرض البيانات كقيم متوسطةتم إجراء التحاليل في ثلاث نسخ فنية ). هيكل مجال C لبكتيريا ب. يونيكوميس GH26. D تم التنبؤ بالهيكل العام لبكتيريا ب. يونيكوميس GH26 بواسطة AlphaFold2 استنادًا إلى تسلسل البروتين. تم تلوين المجال الشبيه بـ Ig باللون الأخضر، ومجال GH26 باللون الأحمر، والليكتين المرتبط بالغالاكتوز باللون الأرجواني، على التوالي. E تم تراكب هيكل مجال ب. يونيكوميس GH26 (الأحمر) على هيكل ب. أوفاتوس GH26 (الرمادي) (رمز PDB 6HF4) (درجة TMRMSDالمواقع النشطة (E415 و E504)، مواقع ربط الكالسيوم (L198 و S201)، ومواقع ربط الركيزة (V170 و A200 و W207 و Y342 و W527 و Y528) من B. uniformis GH26 (نماذج عصا حمراء)، والمواقع النشطة (E201 و E291)، مواقع ربط الكالسيوم (L105 و S108)، ومواقع ربط الركيزة (V77 و A107 و W112 و Y148 و W314 و Y315) من B. ovatus GH26 (نماذج عصا رمادية) محفوظة.
الشكل 6 | النشاط الإنزيمي والخصوصية لإنزيم B. uniformis GH26. A يوضح الرسم البياني العمودي المانوز الناتج من الطرف المختزل الذي تم إنتاجه بواسطة GH26 BuDA183 (سماوي) و GH26 BoATCC8483 (رمادي) باستخدام 13 ركيزة مانان. كانت تركيز الإنزيمتركيز الركيزة كاندرجة حرارة التفاعل كانت، وكان وقت التفاعل 30 دقيقة. الطرف المختزل هو نهاية سلسلة الكربوهيدرات التي تحتوي على مجموعة ألدهايد أو كيتون حرة قادرة على اختزال أيونات النحاس في اختبار DNS. يتم تقديم البيانات كقيم متوسطة ± الانحراف المعياري. تم إجراء الاختبارات في ثلاث نسخ فنية ( ). B-D يُظهر مخطط النقاط إنتاج النهاية المختزلة مع مرور الوقت لستة ركائز تم تحللها بواسطة GH26 BuDA183 (النوع البري)، GH26 BuDA183 E415A، وE504A (طفرة في موقع التحفيز). الركائز هي: غلوكان مانان الكونجاك (أصفر)، بوليسكاريدات دندروبيوم أوفيكانال. (أحمر)، جالاكتوسيل مانابيوز (أخضر)، جالاكتومانان الخروب (أزرق)، جالاكتومانان الغوار (وردي)، ومانان جوز العاج (بنفسجي). كانت تركيز الإنزيم 125 نانومتر، وتركيز الركيزة كاندرجة حرارة التفاعل كانتوزمن الاستجابة كانتُعرض البيانات كقيم متوسطةتم إجراء الاختبارات في ثلاث نسخ فنيةتفاعلات E-F لـ GH26 BuDA183 (النوع البري) مع الجالاكتوسيل مانابيوز، و1-كيستوز يتم تصورها في 3D (يسار) و2D (يمين). يتم عرض الركائز كعصي أو دوائر خضراء، والمواد المتفاعلة كعصي أو دوائر رمادية، والمواقع الحفازة كعصي أو دوائر زرقاء. يتم عرض قوى التفاعل كخطوط منقطة. G نظام التفاعل هو نفسه كما في اللوحة (B-D)، باستثناء أن الإنزيم هو GH26 BuDA183 A363K (تبادلات مختلفة في المواد المتفاعلة).
نقاش
تلاعب الميكروبيوم المعوي بالبوليسكاريدات هو استراتيجية علاجية واعدة لمجموعة متنوعة من الأمراض البشرية.فهم كيفية استخدام بكتيريا الأمعاء للبوليسكاريدات الطبية سيوجه اختيار البوليسكاريدات لتعديل الميكروبيوم. ومع ذلك، فإن المعرفة في هذا المجال لا تزال محدودة. لمعالجة هذه الفجوة، قمنا برسم خرائط منهجية لملفات استخدام 20 نوعًا مختلفًا من البوليسكاريدات الطبية بواسطة 28 نوعًا من بكتيرويدس وبارابكتيرويدس البشرية. وجدنا أن الأنواع المختلفة من بكتيرويدس وبارابكتيرويدس أظهرت ملفات استخدام متميزة للبوليسكاريدات الطبية، والتي كانت مرتبطة بالتنوع الجينومي في الإنزيمات النشطة للكربوهيدرات. من خلال التحليل المقارن للتعبير الجيني والتلاعب الجيني، تحققنا من أن موضع استخدام البوليسكاريد PUL34_Bu مكن B. uniformis من استخدام بوليسكاريدات دندروبيوم.
اكتشفنا أن إنزيم GH26 في B. uniformis و B. ovatus، الذي تم تصنيفه كمانوزيداز، كان ضروريًا لاستخدام عدة مانانات مشتقة من النباتات (مثل بوليسكاريدات دندروبيوم). بشكل عام، توفر دراستنا إطارًا عامًا لتوصيف استخدام البوليسكاريدات المشتقة من النباتات بواسطة بكتيريا الأمعاء البشرية وفهم الآليات الجزيئية الأساسية. كما أن الاستخدام المحدد للغاية لبعض البوليسكاريدات الطبية (مثل بوليسكاريدات دندروبيوم) يبرز أيضًا إمكانيات البروبيوتيك والأدوية المعتمدة على الجليكوز لتعديل ميكروبيوم الأمعاء البشرية بشكل مستهدف. ب. يونيكوميس هو عضو بارز في ميكروبيوتا الأمعاء البشرية ويلعب دورًا حاسمًا في الحفاظ على صحة الأمعاء. على وجه التحديد، سلالة CECT 7771 تحسن من الخلل الأيضي والمناعي في الفئران التي تعاني من السمنة الناتجة عن نظام غذائي عالي الدهون.ويعزز الفوائد الأيضية والمناعية عند دمجه مع الألياف في الفئران البدينة.
بالإضافة إلى ذلك، B. uniformis وركيزته المفضلة،-سيكلوديكسترين، يحسن أداء تمارين التحمل في كل من الفئران والذكور البشرالسلالة JCM5828 تنظم مسارات إشارات NF-кB و MAPK في القولون من خلال تثبيط إشارات IL-17، مما يحسن من تطور التهاب القولون.السلالة F18-22 تخفف الالتهاب المعوي من خلال تحسين اختلال ميكروبات الأمعاءعلاوة على ذلك، تنتج بكتيريا B. uniformis حمض الكوليك 3-سوكينيلات (3-sucCA)، الذي يخفف من التهاب الكبد الدهني المرتبط بالخلل الأيضي (MASH).إن تعديل بكتيريا B. uniformis لتعزيز خصائصها البروبيوتيك يحمل وعدًا كبيرًا للتطبيقات العلاجية. من خلال تعديل B. uniformis لتفكيك السكريات المتعددة المحددة بشكل أكثر كفاءة أو إنتاج مستقلبات مفيدة، مثل 3-sucCA، يمكننا تحسين قدرتها على الاستعمار في الأمعاء وتحقيق آثار صحية إيجابية. يمكن أيضًا تصميم سلالات معدلة لتوصيل الجزيئات العلاجية مباشرة إلى الأمعاء، مما يوفر نهجًا مستهدفًا لعلاج الاضطرابات المعوية.
تعكس اختلافات ملفات نمو البولي سكاريد اختلافات في هيكل البولي سكاريد، مما يشير إلى إمكانية تطوير علامات مراقبة الجودة للبولي سكاريد الطبي من خلال مقارنة ملفات النمو. تعتبر مراقبة الجودة في تصنيع أدوية البولي سكاريد تحديًا بسبب تعقيدها وتنوعها الكيميائي واختلافات الدفعات.تم استخدام طرق حالية لتوصيف البوليسكاريدات الطبية، مثل 1) قياس محتوى السكر بناءً على قياس اللون، 2) تحديد الوزن الجزيئي والتوزيع بناءً على الكروماتوغرافيا السائلة عالية الأداء (HPLC)، و3) قياس تركيب السكريات الأحادية بناءً على HPLC. ومع ذلك، فإن هذه الطرق لا تلتقط الهيكل عالي الأبعاد للبوليسكاريدات، والذي يرتبط ارتباطًا وثيقًا بأنشطتها الحيوية. يمكن أن توفر ملفات نمو البوليسكاريدات رؤى قيمة حول الخصائص الهيكلية لها.
في هذه الدراسة، لوحظ أن البوليسكاريد يمكن أن ينظم بشكل متزامن العديد من PULs، كما تم ملاحظة تنظيم جزئي للجينات في PULs الأخرى، مشابهًا لاستجابة B. dorei لسكريات الحليب البشري.. هذه الظواهر قد تكون ناتجة عن التكرار الوظيفي لـ PULs. على سبيل المثال، كل من PUL72 و PUL16 في. الثيتايوتاوميكرون متورطون في معالجة مانوز عاليالجليكوزيداتأظهر إلغاء ستة من PULs التي تم تنظيمها بواسطة تدخل البكتين أنه لم يكن أي منها ضروريًا تمامًا للتفكك الكامل للتحضير المحدد للبكتين.. علاوة على ذلك، يعبر B. thetaiotaomicron عن عدة PULs للاستجابة لمخاط المضيف-الجليكوز، مما يضمن بقائها وعمليات الأيض حتى عندما تكون بعض PULs غير نشطة؛ على سبيل المثال، حذف ECF-الجين BT1053 أو الإدخال في الجين المشابه لـ susC BT1042 لم يؤثر بشكل كبير على النمو في الظروف المحايدة-جليكوزيدات.
من الجدير بالذكر أن عدة سلالات من B. uniformis، بما في ذلك B. uniformis DA183، تحتوي على إنزيمين GH26 في جينوماتها، يقعان في PULs مختلفة. قد تكون هذه الإنزيمات قد تطورت لاستهداف ركائز مختلفة أو للعمل تحت ظروف مختلفة. على سبيل المثال، في B. ovatus ATCC8483، يقوم PUL بترميز إنزيمين GH26.-ماناناز، BoMan26A و BoMan26B، بأنشطة مميزة: BoMan26A تشكل بشكل أساسي مانوبيوز، بينما BoMan26B، التي تكون مكشوفة على السطح، تقوم بتحلل الجالاكتومانان إلى خليط متنوع من الأوليغوسكريدات.تضمن هذه الازدواجية الوظيفية والتخصص الانهيار الفعال واستخدام الجالاكتومانان المعقد.
يجب أن تستكشف الأبحاث المستقبلية اختلافات مستويات السلالات في استخدام الجليكوز، كما هو الحال مع B. ovatus باستخدام الميوسين.مع انخفاض تكلفة التسلسل، يتم اعتبار المجتمعات الاصطناعية (SynComs) لتوصيف ملف النمو بسبب تركيباتها المحددة جيدًا وقابليتها للتكرار.كما يتضح في هذه الدراسة، يمكن أن تعكس الاختلافات في ملفات النمو وتوزيع إنزيمات CAZyme اختلافات في هياكل البوليسكاريد. لذلك، قد تساعد ملفات النمو الجديدة في تحديد البوليسكاريدات ذات الهياكل الجديدة المحتملة. بالإضافة إلى ذلك، فإن تحليل البروتينات التي تقوم بتفكيك هذه البوليسكاريدات يوفر رؤى قيمة حول هياكل سلاسل السكر المعقدة الخاصة بها.قد تحسن التقدم في التنبؤ بوظائف CAZyme فهمنا لهياكل البوليسكاريد الطبية من خلال تحليل ترانسكريبتوم PUL. أخيرًا، يمكن أن تخدم PULs المحددة كعلامات للهندسة الوراثية لبكتيريا الأمعاء (مثل PULs لاستخدام الإينولين )، وأدوات لتعزيز استعمار البروبيوتيك (مثل PULs لاستخدام بوليسكاريد البورفيرين ).
طرق
جمع وحفظ عينات البراز البشري
في هذه الدراسة، قدم جميع المشاركين البالغ عددهم 119 شخصًا موافقة مستنيرة، مع موافقة من معهد شنتشن للتكنولوجيا المتقدمة، الأكاديمية الصينية للعلوم (SIAT-IRB-200315-H0438). تم نقل عينات البراز المجمعة حديثًا من المتبرعين الأصحاء بسرعة إلى غرفة لاهوائية (غرف لاهوائية من الفينيل، كويلاب، الولايات المتحدة الأمريكية). تم إعادة تعليق كل عينة (3 جرام) في 15 مل منغليسرولفي محلول ملحي معزول مع فوسفات، مع-هيدروكلوريد السيستين)، تم خلطه باستخدام جهاز الخلط، وتم تصفيته من خلال شبكة نايلون معقمة لإزالة الجسيمات الكبيرة. تم تخزين كميات من المعلق المعالج في قوارير تجميد معقمة عندللحفظ على المدى الطويل، جاهز للعزل اللاحق لبكتيرويدس الأمعاء وبارابكتيرويدس.
عزل وزراعة بكتيريا بكتيرويدس وبارابكتيرويدس من الأمعاء
تم شراء بكتيرويدس يونيورميس ATCC 8492 وبكتيرويدس أوفاتوس ATCC 8483 من ATCC. تم عزل 28 نوعًا من بكتيرويدس وبارابكتيرويدس من أمعاء الإنسان المستخدمة في هذه الدراسة من عينات براز مجموعة SIAT. تم إذابة عينات البراز منتم تخفيفها ونشرها على أطباق أجار الدهون الحمضية من الخميرة المعدلة (mYCFA) (الجدول التكميلي 12). تم حضانة الأطباق في ظروف لاهوائية عندلـأيام في جو من، و . تم اختيار مستعمرات فردية، وتمت زراعتها على أطباق جديدة، وتمت حضانتها لاحقًا في ظروف لا هوائية عند لمدة أخرى أيام. تم تكرار هذه العملية لتنقية السلالات للحصول على سلالات نقية. ثم تم نقل السلالات المعزولة إلى وسط سائل وتمت حضانتها عند لمدة يومين. ثم قمنا بتكبير وتسلسل الجينات الكاملة 16S rRNA باستخدام زوج من بادئات PCR (27 F 5′-AGAGTTTGATCMTGGCTCAG-3’؛ 1492 R 5′-GGTTACCTTGTTACGACTT-3′). السلالات التي أنتجت قمم مزدوجة في تسلسلاتها تم التخلص منها أو خضعت لجولة أخرى من التنقية. تم تخزين جميع السلالات المنقاة عند في تعليق الجليسرول الذي يحتوي على السيستين. تم تحديد التصنيف الضريبي للعزلات من خلال مقارنة تسلسلات الجينات الكاملة 16S rRNA الخاصة بها بتلك الموجودة في قاعدة بيانات تصنيف الجينوم (إصدار GTDB 207) (الجدول التكميلي S1A). لأغراض الزراعة، تم زراعة أنواع البكتيرويد في وسط مغذي من مغذيات القلب والدماغ (BHIS) معزز بـ الهيمين، فيتامين K3، و-السيستين (الجدول التكميلي 12). تمت الزراعة في غرفة لا هوائية مع تركيبة غازية من ، و، و، maintained at a temperature of
.
اختبار الوسط الأدنىلتحديد وسط أدنى مناسب لنمو أنواع البكتيرويد، اختبرنا ثلاثة أوساط أدنى: وسط بكتيرويد المعدل (mBMM) ، وسط الأحماض الدهنية المعدل من الخميرة كاسيتون (mYCFA) ، ووسط M9 المعدل (mM9) ، جميعها معززة بـ الجلوكوز (الجدول التكميلي 12). باستخدام محطة عمل لا هوائية مع خليط غازي من ، و
، تم توزيع الوسط الأساسي المعدل في طبق 96 بئر. تم غسل الثقافات البكتيرية مرتين بمحلول PBS لا هوائي، وتم ضبط الكثافة الضوئية عند 600 نانومتر (OD600) لمحلول البكتيريا إلى 0.1. أشارت منحنيات النمو إلى أن كل من mBMM وmYCFA دعمت عمومًا نمو أنواع البكتيرويد (الشكل التكميلي 1). مقارنةً بـ mYCFA، قدم mBMM، كونه وسطًا صناعيًا بتركيب واضح، قياسات أكثر استقرارًا لاستخدام البوليسكاريد.
انتشار ووفرة أنواع البكتيرويد تم حساب وفرة وانتشار السلالات المختارة في دراستنا. قمنا بفحص 16282 جينوم بشري صحي من بيانات الميتاجينوم المتاحة للجمهور من خلال البحث باستخدام فلتر محدد
الظروف (experiment_type = ‘Metagenomics’ AND QCStatus = ‘Good runs’ AND host age AND Country=is not null AND Recent Antibiotics Use = ‘No’ AND Phenotype = ‘Health’) في GMrepo لتمثيل الميكروبيوم المعوي البشري الصحي العالمي. تم تعريف الأنواع ذات تكرار الحدوث المتوسط المتساوي > ووفرة نسبية متوسطة
كأنواع أساسية.
قياس ملفات نمو البكتيرياتم إضافة وسط mBMM إلى طبق 96 بئر في محطة عمل لا هوائية ، و. تم تعزيز الوسط بـ (تحكم سلبي)، الجلوكوز (تحكم إيجابي)، أو البوليسكاريدات الطبية (علاج). بعد ذلك، تم غسل البكتيريا مرتين بمحلول PBS لا هوائي، وتم ضبط OD600 لمحلول البكتيريا إلى 0.1. ثم تم تلقيح البكتيريا بنسبة 1:10 في طبق 96 بئر وزراعتها عند
. تم قياس OD النمو وتسجيله كل 12 ساعة باستخدام قارئ الميكرو بلايت.
استخراج البوليسكاريدات الطبيةتم شراء سبعة عشر عشبًا من مدينة بوزهو (أنhui، الصين) وتم التحقق منها وتحديدها بواسطة أحد المؤلفين، الدكتور جي يانغ. تم إيداع عينات التحقق في كلية علوم الصحة (FHS)، جامعة ماكاو. لمزيد من المعلومات التفصيلية حول الأعشاب، انظر الجدول التكميلي 3A. من بين طرق الاستخراج المختلفة (مثل، الماء الساخن/الإيثانول، القلوي، الإنزيمي)، يعتبر استخراج الماء الساخن الأكثر استخدامًا بسبب سهولة تشغيله، وذوبانه العالي في الماء الساخن، والحد الأدنى من الضرر للبوليسكاريدات . تم إجراء استخراج البوليسكاريدات عن طريق نقع القطع الطبية في 10 أضعاف حجمها من الماء المقطر لمدة 30 دقيقة، تليها الغليان على حرارة عالية والتسخين على حرارة منخفضة لمدة 30 دقيقة. ثم تم تصفية السائل الساخن من خلال قماش شبكي 200، مع التخلص من الرواسب. تم تكرار هذه العملية مع 10 أضعاف حجم الماء المقطر، والغليان على حرارة عالية والتسخين على حرارة منخفضة لمدة 25 دقيقة، وتصفيته مرة أخرى أثناء كونه ساخنًا. تم دمج كل من المصفاة وتقليلها إلى درجة حرارة منخفضة باستخدام ضغط الفراغ. بعد التبريد، تمت إضافة ثلاثة أضعاف حجم الإيثانول اللامائي لترسيب البوليسكاريدات مع التحريك المستمر، وتم ترك الخليط ليقعد طوال الليل عند . تم فصل الراسب عن السائل باستخدام فلتر مضخة فراغ، وغسله ثلاث مرات بالإيثانول اللامائي، وتجفيفه بالفراغ عند
للحصول على المنتج النهائي. بالنسبة للبوليسكاريدات الطبية_2 (مثل، DPs_2)، كانت عملية الاستخراج مشابهة لتلك الخاصة بالبوليسكاريدات الطبية_1 (مثل، DPs_1)، مع الخطوة الإضافية للغسيل لإزالة الجزيئات الأصغر من 3.5 كيلو دالتون.
تحديد محتوى السكر الكليتم قياس محتوى السكر الكلي باستخدام البروتوكول كما يلي. على وجه التحديد، تم استخدام الجلوكوز (Mw 180 Da) المستخرج من سيغما (سانت لويس، ميزوري، الولايات المتحدة الأمريكية) لإنشاء منحنى المعايرة. تم إعداد محلول قياسي من الجلوكوز بتركيز . لكل عينة من البوليسكاريدات، تم إعداد محلول البوليسكاريدات الطبية بتركيز . تم تسخين الخليط لتسهيل الذوبان ثم تم تصفيته باستخدام عنصر فلتر حتى أصبح المحلول واضحًا. تم إضافة محلول الفينول إلى محلول الجلوكوز القياسي ومحلول البوليسكاريدات الطبية. تم خلط الخليط جيدًا لضمان الخلط المناسب وتركه لمدة 30 دقيقة. تم تسخين جهاز الطيف الضوئي عند طول موجي 490 نانومتر (باستخدام مصدر ضوء LED وعنصر حساس للضوء CMOS). عادةً ما تكتمل عملية التسخين بحلول نهاية التفاعل، وتصبح قراءة المقياس مستقرة. تم إجراء المعايرة عند نقطة الصفر باستخدام محلول الفينول بدون إضافة الجلوكوز، والمعايرة عند نقطة واحدة مع حاجز أسود (الجدول التكميلي S3). ثم، تم قياس معدل الامتصاص لكل عينة ثلاث مرات. تم حساب المنحنى القياسي باستخدام الانحدار الخطي مع بيانات عينة الجلوكوز القياسية المقاسة أعلاه. تم حساب إجمالي
محتوى السكر عن طريق استبدال معدل الامتصاص للبوليسكاريدات الطبية. في هذه التجربة، مع تركيز عينة من ، يمكن حساب محتوى السكر لعينة باستخدام الصيغة:
عندما يكون تركيز السكر هو ، يكون معدل الامتصاص هو ، المنحنى القياسي هو ، وتركيز العينة هو في هذه التجربة).
تحديد الوزن الجزيئي والتوزيع
تم قياس الوزن الجزيئي والتوزيع وفقًا للطريقة كما يلي. على وجه التحديد، تم استخدام معايير الدكستران بأوزان جزيئية مختلفة ( ) المستخرجة من سيغما (سانت لويس، ميزوري، الولايات المتحدة الأمريكية) لإنشاء منحنى المعايرة. بناءً على مبدأ كروماتوغرافيا العمود الهلامي، هناك علاقة أسية بين الوزن الجزيئي ووقت الاحتفاظ لمركب. تم إنشاء نموذج رياضي بالتعبير لتمثيل هذه العلاقة:
في هذه الدراسة، تم تحليل توزيع الوزن الجزيئي لعينات البوليسكاريد باستخدام نظام HPLC مع المعلمات التالية: الطور المتحرك: محلول NaCl ، معدل التدفق: ، كمية العينة المفردة: ، تركيز محلول العينة: ، العمود الكروماتوغرافي: TSK-gel G3000PWXL ( ) العمود، درجة حرارة العمود: ، الكاشف: كاشف مؤشر الانكسار (RI). تم إذابة عينات السكر القياسية ذات الأوزان الجزيئية المعروفة وعينات البوليسكاريدات الطبية في الماء بتركيز نهائي من ، وتم تصفيتها باستخدام عنصر فلتر حتى تصبح واضحة. تم تسجيل وقت الاحتفاظ لكل عينة. تم حساب المنحنى القياسي باستخدام الانحدار الأسّي مع بيانات العينات القياسية. من خلال استبدال وقت الاحتفاظ للعينات البوليسكاريد الطبية، تم حساب الوزن الجزيئي.
تحديد تركيب السكريات الأحادية
تم قياس تركيب السكريات الأحادية وفقًا للطريقة ، كما هو موضح أدناه. على وجه التحديد، تم تحليل التركيب الأحادي للسكريات لعينة البوليسكاريدات باستخدام HPLC مع المعلمات التالية: الطور المتحرك: أسيتونيتريل مضاف إليهمحلول عازل الفوسفات، معدل التدفق:مقدار العينة الواحدة:عمود الكروماتوغرافيا: كوزموسيل 5C18-PAQ )، درجة الحرارة في العمود: كاشف: كاشف الأشعة فوق البنفسجية (UV). تم خلط كل مونوسكاريد (مانوز، جلوكوز، جالاكتوز، رامنوز، فوكوز، زيلوز، أرايبينوز، حمض الجالاكتورونيك، حمض الجلوكورونيك) المستخرج من سيغما (سانت لويس، ميزوري، الولايات المتحدة الأمريكية) بتركيز 10 مللي مول. قمنا بإعداد سلسلة من حلول المونوسكاريد القياسية وقمنا بمعايرة نظام الكروماتوغرافيا السائلة باستخدام هذه المعايير. تم تحلل عينات البوليسكاريد الطبية بالكامل وأجريت عملية تحويل مسبق للعمود باستخدام 1-فينيل-3-ميثيل-5-بيرازولينون. تم مقارنة زمن الاحتفاظ مع عينات المونوسكاريد القياسية لتحديد أنواع المونوسكاريد الموجودة في عينة البوليسكاريد. من خلال قياس نسبة مساحة الذروة لكل مونوسكاريد، تم حساب نسبة كل مونوسكاريد في عينة البوليسكاريد.
تسلسل وتحليل النسخ الجيني
تم زراعة أنواع البكتيرويد في وسط BHIS المدعوم بـهيمينفيتامين K3، وسيستين في غرفة لاهوائية (كوي؛، و ) في تم غسل البكتيريا ثلاث مرات بمحلول PBS اللاهوائي، وتم قياس الكثافة الضوئية عند
تم ضبط 600 نانومتر (OD600) من محلول البكتيريا إلى 0.1. ثم تم تلقيح البكتيريا بنسبة 1:10 في لوحة 96 بئر وزرعها في. تم قياس OD600 وتسجيله كل 15 دقيقة باستخدام قارئ الميكرو بلايت. بعد الوصول إلى مرحلة النمو اللوغاريتمي (الشكل التوضيحي 6A-C)، تم جمع البكتيريا عن طريق الطرد المركزي عند 5000 × ج لمدة دقيقتين. تم تخزين البكتيريا المجمعة في فترة النمو اللوغاريتمي في قبل تسلسل النسخ.
تم استخراج RNA الكلي من العينة باستخدام طريقة Trizol، بما في ذلك معالجة DNase لإزالة الحمض النووي الجيني، وتم تقييم الجودة باستخدام جهاز NanoDrop One (Thermo, A30221) وجهاز Agilent 4200 TapeStation (Agilent, G2991AA). تم إزالة RNA الريبوسومي باستخدام مجموعة إزالة RNA Ribo-Zero (Epicenter, 15066012) المصممة للبكتيريا. لبناء المكتبة، تم استخدام مجموعة NEBNext Ultra II Directional RNA Library Prep Kit (NEB, e7760) الخاصة بـ Illumina: تم تجزئة mRNA إلى قطع قصيرة، وتم تخليق cDNA أحادي السلسلة باستخدام سداسيات عشوائية، وتم توليد cDNA مزدوج السلسلة مع استبدال dUTP، وتم تنقيته باستخدام كريات AMPure XP (Beckman, A63880)، وتم هضم السلسلة الثانية المحتوية على U باستخدام إنزيم USER (NEB, M5508). تم إجراء إصلاح النهاية، وإضافة ذيل A، وربط المحولات على cDNA المنقى، تلا ذلك اختيار حجم القطع باستخدام كريات AMPure XP. تم إجراء تضخيم PCR، وتم تنقية منتجات PCR باستخدام كريات AMPure XP للحصول على المكتبة النهائية. تم فحص المكتبات باستخدام جهاز Agilent 4200 TapeStation، وتم تسلسل المكتبات المؤهلة فقط على منصة Illumina (Illumina, HiSeq 2000). شملت التحسينات الرئيسية ضمان معالجة DNase أثناء استخراج RNA، واستخدام مجموعة إزالة RNA المحددة للبكتيريا، وتوليد مكتبات محددة السلسلة، والتحقق من توافق المحولات مع RNA بدائيات النوى، وضبط اختيار حجم القطع لطول النسخ بدائيات النوى.
بعد الحصول على بيانات التسلسل لكل عينة، من المهم تقييم جودة البيانات وإزالة أي قراءات منخفضة الجودة لضمان موثوقية التحليلات اللاحقة. البرنامج المستخدم في مراقبة الجودة هو fastp.، وتطبق معايير مراقبة الجودة التالية: 1) الحد الأدنى لدرجة جودة القاعدة 15؛ 2) حجم نافذة الانزلاق 4 مع متوسط درجة جودة القاعدة أكبر من 20؛ 3) الحد الأدنى لطول القراءة 75 نقطة أساسية؛ 4) القيم الافتراضية للمعلمات الأخرى. يتم محاذاة بيانات التسلسل، بعد مراقبة الجودة، إلى تسلسلات الرنا الريبوسومي في قاعدة بيانات Rfam باستخدام Bowtie2.لإزالة قراءات الرنا الريبوسومي. ثم يتم محاذاة القراءات المتبقية إلى الجينوم المرجعي باستخدام Bowtie2، مما ينتج عنه ملف BAM يحتوي على نتائج المحاذاة. يتم معالجة نتائج المحاذاة بشكل إضافي لتوليد عدد القراءات باستخدام RSEM.لتحليل تعبير الجينات، نستخدم عدد القراءاتلتحديد الجينات المعبر عنها بشكل مختلف بين مجموعات العينات المتعددة.
التعليق الجينومي لموقع استخدام البوليسكاريد (PUL)
تم توضيح PULs من بكتيرويدس بواسطة PULpyمن الجدير بالذكر أن الأنواع المختلفة من B. uniformis يمكن أن تحتوي على أعداد وهياكل مختلفة من PULs، ويمكن ملاحظة هذه الاختلافات أيضًا بين طرق التوصيف المختلفة، مثل PULpy.وخوارزمية توقع PULتم استخدامه بواسطة PULDB. تم إعادة تصحيح حدود PUL بناءً على اختلافات التعبير الجيني. على وجه التحديد، من خلال استخدام تحليل النسخ المقارن لأنواع Bacteroides المزروعة في وسط غذائي محدود مضاف إليه السكريات المتعددة أو الجلوكوز، تم تحديد PULs التي كانت لديها أعلى زيادة في التعبير. ثم تم مقارنة النسخ الجينية للجينات الموجودة في أعلى وأسفل هذه PULs. تم تعريف الجينات التي كانت مرتفعة التعبير بشكل مستمر داخل وخارج PULs المتوقعة على أنها PULs المعاد تصحيحها.
بناء طفرات knockout لبكتيريا B. uniformis
نظرًا لصعوبة التلاعب الجيني في سلالات باكتيرويدز البرية، قمنا بإجراء حذف PUL34_Bu في السلالة النموذجية B. uniformis ATCC8492 (الشكل التوضيحي 8). تم زراعة المتلقي B. uniformis ATCC8492 للتزاوج في وسط BHIS في ظروف لاهوائية. غرفة (كوي؛، و ) في تم زراعة المتبرع Escherichia coli S17-1 (Apir) في وسط لوريا-بيرتاني (LB) في. تم استنساخ قطع الحمض النووي (الجدول التكميلي 13A) من sgRNA أو الأذرع المتجانسة لـ sgRNA (قوالب إصلاح بحجم 1 كيلوبايت) في هيكل البلازميد بواسطة مجموعة تركيب الحمض النووي NEBuilder HiFi (NEB، E2621S). تم تصنيع البرايمرات (الجدول التكميلي 13B) المستخدمة لتكبير هيكل البلازميد، sgRNA، قالب الإصلاح واكتشاف الطفرات بواسطة شركة سانغون للتكنولوجيا الحيوية. تم دمج بلازميدات الإزالة في B. uniformis ATCC8492. ثم تم تخفيف سلالات البكتيرويد في وسط BHI بنسبة 1:100 مع تحفيز aTc لمدة 24 ساعة (تركيز نهائي لـ aTcتم تخفيف الثقافات بواسطة و -طوي وانتشر على طبق بكتيريا BHI-aTc وتم حضنه فيلمدة 40 ساعة. تم تحديد المستعمرات بواسطة تفاعل البوليميراز المتسلسل والتسلسل.
محاذاة التسلسل والبنية
تم بناء شجرة النشوء والتطور للإنزيمات من عائلة GH26 باستخدام MUSLEوبنيت شجرة تطورية باستخدام طريقة الاحتمالية القصوىلـ 89 تسلسل بروتين GH26 المميز المودع في قاعدة بيانات CAZyتم توقع الهيكل ثلاثي الأبعاد لبروتين GH26 من سلالة B. uniformis DA183 باستخدام AlphaFold2.تم حساب قيمة اختبار فرق المسافة المحلية المتوقعة (pLDDT) باستخدام AlphaFold2. تم محاذاة المجال التحفيزي لبكتيريا B. uniformis DA183 GH26 مع جين GH26 المجاور في B. ovatus ATCC8483. (رقم الوصول في GenBank ALJ47537.1؛ رمز PDB 6HF4) باستخدام US-align لحساب درجة TM و RMSD.
تعبير وتنقية إنزيم باكتيرويدس GH26
تم تحسين تسلسلات الترميز للبروتينات Bacteroides GH26 كاملة الطول، والطفرات النقطية، والمختصرة CBM (الجدول التكميلي 14) للتعبير في E. coli، وتم تصنيعها كيميائيًا (شركة Tsingke Biotechnology Co., Ltd.)، وتم استنساخها في بلازميد pET28a مع نهاية C.علامة His بين مواقع Ndel و Xhol. تم تحويل البلازميدات إلى E. coli BL21(DE3). تم توقع حدود الببتيد الإشاري باستخدام SignalP (الإصدار 6.0).تمت إزالة ببتيد الإشارة لضمان التعبير القابل للذوبان. تم توقع حدود مجال CBM باستخدام Chainsaw.تم زراعة خلايا E. coli BL21(DE3) التي تحتوي على بلازميد pET28a في 500 مل من وسط LB معكاناميسين فيحتى OD600تم تحفيز التعبير عن طريق إضافة 0.2 مللي مول من IPTG، واستمرت الثقافة لمدة 20 ساعة عندتم تحلل الخلايا في 50 مل من محلول التحللإيميدازول، pH 7.4باستخدام كسارة خلايا عالية الضغط. تم طرد السائل الخلوي في جهاز الطرد المركزي عند، لمدة ساعة واحدة، وتم تحضين السائل العلوي مع 5 مل من راتنج نيكيل NTA (Yeasen, 20502ES50) لمدة ساعة واحدة عندمع دوران الرأس إلى الأسفل. تم صب الراتنج في عمود تدفق الجاذبية وغسله ثلاث مرات بـ 50 مل من محلول الغسيل.، إيميدازول، pH 7.4). تم استبعاد البروتين باستخدام محلول الإخراجإيميدازول، pH 7.4).
اختبار نشاط إنزيم باكتيرويدس GH26
غلوكان كونجاك، غار غم غالاكتومانان، غالاكتومانان الخروب، مانان جوز العاج، غلوكان الشعير، وزيغلوجان التمر كانت من ميغازايم (بري، أيرلندا). زيلان كوز الذرة، 1،1-كستوتيتراوز، و1-كستوز كانت من TCI. غالاكتوسيل مانوبيوز كانت من ACMEC؛ رافينوز كانت من HUICH. تريهالوز كانت من شركة هوي هوي (شنغهاي) للتكنولوجيا الحيوية المحدودة. تم قياس النشاط المحدد باستخدام اختبار السكر المختزل القياسي DNS.يستخدم GH26 BuDA183 و GH26 BoATCC8483 و المواد الأساسية (الجدول التكميلي 3C) في محلول فوسفات البوتاسيوم بتركيز 50 مللي مولار، pH 6.5. كانت مدة الحضانة 30 دقيقة عندتم قياس نشاط الإنزيم للطفرات النقطية و GH26 المقطوع CBM بنفس الطريقة المستخدمة في اختبار DNS القياسي باستخدام 125 نانومتر من إنزيم GH26 وستة ركائز تفاعلية (الشكل 6A) في محلول فوسفات البوتاسيوم بتركيز 50 مللي مولار، pH 6.5، عند. كانت فترة الحضانة 30 دقيقة عند .
تم استخدام المانوز للحصول على منحنى معيار التركيز (الجدول التكميلي 15).
حركية الإنزيمات
تم قياس حركيات ميكاليز-مينتن لـ 125 نانومتر من النوع البري الكامل، والمختصر CBM، والمتغيرات A 363 K في ثلاث تكرارات باستخدام اختبار نشاط DNS.لكن مع تغيير الوقت وتركيز الركيزة. كانت جميع التفاعلات بحجم إجمالي من واحتوى وكونجاك غلوكومانان (1.25، 2.5، 3.75، 5، 6.25، 7.5، 8.75، و ). تم إيقاف التفاعل بواسطة الحرارة لـ لمدة دقيقة واحدة. تم استخدام المعدلات الأولية التي تم الحصول عليها لإنشاء منحنيات ميكايليس-مينتن عبر النمذجة غير الخطية.دالة في R تم من خلالها تقدير قيم Km و Kcat.
الربط الجزيئي والتصورات
تم نمذجة الركائز الأوليغوساكاريدية باستخدام موقع GLYCAM-Web (مركز أبحاث الكربوهيدرات المعقدة، جامعة جورجيا، أثينا، جورجيا (http://www.glycam.com“)) أو مشتق من بنك بيانات البروتين RCSB (PDB). تم إجراء ربط الجزيئات باستخدام AutoDock Vina (الإصدار 1.2.0)تم استخدام معايير قياسية. تم اشتقاق الموقع النشط لـ GH26 من هيكل Bo GH26 (PDB، 6 HF 4)عن طريق التراكب الهيكلي مع محاذاة الولايات المتحدةتم تحليل تسع توافقيات مختلفة للرابطة لكل مركب ربط باستخدام CSM-carbohydrate، حيث تم اختيار التوافق الذي يمتلك أدنى قيمة دلتا-G (طاقة الربط) لدراسة تفاعلات الرابطة-المستقبل بواسطة برنامج Discovery Studio Visualizer..
الإحصائيات وإمكانية التكرار
لم يتم استخدام أي طريقة إحصائية لتحديد حجم العينة مسبقًا. لم يتم استبعاد أي بيانات من التحليلات. تم تحديد الدلالة الإحصائية لاختبارات ملف النمو في الشكل 1E والشكل التكميلي 3A باستخدام اختبار t غير المقترن ذو الجانبين بين كل مجموعة تجريبية والضبط السلبي، مع تعديل المقارنات المتعددة باستخدام طريقة بونفيروني؛ تم إجراء كل تجربة ثلاث مرات لضمان الموثوقية. بالنسبة لتحليل النسخ المقارن، تم تقييم التعبير التفاضلي باستخدام DESeq2 مع اختبار والد ذو الجانبين، وتم تعديله.تم حساب القيم باستخدام طريقة بنجاميني وهوشبرغ للتحكم في معدل الاكتشاف الخاطئ (FDR)؛ الجينات التي تم تعديلها-قيمةاعتُبرت ذات دلالة إحصائية. تُظهر الصور التمثيلية من التجارب الفردية تحليل هلام الحمض النووي الكهربائي و SDS-PAGE للبروتين، بينما تُعرض بيانات توصيف نشاط الإنزيم في المختبر كقيم متوسطة ± الانحراف المعياري، مع إجراء الاختبارات في ثلاث نسخ فنية. ) لضمان الموثوقية.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
تم تقديم جميع بيانات التسلسل التي تم إنشاؤها في هذه الدراسة إلى قاعدة بيانات ENA BioProject (https://www.ebi.ac.uk/ena/browser/search) تحت رقم الوصول PRJEB75723. تتوفر بيانات أخرى في المعلومات التكميلية والبيانات التكميلية. يتم توفير بيانات المصدر مع هذه الورقة.
References
Garrett, W. S. The gut microbiota and colon cancer. Science 364, 1133-1135 (2019).
Sun, L. et al. Bile salt hydrolase in non-enterotoxigenic Bacteroides potentiates colorectal cancer. Nat. Commun. 14, 755 (2023).
Huang, J. et al. Ginseng polysaccharides alter the gut microbiota and kynurenine/tryptophan ratio, potentiating the antitumour effect of antiprogrammed cell death 1/programmed cell death ligand 1 (anti-PD-1/PD-L1) immunotherapy. Gut 71, 734-745 (2022).
Tanoue, T. et al. A defined commensal consortium elicits CD8 T cells and anti-cancer immunity. Nature 565, 600-605 (2019).
Zafar, H. & Saier, M. H. Jr Gut Bacteroides species in health and disease. Gut Microbes 13, 1848158 (2021).
Beller, Z. W. et al. Inducible CRISPR-targeted “knockdown” of human gut Bacteroides in gnotobiotic mice discloses glycan utilization strategies. Proc. Natl Acad. Sci. 120, e2311422120 (2023).
Hehemann, J.-H. et al. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 464, 908-912 (2010).
Kijner, S., Ennis, D., Shmorak, S., Florentin, A. & Yassour, M. CRISPR-Cas-based identification of a sialylated human milk oligosaccharides utilization cluster in the infant gut commensal Bacteroides dorei. Nat. Commun. 15, 105 (2024).
Panwar, D., Shubhashini, A. & Kapoor, M. Complex alpha and beta mannan foraging by the human gut bacteria. Biotechnology Advances, 108166 (2023).
Buzun, E. et al. A bacterial sialidase mediates early-life colonization by a pioneering gut commensal. Cell Host Microbe 32, 181-190. e189 (2024).
Kou, F. et al. A review of Ganoderma lucidum polysaccharides: Health benefit, structure-activity relationship, modification, and nanoparticle encapsulation. International journal of biological macromolecules, 125199 (2023).
Lin, T.-L. et al. Role of gut microbiota in identification of novel TCMderived active metabolites. Protein Cell 12, 394-410 (2021).
Yu, Y., Shen, M., Song, Q. & Xie, J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydr. Polym. 183, 91-101 (2018).
Shan-Shan, S., Kai, W., Ke, M., Li, B. & Hong-Wei, L. An insoluble polysaccharide from the sclerotium of Poria cocos improves hyperglycemia, hyperlipidemia and hepatic steatosis in ob/ob mice via modulation of gut microbiota. Chin. J. Nat. Med. 17, 3-14 (2019).
Shi, L. Bioactivities, isolation and purification methods of polysaccharides from natural products: A review. Int. J. Biol. Macromolecules 92, 37-48 (2016).
Wu, T.-R. et al. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 68, 248-262 (2019).
Zhang, Y. et al. Identification of the core active structure of a Dendrobium officinale polysaccharide and its protective effect against dextran sulfate sodium-induced colitis via alleviating gut microbiota dysbiosis. Food Res. Int. 137, 109641 (2020).
Martens, E. C., Chiang, H. C. & Gordon, J. I. Mucosal glycan foraging enhances fitness and transmission of a saccharolytic human gut bacterial symbiont. Cell host microbe 4, 447-457 (2008).
Lapébie, P., Lombard, V., Drula, E., Terrapon, N. & Henrissat, B. Bacteroidetes use thousands of enzyme combinations to break down glycans. Nat. Commun. 10, 2043 (2019).
Martens, E. C. et al. Recognition and degradation of plant cell wall polysaccharides by two human gut symbionts. PLOS Biol. 9, e1001221 (2011).
Foley, M. H., Cockburn, D. W. & Koropatkin, N. M. The Sus operon: a model system for starch uptake by the human gut Bacteroidetes. Cell. Mol. Life Sci. 73, 2603-2617 (2016).
Luis, A. S. et al. A single sulfatase is required to access colonic mucin by a gut bacterium. Nature 598, 332-337 (2021).
Wu, S. et al. GMrepo: a database of curated and consistently annotated human gut metagenomes. Nucleic acids Res. 48, D545-D553 (2020).
Liu, C. et al. Enlightening the taxonomy darkness of human gut microbiomes with a cultured biobank. Microbiome 9, 119 (2021).
Drula, E. et al. The carbohydrate-active enzyme database: functions and literature. Nucleic acids Res. 50, D571-D577 (2022).
Qi, H. et al. Comparisons of isolation methods, structural features, and bioactivities of the polysaccharides from three common Panax species: A review of recent progress. Molecules 26, 4997 (2021).
Kuang, M.-T. et al. Structural characterization and hypoglycemic effect via stimulating glucagon-like peptide-1 secretion of two polysaccharides from Dendrobium officinale. Carbohydr. Polym.
241, 116326 (2020).
Yin, J.-Y. et al. Separation, structure characterization, conformation and immunomodulating effect of a hyperbranched heteroglycan from Radix Astragali. Carbohydr. Polym. 87, 667-675 (2012).
Wardman, J. F., Bains, R. K., Rahfeld, P. & Withers, S. G. Carbohydrate-active enzymes (CAZymes) in the gut microbiome. Nat. Rev. Microbiol. 20, 542-556 (2022).
Yin, Y. et al. dbCAN: a web resource for automated carbohydrateactive enzyme annotation. Nucleic acids Res. 40, W445-W451 (2012).
Stewart, R. D., Auffret, M. D., Roehe, R. & Watson, M. Open prediction of polysaccharide utilisation loci (PUL) in 5414 public Bacteroidetes genomes using PULpy. bioRxiv, 421024, https://doi.org/ 10.1101/421024 (2018).
Guo, M., Shao, S., Wang, D., Zhao, D. & Wang, M. Recent progress in polysaccharides from Panax ginseng CA Meyer. Food Funct. 12, 494-518 (2021).
Li, J. et al. Purification, structural characterization, and immunomodulatory activity of the polysaccharides from Ganoderma lucidum. Int. J. Biol. macromolecules 143, 806-813 (2020).
Zheng, L. et al. CRISPR/Cas-based genome editing for human gut commensal bacteroides species. ACS Synth. Biol. 11, 464-472 (2022).
García-Bayona, L. & Comstock, L. E. Streamlined genetic manipulation of diverse bacteroides and parabacteroides isolates from the human gut microbiota. MBio 10, 01762-01719 (2019).
Almeida, A. et al. A unified catalog of 204,938 reference genomes from the human gut microbiome. Nat. Biotechnol. 39, 105-114 (2021).
Buchfink, B., Reuter, K. & Drost, H.-G. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat. methods 18, 366-368 (2021).
Zhou, N. et al. Konjac glucomannan: a review of structure, physicochemical properties, and wound dressing applications. J. Appl. Polym. Sci. 139, 51780 (2022).
Mudgil, D., Barak, S. & Khatkar, B. S. Guar gum: processing, properties and food applications-a review. J. food Sci. Technol. 51, 409-418 (2014).
Lazaridou, A., Biliaderis, C. G. & Izydorczyk, M. S. Structural characteristics and rheological properties of locust bean galactomannans: a comparison of samples from different carob tree populations. J. Sci. Food Agriculture 81, 68-75 (2001).
Ghysels, S. et al. Fast pyrolysis of mannan-rich ivory nut (Phytelephas aequatorialis) to valuable biorefinery products. Chem. Eng. J. 373, 446-457 (2019).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583-589 (2021).
Bågenholm, V. et al. A surface-exposed GH26 -mannanase from Bacteroides ovatus: Structure, role, and phylogenetic analysis of BoMan26B. J. Biol. Chem. 294, 9100-9117 (2019).
Okano, H., Kanaya, E., Ozaki, M., Angkawidjaja, C. & Kanaya, S. Structure, activity, and stability of metagenome-derived glycoside hydrolase family 9 endoglucanase with an N-terminal Ig-like domain. Protein Sci. 24, 408-419 (2015).
Liberato, M. V. et al. Molecular characterization of a family 5 glycoside hydrolase suggests an induced-fit enzymatic mechanism. Sci. Rep. 6, 23473 (2016).
Murray, P. A., Kern, D. G. & Winkler, J. R. Identification of a galactosebinding lectin on Fusobacterium nucleatum FN-2. Infect. Immun. 56, 1314-1319 (1988).
Kuhaudomlarp, S., Gillon, E., Varrot, A. & Imberty, A. LecA (PA-IL): A galactose-binding lectin from Pseudomonas aeruginosa. Lectin Purification and Analysis: Methods and Protocols, 257-266 (2020).
Bolam, D. N. et al. Mannanase A from Pseudomonas fluorescens ssp. cellulosa is a retaining glycosyl hydrolase in which E212 and E320 are the putative catalytic residues. Biochemistry 35, 16195-16204 (1996).
Eberhardt, J., Santos-Martins, D., Tillack, A. F. & Forli, S. AutoDock Vina 1.2. O: New docking methods, expanded force field, and python bindings. J. Chem. Inf. modeling 61, 3891-3898 (2021).
Nguyen, T. B., Pires, D. E. & Ascher, D. B. CSM-carbohydrate: protein-carbohydrate binding affinity prediction and docking scoring function. Brief. Bioinforma. 23, bbab512 (2022).
dos Santos, C. R. et al. Molecular insights into substrate specificity and thermal stability of a bacterial GH5-CBM27 endo-1, 4- -Dmannanase. J. Struct. Biol. 177, 469-476 (2012).
Roske, Y., Sunna, A., Pfeil, W. & Heinemann, U. High-resolution crystal structures of Caldicellulosiruptor strain Rt8B. 4 carbohydrate-binding module CBM27-1 and its complex with mannohexaose. J. Mol. Biol. 340, 543-554 (2004).
Zhao, L. et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 359, 1151-1156 (2018).
O’Keefe, S. J. et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 6, 1-14 (2015).
Sorbara, M. T. & Pamer, E. G. Microbiome-based therapeutics. Nat. Rev. Microbiol. 20, 365-380 (2022).
Gauffin Cano, P., Santacruz, A., Moya, Á. & Sanz, Y. Bacteroides uniformis CECT 7771 ameliorates metabolic and immunological dysfunction in mice with high-fat-diet induced obesity. (2012).
López-Almela, I. et al. Bacteroides uniformis combined with fiber amplifies metabolic and immune benefits in obese mice. Gut Microbes 13, 1-20 (2021).
Morita, H. et al. Bacteroides uniformis and its preferred substrate, cyclodextrin, enhance endurance exercise performance in mice and human males. Sci. Adv. 9, eadd2120 (2023).
Yan, Y. et al. Bacteroides uniformis-induced perturbations in colonic microbiota and bile acid levels inhibit TH17 differentiation and ameliorate colitis developments. npj Biofilms Microbiomes 9, 56 (2023).
Dai, W. et al. Discovery of Bacteroides uniformis F18-22 as a Safe and Novel Probiotic Bacterium for the Treatment of Ulcerative Colitis from the Healthy Human Colon. Int. J. Mol. Sci. 24, 14669 (2023).
Nie, Q. et al. Gut symbionts alleviate MASH through a secondary bile acid biosynthetic pathway. Cell 187, 2717-2734. e2733 (2024).
Zeng, P., Li, J., Chen, Y. & Zhang, L. The structures and biological functions of polysaccharides from traditional Chinese herbs. Prog. Mol. Biol. Transl. Sci. 163, 423-444 (2019).
Cuskin, F. et al. Human gut Bacteroidetes can utilize yeast mannan through a selfish mechanism. Nature 517, 165-169 (2015).
Sastre, D. E. et al. Human gut microbes express functionally distinct endoglycosidases to metabolize the same N-glycan substrate. Nat. Commun. 15, 5123 (2024).
Martens, E. C., Roth, R., Heuser, J. E. & Gordon, J. I. Coordinate regulation of glycan degradation and polysaccharide capsule biosynthesis by a prominent human gut symbiont. J. Biol. Chem. 284, 18445-18457 (2009).
Bågenholm, V. et al. Galactomannan catabolism conferred by a polysaccharide utilization locus of Bacteroides ovatus: enzyme synergy and crystal structure of a -mannanase. J. Biol. Chem. 292, 229-243 (2017).
Patnode, M. L. et al. Strain-level functional variation in the human gut microbiota based on bacterial binding to artificial food particles. Cell host microbe 29, 664-673. e665 (2021).
Odamaki, T. et al. Comparative genomics revealed genetic diversity and species/strain-level differences in carbohydrate metabolism of three probiotic bifidobacterial species. Int. J. Genomics 2015, 567809 (2015).
van Leeuwen, P. T., Brul, S., Zhang, J. & Wortel, M. T. Synthetic microbial communities (SynComs) of the human gut: design, assembly, and applications. FEMS Microbiol Rev 47, https://doi. org/10.1093/femsre/fuad012 (2023).
Cheng, A. G. et al. Design, construction, and in vivo augmentation of a complex gut microbiome. Cell 185, 3617-3636.e3619 (2022).
Parks, D. H. et al. GTDB: an ongoing census of bacterial and archaeal diversity through a phylogenetically consistent, rank normalized and complete genome-based taxonomy. Nucleic acids Res. 50, D785-D794 (2022).
Bacic, M. K. & Smith, C. J. Laboratory maintenance and cultivation of bacteroides species. Curr. Protoc. Microbiol. https://doi.org/10. 1002/9780471729259.mc13c01s9 (2008).
Liu, W., Zhu, W. Y., Yao, W. & Mao, S. Y. Isolation and identification of a lactate-utilizing, butyrate-producing bacterium and its primary metabolic characteristics. Wei Sheng Wu Xue Bao 47, 435-440 (2007).
Fischer, E. & Sauer, U. Metabolic flux profiling of Escherichia coli mutants in central carbon metabolism using GC-MS. Eur. J. Biochem 270, 880-891 (2003).
Portnoy, V. A., Herrgård, M. J. & Palsson, B. Aerobic fermentation of D-glucose by an evolved cytochrome oxidase-deficient Escherichia coli strain. Appl Environ. Microbiol 74, 7561-7569 (2008).
DuBois, M., Gilles, K. A., Hamilton, J. K. & Rebers, P. t. & Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350-356 (1956).
Dubois, M., Gilles, K., Hamilton, J., Rebers, P. & Smith, F. A colorimetric method for the determination of sugars. Nature 168, 167-167 (1951).
Sun, L. et al. Structural characterization of rhamnogalacturonan domains from Panax ginseng CA Meyer. Carbohydr. Polym. 203, 119-127 (2019).
Vidal, S., Williams, P., O’neill, M. & Pellerin, P. Polysaccharides from grape berry cell walls. Part I: tissue distribution and structural characterization of the pectic polysaccharides. Carbohydr. Polym. 45, 315-323 (2001).
Zhang, X. et al. Total fractionation and characterization of the watersoluble polysaccharides isolated from Panax ginseng CA Meyer. Carbohydr. Polym. 77, 544-552 (2009).
Kazemi, M., Khodaiyan, F., Labbafi, M., Hosseini, S. S. & Hojjati, M. Pistachio green hull pectin: Optimization of microwave-assisted extraction and evaluation of its physicochemical, structural and functional properties. Food Chem. 271, 663-672 (2019).
Feng, Z. et al. The model polysaccharide potato galactan is actually a mixture of different polysaccharides. Carbohydr. Polym. 313, 120889 (2023).
Yang, X., Zhao, Y., Wang, Q., Wang, H. & Mei, Q. Analysis of the monosaccharide components in Angelica polysaccharides by high performance liquid chromatography. Anal. Sci. 21, 1177-1180 (2005).
Liu, J. et al. Multi-level fingerprinting and cardiomyocyte protection evaluation for comparing polysaccharides from six Panax herbal medicines. Carbohydr. Polym. 277, 118867 (2022).
Chen, S., Zhou, Y., Chen, Y. & Gu, J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884-i890 (2018).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. methods 9, 357-359 (2012).
Li, B. & Dewey, C. N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinforma. 12, 1-16 (2011).
Dillies, M.-A. et al. A comprehensive evaluation of normalization methods for Illumina high-throughput RNA sequencing data analysis. Brief. Bioinforma. 14, 671-683 (2013).
Terrapon, N., Lombard, V., Gilbert, H. J. & Henrissat, B. Automatic prediction of polysaccharide utilization loci in Bacteroidetes species. Bioinformatics 31, 647-655 (2015).
Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic acids Res. 32, 1792-1797 (2004).
Yang, Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol. Evolution 24, 1586-1591 (2007).
Zhang, C., Shine, M., Pyle, A. M. & Zhang, Y. US-align: universal structure alignments of proteins, nucleic acids, and macromolecular complexes. Nat. methods 19, 1109-1115 (2022).
Teufel, F. et al. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 40, 1023-1025 (2022).
Wells, J. et al. Chainsaw: protein domain segmentation with fully convolutional neural networks. Bioinformatics 40, btae296 (2024).
Miller, G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31, 426-428 (1959).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing (Vienna, Austria, 2018). https://www.R-project.org/.
Visualizer, D. Discovery studio visualizer. 2. Accelrys Software Inc. (2005).
Camacho, C. et al. BLAST+: architecture and applications. BMC Bioinforma. 10, 1-9 (2009).
شكر وتقدير
نود أن نشكر ثانه بينه نغوين من جامعة كوينزلاند على مساعدته في حساب قيمة دلتا-جي (طاقة الربط) والتقييم للمجمع الرئيسي للتفاعل باستخدام أداة CSM-الكربوهيدرات. تم تمويل هذا العمل من خلال منح منتظمة (رقم الملف 0063/2022/A2، 0018/2024/RIB1 و 0137/2024/AMJ) مدعومة من صندوق تطوير العلوم والتكنولوجيا في ماكاو ووزارة العلوم والتكنولوجيا لجمهورية الصين الشعبية. كما تم دعم هذا العمل من قبل عالم شاب 2020 كيهوانغ بتمويل من الإدارة الوطنية للطب الصيني التقليدي ونظام تمويل FDCT للباحثين ما بعد الدكتوراه في مؤسسات التعليم العالي (0017/2021/APD). كما يتم دعم هذا العمل مالياً من قبل منحة البحث الابتدائية لجامعة ماكاو (SRG2022-00020FHS)، ومنحة البحث الداخلية لمختبر الدولة الرئيسي لجودة البحث في الطب الصيني (UM) لعام 2023 (IRG) (SKL-QRCM-IRG2023-001 & 005/2023/SKL)، ومشروع FSCPO (النوع 1 والنوع 2) من كلية العلوم الصحية بجامعة ماكاو. حصل هذا العمل على دعم من مختبر شنتشن الرئيسي لتصنيع الأدوية الميكروبية الذكية (ZDSYS20210623091810032). كما يتم تمويل هذا العمل من قبل المؤسسة الوطنية للعلوم الطبيعية في الصين بموجب رقم المنحة 32201313.
مساهمات المؤلفين
قام Z.Q. و H.L. و J.Y. بإجراء التجارب وتحليل البيانات بمساعدة من L.Z. و J.H. و Z.W. و C.X. و W.Z. و X.X. كتب Z.Q. الورقة. قام E.L.L. و L.D. و H.Z. بتصميم وتوجيه المشاريع. قدم L.S. و Y.Z. و Y.X. و J.L. موارد الأعشاب وناقشوا النتائج ووافقوا على الورقة. قدم Y.-Z.Z. و L.Y. و L.L. تعليقات على تحليل البيانات والمخطوطة.
Selective utilization of medicinal polysaccharides by human gut Bacteroides and Parabacteroides species
Received: July 2024
Accepted: 30 December 2024
Published online: 14 January 2025
Check for updates
Zepeng Qu , Hongbin Liu , Ji Yang , Linggang Zheng , Jumin Huang , Ziming Wang , Chun Xie , Wenlong Zuo , Xiong Xia , Lin Sun , Yifa Zhou , Ying Xie , Jingguang Lu¹, Yizhun Zhu¹, Lili Yu¹, Lihua LiuT, Hua Zhou , Lei Dai Elaine Lai-Han Leung
Human gut Bacteroides and Parabacteroides species play crucial roles in human health and are known for their capacity to utilize diverse polysaccharides. Understanding how these bacteria utilize medicinal polysaccharides is foundational for developing polysaccharides-based prebiotics and drugs. Here, we systematically mapped the utilization profiles of 20 different medicinal polysaccharides by 28 human gut Bacteroides and Parabacteroides species. The growth profiles exhibited substantial variation across different bacterial species and medicinal polysaccharides. Ginseng polysaccharides promoted the growth of multiple Bacteroides and Parabacteroides species; in contrast, Dendrobium polysaccharides selectively promoted the growth of Bacteroides uniformis. This distinct utilization profile was associated with genomic variation in carbohydrate-active enzymes, rather than monosaccharides composition variation among medicinal polysaccharides. Through comparative transcriptomics and genetical manipulation, we validated that the polysaccharide utilization locus PUL34_Bu enabled Bacteroides uniformis to utilize Dendrobium polysaccharides (i.e. glucomannan). In addition, we found that the GH26 enzyme in PUL34_Bu allowed Bacteroides uniformis to utilize multiple plant-derived mannan. Overall, our results revealed the selective utilization of medicinal polysaccharide by Bacteroides and Parabacteroides species and provided insights into the use of polysaccharides in engineering the human gut microbiome.
Human gut Bacteroides and Parabacteroides, primary consumers of polysaccharides, play critical roles in human health and disease, such as the development and immunotherapy of cancer, colonization resistance to pathogens , etc. They utilize endogenous (host-derived) and exogenous (e.g. plant-derived) oligo/polysaccharides, providing nutrition and vitamins to the host and other intestinal microbes . Recent studies highlight that polysaccharides impact host health by regulating the growth and metabolic profiles of Bacteroides .
Understanding how Bacteroides and Parabacteroides utilize these polysaccharides is essential for developing novel polysaccharidebased prebiotics and drugs to promote health.
Medicinal polysaccharides from herbs and mushrooms have complex and diverse structures, comprising various glycosidic linkages (e.g., – or -glycosidic linkages) in backbone or branched chains and including ten or more monosaccharides, sometimes with modifications like methylation and
acetylation . Research shows that medicinal polysaccharides and other plant-derived polysaccharides (e.g. from vegetables and fruits) possess various biological activities , including anti-tumor, antiviral, anti-inflammatory, and immunomodulatory effects. Bacteroides and Parabacteroides species can be promoted by multiple medicinal polysaccharides. Ginseng polysaccharides (GPs) have been shown to enrich both Bacteroides vulgatus and Parabacteroides distasonis, thereby enhancing the response rate of PD-1/PD-L1 immunotherapy . Ophiocordyceps sinensis polysaccharides have been found to enrich gut commensal Parabacteroides goldsteinii, leading to improvements in obesity and metabolic disorders . Dendrobium officinale polysaccharide recall the diversity of gut microbiota, and increasing the abundance of Bacteroides, having protective effect against dextran sulfate sodium induced colitis in mice .
Despite the advancement in polysaccharides utilization of Bacteroides, how Bacteroides utilize medicinal polysaccharides is not clear. Bacteroides and Parabacteroides are equipped with hundreds of carbohydrate-active enzymes (CAZymes) responsible for degrading polysaccharides . The CAZymes are typically organized in clusters known as polysaccharide utilization loci (PULs), which are upregulated by specific polysaccharides . Glycoside hydrolase (GH) are enzymes responsible for the hydrolysis of glycosidic bonds. The genomes of Bacteroides species usually harbor multiple PULs for utilizing different polysaccharides. Some of the mechanisms have been elucidated, for example, B. thetaiotaomicron VPI-5482 utilize starch using Sus system ; Bacteroides spp. degrade complex O-glycans found in mucins using sulfatase ; B. plebeius utilize sulphated polysaccharide porphyran utilization using porphyranases , etc.
In this study, we characterized the growth phenotypes of 28 human gut Bacteroides and Parabacteroides species using 20 medicinal polysaccharides. We observed significant variations in growth profiles among species and polysaccharides. Notably, Dendrobium polysaccharides (DPs, a type of glucomannan) specifically induced the growth of B. uniformis DA183. Through transcriptomics and genetic manipulations, we identified the key gene cluster PUL34_Bu and the critical enzyme GH26_BuDA183 involved in DPs utilization. In vitro enzyme activity assays and molecular docking further elucidated the molecular mechanism of GH26 catalysis. These results provide valuable insights into how human gut microbes utilize medicinal polysaccharides and offer important information for developing novel polysaccharide-based drugs.
Results
Mapping the utilization profile of medicinal polysaccharides by human gut Bacteroides and Parabacteroides species
To comprehensively analyze the utilization of medicinal polysaccharides by human gut Bacteroides and Parabacteroides, we developed an in vitro gut bacterial growth profile platform showed in Fig. 1A, starting with testing three minimal media. Growth curves indicated that modified Bacteroides minimal medium (mBMM), a synthetic medium, generally supported the growth of Bacteroides and Parabacteroides species (Supplementary Fig. 1).
Bacteroides and Parabacteroides, which are highly prevalent and abundant in the human gut microbiota, encode numerous PULs. The platform includes 18 Bacteroides species, 6 Parabacteroides species, and 4 Phocaeicola species (formerly Bacteroides spp.). Some strains were chosen repetitively at the species level (e.g., B. fragilis DA486 and DA557) due to their different CAZyme profiles. To evaluate the prevalence and mean relative abundancreactivee of these Bacteroides species in the human gut microbiota, we analyzed publicly available 16 S rRNA gene amplicon datasets ( ) from GMrepo . We found that of the Bacteroides species had a prevalence greater than , and had a mean relative abundance greater than 0.1% (Fig. 1B, Supplementary Tables 1, 2). The coverage of core human gut Bacteroides species (prevalence , mean relative
abundance ) and CAZymes in Bacteroides (CAZy database ) was and , respectively (Fig. 1C, D). These selected Bacteroides ensure a systematic and comprehensive analysis of polysaccharide utilization.
Bacteroides and Parabacteroides were reported as the target of many medicinal polysaccharides in the intestine, but the mechanisms of their utilization remain underexplored. To expand our understanding, we selected 20 phylogenetic different medicinal polysaccharides with distinct structure (i.e. sugar linkage and modification) (Table 1). For instance, GPs from the Araliaceae family contained starch-like glucans, arabinogalactans, -GalA backbone (homogalacturonan), rhamnogalacturonan-rich pectin ; Dendrobium polysaccharides (DPs_1 and DPs_2, prepared using different methods) feature backbones of )- -D-Glcp- – -D-Manp- -2-O-acetyl- -D-Manp- , and -3-O-acetyl- -D-Manp- . These 20 medicinal polysaccharides exhibit multiple biological activities, including anti-tumor, antioxidant, anti-inflammatory, and immune regulation, while maintaining low toxicity and minimal side effects. More details about other medicinal polysaccharides used in this study can be found in Supplementary Table 3A and B.
We found that the bacterial growth data were consistent across three technical replicates ( ) (Supplementary Fig. ) and between two independent experiments ( , ) (Supplementary Fig. 2D), indicating that our method is highly reproducible. Characterizing the growth phenotypes revealed diverse growth profiles among different Bacteroides and medicinal polysaccharides (Fig. 1E, Supplementary Fig. 3A-C). The statistical analysis indicated that each strain can utilize an average of 4.79 types of medicinal polysaccharides, among which some strains can utilize up to 9 medicinal polysaccharides, while others cannot utilize any of the 20 medicinal polysaccharides. Each type of medicinal polysaccharide can support the growth of an average of 6.8 strains. Some polysaccharides, such as GPs, support the growth of multiple Bacteroides and Parabacteroides species, whereas others, like DPs_2, support only a single Bacteroides species. We found that polysaccharides extracted from plants with close phylogeny resulted in similar bacterial growth profiles (Supplementary Fig. 3D). This similarity is likely due to comparable polysaccharide compositions and structures, as seen in RNPs and PQPs, which both contain Ara, Gal, GalA, Glc, GlcA, Man, and Rha, with glycosidic linkages forming their linear backbones .
Growth profiles variation among medicinal polysaccharides may be due to the differences in physicochemical properties of polysaccharides, such as monosaccharide composition, and polysaccharide structure. To determine whether polysaccharide structure or monosaccharides composition causes variations in growth profiles, we selected eight medicinal polysaccharides exhibiting different growth profile. First, we characterized the physical and chemical properties of these polysaccharides (Supplementary Fig. 4A-B, Supplementary Fig. 5A-D, and Supplementary Tables 5-7). The monosaccharide composition analysis revealed that APs, GPs, GLPs_1, and GLPs_2 were mainly composed of glucose (Glc) (92.8%, , and , respectively). DPs_1 and DPs_2 were mainly composed of mannose (Man) ( and 78.7%, respectively). LBPs_1 were mainly composed of arabinose (Ara), galactose, and galacturonic acid (GalA), while LBPs_2 were mainly composed of , and . The selected medicinal polysaccharides showed differences in total sugar content, molecular weight and distribution, and monosaccharide composition, which indicated that the medicinal polysaccharides were diverse in physicochemical properties. Notably, we found almost no correlation between distance of monosaccharide composition and growth profile, indicating that polysaccharides with similar monosaccharide composition did not exhibit similar growth profiles (Supplementary Fig. 4C). For instance, Astragalus polysaccharide (APs) and GPs,
Fig. 1 | The utilization profile of medicinal polysaccharides by human gut Bacteroides and Parabacteroides species. A The flow chart showing the experimental process of growth profile characterization for medicinal polysaccharides, including i) extraction of medicinal polysaccharides using hot water/ethanol, ii) isolation of the human gut Bacteroides species through culturomics, iii) growth profiling of Bacteroides species with minimal medium supplemented with medicinal polysaccharides in an anaerobic chamber , and . Growth optical density (OD) was measured and recorded every 12 h using a microplate reader. B The prevalence (bar chart) and relative abundance (point plot) of Bacteroides species based on public human gut metagenome sequence data ( ). The ‘ ‘ refers to the number of fecal samples collected from healthy individuals in the GMrepo database. Bacteroides species were colored by genus, Bacteroides in blue, Parabacteroides in red, and Phocaeicola in green. Prevalence 0.3 (red dotted line), Relative abundance (green dotted line). Relative abundance data are expressed as mean values ± standard deviation (SD). C The pie chart showed the coverage of core human gut Bacteroides (prevalence> , mean relative abundance ). D The pie chart showed the coverage of human gut Bacteroides carbohydrate active enzymes in CAZy database. E The heat map showed the growth OD600 of 28 Bacteroides species at 48 h under 20
medicinal polysaccharides ( independent experiments). Statistical significance was determined using a two-sided unpaired t-test between each experimental group and the negative control group, with adjustments for multiple comparisons made using the Bonferroni method. The values are indicated as follows: *<0.05; ; *** < 0.001; . Each experiment was conducted in triplicate to ensure reliability. The sub-graph in the right counts the number of medicinal polysaccharides utilized by Bacteroidetes spp. The sub-graph at the bottom counts the number of Bacteroidetes spp. utilizing medicinal polysaccharides. RNPs: Radix Notoginseng Polysaccharides; GPs: Ginseng polysaccharides; PQPs: Panax Quinquefolium Polysaccharides; PSPs: Polygonatum sibiricum Polysaccharides; RAPs: Rhizoma anemarrhenae Polysaccharides; CPPs: Codonopsis Pilosula Polysaccharides; AdPs: Adenophora Polysaccharides; PLPs: Pueraria lobata Polysaccharides; APs: Astragalus polysaccharide; DPs_1 and DPs_2: Dendrobium polysaccharides; GLPs_1 and GLPs_2: Ganoderma lucidum polysaccharides; LBPs_1 and LBPs_2: Lycium barbarum polysaccharides; PPs: Poria Polysaccharides; RPPs: Radix Pseudoxtellariae Polysaccharides; AnPs: Angelica Polysaccharides; HAPs: Hovenia Acerba Polysaccharides; SPs: Scrophulariaceae Polysaccharide; PC: positive control; NC: negative control.
despite having similar monosaccharide compositions, displayed markedly different human gut bacteria growth profiles. It is worth mentioning that APs consist of methylated GalA and diverse side chains , differing from GPs. These findings suggest that polysaccharide structure, rather than monosaccharide composition, drives growth profile variations.
The utilization profile of medicinal polysaccharides is associated with genomic variation in carbohydrate-active enzymes
The characterization of the chemical structure of polysaccharides is a recognized worldwide challenge. However, complex polysaccharides have led to the evolution of numerous diverse carbohydrate-active enzymes (CAZymes) families. In turn, the specificity of degrading
Notes: f_ means family; s_ means species
enzymes can provide information on the structure of the degraded polysaccharides. The genes encoded by different gut bacteria can help us understand the utilization profile. Bacteroides encode hundreds CAZymes to utilize complex polysaccharides . CAZymes involved in carbohydrate degradation include glycoside hydrolases (GHs) for hydrolyzing glycosidic linkages, polysaccharide lyases (PLs) for breaking down polysaccharides containing uronic acids, carbohydrate-binding modules (CBMs) for binding carbohydrate motifs, carbohydrate esterases (CEs) for de-esterification, and auxiliary activity enzymes (AAs) for various redox transformations. Using dbCAN (version 4.1.4) to analyze annotated CAZymes, we observed variations in CAZyme distribution at the genus, species, and strain levels among Bacteroides and Parabacteroides species (Fig. 2A). Clustering the 28 Bacteroides based on the Euclidean distance of CAZymes revealed that Bacteroides and Parabacteroides with similar taxonomy share similar CAZyme distribution patterns (Fig. 2A). GHs were the most widely encoded, whereas AAs were less common due to the gut’s anaerobic environment limiting oxygen-dependent AAs .
To explain the growth profile variations among different Bacteroides and Parabacteroides species, we clustered the 28 stains based on the Euclidean distance of CAZymes (Fig. 2B), and growth profile distribution (Fig. 2C). We found that Bacteroides spp. with similar CAZyme genes distribution had similar polysaccharide metabolic pattern. For example, P. dorei DA26 and P. vulgatus DA57 exhibited similar CAZyme gene distributions and polysaccharide metabolic patterns. A similar phenomenon was observed among Parabacteroides spp. Additionally, a significant correlation was observed between the distances of CAZyme distribution and growth profiles, indicating that Bacteroides and Parabacteroides with similar CAZyme distributions exhibit similar growth profiles (Fig. 2D). These results suggest that CAZyme distribution drives growth profile differences.
Transcriptomics analysis reveals that specific PULs are highly up-regulated by medicinal polysaccharides
Differential gene expression aids in discovering enzymes and pathways for utilizing specific carbon sources. First, we selected Ginseng polysaccharides (GPs) that supported the growth of multiple
Bacteroides. We analyzed the transcriptome data of P. vulgatus DA57 and . distasonis DA104 cultivated with GPs and glucose as the sole carbon sources (Fig. 3A-F, Supplementary Fig. 6A-D, and Supplementary Tables 8, and 9). Comparative transcriptome analysis revealed that the top 10 upregulated carbohydrate metabolism genes were located in PUL19 of P. vulgatus DA57 (PUL19_Pv) (Fig. 3A) and PUL29 of P. distasonis DA104 (PUL29_Pd) (Fig. 3C) as annotated by PULpy . We refined the PUL boundaries based on the expression levels of upstream and downstream genes (Fig. 3C, E, I, and Supplementary Fig. 7C) (see “Methods” for more details). Notably, GH13_10 and GH13_20, described as -amylases by the CAZy database , were upregulated in both PUL19 Pv and PUL29 Pd. Given that neutral polysaccharides, including amyloid mixtures , are the main components of GPs, these results suggest that GH13_10 and GH13_20 are crucial for GPs utilization.
Next, we examined gene expression data of Bacteroides utilizing Ganoderma lucidum polysaccharides (GLPs_1), which supported the growth of a moderate number of Bacteroides. The top 10 upregulated carbohydrate metabolism genes were located in PUL16, PUL25, and PUL39 of B. uniformis DA183 (Supplementary Fig. 6E-F, Supplementary Fig. 7A, and Supplementary Table 10). Analysis of gene cluster structures showed that these PULs encoded GH55 (i.e., exo/endo- -1,3-glucanase) in PUL16, GH9 (i.e., endo- -1,4-glucanase) in PUL16&25, GH3 (i.e., endo- -1,6-glucosidase) in PUL39, and GH16_3 and GH158 (i.e., endo- -1,3-glucanase) in PUL39 (Supplementary Fig. 7B, C). The presence of multiple PULs responsible for the same polysaccharides aligns with previous studies . GLPs are composed of )- -D-GIcp- –-D-Galp- , and – -D-Glcp- . These findings indicate that PUL16, PUL25, and PUL39 are likely responsible for GLP utilization.
We also analyzed gene expression data for Bacteroides utilizing DPs_2, which selectively promoted the growth of B. uniformis. Focusing on B. uniformis DA183, we compared transcriptome data when cultivated with DPs and glucose as the sole carbon sources. The top 10 upregulated carbohydrate metabolism genes were located in PUL34 of B. uniformis DA183 (PUL34_Bu) (Fig. 3G-I, Supplementary Fig. 6E-F, Supplementary Table 11). DPs are mannose oligosaccharide with a backbone consisted of -D-GIc -D-Man
Fig. 2 | Genomic variation in carbohydrate-active enzymes determines the distinct utilization profile of medicinal polysaccharides among Bacteroides and Parabacteroides species. A The heat map showed the distribution of CAZyme genes involved in carbohydrate degradation. Bacteroidetes spp. were clustered based on the Euclidean distance of CAZyme genes. CAZymes genes clustered based on the Euclidean distance of distribution in Bacteroides and Parabacteroides. Glycoside Hydrolases (GHs): hydrolysis and/or rearrangement of glycosidic bonds; Polysaccharide Lyases (PLs): non-hydrolytic cleavage of glycosidic bonds; Carbohydrate-Binding Modules (CBMs): adhesion to carbohydrates; Carbohydrate Esterases (CEs): hydrolysis of carbohydrate esters; Auxiliary Activities (AAs): redox enzymes that act in conjunction with CAZymes. B Bray curtis based principal components analysis (PCA) shows the dimension reduction analysis of
carbohydrate active enzymes in Bacteroides. The dots on the graph are colored according to genus of Bacteroidetes. C Euclidean distance based principal components analysis (PCA) shows the dimension reduction analysis of polysaccharides utilization at 48 h by Bacteroides. The dots on the graph are colored according to genus of Bacteroidetes. D The point diagram displays the correlation between growth profile distance and CAZymes distribution distance. The R and values refer to the most parsimonious model. Statistical significance was determined using a two-sided Pearson correlation test (for R value) and an ordinary least squares (OLS) regression analysis (for the linear fit). The figure also includes a confidence interval around the best linear fit. Light green dots represented high dot density and dark green dots represented low dot density.
O-acetyl- -D-Manp- , and -3-O-acetyl- -D-Manp- . The measured monosaccharide composition of DPs confirms that mannose is the main component (Supplementary Fig. 4A, B). Two significantly upregulated glycoside hydrolases (GHs) in PUL34_Bu, GH26 and GH5_7, are described as exo- -1,4-mannobiose and endo- -1,4mannanase by the CAZy database .
PUL34_Bu gene cluster in B. uniformis is required for Dendrobium polysaccharides utilization
To explore the genetic basis underlying the highly specific utilization of Dendrobium polysaccharides (DPs) by B. uniformis, we utilized CRISPR/Cas-based Bacteroides genome editing tool to construct PUL34 knock-out mutants (Methods). To ensure successful gene
knockout, we targeted two regions in the gene cluster for separate knockout (Fig. 4A). The first region (PUL34M_Bu) was a moderately sized segment with significant upregulation at the transcriptional level and genes transcribed in the same direction. The second region (PUL34S_Bu) contained three key enzymes which were isozymes of three key enzymes involved in inulin utilization , including a SusD-like protein for polysaccharide binding
in the outer membrane, a SusC-like protein for polysaccharide transport, and GH26 for glycosidic bond hydrolysis. Gel electrophoresis and DNA sequencing confirmed the successful knockout of the two regions (Fig. 4B, Supplementary Fig. 9A-D). When grown in mBMM medium with DPs as the sole carbon source, the wild-type strain grew, whereas the mutant strain did not (Fig. 4C, and D).
Fig. 3 | Up-regulation of specific polysaccharide utilization locus genes by medicinal polysaccharides. A Volcano plot of . vulgatus DA57 gene expression comparing minimal media supplemented with GPs to glucose, showing the foldchange ( -axis) versus the differential significance (-log10 adjusted -value, axis). The top 10 up-regulated genes are highlighted in green. Differential expression analysis was performed using DESeq2, employing a two-sided Wald test. Adjusted -values were obtained after correcting for multiple hypothesis testing using the Benjamini and Hochberg method to control the false discovery rate (FDR). Genes with an adjusted -value are considered statistically significant. The -axis represents the fold change in gene expression, while the -axis shows the – of the adjusted -value. Each experiment was conducted in triplicate to ensure reliability. B The schematic diagram shows that PUL19 of P. vulgatus DA57 (PUL19_Pv) contains 8 genes, with a cluster size of 14,518 base pairs. susC: susC-like TonB-dependent transporter; susD: susD-like cell-surface glycan-
binding protein; GH:glycoside hydrolyze; Unk: unknown functions. C Volcano plots of P. distasonis DA104 gene expression comparing minimal media supplemented with GPs to glucose. The statistical analysis is the same as in (A). D The schematic diagram shows that PUL29 of P. distasonis DA104 (PUL29 Pd) contains 8 genes, with a cluster size of 13,063 base pairs. E-F Bubble plot displays the differential expression of the E) PUL19 Pv , and F) PUL29 Pd genes and its neighboring 3 genes, utilizing GPs and glucose. G Volcano plots of B. uniformis DA183 gene expression comparing minimal media supplemented with DPs to glucose. The statistical analysis is the same as in (A). H The schematic diagram shows that PUL34_Bu contains 22 genes, with a cluster size of 39,709 base pairs. CE: carbohydrate esterase. I Bubble plot displays the differential expression of the PUL34_Bu gene and its neighboring 3 genes, utilizing DPs and glucose. The region between two vertical cyan dotted lines indicates significantly upregulated genes with consistent transcription direction.
Fig. 4 | PUL34_Bu gene cluster in B. uniformis is required for Dendrobium polysaccharides utilization. A The schematic diagram shows the two region of PUL34_Bu genes knock out. PUL34M_Bu indicates significantly upregulated genes with consistent transcription direction. PUL34S_Bu indicates the putative three key genes. B Nucleic acid electrophoresis diagram shows the four DNA products: products amplified with primers designed within a 1.5 k region flanking the region of PUL34M_Bu (P1 ~ P4), and genomic DNA of wild-type, and mutate strain; Products amplified with primers designed within a 1.5 k region flanking the region of PUL34S_Bu (P2 ~ P3), and genomic DNA of wild-type, and mutate strain. If the fragment is successfully deleted, a 3000 bp product will be amplified. In contrast, if
the deletion is not successful, no product will be amplified due to the excessive length of the target band. This representative image is from a single experiment. C-D The point plot shows the growth density of C) wild-type and PUL34S knockout strains, and D) wild-type and PUL34M knockout strains in mBMM medium with DPs_2 as the sole carbon source. Data are presented as mean values ± SD. Assays were performed in technical triplicates ) E The heatmap shows the correlation between PUL34_Bu and growth density OD600. PUL34_Bu genes were identified by BLASTP (version 2.6.0) (coverage , identify ). The species carrying PUL34_Bu genes were colored in cyan, otherwise, in white. The species utilizing DPs was colored gray, otherwise, in white.
To investigate the distribution of PUL34_Bu in the genomes of 27 other Bacteroides and Parabacteroides species, we performed sequence alignment with a sequence similarity threshold of and a coverage threshold of (Methods). We found that only B. uniformis encoded a complete PUL34_Bu gene cluster (Fig. 4E). In summary, we revealed that a complete PUL34_Bu gene cluster allowed for the highly specific utilization of DPs by B. uniformis.
GH26 enzymes is necessary for plant-derived mannan utilization in B. uniformis
We constructed a phylogenetic tree for the 89 characterized GH26 protein sequences deposited in the CAZy database (Methods). B. uniformis GH26 is clustered with other enzymes (including GH26 enzymes from B. ovatus ATCC8483) annotated as Mannan endo-1,4- mannosidase (EC 3.2.1.78) (Fig. 5A). To explore the distribution of .
uniformis GH26 in human gut bacteria, we performed sequence alignments in UHGP database using DIAMOND methods . We found that the B. uniformis GH26 orthologs were mainly present in Bacteroides spp. at the functional conserved alignments threshold (identity , coverage ), including 64/66 B. uniformis, 26/32 B. cellulosilyticus, 3/17 B. ovatus, 2/2 B. stercorirosoris, and 2/2 P. dorei (Supplementary Fig. 10A, B).
To investigate the necessity of PUL34_Bu for the utilization of plant-derived mannans by gut Bacteroides, we conducted growth profiling for B. ovatus ATCC8483, B. uniformis DA183, B. uniformis ATCC8492 (wild-type), and B. uniformis ATCC8492 PUL34S_1&_2 (mutate) utilizing various plant-derived mannans, such as Dendrobium officinale polysaccharide (DPs), konjac glucomannan , guar gum galactomannan , carob galactomannan , and ivory nut mannan . We found that wild type B. uniformis and B. ovatus strains were able to utilize these multiple plant-derived mannan, while the PUL34S knockout mutants were not able to grow (Fig. 5B). These results indicated that GH26 in B. uniformis hydrolyzes a wide range of mannan substrates.
To further understand the structural properties of B. uniformis GH26, we predicted its structure using AlphaFold2 (pLDDT=93.04). We found B. uniformis GH26 contained a signal peptide (residues 1 to 30), a bacterial Ig-like domain (residues 31 to 131), a GH26 domain (residues 135 to 564), and galactose-binding lectin (residues 239 to 354) (Fig. 5C, D). The structure of GH26 enzyme from B. ovatus ATCC8483 has been determined by crystallography (GenBank accession ALJ47537.1; PDB code 6HF4) . Comparing with the structure of . ovatus GH26, we found that the structure of B. uniformis GH26 contained two extra modules: 1) Ig-like domain which is related to the stability of the enzyme or serving as connector between two domains ; 2) carbohydrate binding module . We found that the active sites, the calcium-binding sites, and the substrate-binding sites of B. uniformis GH26, and B. ovatus GH26 are conserved (TM-score 0.91 , RMSD ) (Fig. 5E, and Supplementary Fig. 11).
The conserved residues of B. uniformis GH26 (W527 and Y528) and B. ovatus GH26 (W314 and Y315) are responsible for binding mannose by hydrogen bonds , which indicates that the substrate of . uniformis GH26 contains mannose. However, residues K149 responsible for binding galactose are not conserved between the two enzymes, which indicated a different substrate-binding specificity. Indeed, the substrate of B. ovatus GH26 is a galactomannan, which is structurally different from DPs (i.e. glucomannan).
We observed that DPs_1 also induced the growth of B. finegoldii DA347, which encodes a GH26 gene in its genome (Fig. 4E). This observation prompted us to perform sequence and structural alignments of the GH26 from B. finegoldii DA347. Sequence alignment showed low similarity among GH26 from B. uniformis DA183, B. ovatus ATCC8483, and B. finegoldii DA347. The sequence identity is coverage) between B. uniformis DA183 and B. ovatus ATCC8483, 49.1% (52.0% coverage) between B. uniformis DA183 and B. finegoldii DA347, and 48.3% ( coverage) between B. ovatus ATCC8483 and B. finegoldii DA347 (Supplementary Fig. 12A). Structural alignment showed that B. finegoldii DA347 encodes an enzyme structurally similar to B. ovatus ATCC8483 GH26 (TM-score , RMSD ) (Supplementary Fig. 12B-C). Moreover, the active sites (E217 and E321), the calcium-binding sites (L109 and S112), and the substrate-binding sites (W116, W353, and Y354) of GH26 from B. finegoldii DA347 (cyan stick models), and the active sites (E201 and E291), the calcium-binding sites (L105 and S108), and the substrate-binding sites (W112, W314, and Y315) of B. ovatus GH26 (gray stick models) are conserved (Supplementary Fig. 12D). These results may explain its ability to grow using DPs_1.
The enzymatic activity and specificity of Bacteroides GH26 enzyme
To further characterize the enzyme activity and substrate specificity of Bacteroides GH26, we expressed and purified the GH26 enzyme from .
uniformis DA183 and B. ovatus ATCC8483 (Supplementary Fig. 13A, B) and tested them with 13 substrates, including 6 mannans, 3 potential substrates from structurally similar proteins or the same protein family, and 4 well-defined substrates as potential negative controls (Supplementary Table 3C). Reducing-end production was measured using the standard 3,5-dinitrosalicylic acid (DNS) assay with mannose as the standard (Methods). We found that both enzymes hydrolyzed all six mannans, but not the other seven substrates (Fig. 6A, and Supplementary Table 16A, B).
The putative catalytic residues are two Glu residues (E2O1 and E291 in B. ovatus ATCC8483 and E415 and E504 in B. uniformis DA183), which hydrolyze mannans via a double-displacement general acid -base mechanism . We performed point mutations on the two catalytic sites of B. uniformis GH26 (E415A and E504A) and found that mutation of either Glu residue leads to loss of enzyme activity, indicating that both catalytic sites are necessary for enzyme activity (Fig. 6B, and Supplementary Table 17).
To investigate the structural basis of different substrate specificity, we performed molecular docking simulations for 5 structurally defined substrates ( 1 reactive substrate and 4 non-reactive substrates) using AutoDock Vina (v1.2.0) . We predicted their binding affinities and ranked the docking poses of the enzyme-substrate complexes using CSM-carbohydrate . The predicted binding free energy of the non-reactive substrates was only slightly higher than that of the reactive galactosyl mannobiose, and the non-catalyzed trehalose even had a lower binding free energy. This indicates that substrate specificity cannot be solely explained by comparing the binding free energies of the substrates. Notably, the catalytic residues were found in the interactive residues of enzyme-reactive substrate complexes, but not in those of enzyme-non-reactive substrate complexes, suggesting that substrate reactivity depends on their ability to interact with catalytic residues (Fig. 6C, D and Supplementary Fig. 14A-C).
Among the six reactive mannans, GH 26 from B. uniformis DA183 showed the highest activity toward glucomannan (e.g. konjac mannan) (Fig. 6B), while GH26 from B. ovatus ATCC8483 had the highest activity toward galactomannan (Supplementary Fig. 15A). To investigate the role of differential substrate-binding sites in the activity of GH26 from B. uniformis DA183 and B. ovatus ATCC8483, we identified the unique differential site (A363 in B. uniformis DA183 and K149 in B. ovatus ATCC8483) through structural alignment and interchanged the residues. Interestingly, the GH26 BoATCC8483 K149A mutant showed significantly reduced activity toward guar gum galactomannan and became most active toward konjac glucomannan (Supplementary Fig. 15C). Unexpectedly, the enzymatic activity of the GH26 BuDA183 A363K mutant showed a slight increase and did not result in a significant change in substrate specificity, unlike the GH26 BoATCC8483 K149A mutant, suggesting that other unpredicted differences in substrate-binding sites contribute to the distinct substrate preferences of the two enzymes (Fig. 6G, and Supplementary Table 17).
To evaluate the role of CBM domain in GH26 BuDA183, we firstly compared the enzymatic activity of full-length wild type and CBM truncated GH26. Interestingly, no significant difference in enzyme activity was observed between wild type GH26 (Km=3.75 ± 1.44 mg/mL, Kcat and CBM truncated GH26 , Kcat ) (Supplementary Fig. 13A-B, Supplementary Fig. 16A-B, and Supplementary Table 18). Notably, some studies have shown that CBM domain provided thermoprotection for certain carbohydrate-hydrolyzing enzymes . By comparing the enzyme activities of wild-type GH26 and CBMtruncated GH26 at different temperatures, we found that the CBMtruncated GH26 became less thermostable. Specifically, the Tm values were for wild-type GH26 and for the CBM-truncated variant (Supplementary Fig. 16C, and Supplementary Table 19). This indicates that the possible function of the CBM domain of GH26 is related to thermostability.
Fig. 5 | Utilization of plant-derived mannan by Bacteroides GH26 enzymes. A Phylogenetic tree of GH26 protein sequences, including 89 characterized GH26 and . uniformis GH26 in this study. Organism names are indicated to the species level, next to the GenBank accession number of the GH33 protein analyzed. The tree was generated using the Maximum likelihood method. The tree label was colored by EC classification. B. uniformis GH26 in this study was marked with a red five-pointed star. The crystallized GH26 of B. ovatus was marked with a gray fivepointed star. B The point plot shows the growth density of B. ovatus ATCC8483 (blue), B. uniformis DA183 (gray), B. uniformis ATCC8492 (red), mutant strain of B. uniformis ATCC8492 deleted PUL34S (green) and another mutant strain of B. uniformis ATCC8492 deleted PUL34S (orange) utilizing 6 carbon source, such as glucose, Dendrobium officinale polysaccharide (DPs), konjac glucomannan, guar gum
galactomannan, carob galactomannan, and ivory nut mannan. Data are presented as mean values SD. Assays were performed in technical triplicates ( ).
C Domain structure of B. uniformis GH26. D Overall structure of B. uniformis GH26 was predicted by AlphaFold2 based on the protein sequence. The Ig-like domain is colored in in green, GH26 domain in red, galactose-binding lectin in purple, respectively. E The structure of B. uniformis GH26 (red) domain is superimposed on that of B. ovatus GH26 (gray) (PDB code 6HF4) (TM-score , RMSD ). F The active sites (E415 and E504), the calcium-binding sites (L198 and S201), and the substrate-binding sites (V170, A200, W207, Y342, W527, and Y528) of B. uniformis GH26 (red stick models), and the active sites (E201 and E291), the calciumbinding sites (L105 and S108), and the substrate-binding sites (V77, A107, W112, Y148, W314, and Y315) of B. ovatus GH26 (gray stick models) is conserved.
Fig. 6 | The enzymatic activity and specificity of B. uniformis GH26 enzyme. A The bar chart shows the reducing-end mannose produced by GH26 BuDA183 (cyan) and GH26 BoATCC8483 (gray) using 13 mannan substrates. The enzyme concentration was , substrate concentration was , reaction temperature was , and reaction time was 30 min . The reducing end is the terminus of a carbohydrate chain that has a free aldehyde or ketone group capable of reducing copper ions in the DNS assay. Data are presented as mean values ± SD. Assays were performed in technical triplicates ( ). B-D The dot plot shows the reducing-end production over time for six substrates hydrolyzed by GH26 BuDA183 (Wild type), GH26 BuDA183 E415A, and E504A (Catalytic site mutation). The substrates are: konjac glucomannan (yellow), Dendrobium officinale polysaccharides
(red), galactosyl mannabiose (green), carob galactomannan (blue), guar gum galactomannan (pink), and ivory nut mannan (purple). The enzyme concentration was 125 nM , substrate concentration was , reaction temperature was , and reaction time was . Data are presented as mean values SD. Assays were performed in technical triplicates ( ). E-F Interactions of GH26 BuDA183 (Wild type) with galactosyl mannabiose, and 1-kestose are visualized in 3D (left) and 2D (right). Substrates are shown as green sticks or circle, interacting residues as gray sticks or circle, and catalytic sites as cyan sticks or circle. Interaction forces are displayed as dotted lines. G The reaction system is the same as in Panel (B-D), except the enzyme is GH26 BuDA183 A363K (Different residues interchanges).
Discussion
Manipulating the gut microbiome with polysaccharides is a promising therapeutic strategy for various human diseases . Understanding how gut bacteria utilize medicinal polysaccharides will guide the selection of polysaccharides for microbiome manipulation. However, knowledge in this area is still limited. To address this gap, we systematically mapped the utilization profiles of 20 different medicinal polysaccharides by 28 human gut Bacteroides and Parabacteroides species. We found that different Bacteroides and Parabacteroides species showed distinct utilization profiles of medical polysaccharides, which was associated with genomic variation in carbohydrate-active enzymes. Through comparative transcriptomics and genetical manipulation, we validated that the polysaccharide utilization locus PUL34_Bu enabled B. uniformis to utilize Dendrobium polysaccharides.
We discovered that the GH26 enzyme in B. uniformis and B. ovatus, annotated as a mannosidase, was essential for the utilization of multiple plant-derived mannans (e.g. Dendrobium polysaccharides). Overall, our study provides a general framework to profile the utilization of plant-derived polysaccharides by human gut bacteria and to understand the underlying molecular mechanisms. The highly specific utilization of some medicinal polysaccharide (e.g. Dendrobium polysaccharides) also highlight the potential of glycan-based prebiotics and drugs for targeted modulation of the human gut microbiome.
B. uniformis is a prominent member of the human gut microbiota and plays a crucial role in maintaining gut health. Specifically, strain CECT 7771 ameliorates metabolic and immunological dysfunction in mice with high-fat-diet-induced obesity and enhances metabolic and immune benefits when combined with fiber in obese mice .
Additionally, B. uniformis and its preferred substrate, -cyclodextrin, improve endurance exercise performance in both mice and human males . Strain JCM5828 regulates colon NF-кB and MAPK signaling pathways by inhibiting IL-17 signaling, thereby ameliorating colitis development . Strain F18-22 alleviates intestinal inflammation by improving gut dysbiosis . Moreover, B. uniformis produces 3-succinylated cholic acid (3-sucCA), which alleviates metabolic dysfunction-associated steatohepatitis (MASH) . Engineering B. uniformis to enhance its probiotic properties holds significant promise for therapeutic applications. By modifying B. uniformis to more efficiently degrade specific polysaccharides or produce beneficial metabolites, such as 3 -sucCA, we can potentially improve its ability to colonize the gut and exert positive health effects. Engineered strains could also be designed to deliver therapeutic molecules directly to the gut, offering a targeted approach to treating gastrointestinal disorders.
Growth profile variations reflect differences in polysaccharide structure, suggesting the potential for developing quality control markers for medicinal polysaccharides by comparing growth profiles. Quality control in polysaccharide drug manufacturing is challenging due to their complexity, chemical diversity, and batch-to-batch variations . Current methods have been employed for characterizing medicinal polysaccharides, such as 1) quantifying the sugar content based on colorimetry, 2) determining the molecular weight and distribution based on high-performance liquid chromatography (HPLC), and 3) measuring monosaccharide composition based on HPLC. However, such methods do not capture the high-dimensional structure of polysaccharides, which is closely related to their bioactivities. Growth profiles can provide valuable insights into the structural characteristics of polysaccharides.
In this study, it was observed that a polysaccharide can simultaneously up-regulate multiple PULs, and partial gene upregulation in other PULs was also noted, similar to B. dorei’s response to human milk oligosaccharides . These phenomena are potentially due to the functional redundancy of PULs. For example, both PUL72 and PUL16 in . thetaiotaomicron are involved in processing high-mannose glycans . Knockout of six PULs up-regulated by pectin intervention revealed that none were absolutely necessary for complete degradation of the specific pectin preparation . Moreover, B. thetaiotaomicron expresses multiple PULs to respond to host mucin -glycans, ensuring its survival and metabolism even when some PULs are inactive; for instance, deletion of ECF- gene BT1053 or insertion in susC-like gene BT1042 did not significantly affect growth on neutral -glycans .
Notably, several B. uniformis strains, including B. uniformis DA183, contain two GH26 enzymes in their genomes, located in different PULs. These enzymes may have evolved to target different substrates or function under different conditions. For example, in B. ovatus ATCC8483, the PUL encodes two GH26 -mannanases, BoMan26A and BoMan26B, with distinct activities: BoMan26A primarily forms mannobiose, while BoMan26B, which is surface-exposed, degrades galactomannan into a diverse mixture of oligosaccharides . This functional redundancy and specialization ensure efficient degradation and utilization of complex galactomannans.
Future research should explore strain-levels variations in glycan utilization, as seen with B. ovatus using mucin . With decreasing sequencing cost, synthetic communities (SynComs) are being considered for growth profile characterization due to their well-defined compositions and reproducibility . As demonstrated in this study, differences in growth profiles and CAZyme distribution can reflect variations in polysaccharide structures. Therefore, novel growth profiles may aid in identifying polysaccharides with potential novel structures. Additionally, analyzing the proteins that degrade these polysaccharides provides valuable insights into their complex sugar chain architectures . Advances in CAZyme functional prediction may improve our understanding of medicinal polysaccharide structures through PUL transcriptome analysis. Finally, identified PULs can serve
as markers for the genetic engineering of gut bacteria (e.g. PULs for inulin utilization ), and tools for enhancing probiotic colonization (e.g. PULs for porphyrin polysaccharide utilization ).
Methods
Collection and preservation of human stool samples
In this study, all 119 human participants provided informed consent, with approval from the Shenzhen Institute of Advanced Technology, Chinese Academy of Sciences (SIAT-IRB-200315-H0438). Freshly collected stool samples from healthy donors were promptly transferred to an anaerobic chamber (Vinyl Anaerobic Chambers, Coylab, USA). Each sample ( 3 g ) was resuspended in 15 mL of glycerol ( , in sterile phosphate-buffered saline, with -cysteine hydrochloride), vortexed for homogenization, and filtered through sterile nylon mesh to remove large particles. Aliquots of the processed suspensions were stored in sterile cryovials at for long-term preservation, ready for subsequent isolation of gut Bacteroides and Parabacteroides.
Isolation and cultivation of gut Bacteroides and Parabacteroides
Bacteroides uniformis ATCC 8492 and Bacteroides ovatus ATCC 8483 were purchased from ATCC. The 28 human gut Bacteroides and Parabacteroides used in this study were isolated from the feces samples of SIAT cohort. Fecal samples thawed from were diluted and spread onto modified Yeast Casitone Fatty Acids (mYCFA) agar plates (Supplementary Table 12). The plates were incubated anaerobically at for days in an atmosphere of , and . Single colonies were picked up, streaked onto fresh plates, and further incubated anaerobically at for another days. This purification process was repeated once to obtain pure strains. Isolated strains were then transferred into liquid medium and incubated at for 2 days. We then amplified and sequenced the full-length 16S rRNA genes using the PCR primer pair ( 27 F 5′-AGAGTTTGATCMTGGCTCAG-3′; 1492 R 5′-GGTTACCTTGTTACGACTT-3′). Strains that produced double peaks in their sequences were either discarded or subjected to another round of purification. All purified strains were stored at in a glycerol suspension containing cysteine. Taxonomic classification of isolates was determined by comparing their full-length 16 S rRNA gene sequences to those present in the Genome Taxonomy Database (GTDB release 207) (Supplementary Table S1A). For culture purposes, the Bacteroides species were cultivated in brain heart infusion medium (BHIS) supplemented with hemin, vitamin K3, and -cysteine (Supplementary Table 12). The cultivation was carried out in an anaerobic chamber with a gas composition of , and , maintained at a temperature of .
Minimal medium test
To determine a minimal medium suitable for the growth of Bacteroides species, we tested three minimal media: modified Bacteroides minimal medium (mBMM) , modified Yeast Casitone Fatty Acids (mYCFA) medium , and modified M9 minimal (mM9) medium , all supplemented with glucose (Supplementary Table 12). Using an anaerobic workstation with a gas mixture of , and , the modified basic medium was dispensed into a 96-well plate. The bacterial cultures were washed twice with anaerobic PBS, and the optical density at 600 nm (OD600) of the bacterial solution was adjusted to 0.1. Growth curves indicated that both mBMM and mYCFA generally supported the growth of Bacteroides species (Supplementary Fig. 1). Compared to mYCFA, mBMM, being a synthetic medium with a clear composition, provided more stable measurements of polysaccharide utilization.
Prevalence and abundance of Bacteroides species
The abundance and prevalence of the strains chosen in our study was counted. We screened 16282 healthy human metagenome from publicly available metagenomic data by search with defined filter
conditions (experiment_type = ‘Metagenomics’ AND QCStatus = ‘Good runs’ AND host age AND Country=is not null AND Recent Antibiotics Use = ‘No’ AND Phenotype = ‘Health’) in GMrepo to represented the global health human gut microbiota. Species with equally weighted average frequency of occurrence prevalence> and mean relative abundance was defined as core species.
Measurement of bacterial growth profiles
The mBMM medium was added to a 96-well plate in an anaerobic workstation , and . The medium was supplemented with (negative control), glucose (positive control), or medicinal polysaccharides (treatment). Next, the bacteria were washed twice with anaerobic PBS, and the OD600 of the bacteria solution was adjusted to 0.1. The bacteria were then inoculated at a 1:10 ratio into the 96-well plate and cultured at . The growth OD was measured and recorded every 12 h using a microplate reader.
Extraction of medicinal polysaccharides
Seventeen herbs were purchased from Bozhou city (Anhui, China) and authenticated by and identified by one of authors, Dr. Ji Yang. The voucher specimens were deposited at Faculty of Health Sciences (FHS), University of Macau. The more detailed information about herbs see Supplementary Table 3A. Among various extraction methods (e.g., hot water/ethanol, alkaline, enzymatic), hot water extraction is the most widely used due to its ease of operation, high solubility in hot water, and minimal damage to the polysaccharides . Extraction of polysaccharides was performed by soaking the medicinal pieces in 10 times their volume of deionized water for 30 minutes, followed by boiling at high heat and simmering at low heat for 30 minutes. The hot liquid was then filtered through a 200 -mesh cloth, discarding the residue. This process was repeated with another 10 times the volume of deionized water, boiling at high heat and simmering at low heat for 25 minutes, and filtering again while hot. Both filtrates were combined and reduced to a low temperature using vacuum pressure. After cooling, three times the volume of anhydrous ethanol was added to precipitate the polysaccharides while constantly stirring, and the mixture was left to stand overnight at . The precipitate was separated from the liquid using a vacuum pump filter, washed three times with anhydrous ethanol, and vacuumdried at to obtain the final product. For medicinal polysaccharides_2 (e.g., DPs_2), the extraction process was similar to that of medicinal polysaccharides_1 (e.g., DPs_1), with the additional step of dialysis to remove molecules smaller than 3.5 kDa .
Determination of total sugar content
Total sugar content was measured using the protocol as follows. Specifically, glucose (Mw 180 Da) obtained from Sigma (St. Louis, MO, USA) were used to establish the calibration curve. Glucose standard solution with a concentration of was prepared. For each polysaccharide sample, medicinal polysaccharides solution with a concentration of were prepared. The mixture was heated to facilitate dissolution and then was filtered using a filter element until the solution became clear. phenol solution was added to glucose standard and medicinal polysaccharides solution. The mixture was thoroughly shaken to ensure proper mixing and stand for 30 minutes. The spectrophotometer at a wavelength of 490 nm (using an LED light source and CMOS photosensitive element) was preheated. The preheating process is typically completed by the end of the reaction, and the meter reading becomes stable. The zero-point calibration was performed with phenol solution without glucose added, and the one-point calibration with a black baffle (Supplementary Table S3). Then, the absorption rate of each sample was measured three times. The standard curve was calculated using linear regression with the data of glucose standard sample measured above. The total
sugar content was calculated by substituting the absorption rate of medicinal polysaccharides. In this experiment, with a sample concentration of , the sugar content of a sample can be calculated using the formula:
When the sugar concentration is , the absorption rate is , the standard curve is , and the sample concentration is in this experiment).
Determination of molecular weight and distribution
Molecular weight and distribution were measured according to the method as follows. Specifically, dextran standards with different molecular weights ( ) obtained from Sigma (St. Louis, MO, USA) were used to establish the calibration curve. Based on the gel column chromatography principle, there is an exponential relationship between the molecular weight and the retention time of a compound. A mathematical model with the expression is established to represent this relationship:
In this study, the molecular weight distribution of polysaccharide samples was analyzed using an HPLC system with the following parameters: Mobile phase: NaCl solution , Flow rate: , Single sampling amount: , Sample solution concentration: , Chromatographic column: TSK-gel G3000PWXL ( ) column, Column temperature: , Detector: Refractive index (RI) detector. The standard sugar samples with known molecular weights and medicinal polysaccharides samples were dissolved water with final concentration of , and filtered using a filter element until they are clear. The retention time of each sample was recorded. The standard curve was calculated using exponential regression with the data from the standard samples. Substituting the peak retention time of medicinal polysaccharides samples, molecular weight was calculated.
Determination of monosaccharide composition
Monosaccharide composition was measured according to the method , as detailed below. Specifically, the monosaccharide composition of polysaccharides samples was analyzed using HPLC with the following parameters: mobile phase: acetonitrile supplemented with phosphate buffer solution, flow rate: , single sampling amount: , chromatographic column: Cosmosil 5C18-PAQ ( ), column temperature: , detector: Ultraviolet (UV) detector. Each monosaccharide (Man, Glc, Gal, Rha, Fuc, Xyl, Ara, GalA, GlcA) obtained from Sigma (St. Louis, MO, USA) was mixed at a concentration of 10 mM . We prepared a series of monosaccharide standard solutions and calibrated the liquid chromatography system using these standards. The medicinal polysaccharides samples were completely hydrolyzed and performed pre-column derivatization with 1-phenyl-3-methyl-5-pyrazolinone. The retention time was compared with the standard monosaccharide samples to determine the types of monosaccharides present in the polysaccharide sample. Through measuring peak area ratio of each monosaccharide, the proportion of each monosaccharide in the polysaccharide sample was calculated.
Transcriptome sequencing and analysis
Bacteroides species were cultured in BHIS medium supplemented with hemin, vitamin K3, and cysteine in an anaerobic chamber (Coy; , and ) at . The bacteria were washed three times with anaerobic PBS, and the optical density at
600 nm (OD600) of the bacterial solution was adjusted to 0.1. The bacteria were then inoculated at a 1:10 ratio into a 96-well plate and cultured at . The OD600 was measured and recorded every 15 minutes using a microplate reader. After reaching the logarithmic growth phase (Supplementary Fig. 6A-C), the bacteria were collected by centrifugation at 5000 xg for 2 minutes. The collected bacteria in the logarithmic growth period were stored at before transcript sequencing.
Total RNA was extracted from the sample using the Trizol method, including DNase treatment to remove genomic DNA, and assessed for quality using the NanoDrop One (Thermo, A30221) and Agilent 4200 TapeStation (Agilent, G2991AA). Ribosomal RNA was removed using the Ribo-Zero rRNA Removal Kit (Epicenter, 15066012) designed for bacteria. For library construction, the NEBNext Ultra II Directional RNA Library Prep Kit (NEB, e7760) for Illumina was employed: mRNA was fragmented into short segments, singlestranded cDNA was synthesized using random hexamers, doublestranded cDNA was generated with dUTP substitution, purified using AMPure XP beads (Beckman, A63880), and the U-containing second strand was digested using USER enzyme (NEB, M5508). End repair, Atailing, and adapter ligation were performed on the purified cDNA, followed by fragment size selection using AMPure XP beads. PCR amplification was conducted, and PCR products were purified with AMPure XP beads to obtain the final library. Libraries were inspected using the Agilent 4200 TapeStation, and only qualified libraries were sequenced on the Illumina platform (Illumina, HiSeq 2000). Key improvements included ensuring DNase treatment during RNA extraction, using a bacterial-specific rRNA removal kit, generating strand-specific libraries, verifying adapter compatibility with prokaryotic RNA, and adjusting fragment size selection for prokaryotic transcript lengths.
After obtaining the sequencing data for each sample, it is important to assess the quality of the data and remove any low-quality reads to ensure the reliability of subsequent analyzes. The software used for quality control is fastp , and the following quality control criteria are applied: 1) Minimum base quality score of 15; 2) Sliding window size of 4 with an average base quality score greater than 20;3) Minimum read length of 75 bp ;4) Default values for other parameters. The sequencing data, after quality control, is aligned to the ribosomal RNA sequences in the Rfam database using Bowtie2 to remove rRNA reads. The remaining reads are then aligned to the reference genome using Bowtie2, resulting in a BAM file containing the alignment results. The alignment results are further processed to generate read counts using RSEM . For gene expression analysis, we utilize the read counts to identify differentially expressed genes among multiple sample groups.
Genomic annotation of Polysaccharide Utilization Locus (PUL)
PULs of Bacteroides were annotated by PULpy . It is worth noting that different species of B. uniformis can have varying numbers and architectures of PULs, and these differences can also be observed between different annotated methods, such as PULpy and PUL prediction algorithm used by PULDB. The boundaries of the PUL were further recorrected based on gene expression differences. Specifically, by using a comparative transcriptomic analysis of Bacteroides spp. cultured in minimal medium supplemented with polysaccharides or glucose, the PULs with the highest upregulation was identified. Then, the transcriptomes of the upstream and downstream genes of this PULs were compared. Genes that were consistently upregulated within and outside the predicted PULs were defined as recorrected PULs.
Construction of B. uniformis knockout mutants
Due to the difficulty of genetic manipulation in wild Bacteroides strains, we performed PUL34_Bu knockout in the type strain B. uniformis ATCC8492 (Supplementary Fig. 8). Recipient B. uniformis ATCC8492 for conjugation were cultured in BHIS medium in an anaerobic
chamber (Coy; , and ) at . Donor Escherichia coli S17-1 (Apir) were cultured in Luria-Bertani (LB) medium at . The DNA fragments (Supplementary Table 13A) of sgRNA or sgRNAhomologous arms (1 kb repair templates) were cloned into the plasmid backbone by NEBuilder HiFi DNA assembly master mix (NEB, E2621S). The primers (Supplementary Table 13B) used to amplify plasmid skeleton, sgRNA, repair template and mutant detection were synthetized by Sangon Biotech. The knock out plasmids conjugated into B. uniformis ATCC8492. And then the Bacteroides strains were diluted 1:100 in BHI medium with aTc induced for 24 h (aTc final concentration . Cultures were diluted by and -fold and spread onto BHI-aTc agar plate and incubated at for 40 h . Colonies were identified by PCR and sequencing.
Sequence and structure alignment
The phylogenetic tree of enzymes from the GH26 family was constructed with MUSLE and constructed a phylogenetic tree using the Maximum likelihood method for the 89 characterized GH26 protein sequences deposited in the CAZy database . The GH26 protein 3D structure of our B. uniformis DA183 were predicted using AlphaFold2 . The predicted local distance difference test value (pLDDT) was calculated using AlphaFold2. The catalytic domain of B. uniformis DA183 GH26 was aligned to its phylogenetic neighboring GH26 in B. ovatus ATCC8483 (GenBank accession ALJ47537.1; PDB code 6HF4) using US-align to compute the TM-score and RMSD.
Bacteroides GH26 enzyme expression and purification
Sequences coding for full-length wild-type, point mutant, and CBMtruncated Bacteroides GH26 proteins (Supplementary Table 14) were optimized for expression in E. coli, chemically synthesized (Tsingke Biotechnology Co., Ltd.), and cloned into the pET28a plasmid with a C-terminal His-tag between the Ndel and Xhol sites. The plasmids were transformed into E. coli BL21(DE3). The signal peptide boundary was predicted using SignalP (version 6.0) . The signal peptide was removed to ensure soluble expression. The CBM domain boundary was predicted using Chainsaw . E. coli BL21(DE3) cells containing the pET28a plasmid were grown in 500 mL of LB medium with kanamycin at until OD600 . Expression was induced by adding 0.2 mM IPTG, and the culture was continued for 20 h at . Cells were lysed in 50 mL of lysis buffer imidazole, pH 7.4 using a high-pressure cell crusher. The lysate was centrifuged at , for 1 h , and the supernatant was incubated with 5 mL of Ni NTA agarose resin (Yeasen, 20502ES50) for 1 h at with head-overtail rotation. The resin was poured into a gravity flow column and washed three times with 50 mL of wash buffer , imidazole, pH 7.4 ). The protein was eluted with elution buffer imidazole, pH 7.4).
Bacteroides GH26 enzyme activity assay
Konjac glucomannan, guar gum galactomannan, carob galactomannan, ivory nut mannan, barley glucan, tamarind xyloglucan was from Megazyme (Bray, Ireland). Corn cob xylan, 1,1-Kestotetraose, and 1-Kestose was from TCI. Galactosyl mannobiose was from ACMEC; Raffinose was from HUICH. Trehalose was from Huhui (Shanghai) Biotechnology Co., Ltd. The specific activity was measured using the standard DNS-reducing sugar assay using GH26 BuDA183 and GH26 BoATCC8483 and substrates (Supplementary Table 3C) in 50 mM potassium phosphate buffer, pH 6.5. The incubation time was 30 min at . The enzyme activity for point mutant and CBM truncated GH26 was measured in the same way as the standard DNS assay using 125 nM GH26 enzyme and six reactive substrates (Fig. 6A) in 50 mM potassium phosphate buffer, pH 6.5 , at . The incubation time was 30 min at .
Mannose was used to obtain a concentration standard curve (Supplementary Table 15).
Enzyme Kinetics
Michaelis-Menten kinetics was measured for 125 nM full-length wild type, CBM truncated, and the variants A 363 K in triplicate using the DNS activity assay but varying time and substrate concentration. All reactions had a total volume of and contained and konjac glucomannan (1.25, 2.5, 3.75, 5, 6.25, 7.5, 8.75 , and ). The reaction was stopped by heat to for 1 min . The obtained initial rates were used to generate Michaelis-Menten curves via nls function in R from which Km and Kcat values were estimated.
Molecular docking and visualizations
The oligosaccharide substrates were modeled using GLYCAM-Web website (Complex Carbohydrate Research Center, University of Georgia, Athens, GA (http://www.glycam.com)) or derived from the RCSB Protein Data Bank (PDB). Ligand docking was performed using AutoDock Vina (v1.2.0) employing standard parameters. The activesite of GH26 was derived from the Bo GH26 structure (PDB, 6 HF 4 ) by structural superposition with US-align . The produced nine different ligand conformations for each docking complex were analyzed with CSM-carbohydrate, out of which the conformation with the lowest delta-G (binding energy) value was selected for further studying the ligand-receptor interactions by Discovery studio visualizer .
Statistics & reproducibility
No statistical method was used to predetermine sample size. No data were excluded from the analyzes. Statistical significance for growth profile assays in Fig. 1E and Supplementary Fig. 3A was determined using a two-sided unpaired t-test between each experimental group and the negative control, with multiple comparisons adjusted using the Bonferroni method; each experiment was conducted in triplicate to ensure reliability. For comparative transcriptome analysis, differential expression was assessed using DESeq2 with a two-sided Wald test, and adjusted -values were calculated using the Benjamini and Hochberg method to control the false discovery rate (FDR); genes with an adjusted -value were considered statistically significant. Representative images from single experiments are shown for nucleic acid gel electrophoresis and protein SDS-PAGE, while data for in vitro enzyme activity characterization are presented as mean values ± SD, with assays performed in technical triplicates ( ) to ensure reliability.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All sequencing data generated in this study have been submitted to the ENA BioProject database (https://www.ebi.ac.uk/ena/browser/search) under accession number PRJEB75723. Other data are available in the Supplementary information and Supplementary Data. Source data are provided with this paper.
References
Garrett, W. S. The gut microbiota and colon cancer. Science 364, 1133-1135 (2019).
Sun, L. et al. Bile salt hydrolase in non-enterotoxigenic Bacteroides potentiates colorectal cancer. Nat. Commun. 14, 755 (2023).
Huang, J. et al. Ginseng polysaccharides alter the gut microbiota and kynurenine/tryptophan ratio, potentiating the antitumour effect of antiprogrammed cell death 1/programmed cell death ligand 1 (anti-PD-1/PD-L1) immunotherapy. Gut 71, 734-745 (2022).
Tanoue, T. et al. A defined commensal consortium elicits CD8 T cells and anti-cancer immunity. Nature 565, 600-605 (2019).
Zafar, H. & Saier, M. H. Jr Gut Bacteroides species in health and disease. Gut Microbes 13, 1848158 (2021).
Beller, Z. W. et al. Inducible CRISPR-targeted “knockdown” of human gut Bacteroides in gnotobiotic mice discloses glycan utilization strategies. Proc. Natl Acad. Sci. 120, e2311422120 (2023).
Hehemann, J.-H. et al. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 464, 908-912 (2010).
Kijner, S., Ennis, D., Shmorak, S., Florentin, A. & Yassour, M. CRISPR-Cas-based identification of a sialylated human milk oligosaccharides utilization cluster in the infant gut commensal Bacteroides dorei. Nat. Commun. 15, 105 (2024).
Panwar, D., Shubhashini, A. & Kapoor, M. Complex alpha and beta mannan foraging by the human gut bacteria. Biotechnology Advances, 108166 (2023).
Buzun, E. et al. A bacterial sialidase mediates early-life colonization by a pioneering gut commensal. Cell Host Microbe 32, 181-190. e189 (2024).
Kou, F. et al. A review of Ganoderma lucidum polysaccharides: Health benefit, structure-activity relationship, modification, and nanoparticle encapsulation. International journal of biological macromolecules, 125199 (2023).
Lin, T.-L. et al. Role of gut microbiota in identification of novel TCMderived active metabolites. Protein Cell 12, 394-410 (2021).
Yu, Y., Shen, M., Song, Q. & Xie, J. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydr. Polym. 183, 91-101 (2018).
Shan-Shan, S., Kai, W., Ke, M., Li, B. & Hong-Wei, L. An insoluble polysaccharide from the sclerotium of Poria cocos improves hyperglycemia, hyperlipidemia and hepatic steatosis in ob/ob mice via modulation of gut microbiota. Chin. J. Nat. Med. 17, 3-14 (2019).
Shi, L. Bioactivities, isolation and purification methods of polysaccharides from natural products: A review. Int. J. Biol. Macromolecules 92, 37-48 (2016).
Wu, T.-R. et al. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut 68, 248-262 (2019).
Zhang, Y. et al. Identification of the core active structure of a Dendrobium officinale polysaccharide and its protective effect against dextran sulfate sodium-induced colitis via alleviating gut microbiota dysbiosis. Food Res. Int. 137, 109641 (2020).
Martens, E. C., Chiang, H. C. & Gordon, J. I. Mucosal glycan foraging enhances fitness and transmission of a saccharolytic human gut bacterial symbiont. Cell host microbe 4, 447-457 (2008).
Lapébie, P., Lombard, V., Drula, E., Terrapon, N. & Henrissat, B. Bacteroidetes use thousands of enzyme combinations to break down glycans. Nat. Commun. 10, 2043 (2019).
Martens, E. C. et al. Recognition and degradation of plant cell wall polysaccharides by two human gut symbionts. PLOS Biol. 9, e1001221 (2011).
Foley, M. H., Cockburn, D. W. & Koropatkin, N. M. The Sus operon: a model system for starch uptake by the human gut Bacteroidetes. Cell. Mol. Life Sci. 73, 2603-2617 (2016).
Luis, A. S. et al. A single sulfatase is required to access colonic mucin by a gut bacterium. Nature 598, 332-337 (2021).
Wu, S. et al. GMrepo: a database of curated and consistently annotated human gut metagenomes. Nucleic acids Res. 48, D545-D553 (2020).
Liu, C. et al. Enlightening the taxonomy darkness of human gut microbiomes with a cultured biobank. Microbiome 9, 119 (2021).
Drula, E. et al. The carbohydrate-active enzyme database: functions and literature. Nucleic acids Res. 50, D571-D577 (2022).
Qi, H. et al. Comparisons of isolation methods, structural features, and bioactivities of the polysaccharides from three common Panax species: A review of recent progress. Molecules 26, 4997 (2021).
Kuang, M.-T. et al. Structural characterization and hypoglycemic effect via stimulating glucagon-like peptide-1 secretion of two polysaccharides from Dendrobium officinale. Carbohydr. Polym.
241, 116326 (2020).
Yin, J.-Y. et al. Separation, structure characterization, conformation and immunomodulating effect of a hyperbranched heteroglycan from Radix Astragali. Carbohydr. Polym. 87, 667-675 (2012).
Wardman, J. F., Bains, R. K., Rahfeld, P. & Withers, S. G. Carbohydrate-active enzymes (CAZymes) in the gut microbiome. Nat. Rev. Microbiol. 20, 542-556 (2022).
Yin, Y. et al. dbCAN: a web resource for automated carbohydrateactive enzyme annotation. Nucleic acids Res. 40, W445-W451 (2012).
Stewart, R. D., Auffret, M. D., Roehe, R. & Watson, M. Open prediction of polysaccharide utilisation loci (PUL) in 5414 public Bacteroidetes genomes using PULpy. bioRxiv, 421024, https://doi.org/ 10.1101/421024 (2018).
Guo, M., Shao, S., Wang, D., Zhao, D. & Wang, M. Recent progress in polysaccharides from Panax ginseng CA Meyer. Food Funct. 12, 494-518 (2021).
Li, J. et al. Purification, structural characterization, and immunomodulatory activity of the polysaccharides from Ganoderma lucidum. Int. J. Biol. macromolecules 143, 806-813 (2020).
Zheng, L. et al. CRISPR/Cas-based genome editing for human gut commensal bacteroides species. ACS Synth. Biol. 11, 464-472 (2022).
García-Bayona, L. & Comstock, L. E. Streamlined genetic manipulation of diverse bacteroides and parabacteroides isolates from the human gut microbiota. MBio 10, 01762-01719 (2019).
Almeida, A. et al. A unified catalog of 204,938 reference genomes from the human gut microbiome. Nat. Biotechnol. 39, 105-114 (2021).
Buchfink, B., Reuter, K. & Drost, H.-G. Sensitive protein alignments at tree-of-life scale using DIAMOND. Nat. methods 18, 366-368 (2021).
Zhou, N. et al. Konjac glucomannan: a review of structure, physicochemical properties, and wound dressing applications. J. Appl. Polym. Sci. 139, 51780 (2022).
Mudgil, D., Barak, S. & Khatkar, B. S. Guar gum: processing, properties and food applications-a review. J. food Sci. Technol. 51, 409-418 (2014).
Lazaridou, A., Biliaderis, C. G. & Izydorczyk, M. S. Structural characteristics and rheological properties of locust bean galactomannans: a comparison of samples from different carob tree populations. J. Sci. Food Agriculture 81, 68-75 (2001).
Ghysels, S. et al. Fast pyrolysis of mannan-rich ivory nut (Phytelephas aequatorialis) to valuable biorefinery products. Chem. Eng. J. 373, 446-457 (2019).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583-589 (2021).
Bågenholm, V. et al. A surface-exposed GH26 -mannanase from Bacteroides ovatus: Structure, role, and phylogenetic analysis of BoMan26B. J. Biol. Chem. 294, 9100-9117 (2019).
Okano, H., Kanaya, E., Ozaki, M., Angkawidjaja, C. & Kanaya, S. Structure, activity, and stability of metagenome-derived glycoside hydrolase family 9 endoglucanase with an N-terminal Ig-like domain. Protein Sci. 24, 408-419 (2015).
Liberato, M. V. et al. Molecular characterization of a family 5 glycoside hydrolase suggests an induced-fit enzymatic mechanism. Sci. Rep. 6, 23473 (2016).
Murray, P. A., Kern, D. G. & Winkler, J. R. Identification of a galactosebinding lectin on Fusobacterium nucleatum FN-2. Infect. Immun. 56, 1314-1319 (1988).
Kuhaudomlarp, S., Gillon, E., Varrot, A. & Imberty, A. LecA (PA-IL): A galactose-binding lectin from Pseudomonas aeruginosa. Lectin Purification and Analysis: Methods and Protocols, 257-266 (2020).
Bolam, D. N. et al. Mannanase A from Pseudomonas fluorescens ssp. cellulosa is a retaining glycosyl hydrolase in which E212 and E320 are the putative catalytic residues. Biochemistry 35, 16195-16204 (1996).
Eberhardt, J., Santos-Martins, D., Tillack, A. F. & Forli, S. AutoDock Vina 1.2. O: New docking methods, expanded force field, and python bindings. J. Chem. Inf. modeling 61, 3891-3898 (2021).
Nguyen, T. B., Pires, D. E. & Ascher, D. B. CSM-carbohydrate: protein-carbohydrate binding affinity prediction and docking scoring function. Brief. Bioinforma. 23, bbab512 (2022).
dos Santos, C. R. et al. Molecular insights into substrate specificity and thermal stability of a bacterial GH5-CBM27 endo-1, 4- -Dmannanase. J. Struct. Biol. 177, 469-476 (2012).
Roske, Y., Sunna, A., Pfeil, W. & Heinemann, U. High-resolution crystal structures of Caldicellulosiruptor strain Rt8B. 4 carbohydrate-binding module CBM27-1 and its complex with mannohexaose. J. Mol. Biol. 340, 543-554 (2004).
Zhao, L. et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 359, 1151-1156 (2018).
O’Keefe, S. J. et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 6, 1-14 (2015).
Sorbara, M. T. & Pamer, E. G. Microbiome-based therapeutics. Nat. Rev. Microbiol. 20, 365-380 (2022).
Gauffin Cano, P., Santacruz, A., Moya, Á. & Sanz, Y. Bacteroides uniformis CECT 7771 ameliorates metabolic and immunological dysfunction in mice with high-fat-diet induced obesity. (2012).
López-Almela, I. et al. Bacteroides uniformis combined with fiber amplifies metabolic and immune benefits in obese mice. Gut Microbes 13, 1-20 (2021).
Morita, H. et al. Bacteroides uniformis and its preferred substrate, cyclodextrin, enhance endurance exercise performance in mice and human males. Sci. Adv. 9, eadd2120 (2023).
Yan, Y. et al. Bacteroides uniformis-induced perturbations in colonic microbiota and bile acid levels inhibit TH17 differentiation and ameliorate colitis developments. npj Biofilms Microbiomes 9, 56 (2023).
Dai, W. et al. Discovery of Bacteroides uniformis F18-22 as a Safe and Novel Probiotic Bacterium for the Treatment of Ulcerative Colitis from the Healthy Human Colon. Int. J. Mol. Sci. 24, 14669 (2023).
Nie, Q. et al. Gut symbionts alleviate MASH through a secondary bile acid biosynthetic pathway. Cell 187, 2717-2734. e2733 (2024).
Zeng, P., Li, J., Chen, Y. & Zhang, L. The structures and biological functions of polysaccharides from traditional Chinese herbs. Prog. Mol. Biol. Transl. Sci. 163, 423-444 (2019).
Cuskin, F. et al. Human gut Bacteroidetes can utilize yeast mannan through a selfish mechanism. Nature 517, 165-169 (2015).
Sastre, D. E. et al. Human gut microbes express functionally distinct endoglycosidases to metabolize the same N-glycan substrate. Nat. Commun. 15, 5123 (2024).
Martens, E. C., Roth, R., Heuser, J. E. & Gordon, J. I. Coordinate regulation of glycan degradation and polysaccharide capsule biosynthesis by a prominent human gut symbiont. J. Biol. Chem. 284, 18445-18457 (2009).
Bågenholm, V. et al. Galactomannan catabolism conferred by a polysaccharide utilization locus of Bacteroides ovatus: enzyme synergy and crystal structure of a -mannanase. J. Biol. Chem. 292, 229-243 (2017).
Patnode, M. L. et al. Strain-level functional variation in the human gut microbiota based on bacterial binding to artificial food particles. Cell host microbe 29, 664-673. e665 (2021).
Odamaki, T. et al. Comparative genomics revealed genetic diversity and species/strain-level differences in carbohydrate metabolism of three probiotic bifidobacterial species. Int. J. Genomics 2015, 567809 (2015).
van Leeuwen, P. T., Brul, S., Zhang, J. & Wortel, M. T. Synthetic microbial communities (SynComs) of the human gut: design, assembly, and applications. FEMS Microbiol Rev 47, https://doi. org/10.1093/femsre/fuad012 (2023).
Cheng, A. G. et al. Design, construction, and in vivo augmentation of a complex gut microbiome. Cell 185, 3617-3636.e3619 (2022).
Parks, D. H. et al. GTDB: an ongoing census of bacterial and archaeal diversity through a phylogenetically consistent, rank normalized and complete genome-based taxonomy. Nucleic acids Res. 50, D785-D794 (2022).
Bacic, M. K. & Smith, C. J. Laboratory maintenance and cultivation of bacteroides species. Curr. Protoc. Microbiol. https://doi.org/10. 1002/9780471729259.mc13c01s9 (2008).
Liu, W., Zhu, W. Y., Yao, W. & Mao, S. Y. Isolation and identification of a lactate-utilizing, butyrate-producing bacterium and its primary metabolic characteristics. Wei Sheng Wu Xue Bao 47, 435-440 (2007).
Fischer, E. & Sauer, U. Metabolic flux profiling of Escherichia coli mutants in central carbon metabolism using GC-MS. Eur. J. Biochem 270, 880-891 (2003).
Portnoy, V. A., Herrgård, M. J. & Palsson, B. Aerobic fermentation of D-glucose by an evolved cytochrome oxidase-deficient Escherichia coli strain. Appl Environ. Microbiol 74, 7561-7569 (2008).
DuBois, M., Gilles, K. A., Hamilton, J. K. & Rebers, P. t. & Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350-356 (1956).
Dubois, M., Gilles, K., Hamilton, J., Rebers, P. & Smith, F. A colorimetric method for the determination of sugars. Nature 168, 167-167 (1951).
Sun, L. et al. Structural characterization of rhamnogalacturonan domains from Panax ginseng CA Meyer. Carbohydr. Polym. 203, 119-127 (2019).
Vidal, S., Williams, P., O’neill, M. & Pellerin, P. Polysaccharides from grape berry cell walls. Part I: tissue distribution and structural characterization of the pectic polysaccharides. Carbohydr. Polym. 45, 315-323 (2001).
Zhang, X. et al. Total fractionation and characterization of the watersoluble polysaccharides isolated from Panax ginseng CA Meyer. Carbohydr. Polym. 77, 544-552 (2009).
Kazemi, M., Khodaiyan, F., Labbafi, M., Hosseini, S. S. & Hojjati, M. Pistachio green hull pectin: Optimization of microwave-assisted extraction and evaluation of its physicochemical, structural and functional properties. Food Chem. 271, 663-672 (2019).
Feng, Z. et al. The model polysaccharide potato galactan is actually a mixture of different polysaccharides. Carbohydr. Polym. 313, 120889 (2023).
Yang, X., Zhao, Y., Wang, Q., Wang, H. & Mei, Q. Analysis of the monosaccharide components in Angelica polysaccharides by high performance liquid chromatography. Anal. Sci. 21, 1177-1180 (2005).
Liu, J. et al. Multi-level fingerprinting and cardiomyocyte protection evaluation for comparing polysaccharides from six Panax herbal medicines. Carbohydr. Polym. 277, 118867 (2022).
Chen, S., Zhou, Y., Chen, Y. & Gu, J. fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884-i890 (2018).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. methods 9, 357-359 (2012).
Li, B. & Dewey, C. N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinforma. 12, 1-16 (2011).
Dillies, M.-A. et al. A comprehensive evaluation of normalization methods for Illumina high-throughput RNA sequencing data analysis. Brief. Bioinforma. 14, 671-683 (2013).
Terrapon, N., Lombard, V., Gilbert, H. J. & Henrissat, B. Automatic prediction of polysaccharide utilization loci in Bacteroidetes species. Bioinformatics 31, 647-655 (2015).
Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic acids Res. 32, 1792-1797 (2004).
Yang, Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol. Evolution 24, 1586-1591 (2007).
Zhang, C., Shine, M., Pyle, A. M. & Zhang, Y. US-align: universal structure alignments of proteins, nucleic acids, and macromolecular complexes. Nat. methods 19, 1109-1115 (2022).
Teufel, F. et al. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 40, 1023-1025 (2022).
Wells, J. et al. Chainsaw: protein domain segmentation with fully convolutional neural networks. Bioinformatics 40, btae296 (2024).
Miller, G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31, 426-428 (1959).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing (Vienna, Austria, 2018). https://www.R-project.org/.
Visualizer, D. Discovery studio visualizer. 2. Accelrys Software Inc. (2005).
Camacho, C. et al. BLAST+: architecture and applications. BMC Bioinforma. 10, 1-9 (2009).
Acknowledgements
We would like to thank Thanh Binh Nguyen from The University of Queensland for his assistance with the calculation of delta-G (binding energy) value and scoring for the dominant docking complex using the CSM-carbohydrate tool. This work was funded by regular grants (File no. 0063/2022/A2, 0018/2024/RIB1 and 0137/2024/AMJ) supported by the Macau Science and Technology Development Fund and the Ministry of Science and Technology of the People’s Republic of China. This work was also supported by the 2020 Young Qihuang Scholar funded by the National Administration of Traditional Chinese Medicine and the FDCT Funding Scheme for Postdoctoral Researchers of Higher Education Institutions (0017/2021/APD). This work is also financially supported by the Start-up Research Grant of University of Macau (SRG2022-00020FHS), the 2023 State Key Laboratory of Quality Research in Chinese Medicine (UM) Internal Research Grant (IRG) (SKL-QRCM-IRG2023-001 & 005/2023/SKL), and the FSCPO project (type 1 and type 2) from Faculty of Health Sciences, University of Macau. This work received support from Shenzhen Key Laboratory for the Intelligent Microbial Manufacturing of Medicines (ZDSYS20210623091810032). This work is also funded by the National Natural Science Foundation of China under grant number 32201313.
Author contributions
Z.Q., H.L., and J.Y. performed experiments and analyzed data with assistance from L.Z., J.H., Z.W., C.X., W.Z., and X.X.Z.Q. wrote the paper. E.L.L., L.D., and H.Z. conceived and guided the projects. L.S., Y.Z., Y.X., and J.L. provided herb resources and discussed results and approved the paper. Y.-Z.Z., L.Y., and L.L. gave comment to the data analysis and manuscript.
Correspondence and requests for materials should be addressed to Hua Zhou, Lei Dai or Elaine Lai-Han Leung.
Peer review information Nature Communications thanks Diego Sastre, and the other, anonymous, reviewer for their contribution to the peer review of this work. A peer review file is available.