التوصيل الفيروسي لمحرر الجينوم الموجه بواسطة RNA لتحرير الخط الجرثومي بدون نقل جينات في الأرابيدوبسيس Viral delivery of an RNA-guided genome editor for transgene-free germline editing in Arabidopsis
تعديل الجينوم يحول علم الأحياء النباتية من خلال تمكين التعديلات الدقيقة على الحمض النووي. ومع ذلك، فإن توصيل أنظمة التعديل إلى النباتات لا يزال يمثل تحديًا، وغالبًا ما يتطلب طرقًا بطيئة ومحددة الجينوم مثل زراعة الأنسجة أو التحول.تعتبر الفيروسات النباتية، التي تصيب وتنتشر بشكل طبيعي إلى معظم الأنسجة، نظام توصيل واعد لمواد التحرير. ومع ذلك، فإن العديد من الفيروسات لديها سعات شحن محدودة، مما يقيّد قدرتها على حمل أنظمة CRISPR-Cas الكبيرة. هنا قمنا بتعديل فيروس اهتزاز التبغ (TRV) ليحمل إنزيم TnpB الموجه بواسطة RNA ISYmu1 وRNA الدليل الخاص به. سمحت هذه الابتكار بتحرير خالٍ من الجينات الغريبة لنبات الأرابيدوبسيس ثاليانا في خطوة واحدة، مع وراثة التعديلات في الجيل التالي. من خلال التغلب على الحواجز التقليدية لتوصيل المواد، تقدم هذه الطريقة منصة جديدة لتحرير الجينوم، مما يمكن أن يسرع بشكل كبير من التكنولوجيا الحيوية النباتية والبحث الأساسي.
الإنزيمات القابلة للبرمجة المسترشدة بـ RNA، بما في ذلك CRISPR-Cas9، تدفع التقدم في تحرير الجينوم لكل من البحث الأساسي والتكنولوجيا الحيوية. لقد سمحت القدرة على تعديل الجينومات النباتية وراثيًا بإنشاء أنماط ظاهرة مصممة بشكل عقلاني. ومع ذلك، فإن توصيل مواد تحرير الجينوم بكفاءة إلى النباتات لا يزال يمثل تحديًا كبيرًا. الاستراتيجية الأكثر شيوعًا هي ترميز محررات الجينوم المسترشدة بـ RNA (على سبيل المثال، إنزيمات CRISPR-Cas) داخل الجينات المنقولة واستخدام زراعة الأنسجة وطرق تحويل النباتات لتحقيق ذلك. النباتات المعدلة وراثيًا، وبعد ذلك تكون هناك حاجة إلى عمليات تهجين جينية لإزالة المادة المعدلة وراثيًا مع الاحتفاظ بالتعديلات.. ومع ذلك، فإن طرق تحويل النباتات الحالية محدودة بأنواع معينة من النباتات والأنماط الجينية، وغالبًا ما تتطلب وقتًا وموارد وخبرة تقنية كبيرة، ويمكن أن تسبب تغييرات غير مقصودة في الجينوم والإيبيجينوم..
أحد الأساليب لتجاوز هذه القيود هو استخدام ناقلات فيروسية نباتية لتوصيل مواد تحرير الجينوم مثل الميجانوكلياز أو نوكليازات إصبع الزنك (ZFNs) للتطفير المستهدف.. بينما استخدام
كان استخدام الميجانيوكلياز وزينفز لتحرير جينوم النباتات بواسطة الفيروسات تقدمًا ملحوظًا، إلا أن القدرة على ترميز نظام كريسبر الموجه بواسطة RNA والذي يمكن برمجته بسهولة ستكون ميزة كبيرة. بناءً على ذلك، تم تصميم عدة ناقلات فيروسية لترميز RNAs الموجهة (gRNAs) لتوصيلها إلى النباتات المعدلة وراثيًا التي تعبر بالفعل عن Cas9، مما أدى إلى تحرير الخلايا الجسدية والجرثومية ونقل التعديلات إلى الجيل التالي.لأن النباتات قد تطورت آليات للحد من العدوى الفيروسية في الأنسجة القمية وخلايا الجراثيم، فإن معظم الفيروسات نادراً ما تُنقل جنسياً.ومع ذلك، يمكن أن يسمح الغزو العابر لخلايا الميرستيم بواسطة RNAs فيروسية تشفر gRNAs بتحرير هذه الخلايا وأن تكون هذه التعديلات قابلة للنقل عبر البذور.بينما تمثل هذه الأساليب تقدمًا مهمًا، إلا أنها لا تزال تتطلب استخدام نوكليازات قد يكون من الصعب هندستها (مثل الميجانوكليازات و ZFNs)، أو نباتات معدلة وراثيًا تعبر عن بروتين إنزيم كريسبر-كاس.
استراتيجية لتجنب الحاجة إلى مواد نباتية معدلة وراثيًا هي استخدام ناقلات فيروسية ذات سعات تحميل كبيرة، قادرة على التعبير عن أنظمة تحرير موجهة بواسطة RNA كاملة (على سبيل المثال، Cas9 و gRNA). لقد حققت هذه الطريقة بعض النجاح؛ ومع ذلك، لا تزال تتطلب خطوات تجديد نباتي لأن هذه الفيروسات لا تسبب تحرير الخط الجرثومي ووراثة التعديلات.من ناحية أخرى، كان ترميز أنظمة كريسبر بالكامل في الفيروسات القادرة على النقل عبر السلالة الجرثومية تحديًا بسبب سعتها المحدودة..
لتجاوز حد حجم الشحنة هذا، استكشفنا إمكانيات TnpB، وهي فئة من الإنزيمات القاطعة الموجهة بواسطة RNA فائقة الصغر.الأحماض الأمينيةلتشفيرها في ناقل فيروس RNA نباتي. كأجداد لإنزيمات Cas، تستخدم TnpBs بشكل مشابه دليلاً RNA قابلاً للبرمجة، يسمى RNA أوميغا.RNA) ، ليتم توجيهها إلى أي موقع مستهدف وتحفيز تعديلات على الجينوم. سابقًا، تم إظهار أن TnpBs ISDra2 و ISYmu1 و ISAam1 قادرة على تحرير الجينوم المستهدف في خلايا الثدييات، و ISDra2 و ISYmu1 في خلايا نبات الأرز أحادي الفلقة.هنا اختبرنا ISDra2 و ISYmu1 و ISAam1 TnpBs لتحرير الجينوم في النبات ثنائي الفلقة، الأرابيدopsis. نظرًا لموقع الحمولة الفردي في متجه TRV الذي يُستخدم عادةً، سعينا للتعبير عن بروتين TnpB و RNA الدليل الخاص به ضمن نفس النسخة من mRNA تحت محفز واحد، مشابهًا لترتيب التعبير الطبيعي الخاص بهم..
لاختبار أنشطة TnpB وgRNA الخاص بهما المرمز في نص واحد، قمنا أولاً بالتعبير عن هذه الثلاثة TnpBs وقمنا بتقييم أنشطتها في التدخل في البلازميد الموجه بواسطة RNA في البكتيريا. قمنا بالتعبير المشترك عن TnpB وgRNA من نفس المحفز كنص واحد، مع الحفاظ على تسلسلاتها الطبيعية دون تحسين الكودونات. قمنا بمقارنة تكوينين لمنطقة الدليل 3′-: أحدهما ممتد بشكل مستمر دون وجود مُنهي لمحاكاة الحالة الطبيعية لـ TnpB، والآخر مُغلق بواسطة ريبوزيم فيروس التهاب الكبد دلتا (HDV)، كما تم استخدامه سابقًا في البكتيريا. (الشكل 1 من البيانات الموسعة). أظهرت نتائجنا أنه بدون ريبوزيم HDV، أظهر ISDra2 فقط نشاط تداخل البلازميد، بينما مع ريبوزيم HDV، أظهرت جميع TnpBs الثلاثة نشاطًا قويًا في كلا و (الشكل 1أ والشكل الموسع 2). كشفت هذه النتائج أن كاسيتات التعبير عن النسخ الفردية التي تحتوي على تسلسل ريبوزيم HDV في الـنهايات قادرة على قطع الحمض النووي البلازميدي في البكتيريا.
لاختبار كاسيت التعبير الفردي لتحرير الجينوم المستهدف في الأرابيدوبسيس، استخدمنا محفز AtUBQ10 لدفع تعبير TnpB- RNA و gRNA تستهدف منطقة جين PHYTOENE DESATURASE3 (AtPDS3)، تليها ريبوزيم HDV ومُنهِي rbcS-E9 (الشكل 1b). قمنا باختبار 20 موقع ISDra2، و10 مواقع ISYmu1 و7 مواقع ISAam1 لقدرات التحرير في خلايا بروتوبلاست الأرابيدوبسيس (الجدول التكميلي 1) . أظهرت ISDra2 و ISYmu1 تحريرًا نشطًا يتراوح من و على التوالي، كما تم قياسه بواسطة تسلسل الأمبليكون من الجيل التالي (amp-seq) (الشكل 3a من البيانات الموسعة). كانت ISAam1 أقل نشاطًا بكثير، حيث تراوحت كفاءة التحرير بين 0-0.3% (الشكل 3a من البيانات الموسعة). في المتوسط، لاحظنا كفاءات تحرير بنسبة 1% لـ ISDra2، و2.1% لـ ISYmu1 و0.1% لـ ISAam1 (الشكل 1c). تماشيًا مع التقارير السابقة، كانت ملفات إصلاح الحمض النووي تتكون من نتائج إصلاح مهيمنة على الحذف لجميع TnpBs الثلاثة (الشكل 3b من البيانات الموسعة)تظهر هذه البيانات أن ISDra2 و ISYmu1 و ISAam1 جميعها قادرة على تحرير الجينوم المستهدف في خلايا نبات الأرابيدopsis باستخدام تصميم التعبير عن النسخة الواحدة.
لتقييم التحرير الذي يتم بوساطة TnpB في النباتات المعدلة وراثياً، اخترنا ISYmu1، حيث أظهر أعلى متوسط كفاءة تحرير في خلايا بروتوبلاست الأرابيدوبسيس ولم يُظهر أي تحرير غير مستهدف في الأرز.تم اختيار اثنين من gRNAs الأكثر نشاطًا في التحرير، كل منهما يستهدف سياقًا جينيًا فريدًا. استهدف gRNA2 منطقة الترميز لجين AtPDS3، بينما استهدف gRNA12 منطقة المحفز مباشرةً upstream لجين AtPDS3. تم إنشاء نباتات متحولة عبر تحويل الغمر الزهري القياسي باستخدام نفس البلازميدات المستخدمة في تجارب البروتوبلاست.لاختبار الحساسية لدرجة الحرارة، تم زراعة النباتات المعدلة وراثياً التي تعبر عن ISYmu1 إما في درجة حرارة الغرفة أو خضعت لعلاج صدمة حرارية. قمنا باختبار التحرير في النباتات البرية (WT)، بالإضافة إلى الطفرة rna dependent rna polymerase 6 (rdr6) المعروفة بتقليل كبت الجينات المنقولة.أظهر التحليل باستخدام amp-seq كفاءة تحرير متوسطة قدرها و لـ gRNA2 في WT و rdr6، على التوالي (الشكل 1d). أظهر تحليل gRNA12 تحريرًا أكبر من gRNA2، بمتوسط التحرير في WT و في rdr6 (الشكل 1e). أظهر مقارنة كفاءة التحرير في النباتات المزروعة في درجة حرارة الغرفة مع تلك التي تلقت علاج الصدمة الحرارية تفضيلًا لزيادة درجة الحرارة لكلا الموقعين المستهدفين في خلفية WT، مما يدل على زيادات بمقدار 6.3 مرة و1.4 مرة في التحرير لـ gRNA2 وgRNA12، على التوالي (الشكل 1d، e). في rdr6، لاحظنا زيادة بمقدار 13 مرة في التحرير لـ gRNA2، ولكن تغيرًا طفيفًا في التحرير لـ gRNA12 (الشكل 1d، e). كانت نتائج التحرير من نباتات T1 المتحولة التي تعبر عن ISYmu1 تتكون من ملفات شيميرية، مهيمنة على الحذف، وإصلاح الحمض النووي (البيانات الموسعة الشكل 4). هذه البيانات توضح أن ISYmu1، المشفر كجين متحول، قادر على إجراء تحرير فعال للجينوم في نباتات الأرابيدوبسيس، وأن العلاج الحراري والطفرات الصامتة rdr6 يمكن استخدامها لزيادة كفاءة التحرير.
مدفوعين بنشاط ISYmu1 في نباتات الأرابيدopsis المعدلة وراثيًا، اختبرنا بعد ذلك ISYmu1 لتحرير الجينوم بواسطة فيروس TRV. TRV هو فيروس RNA ثنائي الجزء يتكون من TRV1 و TRV2 (الشكل 2أ). أظهرت الأعمال السابقة أنه يمكن هندسة RNA TRV2 عن طريق إدخال كاسيت تعبير الحمولة أسفل محفز فيروس البازلاء المبكر البني (pPEBV) (الشكل 2أ).. لاختبار قدرات ISYmu1 في تحرير الجينوم من خلال توصيل TRV إلى الأرابيدوبسيس، قمنا بتصميم هيكلين لشحنة TRV2. في هيكل TRV2 Architecture_A، تم استخدام tRNA كان مباشرةً في مجرى تيار تسلسل TnpB و gRNA (الشكل 2أ). في بنية TRV_B، قمنا بتضمين تسلسل ريبوزيم HDV بين الدليل و tRNA.تسلسل (الشكل 2أ). قمنا بتضمين tRNAفي كلا التصميمين كما تم إظهاره سابقًا لتعزيز حركة TRV النظامية وانتقال الأليلات المعدلة إلى الجيل التالي.
أولاً، قمنا بتقييم إمكانيات التحرير التي يسببها TRV باستخدام gRNA2 مع كل من بنية TRV2_A و بنية TRV2_B. تم اختيار gRNA2 لأنه يستهدف تسلسل ترميز AtPDS3، مما يسهل الفحص الظاهري للتحرير بسبب تبييض الضوء الأبيض للخلايا التي تحتوي على طفرات ثنائية الأليل.قمنا بتوصيل متجهات TRV لكل من WT وطفره جينية.يلعب دورًا في مسار إصلاح الكسر المزدوج غير المتجانس (NHEJ)يجب أن تكون كفاءة التحرير التي تتوسطها ISYmu1 أكبر فيالنوع الجيني إذا تم إصلاح الكسور مزدوجة الشريط الناتجة عن ISYmu1 من خلال NHEJ. تم توصيل كل بلازميد TRV2 مع بلازميد TRV1 إلى نباتات الأرابيدوبسيس باستخدام طريقة الأجروفلود.تمت ملاحظة بقع بيضاء على بعض الأوراق فيأسابيع بعد الفيضانات الزراعية، مما يشير إلى أن قطاعات من الخلايا تحتوي على طفرات ثنائية الأليل في الجين المستهدف AtPDS3 (الشكل 2ب). كشفت تحليل Amp-seq عن متوسط كفاءة تحرير يبلغ 0.1% و0% في نسيج الأوراق من WT والنباتات التي تعرضت للفيضانات الزراعية باستخدام TRV2 Architecture_A ونمت تحت درجة حرارة الغرفة، على التوالي (الشكل 2c). بالنسبة للنباتات المعالجة بالصدمات الحرارية، لاحظنا متوسط كفاءة التحرير لـ في WT و فينباتات مغمورة بالمياه مع TRV2
الشكل 1| تعبير TnpB و RNA الدليل في ترانسكريبت واحد لتحرير جينوم النبات. أ، مخططات شريطية لاختبار تداخل التعبير عن ناقلات الترانسكريبت الواحد TnpB لقص في E. coli. البيانات مأخوذة من تجارب أجريت في (الأعلى) و في (أسفل). تشير الأشرطة إلى غياب (أسود) أو وجود (أرجواني) PAM على البلازميد المستهدف. الـمحور هومقياس وحدات المستعمرات القابلة للتطبيع. الـيعرض المحور الثلاثة TnpBs التي تم اختبارها باستخدام تصميم النسخة الواحدة للتعبير بدون أو مع ريبوزيم HDV. تم حساب الانحراف المعياري للخطأ لكل تجربة، مع 3 تكرارات لكل تجربة. ب، مخطط للنسخة الواحدة للتعبير TnpB-تصميم بلازميد RNA المستخدم في تعديل جينوم النباتات. السهم الأخضر يرمز إلى محفز AtUBQ10؛ والصناديق الرمادية الداكنة تشير إلى-علم، تسلسلات NLS لـ SV40 وتسلسلات ريبوزيم HDV؛ تشير المربعات الرمادية الفاتحة إلى TnpB-RNA وتسلسلات الدليل؛ المربع الأحمر
يرمز إلى مُنهي rbcS-E9؛ السهم الأسود يشير إلى اتجاه TnpB-كاسيت تعبير RNA. ج، رسم بياني عمودي يعرض متوسط كفاءات التحرير (± خطأ معياري) لتجارب البروتوبلاست باستخدام ISDra2 و ISYmu1 و ISAam1 TnpBs. كل نقطة تمثل متوسط كفاءة التحرير (نسبة قراءات indel) لجين gRNA من البيانات الموسعة الشكل 3a، مع عدد العينات المشار إليه في أعلى الرسم. د، هـ، تحرير ISYmu1 الجسدي في نباتات T1 المعدلة وراثيًا لجين gRNA2 ISYmu1 (د) و gRNA12 ISYmu1 (هـ). يتم رسم الأنماط الجينية على طولالمحور وكفاءات التحرير (نسبة قراءات الإنديل) (± الانحراف المعياري للمتوسط) موضوعة علىالمحور. كل نقطة تشير إلى نبات T1 ترانسجيني واحد. تمثل معالجة الغرفة و HS ظروف نمو النباتات عند درجة حرارة الغرفة والصدمات الحرارية، على التوالي.
الهندسة المعمارية_A (الشكل 2ج). باستخدام TRV2 الهندسة المعمارية_B، لاحظنا متوسطاً من و التحرير في WT و ، على التوالي، للنباتات التي نمت في درجة حرارة الغرفة. بالنسبة للنباتات التي تلقت TRV2 Architecture_B وصدمة حرارية، لاحظنا متوسط التحرير كفاءة في WT و في (الشكل 2ج). تظهر هذه النتائج أن الهيكل_B، الذي يحتوي على ريبوزيم HDV، أنتج تحريرًا أعلى من الهيكل_A، وأن يمكن أن يعزز الطفرات كفاءة التحرير.
الشكل 2 | التحرير الجسدي والوراثي في الأرابيدوبسيس باستخدام TRV لتوصيل ISYmu1 TnpB و RNA الموجه المستهدف AtPDS3. أ، مخطط بلازميدات TRV1 و TRV2. تشير الأسهم الخضراء إلى بوليميراز RNA المعتمد على RNA (RDRP) ومروج pPEBV لـ TRV1 و TRV2، على التوالي؛ تشير الصناديق الرمادية في TRV1 و TRV2 إلى مكونات TRV الأصلية؛ تشير صندوق الحمولة الأحمر في TRV2 إلى موقع إما Architecture_A أو Architecture_B؛ أدناه TRV2 مخططات للمكونات، Architecture_A أو Architecture_B، المدمجة في فتحة الحمولة TRV2. ب، صورة تمثيلية لنبات يظهر قطاعات بيضاء في الأوراق (الأسهم الصفراء).أسابيع بعد تسليم TRV. ج، د، رسم بياني عمودي يعرض كفاءات التحرير الجسدي (نسبة قراءات الإنديل) (المحور) لـ ISYmu1 gRNA2 في WT والخلفيات الجينية (ج) و لـ ISYmu1 gRNA12 في WT (د). تم رسم هياكل حمولات TRV2 على طولمحور مع إما غرفة أو علاج HS. كل نقطة تمثل نباتًا فرديًا خضع لتوصيل TRV عبر الزراعة. تم حساب الانحراف المعياري للخطأ لكل تجربة. e، f، ملف إصلاح indel للحمض النووي لنبات WT فردي خضع لتوصيل TRV Cargo Architecture_B مع ISYmu1 gRNA2 (e) أو مع ISYmu1 gRNA12 (f) تحت علاج الصدمة الحرارية. الأنواع الخمسة الأكثر شيوعًا من الإضافات والحذف مدرجة على اليسار. عدد القراءات لكل إضافة وحذف مدرج على اليمين. يتم تحديد PAM بواسطة المربع الأحمر، والموقع المستهدف محدد بالمربع الأسود، في تسلسل المرجع. العدد الإجمالي للقراءات وكفاءة التحرير مدرجة أسفل كل ملف تعريف للإضافة والحذف. صورة تمثيلية من شتلات البذور البيضاء والخضراء من نبات WT تظهرتحرير جسدي باستخدام تصميم TRV2 Architecture_B مع gRNA2 الذي خضع لعلاج الصدمة الحرارية. h، لقطة شاشة لملف تتبع تسلسل سانجر من إحدى النباتات الألبينو في الشكل 3a. الأعلى: تسلسل المكمل العكسي للنمط البري. الوسط: هدف gRNA2 ISYmu1 وPAM (مربع رمادي). الأسفل: ملف تتبع ab1 يعرض حذفًا متجانسًا بطول 4 قواعد.جدول يلخص نقل الأليلات المعدلة من نباتين فرديين خضعا لتوصيل الأجسام باستخدام ISYmu1 gRNA12. تشير عمود ‘النسل المفحوص’ إلى عدد الشتلات التي تم تحديد جينومها؛ ويشير عمود ‘التعديلات ثنائية الأليل (%)’ إلى عدد الشتلات التي تحتوي على تعديلات ثنائية الأليل؛ ويشير عمود ‘التعديلات أحادية الأليل (%)’ إلى عدد النباتات التي تحمل تعديلات أحادية الأليل.
الشكل 3 | التحرير الجسدي والوراثي في الأرابيدوبسيس باستخدام TRV لتوصيل ISYmu1 TnpB وRNA الموجه المستهدف AtCHLI1. أ، صور تمثيلية لنباتات تظهر قطاعات صفراء.أسابيع بعد تسليم TRV. يتم الإشارة إلى كفاءة تحرير الجينوم المستهدف (gRNA) والتحرير الجسدي في الزاوية العلوية اليسرى من كل صورة. ب، رسم بياني عمودي يعرض كفاءات التحرير الجسدي (± الانحراف المعياري) لـ ISYmu1 gRNA4 و gRNA6 و gRNA9 في WT. يتم رسم موقع هدف gRNA على طول المحور. الـ تشير المحور إلى كفاءات التحرير (نسبة قراءات الإندل). كل نقطة تمثل نباتًا فرديًا خضع لتوصيل TRV عبر الزراعة المائية.
ج، صورة تمثيلية من شتلات النسل الصفراء والخضراء من نبات WT
تظهر 67.4% تحريرًا جسديًا باستخدام تصميم TRV2 Architecture_B مع gRNA4 الذي خضع لعلاج صدمة حرارية. د، جدول يلخص نقل الأليلات المعدلة من أربعة ونبات فردي واحد خضع لتوصيل الزراعة المائية باستخدام gRNA4 وgRNA6 على التوالي. هـ، لقطات شاشة لملفات تسلسل سانجر التمثيلية من نبات أصفر يحمل تعديلًا عند gRNA4 (أعلى) أو gRNA6 (أسفل). لكل لوحة: أعلى، تسلسل النوع البري؛ في المنتصف، هدف ISYmu1gRNA وPAM؛ أسفل، ملف تسلسل ab1 يعرض حذفًا متماثلًا.
بعد ذلك، اختبرنا gRNA12 باستخدام TRV2 Architecture_B، حيث أظهر هذا التصميم أعلى مستويات التحرير لـ gRNA2. باستخدام نفس طريقة توصيل TRV عبر الزراعة المائية إلى نباتات WT، لاحظنا متوسطًا من و كفاءة التحرير في ظروف نمو درجة حرارة الغرفة وعلاج الحرارة، على التوالي (الشكل 2د). علاوة على ذلك، أظهرت 6/57 نباتات تحريرًا أكبر من، مع 4 نباتات تظهر تحريرًا أكبر من عند استخدام علاج درجة حرارة الغرفة (الشكل 2د). مرة أخرى، أظهر تحليل نتائج الإصلاح ملفات تعريف مهيمنة للحذف لـ ISYmu1gRNA2 وgRNA12 (الشكل 2هـ، و2ف).
لاختبار نقل الأليلات المعدلة إلى الجيل التالي، قمنا أولاً بفحص نسل نبات WT يظهر تحريرًا جسديًا باستخدام تصميم TRV2 Architecture_B مع gRNA2 الذي خضع لعلاج صدمة حرارية. في المجموع، تم زراعة 2,318 بذور على ألواح MS تحتوي على سكروز. بعد 10 أيام، تم ملاحظة 68 شتلة ألبينو، مما يشير إلى طفرات ثنائية الأليل في جين PDS3 (الشكل 2ز). لتأكيد أن AtPDS3 تم تحويره، قمنا بإجراء تسلسل سانجر على الشتلتين البيضاء الموضحتين في الشكل 2ز، مما كشف أن كلا النباتين متماثلان لحذف 4-زوج قاعدي (الشكل 2ح).
للمزيد من تحديد نقل الأليلات المعدلة، أظهر تسلسل الأمب على 209 شتلات (41 ألبينو و168 خضراء) أن جميع الشتلات الألبينو تحتوي على طفرات ثنائية الأليل، مع كون الغالبية من الطفرات هي الحذف 4-زوج قاعدي الذي لوحظ في الشكل 2ح (الجدول التكميلي 2). من بين 168 شتلة خضراء، كان 8 منها مغايرة الأليل (حذف 4-زوج قاعدي في WT) (الجدول التكميلي 2).
بعد ذلك، قمنا بتحديد نقل الأليلات المعدلة من سطرين فرديين، النبات 54 ( تحرير جسدي) والنبات 69 (77.1% تحرير جسدي)، اللذان خضعا للزراعة المائية باستخدام gRNA12 TRV2 Architecture B في ظروف درجة حرارة الغرفة. كما هو متوقع، لم نلاحظ أي شتلات ألبينو، ربما لأن موقع الهدف هذا يقع upstream من موقع بدء النسخ AtPDS3. باستخدام تسلسل سانجر، قمنا بتحليل الجينومات لـ 148 و75 شتلة نسل من النباتات 54 و69، على التوالي. كشف تحليل تسلسل سانجر للنسل من النبات 54 عن نباتات معدلة ثنائية الأليل و نباتات معدلة أحادية الأليل (الشكل 2ي، الجدول التكميلي 2 والشكل 5أ من البيانات الموسعة). بالنسبة للنبات 69، لاحظنا نقلًا أعلى للأليلات المعدلة، بلغ إجماليه نباتات معدلة ثنائية الأليل و نباتات معدلة أحادية الأليل
(الشكل 2ي، الجدول التكميلي 2 والشكل 5ب من البيانات الموسعة). هذه البيانات توضح وراثة التعديلات الناتجة عن توصيل TRV لـ ISYmu1 في موقعين مستهدفين متميزين.
لاختبار قابلية تطبيق هذه الطريقة على موضع آخر، قمنا بتصميم ستة gRNAs لـ ISYmu1 تستهدف جين AtCHLl1 (AT4G18480) (الجدول التكميلي 1). تم اختيار AtCHLl1 بسبب القراءة الظاهرة الصفراء الفينوتيبية التي تظهرها نباتات AtCHLl1 المطفرة متماثلة الأليل. تم استخدام طريقة الزراعة المائية لتوصيل فردي لمتجهات TRV Architecture_B التي تستهدف كل من المواقع الستة لـ AtCHLl1. تعرضت النباتات إما لظروف نمو درجة حرارة الغرفة أو صدمة حرارية كما هو موضح سابقًا. بعد حوالي أسبوعين من الزراعة المائية، لاحظنا قطاعات صفراء على بعض النباتات المصابة بـ TRV ISYmu1 gRNA4 وgRNA6 وgRNA9 (الشكل 3أ). لتحديد التحرير الجسدي، تم جمع عينات الأنسجة من النباتات المصابة بـ TRV التي تستهدف gRNA4 وgRNA6 وgRNA9 لتحليل الأمب. لاحظنا متوسطًا من و التحرير الجسدي لـ gRNA4 وgRNA6 وgRNA9 لظروف درجة حرارة الغرفة، على التوالي (الشكل 3ب). بالنسبة للنباتات التي خضعت لصدمة حرارية، اكتشفنا متوسط تكرار التحرير من و0% لـ gRNA4 وgRNA6 وgRNA9، على التوالي (الشكل 3ب). علاوة على ذلك، أظهرت 4/47 (8.5%) و4/12 (33.3%) من النباتات المصابة بـ ISYmu1gRNA4 تحريرًا جسديًا أكبر من لعينات درجة حرارة الغرفة وصدمة الحرارة، على التوالي (الشكل 3ب).
بعد ذلك، قمنا بفحص نسل النباتات المصابة بـ TRV التي تستهدف gRNA4 وgRNA6 لتحديد نقل الأليلات المعدلة. تم زراعة الشتلات على ألواح MS تحتوي على سكروز، وبعد 10 أيام لاحظنا شتلات صفراء، متوافقة مع الجينوم الطفري ثنائي الأليل لهذا الجين (الشكل 3ج). في المجموع، لاحظنا نسل أصفر من النباتات المصابة بـ TRV ISYmu1 gRNA4 (الشكل 3د). بالنسبة لنسل نبات مصاب بـ gRNA6، لاحظنا عددًا أقل من الشتلات الصفراء، بلغ إجماليه 2/145 (1.4%) (الشكل 3د). كشف تسلسل سانجر أن جميع النباتات الصفراء تحمل طفرات ثنائية الأليل في موقع الهدف gRNA4 أو gRNA6 (الشكل 3هـ، الشكل 5ج من البيانات الموسعة والجدول التكميلي 2). بعد ذلك، كشف تسلسل سانجر أن 9/143 () و1/137 (0.7%) من الشتلات الخضراء من النباتات 2154_12-26_HS (gRNA4) و2156_12-65 (gRNA6)، على التوالي، تحتوي على تعديلات أحادية الأليل (الشكل 3د والجدول التكميلي 2). تشير هذه البيانات إلى أن التحرير الذي يتم بوساطة TRV باستخدام ISYmu1 قادر على توليد طفرات جسدية مستهدفة في ثلاثة مواقع مستهدفة من جين AtCHLl1، وأن الأليلات المعدلة يمكن نقلها إلى الجيل التالي.
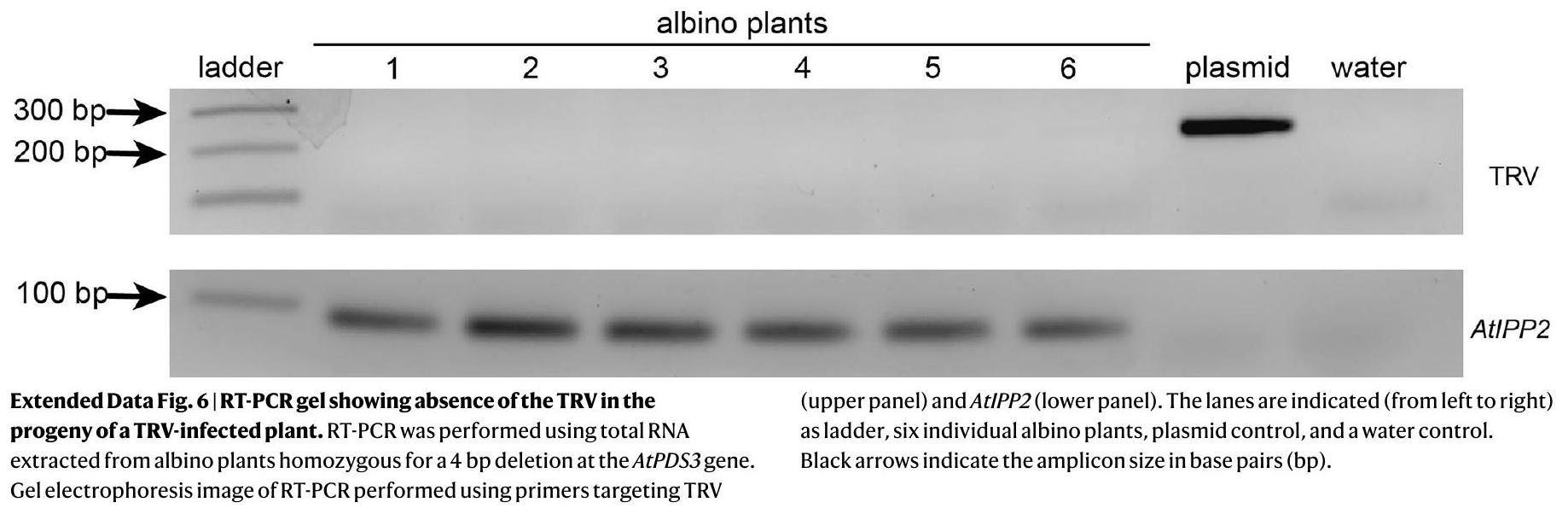
لقد تم إثبات أن TRV لا يتم نقله إلى الجيل التالي بعد تلقيح النباتات بالزراعة المائية. لتأكيد أن TRV لم يكن موجودًا في نسل نبات مصاب بـ TRV، تم إجراء RT-PCR على 5 نباتات ألبينو تحمل حذفًا متماثلًا 4-زوج قاعدي في AtPDS3. متسقة مع الأدبيات، لم يتم الكشف عن TRV في أي من النباتات الألبينو (الشكل 6 من البيانات الموسعة). تشير هذه البيانات إلى أن التعديلات ثنائية الأليل الناتجة عن TRV باستخدام ISYmu1 وراثية وخالية من الفيروسات.
لتقييم التحرير خارج الهدف، قمنا بمسح 3 نباتات ألبينو فردية تحمل طفرات ثنائية الأليل تم إنشاؤها بواسطة ISYmu1 TRV2 Architecture_B gRNA2. تم إجراء تسلسل الجينوم الكامل لتوليد متوسط من تغطية، مع تغطية أكثر من من الجينوم بواسطة قراءات موجهة (الجدول التكميلي 3). في جميع العينات الثلاث، أكدنا الطفرات المستهدفة في جين AtPDS3، كما تم تحديدها سابقًا باستخدام الأمب. بالإضافة إلى ذلك، وجدنا عددًا كبيرًا من الاختلافات المتغيرة مقارنةً بجينوم المرجع Col-O سواء في النباتات الضابطة أو المعدلة (الجدول التكميلي 4)، مما يشير إلى أن معظم المتغيرات المكتشفة ناتجة عن طفرات عفوية موجودة في سلالة الأربيذوبس لدينا. لفحص المتغيرات التي قد تكون ناجمة عن تحرير ISYmu1 خارج الهدف، تمت تصفية جميع المتغيرات في النباتات المعدلة مع المتغيرات الموجودة بالفعل في الخلفية الضابطة. تم أيضًا تصفية المتغيرات التي كانت تغطيتها أقل من 30 مرة. تم التحقق يدويًا من المتغيرات المتبقية لأي استدعاء خاطئ للمتغيرات الإيجابية. في 3 نباتات ألبينو قمنا بتسلسلها، تم الكشف عن 5 و5 و4 متغيرات، وهذه المتغيرات جميعها خارج المواقع المحتملة المتوقعة خارج الهدف بناءً على تشابه التسلسل
إلى تسلسل gRNA2 لـ AtPDS3 (الأشكال 7-10 من البيانات الموسعة، والجدول التكميلي 4 و5. بما يتماشى مع تحليل ISYmu1 خارج الهدف المبلغ عنه في الأرز وخلايا الإنسان، توضح هذه البيانات بشكل أكبر الخصوصية العالية لموقع الهدف لـ ISYmu1.
كان الهدف طويل الأمد لعلماء النبات هو تطوير وسائل سريعة وسهلة لتحرير الجينومات النباتية دون الحاجة إلى زراعة الأنسجة والتحويل الجيني. مؤخرًا، تم إثبات مستويات منخفضة من التحرير الجيني الوراثي الخالي من زراعة الأنسجة من خلال توصيل Cas9 وgRNA إلى Nicotiana benthamiana باستخدام فيروس بقعة التبغ (TRSV)لقد حسّنوا القابلية للتوريث من خلال توصيل Cas9-gRNA TRSV مع تسلسل كتم الجينات المستحث بواسطة فيروس rdr6 على فيروس التفاح الكروي الكامن (ALSV).هنا قمنا بتطوير نهج مبسط وسهل الاستخدام باستخدام محرر الجينوم TnpB المحدد للموقع فائق الصغر، ISYmu1، مع فيروس اهتزاز التبغ، من أجل تحرير جينوم النباتات القابل للتوريث. يجب أن تسرع هذه النتائج من تحرير الجينوم عالي الإنتاجية لكل من الأبحاث الأساسية والتطبيقية. نتوقع أن يكون هذا النهج قابلاً للتطبيق على TnpBs جديدة أخرى، ومجموعة متنوعة من الناقلات الفيروسية وعدد من أنواع النباتات لتحرير الجينوم. وقد كشفت الأعمال الأخيرة عن العديد من أنظمة TnpB من مصادر ميكروبية متنوعة، بما في ذلك الإنزيمات ذات الخصوصيات الفريدة لتسلسل الجوار المحيط بالبروتوسبيسر (PAM).، مما يمكن أن يزيد من نطاق تسلسل الحمض النووي المستهدف الذي يمكن تعديله باستخدام هذا النهج. الفيروس TRV المستخدم في هذه الدراسة له نطاق مضيف واسع يتجاوز 400 نوع، بما في ذلك العديد من النباتات الباذنجانية مثل الطماطم، والنباتات الزينة والمحاصيل الأخرى.. بالإضافة إلى ذلك، الفيروسات النباتية ذات سعات الحمولة المماثلة، مثل فيروس البطاطسومن المحتمل أن يكون فيروس الفسيفساء الشريطي الشعير مناسبًا لهذا النهج، حيث تم إثبات أنه قادر على تحرير الجينات الوراثية بواسطة الفيروس من خلال توصيل gRNA إلى نباتات ترانسجينية تعبر عن Cas9.علاوة على ذلك، لأن هذه الطريقة يمكن أن تخلق قطاعات من الأنسجة تحتوي على تعديلات ثنائية الأليل جسدية، فقد تخدم أيضًا كأداة لتمكين دراسة الجينات التي تسبب الموت الجنيني أو آثار متعددة الأوجه شديدة كطفرات متماثلة الزيجوت. أخيرًا، بالإضافة إلى كونها أداة مهمة لتكنولوجيا المحاصيل الحيوية، يمكن أن يمكّن التسليم الفيروسي لـ TnpBs من إجراء فحوصات CRISPR عالية الإنتاجية في أنواع النباتات النموذجية مثل الأرابيدopsis، مما يفتح المزيد من إمكانيات الاكتشاف الجيني.
طرق
البلازميدات المستخدمة في هذه الدراسة
تم إنتاج البلازميدات المستخدمة في اختبار البكتيريا على النحو التالي. تحتوي الكاسيت التعبيري الفردي على TnpB وتم تصنيع تسلسل RNA ككتل جينية من تقنيات الحمض النووي المتكاملة (IDT) وتم استنساخها باستخدام إنزيم تقييد Bsmbl (E1602S) في متجه (مقاومة الكلورامفينيكول) تحت محفز قابل للتحفيز بالتتراسيكلين (TetR/pTet) لصنع TnpB- بلازميد RNA. تم استنساخ مواقع الهدف مع تسلسلات PAM المختلفة باستخدام إنزيم تقييد BbsI (R3539S) في متجه (مقاومة للأمبيسيلين/الكاربينسيلين).
تم إنشاء البلازميدات لتجارب البروتوبلاست والغمر الزهري في استراتيجية استنساخ من خطوتين. في الخطوة الأولى، تم استخدام تسلسلات ترميز البروتين ISDra2 و ISYmu1 و ISAam1 وتم تصنيع RNAs ككتل جينية بواسطة IDT. ثم، بدءًا من المتجه pC1300_pUB10_pcoCASphi_E9t_MCS_version2، استخدمنا تجميع الحمض النووي NEBuilder HiFi (رقم الكات E2621) وPCR لتجميع TnpB- كتل الجينات RNA في ناقلات التعبير النباتية مع سمّ إدراج محاط بمواقع PaqCI مباشرةً بعدهيكل RNA وسلسلة ريبوزيم HDV السابقة. تم تحويل تفاعلات HiFi بعد ذلك إلى خلايا CcdB Survival 2 T1R Competent Cells (A10460) للحصول على المتجهات الوسيطة pMK003 (ISDra2) وpMK025 (ISYmu1) وpMK024 (ISAam1) لتسهيل استنساخ تسلسل الدليل (الجدول التكميلي 6). في الخطوة الثانية، تم تصنيع تسلسلات الدليل كخيوط علوية وسفلية فردية مع زوائد من 4 أزواج قاعدية من IDT، وتم فوسفتها ودمجها، ثم استخدمت للتجميع باستخدام إنزيم NEB PaqCI (R0745) (الجدول التكميلي 7). عند تحويلها إلى E.coli NEB10-beta القابلة للتنافس (C3019)، كانت المتجهات لا تزال تحتوي على الالجين سيقتل الخلايا، تاركًا فقط المتحولين الذين يمتلكون ناقلات التعبير النباتية TnpB التي تم تجميعها بنجاح وتحمل تسلسل RNA الدليل.
تم إنشاء متجهات TRV المستهدفة لـ AtPDS3 باستخدام بلازميد pDK3888 TRV2 كمتجه أساسي.تم استخدام NEBuilder HiFi DNA Assembly (E2621) لاستنساخ ISYmu1gRNA2 Architecture_A و ISYmu1 gRNA2 Architecture_B و ISYmu1 gRNA12 Architecture_B في فتحة الحمولة TRV2. أولاً، تم هضم pDK3888 باستخدام NEB Zral (R0659) و NEB PmII (R0532) و NEB Quick CIP (M0525) طوال الليل، وتم تنقيته باستخدام عمود تنقية Qiagen QiaQuick (28104). بعد ذلك، تم إجراء ثلاث تفاعلات PCR لتكبير القطع اللازمة لـ NEBuilder HiFi DNA Assembly، تلتها عملية تنقية باستخدام عمود تنقية Qiagen QiaQuick (28104) (الجدول التكميلي 8). ثم، تم استخدام البلازميد pDK3888 المهضوم والمنقى وقطع PCR المنقاة لتجميع البلازميد النهائي TRV2 باستخدام NEBuilder HiFi DNA Assembly وفقًا لبروتوكول الشركة المصنعة. أخيرًا، تم تحويل تفاعل NEBuilder HiFi DNA Assembly إلى E. coli NEB 10-beta القابلة للتحول (C3019). تم إنشاء متجهات TRV2 المستهدفة لـ AtCHLl1 باستخدام تجميع golden-gate.تم فوسفة أوليغو اثنين يتوافقان مع موقع الهدف ودمجهما. ثم تم استخدام الحمض النووي مزدوج الشريطة المدمج في تفاعل بوابة الذهب PaqCI (R0745S) مع pMK435.تم تحويل تفاعل البوابة الذهبية إلى بكتيريا E. coli (C3019) المعتمدة على NEB 10-beta. تم تأكيد البلازميدات الصحيحة باستخدام تسلسل البلازميد الكامل من Primordium. يمكن العثور على البلازميدات ووصفها في الجدول التكميلي 6.
اختبار التداخل البكتيري
لإجراء اختبار التداخل البكتيري، قمنا بالتشارك في نقل 100 نانوجرام من TnpB- بلازميد RNA و 100 نانوجرام من البلازميد المستهدف إلى من خلايا E. coli المعتمدة على NEB 10-beta (C3020K). على وجه التحديد، يحتوي البلازميد المستهدف على موقع مستهدف إما يحيط بـ PAM الكنسي (TTGAT لـ ISYmu1 و ISDra2 و TTTAA لـ ISAam1) أو يحيط بـ PAM غير الكنسي (GGGGG). تم استعادة الخلايا في 1 مل من وسائط NEB 10-Beta Stable/Outgrowth (B9035S) لمدة ساعة واحدة. بعد الاستعادة، تم إعداد سلسلة من تخفيفات 5 أضعاف من ثقافة الاستعادة. كل تخفيفتم وضعه على أطباق LB-أجار تحتوي على مضادين حيويين مزدوجينكلورامفينيكولالكاربينيسيلين و2 نانومتر من الأنهيدروتتراسيكلين) وعلى لوحات التحكم مع مضاد حيوي واحد (كلورامفينيكول و 2 نانومتر من الأنهيدروتتراسيكلين). إذا لم تكن هناك مستعمرات مرئية على لوحات التخفيف التسلسلي،تم زراعة 1 مل من ثقافة الاسترداد بالكامل على طبق المضاد الحيوي المزدوج لتعزيز حساسية الكشف. تم ترك الأطباق طوال الليل في إما أو تم عد وحدات تشكيل المستعمرات (c.f.u.s) على جميع الأطباق في صباح اليوم التالي. تم حساب وحدات c.f.u.s المعدلة من خلال أخذ نسبة وحدات c.f.u.s على الأطباق التي تحتوي على مضاد حيوي مزدوج إلى وحدات c.f.u.s على الأطباق التي تحتوي على مضاد حيوي واحد. تم مقارنة وحدات c.f.u.s المعدلة في ظروف PAM التقليدية بتلك في ظروف PAM غير التقليدية. تم إجراء التجارب ثلاث مرات.
مواد النبات وظروف النمو
لإعداد البروتوبلاست، تم تعليق بذور نوع كولومبيا من الأرابيدوبسيس (Col-0) فيمحلول الأجاروز واحتفظ به فيفي الظلام لمدة 3 أيام للتقسية. بعد التقسية، تم زراعة البذور على أقراص جيفي ونمت تحتفترة ضوء/ظلام مع ظروف إضاءة منخفضة فيلمدة 3-4 أسابيع.
لإنشاء نباتات معدلة وراثيًا، تم استخدام نوع الإيكوتيب Arabidopsis Col-O.تم الحصول على النمط الجيني (SALK_123114) من مختبر فنغ زانغ في جامعة مينيسوتا. تم إنشاء النمط الجيني rdr6 باستخدام تقنية كريسبر-كاس9، مما أدى إلى حذف بطول 616 قاعدة في جسم الجين rdr6. تم تنفيذ تحويل الغمر الزهري وفقًا للبروتوكول كما هو موضح سابقًا باستخدام سلالة أجرومكتريوم AgI0.تم فحص النباتات المتحولة T1 باستخدامأطباق معالهيروجوميسين ب تحتدورة الضوء/الظلام فيبعد أسبوع واحد، تم نقل الشتلات المعدلة وراثيًا التي اجتازت الاختيار إلى التربة ونقلها إلى الدفيئة ) لبقية دورة حياتهم.
للتجارب الزراعية المائية، تم زراعة البذور المعقمة علىأطباق أغار MS وتم طبقتها لمدة 5 أيام. بعد 5 أيام، تم نقل البذور إلى غرفة نمو وزراعتها تحتدورة الضوء/الظلام فيلمدة 8-10 أيام. ثم تم استخدام الشتلات لتوصيل TRV.
تم تعريض مجموعة فرعية من نباتات T1 المعدلة وراثيًا والنباتات التي خضعت لعملية الأجروفlood لعلاج صدمة حرارية تم تعديله من المرجع 37. ثم تم زراعة الشتلات التي اجتازت الاختيار أو خضعت للأجروفlood في التربة ونمت في دفيئة. ) لمدة أسبوع. بعد أسبوع، استمرت النباتات التي لم تتلقَ علاج الصدمة الحرارية في النمو في الدفيئة ( )؛ ومع ذلك، فإن النباتات التي خضعت لعلاج الصدمة الحرارية تعرضت لـ تعرض للحرارة فيكل يوم لمدة 5 أيام، تليها يومان من التعافي في دفيئةاستمر نظام الصدمة الحرارية هذا لمدة أسبوعين.
عزل البروتوبلاست والتحويل الجيني
تم عزل بروتوبلاست الميزوفيل من نبات الأرابيدوبسيس كما هو موصوف سابقًا.تم إجراء نقل البلازميد إلى بروتوبلاستات الأرابيدopsis باستخدام البلازميد وفقًا للمرجع 35. تم تحديد تركيزات البلازميدات باستخدام مقياس الطيف الضوئي نانو دروب. تم إضافة البلازميدات إلى قاع كل أنبوب نقل، وتم تكملة حجم البلازميدات بالماء للوصول إلى البروتوبلاستتمت الإضافة، تلتهامن بولي إيثيلين جلايكول (PEG) الطازج والمعقم –الحل. تم خلط العينات عن طريق النقر برفق على الأنابيب وتم تحضينها في درجة حرارة الغرفة لمدة 10 دقائق. بعدتم إضافة محلول W5 وخلطه مع البروتوبلاست عن طريق قلب الأنبوب من مرتين إلى ثلاث مرات لإيقاف عملية النقل الجيني. بعد ذلك، تم جمع البروتوبلاست عن طريق الطرد المركزي للأنابيب عند قوة طرد مركزي نسبية (RCF) تبلغ 100 لمدة دقيقتين وإعادة تعليقها في 1 مل من محلول WI. ثم تم زرع خلايا البروتوبلاست في أطباق 6 آبار مسبقة الطلاء بـمصل العجل. تم حضانة خلايا البروتوبلاست في أطباق الـ 6 آبار عندلمدة 48 ساعة. خلال فترة الحضانة التي استمرت 48 ساعة، تعرضت خلايا البروتوبلاست لـعلاج الصدمة الحرارية لمدة ساعتين بعد 16 ساعة من النقل الجيني. بعد 48 ساعة من النقل الجيني، تم جمع البروتوبلاستات لاستخراج الحمض النووي الجيني.
توصيل TRV إلى شتلات الأرابيدوبسيس
تم تنفيذ تسليم TRV كما تم وصفه سابقًاتم إدخال متجهات TRV1 و TRV2 أولاً في سلالة GV3101 من الأغروباكتيريوم. ثم تم زراعة الأغروباكتيريوم الحاملة لمتجهات TRV في 200 مل من مرق الليزوجين (LB) مع المضادات الحيوية لمدة 18 ساعة فيتمت طرد ثقافات الأجروبكتيريوم لمدة 20 دقيقة عندتم التخلص من LB وتم إعادة تعليق خلايا Agrobacterium في 200 مل من الماء المعقم. تم طرد Agrobacterium المعاد تعليقه في جهاز الطرد المركزي لمدة 10 دقائق عندتم التخلص من السائل العلوي وتم إعادة تعليق الرواسب في محلول معقم للاختراق الزراعي يحتوي على10 مللي مول 2 -(حمض الإيثان سلفونيك “-مورفولين” وأسيتوسيرينجون إلى الكثافة الضوئيةثم تم حضن خلايا الأجروبكتيريوم فيلمدة 3 ساعات مع اهتزاز بطيء. بعد 3 ساعات، تم خلط الأجروبكتيريوم الحامل لـ TRV1 و TRV2 بنسبة 1:1، وتم توصيل 15 مل من هذا المزيج بنسبة 1:1 من TRV إلى الشتلات التي تتراوح أعمارها بين 8-10 أيام. بعد 4 أيام من زراعة الأجروفlood، تم نقل الشتلات إلى التربة.
فحص نسل النباتات المصابة بفيروس TRV للبحث عن التعديلات
تم حصاد البذور من النباتات المصابة بفيروس TRVأسابيع بعد تسليم TRV. تم زراعة البذور فيألواح MS المدعمة بـ السكروز وتم تخزينه في في الظلام لمدة 5 أيام للتصنيف. بعد 5 أيام، تم نقل البذور إلى غرفة نمو وزراعتها تحتدورة الضوء/الظلام فيلـأيام. بعد ذلك، تم أخذ عينة من مجموعة فرعية من النباتات للتسلسل الجيني. تم أخذ قطعة واحدة من نسيج الورقة، وتم استخراج الحمض النووي باستخدام Invitrogen Platinum Direct PCR Universal Master Mix (A44647500) وفقًا لتعليمات الشركة المصنعة. ثم تم استخدام الحمض النووي لتسلسل amp-seq أو تسلسل سانجر باستخدام البرايمرات المدرجة في الجدول التكميلي 9.
تسلسل الأمبليكون من الجيل التالي
تم استخراج الحمض النووي من عينات البروتوبلاست باستخدام مجموعة كياجين DNeasy النباتية الصغيرة (كياجين، 69106). تم جمع الأنسجة من النباتات المعدلة وراثيًا عن طريق أخذ عينات وتجميع أنسجة الأوراق من 3 أوراق عشوائية على نبات واحد بعد 3 أسابيع من زراعتها في التربة. بالنسبة للنباتات التي خضعت لعملية الأجروفlood، تم أخذ عينات من أنسجة الأوراق عن طريق جمع وتجميع الأنسجة من 3 أوراق عشوائية (ومع ذلك، إذا كانت القطاعات البيضاء أو الصفراء مرئية، تم أخذ عينات منها) على نبات واحد بعيدًا عن موقع توصيل TRV بعد 3 أسابيع من زراعتها في التربة. بمجرد جمع عينات الأنسجة، تم تجميدها فيبين عشية وضحاها. ثم تم طحن العينات واستخراج الحمض النووي باستخدام مجموعة Invitrogen Platinum Direct PCR Universal Master Mix (A44647500) وفقًا لتعليمات الشركة المصنعة. بالنسبة لنسل النباتات التي تعرضت للفيضانات الزراعية، تم أخذ عينة من ورقة واحدة وتم استخراج الحمض النووي باستخدام مجموعة Invitrogen Platinum Direct PCR Universal Master Mix (A44647500) وفقًا لتعليمات الشركة المصنعة. ثم تم استخدام الحمض النووي في تسلسل الأمبليكون من الجيل التالي.
وفقًا للمرجع 35، تم تحديد كفاءة التحرير باستخدام تسلسل الجيل التالي أحادي النهاية على منصة Illumina NovaSeqX. تم إعداد المكتبات من خلال طريقة تضخيم PCR ذات الخطوتين. في الجولة الأولى من التضخيم، تم تضخيم كل موقع مستهدف باستخدام بادئات تحيط بالموقع المستهدف (الجدول التكميلي 9). بعد 25 دورة من التضخيم، تم تنظيف التفاعلات باستخدامتنقية حبات Ampure XP (بيكمان كولتر، A63881). بعد ذلك، مرت كل عينة بـ 12 دورة إضافية من التضخيم باستخدام بادئات فهرسة إلومينا. تم تنظيف العينات باستخدامتنقية حبات أمبيور XP. تم فحص العينات للتأكد من نقائها علىهلام الأجاروز، تم قياسه باستخدام مقياس الطيف الضوئي نانو دروب، تم تطبيعه وتجميعه.
تحليل تسلسل الأمبليكون من الجيل التالي
تم إجراء تحليل تسلسل الأمبليكون وفقًا للمرجع 35. تم استخدام قراءات نهاية واحدة للتحليل. تم تقليم القراءات باستخدام إعدادات Trim Galore الافتراضية. تم تعيين القراءات المتبقية إلى منطقة الجينوم المستهدفة باستخدام محاذي BWA (الإصدار 0.7.17، خوارزمية BWA-MEM). تم استخدام ملفات bam المرتبة والمفهرسة كملفات إدخال لمزيد من التحليل باستخدام حزمة CrispRvariants R (الإصدار 1.14.0). تم تصدير كل نمط طفرات مع عدد القراءات المقابل باستخدام حزمة CrispRvariants R. بعد تقييم جميع عينات التحكم، تم وضع معيار لتصنيف القراءات على أنها معدلة: فقط القراءات التي تحتوي على “-bp حذف أو إدخال (indel) لنفس النمط (indels بنفس الحجم تبدأ من نفس الموقع) معتم احتساب عدد القراءات من عينة كقراءات معدلة. كما تم تصفية المتغيرات أحادية النوكليوتيد.
تحليل خارج الهدف
تم إجراء تحليل خارج الهدف كما تم وصفه سابقًاتم استخراج الحمض النووي من شتلات الأرابيدوبسيس الفردية باستخدام مجموعة كيوجن دي إن إيزي النباتية الصغيرة وتم تقطيعها إلىتمت معالجة الحجم باستخدام جهاز سونكيتور Covaris. تم إعداد المكتبة باستخدام مجموعة Tecan Ovation Ultralow V2 لتسلسل الحمض النووي. بالنسبة لاستدعاء المتغيرات، تم محاذاة قراءات WGS إلى الجينوم المرجعي TAIR10 باستخدام BWA mem (الإصدار 0.7.17).مع المعلمات الافتراضية. GATK (4.2.0.0)تم استخدام MarkDuplicatesSpark لإزالة قراءات النسخ المكررة الناتجة عن تفاعل البوليميراز المتسلسل (PCR). ثم تم استخدام GATK HaplotypeCaller لاستدعاء المتغيرات الخام. تم تصفية تعدد الأشكال النوكليوتيدية المفردة (SNPs) الخام مع QD < 2.0، FS > 60.0، MQ < 40.0 و SOR > 4.0. تم تصفية InDels الخام مع QD < 2.0، FS > 200.0 و SOR.واستخدمت لإعادة معايرة درجة الجودة الأساسية. تم تطبيق ملف البام المعاير لاحقًا على GATK و Strelka (الإصدار 2.9.2) لاستدعاء SNPs/InDels. تم استخدام SNPs/InDels التي تم استدعاؤها بواسطة كل من GATK و Strelka فقط لمزيد من التصفية. تقاطع SNPs/InDel التي تم استدعاؤها بواسطة GATK مع Strelka (الإصدار 2.9.2)تم الحصول عليه باستخدام BedTools (الإصدار 2.26.0)تم تصفية SNPs/InDel باستخدام خلفية النوع البري باستخدام BedTools (الإصدار 2.26.0). تم تصفية المتغيرات التي كانت تغطيتها أقل من 30.
RT-PCR
تم استخراج RNA الكلي من نباتات النسل المصابة بـ TRV باستخدام مجموعة Zymo Research Direct-zol RNA MiniPrep (R2052). تم تحويل RNA الكلي إلى cDNA باستخدام مجموعة Invitrogen SuperScript IV VILO Master Mix (11766050). تم إجراء التحكم في RT-PCR باستخدام بادئات تستهدف جين AtIPP2 (الجدول التكميلي 10). تم إجراء PCR للتحقق من وجود/عدم وجود متجه TRV باستخدام SP9238 و SP9239 (الجدول التكميلي 10).تم إجراء تفاعل البوليميراز المتسلسل باستخدام مجموعة نيو إنجلاند بيو لابز Q5 عالية الدقةماستر ميكس (M0492L) وفقًا لتعليمات الشركة المصنعة، باستخدام من cDNA في تفاعل. تضمنت ظروف تفاعل البلمرة المتسلسل (PCR) خطوة الت denaturation الأولية لـ، ) و لمدة دقيقتين. منتجات تفاعل البوليميراز المتسلسل (PCR) ) تم تحليلها باستخدام الرحلان الكهربائي لهلام الأجاروز.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة ناتشر المرتبط بهذه المقالة.
توفر البيانات
جميع بيانات تسلسل الأمبليكون التي تم إنتاجها في هذه الدراسة متاحة في أرشيف تسلسل NCBI تحت مشروع البيولوجيا PRJNA1124592. بيانات تسلسل الجينوم الكامل متاحة في مشروع البيولوجيا PRJNA1146711. تم توفير بيانات المصدر مع هذه الورقة.
References
Altpeter, F. et al. Advancing crop transformation in the era of genome editing. Plant Cell 28, 1510-1520 (2016).
Chen, K., Wang, Y., Zhang, R., Zhang, H. & Gao, C. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 70, 667-697 (2019).
Nasti, R. A. & Voytas, D. F. Attaining the promise of plant gene editing at scale. Proc. Natl Acad. Sci. USA 118, e2004846117 (2021).
Honig, A. et al. Transient expression of virally delivered meganuclease in planta generates inherited genomic deletions. Mol. Plant 8, 1292-1294 (2015).
Marton, I. et al. Nontransgenic genome modification in plant cells. Plant Physiol. 154, 1079-1087 (2010).
Ali, Z. et al. Efficient virus-mediated genome editing in plants using the CRISPR/Cas9 system. Mol. Plant 8, 1288-1291 (2015).
Ellison, E. E., Chamness, J. C. & Voytas, D. F. Viruses as Vectors for the Delivery of Gene-editing Reagents (Burleigh Dodds Science Publishing, 2021).
Beernink, B. M., Lappe, R. R., Bredow, M. & Whitham, S. A. Impacts of RNA mobility signals on virus induced somatic and germline gene editing. Front. Genome Ed. 4, 925088 (2022).
Uranga, M. et al. Efficient Cas9 multiplex editing using unspaced sgRNA arrays engineering in a Potato virus X vector. Plant J. 106, 555-565 (2021).
Bradamante, G., Mittelsten Scheid, O. & Incarbone, M. Under siege: virus control in plant meristems and progeny. Plant Cell 33, 2523-2537(2021).
Ma, X., Zhang, X., Liu, H. & Li, Z. Highly efficient DNA-free plant genome editing using virally delivered CRISPR-Cas9. Nat. Plants 6, 773-779 (2020).
Liu, Q., Zhao, C., Sun, K., Deng, Y. & Li, Z. Engineered biocontainable RNA virus vectors for non-transgenic genome editing across crop species and genotypes. Mol. Plant 16, 616-631 (2023).
Lee, S.-Y. et al. Development of virus-induced genome editing methods in solanaceous crops. Hortic. Res. 11, uhad233 (2023).
Ariga, H., Toki, S. & Ishibashi, K. Potato virus X vector-mediated DNA-free genome editing in plants. Plant Cell Physiol. 61, 1946-1953 (2020).
Ghoshal, B. et al. A viral guide RNA delivery system for CRISPR-based transcriptional activation and heritable targeted DNA demethylation in Arabidopsis thaliana. PLoS Genet. 16, e1008983 (2020).
Karvelis, T. et al. Transposon-associated TnpB is a programmable RNA-guided DNA endonuclease. Nature 599, 692-696 (2021).
Altae-Tran, H. et al. The widespread IS200/IS605 transposon family encodes diverse programmable RNA-guided endonucleases. Science 374, 57-65 (2021).
Meers, C. et al. Transposon-encoded nucleases use guide RNAs to promote their selfish spread. Nature 622, 863-871(2023).
Li, Q. et al. Genome editing in plants using the TnpB transposase system. aBIOTECH 5, 225-230 (2024).
Xiang, G. et al. Evolutionary mining and functional characterization of TnpB nucleases identify efficient miniature genome editors. Nat. Biotechnol. 42, 745-757 (2024).
Li, Z. et al. Engineering a transposon-associated TnpB-ωRNA system for efficient gene editing and phenotypic correction of a tyrosinaemia mouse model. Nat. Commun. 15, 831 (2024).
Yoo, S.-D., Cho, Y.-H. & Sheen, J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat. Protoc. 2, 1565-1572 (2007).
Zhang, X., Henriques, R., Lin, S.-S., Niu, Q.-W. & Chua, N.-H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 1, 641-646 (2006).
Wang, M. et al. A gene silencing screen uncovers diverse tools for targeted gene repression in Arabidopsis. Nat. Plants 9, 460-472 (2023).
Ellison, E. E. et al. Multiplexed heritable gene editing using RNA viruses and mobile single guide RNAs. Nat. Plants 6, 620-624 (2020).
Nagalakshmi, U., Meier, N., Liu, J.-Y., Voytas, D. F. & Dinesh-Kumar, S. P. High-efficiency multiplex biallelic heritable editing in Arabidopsis using an RNA virus. Plant Physiol. 189, 1241-1245 (2022).
Qi, Y. et al. Increasing frequencies of site-specific mutagenesis and gene targeting in Arabidopsis by manipulating DNA repair pathways. Genome Res. 23, 547-554 (2013).
Brinkman, E. K. & van Steensel, B. Rapid quantitative evaluation of CRISPR genome editing by TIDE and TIDER. Methods Mol. Biol. 1961, 29-44 (2019).
Huang, Y.-S. & Li, H.-M. Arabidopsis CHLI2 can substitute for CHLI1. Plant Physiol. 150, 636-645 (2009).
Bae, S., Park, J. & Kim, J.-S. Cas-OFFinder: a fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 30, 1473-1475 (2014).
Yoshida, T., Ishikawa, M., Toki, S. & Ishibashi, K. Heritable tissue-culture-free gene editing in Nicotiana benthamiana through viral delivery of SpCas9 and sgRNA. Plant Cell Physiol. 65, 1743-1750 (2024).
Badon, I. W., Oh, Y., Kim, H.-J. & Lee, S. H. Recent application of CRISPR-Cas12 and OMEGA system for genome editing. Mol. Ther. 32, 32-43 (2024).
Macfarlane, S. A. Tobraviruses-plant pathogens and tools for biotechnology. Mol. Plant Pathol. 11, 577-583 (2010).
Zhang, C., Liu, S., Li, X., Zhang, R. & Li, J. Virus-induced gene editing and its applications in plants. Int. J. Mol. Sci. 23, 10202 (2022).
Li, Z. et al. Genome editing in plants using the compact editor Cas Ф. Proc. Natl Acad. Sci. USA 120, e2216822120 (2023).
Čermák, T. et al. A multipurpose toolkit to enable advanced genome engineering in plants. Plant Cell 29, 1196-1217 (2017).
LeBlanc, C. et al. Increased efficiency of targeted mutagenesis by CRISPR/Cas9 in plants using heat stress. Plant J. 93, 377-386 (2018).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754-1760 (2009).
McKenna, A. et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297-1303 (2010).
Kim, S. et al. Strelka2: fast and accurate calling of germline and somatic variants. Nat. Methods 15, 591-594 (2018).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841-842 (2010).
شكر وتقدير
تم دعم هذا العمل من قبل منحة برنامج أبحاث جينوم النباتات من NSF (2334027) إلى S.E.J. وJ.A.D. وJ.F.B. S.E.J. وJ.A.D. هما باحثان في معهد هوارد هيوز الطبي. H.S. هو زميل في HHMI من صندوق جين كوفين تشايلدز للبحوث الطبية. تم دعم B.A.A. من قبل m-CAFEs لتحليل المجتمع الميكروبي والتقييم الوظيفي في التربة، وهي منطقة تركيز علمية يقودها مختبر لورانس بيركلي الوطني بناءً على عمل مدعوم من قبل وزارة الطاقة الأمريكية، مكتب العلوم، مكتب الأبحاث البيولوجية والبيئية (DE-ACO2-O5CH11231). نشكر S. Feng وM. Akhavan ومركز أبحاث الخلايا الجذعية Broad على دعم التسلسل؛ C. Picard على المساعدة في تحليل بيانات تسلسل الأمبليكون؛ S. D. Kumar على توفير بلازميد TRV2 pDK3888؛ F. Zhang على توفير (SALK123114) الطفرة؛ وجميع أعضاء مختبر جاكوبسن على الملاحظات المفيدة والاقتراحات.
مساهمات المؤلفين
قام كل من T.W. و M.K. و H.S. و Z.L. و B.A.A. و J.F.B. و J.A.D. و S.E.J. بتصميم البحث. قام T.W. و H.S. و Z.Z. و J.A.D. و S.E.J. بتفسير البيانات. كتب T.W. و S.E.J. الورقة. قام T.W. و M.K. و H.S. و Z.L. و J.A. و M.M.S. و K.V. و G.W. و S.C. و C.A. و N.S. و A.S. و D.S. بإجراء التجارب.
المصالح المتنافسة
قدم T.W. وM.K وJ.A. وZ.L. وH.S. وB.A.A. وJ.A.D. وS.E.J. براءة اختراع (رقم WO/2025/007023، 2025) تغطي جوانب من هذا العمل. S.E.J. هي مؤسسة مشاركة ومستشارة لشركة Inari Agriculture ومستشارة لشركات Terrana Biosciences وInvaio Sciences وSail Biomedicines وSix Street وZymo Research. تمتلك هيئة Regents لجامعة كاليفورنيا براءات اختراع صادرة ومعلقة لتقنيات CRISPR التي يعتبر J.A.D. مخترعًا لها. J.A.D. هي مؤسسة مشاركة لشركات Azalea Therapeutics وCaribou Biosciences وEditas Medicine وEvercrisp وScribe Therapeutics وMammoth Biosciences. J.A.D. هي عضو في مجلس الاستشارات العلمية في Evercrisp وCaribou Biosciences وScribe Therapeutics وMammoth Biosciences وThe Column Group وInari. وهي أيضًا مستشارة لشركة Aditum Bio. J.A.D. هي المستشارة العلمية الرئيسية لشركة Sixth Street، ومديرة في Johnson & Johnson وAltos وTempus، ولديها مشروع بحثي برعاية Apple Tree Partners. يعلن المؤلفون الآخرون عدم وجود مصالح متنافسة.
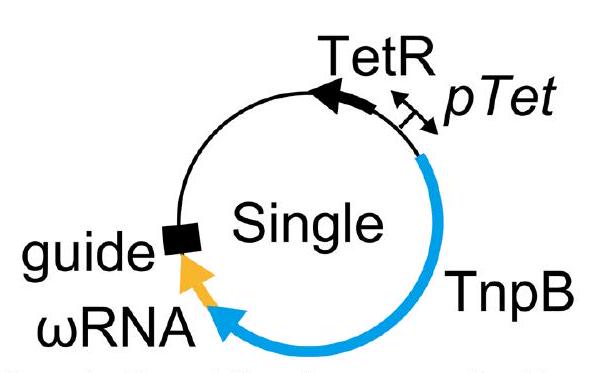
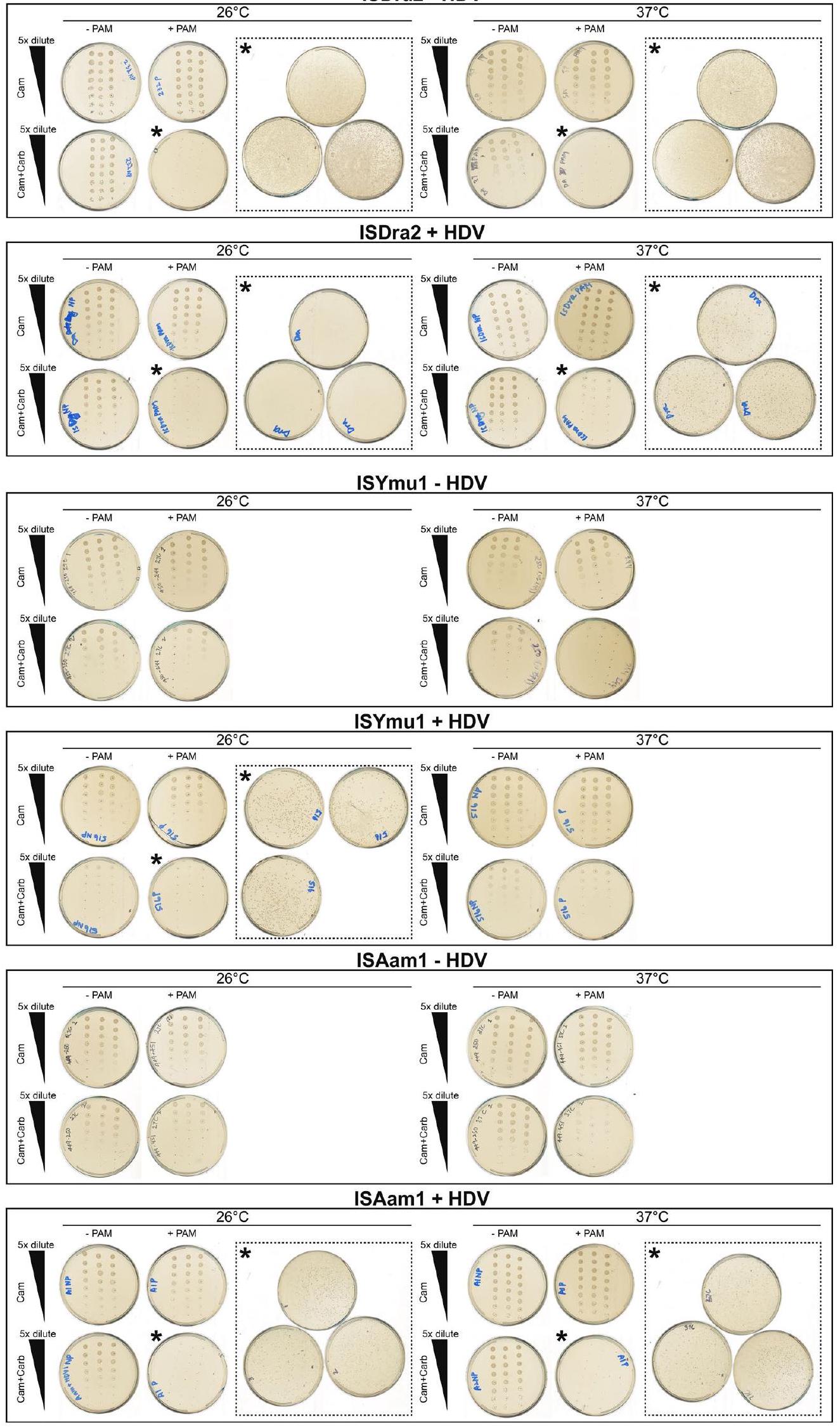
الشكل 1 من البيانات الموسعة | مخطط لمتجهات اختبار التداخل البكتيري التي تحتوي على تصميم TnpB-ωRNA مفرد أو TnpB-ωRNA مفرد-HDV. السهم الأزرق يشير إلى تسلسل TnpB؛ السهم الأصفر يشير إلىتسلسل RNA؛ المستطيل الأسود يشير إلى تسلسل الدليل؛ السهم الأخضر يشير إلى تسلسل ريبوزيم HDV. تحتوي البلازميدات على جين مقاومة التتراسيكلين (TetR). تم استخدام محفز تتراسيكلين (pTet) لتحفيز تعبير TnpB-تسلسلات RNA المفردة أو Single-HDV، وجين مقاومة التتراسيكلين.
ISDra2 – HDV
الشكل 2 من البيانات الموسعة | صور الأطباق لاختبار تداخل البلازميد البكتيري. تخفيفات متسلسلة بنسبة خمسة أضعافتم زراعة 1 مل من ثقافة الاسترداد بعد التحول على كل من أطباق LB-Agar بمضاد حيوي واحد (كام، الصف العلوي) وأطباق LB-Agar بمضادين حيويين (كام + كارب، الصف السفلي).
الأطباق التي لا تحتوي على مستعمرات مرئية (المشار إليها بنجمة) كانتمن الثقافة الأصلية بتركيز 1 مل التي تم زراعتها على أطباق المضادات الحيوية المزدوجة (الإدخالات السهمية). تم إجراء التجارب في ثلاث نسخ.
الشكل 3 من البيانات الموسعة | كفاءة التحرير وملفات إصلاح الحمض النووي لـ ISDra2
ISYmu1 و ISAam1 في تجربة البروتوبلاست. (أ) اسم كل TnpB تم اختباره موجود في الزاوية العليا اليسرى من كل رسم بياني. يتم رسم gRNAs على المحور السيني وكفاءة التحرير (نسبة قراءات indel (%)) على المحور الصادي. كل نقطة تشير إلى عملية نقل واحدة. تم حساب الخطأ المعياري للمتوسط (SEM) لكل موقع مستهدف. تم استخدام نسختين أو ثلاث نسخ لكل موقع مستهدف تم اختباره. (ب) ملفات تعريف إصلاح الحمض النووي indel لعينات النقل الفردية.
تي سي سي إيه آي تي سي تي آي إيه سي آي تي جي تي تي تي سي آي جي تي آي إيه آي جي سي إيه آي
-4 -TTTTAATTTGTGTGGTATTTA
—-ATCTACATGTTTCGTAAGCAA
٥١٦
-14 -TTTTAATTTGTGTGGTATTTAA
————–TCGTAAGCAA
234
-13 -TTTTAATTTGTGTGGTATTTAA
————-TTCGTAAGCAA
228
-8 -TTTTAATTTGTGTGGTATTTA
220
-9 -ITTTTAAT
تي تي جي تي جي جي تي آي تي-
196
أكثر خمسة أنواع شائعة من الإضافات والحذف مدرجة على اليسار. عدد القراءات لكل إضافة أو حذف مدرج على اليمين. يتم تحديد PAM بواسطة المربع الأحمر، والموقع المستهدف محدد بالمربع الأسود، في تسلسل المرجع. يتم إدراج إجمالي عدد القراءات وكفاءة التحرير أسفل كل ملف تعريف للإضافة أو الحذف. يتم عرض اسم كل من TnpB و gRNA فوق تسلسل المرجع لكل ملف تعريف لإصلاح الإضافة أو الحذف.
ISYmu1 gRNA2
المرجع = ITTGAI A AGGGA A TITCIGCCIGC AGA A A AITCCCA A ACACA ACCATI
الشكل 4 من البيانات الموسعة | ملفات إصلاح الحمض النووي التمثيلية للنباتات المتحولة الفردية التي تعبر عن إما ISYmu1 AtPDS3 gRNA2 أو gRNA12. يتم سرد أكثر عشرة أنواع شائعة من الإضافات والحذف على اليسار. يتم سرد عدد القراءات لكل إضافة وحذف على اليمين. يتم تحديد PAM بواسطة المربع الأحمر، وموقع الهدف محدد بالمربع الأسود، في تسلسل المرجع. يتم سرد إجمالي عدد القراءات وكفاءة التحرير أسفل كل ملف إصلاح إضافة وحذف. يتم عرض اسم كل TnpB و gRNA فوق تسلسل المرجع لكل ملف إصلاح إضافة وحذف.
الشكل البياني الممتد 5 | لقطات تسلسل سانجر لأنماط الجينات للنباتات الناتجة. (أ، ب) اللوحات أ وب تتعلق بالنباتات الناتجة المستهدفة بواسطة AtPDS3gRNA12 من النباتات 54 و69، على التوالي. التسلسل في الأعلى هو تسلسل الجينوم البري؛ أسفل ذلك هو هدف ISYmu1gRNA12 وPAM (صندوق رمادي)؛ يعرض ملف تتبع ab1 الطفرة. (ج) كل صندوق يعرض التركيب الجيني لنبات النسل من تجربة تحرير AtCHLl1gRNA4. يتم الإشارة إلى معرف النبات فوق كل مربع. التسلسل في الأعلى هو التسلسل الجيني للنمط البري؛ أسفل ذلك هو هدف ISYmu1 AtCHLl1 gRNA4 وPAM (المربع الأصفر)؛ تعرض ملف ab1 أثر الطفرة.
النبات 43 حذف ثلاثي القواعد غير المتجانس
النباتات 2 و 32 و 43 حاملة حذف 7bp متماثل الزيجوت
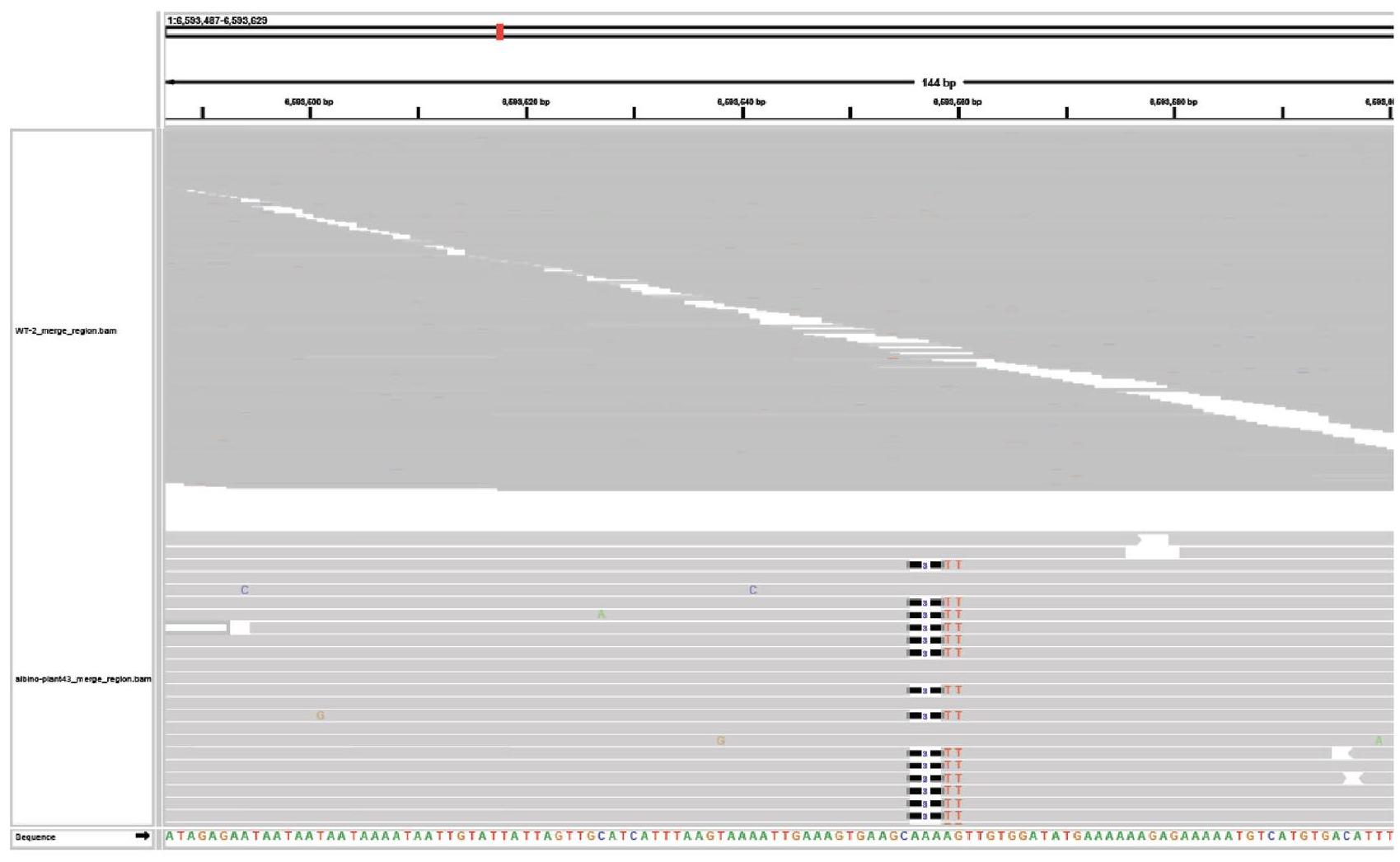
الشكل البياني الموسع 7| الطفرات المحتملة خارج الهدف التي تم تحديدها بواسطة تسلسل الجينوم الكامل. لقطة شاشة لمتصفح الجينوم للتعديلات المحتملة التي تم تحديدها بواسطة تسلسل الجينوم الكامل للنبات 43 (حذف 3 قواعد مزدوجة غير متجانسة) والنباتات 2 و32 و43 (حذف 7 قواعد مزدوجة متجانسة). يتم عرض الطفرة. في المسار السفلي، يتم عرض تسلسل النوع البري في المسار العلوي لكل لقطة شاشة. يتم عرض معرف النبات، ونوع الزيجوت، ونوع الطفرة فوق كل لقطة شاشة. يتم إدراج الموقع الجينومي في الزاوية العلوية اليسرى من كل لقطة شاشة.
النباتات 2 و32 و43 متجانسة الزيجوت SNP (A)
ازرع 32 SNP متغاير الزيجوت (T)
الشكل 8 من البيانات الموسعة | الطفرات المحتملة خارج الهدف التي تم تحديدها بواسطة تسلسل الجينوم الكامل. لقطة شاشة لمتصفح الجينوم للطفرات المحتملة التي تم تحديدها بواسطة تسلسل الجينوم الكامل للنباتات 2 و32 و43 (SNP متماثل الزيجوت (A)) والنبات 32 (SNP غير متماثل الزيجوت (T)). يتم عرض الطفرة على المسار السفلي، وتسلسل النوع البري معروض في المسار العلوي لكل لقطة شاشة. يتم عرض معرف النبات، والزيغوسية، ونوع الطفرة فوق كل لقطة شاشة. يتم إدراج الموقع الجينومي في الزاوية العلوية اليسرى من كل لقطة شاشة.
نبات 2 SNP متغاير الزيجوت (T)
زرع 32 SNP متغاير الزيجوت (T)
الشكل البياني الموسع 9 | الطفرات المحتملة خارج الهدف التي تم تحديدها بواسطة تسلسل الجينوم الكامل. لقطة شاشة لمتصفح الجينوم للتعديلات المحتملة التي تم تحديدها بواسطة تسلسل الجينوم الكامل للنبات 2 (SNP(T) غير المتجانس) والنبات 32 (SNP(T) غير المتجانس). الطفرة معروضة في الشريط السفلي. وتظهر تسلسل النوع البري في المسار العلوي لكل لقطة شاشة. يتم عرض معرف النبات، والزيغوسية، ونوع الطفرة فوق كل لقطة شاشة. يتم إدراج الموقع الجينومي في الزاوية العلوية اليسرى من كل لقطة شاشة.
زرع 2 SNP متماثل الزيجوت (G)
النباتات 2 و 32 و 43 متجانسة الزيجوت SNP (A)
الشكل البياني الممتد 10|الطفرات المحتملة خارج الهدف التي تم تحديدها بواسطة تسلسل الجينوم الكامل. لقطة شاشة لمتصفح الجينوم للطفرات المحتملة التي تم تحديدها بواسطة تسلسل الجينوم الكامل للنبات 2 (SNP متماثل الزيجوت (G)) والنباتات 2 و32 و43 (SNP متماثل الزيجوت (A)). يتم عرض الطفرة على المسار السفلي، وتسلسل النوع البري معروض في المسار العلوي لكل لقطة شاشة. يتم عرض معرف النبات، والزيغوسية، ونوع الطفرة فوق كل لقطة شاشة. الموقع الجينومي مدرج في الزاوية العلوية اليسرى من كل لقطة شاشة.
البحث الذي يتضمن مشاركين بشريين، بياناتهم، أو مواد بيولوجية
معلومات السياسة حول الدراسات التي تشمل مشاركين بشريين أو بيانات بشرية. انظر أيضًا معلومات السياسة حول الجنس، الهوية/التقديم الجنسي، والتوجه الجنسي والعرق، والعرقية والعنصرية.
التقارير عن الجنس والنوع الاجتماعي
غير متوفر
التقارير عن العرق أو الإثنية أو غيرها من التجمعات الاجتماعية ذات الصلة
غير متوفر
خصائص السكان
غير متوفر
التوظيف
غير متوفر
رقابة الأخلاقيات
غير متوفر
يرجى ملاحظة أنه يجب أيضًا تقديم معلومات كاملة حول الموافقة على بروتوكول الدراسة في المخطوطة.
التقارير المتخصصة في المجال
يرجى اختيار الخيار أدناه الذي يناسب بحثك بشكل أفضل. إذا لم تكن متأكدًا، اقرأ الأقسام المناسبة قبل اتخاذ قرارك. علوم الحياة □ العلوم السلوكية والاجتماعية □ العلوم البيئية والتطورية والبيئية لنسخة مرجعية من الوثيقة مع جميع الأقسام، انظرnature.com/documents/nr-reporting-summary-flat.pdf
تصميم دراسة العلوم الحياتية
يجب على جميع الدراسات الإفصاح عن هذه النقاط حتى عندما يكون الإفصاح سلبياً.
حجم العينة
لم يتم إجراء حساب لحجم العينة. بالنسبة لاختبار التداخل البكتيري وتجارب البروتوبلاست، قمنا باختبار ما لا يقل عن نسختين (عادة ثلاث) لكل معالجة للسماح بالمقارنة بين النسخ. بالنسبة لتجارب البروتوبلاست التي تحتوي على نسختين، فشلت إحدى النسخ في التضخيم لتحليل تسلسل الجيل التالي. بالنسبة لتجارب تحرير الجينات المتحولة، قمنا بتحليل البيانات من جميع النباتات المتحولة التي تمكنا من إنشائها باستخدام طريقة غمر الأزهار. بالنسبة لتجارب توصيل TRV، لم نحدد أي أحجام عينة مسبقًا. تم تحديد أحجام عينات النباتات المتحولة وTRV بناءً على كمية المساحة المتاحة في الدفيئة.
استبعاد البيانات
لم يتم استبعاد أي بيانات من هذه الدراسة.
التكرار
للتحقق من قابلية تكرار التجارب، تم إجراء نسخ بيولوجية لكل تجربة.
التوزيع العشوائي
تم جمع جميع العينات بشكل عشوائي من المواد الموضوعية، باستثناء ما تم الإشارة إليه في قسم الطرق. على سبيل المثال، عند جمع الأنسجة لقياس التحرير الجسدي في النباتات التي خضعت لتوصيل TRV، تم جمع ثلاث أوراق عشوائية بعيدة عن موقع التوصيل. ومع ذلك، إذا كانت هناك قطاعات بيضاء أو صفراء من الخلايا موجودة على الأوراق البعيدة عن موقع توصيل TRV، تم جمع الأنسجة من تلك الأوراق.
عمى
كان المحققون غير مدركين لتوزيع المجموعات أثناء جمع البيانات و/أو التحليل لجميع التجارب.
التقارير عن مواد وأنظمة وطرق محددة
نحتاج إلى معلومات من المؤلفين حول بعض أنواع المواد والأنظمة التجريبية والأساليب المستخدمة في العديد من الدراسات. هنا، يرجى الإشارة إلى ما إذا كانت كل مادة أو نظام أو طريقة مدرجة ذات صلة بدراستك. إذا لم تكن متأكدًا مما إذا كان عنصر القائمة ينطبق على بحثك، يرجى قراءة القسم المناسب قبل اختيار رد.
المواد والأنظمة التجريبية
طرق
غير متوفر
مشارك في الدراسة
غير متوفر
–
□
□
〃
ણ
□
□
ણ
نباتات
¹قسم البيولوجيا الجزيئية والخلوية وتطور الحياة، جامعة كاليفورنيا في لوس أنجلوس، لوس أنجلوس، كاليفورنيا، الولايات المتحدة الأمريكية. ²معهد الجينوميات المبتكرة، جامعة كاليفورنيا، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.معهد هوارد هيوز الطبي، جامعة كاليفورنيا، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.معهد كاليفورنيا للعلوم الحيوية الكمية (QB3)، جامعة كاليفورنيا، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.قسم علوم الأرض والكواكب، جامعة كاليفورنيا، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.قسم علوم البيئة والسياسة والإدارة، جامعة كاليفورنيا، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.جامعة ملبورن، ملبورن، أستراليا.قسم البيولوجيا الجزيئية والخلوية، جامعة كاليفورنيا، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.قسم الكيمياء، جامعة كاليفورنيا، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.قسم البيوفيزياء الجزيئية والتصوير الحيوي المتكامل، مختبر لورانس بيركلي الوطني، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.مركز لي كا شينغ لعلم الجينوم الانتقالي، جامعة كاليفورنيا، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.معهد غلادستون لعلوم البيانات والتكنولوجيا الحيوية، سان فرانسيسكو، كاليفورنيا، الولايات المتحدة الأمريكية.معهد غلادستون-يو سي إس إف لعلم المناعة الجينومية، سان فرانسيسكو، كاليفورنيا، الولايات المتحدة الأمريكية.معهد هوارد هيوز الطبي (HHMI)، جامعة كاليفورنيا في لوس أنجلوس، لوس أنجلوس، كاليفورنيا، الولايات المتحدة الأمريكية.العنوان الحالي: وزارة التعليم، المختبر الرئيسي للموارد البيولوجية والبيئة الإيكولوجية، كلية علوم الحياة، المختبر الرئيسي للديناميكا الهيدروليكية وهندسة الأنهار الجبلية، جامعة سيتشوان، تشنغدو، الصين. □ البريد الإلكتروني:jacobsen@ucla.edu
Genome editing is transforming plant biology by enabling precise DNA modifications. However, delivery of editing systems into plants remains challenging, often requiring slow, genotype-specific methods such as tissue culture or transformation . Plant viruses, which naturally infect and spread to most tissues, present a promising delivery system for editing reagents. However, many viruses have limited cargo capacities, restricting their ability to carry large CRISPR-Cas systems. Here we engineered tobacco rattle virus (TRV) to carry the compact RNA-guided TnpB enzyme ISYmu1 and its guide RNA. This innovation allowed transgene-free editing of Arabidopsis thaliana in a single step, with edits inherited in the subsequent generation. By overcoming traditional reagent delivery barriers, this approach offers a novel platform for genome editing, which can greatly accelerate plant biotechnology and basic research.
Programmable RNA-guided endonucleases, including CRISPR-Cas9, are driving advances in genome editing for both fundamental research and biotechnology. The ability to genetically modify plant genomes has allowed for the creation of rationally designed phenotypes. However, efficient delivery of genome editing reagents to plants remains a major challenge. The most common strategy is to encode RNA-guided genome editors (for example, CRISPR-Cas enzymes) within transgenes and use tissue culture and plant transformation approaches to make
transgenic plants, after which genetic crosses are required to remove the transgenic material but retain the edits . However, current plant transformation methods are limited to specific plant species and genotypes, often require considerable time, resources and technical expertise, and can cause unintended changes to the genome and epigenome .
An approach to circumvent these limitations is to use plant viral vectors to deliver genome editing reagents such as meganucleases or zinc finger nucleases (ZFNs) for targeted mutagenesis . While the use
of meganucleases and ZFNs for viral-mediated plant genome editing was a notable advance, the ability to encode an easily programmable RNA-guided CRISPR system would be highly advantageous. As such, several viral vectors have been engineered to encode guide RNAs (gRNAs) for delivery to transgenic plants already expressing Cas9, resulting in somatic and germline editing and transmission of edits to the next generation . Because plants have evolved mechanisms to restrict viral infection of meristem and germ cells, most viruses are rarely sexually transmitted . However, transient invasion of meristem cells by viral RNAs encoding gRNAs can allow these cells to be edited and for these edits to be seed transmissible . While these approaches represent important advances, they still require the use of nucleases that can be challenging to engineer (such as meganucleases and ZFNs), or transgenic plants expressing the CRISPR-Cas endonuclease protein.
A strategy to avoid the need for transgenic plant materials has been the use of viral vectors with large cargo capacities, capable of expressing entire RNA-guided editing systems (for example, Cas9 and the gRNA). This approach has been met with some success; however, it still requires plant regeneration steps because these viruses do not cause germline editing and heritability of the edits . On the other hand, encoding entire CRISPR systems in viruses that are capable of germline transmission has been challenging because of their limited cargo capacity .
To overcome this cargo size limit, we explored the potential of TnpB, a class of ultracompact RNA-guided endonucleases ( amino acids) , to be encoded in a plant RNA viral vector. As ancestors of Cas enzymes, TnpBs similarly utilize a programmable RNA guide, called an omega RNA ( RNA), to be directed to any target site and induce genome edits. Previously, TnpBs ISDra2, ISYmu1 and ISAam1 were shown to be capable of targeted genome editing in mammalian cells, and ISDra2 and ISYmu1 in monocot rice plant cells . Here we tested the ISDra2, ISYmu1 and ISAam1 TnpBs for genome editing in the dicot plant, Arabidopsis. Given the single cargo site in the TRV vector that is typically used, we sought to express both the TnpB protein and its guide RNA within the same mRNA transcript under a single promoter, similar to their natural expression arrangement .
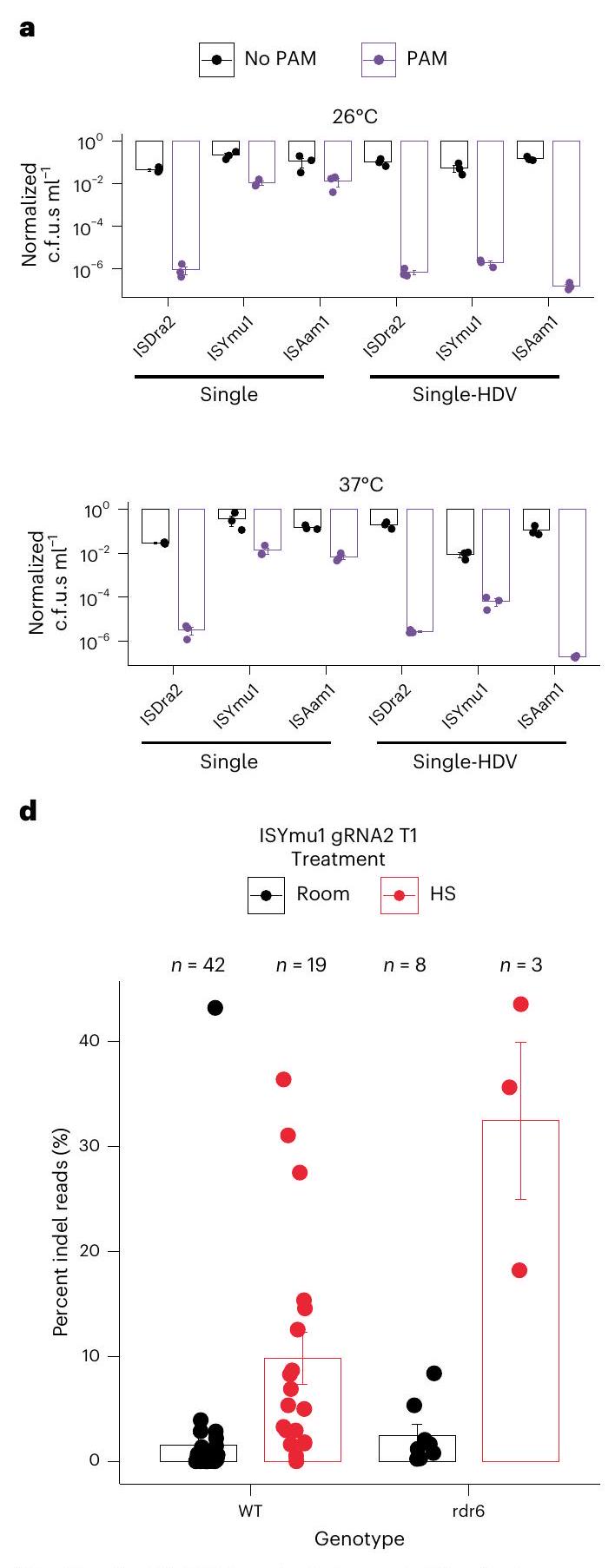
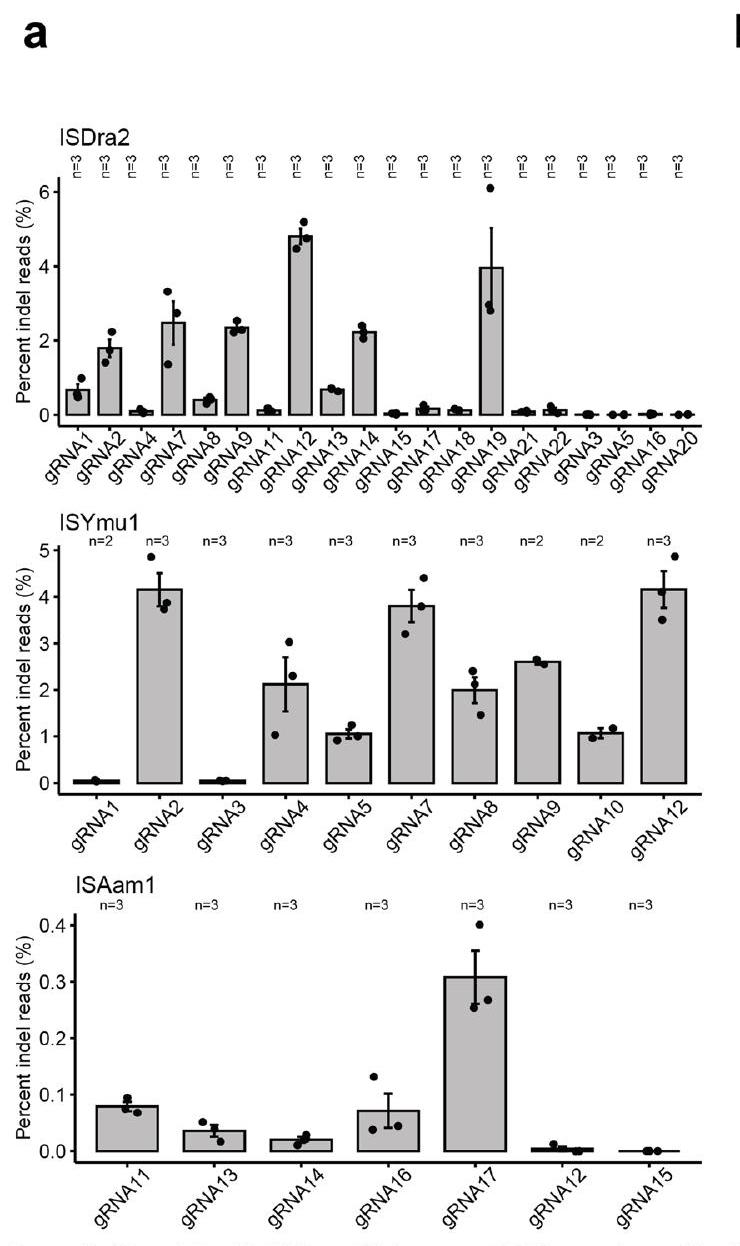
To test the activities of TnpB and its gRNA encoded in a single transcript, we first expressed these three TnpBs and assessed their RNA-guided plasmid interference activities in bacteria. We co-expressed the TnpB and gRNA from the same promoter as a single transcript, maintaining their natural sequences without codon optimization. We compared two configurations of the 3′-guide region: one extended continuously without a terminator to mimic the natural TnpB condition, and another capped by the hepatitis delta virus (HDV) ribozyme, as previously used in bacteria (Extended Data Fig.1). Our results showed that without the HDV ribozyme, only ISDra2 demonstrated plasmid interference activity whereas with the HDV ribozyme, all three TnpBs exhibited robust activity at both and (Fig. 1a and Extended Data Fig. 2). These findings revealed that single transcript expression cassettes with an HDV ribozyme sequence at the end are capable of cleaving plasmid DNA in bacteria.
To test the single expression cassette for targeted genome editing in Arabidopsis, we used the AtUBQ10 promoter to drive expression of the TnpB- RNA and a gRNA targeting the PHYTOENE DESATURASE3 (AtPDS3) gene region, followed by the HDV ribozyme and rbcS-E9 terminator (Fig. 1b). We tested 20 ISDra2 sites, 10 ISYmu1 sites and 7 ISAam1 sites for editing capabilities in Arabidopsis protoplast cells (Supplementary Table 1) . ISDra2 and ISYmu1 demonstrated active editing ranging from and , respectively, as measured by next-generation amplicon sequencing (amp-seq) (Extended Data Fig. 3a). ISAam1 was much less active, with editing efficiency ranging 0-0.3% (Extended Data Fig. 3a). On average, we observed editing efficiencies of 1% for ISDra2, 2.1% for ISYmu1 and 0.1% for ISAam1 (Fig.1c). In line with previous reports, the DNA repair profiles consisted of
deletion-dominant repair outcomes for all three TnpBs (Extended Data Fig. 3b) . These data demonstrate that ISDra2, ISYmu1 and ISAam1 are all capable of targeted genome editing in Arabidopsis plant cells using the single transcript expression design.
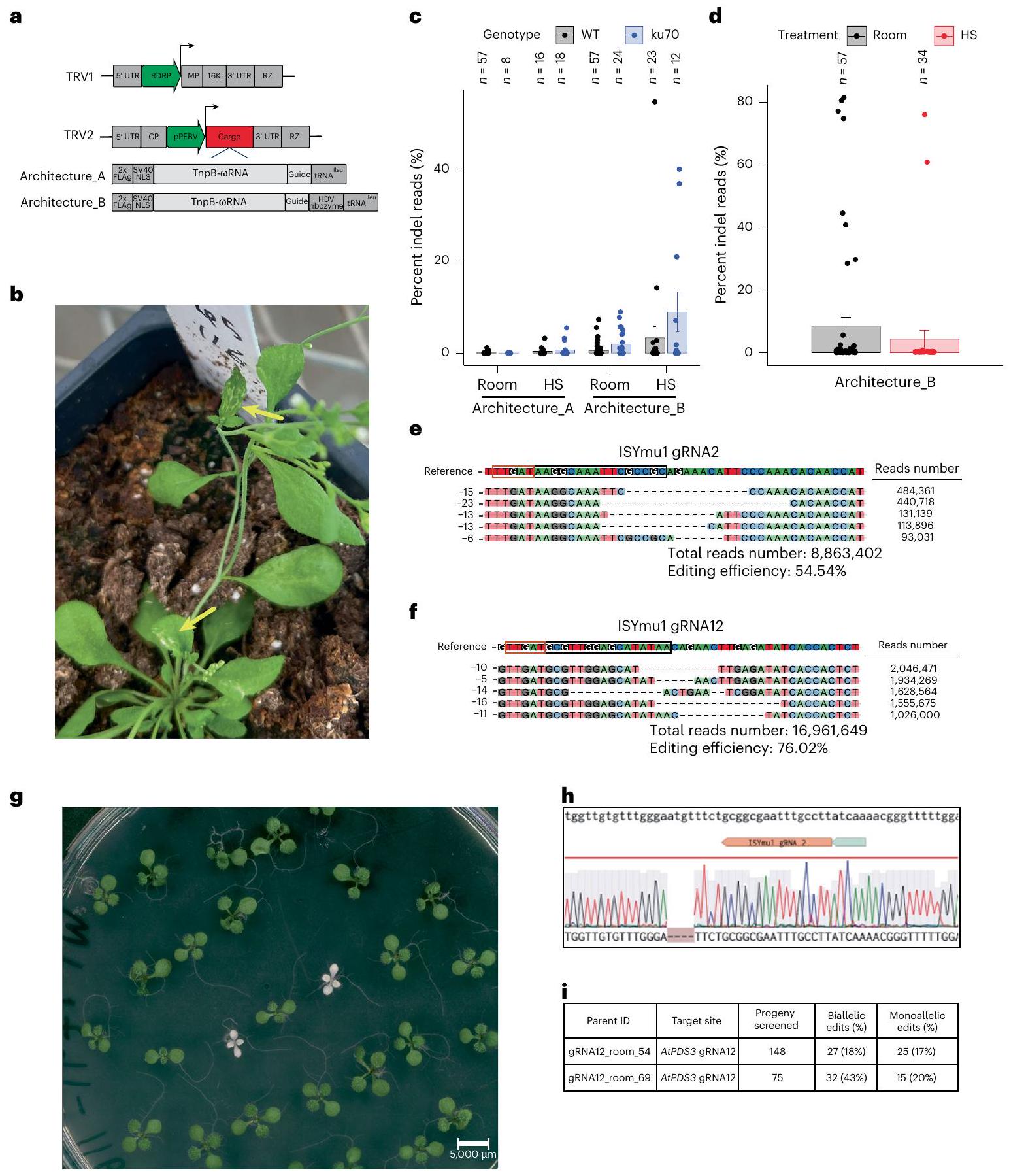
To evaluate TnpB-mediated editing in transgenic plants we selected ISYmu1, as it demonstrated the highest average editing efficiency in Arabidopsis protoplast cells and was shown to exhibit no off-target editing in rice . Two gRNAs with the most active editing were selected, each targeting a unique genomic context. gRNA2 targeted the coding region of AtPDS3, whereas gRNA12 targeted the promoter region directly upstream of the AtPDS3 gene. Transgenic plants were created via standard floral dip transformation utilizing the same plasmids as for the protoplast experiments . To test for sensitivity to temperature, transgenic plants expressing ISYmu1 were either grown at room temperature or subjected to a heat-shock treatment. We tested editing in wild-type (WT) plants, as well as in the rna dependent rna polymerase 6(rdr6) mutant which is known to have reduced transgene silencing . Analysis using amp-seq revealed an average editing efficiency of and for gRNA2 in WT and rdr6, respectively (Fig.1d). Analysis of gRNA12 revealed greater editing than gRNA2, averaging editing in WT and in rdr6 (Fig. 1e). Comparison of editing efficiency in the plants grown at room temperature with those that received the heat-shock treatment revealed a preference for increased temperature for both target sites in the WT background, demonstrating 6.3 -fold and 1.4 -fold increases in editing for gRNA2 and gRNA12, respectively (Fig. 1d,e). In rdr6, we observed a 13-fold increase in editing for gRNA2, but little change in editing for gRNA12 (Fig.1d,e). The editing outcomes from transgenic T1 plants expressing ISYmu1 consisted of chimaeric, deletion-dominant, DNA repair profiles (Extended Data Fig. 4). These data demonstrate that ISYmu1, encoded as a transgene, is capable of performing efficient genome editing in Arabidopsis plants, and that heat treatment and the rdr6 silencing mutant can be used to increase editing efficiency.
Encouraged by the ISYmu1 activity in transgenic Arabidopsis plants, we next tested ISYmu1 for TRV-mediated genome editing. TRV is a bipartite RNA virus composed of TRV1 and TRV2 (Fig. 2a). Previous work has shown that the TRV2 RNA can be engineered by inserting a cargo expression cassette downstream of the pea early browning virus promoter (pPEBV) (Fig. 2a) . To test ISYmu1 for genome editing capabilities via TRV delivery to Arabidopsis, we engineered two TRV2 cargo architectures. In TRV2 Architecture_A, the tRNA was directly downstream of the TnpB and gRNA sequences (Fig. 2a). In TRV Architecture_B, we included an HDV ribozyme sequence between the guide and tRNA sequence (Fig. 2a). We included tRNA in both designs as it was previously shown to promote systemic TRV movement and transmission of edited alleles to the next generation .
First, we evaluated TRV-mediated editing potential with gRNA2 using both TRV2 Architecture_A and Architecture_B. gRNA2 was selected because it targets the AtPDS3 coding sequence, enabling easy phenotypic screening for editing due to white photobleaching of cells containing biallelic mutations . We delivered TRV vectors to both WT and the genetic mutant. plays a role in the non-homologous end joining (NHEJ) double strand break repair pathway . ISYmu1-mediated editing efficiency should be greater in the genotype if double-stranded breaks generated by ISYmu1 are repaired through NHEJ. Each TRV2 plasmid was co-delivered with the TRV1 plasmid to Arabidopsis plants using the agroflood method . White speckles were observed on some of the leaves at weeks post agroflooding, suggesting that sectors of cells contained biallelic mutations in the target AtPDS3 gene (Fig. 2b). Amp-seq analysis revealed an average of 0.1% and 0% editing efficiency in leaf tissue of WT and plants agroflooded with TRV2 Architecture_A and grown under room temperature, respectively (Fig. 2c). For the heat-shock-treated plants, we observed an average editing efficiency of in WT and in plants agroflooded with TRV2
Fig. 1| Expression of TnpB and guide RNA in a single transcript for plant genome editing. a, Barplots of interference assay testing the single transcript expression TnpB vectors for cleavage in E. coli. Data are from experiments performed at (top) and at (bottom). Bars indicate absence (black) or presence (purple) of a PAM on the target plasmid. The axis is a scale of the normalized c.f.u.s . The axis displays the three TnpBs tested using the single expression transcript design without or with an HDV ribozyme. The s.e.m. was calculated for each experiment, with 3 replicates per experiment. b, Schematic of the single expression transcript TnpB- RNA plasmid design used for plant genome editing. The green arrow symbolizes the AtUBQ10 promoter; the dark grey boxes indicate the -FLAG, SV40 NLS and HDV ribozyme sequences; the light grey boxes indicate the TnpB- RNA and guide sequences; the red box
symbolizes the rbcS-E9 terminator; the black arrow indicates the orientation of the TnpB- RNA expression cassette. c, Barplot displaying the average editing efficiencies ( ± s.e.m.) for protoplast experiments using ISDra2, ISYmu1 and ISAam1 TnpBs. Each dot represents the average editing efficiency (percent indel reads) of a gRNA from Extended Data Fig. 3a, with number of samples indicated at the top of the plot. d,e, ISYmu1 somatic editing in T1 transgenic plants for ISYmu1 gRNA2 (d) and ISYmu1 gRNA12 (e). The genotypes are plotted along the axis and the editing efficiencies (percent indel reads) ( ± s.e.m.) are plotted on the axis. Each dot indicates a single T1 transgenic plant. The room and HS treatments stand for room temperature and heat-shock plant growth conditions, respectively.
Architecture_A (Fig. 2c). Using TRV2 Architecture_B, we observed an average of and editing in WT and , respectively, for the room-temperature-grown plants. For the plants that received TRV2 Architecture_B and a heat shock, we observed an average editing
efficiency of in WT and in (Fig. 2c). These results show that Architecture_B, containing the HDV ribozyme, generated higher editing than Architecture_A, and that the mutant can enhance editing efficiency.
Fig. 2 | Somatic and heritable editing in Arabidopsis using TRV to deliver ISYmu1 TnpB and guide RNA targeting AtPDS3. a, Schematic of the TRV1 and TRV2 plasmids. Green arrows indicate the RNA-dependent RNA polymerase (RDRP) and pPEBV promoters for TRV1 and TRV2, respectively; the grey boxes in TRV1 and TRV2 indicate the native TRV components; the red Cargo box in TRV2 indicates the location of either Architecture_A or Architecture_B;below TRV2 are schematics of the components, Architecture_A or Architecture_B, cloned into the TRV2 Cargo slot. b, Representative picture of a plant displaying white sectors in leaves (yellow arrows) weeks after TRV delivery. c,d, Barplot displaying the somatic editing efficiencies (percent indel reads) ( axis) for ISYmu1 gRNA2 in WT and genetic backgrounds (c) and for ISYmu1 gRNA12 in WT (d). The TRV2 cargo architectures are plotted along the axis with either room or HS treatment. Each dot represents an individual plant that underwent agroflood TRV delivery. The s.e.m. was calculated for each experiment. e,f, DNA indel repair profile for an individual WT plant that underwent delivery of TRV Cargo Architecture_B with ISYmu1 gRNA2 (e) or with ISYmu1 gRNA12 (f) under
the heat-shock treatment. The top five most common indel types are listed on the left. The read counts for each indel are listed on the right. The PAM is identified by the red box, and the target site is outlined by the black box, in the Reference sequence. The total read number and editing efficiency are listed below each indel profile.g, Representative image of albino and green progeny seedlings from a WT plant showing somatic editing using the TRV2 Architecture_B design with gRNA2 that underwent heat-shock treatment. h, Sanger sequencing trace file screenshot from one of the albino plants in Fig. 3a. Top: sequence of the wild-type reverse complement. Middle: the ISYmu1 gRNA2 target and PAM (grey box). Bottom: the ab1 trace file displaying a homozygous 4 bp deletion. , Table summarizing the transmission of edited alleles from two individual plants that underwent agroflood delivery using ISYmu1 gRNA12. The ‘Progeny screened’ column indicates the number of seedlings genotyped; the ‘Biallelic edits (%)’ column indicates the number of seedlings containing biallelic edits; and the ‘Monoallelic edits (%)’ column indicates the number of plants harbouring monoallelic edits.
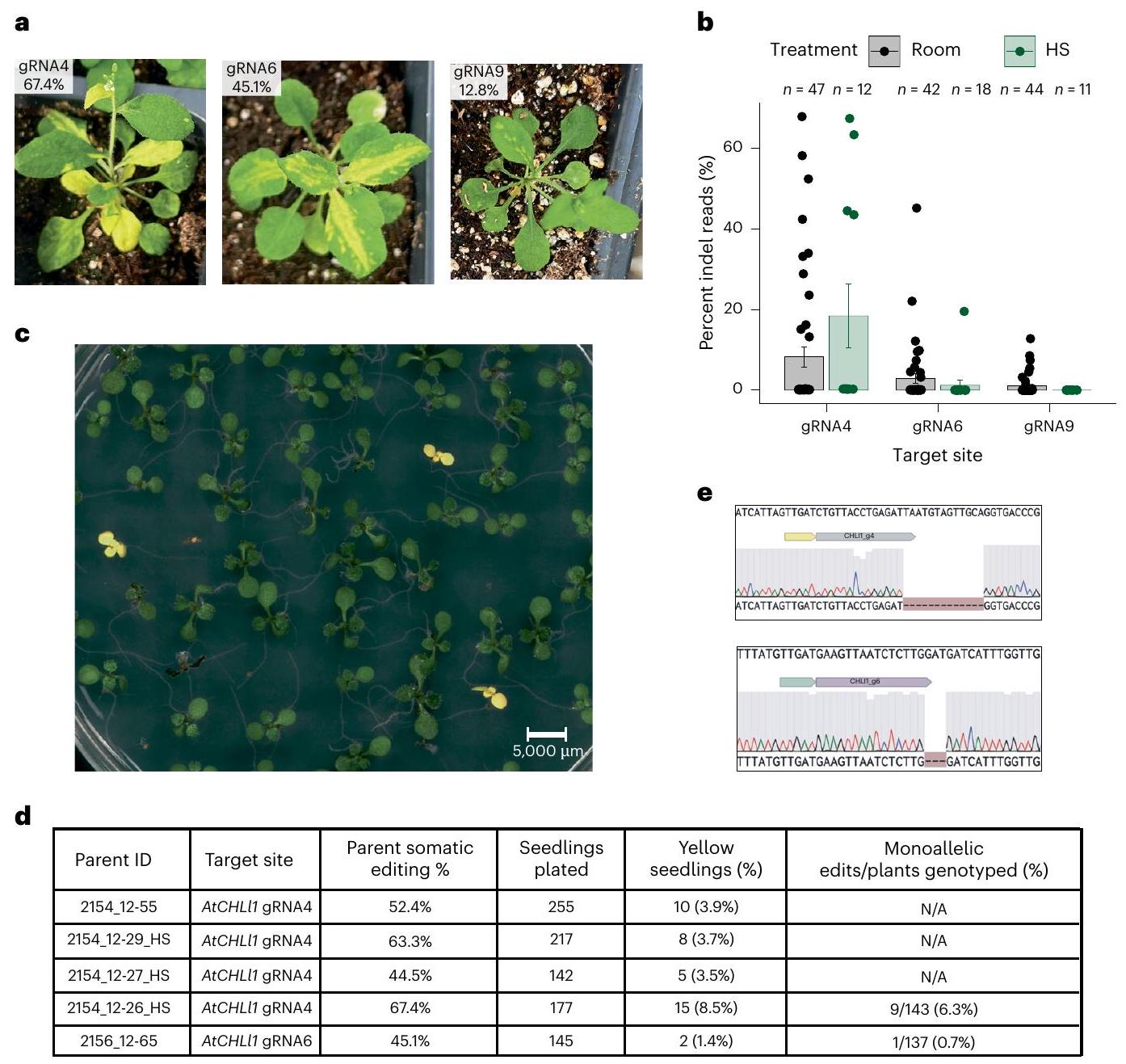
Fig. 3 | Somatic and heritable editing in Arabidopsis using TRV to deliver ISYmu1 TnpB and guide RNA targeting AtCHLI1. a, Representative pictures of plants displaying yellow sectors weeks after TRV delivery. The gRNA and somatic editing efficiency is indicated in the upper left corner of each picture. b, Barplot displaying the somatic editing efficiencies ( ± s.e.m.) for ISYmu1 gRNA4, gRNA6 and gRNA9 in WT. The gRNA target site is plotted along the axis. The axis indicates the editing efficiencies (percent indel reads). Each dot represents an individual plant that underwent agroflood TRV delivery.
c, Representative image of yellow and green progeny seedlings from a WT plant
showing 67.4% somatic editing using the TRV2 Architecture_B design with gRNA4 that underwent heat-shock treatment.d, Table summarizing the transmission of edited alleles from four and one individual plants that underwent agroflood delivery using ISYmu1 gRNA4 and gRNA6, respectively. e, Representative Sanger sequencing trace file screenshots from a yellow plant harbouring an edit at gRNA4 (top) or gRNA6 (bottom). For each panel: top, wild-type sequence; middle, the ISYmu1gRNA target and PAM; bottom, the ab1 trace file displaying a homozygous deletion.
Next, we tested gRNA12 utilizing the TRV2 Architecture_B, since this architecture demonstrated the highest levels of editing for gRNA2. Using the same agroflood TRV delivery method to WT plants, we observed an average of and editing efficiency in room-temperature and heat-treatment growth conditions, respectively (Fig. 2d). Further, 6/57 plants displayed editing greater than , with 4 plants showing greater than editing when room-temperature treatment was used (Fig. 2d). Again, analysis of the repair outcomes showed deletion-dominant profiles for ISYmu1gRNA2 and gRNA12 (Fig. 2e,f).
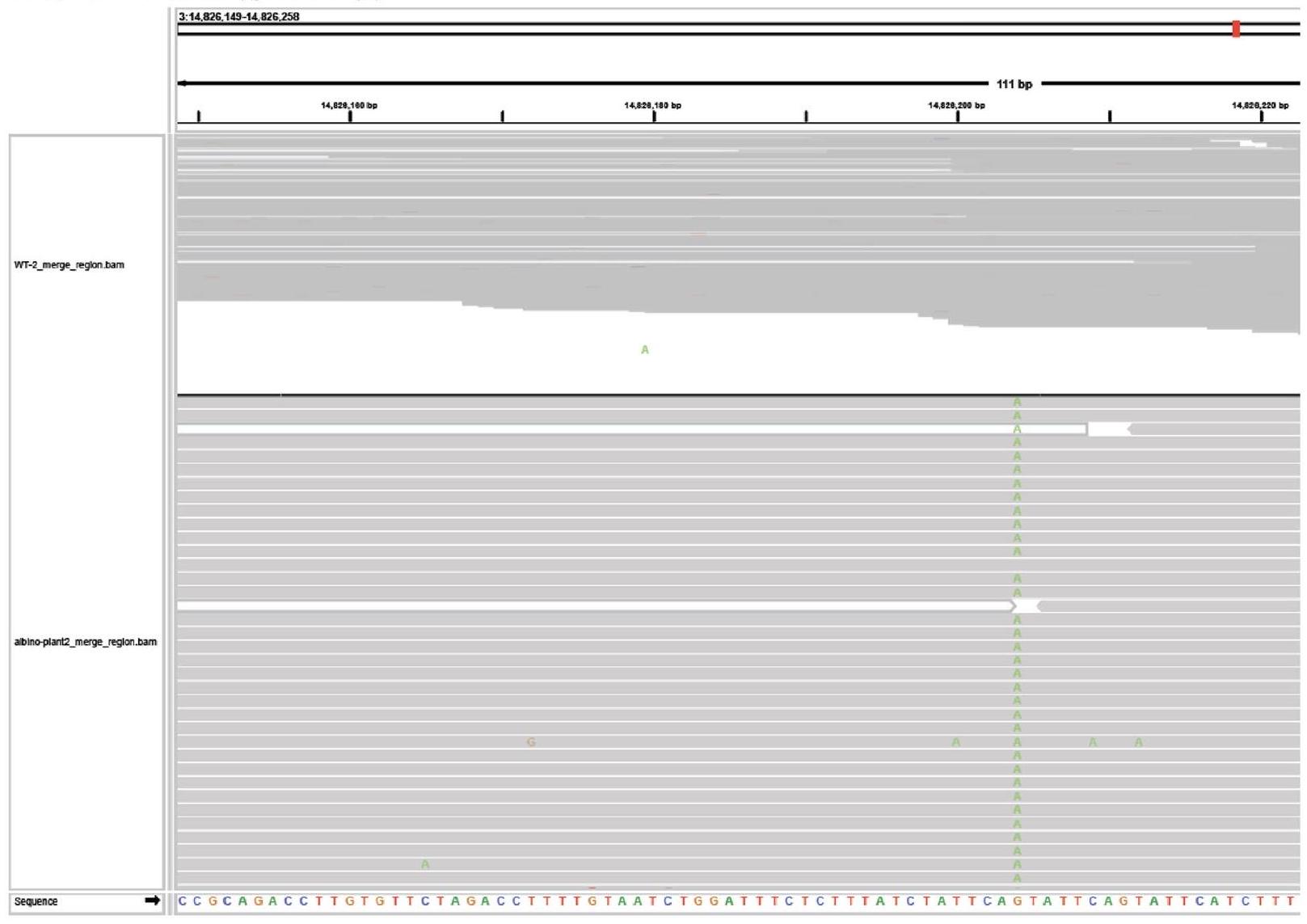
To test for transmission of edited alleles to the next generation, we first screened the progeny of a WT plant showing somatic editing using the TRV2 Architecture_B design with gRNA2 that underwent heat-shock treatment. In total, 2,318 seeds were sown on MS plates containing sucrose. After 10 days, 68 albino seedlings were observed, suggesting biallelic mutations in the PDS3 gene (Fig. 2g). To confirm that AtPDS3 was mutated, we performed Sanger sequencing on the two white seedlings shown in Fig. 2g, which revealed both plants to be homozygous for a 4-bp frame-shift deletion (Fig. 2h).
To further characterize transmission of edited alleles, amp-seq on 209 seedlings ( 41 albino and 168 green) showed that all of the albino seedlings contained biallelic mutations, with the majority of mutations being the 4-bp deletion observed in Fig. 2h (Supplementary Table 2). Of the 168 green seedlings, 8 were heterozygous ( 4 -bp deletion in WT) (Supplementary Table 2).
Next, we characterized transmission of edited alleles from two individual lines, plant 54 ( somatic editing) and plant 69 (77.1% somatic editing), that underwent agroflood using gRNA12 TRV2 Architecture B with the room-temperature condition. As expected, we did not observe any albino seedlings, probably because this target site is located upstream of the AtPDS3 transcription start site. Using Sanger sequencing, we analysed the genotypes of 148 and 75 progeny seedlings from plants 54 and 69, respectively. Sanger sequencing analysis of the progeny from plant 54 revealed biallelic and monoallelic edited plants (Fig. 2i, Supplementary Table 2 and Extended Data Fig. 5a) . For plant 69, we observed higher transmission of edited alleles, totalling biallelic and monoallelic edited plants
(Fig. 2i, Supplementary Table 2 and Extended Data Fig. 5b) . These data demonstrate the heritability of edits generated via TRV delivery of ISYmu1 at two distinct target sites.
To test the applicability of this approach to another locus, we designed six ISYmu1 gRNAs targeting the AtCHLl1 gene (AT4G18480) (Supplementary Table 1).AtCHLl1 was chosen due to the obvious yellow phenotypic readout that AtCHLl1 homozygous mutant plants display . The agroflood method was used to individually deliver TRV Architecture_B vectors targeting each of the six AtCHLl1 sites. Plants were exposed to either room temperature or heat-shock growth conditions as previously described. About 2 weeks post agroflood, we observed yellow sectors on some of the plants infected with TRV ISYmu1 gRNA4, gRNA6 and gRNA9 (Fig.3a). To quantify somatic editing, tissue samples from plants infected with TRV targeting gRNA4, gRNA6 and gRNA9 were collected for amp-seq analysis. We observed an average of and somatic editing for gRNA4, gRNA6 and gRNA9 for the room-temperature condition, respectively (Fig. 3b). For the plants that underwent the heat shock, we detected an average editing frequency of and 0% for gRNA4, gRNA6 and gRNA9, respectively (Fig. 3b). Further, 4/47 (8.5%) and 4/12 (33.3%) plants infected with ISYmu1gRNA4 displayed somatic editing greater than for room-temperature and heat-shock samples, respectively (Fig. 3b).
Next, we screened the progeny from plants infected with TRV targeting gRNA4 and gRNA6 to quantify transmission of edited alleles. Seedlings were grown on MS plates containing sucrose, and after 10 days we observed yellow seedlings, consistent with the biallelic mutation genotype of this gene (Fig. 3c) . In total, we observed yellow progeny from plants infected with TRV ISYmu1 gRNA4 (Fig. 3d). For the progeny of a plant infected with gRNA6, we observed fewer yellow seedlings, totalling 2/145 (1.4%) (Fig. 3d). Sanger sequencing revealed that all of the yellow plants harboured biallelic mutations at the gRNA4 or gRNA6 target site (Fig. 3e, Extended Data Fig. 5c and Supplementary Table 2). Next, Sanger sequencing revealed that 9/143 ( ) and 1/137 (0.7%) green seedlings from plants 2154_12-26_HS (gRNA4) and 2156_12-65 (gRNA6), respectively, contained monoallelic edits (Fig. 3d and Supplementary Table 2). These data indicate that TRV-mediated editing with ISYmu1 is capable of generating targeted somatic mutations at three target sites of the AtCHLl1 gene, and that edited alleles can be transmitted to the next generation.
It has been demonstrated that TRV is not transmitted to the next generation following agroflood inoculation of plants . To confirm that the TRV was not present in the progeny of a TRV-infected plant, RT-PCR was performed on 5 albino plants harbouring homozygous 4-bp deletions at AtPDS3. Consistent with the literature, TRV was not detected in any of the albino plants (Extended Data Fig. 6) . These data indicate that TRV-mediated biallelic edits using ISYmu1 are heritable and virus-free.
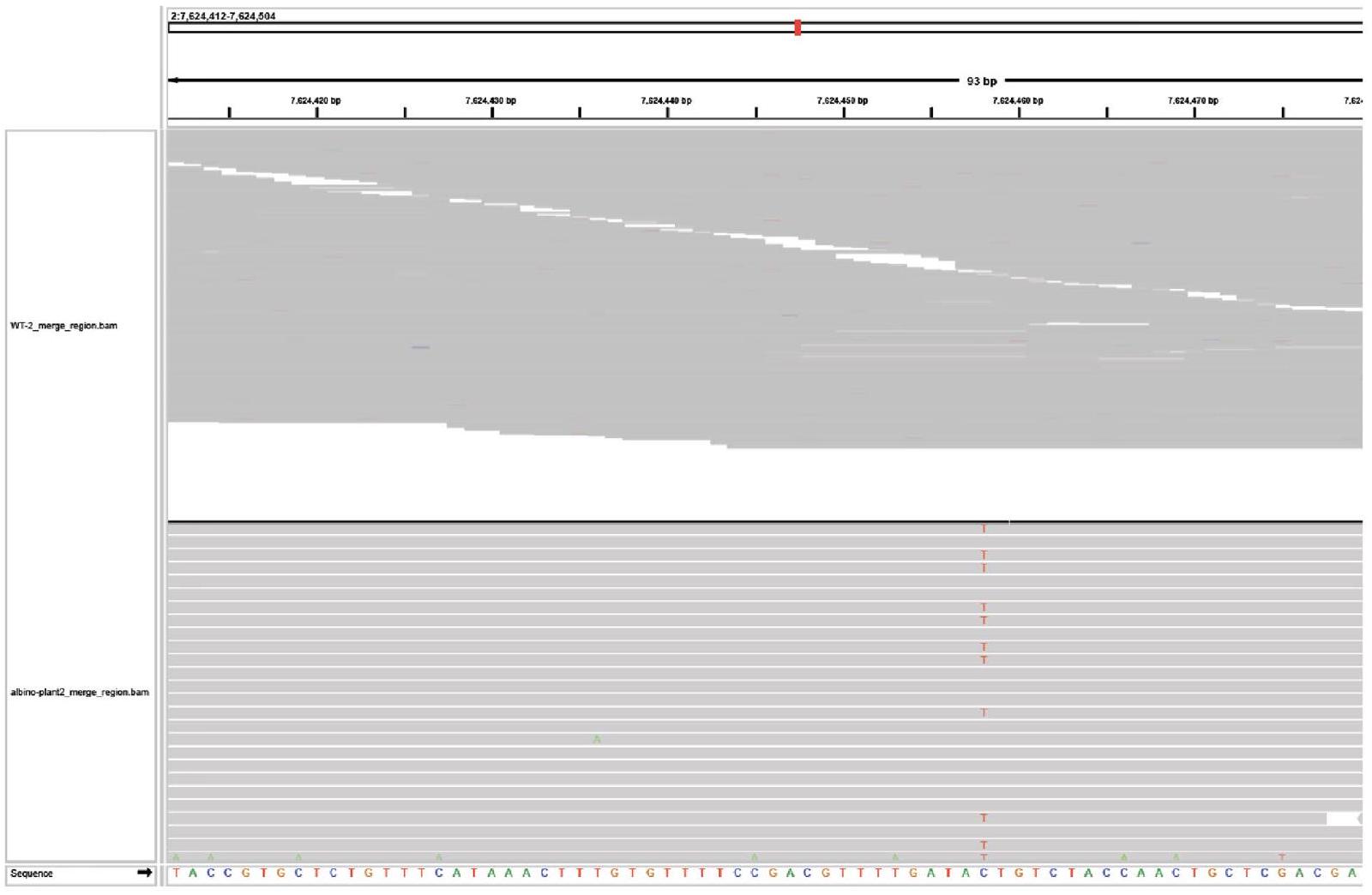
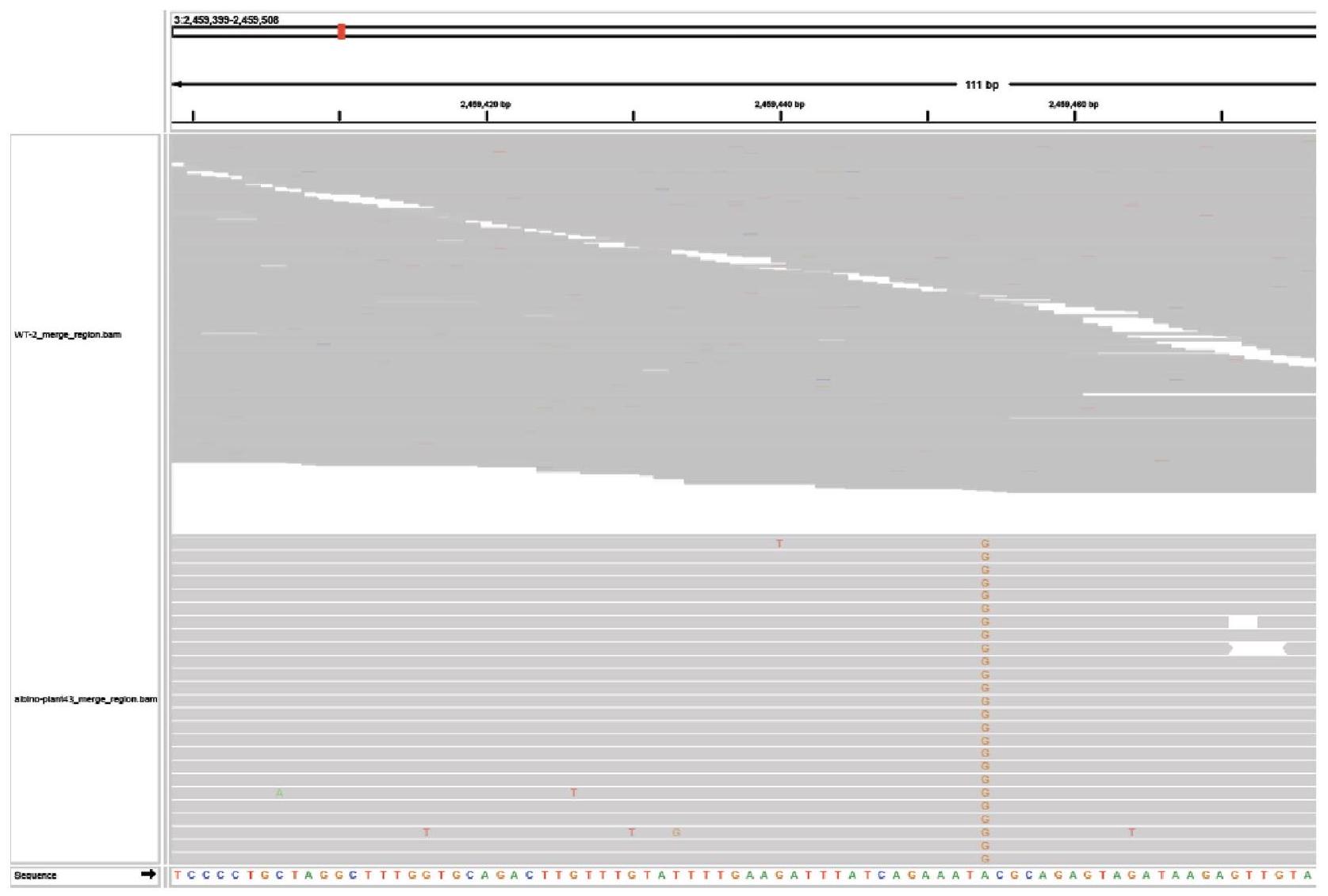
To evaluate off-target editing, we surveyed 3 individual albino plants harbouring biallelic mutations generated by ISYmu1 TRV2 Architecture_B gRNA2. Whole-genome sequencing was performed to generate an average of coverage, with greater than of the genome covered by mapped reads (Supplementary Table 3). In all 3 samples, we confirmed the targeted mutations in the AtPDS3 gene, as previously identified using amp-seq. In addition, we found a large number of variant differences compared with the Col-O reference genome both in the control and the edited plants (Supplementary Table 4), suggesting that most of the variants detected are due to spontaneous mutations present in our lab strain of Arabidopsis. To screen for variants potentially caused by ISYmu1 off-target editing, all variants in the edited plants were filtered with variants already present in the control background. Variants with coverage lower than 30 -fold were also filtered out. The remaining variants were checked manually for any false positive variant calling. In the 3 albino plants we sequenced, only 5, 5 and 4 variants were detected, and these variants are all outside the predicted potential off-target sites based on sequence similarity
to the AtPDS3 gRNA2 sequence (Extended Data Figs. 7-10, and Supplementary Tables 4 and 5. In line with ISYmu1 off-target analysis reported in rice and human cells , these data further demonstrate the high target-site specificity of ISYmu1.
A long-term goal of plant scientists has been the development of fast and easy means of editing plant genomes without the need for tissue culture and transgenesis. Very recently, low levels of tissue-culture-free heritable gene editing was demonstrated by delivering Cas9 and the gRNA to Nicotiana benthamiana using the tobacco ringspot virus (TRSV) . They improved heritability by co-delivering the Cas9-gRNA TRSV with an rdr6virus-induced gene silencing knockdown sequence on the apple latent spherical virus (ALSV) . Here we developed a streamlined and easy-to-use approach utilizing the ultracompact site-specific TnpB genome editor, ISYmu1, together with tobacco rattle virus, for heritable plant genome editing. These results should accelerate high-throughput genome editing for both basic and applied research. We anticipate this approach to be applicable to other novel TnpBs, various viral vectors and a number of plant species for genome editing. Recent work has uncovered many TnpB systems from diverse microbial sources, including enzymes with unique protospacer adjacent motif (PAM) sequence specificities , which can increase the range of target DNA sequences that could be edited using this approach. The TRV virus used in this study has a broad host range of over 400 species, including many solanaceous plants such as tomato, ornamental plants and other crops . In addition, plant viruses with similar cargo capacities, such as potato virus and barley stripe mosaic virus, are likely to be amenable to this approach since it has been demonstrated that they are capable of viral-mediated heritable gene editing by delivering the gRNA to a Cas9-expressing transgenic plant . Further, because this approach can create sectors of tissue harbouring somatic biallelic edits, it may also serve as a tool to enable the study of genes that cause embryonic lethality or severe pleiotropic effects as homozygous mutants. Finally, in addition to being an important tool for crop biotechnology, viral delivery of TnpBs could enable high-throughput CRISPR screens in model plant species such as Arabidopsis, further unlocking their potential for genetic discovery.
Methods
Plasmids used in this study
Plasmids used for bacterial assay were generated as follows. The single expression cassette containing TnpB and RNA sequence were synthesized as geneblocks from Integrated DNA Technologies (IDT) and were golden-gate cloned using Bsmbl restriction enzyme (E1602S) into a vector (chloramphenicol resistance) under a single tetracycline-inducible promoter (TetR/pTet) to make the TnpB- RNA plasmid. Target sites with various PAM sequences and target sites were golden-gate cloned with BbsI restriction enzyme (R3539S) into a vector (ampicillin/carbenicillin resistance).
Plasmids were generated for protoplast and floral dip experiments in a two-step cloning strategy. In step one, the ISDra2, ISYmu1 and ISAam1 protein coding sequences and their RNAs were synthesized as geneblocks by IDT. Then, starting with the pC1300_pUB10_pcoCASphi_E9t_MCS_version2 vector , we used NEBuilder HiFi DNA Assembly (catE2621) and PCR to assemble the TnpB- RNA geneblocks into plant expression vectors with a toxic insert flanked by PaqCI sites immediately downstream of the RNA scaffold and preceding an HDV ribozyme sequence. The HiFi reactions were then transformed into One Shot ccdB Survival 2 T1R Competent Cells (A10460) to obtain the pMK003 (ISDra2), pMK025 (ISYmu1) and pMK024 (ISAam1) intermediate vectors for facile guide sequence cloning (Supplementary Table 6). In step two, guide sequences were synthesized as individual top and bottom strands with 4 base pair overhangs from IDT, phosphorylated and annealed, and then used for golden-gate assembly using the NEB PaqCI (R0745) enzyme (Supplementary Table 7). When transformed into NEB10-beta competent E.coli (C3019), vectors that still contained
the gene would kill the cells, leaving behind only the transformants possessing successfully assembled TnpB plant expression vectors harbouring a guide RNA sequence.
TRV vectors targeting AtPDS3 were created with the pDK3888 TRV2 plasmid as a base vector . NEBuilder HiFi DNA Assembly (E2621) was used to clone the ISYmu1gRNA2 Architecture_A, ISYmu1 gRNA2 Architecture_B and ISYmu1 gRNA12 Architecture_B into the TRV2 cargo slot. First, pDK3888 was digested using NEB Zral (R0659), NEB PmII (R0532) and NEB Quick CIP (M0525) overnight, and purified using Qiagen QiaQuick purification column (28104). Next, three PCR reactions were performed to amplify the fragments needed for NEBuilder HiFi DNA Assembly, followed by purification using a Qiagen QiaQuick purification column (28104) (Supplementary Table 8). Then, the digested and purified pDK3888 plasmid and purified PCR fragments were used to assemble the final TRV2 plasmid using NEBuilder HiFi DNA Assembly according to manufacturer protocol. Finally, the NEBuilder HiFi DNA Assembly reaction was transformed into NEB 10-beta competent E. coli (C3019). TRV2 vectors targeting AtCHLl1 were created using golden-gate assembly . Two oligos corresponding to the target site were phosphorylated and annealed. Then, the annealed double-stranded DNA was used in a PaqCI (R0745S) golden-gate reaction with the pMK435 intermediate vector (Supplementary Table 8). The golden-gate reaction was then transformed into NEB 10-beta competent E. coli (C3019). Correct plasmids were confirmed using Primordium whole-plasmid sequencing. Plasmids and their descriptions can be found in Supplementary Table 6.
Bacterial interference assay
For the bacterial interference assay, we co-transfected 100 ng of the TnpB- RNA plasmid and 100 ng of the target plasmid to of NEB 10-beta electrocompetent E. coli cells (C3020K). Specifically, the target plasmid contains a target site either flanking the canonical PAM (TTGAT for ISYmu1 and ISDra2 and TTTAA for ISAam1) or flanking a non-canonical PAM (GGGGG). The cells were recovered in 1 ml of NEB 10-Beta Stable/Outgrowth media (B9035S) for 1 h . Following recovery, a series of 5-fold dilutions of the recovery culture were prepared. Each dilution was spot plated onto LB-agar plates containing double antibiotics ( chloramphenicol, carbenicillin and 2 nM anhydrotetracycline) and onto control plates with a single antibiotic ( chloramphenicol and 2 nM anhydrotetracycline). If no colonies were visible on the serial dilution plates, of the 1 ml recovery culture was plated entirely on the double antibiotic plate to enhance detection sensitivity. Plates were left overnight at either or , and colony-forming units (c.f.u.s) were counted on all plates the next morning. The normalized c.f.u.s were calculated by taking the ratio of c.f.u.s on the double antibiotic plates to the c.f.u.s on the single antibiotic plates. The normalized c.f.u.s in the canonical PAM conditions were compared to those in the non-canonical PAM conditions. Experiments were performed in triplicate.
Plant materials and growth conditions
For protoplast preparation, Arabidopsis Columbia ecotype (Col-0) seeds were suspended in a agarose solution and kept at in the dark for 3 days to stratify. Following stratification, seeds were planted onJiffy pucks and grown under a light/dark photoperiod with low-light condition at for 3-4 weeks .
For the creation of transgenic plants, the Arabidopsis Col-O ecotype was used. The (SALK_123114) genotype was obtained from Feng Zhang lab at the University of Minnesota. The rdr6genotype was created using CRISPR-Cas9, resulting in a 616-bp deletion in the gene body of rdr6. Floral dip transformation was performed according to the protocol as previously outlined using the AgI0 Agrobacterium strain . Transgenic T1 plants were screened using plates with hygromycin B under a light/dark cycle at . After 1 week,
transgenic seedlings that passed selection were transferred to soil and moved to a greenhouse ( ) for the rest of their life cycle.
For agroflood experiments, sterilized seeds were sown on MS agar plates and stratified for 5 days. After 5 days, the seeds were moved to a growth room and grown under a light/dark cycle at for 8-10 days. The seedlings were then used for TRV delivery.
A subset of transgenic T1 plants and plants that underwent agroflood were subjected to a heat-shock treatment modified from ref. 37. Seedlings that passed selection or underwent agroflood were then transplanted to soil and grown in a greenhouse ( ) for 1 week. After 1 week, plants that did not receive a heat-shock treatment continued to grow in the greenhouse ( ); however, plants that underwent heat-shock treatment were exposed to of heat exposure at every day for 5 days, followed by 2 days of recovery at a greenhouse ( ). This heat-shock regime lasted for 2 weeks.
Protoplast isolation and transfection
Arabidopsis mesophyll protoplast isolation was performed as previously described . Plasmid transfections into Arabidopsis protoplasts were performed using of plasmid following ref. 35. The concentrations of plasmids were determined using a nanodrop spectrophotometer. Plasmids were added to the bottom of each transfection tube, and the volume of the plasmids was supplemented with water to reach . Protoplasts were added, followed by of fresh and sterile polyethylene glycol (PEG)- solution. The samples were mixed by gently tapping the tubes and incubated at room temperature for 10 min . After of W5 solution was added and mixed with the protoplasts by inverting the tube two to three times to stop the transfection. Next, protoplasts were collected by centrifuging the tubes at 100 relative centrifugal force (RCF) for 2 min and resuspended in 1 ml of WI solution. The protoplast cells were then plated in 6-well plates precoated with calf serum. Protoplast cells in the 6-well plates were incubated at for 48 h . During the 48 -h incubation, the protoplast cells were subjected to a heat-shock treatment for 2 h at 16 h post transfection. At 48 h post transfection, protoplasts were collected for genomic DNA extraction.
TRV delivery to Arabidopsis seedlings
TRV delivery was performed as previously described . TRV1 and TRV2 vectors were first introduced into the GV3101Agrobacterium strain. The Agrobacterium harbouring TRV vectors were then grown in 200 ml of lysogeny broth (LB) with antibiotics for 18 h at . Agrobacterium cultures were centrifuged for 20 min at . The LB was discarded and the Agrobacterium cells were resuspended in 200 ml of sterile water. The resuspended Agrobacterium was centrifuged for 10 min at . The supernatant was discarded and the pellet was resuspended in sterile agro-infiltration buffer containing , 10 mM 2 -( -morpholino) ethanesulfonic acid and acetosyringone to otical density . The Agrobacterium cells were then incubated at for 3 h with slow shaking. After 3 h , the Agrobacterium harbouring TRV1 and TRV2 were mixed in a 1:1 ratio, and 15 ml of this 1:1 mixture of TRV was delivered to seedlings at 8-10 days old. After 4 days of agroflood co-culture, seedlings were transplanted to soil.
Screening the progeny of TRV-infected plants for edits
Seeds were harvested from the TRV-infected plants weeks after TRV delivery. The seeds were sown on MS plates supplemented with sucrose and stored at in the dark for 5 days to stratify. After 5 days, the seeds were moved to a growth room and grown under a light/dark cycle at for days. Next, a subset of plants was sampled for genotyping. A single piece of leaf tissue was sampled, and DNA was extracted using Invitrogen Platinum Direct PCR Universal Master Mix (A44647500) according to manufacturer instructions. The DNA was then used for amp-seq or Sanger sequencing using primers listed in Supplementary Table 9.
Next-generation amplicon sequencing
DNA was extracted from protoplast samples with Qiagen DNeasy plant mini kit (Qiagen, 69106). Tissue was collected from transgenic plants by sampling and pooling leaf tissue from 3 random leaves on a single plant 3 weeks after being transplanted to soil. For the plants that underwent agroflood, leaf tissue was sampled by collecting and pooling tissue from 3 random (however, if white or yellow sectors were visible, they were sampled) leaves on a single plant distal to the TRV delivery site 3 weeks after being transplanted to soil. Once tissue samples were collected, they were frozen at overnight. The samples were then ground and DNA was extracted using the Invitrogen Platinum Direct PCR Universal Master Mix (A44647500) according to manufacturer instructions. For the progenies of plants that underwent agroflood, a single leaf tissue was sampled and DNA was extracted using Invitrogen Platinum Direct PCR Universal Master Mix (A44647500) according to manufacturer instructions. The DNA was then used for next-generation amplicon sequencing.
Following ref. 35, editing efficiency was characterized using single-end next-generation sequencing on the Illumina NovaSeqX platform. Libraries were prepared via a 2 -step PCR amplification method. In the first round of amplification, each target site was amplified using primers flanking the target site (Supplementary Table 9). After 25 cycles of amplification, the reactions were cleaned using Ampure XP bead purification (Beckman Coulter, A63881). Next, each sample went through 12 additional cycles of amplification using Illumina indexing primers. The samples were cleaned using Ampure XP bead purification. Samples were checked for purity on a agarose gel, quantified using a nanodrop spectrophotometer, normalized and pooled.
Next-generation amplicon sequencing analysis
Amplicon sequencing analysis was performed following ref. 35. Single-end reads were used for analysis. Reads were adapter trimmed using Trim Galore default settings. Remaining reads were mapped to the target genome region using the BWA aligner (v.0.7.17, BWA-MEM algorithm). Sorted and indexed bam files were used as input files for further analysis using the CrispRvariants R package (v.1.14.0). Each mutation pattern with corresponding read counts was exported using the CrispRvariants R package. After assessing all control samples, a criterion to classify reads as edited was established: only reads with -bp deletion or insertion (indel) of the same pattern (indels of same size starting at the same location) with read counts from a sample were counted as edited reads. Single nucleotide variants were also filtered out.
Off-target analysis
Off-target analysis was performed as previously described . DNA from single Arabidopsis seedlings was extracted with the Qiagen DNeasy plant mini kit and sheared to size with a Covaris sonicator. Library preparation was performed with a Tecan Ovation Ultralow V2 DNA-seq kit. For variant calling, WGS reads were aligned to the TAIR10 reference genome using BWA mem (v.0.7.17) with default parameters. GATK (4.2.0.0) MarkDuplicatesSpark was used to remove PCR duplicate reads. Then GATK HaplotypeCaller was used to call raw variants. Raw single nucleotide polymorphisms (SNPs) were filtered with QD < 2.0, FS > 60.0, MQ < 40.0 and SOR > 4.0. Raw InDels were filtered with QD < 2.0, FS > 200.0 and SOR and used for base quality score recalibration. The recalibrated bam was further applied to GATK and Strelka (v.2.9.2) SNPs/InDel calling. Only SNPs/InDels called by both GATK and Strelka were used for further filtering. The intersection of SNPs/ InDel called by GATK with Strelka (v.2.9.2) was obtained using BedTools (v.2.26.0) . SNPs/InDel were filtered with wild-type background using BedTools (v.2.26.0). Variants with depth coverage lower than 30 were filtered.
RT-PCR
Total RNA from TRV-infected progeny plants was extracted using Zymo Research Direct-zol RNA MiniPrep kit (R2052). Total RNA was converted to cDNA using the Invitrogen SuperScript IV VILO Master Mix (11766050). The RT-PCR control was performed using primers targeting the AtIPP2 gene (Supplementary Table 10). PCR was performed to check for the presence/absence of the TRV vector using SP9238 and SP9239 (Supplementary Table 10) . PCR was performed with New England Biolabs Q5 High-Fidelity Master Mix (M0492L) according to manufacturer instructions, using of cDNA in a reaction. PCR conditions included a initial denaturation step for , ) and for 2 min . PCR amplicons ( ) were analysed using agarose gel electrophoresis.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All the amp-seq data generated in this study are accessible at NCBI Sequence Read Archive under BioProject PRJNA1124592. Whole-genome sequencing data are accessible at BioProject PRJNA1146711. Source data are provided with this paper.
References
Altpeter, F. et al. Advancing crop transformation in the era of genome editing. Plant Cell 28, 1510-1520 (2016).
Chen, K., Wang, Y., Zhang, R., Zhang, H. & Gao, C. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 70, 667-697 (2019).
Nasti, R. A. & Voytas, D. F. Attaining the promise of plant gene editing at scale. Proc. Natl Acad. Sci. USA 118, e2004846117 (2021).
Honig, A. et al. Transient expression of virally delivered meganuclease in planta generates inherited genomic deletions. Mol. Plant 8, 1292-1294 (2015).
Marton, I. et al. Nontransgenic genome modification in plant cells. Plant Physiol. 154, 1079-1087 (2010).
Ali, Z. et al. Efficient virus-mediated genome editing in plants using the CRISPR/Cas9 system. Mol. Plant 8, 1288-1291 (2015).
Ellison, E. E., Chamness, J. C. & Voytas, D. F. Viruses as Vectors for the Delivery of Gene-editing Reagents (Burleigh Dodds Science Publishing, 2021).
Beernink, B. M., Lappe, R. R., Bredow, M. & Whitham, S. A. Impacts of RNA mobility signals on virus induced somatic and germline gene editing. Front. Genome Ed. 4, 925088 (2022).
Uranga, M. et al. Efficient Cas9 multiplex editing using unspaced sgRNA arrays engineering in a Potato virus X vector. Plant J. 106, 555-565 (2021).
Bradamante, G., Mittelsten Scheid, O. & Incarbone, M. Under siege: virus control in plant meristems and progeny. Plant Cell 33, 2523-2537(2021).
Ma, X., Zhang, X., Liu, H. & Li, Z. Highly efficient DNA-free plant genome editing using virally delivered CRISPR-Cas9. Nat. Plants 6, 773-779 (2020).
Liu, Q., Zhao, C., Sun, K., Deng, Y. & Li, Z. Engineered biocontainable RNA virus vectors for non-transgenic genome editing across crop species and genotypes. Mol. Plant 16, 616-631 (2023).
Lee, S.-Y. et al. Development of virus-induced genome editing methods in solanaceous crops. Hortic. Res. 11, uhad233 (2023).
Ariga, H., Toki, S. & Ishibashi, K. Potato virus X vector-mediated DNA-free genome editing in plants. Plant Cell Physiol. 61, 1946-1953 (2020).
Ghoshal, B. et al. A viral guide RNA delivery system for CRISPR-based transcriptional activation and heritable targeted DNA demethylation in Arabidopsis thaliana. PLoS Genet. 16, e1008983 (2020).
Karvelis, T. et al. Transposon-associated TnpB is a programmable RNA-guided DNA endonuclease. Nature 599, 692-696 (2021).
Altae-Tran, H. et al. The widespread IS200/IS605 transposon family encodes diverse programmable RNA-guided endonucleases. Science 374, 57-65 (2021).
Meers, C. et al. Transposon-encoded nucleases use guide RNAs to promote their selfish spread. Nature 622, 863-871(2023).
Li, Q. et al. Genome editing in plants using the TnpB transposase system. aBIOTECH 5, 225-230 (2024).
Xiang, G. et al. Evolutionary mining and functional characterization of TnpB nucleases identify efficient miniature genome editors. Nat. Biotechnol. 42, 745-757 (2024).
Li, Z. et al. Engineering a transposon-associated TnpB-ωRNA system for efficient gene editing and phenotypic correction of a tyrosinaemia mouse model. Nat. Commun. 15, 831 (2024).
Yoo, S.-D., Cho, Y.-H. & Sheen, J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat. Protoc. 2, 1565-1572 (2007).
Zhang, X., Henriques, R., Lin, S.-S., Niu, Q.-W. & Chua, N.-H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 1, 641-646 (2006).
Wang, M. et al. A gene silencing screen uncovers diverse tools for targeted gene repression in Arabidopsis. Nat. Plants 9, 460-472 (2023).
Ellison, E. E. et al. Multiplexed heritable gene editing using RNA viruses and mobile single guide RNAs. Nat. Plants 6, 620-624 (2020).
Nagalakshmi, U., Meier, N., Liu, J.-Y., Voytas, D. F. & Dinesh-Kumar, S. P. High-efficiency multiplex biallelic heritable editing in Arabidopsis using an RNA virus. Plant Physiol. 189, 1241-1245 (2022).
Qi, Y. et al. Increasing frequencies of site-specific mutagenesis and gene targeting in Arabidopsis by manipulating DNA repair pathways. Genome Res. 23, 547-554 (2013).
Brinkman, E. K. & van Steensel, B. Rapid quantitative evaluation of CRISPR genome editing by TIDE and TIDER. Methods Mol. Biol. 1961, 29-44 (2019).
Huang, Y.-S. & Li, H.-M. Arabidopsis CHLI2 can substitute for CHLI1. Plant Physiol. 150, 636-645 (2009).
Bae, S., Park, J. & Kim, J.-S. Cas-OFFinder: a fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 30, 1473-1475 (2014).
Yoshida, T., Ishikawa, M., Toki, S. & Ishibashi, K. Heritable tissue-culture-free gene editing in Nicotiana benthamiana through viral delivery of SpCas9 and sgRNA. Plant Cell Physiol. 65, 1743-1750 (2024).
Badon, I. W., Oh, Y., Kim, H.-J. & Lee, S. H. Recent application of CRISPR-Cas12 and OMEGA system for genome editing. Mol. Ther. 32, 32-43 (2024).
Macfarlane, S. A. Tobraviruses-plant pathogens and tools for biotechnology. Mol. Plant Pathol. 11, 577-583 (2010).
Zhang, C., Liu, S., Li, X., Zhang, R. & Li, J. Virus-induced gene editing and its applications in plants. Int. J. Mol. Sci. 23, 10202 (2022).
Li, Z. et al. Genome editing in plants using the compact editor Cas Ф. Proc. Natl Acad. Sci. USA 120, e2216822120 (2023).
Čermák, T. et al. A multipurpose toolkit to enable advanced genome engineering in plants. Plant Cell 29, 1196-1217 (2017).
LeBlanc, C. et al. Increased efficiency of targeted mutagenesis by CRISPR/Cas9 in plants using heat stress. Plant J. 93, 377-386 (2018).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754-1760 (2009).
McKenna, A. et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297-1303 (2010).
Kim, S. et al. Strelka2: fast and accurate calling of germline and somatic variants. Nat. Methods 15, 591-594 (2018).
Quinlan, A. R. & Hall, I. M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 26, 841-842 (2010).
Acknowledgements
This work was supported by an NSF Plant Genome Research Program grant (2334027) to S.E.J., J.A.D. and J.F.B. S.E.J. and J.A.D. are investigators of the Howard Hughes Medical Institute. H.S. is an HHMI Fellow of The Jane Coffin Childs Fund for Medical Research. B.A.A. was supported by m-CAFEs Microbial Community Analysis and Functional Evaluation in Soils, a Science Focus Area led by Lawrence Berkeley National Laboratory based on work supported by the US Department of Energy, Office of Science, Office of Biological and Environmental Research (DE-ACO2-O5CH11231). We thank S. Feng, M. Akhavan and the Broad Stem Cell Research Center Biosequencing Core for sequencing support; C. Picard for assistance with analysis of amplicon sequencing data; S. D. Kumar for supplying the TRV2 plasmid pDK3888; F. Zhang for providing the (SALK123114) mutant; and all members of the Jacobsen lab for helpful insight and suggestions.
Author contributions
T.W., M.K., H.S., Z.L., B.A.A., J.F.B., J.A.D. and S.E.J. designed the research. T.W., H.S., Z.Z., J.A.D. and S.E.J. interpreted the data. T.W. and S.E.J. wrote the paper. T.W., M.K., H.S., Z.L., J.A., M.M.S., K.V., G.W., S.C., C.A., N.S., A.S. and D.S. performed experiments.
Competing interests
T.W., M.K, J.A., Z.L., H.S., B.A.A., J.A.D. and S.E.J. have filed a patent (no. WO/2025/007023, 2025) covering aspects of this work. S.E.J. is a cofounder and consultant for Inari Agriculture and a consultant for Terrana Biosciences, Invaio Sciences, Sail Biomedicines, Six Street and Zymo Research. The Regents of the University of California have patents issued and pending for CRISPR technologies on which J.A.D. is an inventor. J.A.D. is a cofounder of Azalea Therapeutics, Caribou Biosciences, Editas Medicine, Evercrisp, Scribe Therapeutics and Mammoth Biosciences. J.A.D. is a scientific advisory board member at Evercrisp, Caribou Biosciences, Scribe Therapeutics, Mammoth Biosciences, The Column Group and Inari. She is also an advisor for Aditum Bio. J.A.D. is Chief Science Advisor to Sixth Street, a Director at Johnson & Johnson, Altos and Tempus, and has a research project sponsored by Apple Tree Partners. The other authors declare no competing interests.
Extended Data Fig. 1|Schematic of bacterial interference assay plasmids containing either TnpB-ωRNA Single or TnpB-ωRNA Single-HDV design. The blue arrow indicates the TnpB sequence; the yellow arrow indicates the RNA sequence; the black rectangle indicates the guide sequence; the
green arrow indicates the HDV ribozyme sequence. The plasmids contain the tetracycline resistance gene (TetR). A tetracycline promoter (pTet) was used to drive expression of the TnpB- RNA single or Single-HDV sequences, and the tetracycline resistance gene.
ISDra2 – HDV
Extended Data Fig. 2 | Plate images of bacterial plasmid interference assay. Five-fold serial dilutions from the 1 mL recovery culture post transformation were plated on both single antibiotic LB-Agar plates (Cam, upper row) and double antibiotic LB-Agar plates (Cam + Carb, lower row).
Plates without visible colonies (indicated by an asterisk) had of the original 1 mL recovery culture plated on the double antibiotic plates (dash insets). Experiments were performed in triplicates.
Extended Data Fig. 3 | Editing efficiency and DNA repair profiles for ISDra2,
ISYmu1, and ISAam1 in protoplast experiment. (a) The name of each TnpB tested is at the upper left of each bar plot. The gRNAs are plotted along the X-axis and the editing efficiency (percent indel reads (%)) is plotted on the Y-axis. Each dot indicates a single transfection. The standard error of the mean (SEM) was calculated for each target site. Two or three replicates were used for each target site tested. (b) DNA repair indel profiles for individual transfection samples.
ISDra2 gRNA9
Reads number
-3-TTTGATAGCTTTGAACCGGT—TTCAGGTTTGACACTCTAGCCCT
17162
-10-TTTGATAGCTTTGAACC———-GGTTTGACACTCTAGCCCT
3654
-9 -TTTGATAGCTTTGAACCGGT——— TTTTGACACTCTAGCCCT
2248
-4-TTTGATAGCTTTGAACCGGTTT—-AGGTTTGACACTCTAGCCCT
962
-9 -TTTGATAGCTTTGAACCIG———GGTTTGACACTCTAGCCCT
958
Total reads number: 1799764 Editing efficiency: 2.28%
Total reads number: 2733829 Editing efficiency: 4.10%
ISAam1 gRNA17
ISAam grNA17
Reads number
Reference
□ □
TCCAITCTIACAITGT T T CIGTIA AIGCAA
-4 -TTTTAATTTGTGTGGTATTTA
—-ATCTACATGTTTCGTAAGCAA
516
-14 -TTTTAATTTGTGTGGTATTTAA
————–TCGTAAGCAA
234
-13 -TTTTAATTTGTGTGGTATTTAA
————-TTCGTAAGCAA
228
-8 -TTTTAATTTGTGTGGTATTTA
220
-9 -ITTTTAAT
TTGTGTGGTATT-
196
The top five most common indel types are listed on the left. The read counts for each indel are listed on the right. The PAM is identified by the red box, and the target site is outlined by the black box, in the Reference sequence. The total read number and editing efficiency are listed below each indel profile. The name of each TnpB and gRNA is displayed above the Reference sequence of each indel repair profile.
ISYmu1 gRNA2
Reference =ITTGAI A AGGGA A TITCIGCCIGC AGA A A AITCCCA A ACACA ACCATI
Extended Data Fig. 4 | Representative DNA repair profiles for individual transgenic plants expressing either ISYmu1 AtPDS3 gRNA2 or gRNA12. The top ten most common indel types are listed on the left. The read counts for each indel are listed on the right. The PAM is identified by the red box, and the target site is outlined by the black box, in the Reference sequence. The total read number and editing efficiency are listed below each indel profile. The name of each TnpB and gRNA is displayed above the Reference sequence for each indel repair profile.
Extended Data Fig. 5 | Sanger sequencing screenshots of the progeny plant genotypes. (a,b) Panels a and b correspond the AtPDS3gRNA12 targeted progeny from plants 54 and 69 , respectively. The sequence at the top is the wild type genomic sequence; below that are the ISYmu1gRNA12 target and PAM (gray box); the ab1 trace file displays the mutation. (c) Each box displays the
genotype of a progeny plant from AtCHLl1gRNA4 editing experiment. The plant ID is indicated above each box. The sequence at the top is the wild type genomic sequence; below that are the ISYmu1 AtCHLl1 gRNA4 target and PAM (yellow box); the ab1 trace file displays the mutation.
Plant 43 heterozygous 3bp deletion
Plants 2, 32, and 43 homozygous 7bp deletion
Extended Data Fig. 7| Potential off-target mutations identified by whole genome sequencing. Genome browser screenshot of the potential edits identified by whole genome sequencing for Plant 43 (heterozygous 3 bp deletion) and Plants 2,32, and 43 (homozygous 7 bp deletion). The mutation is displayed
on the bottom track, and the wild type sequence is displayed on the top track of each screenshot. The plant ID, zygosity, and mutation type is displayed above each screenshot. The genomic location is listed in the upper left corner of each screenshot.
Plants 2, 32, and 43 homozygous SNP (A)
Plant 32 heterozygous SNP (T)
Extended Data Fig. 8|Potential off-target mutations identified by whole genome sequencing. Genome browser screenshot of the potential edits identified by whole genome sequencing for Plants 2,32 , and 43 (homozygous SNP (A)) and Plant 32 (heterozygous SNP (T)). The mutation is displayed on
the bottom track, and the wild type sequence is displayed on the top track of each screenshot. The plant ID, zygosity, and mutation type is displayed above each screenshot. The genomic location is listed in the upper left corner of each screenshot.
Plant 2 heterozygous SNP (T)
Plant 32 heterozygous SNP (T)
Extended Data Fig. 9 | Potential off-target mutations identified by whole genome sequencing. Genome browser screenshot of the potential edits identified by whole genome sequencing for Plant 2 (heterozygous SNP(T)) and Plant 32 (heterozygous SNP(T)). The mutation is displayed on the bottom track,
and the wild type sequence is displayed on the top track of each screenshot. The plant ID, zygosity, and mutation type is displayed above each screenshot. The genomic location is listed in the upper left corner of each screenshot.
Plant 2 homozygous SNP (G)
Plants 2, 32, and 43 homozygous SNP (A)
Extended Data Fig. 10|Potential off-target mutations identified by whole genome sequencing. Genome browser screenshot of the potential edits identified by whole genome sequencing for Plant 2 (homozygous SNP (G)) and Plants 2,32, and 43 (homozygous SNP (A)). The mutation is displayed on
the bottom track, and the wild type sequence is displayed on the top track of each screenshot. The plant ID, zygosity, and mutation type is displayed above each screenshot. The genomic location is listed in the upper left corner of each screenshot.
Research involving human participants, their data, or biological material
Policy information about studies with human participants or human data. See also policy information about sex, gender (identity/presentation), and sexual orientation and race, ethnicity and racism.
Reporting on sex and gender
N.A.
Reporting on race, ethnicity, or other socially relevant groupings
N.A.
Population characteristics
N.A.
Recruitment
N.A.
Ethics oversight
N.A.
Note that full information on the approval of the study protocol must also be provided in the manuscript.
Field-specific reporting
Please select the one below that is the best fit for your research. If you are not sure, read the appropriate sections before making your selection.
Life sciences □ Behavioural & social sciences □ Ecological, evolutionary & environmental sciences
For a reference copy of the document with all sections, see nature.com/documents/nr-reporting-summary-flat.pdf
Life sciences study design
All studies must disclose on these points even when the disclosure is negative.
Sample size
No sample size calculation was performed. For the bacterial interference assay and protoplast experiments we tested at least two replicates (usually three) per treatment to allow for comparison between replicates.For protoplast experiments with two replicates, one of the replicates failed to amplify for NGS. For the transgenic editing experiments we analyzed data from all of the transgenic plants we were able to create using the floral dip method. For the TRV delivery experiments we did not predetermine any sample sizes. The trasngenic and TRV plant samples sizes were determined by the amount of available greenhouse space.
Data exclusions
No data was excluded from this study.
Replication
To verify reproducibility of experiments, biological replicates were performed for each experiment.
Randomization
All samples were randomly collected from the subject materials, except where noted in the methods section. For example, when collecting tissue to quantify somatic editing in plants that underwent TRV delivery, three random leaves distal to the delivery site were collected. However, if white or yellow sectors of cells were present on leaves distal to the TRV delivery site, tissue from those leaves was collected.
Blinding
The investigators were blinded to group allocation during data collection and/or analysis for all experiments.
Reporting for specific materials, systems and methods
We require information from authors about some types of materials, experimental systems and methods used in many studies. Here, indicate whether each material, system or method listed is relevant to your study. If you are not sure if a list item applies to your research, read the appropriate section before selecting a response.
Materials & experimental systems
Methods
n/a
Involved in the study
n/a
–
□
□
〃
ણ
□
□
ણ
Plants
¹Department of Molecular, Cell and Developmental Biology, University of California at Los Angeles, Los Angeles, CA, USA. ²Innovative Genomics Institute, University of California, Berkeley, CA, USA. Howard Hughes Medical Institute, University of California, Berkeley, CA, USA. California Institute for Quantitative Biosciences (QB3), University of California, Berkeley, CA, USA. Department of Earth and Planetary Science, University of California, Berkeley, CA, USA. Department of Environmental Science, Policy and Management, University of California, Berkeley, CA, USA. University of Melbourne, Melbourne, Australia. Department of Molecular and Cell Biology, University of California, Berkeley, CA, USA. Department of Chemistry, University of California, Berkeley, CA, USA. Molecular Biophysics and Integrated Bioimaging Division, Lawrence Berkeley National Laboratory, Berkeley, CA, USA. Li Ka Shing Center for Translational Genomics, University of California, Berkeley, CA, USA. Gladstone Institute of Data Science and Biotechnology, San Francisco, CA, USA. Gladstone-UCSF Institute of Genomic Immunology, San Francisco, CA, USA. Howard Hughes Medical Institute (HHMI), University of California at Los Angeles, Los Angeles, CA, USA. Present address: Ministry of Education Key Laboratory for Bio-Resource and Eco-Environment, College of Life Sciences, State Key Laboratory of Hydraulics and Mountain River Engineering, Sichuan University, Chengdu, China. □ e-mail: jacobsen@ucla.edu