التوصيل غير الجراحي للبيومكرومولكولات عبر الجلد باستخدام الكيتوزان المعدل بالفلوركربون لعلاج الميلانوما المناعي واللقاحات الفيروسية Non-invasive transdermal delivery of biomacromolecules with fluorocarbon-modified chitosan for melanoma immunotherapy and viral vaccines
تم اعتبار توصيل الأدوية عبر الجلد بديلاً لتوصيل الأدوية عن طريق الفم والحقن تحت الجلد. ومع ذلك، لا يزال توصيل البيومكرو جزيئات عبر الجلد بدون إبر يمثل تحديًا. هنا، تم تطوير منصة توصيل عبر الجلد تعتمد على الكيتوزان المعدل بالفوركربون المتوافق حيويًا (FCS) لتحقيق توصيل غير جراحي عالي الكفاءة للبيومكرو جزيئات بما في ذلك الأجسام المضادة والمستضدات. تظهر المعقدات النانوية المتكونة قدرة فعالة على الاختراق عبر الجلد من خلال كل من الطرق بين الخلوية وعبر الملاحق. يؤدي توصيل الأجسام المضادة لحجب نقاط التفتيش المناعية عبر الجلد إلى استجابة مناعية أقوى لسرطان الجلد في الفئران الإناث ويقلل من السمية الجهازية مقارنةً بالحقن الوريدي. علاوة على ذلك، يؤدي توصيل لقاح SARS-CoV-2 عبر الجلد في الفئران الإناث إلى مناعة خلوية مماثلة بالإضافة إلى تحسين المناعة الخلوية وذاكرة المناعة مقارنةً بتلك التي تم تحقيقها مع حقن اللقاح تحت الجلد. بالإضافة إلى ذلك، تظهر أنظمة توصيل البروتين المعتمدة على FCS قدرة على التوصيل عبر الجلد لجلود الأرانب والخنازير. وبالتالي، قد توفر أنظمة التوصيل عبر الجلد المعتمدة على FCS فرصة مثيرة للتغلب على حاجز الجلد من أجل توصيل فعال للعلاجات الحيوية.
الإدارة عبر الجلد تشير إلى توصيل الأدوية بدون إبر عبر الجلد دون أضرار جسدية.لقد تم اعتبارها بديلاً جذابًا لتناول الأدوية عن طريق الفم أو الحقن تحت الجلد، نظرًا لمزاياها الفريدة بما في ذلك عدم التداخل، وتجنب تأثير المرور الأول، والإدارة الخالية من الألم، وتحسين التزام المرضى، وتجنب رهاب الإبر وما إلى ذلك.على الرغم من أن مجموعة متنوعة من المعززات عبر الجلد قد ثبت فعاليتها في العيادات، إلا أن الحمولة الموصلة محدودة للغاية بالأدوية التي تمتلك كتل جزيئية تبلغ بضع مئات من الدالتون وتظهر كارهية قوية للماء.في الوقت الحاضر، لا يزال من الصعب تحقيق توصيل فعال عبر الجلد للمواد المحبة للماء. المواد الحيوية الكبيرة مثل الببتيدات أو البروتينات أو الأحماض النوويةبالإضافة إلى ذلك، فإن توصيل اللقاحات يعد حاليًا واحدًا من أكثر مجالات البحث سخونة في كل من المجتمعات السريرية والعلمية نظرًا لوباء فيروس كورونا 2019 (كوفيد-19).بالمقارنة مع الحقن تحت الجلد التقليدي أو الحقن العضلي، قد تكون طريقة توصيل اللقاحات عبر الجلد نهجًا جذابًا نظرًا لإمكانية الإدارة في المنزل ووجود عدد كبير من خلايا المناعة في الجلد..
لتحقيق توصيل الأدوية البيومكرومولكية عبر الجلد، وخاصة البروتينات، تم استخدام معززات كيميائية جديدة مثل الأغشية تم تطوير الببتيدات النافذة، بالإضافة إلى مجموعة متنوعة من أجهزة تعزيز الجسم، بما في ذلك الموجات فوق الصوتية الكافيتية، والكهرو permeation، والتبخر الحراري، والتقشير المجهري، والإبر الدقيقة.على الرغم من أنه يمكن استخدام مثل هذه الاستراتيجيات للتوصيل عبر الجلد لمجموعة متنوعة من الجزيئات الكبيرة، بما في ذلك البروتينات العلاجية، إلا أنها لا تزال تواجه العديد من المخاوف. على سبيل المثال، تم الإبلاغ عن أن الببتيدات القادرة على اختراق الغشاء تمكن من التوصيل عبر الجلد للبروتينات الصغيرة مثل الأنسولين.ولكن مع كفاءة توصيل غير مرضية، ولا تزال غير فعالة بالنسبة للبروتينات ذات الأوزان الجزيئية الكبيرة. في الوقت نفسه، فإن أجهزة التعزيز الفيزيائي مثل التحفيز الكهربائي والاهتزاز الصوتي لا يمكن تشغيلها ذاتيًا تقريبًا، ولكنها قد تؤدي أيضًا إلى تلف الجلد بسبب النبضات عالية الطاقة.تُعتبر الإبر الدقيقة، التي تشير إلى اللصقات التي تحتوي على العديد من الإبر الصغيرة، قد تم تطبيقها على نطاق واسع في توصيل الأدوية عبر الجلد، حيث أظهرت إمكانيات كبيرة في توصيل الأنسولين ولقاحات الإنفلونزا في السنوات الأخيرة.. ومع ذلك، فإن عملية تصنيع لصقات الإبر الدقيقة ومراقبة الجودة، خاصة مع حمولات البيومكرومولكولات، ستكون معقدة. بالإضافة إلى ذلك، قد تتسبب الإبر الدقيقة في بعض الأضرار الجلدية، مما قد يزيد من خطر العدوى. كما تم الإبلاغ مؤخرًا عن بعض المنصات غير الغازية مثل السوائل الأيونية (ILs) وحمض الهيالورونيك (HAs) لفتح الوصلات الضيقة في الطبقة القرنية وتعزيز النقل بين الخلايا.. ومع ذلك، لا تزال تظهر فعالية أقل عبر الجلد. في عدة دراسات سابقة، تم الإبلاغ عن ناقلات الدهون النانوية مثل الإيثوسومات لتوصيل البروتينات عبر الجلد ضد أورام الجلد.ومع ذلك، سيكون من الجذاب تطوير معززات جديدة ذات أمان وكفاءة عالية لتوصيل البروتينات عبر الجلد.
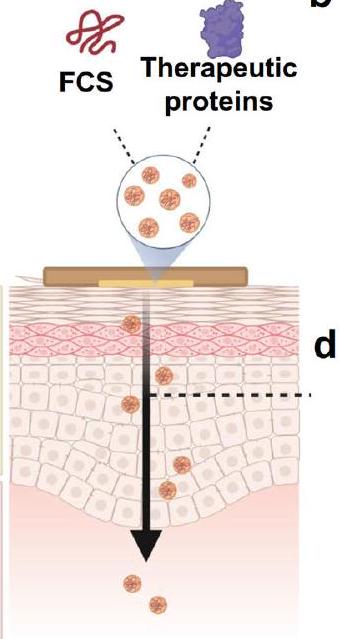
الكيتوزان (CS) هو بوليمر طبيعي كاتيوني قابل للتحلل الحيوي يتمتع بنشاط مضاد للبكتيريا وخصائص لاصقة للمخاط.استلهمت من الفعالية الانتقالية الفعالة للـ CS المعدل بالفلوروكربون (FCS)، كما ورد في دراستنا السابقة لعلاج سرطان المثانة القائم على الحقن داخل المثانة.نحن نفترض أن FCS قد يُستخدم أيضًا لتوصيل البيومكرومولكولات عبر الجلد (الشكل 1أ).
هنا، نكتشف أن FCS يمكن أن يتجمع ذاتيًا مع البيومكرومولكولات مثل البروتينات لتشكيل نانو معقدات، والتي يمكن إضافتها إلى أكوافور.كتركيبة مرهم للتطبيقات الموضعية مع قدرة محسّنة بشكل كبير على اختراق الجلد. ثم نستخدم مثل هذه المنصات القائمة على FCS للتوصيل عبر الجلد لعلاج الأورام المناعية ولقاحات SARS-CoV-2. بمساعدة FCS، يمكن أن يؤدي التوصيل غير الجراحي عبر الجلد لمضاد برنامج الموت المرتبط بالليغاند 1 (aPDL1) إلى تثبيط نمو الأورام المحلية بشكل فعال عند التلامس المباشر مع المرهم المحتوي على FCS/aPDL1. وعند دمجه مع التوصيل المشترك لمضاد بروتين اللمفاويات التائية السامة المرتبطة (aCTLA4)، يمكن أن يحفز هذا المرهم FCS/aPDL1/aCLTA4 استجابات مناعية نظامية قوية لقمع كل من الأورام المحلية والأورام البعيدة. مع استجابات علاجية محسّنة بشكل كبير مقارنة بحقن الأجسام المضادة بنفس الجرعة، قد يؤدي توصيل الأجسام المضادة لمواقع المناعة القائمة على FCS لعلاج الميلانوما إلى تقليل المخاوف بشأن الآثار الجانبية النظامية نظرًا لانخفاض تركيزات المصل نسبيًا من خلال طريق الإدارة الموضعية. علاوة على ذلك، في تجربة إثبات المفهوم، نتحقق من أن FCS يمكن أن يشكل مجمعات نانوية مع بروتين S1 من SARS-CoV-2 كأنتيجين وحمض بولي إينوسينيك: بولي سيتيك (PolyIC)، وهو ربيطة لمستقبلات المناعة الشبيهة بالجراثيم (TLR) 3 كمعزز. يمكن أن يؤدي التطبيق الموضعي لمجمعات FCS/S1/polyIC النانوية إلى تحفيز استجابات مناعية محددة لبروتين S1، تصل إلى مستوى مشابه لذلك الذي تم تحقيقه عن طريق الحقن تحت الجلد لنفس المجمع النانوي. نحن نثبت أيضًا بشكل أولي أن التوصيل عبر الجلد القائم على FCS قد يكون قابلًا للتطبيق على جلود الحيوانات الأكبر مثل الأرانب والخنازير. لذلك، يمثل FCS الذي تم تطويره في هذا العمل ناقلًا فعالًا لتوصيل البيومكرو جزيئات عبر الجلد، مما يوفر إمكانيات لمجموعة واسعة من التطبيقات مثل العلاج المناعي المحلي للميلانوما واللقاحات الموضعية الذاتية ضد الفيروسات (مثل SARS-CoV-2).
النتائج
تحضير مجمعات النانو FCS/البروتين وتقييم قدراتها عبر الجلد خارج الجسم
تم تخليق FCS وفقًا لتقرير سابقباختصار، تم ربط حمض الكربوكسيليك البيرفلوروألكيل (PFCA) بالبوليسكاريد الكاتيوني CS من خلال اقتران الأميد عند استبدال الفلوروكربون. (الشكل S1). ثم تم خلط FCS مع بروتينات مثل الغلوبولين المناعي G (IgG) والألبومين البيض (OVA) بنسب كتلية مختلفة لمدة 30 دقيقة تحت اهتزاز خفيف لتشكيل نانو معقدات (الشكل 1a). كما هو موضح في الشكل 1b و d، فإن كل من FCS/IgG و FCS/OVA بنسبة كتلية عند أظهرت الأحجام حوالي 200 نانومتر في صور المجهر الإلكتروني الناقل (TEM)، متوافقة مع أقطارها الهيدروديناميكية المقاسة بواسطة تشتت الضوء بالليزر الديناميكي (DLS) (الشكل 1c، e). أظهرت الجهود الزيتا (ZP) لكل من FCS/IgG وFCS/OVA شحنات إيجابية عالية، والتي زادت من 6.97 إلى 30.53 مللي فولت و4.38 إلى 36.73 مللي فولت، على التوالي، مع زيادة محتويات FCS أثناء تشكيل النانو مركبات (الشكل 1e). أظهرت قياسات تشتت الضوء الديناميكي (DLS) (الشكل S2) قمة واحدة عندلأجسام النانو المعقدة FCS/IgG، التي كانت أكبر بكثير من أحجام الحرة، مما يشير إلى أن الغالبية من كان ينبغي أن يتم احتواؤه بواسطة FCS. ثم، تم استخدام طيف الانكسار الدائري (CD) للتحقق من هيكل البروتينات قبل وبعد تشكيل النانو معقدات. كما هو موضح في الشكل 1f، أظهر FCS/IgG قمم طيف CD مشابهة عند حوالي 202 و206 و216 نانومتر لتلك الخاصة بـ IgG الحرة، مما يشير إلى أن هيكل البروتين ظل تقريبًا دون تغيير خلال تشكيل مثل هذه النانو معقدات المحتوية على FCS. قد تكون الفروق في الشدة عند 200 نانومتر ناتجة عن المذيب. تم العثور على نتيجة مماثلة أيضًا في المقارنة بين FCS/OVA وOVA الحرة في الشكل 1g. بالإضافة إلى ذلك، ظلت ألفة الأجسام المضادة لـ aPDL1 في تركيبة FCS/aPDL1 تقريبًا دون تغيير كما تم قياسها بواسطة اختبار المناعة المرتبط بالإنزيم (ELISA) (الشكل 1h)، مما يوضح أكثر أن تشكيل النانو معقدات لن يؤثر على نشاط البروتينات المحتواة.
ثم تم التحقيق في الديناميات عبر الجلد والآليات ذات الصلة. أولاً، تم استخدام نظام انتشار فرانز القياسي لقياس كفاءة توصيل FCS المحتوية على النانو معقدات عبر طبقة جلد الفأر. باختصار، تم تثبيت أنسجة الجلد الطازجة بين خليتين زجاجيتين، ثم تم إضافة FCS/IgG أو FCS/OVA، حيث تم وسم IgG وOVA بالفلووريسئين (FITC)، إلى غرفة المتبرع في محاليل ملح الفوسفات المخفف (PBS). تم قياس IgG أو OVA المنقولة من خلال جمع عينات سائلة في غرفة المستقبل في نقاط زمنية مختلفة لقياس فلوريسcence FITC (الشكل 1i). تم قياس جميع مقاومات الجلد قبل وبعد التجربة. اعتُبر الجلد غير مكسور من قبل الشركة المصنعة عندما كانت مقاومته ثلاثة أضعاف أكبر من المذيبات. لتجنب تدمير جلود الفئران، لم يتم إزالة طبقة الدهون في نموذج جلد الفأر. كما هو موضح في الشكل S3، لم تظهر جميع الجلود أي تغيير واضح في المقاومة خلال 12 ساعة. تم غسل الجلود وتحليلها لقياس الاحتفاظ عبر الجلد في منطقة الأدمة. كفاءات توصيل عبر الجلد لمركبات FCS/البروتين بنسب تغذية مختلفة (تم قياسها في نقاط زمنية مختلفة. كما هو موضح في الشكل 1i، k، أظهر FCS/IgG بنسبة كتلة 1:1 أعلى قدرة على الاختراق عبر الجلد مع إجمالي الكمية (مجموع النفاذ والاحتفاظ في الأدمة) يصل إلى حوالي (ستة أضعاف حجم المجموعة الحرة)، والذي قد يُعزى إلى حقيقة أن FCS/IgG المُعد على نسبة 1:1 أظهر أصغر الأحجام مقارنةً بتلك المُعدة بنسب كتلة أخرى. يمكن ملاحظة بوضوح أن الجهد الزتاوي لمركبات FCS/IgG النانوية زاد مع زيادة نسبة FCS، مما قد يكون مفيدًا لاختراق الجلد. ومع ذلك، قد يؤدي زيادة FCS بشكل أكبر إلى تكتل المركبات النانوية. من ناحية أخرى، عندما زادت كمية IgG، تبين أن الجهد الزتاوي لمركب FCS/IgG النانوي أصبح أقل إيجابية مع التكتل، وهو أيضًا غير مناسب لاختراق الجلد. وبالمثل، فإن FCS/OVA مع نسبة الكتلة عندأظهر أعلى قدرة على الاختراق بإجمالي اختراق (مجموع
أ
ب
ج
FCS/IgG
الحجم (نانومتر)
جهد الإمكانية (ملي فولت)
e
FCS/OVA
حجم
إمكانات
ف
ج
ح
أنا
ن
ساي 5.5-أوفا
FITC-FCS
مُدمَج
الشكل 1| توصيف المعقدات النانوية المحتوية على FCS. أ الصورة التخطيطية للمعقدات النانوية المحتوية على FCS للتوصيل عبر الجلد. ب صور TEM تمثيلية لـ FCS/IgG و د FCS/OVA. ج، هـ توزيع الحجم وإمكانات زيتا للمعقدات النانوية المحتوية على FCS بما في ذلك ج FCS/IgG و هـ FCS/OVA. ). طيف الانكسار الدائري (CD) لـإي جي جيطلاء OVA قبل وبعد FCS.الترابط النسبي لمستقبل aPDL1 مع أو بدون FCS تم قياسه بواسطة اختبار ELISA غير المباشر القياسي (iELISA) ). رسم تخطيطي لنظام خلية انتشار فرانز المستخدم في دراسة اختراق الجلد. تراكمي نفاذية واحتفاظ FCS/IgG-FITC وتغلغل FCS/OVA-FITC عبر جلد الفأر بعد الحضانة مع تركيبات مختلفة تحتوي على FCS على مدى الزمنلـ IgG ولـ OVA). الجرعة الكلية: . صور تمثيلية متداخلة لمسام جلد الفئران المعالجة بـ FITC-FCS/OVA-Cy5.5 لـمقياس الرسم:. تم إنشاء جميع الرسوم التوضيحية باستخدام بايو ريندر.كومتُعرض البيانات كمتوسطالانحراف المعياري. يتم توفير بيانات المصدر كملف بيانات المصدر. اختراق واحتباس الأدمة) حتى حوالي ( 11 مرة أكبر من المجموعة الحرة، الشكل 11، م). قد يكون الاختلاف في سلوك الاختراق بين المركبات النانوية المعتمدة على FCS ناتجًا عن الخصائص الفيزيائية والكيميائية المختلفة للبروتينات، مثل الوزن الجزيئي أو نقطة التساوي الكهربائي. بعد ذلك، تم تقطيع الأنسجة في نظام انتشار فرانز بعد التسليم عبر الجلد لعمل مجهرية متزامنة باستخدام FITC-FCS/OVA-Cy5.5 لتقييم اختراق المركبات النانوية FCS بشكل أكبر. كما هو موضح في الشكل 1ن، تم ملاحظة التداخل بين FCS/OVA في منطقة الأدمة من الجلد، مما يدل على أن FCS/OVA يمكن أن تخترق الأدمة الجلدية في شكل مركبات نانوية. بالمقابل، لم يتم ملاحظة اختراق واضح للجلد في مجموعة OVA الحرة. بشكل عام، يمكن أن تقوم المركبات النانوية المعتمدة على FCS بنجاح بتوصيل البروتينات إلى منطقة الأدمة في جلود الفئران.
آلية النقل عبر الجلد للمعقدات النانوية المحتوية على FCS
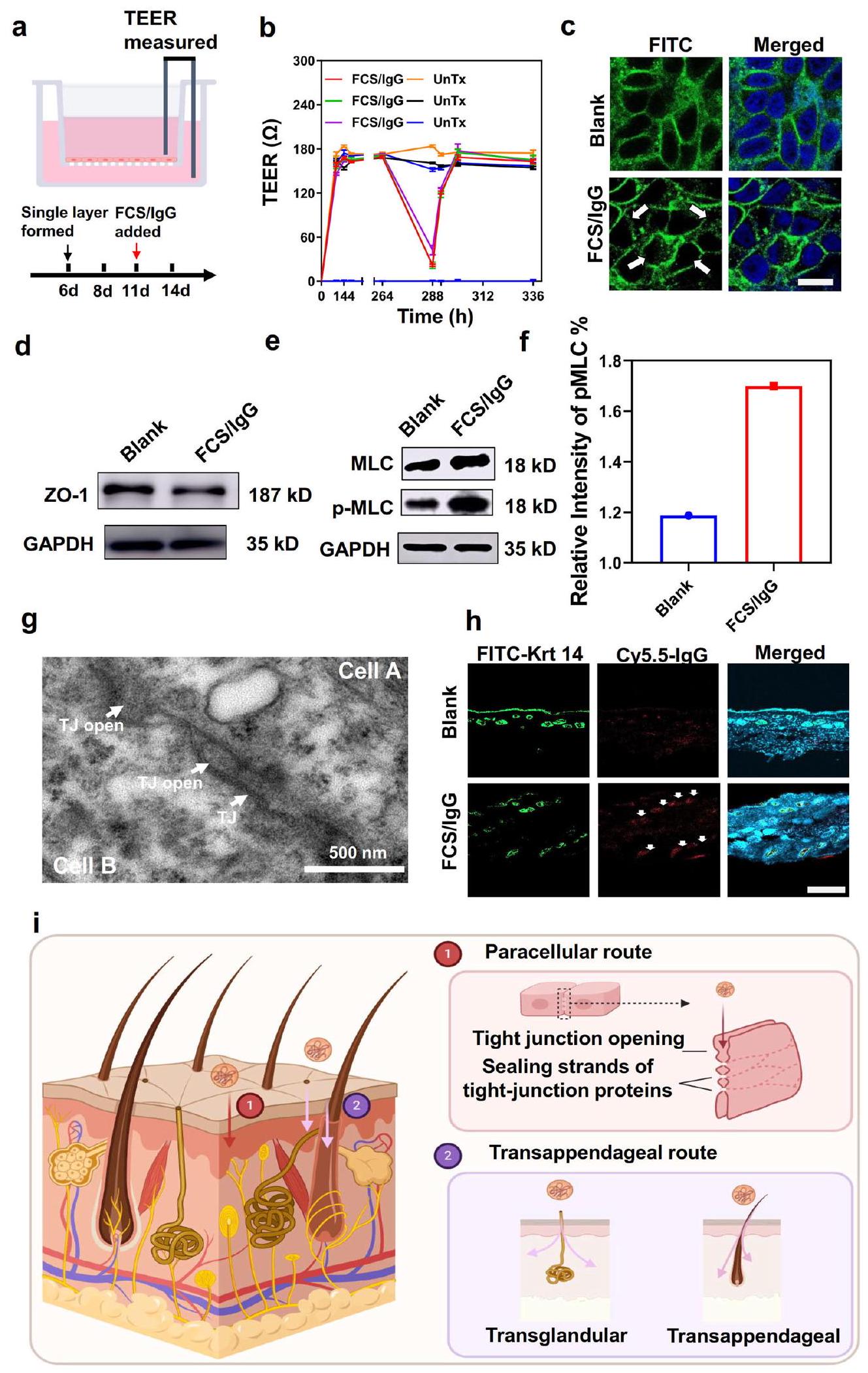
بعد ذلك، قمنا بالتحقيق في الآلية الأساسية عبر الجلد لمثل هذه النانو مركبات المحتوية على FCS. أولاً، تم قياس السمية الخلوية لمركبات FCS النانوية. كما هو موضح في الأشكال S4 و S5، لم يتم ملاحظة سمية خلوية واضحة لمركبات FCS و CS لخلايا HACAT. وفقًا للأدبيات السابقة، هناك ثلاث طرق تقليدية للاختراق لتوصيل عبر الجلد، بما في ذلك الطرق بين الخلوية، وعبر الملحقات، وعبر الخلايا.أولاً، درسنا المسار بين الخلايا، الذي يمكن أن يؤدي من خلاله النانو مركبات إلى توسيع المسافة بين خلايا البشرة وبالتالي المرور من خلالها.لقد تم الإبلاغ عن أن الكيتوزان ومشتقاته يمكن أن تعبر الخلايا الظهارية عن طريق الطريق بين الخلايا.خلال توسيع الفضاء بين الخلايا، نتوقع حدوث تغييرات في مقاومة الخلايا، بالإضافة إلى التعبير وتوزيع البروتينات ذات الصلة.في هذه الحالة، تم استخدام خلايا البشرة البشرية HACAT لتشكيل طبقة خلوية أحادية وتم مراقبة المقاومة الكهربائية عبر الظهارة (TEER) بين جانبي الطبقة الأحادية (الشكل 2أ). كما هو موضح في الشكل 2ب، تم تشكيل طبقة الخلايا الأحادية بعد 6 أيام مع TEER مستقر وعالي. ومن المثير للاهتمام، لوحظ انخفاض واضح في TEER مع إضافة FCS/IgG في اليوم 11، مما يدل على تدمير الطبقة الخلوية الأحادية وفتح القنوات بين الخلايا بعد إضافة المركبات النانوية المحتوية على FCS. والأكثر إثارة للاهتمام، لوحظ إعادة زيادة TEER بعد 4 ساعات من إزالة FCS/IgG، وعاد إلى مستواه الأصلي في 12 ساعة، مما يوضح أن FCS/IgG فتح فقط القناة بين الخلايا بشكل مؤقت. كما كشفت صور TEM للجلد عن فتح الوصلات الضيقة وتوسيع المساحات بين الخلايا بعد العلاج الموضعي للجلد بـ FCS/IgG، مقارنة بالجلد الطبيعي (الأشكال 2ج وS6). لتأكيد توسيع المساحات بين الخلايا، تم تقييم التغيرات في بروتينات الوصلات الضيقة (TJs) مثل زونولا أوكلودين (ZO)-1 بشكل إضافي.مع إضافة FCS/IgG، بينما ظل التعبير الكلي لـ ZO-1 تقريبًا دون تغيير، تم إزعاج توزيعها المستمر بشكل ملحوظ، مما يشير إلى فتح الوصلات الضيقة على طول الواجهة بين الخلايا (الشكل 2c، d). علاوة على ذلك، فإن فسفرة سلسلة الميوسين الخفيفة (MLC)، وهي معلمة مهمة لهيكل الهيكل الخلويتم العثور على أنه تم تنظيمه بشكل إيجابي في الخلايا المعالجة بـ FCS/IgG، مما يدل على أن FCS كان قادرًا على تعزيز فسفرة سلسلة الميوسين الخفيفة لتحفيز انقباض الأكتين وإعادة ترتيب الهيكل الخلوي (الشكل 2e، f).
بالإضافة إلى زيادة نفاذية التجاوز بين الخلايا للمعقدات النانوية المحتوية على FCS، تم أيضًا دراسة المسار عبر الأ appendages، الذي يلعب عادةً دورًا مهمًا في نقل الأدوية الكبيرة القابلة للذوبان في الماء من خلال بصيلات الشعر والغدد العرقية والغدد الدهنية، في تجاربنا.مع تلوين الكيراتين (Krt) 14، تم وسم بصيلات الشعر والغدد العرقية في منطقة الأدمة العميقة. كما هو موضح في الشكل 2g (الأسهم البيضاء)، لوحظ أن FCS/IgG تتواجد مع بصيلات الشعر والغدد العرقية، مما يشير إلى أن المسار عبر الملحقات الجلدية لعب أيضًا دورًا مهمًا في أنظمة توصيل الأدوية عبر الجلد المعتمدة على FCS.
قمنا بدراسة المسار العابر للخلايا بشكل أعمق، والذي يعني مرور الأدوية مباشرة عبر الخلايا الكيراتينية.مختلف عن الطبيعي خلايا الأنسجة، التي تقوم بتحليل الأدوية في الجسيمات الحالة، قد تقوم خلايا قطبية مثل الكيراتينوسيت أحيانًا بطرد الأدوية من خلال الإخراج الخلوي.تم استخدام الإفراز القمي لدراسة هذه الظاهرة في المختبر. باختصار، تم تحضين خلايا HACAT مع FCS/IgG-FITC لعملية الابتلاع الخلوي. بعد 12 ساعة من التحضين، تم غسل FCS/IgG-FITC، وتم تحضين الخلايا لمدة 12 ساعة أخرى. ثم تم قياس الفلورية لـ FITC في السائل الفائق لتقييم الإفراز القمي. كما هو موضح في الشكل S7، كانت نسبة الإفراز القمي لـ FCS/IgG-FITC من خلايا HACAT أقل من، تقريبًا نفس الشيء مقارنةً بتلك المعالجة بـ IgG-FITC الحر، مما يشير إلى أن الطريق الخلوي العابر كان ضئيلاً. بالإضافة إلى ذلك، تم إجراء تقييم إضافي بواسطة نظام انتشار فرانز. كما تم الإبلاغ، فإن الإخراج الخلوي عادة ما يتم بواسطة الكلاثرين. لذلك، تم معالجة الجلد بـكلوربرومازين هيدروكلوريد، المثبط للكلاثرين، لمدة ساعتين لمقارنة قدرة الاختراق مع أو بدون مثبط الكلاثرين. كما هو موضح في الشكل S8، لم يظهر احتفاظ الجلد بـ FCS/IgG-FITC أي تأثير واضح. في الختام، تعتمد عملية توصيل FCS المحتوية على النانو معقدات عبر الجلد بشكل رئيسي على المسارات بين الخلايا وعبر الملحقات (الشكل 2i).
ختامًا، يمكن أن يزيد FCS، كمشتق من الكيتوزان، من المساحة الخلوية من خلال تحفيز فوسفاتة MLC، وهي ظاهرة تم ملاحظتها أيضًا للكيتوزان غير المعدل.يمكن أن تؤدي فسفرة MLC بعد ذلك إلى إعادة ترتيب هيكل السيتوسكلت، مما يحول البروتينات المرتبطة بالوصلات الضيقة إلى السيتوبلازم. بعد ذلك، سيتم فتح الوصلة الضيقة بين الخلايا الظهارية، وسيتم توسيع الفضاء بين الخلايا للسماح بمرور مركباتنا النانوية. من ناحية أخرى، ستجعل الخصائص الفريدة لسلاسل الفلوروكربون غير الكارهة للماء وغير المحبة للماء FCS أقل لزوجة عند اختراقها عبر الحواجز البيولوجية المختلفة.لذلك، أظهرت المعقدات النانوية المعتمدة على FCS أيضًا زيادة في النفاذية عبر بصيلات الشعر من خلال المسار عبر الملحقات.
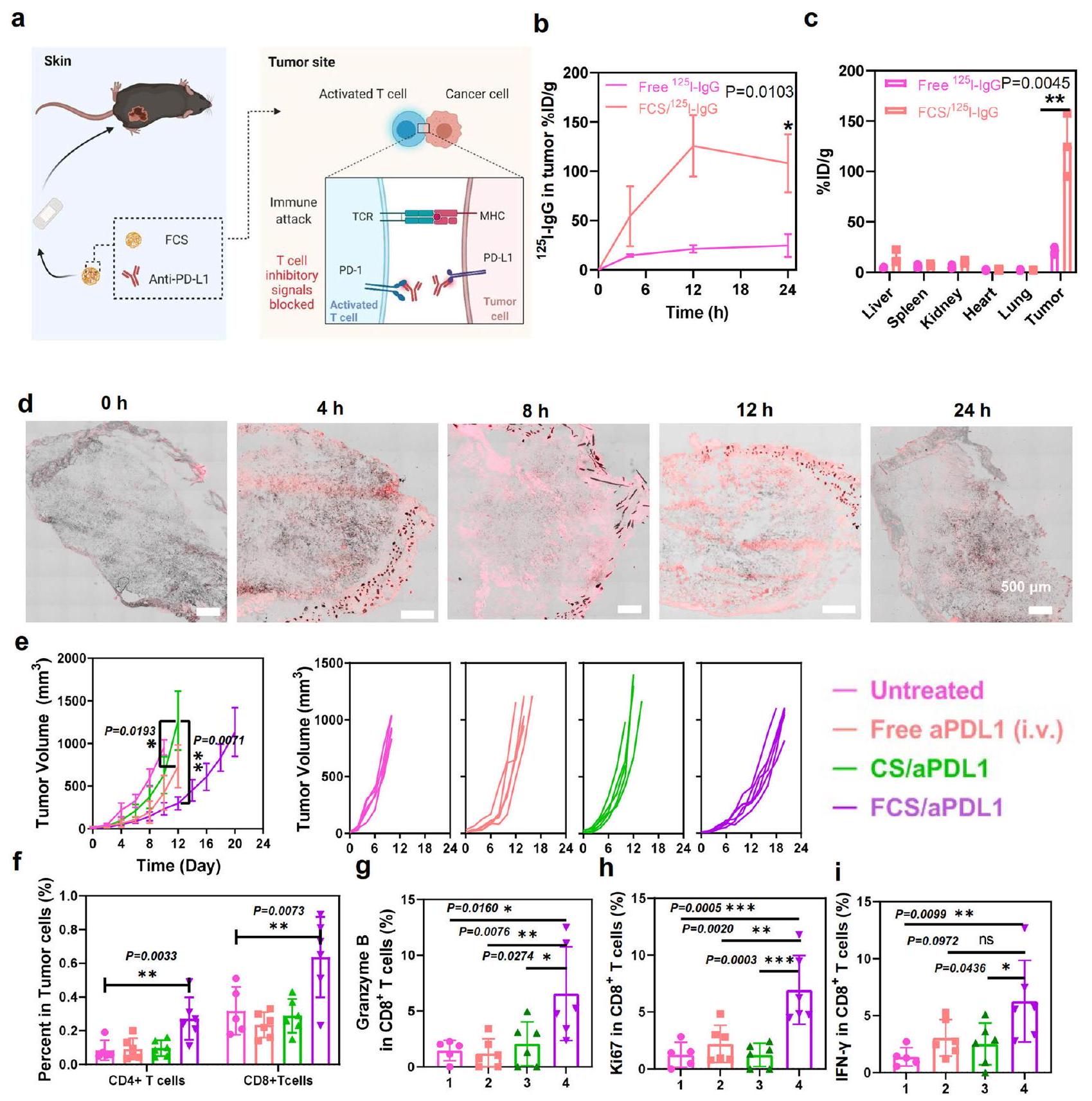
يتطور الميلانوما، كواحد من أكثر الأورام الخبيثة شيوعًا، خاصة بين القوقازيين، في الخلايا الصبغية الموجودة في الطبقة السفلية من البشرة.في علاج الميلانوما، تم استخدام العلاجات المناعية، وخاصة حجب نقاط التفتيش المناعية باستخدام الأجسام المضادة لمستقبل الموت المبرمج-1/رابطةه (aPD1/aPDL1)، على نطاق واسع في العيادات.على الرغم من النتيجة العلاجية المثيرة لاستخدام أجسام مضادة aPD1/aPDL1 لعلاج الميلانوما، لا تزال هناك العديد من القيود، مثل خطر الأمراض المناعية الذاتية بعد الحقن الوريدي.نظرًا لأن FCS يمكن أن يعمل كحامل فعال للتوصيل عبر الجلد للبروتينات، استخدمناه كمنصة للتوصيل عبر الجلد لتوصيل جسم مضاد aPDL1 لعلاج الميلانوما (الشكل 3أ). كان من المتوقع أن الجسم المضاد aPDL1 المنقول يمكن أن يمنع مسار PD1/PDL1 لتحفيز خلايا T السامة وتؤدي إلى تثبيط ملحوظ للأورام.
لإدارة في الجسم الحي لمركبات النانو المعتمدة على FCS الخاصة بنا، مرهم فارغ (أكوافورتم خلط مادة (بشكل رئيسي الفازلين) مع المحلول للحفاظ على الرطوبة لفترة طويلة. ستتحول المراهم الشفافة إلى مراهم تشبه الحليب عند خلطها مع المحاليل. يمكن امتصاص المراهم التي تشبه الحليب خلال 12 ساعة بعد الإعطاء الموضعي لتحقيق توصيل كامل عبر الجلد. لذلك، تم تطبيق الإعطاء لمدة 12 ساعة فقط للعلاج في الجسم الحي للجرعة المطبقة القياسية. لفحص سلوك اختراق الأجسام المضادة المعقدة مع FCS، تم مسح الأورام من الفئران التي تم دهنها بالمراهم المحتوية على نظير مشع.الموسوم IgG ) أو FCS/ تم جمعها لت quantification الأجسام المضادة المنقولة في نقاط زمنية مختلفة (الشكل S9). كما هو موضح في الشكل 3b، مقارنةً بـ freeالأجسام المضادة IgG في المرهم المطبق عبر الجلد،أظهر تراكمًا أعلى بشكل ملحوظ في الورم، بينما بدت النشاطات الإشعاعية في الأعضاء الأخرى أقل بكثير. في الوقت نفسه، كما هو موضح في الشكل 3c، وجدنا أن تراكم FCS/IgG في الورم بلغ ذروته عند أكثر منجرعة الحقن لكل جرام من الأنسجة (%ID/g) بعد 12 ساعة تم تطبيق المرهم (حواليتم تحديد ID في الورم الكلي في الشكل S 10). عند إزالة المرهم بعد 12 ساعة، انخفض مستوى IgG في الورم قليلاً بعد 24 ساعة. في الوقت نفسه، تم إجراء اختبار ELISA أيضًا للكشف عن تراكم IgG في الورم في تركيبة FCS/IgG المطبقة موضعياً على الأورام في المرهم. كما هو موضح في الشكل S11، أظهر اختبار ELISA نتائج مشابهة لبيانات توزيع الإشعاع المعتمدة على التتبع، حيث يمكن توصيل IgG بكفاءة إلى الورم خلال 12 ساعة بمساعدة منصة توصيل عبر الجلد باستخدام مجمعات FCS/IgG النانوية. من ناحية أخرى، تم تضمين الأورام مع FCS/IgG-Cy5.5 لقطع الورم. كما هو موضح في الشكل 3d، كانت الإشارات الفلورية لـ IgG-Cy5.5 في الورم تزداد تدريجياً وتوزع بشكل متساوٍ داخل الورم بأكمله خلال 12 ساعة، مما يدل على النقل المستمر للأجسام المضادة من مرهم FCS/IgG-Cy5.5 إلى الورم. لذلك، فإن التراكم
الشكل 2 | الآلية عبر الجلد للمعقدات النانوية المحتوية على FCS. أ رسم توضيحي لنموذج خلايا HACAT أحادية الطبقة. تأثيرات FCS/IgG على TEER لنموذج خلايا HACAT أحادية الطبقةتم اختبار كل TEER ثلاث مرات). ج صور المناعة الفلورية لتوزيع بروتين ZO-1 المرتبط بالوصلات الضيقة على غشاء خلايا HACAT بعد معالجتها بـ FCS/IgG ( ). تشير الأسهم البيضاء إلى تغيير تخصيص ZO-1. مقياس الرسم: صور التحليل الغربي تظهر ZO-1 ) ومستوى الفسفرة لـ MLC ( MLC ) في الخلايا بعد الحضانة مع FCS/IgG. تم تقديم الأرقام الخام في الأشكال S27 و S28. f التمثيلات البيانية لشدة MLC/ pMLC النسبية مع إضافة FCS/IgG ( صورة TEM تمثيلية لظهارة الجلد بعد معالجتها بـ FCS/IgG. الأسهم البيضاء تشير إلى الروابط الضيقة التقاطعات (TJs) وفتح التقاطعات (TJs) ). صور المناعة الفلورية التمثيلية التي تظهر التوضع المشترك للكيراتين 14 و FCS/IgG-Cy5.5 (الأسهم البيضاء، ). مقياس الرسم: الصورة التخطيطية لآليات النقل عبر الجلد. يمكن أن تخترق النانومركبات المحتوية على FCS البشرة من خلال كل من الطرق بين الخلايا والطريق عبر الملحقات. من خلال الطريق بين الخلايا، يمكن أن يحفز FCS فسفرة MLC وبالتالي يفتح الوصلات الضيقة بين خلايا البشرة عن طريق إغلاق خيوط بروتينات الوصلات الضيقة. من خلال الطريق عبر الملحقات، يمكن أن تعبر النانومركبات المحتوية على FCS البشرة من خلال بصيلات الشعر والغدد العرقية. تم إنشاء جميع الرسوم التوضيحية باستخدامبايو ريندر.كومتُعرض البيانات كمتوسطالانحراف المعياري. يتم توفير بيانات المصدر كملف بيانات المصدر. تم تقييم تأثير أدوية الأجسام المضادة مثل aPDL1 في الورم. كما هو موضح في الشكل S12، أظهر FCS/aPDL1 تراكمًا أعلى بكثير في الورم بعد التطبيق الموضعي مقارنةً بـ free و CS/aPDL1. تم أيضًا تقييم تهيج الجلد للإدارة طويلة الأمد. كما هو موضح في الشكل S13، أظهرت كل من مجموعة الفيلم ومجموعة FCS/IgG مع مجموعة الفيلم ظروف جلدية مشابهة بعد العلاج الموضعي ثلاث مرات (مرة كل يومين)، مما يشير إلى الاستخدام الموضعي الآمن لـ FCS.
استنادًا إلى التراكم الفعال لـ IgG في الورم بعد الإيصال عبر الجلد، قمنا بعد ذلك بإجراء علاج في الجسم الحي لأورام الميلانوما من خلال الإيصال عبر الجلد لـ FCS/aPDL1. تم تقسيم الفئران الحاملة لأورام الميلانوما عشوائيًا إلى أربع مجموعات: (i) غير معالجة، (ii) aPDL1 الحر عن طريق الحقن الوريدي (i.v.)، (iii) CS/PDL1 في المرهم عن طريق الإيصال عبر الجلد، و(iv) FCS/aPDL1 في المرهم عن طريق الإيصال عبر الجلد. بالنسبة للإيصال عبر الجلد، تم خلط محلول CS/aPDL1 أو FCS/aPDL1 مع مرهم فارغ ثم تم تطبيقه على الورم، الذي تم تغطيته بعد ذلك بـفيلم شفاف. تم تكرار هذا العلاج كل يومين ثلاث مرات بجرعة aPDL1 لكل فأر في كل مرة. بالنسبة للحقن الوريدي، تم إعطاء aPDL1 لكل فأر كل يومين ثلاث مرات. كما هو موضح في الشكل 3e، تم تثبيط الأورام في مجموعة المعالجة بـ FCS/aPDL1 بنجاح في وقت قصير بعد العلاج الثالث، بينما أظهرت كل من مجموعة CS/aPDL1 ومجموعة aPDL1 الحرة فعالية مضادة للأورام ضئيلة. خلال 20 يومًا من المراقبة، أظهرت مجموعة المعالجة بـ FCS/aPDL1 أدنى معدل لنمو الأورام وأطول فترة بقاء (الشكل S14).
لفهم آليات تنشيط المناعة للعلاج المناعي الذي يتم توصيله عبر الجلد، تم جمع الأورام من مجموعات مختلفة في اليوم الثاني عشر للتحقيق في أنواع مختلفة من خلايا المناعة، وخاصة خلايا T، باستخدام تحليل تدفق الخلايا. كما هو موضح في الشكل 3f، بالنسبة لمجموعة المعالجة بـ FCS/aPDL1، كانت النسب لكل منوأظهرت خلايا T زيادة واضحة في الورم، مما يشير إلى أن aPDL1 تم توصيله بنجاح إلى الورم لإعادة الأمور إلى نصابها.استنفاد الخلايا. حيث أن إنزيم غرانزيم ب مهم لبرمجة موت الخلايا التي يتم تحفيزها بواسطةالخلايا، Ki 67 هو تكاثر الخلايا، والإنترفيرونهو سيتوكين مرتبط بآليات مؤيدة للموت الخلوي وآليات مضادة للورم، استخدمنا هذه العلامات الثلاثة لتحليل أنشطة خلايا CD8+. كما يتضح في الشكل 3g-i، جميع العلامات الثلاثة فيتم زيادة الخلايا في مجموعة FCS/aPDL1، مما يشير إلى التسلل الفعال وتنشيط الخلايا اللمفاوية التائية السامة للخلايا (CTLs) في تلك الأورام. بالنسبة لحقن aPDL1 عن طريق الوريد وتوصيل CS/aPDL1 عبر الجلد، كانت تسللات الورم لكلا المجموعتينوالخلايا، بالإضافة إلى تعبيرات الجرانيزيم، و ظل تقريبًا دون تغيير، وهو ما قد يكون ناتجًا عن انخفاض تراكم الورم لـ aPDL1 بعد الحقن الوريدي أو التسليم عبر الجلد باستخدام CS. تشير هذه النتائج بوضوح إلى أنه، مقارنةً بالحقن الوريدي لـ aPDL1، أدى التسليم عبر الجلد لـ aPDL1 باستخدام FCS إلى تعزيز الاستجابات المناعية المضادة للورم.
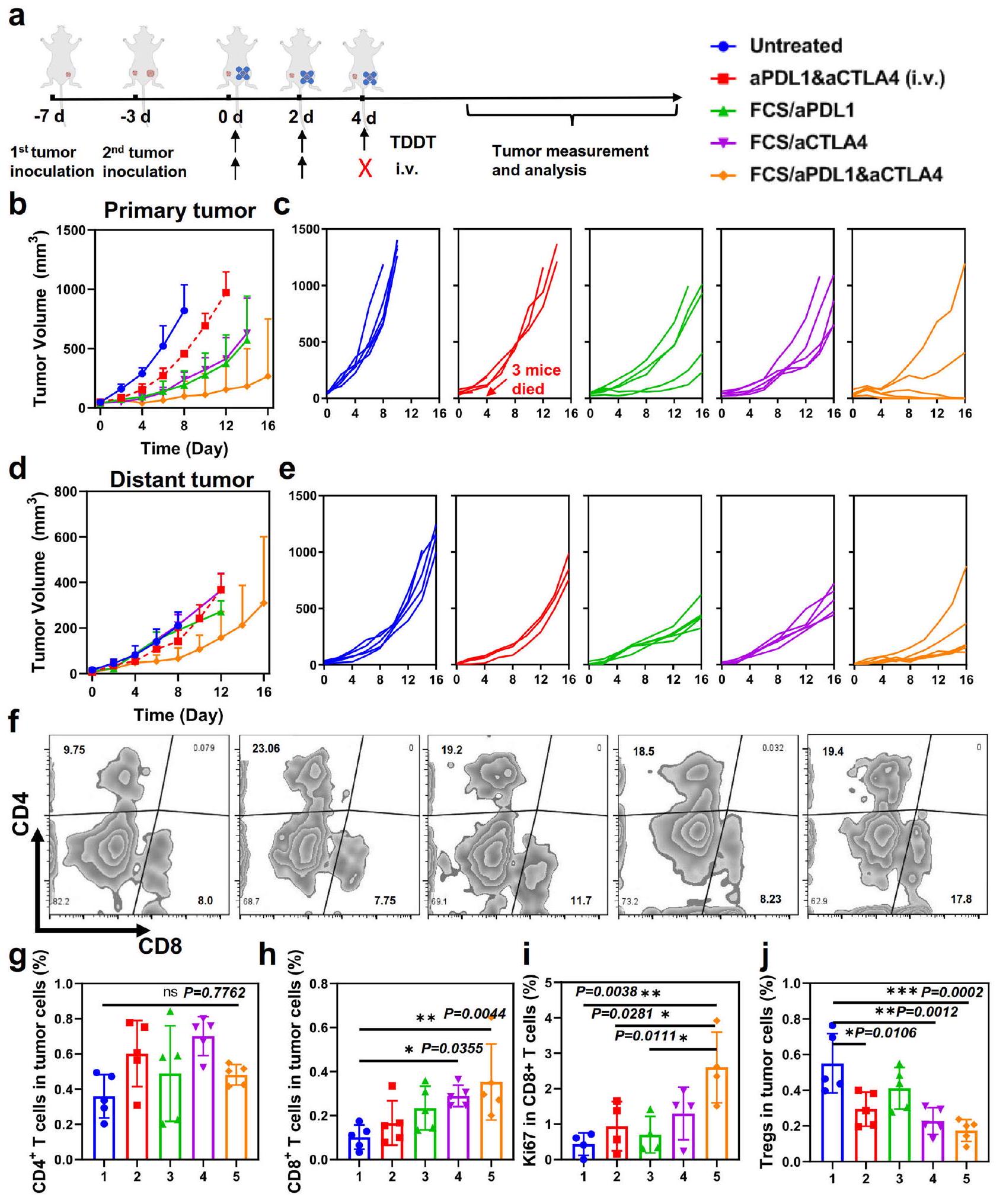
يمكن أيضًا استخدام منصتنا لتوصيل بروتينات علاجية مختلفة بشكل مشترك. بالإضافة إلى aPDL1، فإن الأجسام المضادة المضادة لبروتين 4 المرتبط بالخلايا التائية السامة (aCTLA4) هي جسم مضاد مهم آخر لحجب نقاط التفتيش المناعية لتعطيل الخلايا التائية التنظيمية (Tregs) وتعزيز تنشيط الخلايا التائية الفعال.. وبالتالي، قمنا بمزيد من التحقيق في العلاج المشترك باستخدام aPDL1 و aCTLA4 المقدمين معًا بواسطة حامل توصيل عبر الجلد FCS (الشكل 4أ). في هذه التجربة، تم تلقيح خلايا سرطان B16F10 على الجانب الأيمن من كل فأر كسرطان أولي، وتم تلقيح ورم ثانٍ على الجانب المقابل من نفس الفأر لمحاكاة انتشار السرطان. بعد ثلاثة أيام، تم تقسيم الفئران التي تحمل ورمين من الميلانوما عشوائيًا إلى خمس مجموعات: (1) غير معالجة، (2) aPDL1 و aCTLA4 الحرة (عن طريق الوريد)، (3) FCS/ aPDL1، (4) FCS/aCTLA4، (5) FCS/aPDL1/aCTLA4. تم إعطاء جميع المجموعات ثلاث مرات بـ aPDL1 و aCTLA4 لكل فأر في كل مرة. من الجدير بالذكر أنه بعد الحقن الثاني عن طريق الوريد لـ aPDL1 + aCTLA4، توفي نصف الفئران في هذه المجموعة، على الأرجح بسبب الآثار الجانبية الرهيبة (مثل عاصفة السيتوكين) التي triggered by الإدارة النظامية لكل من aPDL1 و aCTLA4. بالمقابل، أظهرت المجموعات الأخرى التي تلقت الأجسام المضادة لحجب نقاط التفتيش المناعية عن طريق الجلد عدم وجود شذوذ ملحوظ. ومن المثير للاهتمام، مقارنة بالفئران المعالجة بـ FCS/aPDL1 أو FCS/aCTLA4، أظهرت الفئران التي تلقت FCS/aPDL1/aCTLA4 عن طريق الجلد أداءً علاجيًا محسّنًا بشكل أكبر (الشكل 4ب، ج). والأكثر إثارة للاهتمام، أن نمو الأورام البعيدة في الفئران التي تلقت علاج FCS/aPDL1/aCTLA4 تم تثبيته أيضًا (الشكل 4د، هـ). لفهم التأثير البعيد الذي أحدثه FCS/aPDL1/aCTLA4، تم جمع الأورام بعد 12 يومًا من العلاجات المختلفة وتحليلها بواسطة قياس التدفق الخلوي. كما هو موضح في الشكل S19، في الورم الأولي، لم يظهر نسبة خلايا CD4+ T أي زيادة واضحة، بينما كانت نسبة الخلايا التنظيميةتم تقليل الخلايا في الورم الأساسي، والذي قد يكون نتيجة لتوصيل aCTLA4 عبر الجلد بنجاح. من ناحية أخرى، كانت النسب لكل منالخلايا وتم زيادة الخلايا، مما يوضح زيادة تسلل الورم للخلايا السامة للخلاياالخلايا اللمفاوية (CTLs) (الشكل S20). متسقة مع النتائج المذكورة أعلاه، في الورم البعيد، عددالخلايا، خاصةًالخلايا، أظهرت زيادة واضحة في الورم الثاني (الشكل 4f، i)، وانخفضت نسب خلايا Tregs في مجموعة المعالجة بـ FCS/aPDL1/aCTLA4 (الشكل 4j).
يمكن أن يُعزى الفعالية الممتازة ضد الأورام وزيادة الخلايا التائية السامة للخلايا (CTLs) في الورم البعيد إلى الآليات التالية. أولاً، يمكن أن يؤدي تنشيط الخلايا التائية السامة للخلايا في الورم المحلي إلى تحفيز موت مناعي لخلايا الورم وإثارة التعرض المزمن لأنماط الجزيئات المرتبطة بالضرر (DAMPs) بالإضافة إلى مستضدات الورم. ثم يتم تنشيط خلايا تقديم المستضدات (APCs) وتقديم مستضدات الورم.الخلايا، مما يعزز المناعة المضادة للأورام النظامية لمهاجمة الأورام البعيدة. وأخيرًا، كما تم الإبلاغ، فإن حجب PDL1 في العقد اللمفاوية التي تصرف الأورام (TDLNs)، والتي قد يكون من الأكثر كفاءة الوصول إليها عن طريق التسليم عبر الجلد، يمكن أن يدفع بشكل فعال المناعة المضادة للأورام النظامية.مناعة الخلايا حتى في مواقع الأورام البعيدةلذلك، يمكن أن تحقق طريقة توصيل الأجسام المضادة المثبطة للمناعة عبر الجلد فعالية ملحوظة ضد الأورام المحلية والبعيدة.
التطبيق الموضعي لمركبات FCS/S1/polyIC للتطعيم عبر الجلد
التطعيم هو مجال آخر ذو اهتمام كبير لتوصيل الدواء عبر الجلد. بالنسبة للقاح عبر الجلد، بالإضافة إلى تجنب الحقن بواسطة المتخصصين في الرعاية الصحية، يمكن أن يحسن الاستجابات المناعية من خلال استهداف خلايا المناعة الوفيرة تحت طبقة البشرة. منذ تفشي مرض فيروس كورونا 2019 (كوفيد-19)، تم تطوير لقاحات متنوعة
الشكل 3 | الإيصال عبر الجلد لـ aPDL1 لعلاج أورام الميلانوما B16F10. أ رسم تخطيطي يوضح الإدارة الموضعية عبر الجلد لـ FCS/aPDL1 لعلاج أورام الميلانوما B16F10.تراكم FCS/ I-IgG في الورم في فترات زمنية مختلفة ( ). ج توزيع FCS في الجسم I-IgG بعد 12 ساعة بناءً على قياس النشاط الإشعاعي. تم توضيح التحليل الكلي للتراكم والتوزيع الحيوي في الشكل S10 (صور تمثيلية متداخلة تظهر تراكم FCS/IgG-Cy5.5 في الورم في فترات زمنية مختلفة ). مقياس الرسم: . منحنيات نمو الورم للفئران في مجموعات مختلفة (تم إيقاف منحنيات النمو عندما توفيت الفأر الأول في المجموعة المعنية، أو عندما تجاوز حجم ورمها. قياس
الخلايا والخلايا في الورم بعد علاجات مختلفة ). تم توضيح الرسوم البيانية التمثيلية لتدفق الخلايا في الشكل S15. ج-ي قياس الجرانزيم جرانزيم، و IFN- ( إنترفيرونخلايا T في الورم بعد علاجات مختلفة ). تم توضيح الرسوم البيانية التمثيلية لتدفق الخلايا في الأشكال S16-S18. تم إنشاء جميع الرسوم التوضيحية باستخدام بايو ريندر.كومتُعرض البيانات كمتوسطالانحراف المعياري. تم حساب الدلالة الإحصائية من خلال تحليل التباين الأحادي مع اختبار توكي بعد الاختبار.. يتم توفير بيانات المصدر كملف بيانات المصدر.
تم تطويره لقمع عدوى SARS-CoV-2 . حتى الآن، هناك 242 مرشحًا للقاح SARS-CoV-2 في التطوير السريري، وقد تم منح 11 لقاحًا لـ COVID-19 قائمة الاستخدام الطارئ (EUL) من قبل منظمة الصحة العالمية . ومع ذلك، فإن معظم هذه اللقاحات تحتاج إلى حقن تحت الجلد أو في العضل، مما يتطلب ليس فقط طاقم طبي مدرب جيدًا ولكن أيضًا يواجه بعض القضايا الإضافية، مثل التخلص من عدد كبير من الحقن المعقمة. من ناحية أخرى، قد يقدم لقاح SARS-CoV-2 عبر الجلد مساعدة كبيرة في
منع COVID-19 من خلال الإدارة الذاتية، خاصة في المناطق التي تعاني من نقص في الموارد الطبية. نظرًا للانتشار السريع لـ COVID-19 على مستوى العالم والتوصيل الفعال للأجسام المضادة عبر الجلد باستخدام FCS، استكشفنا المزيد ما إذا كان يمكن استخدام FCS لتوصيل لقاح SARS-CoV-2 عبر الجلد في دراسة إثبات المفهوم.
لتركيب لقاح SARS-CoV-2 القائم على FCS، تم خلط FCS مع الوحدة الفرعية S1 من بروتين السنبلة لفيروس SARS-CoV-2 وpolyIC، مما شكل لقاح SARS-CoV-2 عبر الجلد (FCS/S1/polyIC)
الشكل 4 | التأثير الغير مباشر الناتج عن توصيل الأجسام المضادة المشتركة عبر الجلد. أ رسم توضيحي تخطيطي لتوصيل aPDL1 وaCTLA4 عبر الجلد لتثبيط نمو كل من الأورام الأولية والبعيدة. ب-هـ منحنيات نمو الورم للأورام الأولية والبعيدة بعد علاجات مختلفة ( ). تم إيقاف منحنيات النمو عندما ماتت الفأرة الأولى في المجموعة المعنية، أو عندما تجاوز حجم ورم الفأرة الأولى . مخططات تدفق تمثيلية (ف) والتقدير المتعلق لـ
خلايا T وخلايا T في الأورام البعيدة بعد علاجات مختلفة ( ). تقدير لـ خلايا (i) وTregs (CD3 Foxp3 ) (j) في الأورام البعيدة بعد علاجات مختلفة ( ). تم توضيح مخططات تدفق تمثيلية في الشكل S21. تم تقديم البيانات كمتوسط انحراف معياري. تم حساب الدلالة الإحصائية عبر ANOVA أحادي الاتجاه مع اختبار Tukey بعد ذلك. . يتم توفير بيانات المصدر كملف بيانات المصدر.
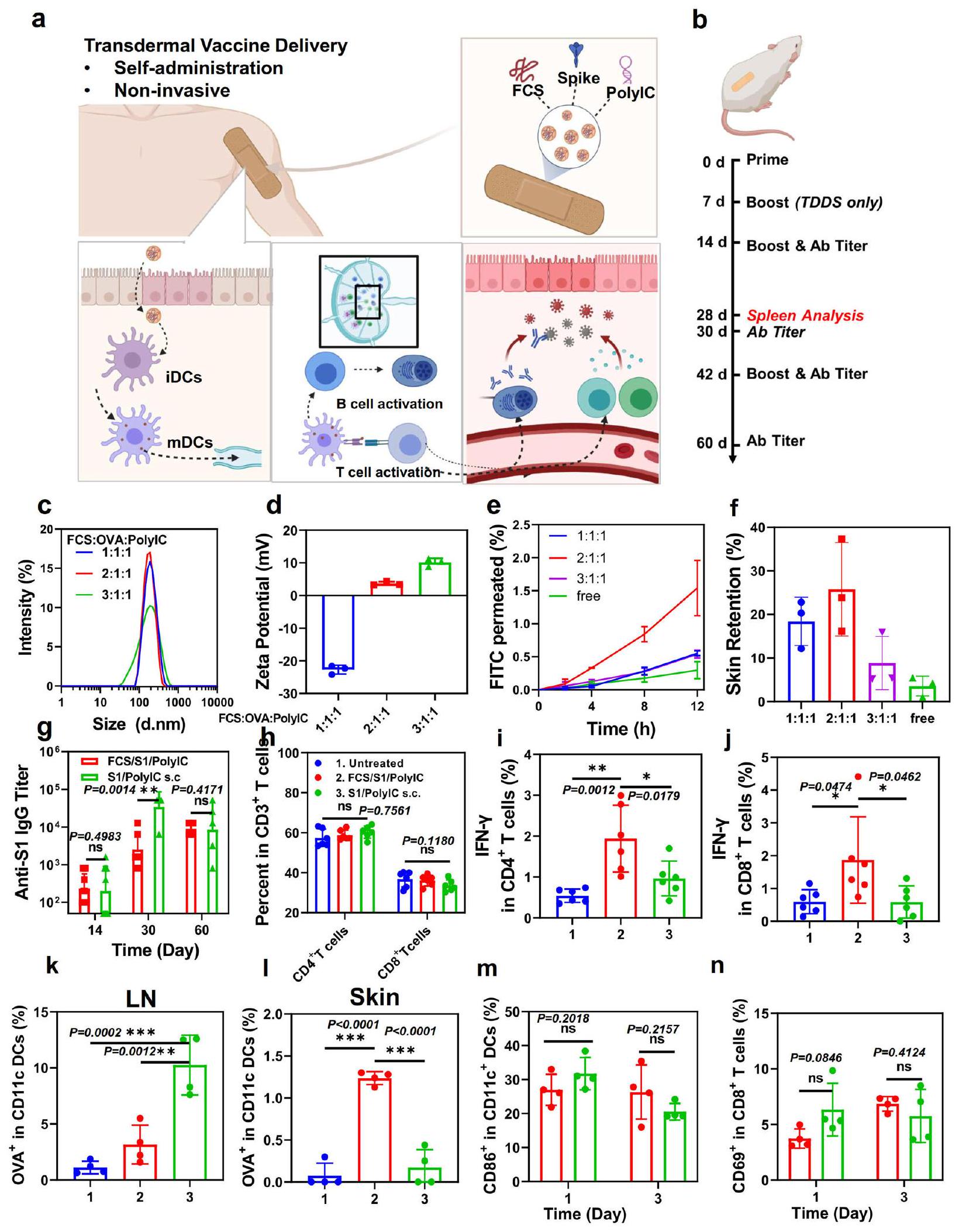
(الشكل 5a). لاحظ أن poly IC كـ RNA مزدوج الشريطة هو منبه لمستقبلات Toll-like 3 (TLR 3) وقد تم استخدامه بشكل شائع كمعزز مناعي . كما قمنا بتحسين نسبة FCS: المستضد: PolyIC في لقاح FCS/S1/ PolyIC. في هذه التجربة، تم استخدام OVA كمستضد معدل لتحسين التركيبة. كما هو موضح في الشكل 5c، d، أظهر FCS/OVA/polyIC بنسب كتلة مختلفة أحجامًا متغيرة بحوالي 200 نانومتر وزيادة في الجهد الزتاوي مع زيادة FCS.
بالنسبة لقدرة اختراق الجلد التي تم قياسها بواسطة نظام انتشار فرانز في الشكل 5e، f، أظهر FCS/OVA/PolyIC المعد في نسبة الكتلة 2:1:1 أعلى نفاذية للجلد. قد يكون ذلك نتيجة للحجم المناسب والجهد الزتاوي حيث أظهرت الجسيمات النانوية المعدة بنسبة 1:1:1 شحنات سالبة بشكل كبير، بينما أظهرت تلك المعدة بنسبة 3:1:1 أحجامًا أكبر بكثير. لذلك، تم استخدام التركيبة بنسبة FCS/OVA/PolyIC عند 2:1:1 للدراسات الإضافية.
الشكل 5 | توصيل لقاح SARS-CoV-2 عبر الجلد. أ رسم توضيحي تخطيطي لتوصيل لقاح SARS-CoV-2 عبر الجلد والاستجابات المناعية المحفزة. بعد التوصيل عبر الجلد، يمكن أن تنشط لقاحات SARS-CoV-2 النانوية مثل هذه خلايا المناعة مثل DCs في الأدمة، أو تهاجر إلى العقد اللمفاوية القريبة للتفعيل المناعي. ب رسم توضيحي تخطيطي لتصميم التجربة يظهر توصيل لقاح SARS-CoV-2 عبر الجلد. ج، د DLS (ج) والجهد الزتاوي (د) للقاحات عبر الجلد القائمة على FCS بنسب كتلة مختلفة من 1:1:1 إلى 3:1:1 ( ). هـ، قدرة اختراق الجلد للقاح عبر الجلد القائم على FCS بنسب كتلة مختلفة ( ). الجرعة الكلية: (غ) عيار الأجسام المضادة IgG المحددة لـ SARS-CoV-2 في فترات زمنية مختلفة تم تحديدها بواسطة ELISA ( ). ح تقدير لـ خلايا، خلايا في الطحال في اليوم تقدير لـ IFN- خلايا تفرز (IFN ) و خلايا (IFN ) في الطحال في اليوم تقدير لـ OVA-Cy5.5 CD45 CD11c Cy5.5 في DCs في العقد اللمفاوية و الجلد ( ). تقدير لـ نضوج DC () و تنشيط مستقبل الخلايا (TCR) () في العقد اللمفاوية ( ). تم إنشاء جميع الرسوم التوضيحية باستخدام BioRender.com. تم توضيح مخططات تدفق تمثيلية في الأشكال S22 وS23. تم تقديم البيانات كمتوسط انحراف معياري. تم حساب الدلالة الإحصائية عبر ANOVA أحادي الاتجاه مع اختبار Tukey بعد ذلك. ; . يتم توفير بيانات المصدر كملف بيانات المصدر.
بالنسبة لتجارب التطعيم في الجسم الحي، تم تقسيم الفئران عشوائيًا إلى ثلاث مجموعات: 1. غير معالجة، 2. توصيل عبر الجلد لـ FCS/ S1/polyIC، و3. حقن تحت الجلد (s.c.) لـ S1/polyIC. كما هو موضح في الشكل 5b، تم إعطاء الفئران في مجموعة التوصيل عبر الجلد 3 مرات في 2 أسابيع (كانت جرعات S1 وpolyIC كلاهما في كل مرة)، بينما تم حقن الفئران في مجموعة الحقن تحت الجلد مرتين بجرعة بروتين S1 عند في كل مرة وجرعة polylC عند في كل مرة. من المثير للاهتمام، أن الفئران التي تلقت توصيلًا عبر الجلد لـ FCS/S1/polyIC أظهرت تقريبًا نفس عيار الأجسام المضادة مثل تلك الخاصة بالفئران التي تلقت حقنًا تحت الجلد لـ S1/ polyIC في غضون أسبوعين، مما يشير إلى أن التوصيل عبر الجلد لـ FCS/ S1/polylC يمكن أن يؤدي إلى مناعة خلوية مماثلة تقريبًا مقارنة باللقاحات المعطاة تحت الجلد (الشكل 5g). علاوة على ذلك، بعد التعزيز في اليوم 14، وصلت عيارات الأجسام المضادة المحددة في كل من مجموعة التوصيل عبر الجلد ومجموعة الحقن تحت الجلد إلى في 30 يومًا، مما يدل على التفعيل الفعال للمناعة الخلوية بواسطة تلك اللقاحات. بعد تعزيز آخر في اليوم 42، تم تطعيم الفئران إما عن طريق التوصيل عبر الجلد لـ FCS/S1/polyIC أو الحقن تحت الجلد لـ S1/ polyIC وظلت عند مستويات عالية من عيارات الأجسام المضادة المضادة لـ S1.
بالإضافة إلى تفعيل المناعة الخلوية من خلال إنتاج أجسام مضادة محددة ضد بروتين S1، تلعب المناعة الخلوية أيضًا دورًا مهمًا في إزالة الفيروس من خلال تدريب خلايا T السامة للتعرف على وقتل خلايا المضيف المصابة بالفيروس . لذلك، تم تقييم مستويات خلايا T السامة في طحال الفئران في اليوم 28 بعد الإدارة الأولية. على الرغم من عدم ملاحظة تغيير واضح في و تسلل الخلايا في المجموعات المختلفة (الشكل 5h)، كانت إفرازات IFN- من خلايا و خلايا قد زادت بشكل واضح في الفئران بعد التوصيل عبر الجلد لـ FCS/S1/PolyIC مقارنة بتلك الموجودة في الفئران التي تلقت حقنًا تحت الجلد لـ S1/polyIC (الشكل 5i، j)، مما يدل على أن استجابات خلايا T السامة كانت أقوى نتيجة للتوصيل عبر الجلد للقاح S1. علاوة على ذلك، كانت نسبة كل من خلايا الذاكرة الفعالة و خلايا و) في طحال الفئران المعالجة بـ FCS/S1/ polylC قد زادت بشكل كبير أيضًا في اليوم 90 مع تعزيز واحد في اليوم 75 (الشكل S24)، بينما كانت تلك في الفئران التي تلقت الإدارة تحت الجلد لـ S1/PolylC تبدو أقل بكثير من تلك في المجموعة عبر الجلد. في اليوم 75 (قبل التعزيز) واليوم 90 (15 يومًا بعد تعزيز واحد)، تم قياس السيتوكينات في مصل الفئران بعد إدارات مختلفة (الشكل S25). كانت مستويات IL-12p40، وهو علامة مهمة للمناعة الفطرية، وIFN- ، العلامات النموذجية للمناعة الخلوية، جميعها أعلى بشكل واضح في الفئران المعالجة بـ FCS/S1/PolyIC، مما يوضح أن توصيل اللقاحات عبر الجلد سيحفز تأثير ذاكرة المناعة التكيفية على المدى الطويل، والذي قد يكون ناتجًا عن الاحتفاظ الطويل للمستضد المعطى مع المعزز.
من أجل التحقيق في القدرة الدقيقة للاختراق للقاحات بعد الإدارة عبر الجلد، قمنا بقياس تراكم المستضدات في العقد اللمفاوية بعد 24 ساعة من التوصيل عبر الجلد أو الحقن تحت الجلد للقاح باستخدام OVA المسمى بـ Cy5.5 كمستضد نموذجي. كما هو متوقع، مقارنةً بالحقن تحت الجلد لـ OVA، أظهرت الفئران بعد التوصيل عبر الجلد للقاح بعد مسح المرهم تمامًا عن سطح الجلد احتفاظًا أقل لـ OVA وفقًا لنظام تصوير IVIS (الشكل S26). ثم، قمنا بتقييم المزيد من امتصاص OVA بواسطة DCs في العقد اللمفاوية والجلد (الشكل 5k، l). من المثير للاهتمام، على الرغم من أن نسبة DCs في العقد اللمفاوية للفئران التي تلقت الحقن تحت الجلد كانت أعلى من تلك الخاصة بالفئران التي تلقت التوصيل عبر الجلد، أظهرت خلايا DCs تحت الجلد امتصاصًا أعلى لمستضد OVA في الفئران المعالجة بالمراهم التي تحتوي على FCS/OVA/polyIC، مما يكشف أن توصيل اللقاح عبر الجلد يمكن أن ينشط خلايا DCs مباشرة في الموقع، وأن خلايا DCs المنشطة يمكن أن تهاجر إلى العقد اللمفاوية لتحفيز استجابات مناعية إضافية. بالمقابل، بالنسبة لمجموعة الحقن تحت الجلد، فإن المستضد والمساعد المناعي سيتجهان إلى العقد اللمفاوية بشكل منفصل ومن المرجح أن يتم استنفادهما بواسطة خلايا المناعة غير النوعية وتنشيط خلايا DCs بفعالية منخفضة..
بالإضافة إلى ذلك، قمنا بقياس نضوج خلايا DC وتنشيطخلايا T بعد يوم واحد وثلاثة أيام من علاجات مختلفة. كما هو موضح في الشكل 5م، ن، أظهرت الفئران التي تلقت لقاحات عبر الجلد نضوجًا مشابهًا تقريبًا لخلايا DC وتنشيط خلايا T مقارنة بالفئران التي تلقت حقن تحت الجلد، مما يتماشى مع الملاحظات التي تم رصدها.تنشيط الخلايا في الطحال. لذلك، على الرغم من أن توصيل اللقاح عبر الجلد المعتمد على FCS أظهر اختراقًا أقل لل antigene مقارنةً بالحقن تحت الجلد، إلا أنه يمكن أن يحفز مناعة خلوية قوية ومناعة هومورالية شبه مماثلة. ومن المزايا الإضافية لمثل هذه اللقاحات المعتمدة على FCS إمكانية الإدارة الذاتية وامتثال المستخدم المفضل. ومع ذلك، لم تُجرَ دراسات إضافية، مثل تحدي فيروس SARS-CoV-2، بسبب قيودنا الحالية في الظروف التجريبية للتعامل مع الفيروسات.
قدرة اختراق المركبات النانوية المعتمدة على FCS في نماذج الأرانب والخنازير
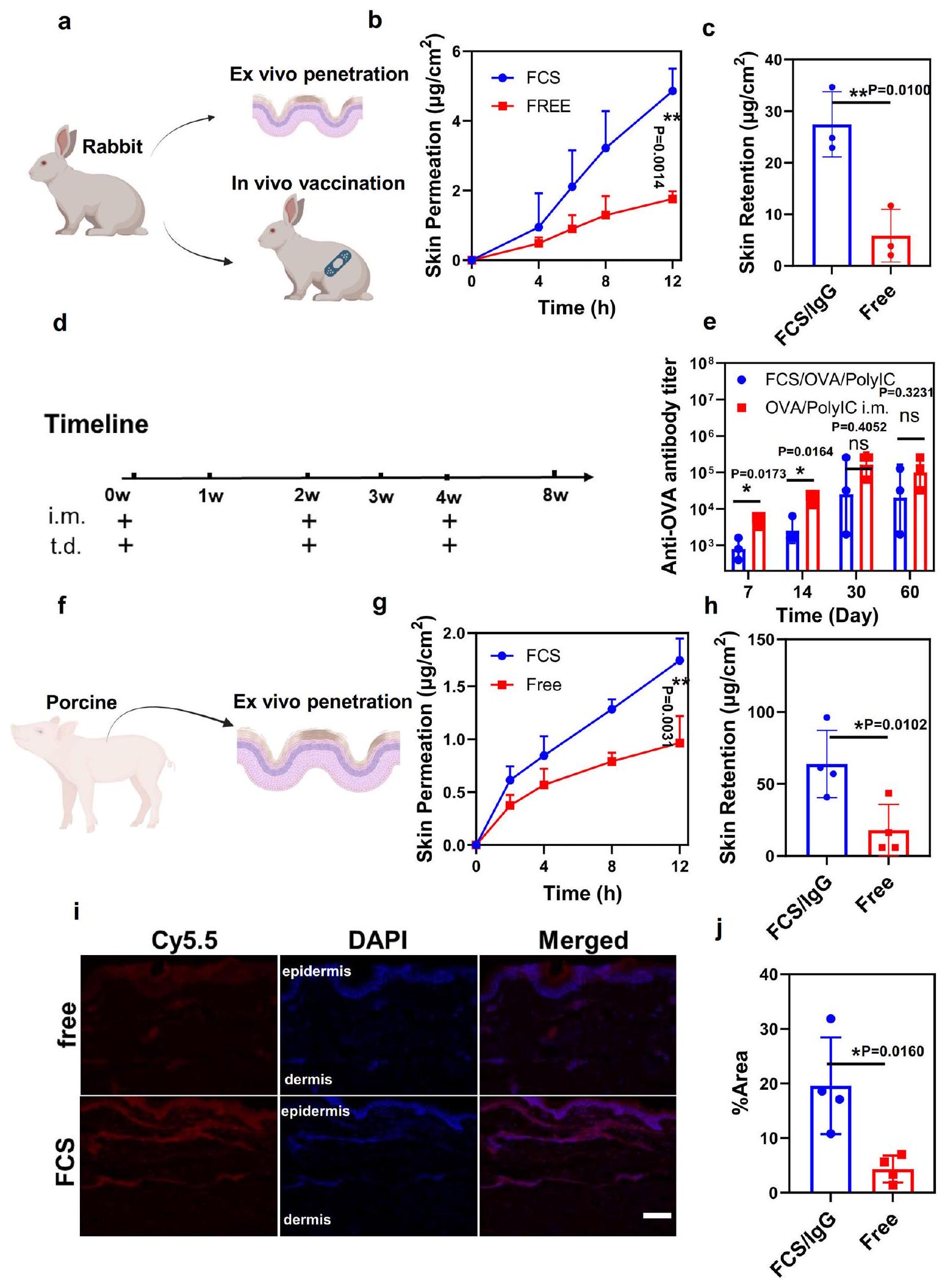
بعد ذلك، قمنا بمزيد من الاختبارات لقدرة FCS على توصيل البروتينات من خلال جلد الحيوانات الأكبر مثل الأرانب (الشكل 6a). أولاً، تم إجراء اختبار انتشار فرانز على جلود الأرانب في المختبر. في هذه الحالة، تمت إزالة طبقة الدهون تحت الجلد من جلود الأرانب لتقييم أكثر دقة. تم قياس مقاومات الجلد قبل وبعد الاختبار لتأكيد سلامة الجلد (الشكل S29). مقارنةً بـ IgG الحر، أظهر FCS/IgG زيادة ملحوظة في اختراق الجلد لجلود الأرانب بكمية تبلغ حوالي (مجموع النفاذ والاحتفاظ في الأدمة، الشكل 6ب، ج). ثم تم إجراء التطعيم على الأرانب. تم تقسيم الأرانب عشوائيًا إلى ثلاث مجموعات: (1) غير معالجة، (2) تطبيق موضعي عبر الجلد لـ FCS/OVA/polyIC، و(3) حقن عضلي (i.m.) لـ OVA/polyIC. كما هو موضح في الشكل 6د، تم تطبيق نفس جرعة OVA وpolyIC على كل من المجموعتين عبر الجلد والعضلي، وفقًا لنفس جدول الإدارة في اليوم 0، اليوم 14، واليوم 30. كما هو موضح في الشكل 6هـ، يبدو أن عيار الأجسام المضادة في أرانب مجموعة التطعيم عبر الجلد قريب من مجموعة التطعيم العضلي، خاصة في النقاط الزمنية اللاحقة (مثل الأيام 30 و60)، مما يدل على نجاح توصيل اللقاحات المعتمدة على FCS عبر الجلد في نموذج الأرانب من أجل تطعيم فعال.
في هذه الأثناء، تم إجراء اختبارات الاختراق خارج الجسم أيضًا على جلود الخنازير. تم الحصول على جلود الخنازير من ظهر الخنازير الصغيرة في عمر ثلاثة أشهر وتمت إزالة جميع طبقة الدهون تحت الجلد لتقييم أكثر دقة. تم قياس مقاومات الجلد قبل وبعد الاختبار لتأكيد سلامة الجلد (الشكل S30). وبالمثل، أظهر اختبار انتشار فرانز أن FCS/IgG حقق أيضًا قدرة على الانتقال عبر الجلد تصل إلى (مجموع البروتينات التي تم اختراقها واحتجازها داخل الجلد). قد تكون الفعالية الأكبر ناتجة عن حجم بصيلات الشعر والغدد العرقية الأكبر في الخنازير. بالإضافة إلى ذلك، أظهرت الصورة المجسمة لشرائح جلد الخنازير أيضًا اختراقًا كبيرًا للأجسام المضادة IgG المعلّمة بالفلوروسنت في منطقة الأدمة لجلود الخنازير التي تم تطبيقها موضعيًا باستخدام FCS/IgG-Cy5.5 (الشكل 6i، j). بشكل جماعي، أظهرت مجمعات البروتينات النانوية FCS/ نجاحًا في توصيل البروتينات عبر الجلد إلى الأدمة والمناطق تحت الجلد لجلود الفئران والأرانب والخنازير.
نقاش
في الختام، قمنا بتطوير نظام توصيل عبر الجلد يعتمد على FCS لتوصيل فعال محليًا للبيومكرومولكولات، بما في ذلك الأجسام المضادة، المستضدات، أو الأحماض النووية (مثل البوليلC)، والتي يمكن خلطها مع FCS وإضافتها إلى مرهم للاستخدام الموضعي. لتوصيل عبر الجلد للأجسام المضادة مثل aPDL1 و aCTLA4، فإننا
الشكل 6 | تقييم قدرة البروتين عبر الجلد على نماذج الأرانب والخنازير. أ رسم توضيحي تخطيطي للتطعيم داخل الجسم على نموذج الأرنب. ب، ج النسب التراكمية للاختراق (ب) والاحتفاظ بالجلد (ج) لبروتين FCS/IgG-FITC الذي اخترق جلد الأرنب مع مرور الوقت. ). الجرعة الكلية: رسم توضيحي تخطيطي لتصميم التجربة يوضح توصيل اللقاح OVA عبر الجلد في نموذج الأرنب. عيار الأجسام المضادة IgG الخاصة بـ OVA في مصل الأرنب عند فترات زمنية مختلفة تم تحديدها بواسطة ELISA. ). رسم توضيحي تخطيطي لاختراق الجلد خارج الجسم على نموذج الخنازير.النسب المئوية التراكمية لـ اختراق (ج) واحتفاظ الجلد (ح) لـ FCS/IgG-FITC الذي تم اختراقه عبر جلد الخنازير مع مرور الوقت ( ). الجرعة الكلية: صور الفلورية التداخلية التمثيلية.تحليل إحصائي لإظهار عمق الاختراق FCS/ IgG-Cy5.5 من خلال جلد الخنازير خلال 12 ساعة. تم استخدام IgG-Cy5.5 الحر كعنصر تحكم في تلك التجارب. ). مقياس الرسم: . تم إنشاء جميع الرسوم التوضيحية باستخدام بايو ريندر.كومتُعرض البيانات كمتوسطالانحراف المعياري. تم حساب الدلالة الإحصائية من خلال تحليل التباين أحادي الاتجاه مع اختبار توكي بعد الاختبار.; تُقدم بيانات المصدر كملف بيانات مصدر.
يمكن أن تؤدي أنظمة التوصيل المعتمدة على FCS إلى تراكم عالٍ للأجسام المضادة في الأورام الميلانينية واستجابات قوية للخلايا التائية، وبالتالي القضاء بنجاح على الأورام الأولية ومنع نمو الأورام البعيدة في الفئران. كما تم الإبلاغ، قد تؤدي التوصيل المشترك لـ aPDL1 و aCTLA4 عن طريق الإدارة الوريدية إلى مجموعة متنوعة من الآثار الجانبية، بما في ذلك السمية الكبدية، والتهاب الرئة المناعي، وحتى الأمراض الغدد الصماء الذاتية.مع استجابة علاجية محسّنة بشكل كبير مقارنةً بحقن الأجسام المضادة النظامية بنفس الجرعة، قد تؤدي طريقة توصيل الأجسام المضادة المثبطة للمناعة المعتمدة على FCS إلى تقليل المخاوف بشأن الآثار الجانبية النظامية نظرًا لانخفاض تركيزات المصل نسبيًا من خلال طريقة الإدارة الموضعية. من ناحية أخرى، فإن توصيل الأدوية عبر الجلد لـ SARS-أدت اللقاحات إلى تحقيق مستوى من الأجسام المضادة المحددة لـ S1 مشابه لذلك الذي تم تحقيقه عن طريق الحقن تحت الجلد، واستجابات أقوى للخلايا التائية. قد يمكّن هذا التسليم عبر الجلد للقاحات SARS-CoV-2 من التطعيم السريع والواسع حتى في المنزل بمجرد إثبات فعالية هذه الاستراتيجية في دراسات لاحقة. وقد وُجد أن تسليم البروتين عبر الجلد القائم على FCS قابل للتطبيق على جلود الحيوانات الأكبر مثل الأرانب والخنازير. وبالتالي، حقق عملنا تسليمًا فعالًا وغير جراحي للبروتينات الكبيرة عبر الجلد دون الحاجة إلى أي تحفيزات جسدية أو كيميائية إضافية، وهو ما كان تحديًا إلى حد ما عبر التقنيات الحالية. بالإضافة إلى توصيل الأجسام المضادة لنقاط التفتيش المناعية واللقاحات، يمكن أيضًا استخدام الناقل الذي تم تطويره لتسليم الجزيئات الكبيرة عبر الجلد لتوصيل جزيئات حيوية علاجية أخرى تهدف إلى تطبيقات طبية متنوعة، ويحتفظ بإمكانات هائلة للتسويق.
طرق
بيان أخلاقي
تم إجراء جميع الدراسات الحيوانية، بما في ذلك التجارب على الفئران والأرانب، وفقًا للبروتوكولات المعتمدة من مركز الحيوانات المختبرية بجامعة سوتشو ولجنة الأخلاقيات بجامعة سوتشو.
المواد، خطوط الخلايا، ونماذج الحيوانات
كيتوزان (DD%)تم شراء ) من شركة العلاء الصناعية (شنغهاي، الصين)، وتم توفير N -(3-(ثلاثي ميثيل أمين) بروبيل)-N-إيثيل كاربودييميد هيدروكلوريد بلوري (EDC) وN-هيدروكسي سوكسينيميد (NHS) من شركة JK Chemical (بكين، الصين). تم الحصول على محلول ملحي معزز بالفوسفات (PBS) من شركة Beijing Solarbio Science Technology Co., Ltd. تم شراء بروتين سبايك لفيروس SARS-CoV-2 من Sino Biological. تم زراعة خط خلايا ورم الميلانوما الفأري B16F10 تحت ظروف قياسية موصى بها من قبل مجموعة الثقافة الأمريكية (ATCC). تم شراء إناث الفئران C57BL/6 (6-8 أسابيع) من شركة Nanjing Pengsheng Biological Technology. تم شراء الأرانب النيوزيلندية (3 كجم) من شركة Suzhou Jinghu Biological Technology. تم جمع جلود الفئران والأرانب في المختبر. تم شراء جلود الخنازير من شركة Taizhou Taihe Biological Technology. تم إجراء جميع الدراسات الحيوانية وفقًا للبروتوكولات المعتمدة من مركز الحيوانات المخبرية بجامعة سوتشو.
تركيب الكيتوزان المعدل بالفلوروكربون
حمض البيرفلوروهيبتانوicتم إذابته في ثنائي ميثيل سلفوكسيد (DMSO، سانغون بيوتيك) ثم تم خلطه مع EDC (78 ملغ، 1.5 مكافئ) و NHS (47.43 ملغ، 1.5 مكافئ) تحت التحريك في الظلام عند درجة حرارة الغرفة لمدة 0.5 ساعة للحصول على حمض البيرفلوروهبتانويك المنشط. ثم، تم إذابة CS (200 ملغ) في محلول حمض الأسيتيك 1%، ومزجه بالتنقيط مع حمض البيرفلوروهبتانويك المنشط، وتحريكه في الظلام لمدة 12 ساعة لتخليق FCS. تم تنقية FCS الناتج عن طريق الغسيل في ماء مقطر مزدوج باستخدام كيس غسيل (MWCO 3500 دالتون، Technologies Co.) لمدة 48 ساعة. ثم، تم تجفيف العينات بالتجميد، وتم تحديد درجة الاستبدال لمجموعات الفلوروالكيل المرتبطة على كل CS بواسطة قياس اللون باستخدام النينهدين وفقًا للإجراء المثبت جيدًا وتم تحليلها باستخدام قارئ الميكرو بلايت (Variskan، THERMO). تم إجراء الرنين المغناطيسي النووي ^19F (جهاز Bruker AV III 600) لـ FCS (5.0 ملغ) المذاب فيمن يحتوي على 1.0 ملغ من تم تطبيقه أيضًا لحساب المستبدلات الفلورو ألكيلية المترافقة.
تحضير النانومركبات المعتمدة على FCS
تم خلط FCS المُركب مع IgG في ماء منزوع الأيونات بنسب وزن مختلفة (من 1:4 إلى 4:1) لمدة 0.5 ساعة لتحضير نانو معقدات FCS/IgG. تم تحضير FCS/aPDL1 وFCS/aCTLA4 وFCS/OVA وFCS/S1 بنفس الطريقة. تم تسجيل طيف الامتصاص للأشعة فوق البنفسجية والمرئية لمختلف المعقدات بواسطة مطياف الأشعة فوق البنفسجية والمرئية-NIR من نوع PerkinElmer Lambda 750 (PerkinElmer، الولايات المتحدة الأمريكية). تم الكشف عن الحجم، والجهد الزتاوي، ومؤشر التشتت للمعقدات النانوية الناتجة بواسطة جهاز Zetasizer Nano ZS (Malvern Instrument). تم استخدام المجهر الإلكتروني الناقل (FEI TECNAI G2) لمراقبة شكل المعقدات النانوية النموذجية.
كفاءة توصيل عبر الجلد
تمت دراسة قدرة توصيل FCS عبر الجلد باستخدام نظام خلية الانتشار فرانز. تم شراء نظام الانتشار فرانز من شركة هوانغهاي المحدودة في شنغهاي. تم جمع جلود الفئران والأرانب والخنازير الطازجة وإضافتها بين غرف المانح والمستقبل. تم جمع عينات من غرفة المستقبل في نقاط زمنية مختلفة وتم إضافة نفس حجم من PBS إلى غرفة المستقبل للحفاظ على حجم ثابت. تم حساب معدل الاختراق النهائي باستخدام الصيغة:
هو حجم غرفة المستقبل (8 مل)هو حجم العينات المجمعة في كل نقطة زمنية (0.4 مل)، Cn هو تركيز FITC في غرفة المستقبل عند كل نقطة زمنية، هو تركيز العينات المجمعة في نقطة زمنية، وA هو تركيز التغذية لـ FITC. تركيز تغذية البروتينات هوفيحل PB (مساحة جلد النفاذية هي.
تحضير مرهم عبر الجلد قائم على FCS وإدارته في الجسم الحي
تم إسقاط المركبات النانوية المركبة من FCS/IgG أو غيرها من FCS/البروتين على سطح المرهم الفارغ (Aquaphor)، بشكل رئيسي الفازلين) بنسبة كتلة 1:1، مثلFCS/بروتين مع 20 ملغ من المرهم. ثم تم خلطهم برفق مع بعضهم البعض. سيصبح المرهم الشفاف الخالي من المواد مثل المرهم الحليبي، والذي ثم تم تطبيقه على جلد الفأر والأرنب للاستخدامات عبر الجلد.
لإدارة الفئران، تم مسح المرهم المختلط الشبيه بالحليب على جلد الفأر باستخدام غطاء دائري منبقطر) لمدة 12 ساعة. تم تغطية الفيلم الشفاف (3 م) أخيرًا لتجنب تأثير لعق الفئران.
لإدارة الأرانب، تم مسح المرهم المختلط الشبيه بالحليب على جلد الأرنب باستخدام غطاء دائري منفي نصف القطر) لمدة 12 ساعة. تم تغطية الفيلم الشفاف (3M) أخيرًا لتجنب تأثير لعق الأرنب.
توسيم، استقرار، وتوزيع حيوي داخل الكائن الحي
تم وسم IgG بـأنا بواسطة طريقة أكسدة الكلورامين-T القياسية. باختصار، IgG،تم خلطي، وكلورامين-T فيلمدة 10 دقائق تحت اهتزاز خفيف. المركب المُصنّعتم غسلها بواسطة الترشيح الفائق 10 كيلودالتون ثلاث مرات. لتحديد استقرار الوسم الإشعاعي،تم خلطه مع المصل لمدة 24 ساعة فيتم أخذ عينات من أجزاء من الخليط في فترات زمنية مختلفة وتم تصفيتها بواسطة الترشيح الفائق 10 كيلودالتون. تم الكشف عن النشاط الإشعاعي المحتجز على المرشحات بواسطة جهاز عدّ غاما Wizard2 (PerkinElmer) لحساب استقرار الوسم الإشعاعي.
للتوزيع الحيوي في الجسم الحي لـتم إعطاء الفئران الحاملة لورم الميلانوما B16F10 موضعياً مع و FCS مرهم I-IgG لنقاط زمنية مختلفة. ثم تم جمع الأعضاء الرئيسية، بما في ذلك الكبد والطحال والكلى والقلب والرئة والورم، وتم قياسها بواسطة جهاز قياس غاما Wizard2 (PerkinElmer). تم حساب فعالية التراكم بواسطة التركيبة.
أينكانت شدة الراديو للعضو في النقطة الزمنية الدقيقة،كان كتلة العضو،كانت شدة الإشعاع للرغوة المطبقة.
علاج في الجسم الحي للورم الميلانيني
تم إنشاء نموذج ورم الميلانوما B16F10 عن طريق الحقن تحت الجلد لـتم حقن خلايا B16F10 في الجانب الأيمن من كل فأرة C57BL/6 أنثوية. تم تقسيم الفئران عشوائيًا إلى خمس مجموعات عندما وصلت أحجام الأورام إلى حوالي. تم معالجة كل مجموعة بجرعة متساوية من aPDL1 و aCTLA4 (تم مسح أورام الفئران بمراهم FCS مختلفة لمدة 12 ساعة، وتم تغطيتها بفيلم شفاف (3M) لتجنب تأثير لعق الفئران. تم قياس حجم الورم بواسطة كاليبر كل يومين:
تم إيقاف منحنيات النمو عندما توفيت الفأر الأول في المجموعة المعنية، أو عندما تجاوز حجم ورم الفأر الأولبموجب الأخلاقيات من مركز الحيوانات المخبرية بجامعة سوتشو ولجنة الأخلاقيات بجامعة سوتشو. في بعض الحالات، تم تجاوز هذا الحد في اليوم الأخير من القياس، وتم euthanized الفئران على الفور.
تلطيخ المناعة الفلورية
لصبغة المناعة الفلورية في المختبر، تم حضن خلايا HACAT في طبق 24 بئر لتشكيل طبقة خلوية واحدة. ثم، تم إضافة FCS/IgG وتم حضنها لمدة 12 ساعة أخرى. بعد إزالة FCS-IgG، تم تثبيت خلايا HACAT بواسطةبارافورمالدهيد، محجوز بواسطة 5% BSA، وتم صبغه باستخدام جسم مضاد مضاد لـ ZO-1. بعد ساعة، تم صبغ الخلايا باستخدام جسم مضاد ثانوي و 4، 6-دياميدينو-2-فينيل إندول (DAPI). بعد التركيب، تم تصوير الشرائح بواسطة ميكروسكوب فلوروسنس تداخلي (لايكا SP5). بالنسبة لصبغ المناعة الفلورية في الجسم الحي، تم جمع أورام B16F10 والجلد من الفئران في نقاط زمنية مختلفة بعد مسحها بمراهم مختلفة وتضمينها في جل OCT المستجيب لدرجة الحرارة لتلوين المناعة الفلورية مع أجسام مضادة مختلفة، بما في ذلك مضاد الكيراتين 14 ومضاد ZO-1. بعد غسل الأجسام المضادة الأولية، تم تلوين شرائح الورم بأجسام مضادة ثانوية موسومة بالفلور وDAPI. بعد التركيب، تم تصوير شرائح الورم بواسطة المجهر الفلوري المجهري (لايكا SP5).
تحليل خلايا T في أعضاء مختلفة
تم جمع الطحال والأورام والجلود والعقد اللمفاوية وتجانسها إلى تعليقات خلوية مفردة وفقًا للبروتوكول القياسي. باختصار، تم معالجتها من خلال التكسير الميكانيكي قبل الهضم لمدة ساعة واحدة عندفي محلول إنزيمي مع RPMI-1640 FBS و 1% PS)، كولاجيناز IV (سيغما)لاجنز I (سيغما)هيالورونيداز (سيغما)، و DNase I (سيغما). ثم تم تمرير العينات من خلال فلاتر نايلون بشبكة 200 للحصول على تعليقات خلوية مفردة. تم حضن التعليقات الخلوية المفردة الناتجة مع مضاد CD16/32 لمدة 30 دقيقة عند . ثم تم صبغ هذه الخلايا بأجسام مضادة مختلفة وفقًا للبروتوكول القياسي.
تم صبغ خلايا DCs في العقد اللمفاوية والجلد باستخدام مضاد CD11c-FITC (Biolegend، الرقم المرجعي 117305)، ومضاد CD80-APC (Biolegend، الرقم المرجعي 104713)، ومضاد CD86-PE (Biolegend، الرقم المرجعي 105007).
تم صبغ مجموعة خلايا T في الأورام والطحال باستخدام مضاد CD3-FITC (Biolegend، الرقم المرجعي 100203)، مضاد CD4-APC (Biolegend، الرقم المرجعي 100411)، مضاد CD45-PerCP (Biolegend، الرقم المرجعي 103130)، ومضاد CD8a-PE (Biolegend، الرقم المرجعي 100707).
لتحليل خلايا T المساعدة في الأورام بشكل أعمق، تم صبغ التعليق باستخدام الأجسام المضادة anti-CD3-FITC (Biolegend، الرقم المرجعي 100203)، anti-CD4-APC (Biolegend، الرقم المرجعي 100411)، وanti-Foxp3-PE (Biolegend، الرقم المرجعي 126403) مع eBioscience.تثبيت/اختراق عامل النسخ Foxp3/تركيز ومخفف وفقًا لبروتوكولات الشركة المصنعة. باختصار، بعد صبغ الخلايا بـ CD16/CD32 وعلامات سطح الخلية، تم تثبيتها باستخدام محلول العمل لتثبيت/اختراق Foxp3 لمدة 30 دقيقة في درجة حرارة الغرفة. ثم، تم غسلها بمحلول الاختراق عند 500 جرام لمدة 5 دقائق وصبغها بـ CD16/CD32 في محلول الاختراق لمدة 30 دقيقة في. ثم تم تلطيخها بمضاد Foxp3-PE لمدة 30 دقيقة أخرى في درجة حرارة الغرفة وأعيد تعليقها في محلول تلوين تدفق الخلايا للتحليل.
لتحليل السمية الخلويةتم صبغ تعليق الخلايا اللمفاوية باستخدام مضاد CD3-APC (Biolegend، الرقم المرجعي 100206)، مضاد CD8a-PE (Biolegend، الرقم المرجعي 100707)، مضاد CD45-PerCP (Biolegend، الرقم المرجعي 103130) ومضاد Ki67-FITC (Biolegend، الرقم المرجعي 652410) لمستوى Ki67 بنفس البروتوكول المستخدم في Foxp3. كما تم صبغ التعليق بمضاد CD45-FITC (Biolegend، الرقم المرجعي 157214)، مضاد CD8a-PE (Biolegend، الرقم المرجعي 100707)، مضاد GranzymeB-PE-Cy7 (Biolegend، الرقم المرجعي 372214) ومضاد CD3-APC (Biolegend، الرقم المرجعي 100206) لمستوى Granzyme B باستخدام eBioscience.مجموعة محلول التثبيت والاختراق داخل الخلايا.
لتحليل IFNالخلايا في الطحال، تم تحضين التعليق أولاً بوسط تحفيز يحتوي على RPMI-1640 (10% FBS و ) ، محلول HEPES (سيغما)، محلول 1 مللي مول من صوديوم بايروفات (سيغما)،2-ميركابتوإيثانولبروتين S1 أو OVA، وإي بايوساينسمحلول بريفيلدين A لمدة 6 ساعات ثم تم صبغه بمضاد CD3-FITC (بايو ليجند، رقم الكاتالوج 100203)، مضاد CD4-PerCP (بايو ليجند، رقم الكاتالوج 100432)، مضاد CD8a-PE (بايو ليجند، رقم الكاتالوج 100707) ومضاد IFN--APC (بيوليجند، رقم الكاتالوج 505810) مع إي بايوساينسمجموعة محلول التثبيت والاختراق داخل الخلايا. لتحليل IFNالخلايا في الأورام، eBioscience cocktail تحفيز الخلايا (بالإضافة إلى مثبطات نقل البروتين) ) تم إضافته في وسط التحفيز لاستبدال البروتين وبريفيلدين أ في البروتوكول أعلاه.
لتحليل خلايا الذاكرة T، تم جمع الطحال من الفئران بعد 90 يومًا من التحفيز والتعزيز وتم صبغه بمضاد CD3-FITC (eBioscience، الرقم المرجعي 11-0031)، مضاد CD8-PerCP-Cy5.5 (eBioscience، الرقم المرجعي 45-0081)، مضاد CD44-PE (eBioscience، الرقم المرجعي 12-0441)، ومضاد CD62L-
APC (eBioscience، الرقم المرجعي 17-0621). تم تخفيف جميع هذه الأجسام المضادة المستخدمة في تجاربنا بحوالي 200 مرة.
مستوى الأجسام المضادة الخاصة بـ S1
تم قياس عيار الأجسام المضادة الخاصة بـ S1 بواسطة طريقة ELISA غير المباشرة القياسية (iELISA). باختصار، تم طلاء ألواح ELISA المكونة من 96 بئرًا (Nunc، Thermo) بين عشية وضحاها بـ 1 نانوغرام لكل بئر من بروتين S1 في محلول الطلاء. ثم تم حجب الألواح بـمن محلول عازل اختبار ELISA (ثيرمو) وتم حضنه في درجة حرارة الغرفة لمدة ساعتين. تم اختبار كل عينة مصل في نسختين بتخفيف من 1:100 إلى 1:640,000 مرتين في محلول عازل اختبار ELISA (ثيرمو)، حيثثم أضيف إلى آبار كل طبق لمدة ساعتين من الحضانة في درجة حرارة الغرفة. تم إضافة إنزيم البيروكسيداز المستخلص من الفجل (HRP) المرتبط بأجسام مضادة من الماعز ضد IgG الفأر (1:10,000، Abcam) للحضانة لمدة ساعة واحدة في درجة حرارة الغرفة. ثم،تم إضافة محلول TMB (Thermo) إلى كل بئر. بعد 15 دقيقة من الحضانة،تم إضافة محلول لإيقاف التفاعل. تم قراءة امتصاص كل بئر عند طول موجي 450 نانومتر (PerkinElmer). تم تعريف العتبة على أنها ضعف متوسط الامتصاص في المجموعة غير المعالجة. يتم تعريف آخر تخفيف لعينة يكون أكبر قليلاً من العتبة على أنه عيار الأجسام المضادة لهذه العينة.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
يعلن المؤلفون أن جميع البيانات اللازمة لتقييم استنتاجات هذا العمل موجودة في المقالة أو المعلومات التكميلية أو ملف بيانات المصدر. تم توفير بيانات المصدر في هذه الورقة.
References
Prausnitz, M. R. & Langer, R. Transdermal drug delivery. Nat. Biotechnol. 26, 1261-1268 (2008).
Prausnitz, M. R., Mitragotri, S. & Langer, R. Current status and future potential of transdermal drug delivery. Nat. Rev. Drug Discov. 3, 115-124 (2004).
Alkilani, A., McCrudden, M. T. & Donnelly, R. Transdermal drug delivery: innovative pharmaceutical developments based on disruption of the barrier properties of the stratum corneum. Pharmaceutics 7, 438-470 (2015).
Prausnitz, M. R. Microneedles for transdermal drug delivery. Adv. Drug Deliv. Rev. 56, 581-587 (2004).
Naik, A., Kalia, Y. N. & Guy, R. H. Transdermal drug delivery: overcoming the skin’s barrier function. Pharm. Sci. Technol. Today 3, 318-326 (2000).
Karande, P., Jain, A., Ergun, K., Kispersky, V. & Mitragotri, S. Design principles of chemical penetration enhancers for transdermal drug delivery. Proc. Natl Acad. Sci. USA 102, 4688-4693 (2005).
Amjadi, M., Mostaghaci, B. & Sitti, M. Recent advances in skin penetration enhancers for transdermal gene and drug delivery. Curr. Gene Ther. 17, 139-146 (2017).
Ahn, D.-G. et al. Current status of epidemiology, diagnosis, therapeutics, and vaccines for novel coronavirus disease 2019 (COVID19). J. Microbiol. Biotechnol. 30, 313-324 (2020).
Ita, K. Transdermal delivery of vaccines-recent progress and critical issues. Biomed. Pharmacother. 83, 1080-1088 (2016).
Nasrollahi, S. A., Taghibiglou, C., Azizi, E. & Farboud, E. S. Cellpenetrating peptides as a novel transdermal drug delivery system. Chem. Biol. Drug Des. 80, 639-646 (2012).
Desai, P., Patlolla, R. R. & Singh, M. Interaction of nanoparticles and cell-penetrating peptides with skin for transdermal drug delivery. Mol. Membr. Biol. 27, 247-259 (2010).
Tadros, A. R. et al. STAR particles for enhanced topical drug and vaccine delivery. Nat. Med. 26, 341-347 (2020).
Zheng, D. et al. Topical delivery of siRNA-based spherical nucleic acid nanoparticle conjugates for gene regulation. Proc. Natl Acad. Sci. USA 109, 11975-11980 (2012).
Chen, Y. et al. Transdermal protein delivery by a coadministered peptide identified via phage display. Nat. Biotechnol. 24, 455-460 (2006).
Marwah, H., Garg, T., Goyal, A. K. & Rath, G. Permeation enhancer strategies in transdermal drug delivery. Drug Deliv. 23, 564-578 (2016).
Cross, S. & Roberts, M. Physical enhancement of transdermal drug application: is delivery technology keeping up with pharmaceutical development? Curr. Drug Deliv. 1, 81-92 (2004).
Nanda, A., Nanda, S. & Khan Ghilzai, N. Current developments using emerging transdermal technologies in physical enhancement methods. Curr. Drug Deliv. 3, 233-242 (2006).
Smith, T. R. F. et al. Immunogenicity of a DNA vaccine candidate for COVID-19. Nat. Commun. 11, 2601 (2020).
Xia, D. et al. An ultra-low-cost electroporator with microneedle electrodes (ePatch) for SARS-CoV-2 vaccination. Proc. Natl Acad. Sci. USA 118, e2110817118 (2021).
Tran, K. T. M. et al. Transdermal microneedles for the programmable burst release of multiple vaccine payloads. Nat. Biomed. Eng. 5, 998-1007 (2020).
Lu, B. et al. lonic liquid transdermal delivery system: progress, prospects, and challenges. J. Mol. Liq. 351, 118643 (2022).
Song, X. et al. Transcutaneous tumor vaccination combined with anti-programmed death-1 monoclonal antibody treatment produces a synergistic antitumor effect. Acta Biomater. 140, 247-260 (2022).
Yang, X. et al. Galactosylated chitosan-modified ethosomes combined with silk fibroin nanofibers is useful in transcutaneous immunization. J. Controlled Release 327, 88-99 (2020).
Shariatinia, Z. Pharmaceutical applications of chitosan. Adv. Colloid Interface Sci. 263, 131-194 (2019).
Li, G. et al. Fluorinated chitosan to enhance transmucosal delivery of sonosensitizer-conjugated catalase for sonodynamic bladder cancer treatment post-intravesical instillation. ACS Nano 14, 1586-1599 (2020).
Zhou, X. et al. Nano-formulations for transdermal drug delivery: a review. Chin. Chem. Lett. 29, 1713-1724 (2018).
Niu, X.-Q. et al. Mechanism investigation of ethosomes transdermal permeation. Int J. Pharm. X 1, 100027 (2019).
Lueßen, H. L. et al. Bioadhesive polymers for the peroral delivery of peptide drugs. J. Controlled Release 29, 329-338 (1994).
Illum, L., Farraj, N. F. & Davis, S. S. Chitosan as a novel nasal delivery system for peptide drugs. Pharm. Res. 11, 1186-1189 (1994).
Antonescu, I. E. et al. The permeation of acamprosate is predominantly caused by paracellular diffusion across cell monolayers: a paracellular modeling approach. Mol. Pharm. 16, 4636-4650 (2019).
Bittermann, K. & Goss, K.-U. Predicting apparent passive permeability of CaCo-2 and MDCK cell-monolayers: a mechanistic model. PLoS ONE 12, e0190319 (2017).
Ahmadi, S. et al. A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis. JCI Insight 5, e132055 (2020).
Robert-Paganin, J., Pylypenko, O., Kikuti, C., Sweeney, H. L. & Houdusse, A. Force generation by myosin motors: a structural perspective. Chem. Rev. 120, 5-35 (2020).
Kováčik, A., Kopečná, M. & Vávrová, K. Permeation enhancers in transdermal drug delivery: benefits and limitations. Expert Opin. Drug Deliv. 17, 145-155 (2020).
Fan, K. et al. Ferritin nanocarrier traverses the blood brain barrier and kills glioma. ACS Nano 12, 4105-4115 (2018).
Liu, Y. et al. A brief review for fluorinated carbon: synthesis, properties and applications. Nanotechnol. Rev. 8, 573-586 (2019).
Lee, Y.-S. Syntheses and properties of fluorinated carbon materials. J. Fluor Chem. 128, 392-403 (2007).
Miller, A. J. & Mihm, M. C. Melanoma. N. Engl. J. Med. 355, 51-65 (2006).
Franklin, C., Livingstone, E., Roesch, A., Schilling, B. & Schadendorf, D. Immunotherapy in melanoma: recent advances and future directions. Eur. J. Surg. Oncol. 43, 604-611 (2017).
Martin-Liberal, J., Kordbacheh, T. & Larkin, J. Safety of pembrolizumab for the treatment of melanoma. Expert Opin. Drug Saf. 14, 957-964 (2015).
Cao, . et al. Granzyme and perforin are important for regulatory cell-mediated suppression of tumor clearance. Immunity 27, 635-646 (2007).
Wei, S. C. et al. Distinct cellular mechanisms underlie anti-CTLA-4 and anti-PD-1 checkpoint blockade. Cell 170, 1120-1133.e17 (2017).
Seidel, J. A., Otsuka, A. & Kabashima, K. Anti-PD-1 and anti-CTLA-4 therapies in cancer: mechanisms of action, efficacy, and limitations. Front Oncol. 8, 86 (2018).
Dammeijer, F. et al. The PD-1/PD-L1-checkpoint restrains T cell immunity in tumor-draining lymph nodes. Cancer Cell 38, 685-700.e8 (2020).
Jiang, W. et al. Exhausted CD8+T cells in the tumor immune microenvironment: new pathways to therapy. Front Immunol. 11, 622509 (2021).
Pielenhofer, J., Sohl, J., Windbergs, M., Langguth, P. & Radsak, M. P. Current progress in particle-based systems for transdermal vaccine delivery. Front. Immunol. 11, 266 (2020).
Corbett, K. S. et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 586, 567-571 (2020).
Elia, U. et al. Design of SARS-CoV-2 hFc-Conjugated ReceptorBinding Domain mRNA Vaccine Delivered via Lipid Nanoparticles. ACS Nano 15, 9627-9637 (2021).
Zhang, N.-N. et al. A thermostable mRNA vaccine against COVID-19. Cell 182, 1271-1283.e16 (2020).
Chakraborty, C., Bhattacharya, M. & Dhama, K. SARS-CoV-2 vaccines, vaccine development technologies, and significant efforts in vaccine development during the pandemic: the lessons learned might help to fight against the next pandemic. Vaccines 11, 682 (2023).
Cheng, Y. & Xu, F. Anticancer function of polyinosinic-polycytidylic acid. Cancer Biol. Ther. 10, 1219-1223 (2010).
Rydyznski Moderbacher, C. et al. Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell 183, 996-1012.e19 (2020).
Shi, J., Li, Y., Chang, W., Zhang, X. & Wang, F.-S. Current progress in host innate and adaptive immunity against hepatitis C virus infection. Hepatol. Int 11, 374-383 (2017).
Rahe, M. & Murtaugh, M. Mechanisms of adaptive immunity to porcine reproductive and respiratory syndrome virus. Viruses 9, 148 (2017).
Schudel, A., Francis, D. M. & Thomas, S. N. Material design for lymph node drug delivery. Nat. Rev. Mater. 4, 415-428 (2019).
Davies, M. & Duffield, E. A. Safety of checkpoint inhibitors for cancer treatment: strategies for patient monitoring and management of immune-mediated adverse events. Immunotargets Ther. 6, 51-71 (2017).
شكر وتقدير
تم دعم هذه المقالة جزئيًا من قبل البرامج الوطنية للبحث في الصين (رقم 2021YFF0701800، 2020YFA0211100)، والعلوم الطبيعية الوطنية
مؤسسة العلوم في الصين (الأرقام: T2321005، 52032008)، مشروع مؤسسة ما بعد الدكتوراه في الصين (رقم: 2023M732543)، صندوق تطوير العلوم والتكنولوجيا (FDCT 0002/2022/AKP)، مشروع البحث الطارئ حول COVID-19 من جامعة تشجيانغ، المركز الوطني للابتكار التكنولوجي في الأدوية الحيوية، مركز الابتكار التعاوني لعلوم وتكنولوجيا النانو في سوتشو، وبرنامج 111 من وزارة التعليم في الصين. تم دعم Z Liu من قبل مؤسسة نيو كورنرستون للعلوم من خلال برنامج الباحثين في نيو كورنرستون وجائزة XPLORER.
مساهمات المؤلفين
وِنْجُون زُو، وْشِيَان شِين، وزُوانغ ليو صَمَّموا وأَخْرَجوا التجارب. وِنْجُون زُو وتِينغ وي صَمَّموا واصطَنَعوا المواد والمركبات النانوية. وِنْجُون زُو وْكِيُوتُونغ جِين أَجْرَوْا التجارب في المختبر. وِنْجُون زُو، ويُوتشُون شُو ويو تشاو أَجْرَوْا التجارب في الجسم الحي على الميلانوما. وِنْجُون زُو، جِيَاقِي لُو، وجُون شُو أَجْرَوْا التجارب في الجسم الحي على اللقاحات. وِنْجُون زُو، جِيَافِي زُو ومُوتشَاو تشين أَجْرَوْا تجارب تحليل تدفق الخلايا. وِنْجُون زُو، ويُوتشُون شُو، ويَشَاو يِين أَجْرَوْا جميع التجارب المعدلة. وِنْجُون زُو، وْشِيَان شِين، وزُوانغ ليو حَلَّلوا جميع البيانات وكتبوا الورقة. جميع المؤلفين ناقشوا النتائج التجريبية وحرروا الورقة.
Transdermal drug delivery has been regarded as an alternative to oral delivery and subcutaneous injection. However, needleless transdermal delivery of biomacromolecules remains a challenge. Herein, a transdermal delivery platform based on biocompatible fluorocarbon modified chitosan (FCS) is developed to achieve highly efficient non-invasive delivery of biomacromolecules including antibodies and antigens. The formed nanocomplexes exhibits effective transdermal penetration ability via both intercellular and transappendageal routes. Non-invasive transdermal delivery of immune checkpoint blockade antibodies induces stronger immune responses for melanoma in female mice and reduces systemic toxicity compared to intravenous injection. Moreover, transdermal delivery of a SARS-CoV-2 vaccine in female mice results in comparable humoral immunity as well as improved cellular immunity and immune memory compared to that achieved with subcutaneous vaccine injection. Additionally, FCS-based protein delivery systems demonstrate transdermal ability for rabbit and porcine skins. Thus, FCS-based transdermal delivery systems may provide a compelling opportunity to overcome the skin barrier for efficient transdermal delivery of bio-therapeutics.
Transdermal administration refers to needleless drug delivery across the skin without physical damages . It has been regarded as an attractive alternative to oral delivery or subcutaneous injection of drugs, due to its unique advantages including non-invasiveness, avoidance of the firstpass effect, painless administration, better patient compliance, avoidance of needle phobia and so . Although a variety of transdermal enhancers have been proven to be effective in clinics, the delivered payloads are greatly limited to drugs that have molecular masses around a few hundred Daltons and exhibit strong hydrophobicity . Nowadays, it is still difficult to realize efficient transdermal delivery of hydrophilic
biomacromolecules such as peptides, proteins, or nucleic acids . Besides, the delivery of vaccines is currently one of the hottest research areas in both clinical and scientific communities considering the Coronavirus Disease 2019 (COVID-19) epidemic . Compared to conventional subcutaneous injection or intramuscular injection, transdermal delivery of vaccines may be an attractive approach due to its possibility in athome administration and the existence of abundant immune cells in the skin .
To realize transdermal delivery of biomacromolecule drugs, especially proteins, novel chemical enhancers such as membrane
penetrating peptides, as well as various physical enhancement devices, including cavitational ultrasound, electroporation, thermal ablation, microdermabrasion, and microneedles, have been developed . Although such strategies could be used for transdermal delivery of various macromolecules, including therapeutic proteins, they still face several concerns. For example, the membrane penetrating peptides have been reported to enable transdermal delivery of small proteins such as insulin , but with unsatisfactory delivery efficiency , and remain to be ineffective for proteins with large molecular weights. Meanwhile, physical enhancement devices such as electroporation and sonophoresis not only can hardly be self-operated, but also could lead to skin damage by high energy pulses . Microneedles, which refer to patches with many small needles, have been widely applied in transdermal delivery, showing great potential to deliver insulin and influenza vaccines in recent years . However, the manufacturing process and quality control of microneedle patches, especially with biomacromolecular payloads, would be complicated. Additionally, the microneedles could still induce certain skin damage, which might increase the risk of infections. Some non-invasive platforms such as ionic liquids (ILs) and hyaluronic acids (HAs) were also reported recently to open tight junctions in the stratum corneum and promote paracellular transport . However, they still showed less transdermal efficacy. In several previous studies, lipid nanocarriers such as ethosomes were also reported for the transdermal delivery of proteins against skin tumors . Nevertheless, it would still be appealing to develop novel enhancers with high safety and efficiency for transdermal delivery of proteins.
Chitosan (CS) is a biodegradable natural cationic polymer with antibiotic activity and mucoadhesive property . Inspired by the efficient transmucosal efficacy of fluorocarbon-modified CS (FCS), as reported in our previous study for intravesical-instillation-based bladder cancer treatment , we speculated that FCS might also be employed for transdermal delivery of biomacromolecules (Fig. 1a).
Herein, we discover that FCS could self-assemble with biomacromolecules such as proteins to form nanocomplexes, which could be added into Aquaphor as an ointment formulation for topical applications with greatly enhanced transdermal penetration ability. We then employ such FCS-based transdermal delivery platforms for tumor immunotherapy and SARS-CoV-2 vaccines. With the help of FCS, non-invasive transdermal delivery of anti-programmed death-ligand 1(aPDL1) antibody could effectively inhibit the growth of local tumors with direct contact with the FCS/aPDL1-containing ointment. While combined with the co-delivery of anti-cytotoxic T-lymphocyteassociated protein 4 (aCTLA4), such FCS/aPDL1/aCLTA4 ointment could induce strong systemic immune responses to suppress both local and abscopal distant tumors. With significantly enhanced therapeutic responses compared to systemic injection of antibodies at the same dose, our FCS-based local delivery of immune checkpoint antibodies to treat melanoma may lead to fewer concerns in systemic side effects considering the relatively low serum concentrations by the topical administration route. Furthermore, in a proof-of-concept experiment, we verify that FCS could form nanocomplexes with the S1 protein of SARS-CoV-2 as the antigen and polyinosinic: polycytidylic acid (PolyIC), a ligand for toll-like receptor (TLR) 3 as the adjuvant. Topical application of FCS/S1/polyIC nanocomplexes could trigger S1-protein-specific immune responses, reaching a level comparable to that achieved by subcutaneous injection of the same nanocomplex. We further preliminarily demonstrate that FCS-based transdermal delivery may be applicable for skins of larger animals such as rabbits and pigs. Therefore, FCS developed in this work represents a rather effective carrier for transdermal delivery of biomacromolecules, offering possibilities for a wide range of applications such as localized melanoma immunotherapy and self-administrated transdermal vaccines against viruses (e.g., SARS-CoV-2).
Results
Preparation of FCS/protein nanocomplexes and ex vivo evaluation of their transdermal abilities
FCS was synthesized following a previous report . Briefly, perfluoroalkyl carboxylic acid (PFCA) was grafted to cationic polysaccharide CS through amide coupling at a fluorocarbon substitution of (Fig. S1). Then, FCS was mixed proteins such as immunoglobulin G (IgG) and ovalbumin (OVA) at different mass ratios for 30 min under mild shaking to form nanocomplexes (Fig. 1a). As shown in Fig. 1b, d, both FCS/IgG and FCS/OVA with mass ratio at showed sizes around 200 nm in the transmission electron microscopy (TEM) images, consistent with their hydrodynamic diameters measured by dynamic laser light scattering (DLS) (Fig. 1c, e). The zeta potentials (ZP) of both FCS/IgG and FCS/OVA showed high positive charges, which increased from 6.97 to 30.53 mV and 4.38 to 36.73 mV , respectively, as the increase of FCS contents during the formation of nanocomplexes (Fig. 1e). Dynamic light scatter (DLS) measurement (Fig. S2) showed a single peak at for FCS/IgG nano-complexes, which were much larger than the sizes of free , indicating that the majority of should have been encapsulated by FCS. Then, the circular dichroism (CD) spectra were used to verify the structure of proteins before and after forming nanocomplexes. As shown in Fig. 1f, FCS/IgG showed similar CD spectrum characteristic peaks at around 202, 206, and 216 nm to that of free IgG, indicating that the structure of protein remained nearly unchanged during the formation of such FCScontaining nanocomplexes. The intensity difference on 200 nm might result from the solvent. A similar result also was found in the comparison between FCS/OVA and free OVA in Fig. 1g. Additionally, the antibody affinity of aPDL1 in the formulation of FCS/aPDL1 remained nearly unchanged as measured by the competition-enzymelinked immunosorbent assay (ELISA) (Fig. 1h), further demonstrating that the formation of nanocomplexes wound not affect the activity of contained proteins.
Then, the transdermal kinetics and related mechanisms were investigated. Firstly, the standard Franz diffusion system was used to measure the transdermal delivery efficiency of FCS-containing nanocomplexes across the mouse skin layer. Briefly, fresh skin tissues were fixed between two glass cells, and then FCS/IgG or FCS/OVA, in which IgG and OVA were labeled with fluorescein (FITC), were added into the donor chamber in phosphate-buffered saline (PBS) solutions. The transmitted IgG or OVA was measured by collecting liquid samples in the receptor chamber at different time points to measure the FITC fluorescence (Fig. 1i). All skin resistances were measured pre and post experiment. Skin was regarded as unbroken by the manufactory when its resistance was three times larger than the solvents. In order to avoid destroying the mouse skins, the fat layer was not removed in the mouse skin model. As shown in Figure S3, all skins showed no obvious change in resistance in 12 h . Skins were washed and pyrolyzed to measure the transdermal retention in the dermis region. Transdermal delivery efficiencies of FCS/protein nanocomplexes with different feeding ratios ( ) were measured at different time points. As shown in Fig. 1i, k, FCS/IgG with the mass ratio at 1:1 showed the highest penetration ability across the skin with the total amount (the sum of permeation and dermis retention) up to about (six times larger than the free group), which may be attributed to the fact that FCS/IgG prepared at 1:1 showed the smallest sizes compared with those prepared at other mass ratios. It could be clearly observed that the zeta potentials of FCS/IgG nanocomplexes increased along with increasing the ratio of FCS, which may be beneficial for skin penetration. However, a further increase of FCS might lead to the aggregation of nanocomplexes. On the other hand, when the amount of IgG increased, the zeta potential of FCS/IgG nanocomplex turned out to be less positive with aggregation, which is also unsuitable for skin penetration. Similarly, FCS/OVA with the mass ratio at showed the highest penetration ability with a total penetration (the sum of
a
b
c
FCS/IgG
Size (nm)
了 Potential (mV)
e
FCS/OVA
Size
Potential
f
g
h
i
n
Cy5.5-OVA
FITC-FCS
Merged
Fig. 1| The characterization of FCS-containing nanocomplexes. a The schematic image of FCS-containing nanocomplexes for transdermal delivery. b Representative TEM images of FCS/IgG and d FCS/OVA. c, e Size distribution and zeta potential of FCS-containing nanocomplexes including c FCS/IgG and e FCS/ OVA ( ). Circular Dichroism (CD) spectra of IgG and OVA pre and post FCS coating. The relative binding affinity of aPDL1 with or without FCS measured by the standard indirect ELISA (iELISA) assay ( ). i Schematic illustration of Franz diffusion cell system used for the skin permeation study. Cumulative
permeation and retention of FCS/IgG-FITC and FCS/OVA-FITC permeated across the mouse skin after incubation with different FCS-containing formulations over time ( for IgG and for OVA). Total dosage: .
n Representative confocal images of mice skin treated with FITC-FCS/OVA-Cy5.5 for . Scale bar: . All illustrations were created with BioRender.com. Data are presented as mean standard deviation. Source data are provided as a Source Data file.
permeation and dermis retention) up to about ( 11 times larger than the free group, Fig. 11, m). The difference in the penetration behaviors of the two FCS-based nano-complexes might be due to the different physical and chemical properties of the proteins, such as the molecular weight or isoelectric point. Afterward, the skins in the Franz diffusion system post transdermal delivery were sliced for confocal microscopy with FITC-FCS/OVA-Cy5.5 to further evaluate the penetration of FCS nanocomplexes. As shown in Fig. 1n, the colocalization of FCS/OVA was observed in the dermis region of the skin, demonstrating that FCS/OVA could penetrate into the skin dermis in the nanocomplex forms. In contrast, no obvious skin penetration was observed in the free OVA group. Collectively, FCS-based nanocomplexes could successfully deliver proteins into the dermis region of mouse skins.
Transdermal mechanism of FCS-containing nanocomplexes
Next, we investigated the underlying transdermal mechanism of such FCS-containing nanocomplexes. Firstly, the cytotoxicity of FCS nanocomplexes was measured. As shown in Figs. S4 and S5, no obvious cytotoxicity of FCS and CS was observed for HACAT cells. According to previous literature, there are three classical permeation routes for transdermal delivery, including intercellular, transappendageal, and transcellular routes . Firstly, we studied the intercellular route, by which the nanocomplexes could enlarge the space between epidermis cells and thus pass through them . It has been reported that chitosan and its derivatives could cross epithelial cells by intercellular route . During the enlargement of intercellular space, we would expect changes in cell resistance, as well as the expression and allocation of related proteins . In this case, human skin epidermis cells HACAT were used to form a cell monolayer and the transepithelial electrical resistance (TEER) between the two sides of the monolayer was monitored (Fig. 2a). As shown in Fig. 2b, the cell single layer was formed 6 days later with stable and high TEER. Interestingly, an obvious decrease of TEER was observed with the addition of FCS/IgG on day 11, indicating the destruction of the cell monolayer and opening of intercellular channels after adding FCS-containing nanocomplexes. More interestingly, the re-increase of TEER was observed 4 h later after the removal of FCS/IgG, and returned to its original level in 12 h , demonstrating that FCS/IgG only temporarily opened the intercellular channel. TEM imaging of the skin also revealed the opening of the tight junctions and enlarged intercellular spaces after topical treatment of the skin with FCS/IgG, in comparison with the normal skin (Figs. 2g and S6). To confirm the enlargement of intercellar spaces, the changes of tight junctions (TJs) related proteins such as zonula occludin (ZO)-1 were further evaluated . With the addition of FCS/IgG, while the total expression of ZO-1 remained nearly unchanged, their continuous distribution was distinctly disturbed, indicating the opening of tight junctions along the interface between cells (Fig. 2c, d). Moreover, the phosphorylation of myosin light chain (MLC), an important parameter for cytoskeletal structure , was found to be up-regulated in FCS/IgG treated cells, demonstrating that FCS was able to promote the phosphorylation of myosin light chain to induce the contraction of actin and the rearrangement of the cytoskeleton (Fig. 2e, f).
In addition to the enhanced intercellular bypass permeability of FCS-containing nanocomplexes, the transappendageal pathway, which usually plays an important role in the transport of large and watersoluble drugs through the hair follicles, sweat glands, and sebaceous glands, was also investigated in our experiments . With the counterstaining of Keratin (Krt) 14, the hair follicles and sweat glands in the deep dermis region were labeled. As shown in Fig. 2g (white arrows), it was observed that FCS/IgG colocalized with hair follicles and sweat glands, indicating that the transappendageal pathway also played an important role in FCS-based transdermal delivery systems.
We further studied the transcellular route, which signifies the passage of drugs directly across keratinocytes . Different from normal
tissue cells, which decompose drugs in lysosomes, polarized cells such as keratinocytes sometimes might expel drugs through exocytosis . Apical exocytosis was used to investigate this phenomenon in vitro. Briefly, HACAT cells were incubated with FCS/IgG-FITC for endocytosis. After 12 h incubation, FCS/IgG-FITC was washed away, and the cells were incubated for another 12 h . Then, the fluorescence of FITC in the supernatant was measured to evaluate the apical exocytosis. As shown in Fig. S7, the apical exocytosis rate of FCS/IgG-FITC from HACAT cells was less than , almost the same compared with that treated with free IgG-FITC, indicating negligible transcellular route was involved. Besides, further evaluation was conducted by the Franz diffusion system. As reported, exocytosis was usually mediated by clathrin. Therefore, skins were treated with chlorpromazine hydrochloride, the inhibitor for clathrin, for 2 h to compare the penetration ability with or without the clathrin inhibitor. As shown in Fig. S8, the skin retention of FCS/IgG-FITC also showed no clear influence. In conclusion, the transdermal delivery of FCS-containing nanocomplexes mainly relied on the intercellular and transappendageal pathways (Fig. 2i).
Conclusively, FCS, as the derivative of chitosan, could also enlarge cellular space by stimulating the phosphorylation of MLC, a phenomenon also observed for unmodified chitosan . The phosphorylation of MLC could then lead to the rearrangement of cytoskeletal structure, transforming tight junction-related protein into the cytoplasm. Afterward, the tight junction between epithelial cells would be opened, and the intercellular space would be enlarged to allow the permeation of our nanocomplexes. On the other side, the unique non-hydrophobic non-hydrophilic properties of fluorocarbon chains would make FCS less sticky when penetrating through various biological barriers . Therefore, FCS-based nanocomplexes also showed enhanced permeation via hair follicles through the transappendageal pathway.
Topical application of FCS/antibody nanocomplexes for melanoma treatment
Melanoma, as one of the most common malignant tumors, especially among Caucasians, develops in the melanocytes located in the bottom layer of the skin’s epidermis . For melanoma treatment, immunotherapies, especially immune checkpoint blockade using antiprogrammed death-1/its ligand (aPD1/aPDL1) antibodies, have been widely used in clinic . Despite the exciting therapeutic result of using aPD1/aPDL1 antibodies for the treatment of melanoma, there are still many limitations, such as the risk of autoimmune diseases after intravenous injection . Considering that FCS could act as the efficient transdermal delivery carrier for proteins, we thus used it as the transdermal delivery platform to deliver aPDL1 antibody for melanoma treatment (Fig. 3a). It was expected that the transmitted aPDL1 could block the PD1/PDL1 pathway to stimulate cytotoxic T cells and lead to remarkable inhibition of tumors.
For in vivo administration of our FCS-based nanocomplexes, blank ointment (Aquaphor , mainly petrolatum) was mixed with the solution to keep moisture for a long time. The transparent blank ointment would turn into milk-like ointment when mixed with solutions. The milk-like ointment could be absorbed in 12 h after topical administration for full transdermal delivery. Therefore, the administration was applied only for 12 h for in vivo treatment for a standard applied dosage. To examine the topical penetration behavior of antibodies complexed with FCS, tumors from the mice swiped with ointment containing radioisotope labeled IgG ( ) or FCS/ were collected for quantification of transmitted antibodies at different time points (Fig. S9). As shown in Fig. 3b, compared with free I-IgG in the transdermally applied ointment, showed dramatically higher accumulation in the tumor, while the radioactivities in other organs appeared to be much lower. Meanwhile, as shown in Fig. 3c, we found that the tumor accumulation of FCS/IgG showed the peaked level at over of injected dose per gram tissue (%ID/g) at 12 h post
the ointment was applied (about ID in the total tumor in Figure S 10 ). As the ointment was removed at 12 h , the IgG level in the tumor decreased slightly at 24 h . Meanwhile, the ELISA assay was also conducted to detect the tumor accumulation of IgG in the FCS/IgG formulation topically applied to tumors in the ointment. As shown in Fig. S11, the ELISA assay showed similar results with the radiolabelingbased biodistribution data that IgG could be efficiently delivered into
the tumor within 12 h with the aid of transdermal delivery platform using FCS/IgG nanocomplexes. On the other hand, the tumors with FCS/IgG-Cy5.5 were imbedded for tumor slicing. As illustrated in Fig. 3d, the fluorescent signals of IgG-Cy5.5 in the tumor were gradually increased and evenly disbursed inside the whole tumor within 12 h , demonstrating the continuous transmission of antibodies from the FCS/IgG-Cy5.5 ointment into the tumor. Therefore, the accumulation
Fig. 2 | The transdermal mechanism of FCS-containing nanocomplexes. a Illustration of the HACAT monolayer cell model. Effects of FCS/IgG on the TEER of the HACAT monolayer cell model ( , each TEER was tested 3 times). c Immunofluorescence images of the distribution of tight junction-related protein ZO-1 on the HACAT cell membrane after being treated with FCS/IgG ( ). The white arrows indicated the allocation change of ZO-1. Scale bar: Western blotting images showing ZO-1 ( ) and the phosphorylated level of MLC ( MLC, ) in cells after incubation with FCS/IgG. The raw figures were provided in Figs. S27 and S28. f The graphical representations of the relative intensity of MLC/ pMLC with the addition of FCS/IgG ( ). g Representative TEM image of skin epithelium after being treated with FCS/IgG. The white arrows indicated the tight
junctions (TJs) and the opening of TJs ( ). h Representative immunofluorescence images exhibiting the colocalization of keratin 14 and FCS/IgG-Cy5.5 (white arrows, ). Scale bar: . i The schematic image of the transdermal mechanisms. FCS-containing nanocomplexes could penetrate the skin epidermis through both paracellular and transappendageal routes. By the paracellular route, FCS could stimulate the phosphorylation of MLC and thus open the tight junction between epidermis cells by sealing strands of tight junction proteins. By the transappendageal route, FCS-containing nanocomplexes could cross the epidermis through hair follicles and sweat glands. All illustrations were created with BioRender.com. Data are presented as mean standard deviation. Source data are provided as a Source Data file.
of antibody drugs such as aPDL1 in the tumor was evaluated. As shown in Fig. S12, FCS/aPDL1 showed obviously higher tumor accumulation after topical application compared with free and CS/aPDL1. The skin irritation was also evaluated for long-term administration. As shown in Fig. S13, both the film group and the FCS/IgG with film group showed similar skin conditions after topical treatment three times (once every two days), suggesting the safe topical use of FCS.
Inspired by the effective accumulation of IgG in the tumor after transdermal delivery, we then carried out in vivo treatment for melanoma tumors by transdermal delivery of FCS/aPDL1. Mice-bearing melanoma tumors were randomly divided into four groups: (i) Untreated, (ii) Free aPDL1 by intravenous (i.v.) injection, (iii) CS/PDL1 in the ointment by transdermal delivery, and (iv) FCS/aPDL1 in the ointment by transdermal delivery. For transdermal delivery, CS/aPDL1 or FCS/aPDL1 solution was mixed with blank ointment and then applied to the tumor, which was subsequently covered with a transparent film. Such treatment was repeated every 2 days three times at the dose of aPDL1 per mouse each time. For i.v. injection, aPDL1 was administrated into each mouse every 2 days three times. As shown in Fig. 3e, the tumors in the FCS/aPDL1 treated group were successfully inhibited in a short time after the third treatment, while both the CS/ aPDL1 group and the free aPDL1 group showed negligible anti-tumor efficacy. During 20 days of observation, the FCS/aPDL1 treated group exhibited the lowest tumor growth rate and the longest survival (Fig. S14).
To understand the immune activation mechanisms of such transdermally delivered immunotherapy, tumors were collected from different groups on day 12 to investigate different types of immune cells, especially T cells, by flow cytometry. As shown in Fig. 3f, for the FCS/aPDL1 treated group, the percentages of both and T cells showed an obvious increase in the tumor, indicating that aPDL1 was successfully delivered into the tumor to revert the cell exhaustion. As Granzyme B is important for cell programming death triggered by cells, Ki 67 is cell proliferation, and Interferon is a cytokine associated with pro-apoptotic and antitumor mechanisms , we used these three markers to analyze the activities of CD8+ cells. As can be seen in Fig. 3g-i, all of the three markers in cells was increased in the FCS/aPDL1 group, suggesting the effective infiltration and activation of cytotoxic T lymphocytes (CTLs) in those tumors. For i.v. injection of aPDL1 and transdermal delivery of CS/aPDL1 groups, the tumor infiltrations of both and cells, as well as the expressions of granzyme , and , remained nearly unchanged, which might be resulted from the low tumor accumulation of aPDL1 after intravenous injection or transdermal delivery using CS. These results clearly indicated that, compared to i.v. injection of aPDL1, transdermal delivery of aPDL1 using FCS resulted in enhanced antitumor immune responses.
Our platform could also be utilized to co-deliver different therapeutic proteins. In addition to aPDL1, anti-cytotoxic T-lymphocyteassociated protein 4 (aCTLA4) is another important antibody for immune checkpoint blockade to disable regulatory T cells (Tregs) and promote effective T-cell activation . Thus, we further investigated the combination therapy using aPDL1 and aCTLA4 co-delivered by the
transdermal delivery carrier FCS (Fig. 4a). In this experiment, B16F10 cancer cells were inoculated on the right flank of each mouse as the primary tumor, and a second tumor was inoculated on the opposite side of the same mouse to mimic the cancer metastasis. Three days later, mice bearing two melanoma tumors were randomly divided into five groups: (i) Untreated, (ii) Free aPDL1 and aCTLA4 (i.v.), (iii) FCS/ aPDL1, (iv) FCS/aCTLA4, (v) FCS/aPDL1/aCTLA4. All the groups were administrated three times with aPDL1 and aCTLA4 per mouse each time. It is worth pointing out that after the second i.v. injection of aPDL1 + aCTLA4, half of the mice in this group died, likely due to the horrible side effects (e.g., cytokine storm) triggered by systemic administration of both aPDL1 and aCTLA4. In contrast, the other groups with transdermal delivery of immune checkpoint blockade antibodies showed negligible abnormality. Excitingly, compared to FCS/aPDL1 or FCS/aCTLA4 treated mice, transdermally delivered FCS/aPDL1/aCTLA4 showed further improved therapeutic performance (Fig. 4b, c). More interestingly, the growth of distant tumors in mice with FCS/aPDL1/aCTLA4 treatment also was inhibited (Fig. 4d, e). To understand the abscopal effect induced by FCS/aPDL1/aCTLA4, tumors were collected 12 days after different treatments and analyzed by flow cytometry. As shown in Fig. S19, in the primary tumor, the percentage of CD4+ T cells showed no obvious increase, while that of regulatory cells was decreased in the primary tumor, which could be the result of the succuss transdermal delivery of aCTLA4. On the other hand, the percentages of both cells and cells were increased, illustrating the enhanced tumor infiltration of cytotoxic lymphocytes (CTLs) (Fig. S20). Consistent with the above results, in the distant tumor, the number of cells, especially cells, exhibited an obvious increase in the second tumor (Fig. 4f, i), and the percentages of Tregs were decreased in FCS/aPDL1/aCTLA4 treated group (Fig. 4j).
The excellent anti-tumor efficacy and the increased CTLs in the distant tumor might be attributed to the following mechanisms. Firstly, the activation of CTLs in the local tumor could induce immunogenic death of tumor cells and trigger the chronic exposure of damage-associated molecular patterns (DAMPs) as well as tumor antigens. Antigen-presenting cells (APCs) are then activated and subsequently present tumor antigens to cells, further amplifying systemic antitumor immunity to attack distant tumors. Lastly, as reported, the blockade of PDL1 in the tumor-draining lymph nodes (TDLNs), which may be more efficient to be reached by transdermal delivery, could effectively propel systemic anti-tumor cell immunity even in distant tumor sites . Therefore, our transdermal immune checkpoint antibody delivery could achieve remarkable antitumor efficacies against both local and distant tumors.
Topical application of FCS/S1/polyIC nanocomplexes for transdermal vaccination
Vaccination is another area of great interest for transdermal delivery . For the transdermal vaccine, in addition to avoiding syringes by health professionals, it could improve immune responses by targeting abundant immune cells beneath the epidermis layer. Since the outbreak of Coronavirus Disease 2019 (COVID-19), various vaccines have
Fig. 3 | Transdermal delivery of aPDL1 for the treatment of B16F10 melanoma tumors. a Schematic illusions illustrating the localized transdermal administration of FCS/aPDL1 for the treatment of B16F10 melanoma tumors. The accumulation of FCS/ I-IgG in the tumor at different time intervals ( ). c Biodistribution of FCS I-IgG at 12 h based on radioactivity measurement. The total accumulation and biodistribution analysis was illustrated in Fig. S10 ( ). d Representative confocal images showing the accumulation of FCS/IgG-Cy5.5 in the tumor at different time intervals ( ). Scale bar: . e Tumor growth curves of mice in different groups ( ). Growth curves were stopped when the first mouse in the related group was dead, or its tumor size exceeded . f Quantification of
cells and cells in the tumor after different treatments ( ). The representative flow cytometric plots were illustrated in Fig. S15. g-i Quantification of granzyme Granzyme , and IFN- ( IFN ) T cells in the tumor after different treatments ( ). The representative flow cytometric plots were illustrated in Figs. S16-S18. All illustrations were created with BioRender.com. Data are presented as mean standard deviation. Statistical significance was calculated via one-way ANOVA with a Tukey post-hoc test. . Source data are provided as a Source Data file.
been developed to suppress the infection of SARS-CoV-2 . Up to now, 242 SARS-CoV-2 vaccine candidates are in clinical development, and 11 COVID-19 vaccines have been granted an Emergency Use Listing (EUL) by the WHO . However, most of these vaccines need a subcutaneous or intramuscular injection, which not only requires welltrained medical staff but also faces some additional issues, such as the disposal of a large number of sterile syringes. The transdermal SARS-CoV-2 vaccine, on the other hand, may offer great assistance in
preventing COVID-19 by self-administration, especially in areas with tight medical resources. Considering the rapid global spread of COVID19 and the efficient transdermal delivery of antibodies using FCS, we further explored whether FCS could be utilized to transdermally deliver the SARS-CoV-2 vaccine in a proof-of-concept study.
To synthesize FCS-based subunit SARS-CoV-2 vaccine, FCS were mixed with the S1 subunit of spike protein of SARS-CoV-2 virus and polyIC, forming the transdermal SARS-CoV-2 vaccine (FCS/S1/polyIC)
Fig. 4 | The abscopal effect induced by transdermal delivery of combined immune checkpoint antibodies. a Schematic illustration of transdermal codelivery of aPDL1 and aCTLA4 to inhibit the growth of both primary and distant tumors. b-e Tumor growth curves of primary and distant tumors after different treatments ( ). Growth curves were stopped when the first mouse in the related group was dead, or the first mouse’s tumor size exceeded . Representative flow cytometry plots ( f ) and the related quantification of
T cells and T cells in distant tumors after different treatments ( ). Quantification of cells (i) and Tregs (CD3 Foxp3 ) (j) in distant tumors after different treatments ( ). The representative flow cytometric plots were illustrated in Fig. S21. Data are presented as mean standard deviation. Statistical significance was calculated via one-way ANOVA with a Tukey post-hoc test. . Source data are provided as a Source Data file.
(Fig. 5a). Note that poly IC as a double-strand RNA is a toll-like receptor 3 (TLR 3) agonist that has been commonly used as immune adjuvant . We also optimized the ratio of FCS: antigen: PolyIC in the FCS/S1/ PolyIC vaccine. In this experiment, OVA was used as a modulated antigen for the optimization of the formulation. As shown in Fig. 5c, d, FCS/OVA/polyIC with different mass ratios showed variant sizes at about 200 nm and increased zeta potential with the increase of FCS.
For the skin penetration ability measured by the Franz diffusion system in Fig. 5e, f, FCS/OVA/PolyIC prepared at the mass ratio of 2:1:1 showed the highest skin permeability. This might result from the proper size and zeta potential since nanoparticles prepared at the ratio of 1:1:1 showed dramatically negative charges, while those prepared at the ratio of 3:1:1 showed much larger sizes. Therefore, the formulation with FCS/OVA/PolyIC ratio at 2:1:1 was used for further studies.
Fig. 5 | Transdermal delivery of SARS-CoV-2 vaccine. a A schematic illustration for transdermal delivery of the SARS-CoV-2 vaccine and the triggered immune responses. After transdermal delivery, such SARS-CoV-2 nano-vaccines could activate immune cells such as DCs in the dermis, or migrate to the nearby lymph nodes for immune activation. b Schematic illustration of the experimental design showing transdermal delivery of SARS-CoV-2 vaccine. c, d DLS (c) and zeta potential (d) of FCS-based transdermal vaccines with different mass ratios from 1:1:1 to 3:1:1 ( ). e, The skin penetration ability of FCS-based transdermal vaccine with different mass ratios ( ). Total dosage: (g) SARS-CoV-2 specific IgG antibody titers at different time intervals determined by ELISA ( ). h Quantification of cells, cells in the spleen at day Quantification of IFN- secreting cells ( IFN ) and cells ( IFN ) in the spleen at day Quantification of OVA-Cy5.5 CD45 CD11c Cy5.5 in DCs in lymph nodes and skin ( ). Quantification of DC maturation ( ) and cell receptor (TCR) activation ( ) in lymph nodes ( ). All illustrations were created with BioRender.com. The representative flow cytometric plots were illustrated in Figs. S22 & S23. Data are presented as mean standard deviation. Statistical significance was calculated via one-way ANOVA with a Tukey post-hoc test. ; . Source data are provided as a Source Data file.
For the in vivo vaccination experiments, mice were randomly divided into three groups: i. untreated, ii. transdermal delivery of FCS/ S1/polyIC, and iii. subcutaneous (s.c.) injection of S1/polyIC. As shown in Fig. 5b, mice in the transdermal delivery group were administrated 3 times in 2 weeks (the doses of S1 and polyIC were both per time), while mice in s.c. injection group was injected twice with the S1 protein dose at per time and the polylC dose at per time. Interestingly, mice with transdermal delivery of FCS/S1/polyIC showed almost similar antibody titer to that of mice with s.c. injection of S1/ polyIC in two weeks, indicating that the transdermal delivery of FCS/ S1/polylC could result in almost the same humoral immunity compared with s.c. administrated vaccines (Fig. 5g). Furthermore, after boosting on day 14, the specific antibody titers in both the transdermal delivery group and s.c. injection group reached up to in 30 days, further indicating the effective activation of humoral immunity by those vaccines. After another boost on day 42, mice were vaccinated by either transdermal delivery of FCS/S1/polyIC or s.c. injection of S1/ polyIC showed remained at high levels of anti-S1 antibody titers.
In addition to the activation of humoral immunity by generating specific antibodies against the S1 protein, cell immunity also plays an important role in virus clearance by training cytotoxic T cells to recognize and kill virus-infected host cells . Therefore, the levels of cytotoxic T cells in the mouse spleen were evaluated on day 28 after the primeval administration. Although no obvious change of and cell infiltration was observed in different groups (Fig. 5h), the secretion of IFN- by cells and cells was obviously increased in mice after transdermal delivery of FCS/S1/PolyIC compared to that in mice by s.c. injection of S1/polyIC (Fig. 5i, j), indicating that stronger cytotoxic T-cell responses were induced by transdermal delivery of S1 vaccine. Moreover, the percentage of both effector memory and cells and ) in the spleen of mice treated with FCS/S1/ polylC was also dramatically increased on day 90 with a single boost on day 75 (Fig. S24), while those in mice with subcutaneous administration of S1/PolylC appeared to be much lower than those in the transdermal group. On day 75 (pre boost) and day 90 (15 days after a single boost), cytokines in the sera of mice after different administrations were measured (Fig. S25). The levels of IL-12p40, an important marker of innate immunity, and IFN- , the typical markers of cellular immunity, were all obviously higher in mice treated with FCS/S1/PolyIC, demonstrating that the transdermal delivery of vaccines would trigger long-term adaptive immune memory effect, which might be resulted from the long retention of co-delivered antigen and adjuvant.
In order to investigate the exact permeation ability of vaccines after transdermal administration, we measured the accumulation of antigens in the lymph node 24 h post transdermal delivery or subcutaneous injection of vaccine using Cy5.5-labeled OVA as the modal antigen. As expected, compared to s.c. injection of OVA, mice after transdermal delivery of vaccine after completely swiping the ointment off the skin surface showed lower OVA retention according to the IVIS imaging system (Fig. S26). Then, we further evaluated the uptake of OVA by DCs in lymph nodes and skin (Fig. 5k, l). Interestingly, even though the percentage of DCs in the lymph node of mice with s.c. the injection was higher than that of mice with transdermal delivery,
the DCs beneath skins showed higher OVA uptake in mice treated by the ointment containing FCS/OVA/polyIC, revealing that the transdermal delivery of vaccine could directly activate DCs in situ and the activated DCs could migrate to lymph nodes to trigger further immune responses. In contrast, for the s.c. injection group, antigen, and adjuvant would migrate to lymph nodes separately and be more likely to be depleted by non-specific immune cells and activate DCs with low effectiveness .
Additionally, we measured the maturation of DC and the activation of T cells on one and three days after different treatments. As shown in Fig. 5m, n, the mice with transdermal delivery of vaccines showed almost similar DC maturation and T cell activation to that of mice with s.c. injection, consistent with the observed cell activation in the spleen. Therefore, although FCS-based transdermal delivery of vaccine showed less absolute antigen penetration compared to that with s.c. injection, it could trigger almost similar humoral immunity and stronger cell immunity. The additional advantages of such FCSbased transdermal vaccines would be the possibility in selfadministration and preferable user compliance. However, further studies, such as the SARS-CoV-2 virus challenge, were not conducted due to our current limitations of experimental conditions in handling viruses.
Penetration ability of FCS-based nanocomplexes on rabbit and porcine models
Next, we further tested the ability of FCS for protein delivery through the skins of larger animals such as rabbits (Fig. 6a). Firstly, the Franz diffusion assay was conducted on rabbit skins in vitro. In this case, the subcutaneous fat layer was removed from rabbit skins for more accurate evaluation. The skin resistances were measured before and after the assay to confirm the skin integrity (Fig. S29). Compared to free IgG, FCS/IgG showed a significant increase in skin permeation for rabbit skins at an amount of about (the sum of permeation and dermis retention, Fig. 6b, c). Vaccination on rabbits was then conducted. Rabbits were randomly divided into three groups: (i) untreated, (ii) topical transdermal application of FCS/OVA/polyIC, and (iii) intramuscular (i.m.) injection of OVA/polyIC. As shown in Fig. 6d, both the transdermal and intramuscular groups were applied with the same dose of OVA and polyIC, following the same administration schedule on day 0 , day 14 , and day 30 . As shown in Fig. 6e, the antibody titer in rabbits of the transdermal vaccination group appeared to be close to that of the i.m. vaccination group, especially at later time points (e.g., days 30 and 60), demonstrating the successful FCS-based transdermal delivery of vaccines in rabbit model for effective vaccination.
Meanwhile, the ex vivo penetration was also conducted on porcine skins. Porcine skins were achieved from the back of little pigs at three months of age and all the subcutaneous fat layer was removed for more accurate evaluation. The skin resistances were measured before and after the assay to confirm the skin integrity (Fig. S30). Similarly, the Franz diffusion assay showed that FCS/IgG also achieved a transdermal ability up to (the sum of proteins penetrated through and retained inside the skin). The larger efficacy might result from the larger hair follicles and sweat glands on porcine. Additionally,
the confocal image of porcine skin slices also showed significant permeation of fluorescently labeled IgG in the dermis region of porcine skins topically applied with FCS/IgG-Cy5.5 (Fig. 6i, j). Collectively, FCS/ protein nanocomplexes showed successful transdermal delivery of proteins into the skin dermis and subcutaneous regions for mouse, rabbit, and porcine skins.
Discussion
In conclusion, we developed a transdermal delivery system based on FCS for efficient local delivery of biomacromolecules, including antibodies, antigens, or nucleic acids (e.g., polylC), which could be mixed with FCS and added into an ointment for topical applications. For transdermal delivery of antibodies such as aPDL1 and aCTLA4, our
Fig. 6 | Evaluation of transdermal protein ability on rabbit and porcine models. a Schematic illustration of in vivo vaccination on the rabbit model. b, c Cumulative percentages of penetration (b) and skin retention (c) of FCS/IgG-FITC permeated across the rabbit skin over time ( ). Total dosage: . d Schematic illustration of the experimental design showing transdermal delivery of OVA vaccination in a rabbit model. e OVA-specific IgG antibody titers in rabbit sera at different time intervals determined by ELISA ( ). Schematic illustration of ex vivo skin penetration on the porcine model. Cumulative percentages of
penetration (g) and skin retention (h) of FCS/IgG-FITC permeated across the porcine skin over time ( ). Total dosage: . i Representative confocal fluorescence images and statistical analysis to show the permeation depth FCS/ IgG-Cy5.5 through the porcine skin in 12 h . Free IgG-Cy5.5 was used as a control in those experiments ( ). Scale bar: . All illustrations were created with BioRender.com. Data are presented as mean standard deviation. Statistical significance was calculated via one-way ANOVA with a Tukey post-hoc test. ; . Source data are provided as a Source Data file.
FCS-based delivery systems could result in high local antibody accumulation in melanomas and rather strong T cell responses, and thus successfully eliminated primary tumors and inhibited the growth of distant tumors on mice. As reported, the co-delivery of aPDL1 and aCTLA4 by intravenous administration might result in a variety of side effects, including hepatotoxicity, immune-mediated pneumonitis, and even autoimmune endocrinopathies . With significantly enhanced therapeutic responses compared to systemic injection of antibodies at the same dose, our FCS-based local delivery of immune checkpoint antibodies may lead to fewer concerns in systemic side effects considering the relatively low serum concentrations by the topical administration route. On the other hand, transdermal delivery of SARS- vaccines resulted in S1-specific antibody titer similar to that achieved by s.c. injection, and even stronger T-cell responses. Such transdermal delivery of SARS-CoV-2 vaccines might enable fast and vast vaccination even at home once this strategy is proven to be effective in further studies. Such FCS-based transdermal protein delivery is found to be applicable to the skins of larger animals such as rabbits and pigs. Our work thus realized effective noninvasive needleless transdermal delivery of macromolecular therapeutics without the need for any additional physical or chemical stimulations, which has been rather challenging via existing techniques. In addition to delivering immune checkpoint antibodies and vaccines, the macromolecular transdermal delivery carrier developed in this work may also be employed for transdermal delivery of other therapeutic biomacromolecules aiming at diverse medical applications and holds tremendous potential for commercialization.
Methods
Ethical statement
All animal studies, including experiments in mice and rabbits, were conducted with the protocols approved by the Soochow University Laboratory Animal Center and the Soochow University Ethics Committee.
Materials, cell lines, and animal models
Chitosan (DD% ) was purchased from Aladdin Industrial Co. (Shanghai, China), N -(3-(Dimethylamino)propyl)-N-ethylcarbodiimide hydrochloride crystalline (EDC) and N-hydroxysuccinimide (NHS) were provided by JK Chemical Company (Beijing, China). Phosphatebuffered saline (PBS) was obtained from Beijing Solarbio Science Technology Co., Ltd. SARS-CoV-2 Spike protein was purchased from Sino Biological. Murine melanoma tumor cell line B16F10 cells were cultured under standard conditions recommended by the American Type Culture Collection (ATCC). Female C57BL/6 mice (6-8 weeks) were purchased from Nanjing Pengsheng Biological Technology Co. New Zealand Rabbits ( 3 kg ) were purchased from Suzhou Jinghu Biological Technology Co. Both mice and rabbit skins were collected in the lab. Porcine skins were purchased from Taizhou Taihe Biological Technology Co. All animal studies were conducted with the protocols approved by Soochow University Laboratory Animal Center.
Synthesis of fluorocarbon-modified chitosan
Perfluoroheptanoic acid ( ) was dissolved in dimethyl sulfoxide (DMSO, Sangon Biotech) and then mixed with EDC ( 78 mg ,
1.5 equiv) and NHS ( 47.43 mg , 1.5 equiv) under stirring in the dark at room temperature for 0.5 h to obtain activated perfluoroheptanoic acid. Then, CS ( 200 mg ) was dissolved in 1% acetic acid solution, mixed dropwise with activated perfluoroheptanoic acid, and stirred in the dark for 12 h to synthesize FCS. The obtained FCS was purified by dialyzing in double-distilled water using a dialysis bag (MWCO 3500 Da, Technologies Co.) for 48 h . Then, the samples were lyophilized, and the degree of substitution of conjugated fluoroalkyl substituents on each CS was characterized by ninhydrin colorimetry following the well-evidenced procedure and analyzed using the microplate reader (Variskan, THERMO). The 19 F NMR (Bruker AV III 600 spectrometers) of FCS ( 5.0 mg ) dissolved in of containing 1.0 mg of was also applied for the calculation of conjugated fluoroalkyl substituents.
Preparation of FCS-based nanocomplexes
The synthesized FCS was mixed with IgG in deionized water at different weight ratios (from 1:4 to 4:1) for 0.5 h to prepare FCS/IgG nanocomplexes. FCS/aPDL1, FCS/aCTLA4, FCS/OVA and FCS/S1 were prepared in the same way. The UV-visible absorbance spectra of various complexes were recorded by a PerkinElmer Lambda 750 ultraviolet-visible-NIR spectrophotometer (PerkinElmer, USA). The size, zeta-potential, and dispersibility index of the obtained nanocomplexes were detected by the Zetasizer Nano ZS (Malvern Instrument). Transmission electron microscopy (FEI TECNAI G2) was applied to observe the morphology of representative nanocomplexes.
Transdermal delivery efficiency
The transdermal delivery ability of FCS was investigated using the Franz diffusion cell system. The Franz diffusion system was purchased from Huanghai Ltd. Shanghai. Fresh mice, rabbits, and porcine back skins were collected and added between the donor and receptor chambers. Samples were collected from the receptor chamber at different time points and the same volume of PBS was added to the receptor chamber to keep the volume unchanged. The final permeation rate was calculated by the formula:
is the volume of the receptor chamber ( 8 mL ), is the volume of the samples collected at each time point ( 0.4 mL ), Cn is the concentration of FITC in the receptor chamber at each time point, is the concentration of the collected samples at the time point, and A is the feeding concentration of FITC. The feeding concentration of proteins is in PB solution ( ). The permeation skin area is .
Preparation of FCS-based transdermal ointment and in vivo administration
The synthesized FCS/IgG or other FCS/protein nanocomplexes were dropped onto the surface of blank ointment (Aquaphor , mainly petrolatum) with a mass ratio at 1:1, such as FCS/protein with 20 mg ointment. Then, they were gently mixed up with each other. The transparent blank ointment would turn into milk-like ointment, which
was then applied onto the mouse and rabbit skin for transdermal applications.
For mice administration, the mixed milk-like ointment was then swiped on the mouse skin with a round cover of in radius) for 12 h . Transparent film ( 3 M ) was covered finally to avoid the influence of mice lick.
For rabbit administration, the mixed milk-like ointment was then swiped on the rabbit skin with a round cover of in radius) for 12 h . Transparent film ( 3 M ) was covered finally to avoid the influence of rabbit lick.
I labeling, stability, and in vivo biodistribution
IgG was labeled with I by the standard chloramine-T oxidation method. Briefly, IgG, I, and chloramine-T were mixed at for 10 min under slight shaking. The synthesized was washed by 10 kDa ultrafiltration three times. To determine radiolabeling stability, was mixed with serum for 24 h at . Portions of the mixture were sampled at different time intervals and filtered by 10 kDa ultrafiltration. The radioactivities retained on the filters were detected by Wizard2 Gamma Counter (PerkinElmer) to calculate radiolabeling stability.
For in vivo biodistribution of I-IgG assay, B16F10 melanoma tumor-bearing mice were topically administrated with free and FCS I-IgG ointment for different time points. Then, the major organs, including the liver, spleen, kidney, heart, lung, and tumor, were collected and measured by Wizard2 Gamma Counter (PerkinElmer). The cumulation efficacy was calculated by the formulation
Where was the radio intensity of the organ at the exact time point, was the mass of the organ, was the radio intensity of the applied ointment.
In vivo treatment for melanoma
The B16F10 melanoma tumor model was established by subcutaneous injection of B16F10 cells into the right flank of each female C57BL/6 mouse. The mice were randomly divided into five groups when their tumor volumes reached about . Each group was treated with an equal dose of aPDL1 and aCTLA4 ( each mouse). The tumors of mice were swiped with different FCS ointment for 12 h , and covered by Transparent film ( 3 M ) to avoid the influence of mice lick. The size of the tumor was measured by a caliper every 2 days:
Growth curves were stopped when the first mouse in the related group was dead, or the first mouse’s tumor size exceeded by ethics from Soochow University Laboratory Animal Center and Soochow University Ethics Committee. In some cases, this limit was exceeded by the last day of measurement, and the mice were immediately euthanized.
Immunofluorescence staining
For in vitro immunofluorescence staining, HACAT cells were incubated in a 24 -well plate to form a single cell layer. Then, FCS/IgG was added and incubated for another 12 h . After removing FCS-IgG, the HACAT cells were fixed by paraformaldehyde, blocked by 5% BSA, and stained with an anti-ZO-1 antibody. One hour later, the cells were stained with a secondary antibody and 4, 6-diamidino-2-phenylindole (DAPI). After mounting, slices were imaged by a confocal fluorescence microscope (Leica SP5). For in vivo immunofluorescence staining, B16F10 tumors and skin were collected from mice at different time points after being swiped with different ointments and embedded in a
temperature-responsive OCT gel for immunofluorescence staining with different antibodies, including anti-Keraten 14 and anti-ZO-1. After washing the primary antibodies, the tumor slides were stained with fluorescently labeled secondary antibodies and DAPI. After mounting, the tumor slices were imaged by the confocal fluorescence microscope (Leica SP5).
Analysis of T cells in different organs
Spleens, tumors, skins, and lymph nodes were collected and homogenized into single-cell suspensions following the standard protocol. Briefly, they were processed through mechanical disruption before digestion for 1 h at in an enzymatic solution with RPMI-1640 ( FBS and 1% PS), collagenase IV (Sigma), lagenase I (Sigma), hyaluronidase (Sigma), and DNase I (Sigma). The samples were then passed through 200-mesh nylon mesh filters to obtain single-cell suspensions. The obtained single-cell suspensions were incubated with anti-CD16/32 for 30 min at . Then, these cells were stained with different antibodies according to the standard protocol.
DCs in lymph nodes and skins were stained with anti-CD11c-FITC (Biolegend, Cat. 117305), anti-CD80-APC (Biolegend, Cat. 104713), and anti-CD86-PE (Biolegend, Cat. 105007).
T cell population in tumors and spleens were stained with anti-CD3-FITC (Biolegend, Cat. 100203), anti-CD4-APC (Biolegend, Cat. 100411), anti-CD45-PerCP (Biolegend, Cat. 103130), and anti-CD8a-PE (Biolegend, Cat. 100707).
To further analyze helper T cells in tumors, the suspension was stained with anti-CD3-FITC (Biolegend, Cat. 100203), anti-CD4-APC (Biolegend, Cat. 100411), anti-Foxp3-PE (Biolegend, Cat. 126403) antibodies with eBioscience Foxp3/Transcription Factor Fixation/PermeabilizationConcentrate and Diluent according to the manufacturer’s protocols. Briefly, after the cells were stained with CD16/CD32 and cell surface markers, they were fixed with the Foxp3 Fixation/Permeabilization working solution for 30 min at room temperature. Then, they were washed with permeabilization buffer at 500 g for 5 min and stained with CD16/CD32 in the permeabilization buffer for 30 min at . Then, they were stained with anti-Foxp3-PE for another 30 min at room temperature and resuspended in the flow cytometry staining buffer for analysis.
For the analysis of cytotoxic cell lymphocytes, the suspension was stained with anti-CD3-APC (Biolegend, Cat. 100206), anti-CD8aPE (Biolegend, Cat. 100707), anti-CD45-PerCP (Biolegend, Cat. 103130) and anti-Ki67-FITC (Biolegend, Cat. 652410) for the level of Ki67 with the same protocol as the Foxp3. The suspension was also stained with anti-CD45-FITC (Biolegend, Cat. 157214), anti-CD8a-PE (Biolegend, Cat. 100707), anti-GranzymeB-PE-Cy7 (Biolegend, Cat. 372214) and anti-CD3-APC (Biolegend, Cat. 100206) for the level of Granzyme B with eBioscience Intracellular Fixation & Permeabilization Buffer Set.
For the analysis of IFN cells in spleens, the suspension was firstly incubated with a stimulation medium with RPMI-1640 (10% FBS and ), HEPES solution (Sigma), 1 mM Sodium pyruvate solution (Sigma), 2-Mercaptoethanol, S1 or OVA protein, and eBioscience Brefeldin A solution for 6 h and then stained with anti-CD3-FITC (Biolegend, Cat. 100203), anti-CD4-PerCP (Biolegend, Cat. 100432), anti-CD8a-PE (Biolegend, Cat. 100707) and anti- IFN- -APC (Biolegend, Cat. 505810) with eBioscience Intracellular Fixation & Permeabilization Buffer Set. For the analysis of IFN cells in tumors, eBioscience Cell Stimulation Cocktail (plus protein transport inhibitors) ( ) was added in the stimulation medium to replace protein and Brefeldin A in the above protocol.
For the analysis of memory T cells, spleens was collected from mice 90 days after the prime-boost and stained with anti-CD3-FITC (eBioscience, Cat. 11-0031), anti-CD8-PerCP-Cy5.5 (eBioscience, Cat. 45-0081), anti-CD44-PE (eBioscience, Cat. 12-0441), and anti-CD62L-
APC (eBioscience, Cat. 17-0621). All these antibodies used in our experiments were diluted by ~200 times.
S1-specific antibody titer
The S1-specific antibody titer was measured by a standard indirect ELISA (iELISA) method. Briefly, 96 -well ELISA plates (Nunc, Thermo) were coated overnight with 1 ng per well of the S1 protein in a coating buffer. The plates were then blocked with of ELISA assay buffer (Thermo) and incubated at room temperature for 2 h . Each serum sample was tested in duplicate at a dilution from 1:100 to 1:640,000 twice in ELISA assay buffer (Thermo), in which was then added into the wells of each plate for 2 h incubation at room temperature. The horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (1:10,000, Abcam) was added for 1 h incubation at room temperature. Then, TMB solution (Thermo) was added to each well. After 15 min incubation, solution was added to stop the reaction. The absorbance of each well was read by the 450 nm wavelength (PerkinElmer). The threshold was defied as the double of the average absorbance in the untreated group. The last dilution of a sample that is just larger than the threshold is defined as the antibody titer of this sample.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The authors declare that all data needed to evaluate the conclusion of this work are within the Article, Supplementary Information, or Source Data file. Source data are provided in this paper.
References
Prausnitz, M. R. & Langer, R. Transdermal drug delivery. Nat. Biotechnol. 26, 1261-1268 (2008).
Prausnitz, M. R., Mitragotri, S. & Langer, R. Current status and future potential of transdermal drug delivery. Nat. Rev. Drug Discov. 3, 115-124 (2004).
Alkilani, A., McCrudden, M. T. & Donnelly, R. Transdermal drug delivery: innovative pharmaceutical developments based on disruption of the barrier properties of the stratum corneum. Pharmaceutics 7, 438-470 (2015).
Prausnitz, M. R. Microneedles for transdermal drug delivery. Adv. Drug Deliv. Rev. 56, 581-587 (2004).
Naik, A., Kalia, Y. N. & Guy, R. H. Transdermal drug delivery: overcoming the skin’s barrier function. Pharm. Sci. Technol. Today 3, 318-326 (2000).
Karande, P., Jain, A., Ergun, K., Kispersky, V. & Mitragotri, S. Design principles of chemical penetration enhancers for transdermal drug delivery. Proc. Natl Acad. Sci. USA 102, 4688-4693 (2005).
Amjadi, M., Mostaghaci, B. & Sitti, M. Recent advances in skin penetration enhancers for transdermal gene and drug delivery. Curr. Gene Ther. 17, 139-146 (2017).
Ahn, D.-G. et al. Current status of epidemiology, diagnosis, therapeutics, and vaccines for novel coronavirus disease 2019 (COVID19). J. Microbiol. Biotechnol. 30, 313-324 (2020).
Ita, K. Transdermal delivery of vaccines-recent progress and critical issues. Biomed. Pharmacother. 83, 1080-1088 (2016).
Nasrollahi, S. A., Taghibiglou, C., Azizi, E. & Farboud, E. S. Cellpenetrating peptides as a novel transdermal drug delivery system. Chem. Biol. Drug Des. 80, 639-646 (2012).
Desai, P., Patlolla, R. R. & Singh, M. Interaction of nanoparticles and cell-penetrating peptides with skin for transdermal drug delivery. Mol. Membr. Biol. 27, 247-259 (2010).
Tadros, A. R. et al. STAR particles for enhanced topical drug and vaccine delivery. Nat. Med. 26, 341-347 (2020).
Zheng, D. et al. Topical delivery of siRNA-based spherical nucleic acid nanoparticle conjugates for gene regulation. Proc. Natl Acad. Sci. USA 109, 11975-11980 (2012).
Chen, Y. et al. Transdermal protein delivery by a coadministered peptide identified via phage display. Nat. Biotechnol. 24, 455-460 (2006).
Marwah, H., Garg, T., Goyal, A. K. & Rath, G. Permeation enhancer strategies in transdermal drug delivery. Drug Deliv. 23, 564-578 (2016).
Cross, S. & Roberts, M. Physical enhancement of transdermal drug application: is delivery technology keeping up with pharmaceutical development? Curr. Drug Deliv. 1, 81-92 (2004).
Nanda, A., Nanda, S. & Khan Ghilzai, N. Current developments using emerging transdermal technologies in physical enhancement methods. Curr. Drug Deliv. 3, 233-242 (2006).
Smith, T. R. F. et al. Immunogenicity of a DNA vaccine candidate for COVID-19. Nat. Commun. 11, 2601 (2020).
Xia, D. et al. An ultra-low-cost electroporator with microneedle electrodes (ePatch) for SARS-CoV-2 vaccination. Proc. Natl Acad. Sci. USA 118, e2110817118 (2021).
Tran, K. T. M. et al. Transdermal microneedles for the programmable burst release of multiple vaccine payloads. Nat. Biomed. Eng. 5, 998-1007 (2020).
Lu, B. et al. lonic liquid transdermal delivery system: progress, prospects, and challenges. J. Mol. Liq. 351, 118643 (2022).
Song, X. et al. Transcutaneous tumor vaccination combined with anti-programmed death-1 monoclonal antibody treatment produces a synergistic antitumor effect. Acta Biomater. 140, 247-260 (2022).
Yang, X. et al. Galactosylated chitosan-modified ethosomes combined with silk fibroin nanofibers is useful in transcutaneous immunization. J. Controlled Release 327, 88-99 (2020).
Shariatinia, Z. Pharmaceutical applications of chitosan. Adv. Colloid Interface Sci. 263, 131-194 (2019).
Li, G. et al. Fluorinated chitosan to enhance transmucosal delivery of sonosensitizer-conjugated catalase for sonodynamic bladder cancer treatment post-intravesical instillation. ACS Nano 14, 1586-1599 (2020).
Zhou, X. et al. Nano-formulations for transdermal drug delivery: a review. Chin. Chem. Lett. 29, 1713-1724 (2018).
Niu, X.-Q. et al. Mechanism investigation of ethosomes transdermal permeation. Int J. Pharm. X 1, 100027 (2019).
Lueßen, H. L. et al. Bioadhesive polymers for the peroral delivery of peptide drugs. J. Controlled Release 29, 329-338 (1994).
Illum, L., Farraj, N. F. & Davis, S. S. Chitosan as a novel nasal delivery system for peptide drugs. Pharm. Res. 11, 1186-1189 (1994).
Antonescu, I. E. et al. The permeation of acamprosate is predominantly caused by paracellular diffusion across cell monolayers: a paracellular modeling approach. Mol. Pharm. 16, 4636-4650 (2019).
Bittermann, K. & Goss, K.-U. Predicting apparent passive permeability of CaCo-2 and MDCK cell-monolayers: a mechanistic model. PLoS ONE 12, e0190319 (2017).
Ahmadi, S. et al. A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis. JCI Insight 5, e132055 (2020).
Robert-Paganin, J., Pylypenko, O., Kikuti, C., Sweeney, H. L. & Houdusse, A. Force generation by myosin motors: a structural perspective. Chem. Rev. 120, 5-35 (2020).
Kováčik, A., Kopečná, M. & Vávrová, K. Permeation enhancers in transdermal drug delivery: benefits and limitations. Expert Opin. Drug Deliv. 17, 145-155 (2020).
Fan, K. et al. Ferritin nanocarrier traverses the blood brain barrier and kills glioma. ACS Nano 12, 4105-4115 (2018).
Liu, Y. et al. A brief review for fluorinated carbon: synthesis, properties and applications. Nanotechnol. Rev. 8, 573-586 (2019).
Lee, Y.-S. Syntheses and properties of fluorinated carbon materials. J. Fluor Chem. 128, 392-403 (2007).
Miller, A. J. & Mihm, M. C. Melanoma. N. Engl. J. Med. 355, 51-65 (2006).
Franklin, C., Livingstone, E., Roesch, A., Schilling, B. & Schadendorf, D. Immunotherapy in melanoma: recent advances and future directions. Eur. J. Surg. Oncol. 43, 604-611 (2017).
Martin-Liberal, J., Kordbacheh, T. & Larkin, J. Safety of pembrolizumab for the treatment of melanoma. Expert Opin. Drug Saf. 14, 957-964 (2015).
Cao, . et al. Granzyme and perforin are important for regulatory cell-mediated suppression of tumor clearance. Immunity 27, 635-646 (2007).
Wei, S. C. et al. Distinct cellular mechanisms underlie anti-CTLA-4 and anti-PD-1 checkpoint blockade. Cell 170, 1120-1133.e17 (2017).
Seidel, J. A., Otsuka, A. & Kabashima, K. Anti-PD-1 and anti-CTLA-4 therapies in cancer: mechanisms of action, efficacy, and limitations. Front Oncol. 8, 86 (2018).
Dammeijer, F. et al. The PD-1/PD-L1-checkpoint restrains T cell immunity in tumor-draining lymph nodes. Cancer Cell 38, 685-700.e8 (2020).
Jiang, W. et al. Exhausted CD8+T cells in the tumor immune microenvironment: new pathways to therapy. Front Immunol. 11, 622509 (2021).
Pielenhofer, J., Sohl, J., Windbergs, M., Langguth, P. & Radsak, M. P. Current progress in particle-based systems for transdermal vaccine delivery. Front. Immunol. 11, 266 (2020).
Corbett, K. S. et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 586, 567-571 (2020).
Elia, U. et al. Design of SARS-CoV-2 hFc-Conjugated ReceptorBinding Domain mRNA Vaccine Delivered via Lipid Nanoparticles. ACS Nano 15, 9627-9637 (2021).
Zhang, N.-N. et al. A thermostable mRNA vaccine against COVID-19. Cell 182, 1271-1283.e16 (2020).
Chakraborty, C., Bhattacharya, M. & Dhama, K. SARS-CoV-2 vaccines, vaccine development technologies, and significant efforts in vaccine development during the pandemic: the lessons learned might help to fight against the next pandemic. Vaccines 11, 682 (2023).
Cheng, Y. & Xu, F. Anticancer function of polyinosinic-polycytidylic acid. Cancer Biol. Ther. 10, 1219-1223 (2010).
Rydyznski Moderbacher, C. et al. Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell 183, 996-1012.e19 (2020).
Shi, J., Li, Y., Chang, W., Zhang, X. & Wang, F.-S. Current progress in host innate and adaptive immunity against hepatitis C virus infection. Hepatol. Int 11, 374-383 (2017).
Rahe, M. & Murtaugh, M. Mechanisms of adaptive immunity to porcine reproductive and respiratory syndrome virus. Viruses 9, 148 (2017).
Schudel, A., Francis, D. M. & Thomas, S. N. Material design for lymph node drug delivery. Nat. Rev. Mater. 4, 415-428 (2019).
Davies, M. & Duffield, E. A. Safety of checkpoint inhibitors for cancer treatment: strategies for patient monitoring and management of immune-mediated adverse events. Immunotargets Ther. 6, 51-71 (2017).
Acknowledgements
This article was partially supported by the National Research Programs of China (No. 2021YFF0701800, 2020YFA0211100), the National Natural
Science Foundation of China (Nos. T2321005, 52032008), the China Postdoctoral Science Foundation funded project (No. 2023M732543), Fundo para o Desenvolvimento das Ciências e da Tecnologia (FDCT 0002/2022/AKP), Emergency Research Project of COVID-19 from Zhejiang University, National Center of Technology Innovation for Biopharmaceuticals, Collaborative Innovation Center of Suzhou Nano Science and Technology, and the 111 Program from the Ministry of Education of China. Z Liu has been supported by the New Cornerstone Science Foundation through the New Cornerstone Investigator Program and the XPLORER PRIZE.
Author contributions
Wenjun Zhu, Qian Chen, and Zhuang Liu conceived and designed the experiments. Wenjun Zhu and Ting Wei designed and synthesized the materials and nano-complexes. Wenjun Zhu and Qiutong Jin performed the in vitro experiments. Wenjun Zhu, Yuchun Xu and Yu Chao performed the in vivo experiments on melanoma. Wenjun Zhu, Jiaqi Lu, Jun Xu performed the in vivo experiments on vaccines. Wenjun Zhu, Jiafei Zhu and Muchao Chen performed the flow cytometry experiments. Wenjun Zhu, Yuchun Xu and Xiaoying Yan performed all the revised experiments. Wenjun Zhu, Qian Chen and Zhuang Liu analyzed all the data and wrote the paper. All authors discussed the experimental results and edited the paper.
Correspondence and requests for materials should be addressed to Qian Chen or Zhuang Liu.
Peer review information Nature Communications thanks Hongsheng Wang, Arun Arun Iyer, and the other anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.