الجروح المزمنة الناتجة عن السكري معرضة لخطر دائم لتطوير قرحات القدم السكري بسبب نقص الأكسجين الشديد، ووجود أنواع الأكسجين التفاعلية (ROS) بشكل مفرط، وبيئة التهابية معقدة، وإمكانية الإصابة بالبكتيريا. هنا نطور استراتيجية علاج مبرمجة تستخدم هايماتوكوكوس الحي (HEA). من خلال تعديل شدة الضوء، يمكن برمجة HEA لأداء مجموعة متنوعة من الوظائف، مثل النشاط المضاد للبكتيريا، وتوفير الأكسجين، والتخلص من ROS، وتنظيم المناعة، مما يشير إلى إمكانيته للاستخدام في العلاج المبرمج. تحت شدة الضوء العالية ( )، HEA الأخضر (GHEA) مع تحويل ضوئي حراري فعال يقوم بتعقيم سطح الجرح. من خلال تقليل شدة الضوء ( )، يمكن للنظام الضوئي لـ GHEA أن ينتج الأكسجين باستمرار، مما يحل بفعالية مشاكل نقص الأكسجين ويعزز تجديد الأوعية الدموية. يؤدي الإشعاع الضوئي المستمر إلى تراكم الأستازانتين (AST) في خلايا HEA، مما يؤدي إلى تحول تدريجي من اللون الأخضر إلى الأحمر (RHEA). يقوم RHEA بفعالية بالتخلص من ROS الزائد، ويعزز تعبير إنزيمات مضادات الأكسدة داخل الخلايا، ويوجه الاستقطاب نحو البلعميات M2 عن طريق إفراز حويصلات AST عبر الإكسوسومات. يمكن للهلام الحي HEA تعقيم وتعزيز تكاثر الخلايا وهجرتها وتعزيز تكوين الأوعية الجديدة، مما قد يحسن شفاء الجروح المصابة بالسكري في الفئران الإناث.
يعاني حوالي 537 مليون شخص حول العالم من مرض السكري، ومن المتوقع أن تزداد نسبة حدوثه بنسبة بحلول . ربع مرضى السكري لديهم تهديد مدى الحياة لجروح غير شافية مثل قرحات القدم السكري، مما يجبر المرضى على مواجهة خطر البتر، و من هؤلاء الأفراد لديهم متوسط عمر أقل من 5 سنوات . أظهرت الدراسات أن تضرر تكوين الأوعية الدموية استجابةً لنقص الأكسجين هو أحد الأسباب الأكثر خطورة لتدهور الجروح المزمنة لدى مرضى السكري . يؤدي التعرض لمستويات عالية من الجلوكوز إلى تحفيز هيدروكسلة ما بعد الترجمة السريعة وتدهور عامل نقص الأكسجين 1 ألفا. عند الاستجابة لنقص تروية الأنسجة الرخوة، تجعل هذه التغيرات الجروح السكري غير قادرة على زيادة مستوى عامل نمو بطانة الأوعية الدموية
(VEGF)، مما يخلق عقبات أمام تكوين الأوعية وشفاء الجروح . بالإضافة إلى ذلك، فإن الأنواع المفرطة من الأكسجين التفاعلية (ROS) هي عقبة مهمة أخرى أمام عملية شفاء الجروح السكري، مما يتسبب في أضرار دائمة وغير قابلة للإصلاح للجزيئات الحيوية ويستمر في الحفاظ على البلعميات في نمط M1 مما يؤدي إلى تفاقم الاستجابة الالتهابية . نتيجة لذلك، فإن تجنيد البلعميات M1 يخلق بيئة إصابة فريدة في الجروح تتميز بزيادة التحلل البروتيني والأضرار الخلوية الناتجة عن الأكسدة . علاوة على ذلك، فإن الجروح المفتوحة معرضة بشدة للإصابة بالبكتيريا، مما يؤدي إلى تفاقم نقص الأكسجين والالتهاب في الجروح . لذلك، فإن تصميم جهاز شامل بإطلاق الأكسجين القابل للتحكم في الزمان والمكان، والتخلص من ROS، وتعديل
استقطاب البلعميات، وخصائص مضادة للبكتيريا هو موضوع ساخن لعلاج الجروح المزمنة الناتجة عن السكري.
الهلامات هي شبكات بوليمرية ثلاثية الأبعاد مترابطة يمكنها امتصاص والاحتفاظ بكمية كبيرة من الماء أو السوائل البيولوجية. لديها مجموعة من الخصائص الفيزيائية والكيميائية التي تجعلها مناسبة لمجموعة متنوعة من التطبيقات في مجالات مختلفة في الحياة الواقعية . تم تطبيق الهلامات بشكل واسع في المجال الطبي بسبب توافقها الحيوي. يمكن استخدامها كدعائم لهندسة الأنسجة، وأنظمة توصيل الأدوية، وعدسات الاتصال. يمكن أيضًا تصميم الهلامات للاستجابة للمؤثرات الخارجية مثل درجة الحرارة، ودرجة الحموضة، أو الضوء، مما يتيح إطلاق الأدوية بشكل متحكم فيه. في الزراعة، يمكن دمج الهلامات في التربة لتحسين احتباس الماء وتوافر المغذيات للنباتات. تساعد في تقليل استخدام المياه، ومنع تآكل التربة، وتعزيز نمو النباتات من خلال توفير بيئة ملائمة للجذور . في مجال معالجة البيئة، يمكن استخدام الهلامات في معالجة مياه الصرف الصحي وتنظيف البيئة. يمكنها امتصاص وإزالة الملوثات من الماء، بما في ذلك المعادن الثقيلة والملوثات العضوية . تعتبر الهلامات ضمادات جروح واعدة بسبب نعومتها التي تعادل نعومة المصفوفة خارج الخلوية (ECM) وقدرتها على امتصاص السوائل بشكل معتدل . نظرًا لهيكلها المجهري المسامي، تتمتع الهلامات بقدرة تحميل عالية . لذلك، من المرغوب فيه للغاية إنشاء ضمادة هلامية محملة بمواد أو أدوية متعددة الوظائف لمعالجة القضايا المذكورة أعلاه في وقت واحد . في الواقع، تم تطوير مجموعة متنوعة من الهلامات متعددة الوظائف لإصلاح الجروح تحتوي على كميات عالية من الأكسجين لحل نقص الأكسجين، ومواد بوليفينولية أو نانوإنزيمات للتخلص من ROS، وجزيئات تعديل استقطاب البلعميات لتنظيم البيئة المناعية . على سبيل المثال، طور زهاو وآخرون ضمادة جروح علاجية، وهي , باستخدام نظام هلامي بيوميميتيك ونانوإنزيمات مقلدة لبيروكسيد الهيدروجين المعدلة. تم تصميم الهلام ليتناسب في الوقت نفسه مع الإشارات الميكانيكية والكهربائية للجلد بينما يمتلك القدرة المؤكسدة التي تنشط بواسطة . أعد وو وآخرون هلامًا ديناميكيًا متعدد الاستخدامات من قاعدة شيف وإستر البورات متقاطع الارتباط يمكنه توليد الأكسجين باستمرار، وتعزيز استقطاب M2 للبلعميات، والقضاء على الأنواع التفاعلية من الأكسجين والنيتروجين . أعد زانغ وآخرون هلامًا قابلًا للحقن يعتمد على بلازما غنية بالصفيحات ولابونايت يمكن أن يسرع شفاء الجروح من خلال تعزيز استقطاب البلعميات وتكوين الأوعية في الجلد الكامل . من الواضح أن التصميم الحالي للهلامات متعددة الوظائف يتضمن قضايا كبيرة، مثل الفصل المعقد، والتحضير الممل، والكفاءة التآزرية المنخفضة، والسيطرة المحدودة على الزمان والمكان. لذلك، هناك حاجة ملحة لضمادة هلامية ذات تركيبة بسيطة ولكنها تتبع استراتيجية علاجية إجرائية. الجيلاتين الميثاكريلي (GelMA) هو جيلاتين مزدوج الوظيفة يتم الحصول عليه من خلال تفاعل الجيلاتين الأميني مع الأنهدريد الميثاكريلي. توفر مجموعات الأمين الوفيرة الموزعة على طول السلسلة الرئيسية للجيلاتين مواقع تفاعلية غنية للأنهدريد الميثاكريلي. يمكن أن يتفاعل الأنهدريد الميثاكريلي، المرتبط بمجموعات الأمين، مع بعضها البعض لتشكيل هياكل ثلاثية الأبعاد مناسبة لنمو الخلايا وتمايزها في الأبحاث العلمية المتعلقة بالتكنولوجيا. لقد ثبت أن GelMA يتمتع بتوافق حيوي ممتاز، وخصائص التصاق الخلايا، وأداء ميكانيكي. يتم تطبيقه على نطاق واسع في هندسة الأنسجة، وأنظمة توصيل الأدوية، والطباعة ثلاثية الأبعاد، وغيرها من المجالات .
مقارنةً بإعادة تركيب المكونات النشطة إلى خلية ميكروبية نشطة واحدة، فإن الأخيرة تقدم مجموعة أوسع من الوظائف وأسهل في البرمجة . لعبت الكائنات الدقيقة دورًا حاسمًا في تقدم الأبحاث الزراعية والصناعية والصحية العامة من خلال تزويد البشر بمبادئ ومنهجيات متقدمة . أولاً، في حالات عدم التعايش، تشارك أنواع مختلفة من الكائنات الدقيقة في تفاعلات تمنع نمو بعضها البعض، مما يساهم في النشاط المضاد للبكتيريا في شفاء الجروح. ثانيًا، يلعب الأكسجين دورًا حاسمًا في جوانب متعددة من شفاء الجروح، مثل تكاثر الخلايا،
الهجرة، الالتصاق، تطوير الأوعية الدموية، وتجديد الأنسجة. يُعتقد على نطاق واسع أن الكائنات الدقيقة، مثل الكلوروفايتا، هايماتوكوكوس (HEA)، والأركيا، لعبت دورًا أساسيًا في ظهور وتطور الحياة على الأرض من خلال إنتاج الأكسجين عبر عملية التمثيل الضوئي. بدلاً من الاعتماد على مواد إمداد الأكسجين التقليدية، مثل الهيموغلوبين، وأكاسيد المعادن، والبيروكسيدات، وأنظمة توصيل الأكسجين الأخرى ، والتي لديها سعات تحميل محدودة وغير قابلة للتحكم ومعدلات إفراج، هذه الكائنات ذاتية التغذية ضوئيًا. تستخدم أنظمة ضوئية لعملية التمثيل الضوئي، مما يمكنها من إنتاج الأكسجين. على سبيل المثال، طور تشو وآخرون استراتيجية هيدروجيل حيوي نشط لتثبيط المكورات العنقودية الذهبية المقاومة للميثيسيلين. كان الهيدروجيل الحيوي النشط مكونًا من الطحالب الدقيقة الحية سبيرولينا بلاتنسيس المحملة بالبيربرين وكاربوكسي ميثيل الكيتوزان/ألغينات الصوديوم، والتي يمكن أن تطلق BBR ووإنتاج أنواع الأكسجين التفاعلية، مما يسهم في العلاج الكيميائي الضوئي الديناميكي لعدوى المكورات العنقودية الذهبية المقاومة للميثيسيلين. بالإضافة إلى ذلك، يمكن للعديد من المستقلبات والخلايا من الكائنات الدقيقة تنظيم الميكروبيوم المناعي والعمل كمضادات للأكسدة؛ كما تُستخدم هذه المواد في مجموعة متنوعة من إضافات الطعام والأدوية.الأستازانتين، وهو ناتج أيضي لـ HEA (3،3′-ثنائي الهيدروكسي، -كاروتين-4،4′-ديون، AST)، لديه قدرة عالية على التخلص من الجذور الحرة النشطة وخصائص مضادة للأكسدة.لقد تم الإبلاغ عن أن الأنزيمات المضادة للأكسدة، مثل الجلوتاريدوكسي (monothiol glutaredoxin)، الجلوتاريدوكسي (GRX)، الثيوريدوكسي (TRX)، مختزل الثيوريدوكسي (TrxR)، الفيريتين، مختزل المونوهيدروأسكوربات (MDAR)، البيروكسيريدوكسي (PrxR)، بيروكسيداز الجلوتاثيون (GPX)، مختزل الجلوتاثيون (GR)، الكاتلاز (CAT)، ديسموتاز السوبر أكسيد (SOD)، وبيروكسيداز الأسكوربات (APX)، تمتلك أنشطة إنزيمية متنوعة لمكافحة الجذور الحرة.“. بالإضافة إلى ذلك، يمكن أن يثبط AST تعبير السيتوكينات الالتهابية (COX-2، TNF-IL-6 و IL-1 ) . وبالتالي، تمتلك ASTs إمكانيات كبيرة وتقدم تطبيقات واعدة في صحة الإنسان ضد مجموعة واسعة من الأمراض. تقوم AST بتنظيم مستقبل البروتين المرتبط بالبروتين الدهني منخفض الكثافة-1، الذي يمكن أن يرتبط ويقلل من p65 وp-c-Jun وNF-Kb، مما يمنع تفعيل مسارات JNK وNF-кB. تثبيط تفعيل JNK1 وNF-кB يعزز من استقطاب M2 للبلعميات.في السنوات الأخيرة، كان هناك اهتمام متزايد في استخدام الطحالب الدقيقة، مثل HEA، لإنتاج مضاد الأكسدة الطبيعي AST. ويعزى ذلك بشكل أساسي إلى معدلات نموها الأسرع وقدراتها الأعلى في إنتاج الكتلة الحيوية.بالإضافة إلى ذلك، يُعتقد أن HEA تمتلك أكبر قدرة على تراكم AST الطبيعي في ظل ظروف الضغط البيئي.استنادًا إلى تطور الأكسجين التدريجي والقابل للتحكم، بالإضافة إلى قدرات إطلاق AST للسبائك المعدنية عالية الأداء، هناك إمكانيات هائلة على المدى الطويل لتطبيقها في الهياكل الاصطناعية لشفاء الأنسجة المعقدة، وأجهزة الأكسجين القلبية، وكملحقات في علاج الأورام. وحسب علمنا، لم تكن هناك تقارير عن استخدام الكائنات الدقيقة من أجل الأكسجة التدريجية وتنظيم البيئة الدقيقة في علاج الجروح المزمنة الناتجة عن السكري.
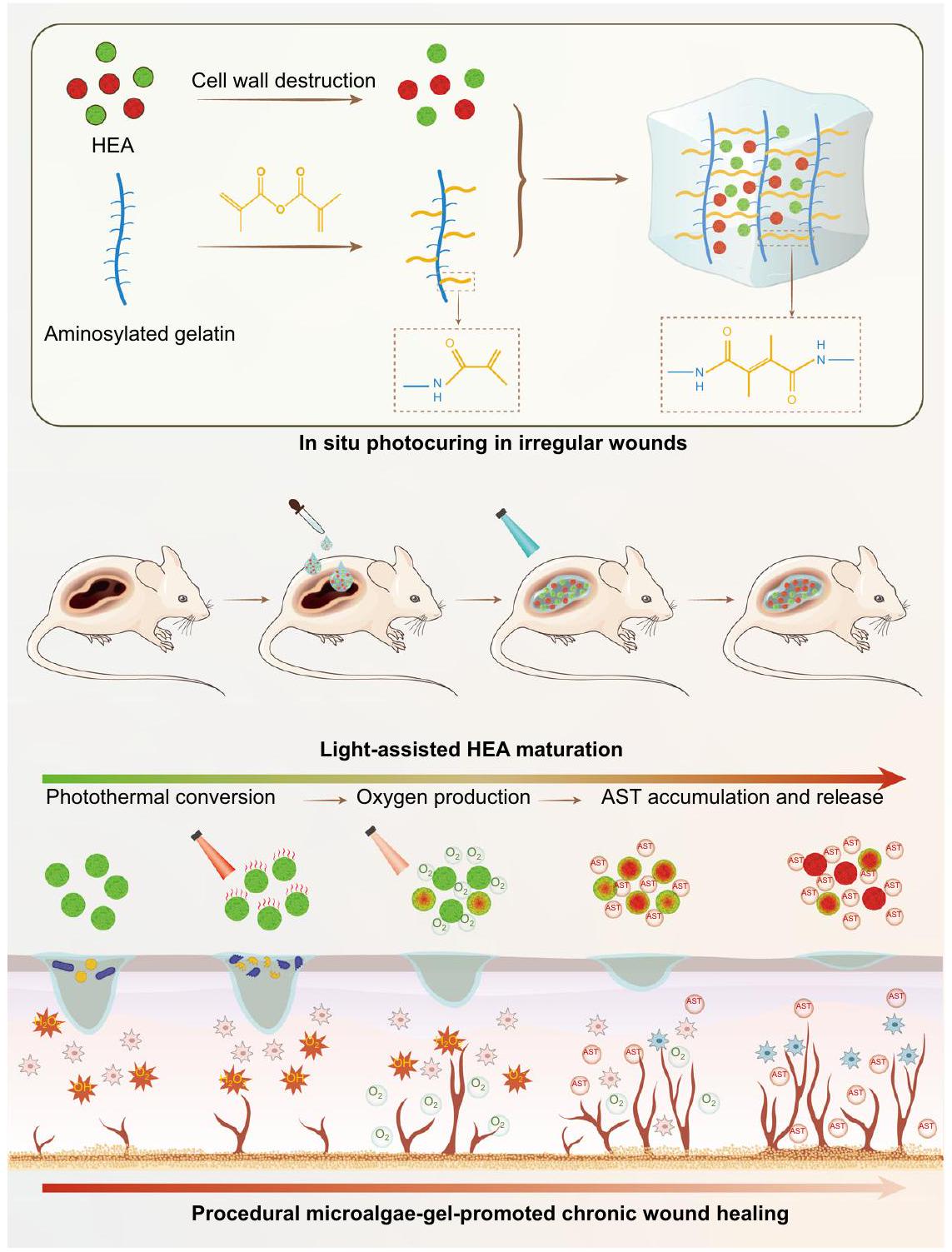
هنا، نقوم بتطوير هيدروجيل بسيط عن طريق احتواء خلايا HEA النشطة في هلام GelMA التقليدي ونؤسس نظامًا للعلاج الإجرائي المحلي (الشكل 1). لزيادة كفاءة إطلاق الأكسجين وAST، يتم أولاً تحضير كريات HEA، وتحميلها بمحلول GelMA، وتثبيتها ضوئيًا في الموقع لتأسيس جروح غير منتظمة بشكل أفضل. وفقًا لكثافة طاقة الضوء، يتوسط وفرة الكلوروفيل الانتقال بين إنتاج الأكسجين عبر عملية التمثيل الضوئي وإنتاج الحرارة عبر تأثير التحويل الضوئي الحراري في خلايا HEA الخضراء (GHEA). لذلك، في المرحلة الأولية من العلاج، يحدث تحويل ضوئي حراري فعال تحت إشعاع ليزر 658 نانومتر مع يمكن أن تقضي كثافة الطاقة بسرعة على البكتيريا. في المرحلة الثانية من العلاج، يتم حل نقص الأكسجين الشديد في الجرح من خلال توفير إمداد مستمر من الأكسجين لعملية التمثيل الضوئي تحت إشعاع ليزر 658 نانومتر معكثافة الطاقة. مع تدهور بيئة الثقافة (المربوطة في الهيدروجيل) أو التحفيز الخارجي (إشعاع الليزر)، تتراكم AST تدريجياً، مما يحول خلايا HEA إلى اللون الأحمر (RHEA) ويمنحها قدرة قوية على تعديل بيئة الجرح الدقيقة. بعد ذلك، تطلق خلايا HEA بشكل روتيني
الشكل 1 | توضيح تخطيطي لتحضير وآلية استراتيجية الشفاء من الجروح المعتمدة على HEA@Gel. تم الحصول على ضمادات جروح غير منتظمة من خلال التصلب الضوئي في الموقع. من خلال تعديل شدة الضوء، يمكن أن يكون HEA@Gel برمجة لأداء النشاط المضاد للبكتيريا، توفير الأكسجين، إزالة الجذور الحرة، وتنظيم المناعة. HEA هيماتوكوكوس، AST أستازانتين.
يعمل AST على جمع الجذور الحرة بشكل مستمر وتنظيم استقطاب البلعميات. يتم تسريع شفاء الجروح المصابة لدى مرضى السكري بشكل كبير من خلال إجراء شامل يقضي على البكتيريا، ويزود الأكسجين، ويجمع الجذور الحرة، ويعزز تكوين الأوعية الدموية، ويسهل التأثيرات المضادة للالتهابات من خلال استقطاب البلعميات M2.
النتائج
تحضير وخصائص هلام HEA
قمنا بتصميم ضمادة هيدروجيل إجرائية قائمة على الطحالب الدقيقة الحية (HEA@Gel) لتسهيل تغطية الجرح وتوصيل الأكسجين القابل للذوبان وAST إلى سرير الجرح عبر التصلب الضوئي في الموقع (الشكل 2أ). للتحقق من مفهوم العلاج الإجرائي، تم تحديد أكثر الأدلة وضوحًا، وهو تغيير لون خلايا HEA، ويظهر في الشكل 2ب-هـ. عندما تكون العناصر الغذائية الضرورية وفيرة، تنمو خلايا HEA بسرعة وتظهر باللون الأخضر بسبب وجود الكلوروفيل (الشكل 2ب). مع استهلاك العناصر الغذائية والتعرض للضوء، انخفض معدل نمو خلايا HEA، وتراكم AST تدريجيًا (الشكل 2ج، د). مع استمرار تراكم AST، أصبحت خلايا HEA داكنة حمراء بشكل ملحوظ (الشكل 2هـ). الجدار الخلوي الصلب والسميك لخلايا HEA يؤدي إلى انخفاض نفاذيتها، مما يمنع إطلاق AST بشكل فعال. من داخل الخلايا (الشكل 2 فلإزالة جدران خلايا HEA، قمنا بتحضير بروتوبلاست HEA باستخدام طريقة إنزيمية خفيفة (سيلولاز وبيكتيناز). كما هو موضح في الشكل 2g والشكل التكميلي 1، احتفظت بروتوبلاست HEA المعالجة بشكلها وبنيتها ونشاطها بعد إزالة جدران خلاياها. ومن المثير للاهتمام، أنه بعد فترة من الثقافة، أفرزت بروتوبلاست HEA عددًا كبيرًا من الحويصلات بقطر 200 نانومتر، والتي عززت الإفراج الفعال وتوصيل AST غير القطبي (الشكل 2h، i). لإثبات وجود AST داخل الحويصلات الإفرازية، تم صبغ الحويصلات بصبغة Dil لتوسيم الغشاء. نظرًا للفلورية الفطرية لـ AST، تم ملاحظة AST والحويصلات تحت مجهر ليزر متداخل، كما هو موضح في الشكل التكميلي 2، مما يكشف عن تداخل واضح بين AST والحويصلات. لمقارنة تأثير وجود جدار الخلية على إفراز AST بشكل أفضل، تم تفريق كميات متساوية من HEA (مع جدار الخلية) وبروتوبلاست HEA (بدون جدار الخلية) بشكل منفصل في 100 مل من الماء المقطر. تم قياس محتوى AST المفرج باستخدام المنحنى القياسي لـ AST الموضح في الشكل التكميلي 3. كما هو موضح في الشكل التكميلي 4a، كانت نسبة إفراز AST من بروتوبلاست HEA أكبر بكثير من تلك التي من بروتوبلاست HEA مع جدران الخلايا المحتفظ بها. علاوة على ذلك، مع مرور الوقت، كان هناك زيادة مستمرة في
الشكل 2 | تحضير HEA@Gel وخصائصه. أ تمثيل تخطيطي لفوائد معالجة GelMA بالضوء. ب-هـ صور لمراحل مختلفة من
صور لـ (ل) جل، (م) GHEA@Gel، و (ن) RHEA@Gel. قضبان القياس،تمثل الصور في (ل-ن) صور Gel و GHEA@Gel و RHEA@Gel، على التوالي. بالنسبة لهذه التوصيفات الشكلية لـ Gel و GHEA@Gel و RHEA@Gel المُصنّعة، تم تكرار كل تجربة ثلاث مرات بشكل مستقل مع نتائج مشابهة. كمية AST المفرج عنها من بروتوبلاستات HEA. على النقيض من ذلك، في HEA مع جدران خلوية محتفظ بها، انخفض معدل إفراز AST بشكل ملحوظ بعد 6 ساعات. قد يُعزى هذا الظاهرة إلى عرقلة الإفراج الإضافي عن AST بسبب وجود جدران خلوية بعد إفراج بعض AST القابل للإفراج بسهولة في المرحلة السابقة. بالإضافة إلى ذلك، من خلال جمع AST المفرج عنها في أنبوب طرد مركزي، تم ملاحظة تغير اللون مع مرور الوقت بصريًا (الشكل التوضيحي 4b).
أظهرت GelMAs ذات الشحنات الإيجابية، والعقاقير القابلة للحقن، والتصلب الضوئي في الموقع إمكانات هائلة لعلاج الجروح غير المنتظمة في هذه الدراسة. وبالتالي، تم دمج محلول GelMA مع خلايا HEA ثم تم تغطيته على الجرح؛ بعد 5 ثوانٍ من التعرض للأشعة الزرقاء، تم تصلب الهيدروجيل وناسب تمامًا موقع الجرح. تُظهر الأشكال 2j وk صورًا لـ Gel النقي وGHEA@Gel وRHEA@Gel، بالإضافة إلى صور آلية التصلب الضوئي. تم فحص الشكل والمظهر لـ Gel وHEA@Gel المُعد مسبقًا عبر المجهر الإلكتروني الماسح بالتبريد (FEI Quanta 450). كما هو موضح في الشكل 2l، أظهر عينة الجل هيكلًا مساميًا واضحًا. احتفظت GHEA@Gel وRHEA@Gel بهيكلهما متعدد المسام، مما يضمن النقل وAST (الشكل 2m وn).
النشاط المضاد للبكتيريا لـ GHEA@Gel
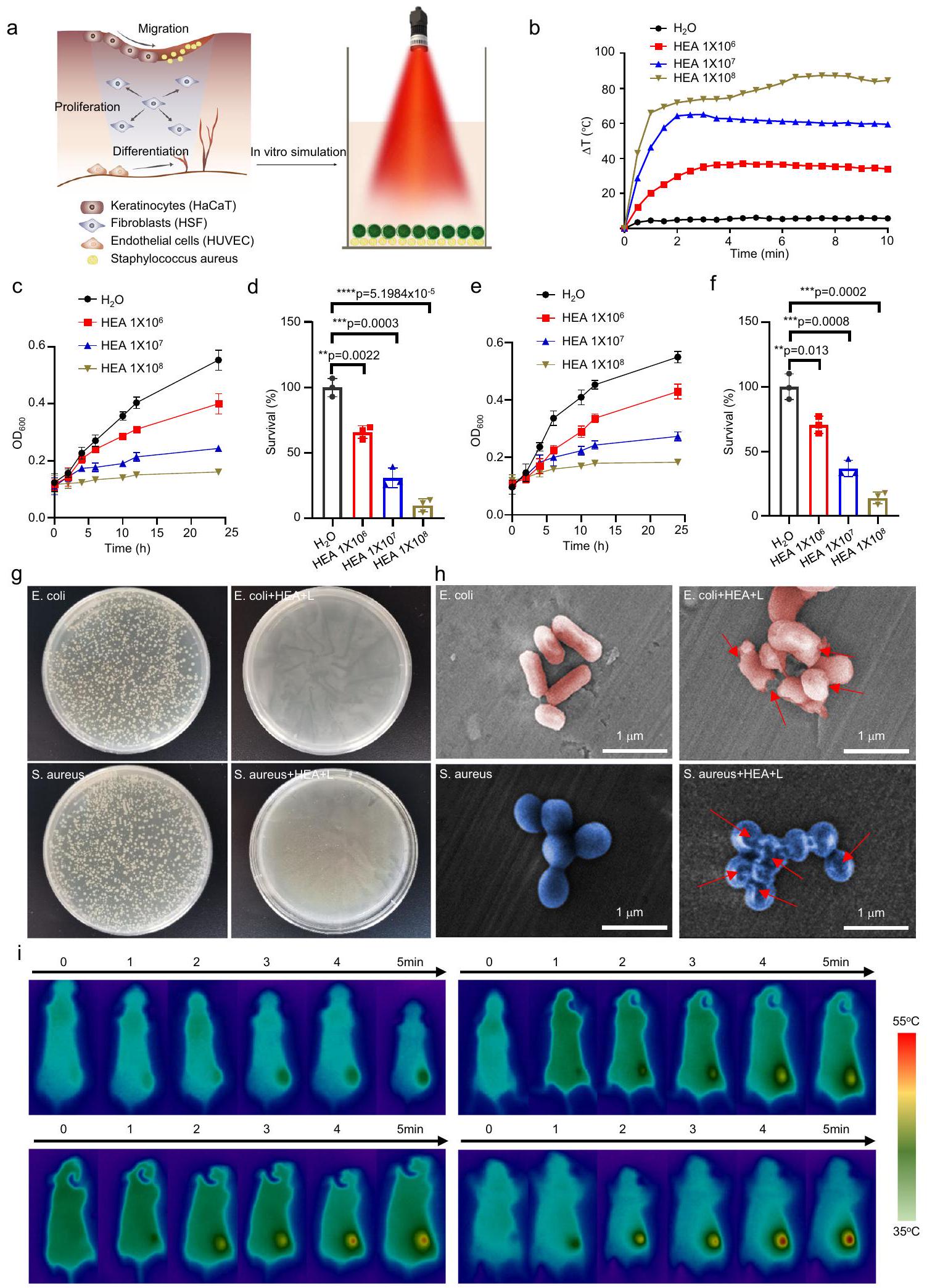
نظرًا للتعرض المستمر لجروح DU للبيئة الخارجية، هناك خطر كبير من العدوى البكتيرية الخارجية وتأخر شفاء الجروح بشكل كبير. كما تم الإبلاغ سابقًا , تتفاعل أنواع مختلفة من الكائنات الدقيقة مع بعضها البعض لتثبيط نمو بعضها البعض في حالة عدم التعايش، مما يساهم في النشاط المضاد للبكتيريا للجروح. للتحقق من تأثير عدم التعايش لعلاج الدمج، تم قياس تكاثر الإشريكية القولونية (E. coli) والمكورات العنقودية الذهبية (S. aureus) بعد الزراعة المشتركة مع GHEA دون التعرض للأشعة الضوئية. كما هو موضح في الشكل التكميلي 5، لوحظ تثبيط واضح لنمو البكتيريا لكل من E. coli وS. aureus، حيث ظلت معدلات بقاء E. coli وS. aureus، على التوالي، عند و، على التوالي، عندما وصلت كثافة خلايا GHEA إلى . بالإضافة إلى ذلك، نظرًا لإمكانية التحويل الضوئي الحراري لخلايا HEA، قمنا بتقييم النشاط المضاد للبكتيريا في المختبر لـ GHEA من خلال تقييم التأثير الضوئي الحراري تحت إشعاع ليزر 658 نانومتر (الشكل 3a). كما هو موضح في الشكل 3b، كان التأثير الضوئي الحراري لـ HEA@Gel متناسبًا مع كثافة خلايا HEA. زادت درجة حرارة الماء فقط بمقدار تحت إشعاع ليزر 658 نانومتر.
عندما وصلت كثافة خلايا GHEA إلى ، زادت درجة حرارة GHEA@Gel بمقدار ، مما يدل على تأثير ضوئي حراري قوي. تم اختيار E. coli وS. aureus بعد ذلك كالبكتيريا النموذجية للتحقيق في النشاط المضاد للبكتيريا الذي يتم بوساطة PTT لـ GHEA@Gel. بعد خطوات معالجة مختلفة، تم قياس الكثافة الضوئية (OD) لتعليق البكتيريا عند 600 نانومتر لتقييم النشاط البكتيري. كما هو موضح في الأشكال 3c-f، تم تثبيط النمو البكتيري بشكل كبير من خلال التأثير الضوئي الحراري لـ GHEA@Gel بالاشتراك مع إشعاع الليزر. لتحديد النشاط المضاد للبكتيريا لـ GHEA@Gel بشكل أكبر، تم إجراء اختبار صفيحة الأجار (الشكل 3g). بعد GHEA@Gel وإشعاع ليزر 658 نانومتر، كان عدد مستعمرات البكتيريا على صفيحة الأجار ضئيلًا. افترضنا أن الانفجار السريع لأغشية البكتيريا قد يكون نتيجة لتأثير حراري محلي ناتج عن التأثير الضوئي الحراري القوي لـ GHEA@Gel. ثم قمنا بتقييم سلامة غشاء البكتيريا باستخدام SEM (الشكل 3h). لوحظ أن البكتيريا الطبيعية كانت لها شكل رهابي أو كروي مع سطح أملس. بعد العلاج بـ GHEA@Gel وإشعاع الليزر عند 658 نانومتر، كانت أغشية خلايا البكتيريا متضررة بشدة، مع انهيار واضح في السطح. من خلال تدمير غشاء البكتيريا، تم تعقيم GHEA@Gel، الذي له تأثير ضوئي حراري بوساطة ليزر 658 نانومتر، بسرعة. تم تقييم التحويل الضوئي الحراري لـ GHEA@Gel بعد ذلك بشكل أكبر في الجسم الحي. كما هو موضح في الشكل 3i، لوحظ تحويل ضوئي حراري واضح وممتاز في جروح DU للفئران، حيث يمكن لليزر 658 نانومتر زيادة درجة الحرارة إلى في
5 دقائق. كانت درجة حرارة تضمن قتل البكتيريا في الجرح بشكل فعال، مع تأثير عدم التعايش على خلايا HEA. ومع ذلك، يجب أيضًا مراعاة عيوب هذه العملية، مثل الأضرار التي تسببها درجات الحرارة العالية على الأنسجة الجلدية الطبيعية في موقع الجرح. للتحقيق في الأضرار التي لحقت بالجلد الطبيعي عند ، تم حرق جلد الفئران بدرجة حرارة ثابتة من مكواة اللحام الكهربائية عند لمدة 5 دقائق. بعد ذلك، تم تسجيل الأضرار الجلدية في موقع الحرق. أظهرت النتائج أن علامات حروق حمراء طفيفة تركت على جلد الفئران بعد حرقها عند ، مما يشير إلى أن يمكن أن تسبب ضررًا لخلايا الجلد لدى الفئران. بعد 4 أيام من علاج الحروق، تلاشت الفلورسنت الحمراء تدريجيًا (الشكل التكميلي 6)، مما يدل على أن الكائن الحي لديه قدرة جيدة على الشفاء الذاتي عند على الحروق الطفيفة. للتحقيق بشكل أكبر في الأضرار التي تسببها على خلايا الجلد الطبيعية، تم تعريض الخلايا الظهارية لـ لمدة 5 دقائق، بعد ذلك تم استخدام FCM للكشف عن موت الخلايا. أظهرت النتائج أنه، مقارنة بالعلاج الضابط، تسبب في موت خلايا يمكن تحمله (الشكل التكميلي 7). باختصار، على الرغم من أن درجة الحرارة العالية لـ كان لها بعض التأثيرات الطفيفة على الخلايا الطبيعية أثناء قتل البكتيريا المصابة، خلال عملية التشغيل الفعلية، لأن مدة كانت قصيرة جدًا (أقل من 1 دقيقة) ولأن الجسم لديه قدرة قوية على الشفاء الذاتي لـ الحروق الطفيفة، فإن هذه الآثار الجانبية مقبولة تمامًا في علاج عدوى الجروح.
الإفراج عن GHEA@Gel
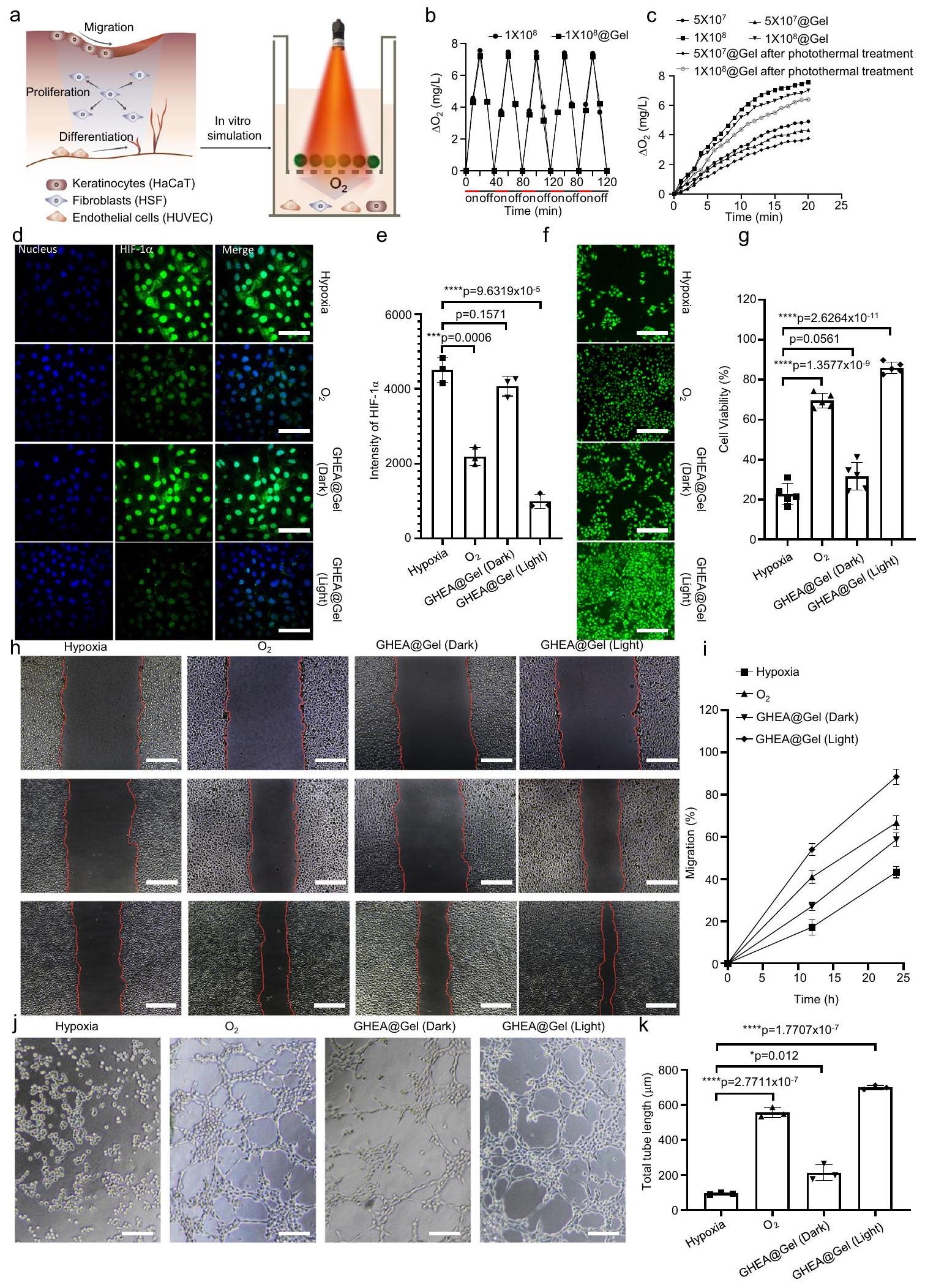
تم تأكيد نشاط GHEA@Gel في البداية من خلال تعريضه للضوء الأحمر ( ) وقياس تركيز الأكسجين القابل للذوبان باستخدام ميكروإلكترود (الشكل 4a). زاد تركيز الأكسجين القابل للذوبان في GHEA@Gel من 0 إلى أكثر من خلال 20 دقيقة تحت ظروف ضوئية كافية، بينما في الظلام، انخفض تركيز الأكسجين القابل للذوبان من 8 إلى خلال 20 دقيقة (الشكل 4b). تشير هذه النتائج إلى أن GHEA في الجل نشط تمامًا وقادر على التمثيل الضوئي والتنفس. في نطاق معين، كانت معدل إنتاج الأكسجين مرتبطًا إيجابيًا بتركيز GHEA@Gel، وكان التركيز الأمثل هو مل (الشكل 4c). تم إجراء تجربة إنتاج الأكسجين أعلاه تحت شدة ضوء قدرها . للتحقيق فيما إذا كان GHEA يمكن أن ينتج الأكسجين تحت إشعاع ليزر بشدة قدرها ، تم تعريض GHEA وGHEA@Gel لإشعاع ليزر 658 نانومتر بشدة قدرها ، وتم قياس التغيرات في توفر الأكسجين في النظام كل دقيقتين لمدة إجمالية قدرها 30 دقيقة. أظهرت النتائج، كما هو موضح في الشكل التكميلي 8، أن منحنى إنتاج كان مشابهًا جدًا للمنحنى عند . للتحقيق فيما إذا كان التأثير الضوئي الحراري لشدة الضوء العالية قدرها يؤثر على قدرة إنتاج الأكسجين لـ GHEA في GHEA@Gel، تم وضع عينات GHEA@Gel التي تحتوي على خلايا GHEA بتركيزات مختلفة تحت إشعاع ليزر 658 نانومتر لمدة 5 دقائق. بعد ذلك، بعد أن انخفضت درجة حرارة GHEA إلى درجة حرارة الغرفة، تم استخدام ليزر منخفض الشدة ( ) لإشعاع GHEA@Gel، بعد ذلك تم قياس إنتاج الأكسجين خلال عملية الإشعاع. كما هو موضح في الشكل 4c، على الرغم من أن التأثير الضوئي الحراري يضعف معدل إنتاج الأكسجين في GHEA، يمكن الكشف عن إنتاج أكسجين ثابت ومستقر. تعتبر قدرة GHEA على إنتاج الأكسجين على مدى فترة طويلة من علاج شفاء الجروح أمرًا حاسمًا لفعاليتها. وفقًا لعملية إنتاج الأكسجين لـ GHEA التي تم محاكاتها في المختبر على مدى 20 يومًا، تميل قدرة خلايا GHEA على إنتاج الأكسجين إلى الضعف ببطء مع مرور الوقت خلال الإشعاع المتكرر بالليزر. ومع ذلك، بعد 20 يومًا من الإشعاع المتقطع بالليزر، لا تزال خلايا GHEA تحتفظ بقدرة جيدة على إنتاج الأكسجين (الشكل التكميلي 9). تشارك أنشطة خلوية متعددة، مثل تكاثر الخلايا الليفية، وهجرة الخلايا الكيراتينية، وتمايز خلايا بطانة الأوعية الدموية، بشكل متزامن في شفاء الجروح؛ لذلك، قمنا بمزيد من فحص تأثير GHEA@Gel على الخلايا الليفية الجلدية البشرية (HSFs)، والخلايا الكيراتينية البشرية الخالدة (HaCaTs)، وخلايا بطانة الأوعية الدموية السليمة من الحبل السري البشري (HUVECs). اكتشفنا أولاً أن التعرض لـ
الشكل 3 | تحويل الحرارة الضوئية والتعقيم لـ GHEA@Gel. أ رسم تخطيطي لتحويل الحرارة الضوئية والتعقيم لـ GHEA@Gel. ب التحول الحراري الضوئي لـ GHEA@Gel. تُعرض البيانات كمتوسط س.د. ( تجارب مستقلة).القياس الكمي لـخلايا الإشريكية القولونية المعالجة بـ GHEA ولايزر 658 نانومتر ). يتم تقديم البيانات كمتوسط س.د. ( الخلايا المستقلة بيولوجيًا). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار. هـ، القياس الكمي لـخلايا المكورات الذهبية المعالجة بـ GHEA وليزر 658 نانومترتُعرض البيانات على أنها
المتوسط س.د. ( الخلايا المستقلة بيولوجيًا). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار. ج الصور الرقمية المقابلة لـ. أوريوس ومستعمرات بكتيريا الإشريكية القولونية التي نمت على أطباق أجار LB وتم تعريضها لمعاملة مختلفة. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة.الصور المجهرية الإلكترونية الماسحة المقابلة لبكتيريا S. aureus و E. coli التي خضعت لعلاجات مختلفة. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. صور الفوتوحرارية للفئران التي تلقت علاجات مختلفة. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة.
33 مللي مول من الجلوكوز ونقص الأكسجين المستحث المصاحب لزيادة التعبير عن عامل نقص الأكسجين ( )، مما يدل على تأثير فرط سكر الدم على نقص الأكسجة (الشكل 4d والأشكال التكميلية 10-13). بعد ذلك، قمنا بتحديد ما إذا كان GHEA@Gel يمكن أن يعكس نقص الأكسجة في الخلايا بشكل كبير ويؤدي إلى خفض في HIF-1في HSFs (الشكل 4e). لوحظ نفس التأثير على HUVECs وHaCaTs المعالجة بـ GHEA@Gel (الأشكال التكميلية 10-13). في بالإضافة إلى التحقق من صحة بيانات الفلورية النوعية، فإن التحليل الكمي عبر مقياس التدفق الخلوي مرغوب فيه. كما هو موضح في الشكل التكميلي 14، أظهرت بيانات مقياس التدفق الخلوي اتجاهًا مشابهًا لنتائج تصوير الفلورية. أظهرت خلايا HSF المعالجة بـ GHEA@Gel تحت ظروف الضوء معدل تكاثر أكبر بشكل ملحوظ مقارنة بتلك المعالجة بالجلوكوز العالي أو حتى (الشكل 4f، g)، مما يشير إلى الإمكانات الكبيرة لـ GHEA@Gel في تشكيل نسيج التندب
الشكل 4 |إصدار من GHEA@Gel. تمثيل تخطيطي لـإصدار من GHEA@Gel. ب مقارنة الذائبإصدار GHEA@Gel تحت ظروف الضوء والظلام. يتم تقديم البيانات كمتوسط س.د. ( تجارب مستقلة).إطلاق المذاب عند تركيزات مختلفة من GHEA وشدات الليزر. د، هـ GHEA@Gel قلل من HIF-1 التعبير في HSFs الناتج عن تركيزات عالية من الجلوكوز. قضبان القياس، . تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. تُعرض البيانات كمتوسط س.د. ( الخلايا المستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار. ف، ج تكاثر خلايا HSF المعالجة بـ 33 مليمول/لتر من الجلوكوز تحت نقص الأكسجين (6 ساعات) لفترات زمنية مختلفة. شريط القياس،. كانت كل تجربة تم تكرارها بشكل مستقل ثلاث مرات مع نتائج مشابهة. تُعرض البيانات كمتوسط س.د. ( الخلايا المستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار.صور تمثيلية وقياس لهجرة خلايا HaCaT. القضبان المقياسية هي. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. تُعرض البيانات كمتوسط س.د. ( خلايا مستقلة).صور تمثيلية وقياس لتكوين أنابيب HUVEC. مقياس الأشرطة،. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. تُعرض البيانات كمتوسط س.د. ( الخلايا المستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار. من خلال إطلاق الأكسجين المذاب لتعزيز تكاثر HSF. بالإضافة إلى ذلك، عزز GHEA@Gel تكاثر الخلايا في HUVECs وHaCaTs (الأشكال التكميلية 15، 16). يعتبر شفاء الخدوش في الكيراتينوسيتات جوهر عمليات إعادة الظهارة خلال شفاء الجروح المزمنة. في اختبار الخدش، قمنا بمقارنة هجرة الخلايا تحت ظروف نقص الأكسجين،ظروف الإمداد، والأكسجين القابل للذوبان الناتج عن GHEA@Gel تحت إشعاع الضوء. الشكل 4h و i يظهران أنه، بالمقارنة معفي بيئة نقص الأكسجين، تم تثبيط هجرة خلايا HaCaT بشكل كبير. ومن الجدير بالذكر أن زراعة الخلايا مع GHEA@Gel في بيئة نقص الأكسجين والتعرض اللاحق للضوء زاد بشكل كبير من معدل هجرة الخلايا، مما يشير إلى أن الأكسجين القابل للذوبان الذي تنتجه GHEA@Gel يمكن أن يعزز التقرن، وبالتالي يسهل شفاء الجروح المزمنة. تعتبر الخلايا البطانية جزءًا لا يتجزأ من الأوعية الدموية وتلعب دورًا حاسمًا في تكوين الأوعية. قمنا بإجراء اختبارات تشكيل الأنابيب باستخدام المجهر. أظهر اختبار تشكيل الأنابيب باستخدام Matrigel، الذي تم إجراؤه لتقييم القدرة على تكوين الأوعية، طول أنبوب وعدد فروع أكبر بشكل ملحوظ بعد 6 ساعات في خلايا HUVEC المعالجة بـ GHEA@Gel مقارنةً بـ-تم معالجة خلايا HUVECs (الشكل 4j، k). أكدت نتائج سلسلة من التجارب المخبرية أن الزيادة في تكوين الأوعية الدموية كانت ناتجة عن الأكسجين القابل للذوبان الذي تنتجه GHEA@Gel.
استغلال الجذور الحرة بواسطة RHEA@Gel
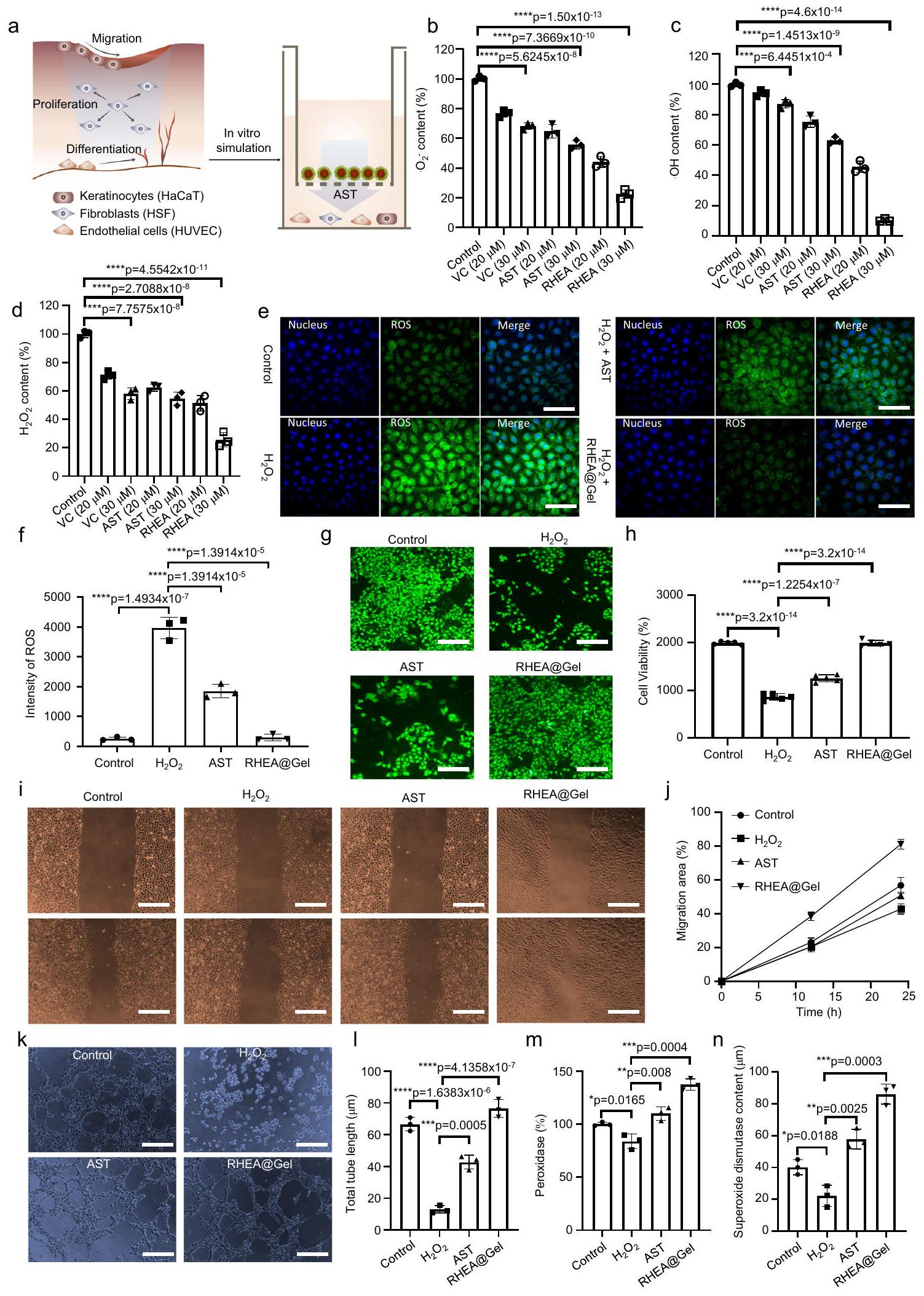
تتميز جروح السكري بمستويات مرتفعة من الجذور الحرة للأكسجين وإجهاد أكسدي مستمر. لقد أظهرت الدراسات السابقة أن إضافة مضادات الأكسدة إلى الهلاميات المائية يقلل من تلف الخلايا الناتج عن الأكسدة، مما يسرع من شفاء الجروح المزمنة. نظرًا للبنية الجزيئية الفريدة لـ AST، فإن الهلاميات المائية المعتمدة على RHEA تعد واعدة لتعزيز شفاء جروح السكري. AST هو أقوى مضاد أكسدة طبيعي (الشكل 5أ). إزالة، و تم قياسه لاحقًا لتحديد الأداء المضاد للأكسدة لـ RHEA@Gel (الشكل 5ب-د). لمقارنة قدرة RHEA@Gel على القضاء على أنواع الأكسجين التفاعلية بدقة أكبر، تم اختيار مضاد الأكسدة الطبيعي فيتامين C (VC) وAST الكيميائي الاصطناعي للمقارنة. كما هو موضح في الشكل 5ب-د، أظهر RHEA@Gel الذي قمنا بتحضيره أقوى قدرة على القضاء على ROS في شكل، و بنفس التركيز. باستخدام مجهر الفلورية، تم تأكيد قدرة RHEA@Gel على التخلص من الجذور الحرة. لمحاكاة البيئة الميكروبية الالتهابية للجروح السكرية، تم اختياره لمزيد من التجارب. على وجه التحديد،تمت إضافة المحلول خلال عملية زراعة خلايا HSF. بعد ذلك، بعد إضافة مادة إزالة الجذور الحرة (AST أو RHEA@Gel)، تم حضن الخلايا لفترة معينة من الوقت لقياس مستوى الجذور الحرة داخل الخلايا. الشكلتوضح أن RHEA@Gel هو منظم فعال لمستويات ROS داخل الخلايا في HSFs. من الواضح من النتائج أنه بعدتمت إضافة إلى زراعة الخلايا، كان هناك زيادة ملحوظة في مستويات ROS داخل الخلايا، كما يتضح من زيادة شدة الفلورية الخضراء. ومع ذلك، عندما تم إضافة AST أو RHEA@Gel إلى الخلايا التي تم حضنها مع، انخفضت تركيزات ROS داخل الخلايا مقارنة بتلك في مجموعة. بالإضافة إلى ذلك، لوحظت ظواهر مماثلة لخلايا HUVECs وHaCaTs (الأشكال التكميلية 17، 18). تشير هذه النتائج إلى أن قدرة RHEA@Gel على امتصاص الجذور الحرة ليست فقط أكبر بكثير من تلك الخاصة بفيتامين C الطبيعي، ولكنها أيضًا أكبر من تلك الخاصة بالفيتامين C المصنع كيميائيًا.
AST. الأسباب التالية هي التي أدت إلى هذه النتيجة. أولاً، AST هو مضاد أكسدة غير قطبي يتجمد في المحاليل المائية والسوائل البيولوجية، مما يقلل من توافره الحيوي. ثانياً، يرتبط نشاط مضادات الأكسدة لـ AST بتركيبه الجزيئي، كما أن التركيب الجزيئي غير الصحيح لـ AST المُصنّع كيميائياً يؤثر أيضاً على نشاطه كمضاد أكسدة. على الرغم من أن AST الطبيعي الذي تنتجه RHEA مُشكل بشكل مثالي، إلا أن خصائصه غير القطبية تؤثر أيضاً على قدرته كمضاد أكسدة. في التوصيف الأولي، تم تحديد أن تحميل بروتوبلاست RHEA في الهيدروجيل أنتج العديد من الحويصلات المغلفة بالغشاء تقريباًفي الحجم، مشابهة للإكسوزومات (الشكل 2g، h). السبب في أن RHEA@Gel لديه أقوى نشاط مضاد للأكسدة هو على الأرجح بسبب التكوين الدقيق لـ AST المفرز طبيعياً، ووجود حويصلات الإكسوزوم المغلفة بالخلايا في RHEA@Gel يزيد بشكل كبير من قابلية ذوبان AST في الماء ويعزز توافره البيولوجي.
استنادًا إلى هذه النتائج، تم التحقيق بشكل أكبر في تأثير RHEA@Gel على تكاثر HSF، وقد أدى RHEA@Gel إلى زيادة كبيرة في معدل التكاثر في HSFs مقارنةً بـ و AST (الشكل 5g، h)، مما يشير إلى أن AST المفرج عنه من RHEA@Gel لديه إمكانيات كبيرة لتشكيل نسيج حبيبي من خلال تعزيز تكاثر HSF. بالإضافة إلى ذلك، تم ملاحظة قدرة RHEA@Gel على تعزيز تكاثر الخلايا أيضًا بالنسبة لـ HUVECs و HaCaTs (الأشكال التكميلية 19، 20). بعد ذلك، قمنا بفحص تأثير AST من RHEA@Gel على خلايا HaCaT المستحثة بواسطة ROS. حيث أن ROS قد أوقف بشكل كبير هجرة خلايا HaCaT، بينما زادت تحفيزات RHEA@Gel بشكل كبير من هجرة خلايا HaCaT (الشكل 5i، j). كان لـ RHEA@Gel و AST تأثيرات كبيرة على هجرة الخلايا. قمنا بإجراء اختبارات تشكيل الأنابيب باستخدام المجهر. أظهر اختبار تشكيل الأنابيب باستخدام Matrigel، الذي تم إجراؤه لتقييم القدرة على تكوين الأوعية، طول أنبوب أكبر وعدد أكبر من الفروع بعد 6 ساعات في HUVECs المعالجة بـ RHEA@Gel مقارنة بـ HUVECs المعالجة بـ AST (الشكل 5k، l). بالإضافة إلى ذلك، اكتشفنا أن AST المفرج عنه من RHEA@Gel يمكن أن يزيد من تعبير البيروكسيداز و SOD في الخلايا، مما يساعد في التخلص من ROS داخل الخلايا. تشير هذه النتائج إلى أن الزيادة في تكوين الأوعية التي لوحظت في سلسلة من الاختبارات في المختبر كانت نتيجة لإفراز AST من RHEA@Gel.
خصائص مضادة للالتهابات لـ RHEA@Gel
البلاعم هي نوع مهم من خلايا المناعة في جهاز المناعة تعمل من خلال البلعمة، السمية الخلوية، وإفراز السيتوكينات. يمكن تنشيط هذه الخلايا وتوجيهها إلى أنماط ظاهرة مختلفة للتكيف مع ظروف فسيولوجية ومرضية متنوعة. تمثل البلاعم M1 نمطًا ظاهريًا التهابيًا نموذجيًا وتلعب دورًا حاسمًا في المناعة المضادة للميكروبات والأورام. يتم تنشيط هذه السيتوكينات بشكل أساسي بواسطة مكونات ميكروبية (مثل الليبوسكرياتيد) وسيتوكينات (مثل IFNتقوم البلعميات M1 بقتل الكائنات الدقيقة المسببة للأمراض من خلال إنتاج جزيئات ضارة مثل أنواع الأكسجين والنيتروجين التفاعلية وإفراز السيتوكينات المؤيدة للالتهابات، مما يؤدي إلى تحفيز استجابات التهابية محلية. تمثل البلعميات M2 نمطًا غير التهابي وتشارك بشكل أساسي في إصلاح الأنسجة وتجديدها وتعديل الاستجابات المناعية. يتم تنشيط البلعميات M2 بشكل رئيسي بواسطة السيتوكينات (مثل الإنترلوكين-4 والإنترلوكين-13) وجزيئات أخرى (مثل VEGF وعامل النمو المحول بيتا). تمارس البلعميات M2 تأثيراتها من خلال قمع الالتهاب، وتعزيز إصلاح الأنسجة، وتنظيم توازن المناعة. يتأثر تنظيم هذه الأنماط الماكروفاجية بشكل رئيسي بواسطة مستقبلات ومسارات إشارة مختلفة. على سبيل المثال، يمكن لمستقبلات Toll-like، والمستقبلات النووية، ومُستقبلات السيتوكين، وغيرها، تنشيط مسارات إشارة محددة لتنظيم استقطاب الماكروفاج ووظائفه. كما أن أنواعًا مختلفة من مسببات الأمراض، وإشارات تلف الخلايا، وجزيئات تنظيم المناعة أيضًا المشاركة في هذه العملية، مما يشكل شبكة إشارات معقدة. اعتمادًا على دور أنماط الماكروفاج المختلفة، يجب استقطاب ماكروفاجات إضافية إلى النمط M2 خلال شفاء الجروح. ومع ذلك، فإن التحول الظاهري من M1 إلى M2 يتعرض للخطر في الجروح السكرية، مما يؤدي إلى التهاب مستمر وشفاء بطيء للجروح. AST هو مركب طبيعي مستخرج من HEA وله خصائص مضادة للالتهابات. لقد سعينا إلى
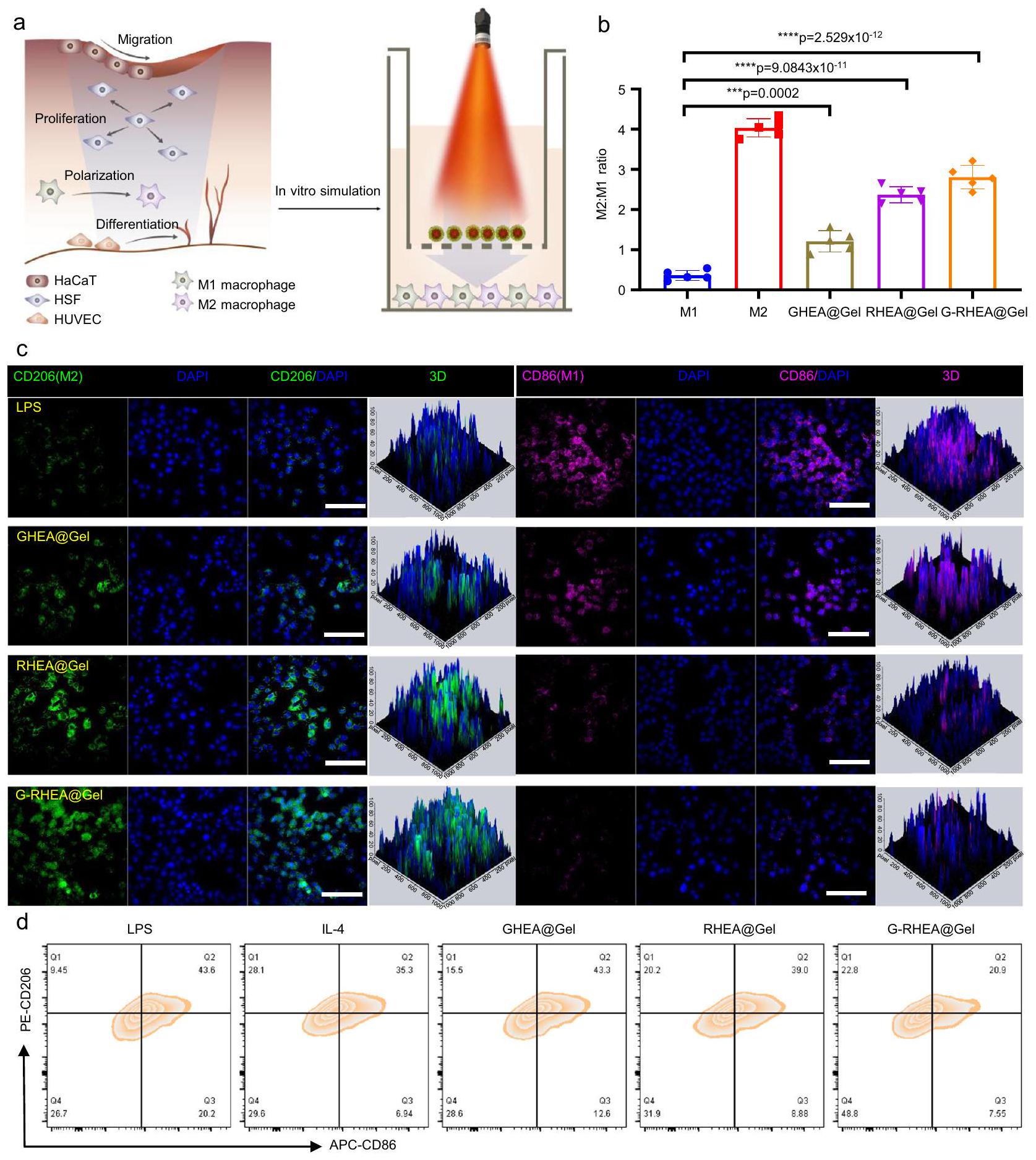
الشكل 5 | قدرة RHEA@Gel على التخلص من الجذور الحرة. أ تمثيل تخطيطي للتخلص من الجذور الحرة بواسطة RHEA@Gel. ب، (ج) OH -، و (د) -أداء التقاط الجذور الحرة استجابةً لعلاجات مختلفة. تُعرض البيانات كمتوسط س.د. ( الخلايا المستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبيناختبار. قدرة RHEA@Gel على التخلص من ROS في HSF. قضبان القياس،. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. تُعرض البيانات كمتوسط س.د. ( الخلايا المستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار.انتشار خلايا HSF المعالجة بـ 33 مللي مول من الجلوكوز وفي مجموعات مختلفة. قضبان القياس،. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. تُعرض البيانات كمتوسط س.د. ( خلايا مستقلة). الفروق الإحصائية تم تحليلها بواسطة اختبار الطالب ذو الجانبين-اختبار.صور وقياس هجرة خلايا HaCaT. قضبان القياس،. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. تُعرض البيانات كمتوسط س.د. ( خلايا مستقلة).صور تمثيلية وتحليل كمي لتكوين الأنابيب في خلايا HUVECs. قضبان القياس،. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. تُعرض البيانات كمتوسط س.د. ( خلايا مستقلة).البيروكسيداز و(تعبير سوبر أكسيد ديسموتاز في المجموعات المختلفة. تُعرض البيانات كمتوسط س.د. ( الخلايا المستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار. تحقيق ما إذا كان تأثير AST المضاد للالتهابات ناتجًا عن تنظيم انتقال النمط الظاهري للبلاعم وتعديل البيئة الدقيقة المناعية في الجروح (الشكل 6أ). باختصار، تم تحفيز البلاعم M0 بواسطة الليببوليسكاريد (LPS) والإنترفيرونإنترفيرونللتبديل إلى النمط الظاهري M1 كتحكم إيجابي، بينما تم تحفيز تلك الخلايا باستخدام الإنترلوكين-4 (IL-4) للتبديل إلى النمط الظاهري M2 كتحكم سلبي. تم تحديد البلعميات CD206 (النمط الظاهري M2) وCD86 (النمط الظاهري M1) من خلال صبغة المناعية الفلورية لتأكيد حالة الاستقطاب للبلعميات تحت ظروف التحفيز المختلفة. تم الكشف عن المزيد من البلعميات CD206+ وأقل من البلعميات CD86+ في مجموعة GHEA@Gel مقارنة بمجموعة التحكم، مما يشير إلى أن قد يكون الإفراز قد زاد من تنشيط M2 (الشكل 6ب، ج والشكل التكميلي 21). مقارنةً بالمجموعات الضابطة وGHEA@Gel، أظهرت البلعميات المعالجة بـ RHEA@Gel مستويات أعلى بشكل ملحوظ من CD206 وأقل من CD86، مما يشير إلى أن AST المفرج عنه من RHEA ضروري لاستقطاب البلعميات. علاوة على ذلك، أظهرت مجموعة GRHEA@Gel أعلى تلوين لـ CD206 وأقل إشارة لـ CD86، مما يثبت أكثر أنأدت عملية التحرير وAST بشكل متزامن إلى تحفيز استقطاب M2 للخلايا البالعة. للتحقق بشكل أكبر من قدرة HEA على تعزيز تحول الخلايا البالعة M1 إلى خلايا بالعة M2، تم استخدام تحليل تدفق الخلايا للكشف عن الخلايا البالعة في مجموعات العلاج المختلفة. كانت النتائج الموضحة في الشكل 6d متوافقة مع نتائج المجهر الفلوري (الشكل 6d والشكل التكميلية 22).
التقييم الحي لشفاء الجروح المصابة لدى مرضى السكري
يتم تصوير النهج العلاجي في الشكل 7أ. الاستفادة من تأثير التحويل الضوئي الحراري لـ HEA@Gel تحت إشعاع ليزر بزاوية 658 نانومتر عند كثافة وتأثير عدم التعايش بين HEA و S. aureus، تم تعقيم الجرح المصاب بالسكري أولاً. بعد العلاج القاتل للبكتيريا، تم استخدام ليزر بطول موجي 658 نانومتر لإضاءة الجرح كل يوم لاحق عند شدة ضوء لمدة 30 دقيقة في الأيام العشرة التالية. للإشعاع بالليزر منخفض الكثافة تأثيران. أولاً، يقوم GHEA بتنشيط عملية التمثيل الضوئي لإنتاج لتحسين البيئة الميكروية للجروح الناجمة عن نقص الأكسجين وتسريع تجديد الأوعية الدموية وشفاء الجروح. والثاني هو تعزيز إطلاق AST من RHEA لتقليل مستويات ROS والعوامل الالتهابية المفرطة التعبير وتنظيم استقطاب البلعميات M2 في الجروح. ومن ثم، من المحتمل أن يكون HEA@Gel قد عزز تكاثر الخلايا الليفية، وهجرة الخلايا الكيراتينية، وتمايز الخلايا البطانية، والتسريع التدريجي لشفاء الجروح المصابة لدى مرضى السكري.
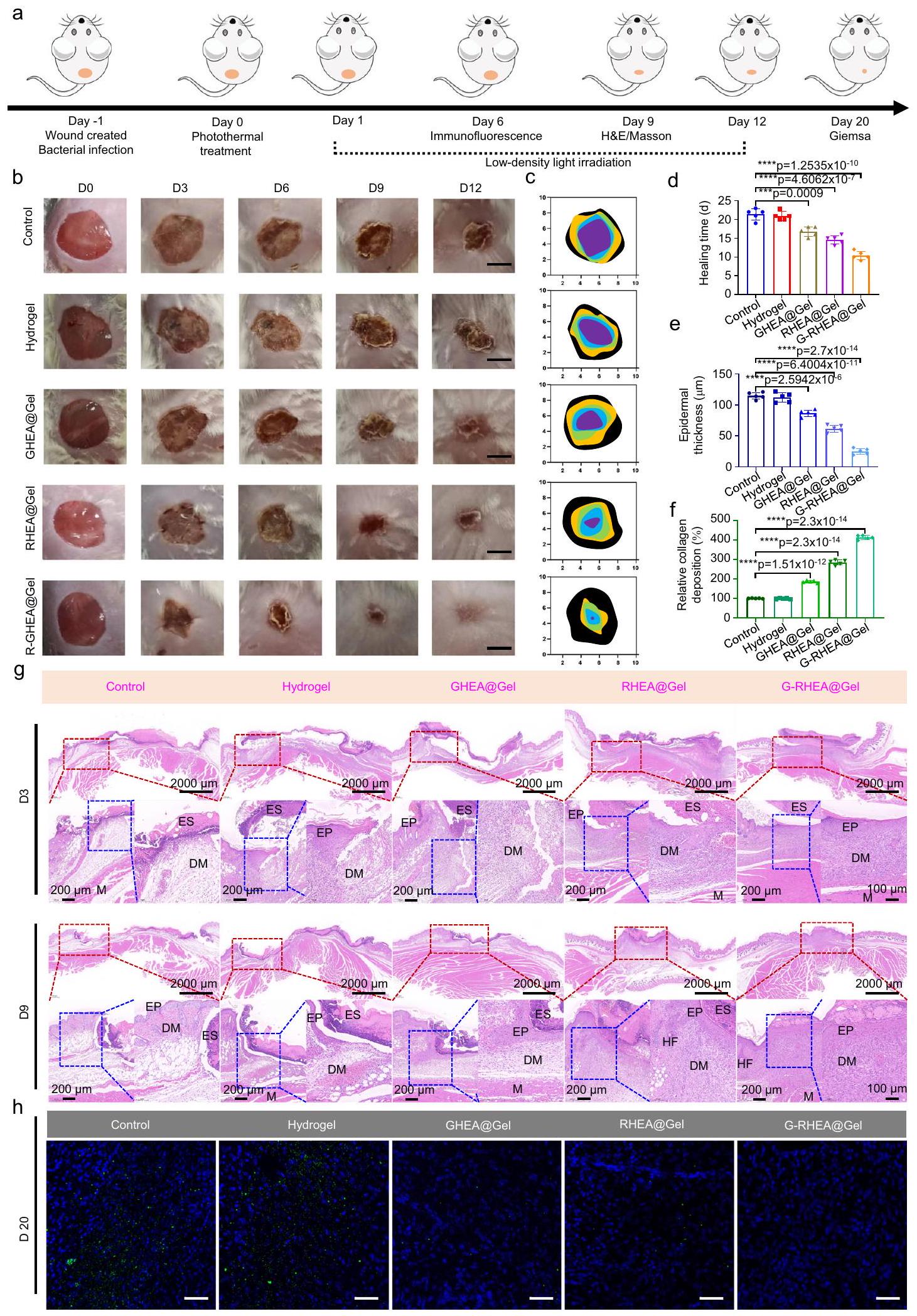
شكلتظهر الصور الماكروسكوبية للجروح، آثار إغلاق الجروح في نقاط زمنية مختلفة، وأوقات الشفاء الكمي للجروح المقابلة. في اليوم الثالث، كانت الجروح في مجموعة GRHEA@Gel مع الليزر أصغر من تلك الموجودة في المجموعات الأخرى. في اليوم السادس، انخفضت مساحة الجرح في مجموعة G-RHEA@Gel مع الليزر بشكل ملحوظ، بينما ظلت أحجام الجروح في مجموعات التحكم، Gel، GHEA@Gel، وRHEA@Gel مع الليزر عند، ، و ، على التوالي. في اليوم الجروح المصابة في المجموعة المعالجة بـ G-RHEA@Gel والليزر قد أغلقت. بالمقارنة، كانت هناك مناطق جروح كبيرة لا تزال موجودة في مجموعتي التحكم والهلام، بينما كانت مجموعتا GHEA@Gel و RHEA@Gel تشهدان شفاءً أسرع. أظهرت النتائج أن العلاج بـ G-RHEA@Gel وإشعاع الليزر عزز شفاءالجروح المصابة بالبكتيريا الذهبية لدى مرضى السكري.
كما هو موضح في الشكل 7g، تم إجراء الفحص النسيجي للجروح الملتئمة باستخدام صبغة الهيماتوكسيلين-إيوزين (H&E) وصبغة ماسون الثلاثية (MT). في اليوم الثالث، لم تظهر أي من المجموعات تكوين البشرة أو ترسب الكولاجين. جميع المجموعات أظهرت تجديد البشرة في اليوم التاسع، لكن البشرة في مجموعة GRHEA@Gel مع الليزر كانت أكثر طبيعية ونضجًا من تلك الموجودة في المجموعات الأخرى. كما هو موضح في الشكل 7e، كانت سماكات البشرة في مجموعات التحكم، Gel، GHEA@Gel، RHEA@Gel، وG-RHEA@Gel ، و “، على التوالي. أظهر صبغ H&E أن الجروح المصابة لدى مرضى السكري في مجموعة G-RHEA@Gel المعالجة بالليزر أظهرت شفاءً مرضيًا للجروح مع تسريع في تكوين البشرة والأدمة. بالإضافة إلى ذلك، تم عرض مستوى ترسب الكولاجين في اليوم التاسع في الشكل 7f. كان ترسب الكولاجين أكبر في مجموعة GRHEA@Gel المعالجة بالليزر مقارنةً بالمجموعات الأخرى. أشارت نتائج صبغ H&E وMT إلى أن معالجة G-RHEA@Gel باستخدام ليزر 658 نانومتر يمكن أن تسرع شفاء الجروح المصابة لدى مرضى السكري من خلال تعزيز تكوين البشرة، وتسهيل تجديد الأنسجة الجلدية، وزيادة ترسب الكولاجين. بالإضافة إلى ذلك، تم استخدام صبغ المناعية الكيميائية باستخدام جسم مضاد ضد المكورات العنقودية الذهبية لتقييم التلوث البكتيري للجروح. كما هو موضح في الشكل 7h، في اليوم 20، احتوى نسيج الجروح في مجموعة التحكم ومجموعة الهلام النقي على عدد كبير من البكتيريا (فلوروسينس أخضر). عندما تم دمج HEA@Gel مع إشعاع ليزر 658 نانومتر، كان من الصعب بشكل خاص اكتشاف البكتيريا في مجموعات GHEA@Gel وRHEA@Gel وG-RHEA@Gel بسبب التأثير المضاد للبكتيريا القوي للعلاج. يمكن أن يُعزى التأثير المضاد للبكتيريا الممتاز الذي تموسطه HEA بشكل رئيسي إلى التأثير غير التبادلي بين البكتيريا وHEA وتأثير HEA القاتل للبكتيريا عند الإشعاع باستخدام ليزر 658 نانومتر. للتحقق من هذا الاستنتاج بشكل أكبر، تم جمع جلود النباتات التي خضعت لعلاجات مختلفة وتجانست في اليوم 20. تم زرع المعلقات المخففة بالتسلسل على أجار LB لتحديد عدد المستعمرات البكتيرية. كما هو موضح في الشكل التكميلية 23، ظهرت عدد كبير من مستعمرات S. aureus في مجموعة التحكم ومجموعة الهلام. كان عدد المستعمرات في مجموعة GHEA@Gel المعالجة بالليزر 658 نانومتر ( ) الإشعاع ومجموعة RHEA@Gel المعالجة بالليزر 658 نانومتر ( تم تقليل تأثير الإشعاع بشكل كبير بسبب التأثير البكتيري الحراري الضوئي والتأثير غير التبادلي، على التوالي. لذلك، في ضوء التأثيرين المضادين للبكتيريا المذكورين أعلاه، لم يكن هناك تقريبًا أي تشكيل مستعمرات في مجموعة G-RHEA@Gel بعد العلاج.
تحليل المناعة الفلورية
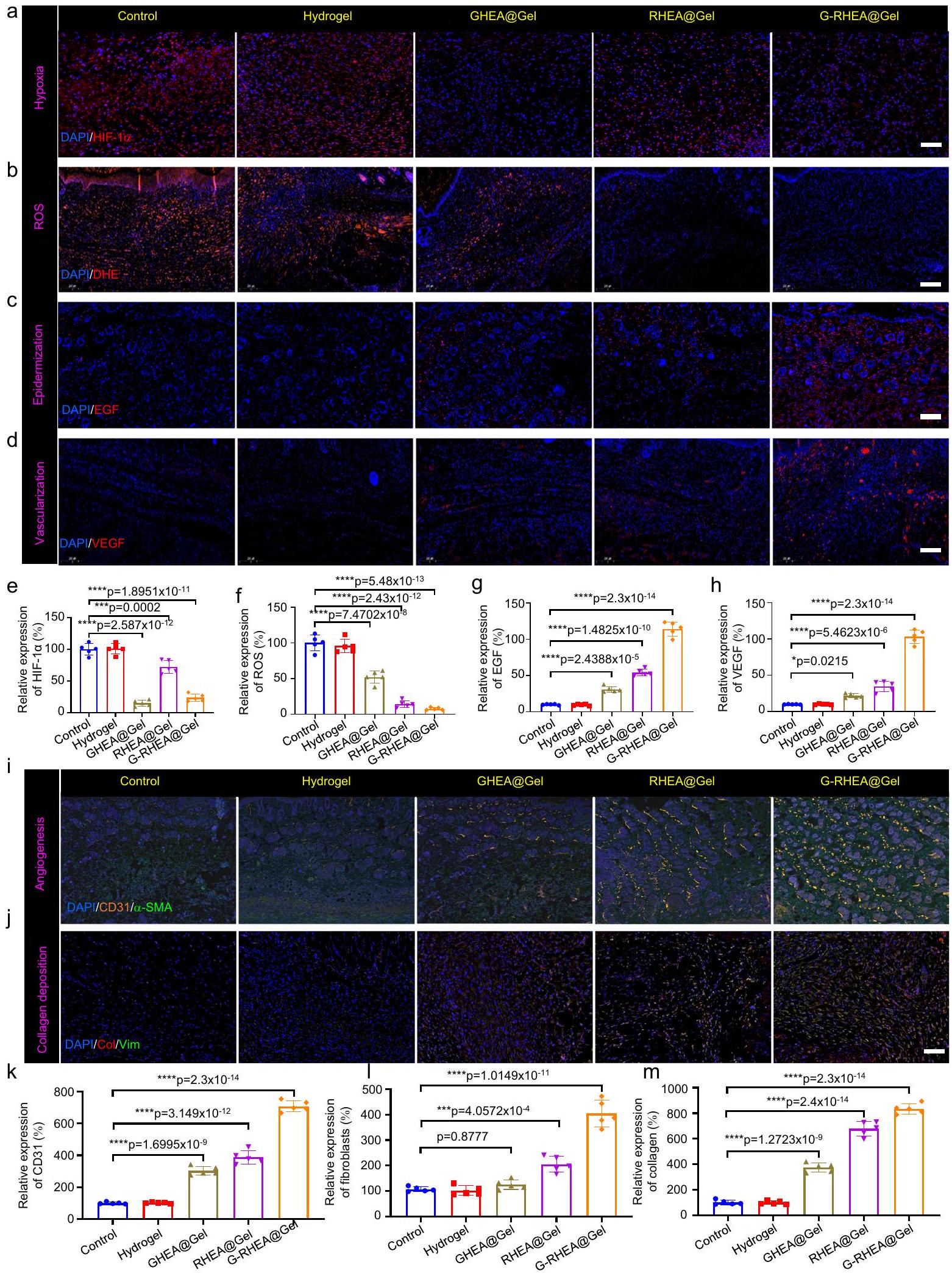
لإظهار فعالية HEA@Gel، تم فحص قدرة GHEA على تعزيز شفاء الجروح من خلال HIF-1تلطيخ المناعة الفلورية. بالنسبة لمجموعتي التحكم والهلام، كان هناك HIF-1 كبيرتم ملاحظة التعبير. بسبب عملية التمثيل الضوئي الفعالة لـ GHEA، فإن HIF-1كان مستوى GHEA@Gel أقل بكثير من المستوى في مجموعة التحكم، مما يشير إلى إمداد أكسجين أكبر بشكل ملحوظ (الشكل 8a، e). بالمقارنة مع أولئك في مجموعة GHEA@Gel، كان إمداد الأكسجين في مجموعة RHEA@Gel أقل بسبب تراكم AST العالي وتقليل عملية التمثيل الضوئي. تقريبًا لا يوجد HIF-1تم الكشف عن التعبير في
الشكل 6 | تنظيم استقطاب البلعميات بواسطة RHEA@Gel. أ رسم توضيحي تخطيطي لتنظيم استقطاب البلعميات بواسطة RHEA@Gel. ب الكمية النسبية لنسبة البلعميات M2 إلى M1. تُعرض البيانات كمتوسط س.د. ( الخلايا المستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار. صور الفلورسنت المميزة لخلايا Raw264.7
ملطخة بـ CD206 (أخضر) و CD86 (وردي) تحت التحفيز الالتهابي. مقياس الرسم:تحليل تدفق الخلايا لتحديد استقطاب البلعميات بعد علاجات مختلفة. تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. مجموعة G-RHEA@Gel، مما يؤكد على الإمداد العالي بالأكسجين من GRHEA@Gel. من خلال إطلاق الأكسجين المذاب، كان GHEA@Gel مع إشعاع ليزر 658 نانومتر قادرًا على تخفيف نقص الأكسجين المحلي. لفحص قدرة GHEA@Gel على القضاء على ROS في الجسم الحي، تم استخدام مجس دihydroethidium (DHE) لقياس تركيز ROS في منطقة الجرح. أظهرت مجموعتا التحكم والهلام إشارات فلورية حمراء ساطعة وعالية الكثافة، بينما انخفضت شدة الفلورية بشكل ملحوظ في مجموعة GHEA@Gel، كما هو موضح في الشكل 8b، f. في
لم يتم الكشف عن أي فلوريسنس أحمر تقريبًا في مجموعات RHEA@Gel و G-RHEA@Gel. وفقًا لهذه النتائج، فإن RHEA@Gel قادر على التخلص بكفاءة من الجذور الحرة لتخفيف الإجهاد التأكسدي.
كما هو موضح في الشكل 8c و g، تم التحقيق في الآليات البيولوجية الأساسية لشفاء الجروح من خلال صبغة المناعية لعامل نمو البشرة (EGF) في اليوم السادس. تم ملاحظة تعبير EGF في كل من مجموعتي GHEA@Gel و RHEA@Gel، مما يشير إلى أن كل من قدرة GHEA@Gel على توفير الأكسجين وقدرتها على القضاء على الجذور الحرة.
الشكل 7 | تأثيرات HEA@Gel على تعزيز شفاء الجروح في الحالات المصابة
فئران مصابة بالسكري. أ رسم تخطيطي يوضح تسلسل التجارب الحيوانية التي أجريت لتقييم الفعالية العلاجية لـ HEA@Gel. ب صور لجروح تمثيلية من مجموعات علاجية مختلفة. ج تحقيق كمي لمساحة الجرح. تم تكرار كل تجربة خمس مرات بشكل مستقل مع نتائج مشابهة. د تحليل إحصائي لوقت الشفاء. تُعرض البيانات كمتوسط. س.د. ( فئران مستقلة). تم تحليل الفروق الإحصائية بواسطة
وجهين للطالب-اختبار. e، f التحليل الكمي النسبي لترسب الكولاجين وسمك البشرة في اليوم التاسع. يتم تقديم البيانات كمتوسط س.د. ( فئران مستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبيناختبار. صبغة HE في الأيام 3 و 9. تم تكرار كل تجربة خمس مرات بشكل مستقل مع نتائج مشابهة.تلوين المناعية النسيجية للجروح في اليوم العشرين. مقياس الشريط هوتم تكرار كل تجربة خمس مرات بشكل مستقل مع نتائج مشابهة.
الشكل 8 | تحليل العمليات والآليات لشفاء الجروح لمختلف العلاجات. أ HIF-1 التعبير، (ب) محتوى ROS، (ج) تعبير EGF، و (د) تعبير VEGF في اليوم السادس تحت علاجات مختلفة. مقياس الشريط هو تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة.تحقيق كمي لـ HIF-1تعبير ROS و EGF و VEGF في اليوم السادس استجابةً لعلاجات مختلفة. تُعرض البيانات كمتوسط س.د. ( فئران مستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبيناختبار. صبغة المناعة المزدوجة باستخدام الفلورسنت لـ (i)-SMA (أصفر) و CD31 (أخضر) و (ج) علامات الخلايا الليفية الفيمنتين (أخضر) والكولاجين (أحمر) استجابةً لعلاجات مختلفة في اليوم السادس. مقياس الرسم،تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة.تحقيق كمي في تعبير CD31، وتكوين الأوعية الدموية الجديدة، وترسيب الكولاجين I في اليوم السادس استجابةً لعلاجات مختلفة. تُعرض البيانات كمتوسط. س.د. ( فئران مستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار. قد تعزز قدرة RHEA@Gel تعبير EGF. لوحظ أعلى تعبير لـ EGF في مجموعة علاج G-RHEA@Gel، مما يشير إلى أن إمداد الأكسجين والتخلص من ROS هما شرطان ضروريان لتكوين البشرة. بالإضافة إلى ذلك، يعتبر VEGF علامة هامة في مجرى HIF، ويقوم نقص الأكسجين بتثبيط تعبير VEGF. تُظهر الشكل 8d، h نتائج صبغة المناعية لـ VEGF، والتي كشفت عن مستوى أعلى من VEGF في مجموعة G-RHEA@Gel مقارنةً بمجموعة التحكم، مما يشير إلى زيادة في تكوين الأوعية الدموية بسبب الهيدروجيل، وإمداد الأكسجين، والتخلص من ROS.
تعتبر تكوين الأوعية الدموية مؤشرًا حاسمًا على شفاء الجروح لدى مرضى السكري. تم تقييم تكوين الأوعية الدموية من خلال صبغة المناعية للـ a-أكتين العضلي الملساء و CD31. كما هو موضح في الشكل 8i و k، كانت تعبيرات CD31 في مجموعتي GHEA@Gel و RHEA@Gel أكبر بكثير من تلك الموجودة في مجموعة التحكم ومجموعة الجل. كان تعبير CD31 هو الأعلى في مجموعة G-RHEA@Gel. أظهرت النتائج أن G-RHEA@Gel المعالج بالإشعاع بالليزر يمتلك قدرة تفوق في تكوين الأوعية الدموية وعزز شفاء الجروح بشكل أكثر فعالية من الهلامات الأخرى. لذلك، يمكن تسريع شفاء الجروح لدى مرضى السكري من خلال تعزيز تكوين البشرة، وترسيب الكولاجين، وتكوين الأوعية الجديدة. بالإضافة إلى ذلك، تم إجراء صبغة مناعية للكولاجينات والفيمينتين (علامة الخلايا الليفية) لتقييم ترسيب الكولاجين في الأنسجة الحبيبية (الشكل 8j و l و m). تم تسهيل تكوين ECM من خلال زيادة ترسيب المصفوفة والمحاذاة الاتجاهية لألياف الكولاجين. كانت مجموعة التحكم تحتوي على أقل كمية من الكولاجين، بينما كانت مجموعة GRHEA@Gel تحتوي على مستويات أعلى من ترسيب الكولاجين والمحاذاة الاتجاهية، مما يشير إلى إنتاج أفضل للكولاجين مما يعزز إصلاح الجروح.
تنظيم في الجسم لقطبية البلعميات
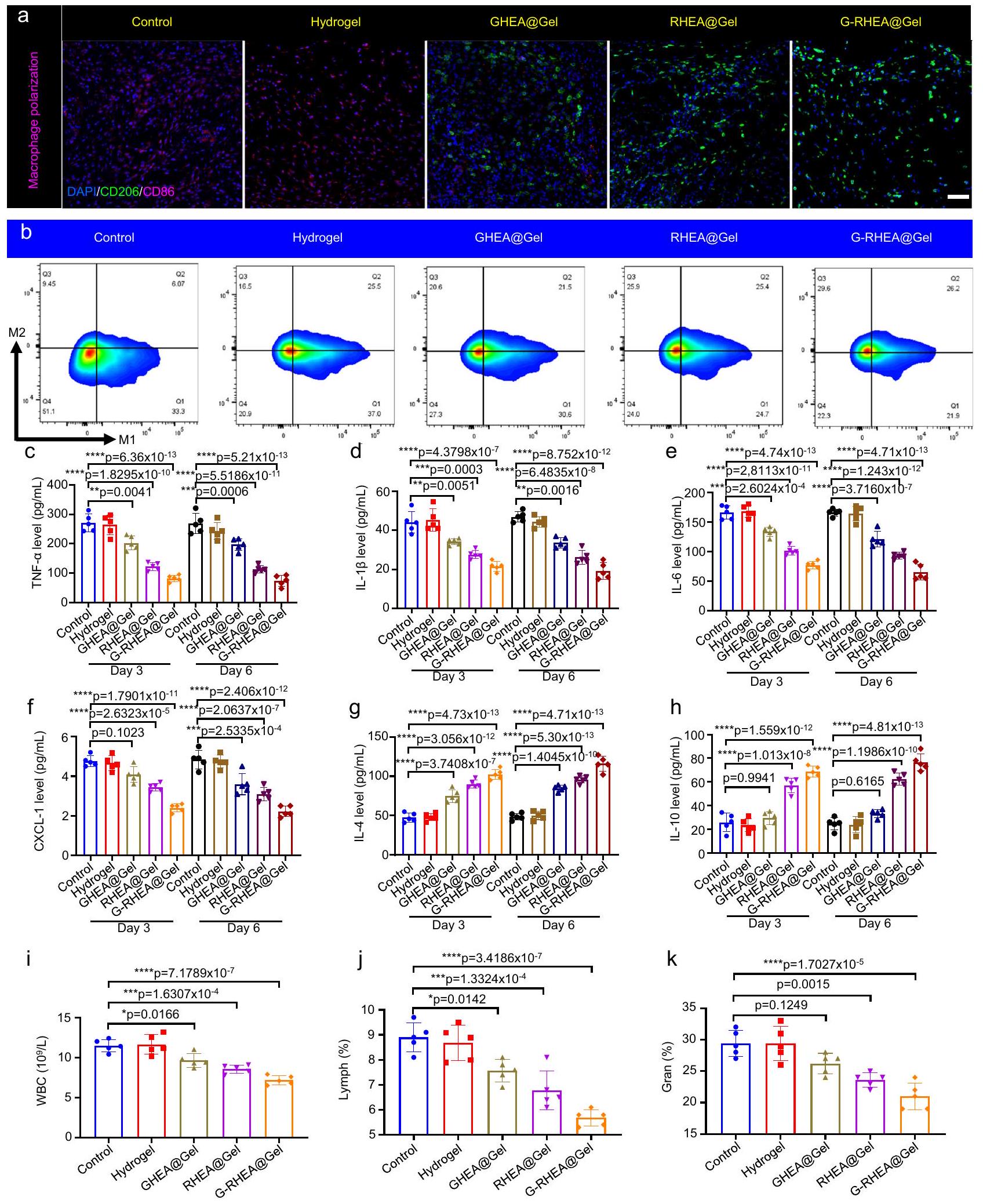
تلعب البلعميات دورًا حاسمًا في عملية شفاء الجروح. تم تأكيد نسبة البلعميات M2/M1 في الجسم من خلال صبغة مناعية لـ CD206 و CD86 (الشكل 9a). تم الكشف عن عدد أكبر من البلعميات M2 وعدد أقل من البلعميات M1 في مجموعتي GHEA@Gel و RHEA@Gel، مما أدى إلى نسبة أكبر من البلعميات M2/M1 (الشكل التكميلية 24)، مما يدل على انخفاض في القدرات المؤيدة للالتهابات وزيادة في القدرات المضادة للالتهابات. على وجه الخصوص، أظهرت مجموعة G-RHEA@Gel أعلى قدرة مضادة للالتهابات، والتي تم نسبها إلى التأثيرات المشتركة المضادة للبكتيريا، وتوفير الأكسجين، وإزالة ROS، والإجراءات المضادة للالتهابات لـ GHEA و RHEA. تم قياس CD206 (البلعميات M2) و (البلعميات M1) بواسطة قياس التدفق، وكانت النتائج متوافقة مع صور الصبغة المناعية (الشكل 9b والشكل التكميلية 25). يعتبر عامل النسخ الكابا B (NF-кB) هو المسار الإشاري المركزي الذي ينظم الالتهاب من خلال التوسط في تعبير السيتوكينات المؤيدة للالتهابات في مجرى الدم، بما في ذلك TNF, IL-6، و IL-1 , مما يعيق استعادة الأنسجة. تعتبر العدلات هي الخلايا الفعالة السائدة في العدوى البكتيرية، وتتراكم باستمرار في جروح مرضى السكري، مما يعيق عملية الشفاء. CXCL-1، وهو عضو في عائلة كيموكين CXC، يشارك في هجرة وتفعيل العدلات. في الأيام 3 و 6، تم تقليل مستويات IL- , IL- 6، TNF- و CXCL- 1 في الجروح في جميع مجموعات الجل (الشكل 9c-f). أظهرت مجموعة G-RHEA@Gel انخفاضات ملحوظة في مستويات IL- , IL- 6، TNF- و CXCL- 1 مقارنة بتلك الموجودة في المجموعات الأخرى، مما يشير إلى فعالية جيدة للعلاج المشترك في تخفيف الالتهاب. بالمقابل، IL-4 و IL-10 هما سيتوكينات تعزز التجديد وتساعد في إصلاح الأنسجة، وشفاء الجروح، وتجديد المحاور، وقطبية البلعميات M2. أظهرت مجموعة G-RHEA@Gel أعلى مستويات من IL-4 و IL-10 بين جميع مجموعات الجل مقارنة بتلك الموجودة في مجموعة التحكم، مما يشير إلى تأثير ملحوظ في تعزيز التجديد (الشكل ). في اليوم الثالث بعد العلاج، كانت عدد كريات الدم البيضاء (WBC) وعدد اللمفاويات (Lymph) ونسبة العدلات (Gran) في الدم المحيطي لمجموعات GHEA@Gel و RHEA@Gel و GRHEA@Gel أقل بكثير من تلك الموجودة في مجموعة التحكم
ومجموعات الهلام (الشكل 9i-k). وبالتالي، يمكن أن تساعد التأثيرات المضادة للبكتيريا، وإزالة ROS، وتوفير الأكسجين، والتأثيرات المضادة للالتهابات في مجموعة G-RHEA@Gel في تقليل التهاب الجروح بشكل كبير. تم إجراء صبغة H&E للكشف عن تلف الأنسجة، ويظهر الشكل التكميلية 26 سلامة جميع الهلامات للتجارب في الجسم. أظهرت هذه النتائج أن العلاج بـ G-RHEA@Gel وتحفيز الليزر هو العلاج الأمثل لشفاء الجروح. علاوة على ذلك، لتقييم ملف سلامة الهلام، تم حضن GRHEA@Gel مع خلايا HaCaT. أظهرت النتائج سلامة ممتازة، كما يتضح من نشاط الخلايا بعد التداخل مع G-RHEA@Gel. يظهر الشكل التكميلية 27a أنه، مقارنة بالعلاج التحكم، لم يؤثر علاج G-RHEA@Gel سلبًا على نشاط الخلايا. بالإضافة إلى ذلك، أظهرت صبغة الخلايا باستخدام صبغات الكالسيتين-AM و PI وملاحظتها باستخدام ميكروسكوب ليزر مسح ضوئي أن الخلايا حافظت على نشاط جيد بعد التداخل مع G-RHEA@Gel (الشكل التكميلية 27b). علاوة على ذلك، تم تغطية جلد الفئران السليم بهلام G-RHEA@Gel لمدة 30 يومًا. كشفت الفحوصات النسيجية عبر صبغة H&E وصبغة ماسون عن عدم وجود دليل على تلف الجلد بعد التعرض المطول لـ G-RHEA@Gel على سطح الجلد (الشكل التكميلية 28). لاستكشاف سلامة G RHEA@Gel على المدى الطويل، تم الكشف عن مستويات العوامل الالتهابية، بما في ذلك TNF- , IL- , IL-4، IL-6، IL-10، و CXCL-1، ومؤشرات الدم، بما في ذلك عدد كريات الدم البيضاء، وعدد اللمفاويات، وعدد الحبيبات، في الدم المحيطي للفئران. كما هو موضح في الشكل التكميلية 29، لم يكن هناك فرق كبير بين الفئران المعالجة بـ G-RHEA@Gel ومجموعة التحكم.
نقاش
تشكل الجروح المزمنة لدى مرضى السكري تهديدًا خطيرًا لحياة الإنسان وصحته. إن علاج الجروح المصابة لدى مرضى السكري هو إجراء معقد للغاية يتضمن العدوى البكتيرية، ونقص الأكسجين، وزيادة التعبير عن ROS، والالتهاب، من بين عوامل أخرى. هناك حاجة ماسة لاستراتيجية علاجية بسيطة وفعالة. إن الجمع بين العلاجات الميكروبية متعددة الوظائف والهلامات المتوافقة حيويًا يحمل وعدًا كبيرًا لعلاج جروح السكري. نظرًا لكفاءتها العالية في إنتاج الأكسجين وإفراز AST، ظهرت HEA كأكثر الكائنات الدقيقة قيمة لعلاج جروح السكري. ومع ذلك، لم يتم التحقيق بعد في استراتيجيات تعتمد على HEA الحية.
في هذه الدراسة، وضعنا استراتيجية علاجية إجرائية تعتمد على هلامات HEA الحية المبرمجة لتنفيذ مضادات البكتيريا، وإطلاق الأكسجين، وإزالة ROS، وتنظيم قطبية البلعميات لتعزيز الشفاء السريع للجروح لدى مرضى السكري بشكل شامل. باستخدام تقنية التصلب الضوئي البسيطة في الموقع، يمكن أن يتكيف HEA@Gel مع أي جروح غير متماثلة. عند التعرض للضوء عالي الكثافة ( )، كان للتحويل الضوئي لـ GHEA@Gel تأثيرات مضادة للبكتيريا فعالة، مما يكشف عن المرحلة الأولية من علاج الجروح. من خلال تنظيم الضوء منخفض الكثافة ( )، يمكن لنظام التمثيل الضوئي لـ GHEA@Gel أن ينتج الأكسجين باستمرار، مما يحل بشكل فعال مشكلة نقص الأكسجين ويعزز تجديد الأوعية، وبالتالي تحقيق الهدف من المرحلة الثانية. أدى الإشعاع الضوئي المستمر وتأثير الربط لشبكة الهلام إلى تراكم AST في خلايا HEA، مما أدى إلى تغيير تدريجي في اللون من الأخضر إلى الأحمر (RHEA). من خلال إفراز حويصلات AST عبر الإكسوزومات، يمكن لـ RHEA إزالة ROS الزائدة بكفاءة، وتعزيز تعبير الإنزيمات المضادة للأكسدة داخل الخلايا، واستقطاب البلعميات M2 مباشرة، مما يكمل المرحلة الثالثة من جروح السكري المصابة. من خلال التحكم في كثافة الضوء في المختبر، يمكن تعديل الأيض ووظيفة HEA حسب الطلب، مما يسهل العلاج المبرمج للجروح. تشير التجارب إلى أن هلام HEA الحي يمكن أن يعزز تكاثر الخلايا وهجرتها، ويعزز تشكيل الأنابيب في المختبر، ويعزز شفاء الجروح المزمنة لدى مرضى السكري في الجسم. يوفر العلاج القائم على هلام الطحالب استراتيجية متقدمة لتحسين شفاء الجروح المزمنة.
الشكل 9 | تنظيم قطبية البلعميات والبيئة المناعية بواسطة HEA@Gel في الجسم. أ صبغة مناعية مزدوجة لـ CD206 (أخضر) و CD86 (وردي) في اليوم 6 استجابة لمختلف العلاجات. شريط القياس، . تم تكرار كل تجربة بشكل مستقل ثلاث مرات مع نتائج مشابهة. ب تحليل نسبة البلعميات M2 إلى M1 استجابةً لعلاجات مختلفة بواسطة تحليل تدفق الخلايا في اليوم السادس. تم تكرار كل تجربة
بشكل مستقل ثلاث مرات مع نتائج مشابهة. تركيزات الجروح في الجسم الحي من (ج) TNF- ، (د) IL-1 “، (هـ) IL-6، (و) CXCL-1، (ز) IL-4، و(ح) IL-10 بعد 3 و6 أيام من العلاج. (ط) عدد كريات الدم البيضاء، (ي) عدد اللمفاويات، و(ك) نسبة العدلات (gran%) في الدم المحيطي. تُعرض البيانات كمتوسط س.د. ( فئران مستقلة). تم تحليل الفروق الإحصائية بواسطة اختبار الطالب ذو الجانبين-اختبار.
الهيدروجيل المحمّل بخلايا الطحالب الدقيقة النشطة يحمل وعدًا كمواد مساعدة للجروح لدى مرضى السكري، حيث يقدم مزايا مثل تعزيز شفاء الجروح، وخصائص مضادة للميكروبات، والتوافق الحيوي. ومع ذلك، فإن معالجة التحديات المتعلقة بالبحث المحدود، والموافقة التنظيمية، واعتبارات التكلفة أمر حاسم لتحقيق التحول السريري الناجح. هناك حاجة إلى مزيد من جهود البحث والتعاون بين الأوساط الأكاديمية والصناعة والهيئات التنظيمية لتحسين هذه المقاربة المبتكرة وتسهيل دمجها في استراتيجيات إدارة جروح السكري الروتينية.
طرق
بيان أخلاقي
تتوافق هذه الدراسة مع جميع اللوائح الأخلاقية ذات الصلة. تم إجراء جميع الدراسات على الحيوانات وفقًا لدليل المعهد الوطني لرعاية واستخدام الحيوانات المخبرية. تم الموافقة على جميع الإجراءات، بما في ذلك رعاية الحيوانات، ونمذجة الجروح، والجرعات، وإنهاء التجارب، من قبل لجنة الأخلاقيات الحيوانية في جامعة تيانجين مع رقم الموافقة المعين TJUE-2023-236.
المواد
تم تزويد خلايا HEA من مجموعة ثقافة الطحالب العذبة في معهد علم الأحياء المائية، FACHB. تم شراء GelMA والمبادر الضوئي LAP من شركة سوزهو يونغتشينتشوان للمعدات الذكية المحدودة. AST (تم شراء (HPLC) من سولار بيو. تم شراء 2,7-ثنائي كلوريد ثنائي هيدروفلوريسئين داي أستات (DCFH-DA) ومجموعة اختبار حيوية الخلايا/السُمية الخلوية Calcein/PI من بييوتيم. تم تزويد DMEM وRPMI 1640 و مصل جنين العجل (FBS) وتربسين-EDTA ومحلول ملحي مخفف بالفوسفات (PBS، pH 7.4 و pH 5.5) من جيبكو لايف تكنولوجيز.
تحضير وتوصيف هيدروجيل HEA@Gel
خلال فترة التذبذب الكاملة، تم إذابة كمية معينة من GelMA والمبادر الضوئي LAP في كمية معينة من PBS وخلطها بشكل موحد. ثم تم تسخين المزيج وإذابته عندلمدة 30 دقيقة. تم تبريد المحلول إلىقبل إضافة خلايا HEA والحفاظ عليها في. بعد عند التعرض للضوء الأزرق، تجمدت الحلول المذكورة بسرعة. تم توصيف الشكل الخارجي والبنية الداخلية للهلاميات المائية باستخدام المجهر الإلكتروني الماسح بالتبريد (FEI Quanta 450).
تم إصدار AST من RHEA
تم تفريق خمسة جرامات من HEA (مع جدران الخلايا) أو بروتوبلاست HEA (بدون جدران الخلايا) بشكل منفصل في 100 مل من الماء المقطر. تم وضع المحاليل في بيئة بدرجة حرارة الغرفة وتم تحريكها ببطء عبر التحريك المغناطيسي. تم استخراج ملليلتر واحد كل ساعة وتم الطرد المركزي عندلمدة 5 دقائق لإزالة الطحالب الدقيقة، بعد ذلك تم جمع السائل العلوي. تم استخراج AST في السائل العلوي باستخدام DMSO، وتم تحديد محتوى AST في السائل العلوي من خلال منحنى قياسي لـ AST. بالإضافة إلى ذلك، لمراقبة ومقارنة إطلاق AST بشكل أفضل، تم تسجيل صور لـ AST المستخرج في مذيب DMSO.
التأثير الضوئي الحراري لـ HEA
تم تعريض تركيزات مختلفة من محلول خلايا GHEA (1 مل) لليزر بزاوية 658 نانومتر ) لمدة 10 دقائق. باستخدام مقياس حرارة بالأشعة تحت الحمراء (كاميرا الأشعة تحت الحمراء TiS40 FLUKE-TiS40 9 هرتز، FLUKE، الولايات المتحدة الأمريكية)، تم الكشف عن زيادة درجة حرارة محلول خلية GHEA الناتجة عن التحويل الضوئي الحراري كل 30 ثانية لمدة 10 دقائق.
أداء مضاد للبكتيريا
تم استخدام استراتيجية عد الأطباق لتقييم القدرة المضادة للبكتيريا لـ GHEA. أولاً، تم أخذ 1 مل من E. coli أو S. aureus ( ) تم حضنه مع GHEA@Gel عند لمدة 12 ساعة. تم إشعاع هلام GHEA@Gel عند 658 نانومتر لمدة 5 دقائق لمجموعات الليزر. ثم، تم وضع 0.1 مل من تعليق البكتيريا على الأجار وتم حضنه لمدة 24 ساعة. عندأخيرًا، تم تحديد عدد المستعمرات البكتيرية، وتم تحديد القدرة المضادة للبكتيريا. لتوضيح القدرة المضادة للبكتيريا لـ GHEA بشكل أكبر، تم فحص صور SEM للأشكال البكتيرية تحت علاجات مختلفة.
خارجيإنتاج
لتحقيق في قدرة خلايا GHEA على إنتاج الأكسجين، تم إعداد حجم نهائي قدره 15 مل من خلايا GHEA في محلول PBS مع أو خلايا GHEAتم التحضير. قبل الكشف، تم ضخ غاز الأرجون في المحلول لمدة 10 دقائق، بعد ذلك تم ترك الخليط في الظلام لمدة ساعة واحدة لاستقرار الظروف الناقصة الأكسجين. باستخدام كاشف الأكسجين، تم الكشف عن الضوء ( أو تم قياس الأكسجين المنتج بواسطة GHEA المدعوم كل دقيقة مع التحريك. بالإضافة إلى ذلك، تم تعريض المحلول لأشعة ليزر عالية الكثافة. ) تُركت في درجة حرارة الغرفة لمدة ساعتين لتبرد. ثم تم تعريض المحلول لأشعة ليزر منخفضة الكثافة ( ) لمدة 20 دقيقة، بعد ذلك تم مراقبة إنتاج الأكسجين. لتحديد ما إذا كان GHEA يمكن أن ينتج الأكسجين خلال عملية الشفاء الطويلة للجروح، تم إعداد محلول خلايا GHEA بحجم نهائي قدره 15 مل خلايا GHEAتم تحضيرها. ثم، تم تعريض المحلول لأشعة ليزر عالية الكثافة ( ) لمدة 5 دقائق. في كل يوم لاحق، تم تعريض المحلول لأشعة ليزر منخفضة الكثافة ( ) لمدة 30 دقيقة، وتمت مراقبة إنتاج الأكسجين بواسطة كاشف الأكسجين.
قدرة استغلال الجذور الحرة
تم تقييم قدرة RHEA@Gel على التخلص من الجذور الحرة باستخدام، ، و كممثل لـ ROS.تم تحديد قدرة التجميع عن طريق خلط 10 مل منمع الهلاميات. بعد ساعتين،تم خلط السائل العلوي لمدة 30 دقيقة معمنتم قياس الامتصاص عند 405 نانومتر.
القدرة على البحث عن الطعامتم تقييمه من خلال حساب نسبة تثبيط اختزال NBT. تحت شدة ضوء ثابتة، NBT ( )، ميثيونين ( 12.5 مللي مولار )، ريبوفلافين ( تم دمج RHEA@Gel لمدة 10 دقائق. تم الكشف عن منحنى المسح الكامل لعدد الموجات في المحلول، وتم تسجيل امتصاص المحلول عند 560 نانومتر.
باستخدام مسبار SA،تم تحديد قدرة RHEA@Gel على التجميع. باختصار،، وتم إضافة SA إلى PBS، وتم تحضين المزيج لمدة 30 دقيقة فيتم الكشف عن منحنى المسح الكامل لعدد الموجات للمحلول، وتم تسجيل امتصاص المحلول عند 510 نانومتر.
مصادر خطوط الخلايا
تم الحصول على الخلايا الليفية الجلدية البشرية (HSF، رقم الكتالوج: PCS-201-012)، وخلايا اللوكيميا أحادية النواة الماوسية (RAW264.7، رقم الكتالوج: TIB-71)، وخلايا بطانة الوريد السري البشري (HUVEC، رقم الكتالوج: PCS-100-013) وخلايا الكيراتينocytes البشرية الخالدة (HaCaT، رقم الكتالوج: PCS-200-011) من مجموعة الثقافة الأمريكية (ATCC).
زراعة الخلايا
تم الحصول على HSFs وHaCaTs وHUVECs من ATCC. تم استخدام وسط Eagle المعدل من دولبيكو (DMEM؛ جيبكو) معزز بـ 10% من مصل العجل الجنيني (FBS) لزراعة HSFs. تم استخدام وسط RPMI 1640 (جيبكو) معزز بـتم استخدام FBS لاحتضان خلايا HUVECs وHaCaTs وRAW264.7.
الأجسام المضادة
أجسام مضادة أحادية النسيلة من نوع الفأر ضد CD86، Abcam (رقم المنتج # 130-122-1)، تخفيف 1:200؛ أجسام مضادة أحادية النسيلة من نوع الفأر ضد CD206، Abcam (رقم المنتج # 141706)، تخفيف 1:200؛ أجسام مضادة أحادية النسيلة من نوع الفأر ضد VEGF، Santa Cruz (رقم المنتج # 57496)، تخفيف 1:200؛ أجسام مضادة أحادية النسيلة من نوع الفأر ضد EGF، Abcam (رقم المنتج # EPR19173)، تخفيف 1:500؛ أجسام مضادة أحادية النسيلة من نوع الفأر ضد HIF-1 ألفا، Abcam (رقم المنتج # EP1215Y)، استخدم تركيزاً من; الأجسام المضادة وحيدة النسيلة من نوع الفأر –
أجسام مضادة CD31، Abcam (رقم المنتج 222783)، تخفيف 1:1000؛ أجسام مضادة أحادية النسيلة ضد الأكتين العضلي الأملس ألفا (SMA) من الفأر، Abcam (رقم المنتج ab7817)، تخفيف 1:500؛ أجسام مضادة أحادية النسيلة ضد VIM من الفأر، Absin (رقم المنتج abs136555)، تخفيف 1:1000؛ أجسام مضادة أحادية النسيلة ضد COL من الفأر، Santa Cruz (رقم المنتج sc-59772)، تخفيف 1:500؛ أجسام مضادة متعددة النسائل ضد المكورات العنقودية الذهبية، Absin (رقم المنتج ab20920)، تخفيف 1:1000؛ أجسام مضادة ثانوية عالية الامتصاص ضد IgG من الماعز (H + L) من ThermoFisher (رقم الكتالوج A-11034)، تخفيف 1:1000؛ أجسام مضادة ضد IgG من الفأر من الماعز (أجسام مضادة ثانوية عالية الامتصاص المتقاطع، ثيرمو فيشر (رقم الكتالوج A-21236)، تخفيف 1:1000.
تحديد إزالة الجذور الحرة داخل الخلوية
تم استخدام مجس DCFH-DA لقياس قدرة العينات على التخلص من الجذور الحرة للأكسجين (ROS).وتم إضافة العينات إلى لوحة تحتوي على 6 آبار تحتوي على خلايا مزروعةتم إضافة مجس DACH-DA بعد 12 ساعة، وتم تحضين المزيج لمدة 20 دقيقة أخرى. ثم تم التقاط صور الفلورسنت للمجموعات المختلفة.
داخل الخليةجيل
تم تحديد القدرة على توليد الأكسجين داخل الخلايا للهلاميات باستخداماستقصاء. تم زراعة الخلايا في طبق 6 آبار (الخلايا/بئر) وتم تغطيتها بالهيدروجيل، الذي تم معالجته إما بالليزر أو لم يتم. بعدتم إضافة إلى الخلايا، وتم حضن الخلايا لمدة 20 دقيقة. ثم، تم التقاط صور الفلورسنت للمجموعات المختلفة.
سمية الخلايا والهجرة
تم استخدام HaCaTs لتقييم السمية الخلوية للهلامات المائية من خلال اختبار MTT. تم زراعة الخلايا في الحجرة السفلية من لوحة ترانسويل ذات 24 بئرًا (كل بئر يحتوي علىالخلايا) لمدة 12 ساعة. بعد ذلك، تم إضافة هيدروجيل G-RHEA@Gel إلى كل حجرة علوية وتم حضنه لمدة 12 ساعة أخرى. ثم، تم إضافة محلول MTT، وتم حضن الخلايا لمدة ساعتين. تم قياس الامتصاص عند 490 نانومتر من خلال عملية موحدة. بالإضافة إلى ذلك، تم استخدام الكالسيفين-AM ( ) و PI تم استخدام صبغات لتحديد توزيع الخلايا الحية والميتة. تم تحليل موت الخلايا عبر المجهر الضوئي الماسح بالليزر. بالنسبة لتجربة هجرة الخلايا،تم زراعة الخلايا في أطباق 24 بئرًا وسمح لها بالنمو لمدة 24 ساعة قبل أن يتم خدشها.تم استخدام الماصة بعد ذلك لرسم خط عريض بشكل موحد في وسط كل بئر من آبار الـ 24 لإزالة الخلايا من تلك المنطقة. ثم تم وضع الخلايا في بيئات مختلفة وتقسيمها إلى أربع مجموعات: مجموعة البيئة ناقصة الأكسجين،مجموعة بيئة الإمداد، البيئة منخفضة الأكسجين مع التعايش مع مجموعة GHEA@Gel، والبيئة منخفضة الأكسجين مع التعايش مع مجموعة GHEA@Gel والتعرض للضوء. تم تسجيل هجرة الخلايا بعد 12 و 24 ساعة.
اختبار تشكيل الأنبوب
لتحقيق تشكيل الأنابيب،تم إضافة 100 ميكرولتر من ماتريجيل المذاب (abs9493، أبسين) إلى كل حجرة سفلية من لوحة 24 بئر. بعد الحضانة فيلـتم زرع خلايا HUVEC في الحجرة السفلية المغلفة بمادة ماتريجيل وتعرضت لعلاجات مختلفة. في مجموعة الليزر، تم تعريض الهيدروجيل لضوء ليزر بطول موجي 658 نانومتر لمدة 10 دقائق. تم تسجيل صور أنابيب HUVEC بواسطة ميكروسكوب وتحليلها باستخدام برنامج ImageJ لتقييم تشكيل الأنابيب.
القدرة المضادة للالتهابات
التأثير المضاد للالتهابات لـ LPS + IFN- أو IL-4 تم تقييمه في نموذج البلعميات. تم زرع البلعميات في الحجرة السفلية من لوحة ترانسويل ذات 24 بئرًا (كل بئر يحتوي على الخلايا) لمدة 12 ساعة. خلال زراعة البلعميات، LPS و إنترفيرونتمت إضافة إلى وسط الثقافة، بعد ذلك تم زراعة الخلايا لمدة 12 ساعة. بعد ذلك، تم إضافة GHEA@Gel أو RHEA@Gel أو G-RHEA@Gel إلى وسط الثقافة، وتم السماح للخلايا بالنمو. لمدة 24 ساعة. تم تثبيت البلعميات بـبارافورمالدهيد. ثم تم إجراء صبغ المناعية الفلورية وتحليل FCM لـ CD206 و CD86. خلال هذه العملية، تم حضن البلعميات معأدى IL-4 إلى تحفيز النمط الظاهري M2 كتحكم.
دراسة حيوانية
إناث فئران BALB/c (6 أسابيع، ) المقدمة من شركة بكين HFK للعلوم البيولوجية تم استخدامها في هذه الدراسة. تم إيواء جميع الحيوانات في منشأة حيوانات خالية من مسببات الأمراض المحددة (SPF) لمدة أسبوعين للتكيف البيئي وتم السماح لها بالوصول الحر إلى الطعام والماء. خلال التجربة، تم الاحتفاظ بجميع الحيوانات في نفس البيئة القياسية ( الرطوبة، دورة ضوء-ظلام لمدة 12 ساعة، وأربعة فئران/قفص). تم تنفيذ جميع الإجراءات، بما في ذلك رعاية الحيوانات، نمذجة الجروح، الجرعات، وإنهاء التجربة، وفقًا لإرشادات الحيوانات التجريبية للمراجعة الأخلاقية لرفاهية الحيوان (GB/T 35892-2018) وتمت الموافقة عليها من قبل لجنة الأخلاقيات الحيوانية في جامعة تيانجين مع رقم الموافقة المعين TJUE-2023-236. تم تخدير الفئران بمخدر الإيزوفلوران قبل أي إجراء قد يسبب الألم. بعد التجربة، تم euthanized الفئران بواسطةاستنشاق يليه خلع عنق الرحم.
إنشاء نموذج جرح مصاب بالسكري
لتقييم الفعالية العلاجية لـ HEA@Gel، تم إنشاء نموذج فأر لجروح السكري المصابة. تم استخدام إناث من فئران BALB/c التي تزنتم الصيام لمدة 12 ساعة، تلاها حقن داخل الصفاق بالستربتوزوتوسين )، وتم تكرار العملية ثلاث مرات خلال 3 أيام. تم إبقاء الفئران تحت التخدير أثناء الحقن داخل الصفاق. تم تزويد جميع الفئران بـ ماء السكروز. بعد أسبوعين من الحقنة الثالثة، تم تحديد الفئران المصابة بالسكري على أنها تعاني من مستويات سكر الدم التي تتجاوزلقياسين متتاليين خلال يومين. تم تخدير الفئران المصابة بالسكري باستخدام مخدر الإيزوفلوران، وتم حلاقة شعر ظهرها بالكامل. ثم تم استخدام مثاقب خزعة بقطر 10 مم لإنشاء جروح كاملة السماكة، وتعليق S. aureus (تم إسقاطه على سطح الجروح لإنشاء نموذج جرح مصاب. تم إبقاء الفئران تحت التخدير خلال هذه العملية. بعد يوم واحد من العدوى، زاد عدد المستعمرات البكتيرية منإلىCFUs، مما يدل على أن نموذج العدوى تم إنشاؤه بنجاح.
شفاء الجروح المصابة لدى مرضى السكري
بعد إنشاء نموذج الجرح السكري المصاب، تم إضافة قطرات من محلول الهيدروجيل، بما في ذلك الهيدروجيل، GHEA@Gel، RHEA@Gel، أو GRHEA@Gel، إلى الجرح. تم استخدام السائل من الهيدروجيل لتغطية منطقة الجرح بالكامل بسرعة، بعد ذلك تم تعريض الهيدروجيل لأشعة فوق بنفسجية تحت مصباح للأشعة فوق البنفسجية لمدة 10 ثوانٍ لتحقيق تصلب الهيدروجيل. تم إبقاء الفئران تحت التخدير أثناء تغطية الهيدروجيل وعملية التصلب الضوئي. تم معالجة الجروح بمحلول PBS كمجموعة تحكم. بعد ذلك، تم تعريض الجروح التي تغطيها هيدروجيلات مختلفة ليزر بزاوية 658 نانومتر مع شدة لمدة 5 دقائق لتحقيق التعقيم من خلال التحويل الضوئي الحراري. تم إبقاء الفئران تحت التخدير أثناء استخدام الليزر عالي القدرة ( ) الإشعاع. في كل يوم لاحق، تم تخدير الفئران باستخدام مخدر الإيزوفلوران، وتم استخدام ليزر بزاوية 658 نانومتر لإضاءة الجرح عند شدة ضوء لمدة 30 دقيقة في الأيام العشرة التالية. للإشعاع بالليزر منخفض الكثافة تأثيران. أولاً، يقوم GHEA بتنشيط عملية التمثيل الضوئي لإنتاج الأكسجين لتحسين البيئة الدقيقة للجروح ذات نقص الأكسجين وتسريع تجديد الأوعية الدموية وشفاء الجروح. الثاني هو تعزيز إفراز AST من RHEA لتقليل مستويات ROS والعوامل الالتهابية المفرطة التعبير وتنظيم البيئة الدقيقة المناعية في الجروح. مع شفاء الجرح، بدأت حواف الهيدروجيل تنفصل تدريجياً عن نسيج الجرح الذي تم شفاؤه، وتمت إزالة الهيدروجيل ببطء من الجلد في اليوم الثاني عشر بعد تخدير الفئران باستخدام الإيزوفلوران.
في الأيام، وتم التقاط 12 صورة للجروح. في اليومين 3 و 9، تم euthanized الفئران بواسطةتم استنشاق الهواء تلاه خلع عنق الرحم، وتم جمع الجلد من موقع الجرح لإجراء صبغة HE وصبغة ماسون. تم تحليل مستوى نقص الأكسجة، ومستوى ROS، والتوعية الوعائية، وترسب الكولاجين، وتكوين البشرة، واستقطاب البلعميات في الجلد من موقع الجرح بواسطة صبغة المناعية الفلورية لـ HIF-دي إتش إي، سي دي-SMA، الكولاجين/الفيمنتين، EGF، VEGF، وCD206/CD86 في اليوم السادس. لتقييم التوافق الحيوي والسلامة الحيوية لعلاجات HEA@Gel، تم جمع الأعضاء الرئيسية للفئران، بما في ذلك القلب، الكبد، الطحال، الرئة والكلى، لإجراء تحليل صبغة HE في اليوم الثامن بعد علاج HEA@Gel. لتقييم القدرة المضادة للبكتيريا على المدى الطويل لـ HEA@Gel، تم جمع الجلد في موقع الجرح في اليوم العشرين، وتم إجراء صبغة مناعية كيميائية باستخدام جسم مضاد ضد المكورات العنقودية الذهبية. بالإضافة إلى ذلك، لتحليل التأثير المضاد للبكتيريا بصريًا، تم جمع الجلد وهرسه. تم تخفيف الهريس بشكل متسلسل وزرعه على أجار LB لت quantifying مستعمرات البكتيريا.
الأثر طويل الأمد لـ G-RHEA@Gel على الجلد
إمكانات السلامة لمادة ما تعتبر حاسمة لتطبيقها. لاستكشاف التأثير طويل الأمد لـ HEA@Gel على الجلد، تم استخدام جلد إناث الفئران BALB/c الصحية (6 أسابيع، ) تم تغطيته بهلام GRHEA@Gel لمدة 30 يومًا. ثم تم euthanized الفئران عن طريق الاستنشاق يليه خلع العنق، وتم جمع الجلد المغطى بالهلام لإجراء صبغة H&E وMasson. بالإضافة إلى ذلك، تم الكشف عن مستويات TNF- , IL-1 , IL-4، IL-6، IL-10، وCXCL-1، وكذلك تم الكشف عن عدد كريات الدم البيضاء واللمفاويات والجرانول.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقرير Nature Portfolio المرتبط بهذه المقالة.
توفر البيانات
يعلن المؤلفون أن جميع البيانات التي تدعم نتائج هذه الدراسة متاحة ضمن المقالة والمعلومات التكميلية. جميع البيانات الأخرى متاحة من المؤلفين المراسلين عند الطلب. يتم توفير بيانات المصدر مع هذه الورقة.
References
Chen, H. et al. Dissolved oxygen from microalgae-gel patch promotes chronic wound healing in diabetes. Sci. Adv. 6, eaba4311 (2020).
Hart, T., Milner, R. & Cifu, A. Management of a diabetic foot. JAMA 318, 1387-1388 (2017).
Botusan, I. R. et al. Stabilization of HIF-1 is critical to improve wound healing in diabetic mice. Proc. Natl Acad. Sci. 105, 19426-19431 (2008).
Thangarajah, H. et al. HIF-1 dysfunction in diabetes. Cell Cycle 9, 75-79 (2010).
Thangarajah, H. et al. The molecular basis for impaired hypoxiainduced VEGF expression in diabetic tissues. Proc. Natl Acad. Sci. 106, 13505-13510 (2009).
Zhu, Y., Wang, Y., Jia, Y., Xu, J. & Chai, Y. Roxadustat promotes angiogenesis through HIF-1 VEGF/VEGFR2 signaling and accelerates cutaneous wound healing in diabetic rats. Wound Repair Regen. 27, 324-334 (2019).
Zhang, F. et al. Infected wound repair with an ultrasoundenhanced nanozyme hydrogel scaffold. Mater. Horiz. 10, 5474 (2023).
Liu, S. et al. Absorbable thioether grafted hyaluronic acid nanofibrous hydrogel for synergistic modulation of inflammation microenvironment to accelerate chronic diabetic wound healing. Adv. Healthc. Mater. 9, 2000198 (2020).
Qian, Y. et al. Immunoregulation in diabetic wound repair with a photoenhanced glycyrrhizic acid hydrogel scaffold. Adv. Mater. 34, 2200521 (2022).
Krzyszczyk, P., Schloss, R., Palmer, A. & Berthiaume, F. The role of macrophages in acute and chronic wound healing and interventions to promote pro-wound healing phenotypes. Front. Phys. 9, 419 (2018).
Wang, T., Li, Y., Cornel, E. J., Li, C. & Du, J. Combined antioxidantantibiotic treatment for effectively healing infected diabetic wounds based on polymer vesicles. ACS Nano 15, 9027-9038 (2021).
Ouyang, J. et al. In situ sprayed NIR-responsive, analgesic black phosphorus-based gel for diabetic ulcer treatment. Proc. Natl Acad. Sci. 117, 28667-28677 (2020).
Talebian, S. et al. Self-healing hydrogels: the next paradigm shift in tissue engineering? Adv. Sci. 6, 1801664 (2019).
Liu, X., Inda, M. E., Lai, Y., Lu, T. K. & Zhao, X. Engineered living hydrogels. Adv. Mater. 34, e2201326 (2022).
Louf, J.-F., Lu, N. B., O’Connell, M. G., Cho, H. J. & Datta, S. S. Under pressure: hydrogel swelling in a granular medium. Sci. Adv. 7, eabd2711 (2021).
Xu, Y. et al. Robust and multifunctional natural polyphenolic composites for water remediation. Mater. Horiz. 9, 2496-2517 (2022).
Cao, H. et al. Versatile polyphenolic platforms in regulating cell biology. Chem. Soc. Rev. 51, 4175-4198 (2022).
Fang, Y. et al. Dissecting biological and synthetic soft-hard interfaces for tissue-like systems. Chem. Rev. 122, 5233-5276 (2022).
Hamedi, H., Moradi, S., Hudson, S. M., Tonelli, A. E. & King, M. W. Chitosan based bioadhesives for biomedical applications: a review. Carbohyd. Polym. 282, 119100 (2022).
Peng, W. et al. Recent progress of collagen, chitosan, alginate and other hydrogels in skin repair and wound dressing applications. Int. J. Biol. Macromol. 208, 400-408 (2022).
Xue, X. et al. Fabrication of physical and chemical crosslinked hydrogels for bone tissue engineering. Bioact. Mater. 12, 327-339 (2022).
Kang, Y. et al. The marriage of Xenes and hydrogels: fundamentals, applications, and outlook. Innovation 3, 100327 (2022).
Webber, M. J. & Tibbitt, M. W. Dynamic and reconfigurable materials from reversible network interactions. Nat. Rev. Mater. 7, 541-556 (2022).
Zeng, Q., Qi, X., Shi, G., Zhang, M. & Haick, H. Wound dressing: from nanomaterials to diagnostic dressings and healing evaluations. ACS Nano 16, 1708-1733 (2022).
Zhao, Y. et al. Supramolecular adhesive hydrogels for tissue engineering applications. Chem. Rev. 122, 5604-5640 (2022).
Dong, R. & Guo, B. Smart wound dressings for wound healing. Nano Today 41, 101290 (2021).
Liang, Y., He, J. & Guo, B. Functional hydrogels as wound dressing to enhance wound healing. ACS Nano 15, 12687-12722 (2021).
Zhang, S. et al. Recent advances in responsive hydrogels for diabetic wound healing. Mater. Today Bio 18, 100508-100508 (2023).
Cheng, S. et al. Dendritic hydrogels with robust inherent antibacterial properties for promoting bacteria-infected wound healing. ACS Appl. Mater. Interfaces 14, 11144-11155 (2022).
Hao, Y., Zhao, W., Zhang, H., Zheng, W. & Zhou, Q. Carboxymethyl chitosan-based hydrogels containing fibroblast growth factors for triggering diabetic wound healing. Carbohyd. Polym. 287, 119336 (2022).
Liang, Y. et al. Glucose dual responsive metformin release hydrogel dressings with adhesion and self-healing via dualdynamic bonding for athletic diabetic foot wound healing. ACS Nano 16, 3194-3207 (2022).
Sabbagh, F., Muhamad, I. I., Niazmand, R., Dikshit, P. K. & Kim, B. S. Recent progress in polymeric non-invasive insulin delivery. Int. J. Biol. Macromol. 203, 222-243 (2022).
Yang, J. et al. Glucose-responsive multifunctional metal-organic drug-loaded hydrogel for diabetic wound healing. Acta Biomater. 140, 206-218 (2022).
Zhao, Y. et al. Biomimetic nanozyme-decorated hydrogels with H2O2-activated oxygenation for modulating immune microenvironment in diabetic wound. ACS Nano 17, 16854-16869 (2023).
Wu, Y. et al. A versatile glycopeptide hydrogel promotes chronic refractory wound healing through bacterial elimination, sustained oxygenation, immunoregulation, and neovascularization. Adv. Funct. Mater. 33, 2305992 (2023).
Zhang, J. et al. An injectable bioactive dressing based on plateletrich plasma and nanoclay: sustained release of deferoxamine to accelerate chronic wound healing. Acta Pharm. Sin. B 13, 4318-4336 (2023).
Liu, B. et al. Hydrogen bonds autonomously powered gelatin methacrylate hydrogels with super-elasticity, self-heal and underwater self-adhesion for sutureless skin and stomach surgery and E-skin. Biomaterials 171, 83-96 (2018).
Kurian, A. G., Singh, R. K., Patel, K. D., Lee, J.-H. & Kim, H.-W. Multifunctional GelMA platforms with nanomaterials for advanced tissue therapeutics. Bioact. Mater. 8, 267-295 (2022).
Koo, H., Allan, R. N., Howlin, R. P., Stoodley, P. & Hall-Stoodley, L. Targeting microbial biofilms: current and prospective therapeutic strategies. Nat. Rev. Microbiol. 15, 740-755 (2017).
Coban, O., De Deyn, G. B. & van der Ploeg, M. Soil microbiota as game-changers in restoration of degraded lands. Science 375, 990 (2022).
Jansson, J. K. & Hofmockel, K. S. Soil microbiomes and climate change. Nat. Rev. Microbiol. 18, 35-46 (2020).
van Oppen, M. J. H. & Blackall, L. L. Coral microbiome dynamics, functions and design in a changing world. Nat. Rev. Microbiol. 17, 557-567 (2019).
Zhang, H. et al. Bacteria photosensitized by intracellular gold nanoclusters for solar fuel production. Nat. Nanotechnol. 13, 900-905 (2018).
Huo, M. et al. Photosynthetic tumor oxygenation by photosensitizer-containing cyanobacteria for enhanced photodynamic therapy. Angew. Chem. Int. Ed. 59, 1906-1913 (2020).
Huang, J. et al. Bio-inspired synthesis of metal nanomaterials and applications. Chem. Soc. Rev. 44, 6330-6374 (2015).
Logan, B. E., Rossi, R., Ragab, A. A. & Saikaly, P. E. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 17, 307-319 (2019).
Singh, P., Kim, Y.-J., Zhang, D. & Yang, D.-C. Biological synthesis of nanoparticles from plants and microorganisms. Trends Biotechnol. 34, 588-599 (2016).
Yang, H. et al. Preparation of nickel-iron hydroxides by microorganism corrosion for efficient oxygen evolution. Nat. Commun. 11, 5075 (2020).
Chen, J. et al. Oxygen-self-produced nanoplatform for relieving hypoxia and breaking resistance to sonodynamic treatment of pancreatic cancer. ACS Nano 11, 12849-12862 (2017).
Li, X., Kwon, N., Guo, T., Liu, Z. & Yoon, J. Innovative strategies for hypoxic-tumor photodynamic therapy. Angew. Chem. Int. Ed. 57, 11522-11531 (2018).
Saravanakumar, G., Kim, J. & Kim, W. J. Reactive-oxygen-speciesresponsive drug delivery systems: promises and challenges. Adv. Sci. 4, 1600124 (2017).
Xuan, M. et al. Magnetic mesoporous silica nanoparticles cloaked by red blood cell membranes: applications in cancer therapy. Angew. Chem. Int. Ed. 57, 6049-6053 (2018).
Zhang, X. et al. Black phosphorus-loaded separable microneedles as responsive oxygen delivery carriers for wound healing. ACS Nano 14, 5901-5908 (2020).
Zhao, L.-P. et al. Self-delivery nanomedicine for O-2-economized photodynamic tumor therapy. Nano Lett. 20, 2062-2071 (2020).
Hu, H. et al. Microalgae-based bioactive hydrogel loaded with quorum sensing inhibitor promotes infected wound healing. Nano Today 42, 101368 (2022).
Ou, M. et al. Heterojunction engineered bioactive chlorella for cascade promoted cancer therapy. J. Control. Release 345, 755-769 (2022).
Khoo, K. S. et al. Recent advances in biorefinery of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 288, 121606 (2019).
Yang, L. et al. Influence of molecular structure of astaxanthin esters on their stability and bioavailability. Food Chem. 343, 128497(2021).
Zhao, X. et al. Physical and oxidative stability of astaxanthin microcapsules prepared with liposomes. J. Sci. Food Agric. 102, 4909-4917 (2022).
Gwak, Y. et al. Comparative analyses of lipidomes and transcriptomes reveal a concerted action of multiple defensive systems against photooxidative stress in Haematococcus Pluvialis. J. Exp. Bot. 65, 4317-4334 (2014).
Hu, C. et al. Transcriptomic analysis unveils survival strategies of autotrophic Haematococcus pluvialis against high light stress. Aquaculture 513, 734430 (2019).
Zhang, X. et al. ROS-triggered self-disintegrating and pHresponsive astaxanthin nanoparticles for regulating the intestinal barrier and colitis. Biomaterials 292, 121937 (2023).
Wen, X. et al. Astaxanthin acts via LRP-1 to inhibit inflammation and reverse lipopolysaccharide-induced M1/M2 polarization of microglial cells. Oncotarget 8, 69370-69385 (2017).
Ambati, R. R., Phang, S. M., Ravi, S. & Aswathanarayana, R. G. Astaxanthin: sources, extraction, stability, biological activities and its commercial applications-a review. Mar. Drugs 12, 128-152 (2014).
Davinelli, S., Nielsen, M. E. & Scapagnini, G. Astaxanthin in skin health, repair, and disease: a comprehensive review. Nutrients 10, 522 (2018).
Lim, K. C., Yusoff, F. M., Shariff, M. & Kamarudin, M. S. Astaxanthin as feed supplement in aquatic animals. Rev. Aquac. 10, 738-773 (2018).
Oslan, S. N. H. et al. A review on Haematococcus pluvialis bioprocess optimization of green and red stage culture conditions for the production of natural astaxanthin. Biomolecules 11, 256 (2021).
Perozeni, F. et al. Turning a green alga red: engineering astaxanthin biosynthesis by intragenic pseudogene revival in Chlamydomonas reinhardtii. Plant Biotechnol. J. 18, 2053-2067 (2020).
Rammuni, M. N., Ariyadasa, T. U., Nimarshana, P. H. V. & Attalage, R. A. Comparative assessment on the extraction of carotenoids from microalgal sources: Astaxanthin from H-pluvialis and beta-carotene from D. salina. Food Chem. 277, 128-134 (2019).
Shah, M. M. R., Liang, Y., Cheng, J. J. & Daroch, M. Astaxanthinproducing green microalga Haematococcus pluvialis: from single Cellto high value commercia products. Front. Plant Sci. 7, 531 (2016).
Huang, W.-C., Liu, H., Sun, W., Xue, C. & Mao, X. Effective astaxanthin extraction from wet Haematococcus pluvialis using switchable hydrophilicity solvents. ACS Sustain. Chem. Eng. 6, 1560-1563 (2018).
Yang, H. E., Yu, B. S. & Sim, S. J. Enhanced astaxanthin production of Haematococcus pluvialis strains induced salt and high light resistance with gamma irradiation. Bioresour. Technol. 372, 128651 (2023).
Xu, R., Zhang, L., Yu, W. & Liu, J. A strategy for interfering with the formation of thick cell walls in Haematococcus pluvialis by downregulating the mannan synthesis pathway. Bioresour. Technol. 362, 127783 (2022).
Fulbright, S. P. et al. Bacterial community changes in an industrial algae production system. Algal Res. 31, 147-156 (2018).
Mickalide, H. & Kuehn, S. Higher-order interaction between species inhibits bacterial invasion of a phototroph-predator microbial community. Cell Syst. 9, 521-533 (2019).
الشكر والتقدير
تم دعم هذه الدراسة ماليًا من خلال منحة من المؤسسة الوطنية للعلوم الطبيعية في الصين (رقم المنحة 32071322، X.J.)، وصناديق العلوم الطبيعية الوطنية للعلماء الشباب المتميزين (رقم المنحة 32122044، X.J.)، ومؤسسة العلوم ما بعد الدكتوراه في الصين (رقم المنحة 2023T160479، 2023M742603، Y.K.)، ولجنة التكنولوجيا والابتكار لبلدية شنتشن (رقم المنحة.
JCYJ20210324113004010، X.J.).
مساهمات المؤلفين
صمم X. Ji وB. Liu وJ. Xie وأشرفوا على المشروع. صمم X. Ji وY. Kang وL. Xu وJ. Dong استراتيجيات التجارب. قام Y. Kang وL. Xu وJ. Dong وX. Yuan وJ. Ye وY. Fan بإجراء التجارب وتحليل البيانات. كتب X. Ji وB. Liu وJ. Xie المخطوطة.
أكاديمية الهندسة الطبية والطب الانتقالي، كلية الطب، جامعة تيانجين، تيانجين 300072، الصين. قسم السيطرة على الأمراض والوقاية منها، مركز الطب المميز لقوة الصواريخ، بكين 10088، الصين. قسم الحروق، المستشفى الأول التابع لجامعة صن يات سين، قوانغتشو 510080، الصين. كلية الطب، جامعة لين يي، لين يي 276000، الصين. البريد الإلكتروني: neaucn@126.com; xiejulin@mail.sysu.edu.cn; jixiaoyuan@tju.edu.cn
Chronic diabetic wounds are at lifelong risk of developing diabetic foot ulcers owing to severe hypoxia, excessive reactive oxygen species (ROS), a complex inflammatory microenvironment, and the potential for bacterial infection. Here we develop a programmed treatment strategy employing live Haematococcus (HEA). By modulating light intensity, HEA can be programmed to perform a variety of functions, such as antibacterial activity, oxygen supply, ROS scavenging, and immune regulation, suggesting its potential for use in programmed therapy. Under high light intensity ( ), green HEA (GHEA) with efficient photothermal conversion mediate wound surface disinfection. By decreasing the light intensity ( ), the photosynthetic system of GHEA can continuously produce oxygen, effectively resolving the problems of hypoxia and promoting vascular regeneration. Continuous light irradiation induces astaxanthin (AST) accumulation in HEA cells, resulting in a gradual transformation from a green to red hue (RHEA). RHEA effectively scavenges excess ROS, enhances the expression of intracellular antioxidant enzymes, and directs polarization to M2 macrophages by secreting AST vesicles via exosomes. The living HEA hydrogel can sterilize and enhance cell proliferation and migration and promote neoangiogenesis, which could improve infected diabetic wound healing in female mice.
Approximately 537 million people worldwide suffer from diabetes, and its incidence is projected to increase by by . A quarter of diabetic patients have a lifetime threat of persistent nonhealing wounds such as diabetic foot ulcers, which force patients to be in danger of amputation, and of these individuals have a life expectancy of less than 5 years . Studies have shown that damaged neovascularization in response to hypoxia is one of the most severe causes of chronic wound deterioration in patients with diabetes . Exposure to high glucose triggers rapid posttranslational hydroxylation and degradation of hypoxia-inducible factor-1alpha. When responding to soft tissue ischemia, this alteration renders diabetic wounds incapable of upregulating vascular endothelial growth factor
(VEGF), creating obstacles to angiogenesis and wound healing . Additionally, excessive reactive oxygen species (ROS) are another important impediment to the diabetic wound healing process, causing perpetual and irreversible damage to biomolecules and sustaining macrophages in the M1 phenotype to exacerbate the inflammatory response . As a result, the recruitment of M1 macrophages creates a unique injury microenvironment in wounds characterized by augmented proteolysis and oxidative cellular damage . Moreover, open wounds are highly susceptible to bacterial infection, which exacerbates wound hypoxia and inflammation . Thus, designing an all-rounder with space-time controllable oxygen release, ROS scavenging, modulation of
macrophage polarization, and antibacterial properties is a hot topic for the clinical treatment of chronic diabetic wounds.
Hydrogels are three-dimensional cross-linked polymer networks that can absorb and retain a large amount of water or biological fluids. They have a range of physical and chemical properties that make them suitable for various applications in different fields in real life . Hydrogels have been extensively applied in the biomedical field due to their biocompatibility. They can be used as scaffolds for tissue engineering, drug delivery systems, and contact lenses. Hydrogels can also be engineered to respond to external stimuli such as temperature, pH , or light, enabling controlled drug release. In agriculture, hydrogels can be incorporated into soil to improve water retention and nutrient availability for plants. They help reduce water usage, prevent soil erosion, and promote plant growth by providing a favorable environment for roots . In the environmental remediation field, hydrogels can be utilized for wastewater treatment and environmental cleanup. They can absorb and remove contaminants from water, including heavy metals and organic pollutants . Hydrogels are considered promising wound dressings because of their softness comparable to that of the extracellular matrix (ECM) and their ability to mildly absorb exudates . Due to their porous microstructure, hydrogels in particular have a high loading capacity . Therefore, it is highly desirable to construct a hydrogel dressing loaded with multifunctional materials or drugs to simultaneously address the aforementioned issues . In fact, a variety of multifunctional hydrogels for wound repair have been developed that contain high amounts of oxygen for resolving hypoxia, polyphenolic substances or nanozymes for ROS scavenging, and macrophage polarity-modulating molecules for regulating the immune microenvironment . For example, Zhao et al. developed a therapeutic wound dressing, namely, , utilizing a biomimetic hydrogel system and modified hydrogen peroxidemimicking nanozymes. The hydrogel is engineered to simultaneously match the mechanical and electrical signals of the skin while possessing oxidative capability activated by . Wu et al. prepared a versatile dynamic Schiff base and borate ester cross-linked glycopeptide hydrogel that could continuously generate oxygen, promote M2 polarization of macrophages, and eliminate reactive oxygen and nitrogen species . Zhang et al. prepared an injectable hydrogel based on platelet-rich plasma and laponite that could accelerate wound healing by promoting macrophage polarization and angiogenesis in full-thickness skin . Clearly, the current design for multifunctional hydrogels entails significant issues, such as complex separation, tedious preparation, low synergistic efficiency, and limited space-time control. Therefore, there is an urgent need for a hydrogel dressing with a simple composition but a procedural therapeutic strategy. Gelatin methacryloyl (GelMA) is a dual-functionalized gelatin obtained through the reaction between aminosylated gelatin and methacrylic anhydride. The abundant amino groups distributed along the main chain of gelatin provide rich reactive sites for methacrylic anhydride. Methacrylic anhydride, bound to amino groups, can further react with each other to form three-dimensional structures suitable for cell growth and differentiation in scientific research related to technology. GelMA has been demonstrated to possess excellent biocompatibility, cell adhesion properties, and mechanical performance. It is widely applied in tissue engineering, drug delivery, 3D printing, and other fields .
Comparing the recombination of active components to a single microbial active cell, the latter offers a wider range of functions and is more easily programmable . Microorganisms have played a crucial role in advancing agricultural, industrial, and public health research by providing humans with advanced principles and methodologies . First, in cases of nonsymbiosis, various types of microorganisms engage in interactions that inhibit the growth of one another, thereby contributing to antibacterial activity in wound healing. Second, oxygen plays a critical role in multiple aspects of wound healing, such as cell
proliferation, migration, adhesion, blood vessel development, and tissue regeneration. It is widely believed that microorganisms, such as Chlorophyta, Haematococcus (HEA), and Archaea, played a fundamental role in the emergence and evolution of life on Earth by producing oxygen through photosynthesis. Rather than relying on conventional oxygen supply materials, such as hemoglobin, metal oxides, peroxides, and other oxygen delivery systems , which have limited and uncontrollable load capacities and release rates, these organisms are photoautotrophic. They utilize photosystems for photosynthesis, enabling them to produce oxygen. For example, Zhou et al. developed a bioactive hydrogel strategy for inhibition of methicillin-resistant Staphylococcus aureus. The bioactive hydrogel was composed with the berberine loaded living microalga Spirulina platensis and carboxymethyl chitosan/sodium alginate, which could release BBR and and produce ROS, mediating chemophotodynamic therapy for methicillin-resistant Staphylococcus aureus infection . In addition, numerous metabolites and cells from microorganisms can regulate the immune microenvironment and act as antioxidants; these substances are also used in a variety of food and drug additives . Astaxanthin, a metabolite of HEA (3,3′-dihydroxy-, -carotene-4,4′-dione, AST), has high ROS scavenging and antioxidant activity . It has been reported that ASTs, such as monothiol glutaredoxin, glutaredoxin (GRX), thioredoxin (TRX), thioredoxin reductase (TrxR), ferritin, monodehydroascorbate reductase (MDAR), peroxiredoxin (PrxR), glutathione peroxidase (GPX), glutathione reductase (GR), catalase (CAT), superoxide dismutase (SOD), and ascorbate peroxidase (APX), have various ROS scavenging enzyme activities . In addition, AST can inhibit the expression of inflammatory cytokines (COX-2, TNF- , IL-6, and IL-1 ) . Consequently, ASTs possess considerable potential and offer promising applications in human health against a broad spectrum of diseases. AST upregulates low-density lipoprotein receptor-related protein-1, which can bind to and downregulate p65, p-c-Jun, and NF-Kb, thereby blocking activation of the JNK and NF-кB pathways. Inhibiting the activation of JNK1 and NF-кB and promotes M2 polarization of macrophages . In recent years, there has been an increasing interest in utilizing microalgae, such as HEA, for the production of the natural antioxidant AST. This is primarily attributed to their faster growth rates and higher biomass production capabilities . Additionally, HEA is thought to possess the greatest ability to accumulate natural AST under conditions of environmental stress . Based on the progressive and controllable oxygen evolution, as well as AST release capabilities of HEAs, there is tremendous long-term potential for their application in artificial scaffolds for complex tissue healing, myocardial oxygenators, and as adjuncts in tumor therapy. As far as we know, there have been no reports of utilizing microorganisms for progressive oxygenation and microenvironmental regulation in the treatment of chronic diabetic wounds.
Here, we develop a simple hydrogel by encapsulating active HEA cells in conventional GelMA gel and established a system for local procedural therapy (Fig. 1). To increase the efficiency of oxygen and AST release, HEA spheroplasts are first prepared, loaded with GelMA solution, and photocured in situ to better establish irregular wounds. According to the light power density, an abundance of chlorophyll mediates the transition between oxygen production via photosynthesis and heat production via the photothermal conversion effect in green HEA (GHEA) cells. Therefore, in the initial phase of treatment, efficient photothermal conversion under 658 nm laser irradiation with a power density can swiftly eliminate bacteria. At the second stage of treatment, severe hypoxia in the wound is resolved by providing a constant oxygen supply for photosynthesis under 658 nm laser irradiation with a power density. With deterioration of the culture environment (bound in hydrogel) or external stimulation (laser irradiation), AST gradually accumulates, turning HEA cells red (RHEA) and endowing them with a potent capacity to modulate the wound microenvironment. Subsequently, HEA cells routinely release
Fig. 1 | Schematic illustration of the preparation and mechanism of the HEA@Gel-based wound healing strategy. Irregular wound dressings were obtained by in-situ photocuring. By modulating light intensity, HEA@Gel can be
programmed to perform antibacterial activity, oxygen supply, ROS scavenging, and immune regulation. HEA Haematococcus, AST astaxanthin.
AST to persistently scavenge ROS and regulate macrophage polarization. The healing of infected diabetic wounds is significantly accelerated by an all-in-one procedure that eliminates bacteria, supplies oxygen, scavenges ROS, promotes angiogenesis and facilitates antiinflammatory effects via the polarization of M2 macrophages.
Results
HEA@Gel preparation and characterization
We designed a living microalgal-based procedural hydrogel dressing (HEA@Gel) to easily mask the wound and deliver soluble oxygen and AST into the wound bed via in situ photocuring (Fig. 2a). To verify the concept of procedural therapy, the most obvious evidence, the change in color of HEA cells, was identified and is displayed in Fig. 2b-e. When the necessary nutrients are plentiful, HEA cells grow rapidly and appear green due to the presence of chlorophyll (Fig. 2b). With the consumption of nutrients and exposure to light, the growth rate of HEA cells decreased, and AST gradually accumulated (Fig. 2c, d). As AST continued to accumulate, the HEA cells became distinctly darker red (Fig. 2e). The solid and thick cell wall of HEA cells results in their low permeability, which prevents AST from being effectively released
from within the cells (Fig. 2 f. To remove the walls of HEA cells, we prepared HEA protoplasts using a mild enzymatic method (cellulase and pectinase). As shown in Fig. 2g and Supplementary Fig. 1, HEAtreated protoplasts retained their morphology, structure and activity after their cell walls were removed. Interestingly, after a period of culture, HEA protoplasts secreted numerous 200 nm diameter vesicles, which promoted the effective release and delivery of hydrophobic AST (Fig. 2h, i). To demonstrate the presence of AST within secretory vesicles, the vesicles were stained with Dil dye for membrane labeling. Due to the inherent fluorescence of AST, AST and the vesicles were observed under a laser confocal microscope, as shown in Supplementary Fig. 2, revealing clear colocalization of AST and the vesicles. To better compare the effects of the presence of the cell wall on AST release, equal amounts of HEA (with the cell wall) and HEA protoplasts (without the cell wall) were separately dispersed in 100 mL of deionized water. The released AST content was measured by using the standard curve of AST shown in Supplementary Fig. 3. As shown in Supplementary Fig. 4a, the rate of AST release from HEA protoplasts was significantly greater than that from HEA protoplasts with retained cell walls. Furthermore, over time, there was a continuous increase in
Fig. 2 | HEA@Gel preparation and characterization. a Schematic representation of the benefits of photocuring GelMA. b-e Photographs of various stages of the
images of (l) Gel, (m) GHEA@Gel, and (n) RHEA@Gel. Scale bars, . The insets in (l-n) represented the photographs of of Gel, GHEA@Gel, and RHEA@Gel, respectively. For these morphological characterizations of the fabricated Gel, GHEA@Gel, and RHEA@Gel, each experiment was repeated three times independently with similar results.
the amount of AST released from HEA protoplasts. In contrast, in HEA with retained cell walls, the rate of AST release significantly decreased after 6 h . This phenomenon may be attributed to the hindrance of further AST release by the presence of cell walls following the release of some easily releasable AST in the earlier stage. Additionally, by collecting the released AST in a centrifuge tube, the color change over time was visually observed (Supplementary Fig. 4b).
GelMAs with positive charges, injectables, and in situ photocuring demonstrated enormous potential for the treatment of irregular wounds in this study. Consequently, the GelMA solution was combined with HEA cells and then coated on the wound; after 5 s of blue light irradiation, the hydrogel was cured and fit perfectly into the wound site. Figure 2j, k show images of pure Gel, GHEA@Gel, and RHEA@Gel, as well as images of the photocuring mechanism. The morphology and structure of the as-prepared Gel and HEA@Gel were subsequently examined via cryo-scanning electron microscopy (FEI Quanta 450). As shown in Fig. 2l, the gel sample exhibited an evidently porous structure. GHEA@Gel and RHEA@Gel retained their polyporous structure, guaranteeing and AST transportation (Fig. 2m, n).
Antibacterial activity of the GHEA@Gel
Due to the constant exposure of DU wounds to the external environment, there is a high risk of external bacterial infection and significantly delayed wound healing. As previously reported , different kinds of microorganisms interact with each other to inhibit each other’s growth in the case of nonsymbiosis, which contributes to wound antibacterial activity. To verify the nonsyniotic effect of the combination treatment, the proliferation of Escherichia coli (E. coli) and Staphylococcus aureus (S. aureus) after cocultivation with GHEA without light irradiation was measured. As shown in Supplementary Fig. 5, obvious inhibition of bacterial growth was observed for both E. coli and S. aureus, in which the survival rates of E. coli and S. aureus, respectively, remained at only and , respectively, when the density of the GHEA cells reached . In addition, in light of the possibility of photothermal conversion of HEA cells, we further evaluated the in vitro antibacterial activity of GHEA by assessing the photothermal effect under 658 nm laser irradiation (Fig. 3a). As shown in Fig. 3b, the photothermal effect of HEA@Gel was proportional to the HEA cell density. The temperature of the water only increased by under 658 nm laser irradiation.
When the density of the GHEA cell reached , the temperature of the GHEA@Gel increased by , indicating a robust photothermal effect. E. coli and S. aureus were subsequently chosen as typical bacteria for investigating the PTT-mediated antibacterial activity of GHEA@Gel. After various processing steps, the optical density (OD) of the bacterial suspension at 600 nm was measured to assess bacterial activity. As shown in Fig. 3c-f, the photothermal effect of GHEA@Gel in combination with laser irradiation significantly inhibited bacterial growth. To further determine the antibacterial activity of the GHEA@Gel, an agar plate test was performed (Fig. 3g). After GHEA@Gel and 658 nm laser irradiation, the number of bacterial colonies on the agar plate was negligible. We hypothesized that the rapid rupture of the bacterial membranes could be the result of a local hyperthermal effect triggered by the powerful photothermal effect of the GHEA@Gel. We then evaluated the integrity of the bacterial membrane using SEM (Fig. 3h). Normal bacteria were observed to have a rhabditiform or spherical morphology with a smooth surface. Following treatment with GHEA@Gel and laser irradiation at 658 nm , the cell membranes of the bacteria were severely compromised, with obvious surface collapse. By destroying the bacterial membrane, GHEA@Gel, which has a 658 nm laser-mediated photothermal effect, was rapidly sterilized. The photothermal conversion of the GHEA@Gel was then evaluated further in vivo. As depicted in Fig. 3i, obvious and excellent photothermal conversion was observed in the DU wounds of mice, where a 658 nm laser could increase the temperature to in
5 min . A temperature of ensured that the bacteria in the wound were effectively killed, coupled with a nonsymbiosis effect on the HEA cells. However, the disadvantages of this process must also be considered, such as the damage caused by high temperature to normal skin tissue at the wound site. To investigate the damage to normal skin at , the skin of the mice was scalded with a constant temperature of electric soldering iron at for 5 min . Subsequently, the skin damage at the site of the burn was recorded. The results showed that slight red scald marks were left on the skin of the mice after they were scalded at , indicating that could cause damage to the skin cells of the mice. After 4 days of burn treatment, the red fluorescence gradually faded (Supplementary Fig. 6), demonstrating that the organism has good self-healing ability at on mild scald. To further investigate the damage caused by to normal skin cells, the epithelial cells were exposed to for 5 min , after which FCM was used to detect cell death. The results showed that, compared with the control treatment, caused bearable cell death (Supplementary Fig. 7). In summary, although a high temperature of had some slight effects on normal cells while killing infected bacteria, during the actual operation process, because the duration of was very short (less than 1 min ) and because the body has a strong ability to self-heal minor burns, these side effects are completely acceptable in the treatment of wound infection.
release from the GHEA@Gel
The activity of the GHEA@Gel was initially confirmed by exposing it to red light ( ) and measuring the soluble oxygen concentration with a microelectrode (Fig. 4a). The soluble oxygen concentration in the GHEA@Gel increased from 0 to more than within 20 min under adequate light conditions, while in the dark, the soluble oxygen concentration decreased from 8 to within 20 min (Fig. 4b). These findings indicate that the GHEA in the gel is fully active and capable of photosynthesis and respiration. In a certain range, the oxygen production rate was positively correlated with the GHEA@Gel concentration, and the optimal concentration was mL (Fig. 4c). The above oxygen generation experiment was conducted under a light intensity of . To investigate whether GHEA can produce oxygen under laser irradiation at an intensity of , GHEA and GHEA@Gel were subjected to 658 nm laser irradiation at an intensity of , and the changes in oxygen availability in the system were measured every 2 min for a total duration of 30 min . The results, as shown in Supplementary Fig. 8, indicated that the generation curve was very similar to the curve at . To investigate whether the photothermal effect of a high light intensity of affects the oxygen production capacity of GHEA in GHEA@Gel, GHEA@Gel samples containing GHEA cells at different concentrations were placed under 658 nm laser irradiation for 5 min . Subsequently, after the GHEA temperature decreased to room temperature, a lowintensity laser ( ) was used to irradiate the GHEA@Gel, after which the generation of oxygen during the irradiation process was measured. As shown in Fig. 4c, although the photothermal effect weakens the rate of oxygen production in the GHEA, a consistent and stable oxygen output can be detected. The ability of GHEA to produce oxygen over a long period of wound healing treatment is critical to its effectiveness. According to the GHEA oxygen-producing process simulated in vitro over a period of 20 days, the ability of GHEA cells to produce oxygen tends to slowly weaken over time during repeated laser irradiation. However, after 20 days of intermittent laser irradiation, the GHEA cells still retained good oxygen production capacity (Supplementary Fig. 9). Multiple cell activities, such as fibroblast proliferation, keratinocyte migration, and endothelial cell differentiation, are concurrently involved in wound healing; therefore, we further examined the effect of GHEA@Gel on human skin fibroblasts (HSFs), human immortalized keratinocytes ( HaCaTs ), and human umbilical vein endothelial cells (HUVECs). We first discovered that exposure to
Fig. 3 | GHEA@Gel photothermal conversion and sterilization. a Schematic illustration of the photothermal conversion and sterilization of GHEA@Gel. b Photothermal transformation of GHEA@Gel. The data are presented as the mean s.d. ( independent experiments). Quantitative measurement of . coli cells treated with GHEA and a 658 nm laser ( ). The data are presented as the mean s.d. ( biologically independent cells). Statistical differences were analyzed by Student’s two-sided -test. e, Quantitative measurement of . aureus cells treated with GHEA and a 658 nm laser . The data are presented as
the mean s.d. ( biologically independent cells). Statistical differences were analyzed by Student’s two-sided -test. g The corresponding digital images of . aureus and . coli bacterial colonies grown on LB agar plates subjected to various treatments. Each experiment was repeated independently three times with similar results. The corresponding SEM images of S. aureus and E. coli bacteria subjected to various treatments. Each experiment was repeated independently three times with similar results. i Photothermal images of mice receiving various treatments. Each experiment was repeated independently three times with similar results.
33 mM glucose and induced hypoxia accompanied by overexpression of hypoxia-inducible factor- ( ), indicating the effect of hyperglycemia on hypoxia (Fig. 4d and Supplementary Figs. 10-13). We subsequently determined whether GHEA@Gel could significantly reverse cell hypoxia and result in an reduction in HIF-1 in HSFs (Fig. 4e). The same effect was observed for HUVECs and HaCaTs treated with GHEA@Gel (Supplementary Figs. 10-13). In
addition to qualitative fluorescence data validation, quantitative analysis via a flow cytometer is desirable. As shown in Supplementary Fig. 14, the flow cytometry data exhibited a trend similar to that of the fluorescence imaging results. The HSFs treated with GHEA@Gel under light conditions exhibited a significantly greater proliferation rate than did those treated with high glucose or even (Fig. 4f, g), indicating the significant potential of GHEA@Gel for forming granulation tissue
Fig. 4 | release from the GHEA@Gel. a Schematic representation of release from GHEA@Gel. b Comparison of the dissolved release of GHEA@Gel under light and dark conditions. The data are presented as the mean s.d. ( independent experiments). The release of dissolved at various GHEA concentrations and laser intensities. d, e GHEA@Gel decreased HIF-1 expression in HSFs induced by high glucose concentrations. Scale bars, . Each experiment was repeated independently three times with similar results. The data are presented as the mean s.d. ( independent cells). Statistical differences were analyzed by Student’s two-sided -test. f, g Proliferation of HSF cells treated with 33 mM glucose under hypoxia ( 6 h ) for various durations. Scale bars, . Each experiment was
repeated independently three times with similar results. The data are presented as the mean s.d. ( independent cells). Statistical differences were analyzed by Student’s two-sided -test. Representative images and quantification of the migration of HaCaT cells. The scale bars are . Each experiment was repeated independently three times with similar results. The data are presented as the mean s.d. ( independent cells). Representative images and quantification of HUVEC tube formation. Scale bars, . Each experiment was repeated independently three times with similar results. The data are presented as the mean s.d. ( independent cells). Statistical differences were analyzed by Student’s two-sided -test.
by releasing dissolved oxygen to promote HSF proliferation. In addition, GHEA@Gel promoted cell proliferation in HUVECs and HaCaTs (Supplementary Figs. 15, 16). Scratch healing of keratinocytes is ideally the core of re-epithelialization processes during chronic wound healing. In the scratch test, we compared cell migration under hypoxic conditions, supply conditions, and soluble oxygen generated by GHEA@Gel under light irradiation. Fig. 4h, i show that, compared with an supply, a hypoxic environment significantly inhibited the migration of HaCaT cells. Notably, coculturing cells with GHEA@Gel in a hypoxic environment and subsequent exposure to light significantly increased the migration rate of the cells, indicating that the soluble oxygen produced by GHEA@Gel could promote epithelialization, thus facilitating chronic wound healing. Endothelial cells are an integral part of blood vessels and play a crucial role in vascularization. We performed tube formation tests using microscopy. The Matrigel tube formation assay, performed to assess angiogenic capacity, revealed a significantly greater tube length and number of branches after 6 h in the GHEA@Gel-treated HUVECs than in the -treated HUVECs (Fig. 4j, k). The results of a series of in vitro assays confirmed that the increase in angiogenesis was caused by the soluble oxygen produced by the GHEA@Gel.
ROS scavenging by RHEA@Gel
Diabetic wounds are characterized by enhanced ROS levels and sustained oxidative stress. The addition of antioxidants to hydrogels has been shown previously to reduce oxidative cellular damage, which speeds up chronic wound healing. Due to the unique molecular structure of AST, RHEA-based hydrogels are promising for promoting diabetic wound healing. AST is the strongest natural antioxidant (Fig. 5a). The clearance of , and was subsequently measured to determine the antioxidative performance of RHEA@Gel (Fig. 5b-d). To compare the ROS scavenging ability of RHEA@Gel with greater precision, the natural antioxidant vitamin C (VC) and the chemosynthetic AST were chosen for comparison. As shown in Fig. 5b-d, our prepared RHEA@Gel exhibited the strongest ability to scavenge ROS in the form of , and at the same concentration. Using fluorescence microscopy, the ROS scavenging ability of the RHEA@Gel was subsequently confirmed. To simulate the inflammatory microenvironment of diabetic wounds, was chosen for further experiments. Specifically, solution was added during the HSF cell culture process. Subsequently, after the addition of an ROS scavenger (AST or RHEA@Gel), the cells were incubated for a certain period of time to measure the level of intracellular ROS. Figure illustrate that RHEA@Gel is an effective regulator of intracellular ROS levels in HSFs. It is evident from the results that after was added to the cell culture, there was a significant increase in the intracellular ROS levels, as indicated by the increase in the intensity of the green fluorescence. However, when AST or RHEA@Gel was added to cells incubated with , the intracellular ROS concentrations decreased compared with those in the group. Additionally, comparable phenomena were observed for HUVECs and HaCaTs (Supplementary Figs. 17, 18). These results indicate that the ability of RHEA@Gel to absorb free radicals is not only significantly greater than that of natural VC but also greater than that of chemically synthesized
AST. The following are the reasons for this outcome. First, AST is a hydrophobic antioxidant that coagulates in aqueous solutions and biological fluids, reducing its bioavailability. Second, the antioxidant activity of AST is related to its molecular structure, and an incorrect molecular structure of chemically synthesized AST also affects its antioxidant activity. Even though the naturally occurring AST produced by RHEA is perfectly formed, its hydrophobicity also affects its antioxidant capacity. In the preliminary characterization, it was determined that loading RHEA protoplasts into the hydrogel generated numerous membrane-wrapped vesicles approximately in size, similar to exosomes (Fig. 2g, h). The reason why RHEA@Gel has the strongest antioxidant activity is likely due to the precise configuration of naturally secreted AST, and the presence of cell-wrapped exosome vesicles in RHEA@Gel significantly increases the water solubility of AST and enhances its bioavailability.
Based on these findings, the effect of RHEA@Gel on HSF proliferation was further investigated, and RHEA@Gel induced a dramatically greater proliferation rate in HSFs than in and AST (Fig. 5g, h), indicating that the AST released from RHEA@Gel has significant potential to form granulation tissue by promoting HSF proliferation. In addition, the ability of RHEA@Gel to promote cell proliferation was also observed for HUVECs and HaCaTs (Supplementary Figs. 19, 20). Next, we examined the influence of AST from RHEA@Gel on ROSinduced HaCaT cells. ROS significantly inhibited HaCaT cell migration, whereas RHEA@Gel stimulation significantly increased HaCaT cell migration (Fig. 5i, j). RHEA@Gel and AST had significant effects on cell migration. We performed tube formation tests using microscopy. The Matrigel tube formation assay, performed to assess angiogenic capacity, revealed a significantly greater tube length and greater number of branches after 6 h in the RHEA@Gel-treated HUVECs than in the AST-treated HUVECs (Fig. 5k, l). In addition, we discovered that the AST released by RHEA@Gel could increase the expression of peroxidase and SOD in cells, thereby scavenging intracellular ROS. These results suggested that the increase in angiogenesis observed in a series of in vitro assays was a result of the release of AST from the RHEA@Gel.
Anti-inflammatory properties of RHEA@Gel
Macrophages are an important type of immune cell in the immune system that functions by phagocytosis, cytotoxicity, and cytokine secretion. These cells can be activated and polarized into different phenotypes to adapt to various physiological and pathological conditions. M1 macrophages represent a typical proinflammatory phenotype and play a crucial role in antimicrobial and antitumor immunity. The activation of these cytokines is mediated primarily by microbial components (such as lipopolysaccharides) and cytokines (such as IFN ). M1 macrophages kill pathogenic microorganisms by producing damaging molecules such as reactive oxygen and nitrogen species and secreting proinflammatory cytokines, thereby triggering local inflammatory responses. M2 macrophages represent a noninflammatory phenotype and are involved primarily in tissue repair, regeneration, and modulation of immune responses. The activation of M2 macrophages is mediated mainly by cytokines (such as interleukin-4 and interleukin-13) and other molecules (such as VEGF and transforming growth factor-beta). M2 macrophages exert their effects by
suppressing inflammation, promoting tissue repair, and regulating immune balance. The regulation of these macrophage phenotypes is influenced mainly by various receptors and signaling pathways. For example, Toll-like receptors, nuclear receptors, cytokine receptors, and others can activate specific signaling pathways to regulate macrophage polarization and functions. Different types of pathogens, cellular damage signals, and immune regulatory molecules also
participate in this process, forming a complex signaling network. Depending on the role of different macrophage phenotypes, additional macrophages must be polarized to the M2 phenotype during wound healing. However, the phenotypic switch from M1 to M2 is compromised in diabetic wounds, resulting in sustained inflammation and sluggish wound healing. AST is a natural compound extracted from HEA that has anti-inflammatory properties. We sought to
Fig. 5 | ROS scavenging ability of RHEA@Gel. a Schematic representation of ROS scavenging by RHEA@Gel. b , (c) OH -, and (d) -scavenging performance in response to various treatments. The data are presented as the mean s.d. ( independent cells). Statistical differences were analyzed by Student’s two-sided test. e, f ROS scavenging ability of RHEA@Gel in HSF. Scale bars, . Each experiment was repeated independently three times with similar results. The data are presented as the mean s.d. ( independent cells). Statistical differences were analyzed by Student’s two-sided -test. The proliferation of HSF cells treated with 33 mM glucose and in different groups. Scale bars, . Each experiment was repeated independently three times with similar results. The data are presented as the mean s.d. ( independent cells). Statistical differences
were analyzed by Student’s two-sided -test. Images and quantification of the migration of HaCaT cells. Scale bars, . Each experiment was repeated independently three times with similar results. The data are presented as the mean s.d. ( independent cells). , I Representative images and quantitative analysis of tube formation in HUVECs. Scale bars, . Each experiment was repeated independently three times with similar results. The data are presented as the mean s.d. ( independent cells). Peroxidase and ( ) superoxide dismutase expression in the different groups. The data are presented as the mean s.d. ( independent cells). Statistical differences were analyzed by Student’s two-sided -test.
investigate whether the anti-inflammatory effect of AST is due to the regulation of macrophage phenotypic transition and the modification of the immune microenvironment in wounds (Fig. 6a). Briefly, M0 macrophages were stimulated with lipopolysaccharide (LPS) and interferon IFN to switch to the M1 phenotype as a positive control, while those stimulated with interleukin-4 (IL-4) to switch to the M2 phenotype served as a negative control. CD206 (M2 phenotype) and CD86 (M1 phenotype) macrophages were identified by immunofluorescence staining to confirm the polarization status of the macrophages under different stimulation conditions. More CD206+ macrophages and fewer CD86+ macrophages were detected in the GHEA@Gel group than in the control group, indicating that release may have increased M2 activation (Fig. 6b, c and Supplementary Fig. 21). Compared to those in the control and GHEA@Gel groups, macrophages treated with RHEA@Gel displayed significantly more CD206 and less CD86 signaling, indicating that the AST released by RHEA is essential for macrophage polarization. Moreover, the GRHEA@Gel group displayed the highest CD206 staining and the lowest CD86 signal, further validating that liberation and AST synergistically induced M2 polarization of macrophages. To further validate the ability of HEA to promote the transformation of M1 macrophages into M2 macrophages, flow cytometry was used to detect macrophages in the different treatment groups. The results shown in Fig. 6d were consistent with the confocal microscopy results (Fig. 6d and Supplementary Fig. 22).
In vivo evaluation of infected diabetic wound healing
The therapeutic approach is depicted in Fig. 7a. Benefiting from the photothermal conversion effect of HEA@Gel under 658 nm laser irradiation at an intensity of and the nonsymbiosis effect between HEA and S. aureus, the infected diabetic wound was sterilized first. After bactericidal treatment, every subsequent day, a 658 nm laser was used to illuminate the wound at a light intensity of for 30 min in the following 10 days. Low-intensity laser irradiation has two effects. First, GHEA activates photosynthesis to produce to improve the hypoxic wound microenvironment and accelerate vascular regeneration and wound healing. The second is to promote the release of AST from RHEA to reduce the levels of overexpressed ROS and inflammatory factors and regulate macrophage M 2 polarization in wounds. Hence, HEA@Gel likely promoted fibroblast proliferation, keratinocyte migration, endothelial cell differentiation, and the progressive acceleration of infected wound healing in diabetic patients.
Figure shows macroscopic images of the wounds, the wound closure traces at various time points, and the corresponding quantitative wound healing times. On day 3, the wounds in the GRHEA@Gel with laser group were smaller than those in the other groups. On day 6, the wound area of the G-RHEA@Gel with laser group decreased significantly, whereas the wound sizes in the control, Gel, GHEA@Gel, and RHEA@Gel with laser groups remained at , , and , respectively. On day of the infected wounds in the group treated with G-RHEA@Gel and the laser had closed. In comparison, the control and gel groups still had substantial wound areas, while the GHEA@Gel and RHEA@Gel groups had faster healing. The
results indicated that treatment with G-RHEA@Gel and laser irradiation promoted the healing of . aureus-infected diabetic wounds.
As depicted in Fig. 7g, histological examination of the healed wound was performed by both hematoxylin-eosin (H&E) and Masson’s trichrome (MT) staining. On day 3, none of the groups exhibited epidermal formation or collagen deposition. All the groups exhibited epidermal regeneration on day 9, but the epidermis of the GRHEA@Gel with laser group was more natural and mature than that of the other groups. As shown in Fig. 7e, the epidermal thicknesses of the control, Gel, GHEA@Gel, RHEA@Gel, and G-RHEA@Gel groups were , and , respectively. H&E staining demonstrated that infected diabetic wounds in the G-RHEA@Gel group treated with a laser exhibited satisfactory wound healing with accelerated epidermis and dermis formation. In addition, the collagen deposition level on day 9 is shown in Fig. 7f. Collagen deposition was greater in the GRHEA@Gel with laser group than in the other groups. The H&E and MT staining results suggested that treatment of G-RHEA@Gel with a 658 nm laser could expedite the healing of infected diabetic wounds by promoting the formation of epidermis, facilitating dermal tissue regeneration, and increasing collagen deposition. In addition, immunohistochemical staining using an anti-Staphylococcus aureus antibody was applied to assess the bacterial contamination of the wounds. As shown in Fig. 7h, on day 20, the control group and pure gel group wound tissue contained a high number of bacteria (green fluorescence). When HEA@Gel was combined with 658 nm laser irradiation, it was particularly difficult to detect bacteria in the GHEA@Gel, RHEA@Gel, and G-RHEA@Gel groups due to the powerful antibacterial effect of the treatment. The excellent antibacterial effect mediated by HEA could be attributed mainly to the nonsymbiotic effect between bacteria and HEA and the photothermal bactericidal effect of HEA upon irradiation with a 658 nm laser. To further validate this conclusion, the skins of plants subjected to different treatments were collected and homogenized on day 20. Serially diluted homogenates were plated on LB agar to quantify the bacterial colonies. As shown in Supplementary Fig. 23, a large number of S. aureus colonies appeared in the control group and the hydrogel group. The number of colonies in the GHEA@Gel group treated with 658 nm laser ( ) irradiation and the RHEA@Gel group treated with 658 nm laser ( ) irradiation were greatly reduced due to the photothermal bactericidal effect and nonsymbiotic effect, respectively. Therefore, in view of the above two antibacterial effects, there was almost no colony formation in the G-RHEA@Gel group after treatment.
Immunofluorescence analysis
To demonstrate the efficacy of HEA@Gel, the ability of GHEA to promote wound healing was examined via HIF-1 immunofluorescence staining. For the control and gel groups, significant HIF-1 expression was noted. Due to the efficient photosynthesis of GHEA, the HIF-1 level in the GHEA@Gel group was significantly lower than that in the control group, indicating a significantly greater oxygen supply (Fig. 8a, e). Compared to those in the GHEA@Gel group, the oxygen supply in the RHEA@Gel group was lower due to the high AST accumulation and reduced photosynthesis. Almost no HIF-1 expression was detected in
Fig. 6 | Regulation of macrophage polarization by RHEA@Gel. a Schematic illustration of the regulation of macrophage polarization by RHEA@Gel. b Relative quantification of the ratio of M2 to M1 macrophages. The data are presented as the mean s.d. ( independent cells). Statistical differences were analyzed by Student’s two-sided -test. c Characteristic fluorescence images of Raw264.7 cells
stained with CD206 (green) and CD86 (pink) under inflammatory stimulation. Scale bar: . d Flow cytometry analysis of macrophage polarization after different treatments. Each experiment was repeated independently three times with similar results.
the G-RHEA@Gel group, confirming the high oxygen supply of GRHEA@Gel. By releasing dissolved oxygen, GHEA@Gel with 658 nm laser irradiation was able to alleviate local hypoxia. To examine the ability of GHEA@Gel to scavenge ROS in vivo, a dihydroethidium (DHE) probe was used to measure the ROS concentration in the wound area. The control and gel groups exhibited bright and high-intensity red fluorescent signals, while the intensity of fluorescence decreased significantly in the GHEA@Gel group, as shown in Fig. 8b, f. In the
RHEA@Gel and G-RHEA@Gel groups, hardly any red fluorescence was detected. According to these results, RHEA@Gel is capable of efficiently scavenging ROS to alleviate oxidative stress.
As depicted in Fig. 8c, g, the underlying biological mechanisms of wound healing were investigated by epidermal growth factor (EGF) immunofluorescence staining on day 6. EGF expression was observed in both the GHEA@Gel and RHEA@Gel groups, indicating that both the oxygen supply capability of GHEA@Gel and the ROS scavenging
Fig. 7 | Effects of HEA@Gel on enhancing wound healing in infected
diabetic mice. a Schematic depiction of the sequence of animal experiments conducted to evaluate the therapeutic efficacy of HEA@Gel. b Photographs of representative wounds from various treatment groups. c Quantitative investigation of the wound area. Each experiment was repeated five times independently with similar results. d Statistical analysis of the healing time. The data are presented as the mean s.d. ( independent mice). Statistical differences were analyzed by
Student’s two-sided -test. e, f Relative quantitative analysis of collagen deposition and epidermal thickness on day 9. The data are presented as the mean s.d. ( independent mice). Statistical differences were analyzed by Student’s two-sided test. g HE staining on days 3 and 9. Each experiment was repeated five times independently with similar results. Immunohistochemical staining of the wound on day 20. The scale bar is . Each experiment was repeated five times independently with similar results.
Fig. 8 | Analysis of the processes and mechanisms of wound healing for various treatments. a HIF-1 expression, (b) ROS content, (c) EGF expression, and (d) VEGF expression on day 6 under various treatments. The scale bar is . Each experiment was repeated independently three times with similar results. Quantitative investigation of HIF-1 , ROS, EGF, and VEGF expression on day 6 in response to various treatments. The data are presented as the mean s.d. ( independent mice). Statistical differences were analyzed by Student’s two-sided test. Double immunofluorescence staining of (i) -SMA (yellow) and CD31 (green)
and (j) the fibroblast markers vimentin (green) and collagen (red) in response to various treatments on day 6. Scale bar, . Each experiment was repeated independently three times with similar results. Quantitative investigation of CD31 expression, neovascularization, and collagen I deposition on day 6 in response to various treatments. The data are presented as the mean s.d. ( independent mice). Statistical differences were analyzed by Student’s two-sided -test.
capability of RHEA@Gel could promote EGF expression. The highest EGF expression was observed in the G-RHEA@Gel treatment group, indicating that oxygen supply and ROS scavenging are necessary conditions for epidermis formation. In addition, VEGF is an important downstream marker of HIF, and hypoxia suppresses VEGF expression. Figure 8d, h shows the results of VEGF immunofluorescence staining, which revealed a greater level of VEGF in the G-RHEA@Gel group than in the control group, indicating increased blood vessel formation due to the hydrogel, oxygen delivery, and ROS scavenging.
Angiogenesis is a crucial indicator of diabetic wound healing. Neovascularization was evaluated by immunofluorescence staining of a-smooth muscle actin and CD31. As depicted in Fig. 8i, k, the expression of CD31 in the GHEA@Gel and RHEA@Gel groups was significantly greater than that in the control and Gel groups. CD31 expression was highest in the G-RHEA@Gel group. The results demonstrated that G-RHEA@Gel-treated with laser irradiation possessed superior pro-vascularization capacity and promoted wound healing more effectively than the other hydrogels. Therefore, diabetic wound healing can be accelerated by promoting epidermal formation, collagen deposition, and neoangiogenesis. In addition, immunofluorescence staining for collagens and vimentin (a fibroblast marker) was performed to assess collagen deposition in granulation tissue (Fig. 8j, l, and m). ECM formation was facilitated by the increased deposition of matrix and the directional alignment of collagen fibers. The control group had the least amount of collagen, whereas the GRHEA@Gel group had higher levels of collagen deposition and orientational alignment, suggesting better collagen production favoring wound repair.
In vivo regulation of macrophage polarization
In the process of wound healing, macrophages play a critical role. The in vivo M2/M1 macrophage ratio was confirmed by immunofluorescence staining for CD206 and CD86 (Fig. 9a). Higher populations of M2 macrophages and lower populations of M1 macrophages were detected in the GHEA@Gel and RHEA@Gel groups, which resulted in a greater M2/M1 macrophage ratio (Supplementary Fig. 24), demonstrating decreased proinflammatory and enhanced antiinflammatory capacities. In particular, the G-RHEA@Gel group displayed the highest anti-inflammatory capacity, which was attributed to the combined antibacterial, oxygen supply, ROS scavenging, and antiinflammatory actions of GHEA and RHEA. CD206 (M2 macrophages) and (M1 macrophages) cells were quantified by flow cytometry, and the results were consistent with the immunofluorescence images (Fig. 9b and Supplementary Fig. 25). Nuclear factor-кB (NF-кB) is the central signaling pathway regulating inflammation by mediating the expression of downstream proinflammatory cytokines, including TNF, IL-6, and IL-1 , which further impedes tissue recovery. Neutrophils are the predominant effector cells in bacterial infections, and they persistently accumulate in diabetic wounds, thereby retarding the healing process. CXCL-1, a member of the CXC chemokine family, is involved in the migration and activation of neutrophils. At days 3 and 6 , the IL- , IL- 6 , TNF- and CXCL- 1 levels in the wounds were reduced in all the gel groups (Fig. 9c-f). The G-RHEA@Gel group exhibited marked decreases in the levels of IL- , IL- 6 , TNF- and CXCL- 1 compared with those in the other groups, indicating the good effectiveness of the combination treatment in alleviating inflammation. In contrast, IL-4 and IL-10 are regeneration-promoting cytokines that facilitate tissue repair, wound healing, axonal regeneration, and M2 macrophage polarization. The G-RHEA@Gel group expressed the highest levels of IL-4 and IL-10 among all the gel groups compared to those in the control group, indicating a remarkable pro-regenerative effect (Fig. ). On day 3 following treatment, the white blood cell (WBC) count, lymphocyte (Lymph) count, and neutrophil percentage (Gran) in the peripheral blood of the GHEA@Gel, RHEA@Gel, and GRHEA@Gel groups were significantly lower than those in the control
and hydrogel groups (Fig. 9i-k). Consequently, the synergistic antibacterial, ROS scavenging, oxygenation, and anti-inflammatory effects in the G-RHEA@Gel group could aid in significantly reducing wound inflammation. H&E staining was performed to detect tissue damage, and Supplementary Fig. 26 shows the biosafety of all the hydrogels for in vivo experiments. These results demonstrated that treatment with G-RHEA@Gel and laser stimulation is the optimal wound healing therapy. Furthermore, to assess the safety profile of the hydrogel, GRHEA@Gel was incubated with HaCaT cells. The results demonstrated excellent safety, as evidenced by the cell activity following coincubation with G-RHEA@Gel. Supplementary Fig. 27a shows that, compared with the control treatment, the G-RHEA@Gel treatment did not adversely affect cell activity. Additionally, staining the cells with calcein-AM and PI dyes and observing them using a confocal laser scanning microscope intuitively revealed that the cells maintained good activity after coincubation with G-RHEA@Gel (Supplementary Fig. 27b). Moreover, the healthy skin of the mice was covered with the G-RHEA@Gel hydrogel for 30 days. Histological examination via H&E and Masson staining revealed no evidence of skin damage following prolonged exposure to G-RHEA@Gel on the skin surface (Supplementary Fig. 28). To further explore the long-term safety of G RHEA@Gel, the levels of inflammatory factors, including TNF- , IL- , IL-4, IL-6, IL-10, and CXCL-1, and hematological indices, including WBC, lymphocyte, and granule counts, in the peripheral blood of the mice were detected. As shown in Supplementary Fig. 29, there was no significant difference between the G-RHEA@Gel-treated mice and the control group.
Discussion
Chronic diabetic wounds pose a grave threat to human life and health. The treatment of infected diabetic wounds is a very complex procedure involving bacterial infection, hypoxia, ROS hyperexpression, and inflammation, among other factors. A treatment strategy that is simple and effective is urgently needed. The combination of multifunctional microbial therapies and biocompatible hydrogels holds great promise for treating diabetic wounds. Due to its high efficiency in oxygen production and AST secretion, HEA has emerged as the most valuable microorganism for the treatment of diabetic wounds. However, strategies based on living HEA have not yet been investigated.
In this study, we devised a procedural treatment strategy based on living HEA hydrogels programmed to implement antibacterial, oxygen release, ROS scavenging, anti-inflammatory, and macrophage polarization regulation to promote rapid healing of diabetic wounds in a comprehensive manner. Using simple in situ photocuring technology, HEA@Gel could conform to any asymmetrical wounds. Upon exposure to high-intensity light ( ), the photothermal conversion of GHEA@Gel had effective antibacterial effects, revealing the initial phase of wound treatment. By regulating low-intensity light ( ), the GHEA@Gel photosynthetic system can continuously produce oxygen, effectively resolving the problem of hypoxia and promoting vascular regeneration, thereby achieving the objective of the second phase. Continuous light irradiation and the binding effect of the hydrogel network induced AST accumulation in HEA cells, resulting in a gradual change in color from green to red (RHEA). By secreting AST vesicles via exosomes, RHEA is able to efficiently remove excess ROS, enhance the expression of intracellular antioxidant enzymes, and directly polarize M2 macrophages, completing the third phase of infected diabetic wounds. By controlling the light intensity in vitro, the metabolism and function of HEA could be modulated on demand, facilitating programmed wound treatment. Experiments indicate that the living HEA hydrogel can enhance the proliferation and migration of cells, promote tube formation in vitro, and reinforce chronic diabetic wound healing in vivo. Microalgae gelbased treatment provides an advanced strategy for ameliorating chronic wound healing.
Fig. 9 | Regulation of macrophage polarization and the immune microenvironment by HEA@Gel in vivo. a Double immunofluorescence staining of CD206 (green) and CD86 (pink) on day 6 in response to various treatments. Scale bar, . Each experiment was repeated independently three times with similar results. b Analysis of the ratio of M2 to M1 macrophages in response to various treatments by flow cytometry on day 6. Each experiment was repeated
independently three times with similar results. In vivo wound concentrations of (c) TNF- , (d) IL-1 , (e) IL-6, (f) CXCL-1, (g) IL-4, and (h) IL-10 after 3 and 6 days of treatment. i White blood cell count, (j) lymphocyte count, and (k) neutrophil percentage (gran%) in peripheral blood. The data are presented as the mean s.d. ( independent mice). Statistical differences were analyzed by Student’s twosided -test.
Hydrogels loaded with active microalgal cells hold promise as diabetic wound excipients, offering advantages such as enhanced wound healing, antimicrobial properties, and biocompatibility. However, addressing the challenges of limited research, regulatory approval, and cost considerations is crucial for successful clinical transformation. Further research efforts and collaborations between academia, industry, and regulatory bodies are necessary to optimize this innovative approach and facilitate its integration into routine diabetic wound management strategies.
Methods
Ethical statement
This research complies with all relevant ethical regulations. All animal studies were conducted in accordance with the National Institute Guide for the Care and Use of Laboratory Animals. All procedures, including animal care, wound modeling, dosing, and termination, were approved by the Tianjin University Institute Animal Ethics Committee with the assigned approval number TJUE-2023-236.
Materials
HEA cells were supplied by Freshwater Algae Culture Collection at the Institute of Hydrobiology, FACHB. GelMA and photoinitiator LAP were purchased from Suzhou Yongqinquan intelligent equipment Co., LTD. AST ( (HPLC)) were purchased from Solarbio. 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA), and Calcein/PI Cell Viability/ Cytotoxicity Assay Kit were purchased from Beyotime. DMEM, RPMI 1640, fetal bovine serum (FBS), trypsin-EDTA, and phosphate-buffered saline (PBS, pH 7.4 and pH 5.5) were supplied by Gibco Life Technologies.
Preparation and characterization HEA@Gel hydrogel
During the full oscillation period, a certain amount of GelMA and the photoinitiator LAP were dissolved in a certain amount of PBS and mixed uniformly. The mixture was then heated and dissolved at for 30 min . The solution was cooled to before HEA cells were added and maintained at . After of exposure to blue light, the above solution solidified rapidly. The morphology and internal structure of the hydrogels were characterized by cryo-scanning electron microscopy (FEI Quanta 450).
AST released from RHEA
Five grams of HEA (with cell walls) or HEA protoplast (without cell walls) was separately dispersed in 100 mL of deionized water. The solutions were placed in a room temperature environment and stirred slowly via magnetic stirring. One milliliter was extracted every hour and centrifuged at for 5 min to remove the microalgae, after which the supernatant was collected. The AST in the supernatant was extracted with DMSO, and the content of AST in the supernatant was determined by an AST standard curve. Additionally, to better observe and compare AST release, images of the extracted AST in DMSO solvent were also recorded.
The photothermal effect of HEA
Different concentrations of GHEA cell solution ( 1 mL ) were exposed to a 658 nm laser ( ) for 10 min . Using an infrared thermometer (TiS40 Infrared Camera FLUKE-TiS40 9 Hz , FLUKE, USA), the increase in temperature of the GHEA cell solution from photothermal conversion was detected every 30 s for 10 min .
Antibacterial performance
A plate count strategy was used to evaluate the antibacterial ability of the GHEA. First, 1 mL of E. coli or S. aureus ( ) was incubated with GHEA@Gel at for 12 h . The GHEA@Gel hydrogels were irradiated at 658 nm for 5 min for the laser groups. Then, 0.1 mL of the bacterial suspension was plated on agar and incubated for 24 h
at . Finally, the number of bacterial colonies was determined, and the antibacterial capacity was determined. To further demonstrate the antibacterial capability of the GHEA, SEM images of the bacterial morphologies under different treatments were examined.
Extracellular production
To investigate the oxygen-producing ability of the GHEA cells, a 15 mL final volume of GHEA cell in PBS solution with or GHEA cells was prepared. Before detection, argon gas was pumped into the solution for 10 min , after which the mixture was left in the dark for 1 h to stabilize the hypoxic conditions. Using an oxygen detector, the light ( or )-assisted GHEA-producing oxygen was measured every minute with stirring. In addition, the solution irradiated by a high-intensity laser ( ) was left at room temperature for 2 h to cool. The solution was then irradiated with a low-intensity laser ( ) for 20 min , after which oxygen production was monitored. To determine whether GHEA can produce oxygen during long-term wound healing, a 15 mL final volume of GHEA cell solution with GHEA cells was prepared. Then, the solution was irradiated by a high-intensity laser ( ) for 5 min . Every subsequent day, the solution was irradiated with a low-intensity laser ( ) for 30 min , and oxygen production was monitored by an oxygen detector.
ROS scavenging capacity
The ROS scavenging capacity of RHEA@Gel was evaluated using , , and as representative ROS . The scavenging capacity was determined by mixing 10 mL of with the hydrogels. 2 h later, of the supernatant was mixed for 30 min with of solution. The 405 nm absorbance was measured.
The ability to scavenge was evaluated by calculating the inhibition ratio of NBT photoreduction. Under a constant light intensity, NBT ( ), methionine ( 12.5 mM ), riboflavin ( ), and RHEA@Gel were combined for 10 min . The solution’s full wavenumber scanning curve was detected, and the solution’s absorbance at 560 nm was recorded.
Using an SA probe, the scavenging ability of RHEA@Gel was determined. Briefly, , and SA were added to PBS , and the mixture was incubated for 30 min at . The full wavenumber scanning curve of the solution was detected, and the absorbance of the solution at 510 nm was recorded.
Cell line sources
Human skin fibroblasts (HSF, catalog number: PCS-201-012), mouse monocyte macrophage leukemia cells (RAW264.7, catalog number: TIB-71), human umbilical vein endothelial cells (HUVEC, catalog number: PCS-100-013) and human immortalized keratinocytes (HaCaT, catalog number: PCS-200-011) were obtained from the American Type Culture Collections (ATCC).
Cell culture
HSFs, HaCaTs, and HUVECs were obtained from ATCC. Dulbecco’s modified Eagle’s medium (DMEM; Gibco) supplemented with 10% FBS was used to incubate the HSFs. RPMI 1640 medium (Gibco) supplemented with FBS was used to incubate HUVECs, HaCaTs, and RAW264.7.
The DCFH-DA probe was used to measure the ability of the samples to scavenge ROS. and the samples were added to a 6 -well plate containing cultured cells ( cells/well). The DACH-DA probe was added after 12 h , and the mixture was incubated for another 20 min . Then, fluorescence images were taken for the various groups.
Intracellular generation
The intracellular oxygen generation capacity of the hydrogels was determined using the probe. The cells were cultured in 6 -well plate ( cells/well) and covered with hydrogels, which were either treated with lasers or not. After was added to the cells, the cells were incubated for 20 min . Then, fluorescence images were taken for the various groups.
Cell toxicity and migration
HaCaTs were utilized to assess the cytotoxicity of the hydrogels via an MTT assay. The cells were seeded into the lower chamber of a 24 -well transwell plate (each well containing cells) for 12 h . Subsequently, the G-RHEA@Gel hydrogel was added to each upper chamber and incubated for another 12 h . Then, MTT solution was added, and the cells were incubated for 2 h . The absorbance at 490 nm was measured via a standardized process. Additionally, calcein-AM ( ) and PI staining were used to distinguish the distribution of living and dead cells. Cell apoptosis was analyzed via confocal laser scanning microscopy. For the cell migration experiment, cells were seeded in 24 -well plates and allowed to grow for 24 h before being scratched. A pipette was subsequently used to draw a uniformly wide line in the center of each well of the 24 -well plate to remove cells from that area. The cells were then placed in different environments and divided into four groups: the hypoxic environment group, the supply environment group, the hypoxic environment with coincubation with GHEA@Gel group, and the hypoxic environment with coincubation with GHEA@Gel and exposure to light group. Cell migration was recorded at 12 and 24 h thereafter.
Tube formation assay
To investigate tube formation, of thawed Matrigel (abs9493, Absin) was added to each lower chamber of a 24 -well plate. After incubation at for HUVECs were seeded into the Matrigel-coated lower chamber and exposed to various treatments. In the laser group, the hydrogel was exposed to 658 nm laser light for 10 min . HUVEC tube images were recorded by a microscope and analyzed using ImageJ software to evaluate tube formation.
Anti-inflammatory capacity
The anti-inflammatory effect of LPS + IFN- or IL-4 was evaluated in a macrophage model. The macrophage was seeded into the lower chamber of a 24 -well transwell plate (each well containing cells) for 12 h . During macrophage culture, LPS and IFN were added to the culture medium, after which the cells were cultured for 12 h . Subsequently, GHEA@Gel, RHEA@Gel, or G-RHEA@Gel was added to the culture medium, and the cells were allowed to grow
for 24 h . Macrophages were fixed with paraformaldehyde. Then, immunofluorescence staining and FCM analysis of CD206 and CD86 were performed. During this process, macrophages coincubated with IL-4 induced the M2 phenotype as a control.
Animal study
Female BALB/c mice ( 6 weeks, ) provided by Beijing HFK Bioscience Company were used in this study. All animals were housed in a specific pathogen-free (SPF) animal facility for 2 weeks for environmental adaptation and allowed free access to food and water. During the experiment, all animals were kept in the same standard environment ( humidity, 12 h light-dark cycle, and four mice/cage). All procedures, including animal care, wound modeling, dosing, and termination, were performed according to the Experimental Animal Guidelines for Ethical Review of Animal Welfare (GB/T 35892-2018) and approved by the Tianjin University Institute Animal Ethics Committee with the assigned approval number TJUE-2023-236. The mice were anesthetized with isoflurane anesthetic before any procedure that would cause pain. After the experiment, the mice were euthanized by inhalation followed by cervical dislocation.
Establishment of an infected diabetic wound model
To evaluate the therapeutic efficacy of HEA@Gel, a mouse model of infected diabetic wounds was created. Female BALB/c mice weighing were fasted for 12 h , followed by intraperitoneal injection of streptozotocin ( ), and the process was repeated three times within 3 days. The mice were kept under anesthesia during intraperitoneal injection. All mice were provided with sucrose water. Two weeks after the third injection, diabetic mice were identified as having blood glucose levels exceeding for two consecutive measurements within 2 days. The diabetic mice were anesthetized with isoflurane anesthetic, and their back fur was completely shaved. Then, 10 mm -diameter skin biopsy punches were used to create fullthickness wounds, and of S. aureus suspension ( was dropped on the surface of the wounds to create an infected wound model. The mice were kept under anesthesia during this procedure. After 1 day of infection, the number of bacterial colonies increased from to CFUs, indicating that the infection model was successfully established.
Infected diabetic wound healing
After the infected diabetic wound model was established, droplets of hydrogel solution, including hydrogel, GHEA@Gel, RHEA@Gel, or GRHEA@Gel, were added to the wound. The fluid from the hydrogel was used to quickly cover the entire wound area, after which the hydrogel was irradiated under an ultraviolet lamp for 10 s to achieve hydrogel curing. The mice were kept under anesthesia while the hydrogel was covered and photocuring. The wounds were treated with PBS as a control group. After that, the wounds with different hydrogel covers were exposed to a 658 nm laser with an intensity of for 5 min to achieve sterilization through photothermal conversion. The mice were kept under anesthesia during high-power laser ( ) irradiation. Every subsequent day, the mice were anesthetized with isoflurane anesthetic, and a 658 nm laser was used to illuminate the wound at a light intensity of for 30 min in the following 10 days. Low-intensity laser irradiation has two effects. First, GHEA activates photosynthesis to produce oxygen to improve the hypoxic wound microenvironment and accelerate vascular regeneration and wound healing. The second is to promote the release of AST from RHEA to reduce the levels of overexpressed ROS and inflammatory factors and regulate the immune microenvironment in wounds. As the wound healed, the edge of the hydrogel gradually separated from the healed wound tissue, and the hydrogel was slowly removed from the skin on the 12th day after the mice were anesthetized with isoflurane.
On days , and 12 , wound photographs were taken. On days 3 and 9, the mice were euthanized by inhalation followed by cervical dislocation, and the skin from the wound site was collected for HE and Masson staining. The hypoxia level, ROS level, vascularization, collagen deposition, epidermis formation, and macrophage polarization of the skin from the wound site were analyzed by immunofluorescence staining for HIF- , DHE, CD -SMA, collagen/vimentin, EGF, VEGF, and CD206/CD86 on day 6. To evaluate the biocompatibility and biosafety of HEA@Gel-based treatments, the major organs of the mice, including the heart, liver, spleen, lung and kidney, were collected for HE staining analysis on day 8 after HEA@Gel treatment. To evaluate the long-term antibacterial ability of HEA@Gel, the skin at the wound site was collected on day 20, and immunohistochemical staining was performed with an anti-Staphylococcus aureus antibody. In addition, to analyze the antibacterial effect visually, the skin was collected and homogenized. Serially diluted homogenates were plated on LB agar to quantify the bacterial colonies.
Long-term impact of the G-RHEA@Gel on the skin
The safety potential of a material is critical to its application. To explore the long-term impact of HEA@Gel on skin, the healthy skin of female BALB/c mice ( 6 weeks, ) was covered with the GRHEA@Gel hydrogel for 30 days. Then, the mice were euthanized by inhalation followed by cervical dislocation, and the skin covered with hydrogel was collected for H&E and Masson staining. In addition, the TNF- , IL-1 , IL-4, IL-6, IL-10, and CXCL-1 levels and WBC and Lymph and Gran counts were also detected.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The authors declare that all data supporting the findings of this study are available within the article and the Supplementary Information. All other data are available from the corresponding authors upon request. Source data are provided with this paper.
References
Chen, H. et al. Dissolved oxygen from microalgae-gel patch promotes chronic wound healing in diabetes. Sci. Adv. 6, eaba4311 (2020).
Hart, T., Milner, R. & Cifu, A. Management of a diabetic foot. JAMA 318, 1387-1388 (2017).
Botusan, I. R. et al. Stabilization of HIF-1 is critical to improve wound healing in diabetic mice. Proc. Natl Acad. Sci. 105, 19426-19431 (2008).
Thangarajah, H. et al. HIF-1 dysfunction in diabetes. Cell Cycle 9, 75-79 (2010).
Thangarajah, H. et al. The molecular basis for impaired hypoxiainduced VEGF expression in diabetic tissues. Proc. Natl Acad. Sci. 106, 13505-13510 (2009).
Zhu, Y., Wang, Y., Jia, Y., Xu, J. & Chai, Y. Roxadustat promotes angiogenesis through HIF-1 VEGF/VEGFR2 signaling and accelerates cutaneous wound healing in diabetic rats. Wound Repair Regen. 27, 324-334 (2019).
Zhang, F. et al. Infected wound repair with an ultrasoundenhanced nanozyme hydrogel scaffold. Mater. Horiz. 10, 5474 (2023).
Liu, S. et al. Absorbable thioether grafted hyaluronic acid nanofibrous hydrogel for synergistic modulation of inflammation microenvironment to accelerate chronic diabetic wound healing. Adv. Healthc. Mater. 9, 2000198 (2020).
Qian, Y. et al. Immunoregulation in diabetic wound repair with a photoenhanced glycyrrhizic acid hydrogel scaffold. Adv. Mater. 34, 2200521 (2022).
Krzyszczyk, P., Schloss, R., Palmer, A. & Berthiaume, F. The role of macrophages in acute and chronic wound healing and interventions to promote pro-wound healing phenotypes. Front. Phys. 9, 419 (2018).
Wang, T., Li, Y., Cornel, E. J., Li, C. & Du, J. Combined antioxidantantibiotic treatment for effectively healing infected diabetic wounds based on polymer vesicles. ACS Nano 15, 9027-9038 (2021).
Ouyang, J. et al. In situ sprayed NIR-responsive, analgesic black phosphorus-based gel for diabetic ulcer treatment. Proc. Natl Acad. Sci. 117, 28667-28677 (2020).
Talebian, S. et al. Self-healing hydrogels: the next paradigm shift in tissue engineering? Adv. Sci. 6, 1801664 (2019).
Liu, X., Inda, M. E., Lai, Y., Lu, T. K. & Zhao, X. Engineered living hydrogels. Adv. Mater. 34, e2201326 (2022).
Louf, J.-F., Lu, N. B., O’Connell, M. G., Cho, H. J. & Datta, S. S. Under pressure: hydrogel swelling in a granular medium. Sci. Adv. 7, eabd2711 (2021).
Xu, Y. et al. Robust and multifunctional natural polyphenolic composites for water remediation. Mater. Horiz. 9, 2496-2517 (2022).
Cao, H. et al. Versatile polyphenolic platforms in regulating cell biology. Chem. Soc. Rev. 51, 4175-4198 (2022).
Fang, Y. et al. Dissecting biological and synthetic soft-hard interfaces for tissue-like systems. Chem. Rev. 122, 5233-5276 (2022).
Hamedi, H., Moradi, S., Hudson, S. M., Tonelli, A. E. & King, M. W. Chitosan based bioadhesives for biomedical applications: a review. Carbohyd. Polym. 282, 119100 (2022).
Peng, W. et al. Recent progress of collagen, chitosan, alginate and other hydrogels in skin repair and wound dressing applications. Int. J. Biol. Macromol. 208, 400-408 (2022).
Xue, X. et al. Fabrication of physical and chemical crosslinked hydrogels for bone tissue engineering. Bioact. Mater. 12, 327-339 (2022).
Kang, Y. et al. The marriage of Xenes and hydrogels: fundamentals, applications, and outlook. Innovation 3, 100327 (2022).
Webber, M. J. & Tibbitt, M. W. Dynamic and reconfigurable materials from reversible network interactions. Nat. Rev. Mater. 7, 541-556 (2022).
Zeng, Q., Qi, X., Shi, G., Zhang, M. & Haick, H. Wound dressing: from nanomaterials to diagnostic dressings and healing evaluations. ACS Nano 16, 1708-1733 (2022).
Zhao, Y. et al. Supramolecular adhesive hydrogels for tissue engineering applications. Chem. Rev. 122, 5604-5640 (2022).
Dong, R. & Guo, B. Smart wound dressings for wound healing. Nano Today 41, 101290 (2021).
Liang, Y., He, J. & Guo, B. Functional hydrogels as wound dressing to enhance wound healing. ACS Nano 15, 12687-12722 (2021).
Zhang, S. et al. Recent advances in responsive hydrogels for diabetic wound healing. Mater. Today Bio 18, 100508-100508 (2023).
Cheng, S. et al. Dendritic hydrogels with robust inherent antibacterial properties for promoting bacteria-infected wound healing. ACS Appl. Mater. Interfaces 14, 11144-11155 (2022).
Hao, Y., Zhao, W., Zhang, H., Zheng, W. & Zhou, Q. Carboxymethyl chitosan-based hydrogels containing fibroblast growth factors for triggering diabetic wound healing. Carbohyd. Polym. 287, 119336 (2022).
Liang, Y. et al. Glucose dual responsive metformin release hydrogel dressings with adhesion and self-healing via dualdynamic bonding for athletic diabetic foot wound healing. ACS Nano 16, 3194-3207 (2022).
Sabbagh, F., Muhamad, I. I., Niazmand, R., Dikshit, P. K. & Kim, B. S. Recent progress in polymeric non-invasive insulin delivery. Int. J. Biol. Macromol. 203, 222-243 (2022).
Yang, J. et al. Glucose-responsive multifunctional metal-organic drug-loaded hydrogel for diabetic wound healing. Acta Biomater. 140, 206-218 (2022).
Zhao, Y. et al. Biomimetic nanozyme-decorated hydrogels with H2O2-activated oxygenation for modulating immune microenvironment in diabetic wound. ACS Nano 17, 16854-16869 (2023).
Wu, Y. et al. A versatile glycopeptide hydrogel promotes chronic refractory wound healing through bacterial elimination, sustained oxygenation, immunoregulation, and neovascularization. Adv. Funct. Mater. 33, 2305992 (2023).
Zhang, J. et al. An injectable bioactive dressing based on plateletrich plasma and nanoclay: sustained release of deferoxamine to accelerate chronic wound healing. Acta Pharm. Sin. B 13, 4318-4336 (2023).
Liu, B. et al. Hydrogen bonds autonomously powered gelatin methacrylate hydrogels with super-elasticity, self-heal and underwater self-adhesion for sutureless skin and stomach surgery and E-skin. Biomaterials 171, 83-96 (2018).
Kurian, A. G., Singh, R. K., Patel, K. D., Lee, J.-H. & Kim, H.-W. Multifunctional GelMA platforms with nanomaterials for advanced tissue therapeutics. Bioact. Mater. 8, 267-295 (2022).
Koo, H., Allan, R. N., Howlin, R. P., Stoodley, P. & Hall-Stoodley, L. Targeting microbial biofilms: current and prospective therapeutic strategies. Nat. Rev. Microbiol. 15, 740-755 (2017).
Coban, O., De Deyn, G. B. & van der Ploeg, M. Soil microbiota as game-changers in restoration of degraded lands. Science 375, 990 (2022).
Jansson, J. K. & Hofmockel, K. S. Soil microbiomes and climate change. Nat. Rev. Microbiol. 18, 35-46 (2020).
van Oppen, M. J. H. & Blackall, L. L. Coral microbiome dynamics, functions and design in a changing world. Nat. Rev. Microbiol. 17, 557-567 (2019).
Zhang, H. et al. Bacteria photosensitized by intracellular gold nanoclusters for solar fuel production. Nat. Nanotechnol. 13, 900-905 (2018).
Huo, M. et al. Photosynthetic tumor oxygenation by photosensitizer-containing cyanobacteria for enhanced photodynamic therapy. Angew. Chem. Int. Ed. 59, 1906-1913 (2020).
Huang, J. et al. Bio-inspired synthesis of metal nanomaterials and applications. Chem. Soc. Rev. 44, 6330-6374 (2015).
Logan, B. E., Rossi, R., Ragab, A. A. & Saikaly, P. E. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 17, 307-319 (2019).
Singh, P., Kim, Y.-J., Zhang, D. & Yang, D.-C. Biological synthesis of nanoparticles from plants and microorganisms. Trends Biotechnol. 34, 588-599 (2016).
Yang, H. et al. Preparation of nickel-iron hydroxides by microorganism corrosion for efficient oxygen evolution. Nat. Commun. 11, 5075 (2020).
Chen, J. et al. Oxygen-self-produced nanoplatform for relieving hypoxia and breaking resistance to sonodynamic treatment of pancreatic cancer. ACS Nano 11, 12849-12862 (2017).
Li, X., Kwon, N., Guo, T., Liu, Z. & Yoon, J. Innovative strategies for hypoxic-tumor photodynamic therapy. Angew. Chem. Int. Ed. 57, 11522-11531 (2018).
Saravanakumar, G., Kim, J. & Kim, W. J. Reactive-oxygen-speciesresponsive drug delivery systems: promises and challenges. Adv. Sci. 4, 1600124 (2017).
Xuan, M. et al. Magnetic mesoporous silica nanoparticles cloaked by red blood cell membranes: applications in cancer therapy. Angew. Chem. Int. Ed. 57, 6049-6053 (2018).
Zhang, X. et al. Black phosphorus-loaded separable microneedles as responsive oxygen delivery carriers for wound healing. ACS Nano 14, 5901-5908 (2020).
Zhao, L.-P. et al. Self-delivery nanomedicine for O-2-economized photodynamic tumor therapy. Nano Lett. 20, 2062-2071 (2020).
Hu, H. et al. Microalgae-based bioactive hydrogel loaded with quorum sensing inhibitor promotes infected wound healing. Nano Today 42, 101368 (2022).
Ou, M. et al. Heterojunction engineered bioactive chlorella for cascade promoted cancer therapy. J. Control. Release 345, 755-769 (2022).
Khoo, K. S. et al. Recent advances in biorefinery of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 288, 121606 (2019).
Yang, L. et al. Influence of molecular structure of astaxanthin esters on their stability and bioavailability. Food Chem. 343, 128497(2021).
Zhao, X. et al. Physical and oxidative stability of astaxanthin microcapsules prepared with liposomes. J. Sci. Food Agric. 102, 4909-4917 (2022).
Gwak, Y. et al. Comparative analyses of lipidomes and transcriptomes reveal a concerted action of multiple defensive systems against photooxidative stress in Haematococcus Pluvialis. J. Exp. Bot. 65, 4317-4334 (2014).
Hu, C. et al. Transcriptomic analysis unveils survival strategies of autotrophic Haematococcus pluvialis against high light stress. Aquaculture 513, 734430 (2019).
Zhang, X. et al. ROS-triggered self-disintegrating and pHresponsive astaxanthin nanoparticles for regulating the intestinal barrier and colitis. Biomaterials 292, 121937 (2023).
Wen, X. et al. Astaxanthin acts via LRP-1 to inhibit inflammation and reverse lipopolysaccharide-induced M1/M2 polarization of microglial cells. Oncotarget 8, 69370-69385 (2017).
Ambati, R. R., Phang, S. M., Ravi, S. & Aswathanarayana, R. G. Astaxanthin: sources, extraction, stability, biological activities and its commercial applications-a review. Mar. Drugs 12, 128-152 (2014).
Davinelli, S., Nielsen, M. E. & Scapagnini, G. Astaxanthin in skin health, repair, and disease: a comprehensive review. Nutrients 10, 522 (2018).
Lim, K. C., Yusoff, F. M., Shariff, M. & Kamarudin, M. S. Astaxanthin as feed supplement in aquatic animals. Rev. Aquac. 10, 738-773 (2018).
Oslan, S. N. H. et al. A review on Haematococcus pluvialis bioprocess optimization of green and red stage culture conditions for the production of natural astaxanthin. Biomolecules 11, 256 (2021).
Perozeni, F. et al. Turning a green alga red: engineering astaxanthin biosynthesis by intragenic pseudogene revival in Chlamydomonas reinhardtii. Plant Biotechnol. J. 18, 2053-2067 (2020).
Rammuni, M. N., Ariyadasa, T. U., Nimarshana, P. H. V. & Attalage, R. A. Comparative assessment on the extraction of carotenoids from microalgal sources: Astaxanthin from H-pluvialis and beta-carotene from D. salina. Food Chem. 277, 128-134 (2019).
Shah, M. M. R., Liang, Y., Cheng, J. J. & Daroch, M. Astaxanthinproducing green microalga Haematococcus pluvialis: from single Cellto high value commercia products. Front. Plant Sci. 7, 531 (2016).
Huang, W.-C., Liu, H., Sun, W., Xue, C. & Mao, X. Effective astaxanthin extraction from wet Haematococcus pluvialis using switchable hydrophilicity solvents. ACS Sustain. Chem. Eng. 6, 1560-1563 (2018).
Yang, H. E., Yu, B. S. & Sim, S. J. Enhanced astaxanthin production of Haematococcus pluvialis strains induced salt and high light resistance with gamma irradiation. Bioresour. Technol. 372, 128651 (2023).
Xu, R., Zhang, L., Yu, W. & Liu, J. A strategy for interfering with the formation of thick cell walls in Haematococcus pluvialis by downregulating the mannan synthesis pathway. Bioresour. Technol. 362, 127783 (2022).
Fulbright, S. P. et al. Bacterial community changes in an industrial algae production system. Algal Res. 31, 147-156 (2018).
Mickalide, H. & Kuehn, S. Higher-order interaction between species inhibits bacterial invasion of a phototroph-predator microbial community. Cell Syst. 9, 521-533 (2019).
Acknowledgements
This study was financially supported by a grant from the National Natural Science Foundation of China (Grant No. 32071322, X.J.), National Natural Science Funds for Excellent Young Scholar (Grant No. 32122044, X.J.), China Postdoctoral Science Foundation (Grant No. 2023T160479, 2023M742603, Y.K.), and Technology & Innovation Commission of Shenzhen Municipality (Grant No.
JCYJ20210324113004010, X.J.).
Author contributions
X. Ji, B. Liu, and J. Xie designed and supervised the project. X. Ji, Y. Kang, L. Xu, and J. Dong designed the experimental strategies. Y. Kang, L. Xu, J. Dong, X. Yuan, J. Ye, and Y. Fan performed the experiments and analyzed the data. X. Ji, B. Liu, and J. Xie wrote the manuscript.
Correspondence and requests for materials should be addressed to Bing Liu, Julin Xie or Xiaoyuan Ji.
Peer review information Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Academy of Medical Engineering and Translational Medicine, Medical College, Tianjin University, Tianjin 300072, China. Department of Disease Control and Prevention, Rocket Force Characteristic Medical Center, Beijing 10088, China. Department of Burns, The First Affiliated Hospital of Sun Yat-Sen University, Guangzhou 510080, China. Medical College, Linyi University, Linyi 276000, China. e-mail: neaucn@126.com; xiejulin@mail.sysu.edu.cn; jixiaoyuan@tju.edu.cn