DOI: https://doi.org/10.1038/s41561-024-01384-7
تاريخ النشر: 2024-02-20
الحساسية الناشئة لدرجة حرارة الكربون العضوي في التربة المدفوعة بالارتباطات المعدنية
تاريخ القبول: 23 يناير 2024
تاريخ النشر على الإنترنت: 20 فبراير 2024
(D) تحقق من التحديثات
الملخص
تعتمد تحلل المادة العضوية في التربة وتفاعلاتها مع المناخ على ما إذا كانت المادة العضوية مرتبطة بالمعادن التربة. ومع ذلك، فإن قيود البيانات قد أعاقت التحليلات على نطاق عالمي لبرك الكربون العضوي المرتبط بالمعادن والجزيئات ومرجعيتها في نماذج النظام الأرضي المستخدمة لتقدير ردود الفعل بين دورة الكربون والمناخ. هنا نقوم بتحليل التقديرات العالمية المستمدة من الملاحظات لبرك الكربون في التربة لتحديد نسبها النسبية وحساب حساسياتها الحرارية المناخية كتناقص في الكربون مع زيادة درجة الحرارة. نجد أن الحساسية الحرارية المناخية للكربون الجزيئي أعلى في المتوسط
أثر درجات الحرارة على مخزونات الكربون في التربة الكلية ومعدلات الدوران على نطاقات النظام البيئي إلى النطاق العالمي
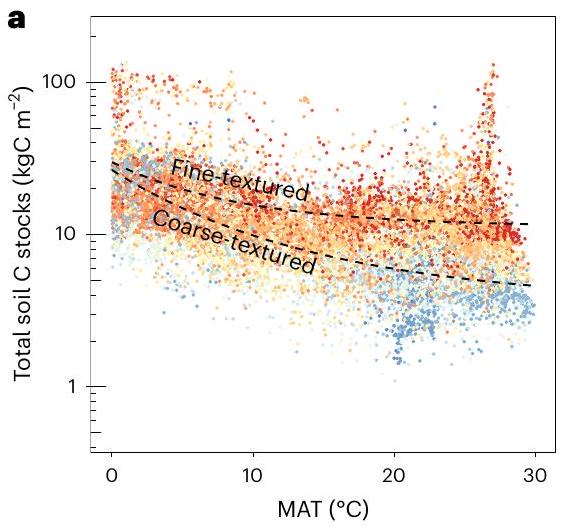
والكربون الجزيئي في التربة (المستمد من تجميع ملاحظات أجزاء التربة
الحساسية الحرارية المناخية لبرك الكربون في التربة
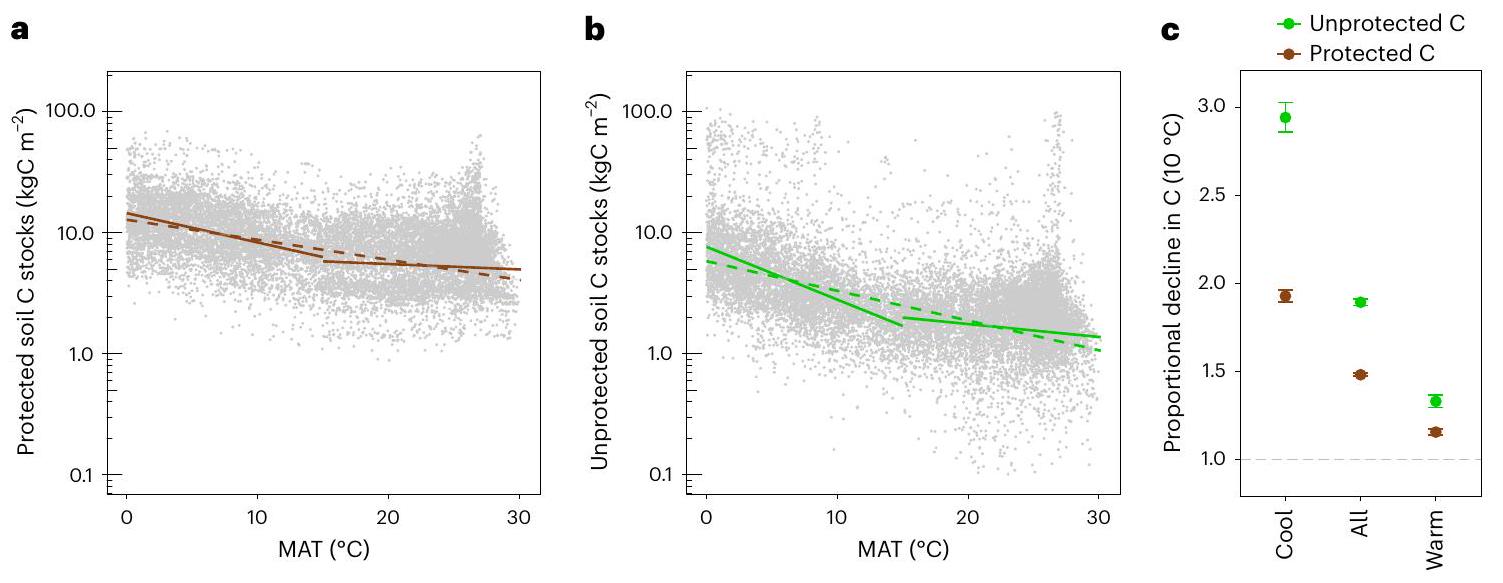
محاسبة المتغيرات المربكة المحتملة (انظر ‘تحليل البيانات’ في الطرق). النقاط تمثل الانخفاض النسبي لكل مجموعة من مجموعات الكربون في التربة (المستمدة من انحدارات الانحدار؛
التربة التي تحتوي على مخزونات كربون غير محمية أعلى تؤدي إلى حساسية أكبر لدرجة الحرارة. تظهر حساسية درجة الحرارة المناخية لكربون التربة الكلي من توزيعات المخزونات الأساسية واعتمادياتها على درجة الحرارة.
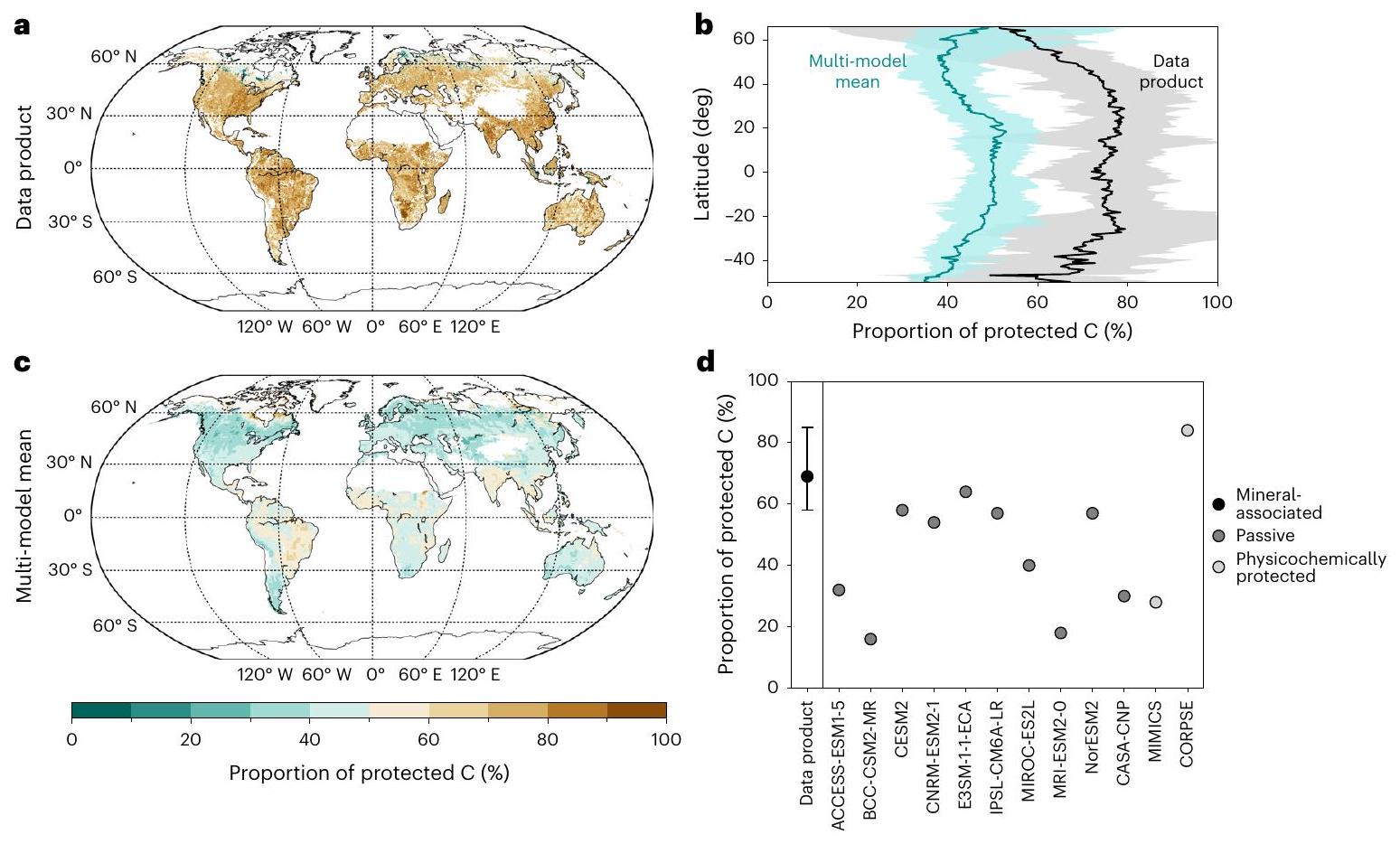
أنماط عالمية في خزانات الكربون في التربة عبر البيانات والنماذج
التحكمات المعدنية، أوقات التدوير والستيوكيومترية للكربون المحمي الذي يُستخدم لتقريب تشكيل وخصائص المادة العضوية المرتبطة بالمعادن (انظر ‘قابلية تفسير نموذج الحوض’ في الطرق). وبالتالي، نتبنى هذا التقليد هنا، مع الاعتراف بأن هناك لا تزال عدم يقين جوهري في قابلية تفسير الأحواض المودلّة والكسور المعرفة عمليًا. نفترض أن المعايير المستندة إلى توزيعات الأحواض النسبية، جنبًا إلى جنب مع أعمار الكربون المشع
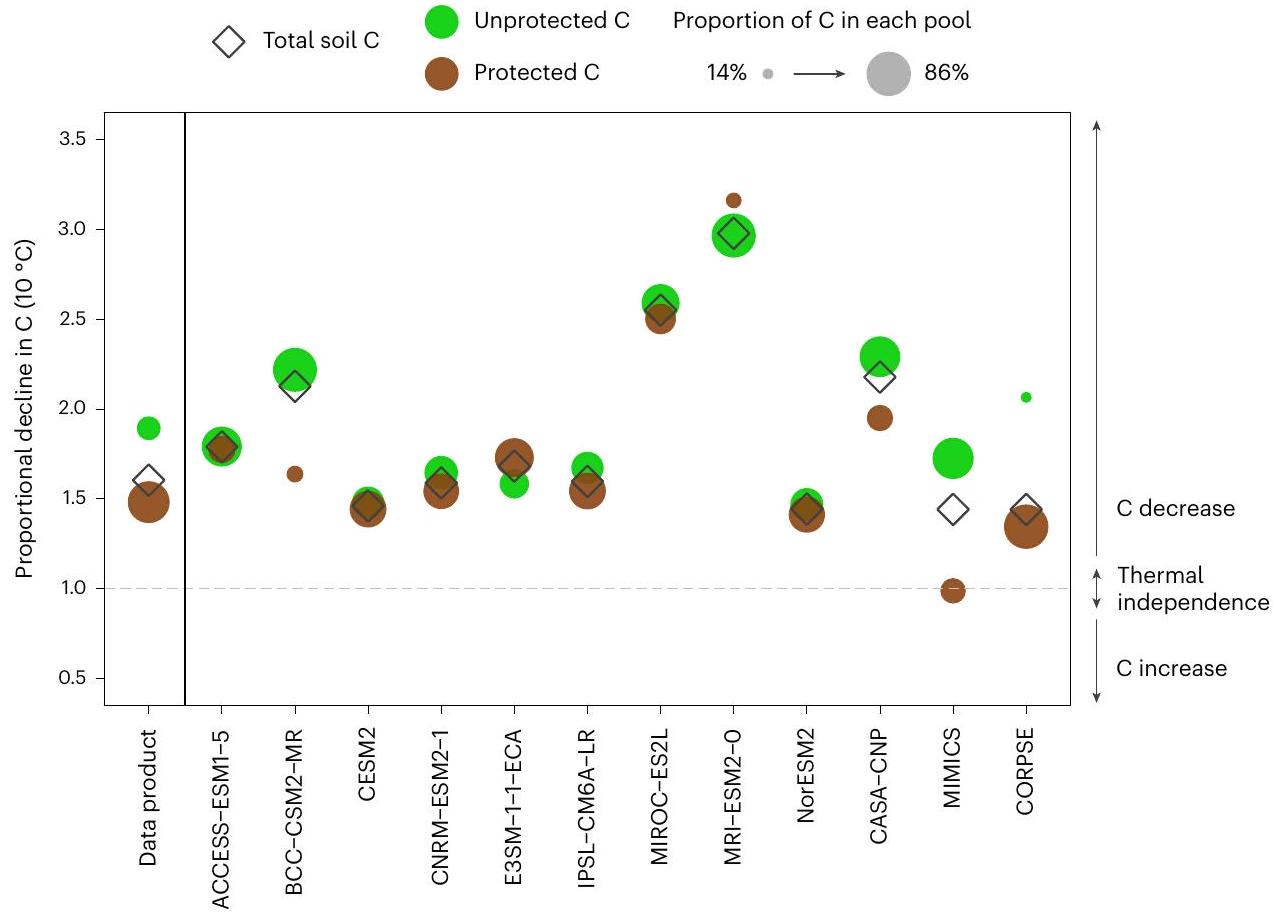
حساسية الكربون في التربة لدرجة الحرارة في النماذج العالمية
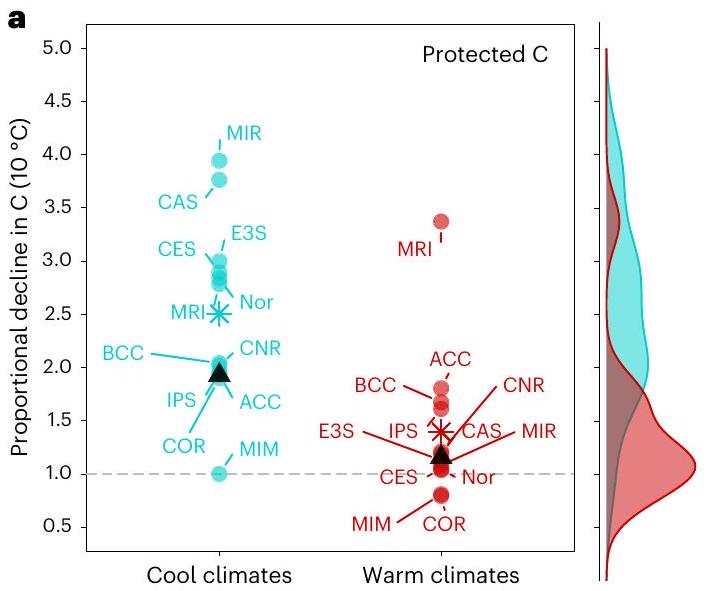
قد تبدو بعض النماذج قادرة على التقاط حساسية درجة الحرارة المناخية المستمدة من الملاحظات لكتل الكربون في التربة، لكن هذا الاتفاق لا يحدث دائمًا للأسباب الصحيحة وقد ينهار عند النظر في مساهمة خزانات الكربون في التربة المحمية وغير المحمية. على سبيل المثال، ستة من نماذج CMIP6 ESMs (ACCESS-ESM1-5، CESM2، CNRM-ESM2-1، E3SM-1-1-ECA، IPSL-CM6A-LR وNorESM2) ونموذجان محددان ميكروبيًا (MIMICS وCORPSE) تلتقط تقريبًا حساسية درجة الحرارة لكتل الكربون في التربة (الشكل 4 والجدول التكميلي 6). ومع ذلك، تظهر هذه النماذج نفسها توزيعات متباينة وحساسيات درجة حرارة مناخية لخزانات الكربون في التربة المحمية وغير المحمية (الشكل 4). فقط عدد قليل من النماذج تلتقط تقريبًا حساسية درجة الحرارة المناخية للكربون المحمي (CESM2، CNRM-ESM2-1، IPSL-CM6A-LR، NorESM2 وCORPSE) وأقل منها للكربون غير المحمي (ACCESS-ESM1-5، MIMICS وCORPSE).
للعوامل المحتملة المربكة (انظر ‘تحليل البيانات’ في الطرق). تظهر النتائج لمنتج البيانات ومجموعة من نماذج ESM ونماذج الأراضي غير المتصلة عبر خلايا الشبكة مع MAT
لـ C المحمية في نماذج مختارة
كلا المناخين البارد والدافئ، وكلا برك الكربون في المناخات الدافئة (الشكل 5). هذا صحيح بشكل خاص بالنسبة لـ MIROC-ES2L و CASA-CNP و MRI-ESM2-0، التي تفرط في تقدير الحساسية المناخية للمناطق المحمية.
الآثار ووجهات النظر المستقبلية
| منتج بيانات | ACC
|
E3S
|
ولا
|
| * متوسط النماذج المتعددة | نسخة مخفية
|
نظام تحديد المواقع الداخلي
|
كاس
|
| * مخرجات النموذج | CES
|
مير
|
ميم
|
| سي إن آر
|
الرنين المغناطيسي
|
كور
|
دوائر؛ مُعلمة بالاختصارات)؛ تم تقديم فترات الثقة 95% في الجدول التكميلية 6. تُظهر تقديرات كثافة النواة لكل لوحة، موضحة توزيع حساسية درجات الحرارة المناخية عبر مجموعة النماذج لكل مجموعة وبيئة حرارية. القيم الأعلى (
تلك البرك غير المحمية مع الاحترار، هي تقدير غير دقيق لدورة الكربون العالمية – ردود الفعل المناخية وتوقعات تراكم الكربون المدفوعة بالإنتاجية العالية في هذه المناطق. بينما قد يتم تعويض هذا التحيز جزئيًا في نماذج النظام البيئي التي تفرط في تقدير حساسية درجة الحرارة المناخية للكربون المحمي عبر المناخات، فإن تأثير هذا التقدير المفرط على توقعات ردود الفعل لدورة الكربون والمناخ ربما يكون أضعف لأن هذه البرك تتسم بطبيعتها ببطء الدورة في نماذج التربة من الدرجة الأولى وبالتالي تتغير قليلاً جدًا على فترات زمنية ذات صلة بسياسة المناخ (الأشكال التكميلية 6-8). يمكن أن تؤدي الاستجابات الخاصة بالبرك المتباينة في النهاية إلى اختلافات في تركيبة ووظيفة الكربون في التربة. لذلك، من الضروري أن تقوم النماذج بتقييم ليس فقط حساسية المناخ الكلية ولكن أيضًا تلك الخاصة بالبرك الأساسية للكربون في التربة، من أجل التنبؤ بثقة بديناميات الكربون في التربة في المستقبل.
المحتوى عبر الإنترنت
References
- Friedlingstein, P. et al. Uncertainties in CMIP5 climate projections due to carbon cycle feedbacks. J. Clim. 27, 511-526 (2014).
- Davidson, E. A. & Janssens, I. A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440, 165-173 (2006).
- García-Palacios, P. et al. Evidence for large microbial-mediated losses of soil carbon under anthropogenic warming. Nat. Rev. Earth Environ. 2, 507-517 (2021).
- Koven, C. D., Hugelius, G., Lawrence, D. M. & Wieder, W. R. Higher climatological temperature sensitivity of soil carbon in cold than warm climates. Nat. Clim. Chang. 7, 817-822 (2017).
- Crowther, T. et al. Quantifying global soil C losses in response to warming. Nature 104, 104-108 (2016).
- Todd-Brown, K., Zheng, B. & Crowther, T. W. Field-warmed soil carbon changes imply high 21st-century modeling uncertainty. Biogeosciences 15, 3659-3671 (2018).
- Van Gestel, N. et al. Predicting soil carbon loss with warming. Nature 554, E4-E5 (2018).
- Hicks Pries, C. E., Castanha, C., Porras, R. & Torn, M. S. The whole-soil carbon flux in response to warming. Science 355, 1420-1423 (2017).
- Rocci, K. S., Lavallee, J. M., Stewart, C. E. & Cotrufo, M. F. Soil organic carbon response to global environmental change depends on its distribution between mineral-associated and particulate organic matter: a meta-analysis. Sci. Total Environ. 793, 148569 (2021).
- Conant, R. T. et al. Temperature and soil organic matter decomposition rates – synthesis of current knowledge and a way forward. Glob. Chang. Biol. 17, 3392-3404 (2011).
- Soong, J. L. et al. Five years of whole-soil warming led to loss of subsoil carbon stocks and increased
efflux. Sci. Adv. 7, eabd1243 (2021). - Nottingham, A. T., Meir, P., Velasquez, E. & Turner, B. L. Soil carbon loss by experimental warming in a tropical forest. Nature 584, 234-237 (2020).
- Pellegrini, A. F. A. et al. Low-intensity frequent fires in coniferous forests transform soil organic matter in ways that may offset ecosystem carbon losses. Glob. Chang. Biol. 27, 3810-3823 (2021).
- Lugato, E., Lavallee, J. M., Haddix, M. L., Panagos, P. & Cotrufo, M. F. Different climate sensitivity of particulate and mineral-associated soil organic matter. Nat. Geosci. 14, 295-300 (2021).
- Sokol, N. W. et al. Global distribution, formation and fate of mineral-associated soil organic matter under a changing climate: a trait-based perspective. Funct. Ecol. 36, 1411-1429 (2022).
- Lavallee, J. M., Soong, J. L. & Cotrufo, M. F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Chang. Biol. 26, 261-273 (2020).
- Kögel-Knabner, I. et al. Organo-mineral associations in temperate soils: integrating biology, mineralogy, and organic matter chemistry. J. Plant Nutr. Soil Sci. 171, 61-82 (2008).
- Heckman, K. et al. Beyond bulk: density fractions explain heterogeneity in global soil carbon abundance and persistence. Glob. Chang. Biol. 28, 1178-1196 (2022).
- Wieder, W. R., Allison, S. D., Davidson, E. A., Georgiou, K. & Hararuk, O. Explicitly representing soil microbial processes in Earth system models. Glob. Biogeochem. Cycles 29, 1782-1800 (2015).
- Sulman, B. N., Phillips, R. P., Oishi, A. C., Shevliakova, E. & Pacala, S. W. Microbe-driven turnover offsets mineral-mediated storage of soil carbon under elevated
. Nat. Clim. Chang. 4, 1099-1102 (2014). - Ahrens, B., Braakhekke, M. C., Guggenberger, G., Schrumpf, M. & Reichstein, M. Contribution of sorption, DOC transport and microbial interactions to the
age of a soil organic carbon profile: Insights from a calibrated process model. Soil Biol. Biochem. 88, 390-402 (2015). - Sulman, B. N. et al. Multiple models and experiments underscore large uncertainty in soil carbon dynamics. Biogeochemistry 141, 109-123 (2018).
- Wieder, W. R., Sulman, B. N., Hartman, M. D., Koven, C. D. & Bradford, M. A. Arctic soil governs whether climate change drives global losses or gains in soil carbon. Geophys. Res. Lett. 46, 14486-14495 (2019).
- Berardi, D. et al. 21st-century biogeochemical modeling: challenges for Century-based models and where do we go from here? Glob. Chang. Biol. Bioenergy 12, 774-788 (2020).
- Kleber, M. et al. Dynamic interactions at the mineral-organic matter interface. Nat. Rev. Earth Environ. 2, 402-421 (2021).
- Melillo, J. M. et al. Long-term pattern and magnitude of soil carbon feedback to the climate system in a warming world. Science 358, 101-105 (2017).
- Walker, T. W. N. et al. A systemic overreaction to years versus decades of warming in a subarctic grassland ecosystem. Nat. Ecol. Evol. 4, 101-108 (2020).
- Bradford, M. Thermal adaptation of decomposer communities in warming soils. Front. Microbiol. 4, 333 (2013).
- Hartley, I. P., Hill, T. C., Chadburn, S. E. & Hugelius, G. Temperature effects on carbon storage are controlled by soil stabilisation capacities. Nat. Commun. 12, 6713 (2021).
- Doetterl, S. et al. Soil carbon storage controlled by interactions between geochemistry and climate. Nat. Geosci. 8, 780-783 (2015).
- Abramoff, R. Z., Torn, M. S., Georgiou, K., Tang, J. & Riley, W. J. Soil organic matter temperature sensitivity cannot be directly inferred from spatial gradients. Glob. Biogeochem. Cycles 33, 761-776 (2019).
- Luo, Z. et al. Convergent modelling of past soil organic carbon stocks but divergent projections. Biogeosciences 12, 4373-4383 (2015).
- Georgiou, K. et al. Global stocks and capacity of mineral-associated soil organic carbon. Nat. Commun. 13, 3797 (2022).
- Eyring, V. et al. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 9, 1937-1958 (2016).
- Jobbágy, E. G. & Jackson, R. B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 10, 423-436 (2000).
- Ahrens, B. et al. Combination of energy limitation and sorption capacity explains
depth gradients. Soil Biol. Biochem. 148, 107912 (2020). - Gentsch, N. et al. Temperature response of permafrost soil carbon is attenuated by mineral protection. Glob. Chang. Biol. 24, 3401-3415 (2018).
- Giardina, C. P. & Ryan, M. G. Evidence that decomposition rates of organic carbon inmineral soil do not vary with temperature. Nature 404, 858-861 (2000).
- Giardina, C. P., Litton, C. M., Crow, S. E. & Asner, G. P. Warming-related increases in soil
efflux are explained by increased below-ground carbon flux. Nat. Clim. Chang. 4, 822-827 (2014). - Wang, C. et al. The temperature sensitivity of soil: microbial biodiversity, growth, and carbon mineralization. ISME J. 15, 2738-2747 (2021).
- Shi, Z. et al. The age distribution of global soil carbon inferred from radiocarbon measurements. Nat. Geosci. 13, 555-559 (2020).
- He, Y. et al. Radiocarbon constraints imply reduced carbon uptake by soils during the 21st century. Science 353, 1419-1424 (2016).
- Todd-Brown, K. E. O. et al. Causes of variation in soil carbon simulations from CMIP5 Earth system models and comparison with observations. Biogeosciences 10, 1717-1736 (2013).
- Ito, A. et al. Soil carbon sequestration simulated in CMIP6-LUMIP models: implications for climatic mitigation. Environ. Res. Lett. 15, 124061 (2019).
- Abramoff, R. Z. et al. Improved global-scale predictions of soil carbon stocks with Millennial Version 2. Soil Biol. Biochem. 164, 108466 (2022).
- Zimmermann, M., Leifeld, J., Schmidt, M. W. I., Smith, P. & Fuhrer, J. Measured soil organic matter fractions can be related to pools in the RothC model. Eur. J. Soil Sci. 58, 658-667 (2007).
- Pierson, D. et al. Optimizing process-based models to predict current and future soil organic carbon stocks at high-resolution. Sci. Rep. 12, 10824 (2022).
- Koven, C. et al. On the formation of high-latitude soil carbon stocks: effects of cryoturbation and insulation by organic matter in a land surface model. Geophys. Res. Lett. https://doi. org/10.1029/2009GL040150 (2009).
- Koven, C. D. et al. The effect of vertically resolved soil biogeochemistry and alternate soil C and N models on C dynamics of CLM4. Biogeosciences 10, 7109-7131 (2013).
- Plaza, C. et al. Direct observation of permafrost degradation and rapid soil carbon loss in tundra. Nat. Geosci. 12, 627-631 (2019).
- Parton, W. J., Schimel, D. S., Cole, C. V. & Ojima, D. S. Analysis of factors controlling soil organic matter levels in Great Plains grasslands. Soil Sci. Soc. Am. J. 51, 1173-1179 (1987).
- Haaf, D., Six, J. & Doetterl, S. Global patterns of geo-ecological controls on the response of soil respiration to warming. Nat. Clim. Chang. 11, 623-627 (2021).
- Jones, C. D. et al. C4MIP – the Coupled Climate-Carbon Cycle Model Intercomparison Project: experimental protocol for CMIP6. Geosci. Model Dev. 9, 2853-2880 (2016).
- Bouskill, N. J., Riley, W. J., Zhu, Q., Mekonnen, Z. A. & Grant, R. F. Alaskan carbon-climate feedbacks will be weaker than inferred from short-term experiments. Nat. Commun. 11, 5798 (2020).
adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons. org/licenses/by/4.0/.
(c) The Author(s) 2024
طرق
مصادر البيانات والمعالجة
ناتج نموذج الأرض العالمي
قابلية تفسير خزانات النموذج
حوض رينولدز كريك التجريبي
تحليل البيانات
التحليلات على توليف ملاحظي
توفر البيانات
توفر الكود
References
- Harris, I., Jones, P. D., Osborn, T. J. & Lister, D. H. Updated high-resolution grids of monthly climatic observations – the CRU TS3.10 Dataset. Int. J. Climatol. 34, 623-642 (2014).
- Schneider, U. et al. GPCC Full Data Reanalysis Version 6.0 at
: Monthly Land-Surface Precipitation from Rain-Gauges built on GTS-based and Historic Data (GPCC, 2011); https://doi. org/10.5676/DWD_GPCC/FD_M_V6_050 - Friedl, M. A. et al. MODIS Collection 5 global land cover: algorithm refinements and characterization of new datasets. Remote Sens. Environ. 114, 168-182 (2010).
- Zhao, M., Heinsch, F. A., Nemani, R. R. & Running, S. W. Improvements of the MODIS terrestrial gross and net primary production global data set. Remote Sens. Environ. 95, 164-176 (2005).
- Wieder, W. R., Boehnert, J., Bonan, G. B. & Langseth, M. Regridded Harmonized World Soil Database v1.2 (ORNL DAAC, 2014); https:// doi.org/10.3334/ORNLDAAC/1247
- Hengl, T. et al. SoilGrids250m: global gridded soil information based on machine learning. PLoS ONE https://doi.org/10.1371/ journal.pone. 0169748 (2017).
- Georgiou, K. et al. Globally-gridded data for manuscript: global stocks and capacity of mineral-associated soil organic carbon. Zenodo https://doi.org/10.5281/zenodo. 6539765 (2022)
- Ploton, P. et al. Spatial validation reveals poor predictive performance of large-scale ecological mapping models. Nat. Commun. 11, 4540 (2020).
- Meyer, H. & Pebesma, E. Machine learning-based global maps of ecological variables and the challenge of assessing them. Nat. Commun. 13, 2208 (2022).
- Thornton, P. E. & Rosenbloom, N. A. Ecosystem model spin-up: estimating steady state conditions in a coupled terrestrial carbon and nitrogen cycle model. Ecol. Modell. 189, 25-48 (2005).
- Lawrence, D. M. et al. Parameterization improvements and functional and structural advances in Version 4 of the Community Land Model. J. Adv. Model. Earth Syst. https://doi. org/10.1029/2011MS00045 (2011).
- Lee, W. L. et al. Taiwan Earth System Model Version 1: description and evaluation of mean state. Geosci. Model Dev. 13, 3887-3904 (2020).
- Christensen, B. T. in Evaluation of Soil Organic Matter Models Vol. 38 (eds Powlson, D. S. et al.) 143-159 (Springer, 1996).
- Elliot, E. T., Paustian, K. & Frey, S. D. in Evaluation of Soil Organic Matter Models Vol. 38 (eds Powlson, D. S. et al.) 161-179 (Springer, 1996).
- Lehmann, J. & Kleber, M. The contentious nature of soil organic matter. Nature 528, 60-68 (2015).
- Zhang, Y. et al. Simulating measurable ecosystem carbon and nitrogen dynamics with the mechanistically defined MEMS 2.0 model. Biogeosciences 18, 3147-3171 (2021).
- Dangal, S. R. S. et al. Improving soil carbon estimates by linking conceptual pools against measurable carbon fractions in the DAYCENT model version 4.5. J. Adv. Model. Earth Syst. 14, e2021MSOO2622 (2022).
- Abramoff, R. et al. The Millennial model: in search of measurable pools and transformations for modeling soil carbon in the new century. Biogeochemistry 137, 51-71 (2018).
- Parton, W. J. & Rasmussen, P. E. Long-term effects of crop management in wheat-fallow: II. CENTURY model simulations. Soil Sci. Soc. Am. J. 58, 530-536 (1994).
- Cotrufo, M. F., Ranalli, M. G., Haddix, M. L., Six, J. & Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 12, 989-994 (2019).
- Rocci, K. S. et al. Aligning theoretical and empirical representations of soil carbon-to-nitrogen stoichiometry with process-based terrestrial biogeochemistry models. Soil Biol. Biochem. 189, 109272 (2024).
- Lawrence, C. R. et al. An open-source database for the synthesis of soil radiocarbon data: International Soil Radiocarbon Database (ISRaD) version 1.0. Earth Syst. Sci. Data 12, 61-76 (2020).
- Kyker-Snowman, E., Wieder, W. R., Frey, S. D. & Grandy, A. S. Stoichiometrically coupled carbon and nitrogen cycling in the MIcrobial-MIneral Carbon Stabilization model version 1.0 (MIMICS-CN v1.0). Geosci. Model Dev. 13, 4413-4434 (2020).
- Georgiou, K. et al. Divergent controls of soil organic carbon between observations and process-based models. Biogeochemistry 2, 775-792 (2021).
- Angst, G., Mueller, K. E., Nierop, K. G. J. & Simpson, M. J. Plant- or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol. Biochem. 156, 108189 (2021).
الشكر والتقدير
مساهمات المؤلفين
الصيغ، وقدم J.P.-R. وN.J.B. وE.W.S. وA.F.A.P. ملاحظات حول تحليلات البيانات. قام K.G. بإجراء جميع التحليلات وكتب المسودة الأولى من الورقة. ساهم جميع المؤلفين في مراجعات الورقة اللاحقة.
المصالح المتنافسة
معلومات إضافية
DOI: https://doi.org/10.1038/s41561-024-01384-7
Publication Date: 2024-02-20
Emergent temperature sensitivity of soil organic carbon driven by mineral associations
Accepted: 23 January 2024
Published online: 20 February 2024
(D) Check for updates
Abstract
Soil organic matter decomposition and its interactions with climate depend on whether the organic matter is associated with soil minerals. However, data limitations have hindered global-scale analyses of mineral-associated and particulate soil organic carbon pools and their benchmarking in Earth system models used to estimate carbon cycle-climate feedbacks. Here we analyse observationally derived global estimates of soil carbon pools to quantify their relative proportions and compute their climatological temperature sensitivities as the decline in carbon with increasing temperature. We find that the climatological temperature sensitivity of particulate carbon is on average
studies have explored the effects of temperature on bulk soil C stocks and turnover rates at ecosystem to global scales
and particulate soil C (derived from an observational synthesis of soil fractions
Climatological temperature sensitivity of soil carbon pools
accounting for potential confounding variables (see ‘Data analysis’ in the Methods). Points depict the proportional decline of each soil C pool (derived from the regression slopes;
soils where higher unprotected C stocks drive a higher temperature sensitivity. The climatological temperature sensitivity of bulk soil C emerges from the underlying pool distributions and their respective temperature dependencies.
Global patterns in soil carbon pools across data and models
mineralogical controls, turnover times and stoichiometry of protected C that are used to approximate the formation and characteristics of mineral-associated organic matter (see ‘Model pool interpretability’ in the Methods). We thus adopt this convention here, while recognizing that there are still inherent uncertainties in the interpretability of modelled pools and operationally defined fractions. We posit that benchmarks based on relative pool distributions, in tandem with radiocarbon ages
Temperature sensitivity of soil carbon in global models
some models may appear to capture the observationally derived climatological temperature sensitivity of bulk soil C stocks, this agreement does not always occur for the right reasons and may break down when considering the contribution of underlying protected and unprotected soil C pools. For example, six of the CMIP6 ESMs (ACCESS-ESM1-5, CESM2, CNRM-ESM2-1, E3SM-1-1-ECA, IPSL-CM6A-LR and NorESM2) and two microbially explicit models (MIMICS and CORPSE) roughly capture the temperature sensitivity of bulk soil C stocks (Fig. 4 and Supplementary Table 6). However, these same models show divergent distributions and climatological temperature sensitivities of underlying protected and unprotected soil C pools (Fig. 4). Only a few models roughly capture the climatological temperature sensitivity of protected C (CESM2, CNRM-ESM2-1, IPSL-CM6A-LR, NorESM2 and CORPSE) and even fewer of unprotected C (ACCESS-ESM1-5, MIMICS and CORPSE).
for potential confounding variables (see ‘Data analysis’ in the Methods). Results are shown for the data product and an ensemble of ESMs and offline land models across gridcells with MAT
of protected C in select models
both cool and warm climates, and of both C pools in warm climates (Fig. 5). This is especially true of MIROC-ES2L, CASA-CNP and MRI-ESM2-0, which overestimate the climatological sensitivity of protected
Implications and future perspectives
| A Data product | ACC
|
E3S
|
Nor
|
| * Multi-model mean | BCC
|
IPS
|
CAS
|
| * Model output | CES
|
MIR
|
MIM
|
| CNR
|
MRI
|
COR
|
circles; labelled with abbreviations); 95% CIs are given in Supplementary Table 6. Kernel density estimates are shown for each panel, illustrating the distribution of climatological temperature sensitivities across the model ensemble for each pool and temperature regime. Higher values (
these unprotected pools with warming, an underestimation of the global carbon cycle-climate feedback and compounding projections of higher productivity-driven C accumulation in these regions. Whereas this bias may be partially offset in ESMs that overestimate the climatological temperature sensitivity of protected C across climates, the effect of this latter overestimate on projections of carbon cycle-climate feedbacks is probably weaker because these pools are intrinsically slow-cycling in first-order soil models and consequently change very little on timescales relevant to climate policy (Supplementary Figs. 6-8). Divergent pool-specific responses can ultimately lead to differences in soil C composition and functioning. It is therefore imperative that models benchmark not only bulk climatological sensitivities but also those of underlying soil C pools, to confidently project soil C dynamics into the future.
Online content
References
- Friedlingstein, P. et al. Uncertainties in CMIP5 climate projections due to carbon cycle feedbacks. J. Clim. 27, 511-526 (2014).
- Davidson, E. A. & Janssens, I. A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440, 165-173 (2006).
- García-Palacios, P. et al. Evidence for large microbial-mediated losses of soil carbon under anthropogenic warming. Nat. Rev. Earth Environ. 2, 507-517 (2021).
- Koven, C. D., Hugelius, G., Lawrence, D. M. & Wieder, W. R. Higher climatological temperature sensitivity of soil carbon in cold than warm climates. Nat. Clim. Chang. 7, 817-822 (2017).
- Crowther, T. et al. Quantifying global soil C losses in response to warming. Nature 104, 104-108 (2016).
- Todd-Brown, K., Zheng, B. & Crowther, T. W. Field-warmed soil carbon changes imply high 21st-century modeling uncertainty. Biogeosciences 15, 3659-3671 (2018).
- Van Gestel, N. et al. Predicting soil carbon loss with warming. Nature 554, E4-E5 (2018).
- Hicks Pries, C. E., Castanha, C., Porras, R. & Torn, M. S. The whole-soil carbon flux in response to warming. Science 355, 1420-1423 (2017).
- Rocci, K. S., Lavallee, J. M., Stewart, C. E. & Cotrufo, M. F. Soil organic carbon response to global environmental change depends on its distribution between mineral-associated and particulate organic matter: a meta-analysis. Sci. Total Environ. 793, 148569 (2021).
- Conant, R. T. et al. Temperature and soil organic matter decomposition rates – synthesis of current knowledge and a way forward. Glob. Chang. Biol. 17, 3392-3404 (2011).
- Soong, J. L. et al. Five years of whole-soil warming led to loss of subsoil carbon stocks and increased
efflux. Sci. Adv. 7, eabd1243 (2021). - Nottingham, A. T., Meir, P., Velasquez, E. & Turner, B. L. Soil carbon loss by experimental warming in a tropical forest. Nature 584, 234-237 (2020).
- Pellegrini, A. F. A. et al. Low-intensity frequent fires in coniferous forests transform soil organic matter in ways that may offset ecosystem carbon losses. Glob. Chang. Biol. 27, 3810-3823 (2021).
- Lugato, E., Lavallee, J. M., Haddix, M. L., Panagos, P. & Cotrufo, M. F. Different climate sensitivity of particulate and mineral-associated soil organic matter. Nat. Geosci. 14, 295-300 (2021).
- Sokol, N. W. et al. Global distribution, formation and fate of mineral-associated soil organic matter under a changing climate: a trait-based perspective. Funct. Ecol. 36, 1411-1429 (2022).
- Lavallee, J. M., Soong, J. L. & Cotrufo, M. F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Chang. Biol. 26, 261-273 (2020).
- Kögel-Knabner, I. et al. Organo-mineral associations in temperate soils: integrating biology, mineralogy, and organic matter chemistry. J. Plant Nutr. Soil Sci. 171, 61-82 (2008).
- Heckman, K. et al. Beyond bulk: density fractions explain heterogeneity in global soil carbon abundance and persistence. Glob. Chang. Biol. 28, 1178-1196 (2022).
- Wieder, W. R., Allison, S. D., Davidson, E. A., Georgiou, K. & Hararuk, O. Explicitly representing soil microbial processes in Earth system models. Glob. Biogeochem. Cycles 29, 1782-1800 (2015).
- Sulman, B. N., Phillips, R. P., Oishi, A. C., Shevliakova, E. & Pacala, S. W. Microbe-driven turnover offsets mineral-mediated storage of soil carbon under elevated
. Nat. Clim. Chang. 4, 1099-1102 (2014). - Ahrens, B., Braakhekke, M. C., Guggenberger, G., Schrumpf, M. & Reichstein, M. Contribution of sorption, DOC transport and microbial interactions to the
age of a soil organic carbon profile: Insights from a calibrated process model. Soil Biol. Biochem. 88, 390-402 (2015). - Sulman, B. N. et al. Multiple models and experiments underscore large uncertainty in soil carbon dynamics. Biogeochemistry 141, 109-123 (2018).
- Wieder, W. R., Sulman, B. N., Hartman, M. D., Koven, C. D. & Bradford, M. A. Arctic soil governs whether climate change drives global losses or gains in soil carbon. Geophys. Res. Lett. 46, 14486-14495 (2019).
- Berardi, D. et al. 21st-century biogeochemical modeling: challenges for Century-based models and where do we go from here? Glob. Chang. Biol. Bioenergy 12, 774-788 (2020).
- Kleber, M. et al. Dynamic interactions at the mineral-organic matter interface. Nat. Rev. Earth Environ. 2, 402-421 (2021).
- Melillo, J. M. et al. Long-term pattern and magnitude of soil carbon feedback to the climate system in a warming world. Science 358, 101-105 (2017).
- Walker, T. W. N. et al. A systemic overreaction to years versus decades of warming in a subarctic grassland ecosystem. Nat. Ecol. Evol. 4, 101-108 (2020).
- Bradford, M. Thermal adaptation of decomposer communities in warming soils. Front. Microbiol. 4, 333 (2013).
- Hartley, I. P., Hill, T. C., Chadburn, S. E. & Hugelius, G. Temperature effects on carbon storage are controlled by soil stabilisation capacities. Nat. Commun. 12, 6713 (2021).
- Doetterl, S. et al. Soil carbon storage controlled by interactions between geochemistry and climate. Nat. Geosci. 8, 780-783 (2015).
- Abramoff, R. Z., Torn, M. S., Georgiou, K., Tang, J. & Riley, W. J. Soil organic matter temperature sensitivity cannot be directly inferred from spatial gradients. Glob. Biogeochem. Cycles 33, 761-776 (2019).
- Luo, Z. et al. Convergent modelling of past soil organic carbon stocks but divergent projections. Biogeosciences 12, 4373-4383 (2015).
- Georgiou, K. et al. Global stocks and capacity of mineral-associated soil organic carbon. Nat. Commun. 13, 3797 (2022).
- Eyring, V. et al. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 9, 1937-1958 (2016).
- Jobbágy, E. G. & Jackson, R. B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 10, 423-436 (2000).
- Ahrens, B. et al. Combination of energy limitation and sorption capacity explains
depth gradients. Soil Biol. Biochem. 148, 107912 (2020). - Gentsch, N. et al. Temperature response of permafrost soil carbon is attenuated by mineral protection. Glob. Chang. Biol. 24, 3401-3415 (2018).
- Giardina, C. P. & Ryan, M. G. Evidence that decomposition rates of organic carbon inmineral soil do not vary with temperature. Nature 404, 858-861 (2000).
- Giardina, C. P., Litton, C. M., Crow, S. E. & Asner, G. P. Warming-related increases in soil
efflux are explained by increased below-ground carbon flux. Nat. Clim. Chang. 4, 822-827 (2014). - Wang, C. et al. The temperature sensitivity of soil: microbial biodiversity, growth, and carbon mineralization. ISME J. 15, 2738-2747 (2021).
- Shi, Z. et al. The age distribution of global soil carbon inferred from radiocarbon measurements. Nat. Geosci. 13, 555-559 (2020).
- He, Y. et al. Radiocarbon constraints imply reduced carbon uptake by soils during the 21st century. Science 353, 1419-1424 (2016).
- Todd-Brown, K. E. O. et al. Causes of variation in soil carbon simulations from CMIP5 Earth system models and comparison with observations. Biogeosciences 10, 1717-1736 (2013).
- Ito, A. et al. Soil carbon sequestration simulated in CMIP6-LUMIP models: implications for climatic mitigation. Environ. Res. Lett. 15, 124061 (2019).
- Abramoff, R. Z. et al. Improved global-scale predictions of soil carbon stocks with Millennial Version 2. Soil Biol. Biochem. 164, 108466 (2022).
- Zimmermann, M., Leifeld, J., Schmidt, M. W. I., Smith, P. & Fuhrer, J. Measured soil organic matter fractions can be related to pools in the RothC model. Eur. J. Soil Sci. 58, 658-667 (2007).
- Pierson, D. et al. Optimizing process-based models to predict current and future soil organic carbon stocks at high-resolution. Sci. Rep. 12, 10824 (2022).
- Koven, C. et al. On the formation of high-latitude soil carbon stocks: effects of cryoturbation and insulation by organic matter in a land surface model. Geophys. Res. Lett. https://doi. org/10.1029/2009GL040150 (2009).
- Koven, C. D. et al. The effect of vertically resolved soil biogeochemistry and alternate soil C and N models on C dynamics of CLM4. Biogeosciences 10, 7109-7131 (2013).
- Plaza, C. et al. Direct observation of permafrost degradation and rapid soil carbon loss in tundra. Nat. Geosci. 12, 627-631 (2019).
- Parton, W. J., Schimel, D. S., Cole, C. V. & Ojima, D. S. Analysis of factors controlling soil organic matter levels in Great Plains grasslands. Soil Sci. Soc. Am. J. 51, 1173-1179 (1987).
- Haaf, D., Six, J. & Doetterl, S. Global patterns of geo-ecological controls on the response of soil respiration to warming. Nat. Clim. Chang. 11, 623-627 (2021).
- Jones, C. D. et al. C4MIP – the Coupled Climate-Carbon Cycle Model Intercomparison Project: experimental protocol for CMIP6. Geosci. Model Dev. 9, 2853-2880 (2016).
- Bouskill, N. J., Riley, W. J., Zhu, Q., Mekonnen, Z. A. & Grant, R. F. Alaskan carbon-climate feedbacks will be weaker than inferred from short-term experiments. Nat. Commun. 11, 5798 (2020).
adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons. org/licenses/by/4.0/.
(c) The Author(s) 2024
Methods
Data sources and processing
Global land model output
Model pool interpretability
the Reynolds Creek Experimental Watershed
Data analysis
analyses were performed on an observational synthesis
Data availability
Code availability
References
- Harris, I., Jones, P. D., Osborn, T. J. & Lister, D. H. Updated high-resolution grids of monthly climatic observations – the CRU TS3.10 Dataset. Int. J. Climatol. 34, 623-642 (2014).
- Schneider, U. et al. GPCC Full Data Reanalysis Version 6.0 at
: Monthly Land-Surface Precipitation from Rain-Gauges built on GTS-based and Historic Data (GPCC, 2011); https://doi. org/10.5676/DWD_GPCC/FD_M_V6_050 - Friedl, M. A. et al. MODIS Collection 5 global land cover: algorithm refinements and characterization of new datasets. Remote Sens. Environ. 114, 168-182 (2010).
- Zhao, M., Heinsch, F. A., Nemani, R. R. & Running, S. W. Improvements of the MODIS terrestrial gross and net primary production global data set. Remote Sens. Environ. 95, 164-176 (2005).
- Wieder, W. R., Boehnert, J., Bonan, G. B. & Langseth, M. Regridded Harmonized World Soil Database v1.2 (ORNL DAAC, 2014); https:// doi.org/10.3334/ORNLDAAC/1247
- Hengl, T. et al. SoilGrids250m: global gridded soil information based on machine learning. PLoS ONE https://doi.org/10.1371/ journal.pone. 0169748 (2017).
- Georgiou, K. et al. Globally-gridded data for manuscript: global stocks and capacity of mineral-associated soil organic carbon. Zenodo https://doi.org/10.5281/zenodo. 6539765 (2022)
- Ploton, P. et al. Spatial validation reveals poor predictive performance of large-scale ecological mapping models. Nat. Commun. 11, 4540 (2020).
- Meyer, H. & Pebesma, E. Machine learning-based global maps of ecological variables and the challenge of assessing them. Nat. Commun. 13, 2208 (2022).
- Thornton, P. E. & Rosenbloom, N. A. Ecosystem model spin-up: estimating steady state conditions in a coupled terrestrial carbon and nitrogen cycle model. Ecol. Modell. 189, 25-48 (2005).
- Lawrence, D. M. et al. Parameterization improvements and functional and structural advances in Version 4 of the Community Land Model. J. Adv. Model. Earth Syst. https://doi. org/10.1029/2011MS00045 (2011).
- Lee, W. L. et al. Taiwan Earth System Model Version 1: description and evaluation of mean state. Geosci. Model Dev. 13, 3887-3904 (2020).
- Christensen, B. T. in Evaluation of Soil Organic Matter Models Vol. 38 (eds Powlson, D. S. et al.) 143-159 (Springer, 1996).
- Elliot, E. T., Paustian, K. & Frey, S. D. in Evaluation of Soil Organic Matter Models Vol. 38 (eds Powlson, D. S. et al.) 161-179 (Springer, 1996).
- Lehmann, J. & Kleber, M. The contentious nature of soil organic matter. Nature 528, 60-68 (2015).
- Zhang, Y. et al. Simulating measurable ecosystem carbon and nitrogen dynamics with the mechanistically defined MEMS 2.0 model. Biogeosciences 18, 3147-3171 (2021).
- Dangal, S. R. S. et al. Improving soil carbon estimates by linking conceptual pools against measurable carbon fractions in the DAYCENT model version 4.5. J. Adv. Model. Earth Syst. 14, e2021MSOO2622 (2022).
- Abramoff, R. et al. The Millennial model: in search of measurable pools and transformations for modeling soil carbon in the new century. Biogeochemistry 137, 51-71 (2018).
- Parton, W. J. & Rasmussen, P. E. Long-term effects of crop management in wheat-fallow: II. CENTURY model simulations. Soil Sci. Soc. Am. J. 58, 530-536 (1994).
- Cotrufo, M. F., Ranalli, M. G., Haddix, M. L., Six, J. & Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 12, 989-994 (2019).
- Rocci, K. S. et al. Aligning theoretical and empirical representations of soil carbon-to-nitrogen stoichiometry with process-based terrestrial biogeochemistry models. Soil Biol. Biochem. 189, 109272 (2024).
- Lawrence, C. R. et al. An open-source database for the synthesis of soil radiocarbon data: International Soil Radiocarbon Database (ISRaD) version 1.0. Earth Syst. Sci. Data 12, 61-76 (2020).
- Kyker-Snowman, E., Wieder, W. R., Frey, S. D. & Grandy, A. S. Stoichiometrically coupled carbon and nitrogen cycling in the MIcrobial-MIneral Carbon Stabilization model version 1.0 (MIMICS-CN v1.0). Geosci. Model Dev. 13, 4413-4434 (2020).
- Georgiou, K. et al. Divergent controls of soil organic carbon between observations and process-based models. Biogeochemistry 2, 775-792 (2021).
- Angst, G., Mueller, K. E., Nierop, K. G. J. & Simpson, M. J. Plant- or microbial-derived? A review on the molecular composition of stabilized soil organic matter. Soil Biol. Biochem. 156, 108189 (2021).
Acknowledgements
Author contributions
formulations, and J.P.-R., N.J.B., E.W.S. and A.F.A.P. provided feedback on the data analyses. K.G. performed all analyses and wrote the first draft of the paper. All authors contributed to subsequent paper revisions.