اللاكتات ينشط المناعة المدربة من خلال تغذية دورة حمض الثلاثي الكربوكسيليك وتنظيم اللاكتيل في الهيستونات Lactate activates trained immunity by fueling the tricarboxylic acid cycle and regulating histone lactylation
اللاكتات ينشط المناعة المدربة من خلال تغذية دورة حمض الثلاثي الكربوكسيليك وتنظيم اللاكتيل في الهيستونات
تاريخ الاستلام: 6 أغسطس 2024
تم القبول: 24 مارس 2025
نُشر على الإنترنت: 04 أبريل 2025 تحقق من التحديثات
Huanhuan Cai ${ }^{1,2,11}$, Xueyuan Chen ${ }^{3,11}$, Yan Liu ${ }^{3,11}$, Yingbo Chen ${ }^{4,5}$, Gechang Zhong ${ }^{4,5}$, Xiaoyu Chen ${ }^{4,5}$, Shuo Rong ${ }^{4,5}$, Hao Zeng ${ }^{6}$, Lin Zhang ${ }^{1,2}$, Zelong Li ${ }^{7,8}$, Aihua Liao ${ }^{7,8}$, Xiangtai Zeng ${ }^{7,8}$, Wei Xiong ${ }^{1,2}$, Cihang Guo ${ }^{1,2}$, Yanfang Zhu ${ }^{1,2}$, Ke-Qiong Deng ${ }^{1,2}$, Hong Ren ${ }^{9}$, Huan Yan ${ }^{(1)}{ }^{4,5}$, Zeng Cai ${ }^{4,5}$, Ke Xu ${ }^{4,5}$, Li Zhou ${ }^{(1)}{ }^{4,5}$, Zhibing Lu ${ }^{(1)}{ }^{1,2}$, Fubing Wang ${ }^{(1)}{ }^{10}$ & Shi Liu ${ }^{(1)}{ }^{1,2,4,5}$ ®
تشير المناعة المدربة إلى الذاكرة طويلة الأمد لخلايا المناعة الفطرية. ومع ذلك، لا يُعرف الكثير عن كيفية تأثير توفر المغذيات البيئية على المناعة المدربة. تجد هذه الدراسة أن المصادر الكربونية الفسيولوجية تؤثر على مساهمة الجلوكوز في دورة حمض التريكربوكسيليك (TCA) وتعزز إنتاج السيتوكينات من وحيدات النوى المدربة. تُظهر تجاربنا أن وحيدات النوى المدربة تفضل استخدام اللاكتات بدلاً من الجلوكوز كمادة أساسية لدورة TCA، وأن استقلاب اللاكتات ضروري لاستجابة خلايا المناعة المدربة للعدوى البكتيرية والفطرية. باستثناء المساهمة في دورة TCA، فإن اللاكتات الذاتية أو اللاكتات الخارجية تدعم أيضًا المناعة المدربة من خلال تنظيم اللاكتيل في الهيستونات. تُظهر المزيد من تحليلات النسخ الجيني، وATAC-seq، وCUT&Tag-seq أن اللاكتات تعزز من وصول الكروماتين بطريقة تعتمد على اللاكتيل في الهيستونات. إن تثبيط استقلاب اللاكتات المعتمد على اللاكتات عن طريق كتم إنزيم لاكتات ديهيدروجيناز A (LDHA) يعيق كل من دورة TCA المدفوعة باللاكتات واللاكتيل في الهيستونات. تشير هذه النتائج إلى أن اللاكتات هي محور البرامج المناعية الأيضية والوراثية في المناعة المدربة.
لقد ثبت مؤخرًا أن إعادة برمجة العمليات الأيضية الخلوية ضرورية للخلايا المناعية.مفهوم مقبول على نطاق واسع هو أن العوامل الداخلية للخلايا، مثل الجزيئات الحيوية أو الإنزيمات الأيضية، تلعب دورًا حاسمًا في المسار الأيضي في خلايا المناعة.. ومع ذلك، فإن توفر العناصر الغذائية في البيئة أيضًا بشكل كبير يؤثر على الاستخدام الأيضي في خلايا المناعةعلى سبيل المثال، أثرت تركيزات ركيزة الخميرة على الناتج الأيضي مرتين أكثر من تعبير الإنزيم.تظهر خلايا السرطان تغييرات في نمو الخلايا ومقاومة الإجهاد إذا تم زراعة خلايا السرطان في وسط فسيولوجي بسبب الاختلافات في استخدام الركيزة.. من بين العناصر الغذائية في
في البيئة، يعتبر الجلوكوز مغذٍّ هامٍّ يغذي الأنشطة الأيضية الخلوية. بعد امتصاصه من خلال ناقل الجلوكوز، يتدفق الجلوكوز عبر ثلاث مسارات أيضية متميزة: التحلل السكري، ومسار الفوسفات الخماسي، ومسار تخليق الهيكسوزامين.يتم استقلاب الجلوكوز عبر التحلل السكري إلى البيروفات، الذي يدخل دورة TCA عندما يكون الأكسجين متاحًا.عندما يكون الأكسجين غير متوفر، يتحول البيروفات إلى التحلل السكري، حيث يتحول البيروفات إلى حمض اللبنيك.تقليديًا، كان يُنظر إلى اللاكتات على أنها نفايات من عملية استقلاب الجلوكوز.. ومع ذلك، تظهر مجموعة متزايدة من الأبحاث أن اللاكتات هي المستقلب الرئيسي المتداول الذي يغذي دورة TCA وركيزة هامة للجلوكوجين.حدد دراسة حديثة تعديلًا وراثيًا جديدًا، وهو استحلاب الهيستون، الذي ينظم الهيستونات عن طريق إضافة مجموعات لاكتيل إلى بقايا الليسين.تتميز لاكتيلation الهيستون بديناميات زمنية مختلفة عن الأسيتيلation وتسبب تعبير الجينات المقابل.. بعد ذلك، تم تحديد أكثر من 30 موقعًا لكتلة الهيستون في عينات فطرية ونباتية وفأرية وطفيليات (أوليات) .
تقول الحكمة التقليدية إن جهاز المناعة مقسم إلى جهاز المناعة الفطري وجهاز المناعة التكيفي. واحدة من الأسس الرئيسية لهذا التقسيم هي أن جهاز المناعة التكيفي أبطأ نسبيًا ويمكنه بناء ذاكرة مناعية محددة.على النقيض من ذلك، فإن الجهاز المناعي الفطري سريع وغير محدد ولا يمكنه بناء ذاكرة مناعية.ومع ذلك، أظهرت الدراسات الحديثة أن خلايا المناعة الفطرية، مثل خلايا القاتل الطبيعي والوحيدات، يمكن أن تشكل ذاكرة غير محددة تتحدى هذا النموذج.على وجه التحديد، فإن التكيف الوظيفي للمناعة الفطرية بعد إصابة أولية، بما في ذلك تطعيم باسيلي كالميت-غيران أو عدوى الميكروبات، يمثل ذاكرة مناعية فطرية بحكم الواقع.بمجرد أن تواجه هذه الخلايا المناعية الفطرية محفزًا ثانويًا غير محدد (غير متجانس)، يتغير استجابتها، مما يؤدي إلى استجابة أكثر قوة.يؤدي تحفيز ذاكرة غير محددة إلى زيادة وظيفة خلايا المناعة الفطرية المعروفة بالمناعة المدربة.الأهمية البيولوجية للذاكرة المناعية الفطرية، مثل الرخويات، والنباتات، والحشرات، والثدييات، واسعة الانتشار.تحليل المسارات المنظمة بشكل مختلف في المناعة المدربة أظهر أن mTOR-HIF1تنظيم الفسفرة التأكسدية إلى التحلل السكري، الآلية التنظيمية المركزية لـ-غلوكان، مكون جدار الخلية الفطرية، أدى إلى تحفيز المناعة المدربةومع ذلك، لا يزال تأثير توفر العناصر الغذائية البيئية على استقلاب المناعة المدربة ووظيفتها غير مفهوم بشكل جيد.
في هذه الدراسة، وجدنا أن المصادر الكربونية الفسيولوجية (PCSs) تؤثر على استخدام الجلوكوز من قبل المونوسيتات المدربة. لقد أظهرت تجاربنا أن اللاكتات هي وقود فسيولوجي للمناعة المدربة، ويفضل على الجلوكوز، وتنظم إنتاج السيتوكينات من قبل خلايا المناعة المدربة. من الناحية الآلية، تعتبر بعض وظائف اللاكتات الداخلية ركيزة هامة لتخليق الأسيتيل-CoA وتغذي دورة TCA. بينما تنتقل اللاكتات الأخرى إلى نواة الخلية وتنظم إنتاج السيتوكينات من خلال تأثيرات أسيتيل اللاكتات على الهيستونات. تحدد بياناتنا المستقلب السائد (اللاكتات) في المناعة المدربة، وتساهم في فهمنا لذاكرة المناعة الفطرية، وتفتح آفاق علاجية جديدة.
النتائج
تؤثر PCS على استخدام الجلوكوز في وحيدات النواة خلال تحفيز المناعة المدربة
تم تصميم وسائط زراعة الخلايا التقليدية، مثل وسط معهد روسويل بارك التذكاري (RPMI) 1640، لتعزيز نمو الخلايا وبقائها خارج الجسم.. وبالتالي، فإن تركيز بعض العناصر الغذائية في وسط زراعة الخلايا التقليدي هو فوق الفسيولوجي (الشكل التوضيحي 1a). بالإضافة إلى ذلك، فإن الوسط التقليدي يفتقر أيضًا إلى معظم وسائط الزراعة القياسية، مثل اللاكتات، والسيترات، والأسيتات،-هيدروكسي بيوتيراتوالبيروفاتفي هذه الدراسة، أطلقنا على هذه المستقلبات اسم PCSs (الشكل التوضيحي 1a). تظهر الدراسات السابقة أن هذه المصادر الكربونية هي ركائز قابلة للأكسدة في الجسم الحي.من الجدير بالذكر أنه في بعض الحالات، تظهر هذه المصادر الكربونية تدفقات دوران دورية. تجاوز المصادر التقليدية، مثل الجلوكوز والجلوتامينلذا، قمنا بتصميم وسط RPMI المعدل (MM) بتركيزات تقريبية من المستقلبات القطبية في مصل الفئران (الشكل التوضيحي 1a).
للحصول على خلايا مناعية مدربة، قمنا بمعالجة المونوسيتات المنقاة بـ-غلوكان لمدة 24 ساعة، تلاه غسل وراحة لمدة خمسة أيام في وسط زراعة. ثم تم إعادة تحفيز الخلايا بـ-غلوكان أو LPS لمدة 24 ساعة كما هو موصوف سابقًا (الشكل التوضيحي التكميلي 1ب). توسيم الجلوكوز إلى اللاكتات ووسائط دورة حمض الستريك داخل الخلايا، بما في ذلك السترات والفومارات، ضمنفيالمونوسيتات المعالجة بـ -غلوكان (الأشكال التكميلية 1c). أظهر تحليل مستويات المستقلبات في وسط الثقافة على مر الزمن عدة اتجاهات (الأشكال التكميلية 1d-1f). كما هو متوقع، تم استهلاك الجلوكوز والجلوتامين خلال 12-24 ساعة أثناء عملية المناعة المدربة، وأثر علاج PCS بشكل طفيف على استهلاك الجلوكوز والجلوتامين فيتمت معالجة وحيدات النوى بالغلوكوز (الأشكال التكميلية 1d و1e). لوحظ انخفاض أبطأ في مصادر الكربون الأخرى، بما في ذلك OHB، السترات، والبيروفات، في الوسط المدعوم بـ PCS مقارنة بالوسط المدعوم بالغلوكوز (الشكل التكميلية 1f). ومن المثير للاهتمام، أننا لاحظنا تراكم عدة وسائط من دورة TCA (و الفومارات) في الوسط خارج الخلوي مع مرور الوقت، مما يدل على تصدير هذه المستقلبات (الشكل التوضيحي التكميلي 1f).
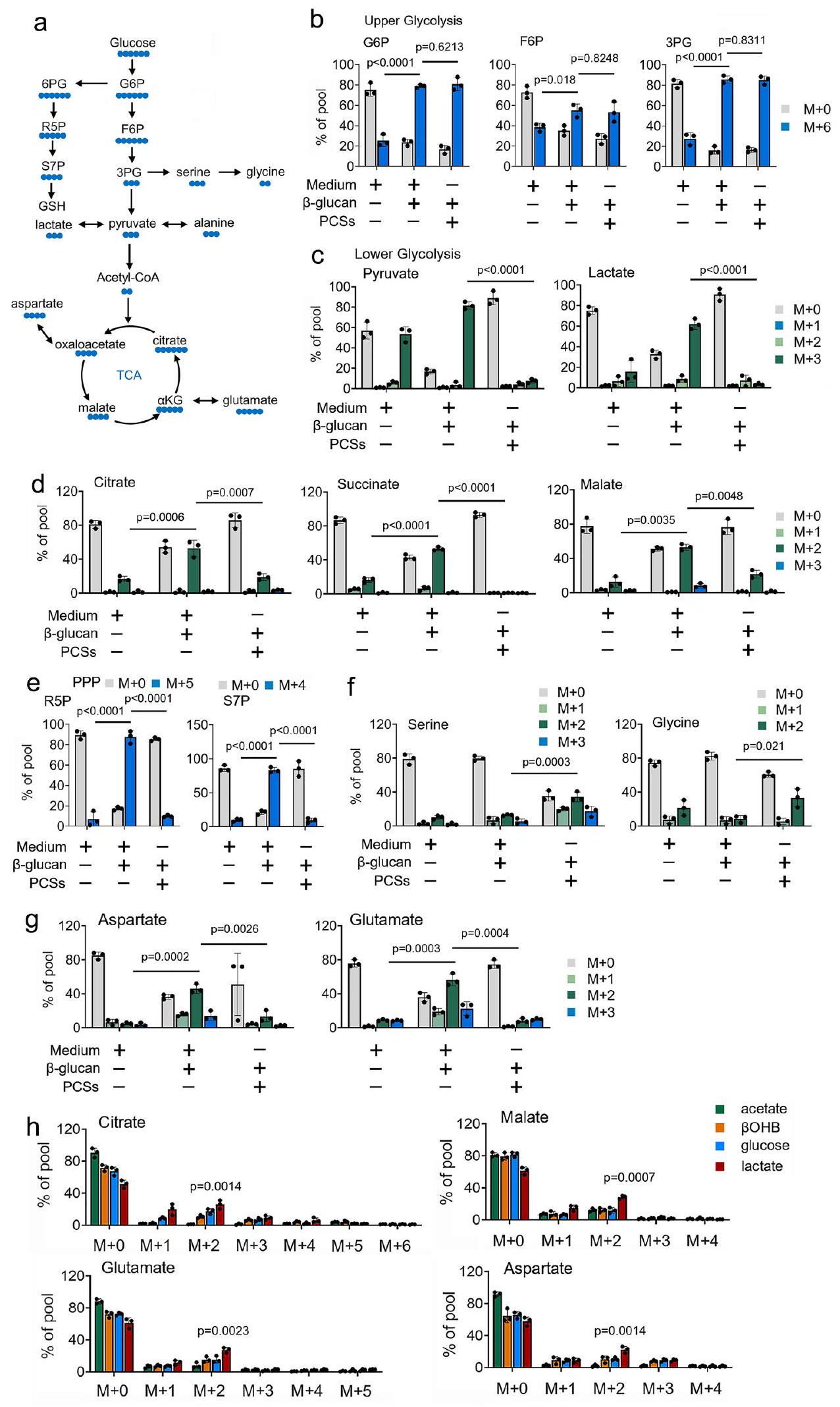
استخدامالمتتبعات الأيضية المعتمدة على -، نحقق في دور PCSs في استقلاب الجلوكوز في المناعة المدربة (الشكل 1a). تظهر تجربة مطيافية الكتلة (MS) أن-الجلوكوز يوسم بشكل واسع الوسائط في عملية التحلل السكري ودورة حمض الستريك فيالمونوسيتات المعالجة بـ -غلوكان (الأشكال 1ب-1د). على النقيض من ذلك، في وجود PCS عند تركيز فسيولوجي فيالمونوسيتات المعالجة بـ -غلوكان،تم تثبيط مساهمة الجلوكوز في وسائط دورة TCA بشكل كبير (الأشكال 1ب-1د). من المثير للاهتمام أن وجود PCSs بتركيزات فسيولوجية يوجه تدفق الجلوكوز إلى PPP وطريق تخليق السيرين (الأشكال 1هـ و1د). تم الحصول على نتائج مماثلة في المستقلبات الناتجة عن تدهور TCA، مثل الغلوتامات والأسبارتات (الشكل 1ز). نلاحظ نتائج مماثلةأنماط التسمية فيوحيدات النواة المعالجة بـ -غلوكان المزروعة في PCS و-الجلوكوز لفترة طويلة (6 أيام) (الشكل التوضيحي 1g). للتحقيق في مساهمة PCSs في استقلاب دورة TCA،تم زراعة وحيدات النواة المعالجة بـ -غلوكان في وسط MM المدعوم بـ “-المركبات المعلمة عند تركيزات فسيولوجية. نلاحظ أن-جلوكوز و-جلوتامين ساهم في توفير الكربون لوسائط دورة TCA (الشكل التوضيحي 1h). ومن المثير للاهتمام أن وسائط دورة TCA تم وسمها أيضًا من بعض PCSs، بما في ذلكواللاكتات، فيالمونوسيتات المعالجة بـ -غلوكان (الشكل التوضيحي 1h). أظهرت النتائج أنويساهم اللاكتات في إضافة الكربون إلى دورة TCA من خلال دخول 2 كربون عبر إنزيم سيترات سينثاز (سترات) وتحمل خلال الدورة بأكملها فيالمونوسيتات المعالجة بـ -غلوكان (الشكل 1h). من الجدير بالذكر أنه عند النظر في التركيز المنخفض لللاكتيتمقارنة بتركيز الجلوكوزتفضل المونوسيتات المدربة استخدام الأحماض الدهنية بدلاً من الجلوكوز لعملية الأيض في دورة حمض الستريك (الشكل 1h). لأنه قد تم الإبلاغ عن أن الألانين لم يساهم في دورة حمض الستريك.وجودتم تضمين -ألانين كعنصر تحكم سلبي للمقارنة (الشكل التوضيحي التكميلي 1h). مجتمعة، تشير هذه النتائج إلى أن PCSs قللت من مساهمة-الجلوكوز إلى دورة TCA، ويدفع-تحويل الجلوكوز من التحلل السكري إلى مسار الفوسفات pentose و SSP، في المناعة المدربة.
اللاكتات ينشط المناعة المدربة
نظرًا لأن اللاكتات لديها التنظيم الأكثر أهمية على دورة TCA بين هذه الـ PCSs المذكورة أعلاه (الشكل 1h)، قمنا بعد ذلك بالتحقيق في دور اللاكتات في المناعة المدربة استجابةً لعدوى C. albicans (الشكل 2a). بما يتماشى مع دراسة سابقةتزيد عدوى C. albicans من إنتاج اللاكتات بطريقة تعتمد على الوقت والجرعة (الشكل التوضيحي 2a). نظرًا لأن إنزيم لاكتات ديهيدروجيناز (Ldh) يقوم بتحويل اللاكتات إلى البيروفات باستخدام NAD/NADH كعامل مساعد، سألنا عما إذا كانت LDHA تلعب دورًا في المناعة المدربة. أولاً، استخدمنا الفئران المعدلة وراثيًا التي تم حذف LDHA فيها بواسطة التاموكسيفين ()، و
الشكل 1 | تؤثر PCSs على استخدام الجلوكوز في المناعة المدربة. أ رسم تخطيطي لـ-توسيم الكربون للجلوكوز من خلال دورة TCA، ومسار الفوسفات الخماسي (PPP)، ومسار تخليق السيرين (SSP). تم زراعة البا-ج الكريات البيضاء الأحادية في وسط MM يحتوي على-جلوكوز ( ) وعولجت بـ -غلوكان ( ) لمدة 4 ساعات، تليها تحليلات الطيف الكتلي. تم زراعة وحيدات النوى في MM تحتوي على ما هو محددمصدر الكربون المعلم و المعالج بـ-غلوكان (مل) لمدة 4 ساعات. المساهمات النسبية لـمن المشار إليهالمواد الفرعية المعلّمة إلى المستقلبات في عملية الأيض الكربوني المركزي موضحة. البيانات في (ب-ح) معبّرة كمتوسطات ± الانحراف المعياري.الفئران لكل حالة، تحليل التباين ثنائي الاتجاه. الوسط: محلول ملحي مخفف بالفوسفات. انظر أيضًا الشكل التكميلية 1.
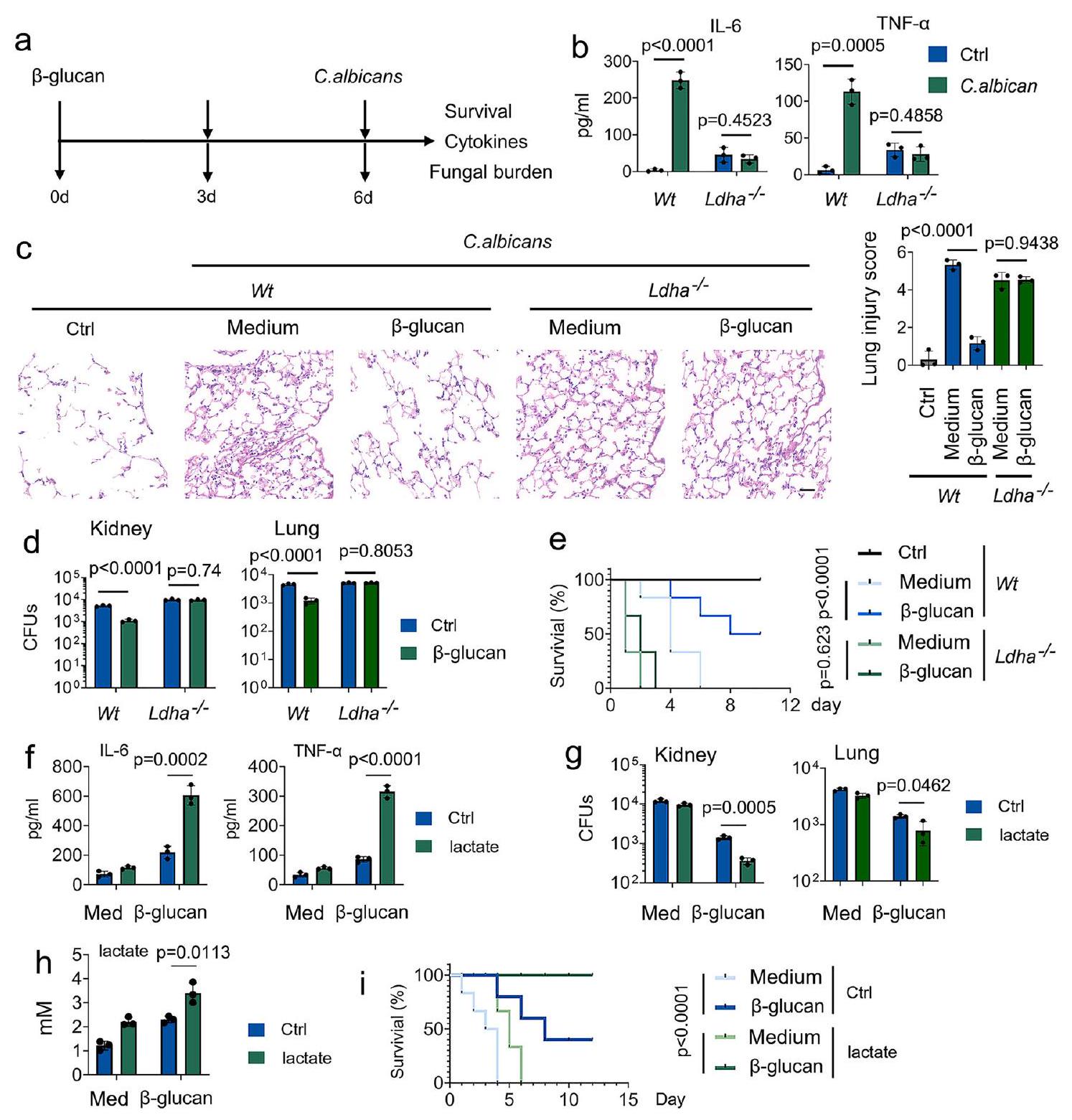
الشكل 2 | اللاكتات تنظم المناعة المدربة. أ تمثيل تخطيطي لإعداد تجربة المناعة المدربة في الجسم الحي. ب-د و تم تدريب الفئران مع أو بدون-غلوكان. بعد ستة أيام، تم إصابة الفئران مع أو بدون ثانويةالجرعة القاتلة من الكانديدا البيضلمدة 24 ساعة، تليها قياس IL-6 و TNF-المستويات (ب)، إصابة الرئة (ج)، وحمل الفطريات في الكلى والرئة (د). هـ و تم تدريب الفئران مع أو بدون-غلوكان. بعد ستة أيام، تم إصابة الفئران بجرعة قاتلة ثانوية من C. albicans أو بدونها ( ) للأوقات المحددة. تظهر منحنيات البقاء البيانات التي تم جمعها حتى اليوم العاشر بعد العدوى. تم إجراء التحليل الإحصائي باستخدام اختبار لوغ-رانك ( لكل مجموعة).تم تدريب فئران C57BL/6 مع أو بدون-غلوكان. بعد ستة أيام، تم إصابة الفئران ثانويًا بدون أو مع C. albicans ( ) وتم حقنها مع أو بدون لاكتات ( 1 مللي مول ) لمدة 24 ساعة، تليها قياس IL-6 و TNF-المستويات (ف)، عبء الفطريات في الكلى (ز)، ومستويات اللاكتات في المصل (ح). تم تدريب الفئران من سلالة C57BL/6 مع أو بدونغلوكان. بعد ستة أيام، تم إصابة الفئران بجرعة قاتلة ثانوية من C. albicans أو بدونها. ) وتمت إضافة اللاكتات ( 1 مللي مول ) مع أو بدونها لفترات زمنية محددة. تظهر منحنيات البقاء البيانات المجمعة حتى اليوم 12 بعد العدوى. تم إجراء التحليل الإحصائي باستخدام اختبار لوغ-رانك ( لكل مجموعة). البيانات في (ب-د، و ) يتم التعبير عنها كمتوسطات ± الانحراف المعياري للخطأ، الفئران لكل حالة، تحليل التباين ثنائي الاتجاه. انظر أيضًا الأشكال التكميلية 2.
تعبير بروتين LDHA في أعضاء متنوعة منتم إلغاء الفئران عند علاج التاموكسيفين (الأشكال التكميلية 2ب و2ج). للتحقيق في دور اللاكتات على وظيفة وحيدات النوى المدربة، قمنا بتدريب الفئران مع-غلوكان تليه تحدي مع. الكانديدا (الشكل 2أ). كما هو متوقع، IL-6 و TNF-تم تحفيز الإنتاج فيمدرب على -غلوكانفئران، ولكن ليس فيالفئران (الشكل 2ب). أظهر التحليل النسيجي لنسج الرئة أن قبلعلاج -غلوكان يحد من تلف الرئة خلال العدوى اللاحقة (الشكل 2ج). مقارنةً بـمدرب على -غلوكانالفئران، تم إثبات إصابة أكثر شدة في الرئة فيالفئران (الشكل 2ج). في هذا السياق،الفئران عرضة بشكل كبير للفتك الناتج عن C. albicans، مصحوبة بزيادة الحمل البكتيري في أنسجة الرئة والكلى (الأشكال 2د و2هـ). دراسة سابقة أُبلغ أن المناعة المدربة هي مضيف واقٍ ضد عدوى C. albicans الثانوية التي تعتمد على وحيدات النوى المستحثة بواسطة نفس العامل الممرض.. تم الحصول على نتائج مماثلة في سلالات C. albicans المدربة Wt والفئران (الأشكال التكميلية 2d-2g). لتحديد ما إذا كان LDHA ينظم المناعة المدربة في المختبر، استخدمنا نوعين محددين من الأحماض النووية الصغيرة المداخلة (siRNAs) لـ LDHA. قمنا بتأكيد كفاءة siRNA (الشكل التكميلية 2h). تم اختيار siRNA-LDHA #2 للتجارب التالية. كما هو موضح في الشكل التكميلية 2i، فإن تقليل LDHA يمنعتحفيز التعبير عن IL-6 و TNF- بواسطة -غلوكانفي خلايا الدم المحيطية الوحيدة النواة البشرية (PBMCs).
بعد ذلك، بحثنا في دور اللاكتات في المناعة المدربة استجابةً لعدوى C. albicans. كما هو موضح في الشكل التوضيحي الإضافي 2j، اللاكتات يعززتحفيز التعبير عن IL-6 و TNF- بواسطة -غلوكان بطريقة تعتمد على الجرعة في خلايا الدم البيضاء المحيطية البشرية. ومن الجدير بالذكر أنه لاستبعاد احتمال أن يؤثر اللاكتات على المناعة المدربة من خلال تغيير الحموضةتم تعديل درجة الحموضة في الوسط إلى مستوى متساوٍ في جميع التجارب المخبرية في هذا المخطوط. زادت وجود اللاكتات من مستويات IL-6 و TNF- التي تسببها C. albicans.، مصحوبًا بعبء فطري أقل في أنسجة الكلى والرئة (الأشكال 2f-2h). بشكل لافت، زاد اللاكتات من الاستجابة الوقائية للوحيدات المدربة ضد عدوى C. albicans المميتة (الشكل 2i). تشير هذه النتائج إلى أن اللاكتات ضرورية للتفاعل الوقائي للمناعة المدربة ضد عدوى C. albicans.
اللاكتات هو وقود فسيولوجي خلال تحفيز المناعة المدربة
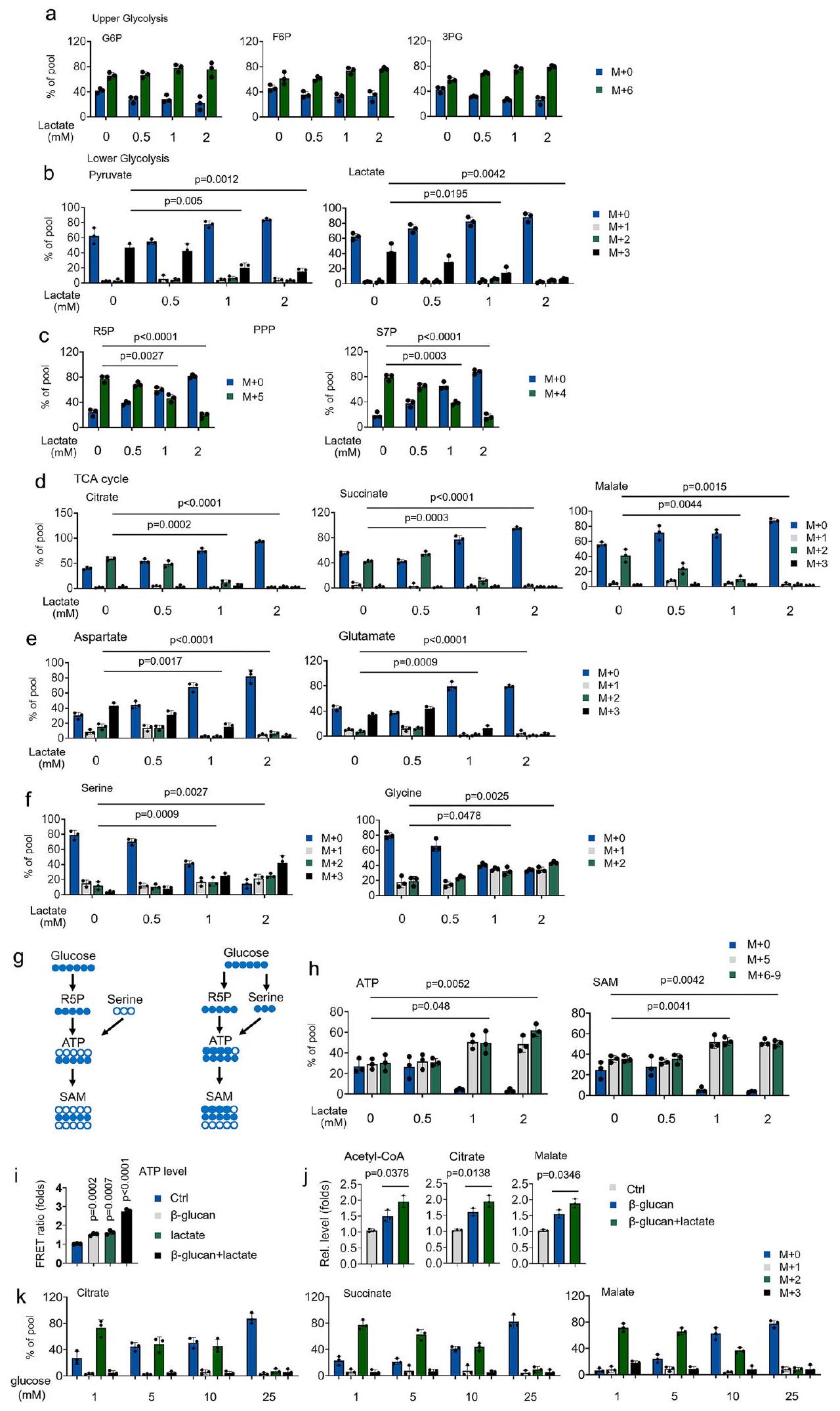
بعد ذلك، نحقق في الآليات التي ينظم بها اللاكتات المناعة المدربة. أولاً، قمنا بقياس الجدول الزمني لـدمج الجلوكوز خلالعلاج -غلوكان.أحادية النواة المعالجة بـ -غلوكان أظهرت مستويات أعلىتشير دمج الجلوكوز في الجلوكوز-6-فوسفات (G6P)، الفركتوز-6-فوسفات (F6P)، 3-فوسفوغليسرات (3PG)، البيروفات، اللاكتات، السترات، السكسينات، الماليك، الغلوتامات والأسبارتات خلال 0-4 ساعات (الأشكال التكميلية 3a-3f) إلى زيادة تدفق الجليكوليز إلى دورة TCA.تدفق دمج الجلوكوز من الفسفرة التأكسدية (OxPhos) إلى SSP وPPP، بالإضافة إلى تخليق ATP وتوليد S-أدينوزيل ميثيونين (SAM)، بعد 4 ساعات منعلاج الجلوكان (الأشكال التكميلية 3a-3g). ومع ذلك،استمر دمج الجلوكوز في عملية التحلل السكري في الزيادة طوال التجربة (الشكل التكميلي 3ب). بالإضافة إلى ذلك،-غلوكان أيضًا حفز إنزيمات هيكسوكيناز 2 (HK2) ونشاط إنزيم LDHA بطريقة تعتمد على الوقت والجرعة (الأشكال التكميلية 3h و3i). تشير هذه البيانات إلىتفضل وحيدات النواة المعالجة بـ -غلوكان استخدام الجلوكوز كوقود فسيولوجي في البداية. ومع ذلك،تفضل وحيدات النواة المعالجة بـ -غلوكان استخدام الجلوكوز للتخليق الحيوي لاحقًا.
لتقييم المنافسة بين الجلوكوز واللاكتات في استقلاب دورة TCA، تم زراعة وحيدات النواة المعالجة مع-الجلوكوز واللاكتات غير المعلمة مع زيادة تدريجية في التركيز. حتى في وجود تركيزات الجلوكوز الفسيولوجية (5 مللي مول) ،تستخدم وحيدات النوى المعالجة بـ -غلوكان تركيزات منخفضة من اللاكتات (كمصدر للكربون للوسائط في دورة TCA (الأشكال 3a-3d). بالمقابل، فإن تركيز منخفض من اللاكتاتيوجه الجلوكوزالتدفق من OxPhos إلى SSP و PPP (الأشكال 3a-3f). جزء كبير من SAM المعلم المشتق من الجلوكوز، بما في ذلك (عبر برنامج الشراكة بين القطاعين العام والخاص) و (عبر كل من PPP و SSP)، زادت بشكل ملحوظ في وحيدات النواة المعالجة باللاكتات (الأشكال 3g و 3h)، مما يشير إلى أن الجلوكوز يُستخدم في التخليق الحيوي – في وجود اللاكتات. نظرًا لأن اللاكتات تساهم في استقلاب دورة TCA، قمنا بعد ذلك بالتحقيق في دور اللاكتات على الطاقة الحيوية لوحيدات النواة المدربة. باستخدام مجسات ATP المعتمدة على نقل الطاقة الرنانة فورستر (FRET) المستهدفة إلى الميتوكوندريا (Mito، mtAT1.03)، وجدنا أن وجود اللاكتات أوعلاج -غلوكان يعزز إنتاج ATP (الشكل 3i). مجموعة منعلاج -غلوكان واللاكتات أدى إلى تحفيز تآزري كبير لإنتاج ATP (الشكل 3i). وبالمثل، فإن وجود اللاكتات يعزز معدل استهلاك الأكسجين الأساسي (OCR) وإنتاج ATP من OxPhos (الشكل التوضيحي 3j). يتماشى معبيانات الأيض، زيادة اللاكتاتمستويات الأسيتيل-CoA، السترات، والمالات المستحثة بواسطة -غلوكان (الشكل 3j). أخيرًا، نحقق في دور الجلوكوز في عملية الأيض لدورة TCA المساهمة في اللاكتات خلالعلاج -غلوكان. كما هو موضح في الشكل 3k، فإن تركيز اللاكتات المنخفض (1 مللي مول) لا يزال يساهم في وسائط دورة TCA في الجلوكوز. ) احتوى على وسط ثقافي خلال علاج -غلوكان. عندما تكون تركيزات الجلوكوز أعلى بـ 25 مرة من مستويات اللاكتات،لم يتمكن اللاكتات من المساهمة في الكربون لدورة TCA (الشكل 3k). تشير هذه النتائج إلى أن اللاكتات هي ركيزة لدورة TCA للوحيدات المنشطة تحت الظروف الفسيولوجية، حتى عند توفر الجلوكوز.
اللاكتات يغذي المناعة المدربة عبر LDHA
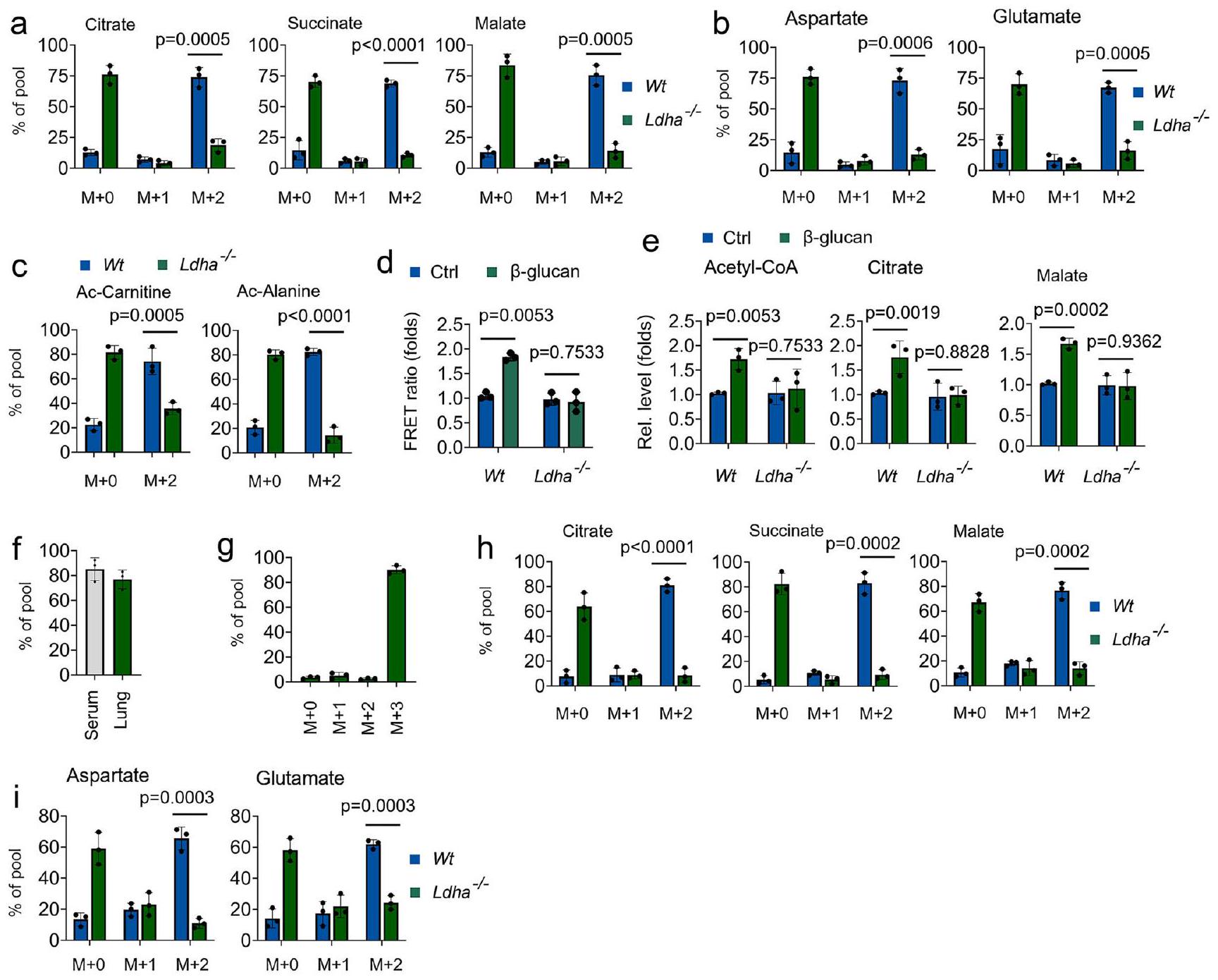
بعد ذلك، نحقق في دور LDHA في الوظيفة الحيوية لللاكتيت في المناعة المدربة. كما هو موضح في الأشكال 4a و 4b، انخفضت وفرةتمت ملاحظة السترات والسكسينات والمالات والجلوتامات والأسبارتات المشتقة من اللاكتات فيالخلايا الوحيدة استجابةً لـعلاج الجلوكاني-اللاكتات ساهم أيضًا في الميتابوليتات المرتبطة بأسيتيل-CoA، بما في ذلك أسيتيل-كارنيتين وأسيتيل-ألانين، فيمعالج بـ -غلوكانوحيدات النواة، ولكن ليس فيمعالج بـ -غلوكانالخلايا الوحيدة (الشكل 4ج). من خلال استخدام مجس ATP الميتوكوندري، علاج -غلوكان يعزز إنتاج ATP فيوحيدات النواة، ولكن ليس فيمعالج بـ -غلوكانالخلايا الوحيدة (الشكل 4د). بشكل متسق، أدى حذف LDHA إلى تثبيطمستويات الأسيتيل-CoA، السترات، الماليك المستحثة بواسطة -غلوكان (الشكل 4e). يُستخدم أكسامات الصوديوم (Oxa) على نطاق واسع لتثبيط إنتاج اللاكتات من خلال تنظيم أنشطة LDHA.. باستمرار، تم تثبيط مستويات الأسيتيل-CoA، والسترات، والمالات المستحثة بواسطة -غلوكان في وحيدات النوى المدربة المعالجة بأوكسات (الشكل التوضيحي 4a). بعد ذلك، استكشفنا ما إذا كان المضيف يستخدم اللاكتات كوقود في المناعة المدربة في الجسم الحي من خلال ضخالمستقلب. في نظامنا الحي،إثراء الموصوف بالكامل ) -لاكتات في المصل والرئة (الأشكال 4 ف و 4 ج). متسق مع بيانات خارج الجسم الحي، تم حقن-اللاكتات تم وسمه بسهولة بمستقلبات دورة TCA في الجسم الحي استجابةً لـمعالج بـ -غلوكانالفئران، ولكن ليس فيالفئران (الأشكال 4هـ و4ي). قمنا بمزيد من التحقيقتم تنظيم استقلاب الجلوكوز بواسطة -غلوكان في نظام استقلاب اللاكتات المثبط. ومن المثير للاهتمام،علاج -غلوكان يوجه تدفق الجلوكوز من PPP و SSP مرة أخرى إلى OxPhos عندما يتم تثبيط استقلاب اللاكتات (الأشكال التكميلية 4b-4f)، مما يشير إلى أن الجلوكوز هو مصدر كربوني بديل لدورة TCA في المناعة المدربة. تشير هذه النتائج إلى أن معالجة اللاكتات عبر LDHA هي نقطة التمثيل الغذائي الحرجة في المناعة المدربة.
اللاكتات ينظم أسيتيل الهستون عبر LDHA في وحيدات النواة خلال تحفيز المناعة المدربة
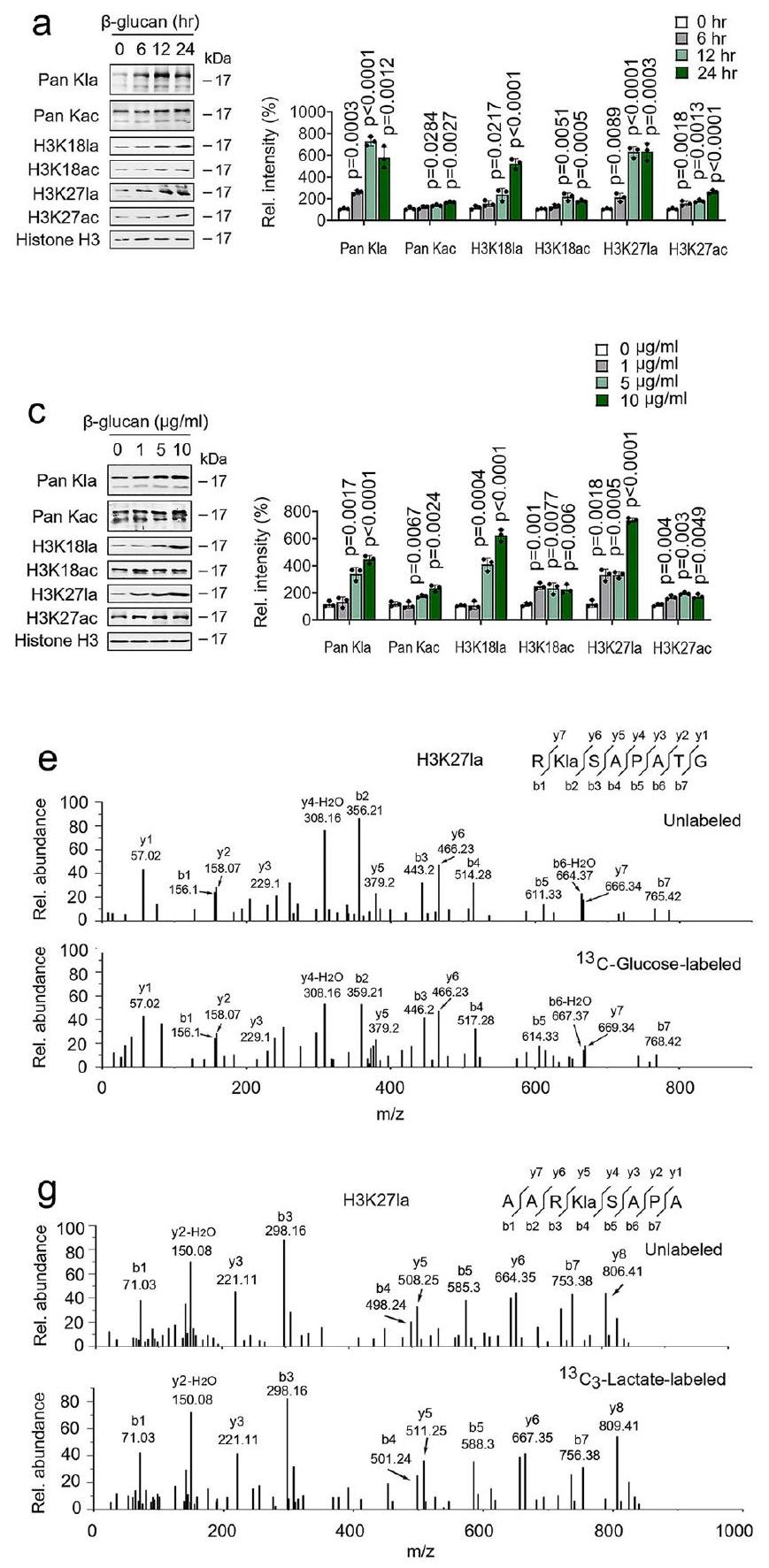
أظهرت دراسة سابقة أن تعديل اللايسين باللاكتيل (Kla) هو نوع جديد من علامات الهيستون يمكن تحفيزه بواسطة اللاكتات. (الشكل التوضيحي الإضافي 5a)؛ كنا نشك في أن-غلوكاني تم تنظيم مستوى عالٍ من اللاكتات وتأثيره على اللاكتيلation الهيستونية في المناعة المدربة. أظهرت تحليل المناعية أن مستويات الهيستون Kla قد زادت؛ ومع ذلك، كانت مستويات الأسيتيل الهيستوني (Kac) قد زادت بشكل طفيف، بطريقة تعتمد على الوقت في وحيدات النوى المعالجة بـ β-غلوكاني (الشكل 5a والشكل التوضيحي 5b). تم الكشف عن ارتفاع مستويات الهيستون Kla في وقت مبكر يصل إلى 6 ساعات بعدعلاج -غلوكان (الشكل 5أ والشكل التكميلي 5ب). من المثير للاهتمام أن مستويات الهيستون Kla العالية تم الحفاظ عليها من اليوم 1 إلى 9 (الشكل 5ب والشكل التكميلي 5ج). كما هو متوقع،تحدي -غلوكان يحفز مستويات الهيستون Kla، ويزيد بشكل طفيف من مستويات Kac، بطريقة تعتمد على الجرعة (الشكل 5c والشكل التوضيحي 5d). نظرًا لأن تجديد الخلايا بوسائط جديدة لم يؤثر على مستويات Kla، فإن زيادة هيستون Kla خلال المناعة المدربة هي داخلية وليست نتيجة لتأثيرات باراكرين (الشكل التوضيحي 5e). يتماشى ذلك مع النتائج السابقة.أظهر تحليل MS/MS أن هناك تغييرًا في الكتلة قدره 72.021 دالتون على بقايا الليسين في الهيستون تم ملاحظته فيالمونوسيتات المعالجة بـ -غلوكان (الأشكال التكميلية 5f و5g). في هذا السياق، تجارب الوسم الأيضي باستخدام-جلوكوز أوتمت متابعة تحليل MS/MS بواسطة -لاكتات مما أظهر أن-غلوكان يعزز اللاكتيل من الهيستون مباشرة من الجلوكوز أو اللاكتات الخارجية (الأشكال 5د-5ز). بالإضافة إلى ذلك، زادت معالجة β-غلوكان من مستويات هيستون Kla فيوحيدات النواة ولكن ليس فيالخلايا الوحيدة (الشكل 5h والشكل التوضيحي 5h). تشير هذه النتائج إلى مستويات عالية من اللاكتات داخل الخلايا في تنظيم المناعة المدربة لأسيتيل الهيستون.
يحافظ اللاكتات على إمكانية الوصول إلى الكروماتين من خلال أسيتيل اللاكتيل في الخلايا الوحيدة خلال تحفيز المناعة المدربة
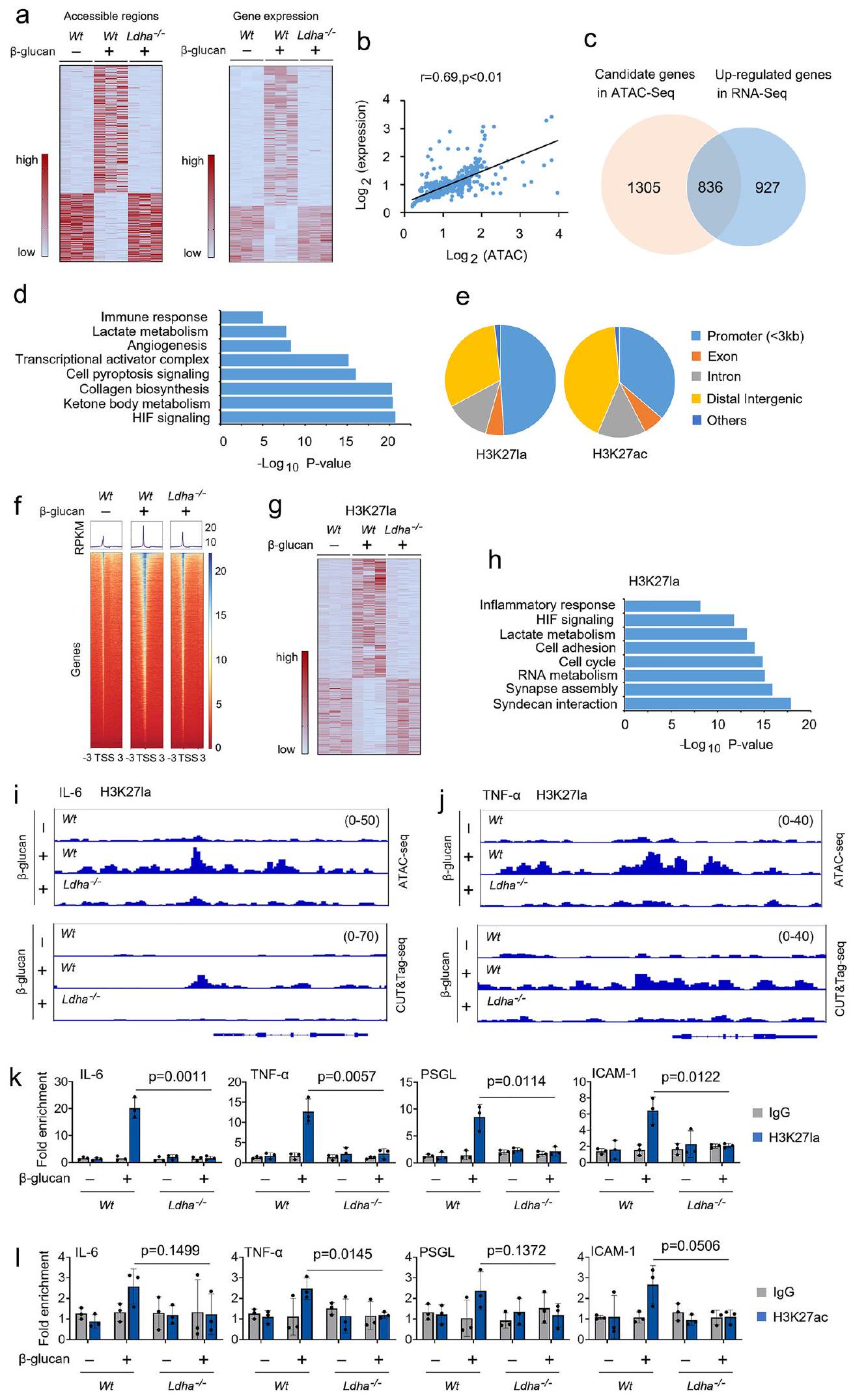
للتحقيق في التأثير الميكانيكي لإنتاج اللاكتات على المناعة المدربة، قمنا بإجراء تحليل اختبار الوصول إلى الكروماتين بواسطة ترانسبوزاز مع تسلسل عالي الإنتاجية (ATAC-seq) لمقارنة التغيرات في وصول مناطق الكروماتين بينوحيدات النواة ، و المونوسيتات المدربة. حددنا 2,848 منطقة كروماتينية تظهر تغييرات في الوصول (الشكل 6أ). لتقييم التأثير الوظيفي لللاكتيت على النسخ، قمنا بإجراء تسلسل RNA فيوحيدات النواة،، و المونوسيتات المدربة في النسخ البيولوجية المكررة (الشكل 6أ). استكشفنا أولاً العلاقة بين التغيرات في وصول الكروماتين والتعبير الجيني. قمنا بتعيين كل المناطق القابلة للوصول بشكل مختلف (DARs) إلى أقرب موقع لبدء النسخ وفحصنا الفروق في تعبير الجينات. لاحظنا ارتباطات كبيرة بين التغيرات في وصول الكروماتين وتعبير الجين المجاور (الشكل 6b). بعد ذلك، حددنا 836 جينًا تم تنظيمها بشكل مرتفع فيوحيدات النواة،، و وحيدات النواة المدربة، التي تم تمييزها أيضًا بالكروماتين
الشكل 3 | اللاكتات هو وقود فسيولوجي للمناعة المدربة. أ-و تم زراعة وحيدات النوى في وسط MM يحتوي على-جلوكوز (5 مللي مولار)، مما يدل على تركيز اللاكتات غير المعلم، وتم معالجته بـ-غلوكان ( ) لمدة 4 ساعات، تليها تحليلات الطيف الكتلي. ج الرسم التخطيطي لاشتقاق ومساهمة ذرات الكربون في تخليق SAM.تم إجراء التجارب كما هو موضح في (أ).تم توضيح تحويل -الجلوكوز إلى ATP أو SAM. تم معالجة وحيدات النوى بـ-غلوكان ولاكتاتلمدة 30 دقيقة؛ تم قياس ATP الميتوكوندري بواسطة mtAT 1.03 استقصاء.تم معالجة وحيدات النوى مع أو بدون-غلوكان ولاكتاتلمدة 12 ساعة، ثم تم قياس مستويات الأسيتيل-CoA، والسترات، والمالات.تم زراعة وحيدات النوى في وسط MM يحتوي على-لاكتات (1 مللي مول)، مما يدل على تركيز الجلوكوز غير المعلم، وتمت معالجته بـ-غلوكان ( ) لمدة 4 ساعات، تليها تحليلات الطيف الكتلي. البيانات في ( و ) يتم التعبير عنها كمتوسطات ± الانحراف المعياري للخطأ، الفئران لكل حالة، تحليل التباين ثنائي الاتجاه. انظر أيضًا الأشكال التكميلية 3.
الشكل 4 | يؤثر Ldha على الأيض ووظيفة المناعة المدربة. أ-ج تم زراعة وحيدات النوى من فئران Wt وLdha// في وسط MM يحتوي على-لاكتاتوعولجت بـ-غلوكانلمدة 12 ساعة، تليها تحليلات مطيافية الكتلة. د الخلايا الوحيدة من و تم زراعة الفئران في وسط يحتوي على اللاكتات (0.5 مللي مول من الجلوكوز، 0.5 مللي مول من الجلوتامين، و2 مللي مول من اللاكتات)، وتم معالجتها بـغلوكوزان لمدة 30 دقيقة، تم قياس ATP الميتوكوندري بواسطة مجس mtAT 1.03. الخلايا الوحيدة من و تم معالجة الفئران مع أو بدون-غلوكان و اللاكتيت (1 مللي مول) لمدة 12 ساعة، تليها قياس مستويات الأسيتيل-CoA، والسترات، والمالات. ف، ج تم تدريب فئران C57BL/6 مع أو بدون-غلوكان. بعد ستة أيام، تم حقن الفئران بـ-لاكتات لمدة 12 ساعة، تليها تحليلات الطيف الكتلي. ح، ط و تم تدريب الفئران مع أو بدون-غلوكان. بعد ستة أيام، تم حقن الفئران بـ-لاكتات لمدة 12 ساعة؛ تم عزل وحيدات النوى وتبعتها تحليلات الطيف الكتلي. جميع البيانات معبّر عنها كمتوسطات ± الانحراف المعياري للخطأ،فئران لكل حالة، تحليل التباين ثنائي الاتجاه. انظر أيضًا الشكل التكميلية 4. تعديلات الوصول (الشكل 6ج). أظهر تحليل عملية البيولوجيا لجينات مصفوفة الجينات (GO) غنىً كبيرًا للمسارات البيولوجية، بما في ذلك استجابة المناعة، واستقلاب اللاكتات، وإشارات HIF (الشكل 6د). اخترنا تعديل الهيستون H3 ليسين 27 (H3K27la)، حيث يتم أيضًا أسيتيل هذا المتبقي من الليسين (H3K27ac)، لتوضيح الأهمية الوظيفية المحتملة لأسيتيل الهيستون في المناعة المدربة باستخدام CUT&Tag، وهو اختبار جديد لتثبيت المناعة على مستوى الجينوم. يكشف تحليل توزيع الجينوم أن تعديلات الهيستون H3K27la وH3K27ac كانت موجودة بشكل رئيسي ضمن مناطق المحفزات (الشكل 6هـ). كما هو متوقع، زادت قمم H3K27la بالقرب من مواقع بدء النسخ (TSSs) في-المونوسيتات المدربة، ولكن ليسالمونوسيتات المدربة (الشكل 6f). ومع ذلك، كانت قمم H3K27ac مرتفعة قليلاً بالقرب من مواقع بدء النسخ (TSSs) فيالخلايا الوحيدة المدربة تقارن معالخلايا الوحيدة غير المدربة (الشكل التوضيحي 6أ). قمنا بمقارنة تعبير الجينات المعلمة بـ H3K27la فيوحيدات غير مدربة و الخلايا الوحيدة المدربة. من بين إجمالي 7,547 قمة مشتركة بين المجموعات الثلاث، حصلنا على 248 قمة تغيرت بشكل ملحوظ (الشكل 6g). أظهر تحليل مسار GO أن العمليات البيولوجية مثل الاستجابة الالتهابية، واستقلاب اللاكتات، والتصاق الخلايا، تتوافق مع بيانات ATAC-seq (الشكل 6h). إلى
شكل.-غلوكوزان ينظم اللاكتيل في الهيستون عبر Ldha. أ، ب تم معالجة وحيدات النواة مع أو بدون-غلوكانللفترة المحددة قبل التحليل الغربي. يتم عرض قياس المناعية في اللوحة اليمنى. تم معالجة وحيدات النوى مع أو بدون-غلوكان بتركيزات محددة لمدة 24 ساعة قبل إجراء التحليل الغربي. يتم عرض تقدير المناعية في اللوحة اليمنى. د، هـ طيف MS/MS لـ-ببتيد هيستون كلاس المسمى بالجلوكوز ونظيره غير المسمى منوحيدات النواة المعالجة بـ -غلوكان. ف،كانت التجارب
تم التنفيذ كما هو موصوف في (د، هـ)، باستثناءتم استخدام -لاكتات.الخلايا الوحيدة من Wt وتم معالجة الفئران مع أو بدون-غلوكان لمدة 24 ساعة قبل إجراء التحليل الغربي. يتم عرض قياس المناعية في اللوحة اليمنى. تم تكرار جميع التجارب ثلاث مرات على الأقل. البيانات في (أ-ج و ح) معبّرة كمتوسطات SEMs، فئران لكل حالة، تحليل التباين ثنائي الاتجاه. انظر أيضًا الشكل التوضيحي التكميلي 5. اختبرنا دور H3K27la و H3K27ac في وظيفة المونوسيت المدرب، قمنا بدمج مجموعات بيانات ATAC-seq ومجموعات بيانات CUT&Tag في عارض الجينوم التكاملي لفحص ملفات ارتباطها على الجينات. كما هو موضح في الشكل التكميلية 6b، كانت قمم H3K27ac معززة قليلاً عند المحفزات الخاصة بـ IL-6 و TNF-في وحيدات النوى المدربة مقارنة بـالمونوسيتات غير المدربة. ومع ذلك، كانت قمم H3K27la غنية للغاية عند المحفزات للجينات الحرجة لعوامل الالتهاب وجزيئات التصاق الخلايا، بما في ذلك
IL-6، TNF-PSGL و ICAM-1، في-المونوسيتات المدربة، ولكن ليس فيتم تدريب وحيدات النوى (الأشكال 6i، 6j والأشكال التكميلية 6c و6d). ثم استخدمنا اختبار ترسيب المناعة للكروماتين (ChIP)-qPCR لتقييم تأثيرات H 3 K 27 la وH 3 K 27 ac على التعبير عن جينات المؤثر المستقرة. في الواقع، تم تجنيد H3K27la إلى مناطق المحفز لجينات IL-6 وTNF-، PSGL، و ICAM-1 زادت في-المونوسيتات المدربة ولكن ليس فيالخلايا الوحيدة المدربة (الشكل 6k). كانت هناك اختلافات طفيفة فقط بين من H3K27ac التوظيف في مناطق المحفز لجين IL-6 و TNF-PSGL و ICAM-1 فيالمونوسيتات المدربة تقارن معالمونوسيتات غير المدربة (الشكل 61). دور تعديلات الهيستون على تنشيط المحفز ليس مقصورًا على ليسين واحد، حيث تم الحصول على نتائج مماثلة بواسطة H3K18la وH3K18ac (الأشكال التكميلية 6e و6f). تماشيًا مع هذه الملاحظات، فإن حجب هيستون CBP/p300 نشاط الأستيل ترانسفيراز (HAT) مع مثبط HAT (HATi) قلل أيضًا من تفاعل H3K27la وH3K18la مع المحفز لـ IL-6 وTNF-تشير هذه النتائج إلى أن إنتاج اللاكتات الداخلي يغير البرمجة الجينية المعتمدة على الأسيتيل عند المحفز لعوامل الالتهاب وجزيئات التصاق الخلايا في المونوسيتات المدربة.
الشكل 6 | ينظم اللاكتات تعديل الهيستون باللاكتيل عبر Ldha في وحيدات النواة خلال تحفيز المناعة المدربة. أ خريطة حرارية تمثل المناطق القابلة للوصول بشكل مختلف (DARs) من ATAC-seq (اللوحة اليسرى) والتعبير الجيني المختلف من RNA-seq (اللوحة اليمنى) في وحيدات النواة منوحيدات النواة،وحيدات النواة المدربة، وتم تدريب وحيدات النوى كما هو موضح في الشكل S1b. ب DARs من ATAC-seq والتعبير الجيني التفاضلي من RNA-seq خضعت لتحليل ارتباط بيرسون. معامل الارتباط ( ) و تم حساب القيم بواسطة تحليل الارتباط لبيرسون. ج. مخطط فين يوضح عدد الجينات المرشحة في ATAC-seq والجينات المرتفعة التعبير في RNA-seq لـوحيدات النواة،وحيدات النواة المدربة، والمونوسيتات المدربة. تحليل GO للجينات المختلفة بشكل ملحوظ في (ج). يتم عرض أهم العمليات البيولوجية غير المتكررة، مع أعداد الجينات المعنية.القيم.القيم بواسطة اختبار t لطلاب ذو طرفين. الرسم البياني الدائري الذي يوضح توزيع H 3 K 27 la و H 3 K 27 ac في المناطق الجينومية المعلّمة في وحيدات النوى المدربة. خريطة الحرارة التي توضح التواجد الجينومي لـ
H3K27laتسلسلات جينية محيطة بمواقع بدء النسخ في أحادية النواة المدربة من و الفئران. الجينات المعروضة في الصفوف مرتبة بترتيب تنازلي حسب قوة الإشارة. خريطة حرارية لوصول قمة الكروماتين للقمم المختلفة بشكل ملحوظ في وحيدات النواة المدربة من و الفئران. كل صف يمثل درجة Z لعدد القراءات المعادلة المنقولة باستخدام log2 ضمن كل عينة باستخدام CUT&Tag-seq. تحليل GO للجينات المختلفة بشكل ملحوظ في (g). أكثر العمليات البيولوجية دلالة وغير المتكررة مع أرقام الجينات المعنية وتظهر القيم.قيم اختبار t لطلاب ذو طرفين. i، j مسارات متصفح الجينوم التمثيلية لـ ATAC-seq و CUT&Tag-seq لتراكم قمة H3K27la عند مواقع جينات IL-6 و TNF-a لعدلات الدم المدربة من و فئران. ك،الخلايا الوحيدة من و تم تدريب الفئران كما هو موضح في الشكل S1b، تلا ذلك تحليل ChIP. البيانات في ( و ) يتم التعبير عنها كوسائل SEMs، الفئران لكل حالة، تحليل التباين ثنائي الاتجاه. انظر أيضًا الشكل التوضيحي التكميلي 6.
اللاكتات الخارجي يدعم المناعة المدربة من خلال المساهمة في دورة TCA وخلط الهيستون باللاكتات
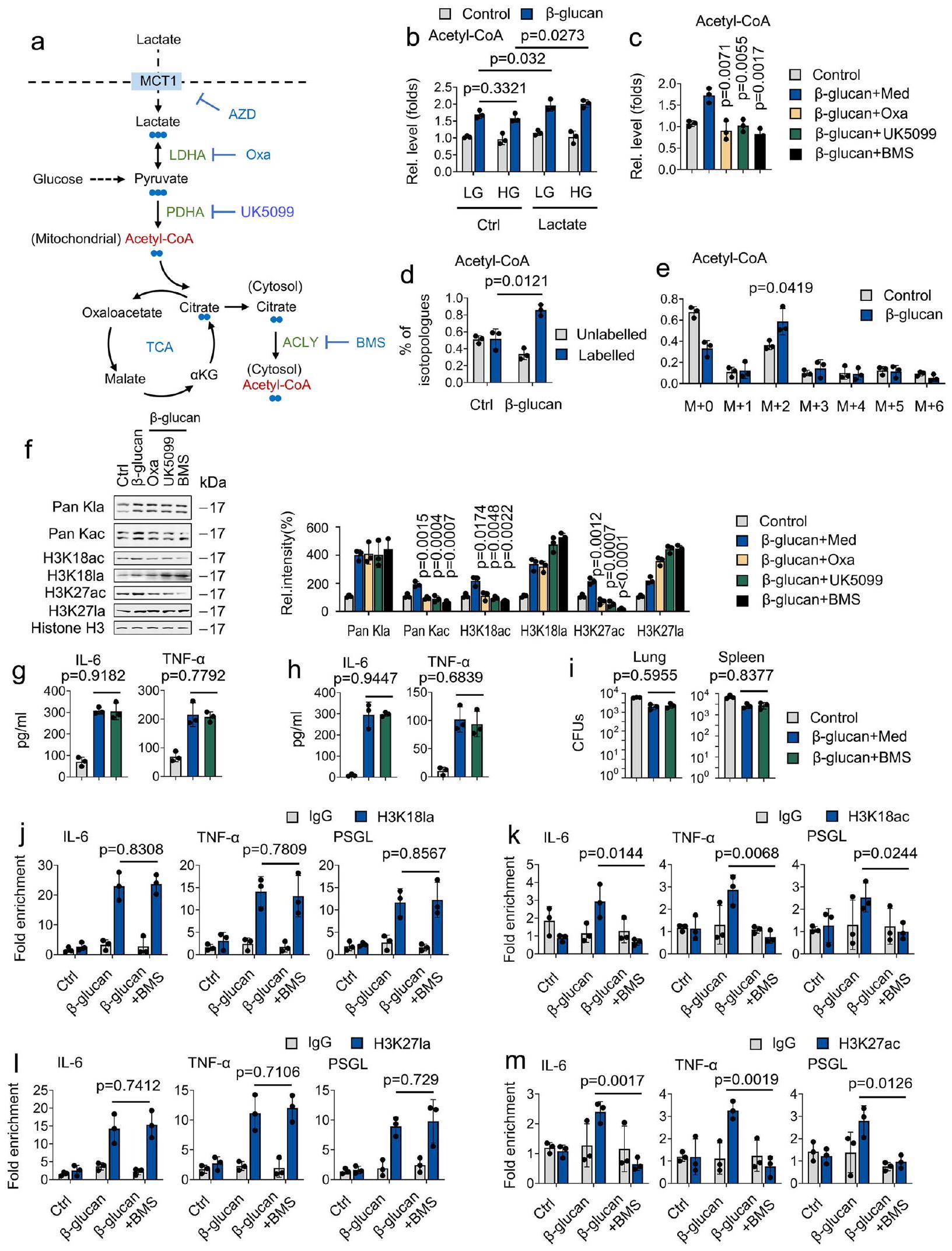
يمكن إنتاج اللاكتات داخل الخلايا بواسطة LDHA أو استيرادها بواسطة الناقل أحادي الكربوكسيلات 1 (MCT1؛ المعروف أيضًا باسم SLC16A1). لذلك، سعينا لاستكشاف تأثير MCT1 على المناعة المدربة. كما هو موضح في الشكل التوضيحي 7a، IL-6 و TNF-تم تحفيز الإنتاج فيمدرب على -غلوكانالفئران، ولكن ليس في الفئران المعالجة بمثبط MCT1 AZD3965 (AZD). في هذا السياق، كانت الفئران المعالجة بـ AZD أكثر عرضة للفتك الناتج عن C. albicans، مع زيادة الحمل البكتيري في أنسجة الرئة والكلى (الأشكال التكميلية 7b و7c). بعد ذلك، نقوم بتقييم دور MCT1 في دورة TCA المدفوعة باللاكتات من خلال معالجة وحيدات النوى المدربة بـ-لاكتات. كما هو موضح في الأشكال التكميلية 7d-7f، ألغى علاج AZD دخول اللاكتات إلى دورة TCA. كما هو متوقع،علاج -غلوكان زاد من مستويات هيستون كلاس فيالمونوسيتات ولكن ليس في المونوسيتات المعالجة بـ AZD (الشكل التوضيحي 7g). أظهرت تجربة ChIP-qPCR أن معالجة AZD تمنع تجنيد H3K27la وH3K18la إلى مناطق المحفزات لـ IL-6 وTNF-، PSGL، و ICAM-1 (الأشكال التكميلية 7h و 7i). وبالتالي، فإن امتصاص اللاكتات بواسطة MCT1 يعمل كركيزة لدورة TCA وينظم إنتاج السيتوكينات عبر استيلاء الهيستون على اللاكتات.
بياناتنا كشفت أن-غلوكاني تسبب بشكل خفيف في تحفيز أسيتيل الهستون. افترضنا أن اللاكتات يتم امتصاصها وتحويلها بسهولة إلى حمض البيروفيك من خلال LDHA، ومن ثم يتم تحويلها إلى أسيتيل-CoA سيتوزولي في تفاعلين محفزين إنزيمياً بواسطة ديهيدروجيناز البيروفات (PDHA) و ATP-سيترات لاييز (ACLY) على التوالي. (الشكل 7أ). لهذا الغرض؛ نقارن دور الجلوكوز واللاكتات في توليد الأسيتيل-CoA. كما هو موضح في الشكل 7ب، لا يؤثر كل من الجلوكوز العالي (HG) أو الجلوكوز المنخفض (LG) على توليد الأسيتيل-CoA المنظم بواسطة -غلوكان- ومع ذلك، فإن معالجة اللاكتات أدت إلىأسيتيل منظم بواسطة -غلوكانالجيل، مما يشير إلى أن-غلوكان يعزز توليد الأسيتيل-CoA عبر اللاكتات. ومع ذلك، ليس الجلوكوز (الشكل 7ب). لتفكيك الخطوة الرئيسية في استقلاب اللاكتات المعنية في توليد الأسيتيل-CoA، استخدمنا سلسلة من الأدوية المستهدفة LDHA وPDHA وACLY بواسطة مثبط LDHA (أوكزامات الصوديوم، Oxa)، ومثبط ناقل البيروفات (UK5099)، ومثبط ACLY (BMS303141، BMS)، على التوالي (الشكل 7أ). من المثير للاهتمام، أن معالجة Oxa وUK5099 وBMS تمنعتوليد الأسيتيل-CoA الناتج عن الجلوكاني (الشكل 7c). تحليل LC/MS الذي يركز على النظائر المتماثلة للأسيتيل-CoA يظهر أن كربونات اللاكتات وسمت الغالبية العظمى من الأسيتيل-CoA.كشف نظير الأسيتيل-CoA عن نمط وسم ملحوظ ناتج عن كربونات اللاكتات (الأشكال 7d و 7e). لتحديد المسار الذي يسهل من خلاله اللاكتات أسيتيلation الهيستون، افترضنا أن هذا ينطوي على الأيض المؤكسد، وخاصة ACLY. ومن المثير للاهتمام، أن BMS عارضت-تحفيز الأسيتيل للهيستون بواسطة -غلوكان ولكن ليس اللاكتيل للهيستون (الشكل 7f). من المدهش أن مثبط ACLY لم يؤثر علىIL-6 و TNF- المستندة إلى -غلوكانالإنتاج، والحمل البكتيري في أنسجة الرئة والكلى في المختبر وفي الجسم الحي (الأشكال 7g-7i). بالإضافة إلى ذلك، فإن علاج BMS يثبط H3K27ac وH3K18ac، ولكن ليس H3K27la وH3K18la، من الانضمام إلى مناطق المحفز لـ IL-6 وTNF-“، و PSGL (الأشكال.تشير هذه النتائج إلى أن اللاكتات هي ركيزة هامة لإنتاج الأسيتيل-CoA. بالمقارنة مع الأسيتيل الحمض النووي الهيستوني الذي يتم بوساطة اللاكتات، تلعب اللاكتات-mediated هيستون لاكتيلات دورًا رئيسيًا في المناعة المدربة.
نقاش
المناعة المدربة تشير إلى الصفات الذاكرية طويلة الأمد للمناعة الفطرية.كيف يؤثر توفر العناصر الغذائية البيئية على المناعة المدربة لا يزال بحاجة إلى فهم أفضل. هنا، نعرض للمرة الأولى أن وحيدات النوى المدربة تظهر درجة عالية من المرونة في اختيار الوقود، ليس فقط الوقود التقليدي مثل الجلوكوز ولكن أيضًا PCSs. من خلال استكشاف ذلك، أظهرنا أن اللاكتات الداخلية للخلايا هي وقود بارز في وحيدات النوى المدربة، مما يساهم في دورة TCA، حتى عندما يكون الجلوكوز متاحًا. من الناحية الآلية، كانت اللاكتات مادة مهمة لإنتاج أسيتيل-CoA في وحيدات النوى المدربة وتنظمت وظيفة وحيدات النوى المدربة من خلال تأثيراتها على اللاكتيل في الهيستونات. تبرز دراستنا دورًا لم يُقدَّر سابقًا للاكتات في المناعة المدربة.
لقد تم نسب الذاكرة المناعية لفترة طويلة فقط إلى خلايا T التكيفية و خلايا المفهوم الذي ظهر مؤخرًا للمناعة المدربة يُعرف بأنه يسبب تغييرات طويلة الأمد في خلايا المناعة الفطرية، مما يمكّن استجابة مضيف فطرية قوية لتحفيز ثانوي.أظهر تحليل المسارات المنظمة بشكل مختلف في وحيدات النوى المدربة دورًا أساسيًا في التغيرات في الأيض داخل الخلايا، خاصةً في أيض الجلوكوز.المTOR-HIF1المسار المنظم أدى إلى تحول من الفسفرة التأكسدية إلى التحلل السكري، وهو الآلية التنظيمية المركزية للمناعة المدربة. هذه الدراسة رصدت مستويات عالية من اللاكتات، وهو منتج نهائي من التحلل السكري، في المناعة المدربة، لكن الأهمية الفسيولوجية للاكتات المفرط لم يتم استكشافها. نحن نوضح أن اللاكتات هي وقود فسيولوجي للمناعة المدربة، حيث تعمل كركيزة مفضلة لدورة TCA والبناء الحيوي. بالإضافة إلى ذلك، تنظم اللاكتات إنتاج السيتوكينات من خلال تأثيراتها على أسيتلة الهيستون باللاكتات. نتائجنا تكمل بشكل قوي العمل السابق، مما يملأ مسارات الإشارات اللاحقة للمناعة المدربة. نقترح نموذج عمل لأبرز المسارات المنظمة في المناعة المدربة بناءً على الدراسات السابقة وبياناتنا الحالية. وفقًا لهذا النموذج، تقوم خلايا المناعة الفطرية المدربة بتنشيط مسار Akt-mTOR-HIF-1.مسار، يؤدي إلى تحول في الأيض من الفسفرة التأكسدية إلى التحلل السكري وتراكم عالي لللاكتات. جزء من مستويات اللاكتات المرتفعة يعمل كوقود فسيولوجي لدورة TCA. الجزء الآخر من مستويات اللاكتات المرتفعة يدخل النواة ويرتبط بمواقع محفزات السيتوكينات، مما يؤدي إلى الحفاظ على إمكانية الوصول إلى الكروماتين. إذا واجهت خلايا المناعة الفطرية المدربة محفزًا ثانويًا، فإن الكروماتين القابل للوصول يسمح لعوامل النسخ بالارتباط بسرعة مع المحفز المقابل، مما يؤدي إلى استجابة أسرع وأكثر قوة.
مؤخراً، كشفت بعض الأبحاث عن العلاقة بين تحفيز مسارات المناعة الفطرية واستحثاث التغيرات الوراثية والتمثيلية في خلايا المناعة المدربة.روب وآخرون يظهرون أن التحلل الجلوتاميني هو مسار أيضي آخر متورط فيالمناعة المدربة المستحثة بواسطة -غلوكان، المرتبطة بالزيادة في H3K4me3، والتي هي أساسي للمناعة المدربة. ومع ذلك، لم يكن هناك فرق ملحوظ في تعبير الميثيل ترانسفيراز بين الخلايا المناعية غير المدربة والمدربة. يظهر سيران وآخرون أن مسار تخليق الكوليسترول/الميفالونات مرتفع فيفئران مدربة على -غلوكان، والتي ترتبط بالثراء في. ومع ذلك، فإن دراسة الجينوم الكامل H3K27ac لا تكشف عن تداخل كامل بين – المناعة المدربة الناتجة عن الجلوكاني والميفالونات. نشك في أن جزءًا من هذه الظاهرة يعود إلى وسط زراعة الخلايا. تم تصميم وسط زراعة الخلايا الحالي لتعزيز نمو الخلايا وبقائها. وبالتالي، يحتوي وسط زراعة الخلايا الحالي على تركيزات فوق الفسيولوجية لبعض العناصر الغذائية بينما يفتقر إلى عناصر غذائية أخرى ذات صلة بتمثيل الخلايا. إحدى النتائج المهمة لهذه الدراسة هي أن
الشكل 7 | اللاكتات الخارجية تدعم المناعة المدربة من خلال المساهمة في دورة TCA وتعديل الهيستون باللاكتيل. أ رسم تخطيطي لـتوسيم الكربون -لاكتات من خلال دورة TCA. تم زراعة وحيدات النوى مع جلوكوز عالي (HG) (15 مللي مول)، جلوكوز منخفض (LG) (3 مللي مول)، و/أو لاكتات (1 مللي مول)، وتم علاجها مع أو بدونغلوكانلمدة 24 ساعة، تليها قياس مستويات الأسيتيل-CoA. تم زراعة الكريات الوحيدة في وسط يحتوي على اللاكتات (0.5 مللي مول من الجلوكوز، 0.5 مللي مول من الجلوتامين، و2 مللي مول من اللاكتات) وتم معالجتها مع أو بدون-غلوكان ( )، أكسامات الصوديوم ( 20 مللي مول )، UK5099 ب.م.س 303141 (ب.م.س)لمدة 24 ساعة، تليها قياس مستويات الأسيتيل-CoA. د، هـ تم زراعة وحيدات النواة في وسط MM يحتوي على-لاكتاتوعولجت بـ-غلوكانلمدة 4 ساعات، تلتها تحليل LC/MS. تظهر الفقرات المعلمة وغير المعلمة من الأسيتيل-CoA. تم زراعة وحيدات النوى في وسط يحتوي على اللاكتات (0.5 مللي مول من الجلوكوز، 0.5 مللي مول من الجلوتامين، و2 مللي مول من اللاكتات) و معالج أو بدون-غلوكانأوكسامات الصوديومالمملكة المتحدة 5099نظام إدارة البطارية لمدة 24 ساعة قبل إجراء التحليل الغربي. يتم عرض قياس المناعية في اللوحة اليمنى. تم تكرار التجارب ثلاث مرات على الأقل. تم زراعة الكريات البيضاء الأحادية في وسط يحتوي على اللاكتات (0.5 مللي مول من الجلوكوز، 0.5 مللي مول من الجلوتامين، و2 مللي مول من اللاكتات)، وتم معالجتها مع أو بدون -غلوكان ( ) ، نظام إدارة المباني ( ) لمدة 24 ساعة، ثم IL-6 و TNF-تم قياس المستويات. ح، طتم تدريب الفئران مع أو بدون-غلوكان. بعد ستة أيام، تم إصابة الفئران بجرعة قاتلة ثانوية من C. albicans أو بدونها ( ) و/أو BMS ( ) لمدة 24 ساعة، تليها قياس IL-6 و TNF- المستويات ( ) وحمل الفطريات (ي). الخلايا الوحيدة منتمت معالجة الفئران كما هو موضح في (ج)، تلاها تحليل ChIP. تُعبر البيانات عن المتوسطات ± الانحراف المعياري.الفئران لكل حالة، تحليل التباين الثنائي الاتجاه. انظر أيضًا الشكل التوضيحي 7. استخدام الجلوكوز بواسطة المونوسيتات المدربة يتأثر بشكل كبير بالركائز غير الجلوكوزية الأخرى، وبالتحديد بعض العناصر الغذائية الغائبة عن معظم تركيبات الوسط اليوم. أظهرت دراستنا أن اللاكتات هي مصدر رئيسي لإنتاج الأسيتيل-CoA في المونوسيتات المدربة. وهذا يساهم مباشرة في اللاكتيلation الهيستونية ويعزز اللاكتيلation المسموح بها في الهيستونات عند مواقع جينات السيتوكينات. من الناحية الآلية، نوضح أن LDHA، الذي يتحكم في استقلاب اللاكتات من خلال تنظيم تحويل اللاكتات/البيروفات، مطلوب لدورة TCA المدفوعة باللاكتات وتنظيم اللاكتيلation الهيستونية. توضح دراستنا أن اللاكتات هي المحور بين الاستقلاب والمناعة وعلم الوراثة اللاجينية في المونوسيتات المدربة.
لم يغفل انتباهنا بعد أن هناك عدة تحذيرات مهمة. (ط) لا تزال الفروق والروابط بين اللاكتيل في الهيستون، والميثيل في الهيستون، واللاكتيل في الهيستون في المناعة المدربة بحاجة إلى الاستكشاف. (2) نحن نركز فقط على دور التركيزات العالية (التي تتجاوز ) PCS على استخدام الجلوكوز في المناعة المدربة. قد تعمل تركيزات PCS المنخفضة أيضًا كوقود حيوي في خلايا المناعة المدربة عندما تكون المغذيات غير متاحة. (iii) لم نستكشف ما إذا كان استقلاب اللاكتات هدفًا فعالًا في العلاج السريري. (iv) من الصعب إثبات التأثير السببي المباشر للاكتيل على المناعة المدربة، حيث إن مثبطات p300 تمنع أيضًا أسيتيل الهستون. (v) يؤثر نقص LDHA بشكل كبير على البقاء في هذا النموذج، بغض النظر عنعلاج -غلوكاني (الشكل 2e). من المحتمل أن ينشأ هذا من الدور الحاسم لـ LDHA في دعم الاستجابات الجليكولية للعدوى في جميع خلايا المناعة الفطرية (في غياب تدريب المناعة). (vi) مقارنة وصول الهيستون بين خلايا WT و LDHA لاستنتاج التغيرات التي يسببها اللاكتات (دون ضوابط معالجة اللاكتات) تمثل تحديًا، لأن فقدان LDHA سيغير بشكل كبير الحالة الأيضية والوظيفية لهذه الخلايا (على سبيل المثال من خلال تقييد تدفق الجليكوليز و NAD/H الأكسدة والاختزال) بالإضافة إلى دوره في السماح بتوليد اللاكتات واستخدامها لاحقًا. عند النظر في الخطوة التالية، ستساعد الدراسات التي تستكشف هذه الأسئلة بشكل كبير في توضيح دور اللاكتات في المناعة المدربة.
طرق
بيان الأخلاقيات
تماشيًا مع إعلان هلسنكي، وافق مجلس المراجعة المؤسسية في جامعة ووهان على جمع العينات السريرية وفقًا للإرشادات لحماية المشاركين البشريين. قدم جميع المشاركين في الدراسة موافقة خطية مستنيرة لجمع العينات والتحليلات اللاحقة.
تبع جميع تجارب الحيوانات دليل المعاهد الوطنية للصحة لرعاية واستخدام الحيوانات المخبرية. تم اعتماد البروتوكولات والإجراءات من قبل لجنة رعاية واستخدام الحيوانات المؤسسية في جامعة ووهان (ترخيص المشروع WDSKY0201302).
عزل الخلايا الوحيدة والنوى الوحيدة في الدم المحيطي
تم عزل الخلايا الوحيدة النواة في الدم المحيطي (PBMCs) من دم المتبرعين الأصحاء بواسطة الطرد المركزي القياسي باستخدام تدرج فيكول وزرعها في RPMI 1640 بدون مصل جنيني بقر ومضادات حيوية. تم إجراء عزل وحيدات النوى باستخدام بيركول (SigmaAldrich) كما هو موصوف سابقًا.باختصار،تم وضع خلايا PBMCs فوق محلول شخصي مفرط التوتربيركول0.16 م NaCl) وتم الطرد المركزي لمدة 15 دقيقة عندتم عزل طبقة التداخل، وتم غسل الخلايا بمحلول فوسفات البفر البارد (PBS).
تحليل البقعة الغربية
تم إجراء تحليلات Western blot كما هو موصوف سابقًاباختصار، تم جمع الخلايا بواسطة الطرد المركزي منخفض السرعة وغسلها بمحلول PBS. تم تحلل الخلايا في محلول RIPA (تكنولوجيا الإشارات الخلوية، بوسطن، ماساتشوستس، الولايات المتحدة)، وتم تحديد تركيزات البروتين باستخدام اختبارات BCA. تم فصل أربعين ميكروغرامًا من كل عينة بروتين باستخدام SDS-PAGE بنسبة 12% ونقلها إلى أغشية نيتروسليلوز (بايو راد). تم حجب الأغشية بـمحلول ملحي مخفف بفر تريس مع توين 20 (TBST) وحليب خالي من الدسم لمدة ساعة واحدة في درجة حرارة الغرفة. ثم تم تحضين الأغشية مع الأجسام المضادة الأولية طوال الليل في. بعد ذلك، تم تحضين البقع مع أجسام مضادة ثانوية مرتبطة ببيروكسيداز الفجل (جاكسون إيمونوريسيرش) لمدة ساعة إضافية. تم تصور الأشرطة المناعية باستخدام نظام كيمياء مضيئة معزز (GE Healthcare).
قياسات السيتوكينات
تم تحديد السيتوكينات في سوائل زراعة PBMC بواسطة اختبار Procartaplex المتعدد (Thermo Fisher Scientific، الولايات المتحدة الأمريكية) وتم الحصول عليها على نظام Luminex 200 (Thermo Fisher Scientific، الولايات المتحدة الأمريكية) وفقًا لبروتوكول الشركة المصنعة.
تحليل تسلسل المناعة الكروماتينية (ChIP-seq)
تم ربط العينات بشكل متقاطع في التعليق لمدة 5 دقائق في 10 مل منالفورمالديهايد (بيرس، #28906) في PBS عند درجة حرارة الغرفة. تم فصل النوى باستخدام مجموعة عزل نواة الخلية (ثيرمو فيشر ساينتيفيك، #78835). تم تخفيف المستخلصات إلى 500 مل عن طريق إضافة محلول الربط (25 مليمول HEPES pH 7.5،تم إضافة RNAse A/T1 (ثيرمو فيشر ساينتيفيك، #EN0551)، وتم تحضين العينة في لمدة 25 دقيقة. ثم، تمت إضافته إلى تركيز نهائي قدره 40 مللي مول، تلاه 75 وحدة من نوكلياز الميكروكوكال (وورثينجتون للكيماويات الحيوية)، وتم حضنه في لمدة 5 دقائق. تم إيقاف نشاط MNase بإضافة 40 مللي مول من EDTA، وتم رفع الحجم الكلي إلى 1.2 مل مع محلول الربط. بعد ذلك، تم إزالة المواد غير القابلة للذوبان عن طريق الطرد المركزي بأقصى سرعة ( ) في لمدة 5 دقائق، وتم جمع السائل العلوي الذي يحتوي على الكروماتين القابل للذوبان. في هذه المرحلة، تم قياس 5 مل من الكروماتين باستخدام مجموعة اختبار Qubit dsDNA HS (Invitrogen، #Q32851).
تم تخفيف العينات باستخدام محلول الربط لضمان تركيزات كروماتين متشابهة ولتطابق ظروف IP. تم تخصيص 50 مل من الكروماتين كمدخل لكل عينة. لكل IP، تم غسل 25 مل من كرات مغناطيسية مغطاة ببروتين A (Invitrogen، #10008D) مرة واحدة بمحلول الربط وتم تحضينها مع إما، أو 10 مل من جسم مضاد H3K27Ac. تم ضبط الحجم الكلي للحبيبات بالإضافة إلى الجسم المضاد إلى 200 مل باستخدام محلول الربط وتم تدويره في درجة حرارة الغرفة لمدة 30 دقيقة. لمقارنة التعديلات بعد الترجمة بين العينات، قمنا بإجراء تكرار بيولوجي واحد لجميع العينات في نفس اليوم. قمنا بإعداد مزيج رئيسي من الحبيبات بالإضافة إلى الأجسام المضادة، تم تكبير جميع المكونات بعدد العينات. تم إزالة العازل المحتوي على الأجسام المضادة، وتم إعادة تعليق الخرز مع الأجسام المضادة في 200 مل من الكروماتين القابل للذوبان، تلا ذلك دوران لمدة 30 دقيقة في درجة حرارة الغرفة. تم إزالة الكروماتين غير المرتبط، وتم خلط الخرز مع 500 مل من عازل الربط. تمت إزالة العازل، وتم استرجاع المادة المرتبطة من الخرز عن طريق الخلط في 133 مل من عازل الاسترجاع. في هذه المرحلة، تم رفع المدخل إلى 133 مل عن طريق إضافة 83 مل من عازل الاسترجاع. تم إضافة بروتيناز K (إنفيتروجين، #25530015) إلى تركيز نهائي قدره 15 مللي مول، وتم حضن العينة طوال الليل في.
في صباح اليوم التالي، تم تنقية كل عينة من الحمض النووي باستخدام مجموعة MinElute PCR (Qiagen، #28004) وتم إزالتها في 30 مل من محلول EB. تم قياس 5 مل من الحمض النووي باستخدام مجموعة Qubit dsDNA HS Assay. تم تجميد الـ 25 مل المتبقية من الحمض النووي فيحتى إعداد المكتبة. تم تسلسل المكتبات باستخدام جهاز NextSeq 500 (إيلومينا) باستخدام تسلسل مزدوج الطرف بطول 75 قاعدة.قراءات/عينات لعناوين IPيقرأ/ عينة للإدخال). تم محاذاة بيانات التسلسل من الجيل التالي (NGS) إلى جينوم mm 10. تم معالجة ملفات Bed التي تم إنشاؤها بواسطة محاذاة NGS باستخدام أحدث إصدار من siQ-ChIP (https://github. com/BradleyDickson/siQ-ChIP)، وتم إجراء قياس siQ-ChIP. تم حساب الاستجابات تلقائيًا بواسطة برنامج siQ-ChIP كنسبة المساحة تحت القمم المتداخلة لأي زوج من المسارات التي يتم مقارنتها. تم تصور مسارات ChIP الفردية باستخدام IGV.81.
تحليل تسلسل CUT&Tag
تم إعداد المكتبة لـ CUT&Tag كما تم الإبلاغ عنه سابقًا. باختصار، تم غسل الخلايا مرتين بمحلول PBS، مع كريات مغناطيسية مغطاة بالكونكانافالين A (مختبرات بانغز، BP531) فيمحلول الغسيلهيبسسبيرميدين، ومثبط البروتياز تم حضانة الخلايا معالمزيج المنشط (روش) لمدة 10 دقائق في درجة حرارة الغرفة. تم جمع الكريات المرتبطة بالخلايا وإعادة تعليقها بـمحلول عازل دج-واش (20 مللي مولار هيبس pH 7.5،سبيرميدين، خليط مثبطات البروتياز، وديجيتونين) يحتوي على 2 مللي مول من EDTA،، وتخفيف 1:50 من الأجسام المضادة الأولية، وتم الحضانة فيبين عشية وضحاها. جسم مضاد ثانوي مخفف بنسبة 1:100 فيتم بعد ذلك إضافة محلول Dig-Wash Buffer إلى الكريات وتم حضنه لمدة 60 دقيقة في درجة حرارة الغرفة بعد إزالة الأجسام المضادة الأولية باستخدام حامل مغناطيسي (Vazyme, CM101). تم إعداد مجمع موصل pG-Tn5 وفقًا لتعليمات الشركة المصنعة باستخدام إنزيم pG-Tn5 Transposase النشط للغاية لتقنية CUT&Tag (Vazyme, S602). تم إجراء تقسيم قياسي وتضخيم كما تم الإبلاغ عنه سابقًا.تم تنقية مكتبات الحمض النووي المضخم باستخدام كرات تنظيف الحمض النووي VAHTS (Vazyme، N411) وشحنت للتسلسل بواسطة تقنية الجيل التالي (NGS) من قبل شركة Annoroad Gene Technology. تم تصفية القراءات ورسمها على الجينوم mm10، ثم تم تحديد القمم لإنشاء مصفوفة قمم باستخدام خط أنابيب مشابه لتحليل ATAC-seq المذكور أعلاه. تم تحديد القمم التفاضلية بواسطة، وعدد القراءات المعايرة.
تحليل تسلسل ATAC
تم إجراء تسلسل ATAC كما تم الإبلاغ عنه سابقًاباختصار، تم تحلل الخلايا باستخدام محلول التحلل (10 مللي مول من تريس-هيدروكلوريد، الرقم الهيدروجيني)، و إيجيبال ) في لمدة 10 دقائق. تم إجراء تقسيم وتضخيم وفقًا لتعليمات الشركة المصنعة باستخدام مجموعة تحضير مكتبة الحمض النووي TruePrep V2 (Vazymes، TD501). قامت شركة Annoroad Gene Technology بإجراء تسلسل أحادي الطرف، وتم محاذاة القراءات النظيفة مع المحولات المقطوعة إلى الجينوم المرجعي mm 10 باستخدام حزمة Bowtie2 (2.3.5.1).تمت تسمية القمم العريضة بواسطة حزمة Macs2 (2.1.2)باستخدام المعامل (-nomodel-shift-100-extsize 200 -B-broad)، وتم استدعاء القمم التي تم الوصول إليها بشكل مختلف باستخدام حزمة Deseq2 (الإصدار 1.20.0)تم استدعاء القمم ذات الوصول التفاضلي بواسطة و وأعداد القراءة المعياريةتحليل إثراء الأنماط وقمة- تم إجراء توضيح الجينات المرتبطة باستخدام HOMER (الإصدار 4.11.1) باستخدام القمم المفلترة بواسطةوأعداد القراءة المعياريةتمت تصور مسارات المتصفح بواسطة متصفح IGV (الإصدار 2.8.2) بعد تطبيع القراءة من كل عينة إلى حجم مكتبتها. Ataqv (الإصدار 1.0.0،https://github.com/ParkerLab/ataqvتم استخدام حزمة ) التي طورتها مختبر باركر من جامعة ميتشيغان لإجراء تحليل جودة بيانات ATAC-seq.
فئران
النوع البريتم شراء فئران C57BL/6 من جامعة سانشيا (ييتشانغ، هوبي). Ldhaتم وصف الفئران على خلفية C57BL/6J سابقًاتم تعيين أشقاء من نفس الجنس عشوائيًا إلى مجموعات تجريبية. تم تخصيص الفئران عشوائيًا إلى مجموعات تجريبية، وكان الباحث غير مدرك لتخصيص المجموعات خلال التجربة. تم علاج الفئران التي تتراوح أعمارها بين 6-8 أسابيع بالتاموكسيفين.في زيت الذرة) أو زيت العملة عن طريق الحقن داخل البطن. وفقًا للبروتوكولات المقدمة من مركز كائنات النماذج في شنغهاي (شنغهاي، الصين) لخطوط محركات Cre القابلة للتحفيز، تم تأكيد حذف الإكسون 2 من LDHA عن طريق تحديد النمط الجيني بعد 10 أيام من الحقنة النهائية. تم إجراء جميع التحليلات بشكل أعمى. Ldhaتم تخصيص الفئران عشوائيًا إلى مجموعات تجريبية لمزيد من العلاج، وتم تعيين عينات الخلايا بناءً على النمط الجيني المعني. تم تربية جميع الفئران في ظروف خالية من مسببات الأمراض المحددة في المنشأة الحيوانية المعتمدة في جامعة ووهان.
زراعة بكتيرية
تم زراعة المتفطرة السلية (Mtb) H37Rv في وسط 7H9 (BD) معزّز بـالجلسرين (ويزنت)، 0.05% توين 80 (فيشر)، والألبومين-ديكستروز-كاتالاز تحت اهتزاز مستمر عند. في التجارب في المختبر وفي الجسم الحي، تم طرد بكتيريا Mtb (H37Rv) في مرحلة النمو اللوغاريتمي (OD 0.4-0.9) باستخدام الطرد المركزي ( ) وإعادة تعليقها في RPMI بدون بنسلين/ستربتوميسين أو PBS معقم.
تم إضافة الفورمالديهايد إلى وسط الثقافة بتركيز نهائي قدرهثم تم غسل الخلايا مرتين بمحلول PBS، وتم كشطها، وتحليلها في محلول التحليل. SDS، 10 مللي مول من تريس-هيدروكلوريد [رقم الهيدروجين 8.0]، 10% خليط مثبطات البروتياز، كل من الأبرتينين والليوبيبتين) لمدة 10 دقائق فيتمت معالجة المستخلصات بالموجات فوق الصوتية على الثلج، وتمت إزالة الحطام عن طريق الطرد المركزي عندلمدة 15 دقيقة عندتم استخدام ربع السائل العلوي كتحكم في إدخال الحمض النووي. تم تخفيف العينة المتبقية بمقدار 10 مرات باستخدام محلول التخفيف. SDS، ترايتون X-100، 1 مللي مول من EDTA، 10 مللي مول من تريس-HCl [pH 8.0]، و150 مللي مول من NaCl) تليها حضانة مع الأجسام المضادة طوال الليل فيتم جمع المجمعات المناعية باستخدام بروتين A/GSepharose. تم غسل الرواسب أربع مرات باستخدام محلول الغسيل الذي يحتوي على 2 مللي مول من EDTA و 50 مللي مول من Tris-HCl، pH 8.0. بعد الغسل، تم حاضنة الرواسب مع محلول الإزالة ( SDS و 0.1 م ) في درجة حرارة الغرفة. تم نقل السائل العلوي إلى أنابيب نظيفة، وتم إضافة RNase A لتدمير RNA المرتبط في العينة. تم حضن العينات عند لمدة 5 ساعات لعكس الروابط المتقاطعة للفورمالدهيد، وتم ترسيب الحمض النووي باستخدام الإيثانول واستخراجه مرتين باستخدام الفينول-كلوروفورم. أخيرًا، تم إعادة تعليق الحبيبات في محلول تريس-EDTA (TE) وخضعت لتضخيم PCR. تم إجراء RT-PCR الكمي في الوقت الحقيقي باستخدام جهاز LightCycler 480 (روش) ونظام SYBR الأخضر (أبلايد بيوسيستمز). تم تضخيم GAPDH كعنصر تحكم داخلي.
تحليل HPLC-MS/MS
تم إجراء قياس الطيف الكتلي وتحديد المستقلبات علىالميثانول والمستقلبات المستخرجة من الماء بدرجة LC/MS. تم إجراء التحليلات باستخدام الكروماتوغرافيا السائلة عالية الأداء، وطيف الكتلة عالي الدقة، وطيف الكتلة المتسلسل (HPLC-MS/MS). كان النظام يتكون من جهاز Thermo
Q-Exactive متصل بمصدر رذاذ كهربائي وسلسلة HPLC Ultimate 3000 (ثيرمو) تتكون من جهاز إزالة الغاز، ومضخة ثنائية، وعينة تلقائية مزودة بعمود Xbridge Amide (أبعاد و حجم الجسيمات). المرحلة المتحركة A: 20 مللي مول من أسيتات الأمونيوم (pH 9.0)، 20 مللي مول من هيدروكسيد الأمونيوم،ماء، وأسيتونيتريل. المرحلة المتحركة ب:أسيتونيتريل. كان التدرج كما يلي: ( 2.5 دقيقة ) ; بمعدل تدفق تم ضبط الشعيرة في مصدر ESI على، مع غاز الغلاف عند 45 وحدة عشوائية، وغاز مساعد عند خمس وحدات عشوائية، وجهد الرش عند 4.0 كيلو فولت. تم تكسير أعلى 5 أيونات سابقة باستخدام خلية التفكك بالاصطدام عالي الطاقة المضبوطة علىطاقة الاصطدام العادية في MS2. تم إجراء جمع البيانات وتحليلها بواسطة برنامج Tracefinder 2.1 (ثيرمو فيشر العلمية) وبرنامج Xcalibur 4.0 (ثيرمو فيشر العلمية).
في الجسم الحي-تسريبات المتعقب
تم حقن الفئران عن طريق الوريد بـ-المتتبعات المعلّمة.معدل تسريب اللاكتات هولمدة 4 ساعات لفأر وزنه 20 جرام. ثم تم عزل المصل والرئة ومعالجتهما للتحليل الأيضي.
تحديد مستوى ATP الميتوكوندري بواسطة تحليل تدفق الخلايا
لتحديد ATP المعتمد على FRET، تم استخدام جهاز قياس التدفق Novocyte 3000 (ACEA BioSciences) لتسجيل فلوريسcence ERAT عند قنوات مع فلاتر Ex/Em تم ضبطها كما يلي: (1) (عرض النطاق: 45 ). (2) تم تعريف مستويات ATP الميتوكوندري للأفراد الخلوية على أنها نسبة شدة الفلورسنت للقناة 2 (Ex/Em: 405/530) مقسومة على تلك الخاصة بالقناة 1 (Ex/Em: 405/445)، مشابهة للمعايير المستخدمة سابقًا..
المواد الكيميائية ومجموعات الفحص التجارية الحيوية الهامة
تفاصيل المواد الكيميائية الحرجة، والأجسام المضادة، ومجموعات الفحص التجارية مدرجة في الجدول التكميلي 1 والجدول التكميلي 2. ما لم يُذكر خلاف ذلك، تم شراء جميع المواد الكيميائية الحيوية من سيغما-ألدريش.
التحليل الإحصائي
تم الحصول على البيانات من ثلاثة تجارب مستقلة قابلة للتكرار. تم التعبير عن البيانات كمتوسط ± الانحرافات المعيارية أو متوسط ± الخطأ المعياري للمتوسط. تم تحديد الدلالة الإحصائية باستخدام اختبار t غير المتزاوج ذو الطرفين لطالب، واختبارات ANOVA أحادية الاتجاه أو ثنائية الاتجاه كما هو موضح في الأسطورة.تم اعتبار القيمة < 0.05 دالة إحصائيًا وتم تمييزها بنجمة (*).
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة ناتشر المرتبط بهذه المقالة.
Stienstra, R., Netea-Maier, R. T., Riksen, N. P., Joosten, L. A. B. & Netea, M. G. Specific and complex reprogramming of cellular metabolism in myeloid cells during innate immune responses. Cell Metab. 26, 142-156 (2017).
Zhou, L. et al. Hepatitis B virus rigs the cellular metabolome to avoid innate immune recognition. Nat. Commun. 12, 98 (2021).
He, Q. et al. MAVS integrates glucose metabolism and RIG-I-like receptor signaling. Nat. Commun. 14, 5343 (2023).
Pan, C., Li, B. & Simon, M. C. Moonlighting functions of metabolic enzymes and metabolites in cancer. Mol. Cell 81, 3760-3774 (2021).
Muir, A., Danai, L. V. & Vander Heiden, M. G. Microenvironmental regulation of cancer cell metabolism: implications for experimental design and translational studies. Dis. Model Mech. 11, dmmO35758 (2018).
Hackett, S. R. et al. Systems-level analysis of mechanisms regulating yeast metabolic flux. Science 354, aaf2786 (2016).
Rossiter, N. J. et al. CRISPR screens in physiologic medium reveal conditionally essential genes in human cells. Cell Metab. 33, 1248-1263.e9 (2021).
Puleston, D. J., Villa, M. & Pearce, E. L. Ancillary activity: beyond core metabolism in immune cells. Cell Metab. 26, 131-141 (2017).
. et al. Interleukin- 6 classic and trans-signaling utilize glucose metabolism reprogramming to achieve anti- or pro-inflammatory effects. Metabolism 155, 155832 (2024).
Peng, M. et al. Aerobic glycolysis promotes T helper 1 cell differentiation through an epigenetic mechanism. Science 354, 481-484 (2016).
Lin, J., Liu, G., Chen, L., Kwok, H. F. & Lin, Y. Targeting lactate-related cell cycle activities for cancer therapy. Semin Cancer Biol. 86, 1231-1243 (2022).
Faubert, B. et al. Lactate metabolism in human lung tumors. Cell 171, 358-371.e9 (2017).
Zhang, W. et al. Lactate is a natural suppressor of RLR signaling by targeting MAVS. Cell 178, 176-189.e15 (2019).
Hui et al. Glucose feeds the TCA cycle via circulating lactate. Nature 551, 115-118 (2017).
Zhang, D. et al. Metabolic regulation of gene expression by histone lactylation. Nature 574, 575-580 (2019).
Chen, A.-N. et al. Lactylation, a novel metabolic reprogramming code: current status and prospects. Front Immunol. 12, 688910 (2021).
Khan, N. et al. M. tuberculosis reprograms hematopoietic stem cells to limit myelopoiesis and impair trained immunity. Cell 183, 752-770.e22 (2020).
Mitroulis, I. et al. Modulation of myelopoiesis progenitors is an integral component of trained immunity. Cell 172, 147-161.e12 (2018).
Kalafati, L. et al. Innate immune training of granulopoiesis promotes anti-tumor activity. Cell 183, 771-785.e12 (2020).
Cirovic, B. et al. BCG vaccination in humans elicits trained immunity via the hematopoietic progenitor compartment. Cell Host Microbe 28, 322-334.e5 (2020).
Netea, M. G. et al. Trained immunity: a program of innate immune memory in health and disease. Science 352, aaf1098 (2016).
Netea, M. G. & van der Meer, J. W. M. Trained immunity: an ancient way of remembering. Cell Host Microbe 21, 297-300 (2017).
Netea, M. G. et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 20, 375-388 (2020).
Cheng, S.-C. et al. mTOR- and HIF- -mediated aerobic glycolysis as metabolic basis for trained immunity. Science 345, 1250684 (2014).
Cantor, J. R. et al. Physiologic medium rewires cellular metabolism and reveals uric acid as an endogenous inhibitor of UMP synthase. Cell 169, 258-272.e17 (2017).
Kaymak, I. et al. Carbon source availability drives nutrient utilization in CD8+ T cells. Cell Metab. 34, 1298-1311.e6 (2022).
Luda, K. M. et al. Ketolysis drives CD8+ T cell effector function through effects on histone acetylation. Immunity 56, 2021-2035.e8 (2023).
Ron-Harel, N. et al. T cell activation depends on extracellular alanine. Cell Rep. 28, 3011-3021.e4 (2019).
Su, H. et al. Glutathione synthesis primes monocytes metabolic and epigenetic pathway for -glucan-trained immunity. Redox Biol. 48, 102206 (2021).
Luengo, A. et al. Increased demand for NAD+ relative to ATP drives aerobic glycolysis. Mol. Cell 81, 691-707.e6 (2021).
Peignier, A. & Parker, D. Trained immunity and host-pathogen interactions. Cell Microbiol 22, e13261 (2020).
Zhao, Y. et al. In vivo monitoring of cellular energy metabolism using SoNar, a highly responsive sensor for NAD(+)/NADH redox state. Nat. Protoc. 11, 1345-1359 (2016).
Cheng, S.-C. et al. Broad defects in the energy metabolism of leukocytes underlie immunoparalysis in sepsis. Nat. Immunol. 17, 406-413 (2016).
Saeed, S. et al. Epigenetic programming of monocyte-tomacrophage differentiation and trained innate immunity. Science 345, 1251086 (2014).
Farber, D. L., Netea, M. G., Radbruch, A., Rajewsky, K. & Zinkernagel, R. M. Immunological memory: lessons from the past and a look to the future. Nat. Rev. Immunol. 16, 124-128 (2016).
Netea, M. G., Quintin, J. & van der Meer, J. W. M. Trained immunity: a memory for innate host defense. Cell Host Microbe 9, 355-361 (2011).
Arts, R. J. W. et al. Glutaminolysis and fumarate accumulation integrate immunometabolic and epigenetic programs in trained immunity. Cell Metab. 24, 807-819 (2016).
Bekkering, S. et al. Metabolic induction of trained immunity through the mevalonate pathway. Cell 172, 135-146.e9 (2018).
Repnik, U., Knezevic, M. & Jeras, M. Simple and cost-effective isolation of monocytes from buffy coats. J. Immunol. Methods 278, 283-292 (2003).
Yu, H. et al. Major vault protein promotes hepatocellular carcinoma through targeting interferon regulatory factor 2 and decreasing p53 activity. Hepatology 72, 518-534 (2020).
Kaya, H. S. et al. CUT&Tag for efficient epigenomic profiling of small samples and single cells. Nat. Commun. 29, 1930 (2019).
Buenrostro, J. D., Giresi, P. G., Zaba, L. C., Chang, H. Y. & Greenleaf, W. J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 10, 1213-1218 (2013).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357-359 (2012).
Zhang, Y. et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 9, R137 (2008).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Xie, H. et al. Targeting lactate dehydrogenase-a inhibits tumorigenesis and tumor progression in mouse models of lung cancer and impacts tumor-initiating cells. Cell Metab. 19, 795-809 (2014).
Imamura, H. et al. Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encodedindicators. PNAS 106, 15651-15656 (2009).
Vishnu, N. et al. ATP increases within the lumen of the endoplasmic reticulum upon intracellular release. Mol. Biol. Cell 25, 368-379 (2014).
شكر وتقدير
تم دعم هذا العمل من قبل البرنامج الوطني الرئيسي للبحث والتطوير في الصين (أرقام المنح: 2021YFC2701800، 2021YFC2701804، 2023YFC2307800)، مؤسسة العلوم الطبيعية الوطنية في الصين (رقم المنحة U22A2O335)، صناديق البحث الأساسية للجامعات المركزية (رقم المنحة: 2042024kf0011، 2042022dx0003)، مؤسسة العلوم الطبيعية في ووهان (رقم المنحة: 2024040701010031)، تجمع المواهب عالية المستوى في هوانغ (رقم المنحة: 2021RC5006)، الأموال من إدارة العلوم والتكنولوجيا في مقاطعة هوبى (رقم المنحة: 2022CFB624)، برنامج الابتكار المعرفي في ووهان (رقم المنحة: 2022020801010116)، صناديق البحث الأساسية للجامعات المركزية (رقم المنحة: 2042021kf023).
مساهمات المؤلفين
H.C.، X.C.، Y.L.: تنسيق البيانات، إدارة المشروع، كتابة المسودة الأصلية. Y.C.، G.Z.: تنسيق البيانات، إدارة المشروع، الموارد. X.C.: الموارد. S.R.، H.Z.: تنسيق البيانات. L.Z.: إدارة المشروع.Z.Li.: الإشراف. A.L.: التصور. X.Z.: إدارة المشروع. W.X.، Li.Z.: تنسيق البيانات، C.G.: الموارد. Y.Z.: الموارد. K.D.: الموارد.Z.Lu., H.R.: الموارد. H.Y.، Z.C.، K.Xu.: التصور، الحصول على التمويل، الموارد، الإشراف. F.W.، S.L.: التصور، الحصول على التمويل، الموارد، الإشراف.
ملاحظة الناشر تظل Springer Nature محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
الوصول المفتوح هذه المقالة مرخصة بموجب ترخيص المشاع الإبداعي النسب-غير التجارية-لا اشتقاقات 4.0 الدولية، والذي يسمح بأي استخدام غير تجاري، ومشاركة، وتوزيع، وإعادة إنتاج في أي وسيلة أو صيغة، طالما أنك تعطي الائتمان المناسب للمؤلفين الأصليين والمصدر، وتوفر رابطًا لترخيص المشاع الإبداعي، وتوضح إذا قمت بتعديل المادة المرخصة. ليس لديك إذن بموجب هذا الترخيص لمشاركة المواد المعدلة المشتقة من هذه المقالة أو أجزاء منها. الصور أو المواد الأخرى من طرف ثالث في هذه المقالة مشمولة في ترخيص المشاع الإبداعي للمقالة، ما لم يُشار إلى خلاف ذلك في سطر الائتمان للمادة. إذا لم تكن المادة مشمولة في ترخيص المشاع الإبداعي للمقالة واستخدامك المقصود غير مسموح به بموجب اللوائح القانونية أو يتجاوز الاستخدام المسموح به، ستحتاج إلى الحصول على إذن مباشرة من صاحب حقوق الطبع والنشر. لعرض نسخة من هذا الترخيص، قم بزيارة http:// creativecommons.org/licenses/by-nc-nd/4.0/.
¹قسم أمراض القلب، مستشفى تشونغنان بجامعة ووهان، ووهان 430072، الصين. ²معهد إصابة القلب وإصلاحه، جامعة ووهان، ووهان 430072، الصين. قسم الأمراض المعدية، المركز الطبي الخامس للمستشفى العام لجيش التحرير الشعبي الصيني، بكين 100039، الصين. المختبر الوطني الرئيسي لعلم الفيروسات، مركز أبحاث الفيروسات الحديثة، كلية علوم الحياة، جامعة ووهان، ووهان 430072، الصين. مركز العلوم الحدودية لعلم المناعة والتمثيل الغذائي، مركز تايكانغ للعلوم الطبية والحياتية، جامعة ووهان، ووهان 430072، الصين. قاعدة التربية للمختبر الوطني الرئيسي لعلوم طب الأسنان الأساسية (هوبى-موست) & المختبر الرئيسي للطب الحيوي الفموي وزارة التعليم، كلية ومستشفى طب الأسنان، جامعة ووهان، ووهان 430072، الصين. قسم الجراحة العامة، المستشفى الأول التابع لجامعة غاننان الطبية، غانزو 341000، الصين. قسم الأمراض المعدية، مستشفى الشعب الأول في لونغنان، لونغنان 341700، الصين. مركز شنجهاي الطبي للأطفال، المستشفى التابع لجامعة شنجهاي جياو تونغ كلية الطب، شنجهاي 200240، الصين. مركز ووهان للأبحاث حول الأمراض المعدية والسرطان، الأكاديمية الصينية للعلوم الطبية، ووهان 430072، الصين. ساهم هؤلاء المؤلفون بالتساوي: هوانهوان كاي، شيويوان تشين، يان ليو. البريد الإلكتروني: luzhibing222@163.com; wfb20042002@sina.com; liushi_liushi@whu.edu.cn
Lactate activates trained immunity by fueling the tricarboxylic acid cycle and regulating histone lactylation
Received: 6 August 2024
Accepted: 24 March 2025
Published online: 04 April 2025
Check for updates
Huanhuan Cai ${ }^{1,2,11}$, Xueyuan Chen ${ }^{3,11}$, Yan Liu ${ }^{3,11}$, Yingbo Chen ${ }^{4,5}$, Gechang Zhong ${ }^{4,5}$, Xiaoyu Chen ${ }^{4,5}$, Shuo Rong ${ }^{4,5}$, Hao Zeng ${ }^{6}$, Lin Zhang ${ }^{1,2}$, Zelong Li ${ }^{7,8}$, Aihua Liao ${ }^{7,8}$, Xiangtai Zeng ${ }^{7,8}$, Wei Xiong ${ }^{1,2}$, Cihang Guo ${ }^{1,2}$, Yanfang Zhu ${ }^{1,2}$, Ke-Qiong Deng ${ }^{1,2}$, Hong Ren ${ }^{9}$, Huan Yan ${ }^{(1)}{ }^{4,5}$, Zeng Cai ${ }^{4,5}$, Ke Xu ${ }^{4,5}$, Li Zhou ${ }^{(1)}{ }^{4,5}$, Zhibing Lu ${ }^{(1)}{ }^{1,2}$, Fubing Wang ${ }^{(1)}{ }^{10}$ & Shi Liu ${ }^{(1)}{ }^{1,2,4,5}$ ®
Trained immunity refers to the long-term memory of the innate immune cells. However, little is known about how environmental nutrient availability influences trained immunity. This study finds that physiologic carbon sources impact glucose contribution to the tricarboxylic acid (TCA) cycle and enhance cytokine production of trained monocytes. Our experiments demonstrate that trained monocytes preferentially employe lactate over glucose as a TCA cycle substrate, and lactate metabolism is required for trained immune cell responses to bacterial and fungal infection. Except for the contribution to the TCA cycle, endogenous lactate or exogenous lactate also supports trained immunity by regulating histone lactylation. Further transcriptome analysis, ATAC-seq, and CUT&Tag-seq demonstrate that lactate enhance chromatin accessibility in a manner dependent histone lactylation. Inhibiting lactatedependent metabolism by silencing lactate dehydrogenase A (LDHA) impairs both lactate fueled the TCA cycle and histone lactylation. These findings suggest that lactate is the hub of immunometabolic and epigenetic programs in trained immunity.
Reprogramming cellular metabolic processes has recently been proven essential for immune cells . A widely accepted concept is that cell-intrinsic factors, such as critical molecules or metabolic enzymes, play a crucial role in the metabolic pathway in immune cells . However, the availability of nutrients in the environment also significantly
impacts metabolic utilization in immune cells . For example, yeast substrate concentrations impacted metabolic output twice more than enzyme expression . Cancer cells show changes in cell growth and stress resistance if cancer cells are cultured with a physiologic medium due to differences in substrate utilization . Among nutrients in the
environment, glucose is a significant nutrient that fuels cellular metabolic activities. After uptake through the glucose transporter, glucose flows through three distinctly metabolic pathways: glycolysis, the pentose phosphate pathway (PPP), and the hexosamine biosynthesis pathway . Glucose is metabolized via glycolysis into pyruvate, which enters the TCA cycle when oxygen is available . When oxygen is unavailable, pyruvate shifts to glycolysis, in which pyruvate converts to lactate . Traditionally, lactate has been seen as a garbage of glucose metabolism . However, a growing body of research shows that lactate is the primary circulating metabolite that fuels the TCA cycle and a significant substrate for gluconeogenesis . A recent study identified a new epigenetic modification, histone lactylation, that regulates histones by adding lactyl groups to their lysine residues . Histone lactylation has different temporal dynamics from acetylation and induces corresponding gene expression . Subsequently, more than 30 histone lactylation sites have been identified in fungal, plant, mouse, and parasitic (protozoic) samples .
Conventional wisdom says that the immune system is divided into the innate and the adaptive immune system. One of the main bases for this division is that the adaptive immune system is relatively slower and can build specific immunological memory . By contrast, the innate immune system is rapid and non-specific and cannot build immunological memory . However, recent studies have shown that innate immune cells, such as natural killer cells and monocytes, can make a non-specific memory that challenges this paradigm . Specifically, the functional adaptation of innate immunity after a primary insult, including Bacille Calmette-Guerin vaccination or microorganism infection, represents a de facto innate immune memory . Once these innate immune cells encounter a secondary non-specific (heterologous) stimulus, their response is altered, which results in a more robust response . The induction of a non-specific memory results in a stepped-up function of the innate immune cells termed trained immunity . The biological relevance of innate immune memory, such as cephalopods, plants, insects, and mammals, is widespread . Analysis of differentially regulated pathways in trained immunity showed that mTOR-HIF1 regulated oxidative phosphorylation to glycolysis, the central regulatory mechanism for -glucan, a fungal cell wall component, induced trained immunity . Nevertheless, how environmental nutrient availability impacts trained immunity metabolism and function remains poorly understood.
In this study, we found that physiologic carbon sources (PCSs) influence the glucose utilization of trained monocytes. Our experiments have demonstrated that lactate is a physiological fuel for trained immunity, is preferred over glucose, and regulates the cytokine production of trained immune cells. Mechanistically, some intrinsic lactate function is a significant substrate for acetyl-CoA synthesis and fuels the TCA cycle. The other inherent lactate moves into the cell nucleus and regulates cytokine production through the effects of histone lactylation. Our data identifies the dominant metabolite (lactate) in trained immunity, contributes to our understanding of innate immune memory, and opens new therapeutic avenues.
Results
PCSs impact glucose utilization in monocytes during the induction of trained immunity
Traditional cell culture mediums, such as the Roswell Park Memorial Institute (RPMI) 1640 medium, are designed to promote cell growth and survival ex vivo . Thus, the concentration of some nutrients in traditional cell culture medium is supraphysiological (Supplementary Fig. 1a). In addition, the conventional medium also lacks most standard culture media, such as lactate, citrate, acetate, -hydroxybutyrate , and pyruvate . In this study, we referred to those metabolites as PCSs (Supplementary Fig. 1a). Previous studies show that these carbon sources are oxidizable substrates in vivo . Notably, in some situations, these carbon sources exhibit circulatory turnover fluxes
exceeding traditional sources, such as glucose and glutamine . Thus, we designed the modified RPMI medium (MM) with approximate polar metabolite concentrations in mouse serum (Supplementary Fig. 1a).
To obtain trained immune cells, we treated purified monocytes with -glucan for 24 hrs , followed by washing and resting for five days in a culture medium. Then, cells were restimulated with -glucan or LPS for 24 hrs as previously described (Supplementary Fig. 1b). -glucose labeling into lactate and intracellular TCA cycle intermediates, including citrate and fumarate, within in -glucan treated monocytes (Supplementary Figs. 1c). Analysis of metabolite levels in culture medium over time revealed several trends (Supplementary Figs. 1d-1f). As expected, glucose and glutamine were consumed within 12-24 hrs during the trained immunity process, and PCS treatment mildly affected glucose and glutamine consumption in glucan treated monocytes (Supplementary Figs. 1d and 1e). A slower decrease of other carbon sources, including OHB, citrate, and pyruvate, was observed in the PCS-supplemented medium compared to the glucose-supplemented medium (Supplementary Fig. 1f). Interestingly, we observed accumulation of several TCA cycle intermediates ( and fumarate) in extracellular medium over time, indicated the export of these metabolites (Supplementary Fig. 1f).
Using -based metabolic tracers, we investigate the role of PCSs on glucose metabolism in trained immunity (Fig. 1a). Mass spectrometry (MS) assay shows that -glucose extensively labels intermediates of glycolysis and the TCA cycle in -glucan treated monocytes (Figs. 1b-1d). By contrast, in the presence of PCSs at a physiologic concentration in -glucan treated monocytes, -glucose contribution into TCA cycle intermediates was significantly inhibited (Figs. 1b-1d). Intriguingly, the presence of PCSs at physiologic concentration directs glucose flux to the PPP and the serine synthesis pathway (SSP) (Figs. 1e and 1d). Similar results were obtained in metabolites arising from TCA cataplerosis, such as glutamate and aspartate (Fig. 1g). We observe similar labeling patterns in -glucan treated monocytes cultured PCSs and -glucose for long (6 days) (Supplementary Fig. 1g). To investigate the contribution of PCSs to TCA cycle metabolism, -glucan treated monocytes were cultured in MM supplemented with separated -labeled PCSs at physiologic concentrations. We observe that -glucose and -glutamine contributed carbon to the TCA cycle intermediates (Supplementary Fig. 1h). Intriguingly, TCA cycle intermediates were also labeled from some PCSs, including and lactate, in -glucan treated monocytes (Supplementary Fig. 1h). Results showed that and lactate contribute carbon to the TCA cycle by 2-carbon entry via citrate synthase ( citrate) and carry through the entire cycle in -glucan treated monocytes (Fig. 1h). Of note, when considering the lower concentration of lactate compared with the concentration of glucose , trained monocytes prefer to use PCSs over glucose for TCA cycle metabolism (Fig. 1h). Because it has been reported that alanine did not contribute to the TCA cycle , the presence of -alanine was included as a negative control for comparison (Supplementary Fig. 1h). Taken together, these findings suggest that PCSs reduced the contribution of -glucose to the TCA cycle, and drives -glucose switch from glycolysis to PPP and SSP, in trained immunity.
Lactate activates trained immunity
Because among these above PCSs, lactate has the most significant regulation on the TCA cycle (Fig. 1h), we next investigated the role of lactate on trained immunity in response to C. albicans infection (Fig. 2a). In line with a previous study , C. albicans infection enhances the production of lactate in a time- and dose-dependent manner (Supplementary Fig. 2a). Since lactate dehydrogenase (Ldh) interconverts lactate and pyruvate by using NAD /NADH as a co-factor , we asked whether LDHA plays a role in trained immunity. Firstly, we employed the tamoxifen-inducible LDHA knockout mice ( ), and
Fig. 1 | PCSs impact glucose utilization in trained immunity. a Schematic of -glucose carbon labeling through the TCA cycle, the pentose phosphate pathway (PPP), and the serine synthesis pathway (SSP). b-g Monocytes were cultured in MM containing -glucose ( ) and treated with -glucan ( ) for 4 hrs , followed by mass spectrometry analyzes. h Monocytes were cultured in MM
containing the indicated -labeled carbon source and treated with -glucan ( ml ) for 4 hrs . The relative contributions of from indicated -labeled substrates into metabolites of central carbon metabolism are shown. Data in (b-h) are expressed as means ± SEMs, mice per condition, two-way ANOVA. Medium: Phosphate buffered saline. See also Supplementary Fig. 1.
Fig. 2 | Lactate regulates trained immunity. a Schematic representation of in vivo trained immunity experimental setup. b-d and mice were trained with or without -glucan. Six days later, mice were infected with or without a secondary . albicans lethal dose for 24 hrs , followed by measuring IL- 6 and TNF- levels (b), lung injury(c), and kidney and lung fungal burden (d). e and mice were trained with or without -glucan. Six days later, mice were infected with or without a secondary C. albicans lethal dose ( ) for indicated times. Survival curves show data collected until day ten after infection. Statistical analysis was performed using the log-rank test ( for each group). C57BL/6 mice were trained with or without -glucan. Six days later, mice were secondarily infected
without or with C. albicans ( ) and infused with or without lactate ( 1 mM ) for 24 hrs , followed by measuring IL-6 and TNF- levels (f), kidney fungal burden (g), and lactate levels in serum (h). i C57BL/6 mice were trained with or without glucan. Six days later, mice were infected with or without a secondary C. albicans lethal dose ( ) and infused with or without lactate ( 1 mM ) for indicated times. Survival curves show data collected until day 12 after infection. Statistical analysis was performed using the log-rank test ( for each group). Data in (b-d, and ) are expressed as means ± SEMs, mice per condition, two-way ANOVA. See also Supplementary Figs. 2.
LDHA protein expression in diverse organs of mice was abrogated upon tamoxifen treatment (Supplementary Figs. 2b and 2c). To investigate the role of lactate on trained monocyte function, we trained mice with -glucan followed by a challenge with . albicans (Fig. 2a). As expected, IL-6 and TNF- production was induced in -glucan-trained mice, but not in mice (Fig. 2b). Histological analysis of the lung tissue showed that prior -glucan treatment limits lung damage during subsequent infection (Fig. 2c). Compared with -glucan-trained mice, more severe lung injury was demonstrated in mice (Fig. 2c). In this vein, mice significantly susceptible to C. albicans-induced lethality, accompanied by higher bacterial burden in the lung and kidney tissue (Figs. 2d and 2e). A previous study
reported that trained immunity is a protective host against secondary C. albicans infection that relies on monocytes induced by the same pathogen . Similar results were obtained in C. albicans-trained Wt and mice (Supplementary Figs. 2d-2g). To determine whether LDHA regulates trained immunity in vitro, we employed two specific small interfering RNAs (siRNAs) for LDHA. We confirmed siRNA efficiency (Supplementary Fig. 2h). SiRNA-LDHA #2 was selected for the following experiments. As shown in Supplementary Fig. 2i, LDHA knockdown inhibits -glucan-induced expression of IL-6 and TNF- in human peripheral blood mononuclear cells (PBMCs).
Next, we investigated the role of lactate on trained immunity in response to C. albicans infection. As shown in Supplementary Fig. 2j,
lactate enhances -glucan-induced expression of IL-6 and TNF- in a dose-dependent manner in human PBMCs. Of note, to rule out the possibility that lactate affects trained immunity by changing the acidity , the pH of the medium was adjusted to an equal level in all in vitro experiments of this manuscript. The presence of lactate increased C. albicans-induced levels of IL-6 and TNF- , accompanied by lower fungal burden in the kidney and lung tissue (Figs. 2f-2h). Strikingly, lactate increased the protective response of trained monocytes against lethal systemic C. albicans infection (Fig. 2i). These findings suggest that lactate is indispensable for the protective reaction of trained immunity against C. albicans infection.
Lactate is a physiologic fuel during the induction of trained immunity
Next, we investigate the mechanisms by which lactate regulates trained immunity. Firstly, we measured the timeline of -glucose incorporation during -glucan treatment. -glucan treated monocytes displayed higher -glucose incorporation into glucose-6-phosphate (G6P), fructose-6-phosphate (F6P), 3-phosphoglycerate (3PG), pyruvate, lactate, citrate, succinate, malate, glutamate and aspartate within 0-4 hrs (Supplementary Figs. 3a-3f), indicates increased glycolytic flow to the TCA cycle. -glucose incorporation flow from oxidative phosphorylation (OxPhos) to SSP and PPP, as well as ATP synthesis and S-adenosylmethionine (SAM) generation, after 4 hrs of glucan treatment (Supplementary Figs. 3a-3g). However, -glucose incorporation into glycolysis continued to increase throughout the experiment (Supplementary Fig. 3b). In addition, -glucan also induced hexokinase 2 (HK2) enzymes and LDHA enzyme activity in a time- and dose-dependent manner (Supplementary Figs. 3h and 3i). Those data suggest -glucan treated monocytes prefer to use glucose as physiologic fuel early. Still, -glucan treated monocytes prefer to use glucose for biosynthesis later.
To evaluate the competition between glucose and lactate for TCA cycle metabolism, treated monocytes were cultured with -glucose and unlabeled lactate with gradually increased concentration. Even in the presence of physiologic glucose concentrations ( 5 mM ), -glucan treated monocytes use low concentrations of lactate ( ) as a carbon source for intermediates in the TCA cycle (Figs. 3a-3d). By contrast, a low concentration of lactate directs glucose flux from OxPhos to SSP and PPP (Figs. 3a-3f). A substantial fraction of glucose-derived labeled SAM, including (via the PPP) and (via both the PPP and SSP), significantly increased in lactate-treated monocytes (Figs. 3g and 3 h ), indicates glucose is used for biosynthesis-in the presence lactate. Because lactate contributes to TCA cycle metabolism, we next investigated the role of lactate on trained monocytes bioenergetics. By using Forster resonance energy transfer (FRET)-based ATP sensors targeted to mitochondria (Mito, mtAT1.03), we found that the presence of lactate or -glucan treatment enhances ATP production (Fig. 3i). The combination of -glucan and lactate treatment resulted in a significant synergistic induction of ATP production (Fig. 3i). Similarly, the presence of lactate enhances their basal oxygen consumption rate (OCR) and ATP production from OxPhos (Supplementary Fig. 3j). Consistent with metabolite data, lactate increases -glucan-induced levels of acetyl-CoA, citrate, and malate (Fig. 3j). Finally, we investigate the role of glucose on the lactate contributed TCA cycle metabolism during -glucan treatment. As shown in Fig. 3k, a low lactate concentration ( 1 mM ) still contributes to TCA cycle intermediates in glucose ( ) contained culture medium during -glucan treatment. When the glucose concentration is 25 -fold higher than lactate levels, -lactate could not contribute carbon to the TCA cycle (Fig. 3k). These findings suggest lactate is a TCA cycle substrate for trained monocytes under physiologic conditions, even when glucose is available.
Lactate fuels the trained immunity via LDHA
Next, we investigate the role of LDHA on the bioenergetic function of lactate in trained immunity. As shown in Figs. 4a and 4b, decreased abundance of -lactate-derived citrate, succinate, malate, glutamate, and aspartate was observed in monocytes in response to glucan treatment. -lactate also contributed to acetyl-CoA-related metabolites, including Ac-carnitine and Ac-alanine, in -glucan treated monocytes, but not in -glucan treated monocytes (Fig. 4c). By using the mitochondrial ATP probe , -glucan treatment enhances and ATP production in monocytes, but not in -glucan treated monocytes (Fig. 4d). Consistently, LDHA knockout inhibited -glucan-induced levels of acetyl-CoA, citrate, malate (Fig. 4e). Sodium oxamate (Oxa) is widely used to inhibit lactate production by regulating the activities of LDHA . Consistently, -glucaninduced levels of acetyl-CoA, citrate, and malate were inhibited in Oxatreated trained monocytes (Supplementary Fig. 4a). We next explored whether the host used lactate as a fuel in trained immunity in vivo by infusing metabolite. In our in vivo system, enrichment of fully labeled ( ) -lactate in serum and lung (Figs. 4 f and 4 g ). Consistent with ex vivo data, infused -lactate readily labeled TCA cyclederived metabolites in vivo in response to -glucan treated mice, but not in mice (Figs. 4h and 4i). We further investigated -glucan-regulated glucose metabolism in lactate metabolism inhibited system. Intriguingly, -glucan treatment directs glucose flux from PPP and SSP back to OxPhos when lactate metabolism is inhibited (Supplementary Figs. 4b-4f), suggesting glucose is an alternative carbon source for the TCA cycle in trained immunity. These findings indicate that lactate processing via LDHA is a critical metabolic node in trained immunity.
Lactate regulates histone lactylation via LDHA in monocytes during the induction of trained immunity
A previous study showed that lysine lactylation (Kla) is a new type of histone mark that can be stimulated by lactate (Supplementary Fig. 5a); we suspected that -glucan regulated high level of lactate impact on histone lactylation in trained immunity. Immunoblots analysis revealed that histone Kla levels were increased; however, histone acetylation (Kac) levels were mildly increased, in a time-dependent fashion in β-glucan treated monocytes (Fig. 5a and Supplementary Fig. 5b). Histone Kla levels elevation was detected as early as 6 hrs after -glucan treatment (Fig. 5a and Supplementary Fig. 5b). Intriguingly, high histone Kla levels were maintained from days 1 to 9 (Fig. 5b and Supplementary Fig. 5c). As expected, -glucan challenge induce histone Kla levels, mildly induce Kac levels, in dose-dependent manner (Fig. 5c and Supplementary Fig. 5d). Given that replenishing cells with fresh media did not affect Kla levels, the increase of histone Kla during trained immunity is intrinsic and not due to paracrine effects (Supplementary Fig. 5e). Consistent with previous results , MS/MS analysis showed that a mass shift of 72.021 Da on lysine residues in the histone was observed in -glucan treated monocytes (Supplementary Figs. 5f and 5g). In this vein, metabolic labeling experiments using -glucose or -lactate followed by MS/MS analysis demonstrated that -glucan promotes histone lactylation directly from glucose or exogenous lactate (Figs. 5d-5g). In addition, β-glucan treatment increased histone Kla levels in monocytes but not in monocytes (Fig. 5h and Supplementary Fig. 5h). These findings suggest high intracellular lactate levels in trained immunity-regulated histone lactylation.
Lactate maintains chromatin accessibility via histone lactylation in monocytes during the induction of trained immunity
To investigate the mechanical effect of lactate production on trained immunity, we performed the Assay for Transposase-Accessible Chromatin with high throughput sequencing (ATAC-seq) analysis to compare chromatin region accessibility changes between monocytes, , and trained monocytes. We identified 2,848 chromatin regions exhibiting accessibility changes (Fig. 6a). To evaluate the functional impact of lactate on transcription, we performed RNA-seq in monocytes, , and trained monocytes in biological duplicates (Fig. 6a). We first explored the relationship between changes in chromatin accessibility and gene expression. We assigned each
differentially accessible regions (DARs) to its nearest transcription start site and examined the gene expression differences. We observed significant correlations between changes in chromatin accessibility and expression of the neighboring gene (Fig. 6b). Next, we identified 836 genes that were upregulated in monocytes, , and trained monocytes, which were also marked with chromatin
Fig. 3 | Lactate is a physiologic fuel for trained immunity. a-f Monocytes were cultured in MM containing -glucose ( 5 mM ), indicating unlabeled lactate concentration, and treated with -glucan ( ) for 4 hrs , followed by mass spectrometry analyzes. g Schematic of derivation and contribution of carbon atoms in SAM synthesis. Experiments were performed as described in (a). -glucose labeling into ATP or SAM is shown. i Monocytes were treated with -glucan and lactate for 30 min ; mitochondrial ATP was measured by the mtAT 1.03
probe. Monocytes were treated with or without -glucan and lactate for 12 hrs , then acetyl-CoA, citrate, and malate levels were measured. Monocytes were cultured in MM containing -lactate ( 1 mM ), indicating unlabeled glucose concentration, and treated with -glucan ( ) for 4 hrs , followed by mass spectrometry analyzes. Data in ( and ) are expressed as means ± SEMs, mice per condition, two-way ANOVA. See also Supplementary Figs. 3.
Fig. 4 | Ldha influences the metabolism and function of trained immunity. a-c Monocytes from Wt and Ldha// mice were cultured in MM containing -lactate and treated with -glucan for 12 hrs , followed by mass spectrometry analyzes. d Monocytes from and mice were cultured in lactate media ( 0.5 mM glucose, 0.5 mM glutamine, and 2 mM lactate), and treated with glucan for 30 min , mitochondrial ATP was measured by the mtAT 1.03 probe. e Monocytes from and mice were treated with or without -glucan and
lactate ( 1 mM ) for 12 h , followed by measuring acetyl-CoA, citrate, and malate levels. f, g C57BL/ 6 mice were trained with or without -glucan. Six days later, mice were infused with -lactate for 12 hrs , followed by mass spectrometry analyzes. h, i and mice were trained with or without -glucan. Six days later, mice were infused with -lactate for 12 hrs ; monocytes were isolated and followed by mass spectrometry analyzes. All data are expressed as means ± SEMs, mice per condition, two-way ANOVA. See also Supplementary Fig. 4.
accessibility alterations (Fig. 6c). Gene ontology (GO) biological process analysis of these genes revealed significant enrichment of biological pathways, including immune response, lactate metabolism, and HIF signaling (Fig. 6d). We choose H3 lysine 27 (H3K27la), this lysine residue is also acetylated ( H 3 K 27 ac ), to elucidate the potential functional significance of histone lactylation in trained immunity by using CUT&Tag, a novel genome-wide immunotethering assay. Genomewide distribution analysis reveals that H3K27la and H3K27ac histone modifications were located mainly within the promoter regions (Fig. 6e). As anticipated, H3K27la peaks were increased near
transcription start sites (TSSs) in -trained monocytes, but not trained monocytes (Fig. 6f). However, H3K27ac peaks were slightly increased near TSSs in -trained monocytes compare with non-trained monocytes (Supplementary Fig. 6a). We compared the expression of H3K27la-marked genes in non-trained monocytes, and trained monocytes. Among the total 7,547 shared peaks between the three groups, we obtained 248 peaks that were significantly changed (Fig. 6g). GO pathway analysis revealed that biological processes such as inflammatory response, lactate metabolism, and cell adhesion, which is consistent with ATAC-seq data (Fig. 6h). To
Fig. -glucan regulates histone lactylation via Ldha. a, b Monocytes were treated with or without -glucan for the indicated time before western blotting. Quantification of the immunoblots is shown on the right panel. c Monocytes were treated with or without -glucan at indicated concentrations for 24 hrs before western blotting. Quantification of the immunoblots is shown on the right panel. d, e MS/MS spectra of a -glucose labeled histone Kla peptide and its unlabeled counterpart from -glucan treated monocytes. f, Experiments were
performed as described in (d,e), except -lactate was used. Monocytes from Wt and mice were treated with or without -glucan for 24 hrs before western blotting. Quantification of the immunoblots is shown on the right panel. All experiments were repeated at least three times. Data in (a-c and h) are expressed as means SEMs, mice per condition, two-way ANOVA. See also Supplementary Fig. 5.
test the role of H3K27la and H3K27ac in the function of trained monocytes, we integrated our ATAC-seq datasets and CUT&Tag datasets into the Integrative Genomics Viewer to examine their binding profiles on the genes. As shown in Supplementary Fig. 6b, H3K27ac peaks were slightly enhanced at the promoters of IL-6 and TNF- in Wttrained monocytes compared with non-trained monocytes. However, H3K27la peaks were highly enriched at the promoters of critical genes of inflammatory factors and cell adhesion molecules, including
IL-6, TNF- , PSGL, and ICAM-1, in -trained monocytes, but not in trained monocytes (Figs. 6i, 6j and Supplementary Figs. 6c and 6d). We then used a chromatin immunoprecipitation (ChIP)-qPCR assay to assess the effects of H 3 K 27 la and H 3 K 27 ac on the stabilized effector gene expression. Indeed, H3K27la recruitment to the promoter regions of IL-6, TNF- , PSGL, and ICAM-1 was increased in -trained monocytes but not in trained monocytes (Fig. 6k). Only minor differences existed between the from the H3K27ac
recruitment to the promoter regions of IL-6, TNF- , PSGL, and ICAM-1 in -trained monocytes compare with the non-trained monocytes (Fig. 61). The role of histone modifications on the activation of the promoter is not restricted to one lysine, as similar results were obtained by H3K18la and H3K18ac (Supplementary Figs. 6e and 6f). In line with these observations, blocking CBP/p300 histone
acetyltransferase (HAT) activity with HAT inhibitor (HATi) also decreased H3K27la and H3K18la interaction with the promoter of IL-6, TNF- , PSGL, and ICAM-1 (Supplementary Figs. 6g and 6h). These findings suggest that intrinsic lactate production alters acetylationdependent epigenetic programming at the promoter of inflammatory factors and cell adhesion molecules in trained monocytes.
Fig. 6 | Lactate regulates histone lactylation via Ldha in monocytes during induction of trained immunity. a Heatmap representing the differentially accessible regions (DARs) of ATAC-seq (left panel) and differential gene expression of RNA-seq (right panel) in monocytes from monocytes, trained monocytes, and trained monocytes trained as described in Fig S1b. b DARs of ATAC-seq and differential gene expression of RNA-seq subjected to Pearson’s correlation analysis. Correlation coefficient ( ) and values were calculated by Pearson’s correlation analysis. c Venn diagram depicting the number of candidate genes in ATAC-seq and upregulated genes in RNA-seq of monocytes, trained monocytes, and trained monocytes. d GO analysis of significantly different genes in (c). The most significant and nonredundant biological processes are shown, with respective gene numbers and values. values by two-tailed Student’s t-test. e Pie chart showing the distribution of H 3 K 27 la and H 3 K 27 ac at annotated genomic regions in trained monocytes. f Heatmap showing the genomic occupancy of
H3K27la flanking TSSs in trained monocytes from and mice. The genes shown in rows are sorted in descending order by signal strength. g Heatmap of chromatin peak accessibility for significantly different peaks in trained monocytes from and mice. Each row represents a Z score of log2-transferred normalized read counts within each sample using CUT&Tag-seq. h GO analysis of significantly different genes in (g). Most significant and nonredundant biological processes with respective gene numbers and values are shown. values two-tailed Student’s t-test. i, j Representative ATAC-seq and CUT&Tag-seq genome browser tracks for H3K27la peak enrichment at IL-6 and TNF-a gene loci for trained monocytes from and mice. k, Monocytes from and mice were trained as described in Fig S1b, followed by ChIP analysis. Data in ( and ) are expressed as means SEMs, mice per condition, two-way ANOVA. See also Supplementary Fig. 6.
Exogenous lactate supports trained immunity through contribution to the TCA cycle and histone lactylation
Intracellular lactate can be produced by LDHA or imported by monocarboxylate transporter 1 (MCT1; also known as SLC16A1) . Thus, we sought to explore the effect of MCT1 on the trained immunity. As shown in Supplementary Fig. 7a, IL-6, and TNF- production was induced in -glucan-trained mice, but not in MCT1 inhibitor AZD3965 (AZD) treated mice. In this vein, AZD-treated mice were significantly susceptible to C. albicans-induced lethality, with higher bacterial burden in the lung and kidney tissue (Supplementary Figs. 7b and 7c). Next, we evaluate the role of MCT1 on the lactatefueled TCA cycle by treating trained monocytes with -lactate. As shown in Supplementary Figs. 7d-7f, AZD treatment abolished lactate entry into the TCA cycle. As expected, -glucan treatment increased histone Kla levels in monocytes but not in AZD-treated monocytes (Supplementary Fig. 7g). ChIP-qPCR assay indicated that AZD treatment inhibit H3K27la and H3K18la recruitment to the promoter regions of IL-6, TNF- , PSGL, and ICAM-1 (Supplementary Figs. 7h and 7i). Hence, MCT1-mediated lactate uptake service as a TCA cycle substrate regulated cytokine production via histone lactylation.
Our data revealed that -glucan mildly induced histone acetylation. We hypothesized that lactate is taken up and readily converted to pyruvic acid through LDHA and subsequently converted to cytosolic acetyl-CoA in two enzymatic catalyzed reactions by pyruvate dehydrogenase (PDHA) and ATP-citrate lyase (ACLY), respectively (Fig. 7a). To this purpose; we compare the role of glucose and lactate on acetyl-CoA generation. As shown in Fig. 7b, neither high glucose (HG) nor low glucose (LG) affect -glucan-regulated acetyl-CoA gen-eration-however, lactate treatment induced -glucan regulated acetyl generation, indicating that -glucan promotes acetyl-CoA generation via lactate. Still, not glucose (Fig. 7b). To dissect the key step of lactate metabolism involved in acetyl-CoA generation, we employed a series of pharmacologically targeting LDHA, PDHA and ACLY by the LDHA inhibitor (sodium oxamate, Oxa), pyruvate transporter inhibitor (UK5099), and ACLY inhibitor (BMS303141, BMS), respectively (Fig. 7a). Interesting, treatment of Oxa, UK5099, and BMS inhibit glucan induced acetyl-CoA generation (Fig. 7c). LC/MS analysis focused on isotopologues of acetyl-CoA demonstrate that lactate carbons labeled the majority of acetyl-CoA. The acetyl-CoA isotopologue revealed the most substantial labeling pattern derived from lactate carbons (Figs. 7d and 7e). To pinpoint the pathway in how lactate facilitates histone acetylation, we hypothesized that this involves oxidative metabolism, particularly ACLY. Strikingly, BMS counteracted -glucan-mediated histone acetylation but not histone lactylation (Fig. 7f). Surprising, the ACLY inhibitor did not affect -glucan-mediated IL-6 and TNF- production, and bacterial burden in the lung and kidney tissue in vitro and in vivo (Figs. 7g-7i). In addition, BMS treatment inhibits H3K27ac and H3K18ac, but not H3K27la and H3K18la, recruitment to the promoter regions of IL-6, TNF- , and PSGL (Figs. ). These results suggest that lactate is a significant substrate
for acetyl-CoA production. Compared with lactate-mediated histone acetylation, lactate-mediated histone lactylation plays a dominant role in trained immunity.
Discussion
Trained immunity refers to the long-lasting memory traits of innate immunity . How environmental nutrient availability impacts trained immunity still needs to be better understood. Here, for the first time, we show that trained monocytes exhibit a high degree of plasticity in fuel choice, not only conventional fuels such as glucose but also PCSs. Exploring this, we demonstrated that cell-intrinsic lactate is a prominent fuel in trained monocytes, contributing to the TCA cycle, even when glucose is available. Mechanistically, lactate was a significant substrate for acetyl-CoA production in trained monocytes and regulated trained monocytes’ function through effects on histone lactylation. Our study highlights a previously underappreciated role of lactate in trained immunity.
Immunological memory has long been ascribed only to adaptive T and cells . The recently emerged concept of trained immunity is defined as causing long-term alterations in innate immune cells, enabling a robust innate host response to a secondary stimulus . Analysis of differentially regulated pathways in trained monocytes demonstrated an essential role in changes in intracellular metabolism, especially glucose metabolism . The mTOR-HIF1 pathway regulated a shift from oxidative phosphorylation to glycolysis, the central regulatory mechanism for trained immunity . This study observed the high lactate levels, an end product of glycolysis, in trained immunity, but the physiological significance of overexpressed lactate was not explored. We show lactate is a physiologic fuel for trained immunity, serving as a preferred TCA cycle and biosynthetic substrate. Additionally, lactate regulates cytokine production through its effects on histone lactylation. Our results strongly complement the previous work, filling the downstream signaling pathways of trained immunity. We propose a working model of the most prominent regulated pathways in trained immunity based on prior studies and our present data. According to this model, trained innate immune cells activate the Akt-mTOR-HIF-1 pathway, leading to a shift in metabolism from oxidative phosphorylation to glycolysis and high lactate accumulation. A portion of elevated lactate levels serves as a physiologic fuel for the TCA cycle. The other part of elevated levels of lactate enters into the nucleus and binds the loci of cytokines promoter, leading to maintaining the accessibility of chromatin. If trained innate immune cells encounter a secondary stimulus, accessible chromatin permits transcription factors to quickly bind to the corresponding promoter, leading to a faster and more robust response.
Recently, some research has revealed the link between the stimulation of innate immune pathways and the induction of epigenetic and metabolic changes in trained immune cells . Rob et al. show that glutaminolysis is another metabolic pathway involved in -glucan induced trained immunity, linked to enrichment in H3K4me3, which is
essential for trained immunity . However, there was no detectable difference in the expression of methyltransferases between nontrained and trained immune cells. Siroon et al. show that the cholesterol/mevalonate synthesis pathway is elevated in -glucan-trained mice, which is linked to enrichment in . However, the entire genome H3K27ac study does not reveal a complete overlap between –
glucan and mevalonate-induced trained immunity. We suspect part of this phenomenon is due to the cell culture medium. The current cell culture medium is designed to promote cell growth and survival. Thus, the current cell culture medium contains supraphysiological concentrations of some nutrients while absenting other nutrients relevant to cellular metabolism. An important implication of this study is that
Fig. 7 | Exogenous lactate supports trained immunity through contribution to the TCA cycle and histone lactylation. a Schematic of -lactate carbon labeling through the TCA cycle. b Monocytes were cultured with high glucose (HG) ( 15 mM ), low glucose (LG) ( 3 mM ), and/or lactate ( 1 mM ), and treated with or without glucan for 24 hrs , followed by measuring acetyl-CoA levels. c Monocytes were cultured in lactate media ( 0.5 mM glucose, 0.5 mM glutamine, and 2 mM lactate) and treated with or without -glucan ( ), sodium oxamate ( 20 mM ), UK5099 , BMS303141 (BMS) for 24 hrs , followed by measuring acetyl-CoA levels. d, e Monocytes were cultured in MM containing -lactate and treated with -glucan for 4 hrs , followed by LC/MS analysis. Shown are labeled and unlabeled fractions of acetyl-CoA. f Monocytes were cultured in lactate media ( 0.5 mM glucose, 0.5 mM glutamine, and 2 mM lactate) and
treated with or without -glucan , sodium oxamate , UK5099 , BMS for 24 hrs before western blotting. Quantification of the immunoblots is shown on the right panel. Experiments were repeated at least three times. g Monocytes were cultured in lactate media ( 0.5 mM glucose, 0.5 mM glutamine, and 2 mM lactate), treated with or without -glucan ( ), BMS ( ) for 24 hrs , and then IL- 6 and TNF- levels were measured. h, i mice were trained with or without -glucan. Six days later, mice were infected with or without a secondary C. albicans lethal dose ( ) and/or BMS ( ) for 24 hrs , followed by measuring IL-6 and TNF- levels ( ) and fungal burden (i). Monocytes from mice were treated as described in (g), followed by ChIP analysis. Data are expressed as means ± SEMs, mice per condition, two-way ANOVA. See also Supplementary Fig. 7.
glucose utilization by trained monocytes is highly influenced by other non-glucose substrates, precisely a few of the nutrients absent from most medium formulations today. Our study established that lactate is a primary source of acetyl-CoA production in trained monocytes. This directly contributes to histone lactylation and promotes permissive lysine lactylation in histones at cytokines gene loci. Mechanistically, we show that LDHA, which controls lactate metabolism via regulating lactate/pyruvate conversion, is required for a lactate-fueled TCA cycle and regulated histone lactylation. Our study demonstrates that lactate is the hub between metabolism, immunity, and epigenetics in trained monocytes.
It has yet to slip our attention that there are several important caveats. (i) The distinctions and linkages of histone lactylation, histone methylation, and histone lactylation in trained immunity still need to be explored. (ii) We only focus on the role of higher concentration (exceeding ) PCS on the glucose utilization of trained immunity. Lower concentration PCS may also serve as bioenergetic fuels in trained immune cells when the nutrients are unavailable. (iii) We did not explore whether lactate metabolism is an effective target in clinical treatment. (iv) The direct causal effect of lactylation on trained immunity is challenging to demonstrate, as p300 inhibitors also block histone acetylation. (v) LDHA knockout has a significant impact on survival in this model, irrespective of -glucan treatment (Fig. 2e). This likely arises from the critical role of LDHA in supporting glycolytic responses to infection of all innate immune cells (in the absence of immune training). (vi) Comparison of histone accessibility between WT and LDHA cells to infer lactate-mediated changes (without lactatetreated controls) is challenging, because loss of LDHA will significantly alter the metabolic and functional status of these cells (for example by limiting glycolytic flux and NAD/H redox) in addition to its role permitting lactate generation and subsequent usage. When considering the next step, studies exploring these questions would greatly help clarify the role of lactate in trained immunity.
Methods
Ethics statement
Consistent with the Helsinki Declaration, the Institutional Review Board of Wuhan University approved the collection of clinical samples following guidelines for protecting human subjects. All study participants provided written informed consent for collecting samples and subsequent analyzes.
All animal experiments followed the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The protocols and procedures were approved by the Institutional Animal Care and Use Committee of Wuhan University (project license WDSKY0201302).
Monocyte and peripheral blood mononuclear cell isolation
Peripheral blood mononuclear cells (PBMCs) were isolated from the blood of healthy donors by standard Ficoll gradient centrifugation and cultured in RPMI 1640 without FBS and antibiotics. Percoll (SigmaAldrich) isolation of monocytes was performed as previously described . Briefly, PBMCs were layered on top of a
hyper-osmotic personal solution ( Percoll, , and 0.16 M NaCl ) and centrifuged for 15 min at . The interphase layer was isolated, and cells were washed with cold Phosphate-Buffered Saline (PBS).
Western blot analysis
Western blot analyzes were performed as previously described . Briefly, cells were harvested by low-speed centrifugation and washed with PBS. Cells were lysed in RIPA Buffer (Cell Signaling Technology, Boston, MA, US), and protein concentrations were determined using BCA assays. Forty micrograms of each protein sample were separated using 12% SDS-PAGE and transferred to nitrocellulose membranes (BioRad). Membranes were blocked with Tris-buffered saline with Tween 20 (TBST) and non-fat milk for 1 hour at room temperature. Then, membranes were incubated with primary antibodies overnight at . Subsequently, blots were incubated with horseradish peroxidase-linked secondary antibodies (Jackson ImmunoResearch) for an additional 1 hour. Immunoreactive bands were visualized using an enhanced chemiluminescence system (GE Healthcare).
Cytokine measurements
Cytokines were determined in PBMC culture supernatants by a multiplex Procartaplex assay (Thermo Fisher Scientific, USA) and acquired on a Luminex 200 System (Thermo Fisher Scientific, USA) according to the manufacturer’s protocol.
Samples were cross-linked in suspension for 5 min in 10 mL of formaldehyde (Pierce, #28906) in PBS at room temperature. Nuclei were separated using the cell nucleus isolation kit (Thermo Fisher Scientific, #78835). Lysates were diluted to 500 mL by the addition of binding buffer ( 25 mM HEPES pH 7.5, of RNAse A/T1 (Thermo Fisher Scientific, #EN0551) was added, and the sample was incubated at for 25 min . Then, was added to a final concentration of 40 mM , followed by 75 U of micrococcal nuclease (Worthington Biochemical), and incubated at for 5 min . MNase was quenched by adding 40 mM EDTA, and the total volume was brought to 1.2 mL with binding buffer. Next, insolubilities were removed by centrifugation at max speed ( ) at for 5 min , and the supernatant containing soluble chromatin was collected. At this stage, 5 mL of chromatin was measured using the Qubit dsDNA HS Assay Kit (Invitrogen, #Q32851).
Samples were diluted with binding buffer to ensure similar chromatin concentrations and to match IP conditions. 50 mL of chromatin was set aside as input for each sample. For each IP, 25 mL of Protein A-coated magnetic beads (Invitrogen, #10008D) were washed once with binding buffer and incubated with either , or 10 mL of H3K27Ac antibody. The total volume of bead plus antibody was brought to 200 mL using the binding buffer and was rotated at room temperature for 30 min . To compare post-translational modifications between samples, we performed one biological replicate of all samples on the same day. We made a master mix of bead plus antibodies,
scaling up all components by the number of samples. Buffercontaining antibody was removed, and bead plus antibody was resuspended in 200 mL of soluble chromatin followed by 30 min rotation at room temperature. Unbound chromatin was removed, and beads were vortexed with 500 mL of binding buffer. Buffer was removed, and bound material was eluted from beads by vortexing in 133 mL of elution buffer. At this time, the input was brought to 133 mL by adding 83 mL elution buffer. Proteinase K (Invitrogen, #25530015) was added to a final concentration of 15 mM , and the sample was incubated overnight at .
The following morning, each DNA sample was purified using a MinElute PCR Kit (Qiagen, #28004) and eluted in 30 mL of Buffer EB. 5 mL of DNA was quantified by Qubit dsDNA HS Assay Kit. The remaining 25 mL of DNA was frozen at until library preparation. Libraries were sequenced using a NextSeq 500 (Illumina) using 75 bp paired-end sequencing ( reads/sample for IPs, reads/ sample for input). Next-generation sequencing (NGS) data were aligned to the mm 10 genome. Bed files generated by the NGS alignment were processed using the latest siQ-ChIP release (https://github. com/BradleyDickson/siQ-ChIP), and siQ-ChIP quantification was performed. Responses were computed automatically by the siQ-ChIP software as the area ratio under overlapping peaks for any pair of tracks being compared. Individual ChIP tracks were visualized using IGV.81.
CUT&Tag sequencing analysis
The library preparation for CUT&Tag was performed as previously reported . In brief, cells were washed twice with PBS, concanavalin A-coated magnetic beads (Bangs Laboratories, BP531) in Wash Buffer Hepes, Spermidine, and protease inhibitor Cells were incubated with of the activated mixture (Roche)] for 10 min at room temperature. Cell-bound beads were collected and resuspended with Dig-Wash Buffer ( 20 mM Hepes pH 7.5, Spermidine, protease inhibitor mixture, and digitonin) containing 2 mM EDTA, , and a 1:50 dilution of the primary antibody, and incubated at overnight. A secondary antibody diluted at 1:100 in of Dig-Wash Buffer was then administered into the beads and incubated for 60 min at room temperature following primary antibody removal with a magnet stand (Vazyme, CM101). The pG-Tn5 adapter complex was prepared according to the manufacturer’s instruction with Hyperactive pG-Tn5 Transposase for CUT&Tag (Vazyme, S602). Standard segmentation and amplification were performed as reported previously . Amplified DNA libraries were purified with VAHTS DNA Clean Beads (Vazyme, N411) and shipped for next-generation sequencing (NGS) sequencing by Annoroad Gene Technology. Reads were filtered and mapped to the mm10 genome, and peaks were then called to generate a peak matrix with a pipeline similar to the ATAC-seq analysis mentioned above. Differential peaks were identified by , and normalized read counts .
ATAC sequencing analysis
The ATAC sequencing was performed as previously reported . In brief, cells were lysed with lysis buffer ( 10 mM Tris-HCl, pH , and Igepal ) at for 10 min . Segmentation and amplification were done following the manufacturer’s instructions with TruePrep DNA library Prep Kit V2 (Vazymes, TD501). Annoroad Gene Technology performed single-end sequencing, and clean reads with trimmed adapters were aligned to the mm 10 reference genome with the Bowtie2 (2.3.5.1) package . Broad peaks were called by the Macs2 (2.1.2) package using the parameter (-nomodel-shift-100-extsize 200 -B-broad), and differential accessed peaks were called using the Deseq2 (v.1.20.0) package . Differential accessed peaks were called by and and normalized read counts . Motif enrichment analysis and peak-
associated gene annotation were performed using HOMER (v4.11.1) using peaks filtered by and normalized read counts . Browser tracks were visualized by IGV Browser (v2.8.2) after normalizing the read from each sample to its library size. Ataqv (v1.0.0, https://github.com/ParkerLab/ataqv) package developed by the Parker Laboratory from the University of Michigan was used to perform the ATAC-seq data quality control analysis.
Mice
Wild-type C57BL/6 mice were purchased from Sanxia University (Yichang, Hubei). Ldha mice on a C57BL/6J background were previously described . Littermates of the same sex were randomly assigned to experimental groups. Mice were randomly allocated to experimental groups, and the investigator was blinded to the group allocation during the experiment. 6-8 weeks-old mice were treated with tamoxifen ( in corn oil) or coin oil by intraperitoneal injection. According to protocols provided by the Shanghai Model Organisms Center (Shanghai, China) for inducible Cre driver lines, LDHA exon2 deletion was confirmed by genotyping 10 days after the final injection. All the analyzes were performed blindly. Ldha mice were randomly allocated into experimental groups for further treatment, and cell samples were assigned based on the genotype of interest. All mice were bred under specific pathogen-free conditions in the accredited animal facility at Wuhan University.
Bacterial culture
Mycobacterium tuberculosis (Mtb) H37Rv were grown in 7H9 broth (BD) supplemented with glycerol (Wisent), 0.05% Tween80 (Fisher), and albumin-dextrose-catalase under constant shaking at . For in vitro and in vivo experiments, Mtb (H37Rv) bacteria in log growing phase (OD 0.4-0.9) were centrifuged ( ) and resuspended in RPMI without penicillin/streptomycin or sterile PBS.
Formaldehyde was added to the culture medium to a final concentration of . The cells were then washed twice with PBS, scraped, and lysed in lysis buffer ( SDS, 10 mM Tris-HCl [pH 8.0], 10% protease inhibitor cocktail, each of aprotinin and leupeptin) for 10 min at . The lysates were sonicated on ice, and the debris was removed by centrifugation at for 15 min at . Onefourth of the supernatant was used as the DNA input control. The remaining sample was diluted 10 -fold with dilution buffer ( SDS, Triton X-100, 1 mM EDTA, 10 mM Tris-HCl [pH 8.0], and 150 mM NaCl ) followed by incubation with antibodies overnight at . Immunoprecipitated complexes were collected using protein A/GSepharose. The pellets were washed four times with dialysis buffer containing 2 mM EDTA and 50 mM Tris-HCl, pH 8.0 . After washing, the precipitates were incubated with an elution buffer ( SDS and 0.1 M ) at room temperature. Supernatants were transferred to clean tubes, and RNase A was added to destroy the bound RNA in the sample. Samples were incubated at for 5 hrs to reverse the formaldehyde cross-links, and DNA was precipitated with ethanol and extracted two times with phenol-chloroform. Finally, pellets were resuspended in Tris-EDTA (TE) buffer and subjected to PCR amplification. Quantitative real-time RT-PCR was performed using a LightCycler 480 (Roche) and the SYBR green system (Applied Biosystems). GAPDH was amplified as an internal control.
HPLC-MS/MS analysis
Mass spectrometry and metabolite identification were performed on methanol and LC/MS-grade water-extracted metabolites. Analyzes were performed using High-Performance Liquid Chromatography, High-Resolution Mass Spectrometry, and Tandem Mass Spectrometry (HPLC-MS/MS). The system consisted of a Thermo
Q-Exactive in line with an electrospray source and an Ultimate 3000 (Thermo) series HPLC consisting of a degasser, a binary pump, and an autosampler outfitted with an Xbridge Amide column (dimensions of and a particle size). The mobile phase A: 20 mM ammonium acetate (pH 9.0), 20 mM ammonium hydroxide, water, and acetonitrile. The mobile phase B: Acetonitrile. The gradient was as follows: ( 2.5 min ); ; with a flow rate of . The capillary of the ESI source was set to , with sheath gas at 45 arbitrary units, auxiliary gas at five arbitrary units, and the spray voltage at 4.0 kV . The top 5 precursor ions were fragmented using the higher energy collisional dissociation cell set to normalized collision energy in MS2. Data acquisition and analysis were carried out by Tracefinder 2.1 software (Thermo Fisher Scientific) and Xcalibur 4.0 software (Thermo Fisher Scientific).
In vivo -tracer infusions
Mice were infused intravenously with -labeled tracers. -lactate infusion rate is for 4 h for a 20 g mouse. Then, serum and lung were isolated and processed for metabolomic analysis.
Quantification of mitochondrial ATP level by flow cytometry
For FRET-based ATP determination, a Novocyte 3000 flow cytometer (ACEA BioSciences) was used to record ERAT fluorescence at channels with Ex/Em filters set as follows: (1) (bandwidth: 45 ). (2) (bandwidth: 30). Mitochondrial ATP levels for individual cells were defined as the ratio of fluorescence intensity of channel 2 (Ex/Em: 405/530) divided by that of channel 1 (Ex/Em: 405/ 445 ), similar to parameters used previously .
Reagents and critical commercial assay kits
The details of critical reagents, antibodies, and commercial assay kits are listed in Supplementary Table 1 and Supplementary Table 2. Unless specified otherwise, all biochemical reagents were purchased from Sigma-Aldrich.
Statistical analysis
Data were obtained from three independent reproducible experiments. Data were expressed as mean ± standard deviations or mean ± the standard error of the mean. Statistical significance was determined using Student’s unpaired two-tailed t-test, one-way or two-way ANOVA multiple comparison tests as indicated in the legend. A value < 0.05 was considered significant and was marked with an asterisk (*).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Stienstra, R., Netea-Maier, R. T., Riksen, N. P., Joosten, L. A. B. & Netea, M. G. Specific and complex reprogramming of cellular metabolism in myeloid cells during innate immune responses. Cell Metab. 26, 142-156 (2017).
Zhou, L. et al. Hepatitis B virus rigs the cellular metabolome to avoid innate immune recognition. Nat. Commun. 12, 98 (2021).
He, Q. et al. MAVS integrates glucose metabolism and RIG-I-like receptor signaling. Nat. Commun. 14, 5343 (2023).
Pan, C., Li, B. & Simon, M. C. Moonlighting functions of metabolic enzymes and metabolites in cancer. Mol. Cell 81, 3760-3774 (2021).
Muir, A., Danai, L. V. & Vander Heiden, M. G. Microenvironmental regulation of cancer cell metabolism: implications for experimental design and translational studies. Dis. Model Mech. 11, dmmO35758 (2018).
Hackett, S. R. et al. Systems-level analysis of mechanisms regulating yeast metabolic flux. Science 354, aaf2786 (2016).
Rossiter, N. J. et al. CRISPR screens in physiologic medium reveal conditionally essential genes in human cells. Cell Metab. 33, 1248-1263.e9 (2021).
Puleston, D. J., Villa, M. & Pearce, E. L. Ancillary activity: beyond core metabolism in immune cells. Cell Metab. 26, 131-141 (2017).
. et al. Interleukin- 6 classic and trans-signaling utilize glucose metabolism reprogramming to achieve anti- or pro-inflammatory effects. Metabolism 155, 155832 (2024).
Peng, M. et al. Aerobic glycolysis promotes T helper 1 cell differentiation through an epigenetic mechanism. Science 354, 481-484 (2016).
Lin, J., Liu, G., Chen, L., Kwok, H. F. & Lin, Y. Targeting lactate-related cell cycle activities for cancer therapy. Semin Cancer Biol. 86, 1231-1243 (2022).
Faubert, B. et al. Lactate metabolism in human lung tumors. Cell 171, 358-371.e9 (2017).
Zhang, W. et al. Lactate is a natural suppressor of RLR signaling by targeting MAVS. Cell 178, 176-189.e15 (2019).
Hui et al. Glucose feeds the TCA cycle via circulating lactate. Nature 551, 115-118 (2017).
Zhang, D. et al. Metabolic regulation of gene expression by histone lactylation. Nature 574, 575-580 (2019).
Chen, A.-N. et al. Lactylation, a novel metabolic reprogramming code: current status and prospects. Front Immunol. 12, 688910 (2021).
Khan, N. et al. M. tuberculosis reprograms hematopoietic stem cells to limit myelopoiesis and impair trained immunity. Cell 183, 752-770.e22 (2020).
Mitroulis, I. et al. Modulation of myelopoiesis progenitors is an integral component of trained immunity. Cell 172, 147-161.e12 (2018).
Kalafati, L. et al. Innate immune training of granulopoiesis promotes anti-tumor activity. Cell 183, 771-785.e12 (2020).
Cirovic, B. et al. BCG vaccination in humans elicits trained immunity via the hematopoietic progenitor compartment. Cell Host Microbe 28, 322-334.e5 (2020).
Netea, M. G. et al. Trained immunity: a program of innate immune memory in health and disease. Science 352, aaf1098 (2016).
Netea, M. G. & van der Meer, J. W. M. Trained immunity: an ancient way of remembering. Cell Host Microbe 21, 297-300 (2017).
Netea, M. G. et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 20, 375-388 (2020).
Cheng, S.-C. et al. mTOR- and HIF- -mediated aerobic glycolysis as metabolic basis for trained immunity. Science 345, 1250684 (2014).
Cantor, J. R. et al. Physiologic medium rewires cellular metabolism and reveals uric acid as an endogenous inhibitor of UMP synthase. Cell 169, 258-272.e17 (2017).
Kaymak, I. et al. Carbon source availability drives nutrient utilization in CD8+ T cells. Cell Metab. 34, 1298-1311.e6 (2022).
Luda, K. M. et al. Ketolysis drives CD8+ T cell effector function through effects on histone acetylation. Immunity 56, 2021-2035.e8 (2023).
Ron-Harel, N. et al. T cell activation depends on extracellular alanine. Cell Rep. 28, 3011-3021.e4 (2019).
Su, H. et al. Glutathione synthesis primes monocytes metabolic and epigenetic pathway for -glucan-trained immunity. Redox Biol. 48, 102206 (2021).
Luengo, A. et al. Increased demand for NAD+ relative to ATP drives aerobic glycolysis. Mol. Cell 81, 691-707.e6 (2021).
Peignier, A. & Parker, D. Trained immunity and host-pathogen interactions. Cell Microbiol 22, e13261 (2020).
Zhao, Y. et al. In vivo monitoring of cellular energy metabolism using SoNar, a highly responsive sensor for NAD(+)/NADH redox state. Nat. Protoc. 11, 1345-1359 (2016).
Cheng, S.-C. et al. Broad defects in the energy metabolism of leukocytes underlie immunoparalysis in sepsis. Nat. Immunol. 17, 406-413 (2016).
Saeed, S. et al. Epigenetic programming of monocyte-tomacrophage differentiation and trained innate immunity. Science 345, 1251086 (2014).
Farber, D. L., Netea, M. G., Radbruch, A., Rajewsky, K. & Zinkernagel, R. M. Immunological memory: lessons from the past and a look to the future. Nat. Rev. Immunol. 16, 124-128 (2016).
Netea, M. G., Quintin, J. & van der Meer, J. W. M. Trained immunity: a memory for innate host defense. Cell Host Microbe 9, 355-361 (2011).
Arts, R. J. W. et al. Glutaminolysis and fumarate accumulation integrate immunometabolic and epigenetic programs in trained immunity. Cell Metab. 24, 807-819 (2016).
Bekkering, S. et al. Metabolic induction of trained immunity through the mevalonate pathway. Cell 172, 135-146.e9 (2018).
Repnik, U., Knezevic, M. & Jeras, M. Simple and cost-effective isolation of monocytes from buffy coats. J. Immunol. Methods 278, 283-292 (2003).
Yu, H. et al. Major vault protein promotes hepatocellular carcinoma through targeting interferon regulatory factor 2 and decreasing p53 activity. Hepatology 72, 518-534 (2020).
Kaya, H. S. et al. CUT&Tag for efficient epigenomic profiling of small samples and single cells. Nat. Commun. 29, 1930 (2019).
Buenrostro, J. D., Giresi, P. G., Zaba, L. C., Chang, H. Y. & Greenleaf, W. J. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position. Nat. Methods 10, 1213-1218 (2013).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357-359 (2012).
Zhang, Y. et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 9, R137 (2008).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Xie, H. et al. Targeting lactate dehydrogenase-a inhibits tumorigenesis and tumor progression in mouse models of lung cancer and impacts tumor-initiating cells. Cell Metab. 19, 795-809 (2014).
Imamura, H. et al. Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encodedindicators. PNAS 106, 15651-15656 (2009).
Vishnu, N. et al. ATP increases within the lumen of the endoplasmic reticulum upon intracellular release. Mol. Biol. Cell 25, 368-379 (2014).
Acknowledgements
This work was supported by the National Key Research and Development Program of China (grant numbers: 2021YFC2701800, 2021YFC2701804, 2023YFC2307800), the National Natural Science Foundation of China (grant number U22A2O335), the Fundamental Research Funds for the Central Universities (grant number: 2042024kf0011, 2042022dx0003), the Natural Science Foundation of Wuhan (grant number: 2024040701010031), the Huxiang high-level talents gather engineering innovation and entrepreneurship talents in Hunan province (grant number: 2021RC5006), the funds from Science and Technology Department of Hubei Province (grant number: 2022CFB624), the Knowledge Innovation Program of Wuhan (grant number: 2022020801010116), the Fundamental Research Funds for the Central Universities (grant number: 2042021kf023).
Correspondence and requests for materials should be addressed to Zhibing Lu, Fubing Wang or Shi Liu.
Peer review information Nature Communications thanks, the anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http:// creativecommons.org/licenses/by-nc-nd/4.0/.
¹Department of Cardiology, Zhongnan Hospital of Wuhan University, Wuhan 430072, China. ²Institute of Myocardial Injury and Repair, Wuhan University, Wuhan 430072, China. Department of Infectious Diseases, The Fifth Medical Center of Chinese PLA General Hospital, Beijing 100039, China. State Key Laboratory of Virology, Modern Virology Research Center, College of Life Sciences, Wuhan University, Wuhan 430072, China. Frontier Science Center for Immunology and Metabolism, Taikang Center for Life and Medical Sciences, Wuhan University, Wuhan 430072, China. The State Key Laboratory Breeding Base of Basic Science of Stomatology (Hubei-MOST) & Key Laboratory of Oral Biomedicine Ministry of Education, School & Hospital of Stomatology, Wuhan University, Wuhan 430072, China. Department of General Surgery, First Affiliated Hospital of Gannan Medical University, Ganzhou 341000, China. Department of Infectious Diseases, Longnan First People’s Hospital, Longnan 341700, China. Shanghai Children’s Medical Center, Affiliated Hospital to Shanghai Jiao Tong University School of Medicine, Shanghai 200240, China. Wuhan Research Center for Infectious Diseases and Cancer, Chinese Academy of Medical Sciences, Wuhan 430072, China. These authors contributed equally: Huanhuan Cai, Xueyuan Chen, Yan Liu. e-mail: luzhibing222@163.com; wfb20042002@sina.com; liushi_liushi@whu.edu.cn