DOI: https://doi.org/10.1016/j.resconrec.2024.107578
تاريخ النشر: 2024-04-08
الميكروبلاستيك في أنظمة المياه العذبة: السلوك الديناميكي وعمليات النقل
معلومات المقال
الكلمات المفتاحية:
نقل التلوث
مياه عذبة
تشتت
خلط
غرق
تجميع
الاحتفاظ
إعادة التعليق
الملخص
تُعتبر النظم البيئية للمياه العذبة وسيلة رئيسية لنقل البلاستيك المستند إلى اليابسة إلى المحيطات. تُظهر جزيئات الميكروبلاستيك (MP) في البيئات المائية العذبة ثباتًا عاليًا ونطاقًا واسعًا من توزيعات الحجم والشكل، مما يجعل حركتها وتوزيعها ومصيرها يختلف بشكل ملحوظ اعتمادًا على الظروف البيئية السائدة. تتطور الخصائص الفيزيائية الأساسية لمختلف بوليمرات البلاستيك باستمرار بمعدلات تفاعل محددة مختلفة بسبب عمليات التجوية المعقدة في البيئة. هذا يغير باستمرار الآليات الأساسية التي تحكم ديناميات الميكروبلاستيك ويزيد من تعقيد مصيرها النهائي في النظم المائية الطبيعية. يقوم هذا البحث بإجراء مراجعة شاملة للسلوك الديناميكي للميكروبلاستيك في النظم البيئية للمياه العذبة، مع التركيز على دراسة عمليات الاستقرار والتجمع والاحتفاظ والتعليق التي تحكم نقلها من المصدر إلى المصب. نقدم نظرة شاملة على الأسس النظرية الرئيسية لسلوك الميكروبلاستيك في التدفقات المحيطة والعوامل المؤثرة الرئيسية (مثل الحجم والكثافة والشكل والتركيب). تسلط نتائجنا الضوء على التفاعلات المعقدة بين سلوكيات الميكروبلاستيك الديناميكية والهيدروديناميكا المحلية وكيمياء المياه، مما يؤدي إلى التطور المستمر للخصائص الفيزيائية والكيميائية للميكروبلاستيك (مثل الحجم والشحنة السطحية) من خلال التفاعلات مع المواد الصلبة المعلقة والمواد العضوية الطبيعية والميكروبات تحت تأثير الضوء والرياح. تشكل هذه الديناميكية تحديات كبيرة في التنبؤ بعمليات نقل الميكروبلاستيك ومصيرها النهائي. يبرز تحليل الفجوات التباين بين النماذج الحالية المعتمدة على ظروف المختبر المسيطر عليها والبيئات الطبيعية المعقدة، مما يدل على الحاجة إلى دراسة سلوك الميكروبلاستيك الديناميكي عبر مجموعة واسعة من الظروف البيئية (مثل محاكاة أنماط التدفق المعقدة وكيمياء المحاليل في المسطحات المائية الحقيقية). هناك حاجة إلى مزيد من البحث لتوسيع الدراسات الميدانية لربط الهيدروديناميكا البيئية بوفرة الميكروبلاستيك وإجراء تجارب متوسطة النطاق تعكس بدقة تأثيرات التجوية وهيدروديناميكا التدفق على سلوكيات الميكروبلاستيك. يعد دمج التجارب الفيزيائية التفصيلية مع أدوات النمذجة العددية أمرًا ضروريًا لفهم شامل للتفاعلات بين مختلف الميكروبلاستيك وتأثيرها العام على البيئة. يسهل ذلك تقييم المخاطر البيئية بشكل موثوق وقوي من أجل التحكم في الميكروبلاستيك وإدارة التلوث.
1. المقدمة
(كول وآخرون، 2011؛ فوزر وآخرون، 2021؛ ستينجر وآخرون، 2021؛ خالد وآخرون، 2021)، ربط آثار الميكروبلاستيك بالتأثيرات البيئية في البيئات البحرية والحد من صحة النظام البيئي ووظيفته وخدماته. هناك الآن تقديرات بشأن مدى وجود البلاستيك البحري في المحيطات والمصادر الرئيسية المساهمة (جامبيك وآخرون، 2015)، مما يزيد الضغط لدفع تغييرات سياسية لتحسين جهود إدارة النفايات. في الجمعية العامة للبيئة التابعة للأمم المتحدة (UNEA) في عام 2022، التزم 170 عضوًا بإنتاج اتفاق دولي ملزم قانونيًا بحلول عام 2024 لإنهاء تلوث البلاستيك. على الرغم من الوعي العام بتلوث البلاستيك والأهمية الكبيرة
2. المنهجية
3. السلوك الديناميكي للجزيئات الدقيقة
3.1. عملية غوص الجزيئات الدقيقة
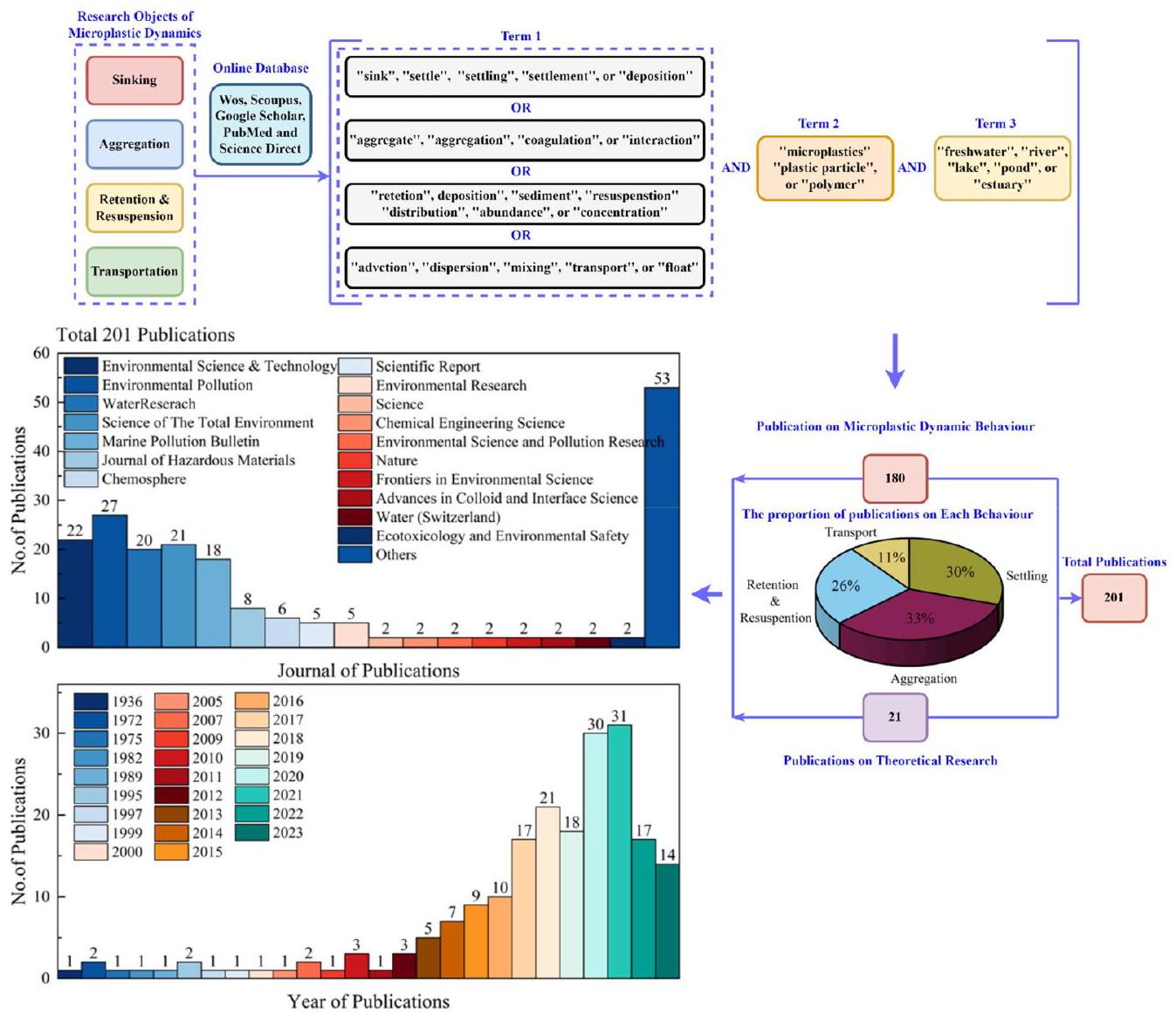
السائل والمسافة التي يتم نقلها بعد إطلاقها من مصادر الانبعاث. وهذا، بدوره، يساهم في فهم أفضل للدور الذي تلعبه أنظمة المياه العذبة مثل الأنهار والبحيرات في نقل الحطام البلاستيكي
من المصادر الأرضية إلى المحيطات (نجوين وآخرون، 2020؛ هوليين وآخرون، 2019).
3.1.1. الخصائص الفيزيائية للجزيئات الدقيقة
العلاقة بين قطر الجسيم ومعدل الاستقرار. يتم التحكم في عملية استقرار الجسيمات الدقيقة في أنظمة التدفق البيئي البطيء جداً أو الساكنة (مثل البرك) من خلال توازن قوة الطفو الجسيمية، والقوة الجاذبية، وقوة السحب. تتناسب شدة القوتين الأوليين بشكل إيجابي مع كثافة الجسيمات وقطرها، بينما تتأثر قوة السحب بشكل أساسي بشكل المقطع العرضي والمساحة السطحية المحددة للجسيمات. وبالتالي، فإن كثافة الجسيمات الأعلى
تحليل عمليات استقرار الميكروبلاستيك والنتائج الرئيسية.
| نوع وشكل البلاستيك | كثافة البلاستيك (
|
حجم البلاستيك (مم) | كيمياء المحاليل | سرعة الاستقرار النهائية (مم/ث) | استنتاج | المراجع |
| حبوب، كرة، ألياف، شظية (PE، EPS، PVC، CoPA) | 1.01-1.31 |
|
ماء مقطر | 3.9-184 | تغيرت سرعات الاستقرار مع تغييرات في حجم الجسيمات وكثافتها وشكلها؛ حيث أن الاستدارة لها تأثير أقل من المعلمات الأخرى. | (والدشلاجلاتر وشوتترومبف، 2019) |
| الميكروبلاستيك، الماكروبلاستيك (PE، PP، PS، PVC، PET) | 0.02-1.5 | 0.58-2.51 5.30 | ماء مقطر | 1.6-35.2 | تؤثر عمليات التجوية والأشكال غير الكروية سلبًا على سرعة استقرار الميكروبلاستيك؛ والصيغة التي تصف استقرار الحبيبات والقطع والميكروبلاستيك الرغوي غير مناسبة للبلاستيكات الرقيقة. | (والدشلاجر وآخرون، 2020) |
| أفلام البوليمر (PE، PS، PET) | 0.92-1.40 | ٤ | ماء منزوع الأيونات | 7.5-28 | لم يتغير معدل استقرار الميكروبلاستيك بشكل كبير بسبب تكوين الأغشية الحيوية؛ كانت بلاستيك PET و PS أكثر ملاءمة للاستعمار مقارنة بميكروبلاستيك PE؛ يمكن أن تسهل الأغشية الحيوية امتصاص المعادن على جزيئات البلاستيك وتحسن من معدل الاستقرار الكلي. | (لايزر وآخرون، 2020) |
| حبيبات أو شظايا (PET، HDPE، PP، PS)، ألياف (PVC)، أفلام (PE) | 0.95-1.432 | 0.63-3.48 | ماء منزوع الأيونات | 5-105 | شكل الجسيمات يؤثر بشكل كبير على عملية الغرق للميكروبلاستيك؛ يمكن أن يعزز التلوث البيولوجي عملية استقرار الجسيمات ذات الكثافة المنخفضة. | (فان ميلكبيك وآخرون، 2020) |
| كرات، شظايا، ألياف (بولي إيثيلين، بولي ستايرين، أكريليك) | 0.95-1.17 | 1-3 | المياه الجوفية | 6.1 (متوسط) | كانت هناك تفاعلات بين تأثير حجم الجسيمات وشكلها وكثافتها على عملية غرق الميكروبلاستيك. | (هويلين وآخرون، 2019) |
| البولي يوريثان غير المنتظم | 1.195 | 0.05-0.50 | مياه نهر مُصفاة | 0-30 | انخفضت سرعة استقرار تجمعات الميكروبلاستيك مع زيادة مستوى الاستعمار الميكروبي. | (نجوين وآخرون، 2020) |
| الميكروبلاستيك غير المنتظم (PA، PMMA، PET) والمستدير (PS) | 1.140، 1.190، و 1.390 | 0.006-0.251 | ماء منزوع الأيونات |
|
كانت سرعة غرق الجسيمات ذات الأشكال غير المنتظمة أقل من تلك الخاصة بالكرات ذات نفس نطاق الحجم؛ يمكن أن تتنبأ تحليل الانحدار التربيعي المتعدد اعتمادًا على حجم الجسيمات وكثافة الجسيمات الزائدة بسرعة غرق الجسيمات الدقيقة النهائية بمتوسط تحديد
|
(كايسر وآخرون، 2019) |
| غير منتظم الشكل (PS) | 1.050 | 0.2-0.9 | مياه الصنبور | 0-2.5 | يمكن أن تؤدي عدم انتظام الشكل فقط إلى تقليل سرعة غرق الجسيمات الدقيقة الأكبر من 0.32 مم؛ العلاقة الإيجابية بين سرعة الاستقرار والحجم غير مهمة مع زيادة عدم الانتظام. | (نجوين، 2021) |
| كرة (PMMA) | 1.190 | 0.150-0.640 | ماء | 0-60 | تتأثر سرعة الاستقرار الصافية بخصائص الجسيمات وخصائص الموجات في الوقت نفسه؛ حيث إن زيادة التأثير الجسيمي وانخفاض رقم رينولدز للجسيمات يزيدان بشكل كبير من سرعة الاستقرار. | (دي ليو وآخرون، 2021) |
| الكرة القريبة والجزء (PS، PA، PET، PVC) | 1.050-1560 | 0.069-3.989 | ماء | 0.85-122.44 | حجم الجسيمات والكثافة يؤثران بشكل إيجابي على سرعة الترسيب؛ بينما تؤثر عدم انتظام الجسيمات، وملوحة المحلول، والظروف الديناميكية بشكل سلبي على ترسيب الجسيمات. | (Z. Wang وآخرون، 2021) |
| قطع غير منتظمة (PS، PA66، PVC، PCL، PLLA، PBAT) | 1.03-1.38 | 0.3-2.2 | مياه البحيرة | 0.30-50 | أحجام الجسيمات وكثافات البوليمر هي العوامل الأكثر أهمية في التحكم في معدلات الاستقرار؛ يؤدي الفيلم الحيوي إلى تحسين طفيف في سرعة الاستقرار بسبب فترات الحضانة القصيرة؛ تزيد خاصية الكارهية للماء للجسيمات من قوى الطفو وتقلل من سرعة الاستقرار. | (إيلاجامي وآخرون، 2020) |
| شظايا وألياف (PET، PVC و NP&A) | 1.01-1.44 | 0.02-4.94 | ماء منزوع الأيونات مع المكالمات | 4-25.6 | كان للبيوفيلم أكبر تأثير على زيادة سرعة الاستقرار للجزيئات الدقيقة غير العائمة بنسبة تصل إلى 130.
|
(مندريك وآخرون، 2023) |
| أقراص (مربعة، مستطيلة، مثلثة) MPs (PVC، PET، PC، ABS) | 1.038-1.343 |
|
مياه الصنبور | 19.6-48.8 | أقراص MP أخف وزناً (
|
(يانغ وآخرون، 2023) |
| الكرات المجهرية (البولي إيثيلين والبوليمر المملوك) | 1.10 و 1.30 | 0.028-0.063 | مياه البحيرة المصفاة | 0.93-2.20 | حجم الجسيمات والديناميكا المائية للبحيرات تؤثر بشكل كبير على زمن إقامة الجسيمات الدقيقة؛ قد يؤدي تجمع الجسيمات الدقيقة مع المواد المعلقة والتفاعلات مع الكائنات الحية في البحيرة إلى زيادة سرعة ترسيب الجسيمات الدقيقة؛ كما أن الحمل المضطرب الناتج عن التغيرات الموسمية في درجة الحرارة يقلل من زمن الإقامة. | (إيلاجامي وآخرون، 2023) |
| نوع وشكل البلاستيك | كثافة البلاستيك (
|
حجم البلاستيك (مم) | كيمياء المحاليل | سرعة الاستقرار النهائية (مم/ث) | استنتاج | المراجع |
| الكرات والقطع (PS) | 1.050 | 0.003 و 0.017 | M4 متوسط |
|
زمن الإقامة للجزيئات الصغيرة (MPs)
|
(غيلفيدير وآخرون، 2023) |
| الألياف (جمع) | 1.308 | 1-4 | مياه الصنبور | 0.1-0.5 | تتأثر سرعة استقرار ألياف MP بشكل كبير باتجاهها وتجعيدها؛ الألياف الطويلة (
|
(نجوين وآخرون، 2022) |
| الألياف (جمع) | 1.308 | 5-15 | ماء | 17-20 | كان استقرار الألياف عملية تعتمد على الاتجاه، وكانت مساراتها متأثرة بشكل عميق بالحركات الثانوية؛ كانت الحركة الثانوية مرتبطة بشكل كبير بأبعاد الألياف وكثافتها؛ وتم اقتراح نموذج سحب جديد لوصف استقرار الألياف. | (تشوي وآخرون، 2022) |
الكثافة:
3.1.2. التلوث البيولوجي
الارتباط، مما يؤدي إلى استجابة أكثر أهمية لزيادة في القطر أو الكثافة الناتجة عن الاستعمار البيولوجي (فازي وريان، 2016). علاوة على ذلك، أشار ريان (2015) إلى أهمية شكل الجسيمات بالنسبة لنسبة المساحة السطحية إلى الحجم، وخاصة بالنسبة للبلاستيكات التي تتجاوز أقطارها 100-200 مم. وهذا يشير إلى أن تأثير التلوث البيولوجي قد يكون أكثر وضوحًا للجسيمات الدقيقة ذات الأشكال المعقدة مقارنة بالجسيمات الكروية ذات نفس الحجم. لتوضيح تأثير التلوث البيولوجي كدالة لحجم وشكل الجسيمات الدقيقة، قام فان ميلكبيك وآخرون (2020) بدراسة دور سمك الفيلم الحيوي (
3.1.3. الحالة البيئية
(دروموند وآخرون، 2022؛ دروموند وآخرون، 2020). بالإضافة إلى ذلك، تعزز التركيزات العالية من الجسيمات المعلقة (مثل الطين والرمل) في الأنهار التفاعلات مع الأفلام الدقيقة، خاصة بعد التلوث الحيوي، لتشكيل الكتل، التي تؤثر بشكل كبير على ديناميات الأفلام الدقيقة من خلال تعزيز ترسيب الرواسب وتراكمها (كوركوران وآخرون، 2020؛ غوليزاده وسيرا، 2022). لاحظ لي وآخرون (2022) أن الاضطراب النهري يمكن أن يعزز تكتل الأفلام الدقيقة، خاصة عند معدل قص أعلى من
3.2. عملية التجميع للأفلام الدقيقة
خصائص السطح (مثل الخشونة السطحية) والخصائص الكيميائية (مثل توزيع الشحنة السطحية والجهد الكهروستاتيكي) (Li et al., 2018; Artham et al., 2009; Michels et al., 2018). تشمل المحركات النظرية لتجمع الجسيمات الدقيقة مجموعة من القوى المتفاعلة مثل التفاعل الكهربائي، الروابط الهيدروجينية، الروابط الكيميائية، والجسور (الشكل 4) (Wang et al., 2015). يوضح الجدول 2 تحليل الدراسات الرئيسية حول تجمع الجسيمات الدقيقة في المحاليل العذبة، ويتم تقديم نظرة عامة مفصلة عن الدراسات المعنية هنا في الجدول S2. يتم مناقشة المعلمات الرئيسية والآليات المعنية في الشكل 4 والجدول 2 في الأقسام التالية (القسم 3.2.1-3.2.6).
3.2.1. خصائص الإلكتروليت
أثر الجسر خلال عملية التجميع، مما يجعل البولي إيثيلين أكثر حساسية لـ
3.2.2. المواد العضوية الطبيعية (NOM)
تحليل عمليات تجميع الميكروبلاستيك والنتائج الرئيسية.
| نوع التجميع | حجم البلاستيك (
|
التركيز (ملغ/لتر) | كيمياء المحاليل | الخاتمة (الآليات) | المراجع |
| التجمع الذاتي (PS شبه كروي) |
|
٢٠.٠ | محلول إلكتروليت
|
توافقت حركيات التجميع مع نظرية DLVO وقاعدة شولز-هاردي. | (لي وآخرون، 2018) |
| التجمع الذاتي (جزيئات نانوية وميكروية كروية من بولي ستيرين) | 0.1، 1.0 | 10 | محلول الكهارل
|
تأثرت العملية الفيزيائية بشكل كبير بآلية تجميع الجسيمات الدقيقة من خلال تغيير استقرارها في المياه العذبة؛ بينما لعبت التركيز، والشحنة الأيونية، والقدرة على الترطيب دورًا رئيسيًا في عملية تجميع الجسيمات النانوية. | (سون وآخرون، 2020) |
| التجمع الذاتي (جزيئات PS) | 0.1 | 10-50 | محلول الإلكتروليت
|
فقط إلكتروليت
|
(Cai وآخرون، 2018) |
| التجمع الذاتي (جزيئات PE و PS) | 0.2-0.75 | 20 | محلول الإلكتروليت (NOM،
|
كان لنوع الملح وقوة الأيونات تأثيرات كبيرة على استقرار البلاستيك النانوي وبالتالي على تركيز التجلط الحرج له؛ حيث اتبعت خصائص تجميعه واستقراره نظرية DLVO وقاعدة شولز-هاردي. | (شمس وآخرون، 2020) |
| التجمع الذاتي (جزيئات PS) | 0.09 | محلول الكهارل (NOM، NaCl، CaCl2، Na2SO4) | كانت تجميع جزيئات PS مهيمنة بواسطة التفاعلات الكهروستاتيكية والنبذ الستيري؛ ولعبت المواد العضوية الطبيعية ودرجة الحموضة الدور الأكثر أهمية في المحاليل ذات القوة الأيونية العالية والمنخفضة ومحتوى المواد العضوية الطبيعية بشكل منفصل. | (ج. وانغ وآخرون، 2021) | |
| التجمع غير المتجانس والمتجانس (جزيئات النانو من بولي ستايرين وجزيئات الميكرو من بولي إيثيلين مع SS) | 0.10، 1000-1200 | 10,160 | محلول الكهارل (HA، NaCl) | أدى امتصاص جزيئات بلاستيك PS على سطح SS الكبير إلى تكوين هيتروأغريغيتس؛ يمكن أن تحسن HA من التنافر الكهروستاتيكي والحواجز الستيرية بين الجزيئات وبالتالي تعيق حدوث التجمع؛ كانت الحركة البراونية وقوة الطفو تهيمنان على حركة PSNPs و PEMPs في الماء على التوالي؛ كانت PEMPs بحجم المليمتر تطفو دائمًا على سطح الماء حتى عندما تم امتصاص سطحها بواسطة SS. | (Y. Li وآخرون، 2019) |
| التجمع غير المتجانس (كرات بولي ستايرين مع جزيئات حيوية) | ٧٠٠-٩٠٠ | 50 خرزة/ل. | مياه البحر | تشكيل الأغشية الحيوية على سطح البلاستيكيات الدقيقة سهل عملية تجمعها و diversified المجتمعات البكتيرية عليها | (ميشيل وآخرون، 2018) |
| التجمع غير المتجانس (قطع HDPE وPP مع الطحالب) | ٤٠٠-١٠٠٠ | ٤٠٠ | وسط الثقافة | تغيرت خصائص سطح قطع البولي بروبيلين والبولي إيثيلين عالي الكثافة نتيجة استعمار الميكرو الطحالب؛ كانت للمواد الخارجية للميكرو الطحالب خصائص تجميع مميزة مع البولي بروبيلين والبولي إيثيلين عالي الكثافة. | (لاجارد وآخرون، 2016) |
| التجمع المتجانس وغير المتجانس (الطحالب الدقيقة، المواد البوليمرية الخارجية، جزيئات PMMA و PS ذات الأشكال غير المنتظمة) | <106, 106-250 | 12.5، 125 | وسط الثقافة | كان التجمع غير المتجانس معتمدًا على إنتاج الغزارة من EPS وحجم ونوع EPS وMPs. | (كونها وآخرون، 2019) |
| التجمع الذاتي (جزيئات PE غير المنتظمة) والتجمع غير الذاتي (جزيئات PE مع الطين الكلوريت، الإيلايت، الكاولينيت، والمونتموريلونيت) | ٢٥.٦ | 20 | ماء منزوع الأيونات مع ظروف هيدروكيميائية مختلفة
|
كانت البوليمرات القديمة أكثر استقرارًا من البوليمرات النقية في محاليل الإلكتروليت أحادية التكافؤ؛ وكان تفاعل البوليمرات والمعادن الطينية يهيمن عليه التنافر الكهروستاتيكي؛ وتم تسهيل التجمع غير المتجانس والترسيب للبوليمرات بزيادة تركيز NaCl. | (وانغ وآخرون، 2023) |
| التجمع غير المتجانس (PP، HDPE، PET، PVC و PS مع الجسيمات النهرية) | 1527-1935 | – | مياه نهر غير مُعالجة | نواب كبار
|
(ستيد وبوند، 2023) |
زعزعة استقرار الميكرو-PS من خلال جذب الجسور والجسور بين الجزيئات
وتجاهلت تنوع المواد العضوية الطبيعية الموجودة في البيئات الطبيعية. بالإضافة إلى الأحماض الهيومينية، فإن المواد الشبيهة بالبروتين، المنتشرة في الأنظمة المائية، تظهر أيضًا تأثيرات تثبيت قوية على جزيئات النانو من PS.
3.2.3. الحل
3.2.4. الخصائص الفيزيائية للميكروبلاستيك
تجمعها من خلال تغيير طاقة تفاعل المستحلب والحركة الكهربائية، مما يؤدي إلى ميل فريد للتجمع في محاليل معينة (أفروز وآخرون، 2013؛ بهاتاشارجي وآخرون، 2000؛ لي وما، 2018). ومع ذلك، حتى الآن، كانت الدراسات التي تركز على تطبيق هذه النتائج على الجسيمات الدقيقة محدودة للغاية، مما يبرز مجالًا حاسمًا للبحث المستقبلي.
3.2.5. الأغشية الحيوية والكتلة الحيوية
نفاذية، أو تم امتصاصها مباشرة على جدران خلايا الطحالب من خلال التفاعلات بين الجزيئات والقوى الامتصاصية. وهذا يؤدي إلى ترسيب سريع للجزيئات الدقيقة من المياه السطحية إلى الرواسب القاعية (نجوين وآخرون، 2020؛ لاجارد وآخرون، 2016؛ بيلر وآخرون، 2021؛ ميشيلز وآخرون، 2018).
3.2.6. التجوية
3.3. الاحتفاظ
3.3.1. الخصائص الفيزيائية للميكروبلاستيك
حجم الجسيمات (Besseling et al., 2017; Nizzetto et al., 2016). علاوة على ذلك، غالبًا ما تظهر الرواسب القاعية انتشارًا للجزيئات الدقيقة ذات الكثافة المنخفضة (مثل PE و PP)، والتي يمكن أن تُعزى إلى القوى القوية بين الجسيمات بما في ذلك جذب الشحنات، التي تحفز التفاعلات مع المعادن، والتلوث البيولوجي، وامتصاص المواد (Corcoran et al., 2015; Ballent et al., 2016; B. He et al., 2020).
تحليل وفرة وخصائص الميكروبلاستيك في رواسب المياه العذبة.
| منطقة المصدر | عمق العينة (سم) | الشكل السائد | الحجم (
|
الكثافة (
|
وفرة (جزيئات/ كجم) | المراجع | ||||||||
| نهر بيجيانغ، الصين | 2 | غير محدد | <5000 | البولي إيثيلين، البولي بروبيلين والكوبيوليمر | 178-544 | (وانغ وآخرون، 2017) | ||||||||
|
|
|
|
0-1.8 | 6350 (قبل الفيضانات) | (هارلي وآخرون، 2018) | ||||||||
|
شظية |
|
التربية البدنية | 165 (ظروف تدفق القاعدة) | (تيبيتيتس وآخرون، 2018) | |||||||||
| 12 |
|
|
100-629 (مارس) |
|
||||||||||
| 0-2 | ألياف، شظايا وكرات | <1000 | PET، PS و PE | 50-195 | (جيانغ وآخرون، 2019) | |||||||||
| أنهار الأمازون، البرازيل | 5-10 | الألياف (فقط) | 0-1000 (رئيسي) | غير محدد | نهر | (جيرولين وآخرون، 2020) | ||||||||
| 0-1000 (رئيسي) | 1524-1738 (نهر سوليمões) | |||||||||||||
| 1000-2000 (رئيسي) | 417-2101 (أنهار الأمازون) | (هو وآخرون، 2020أ) | ||||||||||||
| نهر بريسبان، أستراليا | 0-3 | فيلم، ألياف وقطع | <5000 | البولي إيثيلين، البولي بروبيلين والبولي ستايرين | 10-270 (ديسمبر 2017) | |||||||||
| سطح | شظية وألياف | <5000 | البولي إيثيلين والبولي بروبيلين | 6-2444 (مُعَدل) | (كوركوران وآخرون، 2020) | |||||||||
| نهر التايمز، كندا | ٥ | شظية وفيلم | 1000-5000 | بولي إيثيلين، بولي بروبيلين، بولي فينيل الكحول، بولي فينيل كلوريد | 8-1100 | (كبير وآخرون، 2022) | ||||||||
| بحيرة أونتاريو، كندا | <8 | غير منتظم، رقيق | 250-1000 (رئيسي) | PE، PP و NC | 4635 جزيئًا | (كوركوران وآخرون، 2015) | ||||||||
| بحيرة تايهو، الصين | سطح | ألياف | 100-1000 (رئيسي) | سيلوفان | 11.0-234.6 | (سو وآخرون، 2016) | ||||||||
| بحيرة فيمباناد، الهند | سطح | فيلم ورغوة | 100-500 (رئيسي) | البولي إيثيلين عالي الكثافة، البولي إيثيلين منخفض الكثافة، البولي بروبيلين، البولي ستيرين | 252.8 جزيئات
|
(سروثي وراماسامي، 2017) | ||||||||
| بركة إدجباستون، المملكة المتحدة | سطح | الألياف والفيلم | <5000 | لا شيء | 250-300 | (فوهان وآخرون، 2017) | ||||||||
| بحيرة دونغتينغ، الصين | 0-2 | ألياف | <500 (شائع) | بي إس، بي إي تي | ٢٠٠-١١٥٠ | (جيانغ وآخرون، 2018) | ||||||||
| بحيرة بويانغ، الصين | سطح | الألياف والفيلم | 100-500 (رئيسي) | البولي بروبيلين والبولي إيثيلين | 54-506 | (يوان وآخرون، 2019) | ||||||||
| بحيرة تايهو، الصين | الأعلى، بواسطة بيترسون، امسك | شظية، ألياف، فيلم وكرات |
|
بي في سي، بي إي، بي إس، بي بي | 464.26-1380.69، 893.48 (المتوسط) | (Zhang وآخرون، 2021) | ||||||||
| بحيرة هاواسا، إثيوبيا | أفضل 3 | ألياف (90 %)، قطع (5
|
100-500 (رئيسي) | البولي إيثيلين تيريفثاليت (82 %)، البولي إيثيلين (15 %) والبولي ستيرين (3 %) | 11-74 جزيء/م³ | (جيفاناندام وآخرون، 2022) | ||||||||
| مصب نهر اليانغتسي، الصين | 0-2 | ألياف | 1161.23 (المتوسط) | رايون (الأكثر) | 143 | (F. وو وآخرون، 2020) | ||||||||
| مصب ديروينت، أستراليا | 0-104 | الألياف (87 %) | 63-1000 (رئيسي) | غير محدد | 2430، 4200 | (ويليس وآخرون، 2017) | ||||||||
| مصب نهر يانغتسي، الصين | ٥-١٠ | الألياف (93 %) | <1000 (58 %) | رايون، بوليستر، وأكريليك | ٢٠-٣٤٠ | (بينغ وآخرون، 2017) | ||||||||
| رطوبة أنزلي، إيران | سطح | ألياف | 100-200 (رئيسي) | 0.90-1.40 | 140-2820 (يونيو) 110-3690 (يناير) | (راستا وآخرون، 2020) | ||||||||
| غابات المانغروف، الصين | 1 | الألياف (معظمها) والجزء | 500-1000 (الأعلى) | PP، PE و PS | 273-3520 | (L. Zhang وآخرون، 2020) | ||||||||
| الأراضي الرطبة، الولايات المتحدة الأمريكية | ٥ | الألياف (88 %) | <5000 | البوليسترين، البولي إيثيلين والمطاط الصناعي | 1270 (متوسط) | (هيلكوسكي وآخرون، 2020) | ||||||||
| خزان السدود الثلاثة، الصين خليج جياوزhou، الصين | غير محدد | ورقة وخط | 1000-5000 (رئيسي) | البولي بروبيلين، البولي إيثيلين والبولي إيثيلين تيريفثاليت |
|
(Zhang وآخرون، 2017) | ||||||||
| 0-45 | ألياف | 100-490 (رئيسي) | البولي إيثيلين (المجال)، الرايون و PET | 0-30 | (Zheng وآخرون، 2020) |
3.3.2. البيئة المائية
الحطام في مناطق النشاط الهيدرو ديناميكي الضعيف (براون وآخرون، 2010؛ زانغ وآخرون، 2017). لاحظ فيشر وآخرون (2016) نمط احتفاظ مشابه، مشيرين إلى أن وجود المادة العضوية يعزز التماسك والالتصاق بين الجزيئات الدقيقة والرواسب. بالإضافة إلى ذلك، فإن وفرة المادة العضوية تسرع من تشكيل الأغشية الحيوية على أسطح الجزيئات الدقيقة، مما يحولها من كارهة للماء إلى محبة للماء بشكل قوي ويغير قطبيتها بشكل مشابه للرواسب، مما يزيد من ألفتها مع الرواسب (فان ميلكيبيك وآخرون، 2020). قد يؤدي تشكيل هذه الأغشية الحيوية أيضًا إلى تحفيز التحلل الحيوي في الجزيئات الدقيقة المودعة لفترات طويلة، خاصة في طبقات الرواسب الأعمق، مما يؤدي إلى تفكيكها إلى قطع أصغر تتغلغل أكثر في طبقات الرواسب من خلال مساحات المسام (نيوا وآخرون، 2021). أفاد فيرماير وآخرون (2017) بوجود تفاعل تنافسي بين المادة العضوية والديناميكا المائية، حيث تشجع المادة العضوية على تشكيل تجمعات أكبر وأكثر كثافة من الجزيئات الدقيقة التي من المرجح أن تستقر في عمود الماء، مما يقلل من تأثير الديناميكا المائية على سلوك الجزيئات الدقيقة. بالإضافة إلى ذلك، سلطت بعض الأدبيات الضوء على أهمية تدفقات حبيبات الرواسب وتدفقات الضغط في احتفاظ الجسيمات (فرايز وتاغون، 2010؛ فاريزان وآخرون، 2019). ومع ذلك، تتوفر معلومات قليلة حول كيفية تأثير هذه التدفقات المحددة على احتفاظ الجزيئات الدقيقة.
3.4. إعادة التعليق
3.4.1. الخصائص الفيزيائية للميكروبلاستيك
مضطرب مع
3.4.2. الظروف الهيدروليكية
خلال الفيضانات في مجرى مائي صناعي محكوم، مما يبرز الدور الحاسم لتطور الطبقة النشطة وخصائص نظام التدفق في إعادة تعليق الميكروبلاستيك في الأنهار الحبيبية الخشنة. وبالمثل، لاحظ هيرلي وآخرون (2018) أيضًا زيادة ملحوظة في تركيز الميكروبلاستيك في عمود الماء خلال مواسم الأمطار، والتي تعزى إلى زيادة سرعات التدفق التي ترفع المزيد من الميكروبلاستيك المترسب. نظرًا لأن وفرة الميكروبلاستيك عادة ما تكون أعلى في الرواسب السطحية مقارنة بالطبقات الأعمق (القسم 3.3.1)، فمن المحتمل أن الغالبية العظمى من الميكروبلاستيك التي تدخل الأنظمة المائية الطبيعية تمر بدورة من الاستقرار على سطح الرواسب تحت ظروف تدفق منخفضة وإعادة تعليقها في عمود الماء تحت ظروف تدفق عالية. وهذا يبرز التأثيرات القوية للموسمية على آليات ترسيب وإعادة تعليق الميكروبلاستيك، مما يجعل من الصعب الاحتفاظ بالميكروبلاستيك بشكل دائم في الرواسب النهرية الأعمق (لي وآخرون، 2022).
4. نقل الميكروبلاستيك في أنظمة المياه العذبة
4.1. نقل الميكروبلاستيك في أنظمة الأنهار
4.2. نقل الجسيمات الدقيقة في أنظمة البحيرات
الألياف بشكل أساسي (
نموذج الخصائص البيئية، مثل أنماط التدفق، والتعرض للرياح، والهطول، والتدفق من الأنهار والجداول المجاورة، وخصائص البحيرة، وخصائص الرواسب على سلوك الجسيمات الدقيقة الديناميكي في البحيرات. سيكون هذا النهج متوافقًا مع النظريات الحالية المطبقة على دراسة أنظمة الأنهار، مما يعزز فهمنا لديناميات نقل الجسيمات الدقيقة ومصيرها في أنظمة المياه العذبة في البحيرات.
5. التحديات الحالية وآفاق المستقبل
5.1. نمذجة الظروف البيئية الطبيعية والدراسات المخبرية
5.2. العلاقة بين ديناميات الجسيمات الدقيقة والعمليات البيئية
على أنماط التدفق التي تحرك توزيعات الجسيمات الدقيقة. علاوة على ذلك، فإن فحص كيفية تأثير التغيرات الهيدروديناميكية على ديناميات الجسيمات الدقيقة، خاصة من خلال التأثير على مستويات NOM وSS، وبالتالي التحكم في التلوث البيولوجي والتجوية، هو فجوة معرفية أخرى حاسمة. على الرغم من أن تتبع ديناميات الجسيمات الدقيقة عبر النظام المائي يمثل تحديًا، فإن اعتماد إعدادات الميسوكوزم المماثلة لـ (هارلي وآخرون، 2018) للملاحظات في الموقع يمكن أن يوفر معلومات شاملة حول العمليات المعقدة للتجوية وتأثيرات ديناميات السوائل على سلوك الجسيمات الدقيقة، مما يعوض عن القيود الجوهرية للبيانات المستندة إلى المختبر الحالية.
5.3. التفاعلات بين سلوكيات ديناميات الجسيمات الدقيقة
6. الاستنتاجات
أنظمة المياه العذبة. أجرت هذه الدراسة تحليلًا نقديًا للدراسات الحالية الرائدة حول السلوكيات الديناميكية للجزيئات الدقيقة، بما في ذلك الترسيب، والتجمع، والاحتجاز، وإعادة التعليق، التي تؤثر على نقلها ومصيرها في أنظمة المياه العذبة. من خلال دمج النتائج التجريبية والرؤى النظرية، تم تقديم تحليل شامل للتفاعلات المعقدة بين خصائص الجزيئات الدقيقة والظروف البيئية، مع التأكيد على تعقيد ديناميات الجزيئات الدقيقة ومصيرها في البيئات المائية العذبة. لا توضح نتائجنا فقط الآليات الأساسية الرئيسية التي تحكم السلوك الديناميكي للجزيئات الدقيقة، ولكنها تحدد أيضًا الفجوات البحثية الرئيسية في فهم عمليات نقل الجزيئات الدقيقة ومصيرها البيئي، مما يسهل الإدارة البيئية القوية وتقييم المخاطر البيئية.
التقدم المنهجي ضروري لفك ‘الصندوق الأسود’ لنقل الجسيمات الدقيقة، مما يسهل فهمًا شاملاً لكيفية نقل الجسيمات الدقيقة وتفاعلها تحت مجموعة متنوعة من العوامل الهيدرولوجية والهيدروديناميكية والبيئية.
بيان مساهمة تأليف CRediT
إعلان عن تضارب المصالح
توفر البيانات
شكر وتقدير
المواد التكميلية
References
Alimi, O.S., Farner Budarz, J., Hernandez, L.M., Tufenkji, N., 2018. Microplastics and nanoplastics in aquatic environments: aggregation, deposition, and enhanced contaminant transport. Environ. Sci. Technol. 52 (4), 1704-1724. https://doi.org/ 10.1021/acs.est.7b05559.
Augusto-Silva, P.B., MacIntyre, S., de Moraes Rudorff, C., Cortés, A., Melack, J.M., 2019. Stratification and mixing in large floodplain lakes along the lower Amazon River. J. Great Lakes Res. 45 (1), 61-72. https://doi.org/10.1016/j.jglr.2018.11.001.
Ballent, A., Corcoran, P.L., Madden, O., Helm, P.A., Longstaffe, F.J., 2016. Sources and sinks of microplastics in Canadian Lake Ontario nearshore, tributary and beach
sediments. Mar. Pollut. Bull. 110 (1), 383-395. https://doi.org/10.1016/j. marpolbul.2016.06.037.
Ballent, A., Pando, S., Purser, A., Juliano, M.F., Thomsen, L., 2013. Modelled transport of benthic marine microplastic pollution in the Nazaré Canyon. Biogeosciences 10 (12), 7957-7970.
Ballent, A., Purser, A., de Jesus Mendes, P., Pando, S., Thomsen, L., 2012. Physical transport properties of marine microplastic pollution. Biogeosciences Discuss. 9 (12), 18755-18798. https://doi.org/10.5194/bgd-9-18755-2012.
Barnes, D.K.A., Milner, P., 2005. Drifting plastic and its consequences for sessile organism dispersal in the Atlantic Ocean. Mar. Biol. 146 (4), 815-825. https://doi. org/10.1007/s00227-004-1474-8.
Beletsky, D., Saylor, J.H., Schwab, D.J., 1999. Mean circulation in the Great Lakes. J. Great Lakes Res. 25 (1), 78-93. https://doi.org/10.1016/S0380-1330(99)707185.
Bissell, M.J., Barcellos-Hoff, M.H., 1987. The influence of extracellular matrix on gene expression: is structure the message? J. Cell Sci. 343 (SUPPL. 8), 327-343. https:// doi.org/10.1242/jcs.1987.supplement_8.18.
Boos, J.P., Gilfedder, B.S., Frei, S., 2021. Tracking microplastics across the streambed interface: using laser-induced-fluorescence to quantitatively analyze microplastic transport in an experimental flume. Water Resour. Res. 57 (12), 1-10. https://doi. org/10.1029/2021WR031064.
Browne, M.A., Galloway, T.S., Thompson, R.C., 2010. Spatial patterns of plastic debris along estuarine shorelines. Environ. Sci. Technol. 44 (9), 3404-3409. https://doi. org/10.1021/es903784e.
Buffle, I., Leppard, G.G., 1995. Characterization of aquatic colloids and macromolecules. 1. Structure and behavior of colloidal material. Environ. Sci. Technol. 29 (9), 2169-2175. https://doi.org/10.1021/es00009a004.
Butt, H.J., Jaschke, M., Ducker, W., 1995. Measuring surface forces in aqueous electrolyte solution with the atomic force microscope. Bioelectrochem. Bioenerg. 38 (1), 191-201. https://doi.org/10.1016/0302-4598(95)01800-T.
Campanale, C., et al., 2020. Microplastics and their possible sources: the example of Ofanto river in southeast Italy. Environ. Pollut. 258, 113284 https://doi.org/ 10.1016/j.envpol.2019.113284.
Carpenter, E.J., Smith, K.L., 1972. Plastics on the Sargasso sea surface. Science (80-.) 175 (4027), 1240-1241. https://doi.org/10.1126/science.175.4027.1240.
Chen, Y., et al., 2021. Transport and fate of microplastics in constructed wetlands: a microcosm study. J. Hazard. Mater. 415 (March), 125615 https://doi.org/10.1016/ j.jhazmat.2021.125615.
Chowdhury, I., Mansukhani, N.D., Guiney, L.M., Hersam, M.C., Bouchard, D., 2015. Aggregation and stability of reduced graphene oxide: complex roles of divalent cations, pH, and natural organic matter. Environ. Sci. Technol. 49 (18), 10886-10893. https://doi.org/10.1021/acs.est.5b01866. Sep.
Chubarenko, I., Bagaev, A., Zobkov, M., Esiukova, E., 2016. On some physical and dynamical properties of microplastic particles in marine environment. Mar. Pollut. Bull. 108 (1-2), 105-112. https://doi.org/10.1016/j.marpolbul.2016.04.048.
Chubarenko, I., Efimova, I., Bagaeva, M., Bagaev, A., Isachenko, I., 2020. On mechanical fragmentation of single-use plastics in the sea swash zone with different types of bottom sediments: insights from laboratory experiments. Mar. Pollut. Bull. 150 (November 2019) https://doi.org/10.1016/j.marpolbul.2019.110726.
Cole, M., Lindeque, P., Halsband, C., Galloway, T.S., 2011. Microplastics as contaminants in the marine environment: a review. Mar. Pollut. Bull. 62 (12), 2588-2597. https:// doi.org/10.1016/j.marpolbul.2011.09.025.
Corcoran, P.L., Belontz, S.L., Ryan, K., Walzak, M.J., 2020. Factors controlling the distribution of microplastic particles in benthic sediment of the Thames River, Canada. Environ. Sci. Technol. 54 (2), 818-825. https://doi.org/10.1021/acs. est.9b04896.
Corcoran, P.L., Norris, T., Ceccanese, T., Walzak, M.J., Helm, P.A., Marvin, C.H., 2015. Hidden plastics of Lake Ontario, Canada and their potential preservation in the sediment record. Environ. Pollut. 204, 17-25. https://doi.org/10.1016/j. envpol.2015.04.009.
Cunha, C., Faria, M., Nogueira, N., Ferreira, A., Cordeiro, N., 2019. Marine vs freshwater microalgae exopolymers as biosolutions to microplastics pollution. Environ. Pollut. 249, 372-380. https://doi.org/10.1016/j.envpol.2019.03.046.
Cunha, C., Silva, L., Paulo, J., Faria, M., Nogueira, N., Cordeiro, N., 2020. Microalgalbased biopolymer for nano- and microplastic removal: a possible biosolution for
wastewater treatment. Environ. Pollut. 263 https://doi.org/10.1016/j. envpol.2020.114385.
Daily, J., Hoffman, M.J., 2020. Modeling the three-dimensional transport and distribution of multiple microplastic polymer types in Lake Erie. Mar. Pollut. Bull. 154 (January) https://doi.org/10.1016/j.marpolbul.2020.111024.
Daily, J., Onink, V., Jongedijk, C.E., Laufkötter, C., Hoffman, M.J., 2021. Incorporating terrain specific beaching within a lagrangian transport plastics model for Lake Erie. Microplast. Nanoplast. 1 (1), 1-13. https://doi.org/10.1186/s43591-021-00019-7.
de Carvalho, A.R., et al., 2021. Urbanization and hydrological conditions drive the spatial and temporal variability of microplastic pollution in the Garonne River. Sci. Total Environ. 769 https://doi.org/10.1016/j.scitotenv.2020.144479.
De Leo, A., Cutroneo, L., Sous, D., Stocchino, A., 2021. Settling velocity of microplastics exposed to wave action. J. Mar. Sci. Eng. 9 (2), 1-16. https://doi.org/10.3390/ jmse9020142.
Demiral, D., Albayrak, I., Turowski, J.M., Boes, R.M., 2022. Particle saltation trajectories in supercritical open channel flows: Roughness effect. Earth Surf. Process. Landf. 47 (15), 3588-3610. https://doi.org/10.1002/esp.5475.
Dong, Z., Qiu, Y., Zhang, W., Yang, Z., Wei, L., 2018. Size-dependent transport and retention of micron-sized plastic spheres in natural sand saturated with seawater. Water Res 143, 518-526. https://doi.org/10.1016/j.watres.2018.07.007.
Drago, C., Pawlak, J., Weithoff, G., 2020. Biogenic aggregation of small microplastics alters their ingestion by a common freshwater micro-invertebrate. Front. Environ. Sci. 8 (December) https://doi.org/10.3389/fenvs.2020.574274.
Dris, R., Gasperi, J., Rocher, V., Tassin, B., 2018. Synthetic and non-synthetic anthropogenic fibers in a river under the impact of Paris Megacity: sampling methodological aspects and flux estimations. Sci. Total Environ. 618, 157-164. https://doi.org/10.1016/j.scitotenv.2017.11.009.
Drummond, J.D., Nel, H.A., Packman, A.I., Krause, S., 2020. Significance of hyporheic exchange for predicting microplastic fate in rivers. Environ. Sci. Technol. Lett. 7 (10), 727-732. https://doi.org/10.1021/acs.estlett.0c00595.
Duan, J., et al., 2021. Weathering of microplastics and interaction with other coexisting constituents in terrestrial and aquatic environments. Water Res., 117011
Eerkes-Medrano, D., Thompson, R.C., Aldridge, D.C., 2015. Microplastics in freshwater systems: a review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 75, 63-82. https://doi.org/10.1016/j. watres.2015.02.012.
Egessa, R., Nankabirwa, A., Basooma, R., Nabwire, R., 2020. Occurrence, distribution and size relationships of plastic debris along shores and sediment of northern Lake Victoria. Environ. Pollut. 257, 113442 https://doi.org/10.1016/j. envpol.2019.113442.
Elagami, H., et al., 2022. Measurement of microplastic settling velocities and implications for residence times in thermally stratified lakes. Limnol. Oceanogr. 934-945. https://doi.org/10.1002/lno.12046. Agarwal 2020.
Elagami, H., Frei, S., Boos, J.P., Trommer, G., Gilfedder, B.S., 2023. Quantifying microplastic residence times in lakes using mesocosm experiments and transport modelling. Water Res. 229 (November 2022), 119463 https://doi.org/10.1016/j. watres.2022.119463.
Enfrin, M., Lee, J., Gibert, Y., Basheer, F., Kong, L., Dumée, L.F., 2020. Release of hazardous nanoplastic contaminants due to microplastics fragmentation under shear stress forces. J. Hazard. Mater. 384 (September 2019), 121393 https://doi.org/ 10.1016/j.jhazmat.2019.121393.
Farizan, A., Yaghoubi, S., Firoozabadi, B., Afshin, H., 2019. Effect of an obstacle on the depositional behaviour of turbidity currents. J. Hydraul. Res. 57 (1), 75-89. https:// doi.org/10.1080/00221686.2018.1459891.
Faulstich, L., Prume, J.A., Arendt, R., Reinhardt-Imjela, C., Chifflard, P., Schulte, A., 2022. Microplastics in Namibian river sediments – a first evaluation. Microplastics and Nanoplastics 2 (1), 1-17. https://doi.org/10.1186/s43591-022-00043-1.
Fauser, P., Vorkamp, K., Strand, J., 2021. Residual additives in marine microplastics and their risk assessment – a critical review. Mar. Pollut. Bull. 177, 113467 https://doi. org/10.1016/j.marpolbul.2022.113467. October2022.
Fazey, F.M.C., Ryan, P.G., 2016. Biofouling on buoyant marine plastics: an experimental study into the effect of size on surface longevity. Environ. Pollut. 210, 354-360. https://doi.org/10.1016/j.envpol.2016.01.026.
Ferdowsi, B., Ortiz, C.P., Houssais, M., Jerolmack, D.J., 2017. River-bed armouring as a granular segregation phenomenon. Nat. Commun. 8 (1) https://doi.org/10.1038/ s41467-017-01681-3.
Fischer, E.K., Paglialonga, L., Czech, E., Tamminga, M., 2016. Microplastic pollution in lakes and lake shoreline sediments – a case study on Lake Bolsena and Lake Chiusi (central Italy. Environ. Pollut. 213, 648-657. https://doi.org/10.1016/j. envpol.2016.03.012.
Flemming, H.C., Wingender, J., 2010. The biofilm matrix. Nat. Rev. Microbiol. 8 (9), 623-633. https://doi.org/10.1038/nrmicro2415.
Gao, L., Su, Y., Yang, L., Li, J., Bao, R., Peng, L., 2022. Sorption behaviors of petroleum on micro-sized polyethylene aging for different time in seawater. Sci. Total Environ. 808, 152070 https://doi.org/10.1016/j.scitotenv.2021.152070.
Gerolin, C.R., Pupim, F.N., Sawakuchi, A.O., Grohmann, C.H., Labuto, G., Semensatto, D., 2020. Microplastics in sediments from Amazon rivers, Brazil. Sci. Total Environ. 749, 141604 https://doi.org/10.1016/j.scitotenv.2020.141604.
Gholizadeh, M., Cera, A., 2022. Microplastic contamination in the sediments of Qarasu estuary in Gorgan Bay, south-east of Caspian Sea, Iran. Sci. Total Environ. 838 (February), 155913 https://doi.org/10.1016/j.scitotenv.2022.155913.
Gilfedder, B.S., et al., 2023. Filter feeders are key to small microplastic residence times in stratified lakes: a virtual experiment. Sci. Total Environ. 890 (May), 164293 https:// doi.org/10.1016/j.scitotenv.2023.164293.
Goodarzi, D., Mohammadian, A., Pearson, J., Abolfathi, S., 2022. Numerical modelling of hydraulic efficiency and pollution transport in waste stabilization ponds. Ecol. Eng. 182 (June 2021), 106702 https://doi.org/10.1016/j.ecoleng.2022.106702.
He, B., Goonetilleke, A., Ayoko, G.A., Rintoul, L., 2020a. Abundance, distribution patterns, and identification of microplastics in Brisbane River sediments, Australia. Sci. Total Environ. 700, 134467 https://doi.org/10.1016/j.scitotenv.2019.134467.
He, B., Wijesiri, B., Ayoko, G.A., Egodawatta, P., Rintoul, L., Goonetilleke, A., 2020b. Influential factors on microplastics occurrence in river sediments. Sci. Total Environ. 738, 139901 https://doi.org/10.1016/j.scitotenv.2020.139901.
Helcoski, R., Yonkos, L.T., Sanchez, A., Baldwin, A.H., 2020. Wetland soil microplastics are negatively related to vegetation cover and stem density. Environ. Pollut. 256, 113391 https://doi.org/10.1016/j.envpol.2019.113391.
Hendrickson, E., Minor, E.C., Schreiner, K., 2018. Microplastic abundance and composition in western lake superior as determined via microscopy, Pyr-GC/MS, and FTIR. Environ. Sci. Technol. 52 (4), 1787-1796. https://doi.org/10.1021/acs. est.7b05829.
Hoellein, T.J., Shogren, A.J., Tank, J.L., Risteca, P., Kelly, J.J., 2019. Microplastic deposition velocity in streams follows patterns for naturally occurring allochthonous particles. Sci. Rep. 9 (1), 1-12. https://doi.org/10.1038/s41598-019-40126-3.
Hoffman, M.J., Hittinger, E., 2017. Inventory and transport of plastic debris in the Laurentian Great Lakes. Mar. Pollut. Bull. 115 (1-2), 273-281. https://doi.org/ 10.1016/j.marpolbul.2016.11.061.
Homann, H., Bec, J., Grauer, R., 2013. Effect of turbulent fluctuations on the drag and lift forces on a towed sphere and its boundary layer. J. Fluid Mech. 721, 155-179. https://doi.org/10.1017/jfm.2013.66.
Horton, A.A., Dixon, S.J., 2018. Microplastics: an introduction to environmental transport processes. Wiley Interdiscip. Rev. Water 5 (2), e1268. https://doi.org/ 10.1002/wat2.1268.
Hurley, R., Woodward, J., Rothwell, J.J., 2018. Microplastic contamination of river beds significantly reduced by catchment-wide flooding. Nat. Geosci. 11 (4), 251-257. https://doi.org/10.1038/s41561-018-0080-1.
Jambeck, J., et al., 2015. Plastic waste inputs from land into the ocean. Mar. Pollut. 347 (6223), 768.
Ji, Z.-G., 2017. Hydrodynamics and Water Quality: Modeling Rivers, Lakes, and Estuaries. John Wiley & Sons.
Jiang, C., et al., 2018. Microplastics in sediment and surface water of west dongting lake and south dongting lake: abundance, source and composition. Int. J. Environ. Res. Public Health 15 (10). https://doi.org/10.3390/ijerph15102164.
Jiang, C., et al., 2019. Microplastic pollution in the rivers of the Tibet Plateau. Environ. Pollut. 249, 91-98. https://doi.org/10.1016/j.envpol.2019.03.022.
Junaid, M., Wang, J., 2021. Interaction of nanoplastics with extracellular polymeric substances (EPS) in the aquatic environment: a special reference to eco-corona formation and associated impacts. Water Res. 201 (May), 117319 https://doi.org/ 10.1016/j.watres.2021.117319.
Kaiser, D., Estelmann, A., Kowalski, N., Glockzin, M., Waniek, J.J., 2019. Sinking velocity of sub-millimeter microplastic. Mar. Pollut. Bull. 139 (December 2018), 214-220. https://doi.org/10.1016/j.marpolbul.2018.12.035.
Kaiser, D., Kowalski, N., Waniek, J.J., 2017. Effects of biofouling on the sinking behavior of microplastics. Environ. Res. Lett. 12 (12) https://doi.org/10.1088/1748-9326/ aa8e8b.
Khalid, N., et al., 2021. Linking effects of microplastics to ecological impacts in marine environments. Chemosphere 264, 128541. https://doi.org/10.1016/j. chemosphere.2020.128541.
Khatmullina, L., Isachenko, I., 2017. Settling velocity of microplastic particles of regular shapes. Mar. Pollut. Bull. 114 (2), 871-880. https://doi.org/10.1016/j. marpolbul.2016.11.024.
Kowalski, N., Reichardt, A.M., Waniek, J.J., 2016. Sinking rates of microplastics and potential implications of their alteration by physical, biological, and chemical
factors. Mar. Pollut. Bull. 109 (1), 310-319. https://doi.org/10.1016/j. marpolbul.2016.05.064.
Kukulka, T., Proskurowski, G., Morét-Ferguson, S., Meyer, D.W., Law, K.L., 2012. The effect of wind mixing on the vertical distribution of buoyant plastic debris. Geophys. Res. Lett. 39 (7), 1-6. https://doi.org/10.1029/2012GL051116.
Lagarde, F., Olivier, O., Zanella, M., Daniel, P., Hiard, S., Caruso, A., 2016. Microplastic interactions with freshwater microalgae: hetero-aggregation and changes in plastic density appear strongly dependent on polymer type. Environ. Pollut. 215, 331-339. https://doi.org/10.1016/j.envpol.2016.05.006.
Lead, J.R., Davison, W., Hamilton-Taylor, J., Buffle, J., 1997. Characterizing colloidal material in natural waters. Aquat. Geochem. 3 (3), 213-232. https://doi.org/ 10.1023/A:1009695928585.
Lenaker, P.L., Baldwin, A.K., Corsi, S.R., Mason, S.A., Reneau, P.C., Scott, J.W., 2019. Vertical distribution of microplastics in the water column and surficial sediment from the Milwaukee River Basin to Lake Michigan. Environ. Sci. Technol. 53 (21), 12227-12237. https://doi.org/10.1021/acs.est.9b03850.
Lenaker, P.L., Corsi, S.R., Mason, S.A., 2021. Spatial distribution of microplastics in surficial benthic sediment of lake Michigan and Lake Erie. Environ. Sci. Technol. 55 (1), 373-384. https://doi.org/10.1021/acs.est.0c06087.
Li, J., Song, Y., Cai, Y., 2020. Focus topics on microplastics in soil: analytical methods, occurrence, transport, and ecological risks. Environ. Pollut. 257, 113570 https://doi. org/10.1016/j.envpol.2019.113570.
Li, K., Ma, H., 2018. Deposition dynamics of rod-shaped colloids during transport in porous media under favorable conditions. Langmuir 34 (9), 2967-2980. https://doi. org/10.1021/acs.langmuir.7b03983.
Li, S., Liu, H., Gao, R., Abdurahman, A., Dai, J., Zeng, F., 2018. Aggregation kinetics of microplastics in aquatic environment: complex roles of electrolytes, pH , and natural organic matter. Environ. Pollut. 237, 126-132. https://doi.org/10.1016/j. envpol.2018.02.042.
Li, W., et al., 2019a. Colonization characteristics of bacterial communities on plastic debris influenced by environmental factors and polymer types in the Haihe Estuary of Bohai Bay, China. Environ. Sci. Technol. 53 (18), 10763-10773. https://doi.org/ 10.1021/acs.est.9b03659.
Li, Y., et al., 2019b. Interactions between nano/micro plastics and suspended sediment in water: implications on aggregation and settling. Water Res. 161, 486-495. https:// doi.org/10.1016/j.watres.2019.06.018.
Liu, K., Courtene-Jones, W., Wang, X., Song, Z., Wei, N., Li, D., 2020. Elucidating the vertical transport of microplastics in the water column: a review of sampling methodologies and distributions. Water Res. 186, 116403 https://doi.org/10.1016/ j.watres.2020.116403.
Liu, Y., et al., 2022b. Application of a microplastic trap to the determination of the factors controlling the lakebed deposition of microplastics. Sci. Total Environ. 843 (March), 156883 https://doi.org/10.1016/j.scitotenv.2022.156883.
Liu, Y., Hu, Y., Yang, C., Chen, C., Huang, W., Dang, Z., 2019. Aggregation kinetics of UV irradiated nanoplastics in aquatic environments. Water Res. 163, 114870 https:// doi.org/10.1016/j.watres.2019.114870.
Lu, S., et al., 2018. Impact of water chemistry on surface charge and aggregation of polystyrene microspheres suspensions. Sci. Total Environ. 630, 951-959. https:// doi.org/10.1016/j.scitotenv.2018.02.296.
Mahdian, M., et al., 2023. Modelling impacts of climate change and anthropogenic activities on inflows and sediment loads of wetlands: case study of the Anzali wetland. Sci. Rep. 13 (1), 1-18. https://doi.org/10.1038/s41598-023-32343-8.
Mahdian, M., Noori, R., Salamattalab, M.M., Heggy, E., Bateni, S.M., Nohegar, A., et al., 2024. Anzali wetland crisis: Unraveling the decline of Iran’s ecological gem. J. Geophys. Res. Atmos. 129 https://doi.org/10.1029/2023JD039538 e2023JD039538.
Mancini, M., Serra, T., Colomer, J., Solari, L., 2023. Suspended sediments mediate microplastic sedimentation in unidirectional flows. Sci. Total Environ. 890 (May), 164363 https://doi.org/10.1016/j.scitotenv.2023.164363.
Mani, T., Hauk, A., Walter, U., Burkhardt-Holm, P., 2015. Microplastics profile along the Rhine River. Sci. Rep. 5 (December), 1-7. https://doi.org/10.1038/srep17988.
Mendrik, F., Fernández, R., Hackney, C.R., Waller, C., Parsons, D.R., 2023. Non-buoyant microplastic settling velocity varies with biofilm growth and ambient water salinity. Commun. Earth Environ. 4 (1), 1-9. https://doi.org/10.1038/s43247-023-00690-z.
Miao, L., et al., 2021. Effects of biofilm colonization on the sinking of microplastics in three freshwater environments. J. Hazard. Mater. 413 (December 2020), 125370 https://doi.org/10.1016/j.jhazmat.2021.125370.
Michels, J., Stippkugel, A., Lenz, M., Wirtz, K., Engel, A., 2018. Rapid aggregation of biofilm-covered microplastics with marine biogenic particles. Proc. R. Soc. B Biol. Sci. 285 (1885) https://doi.org/10.1098/rspb.2018.1203.
Mosley, L.M., Hunter, K.A., Ducker, W.A., 2003. Forces between colloid particles in natural waters. Environ. Sci. Technol. 37 (15), 3303-3308. https://doi.org/ 10.1021/es026216d.
Y.W. Naguib and Z. Cui, Nanomedicine: the promise and challenges in cancer chemotherapy, vol. 811. 2014. 10.1007/978-94-017-8739-0_11.
Nakayama, T., Osako, M., 2023. Development of a process-based eco-hydrology model for evaluating the spatio-temporal dynamics of macro- and micro-plastics for the whole of Japan. Ecol. Modell. 476 (November 2022), 110243 https://doi.org/ 10.1016/j.ecolmodel.2022.110243.
Nel, H.A., Dalu, T., Wasserman, R.J., 2018. Sinks and sources: assessing microplastic abundance in river sediment and deposit feeders in an Austral temperate urban river system. Sci. Total Environ. 612, 950-956. https://doi.org/10.1016/j. scitotenv.2017.08.298.
Nguyen, T.H., 2021. Settling behaviour of irregular-shaped polystyrene microplastics. Sci. Technol. Dev. J.1-Eng. Technol. 4 (4), 1219-1228.
Nguyen, T.H., Kieu-Le, T.C., Tang, F.H.M., Maggi, F., 2022. Controlling factors of microplastic fibre settling through a water column. Sci. Total Environ. 838 (December 2021), 156011 https://doi.org/10.1016/j.scitotenv.2022.156011.
Nguyen, T.H., Tang, F.H.M., Maggi, F., 2020. Sinking of microbial-associated microplastics in natural waters. PLoS ONE 15 (2), 1-20. https://doi.org/10.1371/ journal.pone. 0228209.
Niu, L., et al., 2021. New insights into the vertical distribution and microbial degradation of microplastics in urban river sediments. Water Res. 188 https://doi.org/10.1016/j. watres.2020.116449.
Nizzetto, L., Bussi, G., Futter, M.N., Butterfield, D., Whitehead, P.G., 2016. A theoretical assessment of microplastic transport in river catchments and their retention by soils and river sediments. Environ. Sci. Process. Impacts 18 (8), 1050-1059. https://doi. org/10.1039/c6em00206d.
Nouha, K., Kumar, R.S., Balasubramanian, S., Tyagi, R.D., 2018. Critical review of EPS production, synthesis and composition for sludge flocculation. J. Environ. Sci. (China) 66, 225-245. https://doi.org/10.1016/j.jes.2017.05.020.
Ockelford, A., Cundy, A., Ebdon, J.E., 2020. Storm response of fluvial sedimentary microplastics. Sci. Rep. 10 (1), 1-10. https://doi.org/10.1038/s41598-020-58765-2.
Pascolo, S., Petti, M., Bosa, S., 2018. On the wave bottom shear stress in shallow depths: the role of wave period and bed roughness. Water (Switzerland) 10 (10). https://doi. org/10.3390/w10101348.
Peller, J., et al., 2021. Sequestration of microfibers and other microplastics by green algae, Cladophora, in the US Great Lakes. Environ. Pollut. 276, 116695 https://doi. org/10.1016/j.envpol.2021.116695.
Peng, G., Zhu, B., Yang, D., Su, L., Shi, H., Li, D., 2017. Microplastics in sediments of the Changjiang Estuary, China. Environ. Pollut. 225, 283-290. https://doi.org/10.1016/ j.envpol.2016.12.064.
Pohl, F., Eggenhuisen, J.T., Kane, I.A., Clare, M.A., 2020. Transport and burial of microplastics in deep-marine sediments by turbidity currents. Environ. Sci. Technol. 54 (7), 4180-4189. https://doi.org/10.1021/acs.est.9b07527.
Rasta, M., Sattari, M., Taleshi, M.S., Namin, J.I., 2020. Identification and distribution of microplastics in the sediments and surface waters of Anzali Wetland in the Southwest Caspian Sea, Northern Iran. Mar. Pollut. Bull. 160 (August), 111541 https://doi.org/10.1016/j.marpolbul.2020.111541.
Ren, Z., Gui, X., Xu, X., Zhao, L., Qiu, H., Cao, X., 2021. Microplastics in the soilgroundwater environment: aging, migration, and co-transport of contaminants – A critical review. J. Hazard. Mater. 419 https://doi.org/10.1016/j. jhazmat.2021.126455. March.
Righetti, M., Lucarelli, C., 2007. May the Shields theory be extended to cohesive and adhesive benthic sediments? J. Geophys. Res. Ocean. 112 (5), 1-14. https://doi.org/ 10.1029/2006JC003669.
Ryan, P.G., 2015. Does size and buoyancy affect the long-distance transport of floating debris? Environ. Res. Lett. 10 (8) https://doi.org/10.1088/1748-9326/10/8/ 084019.
Schmidt, C., Kumar, R., Yang, S., Büttner, O., 2020. Microplastic particle emission from wastewater treatment plant effluents into river networks in Germany: loads, spatial patterns of concentrations and potential toxicity. Sci. Total Environ. 737, 139544 https://doi.org/10.1016/j.scitotenv.2020.139544.
Shams, M., Alam, I., Chowdhury, I., 2020. Aggregation and stability of nanoscale plastics in aquatic environment. Water Res. 171, 115401 https://doi.org/10.1016/j. watres.2019.115401.
Shang, Q.Q., Fang, H.W., Zhao, H.M., He, G.J., Cui, Z.H., 2014. Biofilm effects on size gradation, drag coefficient and settling velocity of sediment particles. Int. J. Sediment Res. 29 (4), 471-480. https://doi.org/10.1016/S1001-6279(14)60060-3.
Shen, Z., Wang, H., Liang, D., Yan, Y., Zeng, Y., 2022. The fate of microplastics in estuary: a quantitative simulation approach. Water Res. 226 (October), 119281 https://doi.org/10.1016/j.watres.2022.119281.
A. Shields, “Application of similarity principles and turbulence research to bed-load movement,” no. 167, 47, 1936.
Singh, N., Tiwari, E., Khandelwal, N., Darbha, G.K., 2019. Understanding the stability of nanoplastics in aqueous environments: effect of ionic strength, temperature,
dissolved organic matter, clay, and heavy metals. Environ. Sci. Nano 6 (10), 2968-2976. https://doi.org/10.1039/c9en00557a.
Song, Y.K., et al., 2014. Large accumulation of micro-sized synthetic polymer particles in the sea surface microlayer. Environ. Sci. Technol. 48 (16), 9014-9021. https://doi. org/10.1021/es501757s.
Song, Y.K., et al., 2018. Horizontal and vertical distribution of microplastics in Korean coastal waters. Environ. Sci. Technol. 52 (21), 12188-12197. https://doi.org/ 10.1021/acs.est.8b04032.
Stead, J.L., Bond, T., 2023. The impact of riverine particles on the vertical velocities of large microplastics. Sci. Total Environ. 896 (May), 165339 https://doi.org/10.1016/ j.scitotenv.2023.165339.
Stride, B., Abolfathi, S., Bending, G.D., Pearson, J., 2024. Quantifying microplastic dispersion due to density effects. J. Hazard. Mater. 466, 133440 https://doi.org/ 10.1016/j.jhazmat.2024.133440.
Sun, H., Jiao, R., Wang, D., 2020. The difference of aggregation mechanism between microplastics and nanoplastics: role of Brownian motion and structural layer force. Environ. Pollut. 268, 115942 https://doi.org/10.1016/j.envpol.2020.115942.
Tamminga, M., Fischer, E.K., 2020. Microplastics in a deep, dimictic lake of the North German Plain with special regard to vertical distribution patterns. Environ. Pollut. 267, 115507 https://doi.org/10.1016/j.envpol.2020.115507.
Tibbetts, J., Krause, S., Lynch, I., Smith, G.H.S., 2018. Abundance, distribution, and drivers of microplastic contamination in urban river environments. Water (Switzerland) 10 (11). https://doi.org/10.3390/w10111597.
G. Trefalt and M. Borkovec, “Overview of DLVO theory,” 2014.
Vaughan, R., Turner, S.D., Rose, N.L., 2017. Microplastics in the sediments of a UK urban lake. Environ. Pollut. 229, 10-18. https://doi.org/10.1016/j.envpol.2017.05.057.
Vermaire, J.C., Pomeroy, C., Herczegh, S.M., Haggart, O., Murphy, M., 2017. Microplastic abundance and distribution in the open water and sediment of the Ottawa River, Canada, and its tributaries. Facets 2 (1), 301-314.
Wagner, M., Lambert, S., 2018. Freshwater Microplastics. Springer Nature. https://doi. org/10.1007/978-3-319-61615-5.
Waldschlägelatr, K., Schüttrumpf, H., 2019. Effects of particle properties on the settling and rise velocities of microplastics in freshwater under laboratory conditions. Environ. Sci. Technol. 53 (4), 1958-1966. https://doi.org/10.1021/acs.est.8b06794.
Waldschläger, K., Born, M., Cowger, W., Gray, A., Schüttrumpf, H., 2020. Settling and rising velocities of environmentally weathered micro- and macroplastic particles. Environ. Res. 191 (August) https://doi.org/10.1016/j.envres.2020.110192.
Waldschläger, K., Schüttrumpf, H., 2019. Erosion behavior of different microplastic particles in comparison to natural sediments. Environ. Sci. Technol. https://doi.org/ 10.1021/acs.est.9b05394.
Wang, J., et al., 2017. Microplastics in the surface sediments from the Beijiang River littoral zone: composition, abundance, surface textures and interaction with heavy metals. Chemosphere 171, 248-258. https://doi.org/10.1016/j. chemosphere.2016.12.074.
Wang, J., et al., 2021c. Aggregation and stability of sulfate-modified polystyrene nanoplastics in synthetic and natural waters. Environ. Pollut. 268, 114240 https:// doi.org/10.1016/j.envpol.2020.114240.
Wang, X., Bolan, N., Tsang, D.C.W., Sarkar, B., Bradney, L., Li, Y., 2021a. A review of microplastics aggregation in aquatic environment: influence factors, analytical methods, and environmental implications. J. Hazard. Mater. 402 (March 2020), 123496 https://doi.org/10.1016/j.jhazmat.2020.123496.
Wang, Y., Chen, X., Wang, F., Cheng, N., 2023. Influence of typical clay minerals on aggregation and settling of pristine and aged polyethylene microplastics. Environ. Pollut. 316 (P2), 120649 https://doi.org/10.1016/j.envpol.2022.120649.
Wang, Z., Dou, M., Ren, P., Sun, B., Jia, R., Zhou, Y., 2021b. Settling velocity of irregularly shaped microplastics under steady and dynamic flow conditions. Environ. Sci. Pollut. Res. 28 (44), 62116-62132. https://doi.org/10.1007/s11356-021-14654-3.
Willis, K.A., Eriksen, R., Wilcox, C., Hardesty, B.D., 2017. Microplastic distribution at different sediment depths in an urban estuary. Front. Mar. Sci. 4 (DEC), 1-9. https:// doi.org/10.3389/fmars.2017.00419.
Wu, F., Pennings, S.C., Tong, C., Xu, Y., 2020b. Variation in microplastics composition at small spatial and temporal scales in a tidal flat of the Yangtze Estuary, China. Sci. Total Environ. 699, 134252 https://doi.org/10.1016/j.scitotenv.2019.134252.
Wu, J., Jiang, R., Lin, W., Ouyang, G., 2019. Effect of salinity and humic acid on the aggregation and toxicity of polystyrene nanoplastics with different functional groups and charges. Environ. Pollut. 245, 836-843. https://doi.org/10.1016/j. envpol.2018.11.055.
Yang, G., et al., 2023. Settling behaviors of microplastic disks in water. Mar. Pollut. Bull. 188 (February), 114657 https://doi.org/10.1016/j.marpolbul.2023.114657.
Yu, S., et al., 2019. Aggregation kinetics of different surface-modified polystyrene nanoparticles in monovalent and divalent electrolytes. Environ. Pollut. 255, 113302 https://doi.org/10.1016/j.envpol.2019.113302.
Yu, Z., Yao, W., Loewen, M., Li, X., Zhang, W., 2022. Incipient motion of exposed microplastics in an open-channel flow. Environ. Sci. Technol. 56 (20), 14498-14506. https://doi.org/10.1021/acs.est.2c04415.
Yuan, W., Liu, X., Wang, W., Di, M., Wang, J., 2019. Microplastic abundance, distribution and composition in water, sediments, and wild fish from Poyang Lake, China. Ecotoxicol. Environ. Saf. 170 (December 2018), 180-187. https://doi.org/10.1016/ j.ecoenv.2018.11.126.
Zhang, Q., et al., 2021. Distribution and sedimentation of microplastics in Taihu Lake. Sci. Total Environ. 795, 148745 https://doi.org/10.1016/j.scitotenv.2021.148745.
Zhang, Y., et al., 2020b. How climate change and eutrophication interact with microplastic pollution and sediment resuspension in shallow lakes: a review. Sci. Total Environ. 705, 135979 https://doi.org/10.1016/j.scitotenv.2019.135979.
Zhao, J., Shan, T., 2013. Coupled CFD-DEM simulation of fluid-particle interaction in geomechanics. Powder Technol. 239, 248-258. https://doi.org/10.1016/j. powtec.2013.02.003.
Zheng, Y., et al., 2020. Vertical distribution of microplastics in bay sediment reflecting effects of sedimentation dynamics and anthropogenic activities. Mar. Pollut. Bull. 152 (December 2019), 110885 https://doi.org/10.1016/j.marpolbul.2020.110885.
Zhu, H.P., Zhou, Z.Y., Yang, R.Y., Yu, A.B., 2007. Discrete particle simulation of particulate systems: theoretical developments. Chem. Eng. Sci. 62 (13), 3378-3396. https://doi.org/10.1016/j.ces.2006.12.089.
Zhu, X.-T., Yi, J., Qiang, L.-Y., Cheng, J.-P., 2018. Distribution and settlement of microplastics in the surface sediment of Yangtze Estuary. Huan jing ke xue= Huanjing kexue 39 (5), 2067-2074.
- Corresponding author.
E-mail addresses: Soroush.Abolfathi@Warwick.ac.uk, s.abolfathi@warwick.ac.uk (S. Abolfathi).
DOI: https://doi.org/10.1016/j.resconrec.2024.107578
Publication Date: 2024-04-08
Microplastics in freshwater systems: Dynamic behaviour and transport processes
ARTICLE INFO
Keywords:
Pollution transport
Freshwater
Dispersion
Mixing
Sinking
Aggregation
Retention
Resuspension
Abstract
Freshwater ecosystems are viewed as a key medium for the transport of land-based plastics into oceans. Microplastic (MP) particles in freshwater environments demonstrate high persistence and an extensive range of size and shape distributions, which make their mobility, distribution, and fate vary distinctly depending on the prevailing environmental conditions. The inherent physical properties of different plastic polymers are constantly evolving at different specific reaction rates due to the complex weathering processes in the environment. This continuously alters the underlying mechanisms governing MP dynamics and further complicates their ultimate fate in natural aquatic systems. This paper conducts a comprehensive review of the dynamic behaviour of MPs in freshwater ecosystems, focusing on investigating the settling, aggregation, retention, and suspension processes governing their transport from the source to the sink. We provide an in-depth overview of the key theoretical foundations of MP behaviour in ambient flows and the key influential factors (i.e. size, density, shape, composition). Our findings highlight intricate interplays between MP dynamic behaviours and local hydrodynamics and water chemistry, which lead to the continuous evolution of MP physicochemical properties (e.g., size, surface charge) through interactions with suspended solids, organic natural matter, and microorganisms under light and wind exposure. This dynamic poses significant challenges in predicting MP transport processes and ultimate fate. Gap analysis highlights the discrepancy between current models based on controlled laboratory conditions and complex natural environments, signifying the need for investigating MP dynamic behaviour across a wide range of environmental conditions (e.g. simulating complex flow patterns and solution chemistries of real water bodies). Further research is needed to expand field studies to correlate environment hydrodynamics with MP abundance and to conduct mesoscale experiments that accurately reflect the effects of weathering and flow hydrodynamics on MP behaviours. Integrating detailed physical experiments with numerical modelling tools is essential for a comprehensive understanding of the interactions among various MPs and their overall impact on the environment. This facilitates robust and reliable environmental risk assessment for MP control and pollution management.
1. Introduction
(Cole et al., 2011; Fauser et al., 2021; Stenger et al., 2021; Khalid et al., 2021), linking the effects of microplastics to ecological impacts in marine environments and limiting the ecosystem health, function, and services. There are now estimates regarding the extent of marine plastics in oceans and the main contributing sources (Jambeck et al., 2015), putting increased pressure on driving forward policy changes to improve waste management efforts. At the United Nations Environment Assembly (UNEA) in 2022, 170 members committed to generating an internationally legally binding agreement by 2024 to end plastic pollution. Despite the public awareness of plastic pollution and the significant
2. Methodology
3. Dynamic behaviour of microplastics
3.1. Sinking process of microplastics
the fluid and transport distance after their release from emission sources. This, in turn, contributes to a better understanding of the role played by freshwater systems such as rivers and lakes in transporting plastic debris
from terrestrial sources to the oceans (Nguyen et al., 2020; Hoellein et al., 2019).
3.1.1. Physical properties of microplastics
correlation between particle diameter and settling rate. The MP settling process in very slow-moving or static environmental flow systems (e.g. ponds) is governed by the balance of particle buoyancy, gravitational, and drag forces. The magnitude of the first two forces is positively correlated with particle density and diameter, while the drag force is primarily influenced by the cross-sectional shape and specific surface area of the particles. Hence, the higher particle density (
Analysis of microplastic settling processes and key findings.
| Plastic type and shape | Plastic density (
|
Plastic size (mm) | Solution chemistry | Terminal settling velocity (mm/s) | Conclusion | References |
| Pellet, sphere, fibre, fragment (PE, EPS, PVC, CoPA) | 1.01-1.31 |
|
Distilled water | 3.9-184 | Settling velocities varied with changes in particle size, density and shape; roundness has less influence than other parameters. | (Waldschlägelatr and Schüttrumpf, 2019) |
| Microplastics, macroplastics (PE, PP, PS, PVC, PET) | 0.02-1.5 | 0.58-2.51 5.30 | Distilled water | 1.6-35.2 | Weathering and non-sphere shapes harm the settling velocity of microplastics; the formulation describing the settlement of pellet, fragment and foam microplastics is not suitable for film plastics. | (Waldschläger et al., 2020) |
| Polymer foils (PE, PS, PET) | 0.92-1.40 | 4 | Deionized water | 7.5-28 | The settling velocity of microplastics was not changed significantly by the formation of biofilm; PET and PS plastics were more suitable to be colonised than PE microplastics; biofilm could facilitate the sorption of metal on plastic particles and improve the integral velocity. | (Leiser et al., 2020) |
| Granular or fragments (PET, HDPE, PP, PS), fibres (PVC), films (PE) | 0.95-1.432 | 0.63-3.48 | Deionized water | 5-105 | Particle shape strongly affects the sinking process of microplastics; biofouling could promote the settling process of low-density particles. | (Van Melkebeke et al., 2020) |
| Pellets, fragments, fibers (PE, PS, Acrylic) | 0.95-1.17 | 1-3 | Groundwater | 6.1 (average) | There were interactions between the influence of particle size, shape and density on the sinking process of microplastics. | (Hoellein et al., 2019) |
| Irregular polyurethane | 1.195 | 0.05-0.50 | Filtered river water | 0-30 | The settling velocity of microplastic aggregates decreased with the increase of the microbial colonization level. | (Nguyen et al., 2020) |
| Irregularly (PA, PMMA, PET)) and sphere (PS) shaped microplastics | 1.140, 1.190, and 1.390 | 0.006-0.251 | Deionized water |
|
The sinking velocity of irregularly shaped particles was lower than that of spheres of the same size range; a multiple quadratic regression depending on particle size and particle excess density can predict MP terminal sinking velocity with an average determination of
|
(Kaiser et al., 2019) |
| Irregular-shaped (PS) | 1.050 | 0.2-0.9 | Tap water | 0-2.5 | The shape irregularity can only decrease the sinking velocity of microplastics larger than 0.32 mm ; the positive correlation between settling velocity and size is insignificant as irregularity increases. | (Nguyen, 2021) |
| Sphere (PMMA) | 1.190 | 0.150-0.640 | Water | 0-60 | Net settling velocity is affected by particle characteristics and wave characteristics, simultaneously; increased inertial effect and decreased particle Reynolds number greatly increase settling velocity. | (De Leo et al., 2021) |
| Near-sphere and fragment (PS, PA, PET, PVC) | 1.050-1560 | 0.069-3.989 | Water | 0.85-122.44 | Particle size and density positively affect the settling velocity; particle irregularity, solution salinity, and dynamic conditions have negative impacts on particle settling. | (Z. Wang et al., 2021) |
| Irregular fragments (PS, PA66, PVC, PCL, PLLA, PBAT) | 1.03-1.38 | 0.3-2.2 | Lake water | 0.30-50 | Particle sizes and polymer densities are the most critical controls on settling rates; biofilm leads to a slight improvement in settling speed due to the short incubation periods; hydrophobicity of particles increases the buoyancy forces and decreases settling velocity. | (Elagami et al., 2020) |
| Fragment and fibres (PET, PVC and NP&A) | 1.01-1.44 | 0.02-4.94 | Deionised water with calls | 4-25.6 | Biofilm had the greatest impact on the increase of settling velocity of non-buoyant MPs by up to 130
|
(Mendrik et al., 2023) |
| Disk (square, rectangle, and triangle) MPs (PVC, PET, PC, ABS) | 1.038-1.343 |
|
Tap water | 19.6-48.8 | Lighter MP disks (
|
(Yang et al., 2023) |
| Microspheres (PE and Proprietary Polymer) | 1.10 and 1.30 | 0.028-0.063 | Filtered lake water | 0.93-2.20 | Particle size and lake hydrodynamics significantly affect MP residence time; aggregation of MPs with suspended matters and interactions with biota in the lake may increase the settling velocity of MPs; turbulent convection caused by seasonal temperature changes shorten the residence time of | (Elagami et al., 2023) |
| Plastic type and shape | Plastic density (
|
Plastic size (mm) | Solution chemistry | Terminal settling velocity (mm/s) | Conclusion | References |
| Spheres and fragments (PS) | 1.050 | 0.003 and 0.017 | M4 medium |
|
The residence time of small MPs (
|
(Gilfedder et al., 2023) |
| Fibres (PL) | 1.308 | 1-4 | Tap water | 0.1-0.5 | The settling velocity of MP fibres is significantly affected by their orientation and curliness; long fibres (
|
(Nguyen et al., 2022) |
| Fibres (PL) | 1.308 | 5-15 | Water | 17-20 | The settling of fibres was an orientationdependent process and their trajectories were profoundly affected by the secondary motions; the secondary motion was highly associated with fibre dimension and density; a new drag model was proposed to characterise the settling of fibres. | (Choi et al., 2022) |
density:
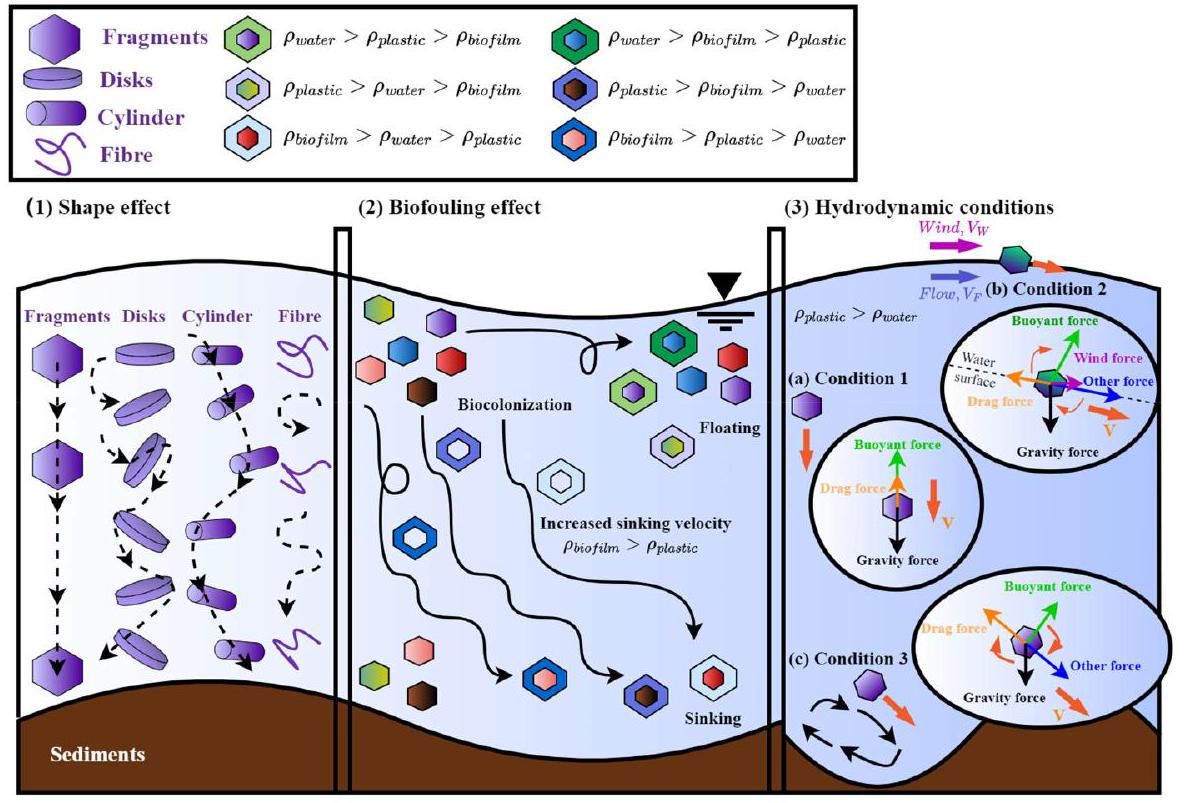
3.1.2. Biofouling
attachment, leading to a more significant response to an increase in diameter or density caused by bio-colonization (Fazey and Ryan, 2016). Furthermore, Ryan (2015) highlighted the importance of particle shape for the surface area-to-volume ratio, specifically for plastics with diameters exceeding 100-200 mm. This suggests that the biofouling effect might be more pronounced for MPs with complex shapes than spherical particles of the same volume. To further clarify the effect of biofouling as a function of MP size and shape, Van Melkebeke et al. (2020) examined the role of biofilm thickness (
3.1.3. Environmental condition
(Drummond et al., 2022; Drummond et al., 2020). In addition, high concentrations of suspended particles (e.g., clay and sand) in rivers foster interactions with MPs, especially after biofouling, to form flocs, which heavily influence MP dynamics by enhancing sediment settling and accumulation (Corcoran et al., 2020; Gholizadeh and Cera, 2022). Li et al. (2022) noted that riverine turbulence can amplify MP flocculation, especially at a shear rate above
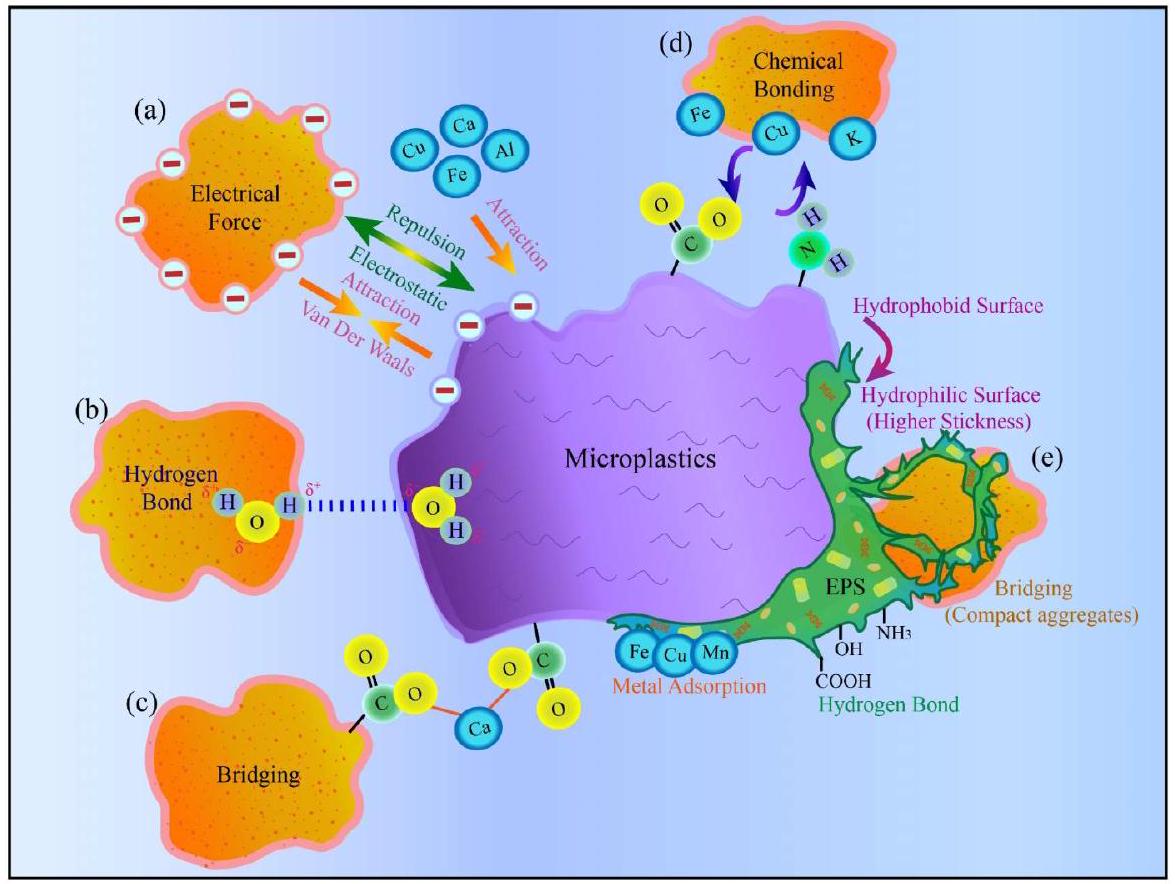
3.2. Aggregation process of microplastics
surface roughness) and chemical properties (e.g., surface charge distribution and electrostatic potential) (Li et al., 2018; Artham et al., 2009; Michels et al., 2018). Theoretical drivers of MP aggregation include a range of interacting forces such as electrical interaction, hydrogen bonding, chemical bonding, and bridging (Fig. 4) (Wang et al., 2015). Table 2 outlines the analysis of the key studies on MP aggregation in freshwater solutions, and a detailed overview of the studies considered here is given in Table S2. Key parameters and mechanisms involved in Fig. 4 and Table 2 are discussed in subsequent sections (Section 3.2.1-3.2.6).
3.2.1. Electrolyte properties
bridging effect during the aggregation process, making PE more sensitive to
3.2.2. Natural organic matter (NOM)
Analysis of microplastic aggregation processes and key findings.
| Aggregation type | Plastic size (
|
Concentration (mg/L) | Solution chemistry | Conclusion (mechanisms) | References |
| Homoaggregation (near-spherical PS) |
|
20.0 | Electrolyte solution
|
Aggregation kinetics agreed with DLVO theory and SchulzeeHardy rule | (Li et al., 2018) |
| Homoaggregation (spherical PS nano- and micro-particles) | 0.1, 1.0 | 10 | Electrolyte solution (
|
Physical process hugely influenced the aggregation mechanism of MPs via changing their stability in freshwater; while concentration, ionic valence, and hydrated ability played a dominant role in the aggregation process of NPs | (Sun et al., 2020) |
| Homoaggregation (PS particles) | 0.1 | 10-50 | Electrolyte solution (
|
Only electrolyte
|
(Cai et al., 2018) |
| Homoaggregation (PE and PS particles) | 0.2-0.75 | 20 | Electrolyte solution (NOM,
|
Salt types and ionic strength had significant effects on the stability of nanoscale plastics and thus their critical coagulation concentration; properties of their aggregation and stability followed DLVO theory and Schulze-Hardy rule | (Shams et al., 2020) |
| Homoaggregation (PS particles) | 0.09 | Electrolyte solution (NOM, NaCl, CaCl2, Na2SO4) | Aggregation of PS particles was dominated by electrostatic interactions and steric repulsions; NOM and pH played the most critical role in solutions with high and low ionic strength and NOM content separately | (J .Wang et al., 2021) | |
| Hetero- and Homoaggregation (PS nanoplastics and PE micropplastics withSS) | 0.10, 1000-1200 | 10,160 | Electrolyte solution (HA, NaCl) | Adsorption of PS plastic particles on the surface of large SS resulted in the formation of heteroaggregates; HA could improve the electrostatic repulsion and steric hindrance between particles and thus hindered the occurrence of aggregation; Brownian motion and buoyant force dominated the movement of PSNPs and PEMPs in water respectively; millimetre-sized PEMPs always floated on water surface even when SS adsorbed their surface | (Y. Li et al., 2019) |
| Heteroaggregation (PS beads with biogenic particles) | 700-900 | 50 beads/L. | Seawater | The formation of biofilms on the surface of microplastics facilitated their aggregation process and diversified the bacterial communities on them | (Michels et al., 2018) |
| Heteroaggregation (HDPE and PP fragments with algae) | 400-1000 | 400 | Culture medium | Microalgae colonization changed the surface properties of PP and HDPE fragments; different microalgae EPS had distinct aggregation properties with PP and HDPE | (Lagarde et al., 2016) |
| Homo- and hetero-aggregation (microalgae, EPS, irregularly shaped PMMA and PS particles) | <106, 106-250 | 12.5, 125 | Culture medium | Heteroaggregation was dependent on the yield production of EPS and the size and type of EPS and MPs | (Cunha et al., 2019) |
| Homoaggregation (irregular PE particles) and hetero-aggregation (PE particles with chlorite, illite, kaolinite, and montmorillonite clays) | 25.6 | 20 | Deionized water with different hydrochemical conditions (
|
Aged PEs were more stable than pristine PEs in monovalent electrolyte solutions; the interaction of PEs and clay minerals was dominated by electrostatic repulsion; hetero-aggregation-settling of PEs was facilitated by the increased NaCl concentration | (Wang et al., 2023) |
| Heteroaggregation (PP, HDPE, PET, PVC and PS with riverine particles) | 1527-1935 | – | Unfiltered river water | Large MPs (
|
(Stead and Bond, 2023) |
destabilization of micro-PS by bridge attraction and intermolecular bridging (
et al., 2019)) and overlooked the diversity of NOM existing in natural environments. Beyond HA, protein-like substances, prevalent in aquatic systems, also exhibit strong stabilization effects on PS nanoparticles
3.2.3. Solution
3.2.4. Physical properties of microplastics
their aggregation by altering colloid interaction energy and electrophoretic mobility, leading to unique aggregation propensity in specific solutions (Afrooz et al., 2013; Bhattacharjee et al., 2000; Li and Ma, 2018). However, to this date, very limited studies focused on applying these findings to MPs, highlighting a crucial area for future research.
3.2.5. Biofilm & biomass
permeability, or directly adsorbed onto the cell walls of algae via intermolecular interactions and adsorptive forces. This leads to rapid sedimentation of MPs from surface water to bottom sediment (Nguyen et al., 2020; Lagarde et al., 2016; Peller et al., 2021; Michels et al., 2018).
3.2.6. Weathering
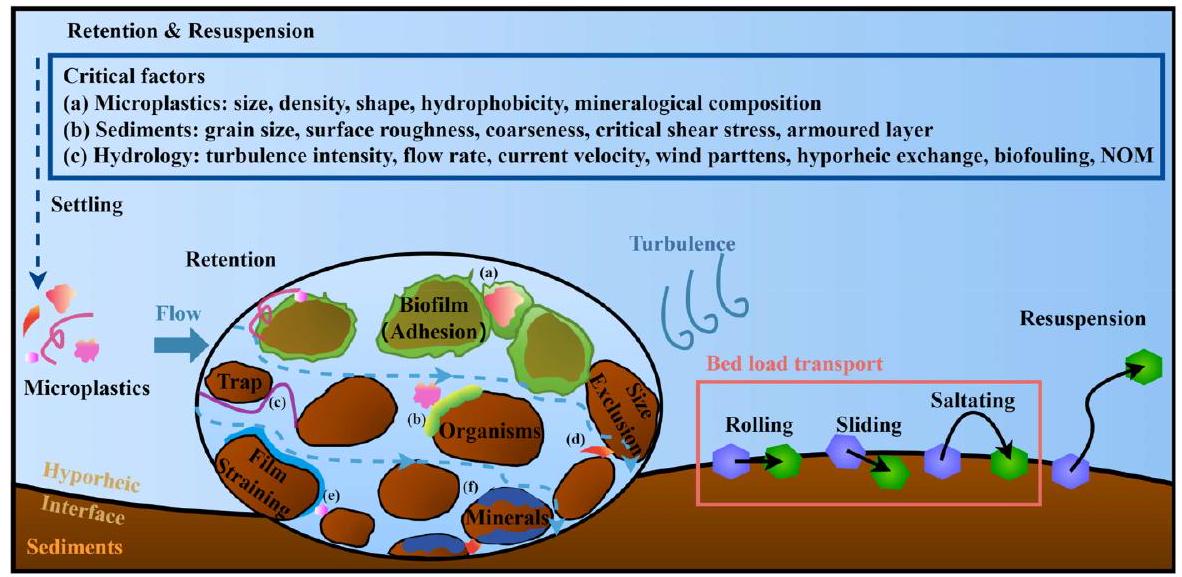
3.3. Retention
3.3.1. Physical properties of microplastics
particle size (Besseling et al., 2017; Nizzetto et al., 2016). Furthermore, bottom sediments frequently exhibit a prevalence of low-density MPs (e. g. PE and PP), which could be attributed to the robust interparticle forces including charge attraction, that induce interactions with minerals, biofouling, and substance adsorption (Corcoran et al., 2015; Ballent et al., 2016; B. He et al., 2020).
Analysis of abundance and characteristics of microplastics in freshwater sediments.
| Source area | Sampling depth (cm) | Dominant shape | Size (
|
Density (
|
Abundance (particles/ kg) | References | ||||||||
| Beijiang River, China | 2 | Not indicated | <5000 | PE, PP and Copolymer | 178-544 | (Wang et al., 2017) | ||||||||
|
|
|
|
0-1.8 | 6350 (pre-flooding) | (Hurley et al., 2018) | ||||||||
|
Fragment |
|
PE | 165 (baseflow conditions) | (Tibbetts et al., 2018) | |||||||||
| 12 |
|
|
100-629 (March) |
|
||||||||||
| 0-2 | Fiber, fragment and pellet | <1000 | PET, PS and PE | 50-195 | (Jiang et al., 2019) | |||||||||
| Amazon Rivers, Brazil | 5-10 | Fiber (only) | 0-1000 (major) | Not indicated | River) | (Gerolin et al., 2020) | ||||||||
| 0-1000 (major) | 1524-1738 (Solimões river) | |||||||||||||
| 1000-2000 (major) | 417-2101 (Amazon rivers) | (He et al., 2020a) | ||||||||||||
| Brisbane River, Australia | 0-3 | Film, fibre and fragment | <5000 | PE, PP and PS | 10-270 (Dec. 2017) | |||||||||
| Surface | Fragment and fiber | <5000 | PE and PP | 6-2444 (normalised) | (Corcoran et al., 2020) | |||||||||
| Thames River, Canada | 5 | fragment and film | 1000-5000 | PE, PP, PVA, PVC | 8-1100 | (Kabir et al., 2022) | ||||||||
| Lake Ontario, Canada | <8 | Irregular, Wispy | 250-1000 (major) | PE, PP and NC | 4635 particles | (Corcoran et al., 2015) | ||||||||
| Taihu Lake, China | Surface | Fibre | 100-1000 (major) | Cellophane | 11.0-234.6 | (Su et al., 2016) | ||||||||
| Vembanad Lake, India | Surface | Film and foam | 100-500 (major) | HDPE, LDPE, PP, PS | 252.8 particles
|
(Sruthy and Ramasamy, 2017) | ||||||||
| Edgbaston Pool, UK | Surface | Fibre and film | <5000 | none | 250-300 | (Vaughan et al., 2017) | ||||||||
| Dongting Lake, China | 0-2 | Fibre | <500 (prevalent) | PS, PET | 200-1150 | (Jiang et al., 2018) | ||||||||
| Poyang Lake, China | Surface | Fibre and film | 100-500 (major) | PP and PE | 54-506 | (Yuan et al., 2019) | ||||||||
| Taihu Lake, China | Top, by a Peterson, grab | Fragment, fibre, film and pellet |
|
PVC, PE, PS, PP | 464.26-1380.69, 893.48 (mean) | (Zhang et al., 2021) | ||||||||
| Lake Hawassa, Ethiopia | top 3 | Fibres (90 %), fragments (5
|
100-500 (major) | PET (82 %), PE (15 %) and PS (3 %) | 11-74 particles/m3 | (Jeevanandam et al., 2022) | ||||||||
| Yangtze Estuary, China | 0-2 | Fibre | 1161.23 (mean) | Rayon (most) | 143 | (F. Wu et al., 2020) | ||||||||
| Derwent Estuary, Australia | 0-104 | Fibre (87 %) | 63-1000 (major) | Not indicated | 2430, 4200 | (Willis et al., 2017) | ||||||||
| Changjiang Estuary, China | 5-10 | Fibre (93 %) | <1000 (58 %) | Rayon, polyester, and acrylic | 20-340 | (Peng et al., 2017) | ||||||||
| Anzali Wetland, Iran | Surface | Fibre | 100-200 (major) | 0.90-1.40 | 140-2820 (June) 110-3690 (January) | (Rasta et al., 2020) | ||||||||
| Mangrove, China | 1 | Fibre (most) and fragment | 500-1000 (highest) | PP, PE and PS | 273-3520 | (L. Zhang et al., 2020) | ||||||||
| Wetland, USA | 5 | Fiber (88 %) | <5000 | PS, PE and synthetic rubber | 1270 (average) | (Helcoski et al., 2020) | ||||||||
| Three Gorges Reservoir, China Jiaozhou Bay, China | Not indicated | Sheet and line | 1000-5000 (major) | PP, PE and PET |
|
(Zhang et al., 2017) | ||||||||
| 0-45 | Fibre | 100-490 (major) | PE (domain), Rayon and PET | 0-30 | (Zheng et al., 2020) |
3.3.2. Aquatic environment
debris in areas of weak hydrodynamic activity (Browne et al., 2010; Zhang et al., 2017). Fischer et al. (2016) observed a similar retention pattern, noting that the presence of organic matter enhances cohesion and adhesion between MPs and sediments. Additionally, the abundance of organic matter expedites biofilm formation on MP surfaces, transitioning them from hydrophobic to strongly hydrophilic and altering their polarities similar to sediments, thereby increasing their affinity with sediments (Van Melkebeke et al., 2020). This biofilm formation may also trigger biodegradation in long-term deposited MPs, particularly in deeper sediment layers, breaking them down into smaller fragments that further penetrate sediment layers through pore spaces (Niu et al., 2021). Vermaire et al. (2017) reported a competitive interaction between organic matter and hydrodynamics, where organic matter encourages the formation of larger, denser MP aggregates that are more likely to settle in the water column, reducing the impact of hydrodynamics on MP behaviour. Besides, some literature has highlighted the significance of sediment grain-scale flows and pressure-driven flows on particle retention (Fries and Taghon, 2010; Farizan et al., 2019). However, little information is available about how these specific flows impact MP retention.
3.4. Resuspension
3.4.1. Physical properties of microplastics
turbulent with
3.4.2. Hydraulic conditions
during floods in a controlled artificial flume, highlighting the critical role of the active layer evolution and flow regime characteristics in MP resuspension in coarse granular rivers. Similarly, Hurley et al. (2018) also observed a marked rise in MP concentration in the water column during the rainy seasons, attributed to the intensified flow velocities that lift more deposited MPs. Given that generally higher abundance of MPs can be found in surface sediments compared to deeper layers (Section 3.3.1), it is plausible that the majority of MPs entering natural aquatic systems undergo a cycle of settling on sediment surface under low flow conditions and being resuspended into the water column under high flow conditions. This highlights the strong seasonality effects on MP sedimentation and resuspension mechanisms, making it difficult to permanently retain MPs in deeper fluvial sediments (Li et al., 2022).
4. Microplastics transport in freshwater systems
4.1. Microplastic transport in riverine systems
4.2. Microplastic transport in lake systems
predominantly fibrous (
model environmental characteristics, such as flow patterns, wind exposure, precipitation, inflow from neighbouring rivers and streams, lake morphology, and sediment properties on MP dynamic behaviour in lakes. This approach would be compatible with current theories applied to the study of river systems, enhancing our understanding of MP transport dynamics and fate in lake freshwater systems.
5. Existing challenges and future perspectives
5.1. Modelling the natural environmental conditions and laboratory studies
5.2. Correlation between microplastic dynamic and environmental processes
on flow patterns driving MP distributions. Furthermore, examining how hydrodynamic changes influence MP dynamics, especially through influencing NOM and SS levels, and thus controlling biofouling and weathering, is another crucial knowledge gap. Although tracking the dynamics of MPs throughout the aquatic system is challenging, adopting mesocosm setups similar to (Hurley et al., 2018) for in-situ observations could provide comprehensive information on intricate weathering processes and fluid dynamics impacts on MP behaviour, thereby compensating for the inherent limitations of existing laboratory-based data.
5.3. Interactions between microplastic dynamic behaviours
6. Conclusions
freshwater systems. This study conducted a critical analysis of the current state-of-the-art studies on the dynamic behaviours of MPs, including sedimentation, aggregation, retention, and resuspension, that influence their transport and fate in freshwater systems. Integrating empirical findings and theoretical insights, a comprehensive analysis of the intricate interactions between MP properties and environmental conditions was provided, emphasising the complexity of MP dynamics and environmental fate in freshwater environments. Our findings not only elucidate the key underlying mechanisms governing dynamic behaviour of MPs, but also identify key research gaps on the understanding of MP transport processes and environmental fate, facilitating robust environmental management and ecological risk assessment.
methodological advances are essential to unravel the ‘black box’ of MP transport, facilitating a comprehensive understanding of how MP transport and interact under a diverse range of hydrology, hydrodynamic, and ecological factors.
CRediT authorship contribution statement
Declaration of competing interest
Data availability
Acknowledgement
Supplementary materials
References
Alimi, O.S., Farner Budarz, J., Hernandez, L.M., Tufenkji, N., 2018. Microplastics and nanoplastics in aquatic environments: aggregation, deposition, and enhanced contaminant transport. Environ. Sci. Technol. 52 (4), 1704-1724. https://doi.org/ 10.1021/acs.est.7b05559.
Augusto-Silva, P.B., MacIntyre, S., de Moraes Rudorff, C., Cortés, A., Melack, J.M., 2019. Stratification and mixing in large floodplain lakes along the lower Amazon River. J. Great Lakes Res. 45 (1), 61-72. https://doi.org/10.1016/j.jglr.2018.11.001.
Ballent, A., Corcoran, P.L., Madden, O., Helm, P.A., Longstaffe, F.J., 2016. Sources and sinks of microplastics in Canadian Lake Ontario nearshore, tributary and beach
sediments. Mar. Pollut. Bull. 110 (1), 383-395. https://doi.org/10.1016/j. marpolbul.2016.06.037.
Ballent, A., Pando, S., Purser, A., Juliano, M.F., Thomsen, L., 2013. Modelled transport of benthic marine microplastic pollution in the Nazaré Canyon. Biogeosciences 10 (12), 7957-7970.
Ballent, A., Purser, A., de Jesus Mendes, P., Pando, S., Thomsen, L., 2012. Physical transport properties of marine microplastic pollution. Biogeosciences Discuss. 9 (12), 18755-18798. https://doi.org/10.5194/bgd-9-18755-2012.
Barnes, D.K.A., Milner, P., 2005. Drifting plastic and its consequences for sessile organism dispersal in the Atlantic Ocean. Mar. Biol. 146 (4), 815-825. https://doi. org/10.1007/s00227-004-1474-8.
Beletsky, D., Saylor, J.H., Schwab, D.J., 1999. Mean circulation in the Great Lakes. J. Great Lakes Res. 25 (1), 78-93. https://doi.org/10.1016/S0380-1330(99)707185.
Bissell, M.J., Barcellos-Hoff, M.H., 1987. The influence of extracellular matrix on gene expression: is structure the message? J. Cell Sci. 343 (SUPPL. 8), 327-343. https:// doi.org/10.1242/jcs.1987.supplement_8.18.
Boos, J.P., Gilfedder, B.S., Frei, S., 2021. Tracking microplastics across the streambed interface: using laser-induced-fluorescence to quantitatively analyze microplastic transport in an experimental flume. Water Resour. Res. 57 (12), 1-10. https://doi. org/10.1029/2021WR031064.
Browne, M.A., Galloway, T.S., Thompson, R.C., 2010. Spatial patterns of plastic debris along estuarine shorelines. Environ. Sci. Technol. 44 (9), 3404-3409. https://doi. org/10.1021/es903784e.
Buffle, I., Leppard, G.G., 1995. Characterization of aquatic colloids and macromolecules. 1. Structure and behavior of colloidal material. Environ. Sci. Technol. 29 (9), 2169-2175. https://doi.org/10.1021/es00009a004.
Butt, H.J., Jaschke, M., Ducker, W., 1995. Measuring surface forces in aqueous electrolyte solution with the atomic force microscope. Bioelectrochem. Bioenerg. 38 (1), 191-201. https://doi.org/10.1016/0302-4598(95)01800-T.
Campanale, C., et al., 2020. Microplastics and their possible sources: the example of Ofanto river in southeast Italy. Environ. Pollut. 258, 113284 https://doi.org/ 10.1016/j.envpol.2019.113284.
Carpenter, E.J., Smith, K.L., 1972. Plastics on the Sargasso sea surface. Science (80-.) 175 (4027), 1240-1241. https://doi.org/10.1126/science.175.4027.1240.
Chen, Y., et al., 2021. Transport and fate of microplastics in constructed wetlands: a microcosm study. J. Hazard. Mater. 415 (March), 125615 https://doi.org/10.1016/ j.jhazmat.2021.125615.
Chowdhury, I., Mansukhani, N.D., Guiney, L.M., Hersam, M.C., Bouchard, D., 2015. Aggregation and stability of reduced graphene oxide: complex roles of divalent cations, pH, and natural organic matter. Environ. Sci. Technol. 49 (18), 10886-10893. https://doi.org/10.1021/acs.est.5b01866. Sep.
Chubarenko, I., Bagaev, A., Zobkov, M., Esiukova, E., 2016. On some physical and dynamical properties of microplastic particles in marine environment. Mar. Pollut. Bull. 108 (1-2), 105-112. https://doi.org/10.1016/j.marpolbul.2016.04.048.
Chubarenko, I., Efimova, I., Bagaeva, M., Bagaev, A., Isachenko, I., 2020. On mechanical fragmentation of single-use plastics in the sea swash zone with different types of bottom sediments: insights from laboratory experiments. Mar. Pollut. Bull. 150 (November 2019) https://doi.org/10.1016/j.marpolbul.2019.110726.
Cole, M., Lindeque, P., Halsband, C., Galloway, T.S., 2011. Microplastics as contaminants in the marine environment: a review. Mar. Pollut. Bull. 62 (12), 2588-2597. https:// doi.org/10.1016/j.marpolbul.2011.09.025.
Corcoran, P.L., Belontz, S.L., Ryan, K., Walzak, M.J., 2020. Factors controlling the distribution of microplastic particles in benthic sediment of the Thames River, Canada. Environ. Sci. Technol. 54 (2), 818-825. https://doi.org/10.1021/acs. est.9b04896.
Corcoran, P.L., Norris, T., Ceccanese, T., Walzak, M.J., Helm, P.A., Marvin, C.H., 2015. Hidden plastics of Lake Ontario, Canada and their potential preservation in the sediment record. Environ. Pollut. 204, 17-25. https://doi.org/10.1016/j. envpol.2015.04.009.
Cunha, C., Faria, M., Nogueira, N., Ferreira, A., Cordeiro, N., 2019. Marine vs freshwater microalgae exopolymers as biosolutions to microplastics pollution. Environ. Pollut. 249, 372-380. https://doi.org/10.1016/j.envpol.2019.03.046.
Cunha, C., Silva, L., Paulo, J., Faria, M., Nogueira, N., Cordeiro, N., 2020. Microalgalbased biopolymer for nano- and microplastic removal: a possible biosolution for
wastewater treatment. Environ. Pollut. 263 https://doi.org/10.1016/j. envpol.2020.114385.
Daily, J., Hoffman, M.J., 2020. Modeling the three-dimensional transport and distribution of multiple microplastic polymer types in Lake Erie. Mar. Pollut. Bull. 154 (January) https://doi.org/10.1016/j.marpolbul.2020.111024.
Daily, J., Onink, V., Jongedijk, C.E., Laufkötter, C., Hoffman, M.J., 2021. Incorporating terrain specific beaching within a lagrangian transport plastics model for Lake Erie. Microplast. Nanoplast. 1 (1), 1-13. https://doi.org/10.1186/s43591-021-00019-7.
de Carvalho, A.R., et al., 2021. Urbanization and hydrological conditions drive the spatial and temporal variability of microplastic pollution in the Garonne River. Sci. Total Environ. 769 https://doi.org/10.1016/j.scitotenv.2020.144479.
De Leo, A., Cutroneo, L., Sous, D., Stocchino, A., 2021. Settling velocity of microplastics exposed to wave action. J. Mar. Sci. Eng. 9 (2), 1-16. https://doi.org/10.3390/ jmse9020142.
Demiral, D., Albayrak, I., Turowski, J.M., Boes, R.M., 2022. Particle saltation trajectories in supercritical open channel flows: Roughness effect. Earth Surf. Process. Landf. 47 (15), 3588-3610. https://doi.org/10.1002/esp.5475.
Dong, Z., Qiu, Y., Zhang, W., Yang, Z., Wei, L., 2018. Size-dependent transport and retention of micron-sized plastic spheres in natural sand saturated with seawater. Water Res 143, 518-526. https://doi.org/10.1016/j.watres.2018.07.007.
Drago, C., Pawlak, J., Weithoff, G., 2020. Biogenic aggregation of small microplastics alters their ingestion by a common freshwater micro-invertebrate. Front. Environ. Sci. 8 (December) https://doi.org/10.3389/fenvs.2020.574274.
Dris, R., Gasperi, J., Rocher, V., Tassin, B., 2018. Synthetic and non-synthetic anthropogenic fibers in a river under the impact of Paris Megacity: sampling methodological aspects and flux estimations. Sci. Total Environ. 618, 157-164. https://doi.org/10.1016/j.scitotenv.2017.11.009.
Drummond, J.D., Nel, H.A., Packman, A.I., Krause, S., 2020. Significance of hyporheic exchange for predicting microplastic fate in rivers. Environ. Sci. Technol. Lett. 7 (10), 727-732. https://doi.org/10.1021/acs.estlett.0c00595.
Duan, J., et al., 2021. Weathering of microplastics and interaction with other coexisting constituents in terrestrial and aquatic environments. Water Res., 117011
Eerkes-Medrano, D., Thompson, R.C., Aldridge, D.C., 2015. Microplastics in freshwater systems: a review of the emerging threats, identification of knowledge gaps and prioritisation of research needs. Water Res. 75, 63-82. https://doi.org/10.1016/j. watres.2015.02.012.
Egessa, R., Nankabirwa, A., Basooma, R., Nabwire, R., 2020. Occurrence, distribution and size relationships of plastic debris along shores and sediment of northern Lake Victoria. Environ. Pollut. 257, 113442 https://doi.org/10.1016/j. envpol.2019.113442.
Elagami, H., et al., 2022. Measurement of microplastic settling velocities and implications for residence times in thermally stratified lakes. Limnol. Oceanogr. 934-945. https://doi.org/10.1002/lno.12046. Agarwal 2020.
Elagami, H., Frei, S., Boos, J.P., Trommer, G., Gilfedder, B.S., 2023. Quantifying microplastic residence times in lakes using mesocosm experiments and transport modelling. Water Res. 229 (November 2022), 119463 https://doi.org/10.1016/j. watres.2022.119463.
Enfrin, M., Lee, J., Gibert, Y., Basheer, F., Kong, L., Dumée, L.F., 2020. Release of hazardous nanoplastic contaminants due to microplastics fragmentation under shear stress forces. J. Hazard. Mater. 384 (September 2019), 121393 https://doi.org/ 10.1016/j.jhazmat.2019.121393.
Farizan, A., Yaghoubi, S., Firoozabadi, B., Afshin, H., 2019. Effect of an obstacle on the depositional behaviour of turbidity currents. J. Hydraul. Res. 57 (1), 75-89. https:// doi.org/10.1080/00221686.2018.1459891.
Faulstich, L., Prume, J.A., Arendt, R., Reinhardt-Imjela, C., Chifflard, P., Schulte, A., 2022. Microplastics in Namibian river sediments – a first evaluation. Microplastics and Nanoplastics 2 (1), 1-17. https://doi.org/10.1186/s43591-022-00043-1.
Fauser, P., Vorkamp, K., Strand, J., 2021. Residual additives in marine microplastics and their risk assessment – a critical review. Mar. Pollut. Bull. 177, 113467 https://doi. org/10.1016/j.marpolbul.2022.113467. October2022.
Fazey, F.M.C., Ryan, P.G., 2016. Biofouling on buoyant marine plastics: an experimental study into the effect of size on surface longevity. Environ. Pollut. 210, 354-360. https://doi.org/10.1016/j.envpol.2016.01.026.
Ferdowsi, B., Ortiz, C.P., Houssais, M., Jerolmack, D.J., 2017. River-bed armouring as a granular segregation phenomenon. Nat. Commun. 8 (1) https://doi.org/10.1038/ s41467-017-01681-3.
Fischer, E.K., Paglialonga, L., Czech, E., Tamminga, M., 2016. Microplastic pollution in lakes and lake shoreline sediments – a case study on Lake Bolsena and Lake Chiusi (central Italy. Environ. Pollut. 213, 648-657. https://doi.org/10.1016/j. envpol.2016.03.012.
Flemming, H.C., Wingender, J., 2010. The biofilm matrix. Nat. Rev. Microbiol. 8 (9), 623-633. https://doi.org/10.1038/nrmicro2415.
Gao, L., Su, Y., Yang, L., Li, J., Bao, R., Peng, L., 2022. Sorption behaviors of petroleum on micro-sized polyethylene aging for different time in seawater. Sci. Total Environ. 808, 152070 https://doi.org/10.1016/j.scitotenv.2021.152070.
Gerolin, C.R., Pupim, F.N., Sawakuchi, A.O., Grohmann, C.H., Labuto, G., Semensatto, D., 2020. Microplastics in sediments from Amazon rivers, Brazil. Sci. Total Environ. 749, 141604 https://doi.org/10.1016/j.scitotenv.2020.141604.
Gholizadeh, M., Cera, A., 2022. Microplastic contamination in the sediments of Qarasu estuary in Gorgan Bay, south-east of Caspian Sea, Iran. Sci. Total Environ. 838 (February), 155913 https://doi.org/10.1016/j.scitotenv.2022.155913.
Gilfedder, B.S., et al., 2023. Filter feeders are key to small microplastic residence times in stratified lakes: a virtual experiment. Sci. Total Environ. 890 (May), 164293 https:// doi.org/10.1016/j.scitotenv.2023.164293.
Goodarzi, D., Mohammadian, A., Pearson, J., Abolfathi, S., 2022. Numerical modelling of hydraulic efficiency and pollution transport in waste stabilization ponds. Ecol. Eng. 182 (June 2021), 106702 https://doi.org/10.1016/j.ecoleng.2022.106702.
He, B., Goonetilleke, A., Ayoko, G.A., Rintoul, L., 2020a. Abundance, distribution patterns, and identification of microplastics in Brisbane River sediments, Australia. Sci. Total Environ. 700, 134467 https://doi.org/10.1016/j.scitotenv.2019.134467.
He, B., Wijesiri, B., Ayoko, G.A., Egodawatta, P., Rintoul, L., Goonetilleke, A., 2020b. Influential factors on microplastics occurrence in river sediments. Sci. Total Environ. 738, 139901 https://doi.org/10.1016/j.scitotenv.2020.139901.
Helcoski, R., Yonkos, L.T., Sanchez, A., Baldwin, A.H., 2020. Wetland soil microplastics are negatively related to vegetation cover and stem density. Environ. Pollut. 256, 113391 https://doi.org/10.1016/j.envpol.2019.113391.
Hendrickson, E., Minor, E.C., Schreiner, K., 2018. Microplastic abundance and composition in western lake superior as determined via microscopy, Pyr-GC/MS, and FTIR. Environ. Sci. Technol. 52 (4), 1787-1796. https://doi.org/10.1021/acs. est.7b05829.
Hoellein, T.J., Shogren, A.J., Tank, J.L., Risteca, P., Kelly, J.J., 2019. Microplastic deposition velocity in streams follows patterns for naturally occurring allochthonous particles. Sci. Rep. 9 (1), 1-12. https://doi.org/10.1038/s41598-019-40126-3.
Hoffman, M.J., Hittinger, E., 2017. Inventory and transport of plastic debris in the Laurentian Great Lakes. Mar. Pollut. Bull. 115 (1-2), 273-281. https://doi.org/ 10.1016/j.marpolbul.2016.11.061.
Homann, H., Bec, J., Grauer, R., 2013. Effect of turbulent fluctuations on the drag and lift forces on a towed sphere and its boundary layer. J. Fluid Mech. 721, 155-179. https://doi.org/10.1017/jfm.2013.66.
Horton, A.A., Dixon, S.J., 2018. Microplastics: an introduction to environmental transport processes. Wiley Interdiscip. Rev. Water 5 (2), e1268. https://doi.org/ 10.1002/wat2.1268.
Hurley, R., Woodward, J., Rothwell, J.J., 2018. Microplastic contamination of river beds significantly reduced by catchment-wide flooding. Nat. Geosci. 11 (4), 251-257. https://doi.org/10.1038/s41561-018-0080-1.
Jambeck, J., et al., 2015. Plastic waste inputs from land into the ocean. Mar. Pollut. 347 (6223), 768.
Ji, Z.-G., 2017. Hydrodynamics and Water Quality: Modeling Rivers, Lakes, and Estuaries. John Wiley & Sons.
Jiang, C., et al., 2018. Microplastics in sediment and surface water of west dongting lake and south dongting lake: abundance, source and composition. Int. J. Environ. Res. Public Health 15 (10). https://doi.org/10.3390/ijerph15102164.
Jiang, C., et al., 2019. Microplastic pollution in the rivers of the Tibet Plateau. Environ. Pollut. 249, 91-98. https://doi.org/10.1016/j.envpol.2019.03.022.
Junaid, M., Wang, J., 2021. Interaction of nanoplastics with extracellular polymeric substances (EPS) in the aquatic environment: a special reference to eco-corona formation and associated impacts. Water Res. 201 (May), 117319 https://doi.org/ 10.1016/j.watres.2021.117319.
Kaiser, D., Estelmann, A., Kowalski, N., Glockzin, M., Waniek, J.J., 2019. Sinking velocity of sub-millimeter microplastic. Mar. Pollut. Bull. 139 (December 2018), 214-220. https://doi.org/10.1016/j.marpolbul.2018.12.035.
Kaiser, D., Kowalski, N., Waniek, J.J., 2017. Effects of biofouling on the sinking behavior of microplastics. Environ. Res. Lett. 12 (12) https://doi.org/10.1088/1748-9326/ aa8e8b.
Khalid, N., et al., 2021. Linking effects of microplastics to ecological impacts in marine environments. Chemosphere 264, 128541. https://doi.org/10.1016/j. chemosphere.2020.128541.
Khatmullina, L., Isachenko, I., 2017. Settling velocity of microplastic particles of regular shapes. Mar. Pollut. Bull. 114 (2), 871-880. https://doi.org/10.1016/j. marpolbul.2016.11.024.
Kowalski, N., Reichardt, A.M., Waniek, J.J., 2016. Sinking rates of microplastics and potential implications of their alteration by physical, biological, and chemical
factors. Mar. Pollut. Bull. 109 (1), 310-319. https://doi.org/10.1016/j. marpolbul.2016.05.064.
Kukulka, T., Proskurowski, G., Morét-Ferguson, S., Meyer, D.W., Law, K.L., 2012. The effect of wind mixing on the vertical distribution of buoyant plastic debris. Geophys. Res. Lett. 39 (7), 1-6. https://doi.org/10.1029/2012GL051116.
Lagarde, F., Olivier, O., Zanella, M., Daniel, P., Hiard, S., Caruso, A., 2016. Microplastic interactions with freshwater microalgae: hetero-aggregation and changes in plastic density appear strongly dependent on polymer type. Environ. Pollut. 215, 331-339. https://doi.org/10.1016/j.envpol.2016.05.006.
Lead, J.R., Davison, W., Hamilton-Taylor, J., Buffle, J., 1997. Characterizing colloidal material in natural waters. Aquat. Geochem. 3 (3), 213-232. https://doi.org/ 10.1023/A:1009695928585.
Lenaker, P.L., Baldwin, A.K., Corsi, S.R., Mason, S.A., Reneau, P.C., Scott, J.W., 2019. Vertical distribution of microplastics in the water column and surficial sediment from the Milwaukee River Basin to Lake Michigan. Environ. Sci. Technol. 53 (21), 12227-12237. https://doi.org/10.1021/acs.est.9b03850.
Lenaker, P.L., Corsi, S.R., Mason, S.A., 2021. Spatial distribution of microplastics in surficial benthic sediment of lake Michigan and Lake Erie. Environ. Sci. Technol. 55 (1), 373-384. https://doi.org/10.1021/acs.est.0c06087.
Li, J., Song, Y., Cai, Y., 2020. Focus topics on microplastics in soil: analytical methods, occurrence, transport, and ecological risks. Environ. Pollut. 257, 113570 https://doi. org/10.1016/j.envpol.2019.113570.
Li, K., Ma, H., 2018. Deposition dynamics of rod-shaped colloids during transport in porous media under favorable conditions. Langmuir 34 (9), 2967-2980. https://doi. org/10.1021/acs.langmuir.7b03983.
Li, S., Liu, H., Gao, R., Abdurahman, A., Dai, J., Zeng, F., 2018. Aggregation kinetics of microplastics in aquatic environment: complex roles of electrolytes, pH , and natural organic matter. Environ. Pollut. 237, 126-132. https://doi.org/10.1016/j. envpol.2018.02.042.
Li, W., et al., 2019a. Colonization characteristics of bacterial communities on plastic debris influenced by environmental factors and polymer types in the Haihe Estuary of Bohai Bay, China. Environ. Sci. Technol. 53 (18), 10763-10773. https://doi.org/ 10.1021/acs.est.9b03659.
Li, Y., et al., 2019b. Interactions between nano/micro plastics and suspended sediment in water: implications on aggregation and settling. Water Res. 161, 486-495. https:// doi.org/10.1016/j.watres.2019.06.018.
Liu, K., Courtene-Jones, W., Wang, X., Song, Z., Wei, N., Li, D., 2020. Elucidating the vertical transport of microplastics in the water column: a review of sampling methodologies and distributions. Water Res. 186, 116403 https://doi.org/10.1016/ j.watres.2020.116403.
Liu, Y., et al., 2022b. Application of a microplastic trap to the determination of the factors controlling the lakebed deposition of microplastics. Sci. Total Environ. 843 (March), 156883 https://doi.org/10.1016/j.scitotenv.2022.156883.
Liu, Y., Hu, Y., Yang, C., Chen, C., Huang, W., Dang, Z., 2019. Aggregation kinetics of UV irradiated nanoplastics in aquatic environments. Water Res. 163, 114870 https:// doi.org/10.1016/j.watres.2019.114870.
Lu, S., et al., 2018. Impact of water chemistry on surface charge and aggregation of polystyrene microspheres suspensions. Sci. Total Environ. 630, 951-959. https:// doi.org/10.1016/j.scitotenv.2018.02.296.
Mahdian, M., et al., 2023. Modelling impacts of climate change and anthropogenic activities on inflows and sediment loads of wetlands: case study of the Anzali wetland. Sci. Rep. 13 (1), 1-18. https://doi.org/10.1038/s41598-023-32343-8.
Mahdian, M., Noori, R., Salamattalab, M.M., Heggy, E., Bateni, S.M., Nohegar, A., et al., 2024. Anzali wetland crisis: Unraveling the decline of Iran’s ecological gem. J. Geophys. Res. Atmos. 129 https://doi.org/10.1029/2023JD039538 e2023JD039538.
Mancini, M., Serra, T., Colomer, J., Solari, L., 2023. Suspended sediments mediate microplastic sedimentation in unidirectional flows. Sci. Total Environ. 890 (May), 164363 https://doi.org/10.1016/j.scitotenv.2023.164363.
Mani, T., Hauk, A., Walter, U., Burkhardt-Holm, P., 2015. Microplastics profile along the Rhine River. Sci. Rep. 5 (December), 1-7. https://doi.org/10.1038/srep17988.
Mendrik, F., Fernández, R., Hackney, C.R., Waller, C., Parsons, D.R., 2023. Non-buoyant microplastic settling velocity varies with biofilm growth and ambient water salinity. Commun. Earth Environ. 4 (1), 1-9. https://doi.org/10.1038/s43247-023-00690-z.
Miao, L., et al., 2021. Effects of biofilm colonization on the sinking of microplastics in three freshwater environments. J. Hazard. Mater. 413 (December 2020), 125370 https://doi.org/10.1016/j.jhazmat.2021.125370.
Michels, J., Stippkugel, A., Lenz, M., Wirtz, K., Engel, A., 2018. Rapid aggregation of biofilm-covered microplastics with marine biogenic particles. Proc. R. Soc. B Biol. Sci. 285 (1885) https://doi.org/10.1098/rspb.2018.1203.
Mosley, L.M., Hunter, K.A., Ducker, W.A., 2003. Forces between colloid particles in natural waters. Environ. Sci. Technol. 37 (15), 3303-3308. https://doi.org/ 10.1021/es026216d.
Y.W. Naguib and Z. Cui, Nanomedicine: the promise and challenges in cancer chemotherapy, vol. 811. 2014. 10.1007/978-94-017-8739-0_11.
Nakayama, T., Osako, M., 2023. Development of a process-based eco-hydrology model for evaluating the spatio-temporal dynamics of macro- and micro-plastics for the whole of Japan. Ecol. Modell. 476 (November 2022), 110243 https://doi.org/ 10.1016/j.ecolmodel.2022.110243.
Nel, H.A., Dalu, T., Wasserman, R.J., 2018. Sinks and sources: assessing microplastic abundance in river sediment and deposit feeders in an Austral temperate urban river system. Sci. Total Environ. 612, 950-956. https://doi.org/10.1016/j. scitotenv.2017.08.298.
Nguyen, T.H., 2021. Settling behaviour of irregular-shaped polystyrene microplastics. Sci. Technol. Dev. J.1-Eng. Technol. 4 (4), 1219-1228.
Nguyen, T.H., Kieu-Le, T.C., Tang, F.H.M., Maggi, F., 2022. Controlling factors of microplastic fibre settling through a water column. Sci. Total Environ. 838 (December 2021), 156011 https://doi.org/10.1016/j.scitotenv.2022.156011.
Nguyen, T.H., Tang, F.H.M., Maggi, F., 2020. Sinking of microbial-associated microplastics in natural waters. PLoS ONE 15 (2), 1-20. https://doi.org/10.1371/ journal.pone. 0228209.
Niu, L., et al., 2021. New insights into the vertical distribution and microbial degradation of microplastics in urban river sediments. Water Res. 188 https://doi.org/10.1016/j. watres.2020.116449.
Nizzetto, L., Bussi, G., Futter, M.N., Butterfield, D., Whitehead, P.G., 2016. A theoretical assessment of microplastic transport in river catchments and their retention by soils and river sediments. Environ. Sci. Process. Impacts 18 (8), 1050-1059. https://doi. org/10.1039/c6em00206d.
Nouha, K., Kumar, R.S., Balasubramanian, S., Tyagi, R.D., 2018. Critical review of EPS production, synthesis and composition for sludge flocculation. J. Environ. Sci. (China) 66, 225-245. https://doi.org/10.1016/j.jes.2017.05.020.
Ockelford, A., Cundy, A., Ebdon, J.E., 2020. Storm response of fluvial sedimentary microplastics. Sci. Rep. 10 (1), 1-10. https://doi.org/10.1038/s41598-020-58765-2.
Pascolo, S., Petti, M., Bosa, S., 2018. On the wave bottom shear stress in shallow depths: the role of wave period and bed roughness. Water (Switzerland) 10 (10). https://doi. org/10.3390/w10101348.
Peller, J., et al., 2021. Sequestration of microfibers and other microplastics by green algae, Cladophora, in the US Great Lakes. Environ. Pollut. 276, 116695 https://doi. org/10.1016/j.envpol.2021.116695.
Peng, G., Zhu, B., Yang, D., Su, L., Shi, H., Li, D., 2017. Microplastics in sediments of the Changjiang Estuary, China. Environ. Pollut. 225, 283-290. https://doi.org/10.1016/ j.envpol.2016.12.064.
Pohl, F., Eggenhuisen, J.T., Kane, I.A., Clare, M.A., 2020. Transport and burial of microplastics in deep-marine sediments by turbidity currents. Environ. Sci. Technol. 54 (7), 4180-4189. https://doi.org/10.1021/acs.est.9b07527.
Rasta, M., Sattari, M., Taleshi, M.S., Namin, J.I., 2020. Identification and distribution of microplastics in the sediments and surface waters of Anzali Wetland in the Southwest Caspian Sea, Northern Iran. Mar. Pollut. Bull. 160 (August), 111541 https://doi.org/10.1016/j.marpolbul.2020.111541.
Ren, Z., Gui, X., Xu, X., Zhao, L., Qiu, H., Cao, X., 2021. Microplastics in the soilgroundwater environment: aging, migration, and co-transport of contaminants – A critical review. J. Hazard. Mater. 419 https://doi.org/10.1016/j. jhazmat.2021.126455. March.
Righetti, M., Lucarelli, C., 2007. May the Shields theory be extended to cohesive and adhesive benthic sediments? J. Geophys. Res. Ocean. 112 (5), 1-14. https://doi.org/ 10.1029/2006JC003669.
Ryan, P.G., 2015. Does size and buoyancy affect the long-distance transport of floating debris? Environ. Res. Lett. 10 (8) https://doi.org/10.1088/1748-9326/10/8/ 084019.
Schmidt, C., Kumar, R., Yang, S., Büttner, O., 2020. Microplastic particle emission from wastewater treatment plant effluents into river networks in Germany: loads, spatial patterns of concentrations and potential toxicity. Sci. Total Environ. 737, 139544 https://doi.org/10.1016/j.scitotenv.2020.139544.
Shams, M., Alam, I., Chowdhury, I., 2020. Aggregation and stability of nanoscale plastics in aquatic environment. Water Res. 171, 115401 https://doi.org/10.1016/j. watres.2019.115401.
Shang, Q.Q., Fang, H.W., Zhao, H.M., He, G.J., Cui, Z.H., 2014. Biofilm effects on size gradation, drag coefficient and settling velocity of sediment particles. Int. J. Sediment Res. 29 (4), 471-480. https://doi.org/10.1016/S1001-6279(14)60060-3.
Shen, Z., Wang, H., Liang, D., Yan, Y., Zeng, Y., 2022. The fate of microplastics in estuary: a quantitative simulation approach. Water Res. 226 (October), 119281 https://doi.org/10.1016/j.watres.2022.119281.
A. Shields, “Application of similarity principles and turbulence research to bed-load movement,” no. 167, 47, 1936.
Singh, N., Tiwari, E., Khandelwal, N., Darbha, G.K., 2019. Understanding the stability of nanoplastics in aqueous environments: effect of ionic strength, temperature,
dissolved organic matter, clay, and heavy metals. Environ. Sci. Nano 6 (10), 2968-2976. https://doi.org/10.1039/c9en00557a.
Song, Y.K., et al., 2014. Large accumulation of micro-sized synthetic polymer particles in the sea surface microlayer. Environ. Sci. Technol. 48 (16), 9014-9021. https://doi. org/10.1021/es501757s.
Song, Y.K., et al., 2018. Horizontal and vertical distribution of microplastics in Korean coastal waters. Environ. Sci. Technol. 52 (21), 12188-12197. https://doi.org/ 10.1021/acs.est.8b04032.
Stead, J.L., Bond, T., 2023. The impact of riverine particles on the vertical velocities of large microplastics. Sci. Total Environ. 896 (May), 165339 https://doi.org/10.1016/ j.scitotenv.2023.165339.
Stride, B., Abolfathi, S., Bending, G.D., Pearson, J., 2024. Quantifying microplastic dispersion due to density effects. J. Hazard. Mater. 466, 133440 https://doi.org/ 10.1016/j.jhazmat.2024.133440.
Sun, H., Jiao, R., Wang, D., 2020. The difference of aggregation mechanism between microplastics and nanoplastics: role of Brownian motion and structural layer force. Environ. Pollut. 268, 115942 https://doi.org/10.1016/j.envpol.2020.115942.
Tamminga, M., Fischer, E.K., 2020. Microplastics in a deep, dimictic lake of the North German Plain with special regard to vertical distribution patterns. Environ. Pollut. 267, 115507 https://doi.org/10.1016/j.envpol.2020.115507.
Tibbetts, J., Krause, S., Lynch, I., Smith, G.H.S., 2018. Abundance, distribution, and drivers of microplastic contamination in urban river environments. Water (Switzerland) 10 (11). https://doi.org/10.3390/w10111597.
G. Trefalt and M. Borkovec, “Overview of DLVO theory,” 2014.
Vaughan, R., Turner, S.D., Rose, N.L., 2017. Microplastics in the sediments of a UK urban lake. Environ. Pollut. 229, 10-18. https://doi.org/10.1016/j.envpol.2017.05.057.
Vermaire, J.C., Pomeroy, C., Herczegh, S.M., Haggart, O., Murphy, M., 2017. Microplastic abundance and distribution in the open water and sediment of the Ottawa River, Canada, and its tributaries. Facets 2 (1), 301-314.
Wagner, M., Lambert, S., 2018. Freshwater Microplastics. Springer Nature. https://doi. org/10.1007/978-3-319-61615-5.
Waldschlägelatr, K., Schüttrumpf, H., 2019. Effects of particle properties on the settling and rise velocities of microplastics in freshwater under laboratory conditions. Environ. Sci. Technol. 53 (4), 1958-1966. https://doi.org/10.1021/acs.est.8b06794.
Waldschläger, K., Born, M., Cowger, W., Gray, A., Schüttrumpf, H., 2020. Settling and rising velocities of environmentally weathered micro- and macroplastic particles. Environ. Res. 191 (August) https://doi.org/10.1016/j.envres.2020.110192.
Waldschläger, K., Schüttrumpf, H., 2019. Erosion behavior of different microplastic particles in comparison to natural sediments. Environ. Sci. Technol. https://doi.org/ 10.1021/acs.est.9b05394.
Wang, J., et al., 2017. Microplastics in the surface sediments from the Beijiang River littoral zone: composition, abundance, surface textures and interaction with heavy metals. Chemosphere 171, 248-258. https://doi.org/10.1016/j. chemosphere.2016.12.074.
Wang, J., et al., 2021c. Aggregation and stability of sulfate-modified polystyrene nanoplastics in synthetic and natural waters. Environ. Pollut. 268, 114240 https:// doi.org/10.1016/j.envpol.2020.114240.
Wang, X., Bolan, N., Tsang, D.C.W., Sarkar, B., Bradney, L., Li, Y., 2021a. A review of microplastics aggregation in aquatic environment: influence factors, analytical methods, and environmental implications. J. Hazard. Mater. 402 (March 2020), 123496 https://doi.org/10.1016/j.jhazmat.2020.123496.
Wang, Y., Chen, X., Wang, F., Cheng, N., 2023. Influence of typical clay minerals on aggregation and settling of pristine and aged polyethylene microplastics. Environ. Pollut. 316 (P2), 120649 https://doi.org/10.1016/j.envpol.2022.120649.
Wang, Z., Dou, M., Ren, P., Sun, B., Jia, R., Zhou, Y., 2021b. Settling velocity of irregularly shaped microplastics under steady and dynamic flow conditions. Environ. Sci. Pollut. Res. 28 (44), 62116-62132. https://doi.org/10.1007/s11356-021-14654-3.
Willis, K.A., Eriksen, R., Wilcox, C., Hardesty, B.D., 2017. Microplastic distribution at different sediment depths in an urban estuary. Front. Mar. Sci. 4 (DEC), 1-9. https:// doi.org/10.3389/fmars.2017.00419.
Wu, F., Pennings, S.C., Tong, C., Xu, Y., 2020b. Variation in microplastics composition at small spatial and temporal scales in a tidal flat of the Yangtze Estuary, China. Sci. Total Environ. 699, 134252 https://doi.org/10.1016/j.scitotenv.2019.134252.
Wu, J., Jiang, R., Lin, W., Ouyang, G., 2019. Effect of salinity and humic acid on the aggregation and toxicity of polystyrene nanoplastics with different functional groups and charges. Environ. Pollut. 245, 836-843. https://doi.org/10.1016/j. envpol.2018.11.055.
Yang, G., et al., 2023. Settling behaviors of microplastic disks in water. Mar. Pollut. Bull. 188 (February), 114657 https://doi.org/10.1016/j.marpolbul.2023.114657.
Yu, S., et al., 2019. Aggregation kinetics of different surface-modified polystyrene nanoparticles in monovalent and divalent electrolytes. Environ. Pollut. 255, 113302 https://doi.org/10.1016/j.envpol.2019.113302.
Yu, Z., Yao, W., Loewen, M., Li, X., Zhang, W., 2022. Incipient motion of exposed microplastics in an open-channel flow. Environ. Sci. Technol. 56 (20), 14498-14506. https://doi.org/10.1021/acs.est.2c04415.
Yuan, W., Liu, X., Wang, W., Di, M., Wang, J., 2019. Microplastic abundance, distribution and composition in water, sediments, and wild fish from Poyang Lake, China. Ecotoxicol. Environ. Saf. 170 (December 2018), 180-187. https://doi.org/10.1016/ j.ecoenv.2018.11.126.
Zhang, Q., et al., 2021. Distribution and sedimentation of microplastics in Taihu Lake. Sci. Total Environ. 795, 148745 https://doi.org/10.1016/j.scitotenv.2021.148745.
Zhang, Y., et al., 2020b. How climate change and eutrophication interact with microplastic pollution and sediment resuspension in shallow lakes: a review. Sci. Total Environ. 705, 135979 https://doi.org/10.1016/j.scitotenv.2019.135979.
Zhao, J., Shan, T., 2013. Coupled CFD-DEM simulation of fluid-particle interaction in geomechanics. Powder Technol. 239, 248-258. https://doi.org/10.1016/j. powtec.2013.02.003.
Zheng, Y., et al., 2020. Vertical distribution of microplastics in bay sediment reflecting effects of sedimentation dynamics and anthropogenic activities. Mar. Pollut. Bull. 152 (December 2019), 110885 https://doi.org/10.1016/j.marpolbul.2020.110885.
Zhu, H.P., Zhou, Z.Y., Yang, R.Y., Yu, A.B., 2007. Discrete particle simulation of particulate systems: theoretical developments. Chem. Eng. Sci. 62 (13), 3378-3396. https://doi.org/10.1016/j.ces.2006.12.089.
Zhu, X.-T., Yi, J., Qiang, L.-Y., Cheng, J.-P., 2018. Distribution and settlement of microplastics in the surface sediment of Yangtze Estuary. Huan jing ke xue= Huanjing kexue 39 (5), 2067-2074.
- Corresponding author.
E-mail addresses: Soroush.Abolfathi@Warwick.ac.uk, s.abolfathi@warwick.ac.uk (S. Abolfathi).