بانوراما مناعية شاملة للالتهاب الرئوي البكتيري تكشفها خريطة تعبير الجينات على مستوى الخلية الواحدة على نطاق واسع A pan-immune panorama of bacterial pneumonia revealed by a large-scale single-cell transcriptome atlas
الالتهاب الرئوي البكتيري هو عبء كبير على الصحة العامة، حيث يساهم في معدلات مرضية ووفاة مرتفعة وتكاليف الرعاية الصحية. الاستراتيجيات العلاجية الحالية بخلاف المضادات الحيوية والعلاجات المساعدة محدودة، مما يبرز الحاجة إلى فهم أعمق لآلية تطور المرض. هنا، استخدمنا تسلسل RNA على مستوى الخلية الواحدة لـ 444,146 خلية من سائل غسل القصبات الهوائية (BALFs) من مجموعة كبيرة تضم 74 فردًا، بما في ذلك 58 مريضًا يعانون من حالات خفيفة. ) وشديد ( ) الأمراض بالإضافة إلى 16 متبرعًا صحيًا. تم تطبيق اختبارات المناعية المرتبطة بالإنزيمات والاختبارات النسيجية للتحقق من صحة هذه المجموعة. لوحظت تباين الاستجابات المناعية في الالتهاب الرئوي البكتيري، مع ملفات تعريف خلايا مناعية مميزة مرتبطة بشدة المرض. تم تمييز الالتهاب الرئوي البكتيري الشديد بعاصفة سيتوكينية التهابية ناتجة عن زيادة تنظيم S100A8/A9 وCXCL8 على مستوى النظام، ويرجع ذلك أساسًا إلى مجموعات معينة من البلعميات والخلايا المتعادلة. في المقابل، يظهر الالتهاب الرئوي البكتيري الخفيف استجابة مناعية خلوية فعالة تتميز بتوسع خلايا T المساعدة الجريبية وخلايا T المساعدة 2، مما يسهل تنشيط خلايا B وإنتاج الأجسام المضادة. على الرغم من أن كلا مجموعتي المرض تظهران إرهاق خلايا T، إلا أن الحالات الخفيفة حافظت على نشاط سيتوتوكسي قوي.وظيفة الخلايا، مما قد يعكس آلية تعويضية. ساهمت استجابات العدلات والبلاعم غير المنظمة بشكل كبير في مسببات الأمراض للمرض الشديد. تعزز العدلات غير الناضجة الالتهاب المفرط وتثبط تنشيط خلايا T، بينما مجموعة فرعية محددة من البلاعم (Macro_03_M1) تظهر ميزات مشابهة لخلايا مثبطة مشتقة من النخاع (M-MDSCs) تثبط خلايا T وتعزز الالتهاب. معًا، تسلط هذه النتائج الضوء على أهداف علاجية محتملة لتعديل الاستجابات المناعية وتحسين النتائج السريرية في الالتهاب الرئوي البكتيري.
التهاب الرئة البكتيري هو قضية هامة في الصحة العامة ويساهم بشكل كبير في المراضة والوفيات وتكاليف الرعاية الصحية.أفادت دراسة أجريت في الصين عام 2021 بمعدل وفيات موحد حسب العمر قدرهوفيات لكل 100,000 بسبب الالتهاب الرئوي البكتيري، مما يجعله السبب الخامس للوفاة.معدلات الوفيات أعلى حتى في أفريقيا جنوب الصحراء الكبرى وجنوب آسيا وجنوب شرق آسيا.لقد زادت حالات الالتهاب الرئوي البكتيري الشديد بسبب عوامل مثل سوء استخدام المضادات الحيوية والمقاومة، وشيخوخة السكان، والاستخدام الواسع للأدوية المثبطة للمناعة، وارتفاع انتشار الأمراض المزمنة.حوالي خُمس المرضى الذين يتم إدخالهم المستشفى بسبب الالتهاب الرئوي يحتاجون إلى دخول وحدة العناية المركزة، وثلث هؤلاء المرضى يتطلبون التهوية الميكانيكية.معدلات الوفيات مرتفعة بشكل خاص بين المرضى الذين تم إدخالهم إلى المستشفى، خاصة بالنسبة لأولئك الذين يعانون من التهاب رئوي شديد، حيث تصل معدلات الوفيات إلىحالياً، تفتقر العلاجات الفعالة بخلاف المضادات الحيوية والعلاج المساعد. لذلك، فإن فهم آلية حدوث الالتهاب الرئوي البكتيري، خاصة في الحالات الشديدة، أمر بالغ الأهمية لتطوير استراتيجيات علاجية جديدة.
عند مواجهة الكائنات الدقيقة المسببة للأمراض، تقوم خلايا الظهارة الهوائية والبلعميات بإفراز كميات كبيرة من السيتوكينات الكيميائية والالتهابية، مثل عامل نخر الورم (TNF-الإنترلوكين (IL)-1، IL-6، وعامل كيميائي جاذب CXC 8 (CXCL8، المعروف أيضًا باسم IL-8).تقوم هذه العوامل بتجنيد وتفعيل الخلايا البلعومية، بما في ذلك العدلات والوحيدات، بالإضافة إلى نظام المكملات، بهدف القضاء على الممرضات. بينما تعتبر الاستجابة المناعية المعتدلة ضرورية لإزالة الممرضات وبقاء المضيف، يمكن أن تؤدي الاستجابة المناعية المفرطة إلى حدوث “عاصفة سيتوكينية”، مما يؤدي إلى التهاب رئوي شديد وزيادة في معدل الوفيات. سريرياً، يتميز التهاب الرئة الشديد بالإفراج المفرط عن الكيموكينات والسيتوكينات، بالإضافة إلى الإفراط في التعبير عن جزيئات الالتصاق.يمكن أن تتسبب هذه العملية الالتهابية في إتلاف سلامة البطانة، مما يسمح بالبلعمة
تاريخ الاستلام: 24 يونيو 2024 تاريخ المراجعة: 4 ديسمبر 2024 تاريخ القبول: 6 ديسمبر 2024 نُشر على الإنترنت: 06 يناير 2025 تدخل الخلايا والمصل وكريات الدم الحمراء إلى تجويف الحويصلات الهوائية، مما يؤدي إلى الوذمة الرئوية. تسهل هذه الوذمة أيضًا تسرب العدلات إلى تجويف الحويصلات الهوائية، مما يعزز إفراز السيتوكينات، وأنواع الأكسجين التفاعلية، والبروتيازات.ومع ذلك، لا يزال الفهم المنهجي لخلل تنظيم المناعة خلال الالتهاب الرئوي البكتيري بعيد المنال. تفتقر الدراسات الشاملة والعميقة حول الكمية، والنمط الظاهري، وخصائص مختلف خلايا المناعة، فضلاً عن مستويات العوامل المؤيدة والمعارضة للالتهاب في مراحل مختلفة من تقدم المرض. كما تتطلب الآليات المتعلقة بحماية المرض أو المسببة له، وخاصة تلك التي تؤدي إلى مرض شديد، مزيدًا من التحقيق.
فهم التغيرات في نظام المناعة لدى المضيف خلال الالتهاب الرئوي البكتيري وتوضيح آليات اختلال تنظيم المناعة أمران حاسمان لتوجيه العلاج السريري، وتقليل حدوث ووفيات الالتهاب الرئوي الشديد، وتحسين نتائج المرضى. لقد ظهرت تقنية تسلسل RNA على مستوى الخلية الواحدة (scRNAseq) كأداة قوية لتفكيك تعقيد الاستجابات المناعية، مما يوفر رؤى حاسمة حول تفاعلات المضيف والجراثيم على مستوى الخلية.بالإضافة إلى ذلك، يمكن أن يوفر تحليل سائل غسل القصبات الهوائية والحويصلات الهوائية (BALF) من البشر رؤى حول الاستجابة المناعية المنسقة للعدوى البكتيرية مثل المتفطرة السلية.لذا، يمكن أن يكشف تحليل تسلسل RNA أحادي الخلية في سوائل الشعب الهوائية (BALFs) عن التركيب الخلوي، وأنماط النسخ، والاستجابات المناعية الديناميكية لدى المرضى الذين يعانون من الالتهاب الرئوي البكتيري، مما يسلط الضوء على المسارات الالتهابية الحيوية والأهداف العلاجية المحتملة.
هنا، استخدمنا تسلسل RNA أحادي الخلية للحصول على تصور غير متحيز للاستجابة المناعية الشاملة في سائل غسيل الشعب الهوائية من مرضى الالتهاب الرئوي البكتيري، بدءًا من الأعراض الخفيفة إلى الشديدة، بالإضافة إلى مجموعة من الأصحاء (الشكل 1أ). تصف دراستنا المشهد النسخي عالي الدقة لسائل غسيل الشعب الهوائية خلال تقدم الالتهاب الرئوي البكتيري. لوحظت ملفات خلايا مناعية مميزة في سوائل غسيل الشعب الهوائية للأفراد المصابين بالالتهاب الرئوي البكتيري. كانت الحالات الشديدة تتميز بزيادة الاستجابات الالتهابية المدفوعة بواسطة العدلات والبلاعم، بينما أظهرت الحالات الخفيفة توسعًا في خلايا المساعد T الجريبية (Tfh) وخلايا المساعد T 2 (Th2)، التي ساعدت في تنشيط خلايا B. لقد ألقى تحديد تفاعلات السيتوكينات والروابط المستقبلية في هذه الملفات الضوء على أهداف علاجية محتملة. هذه النتائج مهمة لفهم الآليات المناعية الكامنة وراء درجات الشدة المختلفة للالتهاب الرئوي البكتيري ويمكن أن توجه تطوير علاجات مستهدفة جديدة لتعديل الاستجابات المناعية وتحسين النتائج السريرية.
النتائج
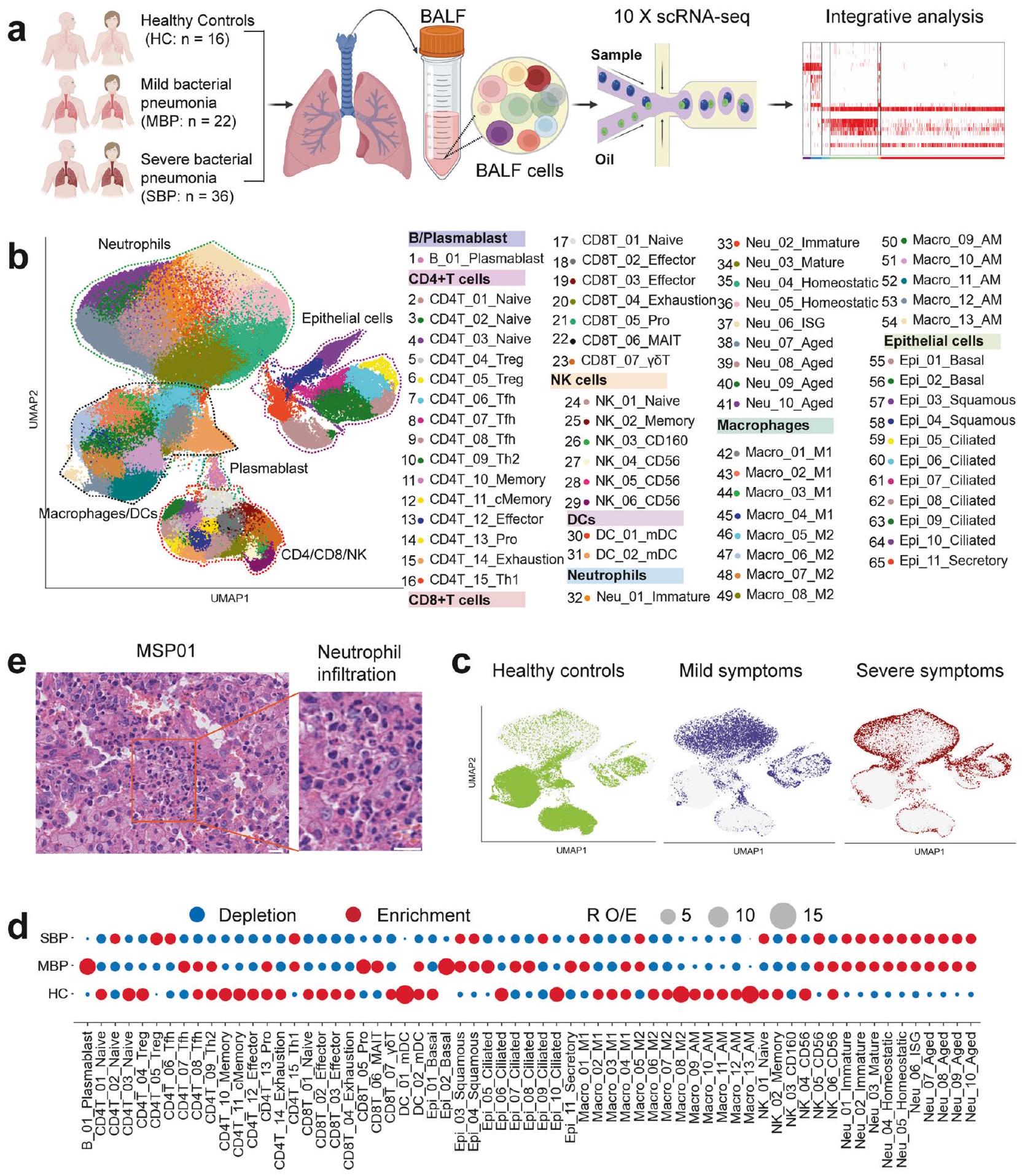
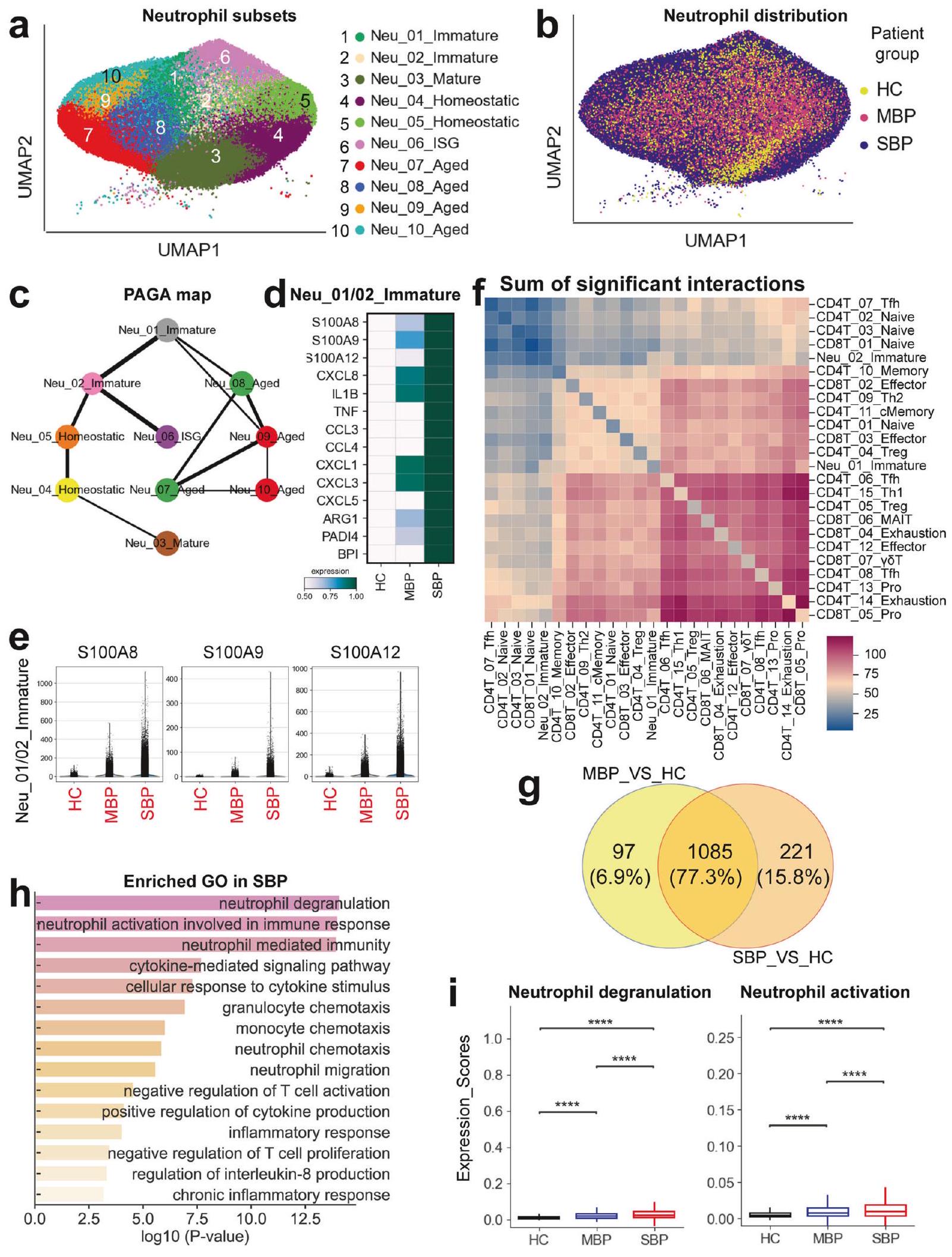
نظرة عامة على تحليل خلايا المناعة في السائل القصبي الهوائي لدى الأفراد المصابين بالالتهاب الرئوي البكتيري لتوصيف المشهد المناعي في الالتهاب الرئوي البكتيري، تم إنشاء ملفات النسخ الجيني على مستوى الخلية الواحدة من عينات سائل غسل القصبات الهوائية (BALF) التي تم الحصول عليها من 58 مريضًا و16 ضابط صحة (HCs) باستخدام تقنية تسلسل RNA أحادي الخلية المعتمدة على القطرات (منصة 10x Genomics) (الشكل 1a). تم تصنيف المرضى الـ 58 الذين تم تشخيصهم بالالتهاب الرئوي البكتيري إلى مجموعتين سريريتين: الالتهاب الرئوي البكتيري الخفيف (MBP، ) والالتهاب الرئوي البكتيري الشديد (SBP، ). إن الغالبية العظمى من العوامل المسببة هي مسببات التهاب الرئة البكتيري الشائعة، مثل Acinetobacter baumannii وPseudomonas aeruginosa وStaphylococcus aureus، من بين آخرين. تم تقديم معلومات سريرية مفصلة ونتائج مختبرية لـ 58 مريضًا تم تسجيلهم في الجدول التكميلي 1 والشكل التكميلي 1. حصلنا على 444,146 خلية مفردة عالية الجودة، بمتوسط 3500 معرف جزيئي فريد (UMIs) يمثل 1164 جينًا (الشكل التكميلي.من هذه الخلايا، 60,084 خليةكانت من حالة HC، 94,590 خليةكانت من حالة MBP، و 289,472 خليةكانت من حالة SBP (الشكل التكميلي S2a-d). بعد التعديل لعمق القراءة وقراءة الميتوكوندريا تم دمج جميع الخلايا عالية الجودة في مجموعة بيانات غير مجمعة وقابلة للمقارنة لتحليل المكونات الرئيسية اللاحق (انظر الطرق).
باستخدام التجميع القائم على الرسوم البيانية لتقنية الاقتراب والتصوير المتجانس (UMAP)، حددنا ملفات التعبير الجيني لثمانية أنواع رئيسية من الخلايا: البلازما، البلعميات، العدلات،تم تمييز خلايا T، وخلايا القاتل الطبيعي (NK)، والخلايا الشجرية (DCs)، والخلايا الظهارية، بناءً على تعبير علامات الجينات الكانونية (الشكل 1ب والشكل التكميلي 2هـ). في فضاء UMAP، تم تمييز الخلايا اللمفاوية (T وNK) بوضوح عن البلعميات والعدلات، في حين قدمت الخلايا الظهارية نمطًا ترنسكريبتوميًا مميزًا عن تلك الخاصة بالخلايا المناعية (الشكل 1ب والشكل التكميلي 2و). عندما تم تلوينها حسب مستوى المرض، كانت تمثيلات UMAP مفصولة حسب الشدة بعد العدوى البكتيرية، خاصة في المرضى الشديدين مقارنةً بالضوابط الصحية، مما يشير إلى تغييرات ترنسكريبتومية ناتجة عن العدوى البكتيرية (الشكل 1ج والشكل التكميلي 3أ). قمنا أولاً بتحديد 45 نوعًا فرعيًا من الخلايا ضمن 8 مجموعات خلوية رئيسية (الشكل التكميلي 2 و). كشفت عملية التصنيف الفرعي الإضافية عن إجمالي 65 نوعًا فرعيًا تشمل أنواعًا متنوعة من خلايا الجهاز التنفسي (الشكل 1ب والشكل التكميلي 4)، والتي تم استخدامها للتحليلات اللاحقة. وبالتالي، قمنا بتعريف تركيبات السكان الفرعيين للخلايا في سوائل غسيل الشعب الهوائية، مما يوفر مجموعة بيانات غنية بالمعلومات للتعليق الدقيق والتحليل الشامل لهذه الأنواع الخلوية عند دقة مختلفة.
عرض كل تجمع خلوي أصول عينات مختلفة وغنى في شدة الأمراض (الشكل 1c والشكل التكميلي 3). لتحليل الملفات المناعية الفريدة لمجموعات الأمراض المختلفة، قمنا بالتحقيق في تركيبة الخلايا المناعية في كل فرد (الشكل 1d والشكل التكميلي 3). ومن الجدير بالذكر أن نسب العدلات في سوائل غسيل الشعب الهوائية كانت مرتفعة بشكل ملحوظ، خاصة في المرضى الذين يعانون من التهاب رئوي بكتيري حاد (الشكل 1d، e والشكل التكميلي 3e).على عكس ارتفاع عدد العدلات في سوائل الشطف الرئوي، انخفضت معظم أنواع الخلايا المناعية الرئيسية الأخرى في المرضى الذين يعانون من الالتهاب الرئوي البكتيري (الشكل 1d والشكل التكميلي 3). من المهم أن تفضيل كل نوع فرعي من الخلايا في مجموعات المرضى المختلفة تم توضيحه أيضًا وفقًا لـ : نسبة الترددات الخلوية المرصودة إلى المتوقعة (تستخدم للتخفيف من تأثير التباين الفني على التركيب الخلوي المرتبط بالمرض)، مع انتشار خلايا B (B_01_Plasmablast)، CD4ت (CD4T_13_Pro) وتتميز خلايا T (CD8T_05_Pro) بزيادة في المرضى الخفيفين مقارنة بالحالات الشديدة والشهود الأصحاء (الشكل 1d). أظهرت خلايا البلازما B (B_01_Plasmablast) في سوائل غسيل القصبات الهوائية تعبيرًا عاليًا عن الجينات التي تشفر المناطق الثابتة من الغلوبولين المناعي G1 (lgG1)، IgG3، IgG4، أو IgGM (الشكل التوضيحي 3f)، مما يشير إلى وظيفتها في إفراز الأجسام المضادة المحددة لمستضدات. تشير هذه النتائج إلى أن تنشيط خلايا B الفعال وإنتاج الأجسام المضادة قد يلعبان دورًا مهمًا في الالتهاب الرئوي البكتيري. بشكل عام، كشفت البيانات أن كل حالة مرضية في المرضى الذين يعانون من الالتهاب الرئوي البكتيري قد تظهر بصمة مناعية فريدة.
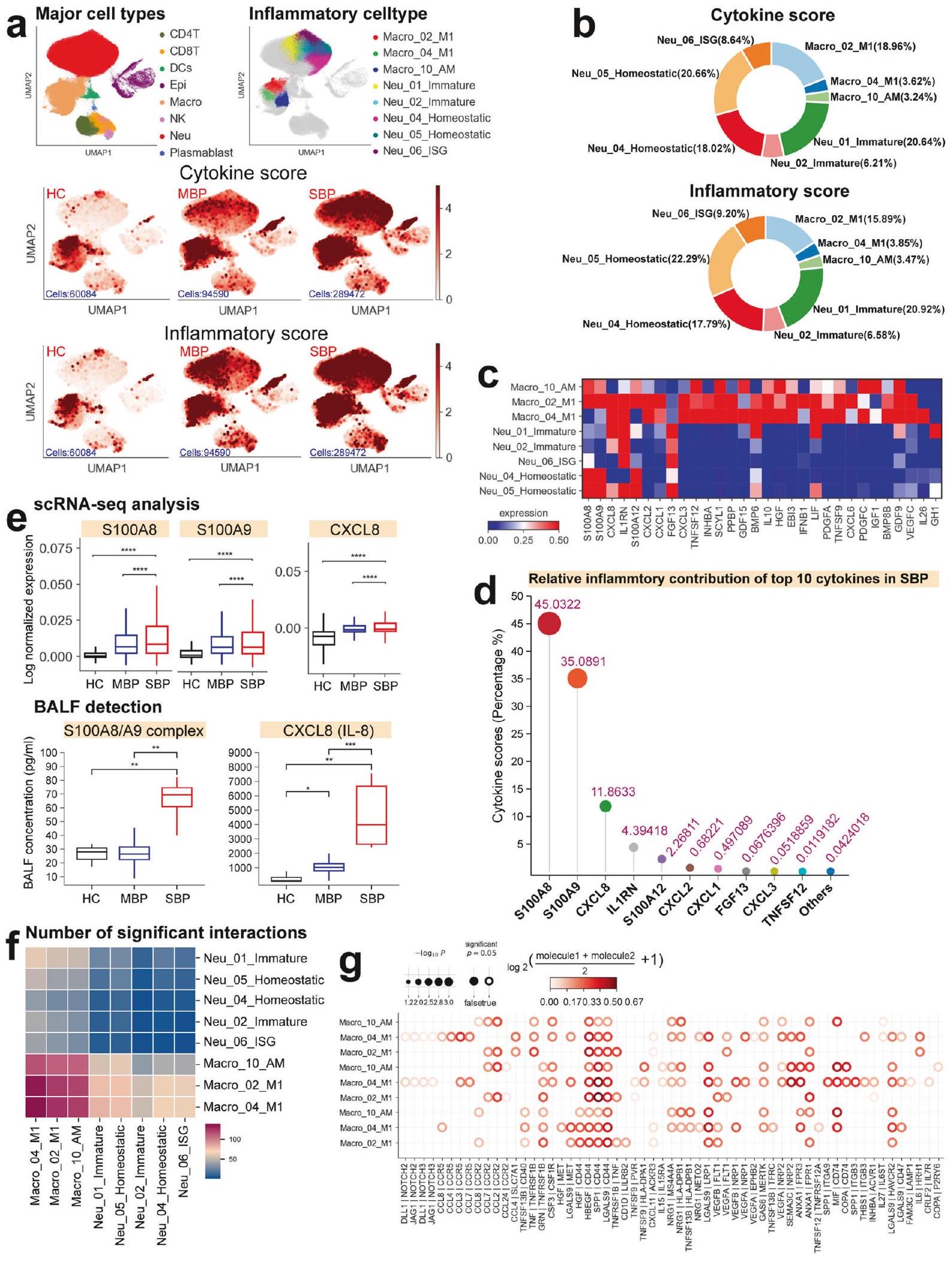
البلاعم والعدلات هما المحركان الرئيسيان للاستجابة الالتهابية في الرئة لدى المرضى الذين يعانون من الالتهاب الرئوي البكتيري. استجابة المضيف الالتهابية ضد الكائنات الدقيقة المسببة هي أمر حاسم لحل الالتهاب الرئوي البكتيري.لتحقيق في الأصول المحتملة لإنتاج السيتوكينات، قمنا بإنشاء درجة سيتوكين ودرجة التهابية لكل خلية من خلايا المناعة من خلال تحليل تعبير جينات السيتوكينات وجينات الاستجابة الالتهابية المعروفة، على التوالي (الجدول التكميلي 2).تم تعريف درجات السيتوكين والالتهاب باستخدام دالة sc.tl.score_genes في Scanpy.تم تحديد تجمعات الخلايا المفرطة الالتهاب من خلال إجراء مقارنات ثنائية لهذه الدرجات عبر جميع الأنماط الفرعية باستخدام اختبارات كروسكال-واليس مع تصحيح بونفيروني. تم استخدام هذين الدرجتين كمؤشرات لتقييم التأثير الالتهابي المحتمل لمجموعات الخلايا المناعية المختلفة. في المرضى الذين يعانون من الالتهاب الرئوي البكتيري، لاحظنا زيادة ملحوظة في تنظيم الجينات المتعلقة بالسيتوكينات والالتهابات.
الشكل 1 نظرة عامة على تصميم الدراسة ونتائج دراسة النسخ الجيني للخلية الواحدة في سائل الشطف الرئوي. أ مخطط يوضح التصميم العام للدراسة، الذي شمل 74 فردًا، بما في ذلك 58 مريضًا (22 مريضًا يعانون من مرض خفيف و36 مريضًا يعانون من مرض شديد) و16 متبرعًا صحيًا. ب نتيجة التجميع (الصف الأيسر) لـ 65 نوعًا فرعيًا من الخلايا (الصف الأيمن) من 74 عينة. كل نقطة تمثل خلية واحدة، ملونة وفقًا لنوع الخلية. ج إسقاط UMAP للمتبرعين الأصحاء، والمرضى الخفيفين والشديدين. د تفضيل المرض لمجموعات الخلايا الرئيسية كما تم تقديره باستخدامصور تمثيلية لصبغة الهيماتوكسيلين والإيوزين لشرائح الرئة من مرضى يعانون من التهاب رئوي بكتيري حاد (MSP01). مقياس الأشرطة، (الصورة الرئيسية؛ اليسار) و (صور مكبرة، يمين)
تشير إلى استجابة التهابية (الشكل 2أ والشكل التوضيحي 5أ). أظهر المرضى الذين يعانون من التهاب رئوي بكتيري شديد مستويات أعلى بشكل ملحوظ من السيتوكينات ودرجات الالتهاب مقارنة بالمرضى الذين يعانون من التهاب رئوي بكتيري خفيف، مما يدل على وجود عاصفة سيتوكينية التهابية في رئتي المرضى المتأثرين بشدة (الشكل التوضيحي 5أ).
أظهرت أربعة عشر نوعًا فرعيًا من الخلايا، بما في ذلك ستة أنواع فرعية من العدلات، وسبعة أنواع فرعية من البلعميات، ونوع فرعي واحد من الخلايا الظهارية، ارتفاعًا ملحوظًا في مستويات السيتوكينات ودرجات الالتهاب استنادًا إلى بيانات تسلسل RNA أحادي الخلية لدينا لسوائل غسيل القصبات الهوائية (الشكل التوضيحي التكميلي 5b)، مما يشير إلى أدوارها المحتملة كمصادر للعاصفة الالتهابية. حددت التحليلات الإضافية ثمانية التهابات تجمعات الخلايا، بما في ذلك ثلاث تجمعات من البلعميات (Macro_02_M1 وMacro_04_M1 وMacro_10_AM) وخمس تجمعات من العدلات (Neu_01_Immature وNeu_02_Immature وNeu_04_Homeostatic وNeu_05_Homeostatic وNeu_06_ISG)، والتي أظهرت درجات أعلى بشكل ملحوظ من السيتوكينات والالتهابات في المرضى الشديدين. مقارنةً بالمرضى الخفيفين والمتبرعين الأصحاء (الشكل 2 أ والشكل التكميلي S6). تشير هذه النتائج إلى أن هذه الكتل الخلوية هي على الأرجح المساهمين الرئيسيين في عاصفة السيتوكينات التي لوحظت في المرضى الشديدين. قمنا بمزيد من التحقيق في الوفرة النسبية لهذه الثمانية مجموعات خلوية في الحالات الشديدة.
الشكل 2 الخلايا النخاعية هي المساهم الرئيسي في إنتاج السيتوكينات المؤيدة للالتهابات في المرضى الشديدين. أ إسقاطات UMAP لسوائل غسيل القصبات الهوائية. ملونة بناءً على ثمانية أنواع رئيسية من الخلايا (أعلى اليسار)، وثمانية أنواع فرعية من الخلايا المفرطة الالتهاب (أعلى اليمين)، ودرجة السيتوكين (الوسط)، ودرجة الالتهاب (الأسفل). ب مخططات دائرية توضح المساهمة النسبية لكل نوع فرعي من الخلايا الالتهابية في درجات السيتوكين والالتهاب في المرضى الشديدين. ج خريطة حرارية توضح تعبير السيتوكينات داخل كل نوع فرعي من الخلايا المفرطة الالتهاب المحددة. د مخطط عصا يظهر المساهمة النسبية لأعلى عشرة سيتوكينات في المرضى الذين يعانون من مرض شديد. هـ مخططات صندوقية لتعبير السيتوكين بناءً على تسلسل RNA أحادي الخلية وتحليل البلازما للضوابط الصحية، والمرضى الخفيفين، والمرضى الشديدين. تم تقييم الأهمية باستخدام اختبار كروسكال-واليس مع تصحيح بونفيروني. ). رسومات خريطة الحرارة لمجموع التفاعل المهم بين ثمانية أنواع فرعية من الخلايا المفرطة الالتهاب.رسم نقطي للتفاعلات بين البلعميات الالتهابية في المرضى الشديدين.تُشير القيم إلى أحجام الدوائر، كما هو موضح في المقياس على اليمين المرضى ولاحظوا زيادة ملحوظة، مما ي implicate لهم في الاستجابة الالتهابية المتزايدة التي لوحظت في الحالات الشديدة (الشكل التوضيحي 5c).
أظهرت التحليلات اللاحقة أن أربعة مجموعات التهابية، بما في ذلك Macro_02_M1 وNeu_01_Immature وNeu_04_Homeostatic وNeu_05_Homeostatic، ساهمت بأكثر منمن درجات الالتهاب والسيتوكينات (الشكل 2ب)، مما يشير إلى أن هذه الخلايا كانت المحركات الرئيسية للالتهاب في المرضى الشديدين. تتماشى هذه النتيجة مع الدراسات السابقة حول أمراض معدية أخرى مثل COVID-19 حيث تم تحديد البلعميات والعدلات أيضًا كمحركات رئيسية لعواصف السيتوكينات.تحليل التوقيعات الالتهابية ضمن المجموعات الفرعية المحددة كشف عن ملفات تعبير جيني مميزة للسيتوكينات المؤيدة للالتهاب لكل مجموعة من خلايا الالتهاب (الشكل 2c)، بما في ذلك CXCL1/2/3/8، S100A8/9/12، IL1RN، PPBP، وغيرها. علاوة على ذلك، لاحظنا زيادة في تعبير السيتوكينات الالتهابية الرئيسية، بما في ذلك CXCL1/2/3 وS100A8/9/12، في المرضى الذين يعانون من أعراض شديدة (الشكل التكميلية 7a). تشير هذه النتائج إلى أن آليات متنوعة قد تدفع العاصفة الالتهابية في الالتهاب الرئوي الشديد. إن زيادة تعبير السيتوكينات المؤيدة للالتهاب الخاصة بنوع الخلية في أربع مجموعات رئيسية من خلايا الالتهاب (الشكل 2c والشكل التكميلية 7b) تعزز من دورها الأساسي في دفع العاصفة الالتهابية التي تُرى في أولئك الذين يعانون من مرض شديد.
عشرة سيتوكينات مؤيدة للالتهابات (CXCL1/2/3/8، S100A8/9/12، IL1RN، FGF13 وTNFSF12)، التي يتم التعبير عنها بشكل أساسي في Macro_02_M1 وNeu_01_Immature وNeu_04_Homeostatic وNeu_05_homeostatic (الشكل التوضيحي 7c)، هي محركات حاسمة للعاصفة الالتهابية، حيث تمثل أكثر مندرجات السيتوكين في المرضى الشديدين (الشكل 2د). كانت S100A8/A9 و CXCL8 من أبرز المساهمين، حيث تشكلان حواليمن درجات السيتوكينات (الشكل التكميلي 2d). أظهر المرضى الذين يعانون من مرض شديد تعبيرات مرتفعة لجينات S100A8/A9 وCXCL8 (الشكل 2e)، مما يدعم فرضيتنا بشكل أكبر. أظهرت تحليل سائل غسيل الشعب الهوائية (BALF) ضمن مجموعتنا تركيزات أعلى بشكل ملحوظ من مركب S100A8/A9 وCXCL8 (المعروف أيضًا باسم IL-8) في المرضى الشديدين، مما يؤكد نتائج تحليل تسلسل RNA أحادي الخلية لدينا (الشكل 2e). وهذا يبرز أهمية مجموعات الخلايا المفرطة الالتهاب والجزيئات (CXCL8 وS100A8/A9) كأهداف علاجية محتملة للتخفيف من المناعة المرضية التي لوحظت في مرضى الالتهاب الرئوي المصابين بشدة.
قد ينتج العاصفة السيتوكينية التي تُرى في المرضى الشديدين عن تواصل خلوي معقد بين أنواع الخلايا المفرطة الالتهاب، يتم توسيعه من خلال إفراز مجموعة متنوعة من السيتوكينات.للمزيد من التحقيق في هذه الظاهرة، قمنا بفحص أنماط تفاعل الليغاند-المستقبل بين ثمانية مجموعات فرعية متميزة من الخلايا المفرطة الالتهاب في هؤلاء المرضى (الشكل 2f والشكل التكميلية 7e، f). كشفت تحليلاتنا عن عدة تفاعلات ملحوظة بين الليغاند والمستقبل ضمن هذه الأنماط الفرعية، مع تسليط الضوء بشكل خاص على التفاعلات المتزايدة بين البلعميات الالتهابية (الشكل 2f والشكل التكميلية 7e). كانت هذه البلعميات الالتهابية تعبر عن عدة مستقبلات، بما في ذلك SLC7A1 وCCR5 وCCR2 وIL15RA وHAVCR2، مما يشير إلى استجابتها لمجموعة متنوعة من السيتوكينات التي تفرزها خلايا أخرى (الشكل 2g والشكل التكميلية 7f). ومن الجدير بالذكر أن تحليلاتنا أكدت أن التفاعلات بين البلعميات الالتهابية ومجموعات الخلايا المفرطة الالتهاب الأخرى كانت في الأساس تعتمد على الكيموكينات ومستقبلاتها المقابلة (الشكل 2g والشكل التوضيحي 7f). توفر هذه النتائج رؤى جزيئية حول الآليات المحتملة التي تكمن وراء الشبكة المعقدة من التفاعلات بين مجموعات الخلايا المفرطة الالتهاب في الالتهاب الرئوي الشديد.
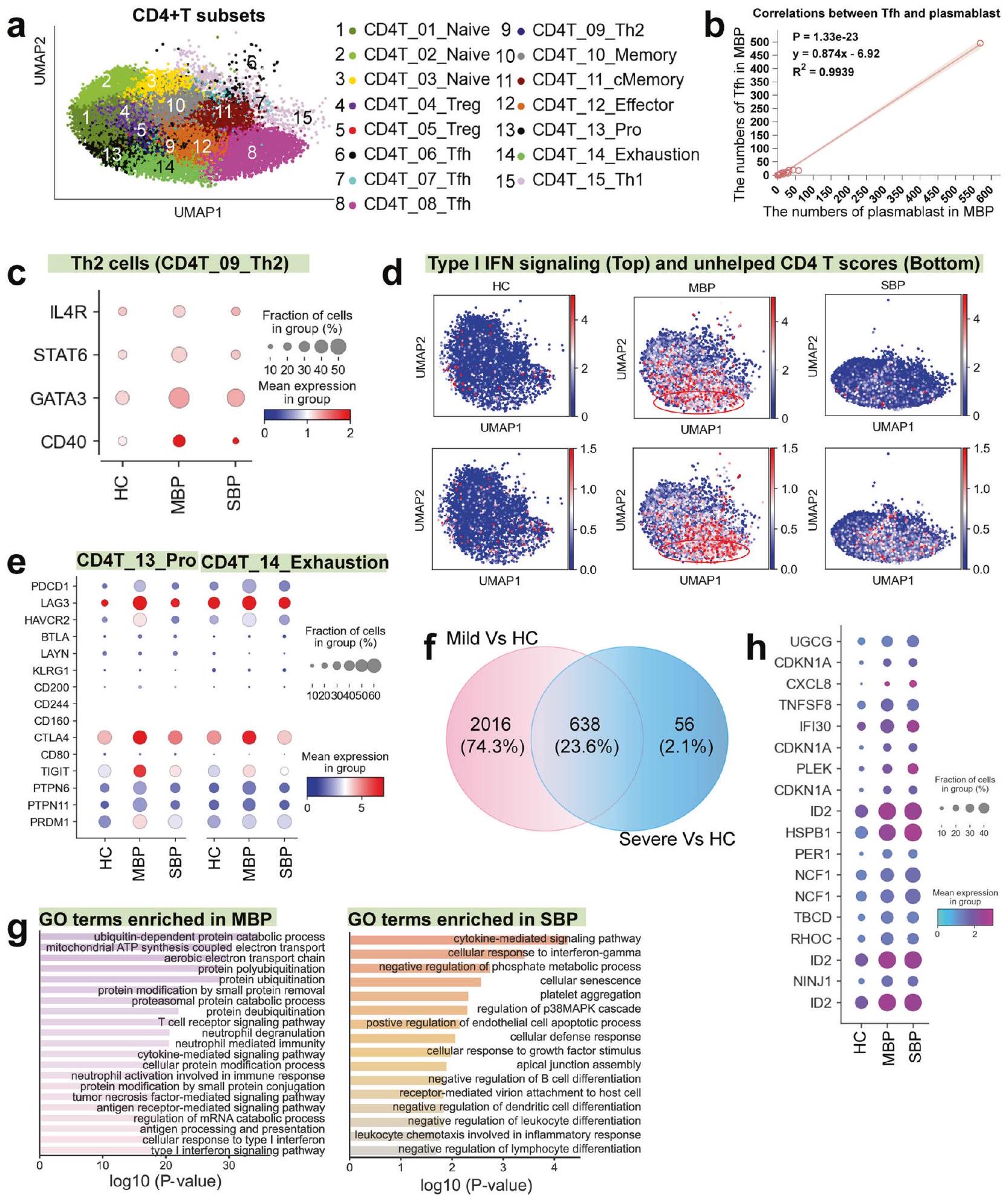
تمت ملاحظة توسع خلايا Tfh و Th2 في المرضى الخفيفين ولكن ليس في المرضى الشديدين إجماليتم تحديد أنواع فرعية من خلايا T، بما في ذلك ثلاثة خلايا غير ناضجة (CD4T_01_Naive، CD4T_02_Naive، وCD4T_03_Naive)، وخليتين تنظيميتين (CD4T_04_Treg وCD4T_05_Treg)، وثلاث خلايا مساعدة جريبية (CD4T_06_Tfh، CD4T_07_Tfh، وCD4T_08_Tfh)، وخلايا Th2 (CD4T_09_Th2)، وخلايا ذاكرة (CD4T_10_Memory)، وذاكرة مركزية (CD4T_11_cMemory)، وخلايا متكاثرة (CD4T_13_Pro)، وخلايا منهكة (CD4T_14_Exhaustion) وخلايا Th1 (CD4T_15_Th1) (الشكل 3أ). أظهر تحليل تفضيل المرض أن خلايا Tfh وTh2 وخلايا CD4 المتكاثرةتم إثراء خلايا T في المرضى الخفيفين، بينما توسع مجموعة فرعية من Treg (CD4_05_Treg) بشكل رئيسي في المرضى الشديدين (الشكل 1d)، مما يشير إلى أن استجابة Tfh وTh2 قد تكون متورطة في السيطرة على العدوى البكتيرية.
أظهرت خلايا CD4T_06/07/08_Tfh ملفًا تعبيريًا يتماشى مع خلايا المساعد التائية الجريبية (Tfh) (على سبيل المثال، التعبير العالي عن المحفز المساعد القابل للتحفيز؛ ICOS) (الشكل التوضيحي 4)، مما يشير إلى وظيفتها المحتملة في تسهيل استجابة مناعية واقية لخلايا B. تعتبر خلايا Tfh ضرورية لتوليد استجابات الأجسام المضادة المثلى وضمان إنتاج أجسام مضادة محايدة عالية الجودة أثناء عدوى مسببات الأمراض.يتماشى مع ذلك، لاحظنا أيضًا توسعًا ملحوظًا في البلازما بلاست لدى الأفراد الذين يعانون من أعراض خفيفة (الشكل 1d)، جنبًا إلى جنب مع ارتباط إيجابي قوي بين زيادة خلايا Tfh وتوسع البلازما بلاست في هؤلاء الأفراد (الشكل 3b). ومع ذلك، يتناقص هذا الارتباط في حالات زيادة الشدة (الشكل التكميلية 8a). الوظيفة الأساسية لخلايا Th2 هي تسهيل تنشيط خلايا B، والسايتوكينات التي تفرزها تعززتكاثر الخلايا، التمايز، وإنتاج الأجسام المضادة.من المثير للاهتمام أنه بين المرضى الخفيفين، لوحظت علاقة قوية بين خلايا Th2 الموسعة وزيادة البلازما بلاست (الشكل التوضيحي 8b). أظهرت الجينات الرئيسية وعوامل النسخ المرتبطة بتوجه Th2 وتنشيط خلايا B مستويات تعبير أعلى في المرضى الخفيفين مقارنة بالمرضى الشديدين والأصحاء (الشكل 3c). مجتمعة، تشير هذه النتائج إلى استجابة منسقة بين خلايا T وخلايا B في المرضى الخفيفين، مما يؤدي إلى مناعة خلطية فعالة ضد البكتيريا، والتي يبدو أنها تصبح غير متصلة في المرضى الشديدين.
يمكن أن تؤدي عدوى الممرضات إلى استنفاد اللمفاويات.لذلك، قمنا بتوصيف تعبير الجينات التي تشفر علامات التعب النموذجية فيخلايا T. من بينأظهرت مجموعات T، نوعان فرعيان (CD4T_13_Pro و CD4T_14_Exhaustion) درجات إرهاق أعلى بشكل ملحوظ (الشكل التوضيحي التكميلي 8c)، مع أعلى درجة إرهاق لوحظت في المرضى الخفيفين (الشكل التوضيحي التكميلي 8d). عدم وجود جوهرياستنفاد خلايا T في المرضى الشديدين (الشكل التوضيحي 8d) يشير إلى أنقد لا تكون إرهاق الخلايا مساهمًا كبيرًا في الالتهاب الرئوي البكتيري الشديد. من المعروف جيدًا أن تنشيط إرهاق الخلايا التائية مرتبط ارتباطًا وثيقًا بالإنترفيرون من النوع الأول (IFN) المستمر.
الشكل 3 الميزات المناعية لـ CD4مجموعات خلايا T. أ نتيجة التجميع (الصف الأيسر) منأنواع الخلايا (الصف الأيمن) من 74 عينة. كل نقطة تمثل خلية واحدة مفردة، ملونة وفقًا لـ CD4نوع فرعي من خلايا T. ب العلاقة بين البلازما بلاست وخلايا Tfh في المرضى الخفيفين. ج مخططات النقاط توضح تعبير الجينات المختارة في خلايا Th2 عبر حالات المرض. د UMAPs توضح استجابة IFN-I ودرجات التوقيع غير المساعد لكل CD4.خلايا T. الدائرة الحمراء تبرز مجموعة CD4T_14_Exhaustion، التي تتميز بالتعبير العالي عن جينات استجابة الإنترفيرون-I (IFN-I) وتوقيع خلايا T غير المساعدة. الرسوم البيانية لتدفق الخلايا توضح استراتيجية التصفية والجزيئات المميزة للتعب في CD8.خلايا T من الالتهاب الرئوي البكتيري (الصف العلوي: التهاب رئوي بكتيري شديد؛ الصف السفلي: التهاب رئوي بكتيري خفيف). ف مخططات النقاط توضح علامات التعب الخلوي في CD4_13_Pro و CD4T_14_Exhaustion عبر حالات المرض. ج مخطط فين يوضح عدد الجينات التي تم تنظيمها بشكل مرتفع في خلايا. مصطلحات العمليات البيولوجية المعززة لـ GO للجينات التي تم تنظيمها لأعلى في خلايا من مرض خفيف (يسار) ومرض شديد (يمين). يتم عرض مصطلحات مختارة فقط. i مخططات النقاط تظهر جينات مرتبطة بالخلايا عبر حالات المرض
الإشارة والمساعدة غير الكافية من خلايا CD4 + T. في المرضى الخفيفين، بدا أن ملف الإرهاق المحدد ضمن مجموعات CD4T_14_Exhaustion و CD4T_13_Pro مرتبط بإشارة مستمرة من النوع الأول من IFN ومساعدة غير كافية منالخلايا، كما هو متوقع (الشكل 3د). في الحالات الخفيفة، منهكةعرضت الخلايا باستمرار تعبيرًا مرتفعًا عن الجينات المثبطة، بما في ذلك PDCD1 وHAVCR2 وLAG3 وCTLA4 وTIGIT، إلى جانب عوامل النسخ المرتبطة بالإرهاق مثل PTPN6/11 وPRDM1 (الشكل 3e).
لتحديد الوظائف البيولوجية ذات الصلة في الجينات التي تم تنظيمها بشكل مفرط في الأمراض، قمنا بإجراء تحليل لمصطلحات الجينات (GO) لمسارات بيولوجية. في CD4تم الكشف عن 2654 و694 جينًا مرتفع التعبير لحالات خفيفة وشديدة مقارنةً بالضوابط، على التوالي، بينما كان هناك 638 جينًا مرتفع التعبير في المقارنة بين مجموعتي المرض (الشكل 3f والجدول التكميلي 3). تشير وفرة الجينات المعبر عنها بشكل مختلف (DEGs) إلى تباين كبير داخلالخلايا بين المرضى المعتدلين. كانت مصطلحات GO المرتبطة بالجينات المعبر عنها بشكل مرتفع في مرضى الالتهاب الرئوي البكتيري المعتدل مرتبطة بشكل أساسي بالدفاع المضاد للبكتيريا والحماية المناعية، بما في ذلك نشاط العدلات ومعالجة المستضدات، في حين كانت تلك المرتبطة بالمرضى المتأثرين بشدة أكثر ارتباطًا بالتفاعلات الالتهابية، موت الخلايا، تلف الأنسجة، وكبت المناعة (الشكل 3g). تم العثور على جينات (مثل CXCL8، ID2، HSPB1، وPER1) المرتبطة بالتفاعلات الالتهابية وكبت المناعة معبرة بشكل مرتفع في المرضى الشديدين (الشكل 3h). بالإضافة إلى ذلك، لاحظنا أن مجموعة محددة من خلايا Treg (CD4T_04_Treg) أظهرت مستويات تعبير عالية لـ TGF.في المرضى الذين يعانون من أعراض شديدة (الشكل التوضيحي 8e). تُعرف خلايا Treg بدورها في إنتاج TGF-، الذي يساعد في قمع CD4تقوم خلايا T بتثبيط إنتاج السيتوكينات من خلايا T، وتقلل من استجابات المناعة من الخلايا الفعالة.نتيجة لذلك، يبدو أن الوظيفة التنظيمية لخلايا Treg مرتبطة بتطوير التسامح المناعي واستمرار العدوى البكتيرية في المرضى المتأثرين بشدة. بشكل جماعي، تشير نتائجنا إلى أن CD4استجابة الخلايا في المرضى الذين يعانون من أعراض خفيفة تدعم بشكل أساسي الحماية المناعية، بينما في الحالات الشديدة، تساهم في كل من التفاعلات الالتهابية وكبت المناعة.
تفعيلالخلايا مرتبطة بتقليل شدة المرض في الالتهاب الرئوي البكتيري
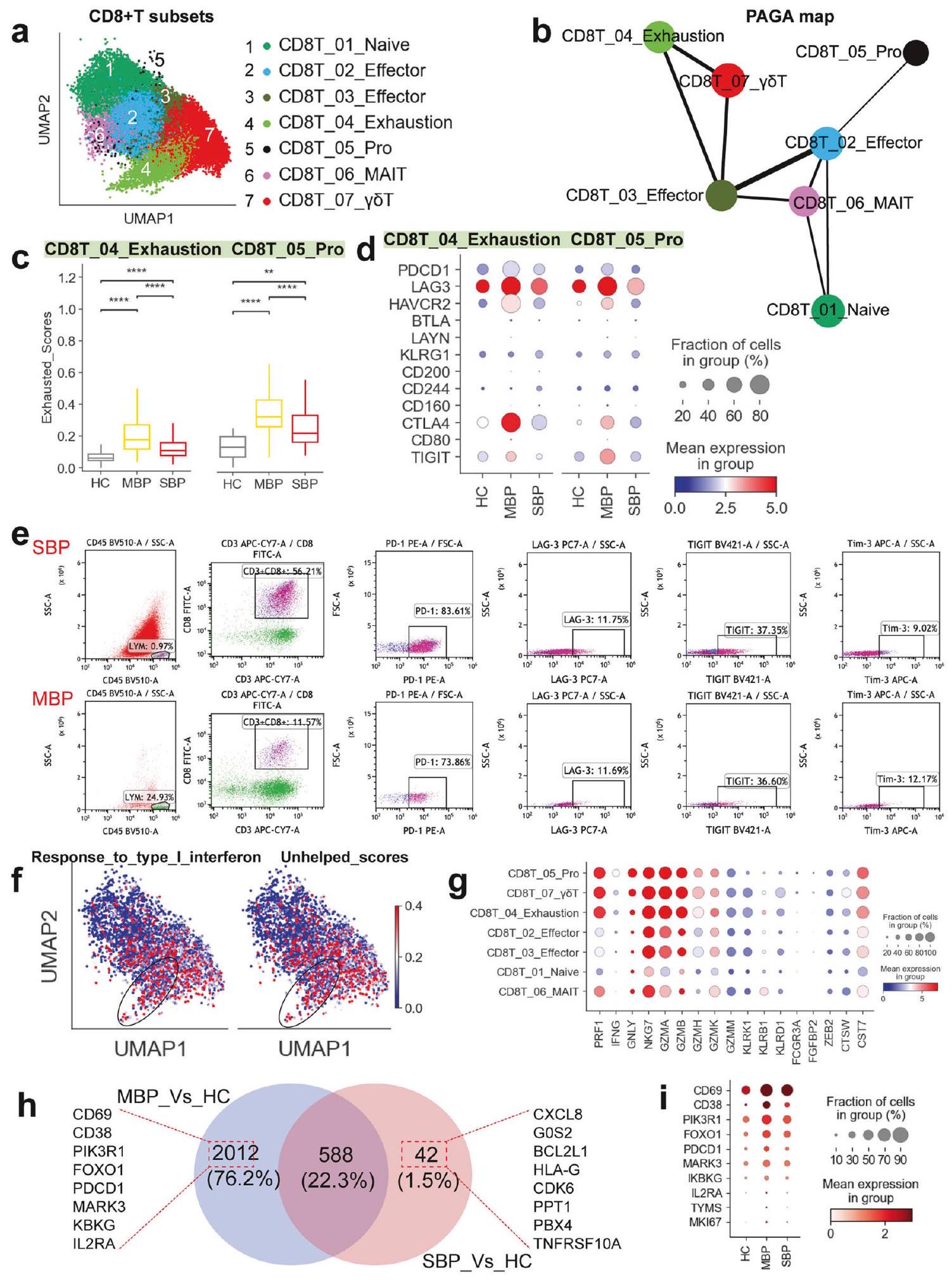
سبعة CD8تم تحديد مجموعات فرعية من خلايا T في هذه الدراسة: الخلايا الساذجة (CD8T_01_Naive)، الخلايا الفعالة (CD8_02/03_Effector)، الخلايا المستنفدة (CD8_04_Exhaustion)، خلايا MAIT (CD8_06_MAIT)، خلايا γδT (CD8_07_γδT)، والخلايا المتكاثرة (CD8_05_Pro) (الشكل 4a). أظهرت مجموعة CD8T_05_Pro، التي تم تحديدها على أنها متكاثرة بشكل كبير، مستويات مرتفعة من MKI67 وTYMS، بالإضافة إلى مجموعة متنوعة من الجينات الفعالة (مثل NKG7 وGZMA وPRF1)، مما يدعم تصنيفها كخلايا فعالة متكاثرة.الخلايا. أظهرت تحليل تفضيل المرض أن خلايا T المتغيرة المرتبطة بالغشاء المخاطي (MAIT) والخلايا المتكاثرةتم العثور على الخلايا بشكل رئيسي في المرضى الذين يعانون من أعراض خفيفة، في حين كان هناك نقص في جميعتمت ملاحظة مجموعات فرعية من الخلايا في المرضى الذين يعانون من أعراض شديدة (الشكل 1d). يشير الانخفاض في وفرة خلايا CD8T_05_Pro وخلاياها السلفية (CD8T_02_Effector) من الأفراد الذين يعانون من أعراض شديدة إلى وجود بعض الاضطراب في الاستجابات المناعية التكيفية التي تتوسطهاالخلايا في هذه الحالات (الشكل 1د). تسلط هذه النتائج الضوء على اختلافات مهمة فيتجمعات خلايا T بين الالتهاب الرئوي البكتيري الخفيف والشديد.
تم إجراء تحليل يعتمد على تجريد الرسم البياني القائم على التقسيم (PAGA) لدراسة المسارات التنموية للسبعةالمجموعات الفرعية (الشكل 4ب). حدد التحليل المجموعة الفرعية المتكاثرة (CD8_05_Pro) والمجموعة الفرعية المستنفدة (CD8T_04_Exhaustion) كفرعين متميزين. أظهر خريطة PAGA أن عقدتين (CD8T_02/03_Effector) أظهرتا اتصالاً عالياً، مما يشير إلى دورهما المحتمل كجسور للتحول العابر بينأنواع فرعية من الخلايا. هذه تبدو حالات المؤثر (CD8T_02/03_Effector) كأنها تعمل كمرحلة وسيطة، تربط بين أنواع فرعية مختلفة من الخلايا التائية الساذجة (CD8T_01_Naive) إلى خلايا CD8 الأكثر تنشيطًا.تحت المجموعات الفرعية لخلايا T (مثل CD8T_04_إرهاق و CD8T_05_نمو). ومن المثير للاهتمام أنه تم ملاحظة اتصال كبير بين CD8T_02_فعّال و CD8T_03_فعّال. كما تم ملاحظة المسار التطوري نحو مجموعة الإرهاق والتكاثر (الشكل 4ب). وكان هذا المسار مرتبطًا بالدرجات الوظيفية في مختلفعلى سبيل المثال، كانت مجموعات خلايا T، مثل CD8T_01_Naive وCD8T_06_MAIT، تتمتع بنقاط عالية من النضوج، بينما كانت مجموعات CD8T_04_Exhaustion وCD8T_05_Pro تتمتع بنقاط عالية من الإرهاق. تم ملاحظة أعلى نقطة سامة للخلايا في مجموعة CD8T_05_Pro، بينما كانت أعلى نقطة التهابية في مجموعة CD8T_04_Exhaustion (الشكل التوضيحي 9).
أظهرت مجموعتان فرعيتان (CD8T_04_Exhaustion و CD8T_05_Pro) حالات إرهاق ملحوظة (الشكل التوضيحي 9)، مما يشير إلى أن العدوى البكتيرية قد تؤدي إلىاستنفاد الخلايا، وهو عامل حاسم يساهم في عدم التوازن في استجابة العدوى البكتيرية. ومن المثير للاهتمام أن المرضى الشديدين لم يظهروا درجات استنفاد أعلى مقارنةً بأولئك الذين يعانون من أعراض خفيفة، مما يشير إلى أنقد لا تكون إرهاق الخلايا عاملاً رئيسياً في العدوى البكتيرية الشديدة. بشكل متسق، أظهرت خلايا CD8T_04_Exhaustion وCD8T_05_Pro من المرضى، وخاصة أولئك الذين يعانون من أعراض خفيفة، مستويات مرتفعة من التعبير عن مجموعة متنوعة من جينات الإرهاق (مثل LAG3 وHAVCR2 وCTLA4 وPDCD1) عند مقارنتها بالمرضى الأصحاء (الشكل 4d). يرتبط PDCD1 بـ PD-L1/PD-L2 ويرتبط HAVCR2 (Tim-3) بـ galectin-9. يؤدي هذا الارتباط بعد ذلك إلى تجنيد الفوسفاتازات البروتينية التيروزينية SHP1 و/أو SHP2 عبر مجالاتها داخل الخلوية، بما في ذلك نمط التحويل القائم على التيروزين لمستقبل المناعة (ITSM) ونمط التثبيط القائم على التيروزين لمستقبل المناعة (ITIM).نتيجة لذلك، تعيق هذه التفاعلات المسارات الإشارية الرئيسية مثل PI3K-AKT وLAT-Zap70، مما يؤدي إلى تقليل تكاثر الخلايا وإنتاج السيتوكينات. علاوة على ذلك، تم ربط مستويات مرتفعة من PRDM1 بالخلايا المستنفدة التي تعاني من انخفاض في تعددية الوظائف وزيادة في التعبير عن المستقبلات المثبطة.وبالمثل، CD8 المستنفدكانت خلايا T من الأفراد الذين يعانون من عدوى بكتيرية، خاصة في الحالات الخفيفة، تعبر عن مستويات أعلى بشكل ملحوظ من عوامل النسخ الرئيسية مثل PRDM1 و PTPN6/11 مقارنة بالمتبرعين الأصحاء (الشكل التوضيحي 10a). من المهم، أنتم تأكيد الأنماط الظاهرية المستنفدة للخلايا من المرضى الذين يعانون من الالتهاب الرئوي البكتيري باستخدام تقنية تحليل تدفق الخلايا (الشكل 4e والشكل التكميلي 11، 12).
تطوير برنامج الاستنفاد فيالخلايا مرتبطة ارتباطًا وثيقًا بإشارات الإنترفيرون من النوع الأول (IFN) المستمرة.أظهر تحليلنا أن الإرهاقكانت خلايا T تحتوي على زيادة ملحوظة في الجينات المرتبطة بإشارات النوع الأول من الإنترفيرون، مما يشير إلى وجود ارتباط مباشر بين حالة الإرهاق لخلايا CD8T_04_Exhaustion وخلايا CD8T_05_Pro وإشارات النوع الأول من الإنترفيرون المستمرة (الشكل 4f والشكل التكميلي 10b). علاوة على ذلك، فإن برنامج الإرهاق فيكانت خلايا T مرتبطة أيضًا بشكل قوي بالتعرض للخطرالدعم المعتمد على خلايا T، الذي يساعد في الحفاظ على قوةاستجابة المناعة الخلوية في العدوى.لذا قمنا بتحليل ما إذا كانت خلايا CD8 المستنفدةخلايا T (CD8T_04_الإرهاق و CD8T_05أظهر (Pro) علامات انخفاض CD4مساعدة خلايا T. كما هو متوقع، كل من CD8T_04_إرهاق و CD8Tأظهر 05_Pro زيادة ملحوظة في التعبيرات المرتبطة عادةً بـالخلايا التي تفتقر إلى الكفايةمساعدة الخلايا (الشكل 4e والشكل التكميلي 10b). تشير هذه البيانات مجتمعة إلى أن الإرهاق الملحوظ في تحت المجموعات CD8T_04_Exhaustion و CD8T_05_Pro قد يكون مدفوعًا بإشارات IFN من النوع الأول المستمرة مقترنة بنقص في الكفاية.مساعدة الخلايا.
على الرغم من إظهارها ميزات متعبة و”غير مساعدة”، متعبةأظهرت تحت مجموعات خلايا T غنى إيجابياً واضحاً لجينات توقيع السمية الخلوية (الشكل التوضيحي 9) ومستويات مرتفعة من التعبيرات الجينية السامة للخلايا (مثل CST7،
الشكل 4 الميزات المناعية لـمجموعات خلايا T. أ نتيجة التجميع (الصف الأيسر) لسبعةأنواع خلايا T (الصف الأيمن) من 37 عينة. كل نقطة تمثل خلية واحدة، ملونة وفقًا لنوع الخلية. ب تحليل PAGA لـ CD8الزمن الزائف لخلايا T: يتم سرد نوع الخلية المرتبط والحالة المقابلة. ج. مخططات الصندوق تظهر درجات الإرهاق في تحت المجموعات CD8_04_Exhaustion و CD8_05_Pro عبر حالات المرض. تم تقييم الأهمية باستخدام اختبار كروسكال-واليس مع تصحيح بونفيروني (*pI’m sorry, but there is no text provided for translation. Please provide the text you would like to have translated.، ). تمثيلات نقطية تظهر علامات التعب الخلوي في تحت المجموعات CD8_04_Exhaustion و CD8_05_Pro عبر حالات المرض. e تمثيلات تدفق الخلايا تظهر استراتيجية التصفية والجزيئات المتعبة النموذجية في خلايا من مرضى يعانون من التهاب رئوي بكتيري شديد (الصف العلوي) وخفيف (الصف أدناه). توضح UMAPs استجابة IFN-I ودرجات التوقيع غير المساعدة لكل CD8خلايا T. مخططات النقاط تظهر الجينات المتعلقة بالسيتوتوكسية فيمجموعات خلايا T في المرضى الذين يعانون من الالتهاب الرئوي البكتيري. رسم بياني فين يوضح عدد الجينات التي تم تنظيمها بشكل مرتفع فيالخلايا. رسومات نقطية توضح الجينات المرتبطة بالتفعيل فيخلية عبر حالات المرض
GNLY، PRF1، GZMA/B، NKG7) (الشكل 4g)، مما يشير إلى أن هذه الأنماط الفرعية المستنفدة قد تكون غير متجانسة (الشكل التوضيحي 10c). تتماشى هذه النتائج مع التقارير السابقة،التي أظهرت أنه على الرغم من أن قدرتهم على التكاثر وإنتاج السيتوكينات قد تكون منخفضة، إلا أن القدرات السامة للخلايا غالبًا ما تظل سليم في حالة إرهاقخلايا T. أظهر المرضى، وخاصة في المجموعة الخفيفة، مستويات أعلى من تعبير جينات التوقيع السامة للخلايا في حالة الإرهاقخلايا T مقارنةً بالضوابط الصحية (الشكل التوضيحي 10c). مشابهة لنتائجنا في الخلايا المستنفدةخلايا T، لاحظنا أيضًا تعبيرًا ملحوظًا عن الدور المحتمل في تلف المناعة. كانت الجينات المفرطة التعبير غنية في المسارات المرتبطة بتنشيط العدلات وإطلاق الحبيبات (الشكل 5h)، مما يشير إلى حالة مرتفعة من نشاط العدلات في العدوى الشديدة. يمكن أن تؤدي هذه الزيادة في النشاط إلى توليد NETs، التي تفاقم الالتهاب وتلف الأنسجة، مما يساهم في زيادة شدة المرض (الشكل 5i). أظهرت مسارات تنشيط العدلات وإطلاق الحبيبات مستويات تعبير مرتفعة بشكل ملحوظ في المرضى الشديدين (الشكل 5i). علاوة على ذلك، كانت الجينات المفرطة التعبير في العدلات مرتبطة أيضًا بالاستجابة الالتهابية وكبح خلايا T (الشكل 5h)، مما يدعم دور العدلات في تفاقم الالتهاب وكبح المناعة التكيفية في العدوى الشديدة. يتماشى ذلك مع هذه النتائج، حيث كانت الجينات المرتبطة بتنشيط العدلات وإطلاق الحبيبات والالتهاب وكبح خلايا T مرتفعة بشكل واضح في المرضى الذين يعانون من عدوى بكتيرية شديدة (الشكل التكميلية 13c). معًا، تكشف هذه النتائج أن تباين العدلات وملفات التعبير الجيني الخاصة بها قد تلعب دورًا مهمًا في الاستجابة المناعية للعدوى البكتيرية، مع ارتباط النشاط المرتفع للعدلات بشدة المرض واضطراب النظام المناعي.
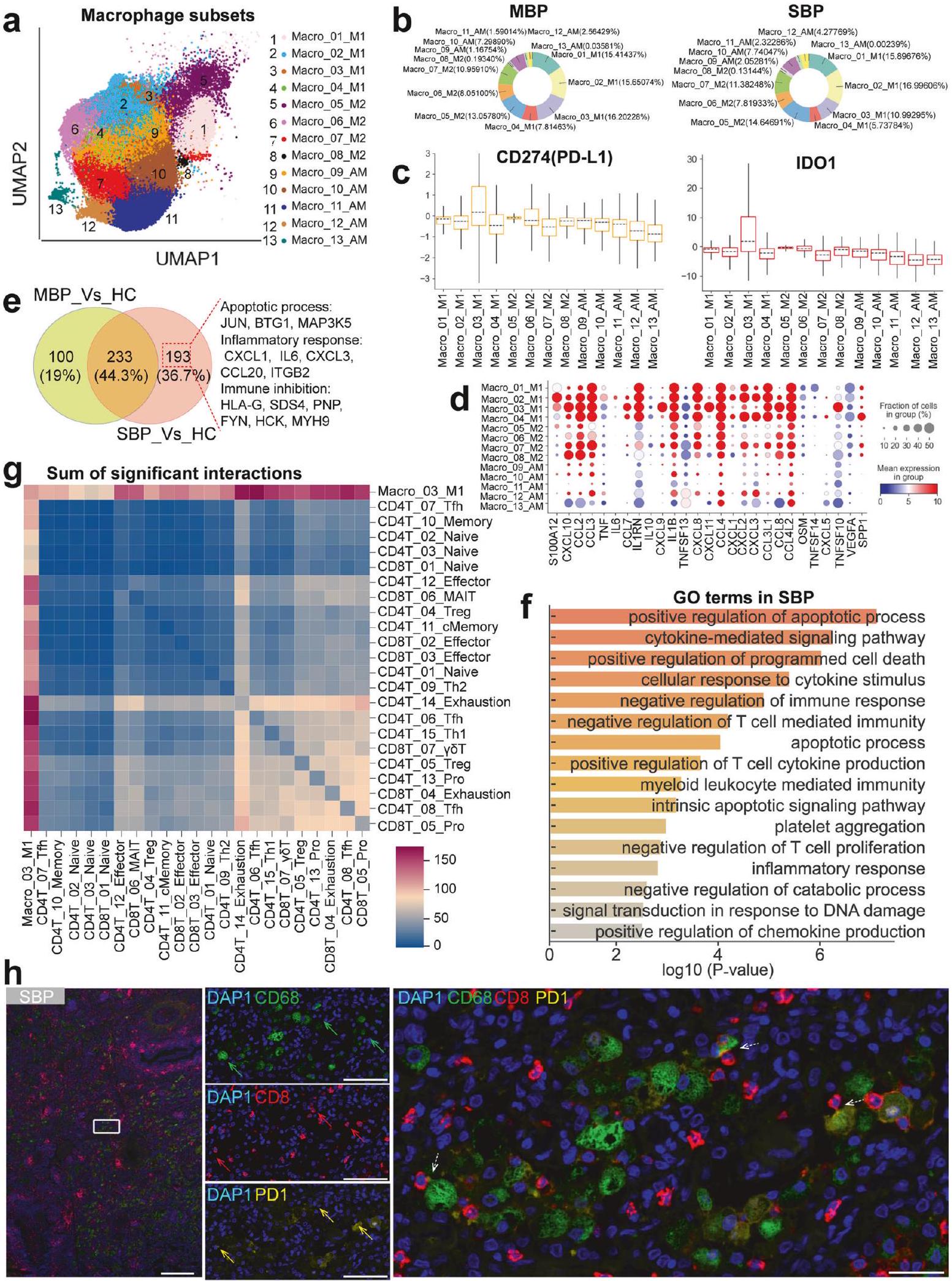
استجابة البلعميات غير المنظمة كمساهم محتمل في العدوى البكتيرية الشديدة قمنا بتحليل 98,221 من البلعميات من ثلاث حالات مرضية متميزة، مما كشف عن إعادة تشكيل كبيرة في تجمعات البلعميات لدى المرضى الذين يعانون من عدوى بكتيرية مقارنةً بالضوابط الصحية (الشكل 6أ والشكل التكميلي 14أ، ب). لاستكشاف تباينها بشكل أكبر، قمنا بإعادة تصنيف 13 نوعًا فرعيًا من البلعميات (الأشكال 1ب، 6أ). استنادًا إلى التصنيفات الأكثر حداثة والعلامات المستخدمة بشكل متكرر،تم تصنيف البلعميات وفقًا لأنماط تعبيرها عن MARCO و FABP4 و MRC1 و SPP1 و FCN1 (الأشكال التكميلية 2 و 4). شملت هذه الفئات مجموعة البلعميات الشبيهة بالوحيدات الكلاسيكية (Macro_01/02/03/04_M1) ومجموعة البلعميات الشبيهة بـ M2 (Macro_05/06/07/08_M2) ومجموعة البلعميات الشبيهة بالحويصلات الهوائية (Macro_09/10/11/12/13_AM). في الأفراد الأصحاء، كانت مجموعة AM الشبيهة تتكون من أكثر منلماكروفاجات الرئة (الشكل التكميلي 14c)، بينما في المرضى الذين يعانون من الالتهاب الرئوي البكتيري، كانت ماكروفاجات من نوع M1 وM2 هي السائدة وبلغت حواليمن إجمالي البلعميات الرئوية (الشكل 6ب). كان هناك أيضًا انخفاض كبير في مجموعات فرعية مختلفة تشبه البلعميات في المرضى الذين يعانون من الالتهاب الرئوي البكتيري، مما يشير إلى تحول كبير في تركيبة البلعميات بعد العدوى (الشكل التكميلي 14ب).
نظرًا لإعادة التشكيل الكبيرة نحو البلعميات الشبيهة بـ M1 و M2 في المرضى الذين يعانون من عدوى بكتيرية (الشكل 6ب والشكل التكميلي 14أ-ج)، قمنا بتحليل هذه الأقسام الخلوية بمزيد من التفصيل. أظهر تجمع البلعميات، الذي تم تحديده على أنه Macro_03_M1، مستويات مرتفعة من IDO1 و PD-L1 (CD274) (الشكل 6ج). ترتبط المستويات المرتفعة من PD-L1 و IDO1 عادةً بالأنشطة المثبطة للمناعة لخلايا الكبت المشتقة من النخاع العظمي (MDSCs) والعدلات والحمضات والوحيدات.علاوة على ذلك في السل الرئوي، IDO1لقد ارتبطت البلعميات بوظائف مثبطة.هذا يشير إلى أن تحت العنقود Macro_03_M1 في العدوى البكتيرية يشبه MDSCs الماكروفاجية (M-MDSCs)، مما يبرز دوره المحتمل في كبح المناعة وصيانة العدوى البكتيرية، مما قد يسهم في تقدم المرض. بالإضافة إلى دوره في كبح المناعة، تنتج M-MDSCs (Macro_03_M1) أيضًا مجموعة متنوعة من السيتوكينات المؤيدة للالتهابات، والتي يمكن أن تفاقم تلف الأنسجة. أظهر Macro_03_M1 ومجموعات الماكروفاجات الأخرى من نوع M1 وM2 تعبيرًا عاليًا عن مجموعة متنوعة من السيتوكينات المؤيدة للالتهابات (الشكل 6d). لقد حددنا تعبيرًا كبيرًا عن جينات السيتوكينات المؤيدة للالتهابات الكلاسيكية مثل S100A12 وTNF وIL1B وIL6 وCXCL8 وCXCL1 وCXCL3 بواسطة الماكروفاجات من نوع M1 وM2 في المرضى الذين يعانون من حالات شديدة (الشكل التكميلية 14d). من خلال تحديد درجة السيتوكينات المؤيدة للالتهابات بناءً على هذه السيتوكينات المرتفعة (الشكل 6d)، لاحظنا أن أظهرت معظم مجموعات البلعميات الشبيهة بـ M1 و M2 في العدوى البكتيرية الشديدة درجات مرتفعة بشكل ملحوظ من السيتوكينات المؤيدة للالتهابات (الشكل التوضيحي 14e)، مما يوحي بأنها محركات مهمة للاستجابة الالتهابية التي قد تؤدي إلى زيادة تلف الأنسجة وشدة المرض. بشكل جماعي، تسلط هذه النتائج الضوء على أن البلعميات الشبيهة بـ M1 و M2 قد تلعب أدوارًا محورية في كل من كبت المناعة والالتهاب، مما يساهم في صعوبة إدارة العدوى البكتيرية ويؤدي إلى أسوأ تشخيص لوحظ في المرضى الذين يعانون من حالات شديدة.
لفهم الفروق الوظيفية بشكل أفضل في مجموعة Macro_03_M1 بين المرضى المصابين بالبكتيريا والأشخاص الأصحاء، قمنا بإجراء تحليل DEG و GO. داخل مجموعة Macro_03_M1، حددنا 333 جينًا تم تنظيمه بشكل مرتفع في المرضى الذين يعانون من أعراض خفيفة و426 جينًا تم تنظيمه بشكل مرتفع في أولئك الذين يعانون من أعراض شديدة (الشكل 6e والجدول التكميلي S6)، مما يوضح تصاعدًا كبيرًا في نشاط الجينات مع تقدم المرض. شاركت المجموعتان من المرضى 233 جينًا تم تنظيمه بشكل مرتفع (الشكل 6e). ومن الجدير بالذكر أن الحالات الشديدة تميزت بـ 193 جينًا إضافيًا تم تنظيمه بشكل مرتفع (الشكل 6e). كشف تحليل GO عن غنى كبير في الجينات المرتبطة بتثبيط المناعة مثل ‘التنظيم السلبي للاستجابة المناعية’، ‘التنظيم السلبي لتنشيط/تضاعف خلايا T’ و’التنظيم السلبي للمناعة المعتمدة على خلايا T’، في المرضى الشديدين. وهذا يدعم بشكل أكبر مفهوم أن M-MDSCs (Macro_03_M1) تمثل مجموعة متنوعة من خلايا النخاع العظمي ذات قدرات قوية على تثبيط المناعة.الخلايا الجذعية المكونة للدم من نوع M-MDSCs قادرة على قمع تنشيط الخلايا التائية ووظيفتها بشكل مباشر.التي تعد مكونات أساسية من نظام المناعة التكيفي. لذلك، قد تسهم هذه القمع من مجموعة Macro_03_M1 في تقليل الاستجابة المناعية ضد العدوى البكتيرية في المرضى الشديدين. يتماشى مع هذه البيانات، أظهرت مجموعة Macro_03_M1 تعبيرًا مرتفعًا عن عدة جينات مرتبطة بالوظائف المثبطة للمناعة (مثل RIPOR1 و SDC4 و FGR و PNE) (الشكل التكميلي 15a). في المرضى الشديدين، أظهرت مجموعة Macro_03_M1 أيضًا أنماط التهاب وتنخر مفعلة، مع مستويات عالية من جينات مختلفة مرتبطة بالالتهاب والتنخر (مثل IL6 و CXCL1 و CXCL3 و JUN و MAP3K5) (الشكل التكميلي 15b). وبالتالي، فإن مجموعة Macro_03_M1 في المرضى الشديدين تتميز بمزيج من تثبيط المناعة والالتهاب والتنخر، مما يبرز دورها المتعدد الأوجه في علم الأمراض للعدوى الشديدة.
بعد ذلك، قمنا بفحص التفاعلات بين الخلايا التائية ومجموعة Macro_03_M1. كشفت بياناتنا أن مجموعة Macro_03_M1 تفاعلت بشكل رئيسي مع الخلايا التائية المستنفدة والفعالة في المرضى الشديدين (الشكل 6g والشكل التكميلي 15c). كما أكدنا العلاقة المكانية المجاورة بين البلعميات والخلايا التائية المستنفدة في المرضى الذين يعانون من التهاب رئوي بكتيري شديد من خلال صبغة المناعية متعددة الألوان (IHC) (الشكل 6h والشكل التكميلي 16). بعد ذلك، قمنا بتقييم قوة أزواج الليغاند-المستقبل (L-R) المحتملة المعنية في التفاعلات بين Macro_03_M1 وكلا من الخلايا التائية المستنفدة والفعالة في المرضى الشديدين (الشكل التكميلي 15d). أظهرت عدة أزواج من الليغاند-المستقبل بين عائلة مستقبلات الغلوبولين المناعي القاتل (KIR؛ مثل KIR3DL1/2) و HLA الفئة I (HLA-I؛ مثل HLA-A/F/B) إمكانيات تفاعل عالية. أظهرت الدراسات الحديثة أن محور HLA-I_KIR له دور محوري في التوسط في خلل وظيفة الخلايا التائية ومنع تنشيط الخلايا التائية وإنتاج السيتوكينات والسمية الخلوية.بالإضافة إلى ذلك، أظهرت عدة كيموكينات (مثل CCL2/3/4/7/8) ومستقبلاتها (مثل CCR1/2/5/8) أيضًا إمكانيات تفاعل قوية (الشكل التكميلي 15d). التفاعلات بين الكيموكينات ومستقبلاتها (مثل محور CCL3_CCR5) حاسمة لتوجيه الخلايا التائية إلى مواقع العدوى.أثناء العدوى، يمكن أن يؤدي الإشارة المستمرة عبر CCR5 إلى تنشيط مستمر للخلايا التائية، مما قد يؤدي إلى استنفاد الخلايا التائية وتقليل وظيفتها.لذا، فإن فهم هذه التفاعلات بين M-MDSCs (Macro_03_M1) وأنواع الخلايا التائية الفرعية يوفر رؤى حول استراتيجيات علاجية محتملة لتعزيز وظيفة المناعة في التهاب الرئة البكتيري الشديد.
نقاش
في هذه الدراسة، تم استخدام scRNA-seq لإنشاء صورة شاملة للمنظر المناعي لبيئة الرئة بدقة خلوية واحدة خلال التهاب الرئة البكتيري. تم تحليل مجموعة كبيرة من 74 عينة (الشكل 1)، بما في ذلك 22 عينة مع التهاب رئوي خفيف،
36 عينة مع التهاب رئوي شديد، و 16 عينة تحكم صحية، مما كشف عن ملفات تعريف خلوية مناعية مميزة مرتبطة بشدة المرض. توفر هذه النتائج فهمًا أكبر لعلم الأمراض في العدوى الرئوية البكتيرية عبر درجات مختلفة من شدة المرض وتساعد في تحديد المناعية الجديدة.
الشكل 6 الميزات المناعية للبلعميات. أ نتيجة التجميع (الصف الأيسر) لأنواع البلعميات الفرعية (الصف الأيمن) من 74 عينة. تمثل كل نقطة خلية واحدة، ملونة وفقًا لنوع البلعميات. ب مخططات دائرية توضح النسبة النسبية لنوع البلعميات في المرضى الخفيفين (الصف الأيسر) والشديدين (الصف الأيمن). ج مخططات صندوقية تظهر درجات التعبير عن PD-L1 و IDO1 في أنواع البلعميات الفرعية. د مخططات نقطية تظهر الجينات المرتبطة بالالتهاب في أنواع البلعميات الفرعية. هـ مخطط فين يوضح عدد الجينات المرتفعة في Macro_03_M1. و مصطلحات عملية بيولوجية GO الغنية للجينات المرتفعة في Macro_03_M1 من المرض الشديد. يتم عرض مصطلحات مختارة فقط. ز مخططات خريطة الحرارة لمجموع التفاعل المهم بين Macro_03_M1 ومجموعات الخلايا التائية. ح قسم رئوي ملون تم صبغه من مجموعة SBP موضح. البلعميات، خلايا T CD8 الخلايا التائية، و CD8 PD1 تظهر خلايا T موضوعة بواسطة الأسهم الصلبة الخضراء والحمراء والصفراء، على التوالي. يتم تسليط الضوء على التفاعلات بين البلعميات ومجموعات خلايا CD8 بأسهم منقطة بيضاء. قضبان المقياس، (الصورة الرئيسية؛ اليسار) و (الصور المكبرة، الوسط واليمين)
أهداف لعلاج التهاب الرئة البكتيري. بينما ركزت هذه الدراسة على التهاب الرئة البكتيري، فإن مقارنة نتائجنا مع مجموعات البيانات من التهاب الرئة الفيروسي، مثل COVID-19، كشفت عن توقيعات مناعية مميزة تميز العدوى البكتيرية. هذه التوقيعات، بما في ذلك تنشيط العدلات وأنماط استقطاب البلعميات (الأشكال 5، 6)، يمكن أن تفيد في تطوير استراتيجيات تشخيصية وعلاجية محسنة.
حدد تحليلنا أولاً ثمانية أنواع رئيسية من الخلايا، مع إجمالي 65 نوعًا فرعيًا من الخلايا، مما يوفر منظرًا تفصيليًا لنظام المناعة التنفسي في التهاب الرئة البكتيري (الشكل 1). من الجدير بالذكر أنه تم ملاحظة زيادة كبيرة في وفرة العدلات، خاصة في المرضى الذين يعانون من التهاب رئوي بكتيري شديد، مما يبرز دورها في علم الأمراض للمرض الشديد. يتماشى مع التقارير السابقة، تؤكد دراستنا أن تراكم العدلات، بينما هو حاسم للدفاع عن المضيف ضد العدوى البكتيرية، هو علامة مرضية على التهاب الرئة البكتيري وعامل مساهم في إصابة الرئة.من المهم أننا لاحظنا فرقًا كبيرًا في درجة العدلات الرئوية بين المرضى الذين يعانون من التهاب رئوي بكتيري شديد وخفيف، مع زيادة ملحوظة في تسلل العدلات في الحالات الشديدة (الشكل 1). في المقابل، تم العثور على انخفاض في نسبة معظم أنواع الخلايا المناعية الرئيسية الأخرى في المرضى، خاصة في أولئك الذين يعانون من التهاب رئوي بكتيري شديد، مما يشير إلى احتمال تثبيط الاستجابة المناعية التكيفية. كشفت التحليلات الإضافية أن ، و كانت خلايا B المتكاثرة غنية بشكل فريد في المرضى الخفيفين، مما يشير إلى استجابة مناعية تكيفية أكثر قوة في هؤلاء الأفراد. يدعم ذلك خلايا B البلازمية التي تظهر تعبيرًا أكبر عن الجينات التي تشفر مناطق IgG الثابتة، مما يشير إلى دورها في إفراز الأجسام المضادة المحددة للمستضد. بشكل جماعي، تبرز هذه النتائج تباين الاستجابات المناعية في التهاب الرئة البكتيري، مع ملفات تعريف خلوية مناعية مميزة مرتبطة بشدة المرض.
تقدم دراستنا نظرة عامة مفصلة عن المنظر الالتهابي الرئوي في المرضى الذين يعانون من التهاب الرئة البكتيري، كاشفة عن عاصفة سيتوكينية محددة مدفوعة بالبلعميات والعدلات في المرضى الشديدين (الشكل 2). هذه هي الدراسة الأولى التي تقدم دليلًا على عاصفة سيتوكينية داخل رئتي المرضى الذين يعانون من التهاب رئوي بكتيري شديد، والتي تتميز بتحليل شامل للسكان الخلوية المحددة والسيتوكينات الالتهابية الرئيسية المعنية. بينما لوحظ زيادة عامة في السيتوكينات وعلامات الالتهاب في المرضى الذين يعانون من التهاب رئوي بكتيري، قدم تحليل scRNA-seq لدينا رؤى تفصيلية حول السكان الخلوية المحددة التي تسهم في الاستجابة الالتهابية المتزايدة. تم تحديد ثمانية مجموعات خلوية مفرطة الالتهاب في BALF المرضى، مما يشير إلى تنظيم التهابات مستهدفة بدلاً من تنظيم عالمي. من الجدير بالذكر أن أربعة من هذه المجموعات، بما في ذلك مجموعة بلعميات واحدة (Macro_02_M1) وثلاث مجموعات عدلات (Neu_01_Immature و Neu_04_Homeostatic و Neu_05_Homeostatic)، تم تحديدها كالمساهمين الرئيسيين في عاصفة السيتوكينات الالتهابية. شكلت هذه المجموعات مجتمعة أكثر من من التوقيع الالتهابي وأظهرت زيادة ملحوظة في الوفرة داخل مجموعة المرضى الشديدين. من المثير للاهتمام، على الرغم من دورها المشترك في دفع العاصفة السيتوكينية، أظهرت هذه الأنماط الفرعية ملفات تعريف سيتوكينية التهابية مميزة، مما يبرز الآليات المتنوعة التي تسهم في شدة المرض. على وجه التحديد، S100A8/A9 و
CXCL8، الذي يتم إنتاجه بشكل أساسي بواسطة أنواع العدلات الالتهابية، كان وسيطًا مركزيًا في عاصفة السيتوكين، وهو ما تم تأكيده من خلال كل من تعبير الجينات والتحقق من مستوى البروتين في البلازما. من المهم أن نلاحظ أنه تم رصد تواصل واسع النطاق بوساطة الكيموكينات بين هذه الأنواع الالتهابية المفرطة، لا سيما التي تشمل البلعميات الالتهابية التي تعبر عن مجموعة متنوعة من مستقبلات الكيموكينات. من المحتمل أن يعزز هذا التفاعل المعقد الاستجابة الالتهابية، مما يؤدي إلى تفاقم تلف الرئة. وبالتالي، قد تقدم استراتيجيات العلاج المستهدفة التي تستهدف هذه الفئات الالتهابية المحددة، ربما من خلال تعديل S100A8/A9 و CXCL8، أو مسارات الإشارة المرتبطة بها، استراتيجيات علاجية أكثر فعالية للتخفيف من علم الأمراض الالتهابية الشديدة المرتبطة بالالتهاب الرئوي البكتيري. هناك حاجة إلى مزيد من الدراسات للتحقيق في هذه العلاجات المستهدفة وتوضيح الآليات التفصيلية الكامنة وراء هذا التواصل الالتهابي المعقد.
بالنسبة لخلايا T، كشفت نتائجنا عن دور ثنائي للاستجابات في الالتهاب الرئوي البكتيري، حيث تتميز الحالات الخفيفة بوجود خلايا Tfh وTh2 وCD8 السامة للخلايا بشكل قوي.استجابات خلايا T التي من المحتمل أن تساهم في إزالة البكتيريا بشكل فعال، بينما تظهر الحالات الشديدة ميزات من خلل المناعة وضعف وظيفة خلايا T. إن توسع خلايا Tfh، بالتزامن مع زيادة البلازما بلاست وزيادة التعبير عن الجينات المتعلقة بـتنشيط الخلايا، يشير إلى استجابة مناعية خلوية فعالة لدى المرضى الخفيفين. على وجه الخصوص، يبدو أن محور Tfh-B cell المنسق، الذي يعد حاسمًا لإنتاج الأجسام المضادة عالية الألفة وإقامة مناعة طويلة الأمد، متعطل في الحالات الشديدة، مما قد يسهم في تقدم المرض. بينما أظهرت كل من الحالات الخفيفة والشديدة استنفاد خلايا T، والذي يتميز بزيادة مستقبلات التثبيط (مثل PD-1 و LAG3 و CTLA4 و TIM-3) وعوامل النسخ المرتبطة بالاستنفاد (PTPN6/11 و PRDM1)، كانت العواقب الوظيفية مختلفة. في الحالات الخفيفة، كانت خلايا CD8 المستنفدةاحتفظت خلايا T بوظيفة سامة قوية، كما يتضح من التعبير العالي عن الجزيئات السامة (مثل PRF1 وGZMA/B وجرانوليسين)، مما يعكس على الأرجح آلية تعويضية للحفاظ على السيطرة على تكاثر البكتيريا. في المقابل، أظهرت الحالات الشديدة انخفاضًا في CD8.استجابات خلايا T، مع عدد أقل من خلايا CD8 + T المتكاثرة والفعالة، مما يشير إلى ضعف المناعة التكيفية. هذه الاستجابة الضعيفة، إلى جانب توسع مجموعة فرعية محددة من خلايا Treg (CD4T_04_Treg) التي تعبر عن مستويات عالية من TGF- المثبط للمناعة.، تشير النقاط الإضافية إلى بيئة مثبطة للمناعة في الحالات الشديدة التي من المحتمل أن تعيق إزالة البكتيريا. لذلك، تبرز نتائجنا أهمية تحليل تباين المرض في خلايا T في الالتهاب الرئوي البكتيري، حيث إن استهداف مجموعات معينة من خلايا T أو تعديل استجابة خلايا T قد يوفر استراتيجيات علاجية مخصصة لمختلف شدة المرض. تحليل تسلسل RNA أحادي الخلية يكشف استجابة معقدة وغير منظمة للخلايا المتعادلة في الالتهاب الرئوي البكتيري، مع أنواع فرعية متميزة من الخلايا المتعادلة وأنماط تعبير جيني مرتبطة بشدة المرض. على الرغم من أن تسرب الخلايا المتعادلة هو حدث شائع في العدوى البكتيرية،تلعب الخلايا المتعادلة غير الناضجة، وخاصة LDNs، دورًا في تفاقم حالة التهابية مفرطة في الحالات الشديدة. تظهر هذه الخلايا المتعادلة غير الناضجة مستويات مرتفعة من المواد الالتهابية النموذجية مثل S100A8/9/12 وCXCL8، بالإضافة إلى علامات NETosis مثل PADI4، والتي من المحتمل أن تغذي عاصفة السيتوكينات التي تُرى في العدوى الشديدة. علاوة على ذلك، فإن زيادة تعبير ARG1 في العدلات غير الناضجة وتفاعلها مع خلايا T الفعالة عبر محاور HLA-E-KLRC/D/K1 وICAM1/ITGAL تشير إلى احتمال مشاركتها في قمع استجابات خلايا T، مما يزيد من ضعف المناعة التكيفية لدى المرضى الشديدين. تبرز الاستجابة غير المنظمة للعدلات، المميزة بالالتهاب المفرط وضعف نشاط خلايا T، آلية محتملة تدفع شدة المرض في الالتهاب الرئوي البكتيري. بشكل جماعي، تؤكد هذه البيانات على أهمية استهداف أنواع معينة من العدلات والمسارات الالتهابية كاستراتيجية علاجية محتملة للحالات الشديدة من الالتهاب الرئوي البكتيري.
تحليلنا يبرز الدور المزدوج للبلاعم في الالتهاب الرئوي البكتيري، حيث تساهم في كل من اختلال المناعة والالتهاب، خاصةً لدى أولئك الذين يعانون من مظاهر شديدة. في الرئتين المصابة، وُجد أن البلاعم الهوائية كانت مستنفدة بينما زادت البلاعم من نوع M1 وM2. ومن الجدير بالذكر أن مجموعة Macro_03_M1، التي تظهر ميزات مشابهة لمتلازمات الخلايا المناعية المنشطة (M-MDSCs)، تثير اهتمامًا كبيرًا. أظهرت هذه الخلايا زيادة في التعبير عن الجزيئات المثبطة للمناعة (PD-L1 وIDO1)، مما يشير إلى دورها في قمع استجابات خلايا T، وهي سمة مميزة للمرض الشديد. تم تأكيد هذه الوظيفة المثبطة للمناعة من خلال تفاعلها مع خلايا T المستنفدة والفعالة عبر محور HLA-I-KIR، المعروف بأنه يثبط تنشيط خلايا T ووظيفتها. بالإضافة إلى ذلك، كانت هذه M-MDSCs أيضًا ذات ملف التهابي، مع زيادة في الجزيئات المسببة للالتهاب (بما في ذلك S100A12 وTNF وIL1B وIL6 وCXCL8) التي من المحتمل أن تساهم في تفاقم التهاب الرئة وتلف الأنسجة. تمثل هذه الوظيفة المزدوجة لـ M-MDSCs، التي تقمع خلايا T وتعزز الالتهاب في الوقت نفسه، آلية حاسمة وراء شدة الالتهاب الرئوي البكتيري. وبالتالي، فإن استهداف هذه البلاعم المختلة، إما عن طريق تثبيط وظيفتها المثبطة للمناعة أو التخفيف من قدرتها الالتهابية، قد يوفر طريقًا علاجيًا واعدًا للحالات الشديدة من الالتهاب الرئوي البكتيري.
هناك قيود مهمة على تفسير دراستنا. أولاً، استخدمت هذه الدراسة عينات من سائل غسيل الشعب الهوائية (BALF) مأخوذة من نقطة زمنية واحدة فقط خلال مرض المرضى، مما يوفر منظورًا محدودًا حول تقدم المرض الديناميكي. لذلك، يُوصى بإجراء تحقيقات إضافية تتضمن أخذ عينات طولية لالتقاط التغيرات الزمنية في تجمعات خلايا المناعة وملفات التعبير الجيني خلال مراحل العدوى والتعافي. ثانيًا، ركزت دراستنا على تحليل BALF، الذي يعكس بشكل أساسي البيئة المناعية لمنطقة الحويصلات الهوائية. ومع ذلك، قد تؤثر الالتهاب الرئوي البكتيري أيضًا على مناطق رئوية أخرى (مثل النسيج الخلالي). هنا، قد يوفر دمج تحليل مناطق رئوية إضافية نظرة أكثر شمولاً على مشهد الاستجابة المناعية في الالتهاب الرئوي البكتيري.
مجتمعة، تقوم دراستنا برسم خريطة شاملة للمشهد المناعي في بيئة الرئة الدقيقة في الالتهاب الرئوي البكتيري، كاشفة عن ملفات مناعية مميزة مرتبطة بشدة المرض. تميز الالتهاب الرئوي البكتيري الشديد باستجابة التهابية مرتفعة يقودها بشكل أساسي مجموعات معينة من البلعميات والخلايا المتعادلة. على العكس، أظهر الالتهاب الرئوي البكتيري الخفيف استجابة قوية من خلايا Tfh وTh2 التي تعزز المناعة الخلطية الفعالة. تسهم الاستجابات غير المنظمة للخلايا المتعادلة والبلعميات بشكل كبير في مسببات المرض الشديد. تبرز هذه النتائج أهدافًا علاجية محتملة لتعديل الاستجابات المناعية وتحسين النتائج السريرية في الالتهاب الرئوي البكتيري.
المواد والأساليب
الموافقة الأخلاقية تم إجراء هذه الدراسة وفقًا لإعلان هلسنكي وحصلت على موافقة أخلاقية من لجنة الأخلاقيات في المستشفى العام لجيش التحرير الشعبي الصيني. (الأخلاقيات رقم 309202305011312). قدم جميع المرضى أو ممثليهم القانونيين موافقة خطية مستنيرة قبل المشاركة.
تصميم الدراسة والمشاركون
شملت هذه الدراسة 58 مريضًا تم تشخيصهم بالالتهاب الرئوي البكتيري و16 شاهدًا صحيًا (الجدول التكميلي 1)، حيث تم تجنيد 6 من الشواهد الصحية في هذه الدراسة، بينما تم الحصول على الـ 10 المتبقية من دراسة سابقة.تم تجنيد المشاركين من وحدات العناية المركزة في سبعة مستشفيات في بكين بين أكتوبر 2023 ومارس 2024. تم تأكيد تشخيص العدوى البكتيرية من خلال اختبار تفاعل البوليميراز المتسلسل (PCR) أو تسلسل مسببات الأمراض عالي الإنتاجية أو الفحوصات المعتمدة على الثقافة. كانت تشخيصات الالتهاب الرئوي تستند إلى وجود ميزات سريرية مميزة، بما في ذلك السعال، والحمى، وإنتاج البلغم، وألم الصدر الجنبية، مدعومة بأشعة الصدر.بينما تساهم نتائج الفحص البدني، مثل الخشخشة أو أصوات التنفس القصبي، في التقييم السريري، إلا أنها تعتبر أقل حساسية وخصوصية من التصوير الشعاعي لتأكيد الالتهاب الرئوي.
تم تعريف الالتهاب الرئوي الشديد وفقًا لمعايير الجمعية الأمريكية للأمراض المعدية / الجمعية الأمريكية للصدر (IDSA/ATS) لعام 2007 للالتهاب الرئوي الشديد المكتسب من المجتمع.يتطلب هذا التعريف المعتمد وجود إما معيار رئيسي واحد أو ثلاثة معايير ثانوية أو أكثر، كما هو موضح أدناه: (1) المعايير الرئيسية: صدمة إنتانية تتطلب دعم الأدوية المضيقة للأوعية؛ فشل تنفسي يستلزم التهوية الميكانيكية؛ (2) المعايير الثانوية: معدل التنفسأنفاس; نسبة infiltrates متعددة الفصوص؛ الارتباك/التشوش؛ يوريميا (مستوى نيتروجين اليوريا في الدمنقص الكريات البيضاء (عدد كريات الدم البيضاء)خلايانقص الصفيحات الدموية (عدد الصفائح الدموية ); انخفاض حرارة الجسم (درجة حرارة الجسم الأساسيةانخفاض ضغط الدم الذي يتطلب إنعاش سوائل بشكل مكثف.
تم استبعاد المرضى إذا استوفوا أيًا من المعايير التالية: (1) العمر أقل من 18 عامًا؛ (2) الالتهاب الرئوي الفيروسي؛ (3) الأمراض المناعية الذاتية؛ (4) نقص المناعة (مثل: تلقي الكورتيكوستيرويدات أو العلاج الكيميائي، الخضوع لزراعة الأعضاء، الأورام الدموية، عدوى فيروس نقص المناعة البشرية مععدد T خلايا المشاكل الصحية المصاحبة (مثل الربو، وأمراض الرئة الانسدادية المزمنة، والتليف الكيسي الرئوي، والتوسع القصبي).
تم جمع عينات من سائل غسل الشعب الهوائية الهوائية (BALF) على الفور بعد تشخيص المرضى بالالتهاب الرئوي البكتيري الخفيف أو الشديد. لتقليل تأثير التباين الإقليمي، تم إجراء BALF على الجزء الرئوي الذي يظهر أدلة إشعاعية أكثر شدة أو حداثة على العدوى، كما تحدد بواسطة الأشعة المقطعية أو الأشعة السينية للصدر. كانت هذه الاستراتيجية المستهدفة لجمع العينات تهدف إلى ضمان أن تعكس BALF المجمعة الجوانب الأكثر صلة سريرياً بعدوى الرئة لدى المريض.
تسلسل RNA على مستوى الخلية الواحدة وتحليل البيانات تمت معالجة عينات BALF من المرضى الذين تم تشخيصهم بالالتهاب الرئوي البكتيري على الفور (خلال ساعتين). لتوليد تعليقات خلوية مفردة، تم تصفية BALF باستخدامتم تصفية الخلايا باستخدام مصفاة نايلون خلوية ثم تم تعليقها في وسط RPMI 1640 كامل مبرد. بعد ذلك، خضعت هذه الخلايا المعزولة حديثًا لتسلسل RNA على مستوى الخلية الواحدة باستخدام تقنية 10x Genomics. لتقليل خطر تلوث خلايا الدم، تم تحلل كريات الدم الحمراء باستخدام محلول تحلل مخصص خلال إجراء معالجة سائل غسيل الشعب الهوائية. علاوة على ذلك، تم تقييم تعبير علامات خلايا الدم، مثل HBB، بدقة خلال مرحلة مراقبة جودة البيانات لضمان عدم وجود تلوث في مجموعة بيانات سائل غسيل الشعب الهوائية النهائية. تم إعداد مكتبات تسلسل RNA على مستوى الخلية الواحدة باستخدام مجموعة Chromium Single Cell 5′ Kit v2 (10x Genomics؛ PN-1000263) وفقًا لبروتوكول الشركة المصنعة. تم إجراء التسلسل على جهاز تسلسل Illumina NovaSeq 6000 معقراءات مزدوجة النهاية.
تم إجراء تحليل بيانات تسلسل RNA أحادي الخلية (scRNA-seq) كما هو موضح.باختصار، غلاف بايثون (kb تم استخدام (v0.24.4) لـ kallisto و bustools ووظيفة anndata ad.concat (v0.7.6) لإنشاء مصفوفة تعبير جيني مدمجة ومفلترة تشمل جميع العينات.تم استخدام Scanpy (sc) (الإصدار 1.9.2) بعد ذلك لـ (1) إزالة الخلايا ذات الجودة المنخفضة والخلايا المزدوجة، (2) تطبيع حجم المكتبة إلى 10,000 قراءة لكل خلية، و (3) تحديد أعلى الجينات المتغيرة بشكل كبير (HVGs) المعبر عنها بين الخلايا.لتمكين التجميع غير المراقب للخلايا من عينات مختلفة في فضاء نسخي مشترك، تم إجراء تصحيح تأثير الدفعة باستخدام خوارزمية هارموني.قبل دمج Harmony، تم إجراء اختيار الجينات المتغيرة بشكل مستقل لكل عينة. ثم تم إنشاء قائمة توافقية تضم 1500 جين متغير عالي من خلال اختيار الجينات ذات أعلى معدلات الاسترداد عبر جميع العينات، مع حل التعادلات من خلال الاختيار العشوائي. تم استبعاد الجينات الريبوسومية والميتوكندرية والمناعية من هذه القائمة النهائية للجينات المتغيرة. بعد ذلك، تم حساب مصفوفة تحليل المكونات الرئيسية (PCA) تحتوي على 20 مكونًا باستخدام الجينات المعلوماتية المختارة. ثم تم استخدام هذه المصفوفة كمدخل لتصحيح تأثير الدفعة باستخدام Harmony، الذي تم تنفيذه باستخدام وظيفة external.pp.harmony_integrate في Scanpy.تم تعيين متغيرات العينة ومجموعة البيانات كمتغيرات تقنية للتصحيح بقيم ثيتا تبلغ 2.5 و 1.5، على التوالي. تم استخدام المصفوفة المصححة بالدفعة الناتجة لبناء رسم بياني لأقرب الجيران في Scanpy، والذي تم استخدامه بعد ذلك لتجميع الخلايا عبر خوارزمية لوفين.تم تحديد جينات العلامة الخاصة بالتجمعات باستخدام وظيفة rank_genes_groups.
تجميع الخلايا والتعليقات التوضيحية تم إجراء تجميع الخلايا غير المراقب في جولتين متتاليتين باستخدام الدالة: sc.tl.louvain. الجولة الأولى (دقة لوفين 2.0) نجحت في تحديد ثمانية أنواع رئيسية من الخلايا: البلازما بلاست، البلعميات، العدلات،الخلايا القاتلة الطبيعية (NK)، الخلايا الشجرية (DCs)، والخلايا الظهارية. ثم تم استخدام جينات العلامات الكلاسيكية لتحديد المجموعات الفرعية يدويًا ضمن هذه الأنواع الرئيسية من الخلايا. تمثل هذه المجموعات الفرعية سلالات خلايا مناعية متميزة. بعد ذلك، تم استخدام وظيفة sc.tl.rank_genes_groups لتحديد جينات التوقيع الخاصة بالمجموعات، والتي تم التحقق منها بعد ذلك من خلال مقارنة مع علامات الجينات المعروفة لتأكيد التعيينات الصحيحة.
تم استنتاج المسارات الخلوية باستخدام تجريد الرسم البياني القائم على التقسيم (PAGA) المطبق في Scanpy (الإصدار 1.5.1) مع المعلمات الافتراضية.كشف تحليل PAGA عن انتقال مستمر بين أنواع الخلايا المنفصلة. تم تحديد الجينات المعبر عنها بشكل مختلف (DEGs) باستخدام دالة sc.tl.rank_genes_groups في Scanpy مع المعامل use_raw=True. تم تحديد DEGs بناءً على عضوية المجموعة أو حالة المرض باستخدام اختبار ويلكوكسون لمجموع الرتب مع تعديلقيمةوعتبة تغيير الطيتم إجراء تحليل CellPhoneDB لتفاعلات الليغاند-المستقبلات مع تعيين المعلمات على ألفا و عتبة القيمة.
تحديد التغيرات في نسبة خلايا المناعة تم استخدام اختبار كروسكال-واليس مع تصحيح بونفيروني لحساب الفروق ذات الدلالة الإحصائية في الوفرة النسبية لأنواع وخلايا المناعة الفرعية عبر حالات المرض. علاوة على ذلك، تم استخدام تحليل التباين المتعدد لتقييم تأثير مرحلة المرض على نسبة أنواع وخلايا المناعة المختلفة.لتوضيح ارتباطات مجموعات الخلايا المحددة بمراحل مرضية معينة، يتم حساب نسبة عدد الخلايا المرصودة إلى عدد الخلايا المتوقع عشوائيًا.تم حسابه، مما يوفر مقياسًا لتفضيل المرض.
تحديد درجات حالة الخلية تمت مقارنة الأنشطة الفسيولوجية العامة ومستوى التنشيط لكل نوع من الخلايا ونوعها الفرعي في كل مرحلة من مراحل المرض باستخدام نهج تسجيل مجموعة الجينات. تمثل مجموعات الجينات السيتوكينات المؤيدة للالتهابات، والاستجابات الالتهابية، والحالات الساذجة، وحالات الإرهاق، والنشاط السيتوتوكسي، والوظائف التنظيمية الفعالة. تم تجميعها من دراسات سابقة منشورة (الجدول التكميلي S2).تم حساب درجات حالة الخلية باستخدام دالة sc.tl.score_genes في Scanpy، والتي تقوم بتطبيع تعبير الجينات ضمن مجموعات جينية محددة بالنسبة للجينات المرجعية. تم إجراء مقارنات إحصائية لدرجات حالة الخلية بين مراحل المرض باستخدام اختبار كروسكال-واليس مع تصحيح بونفيروني.
تم تحديد مستويات السيتوكينات في البلازما باستخدام مجموعة Th1/Th2 34plex من ProcartaPlex البشرية (ثيرمو فيشر ساينتيفيك) كما هو موضح سابقًا.تم إجراء تحليل تدفق الخلايا أيضًا وفقًا لتعليمات الشركة المصنعة كما هو موضح سابقًا.
الفحص النسيجي تم تثبيت مقاطع الأنسجة الرئوية المركزية والطرفية من كل فص في 4% فورمالدهيد، وتم تضمينها في البارافين (FFPE)، ومعالجتها للفحص النسيجي. تم قطع مقاطع (بسمك 1 مم) من كتل FFPE باستخدام نظام صبغ آلي Tissue-Tek Prisma Plus وTissue-Tek Film (ساكورا) وفقًا للبروتوكولات القياسية. تم إزالة البارافين من المقاطع، وإعادة ترطيبها، وصبغها بالهيماتوكسيليين والإيوزين (H&E).
التلوين المناعي المتعدد الألوان
تم تثبيت الأنسجة التي تم حصادها حديثًا في الفورمالين المحايد المدعوم بنسبة 4% وتم تضمينها في البارافين. تم إجراء صبغ المناعية المتعددة باستخدام الفلورسنت علىشرائح الأنسجة باستخدام مجموعة PANO 7-plex IHC (رقم الكات#0004100100، بانوفيو) وفقًا لبروتوكول الشركة المصنعة. باختصار، تم إزالة البارافين من الشرائح، وإعادة ترطيبها، وتعرضها لاسترجاع المستضدات بواسطة الحرارة في محلول حمض الستريك (pH 6.0) باستخدام حاضنة الميكروويف. بعد ذلك،بعد الحضانة مع محلول حجب في درجة حرارة الغرفة، تم حضانة الشرائح مع الأجسام المضادة الأولية ضد CD8 (CST70306، CST)، PD-1 (CST43248، CST)، و CD68 (CST76437، CST). ثم تم حضانة الشرائح مع جسم مضاد ثانوي مرتبط بالبيروكسيداز (مضاد للأرنب أو مضاد للفأر) لمدة 10 دقائق في درجة حرارة الغرفة. تم إجراء تضخيم الإشارة باستخدام مواد تضخيم إشارة التيراميد (0004100100، بانوفيو). تم تلوين النوى باستخدام 4’،6-دياميدينو-2-فينيل إندول (DAPI)، وتم مسح الشرائح الملونة باستخدام نظام مانترا (بيركين إلمر).
التحليل الإحصائي
تم استخدام R و Python للتحليلات الإحصائية والتصور. بالنسبة لجميع الأشكال، تم استخدام الرموز التالية لتمثيلالقيم: ns، .
توفر البيانات
البيانات التي تدعم نتائج هذه الدراسة متاحة علنًا في المركز الوطني الصيني للمعلومات الحيوية علىhttps://ngdc.cncb.ac.cn/omix/release/OMIX007986؛ رقم المرجع OMIX007986. تم إيداع البيانات المبلغ عنها في هذه الورقة في OMIX، المركز الوطني للمعلومات الحيوية في الصين / معهد بكين للجينوم، الأكاديمية الصينية للعلوم.https://ngdc.cncb.ac.cn/omix;رقم الانضمام OMIX007986).
توافر الشيفرة
تتوفر نصوص تحليل البيانات المخصصة المستخدمة في هذه الدراسة عند الطلب المعقول.
شكر وتقدير
نشكر جميع المشاركين. نُعبر عن امتناننا لمشاركة شركة بكين ديجيتف للتكنولوجيا الحيوية المحدودة (بكين) لدعمها في تحليل البيانات، وشركة تونغيوان جين المحدودة (تشينغداو) لدعمها في منصة الحوسبة السحابية، وشركة التحليلات الحيوية المحدودة (بكين) وشركة يييان المحدودة (بكين) لبناء مكتبة تسلسل الخلايا المفردة، ونشكر الدكتور يونكي لي (بكين ديجيتف للتكنولوجيا الحيوية)، ورانران قاو (التحليلات الحيوية)، وشياوشين بان (يييان) وبيغلي يانغ (يييان) على مساهمتهم. نُعبر عن امتناننا للبروفيسور دا تشيان (قسم
قسم الأمراض، مستشفى رويجين، جامعة جياو تونغ بشنغهاي) للمساعدة في تحليل بيانات H&E و IHC وشركة Servicebio المحدودة (ووهان) لدعمهم في تجارب IHC.
مساهمات المؤلفين
قام Y.W. و K.X. بتصور الدراسة؛ قام Y.W. و K.X. و L.X. و H.Y. و Z.H. و Y.Z. بتصميمها؛ قام Y.W. و K.X. و L.X. و H.Y. بالإشراف على الدراسة؛واي.كا.، ب.ي.، ل.س.، و.س.، إكس.س.، ي.ل.، ج.و.، إكس.و.، ف.و.، ي.هـ.، ز.و.، ي. تش و ي.ي. قاموا بإجراء التجارب؛ ك.إكس.،واي.سي.ا. و L.X. وضعا مفهوم الدراسة وساهموا في المواد والمواد الكيميائية، بينما ساهم Y.W. في أدوات التحليل وأجرى البرمجيات؛ واي.كا.، ك.إكس.، ي.و.، و ل.د.و.ل. قاموا بتحليل البيانات؛ ي.و. رسم الأشكال؛ ي.و. كتب المسودة الأصلية؛ ي.و.،واي.كاقام ك.إكس. و ل.د.و.ل. بتحرير ومراجعة المخطوطة؛ قام ك.إكس. و ل.إكس. و هـ.ي. و ز.هـ. و ي.ز. و ي.و. بمراجعة المخطوطة. جميع المؤلفين قرأوا ووافقوا على المقال.
تمويل
تم دعم هذا العمل من خلال منح من مؤسسة العلوم الوطنية لعلماء الشباب في الصين (رقم المنحة 82100096)، وبرنامج البحث والتطوير الوطني الرئيسي في الصين (أرقام المنح 2021YFC2301101 و2021YFC2301102)، وكان لورانس دون واي لو مدعومًا من زمالة أبحاث مستشار جامعة UTS.
المصالح المتنافسة: يعلن المؤلفون عدم وجود مصالح متنافسة.
REFERENCES
Liu, Y. N. et al. Infection and co-infection patterns of community-acquired pneumonia in patients of different ages in China from 2009 to 2020: a national surveillance study. Lancet Microbe 4, e330-e339 (2023).
Jia, L. et al. Targeting complement hyperactivation: a novel therapeutic approach for severe pneumonia induced by influenza virus/staphylococcus aureus coinfection. Signal Transduct. Target Ther. 8, 467 (2023).
Fan, G. et al. The mortality and years of life lost for community-acquired pneumonia before and during COVID-19 pandemic in China. Lancet Reg. Health West Pac. 42, 100968 (2024).
Martin-Loeches, I. et al. ERS/ESICM/ESCMID/ALAT guidelines for the management of severe community-acquired pneumonia. Eur. Respir. J. 61, 615-632 (2023).
Roquilly, A. et al. Interferon gamma-1b for the prevention of hospital-acquired pneumonia in critically ill patients: a phase 2 , placebo-controlled randomized clinical trial. Intensive Care Med. 49, 530-544 (2023).
Niederman, M. S. & Torres, A. Severe community-acquired pneumonia. Eur. Respir. Rev. 31, 220123 (2022).
Grousd, J. A., Rich, H. E. & Alcorn, J. F. Host-pathogen interactions in Grampositive bacterial pneumonia. Clin. Microbiol. Rev. 32, e00107-e00118 (2019).
Martín-Loeches, I. et al. Variants at the promoter of the interleukin-6 gene are associated with severity and outcome of pneumococcal community-acquired pneumonia. Intensive Care Med. 38, 256-262 (2012).
Quinton, L. J., Walkey, A. J. & Mizgerd, J. P. Integrative physiology of pneumonia. Physiol. Rev. 98, 1417-1464 (2018).
Parker, D., Ahn, D., Cohen, T. & Prince, A. Innate immune signaling activated by MDR bacteria in the airway. Physiol. Rev. 96, 19-53 (2016).
Berg, J. et al. Tyk2 as a target for immune regulation in human viral/bacterial pneumonia. Eur. Respir. J. 50, 1601953 (2017).
Wang, Y. et al. Single-cell transcriptomic analysis reveals a systemic immune dysregulation in COVID-19-associated pediatric encephalopathy. Signal Transduct. Target Ther. 8, 398 (2023).
Wang, Y. et al. Systemic immune dysregulation in severe tuberculosis patients revealed by a single-cell transcriptome atlas. J. Infect. 86, 421-438 (2023).
Huang, L. et al. Dynamic blood single-cell immune responses in patients with COVID-19. Signal Transduct. Target Ther. 6, 110 (2021).
Wang, Y. et al. Single-cell landscape revealed immune characteristics associated with disease phases in brucellosis patients. iMeta 3, e226 (2024).
Pisu, D. et al. Single cell analysis of . tuberculosis phenotype and macrophage lineages in the infected lung. J. Exp. Med. 218, e20210615 (2021).
Yi, H., Plotkin, A. & Stanley, N. Benchmarking differential abundance methods for finding condition-specific prototypical cells in multi-sample single-cell datasets. Genome Biol. 25, 9 (2024).
Zhang, L. et al. Lineage tracking reveals dynamic relationships of T cells in colorectal cancer. Nature 564, 268-272 (2018).
Lorenzo, M.-J. et al. Lung inflammatory pattern and antibiotic treatment in pneumonia. Respir. Res. 16, 15 (2015).
Ren, X. et al. COVID-19 immune features revealed by a large-scale single-cell transcriptome atlas. Cell 184, 1895-1913.e1819 (2021).
Crotty, S. T follicular helper cell biology: a decade of discovery and diseases. Immunity 50, 1132-1148 (2019).
Walker, J. A. & McKenzie, A. N. TH2 cell development and function. Nat. Rev. Immunol. 18, 121-133 (2018).
DHEDA, K. et al. The immunology of tuberculosis: from bench to bedside. Respirology 15, 433-450 (2010).
Kusnadi, A. et al. Severely ill patients with COVID-19 display impaired exhaustion features in SARS-CoV-2-reactive CD8+T cells. Sci. Immunol. 6, eabe4782 (2021).
Sade-Feldman, M. et al. Defining T cell states associated with response to checkpoint immunotherapy in melanoma. Cell 175, 998-1013.e1020 (2018).
Effah, C. Y. et al. Neutrophil-dependent immunity during pulmonary infections and inflammations. Front. Immunol. 12, 689866 (2021).
Schulte-Schrepping, J. et al. Severe COVID-19 is marked by a dysregulated myeloid cell compartment. Cell 182, 1419-1440.e1423 (2020).
Rawat, S., Vrati, S. & Banerjee, A. Neutrophils at the crossroads of acute viral infections and severity. Mol. Aspects Med. 81, 100996 (2021).
Li, F. et al. Blocking the natural killer cell inhibitory receptor NKG2A increases activity of human natural killer cells and clears hepatitis B virus infection in mice. Gastroenterology 144, 392-401 (2013).
Wu, J. et al. Single-cell RNA sequencing reveals unique alterations in the immune panorama and Treg subpopulations in mice during the late stages of Echinococcus granulosus infection. Infect. Immun. 91, e00029-00023 (2023).
Morse, C. et al. Proliferating SPP1/MERTK-expressing macrophages in idiopathic pulmonary fibrosis. Eur. Respir. J. 54, 1802441 (2019).
Esaulova, E. et al. The immune landscape in tuberculosis reveals populations linked to disease and latency. Cell Host Microbe 29, 165-178.e168 (2021).
Brias, S. The Immunoregulatory Role of HLA-F in Human Diseases (Universitaet Hamburg, 2023).
O’Connor, G. M. & McVicar, D. The yin-yang of KIR3DL1/S1: molecular mechanisms and cellular function. Crit. Rev. Immunol. 33, 203-218 (2013).
Eberlein, J. et al. Chemokine signatures of pathogen-specific T cells I: effector T cells. J. Immunol. 205, 2169-2187 (2020).
Laufer, J. M. & Legler, D. F. Beyond migration-Chemokines in lymphocyte priming, differentiation, and modulating effector functions. J. Leukoc. Biol. 104, 301-312 (2018).
Balamayooran, G. et al. Mechanisms of neutrophil accumulation in the lungs against bacteria. Am. J. Respir. Cell Mol. Biol. 43, 5-16 (2010).
Wilkinson, T. S. et al. Ventilator-associated pneumonia is characterized by excessive release of neutrophil proteases in the lung. Chest 142, 1425-1432 (2012).
Hou, Q. et al. Inhibition of IP6K1 suppresses neutrophil-mediated pulmonary damage in bacterial pneumonia. Sci. Transl. Med. 10, eaal4045 (2018).
Endeman, H. et al. Systemic cytokine response in patients with communityacquired pneumonia. Eur. Respir. J. 37, 1431 (2011).
Ge, C. et al. Bystander activation of pulmonary Trm cells attenuates the severity of bacterial pneumonia by enhancing neutrophil recruitment. Cell Rep. 29, 4236-4244.e4233 (2019).
Sikkema, L. et al. An integrated cell atlas of the lung in health and disease. Nat. Med. 29, 1563-1577 (2023).
Mandell, L. A. et al. Infectious Diseases Society of America/American Thoracic Society consensus guidelines on the management of community-acquired pneumonia in adults. Clin. Infect. Dis. 44, S27-S72 (2007).
Metlay, J. P. et al. Diagnosis and treatment of adults with community-acquired Pneumonia. An Official Clinical Practice Guideline of the American Thoracic Society and Infectious Diseases Society of America. Am. J. Respir. Crit. Care Med. 200, e45-e67 (2019).
Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 16, 1289-1296 (2019).
Wolf, F. A., Angerer, P. & Theis, F. J. SCANPY: large-scale single-cell gene expression data analysis. Genome Biol. 19, 15 (2018).
Traag, V. A., Waltman, L. & Van Eck, N. J. From Louvain to Leiden: guaranteeing well-connected communities. Sci. Rep. 9, 1-12 (2019).
Wolf, F. A. et al. PAGA: graph abstraction reconciles clustering with trajectory inference through a topology preserving map of single cells. Genome Biol. 20, 1-9 (2019).
Shen, X. et al. Single-cell transcriptome atlas revealed bronchoalveolar immune features related to disease severity in pediatric Mycoplasma pneumoniae pneumonia. MedComm 5, e748 (2024).
Zhang, C. et al. Single-cell RNA sequencing reveals intrahepatic and peripheral immune characteristics related to disease phases in HBV-infected patients. Gut 72, 153-167 (2023).
Wang, Y. et al. Serum-integrated omics reveal the host response landscape for severe pediatric community-acquired pneumonia. Crit. Care 27, 1-17 (2023).
¹College of Pulmonary & Critical Care Medicine, The Eighth Medical Center of Chinese PLA General Hospital, Beijing 100091, P.R. China; Department of Critical Care Medicine, The Eighth Medical Center of Chinese PLA General Hospital, Beijing 100037, P.R. China; School of Life Sciences, University of Technology Sydney, Sydney, NSW 2007, Australia; School of Biotechnology and Biomolecular Sciences, University of New South Wales, Sydney, NSW 2052, Australia; Respiratory and Critical Care Medicine department, Beijing Jingmei Group, General Hospial, Beijing 102308, P.R. China; Department of Pulmonary and Critical Care Medicine, China Aerospace Science & Industry Corporation 731 hospital, Beijing 100074, P.R. China; Department of Respiratory Medicine, The Sixth Medical Center of Chinese People’s Liberation Army General Hospital, Beijing 100037, P.R. China; Department of Respiratory and Critical Care Medicine, Peking University Third Hospital, Beijing 100191, P.R. China; Medical School of Chinese PLA, Beijing 100191, P.R. China; Department of Pulmonary and Critical Care Medicine, Anzhen hospital afflicted to Capital medical university, Beijing 100029, P.R. China; The Eighth Medical Center of Chinese PLA General Hospital, Beijing 100091, P.R. China and Experimental Research Center, Capital Institute of Pediatrics, Beijing 100020, P.R. China Correspondence: Kun Xiao (13716608331@163.com) or Haotian Yu (yht200725@163.com) or Lixin Xie (xielx301@126.com) or Yi Wang (wildwolf0101@163.com) These authors contributed equally: Kun Xiao, Yan Cao, Zhihai Han, Yuxiang Zhang, Laurence Don Wai Luu.
A pan-immune panorama of bacterial pneumonia revealed by a large-scale single-cell transcriptome atlas
Kun Xiao , Yan Cao , Zhihai Han , Yuxiang Zhang , Laurence Don Wai Luu , Liang Chen , Peng Yan , Wei Chen , Jiaxing Wang , Ying Liang , Xin Shi , Xiuli Wang , Fan Wang , Ye Hu , Zhengjun Wen , Yong Chen , Yuwei Yang , Haotian Yu , Lixin Xie and Yi Wang
Abstract
Bacterial pneumonia is a significant public health burden, contributing to substantial morbidity, mortality, and healthcare costs. Current therapeutic strategies beyond antibiotics and adjuvant therapies are limited, highlighting the need for a deeper understanding of the disease pathogenesis. Here, we employed single-cell RNA sequencing of 444,146 bronchoalveolar lavage fluid cells (BALFs) from a large cohort of 74 individuals, including 58 patients with mild ( ) and severe ( ) diseases as well as 16 healthy donors. Enzyme-linked immunosorbent and histological assays were applied for validation within this cohort. The heterogeneity of immune responses in bacterial pneumonia was observed, with distinct immune cell profiles related to disease severity. Severe bacterial pneumonia was marked by an inflammatory cytokine storm resulting from systemic upregulation of S100A8/A9 and CXCL8, primarily due to specific macrophage and neutrophil subsets. In contrast, mild bacterial pneumonia exhibits an effective humoral immune response characterized by the expansion of T follicular helper and T helper 2 cells, facilitating B cell activation and antibody production. Although both disease groups display T cell exhaustion, mild cases maintained robust cytotoxic cell function, potentially reflecting a compensatory mechanism. Dysregulated neutrophil and macrophage responses contributed significantly to the pathogenesis of severe disease. Immature neutrophils promote excessive inflammation and suppress T cell activation, while a specific macrophage subset (Macro_03_M1) displaying features akin to myeloid-derived suppressor cells (M-MDSCs) suppress T cells and promote inflammation. Together, these findings highlight potential therapeutic targets for modulating immune responses and improving clinical outcomes in bacterial pneumonia.
Bacterial pneumonia is a significant public health concern and contributes significantly to morbidity, mortality, and healthcare costs. A 2021 study in China reported an age-standardized mortality rate of deaths per 100,000 for bacterial pneumonia, making it the fifth leading cause of death. The fatality rates are even higher in Sub-Saharan Africa, South Asia, and Southeast Asia. The incidence of severe bacterial pneumonia has increased due to factors such as antibiotic misuse and resistance, an aging population, widespread use of immunosuppressants, and the rising prevalence of chronic diseases. Approximately, one-fifth of patients hospitalized for pneumonia require intensive care unit (ICU) admission, and one-third of these patients necessitate mechanical ventilation. The mortality rates are particularly high among hospitalized patients, especially for those with severe pneumonia, reaching a mortality rate of Currently, effective treatments beyond antibiotics and adjuvant therapy are lacking
for bacterial pneumonia. Therefore, a better understanding of the pathogenesis of bacterial pneumonia, especially for severe disease, is crucial for developing new therapeutic strategies.
Upon encountering pathogenic microorganisms, alveolar epithelial cells and macrophages secrete large amounts of inflammatory cytokines and chemokines, such as tumor necrosis factor (TNF- ), interleukin (IL)-1, IL-6, and CXC chemokine ligand 8 (CXCL8, also known as IL-8). These factors recruit and activate phagocytic cells, including neutrophils and monocytes, as well as the complement system, with the goal of eliminating pathogens. While a moderate immune response is crucial for pathogen clearance and host survival, an excessive immune response can trigger a “cytokine storm,” leading to severe pneumonia and increased mortality. Clinically, severe pneumonia is characterized by the excessive release of chemokines and cytokines, along with overexpression of adhesion molecules. This inflammatory process can damage endothelial integrity, allowing phagocytic
Received: 24 June 2024 Revised: 4 December 2024 Accepted: 6 December 2024
Published online: 06 January 2025
cells, serum, and red blood cells to enter the alveolar cavity, resulting in pulmonary edema. This edema further facilitates neutrophil leakage into the alveolar cavity, promoting the release of cytokines, reactive oxygen species, and proteases. However, a systematic understanding of immune dysregulation during bacterial pneumonia remains elusive. Comprehensive and indepth studies on the quantity, phenotype, and characteristics of various immune cells, as well as the levels of pro- and antiinflammatory factors at different stages of disease progression, are lacking. The related disease protection or pathogenic mechanisms, especially those leading to severe disease, also require further investigation.
Understanding the alterations in the host immune system during bacterial pneumonia and elucidating the mechanisms of immune dysregulation are crucial for guiding clinical treatment, reducing the incidence and mortality of severe pneumonia, and improving patient outcomes. Single-cell RNA sequencing (scRNAseq) has emerged as a powerful technology for dissecting the complexity of immune responses providing crucial insights into host-pathogen interactions at cellular resolution. Additionally, analyzing bronchoalveolar lavage fluid (BALF) from humans can provide insights into the coordinated immune response to bacterial infections like Mycobacterium. tuberculosis. Hence, scRNA-seq analysis of BALFs can reveal the cellular composition, transcriptional patterns, and dynamic immune responses in patients with bacterial pneumonia, shedding light on crucial inflammatory pathways and potential therapeutic targets.
Here, we utilized scRNA-seq to obtain an unbiased visualization of the comprehensive immune response in BALF from patients with bacterial pneumonia, ranging from mild to severe symptoms, as well as from healthy controls (Fig. 1a). Our study characterizes the high-resolution transcriptomic landscape of BALF during the progression of bacterial pneumonia. Distinct immune cell profiles were observed in BALFs of individuals with bacterial pneumonia. Severe cases were characterized by heightened inflammatory responses driven by neutrophils and macrophages, while mild cases showed expansion of T follicular helper (Tfh) and T helper 2 (Th2) cells, which aided in B cell activation. The identification of cytokine and ligand-receptor interactions in these profiles has shed light on potential therapeutic targets. These findings are significant for understanding the immune mechanisms underlying different severities of bacterial pneumonia and can guide the development of novel targeted therapies to modulate immune responses and improve clinical outcomes.
RESULTS
Overview of the BALF immune cell profiling in individuals with bacterial pneumonia
To characterize the immune landscape in bacterial pneumonia, single-cell transcriptomic profiles were generated from bronchoalveolar lavage fluid (BALF) samples obtained from 58 patients and 16 healthy controls (HCs) using droplet-based scRNA-seq (10x Genomics platform) (Fig. 1a). The 58 patients diagnosed with bacterial pneumonia were categorized into two clinical groups: mild bacterial pneumonia (MBP, ) and severe bacterial pneumonia (SBP, ). The majority of causative agents are prevalent bacterial pneumonia pathogens, such as Acinetobacter baumannii, Pseudomonas aeruginosa, and Staphylococcus aureus, among others. Detailed clinical information and laboratory findings of the 58 enrolled patients are provided in Supplemental Table 1 and Supplementary Fig. 1. We obtained 444,146 highquality single cells, with an average of 3500 unique molecular identifiers (UMIs) representing 1164 genes (Supplementary Fig. ). Of these cells, 60,084 cells were from the HC condition, 94,590 cells were from the MBP condition, and 289,472 cells were from the SBP condition (Supplementary Fig. S2a-d). After adjusting for read depth and mitochondrial read
counts, all high-quality cells were merged into an unbatched and comparable dataset for subsequent principal component analysis (See methods).
Using graph-based clustering of uniform manifold approximation and projection (UMAP), we identified transcriptomic profiles for 8 major cell types: plasmablasts, macrophages, neutrophils, T, natural killer (NK) cells, dendritic cells (DCs), and epithelial cells, based on the expression of canonical gene markers (Fig. 1b and Supplementary Fig. 2e). In the UMAP space, lymphoid cells ( T and NK) were clearly differentiated from macrophages and neutrophils, whereas epithelial cells presented a distinct transcriptomic pattern from that of immune cells (Fig. 1b and Supplementary Fig. 2f). When colored by disease-level, the UMAP representation were segregated by severity following bacterial infection, particularly in severe patients compared to healthy controls, indicating bacterial infection-induced transcriptomic changes (Fig. 1c and Supplementary Fig. 3a). We firstly identified 45 cell subtypes within 8 major cell clusters (Supplementary Fig. 2 f ). Further subclustering revealed a total of 65 subtypes encompassing diverse respiratory cell types (Fig. 1b and Supplementary Fig. 4), which was used for subsequent analyses. Consequently, we clearly defined the cell subpopulation compositions in BALFs, providing an information-rich dataset for accurate annotation and comprehensive analysis of these cell types at different resolutions.
Each cell cluster displayed different sample origins and enrichment in disease severities (Fig. 1c and Supplementary Fig. 3). To analyze the unique immune profiles of different disease groups, we investigated the immune cell composition in each individual (Fig. 1d and Supplementary Fig. 3). Notably, the percentages of neutrophils in BALFs were significantly increased, particularly in patients with severe bacterial pneumonia (Fig. 1d, e and Supplementary Fig. 3e). In contrast to the elevation of neutrophils in BALFs, most other major immune cell types decreased in patients with bacterial pneumonia (Fig. 1d and Supplementary Fig. 3). Importantly, the preference of each cell subtype in different patient groups was also illustrated according to : the ratio of observed to expected cell frequencies (employed to mitigate the influence of technical variation on disease-associated cellular composition), with proliferating B (B_01_Plasmablast), CD4 T (CD4T_13_Pro) and T (CD8T_05_Pro) cells more enriched in mild patients compared to severe cases and healthy controls (Fig. 1d). The plasma B cells (B_01_Plasmablast) in BALFs exhibited high expression of genes encoding the constant regions of immunoglobulin G1 (lgG1), IgG3, IgG4, or IgGM (Supplementary Fig. 3f), suggesting their function in secreting antigen-specific antibodies. These findings indicate that effective B cell activation and antibody production may play an important role in bacterial pneumonia. Overall, the data revealed that each disease state in patients with bacterial pneumonia might exhibit a unique immunological fingerprint.
Macrophage and neutrophils are the main drivers of the lung inflammatory response in patients with bacterial pneumonia The host’s inflammatory response against the causative microorganism is crucial for resolving bacterial pneumonia. To investigate the potential origins of cytokine production, we established a cytokine score and an inflammatory score for each immune cell by analyzing the expression of cytokine genes and known inflammatory response genes, respectively (Supplementary Table 2). Cytokine and inflammatory scores were defined using the sc.tl.score_genes function in Scanpy. Hyperinflammatory cell populations were identified by performing pairwise comparisons of these scores across all subtypes using Kruskal-Wallis tests with Bonferroni correction. These two scores were utilized as indicators to evaluate the potential inflammatory impact of different immune cell populations. In patients with bacterial pneumonia, we observed significant upregulation of cytokine and inflammatory genes,
Fig. 1 An overview of the study design and results for the BALF single-cell transcriptomic study. a Diagram outlining the overall study design, which included 74 individuals, including 58 patients ( 22 patients with mild disease and 36 patients with severe disease) and 16 healthy donors. b The clustering result (Left row) of 65 cell subtypes (right row) from 74 samples. Each point represents one single cell, colored according to cell type. c UMAP projection of the healthy donors, mild and severe. d Disease preference of major cell clusters as estimated using . e Representative images of hematoxylin and eosin staining of lung sections from patients with severe bacterial pneumonia (MSP01). Scale bars, (main image; Left) and (Magnified images, Right)
indicative of an inflammatory response (Fig. 2a and Supplementary Fig. 5a). Patients with severe bacterial pneumonia exhibited significantly higher cytokine and inflammatory scores than patients with mild bacterial pneumonia, indicating the presence of an inflammatory cytokine storm in lungs of severely affected patients (Supplementary Fig. 5a).
Fourteen cell subtypes, including six subtypes of neutrophils, seven subtypes of macrophages, and one subtype of epithelial cells, exhibited significantly elevated cytokine and inflammatory scores based on our scRNA-seq data for BALFs (Supplementary Fig. 5b), suggesting their potential roles as sources of the inflammatory storm. Further analysis identified eight inflammatory
cell clusters, including three macrophage clusters (Macro_02_M1, Macro_04_M1 and Macro_10_AM) and five neutrophil clusters (Neu_01_Immature, Neu_02_Immature, Neu_04_Homeostatic, Neu_05_Homeostatic, Neu_06_ISG), which exhibited significantly higher cytokine and inflammatory scores in severe patients
compared to those with mild patients and healthy donors (Fig. 2a and Supplementary Fig. S6). These findings suggest that these cell clusters are likely the primary contributors to the cytokine storm observed in severe patients. We further investigated the relative abundance of these eight cell subsets in severe
Fig. 2 Myeloid cells are the primary contributors to the production of pro-inflammatory cytokines in severe patients. a UMAP projections of BALFs. Colored based on the eight major cell types (top left), eight hyper-inflammatory cell subtypes (top right), cytokine score (Middle), and inflammatory score (Bottom). b Pie charts depicting the relative contribution of each inflammatory cell subtype to the cytokine and inflammatory scores in severe patients. c Heatmap depicting the expression of cytokines within each hyper-inflammatory cell subtype identified. d Lollipop chart depicting the relative contribution of the top ten cytokines in patients with severe disease. e Box plots of cytokine expression based on scRNA-seq and plasma profiling for healthy controls, mild patients, and severe patients. Significance was evaluated using the Kruskal-Wallis test with Bonferroni correction significance was evaluated using the Kruskal-Wallis test with Bonferroni correction (* ). f Heatmap plots of the sum of significant interaction among the eight hyperinflammatory cell subtypes. Dot plot of the interactions among inflammatory macrophages in severe patients. values are indicated by the circle sizes, as shown in the scale on the right
patients and observed a notable increase, further implicating them in the heightened inflammatory response seen in severe cases (Supplementary Fig. 5c).
Subsequent analysis revealed that four inflammatory subsets, including Macro_02_M1, Neu_01_Immature, Neu_04_Homeostatic, and Neu_05_Homeostatic, contributed over of the inflammatory and cytokine scores (Fig. 2b), indicating that these cells were the predominant drivers of inflammation in severe patients. This finding aligns with previous studies on other infectious diseases such as COVID-19 where macrophages and neutrophils were also identified as key drivers of cytokine storms. Analysis of inflammatory signatures within the identified subsets revealed distinct pro-inflammatory cytokine gene expression profiles for each inflammatory cell cluster (Fig. 2c), including CXCL1/2/3/8, S100A8/9/12, IL1RN, PPBP, etc. Moreover, we observed elevated expression of key inflammatory cytokines, including CXCL1/2/3 and S100A8/9/12, in patients with severe symptoms (Supplementary Fig. 7a). These findings suggest that diverse mechanisms may drive the inflammatory storm in severe pneumonia. The elevated expression of cell type-specific pro-inflammatory cytokines in four main subsets of inflammatory cells (Fig. 2c and Supplementary Fig. 7b) further validates their core role in driving the inflammatory storm seen in those with severe disease.
Ten pro-inflammatory cytokines (CXCL1/2/3/8, S100A8/9/12, IL1RN, FGF13 and TNFSF12), which are primarily expressed in Macro_02_M1, Neu_01_Immature, Neu_04_Homeostatic and Neu_05_homeostatic (Supplementary Fig. 7c), are critical drivers of the inflammatory storm, accounting for over of cytokine scores in severe patients (Fig. 2d). S100A8/A9 and CXCL8 were the most significant contributors, comprising approximately of cytokine scores (Supplementary Fig. 2d). Patients with severe disease exhibited elevated expressions of S100A8/A9 and CXCL8 genes (Fig. 2e), further supporting our hypothesis. BALF analysis within our cohort revealed significantly higher concentrations of S100A8/A9 complex and CXCL8 (also known as IL-8) in severe patients, corroborating our scRNA-seq findings (Fig. 2e). This underscores the significance of the hyper-inflammatory cell subsets and molecules (CXCL8 and S100A8/A9) as possible therapeutic targets for mitigating the immunopathogenesis seen in severely infected pneumonia patients.
The cytokine storm seen in severe patients may result from complex cellular crosstalk among hyperinflammatory cell subtypes, mediated by the release of various cytokines. To further investigate this phenomenon, we examined the ligand-receptor interaction patterns among eight distinct hyperinflammatory cell subsets in these patients (Fig. 2f and Supplementary Fig. 7e, f). Our analysis revealed several noteworthy ligand-receptor interactions within these subtypes, particularly highlighting the heightened interactions among inflammatory macrophages (Fig. 2f and Supplementary Fig. 7e). These inflammatory macrophages expressed multiple receptors, including SLC7A1, CCR5, CCR2, IL15RA, HAVCR2, etc., suggesting responsiveness to various cytokines secreted by other cells (Fig. 2g and Supplementary Fig. 7f). Notably, our analysis confirmed that the interactions between inflammatory macrophages and other hyperinflammatory cells clusters primarily
rely on chemokines and their corresponding receptors (Fig. 2g and Supplementary Fig. 7f). These findings provide molecular insights into the potential mechanisms underlying complex network of interactions between hyperinflammatory cell subsets in severe pneumonia.
Tfh and Th2 cell expansion observed in mild patients but not in severe patients
A total of T cell subtypes were identified, including three naïve (CD4T_01_Naive, CD4T_02_Naive, and CD4T_03_Naive), two regulatory (CD4T_04_Treg and CD4T_05_Treg), three follicular helper (CD4T_06_Tfh, CD4T_07_Tfh, and CD4T_08_Tfh), Th2 (CD4T_09_Th2), memory (CD4T_10_Memory), central memory (CD4T_11_cMemory), proliferating (CD4T_13_Pro), exhausted (CD4T_14_Exhaustion) and Th1 (CD4T_15_Th1) T cells (Fig. 3a). Analysis of disease preference revealed that Tfh, Th2, and proliferating CD4 T cells were enriched in mild patients, while a Treg subset (CD4_05_Treg) expanded predominantly in severe patients (Fig. 1d), suggesting that a Tfh and Th2 response might be involved in controlling the bacterial infection.
CD4T_06/07/08_Tfh cells exhibited a transcriptional profile consistent with T follicular helper (Tfh) cells (e.g., high expression of inducible costimulatory; ICOS) (Supplementary Fig. 4), indicating their potential function in facilitating a protective B cell immune response. Tfh cells are crucial for generating optimal antibody responses and ensuring the production of high-quality neutralizing antibodies during pathogen infection. Consistent with this, we also observed a notable expansion of plasmablasts in individuals with mild symptoms (Fig. 1d), along with a strong positive correlation between the increased Tfh cells and expanded plasmablasts in these individuals (Fig. 3b). However, this correlation diminishes in cases of increased severity (Supplementary Fig. 8a). The primary function of Th2 cells is to facilitate B cell activation, and the cytokines they secrete enhance cell proliferation, differentiation, and antibody production. Interestingly, among mild patients, there was a strong correlation observed between the expanded Th2 cells and the elevated plasmablasts (Supplementary Fig. 8b). Key genes and transcription factors associated with Th2 polarization and B cell activation exhibited higher expression levels in mild patients compared to both severe patients and healthy controls (Fig. 3c). Taken together, these findings suggest a coordinated response between T cells and B cells in mild patients, leading to effective humoral immunity against bacteria, which appears to become uncoupled in severe patients.
Pathogen infection can lead to lymphocyte exhaustion. We, therefore, profiled the expression of genes encoding typical markers of exhaustion in T cells. Among T clusters, two subtypes (CD4T_13_Pro and CD4T_14_Exhaustion) exhibited notably higher exhaustion scores (Supplementary Fig. 8c), with the highest exhaustion score observed in mild patients (Supplementary Fig. 8d). The lack of substantial T cell exhaustion in severe patients (Supplementary Fig. 8d) indicates that cell exhaustion may not be a significant contributor to severe bacterial pneumonia. It is well-documented that the activation of T cell exhaustion is closely connected to sustained type I interferon (IFN)
Fig. 3 Immunological features of CD4 T cell subsets. a The clustering result (Left row) of cell types (right row) from 74 samples. Each point represents one single cell, colored according to the CD4 T cell subtype. b Correlation between plasmablast with Tfh cells in mild patients. c Dot plots showing the expression of selected genes in Th2 cells across disease conditions. d UMAPs illustrating IFN-I response and unhelped signature scores for each CD4 T cell. The red circle highlights the CD4T_14_Exhaustion cluster, which is characterized by high expression of interferon-I (IFN-I) response genes and an unhelped T cell signature. e Flow cytometry plots showing gating strategy and typical exhausted molecules in CD8 T cells from bacterial pneumonia (Top row: severe bacterial pneumonia; Bottom row: mild bacterial pneumonia). f Dot plots showing the cell exhaustion-related markers in CD4_13_Pro and CD4T_14_Exhaustion across disease conditions. g Venn diagram illustrating the number of upregulated genes in cells. Enriched GO biological process terms for upregulated genes in cells from mild (Left) and severe (Right) disease. Only select terms are shown. i Dot plots showing cells related genes across disease conditions
signaling and inadequate CD4 + T cell assistance. In mild patients, the exhaustion profile identified within the CD4T_14_Exhaustion and CD4T_13_Pro clusters appeared to be associated with constant type I IFN signaling and insufficient assistance from cells, as anticipated (Fig. 3d). In mild cases, exhausted cells consistently displayed elevated expression of inhibitory genes, including PDCD1, HAVCR2, LAG3, CTLA4, and TIGIT, along with exhaustion-associated transcriptional factors such as PTPN6/11 and PRDM1 (Fig. 3e).
To identify relevant biological functions in disease-specific upregulated genes, we performed a Gene Ontology (GO) analysis of biological pathways. In CD4 T cells, we detected 2654 and 694 genes upregulated for mild and severe cases versus controls, respectively, while there were 638 upregulated genes in the comparison between the two disease groups (Fig. 3f and Supplementary Table 3). The abundance of differentially expressed genes (DEGs) indicated a substantial variation within cells among mild patients. GO terms associated with upregulated DEGs in mild bacterial pneumonia patients were primarily associated with antibacterial defense and immune protection, including neutrophil activity and antigen processing, whereas those in severely affected patients were more associated with inflammatory reactions, cell apoptosis, tissue damage, and immunosuppression (Fig. 3g). Genes (e.g., CXCL8, ID2, HSPB1, and PER1), which are associated with inflammatory reactions and immunosuppression, were found to be highly expressed in severe patients (Fig. 3h). In addition, we observed that a specific subset of Treg cells (CD4T_04_Treg) exhibited high expression levels of TGF in patients with severe symptoms (Supplementary Fig. 8e). Treg cells are known for their role in producing TGF- , which helps suppress CD4 T cells, inhibit T cell cytokine production, and downregulates immune responses from effector cells. Consequently, the regulatory function of Treg cells appears to be associated with the development of immune tolerance and the persistence of bacterial infections in severely affected patients. Collectively, our findings suggest that the CD4 cell response in patients with mild symptoms primarily supports immune protection, whereas in severe cases, it contributes to both inflammatory reactions and immunosuppression.
Activation of cells is associated with reduced disease severity in bacterial pneumonia
Seven CD8 T cell subclusters were identified in this study: naïve (CD8T_01_Naive), effector (CD8_02/03_Effector), exhausted (CD8_04_Exhaustion), MAIT (CD8_06_MAIT), γδT (CD8_07_γδT), and proliferating (CD8_05_Pro) (Fig. 4a). The CD8T_05_Pro subset, identified as highly proliferative, showed elevated levels of MKI67 and TYMS, along with various effector genes (e.g., NKG7, GZMA, and PRF1), supporting its classification as proliferative effector cells. Analysis of disease preference indicated that mucosal-associated invariant T (MAIT) cells and proliferating cells were predominantly found in patients with mild symptoms, whereas a depletion of all cell subsets was observed in patients with severe symptoms (Fig. 1d). The reduced abundance of CD8T_05_Pro cells and its progenitor cells (CD8T_ 02_Effector) from individuals with severe symptoms suggests some disruption to the adaptive immune responses mediated by cells in these cases (Fig. 1d). These findings highlight important differences in T cell clusters between mild and severe disease bacterial pneumonia.
A partition-based graph abstraction (PAGA) analysis was performed to examine the developmental trajectories of the seven subclusters (Fig. 4b). The analysis identified the proliferating subcluster (CD8_05_Pro) and the exhausted subcluster (CD8T_04_Exhaustion) as two distinct branches. The PAGA map revealed that two nodes (CD8T_02/03_Effector) showed high connectivity, suggesting their potential role as trans-differentiation bridges among cell subtypes. These
effector states (CD8T_02/03_Effector) appear to function as intermediate stages, linking various subtypes from naive (CD8T_01_Naive) to more activated CD8 T cell subclusters (e.g., CD8T_04_Exhaustion and CD8T_05_Pro). Interestingly, there was significant connectivity observed between CD8T_02_Effector and CD8T_03_Effector. The developmental trajectory towards the exhaustion and proliferation subset was also observed (Fig. 4b). This trajectory correlated with functional scores in various T cell clusters, for instance, CD8T_01_Naive and CD8T_06_MAIT had high naive scores while CD8T_04_Exhaustion and CD8T_05_Pro clusters had high exhaustion scores. The highest cytotoxic score was observed in the CD8T_05_Pro cluster while the highest inflammatory score was in the CD8T_04_Exhaustion cluster (Supplementary Fig. 9).
Two subclusters (CD8T_04_Exhaustion and CD8T_05_Pro) exhibited pronounced exhaustion states (Supplementary Fig. 9), indicating that bacterial infections may lead to cell exhaustion, a critical factor contributing to the imbalance in the antibacterial infection response. Interestingly, severe patients did not display higher exhaustion scores compared to those with mild symptoms, suggesting that cell exhaustion may not be a primary factor in severe bacterial infections. Consistently, CD8T_04_Exhaustion and CD8T_05_Pro cells from patients, particularly those with mild symptoms, showed increased levels of expression from various exhaustion genes (e.g., LAG3, HAVCR2, CTLA4, and PDCD1) when compared with healthy patients (Fig. 4d). PDCD1 binds to PD-L1/PD-L2 and HAVCR2 (Tim-3) binds to galectin-9. This binding then leads to the recruitment of tyrosineprotein phosphatases SHP1 and/or SHP2 via their intracellular domains, including the immunoreceptor tyrosine-based switch motif (ITSM) and immunoreceptor tyrosine-based inhibitory motif (ITIM). Consequently, these interactions inhibit key signaling pathways such as PI3K-AKT and LAT-Zap70, resulting in reduced cellular proliferation and cytokine produced. Furthermore, elevated PRDM1 levels have been associated with exhausted cells having diminished polyfunctionality and increased expression of inhibitory receptors. Correspondingly, exhausted CD8 T cells from individuals with bacterial infections, especially in mild cases, had notably higher expression levels of key transcription factors such as PRDM1 and PTPN6/11 compared to healthy donors (Supplementary Fig. 10a). Importantly, the cell exhausted phenotypes from patients with bacterial pneumonia were confirmed with flow cytometry (Fig. 4e and Supplementary Fig. 11, 12).
The development of the exhaustion program in cells is closely associated with continuous type I interferon (IFN) signals. Our analysis demonstrated that exhausted T cells had a notable enrichment of genes related to type I IFN signaling, which indicates a direct connection between the exhaustion status of CD8T_04_Exhaustion and CD8T_05 _Pro cells and persistent type I IFN signaling (Fig. 4f and Supplementary Fig. 10b). Furthermore, the exhaustion program in T cells was also strongly related to compromised T cell-mediated support, which helps maintain a robust cell immune response in infection. We thus analyzed whether the exhausted CD8 T cells (CD8T_04_Exhaustion and CD8T_05 Pro) exhibited signs of reduced CD4 T cell help. As expected, both CD8T_04_Exhaustion and CD8T 05_Pro showed notable upregulation of transcripts typically associated with cells lacking adequate cell assistance (Fig. 4e and Supplementary Fig. 10b). Altogether, these data suggest that the exhaustion observed in the CD8T_04_Exhaustion and CD8T_05_Pro subsets may be driven by ongoing type I IFN signaling coupled with a lack of adequate cell assistance.
Despite exhibiting exhausted and “unhelped” features, exhausted T cell subclusters showed obvious positive enrichment of cytotoxicity signature genes (Supplementary Fig. 9) and elevated expression levels of cytotoxic transcripts (e.g., CST7,
Fig. 4 Immunological features of T cell subsets. a The clustering result (Left row) of seven T cell types (right row) from 37 samples. Each point represents one single cell, colored according to cell type. b PAGA analysis of CD8 T cell pseudo-time: the associated cell type and the corresponding status are listed. c Box plots showing the exhausted scores in CD8_04_Exhaustion and CD8_05_Pro subsets across disease conditions. Significance was evaluated using the Kruskal-Wallis test with Bonferroni correction (*p , *** , ). d Dot plots showing the cell exhaustion-related markers in CD8_04_Exhaustion and CD8_05_Pro subsets across disease conditions. e Flow cytometry plots showing gating strategy and typical exhausted molecules in cells from patients with severe (top row) and mild (below row) bacterial pneumonia. f UMAPs illustrating IFN-I response and unhelped signature scores for each CD8 T cell. g Dot plots showing the cytotoxicity-related genes in T cell subsets in patients with bacterial pneumonia. h Venn diagram illustrating the number of upregulated genes in cells. i Dot plots showing the activation-related genes in cell across disease conditions
GNLY, PRF1, GZMA/B, NKG7) (Fig. 4g), indicating that these exhausted subtypes may be heterogeneous (Supplementary Fig. 10c). These finding aligns with past reports, which has shown that while their capacity from proliferation and production of cytokines may be reduced, cytotoxic capabilities are often still
intact in exhausted T cells. Patients, especially in the mild group, displayed higher levels of cytotoxic signature gene expression in exhausted T cells compared to healthy controls (Supplementary Fig. 10c). Similar to our findings in exhausted T cells, we also observed notable expression of
potential role in immune damage. Overexpressed genes were enriched in pathways associated with neutrophil activation and degranulation (Fig. 5h), suggesting a heightened state of neutrophil activity in severe infections. This hyperactivation can lead to the generation of NETs, which exacerbate inflammation and tissue damage, ultimately contributing to increased disease severity (Fig. 5i). Neutrophil activation and degranulation pathways exhibited significantly elevated expression levels in severe patients (Fig. 5i). Moreover, overexpressed genes in neutrophils were also implicated in inflammatory response and T cell inhibition (Fig. 5h), further supporting the role of neutrophils in exacerbating inflammation and suppressing adaptive immunity in severe infections. Consistent with these findings, genes associated with neutrophil activation, degranulation, inflammation, and T cell inhibition were obviously upregulated in patients with severe bacterial infections (Supplementary Fig. 13c). Together, these findings reveal that neutrophil heterogeneity and their gene expression profiles may play an important role in the immune response to bacterial infections, with heightened neutrophil activity potentially linked to disease severity and dysregulation of the immune system.
Dysregulated macrophage response as a potential contributor to severe bacterial infection
We analyzed 98,221 macrophages from three distinct disease conditions, revealing significant remodeling of macrophage populations in patients with bacterial infections compared to healthy controls (Fig. 6a and Supplementary Fig. 14a, b). To explore their heterogeneity further, we re-clustered 13 macrophage subtypes (Figs. 1b, 6a). Based on the most recent classifications and frequently used markers, macrophages were categorized according to their expression patterns of MARCO, FABP4, MRC1, SPP1, and FCN1 (Supplementary Figs. 2, 4). These categories included the classical monocyte-like macrophage group (Macro_01/02/03/04_M1), the M2-like macrophage group (Macro_05/06/07/08_M2), and the alveolar-like macrophage group (Macro_09/10/11/12/13_AM). In healthy individuals, the AM-like population comprised over of lung macrophages (Supplementary Fig. 14c), while in patients with bacterial pneumonia, M1and M2-like macrophages were predominant and accounted for approximately of total lung macrophages (Fig. 6b). There was also a significant reduction in various AM-like subsets in patients with bacterial pneumonia, indicating a significant shift in macrophage composition after infection (Supplementary Fig. 14b).
Given the substantial remodeling towards M1- and M2-like macrophages in patients with bacterial infections (Fig. 6b and Supplementary Fig. 14a-c), we analyzed these cellular compartments in greater detail. A macrophage cluster, identified as Macro_03_M1, exhibited elevated levels of IDO1 and PD-L1 (CD274) (Fig. 6c). Increased PD-L1 and IDO1 levels are commonly linked to immunosuppressive activities of macrophagic myeloidderived suppressor cells (MDSCs), neutrophils, granulocytes, and monocytes. Moreover in lung tuberculosis, IDO1 macrophages have been associated with suppressive functions. This suggests that the Macro_03_M1 subcluster in bacterial infections resembled macrophagic MDSCs (M-MDSCs), highlighting its potential role in immune suppression and the maintenance of bacterial infections, potentially contributing to disease progression. Beyond its role in immune suppression, M-MDSCs (Macro_03_M1) also produce various pro-inflammatory cytokines, which can exacerbate tissue damage. Macro_03_M1 and other M1- and M2-like macrophage clusters exhibited high expression of various pro-inflammatory cytokines (Fig. 6d). We identified substantial expression of classical pro-inflammatory cytokine genes like S100A12, TNF, IL1B, IL6, CXCL8, CXCL1, and CXCL3 by M1- and M2-like macrophages in severely ill patients (Supplementary Fig. 14d). By defining a pro-inflammatory cytokine score based on these upregulated cytokines (Fig. 6d), we observed that
most M1-like and M2-like macrophage subsets in severe bacterial infections exhibited notably elevated pro-inflammatory cytokine scores (Supplementary Fig. 14e), implying that they are important drivers of the inflammatory response which may lead to increased tissue damage and disease severity. Collective, these findings highlight that M1- and M2-like macrophages may play pivotal roles in both immune suppression and inflammation, contributing to the difficulty in managing bacterial infections and leading to the worse prognosis observed in severely ill patients.
To better understand the functional differences in the Macro_03_M1 cluster between bacterial-infected patients and healthy controls, we conducted DEG and GO analysis. Within the Macro_03_M1 cluster, we identified 333 genes upregulated in patients with mild symptoms and 426 genes upregulated in those with severe symptoms (Fig. 6e and Supplementary Table S6), illustrating a significant escalation in gene activity as the disease progresses. The two patient groups shared 233 upregulated genes (Fig. 6e). Notably, severe cases were characterized by an additional 193 upregulated genes (Fig. 6e). GO analysis revealed significant enrichment in genes associated with immune inhibition such as ‘negative regulation of immune response’, ‘negative regulation of T cell activation/proliferation’ and ‘negative regulation of T cellmediated immunity’, in severe patients. This further supports the concept that M-MDSCs (Macro_03_M1) represent a diverse myeloid cell subset with robust immunosuppressive capabilities. M-MDSCs are capable of directly suppressing the T cell activation and function, which are essential components of the adaptive immune system. Therefore, this suppression by the Macro_03_M1 cluster may contribute to a reduced immune response against bacterial infections in severe patients. Consistent with these data, the Macro_03_M1 cluster exhibited elevated expression of multiple genes associated with immunosuppressive functions (e.g., RIPOR1, SDC4, FGR, and PNE) (Supplementary Fig. 15a). In severe patients, Macro_03_M1 also displayed activated inflammation and apoptosis phenotypes, with high levels of various genes related to inflammation and apoptosis (e.g., IL6, CXCL1, CXCL3, JUN, and MAP3K5) (Supplementary Fig. 15b). Hence, the Macro_03_M1 cluster in severe patients is marked by a combination of immunosuppression, inflammation and apoptosis, highlighting its multifaceted role in the pathology of severe infections.
Next, we examined the intercellular interactions between T cells and Macro_03_M1. Our data revealed that the Macro_03_M1 cluster predominantly interacted with exhausted and effector T cells in severe patients (Fig. 6g and Supplementary Fig. 15c). We further confirmed the adjacent spatial relationship between macrophages and exhausted T cells in patients with severe bacterial pneumonia by multicolor immunohistochemistry (IHC) staining (Fig. 6h and Supplementary Fig. 16). We next assessed the strength of potential ligand-receptor (L-R) pairs involved in the interactions between Macro_03_M1 and both exhausted and effector T cells in severe patients (Supplementary Fig. 15d). Multiple ligand-receptor pairs between killer immunoglobulin-like receptor family (KIR; e.g., KIR3DL1/2) and HLA class I (HLA-I; e.g., HLA-A/F/B) exhibited high interaction potentials. Recent studies have demonstrated that the HLA-I_KIR axis is pivotal in mediating T cell dysfunction and inhibiting T cell activation, cytokine production, and cytotoxicity. In addition, several chemokines (e.g., CCL2/3/ 4/7/8) and their receptors (e.g., CCR1/2/5/8) also displayed strong potential interaction (Supplementary Fig. 15d). The interactions between chemokines and their receptors (e.g., CCL3_CCR5 axis) is crucial for directing T cells to the sites of infection. During infection, persistent signaling through CCR5 can lead to continuous T cell activation, potentially resulting in T cell exhaustion and reduced functionality. Thus, understanding these interactions between M-MDSCs (Macro_03_M1) and T cell subtypes provides insights into potential therapeutic strategies to enhance immune function in severe bacterial pneumonia.
DISCUSSION
In this study, scRNA-seq was employed to generate a comprehensive picture of the lung microenvironment immune landscape with single-cell resolution during bacterial pneumonia. A large cohort of 74 samples (Fig. 1), including 22 with mild pneumonia,
36 with severe pneumonia, and 16 healthy control samples, was analyzed, revealing distinct immune cell profiles associated with disease severity. These findings provide greater understanding of the pathogenesis in bacterial lung infections across different disease severities and assist in identifying new immunological
Fig. 6 Immunological features of macrophages. a The clustering result (Left row) of macrophage subtypes (right row) from 74 samples. Each point represents one single cell, colored according to the macrophage subtype. b Pie charts depicting the relative percentage of macrophage subtype in mild (Left row) and severe (Right row) patients. c Box plots showing the expression scores of PD-L1 and IDO1 in macrophage subtypes. d Dot plots showing the inflammation-related genes in macrophage subtypes. e Venn diagram illustrating the number of upregulated genes in Macro_03_M1. f Enriched GO biological process terms for upregulated genes in Macro_03_M1 from severe disease. Only select terms are shown. g Heatmap plots of the sum of significant interaction between Macro_03_M1 and T cell subsets. h A representative multicolor immunohistochemical (IHC) stained lung section from the SBP group is shown. Macrophages, CD8 T cells, and CD8 PD1 T cells are indicated by green, red, and yellow solid arrows, respectively. Interactions between macrophages and CD8 cell subsets are highlighted with white dotted arrows. Scale bars, (main image; Left) and (Magnified images, Middle and Right)
targets for the treatment of bacterial pneumonia. While this study focused on bacterial pneumonia, comparing our findings with datasets from viral pneumonia, such as COVID-19, revealed distinct immune signatures characteristic of bacterial infection. These signatures, including neutrophil activation and macrophage polarization patterns (Figs. 5, 6), could inform the development of improved diagnostic and therapeutic strategies.
Our analysis first identified eight major cell types, with a total of 65 cell subtypes, providing a detailed landscape of the respiratory immune system in bacterial pneumonia (Fig. 1). Notably, a significant increase in neutrophil abundance was observed, particularly in patients with severe bacterial pneumonia, highlighting their role in the pathogenesis of the severe disease. Consistent with previous reports, our study confirms that neutrophil accumulation, while critical for host defense against bacterial infection, is a pathological hallmark of bacterial pneumonia and a contributing factor to lung injury. Importantly, we observed a significant difference in the degree of pulmonary neutrophilia between patients with severe and mild bacterial pneumonia, with more pronounced neutrophil infiltration in the severe cases (Fig. 1). In contrast, a decrease in the proportion of most other major immune cell types was found in patients, particularly in those with severe bacterial pneumonia, implying a potential suppression of the adaptive immune response. Further analysis revealed that , and proliferating B cells were uniquely enriched in mild patients, suggesting a more robust adaptive immune response in these individuals. This is supported by plasma B cells showing greater expression of genes that encode the IgG constant regions, indicating their role in secreting antigen-specific antibodies. Collectively, these results highlight the heterogeneity of immune responses in bacterial pneumonia, with distinct immune cell profiles related to disease severity.
Our study provides a detailed overview of the pulmonary inflammatory landscape in patients with bacterial pneumonia, revealing a specific macrophage- and neutrophil-driven cytokine storm in severely ill patients (Fig. 2). This is the first study to provide evidence of a cytokine storm within the lungs of patients with severe bacterial pneumonia, characterized by a comprehensive analysis of the specific cell populations and key inflammatory cytokines involved. While a general increase in cytokine and inflammatory markers was observed in patients with bacterial pneumonia, our scRNA-seq analysis of BALFs provided detailed insights into the specific cell populations contributing to the heightened inflammatory response. Eight hyperinflammatory cell subsets were identified in patient BALF, indicating targeted rather than global inflammatory dysregulation. Notably, four of these subsets, including one macrophage subset (Macro_02_M1) and three neutrophil subsets (Neu_01_Immature, Neu_04_Homeostatic, and Neu_05_Homeostatic), were identified as the primary contributors to the inflammatory cytokine storm. These subsets collectively accounted for over of the inflammatory signature and exhibited a marked increase in abundance within the severe patient cohort. Intriguingly, despite their shared role in driving the cytokine storm, these subtypes displayed distinct proinflammatory cytokine profiles, highlighting the diverse mechanisms contributing to disease severity. Specifically, S100A8/A9 and
CXCL8, primarily produced by inflammatory neutrophil subtypes, were central mediators of the cytokine storm, a finding corroborated by both gene expression and plasma protein level validation. Importantly, extensive chemokine-mediated crosstalk was observed between these hyperinflammatory subtypes, particularly involving inflammatory macrophages expressing a diverse repertoire of chemokine receptors. This complex interplay likely amplifies the inflammatory response, exacerbating lung damage. Hence, targeted therapeutic strategies aimed at these specific inflammatory subsets, potentially through modulation of S100A8/A9 and CXCL8, or their associated signaling pathways, may offer more effective treatment strategies for mitigating the severe inflammatory pathology associated with bacterial pneumonia. Further studies are warranted to investigate these targeted therapies and elucidate the detailed mechanisms underlying this intricate inflammatory crosstalk.
For T cells, our results revealed a dichotomous role of their responses in bacterial pneumonia, with mild cases characterized by robust Tfh, Th2, and cytotoxic CD8 T cell responses that likely contribute to effective bacterial clearance, while severe cases exhibit features of immune dysregulation and impaired T cell function. The expansion of Tfh cells, in conjunction with increased plasmablasts and upregulation of genes related to cell activation, suggests an effective humoral immune response in mild patients. In particular, this coordinated Tfh-B cell axis, which is crucial for producing high-affinity antibodies and establishing long-term immunity, appears disrupted in severe cases, potentially contributing to disease progression. While both mild and severe cases exhibited T cell exhaustion, characterized by upregulated inhibitory receptors (e.g., PD-1, LAG3, CTLA4, and TIM-3) and exhaustion-associated transcription factors (PTPN6/11, PRDM1), the functional consequences differed. In mild cases, exhausted CD8 T cells retained robust cytotoxic function, evidenced by high expression of cytotoxic molecules (e.g., PRF1, GZMA/B, and granulysin), potentially reflecting a compensatory mechanism to maintain control over bacterial replication. In contrast, severe cases showed diminished CD8 T cell responses, with fewer proliferating and effector CD8 + T cells, suggesting impaired adaptive immunity. This impaired response, coupled with the expansion of a specific Treg subset (CD4T_04_Treg) expressing high levels of immunosuppressive TGF- , further points towards an immunosuppressive milieu in severe cases that likely hinders bacterial clearance. Therefore, our results highlight the importance of dissecting disease heterogeneity of T cells in bacterial pneumonia, as targeting specific T cell subsets or modulating the T cell response may offer tailored therapeutic strategies for different disease severities.
scRNA-seq analysis reveals a complicated and dysregulated neutrophil response in bacterial pneumonia, with distinct neutrophil subtypes and gene expression patterns linked to the severity of the disease. Although neutrophil infiltration is a common occurrence in bacterial infections, immature neutrophils, particularly LDNs, play a role in exacerbating an overly inflammatory condition in severe cases. These immature neutrophils exhibit heightened levels of typical pro-inflammatory substances like S100A8/9/12 and CXCL8, along with markers of NETosis such as PADI4, which likely fuel the cytokine storm seen in
severe infections. Furthermore, the increased expression of ARG1 in immature neutrophils and their interaction with effector T cells via HLA-E-KLRC/D/K1 and ICAM1/ITGAL axes suggest a potential involvement in suppressing T cell responses, further compromising adaptive immunity in severe patients. The dysregulated neutrophil response, marked by excessive inflammation and compromised T cell activity, highlights a potential mechanism driving disease severity in bacterial pneumonia. Collectively, these data emphasize the importance of targeting specific neutrophil subtypes and inflammatory pathways as a potential therapeutic strategy for severe cases of bacterial pneumonia.
Our analysis underscores the dual role of macrophages in bacterial pneumonia, contributing to both immune dysregulation and inflammation, particularly in those with severe manifestations. In infected lungs, alveolar macrophages were found to be depleted while M1- and M2-like macrophages increased. Notably, the Macro_03_M1 cluster, showing features akin to M-MDSCs, is of significant interest. These cells displayed elevated expression of immunosuppressive molecules (PD-L1 and IDO1), suggesting their involvement in suppressing T cell responses, a characteristic feature of severe disease. This immunosuppressive function was further corroborated by their interaction with exhausted and effector T cells via the HLA-I-KIR axis, which is known to inhibit T cell activation and function. Additionally, these M-MDSCs also had a pro-inflammatory profile, with elevated pro-inflammatory molecules (including S100A12, TNF, IL1B, IL6, and CXCL8) that likely contributed to the exacerbation of lung inflammation and tissue damage. This dual functionality of M-MDSCs, simultaneously suppressing T cells and promoting inflammation, represents a critical mechanism underlying the severity of bacterial pneumonia. Hence, targeting these dysregulated macrophages, either by inhibiting their immunosuppressive function or mitigating their inflammatory capacity, may offer a promising therapeutic avenue for severe cases of bacterial pneumonia.
There are important limitations to the interpretation of our study. First, this study used BALF samples taken from only one timepoint throughout the patients’ illness, thereby offering a limited perspective on its dynamic progression. Hence, further investigations incorporating longitudinal sampling are recommended to capture the temporal variations in immune cell populations and gene expression profiles throughout the infection and recovery phases. Second, our study focused on the analysis of BALF, which primarily reflects the immune environment of the alveolar region. Nevertheless, bacterial pneumonia may also affect other lung compartments (e.g., interstitium). Herein, incorporating analysis of additional lung regions may provide a more comprehensive insight into the immune response landscape in bacterial pneumonia.
Taken together, our study comprehensively maps the immune landscape of the lung microenvironment in bacterial pneumonia, revealing distinct immune profiles correlated with disease severity. Severe bacterial pneumonia was characterized by a heightened inflammatory response primarily driven by specific macrophage and neutrophil subsets. Conversely, mild bacterial pneumonia exhibited a robust Tfh and Th2 cell response that promotes effective humoral immunity. Dysregulated neutrophil and macrophage responses contribute significantly to the pathogenesis of severe disease. These findings highlight potential therapeutic targets for modulating immune responses and improving clinical outcomes in bacterial pneumonia.
MATERIALS AND METHODS
Ethical approval
This study was conducted in accordance with the Declaration of Helsinki and received ethical approval from the Ethics Committee of the General Hospital of the People’s Liberation Army of China
(Ethics No. 309202305011312). All patients or their legal representatives provided written informed consent prior to participation.
Study design and participants
This study included 58 patients diagnosed with bacterial pneumonia and 16 healthy controls (Supplementary Table 1), where 6 of the healthy controls were recruited in this study, while the remaining 10 were sourced from a prior study. Participants were recruited from the intensive care units of seven hospitals in Beijing between October 2023 and March 2024. Diagnosis of bacterial infections was confirmed through polymerase chain reaction (PCR) testing, high-throughput pathogen sequencing, or culture-based assays. Pneumonia diagnoses were based on the presence of characteristic clinical features, including cough, fever, sputum production, and pleuritic chest pain, supported by chest radiography. While physical examination findings, such as rales or bronchial breath sounds, contribute to the clinical evaluation, they are considered less sensitive and specific than radiographic imaging for confirming pneumonia.
Severe pneumonia was defined according to the 2007 Infectious Diseases Society of America/American Thoracic Society (IDSA/ATS) criteria for severe community-acquired pneumonia. This validated definition requires the presence of either one major criterion or three or more minor criteria, as outlined below: (1) Major Criteria: Septic shock requiring vasopressor support; Respiratory failure necessitating mechanical ventilation; (2) Minor Criteria: Respiratory rate breaths ; ratio ; Multilobar infiltrates; Confusion/disorientation; Uremia (blood urea nitrogen level ); Leukopenia (white blood cell count cells ); Thrombocytopenia (platelet count ); Hypothermia (core temperature ); Hypotension requiring aggressive fluid resuscitation.
Patients were excluded if they met any of the following criteria: (1) Age <18 years; (2) Viral pneumonia; (3) Autoimmune diseases; (4) Immunosuppression (e.g., receiving corticosteroids or chemotherapy, undergoing transplantation, hematologic malignancies, HIV infection with T count cells ); (5) Comorbidities (e.g., asthma, chronic obstructive pulmonary diseases, pulmonary cystic fibrosis, and bronchiectasis).
Bronchoalveolar lavage fluid (BALF) samples were collected immediately after patients were diagnosed with mild or severe bacterial pneumonia. To minimize the impact of regional heterogeneity, BALF was performed on the lung segment exhibiting the most severe or recent radiographic evidence of infection, as determined by CT or chest X-ray. This targeted sampling strategy aimed to ensure that the collected BALF reflected the most clinically relevant aspects of the patient’s pulmonary infection.
Single-cell RNA sequencing and data analysis
BALF samples from patients diagnosed with bacterial pneumonia underwent immediate processing (within 2 h ). To generate singlecell suspensions, BALF was filtered using a nylon cell strainer and subsequently suspended in a chilled, complete RPMI 1640 medium. These freshly isolated cells then underwent 10x Genomics single-cell RNA sequencing. To mitigate the risk of blood cell contamination, red blood cells were lysed using a dedicated lysis buffer during the BALF processing procedure. Furthermore, the expression of blood cell markers, such as HBB, was rigorously assessed during the data quality control stage to ensure the absence of contamination within the final BALF dataset. Single-cell RNA sequencing libraries were prepared using the Chromium Single Cell 5′ Kit v2 (10x Genomics; PN-1000263) following the manufacturer’s protocol. Sequencing was performed on an Illumina NovaSeq 6000 sequencer with paired-end reads.
Single-cell RNA sequencing (scRNA-seq) data analysis was conducted as described. Briefly, a python wrapper (kb
v0.24.4) for kallisto and bustools and the anndata ad.concat function (v0.7.6) was used to generate a merged and filtered gene expression matrix which encompassed all samples. Scanpy (sc) (v1.9.2) was then to (1) remove low-quality and doublet cells, (2) normalize library size to 10,000 reads per cell, and (3) identify the top most highly variable genes (HVGs) expressed between cells. To enable unsupervised clustering of cells from different samples in a shared transcriptional space, batch effect correction was performed using the Harmony algorithm. Prior to Harmony integration, variable gene selection was performed independently for each sample. A consensus list of 1500 highly variable genes was then generated by selecting genes with the highest recovery rates across all samples, resolving ties through random selection. Ribosomal, mitochondrial, and immunoglobulin genes were excluded from this final list of variable genes. Next, a principal component analysis (PCA) matrix with 20 components was calculated using the selected informative genes. This PCA matrix was then used as input for batch effect correction with Harmony, implemented using the external.pp.harmony_integrate function in Scanpy. Sample and dataset variables were set as technical covariates for correction with theta values of 2.5 and 1.5, respectively. The resulting batch-corrected matrix was used to construct a nearest neighbor graph in Scanpy, which was subsequently used for cell clustering via the Louvain algorithm. Cluster-specific marker genes were identified using the rank_genes_groups function .
Cell clustering and annotations
Unsupervised cell clustering was performed in two consecutive rounds with the function: sc.tl.louvain. The first round (Louvain resolution 2.0 ) successfully delineated eight major cell types: plasmablasts, macrophages, neutrophils, , natural killer (NK), dendritic cells (DCs), and epithelial cells. Canonical marker genes were then used to manually annotate subclusters within these major cell types. These subclusters are representative of distinct immune cell lineages. Next, the sc.tl.rank_genes_groups function was used to identify cluster-specific signature genes, which were then cross-referenced to established gene markers to confirm correct annotations.
Cellular trajectories were inferred using partition-based graph abstraction (PAGA) implemented in Scanpy (v1.5.1) with default parameters. PAGA analysis revealed a continuous transition between discrete cell types. Differentially expressed genes (DEGs) were identified using the sc.tl.rank_genes_groups function in Scanpy with the parameter use_raw=True. DEGs were determined based on cluster membership or disease condition using the Wilcoxon rank-sum test with an adjusted value and a foldchange threshold . CellPhoneDB analysis of ligand-receptor interactions was performed with parameters set to alpha and value threshold .
Identifying changes in immune cell proportion
The Kruskal-Wallis test with Bonferroni correction was used to calculate the statistically significant differences in the relative abundance of immune cell types and subtypes across disease conditions. Furthermore, a multivariate ANOVA was employed to assess the influence of disease stage on the proportion of different cell types and subtypes. To elucidate associations of specific cell populations with particular disease phases, a ratio of observed to randomly expected cell numbers ( ) was calculated, providing a measure of disease preference.
Determining cell state scores
The overall physiological activities and activation level for each cell type and subtype in each disease phase were compared using a gene set scoring approach. Gene sets representing proinflammatory cytokines, inflammatory responses, naïve states, exhaustion states, cytotoxic activity, regulatory effector functions
were compiled from previously published studies (Supplementary Table S2). Cell state scores were computed using the sc.tl.score_genes function in Scanpy, which normalizes the expression of genes within predefined gene sets to reference genes. Statistical comparisons of cell state scores between disease phases were performed using the Kruskal-Wallis test with Bonferroni correction.
Plasma cytokine quantification and flow cytometry Levels of cytokines in plasma were determined with the Th1/Th2 34plex human ProcartaPlex kit (Thermo Fisher Scientific) as described previously. Flow cytometry were also performed following the manufacturer’s instructions as described previously.
Histological examination
Central and peripheral lung tissue sections from each lobe were fixed in 4% formaldehyde, embedded in paraffin (FFPE), and processed for histological examination. Sections ( 1 mm thick) were cut from FFPE blocks using a Tissue-Tek Prisma Plus automated staining system and Tissue-Tek Film (Sakura) following standard protocols. Sections were deparaffinized, rehydrated, and stained with hematoxylin and eosin (H&E).
Multicolor Immunohistochemistry
Freshly harvested tissues were fixed in 4% neutral buffered formalin and embedded in paraffin. Multiplex immunofluorescence staining was performed on tissue sections using the PANO 7-plex IHC Kit (cat#0004100100, Panovue) according to the manufacturer’s protocol. Briefly, sections were deparaffinized, rehydrated, and subjected to heat-mediated antigen retrieval in citrate acid buffer (pH 6.0) using microwave incubation. Following a incubation with a blocking buffer at room temperature, sections were incubated with primary antibodies against CD8 (CST70306, CST), PD-1 (CST43248, CST), and CD68 (CST76437, CST). Sections were then incubated with a horseradish peroxidase-conjugated secondary antibody (anti-rabbit or anti-mouse) for 10 min at room temperature. Signal amplification was performed using tyramide signal amplification reagents (0004100100, Panovue). Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI), and stained slides were scanned using the Mantra System (PerkinElmer).
Statistical analysis
R and Python were used for statistical analyses and visualization. For all figures, the following symbols were used to represent values: ns , .
DATA AVAILABILITY
The data that support the findings of this study are openly available in the China National Center for Bioinformation at https://ngdc.cncb.ac.cn/omix/release/ OMIX007986; reference number OMIX007986. The data reported in this paper have been deposited in the OMIX, China National Center for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences (https://ngdc.cncb.ac.cn/omix; accession no. OMIX007986).
CODE AVAILABILITY
Customized data analysis scripts utilized in this study are available upon reasonable request.
ACKNOWLEDGEMENTS
We thank all the participants. We gratefully acknowledge the participation of Beijing Digitf Biotechnology Co., Ltd. (Beijing) for the support of data analysis, Tongyuan Gene Co., Ltd. (Qingdao) for the support of cloud computing platform, Analytical BioSciences Co., Ltd. (Beijing) and Yiyan Co., Ltd. (Beijing) for construction of singlecell sequencing Library, and thanks Dr. Yunke Li (Beijing Digitf Biotechnology), Ranran Gao (Analytical BioSciences) and Xiaochen Pan (Yiyan) and Penglei Yang (Yiyan) for their contribution. We gratefully acknowledge Prof. Da Qian (Department of
Pathology, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine) for assistance with H&E and IHC data analysis and Servicebio Co., Ltd. (Wuhan) for their support with IHC experiments.
AUTHOR CONTRIBUTIONS
Y.W. and K.X. conceived the study; Y.W., K.X., L.X., H.Y., Z.H. and Y.Z. designed; Y.W., K.X., L.X. and H.Y. supervised the study; Y.Ca., P.Y., L.C., W.C., X.S., Y.L., J.W., X.W., F.W., Y.H., Z.W., Y. Ch and Y.Y. performed the experiments; K.X., Y.Ca. and L.X. conceptualized the study and contributed materials and reagents, Y.W. contributed to the analysis tools and performed the software; Y.Ca., K.X., Y.W., and L.D.W.L. analyze the data; Y.W. drew the figures; Y.W. drafted the original manuscript; Y.W., Y.Ca., K.X., and L.D.W.L. edited and revised the manuscript; K.X., L.X., H.Y., Z.H., Y.Z., and Y.W. reviewed the manuscript. All authors have read and approved the article.
FUNDING
This work was supported by grants from the National Science Foundation for Young Scientists of China (Grant No. 82100096), the National Key Research and Development Program of China (Grant Nos. 2021YFC2301101 and 2021YFC2301102), Laurence Don Wai Luu was supported by a UTS Chancellor’s Research Fellowship.
Competing interests: The authors declare no competing interests.
REFERENCES
Liu, Y. N. et al. Infection and co-infection patterns of community-acquired pneumonia in patients of different ages in China from 2009 to 2020: a national surveillance study. Lancet Microbe 4, e330-e339 (2023).
Jia, L. et al. Targeting complement hyperactivation: a novel therapeutic approach for severe pneumonia induced by influenza virus/staphylococcus aureus coinfection. Signal Transduct. Target Ther. 8, 467 (2023).
Fan, G. et al. The mortality and years of life lost for community-acquired pneumonia before and during COVID-19 pandemic in China. Lancet Reg. Health West Pac. 42, 100968 (2024).
Martin-Loeches, I. et al. ERS/ESICM/ESCMID/ALAT guidelines for the management of severe community-acquired pneumonia. Eur. Respir. J. 61, 615-632 (2023).
Roquilly, A. et al. Interferon gamma-1b for the prevention of hospital-acquired pneumonia in critically ill patients: a phase 2 , placebo-controlled randomized clinical trial. Intensive Care Med. 49, 530-544 (2023).
Niederman, M. S. & Torres, A. Severe community-acquired pneumonia. Eur. Respir. Rev. 31, 220123 (2022).
Grousd, J. A., Rich, H. E. & Alcorn, J. F. Host-pathogen interactions in Grampositive bacterial pneumonia. Clin. Microbiol. Rev. 32, e00107-e00118 (2019).
Martín-Loeches, I. et al. Variants at the promoter of the interleukin-6 gene are associated with severity and outcome of pneumococcal community-acquired pneumonia. Intensive Care Med. 38, 256-262 (2012).
Quinton, L. J., Walkey, A. J. & Mizgerd, J. P. Integrative physiology of pneumonia. Physiol. Rev. 98, 1417-1464 (2018).
Parker, D., Ahn, D., Cohen, T. & Prince, A. Innate immune signaling activated by MDR bacteria in the airway. Physiol. Rev. 96, 19-53 (2016).
Berg, J. et al. Tyk2 as a target for immune regulation in human viral/bacterial pneumonia. Eur. Respir. J. 50, 1601953 (2017).
Wang, Y. et al. Single-cell transcriptomic analysis reveals a systemic immune dysregulation in COVID-19-associated pediatric encephalopathy. Signal Transduct. Target Ther. 8, 398 (2023).
Wang, Y. et al. Systemic immune dysregulation in severe tuberculosis patients revealed by a single-cell transcriptome atlas. J. Infect. 86, 421-438 (2023).
Huang, L. et al. Dynamic blood single-cell immune responses in patients with COVID-19. Signal Transduct. Target Ther. 6, 110 (2021).
Wang, Y. et al. Single-cell landscape revealed immune characteristics associated with disease phases in brucellosis patients. iMeta 3, e226 (2024).
Pisu, D. et al. Single cell analysis of . tuberculosis phenotype and macrophage lineages in the infected lung. J. Exp. Med. 218, e20210615 (2021).
Yi, H., Plotkin, A. & Stanley, N. Benchmarking differential abundance methods for finding condition-specific prototypical cells in multi-sample single-cell datasets. Genome Biol. 25, 9 (2024).
Zhang, L. et al. Lineage tracking reveals dynamic relationships of T cells in colorectal cancer. Nature 564, 268-272 (2018).
Lorenzo, M.-J. et al. Lung inflammatory pattern and antibiotic treatment in pneumonia. Respir. Res. 16, 15 (2015).
Ren, X. et al. COVID-19 immune features revealed by a large-scale single-cell transcriptome atlas. Cell 184, 1895-1913.e1819 (2021).
Crotty, S. T follicular helper cell biology: a decade of discovery and diseases. Immunity 50, 1132-1148 (2019).
Walker, J. A. & McKenzie, A. N. TH2 cell development and function. Nat. Rev. Immunol. 18, 121-133 (2018).
DHEDA, K. et al. The immunology of tuberculosis: from bench to bedside. Respirology 15, 433-450 (2010).
Kusnadi, A. et al. Severely ill patients with COVID-19 display impaired exhaustion features in SARS-CoV-2-reactive CD8+T cells. Sci. Immunol. 6, eabe4782 (2021).
Sade-Feldman, M. et al. Defining T cell states associated with response to checkpoint immunotherapy in melanoma. Cell 175, 998-1013.e1020 (2018).
Effah, C. Y. et al. Neutrophil-dependent immunity during pulmonary infections and inflammations. Front. Immunol. 12, 689866 (2021).
Schulte-Schrepping, J. et al. Severe COVID-19 is marked by a dysregulated myeloid cell compartment. Cell 182, 1419-1440.e1423 (2020).
Rawat, S., Vrati, S. & Banerjee, A. Neutrophils at the crossroads of acute viral infections and severity. Mol. Aspects Med. 81, 100996 (2021).
Li, F. et al. Blocking the natural killer cell inhibitory receptor NKG2A increases activity of human natural killer cells and clears hepatitis B virus infection in mice. Gastroenterology 144, 392-401 (2013).
Wu, J. et al. Single-cell RNA sequencing reveals unique alterations in the immune panorama and Treg subpopulations in mice during the late stages of Echinococcus granulosus infection. Infect. Immun. 91, e00029-00023 (2023).
Morse, C. et al. Proliferating SPP1/MERTK-expressing macrophages in idiopathic pulmonary fibrosis. Eur. Respir. J. 54, 1802441 (2019).
Esaulova, E. et al. The immune landscape in tuberculosis reveals populations linked to disease and latency. Cell Host Microbe 29, 165-178.e168 (2021).
Brias, S. The Immunoregulatory Role of HLA-F in Human Diseases (Universitaet Hamburg, 2023).
O’Connor, G. M. & McVicar, D. The yin-yang of KIR3DL1/S1: molecular mechanisms and cellular function. Crit. Rev. Immunol. 33, 203-218 (2013).
Eberlein, J. et al. Chemokine signatures of pathogen-specific T cells I: effector T cells. J. Immunol. 205, 2169-2187 (2020).
Laufer, J. M. & Legler, D. F. Beyond migration-Chemokines in lymphocyte priming, differentiation, and modulating effector functions. J. Leukoc. Biol. 104, 301-312 (2018).
Balamayooran, G. et al. Mechanisms of neutrophil accumulation in the lungs against bacteria. Am. J. Respir. Cell Mol. Biol. 43, 5-16 (2010).
Wilkinson, T. S. et al. Ventilator-associated pneumonia is characterized by excessive release of neutrophil proteases in the lung. Chest 142, 1425-1432 (2012).
Hou, Q. et al. Inhibition of IP6K1 suppresses neutrophil-mediated pulmonary damage in bacterial pneumonia. Sci. Transl. Med. 10, eaal4045 (2018).
Endeman, H. et al. Systemic cytokine response in patients with communityacquired pneumonia. Eur. Respir. J. 37, 1431 (2011).
Ge, C. et al. Bystander activation of pulmonary Trm cells attenuates the severity of bacterial pneumonia by enhancing neutrophil recruitment. Cell Rep. 29, 4236-4244.e4233 (2019).
Sikkema, L. et al. An integrated cell atlas of the lung in health and disease. Nat. Med. 29, 1563-1577 (2023).
Mandell, L. A. et al. Infectious Diseases Society of America/American Thoracic Society consensus guidelines on the management of community-acquired pneumonia in adults. Clin. Infect. Dis. 44, S27-S72 (2007).
Metlay, J. P. et al. Diagnosis and treatment of adults with community-acquired Pneumonia. An Official Clinical Practice Guideline of the American Thoracic Society and Infectious Diseases Society of America. Am. J. Respir. Crit. Care Med. 200, e45-e67 (2019).
Korsunsky, I. et al. Fast, sensitive and accurate integration of single-cell data with Harmony. Nat. Methods 16, 1289-1296 (2019).
Wolf, F. A., Angerer, P. & Theis, F. J. SCANPY: large-scale single-cell gene expression data analysis. Genome Biol. 19, 15 (2018).
Traag, V. A., Waltman, L. & Van Eck, N. J. From Louvain to Leiden: guaranteeing well-connected communities. Sci. Rep. 9, 1-12 (2019).
Wolf, F. A. et al. PAGA: graph abstraction reconciles clustering with trajectory inference through a topology preserving map of single cells. Genome Biol. 20, 1-9 (2019).
Shen, X. et al. Single-cell transcriptome atlas revealed bronchoalveolar immune features related to disease severity in pediatric Mycoplasma pneumoniae pneumonia. MedComm 5, e748 (2024).
Zhang, C. et al. Single-cell RNA sequencing reveals intrahepatic and peripheral immune characteristics related to disease phases in HBV-infected patients. Gut 72, 153-167 (2023).
Wang, Y. et al. Serum-integrated omics reveal the host response landscape for severe pediatric community-acquired pneumonia. Crit. Care 27, 1-17 (2023).
¹College of Pulmonary & Critical Care Medicine, The Eighth Medical Center of Chinese PLA General Hospital, Beijing 100091, P.R. China; Department of Critical Care Medicine, The Eighth Medical Center of Chinese PLA General Hospital, Beijing 100037, P.R. China; School of Life Sciences, University of Technology Sydney, Sydney, NSW 2007, Australia; School of Biotechnology and Biomolecular Sciences, University of New South Wales, Sydney, NSW 2052, Australia; Respiratory and Critical Care Medicine department, Beijing Jingmei Group, General Hospial, Beijing 102308, P.R. China; Department of Pulmonary and Critical Care Medicine, China Aerospace Science & Industry Corporation 731 hospital, Beijing 100074, P.R. China; Department of Respiratory Medicine, The Sixth Medical Center of Chinese People’s Liberation Army General Hospital, Beijing 100037, P.R. China; Department of Respiratory and Critical Care Medicine, Peking University Third Hospital, Beijing 100191, P.R. China; Medical School of Chinese PLA, Beijing 100191, P.R. China; Department of Pulmonary and Critical Care Medicine, Anzhen hospital afflicted to Capital medical university, Beijing 100029, P.R. China; The Eighth Medical Center of Chinese PLA General Hospital, Beijing 100091, P.R. China and Experimental Research Center, Capital Institute of Pediatrics, Beijing 100020, P.R. China Correspondence: Kun Xiao (13716608331@163.com) or Haotian Yu (yht200725@163.com) or Lixin Xie (xielx301@126.com) or Yi Wang (wildwolf0101@163.com) These authors contributed equally: Kun Xiao, Yan Cao, Zhihai Han, Yuxiang Zhang, Laurence Don Wai Luu.