تتيح تقنية One-Tip تغطية شاملة للبروتيوم في الخلايا الدنيا والزيجوت الفردية One-Tip enables comprehensive proteome coverage in minimal cells and single zygotes
تتضمن سير العمل في البروتيوميات المعتمدة على مطيافية الكتلة (MS) عادةً عمليات معقدة ومتعددة الخطوات، مما يطرح تحديات تتعلق بفقدان العينات، وإمكانية التكرار، ويتطلب استثمارات كبيرة من الوقت والمال، ومهارات متخصصة. هنا نقدم One-Tip، وهي منهجية بروتيوميات تدمج بسلاسة بين إعداد العينات الفعال في وعاء واحد وتحليل الاكتساب المستقل عن البيانات بدقة ونافذة ضيقة (nDIA). تبسط One-Tip بشكل كبير معالجة العينات، مما يمكّن من التعرف القابل للتكرار علىبروتينات منخلايا هيلا. يتم تسليط الضوء على تعددية One-Tip من خلال تحديد nDIA لـالبروتينات في خلايا مفردة من أجنة الفئران المبكرة. بالإضافة إلى ذلك، تتضمن الدراسة جهاز توزيع الخلايا المفردة Uno.، مما يوضح قدرة One-Tip في بروتيوميات الخلايا المفردة مع تحديد أكثر من 3000 بروتين لكل خلية هيلا. كما نقوم بتوسيع سير عمل One-Tip لتحليل الحويصلات خارج الخلوية (EVs) المستخرجة من بلازما الدم، مما يبرز حساسيتها العالية من خلال تحديدبروتينات من تحضير 16 نانوغرام من EV. توسيع One-Tip لقدرات البروتيوميات، مما يوفر عمقًا أكبر وإنتاجية عبر مجموعة من أنواع العينات.
تستند سير العمل التقليدية في البروتيوميات من الأسفل إلى الأعلى إلى عمليات معالجة العينات متعددة الخطوات، التي تشمل تحلل الخلايا بالجملة واستخراج البروتينات من ملايين الخلايا، وتقليل وتثبيت السيستين، وهضم البروتين، وتنظيف العينات وتركيزها، تليها عملية فصل الببتيدات الاختيارية خارج الخط قبل الحصول على قياسات الكروماتوغرافيا السائلة المتزامنة مع مطياف الكتلة (LC-MS/MS).تقدم الخطوات المختلفة العديد من التحديات، بما في ذلك احتمال تلوث العينات وفقدانها مما يؤدي إلى مشاكل في القابلية للتكرار، واستثمارات كبيرة في الوقت والتكلفة، وضرورة وجود خبرة متخصصة وتدريب. وبالتالي، فإن هذه الحواجز غالبًا ما تثني العلماء في مجال العلوم الطبية الحيوية عن القيام بتجارب البروتيوميات المعتمدة على مطيافية الكتلة.
لمعالجة ذلك، ظهرت عدة طرق عمل في وعاء واحد، مما يسهل عملية تحضير العينات لكل من البروتيوميات الكمية.وبروتيوميات الخلايا المفردة (SCP). تهدف هذه العمليات إلى تبسيط عملية إعداد العينات مع الحفاظ على عمق ودقة التحليل البروتيني. على سبيل المثال، قدم كولاك وآخرون بروتوكولًا مبتكرًا في StageTip.، مما يبسط إعداد عينة البروتينات إلى خمس خطوات فقط. هذه الطريقة مريحة بشكل خاص لأنها يمكن أن تتكيف بسهولة مع تنسيق جيد، مما يجعله مناسبًا للتطبيقات ذات الإنتاجية العالية. لقد اكتسب مفهوم الوعاء الواحد قبولًا واسعًا في مجتمع SCP بسبب كفاءته وبساطته. مؤخرًا، اقترح جونستون وآخرون طريقة لتحضير العينات تدمج تحلل الخلايا،
ت denaturation البروتين والهضم في عملية واحدة تستغرق ساعة واحدةقدم ماتزينجر وآخرون سير عمل شامل يتضمن استراتيجيات محسّنة لجميع مراحل إعداد العينات، موحدة لأطباق 384 بئر..
على الرغم من هذه التقدمات، لا تزال التحديات قائمة. في علم البروتيوميات الجماعي، لا تزال الطرق تتطلب إدخال عينات كبيرة، وتفكيك الخلايا بشكل منفصل، والعديد من خطوات التعامل مع السوائل، ونقل العينات، وأوقات تحليل LC-MS/MS الممتدة لتحقيق العمق المطلوب للبروتيوم. وغالبًا ما تؤدي هذه المتطلبات إلى محدودية الإنتاجية، وإعادة الإنتاج، وقابلية التطبيق، وقد لا تلتقط العمق الكامل للبروتيوم. واحدة من القيود الحرجة في طرق SCP هي عدم وجود خطوة لتنظيف العينة، مما يعرض جودة العينة للخطر وبالتالي عمر أنظمة LC-MS. إدراكًا لهذه الفجوة، يقدم دراستنا سير العمل One-Tip، الذي تم تصميمه بدقة للتغلب على القضايا السائدة في علم البروتيوميات الجماعي. تم تصميم One-Tip بشكل أساسي لتحقيق عمق شامل للبروتيوم بدقة وموثوقية عالية، حتى مع عدد قليل من الخلايا. علاوة على ذلك، نوضح مرونة One-Tip من خلال مجموعة واسعة من التطبيقات، بما في ذلك تحليل خلايا جنينية مبكرة فردية وعينات الحويصلات خارج الخلوية من بلازما الإنسان. على الرغم من أنه لم يتم إنشاؤه خصيصًا لـ SCP، فإننا نوضح أيضًا أنه من الممكن تطبيقه مباشرة على SCP باستخدام موزع Uno Single Cell. يعد هذا سير العمل المبتكر والبسيط بتحويل إعداد عينات البروتيوم، مما يوفر كفاءة وعمق في تحليل البروتيوم.
النتائج
تجسد One-Tip أكثر سير عمل مبسط لعلم البروتيوميات
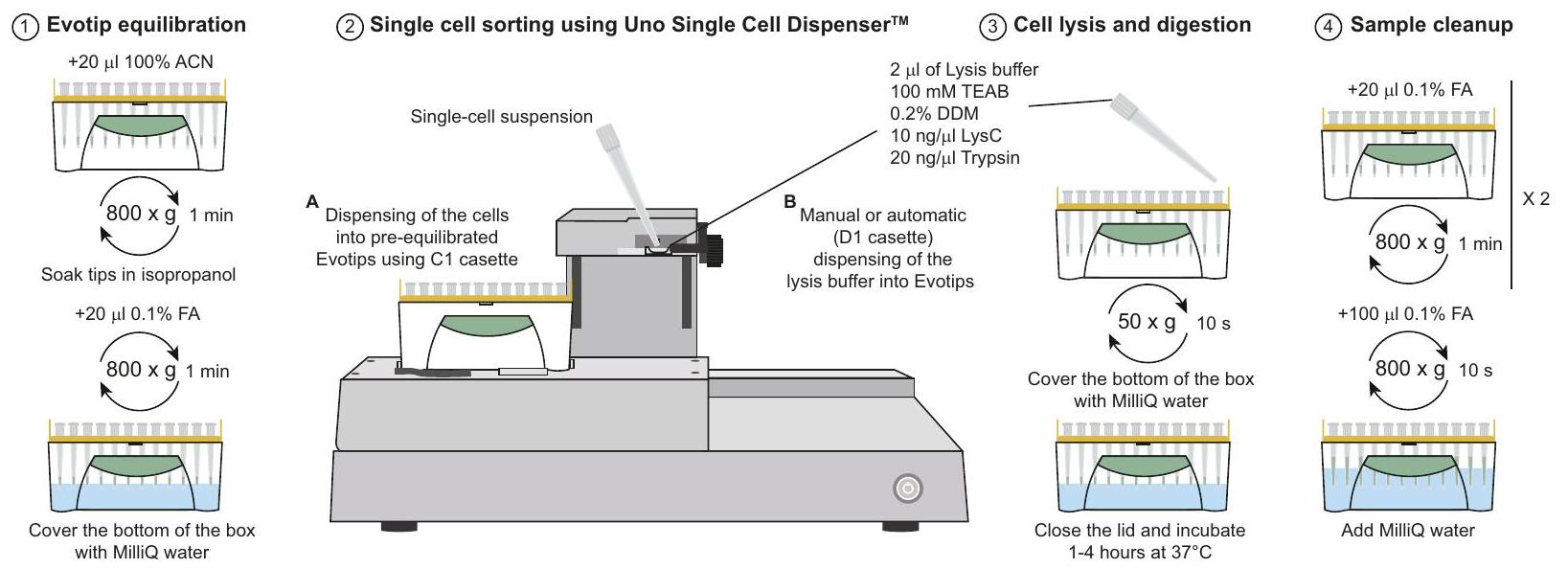
تتطلب منهجية One-Tip خطوتين فقط في عملية السحب، باستخدام جهاز Evotip المتاح تجارياً. : واحدة لتمزق الخلايا ومحلول هضم البروتينات، والأخرى لتعليق الخلايا في PBS (الشكل 1a). بعد ساعة من الحضانة في صندوق Evotip مملوء بالماء مع تحضير Evotip القياسي، تكون عينات One-Tip جاهزة لتحليل LC-MS/MS. يتضمن محلول خلط تمزق الخلايا، واستخراج البروتين، وهضم إنزيم البروتينات Lys-C/تريبسين مزيجًا رئيسيًا يحتوي على مادة خافضة للتوتر السطحي متوافقة مع MS، n-Dodecyl--دي-مالتوسيد (DDM)، وهو منظف غير أيوني قابل للذوبان في الماء فعال في تحلل أغشية الخلايا وإذابة البروتينات دون تغيير طبيعتها.
من الناحية المفاهيمية، يمثل One-Tip أبسط سير عمل للبروتيوميات، حيث لا يتطلب أي معالجة إضافية للعينة ويجمع مباشرة بين إعداد العينة وتحليل LC-MS دون أي فقدان أو تباين في العينة. تؤدي أعداد الخلايا المنخفضة إلى تغطية عالية للبروتيوم وتتجنب الحاجة إلى الصوتنة وتبادلات العازلات الإضافية. وهذا يؤدي إلى وقت إجمالي لإعداد العينة يبلغ حوالي 70 دقيقة، بما في ذلك وقت الهضم الذي يبلغ 60 دقيقة. علاوة على ذلك، فإن سير عمل One-Tip ليس محدودًا في الإنتاجية؛ حتى المعالجة اليدوية يمكن أن تعد بكفاءة آلاف العينات يوميًا باستخدام pipetting متعدد القنوات، كما أنه سهل الأتمتة على روبوتات معالجة السوائل. أخيرًا، نقوم بربط سير عمل One-Tip مع نظام LC-MS/MS من الطراز الأول لضمان أداء تحليلي متفوق. يعتمد ذلك على طريقة Whisper LC الحساسة للغاية على جهاز Evosep One LC وطريقتنا الجديدة لتقنية الاستحواذ المستقل عن البيانات ذات النوافذ الضيقة (nDIA) على مطياف الكتلة Orbitrap Astral..
في التجارب الأولية، قمنا بتقييم نسب مختلفة من المزيج الرئيسي إلى تعليق خلايا هيلا المفردة في PBS، واختبرنا تركيبات مثلخليط رئيسي خلايا، ، و ، باستخدام منصة Orbitrap MS من الجيل السابق، جهاز Orbitrap Exploris 480 (الشكل 1ب). تم تقدير تركيز الخلايا بـ 100 خلية1. كشفت نتائجنا أن حتى أصغر نسبة،أسفر عن تحديد 3172 مجموعة بروتينية ملحوظة (يشار إليها فيما بعد بالبروتين). هذه التغطية البروتينية Comparable لعدد البروتينات التي تم تحديدها في عينات HeLa بتركيز 10 نانوغرام تم إعدادها باستخدام بروتوكول التقاط تجميع البروتين (PAC).، مرجع قياسي. من الجدير بالذكر أن الفرق في الأداء الذي لوحظ بين جهاز Orbitrap Exploris 480 وجهاز Orbitrap Astral عند تحليل عينات One-Tip يعكس الفروق التي تم رؤيتها مع عينات HeLa القياسية. تشير هذه الاتساق إلى أن بروتوكول OneTip لدينا لا يظهر أي انحياز تجاه منصات مطياف الكتلة المحددة.
لاختبار حساسية وإعادة إنتاجية منهجية One-Tip بشكل أكبر، قمنا بتطبيقها على جهاز Orbitrap Astral باستخدام تحليل nDIA مع زيادة عدد خلايا HeLa، حيث تم تحليل 20 و100 و200.
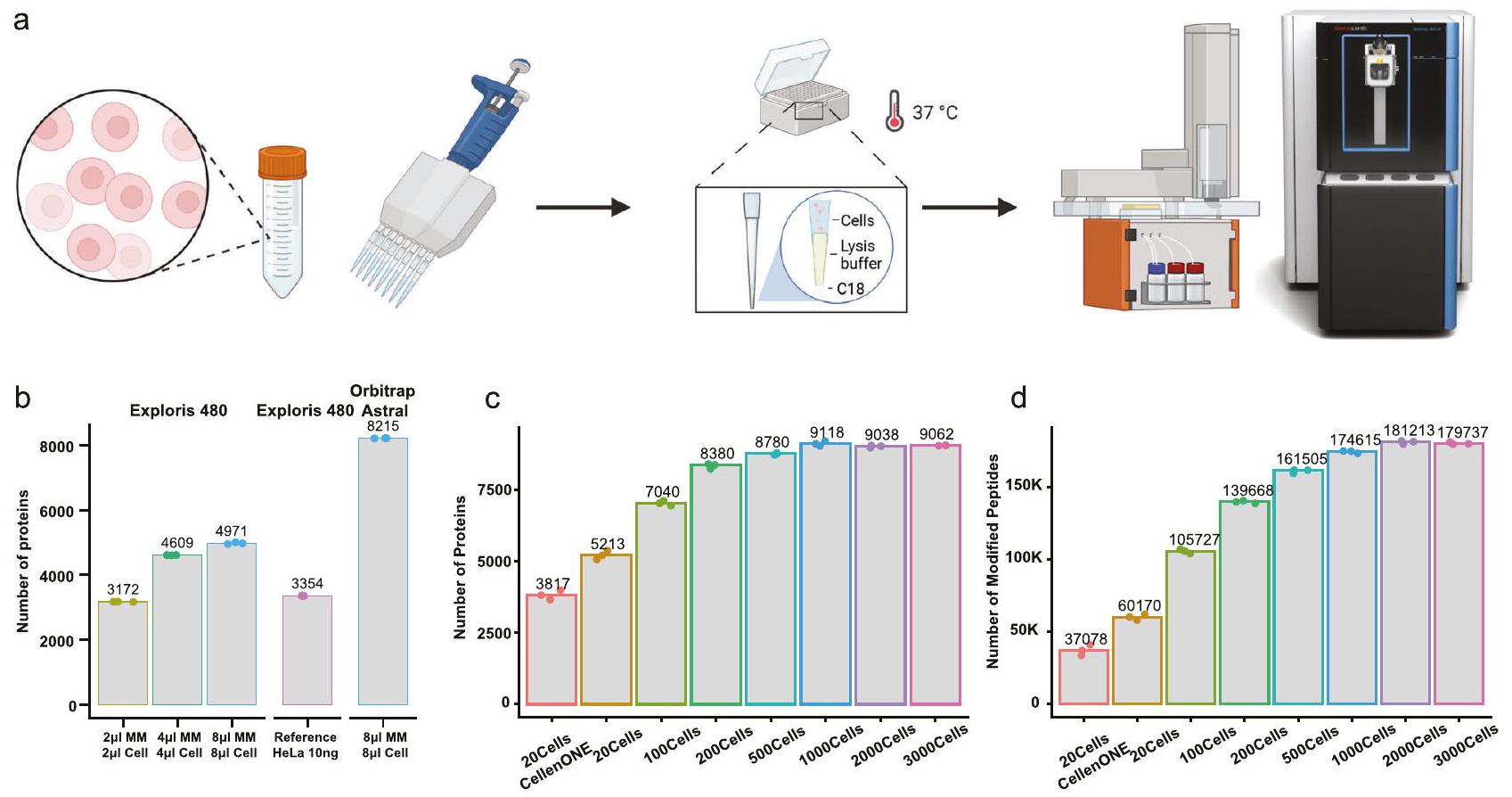
الشكل 1 | تقنية One-tip تحقق عمق بروتيني شبه كامل في التحليل الأحادي. أ رسم بياني يوضح سير عمل One-Tip.عدد البروتينات المحددة مع والبتيدات (د) المحددة في عدد مختلف من الخلايا. كانت مدة الهضم ساعتين في هذه العينات. تم توفير بيانات المصدر كملف بيانات مصدر. أحجام مختلفة من المزيج الرئيسي والخلايا في PBS. ج، د عدد البروتينات (ج)
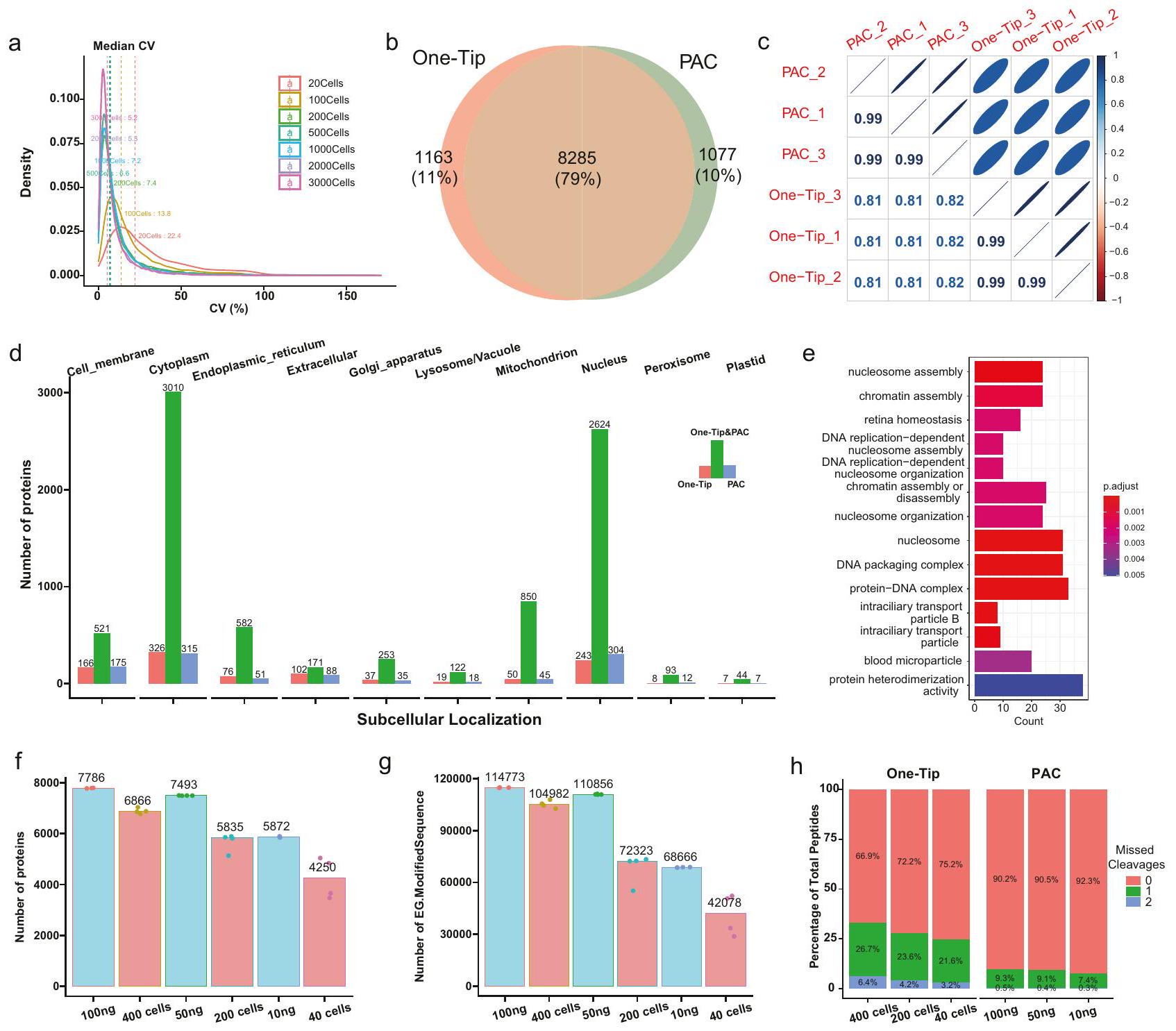
الشكل 2 | One-Tip يظهر قياسًا دقيقًا وتغطية بروتينية قابلة للمقارنة مع PAC. أ توزيع معامل التباين بين النسخ الثلاثة في عدد مختلف من الخلايا.تداخل البروتينات المحددة في سير العمل One-Tip (1000 خلية) و PAC (1000 نانوغرام). ج ارتباط وفرة البروتينات بين عينات OneTip و PAC. د الموقع الفرعي للبروتينات المحددة في عينات One-Tip و PAC. هـ إثراء تمثيل GO للبروتينات المحددة فقط. في عينات One-Tip. تم حساب قيم P باستخدام اختبار فيشر الدقيق أحادي الجانب وتم تعديلها لاحقًا باستخدام طريقة تصحيح بنجاميني-هوشبرغ (BH).عدد البروتينات ) والببتيدات ( تم التعرف عليه في عدد مختلف من الخلايا باستخدام One-Tip وكميات تحميل مختلفة من الببتيدات باستخدام PAC.نسبة الانقسامات المفقودة في عينات One-Tip و PAC. تم توفير بيانات المصدر كملف بيانات مصدر.
500 و 1000 و 2000 و 3000 خلية، على التوالي. لاحظ أن كميات الخلايا هذه كانت مستندة إلى تقديرات عد الخلايا وبالتالي فهي تقريبات. وحجمقد لا يكون المزيج الرئيسي في هذه التجربة مثاليًا لعدد الخلايا المختلفة. أظهر One-Tip عمقًا مثيرًا للإعجاب في البروتيوم لتحليل اللقطة الواحدة في جولات LC-MS/MS التي تستغرق نصف ساعة، حيث تم تحديد أكثر من 5000 مجموعة بروتينية وأكثر من 46000 ببتيد معدل (المشار إليه فيما بعد بالببتيدات) من حوالي 20 خلية (الشكل 1c). تتجاوز هذه الأداء تغطية البروتيوم التي تم تحقيقها حاليًا من خلال عزل 20 خلية بدقة باستخدام CellenONE، وهو نظام إعداد بروتيوميات الخلايا المفردة المخصص، والذي من المحتمل أن يكون مرتبطًا بفقدان العينات في لوحة 96 بئرًا ونقلها إلى نظام LC-MS. علاوة على ذلك، فإن الكمية الابتدائية منغطت الخلايا تقريبًا البروتينوم الكامل، مع تحديد أكثر من 9000 بروتين و170,000 ببتيد (الشكل 1c، d). بعد ذلك، قمنا بتقييم كفاءة هضم الإنزيمات البروتينية Lys-C والتربسين من خلال تغيير وقت الهضم من 1 إلى 4 ساعات. حتى وقت الهضم البروتيني القصير الذي يبلغ ساعة واحدة كان كافيًا لـ هضم البروتينوم بالكامل بشكل فعال، مما يؤدي إلى تغطية تسلسلية عالية مع معدل انقسام مفقود أقل منفي 20 خلية وحواليلكميات خلايا أكبر (الأشكال التكميلية 1-3).
يظهر One-Tip قياسًا دقيقًا وتغطية بروتينية قابلة للمقارنة مع PAC باستخدام 1000 خلية
أظهر One-Tip قابلية إعادة إنتاج عالية ودقة كمية، مع معامل تباين ملحوظ منفي تحليل النسخ المتماثلة لسير العمل مع ارتباط بيرسون يتجاوز 0.99 لـ maxLFQتقدير البروتين القائم على العينات التي تحتوي على أكثر من 100 خلية (الشكل 2أ، الشكل التوضيحي 4). على الرغم من سير العمل البسيط والسريع، فإن الأداء التحليلي لطريقة One-Tip مع ألف خلية قابل للمقارنة مع عينات البروتيوميات الكلية التي تم إعدادها باستخدام طريقة أكثر تعقيدًا، وهي طريقة التقاط تجميع البروتين (PAC).عند دمجه مع طريقة DIA ذات النافذة الضيقة لدينا على مطياف الكتلة Orbitrap Astral. بالمقارنة مع 1000 نانوغرام من lysate HeLa المحلل بواسطة LC-MS/MS فيطريقة النصيحة الواحدة المطبقة على
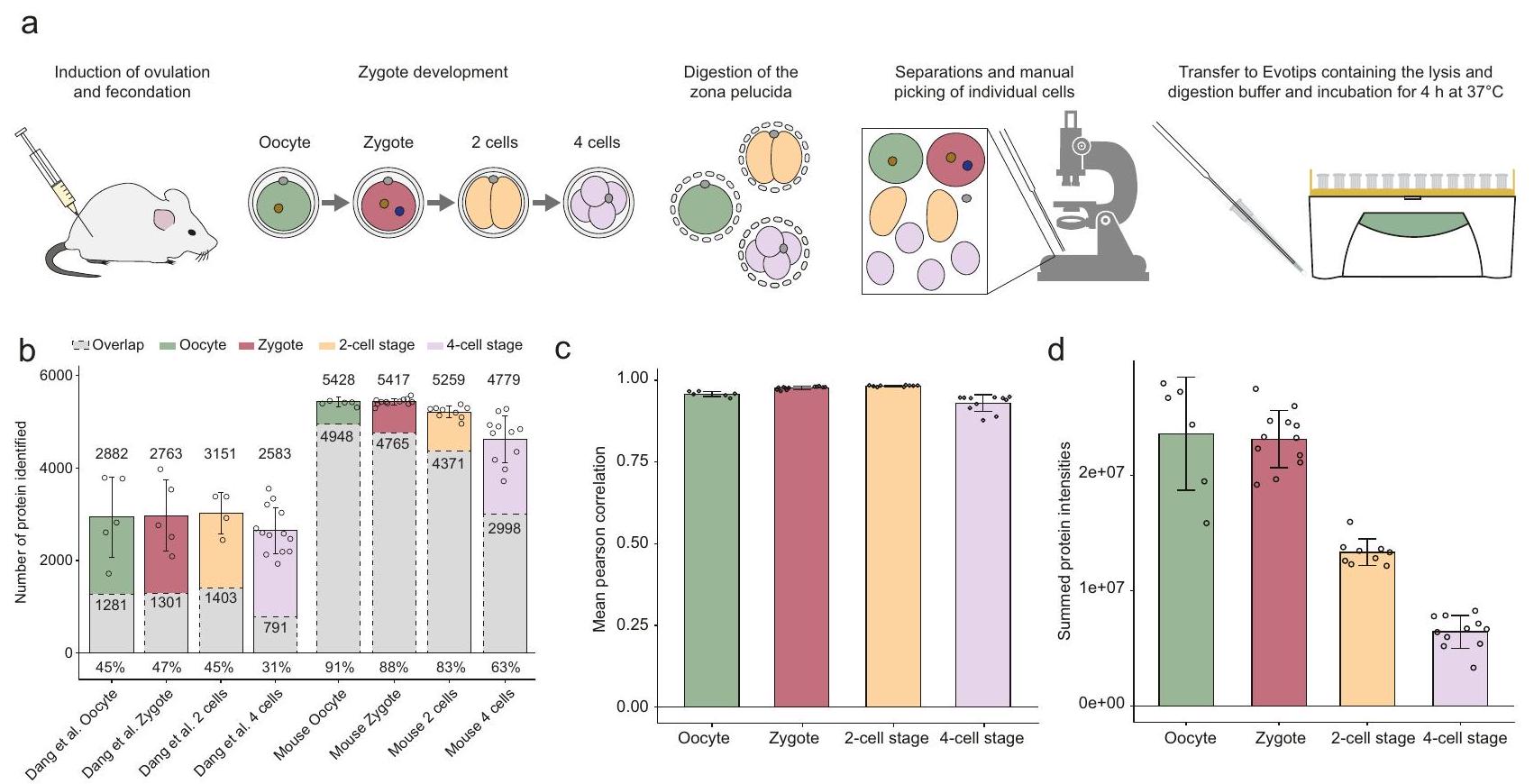
الشكل 3| تحليل One-Tip و LC-MS لجنين الفأر قبل الزرع حتى مرحلة 4 خلايا. أ رسم بياني يوضح سير العمل باستخدام One-Tip لجنين الفأر قبل الزرع.عدد البروتينات ذات القيم الكمية في الدراسة من دانغ وآخرون ودراستنا، وتداخل البروتينات بين العينات (باللون الرمادي) من نفس مراحل التطور. العدد المتوسط للبروتينات لكل حالة، والتداخل والنسبة المئوية المتوسطة للتداخل مقارنة بكل عينة هي مؤشر. ج متوسط الارتباط لكميات البروتين المنضبطة بين العينات داخل كل مجموعة عينات (البويضة، الزيغوت، مرحلة الخلية الثنائية، مرحلة الخلية الرباعية). د مجموع شدة البروتينات لكل مجموعة من العينات. تمثل أشرطة الخطأالانحراف المعياري للمتوسط. عدد النسخ البيولوجية:للبويضة،للكيسة الملقحة،لمرحلة الخلية الثنائيةلمرحلة 4 خلايا. يتم توفير بيانات المصدر كملف بيانات المصدر. عينة الخلايا في تشغيل LC-MS/MS لمدة نصف ساعة حددت عددًا مشابهًا من البروتينات مع تداخل كبير بينها ودقة قياس عالية جدًا (الشكل 2ب، ج). كانت نسبة ارتباط بيرسون الأقل منمن المحتمل أن تكون الفروق في كميات البروتين بين عينات PAC و One-Tip ناتجة عن الاختلاف في معدل الألكلة بالسيستين ومعدلات الانقضاض المفقودة (الشكل التوضيحي 5). ومن المثير للاهتمام أن كل من طريقتي PAC و One-Tip حددتا أكثر من 1000 بروتين فريد قليلاً، ولكن لم تظهر أي انحياز كبير في أنطولوجيا البروتين نحو مواقع فرعية خلوية محددة (الشكل 2d)، ووجود تفضيل طفيف لوظائف بيولوجية معينة مثل تجميع النوكليوسوم في البروتينات التي تم تحديدها فقط في عينات One-Tip (الشكل 2e).
أخيرًا، لتقييم فقدان المادة أثناء تحضير One-Tip، قمنا بمقارنة سلسلة تخفيف من هضم HeLa PAC من 100 نانوغرام إلى 10 نانوغرام، بدءًا من الببتيدات التي تم إزالة الملح عنها والتي تم نقلها إلى Evotips للحقن، إلى سلسلة تخفيف من الخلايا السليمة التي تم فرزها باستخدام cellenONE والتي تت correspond إلى 400 و200 و40 خلية HeLa، والتي تمت معالجتها باستخدام One-Tip وبالتالي تم تحللها فقط في Evotip. كمية البروتين في الخلايا تت correspond تقريبًا إلى 100 و50 و10 نانوغرام من المادة الأصلية، بافتراضكفاءة التحلل والاسترداد. من المهم ملاحظة أن عدد الخلايا التي تم فرزها بواسطة cellenONE يمثل الحد الأقصى لعدد الخلايا المحتمل وجودها في كل عينة، ولكن من المحتمل أن النقل أدى إلى فقدان الخلايا. ومع ذلك، فإن هذا يمثل المقارنة الأكثر عدلاً بين الطريقتين الممكنتين مع أدواتنا الحالية مع عيب لطريقة One-Tip. ثم قمنا بمقارنة عدد البروتينات والببتيدات المحددة في كل عينة (الشكل 2f، g). بشكل عام، كانت أعداد البروتينات والببتيدات في العينات من cellenONE قريبة من مكافئ في تحميل الببتيد أو أقل قليلاً (أي 400 خلية كانت قريبة من 50 نانوغرام). كانت التباينات أعلى في أعداد الخلايا المنخفضة، وهو ما يُحتمل أن يكون بسبب فقدان الخلايا أثناء النقل إلى الأطراف. أخيرًا، كانت نسبة الانشقاقات المفقودة أعلى في عينات One-Tip مقارنةً بالعينات من هضم PAC (الشكل 2h).
تمكن تقنية One-Tip من تحليل بروتينات الخلايا المفردة في الأجنة الفأرية قبل الزرع
لإظهار تعددية وحساسية سير عمل One-Tip، درسنا بعد ذلك تطور الأجنة الفأرية قبل الزرع على مستوى الخلية الواحدة من البويضة والزيغوت إلى مرحلة الخلية 4. تم إزالة المنطقة الشفافة، وهي الغلاف الجليكوبروتيني الخارجي للبويضات والزيغوت ومرحلتين من الخلايا 2 و4، كيميائيًا وتم فصل الخلايا الفردية تحت المجهر وتوزيعها بشكل فردي في Evotips، المحملة مسبقًا بمحلول تحلل الخلايا ومحلول الهضم (الشكل 3أ). تم تنفيذ هذه العملية بالكامل في يوم واحد لكل دفعة من العينات بما في ذلك تحليل LC-MS/MS. في المجموع، تم تحديد 6216 مجموعة بروتينية بوسيط قدره، و4779، لكل مرحلة من مراحل التطور، على التوالي (الشكل 3ب). بالمقارنة مع التحليل الحديث جداً للأجنة البشرية قبل الزرع من دانغ وآخرون.في الدراسة التي استخدم فيها المؤلفون سير عمل متقدم لتحضير العينات وتدرجات LC-MS تستغرق 4 مرات أطول من تلك المستخدمة في هذه الدراسة، نحدد حوالي 2300 بروتين و حوالي 50000 ببتيد إضافي بالإضافة إلىمتوسط عدد الببتيدات لكل بروتين (5.3 مقابل 14.6)، لكل مرحلة (الشكل التوضيحي 5 أ، ب). علاوة على ذلك، تم تحسين التباين في عدد البروتينات، والتداخل (الشكل 3 ب)، وارتباط كميات البروتينات بين العينات من نفس الحالة بشكل كبير (الشكل 3 ج، الشكل التوضيحي 5 ج)، مما يدل على أن سير العمل One-Tip المرتبط بـ Evosep One و Orbitrap Astral أسرع، وأكثر حساسية، وأكثر تكرارية ولديه دقة أفضل في الكمية. من المثير للاهتمام أن كثافات البروتينات المجمعة في كل عينة كانت متساوية بين البويضات والزيجوت، بينما كانت نصف تلك الكثافات في خلايا من مرحلة الخلية 2 وربعها في خلايا من مرحلة الخلية 4، مما يتماشى مع تقليل حجم الخلية خلال الانقسامين المتتاليين للزيجوت (الشكل 3 د). على الرغم من أن تحليلنا استفاد من استخدام أحدث جيل من مطياف الكتلة مقارنة بجيل أقدم في دراسة دانغ وآخرون، والتي قد تضخم هذه الأرقام، فإن دراستنا تبرز أن استخدامنا
الشكل 4 | رؤى بيولوجية من الأجنة الفأرية قبل الزرع.
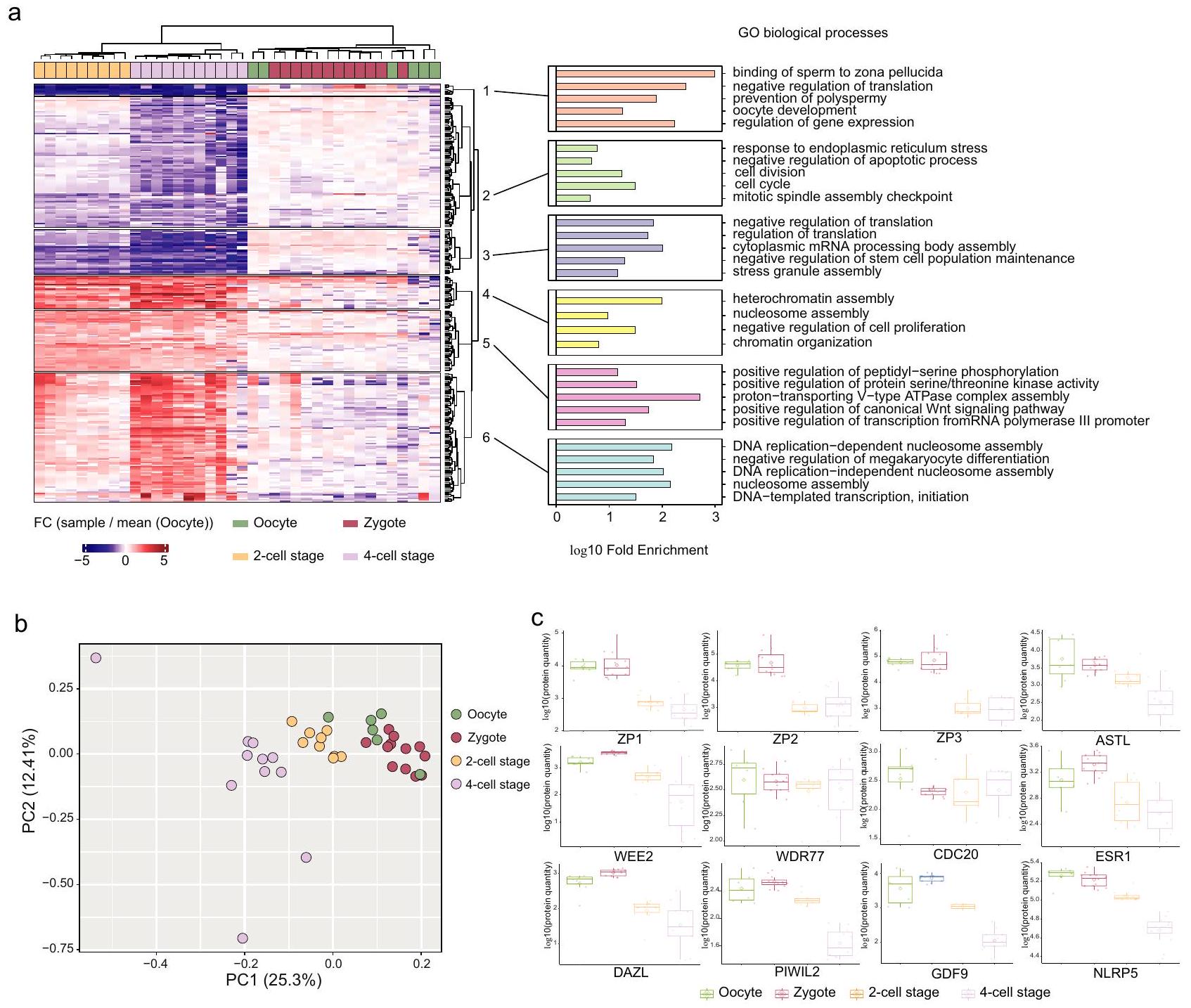
تجميع هرمي غير مشرف باستخدام طرق كانبيرا وورد. د2 للبروتينات التي تظهر تنظيمًا كبيرًا ( من ANOVA أحادي الجانب) و FC مضاعف في أي من مراحل التطور مقارنة بمجموعة البويضات (يسار). تحليل إثراء العمليات البيولوجية GO لمجموعات البروتينات من خريطة الحرارة (يمين). عدد النسخ البيولوجية: للبويضة، للزيجوت، لمرحلة الخلية 2 و لمرحلة الخلية 4. تحليل المكونات الرئيسية لكثافات البروتينات المعيارية لكل عينات دراستنا. تم اعتبار فقط البروتينات التي تم قياسها في جميع
العينات ( ). ج وفرة اثني عشر بروتينًا مرتبطًا بالعمليات التناسلية عبر أربع مراحل تطورية. بالنسبة لمخططات الصندوق، فإن الحواف السفلية والعلوية تتوافق مع الربع الأول والثالث. يمتد الشوكة العلوية من الحافة إلى أكبر قيمة لا تتجاوز IQR من الحافة (حيث IQR هو النطاق بين الربعين). تمتد الشوكة السفلية من الحافة إلى أصغر قيمة في أقصى IQR من الحافة. عدد النسخ البيولوجية: للبويضة، للزيجوت، لمرحلة الخلية 2 و لمرحلة الخلية 4. تم توفير بيانات المصدر كملف بيانات المصدر.
سير العمل البسيط One-Tip يمكّن من الحصول على مجموعات بيانات شاملة مع مواد عينة محدودة بشكل كبير.
أخيرًا، تحققنا مما إذا كان يمكن استرداد معلومات ذات دلالة بيولوجية من تحليل خلايا الأجنة الفأرية. ثم قمنا بإجراء تجميع هرمي غير مشرف للبروتينات المعبر عنها بشكل مختلف التي تم تنظيمها لأعلى أو لأسفل في أي من الحالات مقارنة بالبويضات (الشكل 4 أ). تم فصل كل مجموعة عينة على خريطة الحرارة بعيدًا عن الزيجوت والبويضات كما هو الحال في تحليل المكونات الرئيسية. يكشف تحليل إثراء علم الجينات (GO) للمجموعات عن تنظيم تنازلي للبروتينات المرتبطة بارتباط الحيوانات المنوية بالمنطقة الشفافة، ومنع تعدد الحيوانات المنوية، والاستجابة للإجهاد في الشبكة الإندوبلازمية وزيادة تنظيم البروتينات المرتبطة بتكرار الحمض النووي وتجميع الهيتروكروماتين في مراحل الخلية 2 و4 مقارنة بالبويضات، من بين مسارات أخرى. كانت هذه النتائج متوافقة بشكل عام مع دانغ وآخرون مما يثبت جودة تحليلنا (الشكل 4 أ). أظهر تحليل المكونات الرئيسية (PCA) لكثافات البروتينات المعيارية أن مجموعة البويضات والزيجوت تتجمع معًا وأن مراحل الخلية 2 و4 هي
مفصولة بطريقة زمنية على PC1 (الشكل 4 ب). ثم ركزنا على كميات اثني عشر بروتينًا مرتبطًا بالعمليات التناسلية عبر أربع مراحل تطورية (الشكل 4 ج). من الجدير بالذكر أن ZP3، وهو بروتين حاسم لتكوين المنطقة الشفافة، انخفض خلال مرحلة الخلية 2، متماشيًا مع الانقسام الخلوي الأول، لكنه استقر عند مرحلة الخلية 4، مما يشير إلى تغييرات ديناميكية في التطور الجنيني المبكر. في المقابل، ظلت كميات WDR77، المرتبطة بتنظيم النسخ، و CDC20، الضرورية لعمليات انقسام الخلايا، مستقرة عبر جميع المراحل. تشير هذه الثبات إلى نشاط انقسام ونسخ مستمر من مرحلة الزيجوت، مما يبرز أدوارها الحاسمة في التطور الجنيني المبكر.
توسيع One-Tip إلى خلايا HeLa الفردية مع موزع الخلايا الفردية
مؤخراً، تم وصف موزع الخلايا الفردية Uno (المكافئ لموزع الخلايا الفردية HP D100) لفرز الخلايا الفردية لتحليل البروتيوميات . نظرًا لأن تصميم الأداة يسمح
أ
الشكل 5 | توسيع One-Tip إلى خلايا HeLa الفردية مع موزع الخلايا الفردية.
سير عمل البروتيوميات للخلايا الفردية باستخدام One-Tip وموزع الخلايا الفردية Uno. ب، ج عدد البروتينات (ب) والببتيدات (ج) المحددة في ظروف إعداد عينات مختلفة.
عدد العينات: 0and2_pipette،
0and2_sorter، land2_pipette، 1and2_sorter، carrier. تم توفير بيانات المصدر كملف بيانات المصدر.
من خلال ملاءمة صندوق من Evotips، قمنا بتقييم ما إذا كان سير العمل One-Tip يمكن أن يكون مناسبًا لتحليل الخلايا الفردية واختبرنا إعدادات تجريبية متعددة (الشكل 5 أ). باختصار، إما وضعنا من محلول التحلل والهضم أو لا شيء، قمنا بفرز الخلايا ثم قمنا بتوزيع من محلول التحلل إما يدويًا (بpipette) أو باستخدام الموزع. تظهر النتائج أن تحليل الخلايا الفردية ممكن باستخدام موزع الخلايا الفردية Uno المرتبط بـ One-Tip مع أكثر من 2000 بروتين تم قياسه في المتوسط لأفضل حالة يدويًا) بدون عينة حاملة أو مرجعية مع إشارة أعلى لتعزيز التعرفات أثناء التحليل باستخدام Spectronaut وأكثر من 3000 مع عينة حاملة (الشكل 5 ب، ج). كانت بعض النصائح تحتوي على إشارة قليلة مما يشير إلى أن الخلايا لم تصل إلى قاع Evotips ولم يتم تحللها وهضمها بشكل صحيح. ومع ذلك، تم الكشف عن إشارة متسقة في ما يصل إلى من العينات، حيث كان من المفترض أن تكون الخلايا الفردية موجودة وفقًا لبرنامج الأداة، وهو أقل مما تم الإبلاغ عنه من قبل سانشيز-أفيلا وآخرون. باستخدام لوحة PCR من 384 بئر. بينما كان عدد الببتيدات والبروتينات المحددة والمقاسة أقل من تحليل SCP باستخدام أداة cellenONE ونفس LC-MS ، فإن استخدام مجموعة موزع الخلايا الفردية Uno و One-Tip هو بديل اقتصادي وبسيط لبعض سير العمل الأكثر تعقيدًا وتكلفة المستخدمة حاليًا. أخيرًا، فإن سير العمل المقدم هنا لديه مجال للتحسين سواء من حيث نسبة الخلايا الفردية المستردة أو من حيث أعداد الببتيدات والبروتينات، ويجب تطوير ذلك بشكل أكبر في المستقبل.
سير عمل عالي الحساسية للجزئيات خارج الخلوية باستخدام One-Tip لتسليط الضوء على التطبيق الواسع والحساسية لسير العمل One-Tip، قمنا بمعالجة الجزئيات خارج الخلوية (EV)
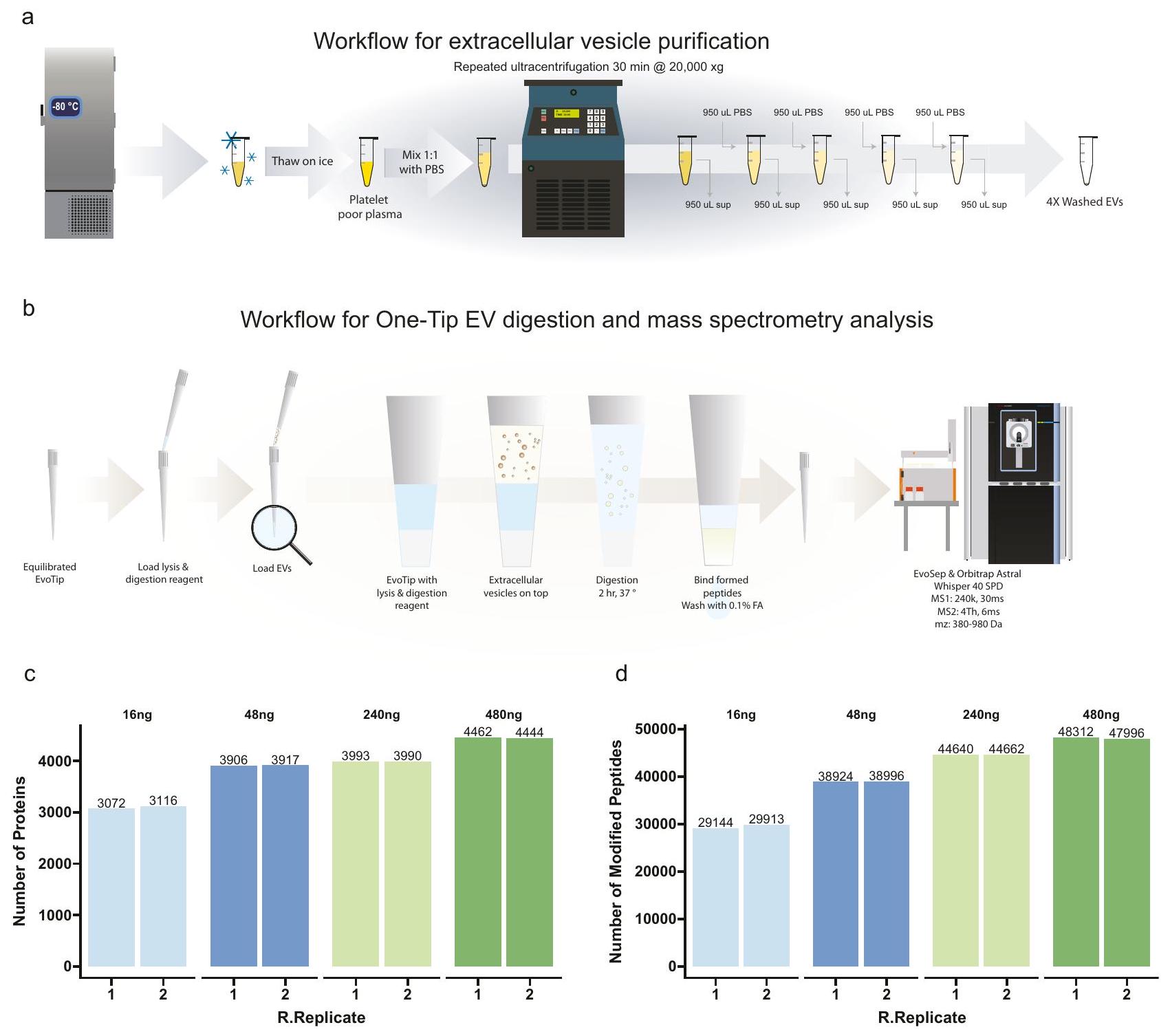
المستخرجة من بلازما الدم بدلاً من الخلايا السليمة. قمنا بعزل EVs من بلازما المتبرعين الذكور كما هو موصوف من قبل كفيرنلاند وآخرون. وقمنا بتحميل سلسلة تخفيف من 480 نانوغرام إلى 16 نانوغرام (مقدرة بناءً على تركيز البروتين في البلازما أثناء تنقية EV) (الشكل 6 أ) على Evotips المعدة لتحليل One-Tip (الشكل 6 ب). قمنا بقياس أكثر من 3000 بروتين و29000 ببتيد مع 16 نانوغرام من المادة الأولية وقريب من 4500 بروتين و48000 ببتيد مع 480 نانوغرام، والأخير يتماشى مع أحدث التقنيات في تحليل EV البلازما (الشكل 6 ج). بالإضافة إلى ذلك، فإن عدد البروتينات والببتيدات المحددة يتناسب مع كمية العينة المحملة، كما لوحظ مع بيانات الأجنة الفأرية. وبالتالي، تظهر هذه البيانات أن One-Tip يمكن تطبيقه بسهولة على أنواع أخرى من العينات بخلاف الخلايا الفردية أو تعليقات الخلايا، وسيمكن من تحليل أكثر حساسية، وسلاسة، وذو إنتاجية أعلى للـ EVs. هذا مهم بشكل خاص في مجال تحليل البلازما واكتشاف العلامات الحيوية وكذلك في مجال EV بشكل عام، حيث يمكن أن تكون كمية الـ EVs المشتقة من البلازما أو الخلايا محدودة.
نقاش
مجتمعة، تظهر نتائجنا أن العمق التحليلي الذي تم تحقيقه باستخدام سير العمل One-tip يتفوق على سير العمل القياسي في مجال البروتيوميات. إن بساطته وسهولة استخدامه وقابليته للتوسع تجعله مناسبًا للغاية لمجموعة متنوعة من الإعدادات التجريبية، بدءًا من تحليل الخلايا الفردية إلى البروتيوميات الجماعية. هذه الطريقة تقضي بشكل فعال على الحاجة إلى منهجيات معقدة، ومعدات محددة، وخبرة متخصصة في البروتيوميات خلال مرحلة إعداد العينة. ونتيجة لذلك، يسد سير العمل One-Tip الفجوة بشكل كبير بين البروتيوميات وعلم الأحياء، مما يبسط تحليلات LC-MS ويجعلها أكثر سهولة وتوافقًا مع
الشكل 6 | تحليل أحادي النصائح للحويصلات خارج الخلوية. أ سير العمل لتنقية الحويصلات خارج الخلوية.سير العمل لتحليل هضم EV باستخدام تقنية One-Tip وقياس الطيف الكتلي.عدد البروتينات (ب) والببتيدات (ج) التي تم تحديدها في كميات تحميل مختلفة.تكرارات فنية لكل حالة. يتم توفير بيانات المصدر كملف بيانات مصدر.
مجموعة أوسع من التطبيقات البيولوجية. يمكن تعديل سير العمل ليتناسب مع منصات مثل cellenONE و Uno Single Cell Dispenser.، مما يعزز من تعدد استخداماته.
بالإضافة إلى ذلك، فإن One-Tip قادر على تحليل مجموعة متنوعة من أنواع العينات، بما في ذلك الخلايا المصنفة بواسطة FACS، والأجنة البشرية قبل الزرع، ومجموعات الخلايا النادرة، وشرائح الأنسجة. هذا الأمر ذو صلة خاصة لأنواع الخلايا الأصغر، مثل خلايا T، حيث من المتوقع تحديد عدد أقل من البروتينات بما يتماشى مع محتوى البروتين (حوالي 25 بيكوجرام لكل خلية). بالنسبة لمثل هذه الخلايا، قد تقدم التحليلات على مستوى الخلية الواحدة رؤى بيولوجية محدودة، ويقدم سير عمل One-Tip بديلاً فعالاً. علاوة على ذلك، نوضح قابلية تطبيق One-Tip على العينات غير الخلوية، مثل الحويصلات خارج الخلوية، مما يبرز فائدته في مجموعة واسعة من سياقات البحث. تجعل هذه القابلية One-Tip أداة قيمة في ترسانة البروتيوميات الحديثة، مما يمكّن من نهج أكثر شمولاً لتحليل العينات ويسهل فهمًا أعمق لمختلف الأنظمة البيولوجية.
طرق
تتوافق هذه الدراسة مع جميع اللوائح الأخلاقية ذات الصلة. تم إجراء العمل على الحيوانات وفقًا للرخصة رقم 2021-15-0201-00851، المعتمدة. بواسطة هيئة التفتيش الوطنية على تجارب الحيوانات الدنماركية، وتمت وفقًا للإرشادات الوطنية والمحلية.
خطوط خلايا هيلا
تم زراعة خلايا سرطان عنق الرحم البشرية HeLa (ATCC CCL-2) في DMEM (Gibco، Invitrogen)، معززة بـمصل الجنين البقريالبنسلين (إنفيتروجن)ستربتوميسين (إنفيتروجن)، عندفي حاضنة مرطبة مع. في تم فصل الخلايا باستخدام التربسين وغسلها مرتين بمحلول فوسفات البفر (PBS) من جيبكو (لايف تكنولوجيز)، قبل إعادة تعليقها في PBS منزوع الغاز.
تدفق عمل إعداد العينة بتفصيل نصيحة واحدة
حدد تركيز خلايا هيلا باستخدام عداد الخلايا وقم بتخفيفها إلى التركيزات المطلوبة باستخدام PBS.
قم بإعداد Evotips وفقًا لتعليمات البائع:
اشطف: اغسل Evotips الجافة بـمن المذيب ب (FA في الأسيتونيتريل) ويدور في جهاز الطرد المركزي بسرعة 800 جرام لمدة 60 ثانية.
الحالة: انقع الإيفوتيبس فيمن 1-بروبانول حتى يتحولوا إلى اللون الأبيض الباهت.
توازن: اشبع Evotips المعالجة بـمن المذيب A (في الماء) ويدور في جهاز الطرد المركزي بسرعة 800 جرام لمدة 60 ثانية.
ماصةمن محلول التحلل والهضم في الإيفوتيبس. يحتوي المحلول على-دوديسيل--دي-مالتوسيد (DDM)، 100 مليمول TEAB،تريبسين، وليس-سي.
ماصةالخلايا في إيفوتيبس.
قم بتدوير الإيفوتيبس بسرعة عند 50 ج لتخلط العازلة والخلايا وتمنع تكون فقاعات الهواء.
أضف الماء إلى صندوق إيفوتيب حتى مستوى راتنج C18 في الإيفوتيب.
قم بحضانة صندوق إيفوتيب عندمن 1 إلى 4 ساعات.
تابع تعليمات البائع مع تعديل طفيف:
تحميل: إضافةمن المذيب A إلى الإيفوتيبس وcentrifuge لمدة 60 ثانية عند 800 ج.
اغسل: اشطف الأطراف بـمن المذيب A ويدور في جهاز الطرد المركزي لمدة 60 ثانية عند 800 ج.
رطب: أضفمن المذيب A إلى الأطراف وcentrifuge لمدة 10 ثوانٍ عند 800 ج للحفاظ على رطوبة الأطراف.
عزل خلايا هيلا باستخدام CellenONE
تم تخفيف خلايا هيلا بمحلول PBS منزوع الغاز إلىخلاياتم عزل الخلايا في لوحة 96 بئر محملة مسبقًا بـتم استخدام PBS مع نظام CellenONE. بعد العزل، تم إعداد العينات وفقًا لعملية One-Tip.
عزل بويضات الفئران، الزيجوت، مراحل الخلية 2 والخلية 4
تم إجراء العمل على الحيوانات وفقًا للرخصة رقم 2021-15-020100851، المعتمدة من قبل هيئة التفتيش الوطنية لتجارب الحيوانات الدنماركية، وتم تنفيذه وفقًا للإرشادات الوطنية والمحلية. تم الاحتفاظ بالفئران في غرف مخصصة في أقفاص ذات تهوية فردية عند درجة حرارة، مع رطوبة قدرهاتم تغيير الهواء في الغرفة من ثماني إلى عشر مرات في الساعة ودورة الظلام/الضوء هيالإضاءة من الساعة 6 صباحًا حتى 6 مساءً. يتم تربية الفئران وفقًا للوائح الدنماركية للتجارب الحيوانية.
تم تحفيز الإباضة لدى إناث C57BL/6NRj قبل البلوغ (عمر 4 أسابيع) (mus musculus) عن طريق حقن داخل البطن (IP) بـ PMSG (HOR-272، Prospec) بجرعة 5 IU/أنثى، تلاها حقن داخل البطن بـ hCG (Chorulon Vet، Pharmacy) بجرعة 5 IU/أنثى بعد 47 ساعة. بعد الحقن الثاني، تم وضع الإناث في تزاوج مع ذكور C57BL/6NRj. في صباح اليوم التالي، تم euthanizing الإناث وتم تشريح قناتي فالوب لجمع الكومولي التي تحتوي على الزيغوتات والبويضات غير المخصبة. تم تفكيك الكومولي عن طريق الحضانة لمدة 10 دقائق في الهيالورونيداز (H4272، Sigma-Aldrich). تم فرز الزيغوتات وتقييمها من خلال وجود الجسم القطبي الثاني وزراعتها في وسط KSOM (MR-106-D، Merc Millipore). في يوم الحصاد، تم معالجة مجموعة من الزيغوتات والبويضات للتحليل، بينما تم حضانة زيغوتات أخرى في KSOM في و لمدة 24 ساعة أو 48 ساعة. في مراحل التطور المختلفة، تم حضانة البويضات والأجنة في محلول تايرود الحمضي (T1788، سيغما-ألدريش) لمدة 5 إلى 15 ثانية لإزالة الغلاف الشفاف. تم غسل البويضات العارية والزيغوت في PBS (20012-027، لايف تكنولوجيز) ونقلها بشكل فردي إلى إيفوتيبس باستخدام كابيلاري زجاجي بقطر. تم تفكيك الأجنة ذات الخلايا 2 و 4 بشكل إضافي عن طريق الحضانة لمدة 5 دقائق في و -خالي من KSOM والأنابيب معقطر الشعيرات الزجاجية في PBS بعد ذلك؛ تم نقل الكتل الفردية بعد ذلك إلى Evotips. كانت Evotips محملة مسبقًا بـمن محلول التحلل والهضم ومن PBS. كانت مدة الحضانة 4 ساعات. عدد النسخ البيولوجية:للبويضة،للزيغوت،لمرحلة الخلية 2 –لمرحلة 4 خلايا. تم استخدام ما مجموعه 20 أنثى متبرعة و8 ذكور للتزاوج في هذه التجارب.
عزل الخلايا باستخدام جهاز توزيع الخلايا الفردية Uno
تم إعداد الخلايا كما هو الحال في الفرز باستخدام CellenOne وتم تمريرها عبرتم قطع الشبكة لضمان عدم وجود كتل قبل الفرز. تم إعداد Evotips لتحليل One-Tip كما كما هو موصوف أعلاه وفي الشكل 5a حتى الخطوة التي يتم فيها إضافة محلول التحلل. نظرًا لأن محلول التحلل مخففمع تعليق الخلايا في PBS في تجربة One-Tip العادية، قمنا بتخفيف محلول التحلل بنسبة 1:1 مع PBS. ثم إماتمت الإضافة يدويًا تليها الطرد المركزي عند 50 جرام لمدة 10 ثوانٍ أو لم يتم إضافة أي محلول تحلل. بعد ذلك، تمت إضافة الماء إلى صندوق Evotip لتغطية طرف Evotips فقط وتم وضع الصندوق على جهاز Uno Single Cell Dispenser.لتوزيع الخلايا المفردة.تم إضافة تعليق الخلايا إلى كاسيت C1 وتم توزيع خلايا فردية في Evotips. بعد توزيع الخلايا،تم إضافة من محلول التحلل المخفف إما عن طريق السحب أو باستخدام الموزع باستخدام كاسيت D1. تمت إزالة الماء وتم طرد النصائح في جهاز الطرد المركزي بسرعة 50 ج لمدة 10 ثوانٍ بحيث يكون محلول التحلل في اتصال مع مرحلة Evotip C18. تمت إضافة الماء مرة أخرى لتغطية قاع الصندوق وتم حضنه فيلمدة ساعتين. تبعت بقية إعداد العينة سير العمل One-Tip. كان عدد العينات هو:أو أند 2 بيبت0and2_sorter،land2_pipette،1and2_sorter، ن = 3 ناقل.
تحضير السيارة الكهربائية
تم جمع عينات الدم الوريدي في أنابيب سيترات (Vacuette، 3.2% سيترات الصوديوم، Greiner Bio-One) من متبرعين ذكور وتم طردها مركزيًا عندلمدة 10 دقائق خلال 30 دقيقة بعد جمع الدم. تم نقل الطور العلوي (البلازما الغنية بالصفائح الدموية) إلى أنبوب جديد وخضع لعملية طرد مركزي ثانية عند 3000 جرام لمدة 10 دقائق لترسيب الصفائح الدموية المتبقية، وتم تقسيم الطور العلوي الفقير بالصفائح الدموية، وتجميده بسرعة، وتخزينه فيفي يوم عزل الحويصلات خارج الخلية، تم إذابة البلازما الفقيرة من الصفائح الدموية على الثلج الرطب ومعالجتها لتكثيف الحويصلات خارج الخلية كما هو موضح في الشكل 6a. باختصار،تم تخفيف البلازما بنسبة 1:1 مع PBS pH 7.2 (Gibco cat# 20012-019) وتم الطرد المركزي عند لمدة 30 دقيقة.تم إزالة السائل العلوي و من PBS تم إضافته فوق الكريات (تخفيف)، يليه الطرد المركزي عندلمدة 30 دقيقة. تم إجراء هذه العملية 4 مرات في المجموع وبعد آخر عملية طرد مركزيتمت إزالة السائل العلوي المتبقيتمت معالجة المحتوى الذي يحتوي على حبيبات EV بشكل إضافي. تم تقدير محتوى البروتين في تحضير EV بناءً على تخفيف تركيز البروتين الكلي في البلازما. ) و 4 ميكروغرام من الحويصلات خارج الخلية (مع استرداد لـ بعد كل عملية طرد مركزي). تم تخفيف التحضيرات النهائية للـ EV بعد 4 عمليات طرد مركزي (3 غسلات) للحصول على كميات تعادلو 480 نانوغرام في. هذه تم إضافة كل تخفيف في نسختين إلى Evotips المعدة لتحليل OneTip التي تحتوي علىمن محلول التحلل وتمت معالجته وفقًا لبروتوكول One-Tip الموضح أعلاه، قبل تحليل LC-MS/MS. تم إجراء التجربة مرتين لكل حالة.
باك
تم إجراء عملية التقاط تجميع البروتين (PAC) كما تم وصفه سابقًا.باختصار، تم تحلل خلايا هيلا في SDS، 100 مللي مولار تريس pH تريس(2-كربوكسي إيثيل) فوسفين (TCEP)، و10 مم 2-كلوروأسيتاميد (CAA) وتم استخدام جهاز سونكيتور بروب برانسون لمدة دقيقة واحدة باستخدام نبضات لمدة 3 ثواني والسعة. تم قياس تركيز البروتين باستخدام اختبار BCA، وتم استخدام البروتينات لهضم PAC. تم إجراء هضم PAC على نظام KingFisher Flex (Apex, Duo Prime) (ثيرمو فيشر ساينتيفيك)، وتم ضبط حجم العينة إلى 300 مع محلول التحلل، ثممن MagReSynتم إضافة كريات الهيدروكسيل (Resyn Biosciences) بنسبة 2:1، كريات/بروتين، ) يتبعه منأسيتونيتريل، تم شطف الكريات 3 مرات بـأسيتونيتريل، ومرتين معتم إجراء هضم الإيثانول على الحبةفي 50 مللي مولار من بيكربونات الأمونيوم تحتوي على ليزيل إندوبيبتيداز (1:500، إنزيم/بروتين، وزن/وزن) وتريبسين (1:250، إنزيم/بروتين، وزن/وزن). تم تحميض العينات باستخدام حمض الفورميك إلىوتم تحميله على Evotips قبل تحليل LC-MS/MS.
LC-MS/MS
تم إجراء تحليل LC-MS/MS على جهاز Orbitrap Astral MS باستخدام برنامج Thermo Tune (الإصدار 0.4 أو أعلى) متصل بجهاز Evosep.
نظام واحد (EvoSep Biosystems). تم تحليل العينات في 40SPD (تدرج 31 دقيقة) باستخدام عمود تحليلي تجاري (Aurora Elite TS، lonOpticks) متصل عبر الإنترنت باستخدام EASY-Spray.المصدر. تم تشغيل جهاز Orbitrap Astral MS بدقة MS كاملة تبلغ 240,000 مع نطاق مسح كامل منعند الإشارة. تم تعيين MS AGC الكامل علىتم تسجيل مسحات MS/MS مع نافذة عزل 2Th، ووقت حقن الأيونات الأقصى (IIT) 3 مللي ثانية لعينات HeLa و4Th و6 مللي ثانية IIT لعينات الأجنة الفأرية، لسلسلة التخفيف من PAC وخلايا CellenONE المفروزة ولتحليل الخلية الواحدة باستخدام موزع Uno. كان نطاق مسح MS/MS منتم استخدام الأيونات المعزولة. تم تجزئة الأيونات باستخدام HCD مع 27% NCE.
تحليل بيانات MS
تم تحليل الملفات الخام في Spectronaut v18 (Biognosys) باستخدام نهج خالٍ من مكتبة الطيف (directDIA +) مع قاعدة بيانات البروتينات البشرية المرجعية (إصدار Uniprot 2022، 20,588 تسلسل) لعينات HeLa، وقاعدة بيانات البروتينات المرجعية للفئران (إصدار Uniprot 2022، 21,989 تسلسل) لخلية الجنين الفأري المفردة مكملة بالملوثات الشائعة (246 تسلسل). تجدر الإشارة إلى أنه نظرًا لأن البروتوكول لا يتضمن الاختزال والقلوية، تم إجراء عمليات البحث في قاعدة البيانات باستخدام ثيولات السيستين الحرة، وبالتالي لم يتم تعيين كرباميلات السيستين كتعديل ثابت، بينما تم تعيين أكسدة الميثيونين وأسيتيل بروتين N-termini كتعديلات متغيرة. تم تعيين تصفية السلفيتر لتتم بناءً على قيم Q، وتم التحقق من تطبيع التشغيل المتقاطع. تم تحليل كل تجربة بعدد مختلف من الخلايا بشكل منفصل، وتم البحث في العينات من أوقات هضم مختلفة مع وتمكين أو عدم تمكين تقييم الطريقة والإشارة إلى الظروف المختلفة (كل واحدة مع التكرارات التجريبية) في علامة إعداد الحالة.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
تم إيداع بيانات بروتيوميات مطيافية الكتلة الناتجة عن هذه الدراسة في اتحاد بروتيوم إكسشينج (http:// proteomecentral.proteomexchange.org) عبر مستودع الشركاء PRIDE مع معرف مجموعة البيانات PXD044991. تم توفير البيانات المصدرية مع هذه الورقة.
References
Zhang, Y., Fonslow, B. R., Shan, B., Baek, M.-C. & Yates, J. R. III Protein analysis by shotgun/bottom-up proteomics. Chem. Rev. 113, 2343-2394 (2013).
Kulak, N. A., Pichler, G., Paron, I., Nagaraj, N. & Mann, M. Minimal, encapsulated proteomic-sample processing applied to copynumber estimation in eukaryotic cells. Nat. Methods 11, 319-324 (2014).
Chen, W. et al. 3D-SISPROT: A simple and integrated spintip-based protein digestion and three-dimensional peptide fractionation technology for deep proteome profiling. J. Chromatogr. A 1498, 207-214 (2017).
Yu, Y. et al. Development of an efficient, effective, and economical technology for proteome analysis. Preprint at Research Square, https://doi.org/10.21203/rs.3.rs-3165690/v1 (2023).
Johnston, S. M. et al. Rapid, one-step sample processing for labelfree single-cell proteomics. J. Am. Soc. Mass Spectrom. 34, 1701-1707 (2023).
Matzinger, M., Müller, E., Dürnberger, G., Pichler, P. & Mechtler, K. Robust and Easy-to-Use One-Pot Workflow for Label-Free SingleCell Proteomics. Anal. Chem. 95, 4435-4445 (2023).
Petrosius, V. et al. Exploration of cell state heterogeneity using single-cell proteomics through sensitivity-tailored data-independent acquisition. Nat. Commun. 14, 5910 (2023).
Ye, Z. et al. High-throughput and scalable single cell proteomics identifies over 5000 proteins per cell. Preprint at bioRxiv, 2023.2011. 2027.568953 (2023).
Guzman, U.H. et al. Ultra-fast label-free quantification and comprehensive proteome coverage with narrow-window data-independent acquisition. Nat. Biotechnol. https://doi.org/10.1038/ s41587-023-02099-7 (2024).
Bekker-Jensen, D. B. et al. A compact quadrupole-orbitrap mass spectrometer with FAIMS interface improves proteome coverage in short LC gradients. Mol. Cell. Proteom. 19, 716-729 (2020).
Batth, T. S. et al. Protein aggregation capture on microparticles enables multipurpose proteomics sample preparation. Mol. Cell. Proteom. 18, 1027-1035 (2019).
Cox, J. et al. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 13, 2513-2526 (2014).
Dang, Y. et al. Functional profiling of stage-specific proteome and translational transition across human pre-implantation embryo development at a single-cell resolution. Cell Discov. 9, 10 (2023).
Aljohani, A., Lai, D. T. C., Bell, P. C. & Edirisinghe, E. A. In Intelligent Computing Methodologies: 13th International Conference, ICIC 2017, Liverpool, UK, August 7-10, 2017, Proceedings, Part III. 3, 13 719-731 (Springer, 2017).
Miller, D. J., Macek, M. B. & Shur, B. D. Complementarity between sperm surface -l, 4-galactosyl-transferase and egg-coat ZP3 mediates sperm-egg binding. Nature 357, 589-593 (1992).
Stopa, N., Krebs, J. E. & Shechter, D. The PRMT5 arginine methyltransferase: many roles in development, cancer and beyond. Cell. Mol. life Sci. 72, 2041-2059 (2015).
Kapanidou, M., Curtis, N. L. & Bolanos-Garcia, V. M. Cdc2O: at the crossroads between chromosome segregation and mitotic exit. Trends Biochemical Sci. 42, 193-205 (2017).
Sanchez-Avila, X. et al. Easy and accessible workflow for label-free single-cell proteomics. J. Am. Soc. Mass Spectrom. 34, 2374-2380 (2023).
Ctortecka, C. et al. Automated single-cell proteomics providing sufficient proteome depth to study complex biology beyond cell type classifications. Preprint at bioRxiv, 2024.2001. 2020.576369 (2024).
Kverneland, A. H., Østergaard, O., Emdal, K. B., Svane, I. M. & Olsen, J. V. Differential ultracentrifugation enables deep plasma proteomics through enrichment of extracellular vesicles. Proteomics 23, 2200039 (2023).
Heil, L. R. et al. Evaluating the performance of the astral mass analyzer for quantitative proteomics using data-independent acquisition. J. Proteome Res. 22, 3290-3300 (2023).
Koenig, C. et al. Protocol for high-throughput semi-automated label-free-or TMT-based phosphoproteome profiling. STAR Protoc. 4, 102536 (2023).
شكر وتقدير
تم دعم Z.Y. من قبل صندوق المعهد المركزي للأبحاث غير الربحية التابع للأكاديمية الصينية للعلوم الطبية (رقم المنحة 2023-RC180-03)، وصندوق الابتكار للأكاديمية الصينية للعلوم الطبية (CAMS) للعلوم الطبية (2022-12M-2-004، 2023-12M-2-005) وصندوق NCTIB لمنصة البحث والتطوير للعلاج بالخلايا والجينات. يتم تمويل العمل في مركز نوفو نورديسك للأبحاث البروتينية (CPR) جزئيًا من خلال تبرع من مؤسسة نوفو نورديسك (NNF14CCOOO1). حصل N.B و J.V.O. على تمويل من صندوق الابتكار الدنماركي بموجب منحة ERA-PerMed-JTC2022-OVA-PDM للمشروع. يتم تمويل P.S. من قبل مجلس البحث السويدي (2022-00323). A.E. و O.Ø. و J.V.O. تلقى الدعم من مجلس البحث الأوروبي منحة التعاون 810057-HighResCells.
مساهمات المؤلفين
قام ز.ي.، ب.س.، و ج.ف.و. بتصميم الدراسة. قام ز.ي. و ب.س. بتنفيذ معظم التجارب وتحليل البيانات. قامت ج.م.-ج. بتنفيذ التجربة على أجنة الفئران. ساهم أ.ع.، د.ب.-ج.، و ن.ب. في تجارب القياس المرجعي. ساهم م.ل.، ي.ج.، ل.س.، ر.ت.، د.ب.-ج.، و ن.ب. في تحسين وتنفيذ تحليل الخلايا الفردية باستخدام جهاز توزيع الخلايا أونو. قام أ.Ø. بتحليل الحويصلات خارج الخلوية. ساعد ج.إكس. في تحليل بيانات أجنة الفئران. كتب ز.ي.، ب.س.، و ج.ف.و. المخطوطة. قرأ جميع المؤلفين وحرروا ووافقوا على النسخة النهائية من المخطوطة.
المصالح المتنافسة
يوان قوه، ليسلي شولتز ورافائيلا تروفر هم موظفون في شركة تيكان، المورد لجهاز توزيع الخلايا الفردية أونو. دورتي ب. بيكر-جنسن، نيكولاي باكه هم موظفون في شركة إيفوسيب بايوسيستمز، المصنعة لأجهزة إيفوسيب وإيفوتيب المستخدمة في هذا العمل. المؤلفون الآخرون يعلنون عدم وجود مصالح متنافسة.
معلومات إضافية
معلومات إضافية النسخة الإلكترونية تحتوي على المواد التكميلية متاحة على https://doi.org/10.1038/s41467-024-46777-9. يجب توجيه المراسلات والطلبات للحصول على المواد إلى زيلو يي أو يسبر ف. أولسن.
تُعرب مجلة Nature Communications عن شكرها لفاديم ديميتشيف، وبنجامين أورسبورن، والمراجعين الآخرين المجهولين، على مساهمتهم في مراجعة هذا العمل. يتوفر ملف مراجعة الأقران.
معلومات إعادة الطباعة والتصاريح متاحة على http://www.nature.com/reprints ملاحظة الناشر: تظل شركة سبرينغر ناتشر محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
مختبر الدولة الرئيسي لأبحاث الآليات الشائعة للأمراض الكبرى، معهد سوتشو للطب النظامي، الأكاديمية الصينية للعلوم الطبية وكلية بكين للطب، سوتشو، الصين. مركز مؤسسة نوفو نورديسك لأبحاث البروتين، جامعة كوبنهاغن، كوبنهاغن، الدنمارك. قسم العلوم الجراحية، جامعة أوبسالا، أوبسالا، السويد. المرافق الأساسية للفئران المعدلة وراثياً، قسم الطب التجريبي، جامعة كوبنهاغن، كوبنهاغن، الدنمارك. مجموعة تيكان المحدودة، مانيندورف، سويسرا. إيفوسيب بيوسيستمز، أودنسه، الدنمارك. ساهم هؤلاء المؤلفون بالتساوي: زيلو يي، بيير ساباتيه. البريد الإلكتروني: yzl@ism.pumc.edu.cn; jesper.olsen@cpr.ku.dk
Mass spectrometry (MS)-based proteomics workflows typically involve complex, multi-step processes, presenting challenges with sample losses, reproducibility, requiring substantial time and financial investments, and specialized skills. Here we introduce One-Tip, a proteomics methodology that seamlessly integrates efficient, one-pot sample preparation with precise, narrow-window data-independent acquisition (nDIA) analysis. One-Tip substantially simplifies sample processing, enabling the reproducible identification of proteins from HeLa cells. The versatility of One-Tip is highlighted by nDIA identification of proteins in single cells from early mouse embryos. Additionally, the study incorporates the Uno Single Cell Dispenser , demonstrating the capability of One-Tip in single-cell proteomics with >3000 proteins identified per HeLa cell. We also extend One-Tip workflow to analysis of extracellular vesicles (EVs) extracted from blood plasma, demonstrating its high sensitivity by identifying proteins from 16 ng EV preparation. One-Tip expands capabilities of proteomics, offering greater depth and throughput across a range of sample types.
Conventional bottom-up proteomics workflows are based on multistep sample-handling processes, incorporating bulk cell lysis and protein extraction from millions of cells, reduction, and alkylation of cysteines, protein digestion, sample cleanup and concentration, followed by optional offline peptide fractionation prior to online liquid chromatography tandem mass spectrometry (LC-MS/MS) acquisition . The different steps introduce several challenges, including potential sample contamination and losses leading to reproducibility problems, considerable time and cost investments, and the necessity for specialized expertise and training. Consequentially, these barriers often discourage biomedical scientists from undertaking MS-based proteomics experiments.
To address this, several one-pot workflows have emerged, streamlining the sample preparation process for both bulk proteomics and single-cell proteomics (SCP) . These workflows aim to streamline the sample preparation process while maintaining the depth and accuracy of proteomic analysis. For instance, Kulak et al. introduced an innovative in-StageTip protocol , which simplifies proteome-sample preparation down to just five pipetting steps. This method is particularly convenient as it can be easily adapted to a well format, making it suitable for high-throughput applications. The one-pot concept has gained widespread acceptance in the SCP community due to its efficiency and simplicity. Recently, Johnston et al. proposed a sample preparation method that integrates cell lysis,
protein denaturation, and digestion into a single hour-long process . Matzinger et al. presented a comprehensive workflow encompassing improved strategies for all stages of sample preparation, standardized for 384-well plates .
Despite these advancements, challenges remain. In bulk proteomics, methods still necessitate substantial sample input, separate cell lysis, multiple liquid handling steps, sample transfers, and extended LC-MS/MS analysis times to achieve the desired proteome depth. These requirements often lead to limited throughput, reproducibility, and applicability, and may not fully capture the proteome depth. One critical limitation in the SCP methods is the lack of a sample cleanup step, which risks compromising the sample quality and hence longevity of LC-MS systems. Recognizing this gap, our study introduces the One-Tip workflow, which is meticulously designed to overcome the prevalent issues in bulk proteomics. One-Tip is primarily tailored to achieve comprehensive proteome depth with high quantitative accuracy and precision, even with a low number of cells. Moreover, we demonstrate the versatility of One-Tip through a wide array of applications, including the analysis of single early embryonic cells and human plasma extracellular vesicle samples. Although not specifically created for SCP, we also show that it is feasible to apply it directly to SCP using the Uno Single Cell dispenser. This innovative and simple workflow promises to transform proteomic sample preparation, offering both efficiency and depth in proteomic analysis.
Results
One-Tip epitomizes the most streamlined workflow for proteomics
The One-Tip methodology only requires two pipetting steps, utilizing a commercially available Evotip : one for the cell lysis and proteolytic digestion buffer, and the other for the cell suspension in PBS (Fig. 1a). After an hour-long incubation in a water-filled Evotip box with standard Evotip preparation, the One-Tip samples are ready for LC-MS/MS analysis. The combined cell lysis, protein extraction, and endoproteinase Lys-C/trypsin digestion master mix buffer includes an MScompatible surfactant, n-Dodecyl- -D-Maltoside (DDM), which is a
water-soluble non-ionic detergent effective in cell membrane lysis and solubilization of proteins without denaturation.
Conceptually, One-Tip represents the simplest workflow for proteomics, as it necessitates no further sample handling and directly integrates sample preparation with the LC-MS analysis without any sample losses and variation. Low cell input numbers result in high proteome coverage and circumvents the need for sonication and additional buffer exchanges. This results in a total sample preparation time of approximately 70 min , including a digestion time of 60 min . Moreover, the One-Tip workflow is not limited in throughput; even manual handling can efficiently prepare thousands of samples daily by using multi-channel pipetting and is also easily automated on liquid handling robots. Lastly, we couple the One-Tip workflow with a top-of-the-line LC-MS/MS system to guarantee superior analytical performance. This is based on the highly sensitive Whisper LC method on the Evosep One LC and our newly developed narrow-window Data Independent Acquisition (nDIA) method on an Orbitrap Astral mass spectrometer .
In the initial experiments, we evaluated different ratios of master mix to HeLa single cell suspension in PBS, testing combinations such as master mix cells, , and , using the previous generation Orbitrap MS platform , the Orbitrap Exploris 480 (Fig. 1b). The cell concentration was estimated at 100 cells l. Our findings revealed that even the smallest ratio, , yielded a notable 3172 protein group (hereafter referred to as protein) identifications. This protein coverage is comparable to the number of proteins identified in 10 ng HeLa samples prepared using the protein aggregation capture (PAC) protocol , a standard reference. Notably, the performance difference observed between the Orbitrap Exploris 480 and the Orbitrap Astral when analyzing One-Tip samples mirrored the differences seen with standard HeLa samples. This consistency indicates that our OneTip protocol exhibits no bias towards specific mass spectrometry platforms.
To further test the sensitivity and reproducibility of the One-Tip methodology, we deployed it on the Orbitrap Astral using nDIA analysis with increasing numbers of HeLa cells, analyzing 20,100,200,
Fig. 1 | One-tip achieves near-complete proteome depth in single-shot analysis. a Graphic depiction of One-Tip workflow. Number of proteins identified with and peptides (d) identified in different number of cells. Digestion time was 2 h in these samples. Source data are provided as a Source Data file. different volumes of the master mix and cells in PBS. c, d Number of proteins (c)
Fig. 2 | One-Tip shows precise quantification and comparable proteome coverage with PAC. a Distribution of coefficient of variance between triplicates in different number of cells. Overlap of proteins identified in One-Tip (1000 cells) and PAC ( 1000 ng ) workflows. c Correlation of protein abundances between OneTip and PAC samples. d Subcellular localization of proteins identified in One-Tip and PAC samples. e GO over-representation enrichment of proteins identified only
in One-Tip samples. P values were calculated using a one-sided Fisher’s Exact Test and subsequently adjusted using the Benjamini-Hochberg (BH) correction method. Number of proteins ( ) and peptides ( ) identified in different number of cells using One-Tip and different peptide loading amounts using PAC. Percentage of missed cleavages in One-Tip and PAC samples. Source data are provided as a Source Data file.
500, 1000, 2000, and 3000 cells, respectively. Note these cell quantities were based on cell counting estimations and therefore are approximations. And the volume of master mix in this experiment might not be optimal for the different number of cells. One-Tip exhibited an impressive proteome depth for single-shot analysis in half-an-hour LC-MS/MS runs, identifying over 5000 protein groups and over 46,000 modified peptides (hereafter referred to as peptides) from approximately 20 cells (Fig. 1c). This performance surpasses the proteome coverage currently achieved by the isolation of precisely 20 cells using the CellenONE, a dedicated single-cell proteomics preparation system, which is likely related to sample losses in a 96 well plate and transfer to the LC-MS system. Furthermore, a starting quantity of cells nearly covered the complete proteome, with over 9000 proteins and 170,000 peptides identified (Fig. 1c, d). We subsequently evaluated the combined endoproteinase Lys-C and trypsin-digestion efficiency by varying the digestion time from 1 to 4 h . Even a proteolytic digestion time as short as 1 h was sufficient to
effectively digest the entire proteome, resulting in high sequence coverage with a missed cleavage rate of less than in 20 cells and approximately for larger cell quantities (Supplementary Figs. 1-3).
One-Tip shows precise quantification and comparable proteome coverage with PAC using 1000 cells
One-Tip demonstrated high reproducibility and quantitative precision, with a remarkable coefficient of variance of in workflow replica analyzes with a Pearson correlation exceeding 0.99 for maxLFQ based protein quantitation in samples with more than 100 cells (Fig. 2a, Supplementary Fig. 4). Despite its simple and swift workflow, the analytical performance of One-Tip with one thousand cells is comparable to that of bulk proteomics samples prepared using a more sophisticated method, Protein Aggregation Capture (PAC) , when combined with our narrow-window DIA method on the Orbitrap Astral mass spectrometer. Compared to 1000-ng PAC-digested HeLa lysate analyzed by LC-MS/MS in , the One-Tip method applied to
Fig. 3|One-Tip and LC-MS analysis of the mouse pre-implantation embryo until the 4-cell stage. a Graphic depiction of the workflow using One-Tip for mouse preimplantation embryos. Number of proteins with quantified values in the study from Dang et al. and our study, and protein overlap between samples (in grey) from the same development stages. The mean number of proteins for each condition, the overlap and the mean percentage of the overlap compared to each sample are
indicated. c Mean correlation of normalized protein abundances between samples within each sample groups (oocyte, zygote, 2-cell stage, 4-cell stage). d Summed protein intensities for each group of samples. Error bars represent the standard deviation of the mean. Number of biological replicates: for oocyte, for zygote, for 2-cell stage and for 4-cell stage. Source data are provided as a Source Data file.
cell samples in a half-an-hour LC-MS/MS run identified a similar number of proteins with substantial overlap between them and very high quantification precision (Fig. 2b, c). The lower Pearson correlation of in protein quantities between PAC and One-Tip samples is likely caused by the difference in cysteine-alkylation and missed-cleavage rates (Supplementary Fig. 5). Interestingly, both the PAC and One-Tip methods identified slightly more than 1000 unique proteins, but exhibiting no considerable bias in protein ontologies towards specific subcellular localizations (Fig. 2d), and a slight preference for particular biological functions such as nucleosome assembly in proteins identified only in One-Tip samples (Fig. 2e).
Lastly, to assess the loss of material during One-Tip preparation, we compared a dilution series of HeLa PAC digest from 100 ng to 10 ng, thus starting from desalted peptides transferred to Evotips for injection, to a dilution series of intact cells sorted using cellenONE corresponding to 400,200 and 40 HeLa cells, which were processed using One-Tip and thus only lysed in the Evotip. The protein amount in the cells corresponds to approximately 100,50 , and 10 ng of starting material, assuming lysis efficiency and recovery. It is important to note that the number of cells sorted by the cellenONE represents the maximum number of cells potentially present in each sample, but the transfer likely resulted in cell loss. Nevertheless, this represents the fairest comparison of the two methods possible with our current instrumentation with a disadvantage for the One-Tip approach. We then compared the protein and peptide numbers identified in each sample (Fig. 2f, g). Overall, the protein and peptide numbers in samples from cellenONE were close to their equivalent in peptide loading or slightly lower (i.e. 400 cells were close to 50 ng ). The variation was higher in lower cell numbers, which is likely due to loss of cells during transfer to the tips. Finally, the percentage of missed cleavages was higher in One-Tip samples than in samples from PAC digestion (Fig. 2h).
One-Tip enables single-cell proteome analysis of mouse preimplantation embryos
To demonstrate the versatility and sensitivity of the One-Tip workflow, we next studied the development of mouse pre-implantation embryos at single-cell level from oocyte and zygote to 4 -cell stage. The zona pellucida, the extracellular glycoproteinaceous coat of oocytes, zygotes, 2 -cell and 4 -cell stages, was chemically removed and single cells were dissociated under a microscope and dispensed individually into Evotips, pre-loaded with cell lysis and digestion buffer (Fig. 3a). This whole procedure was performed in one day for each batch of samples including the LC-MS/MS analysis. In total, 6216 protein groups were identified with a median of , and 4779 , for each developmental stage, respectively (Fig. 3b). In comparison to the very recent analysis of human pre-implantation embryos from Dang et al. in which the authors employed a sophisticated sample preparation workflow and LC-MS gradients of 4 times longer duration than the one used in this study, we identify ~2300 more proteins and ~50,000 more peptides as well as peptides per protein on average ( 5.3 against 14.6), per stage (Supplementary Fig. 5a, b). Moreover, the variation in protein number, the overlap (Fig. 3b), and correlation of protein quantities between samples from a same condition was greatly improved (Fig. 3c, Supplementary Fig. 5c), demonstrating that the One-Tip workflow coupled to Evosep One and Orbitrap Astral is faster, more sensitive, more reproducible and has better quantification accuracy. Interestingly, the summed protein intensities in each sample were equal between oocytes and zygotes, while they were half those intensities in cells from the 2-cell stage and one-fourth in cells from the 4 -cell stage, matching the reduction in cell volume during the two successive cleavages of the zygote (Fig. 3d). Although our analysis benefitted from the use of the latest generation of mass spectrometer compared to an older generation in the study from Dang et al., which would inflate these numbers, our study highlights that using our
Fig. 4 | Biological insights from the mouse pre-implantation embryos.
a Unsupervised hierarchical clustering using canberra and ward.D2 methods of proteins that shows significant regulation ( from one-sided ANOVA) and FC fold in any of the developmental stages compared to the oocyte group (left). GO biological processes enrichment analysis of the protein clusters from the heatmap (right). Number of biological replicates: for oocyte, for zygote, for 2 -cell stage and for 4 -cell stage. PCA of the normalized protein intensities for each samples of our study. Only proteins that were quantified in all
samples were considered ( ). c Abundances of twelve proteins related to reproductive processes across four developmental stages. For the boxplots, the lower and upper hinges correspond to the first and third quartiles. The upper whisker extends from the hinge to the largest value no further than IQR from the hinge (where IQR is the inter-quartile range). The lower whisker extends from the hinge to the smallest value at most IQR of the hinge. Number of biological replicates: for oocyte, for zygote, for 2 -cell stage and for 4 -cell stage. Source data are provided as a Source Data file.
minimalistic One-Tip workflow enables the acquisition of comprehensive datasets with greatly limited sample material.
Finally, we investigated whether biologically meaningful information can be retrieved from the mouse embryo cell analysis. We then performed unsupervised hierarchical clustering of differentially expressed proteins that were up or downregulated in any of the conditions compared to oocytes (Fig. 4a). Each sample group was separated on the heatmap apart from the zygotes and oocytes as on the PCA. Gene ontology (GO) enrichment analysis of clusters reveals downregulation of proteins related to sperm binding to the zona pellucida, prevention of polyspermy, response to ER stress and upregulation of proteins related to DNA replication and heterochromatin assembly in 2 -cell and 4 -cell stages compared to oocytes, among other pathways. These results were globally in agreement with Dang et al. validating the quality of our analysis (Fig. 4a). The principal component analysis (PCA) of normalized protein intensities showed that oocyte and zygote group together and the 2 -cell and 4 -cell stages are
separated in a time-wise manner on PC1 (Fig. 4b). We then focused on the quantities of twelve proteins related to reproductive processes across four developmental stages (Fig. 4c). Notably, ZP3, a protein critical for zona pellucida formation , decreased during the 2 -cell stage, aligning with the first cell division, but stabilized at the 4 -cell stage, indicating dynamic changes in early embryonic development. In contrast, the quantities of WDR77, involved in transcriptional regulation , and CDC20, essential for cell division processes , remained stable across all stages. This consistency suggests continuous mitotic and transcriptional activity from the zygote stage, highlighting their crucial roles in early embryonic development.
Scale One-Tip down to single HeLa cells with single cell dispenser
Recently, the Uno Single Cell Dispenser (equivalent to the HP D100 Single Cell Dispenser) was described for single-cell sorting for proteomics analysis . Since the design of the instrument allows
a
Fig. 5 | Scale One-Tip down to single HeLa cells with single cell dispenser.
a Workflow of single cell proteomics using One-Tip and Uno Single Cell Dispenser. b, c Number of proteins (b) and peptides (c) identified in different sample
preparation conditions. Number of samples: 0and2_pipette,
0and2_sorter, land2_pipette, 1and2_sorter, carrier. Source data are provided as a Source Data file.
fitting a box of Evotips, we evaluated whether the One-Tip workflow could be amenable to single-cell analysis and tested multiple experimental set ups (Fig. 5a). In short, we either placed of lysis and digestion buffer or nothing, sorted the cells and then dispensed of Lysis buffer either manually (pipette) or with the dispenser. The results show that single-cell analysis is feasible using the Uno Single Cell Dispenser coupled to One-Tip with more than 2000 proteins quantified on average for the best condition manually) with no carrier or reference sample with higher signal to boost IDs during analysis with Spectronaut and more than 3000 with a carrier sample (Fig. 5b, c). Some of the tips had little signal suggesting that the cells did not reach the bottom of the Evotips and were not lysed and digested properly. However, consistent signal was detected in up to of the samples, where single cells were supposed to be present according to the instrument software, which is lower than what was reported by Sanchez-Avila et al. using a 384 -well PCR plate. While the number of peptides and proteins identified and quantified are lower than for SCP analysis using the cellenONE instrument and the same LC-MS , using the combination of Uno Single Cell Dispenser and One-Tip is an economical and simple alternative to some of the more complicated and costly workflows used currently. Lastly, the workflow presented here has room for improvement both in terms of percentage of single cells recovered and in terms of peptide and protein numbers, and this should be further developed in the future.
High-sensitivity workflow for extracellular vesicles with One-Tip To further showcase the broad applicability and sensitivity of the One-Tip workflow, we processed extracellular vesicles (EV)
extracted from blood plasma rather than intact cells. We isolated EVs from the plasma of male donors as described by Kverneland et al. and loaded a dilution series from 480 ng to 16 ng (estimated based on the protein concentration of plasma during EV purification) (Fig. 6a) on Evotips prepared for One-Tip analysis (Fig. 6b). We quantified more than 3000 proteins and 29000 peptides with 16 ng of starting material and close to 4500 proteins and 48000 peptides with 480 ng , the latter being on par with the state-of-the-art in plasma EV analysis (Fig. 6c). Additionally, the number of protein and peptides identified scales with the amount of sample loaded, as observed with the mouse embryo data. Thus, this data shows that One-Tip can readily be applied to other types of samples than single cells or cell suspensions and will enable more sensitive, streamlined and higher throughput analysis of EVs. This is particularly relevant in the field of plasma analysis and biomarker discovery as well as in the EV field in general, where the amount of EVs originating from plasma or cells can be limited.
Discussion
Taken together, our results demonstrate that the analytical depth achieved with the One-tip workflow is superior to standard workflows in the field of proteomics. Its simplicity, user-friendliness, and scalability render it highly suitable for a variety of experimental setups, ranging from single-cell analysis to bulk proteomics. This approach effectively eliminates the need for complex methodologies, specific equipment, and specialized proteomics expertise during the sample preparation phase. As a result, the One-Tip workflow substantially bridges the gap between proteomics and biology, simplifying LC-MS analyzes and making them more accessible and compatible with a
Fig. 6 | One-Tip analysis of extracellular vesicles. a Workflow for extracellular vesicle purification. Workflow for One-Tip EV digestion and mass spectrometry analysis. Number of proteins (b) and peptides (c) identified in different loading amounts. technical replicates per condition. Source data are provided as a Source Data file.
broader range of biological applications. The workflow is adaptable to platforms such as cellenONE and Uno Single Cell Dispenser , enhancing its versatility.
Additionally, One-Tip is capable of analyzing a diverse array of sample types, including FACS-sorted cells, human pre-implantation embryos, rare cell populations, and tissue slices. This is particularly relevant for smaller cell types, like T-cells, where lower number of proteins are expected to be identified as it correlates with the protein content (approximately 25 pg per cell). For such cells, single-cell analysis may offer limited biological insights, and the One-Tip workflow presents an effective alternative. Furthermore, we demonstrate the applicability of One-Tip to non-cellular samples, such as extracellular vesicles, showcasing its utility in a wide spectrum of research contexts. This adaptability makes One-Tip a valuable tool in the arsenal of modern proteomics, enabling a more inclusive approach to sample analysis and facilitating a deeper understanding of various biological systems.
Methods
This study complies with all relevant ethical regulations. Animal work was conducted according to license no. 2021-15-0201-00851, approved
by the Danish National Animal Experiments Inspectorate, and performed according to national and local guidelines.
HeLa cell lines
HeLa human cervix carcinoma cells (ATCC CCL-2) were cultured in DMEM (Gibco, Invitrogen), supplemented with fetal bovine serum, penicillin (Invitrogen), streptomycin (Invitrogen), at , in a humidified incubator with . At confluence, cells were detached using trypsin and washed twice with Phosphate Buffered Saline (PBS) from Gibco (Life Technologies), before being resuspended in degassed PBS.
Detailed One-Tip sample preparation workflow
Determine HeLa cell concentration using a cell counter and dilute to the desired concentrations with PBS.
Prepare the Evotips following the vendor’s instructions:
Rinse: Wash dry Evotips with of Solvent B ( FA in acetonitrile) and centrifuge at 800 g for 60 seconds.
Condition: Soak the Evotips in of 1-propanol until they turn pale white.
Equilibrate: Saturate the conditioned Evotips with of Solvent A ( in water) and centrifuge at 800 g for 60 seconds.
Pipette of lysis and digestion buffer into the Evotips. The buffer contains -Dodecyl- -D-Maltoside (DDM), 100 mM TEAB, Trypsin, and Lys-C.
Pipette of cells into the Evotips.
Briefly centrifuge the Evotips at 50 g to mix the buffer and cells and prevent the formation of air bubbles.
Add water to the Evotip box to the level of the C18 resin in the Evotips.
Incubate the Evotip box at for 1 to 4 hours.
Continue with the vendor’s instructions with a slight modification:
Load: add of Solvent A to the Evotips and centrifuge for 60 seconds at 800 g .
Wash: Rinse the tips with of Solvent A and centrifuge for 60 seconds at 800 g .
Wet: Add of Solvent A to the tips and centrifuge for 10 seconds at 800 g to keep the tips wet.
HeLa cell isolation with CellenONE
HeLa cells were diluted with degassed PBS to cells cells were isolated into a 96 well-plate preloaded with PBS using the CellenONE system. After isolation, samples were prepared following the One-Tip workflow.
Isolation of mouse oocytes, zygotes, 2-cell and 4-cell stages
Animal work was conducted according to license no. 2021-15-020100851, approved by the Danish National Animal Experiments Inspectorate, and performed according to national and local guidelines. Mice were kept in designated rooms in individually ventilated cages at a temperature of , with a humidity of , air in the room was changed eight to ten times per hour and dark/light cycle is , light from 6 am to 6 pm . Mouse husbandry is performed according to Danish regulations for animal experiments.
Ovulation of prepubescent (4-week old) C57BL/6NRj females (mus musculus) was induced by intraperitoneal (IP) injection of PMSG (HOR-272, Prospec), 5IU/female, followed by IP injection of hCG (Chorulon Vet, Pharmacy), 5IU/female, 47 h later. After the second injection, females were set in breeding with C57BL/6NRj stud males. Next morning, the females were euthanized and oviducts were dissected to harvest the cumuli containing zygotes and unfertilized oocytes. Cumuli were disaggregated by 10 -minute incubation in Hyaluronidase (H4272, Sigma-Aldrich). Zygotes were sorted out assessed by the presence of the second polar body and cultured in KSOM medium (MR-106-D, Merc Millipore). On the day of harvesting a group of both zygotes and oocytes were processed for analysis, while other zygotes were incubated in KSOM at and for 24 h or 48 h . At the different developmental stages, oocytes and embryos were incubated in Tyrode’s Acidic Solution (T1788, Sigma-Aldrich) for 5 to 15 s in order to remove the zona pellucida. Naked oocyes and zygotes were washed in PBS (20012-027, Life Technologies) and individually transferred to Evotips using a diameter glass capillary. 2-cell and 4 -cell embryos were further disaggregated by 5 -minute incubation in and -free KSOM and pippeting with a diameter glass capillary in PBS afterwards; individual blastomeres were then transferred to Evotips. The Evotips were preloaded with of lysis and digestion buffer and of PBS. Incubation time was 4 h . Number of biological replicates: for oocyte, for zygote, for 2 -cell stage and for 4 -cell stage. A total of 20 donor females and 8 stud males were used in these experiments.
Cell isolation using the Uno Single Cell Dispenser
Cell were prepared as for the sorting with the CellenOne and were passed through a cutoff mesh to ensure that no clumps were present prior to sorting. Evotips were prepared for One-Tip analysis as
described above and in Fig. 5a until the step where the lysis buffer is added. Since the lysis buffer is diluted with the cell suspension in PBS in a regular One-Tip experiment, we pre-diluted the lysis buffer 1:1 with PBS. Then either was added manually followed by centrifugation at 50 g for 10 s or no lysis buffer was added. After this, water was added to the Evotip box to cover only the tip of the Evotips and the box was placed on the Uno Single Cell Dispenser for single-cell dispensing. of cell suspension were added to a C1 cassette and single cells were dispensed into the Evotips. After cell dispensing, of diluted lysis buffer were added either by pipetting or with the dispenser using a D1 cassette. Water was removed and the tips were centrifuged at 50 g for 10 s so that the lysis buffer is in contact with the Evotip C18 phase. Water was added again to cover the bottom of the box and it was incubated at for 2 h . The rest of the sample preparation followed the One-Tip workflow. The number of sample was: Oand2_pipette, 0and2_sorter, land2_pipette, 1and2_sorter, n = 3 carrier.
EV preparation
Venous blood samples were collected in citrate tubes (Vacuette, 3.2% sodium citrate, Greiner Bio-One) from male donors and centrifuged at for 10 min within 30 min after blood collection. The supernatant (platelet rich plasma) was transferred to a new tube and subject to a second centrifugation at 3000 g for 10 min to pellet remaining platelets and the platelet-poor supernatant was aliquoted, snap-frozen, and stored at . At the day of EV isolation, the platelet-poor plasma was thawed on wet ice and processed for EV enrichment as described in Fig. 6a. In short, of plasma was diluted 1:1 with PBS pH 7.2 (Gibco cat# 20012-019) and centrifuged at for 30 min . of supernatant were removed and of PBS were added on top of the pellet ( dilution), followed by centrifugation at for 30 min . This operation was performed 4 times in total and after the last centrifugation of supernatant were removed and the remaining containing the EV-pellet were further processed. The protein content in the EV-preparation was estimated based on the dilution of the total protein concentration in plasma ( ) and 4 ug of EVs in total (with a recovery of after each centrifugation). The final EV preps after 4 centrifugations ( 3 washes) were diluted to obtain amounts equivalent to and 480 ng in . These of each dilution in duplicates were added to Evotips prepared for OneTip analysis containing of lysis buffer and processed following the One-Tip protocol described above, prior to LC-MS/MS analysis. Experiment was performed in duplicate for each condition.
PAC
Protein aggregation capture (PAC) was performed as previously described . Briefly, HeLa cells were lysed in SDS, 100 mM Tris pH tris(2-carboxyethyl)phosphine (TCEP), and 10 mm 2-chloroacetamide (CAA) and sonicated using a Branson probe sonicator 1 min using 3 s pulse and amplitude. Protein concentration was measured using BCA assay, and of proteins were used for PAC digestion. PAC digestion was conducted on a KingFisher Flex (Apex, Duo Prime) system (Thermo Fischer Scientific), sample volume was adjusted to 300 with lysis buffer, then of MagReSyn Hydroxyl beads (Resyn Biosciences) were added (2:1, beads/protein, ) followed by of acetonitrile, beads were rinsed 3 times with acetonitrile, and 2 times with ethanol and on-bead digestion was conducted in 50 mM ammonium bicarbonate containing lysyl endopeptidase (1:500, enzyme/protein, w/w) and trypsin (1:250, enzyme/protein, w/w). Samples were acidified using formic acid to and loaded onto Evotips prior to LC-MS/MS analysis.
LC-MS/MS
LC-MS/MS analysis was performed on an Orbitrap Astral MS using Thermo Tune software (version 0.4 or higher) coupled to an Evosep
One system (EvoSep Biosystems). Samples were analyzed in 40SPD (31min gradient) using a commercial analytical column (Aurora Elite TS, lonOpticks) interfaced online using an EASY-Spray source. The Orbitrap Astral MS was operated at a full MS resolution of 240,000 with a full scan range of when stated. The full MS AGC was set to . MS/MS scans were recorded with 2Th isolation window, 3 ms maximum ion injection time (IIT) for HeLa samples and 4Th and 6 ms IIT for mouse embryonic samples, for the dilution series of PAC and CellenONE-sorted cells and for the single-cell analysis using the Uno dispenser. MS/MS scanning range was from were used. The isolated ions were fragmented using HCD with 27% NCE.
MS data analysis
Raw files were analyzed in Spectronaut v18 (Biognosys) with a spectral library-free approach (directDIA + ) using the human protein reference database (Uniprot 2022 release, 20,588 sequences) for HeLa samples, and the mouse protein reference database (Uniprot 2022 release, 21,989 sequences) for the mouse embryo single cells complemented with common contaminants ( 246 sequences). Note, as the protocol does not involve reduction and alkylation, database searches were performed with free cysteine sulfhydryls and hence cysteine carbamylation was not set as a fixed modification, whereas methionine oxidation and protein N-termini acetylation were set as variable modifications. Precursor filtering was set to perform based on Qvalues, and cross run normalization was checked. Each experiment with different number of cells was analyzed separately, and samples from different digestion times were searched with and without enabling method evaluation and indicating the different conditions (each one with experimental replicates) in the condition setup tab.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The mass spectrometry proteomics data generated in this study have been deposited to ProteomeXchange Consortium (http:// proteomecentral.proteomexchange.org) via the PRIDE partner repository with dataset identifier PXD044991. Source data are provided with this paper.
References
Zhang, Y., Fonslow, B. R., Shan, B., Baek, M.-C. & Yates, J. R. III Protein analysis by shotgun/bottom-up proteomics. Chem. Rev. 113, 2343-2394 (2013).
Kulak, N. A., Pichler, G., Paron, I., Nagaraj, N. & Mann, M. Minimal, encapsulated proteomic-sample processing applied to copynumber estimation in eukaryotic cells. Nat. Methods 11, 319-324 (2014).
Chen, W. et al. 3D-SISPROT: A simple and integrated spintip-based protein digestion and three-dimensional peptide fractionation technology for deep proteome profiling. J. Chromatogr. A 1498, 207-214 (2017).
Yu, Y. et al. Development of an efficient, effective, and economical technology for proteome analysis. Preprint at Research Square, https://doi.org/10.21203/rs.3.rs-3165690/v1 (2023).
Johnston, S. M. et al. Rapid, one-step sample processing for labelfree single-cell proteomics. J. Am. Soc. Mass Spectrom. 34, 1701-1707 (2023).
Matzinger, M., Müller, E., Dürnberger, G., Pichler, P. & Mechtler, K. Robust and Easy-to-Use One-Pot Workflow for Label-Free SingleCell Proteomics. Anal. Chem. 95, 4435-4445 (2023).
Petrosius, V. et al. Exploration of cell state heterogeneity using single-cell proteomics through sensitivity-tailored data-independent acquisition. Nat. Commun. 14, 5910 (2023).
Ye, Z. et al. High-throughput and scalable single cell proteomics identifies over 5000 proteins per cell. Preprint at bioRxiv, 2023.2011. 2027.568953 (2023).
Guzman, U.H. et al. Ultra-fast label-free quantification and comprehensive proteome coverage with narrow-window data-independent acquisition. Nat. Biotechnol. https://doi.org/10.1038/ s41587-023-02099-7 (2024).
Bekker-Jensen, D. B. et al. A compact quadrupole-orbitrap mass spectrometer with FAIMS interface improves proteome coverage in short LC gradients. Mol. Cell. Proteom. 19, 716-729 (2020).
Batth, T. S. et al. Protein aggregation capture on microparticles enables multipurpose proteomics sample preparation. Mol. Cell. Proteom. 18, 1027-1035 (2019).
Cox, J. et al. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 13, 2513-2526 (2014).
Dang, Y. et al. Functional profiling of stage-specific proteome and translational transition across human pre-implantation embryo development at a single-cell resolution. Cell Discov. 9, 10 (2023).
Aljohani, A., Lai, D. T. C., Bell, P. C. & Edirisinghe, E. A. In Intelligent Computing Methodologies: 13th International Conference, ICIC 2017, Liverpool, UK, August 7-10, 2017, Proceedings, Part III. 3, 13 719-731 (Springer, 2017).
Miller, D. J., Macek, M. B. & Shur, B. D. Complementarity between sperm surface -l, 4-galactosyl-transferase and egg-coat ZP3 mediates sperm-egg binding. Nature 357, 589-593 (1992).
Stopa, N., Krebs, J. E. & Shechter, D. The PRMT5 arginine methyltransferase: many roles in development, cancer and beyond. Cell. Mol. life Sci. 72, 2041-2059 (2015).
Kapanidou, M., Curtis, N. L. & Bolanos-Garcia, V. M. Cdc2O: at the crossroads between chromosome segregation and mitotic exit. Trends Biochemical Sci. 42, 193-205 (2017).
Sanchez-Avila, X. et al. Easy and accessible workflow for label-free single-cell proteomics. J. Am. Soc. Mass Spectrom. 34, 2374-2380 (2023).
Ctortecka, C. et al. Automated single-cell proteomics providing sufficient proteome depth to study complex biology beyond cell type classifications. Preprint at bioRxiv, 2024.2001. 2020.576369 (2024).
Kverneland, A. H., Østergaard, O., Emdal, K. B., Svane, I. M. & Olsen, J. V. Differential ultracentrifugation enables deep plasma proteomics through enrichment of extracellular vesicles. Proteomics 23, 2200039 (2023).
Heil, L. R. et al. Evaluating the performance of the astral mass analyzer for quantitative proteomics using data-independent acquisition. J. Proteome Res. 22, 3290-3300 (2023).
Koenig, C. et al. Protocol for high-throughput semi-automated label-free-or TMT-based phosphoproteome profiling. STAR Protoc. 4, 102536 (2023).
Acknowledgements
Z.Y. was supported by the Non-profit Central Research Institute Fund of the Chinese Academy of Medical Sciences (grant no. 2023-RC180-03), the Chinese Academy of Medical Sciences (CAMS) Innovation Fund for Medical Sciences (2022-12M-2-004, 2023-12M-2-005) and the NCTIB Fund for the R&D Platform for Cell and Gene Therapy. Work at The Novo Nordisk Foundation Center for Protein Research (CPR) is funded in part by a donation from the Novo Nordisk Foundation (NNF14CCOOO1). N.B and J.V.O. received funding from the Innovation Fund Denmark under the grant ERA-PerMed-JTC2022-OVA-PDM for the project. P.S. is funded by the Swedish Research Council (2022-00323). A.E., O.Ø., and J.V.O.
received support from the European Research Council ERC Synergy Grant 810057-HighResCells.
Author contributions
Z.Y., P.S., and J.V.O. conceptualized the study. Z.Y. and P.S. performed most of the experiments and data analysis. J. M.-G. performed the experiment on mouse embryos. A.E., D. B.-J., and N. B. contributed to the benchmarking experiments. M. L., Y. G., L. S., R. T., D. B.-J., and N. B. contributed to optimizing and performing the single-cell analysis using the Uno cell dispenser. O. Ø. performed the analysis of extracellular vesicles. J. X. helped with data analysis of the mouse embryo. Z.Y., P.S., and J.V.O. wrote the manuscript. All authors read, edited, and approved the final version of the manuscript.
Competing interests
Yuan Guo, Lesley Schultz and Rafaela Truffer are employees of Tecan, the vendor of the Uno Single Cell Dispenser. Dorte B. Bekker-Jensen, Nicolai Bache are employees of Evosep Biosystems, manufacturer of the Evosep instrumentation and Evotip used in this work. Other authors declare no competing interests.
Additional information
Supplementary information The online version contains
supplementary material available at https://doi.org/10.1038/s41467-024-46777-9.
Correspondence and requests for materials should be addressed to Zilu Ye or Jesper V. Olsen.
Peer review information Nature Communications thanks Vadim Demichev, Benjamin Orsburn and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Reprints and permissions information is available at http://www.nature.com/reprints
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
State Key Laboratory of Common Mechanism Research for Major Diseases, Suzhou Institute of Systems Medicine, Chinese Academy of Medical Sciences & Peking Union Medical College, Suzhou, China. Novo Nordisk Foundation Center for Protein Research, University of Copenhagen, Copenhagen, Denmark. Department of Surgical Sciences, Uppsala University, Uppsala, Sweden. Core Facility for Transgenic Mice, Department of Experimental Medicine, University of Copenhagen, Copenhagen, Denmark. Tecan Group Ltd., Männedorf, Switzerland. Evosep Biosystems, Odense, Denmark. These authors contributed equally: Zilu Ye, Pierre Sabatier. e-mail: yzl@ism.pumc.edu.cn; jesper.olsen@cpr.ku.dk