تثبيط العلاج المناعي للسرطان بواسطة خلايا T CD4 Tr1 السامة المحددة للنيوانتيجينات Neoantigen-specific cytotoxic Tr1 CD4 T cells suppress cancer immunotherapy
يمكن أن تعزز الخلايا أو تثبط مناعة الورم. على الرغم من أن الخلايا التائية التنظيمية معروفة منذ فترة طويلة بأنها تعيق الاستجابات المضادة للورم.آخرتم الإشارة مؤخرًا إلى أن الخلايا تلعب دورًا في تثبيط هذه الاستجابة. ومع ذلك، لا تزال طبيعة ووظيفة الأخيرة غير واضحة. هنا، باستخدام لقاحات تحتوي على نيوأنتيجينات من فئة MHC I (MHC-I) ونسب مختلفة من نيوأنتيجينات MHC-II المستمدة من الورم، اكتشفنا أنه بينما ساهمت لقاحات تحتوي على جرعات منخفضة من الببتيدات المقيدة بـ MHC-II (LDVax) في تعزيز رفض الورم، فإن اللقاحات التي تحتوي على جرعات عالية من نفس نيوأنتيجينات MHC-II (HDVax) منعت الرفض. وقد حددت خصائص الخلايا المثبطة التي تحفزها HDVax على أنها خلايا T تنظيمية من النوع 1 (Tr1) تعبر عن IL-10، وجرانزيم B، وبيرفورين، وCCL5 وLILRB4. قامت خلايا Tr1 الخاصة بالورم بقمع رفض الورم الذي تحفزه مضادات PD1، أو LDVax، أو خلايا T الفعالة الخاصة بالورم التي تم نقلها بشكل تكييفي. من الناحية الآلية، قامت خلايا Tr1 المستحثة بواسطة HDVax بقتل خلايا الدندريتية التقليدية من النوع 1 (cDC1s) التي تقدم مستضدات ورمية من MHC-II بشكل انتقائي، مما أدى إلى انخفاض أعداد cDC1s في الأورام. ثم وثقنا طرقًا للتغلب على هذا التثبيط، تحديدًا من خلال حجب LILRB4، باستخدام ميوتيني IL-2 موجه نحو CD8، أو فقدان مستهدف لـ cDC2/وحيدات النواة. بشكل جماعي، تظهر هذه البيانات أن خلايا Tr1 السامة، التي تحافظ على التسامح المحيطي، تعيق أيضًا الاستجابات المضادة للورم وبالتالي تعمل على إعاقة السيطرة المناعية على السرطان.
نتيجة مرغوبة من العلاج المناعي للسرطان هي إنتاج مستضدات محددة للأورامالخلايا التائية السيتوليتية (CTLs) القادرة على تدمير الأورام. بينما يتم تعزيز تحفيز هذه الخلايا الفعالة غالبًا بواسطة ‘المساعدين’ خلايا يمكن أن يتم تثبيط هذه العملية بواسطةتنظيمي خلايا تشير الدراسات الحديثة إلى أن الأساليب التقليديةقد لا تكون الخلايا هي الوحيدة المثبطة للمناعةسكان خلايا T المقيمين في الأورام التي تنمو بشكل متزايد“، لكن تحديد أصول وطبيعة ووظيفة الأخيرة كان تحديًا. وقد وضع هذا قيودًا نظرية وعملية على العلاج المناعي للسرطان بشكل عام وعلى لقاحات النيوأنتيجين الخاصة بالسرطان بشكل خاص. قدرة CD4تم إثبات أن خلايا Tr1 تحافظ على التسامح في المناعة الذاتية والعدوى المزمنة.وقد تم اقتراح أن تساهم في تثبيط المناعة المرتبطة بالسرطانومع ذلك، لم يتم تعريف خلايا Tr1 بناءً على عامل النسخ الرئيسي، بل تم التعرف عليها على أنها خلايا مثبطة تنتج IL-10 وتكون سلبية لـ FOXP3.. هنا نستخدم نماذج ساركوما الفئران المتجانسة والمميزة لدينا لإظهار أنه، بينما تعزز لقاحات المستضدات الجديدة (SLP) الاصطناعية التي تحتوي على مستضدات جديدة محددة للورم من نوع MHC-I ومدخلات منخفضة من مستضدات جديدة محددة للورم من نوع MHC-II بشكل فعال رفض الورم، فإن اللقاحات المماثلة التعبير عن كميات كبيرة من نفس مستضدات MHC-II الجديدة بشكل غير متوقع يؤدي إلى تثبيط المناعةخلايا متميزة عنالخلايا التي تثبط القضاء على الورم. نحن نميز هذه الخلايا كخلايا Tr1 سيتوليتية، ونقدم رؤى حول تطورها وآليات عملها، ونعتمد استراتيجيات لتجاوز نشاطها المثبط. نحن نظهر أيضًا أن هناك CD4 مشابهةيمكن العثور على مجموعة خلايا T في الأورام المتزايدة في الفئران حيث تصبح غير حساسة لعلاج anti-PD1 وفي المرضى البشر المصابين بالسرطان الذين يستجيبون بشكل ضعيف لعلاج نقاط التفتيش المناعية (ICT) ولقاحات السرطان. نحن نثبت أن القضاء على هذه الخلايا أو تثبيط وظيفتها يجعل العلاج المناعي للسرطان أكثر فعالية. وبالتالي، لا توثق هذه الدراسة فقط وظيفة جديدة لـ CD4 المعروف.تحت مجموعة خلايا T ولكنها توفر أيضًا استراتيجية لتجاوز تأثيراتها المثبطة على رفض الورم.
ملفات جرعات معقدة من MHC-II بواسطة لقاحات الببتيد
لقد حددنا سابقًا المستضدات الجديدة الفريدة من نوع MHC-I و MHC-II المعبر عنها في ثلاثة من خطوط ساركوما الميثيل كولانثرين (MCA) الخاصة بنا.
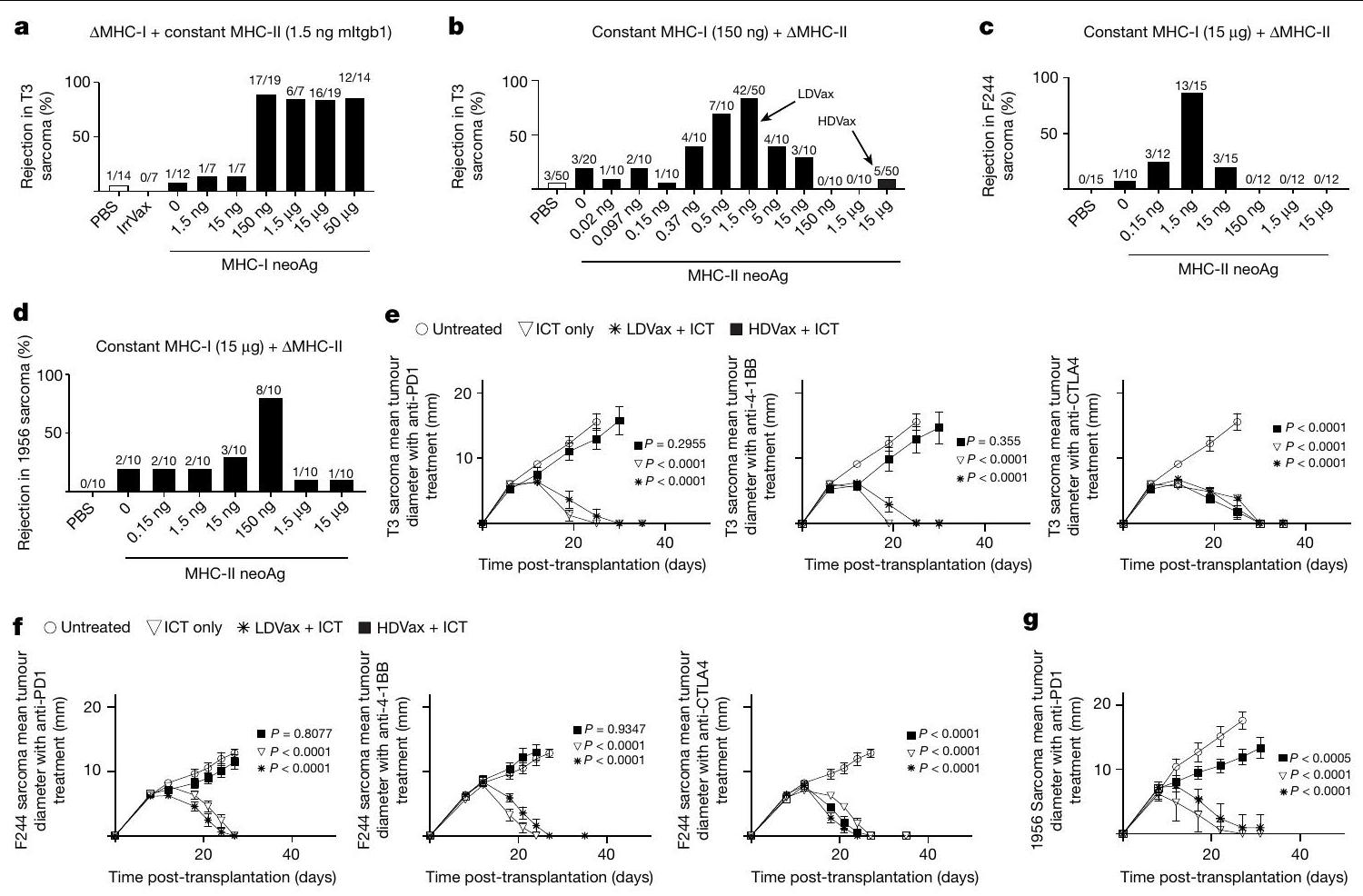
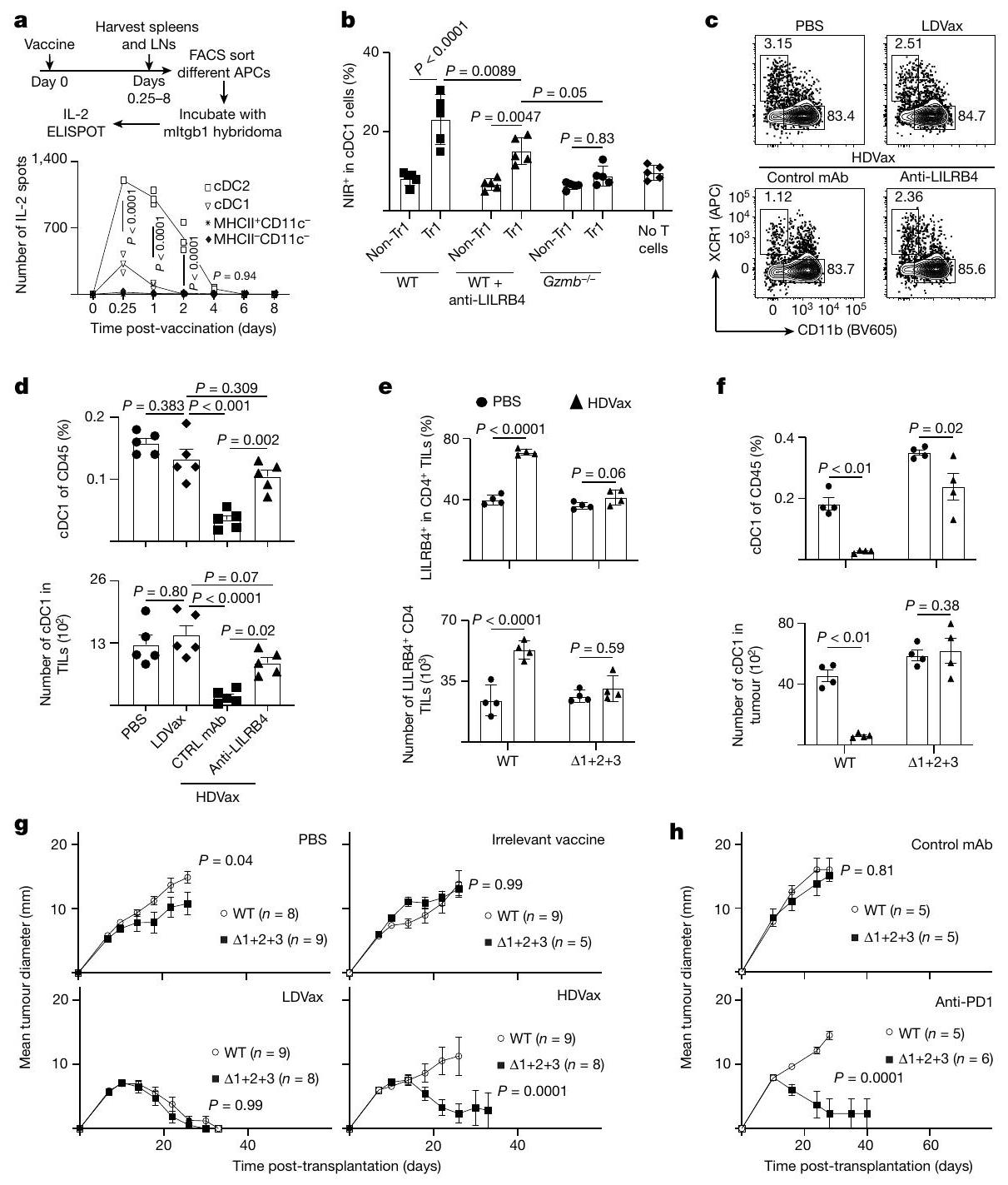
الشكل 1 | جرعات عالية من neoAg MHC-II تثبط فعالية اللقاحات SLP المضادة للورم وبعض ICTs. أ، نسبة بقاء الفئران الحاملة لورم T3 بعد العلاجات باستخدام لقاحات تحتوي على جرعات ثابتة (1.5 نانوغرام) من SLP mItgb1 بالإضافة إلى جرعات مختلفة من SLP mLama4/mAlg8 أو IrrVax. ب، نسبة بقاء الفئران الحاملة لورم T3 بعد العلاجات باستخدام لقاحات تحتوي على جرعات ثابتة منكل بالإضافة إلى جرعات مختلفة من mItgb1SLP.c، نسبة بقاء الفئران الحاملة لورم F244 بعد العلاج باللقاحات التي تحتوي على جرعات ثابتة من mPex14 SLP ( لكل فأر) بالإضافة إلى جرعات مختلفة من mPlec SLP. د، نسبة بقاء الفئران الحاملة للورم 1956 بعد العلاج باللقاحات التي تحتوي على جرعات ثابتة من لكل فأر) بالإضافة إلى جرعات مختلفة من mCs SLP أو IrrVax. الأرقام الموجودة فوق كل عمود تمثل عدد الفئران التي رفضت الأورام من إجمالي عدد الفئران المستخدمة. (الجدول التكميلي 1) أظهر أن الرفض المناعي لكل ورم يعتمد علىواستجابات الخلايا تجاه النيوأنتيجينات المعبر عنها بشكل فريد في كل خط ورمي، وأظهرت أيضًا أن الفعالية المثلى للرفض تتطلب وجود كل من النيوأنتيجينات MHC-I وMHC-II في الأورام واللقاحات.لقد أثارت اهتمامنا التأثيرات العميقة التيالخلايا كانت علىالمناعة الوقائية المعتمدة على الخلايا ضد ساركوما MCA التي لا تعبر عن بروتينات MHC-II، وبالتالي قررنا دراسة الاعتماد المتبادل بين استجابات neoAg لمستضدات MHC-I وMHC-II في تعزيز الفعالية المضادة للورم. اقتربنا من هذا السؤال من خلال تحديد جرعات neoAg لمستضدات MHC-I وMHC-II سواء بشكل منفرد أو معًا التي تحفز الفعالية المضادة للورم بشكل مثالي. كانت لقاحات SLP الخاصة بـ T3 التي تتكون فقط من MHC-I الخاص بـ T3 (G1254V mLama4 بالإضافة إلى A506T mAlg8) أو MHC-II (N710Y mItgb1) بالإضافة إلى مساعد poly-ICLC أو اللقاحات غير ذات الصلة التي تحتوي على SLP لفيروس الورم الحليمي البشري (HPV) (IrrVax) غير فعالة في تحفيز رفض T3 عند إعطائها بعد 6 أيام من حقن خلايا الورم (الشكل 1a، b). ومع ذلك، كانت اللقاحات الخاصة بـ T3 التي تحتوي على neoAgs لمستضدات MHC-I بجرعات تتراوح بين 150 نانوغرام ومع الجرعات المنخفضة المستمرة من mItgb1 SLP (1.5 نانوغرام) تم تحفيز رفض الورم في nearly جميع الفئران الحاملة للورم (الشكل 1a، b). بالمقابل، عندما تم تثبيت جرعات SLPs الخاصة بـ T3 MHC-I neoAg عند 150 نانوغرام وتم تغيير جرعة SLPs الخاصة بـ T3 MHC-II neoAg، لوحظ منحنى استجابة جرعة على شكل جرس يظهر أقصى حماية في 42 من 50 من الفئران الملقحة عند جرعات 1.5 نانوغرام من-لقاح mltgb1 SLP المقيد (المشار إليه فيما بعد باسم LDVax؛ الشكل 1b). اللقاحات التي تحتوي على نفس جرعة SLP MHC-I المحددة لـ T3 بالإضافة إلى جرعات أعلى من SLP mItgb1، على سبيل المثال، 150 نانوغرام، أدى إلى القليل أو عدم وجود فعالية مضادة للورم (يشار إليها فيما بعد باسم HDVax). وبالتالي، فإن الجرعات العالية من MHC-IISLP المحددة للورم تجعل اللقاحات المحددة لـ T3 غير فعالة.
قابلية تعميم عدم فعالية لقاح HDV
لمعالجة ما إذا كانت عدم فعالية لقاح T3 المحدد لـ HDVax قابلة للتعميم على أورام أخرى متميزة من الناحية المستضدية، استخدمنا لقاحات تحتوي على المستضدات الجديدة الرئيسية المقيدة بمركب التوافق النسيجي الرئيسي من الفئة الأولى (MHC-I) والفئة الثانية (MHC-II) لنموذجي ساركوما MCA آخرين: F244 (الذي يعبر عن -طفرة G122A المقيدة في بروتين الغشاء البيروكسيزومي 14 (mPex14) و D2944YI-A-طفرة مقيدة في بليكتين-1 (mPlec) و1956 (تعبير عن -متحور M 50 I المقيد في الوحدة الفرعية للبروتيازوم 26S، غير ATPase 6 (mPsmd6) و I-A-طفرة R144S المقيدة في سينثاز السترات (mCs) (الجدول التكميلي 1). كما هو موضح مع HDVax المحدد لـ T3 وأورام T3، تم ملاحظة نفس نوع المنحنى على شكل جرس لمستضدات النيو (MHC-II) مع أورام F244 و1956 وHDVax المحدد لها (الشكل 1c,d).
HDVax يثبط فعالية ICT المضادة للورم
لتحديد ما إذا كان HDVax يثبط العلاج المناعي بنشاط، قمنا بتقييم تأثير T3-HDVax على فعالية الأجسام المضادة المضادة لـ PD1 و41BB أو CTLA4، حيث أن كل من هذه الأجسام المضادة، عند إعطائها كعلاج وحيد، أدت إلى رفض كامل للأورام T3 في الجسم الحي.
الشكل 2 | تثبيط HDVax محدد لمستضدات النيو (MHC-II) في الورم. أ، نمو ورم T3 و F244 في الجانبين المتقابلين من ذكور الفئران 129S6 WT التي تم تطعيمها وعلاجها بمضاد PD1 (، ممثل لتجربتين). D0، اليومنمو الورم فيالفئران التي تتلقى خلايا T الكلية ) من الفئران المعالجة بـ LDVax فقط أو بالإضافة إلى CD25-negative mltgb1-specific CD4 خلايا Tمرتبة من الفئران المعالجة بـ HDVaxالبيانات في اللوحاتيتم عرضها كمتوسط قطر الورمتم إجراء التحليل الإحصائي باستخدام تحليل التباين الثنائي، مع تصحيح المقارنات المتعددة باستخدام اختبار المقارنات المتعددة لسيداك. ج-هـ، CD4تم تنقية خلايا T وتحفيزها كما هو موضح. تم تقييم إنتاج IL-2 في السائل الفائق باستخدام MultiPlex (، المجمعة من تجربتين؛ د). تُظهر خريطة الحرارة مستويات إنتاج السيتوكينات المختلفة (المشار إليها على المحور؛ هـ). الأرقام في اللوحة تمثل متوسط نسختين بيولوجيتين مستقلتين (تمثل تجربتين). التردد ( ) والأرقام ( ) من -خلايا CD8 المحددةالخلايا التائية المستحثة في الأورام من النوع T3 التي تم جمعها في اليوم الثالث عشر، ممثل لثلاث تجارب) موضحة أيضًا. البيانات في الألواح د، ف، ج معبر عنها كمتوسط s.e.m.; تم إجراء الإحصائيات باستخدام تحليل التباين الأحادي، مع تصحيح المقارنات المتعددة باستخدام طريقة توكي. (الشكل 1e). لم يؤثر دمج ICT و LDVax على فعالية الأجسام المضادة المعطلة للأورام. على العكس من ذلك، ألغى T3-HDVax رفض الورم في جميع الفئران المعالجة بـ anti-PD1 أو anti-4-1BB، لكنه لم يلغي رفض الورم الناتج عن anti-CTLA4. تم الحصول على نتائج مماثلة باستخدام نماذج الورم F244 و 1956 ولقاحاتها النيوأنتيجينية المعنية (الشكل 1f,g)، مما يدل على أن علاج HDVax يمنع أنواع معينة من ICT. وقد تم تفسير الملاحظة الأخيرة من خلال التجارب اللاحقة.
خصوصية المستضد ل inhibiting HDVax
لاختبار خصوصية المستضد لتثبيط HDVax، تم حقن خلايا ساركوما T3 وخلايا ساركوما F244 المتطابقة جنسيًا والمختلفة من الناحية المستضدية في جوانب متقابلة من فئران 129S6 البرية (WT) غير المدربة. بعد ثلاثة أيام، تم علاج الفئران بمضاد PD1 بالإضافة إلى إما HPV-LDVax غير ذي الصلة أو HPV-HDVax، T3-LDVax أو T3-HDVax، أو F244-LDVax أو
تمت مراقبة نمو الورم باستخدام F244-HDVax. لم يمنع كل من LDVax أو علاج HPV-HDVax غير ذي الصلة رفض الأورام T3 أو F244 الناتج عن anti-PD1 (الشكل 2a، الألواح الثلاثة العليا واللوحة السفلى اليسرى). ومع ذلك، فإن علاج T3-HDVax منع بشكل انتقائي رفض الأورام T3 المعتمد على anti-PD1 ولكن ليس الأورام F244 (الشكل 2a، اللوحة السفلى الوسطى). وعلى العكس، فإن علاج F244-HDVax منع بشكل انتقائي رفض الأورام F244 المعتمد على anti-PD1 ولكن ليس الأورام T3 (الشكل 2a، اللوحة السفلى اليمنى). وبالتالي، فإن التأثير المثبط لـ HDVax مرتبط بشكل خاص بـ MHC-II neoAg المعبر عنه بواسطة كل خط ورمي.
الناجم عن HDVaxتقوم خلايا T بتثبيط رفض الورم
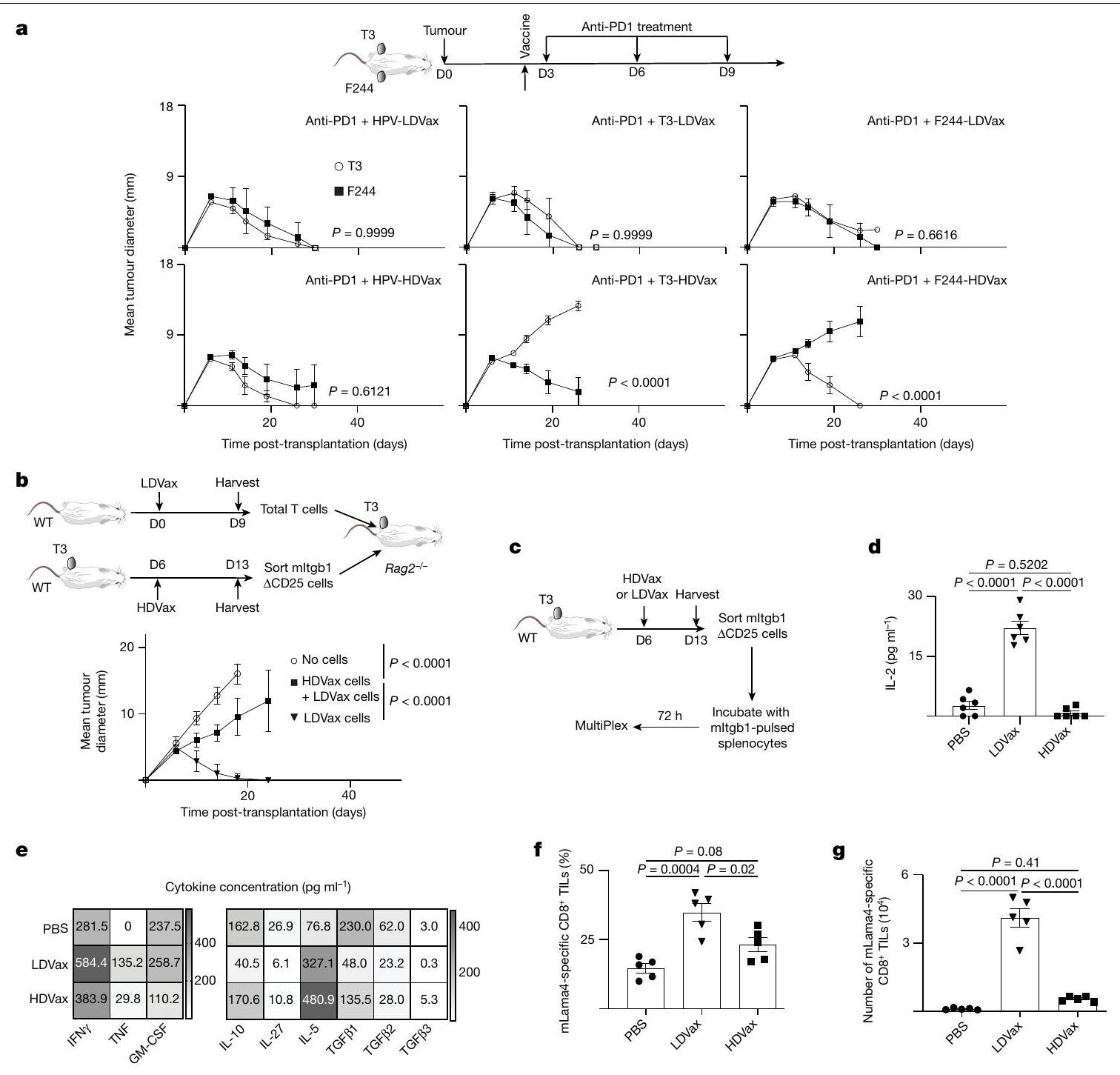
لاختبار رسمي ما إذا كان التثبيط ناتجًا عن تحفيز مثبط للمناعةالخلايا، أنشأنا نظام نقل خلايا T التبني (ACT) في Rag2الفئران (انظر الطرق). ACT منمستنفد من الخلايا نقل خلايا T من فئران WT المعالجة بـ LDVax إلى فئران Rag2-/- الحاملة لورم T3 والمناعية الناقصة كان كافياً لتحفيز رفض T3 (الشكل 2b). ومع ذلك، فإن خلط خلايا T المستحثة بواسطة HDVax،CD4 محدد mItgb1 خالي من الخلاياالخلايا التائية مع خلايا التأثير المستحثة بواسطة LDVax ألغت رفض ورم T3. هذه البيانات اقترحت أن HDVax يحفز سكان الخلايا متميز عن الكلاسيكيالخلايا، التي تمتلك نشاطًا مثبطًا للمناعة قويًا.
خصائص CD4 المستحثة بواسطة HDVaxخلايا
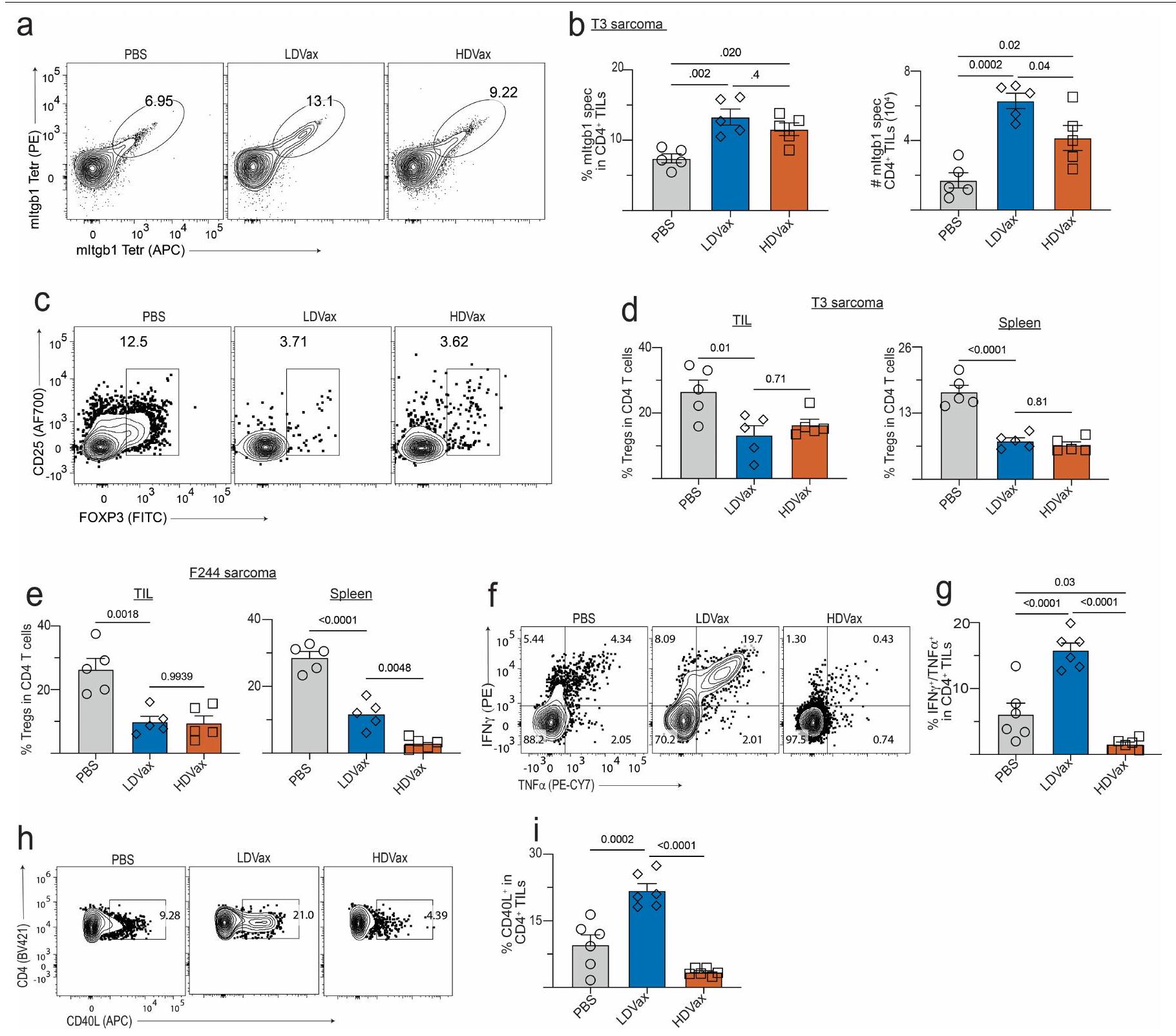
تكرار والأعداد المطلقة لخلايا CD4 المحددة بـ mItgb1كانت الخلايا اللمفاوية المتسللة إلى الورم (TILs) من الفئران المعالجة بـ HDVax أو LDVax مشابهة (الشكل البياني الموسع 1a، b). كشفت تحليلات تدفق السيتومتر أن التردد والأعداد المطلقة لـالخلايا في CD4 المحددة بـ mltgb1انخفضت TILs أو الطحال بشكل مماثل بعد علاج HDVax أو LDVax مقارنةً بالتحكمات المعالجة بـ PBS (الشكل 1c، d من البيانات الموسعة). لوحظت نتائج مماثلة مع الفئران الحاملة لورم F244 المعالجة بـ F244-HDVax أو F244-LDVax (الشكل 1e من البيانات الموسعة). كشفت تحليلات السيتوكين المتعددة عن أنماط مميزة من CD4تم تحفيز TILs بواسطة LDVax مقابل HDVax. بينما تم تحفيز CD4 الخاص بـ mItgb1 بواسطة T3-LDVaxأنتجت خلايا T IL-2 عند تحفيزها بخلايا الطحال المحملة بـ mItgb1، CD4 المحددة بـ mItgb1 الناتجة عن T3-HDVaxفشلت خلايا T في إنتاج IL-2 (الشكل 2c، d). بالإضافة إلى ذلك، بينما أدت لقاحات T3-LDVax إلى تحفيز CD4خلايا T أنتجت مستويات عالية من IFNTNF و GM-CSF (الشكل 2e)، CD4 المستحث بواسطة T3-HDVaxخلايا T أنتجت مستويات عالية من IL-10 و TGF، IL-5 و IL-27. تم تكرار هذه النتائج من خلال تحليلات صبغ الخلايا داخلية (الشكل 1f، g من البيانات الموسعة). علاوة على ذلك، بينما كانت خلايا CD4 الخاصة بـ mItgb1 تم جمع TILs من الفئران المعالجة بـ T3-LDVax والتي أظهرت مستويات مرتفعة من CD40L بعد تحفيز المستضد (LDVax: ; بي بي إس: فقط جزء صغير من CD4 الخاص بـ mItgb1 المستحث بواسطة T3-HDVax المنبه بمولد الضدتعبّر TILs عن CD40L (HDVax: ؛ الشكل التوضيحي الممتد 1h، i).
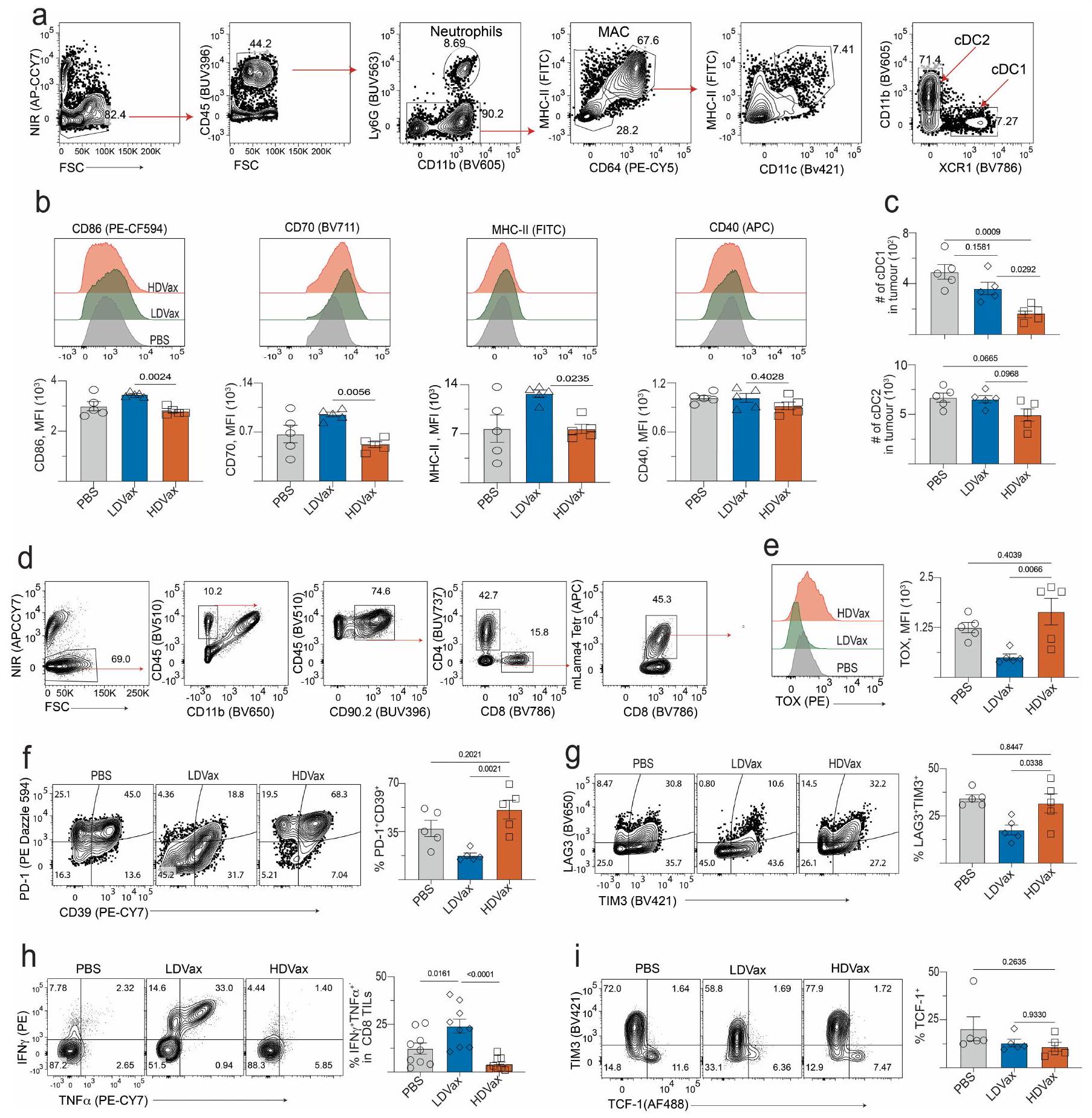
متسق مع التعبير المرتفع المعتمد على T3-LDVax لـ CD40L على CD4أظهرت خلايا الدندريت من نفس الفئران تعبيرًا أعلى قليلاً ولكنه معزز بشكل ملحوظ عن CD86 و CD70 مقارنة بالخلايا المماثلة من الفئران المعالجة بـ T3-HDVax. لم تُلاحظ فروق ذات دلالة إحصائية في مستويات التعبير عن CD40 على خلايا الدندريت (الشكل البياني الممتد 2a، b). ومن الجدير بالذكر أن عدد خلايا cDC1s قد انخفض بمقدار 2.9 مرة في الأورام من الفئران الحاملة لـ T3 المعالجة بـ T3-HDVax مقارنة بـ PBS (الشكل البياني الممتد 2c). لوحظ فقط انخفاض طفيف في عدد خلايا cDC2s، ولم يكن هذا ذا دلالة إحصائية.
توافقًا مع الاستجابة غير الطبيعية للسيتوكينات وانخفاض استعادة خلايا cDC1 من الفئران المعالجة بـ T3-HDVax، فإن الترددات والأعداد المطلقة لخلايا CD8 المحددة لـ mLama4كانت TILs أقل بكثير من CD8تم حصاد TILs من الفئران المعالجة بـ T3-LDVax (الشكل 2f، g والشكل البياني الموسع 2d). علاوة على ذلك، CD8 المحدد لـ mLama4عبّرت TILs من الفئران المعالجة بـ T3-HDVax عن مستويات عالية من علامات الإرهاق (PD1، CD39، TIM3، LAG3 و TOX) وأنتجت كميات منخفضة من IFN. و TNF مقارنة مع مجموعات سكانية مشابهة من الفئران الحاملة للأورام المعالجة بـ T3-LDVax (الشكل البياني الموسع 2d-h). لم تُلاحظ أي اختلافات في TIM3 TCF1السكان الفرعيون (الشكل 2i من البيانات الموسعة). كشفت هذه النتائج أنه بينما ينتج T3-LDVax CD4 محدد للورمتقوم خلايا T التي تنتج مواد تعزز الاستجابة المناعية لمكافحة الأورام، حيث يحفز T3-HDVax خلايا CD4 المحددة للأورام.خلايا T التي تقلل من النشاط المضاد للورم.
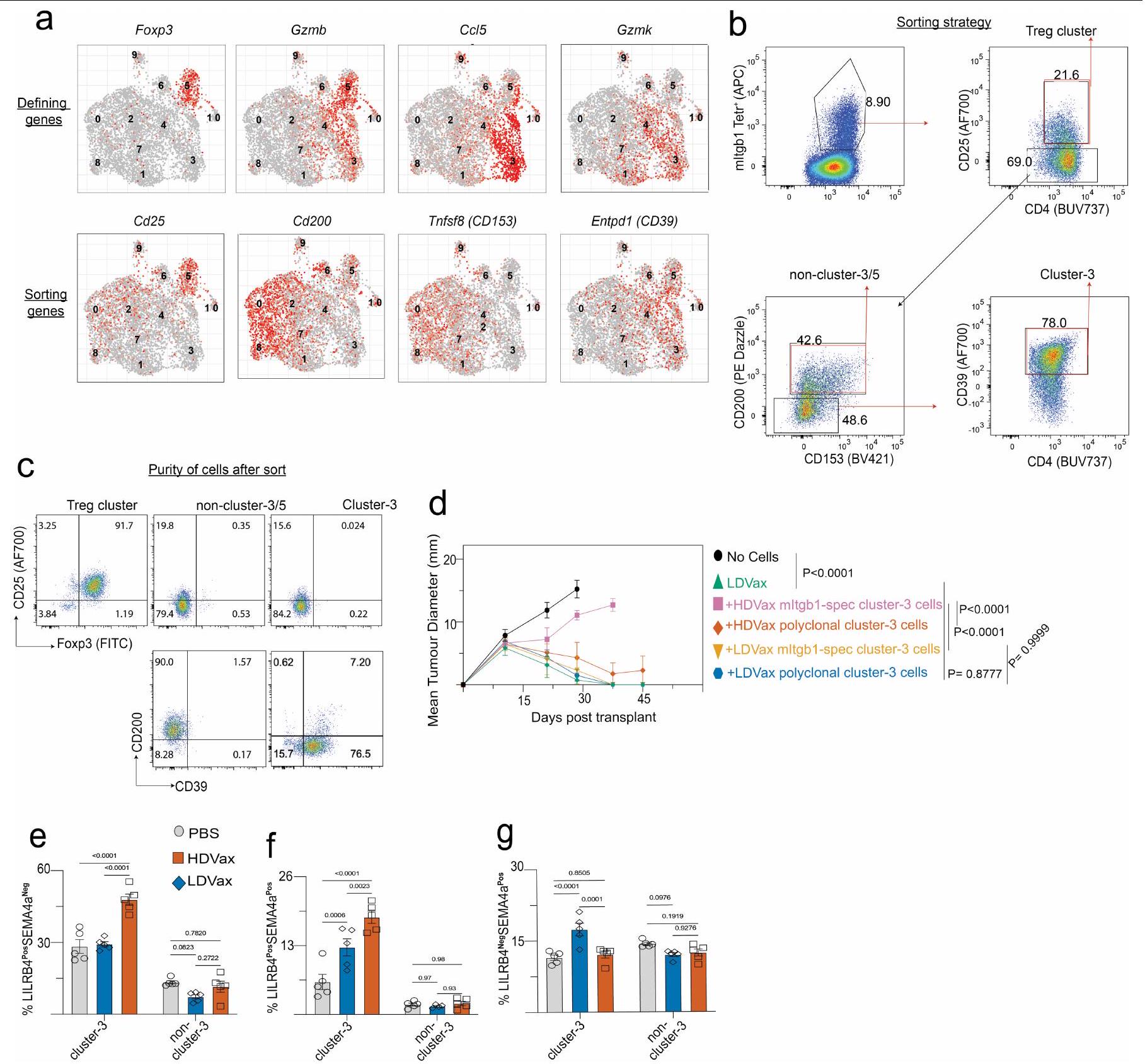
تحديد CD4 المثبطالخلايا التائية المستحثة بواسطة تسلسل RNA أحادي الخلية
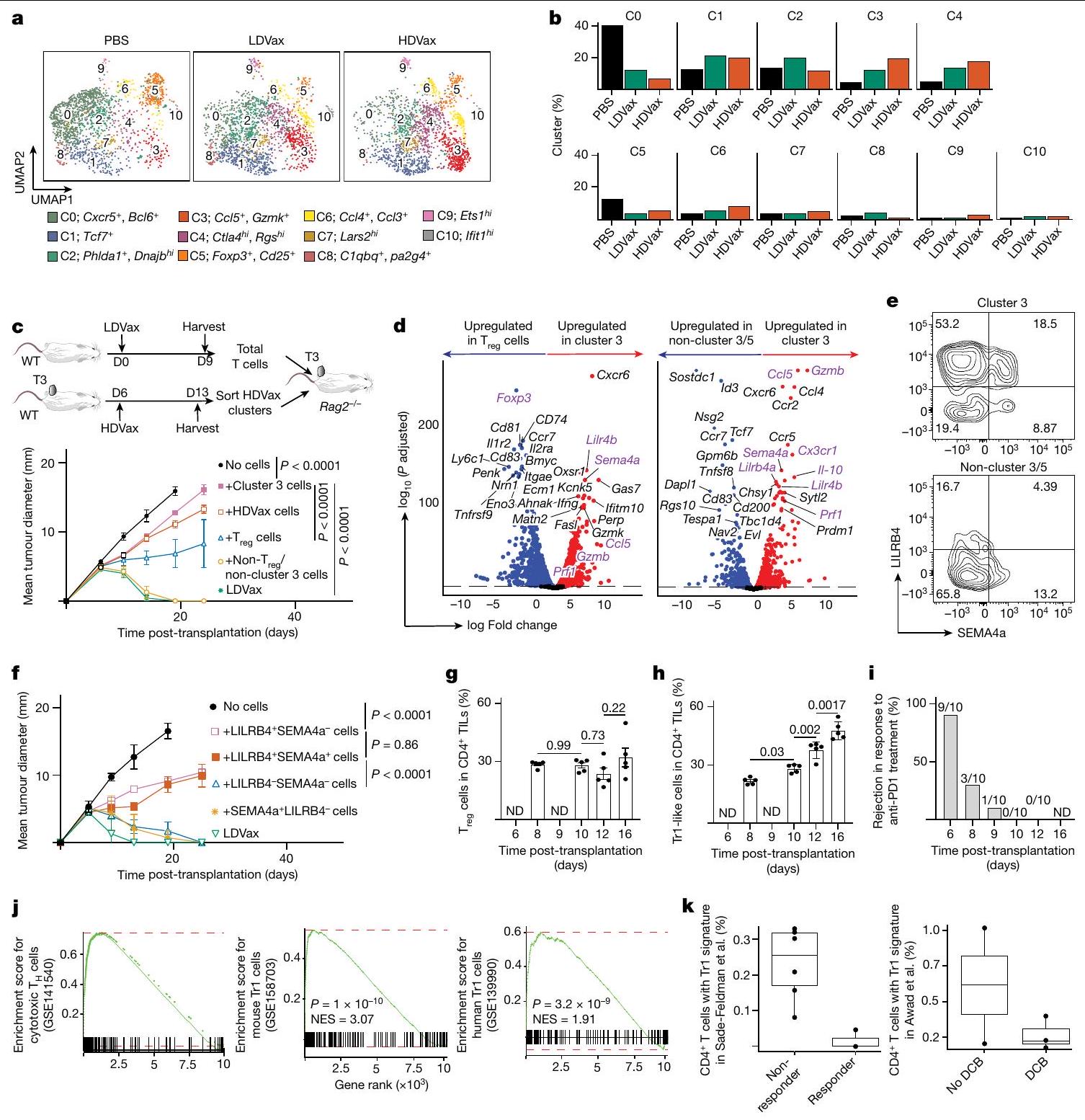
لتعريف أفضل لـ CD4 المثبط الناتج عن HDVaxخلايا T، قمنا بإجراء تسلسل RNA على مستوى الخلية الواحدة (scRNA-seq) على CD4 المحدد بـ mItgb1تيلز من الفئران المعالجة بـ T3-HDVax أو T3-LDVax أو PBS. تجميع غير متحيز باستخدام تقنيات الاقتراب والتقريب الموحد (UMAP) لخلية CD4 المحددة بـ mItgb1كشفت TILs عن 11 مجموعة خلوية (المجموعة 0 إلى المجموعة C10) بتوقيعات جينية مميزة (الشكل 3a,b). وجدنا أن أدى التطعيم بحد ذاته إلى تقليل ثلاثي في تكرارتجمعات TILs في العنقود 0 (مساعدات جريبية)مجموعة خلايا (Bcl6و Cxcr5)) و العنقود 5 ( مجموعة خلايا (Foxp3و Cd25“)) في الأورام من الفئران التي تلقت إما T3-HDVax أو T3-LDVax مقارنةً بـ PBS. على النقيض من ذلك، كانت نسب الخلايا التي تشغل المجموعات 1-4 و6 و8 متأثرة بشكل مختلف بجرعة ببتيد MHC-II المستخدمة في اللقاح. كانت المجموعتان 2 و8 أكثر كثافة بعد T3-LDVax مقارنةً بـ T3-HDVax أو علاج PBS، في حين كانت المجموعات 3 و4 و6 أكثر كثافة بعد علاج T3-HDVax مقارنةً بـ LDVax أو علاج PBS. بينما أحدث T3-HDVax زيادات طفيفة فقط في الكثافة في المجموعتين 4 و6 مقارنةً بـ T3-LDVax أو علاج PBS، كانت كثافة الخلايا في المجموعةزاد إلى أقصى حد وأظهر أكبر الفروقات بين HDVax و LDVax أو PBS.
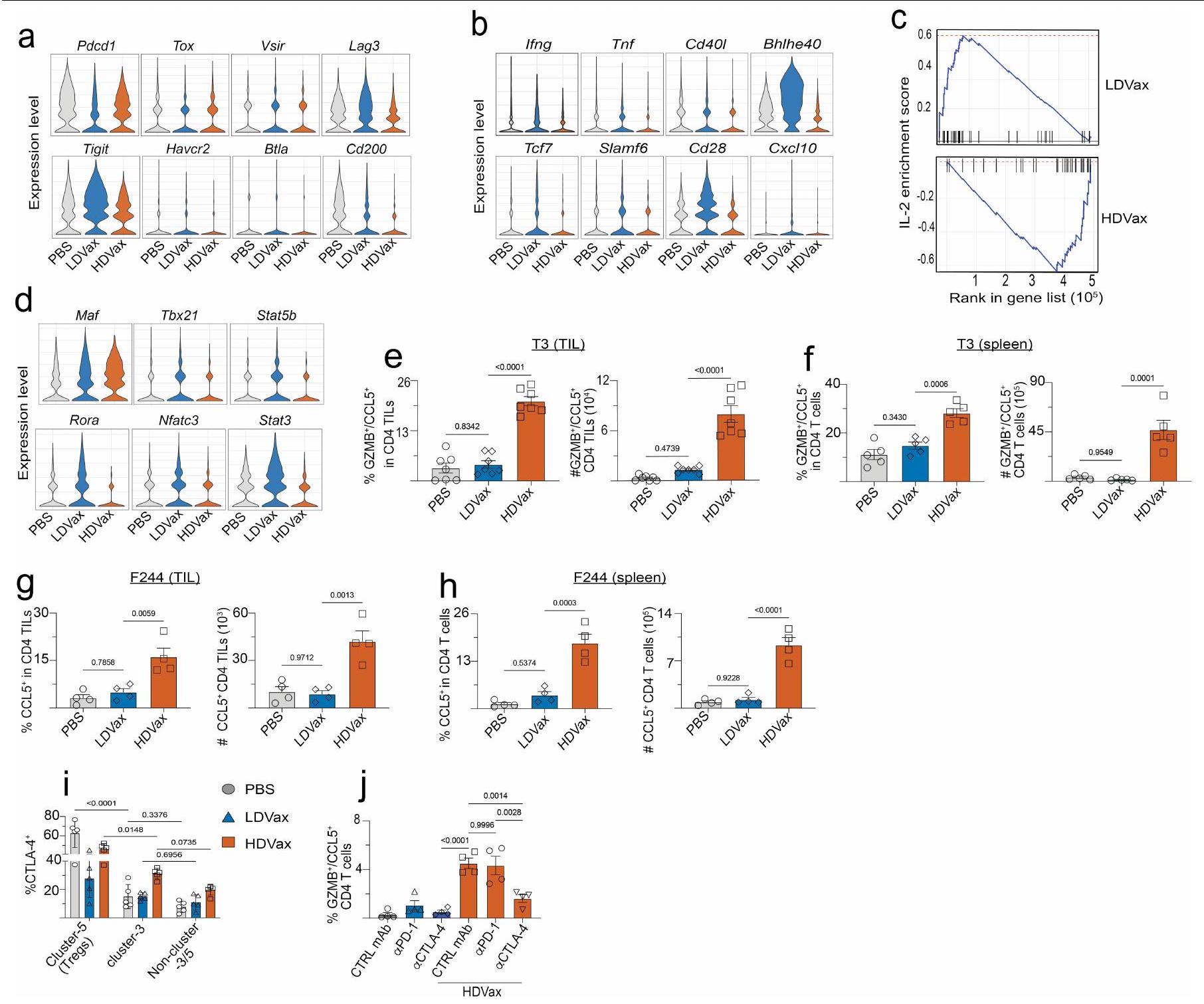
كشفت التحليلات الإضافية أن T3-HDVax أدى إلى CD4تم التعبير عن TILs بمستويات مرتفعة نسبيًا من و السير النسخ الجينية ومستويات منخفضة نسبيًا من نسخ Lag3 وCd2OO وHavcr2 وTigit مقارنة بتلك التي تم تحفيزها بواسطة T3-LDVax أو PBS (الشكل البياني الممتد 3a). CD4أظهرت الخلايا التائية من الفئران المعالجة بـ T3-HDVax أيضًا مستويات أقل من mRNA لجينات Ifng وTnf وCd28 وCd4Ol وعوامل النسخ المرتبطة بالخلايا التائية المساعدة.استقطاب الخلايا، على سبيل المثال، Bhlhe4O، مقارنةً مع TILs من الفئران المعالجة بـ T3-LDVax أو الفئران المعالجة بـ PBS (الشكل البياني الممتد 3b). كما أظهر تحليل إثراء مجموعة جينات Il2 (GSEA) أن الخلايا المستحثة بواسطة HDVa أظهرت عيوبًا في توقيع IL-2 (الشكل البياني الممتد 3c)، وأعربت عن مستويات منخفضة من عوامل النسخ مثل Stat5b وRora وNfatc، وأظهرت زيادة في التعبير عن جين cMaf الذي يثبط إنتاج IL-2.مقارنة مع الخلايا المقابلة بعد علاج T3-LDVax (الشكل البياني الممتد 3d).
للتحقق من هذه الملاحظات، قمنا بتقييم مستويات البروتينات المقابلة في خلايا CD4 المحددة للأورام.تمت دراسة TILs وسكان الخلايا الطحالية من الفئران المعالجة بـ T3-HDVax و T3-LDVax أو PBS. أكدت هذه التحليلات أنCD4 الخاص بـ mItgb1كانت TILs موجودة بترددات وأعداد أعلى بخمسة أضعاف في الفئران المعالجة بـ T3-HDVax مقارنةً بـ TILs من الفئران المعالجة بـ T3-LDVax أو PBS (الشكل البياني الموسع 3e، f). كما لوحظ فرق يتراوح بين 2-3 أضعاف في الخلايا الطحالية من الفئران التي تلقت T3-HDVax مقارنةً بـ T3-LDVax أو PBS. تم ملاحظة نتائج مماثلة باستخدام فئران تحمل ورم F244 المعالجة بـ F244-HDVax، F244-LDVax أو PBS (الشكل البياني الموسع 3g، h). في كلا نموذجَي الورم T3 وF244، كانت خلايا المجموعة 3 المستحثة بواسطة HDVax تعبر عن مستويات أعلى من بروتين CTLA4 مقارنةً بالخلايا في جميع المجموعات الأخرى، باستثناء المجموعة 5 (CD25 الكلاسيكي).الخلايا؛ الشكل البياني الممتد 3i). العلاج بمضاد CTLA4 ولكن ليس بمضاد PD1 قضى على الخلايا من المجموعة 3 والمجموعة 5 في TILs من الفئران المعالجة بـ HDVax (الشكل البياني الممتد 3j). يفسر هذا الاكتشاف لماذا لم يقم HDVax بتثبيط فعالية مضاد الأورام لمضاد CTLA4، لأن الأجسام المضادة الأحادية النسيلة لمضاد CTLA4 (9D9) المستخدمة في الدراسة تستنفد كلا منالخلايا و CD4 المثبط الناتج عن HDVaxخلايا T (الشكل 1e، f).
خلية مجموعة 3 ACT تمنع رفض الورم
على الرغم من أن هذه النتائج أشارت إلى أن خلايا HDVax المثبطة كانت محلية في المجموعة 3، إلا أنها لم تحدد البروتينات السطحية على هذه الخلايا التي ستسمح بعزلها. لذلك، قمنا بإنشاء استراتيجية فرز حيث قمنا بإزالة خلايا T من CD4 المحددة للورم.مجموعة خلايا T التي عبرت عن جينات علامات السطح غير الموجودة في خلايا المجموعة 3 (أي Cd25 و Cd200 و Cd153)، وتم فرز الخلايا المتبقية إيجابيًا للتعبير عن CD39 (الشكل البياني الممتد 4a). سمح لنا هذا النهج بإنشاء ثلاث مجموعات خلوية: (1) خلايا المجموعة 3 المنقاة جزئيًا (CD25CD200و CD39“), (2) CD25عنقود خلايا (أكثر من فوكس بي)، و (3) غير العنقوديالخلايا (CD25CD200CD153و“; الشكل البياني الموسع 4b، c). تم اختبار هذه المجموعات الخلوية بعد ذلك لقدرتها على تثبيط رفض T3. تم رفض أورام T3 في الفئران Rag2-/ التي تتلقىخلايا T الفعالة المستحثة بواسطة T3-LDVax التي تفتقر إلى الخلايا، سواء بمفردها أو بالاشتراك مع خلايا غير متجمعة محددة T3 المستحثة بواسطة HDVax
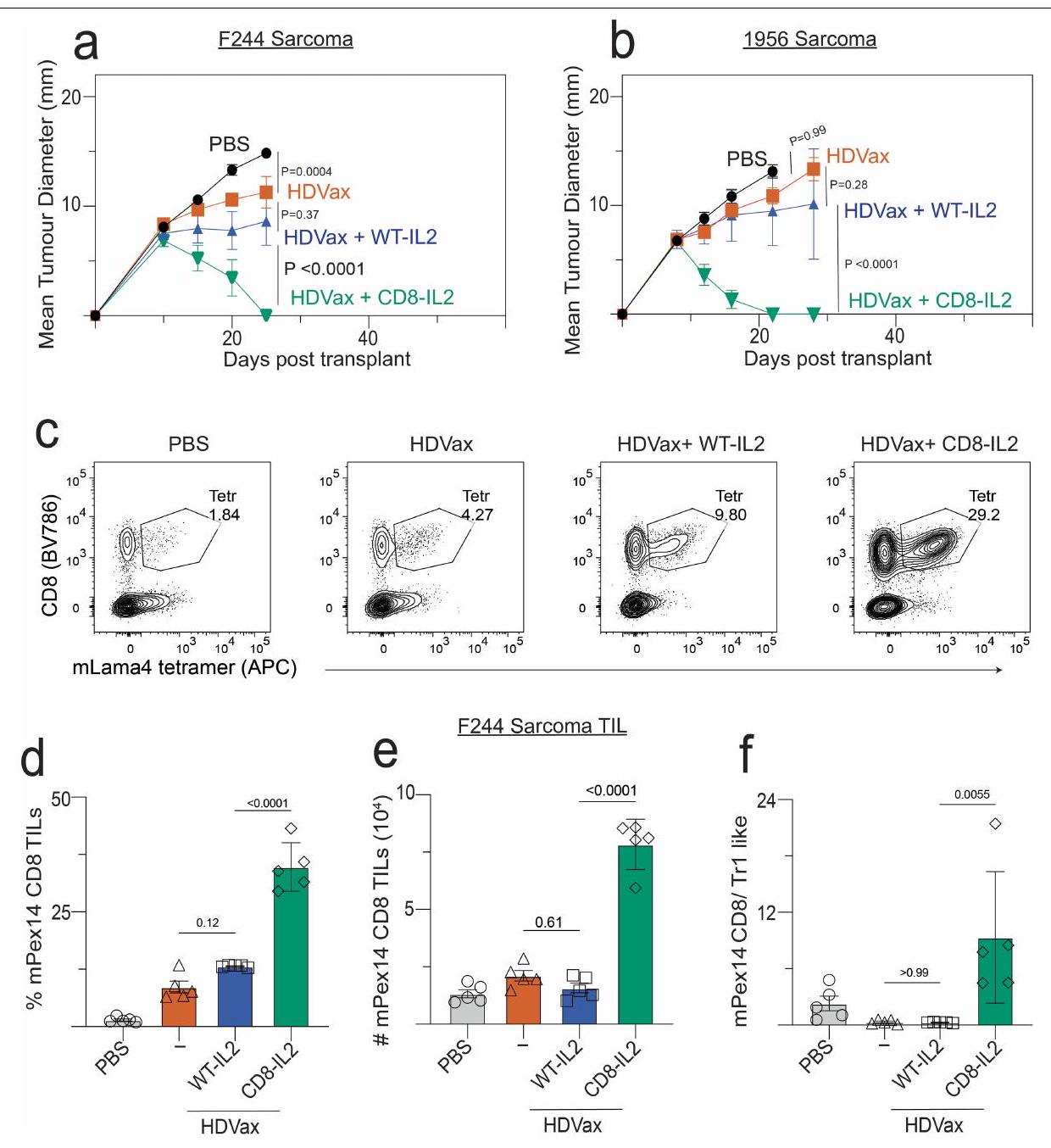
الشكل 3 | الخلايا المستحثة بواسطة HDVax هي خلايا Tr1 سامة للخلايا والتي تعتبر ضرورية وكافية لتثبيط HDVax. أ، UMAP يظهر مجموعات فردية في I--مستضد CD4 الخاص بـ mItgb1تحت ظروف مختلفة. ب، تكرار الخلايا التي تشغل مجموعات فردية. ج، نمو ورم T3 فيالفئران التي تلقت خلايا T من فئران LDVax بالإضافة إلى إجمالي CD25 CD4الخلايا (تقريبًا ) أو مجموعات فرعية مختلفة (حوالي ) مرتبة من الفئران المعالجة بـ HDVax ( ). د، مخطط البركان لبيانات تسلسل RNA الكمي المقارن للتعبير الجيني في خلايا المجموعة 3 بالنسبة إلى الخلايا (يسار) وغير المجمعةالخلايا (يمين).تم الحصول على القيم بعد تطبيق أداة DESeq 2. رسم بياني تمثيلي لتدفق الخلايا يظهر تعبير LILRB4 و SEMA4a في CD4 المحدد بواسطة mItgb1 المستحث بواسطة HDVax.الخلايا التائية في الأورام من النوع T3. نمو الورم من النوع T3 في الفئران Rag2-/- التي تتلقى خلايا T من فئران LDVax ومجموعات فرعية مختلفة (تقريبًا ) مرتبة من الفئران المعالجة بـ HDVax بناءً على تعبير LILRB4 و SEMA4a ( (PBS و LDVax) و للمجموعات الأخرى).تكرارالخلايا (غ) أو CD4 المعبر عن LILRB4 TILs (h) في أورام T3 التي تم جمعها في عدة أيام بعد حقن الورم ( ). ND، لم يتم الانتهاء. أنا، نسبة الفئران الحاملة لورم T3 التي نجت بعد ثلاث جرعات من مضاد PD1 بدأت في أيام مختلفة ( أو 12) زراعة ما بعد الورم، تليها جرعتان لاحقتان تُحقن كل 3 أيام. الأرقام الموجودة فوق كل عمود تمثل عدد الفئران التي رفضت الـ
3/5 خلايا (الشكل 3ج). بالمقابل، لم ترفض خلطات خلايا T الفعالة المستحثة بواسطة LDVax بالإضافة إلى خلايا المجموعة 3 المحددة بواسطة HDVax الأورام T3 (الشكل 3ج). تحتوي الخلطات المماثلة على مجموعة كلاسيكية الأورام على إجمالي الفئران المستخدمة المجمعة من تجربتين. البيانات في الألواح ج، و هي معبّرة كمتوسط قطر الورم س.م. ويمثلان ثلاثة واثنين من التجارب، على التوالي. تم إجراء الإحصائيات باستخدام تحليل التباين الثنائي، مع تصحيح المقارنات المتعددة باستخدام اختبار المقارنة المتعددة لسيداك. البيانات في الألواح تُعبر كمتوسط s.e.m. تمثل تجربتين؛ تم إجراء الإحصائيات باستخدام ANOVA أحادي الاتجاه، مع تصحيح المقارنات المتعددة باستخدام طريقة توكي. ج، GSEA مقارنة توقيع الجين لخلايا الكتلة 3 (تسلسل RNA الكلي) بتوقيع الجين لمختلف مجموعات الخلايا. الخط الأحمر يظهر أقصى انحراف عن الصفر للمنحنى، والذي يعادل بشكل أساسي درجة الإثراء في هذه الحالات. تم استخدام درجة الإثراء لتحديد تحديد القيمة لكل مجموعة جينات بواسطة اختبار التبديل. NES، درجة الإثراء المنظم. k، مخطط صندوقي يوضح تكرار خلايا Tr1-like بين المستجيبين وغير المستجيبين ضمن مجموعة بيانات Sade-Feldman وآخرون (يسار) والفوائد السريرية المستدامة (DCBs) أو غير DCBs ضمن مجموعة بيانات Awad وآخرون (يمين). تم إجراء الإحصائيات باستخدام-اختبارات دون افتراض تساوي التباينات. الـالقيم في مجموعة بيانات سادي-فيلدمن وآخرون كانت، د.ف.؛ لم يتم العثور على فرق كبير في مجموعة بيانات عوض وآخرون (تُعرض البيانات كرسوم بيانية صندوقية تمتد من النسبة المئوية 25 إلى 75، مع وجود الوسيط في المنتصف والشعيرات التي تت correspond إلى القيم الدنيا والقصوى. الخلايا بدلاً من مجموعة 3 من الخلايا التي تم تثبيطها جزئيًا فقط في رفض الورم. كما هو متوقع، فإن خلطات من خلايا T الفعالة المستحثة بواسطة T3-LDVax بالإضافة إلى إما خلايا مجموعة 3 المحددة بواسطة LDVax أو متعددة النسائل لم تقم خلايا T من الفئران المعالجة بـ T3-HDVax والتي تفتقر إلى خصوصية mItgb1 بقمع رفض T3 (الشكل 4d من البيانات الموسعة). وبالتالي، فإن الخلايا المستحثة بواسطة HDVax والمخصصة للورم التي تشغل العنقود 3 ضرورية وكافية لتسهيل تثبيط الفعالية المضادة للورم.
LILRB4 هو علامة مثبطةخلايا
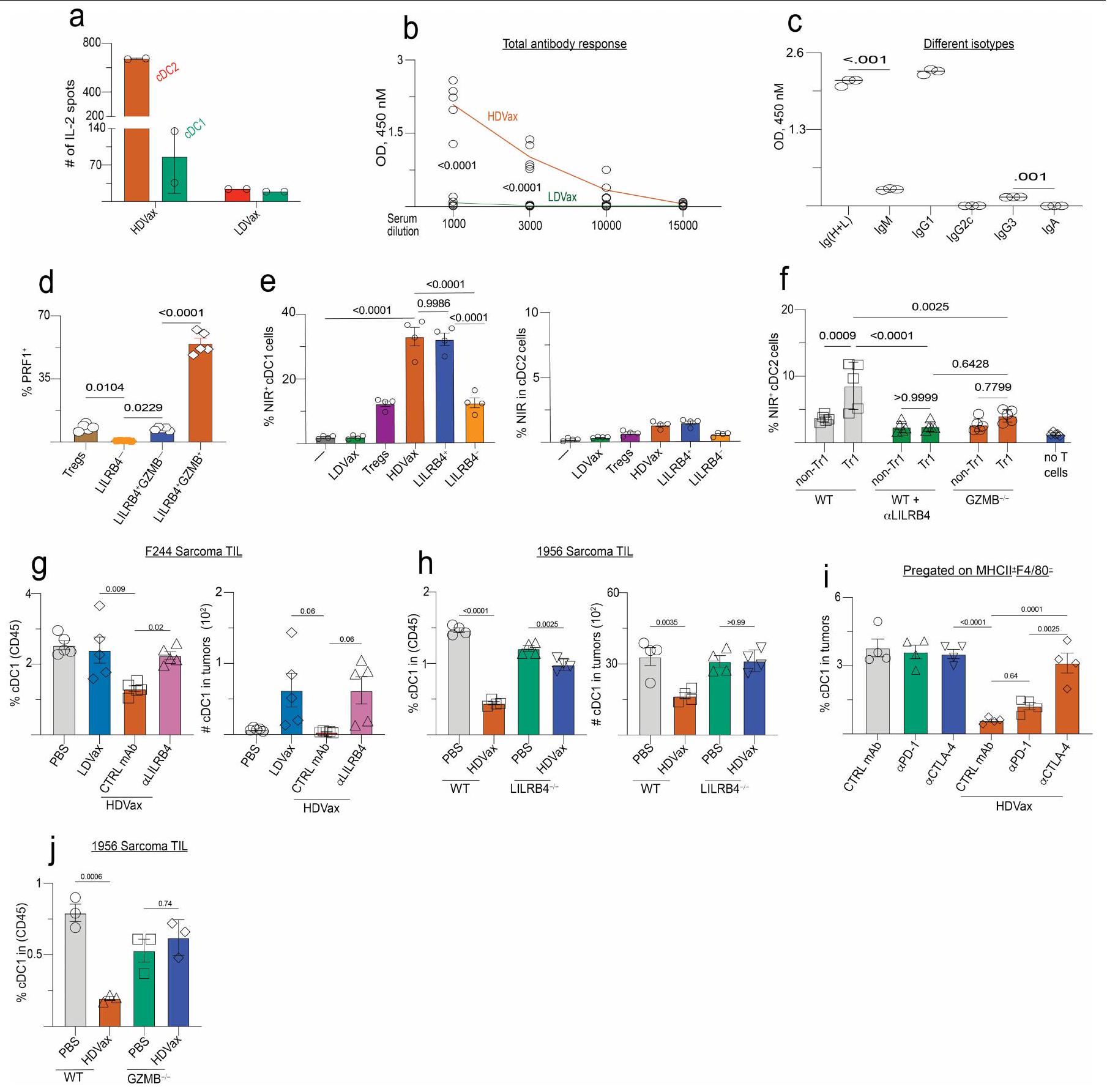
لتعريف أفضل للخلايا المثبطة للمناعة التي تشكل العنقود 3 واستخلاص رؤى حول أصولها وآلية عملها، قمنا بتعريض العنقود 3 المستخلص من HDVax، والعنقود 5 (الخلايا) وغير المجمعةتمت مقارنة تجمعات الخلايا مع تسلسل RNA العميق وقارنت النسخ الجينية لها. أكدت تحليلات مخطط البركان أن خلايا المجموعة 3 تعبر عن مستويات عالية من Gzmb و Ccl5 (الشكل 3d). لم يتم مشاركة هذه السمة من قبل خلايا أو غير متجمعة الخلايا. أظهرت خلايا المجموعة 3 المستحثة بواسطة T3-HDVax أيضًا زيادة انتقائية في التعبير عن علامتين سطحتين معروفتين بأن لهما أنشطة مثبطة للمناعة: Lilrb4 (الذي يشفر ILT3، CD85k أو gp49b)وسما4اأظهرت تحليلات تدفق الخلايا أن خلايا HDVax المجموعة 3 يمكن تقسيمها إلى أربع تحت مجموعات بناءً على التعبير الفردي أو المزدوج لبروتينات LILRB4 وSEMA4a (الشكل 3e والشكل الإضافي 4e-g). نقل الخلايا المعتمد على HDVax المحفز لـ LILRB4SEMA4aأو LILRB4سيما4CD4 المحدد لـ mItgb1تمت إزالة خلايا T بشكل مماثل لرفض ورم T3 في اختبار Rag2-/ ACT (الشكل 3f). بالمقابل، لم تقم الخلايا الإيجابية الوحيدة لـ SEMA4a أو الخلايا السلبية المزدوجة بتثبيط رفض الورم.
بينما أسست هذه النتائج علامات سطحية حددت الخلايا المثبطة، تساءلنا عما إذا كانت موجودة في الأورام المتزايدة بشكل تدريجي التي أصبحت غير حساسة لمضاد PD1. في الواقع، زادت نسبة TILs FOXP3_LILRB4 + في الأورام T3 المتزايدة بشكل تدريجي وغير المعالجة (الشكل 3g، h)، وارتبط ظهورها ببدء المقاومة لمضاد PD1 (الشكل 3i). زيادات مماثلة فيلم يتم ملاحظة الخلايا تحت هذه الظروف. تكشف هذه النتائج، جنبًا إلى جنب مع بيانات تسلسل RNA أحادي الخلية، أن تعبير LILRB4 يحدد غير-CD4 المثبط للخلاياسكان خلايا T التي تحفزها HDVax والتي تتطور أيضًا في الأورام المتزايدة بشكل تدريجي غير المعالجة حيث تتطور مقاومة للعلاج المناعي.
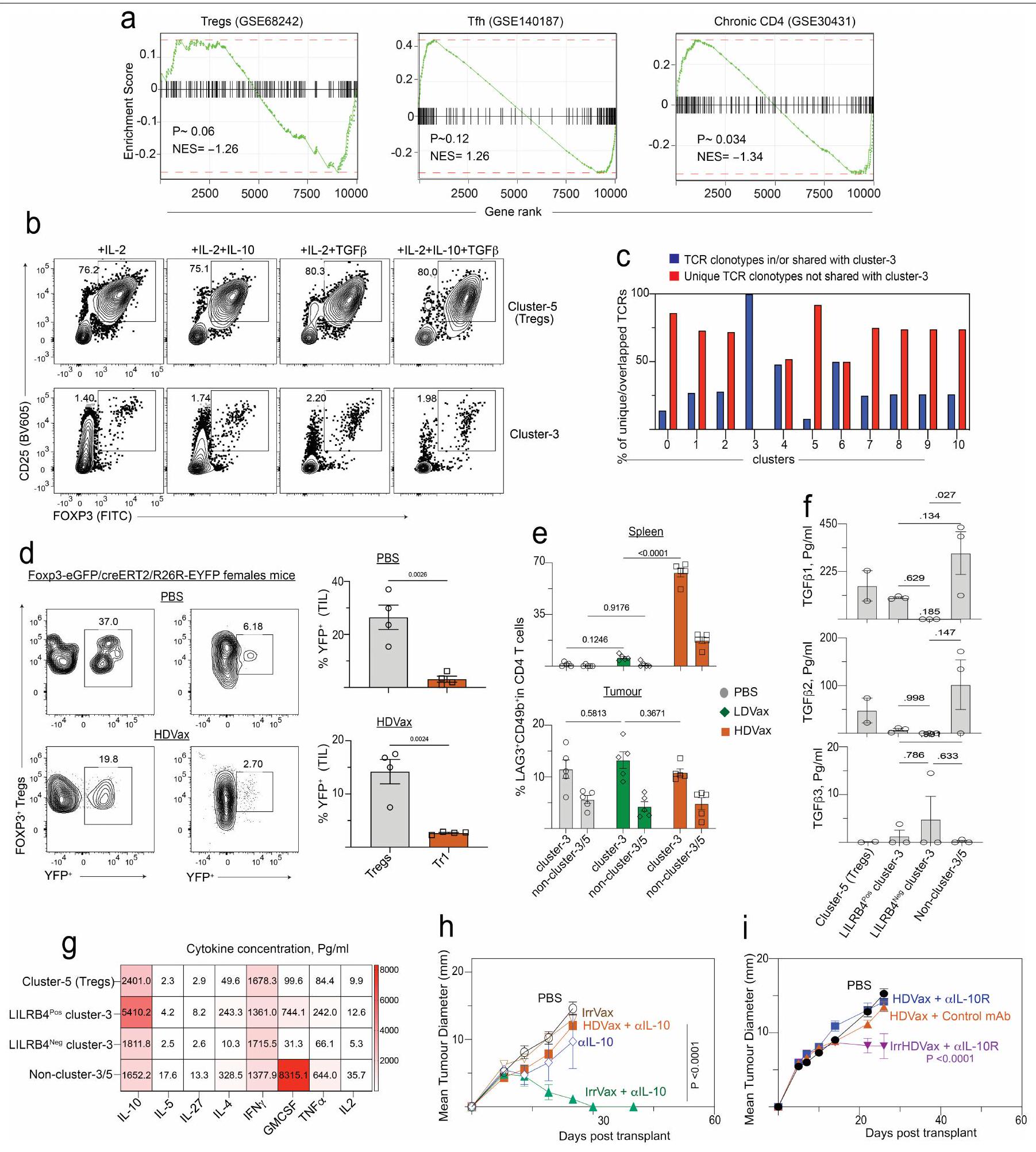
الخلايا المستحثة بواسطة HDVax هي خلايا Tr1
أظهر تحليل GSEA أن خلايا المجموعة 3 تعرض توقيعًا جينيًا مميزًا يميزها عن خلايا FOXP3+خلايا خلايا ومُستَنفَذ مُنبه بشكل مزمنخلايا T (الشكل البياني الموسع 5a). ظلت الخلايا المثبطة المستحثة بواسطة HDVax FOXP3بعد 10 أيام من الحضانة مع سيتوكينات مختلفة في المختبر (الشكل 5b من البيانات الموسعة). بالإضافة إلى ذلك، لم تكن الخلايا المستحثة بواسطة HDVax مشتقة منالخلايا كما يتضح من عدم وجود أنماط تكرارية مشتركة لمستقبلات الخلايا التائية في الكتلة 3 والكتلة 5 ودراسات تتبع السلالة في الجسم الحي باستخدام فئران تتبع سلالة FOXP3 (الشكل 5c,d من البيانات الموسعة). وبالتالي، لم تكن خلايا المجموعة 3 سابقة لـ، ولا متمايزة من، الخلايا. وعلى العكس، فإن توقيع الجين للخلايا في المجموعة 3 تداخل بشكل كبير مع تلك المبلغ عنها لخلايا Tr 1 في الفئران والبشر.و سامة للخلاياخلايا (الشكل 3j). بالإضافة إلى ذلك، كانت نسبة خلايا Tr1-like أعلى في المرضى الذين يظهرون مقاومة للعلاج بمضادات PD1 واللقاحات الشخصية ضد السرطان (الشكل 3k). بينما يتم استخدام تعبير LAG3 و CD49b (الذي يُستخدم أحيانًا لتعريف خلايا Tr1) لم تكن مختلفة في TILs للفئران المعالجة بـ HDVax مقارنة بالفئران المعالجة بـ LDVax، حيث كانت هذه العلامات معبرة بشكل انتقائي على CD4 المحدد بـ mItgb1.خلايا T في طحال الفئران المعالجة بـ T3-HDVax، ولكن ليس في الفئران المعالجة بـ T3-LDVax (الشكل 5e من البيانات الموسعة). LILRB4 المستحثة بواسطة T3-HDVaxخلية المجموعة 3 أنتجت كميات عالية من IL-10 وكميات منخفضة من IL-2 (علامات خلايا Tr1)، بالإضافة إلى مستويات منخفضة من TNF و GM-CSF مقارنة بـ LILRB4.خلايا العنقود 3 خلايا أو غير متجمعة الخلايا (الشكل البياني الموسع 5 ج). TGFتم إنتاجه بشكل رئيسي بواسطة غير العنقوديالخلايا والخلايا (الشكل 5f من البيانات الموسعة). لم يُنقذ حجب IL-10 باستخدام مضاد IL-10 أو مضاد IL-10R تثبيط HDVax (الشكل 5h,i من البيانات الموسعة)، مما يشير إلى أن IL-10 لم يكن كافياً لعرقلة الفعالية المضادة للورم.
تشير هذه البيانات إلى أن HDVax والأورام التي تنمو بشكل تدريجي وتصبح غير حساسة لعلاج anti-PD1 تفعل ذلك جزئيًا بسبب إفراز FOXP3- CD4 المثبط.خلايا Tr1.
مضاد LILRB4 يعكس القمع بواسطة خلايا شبيهة بـ Tr1
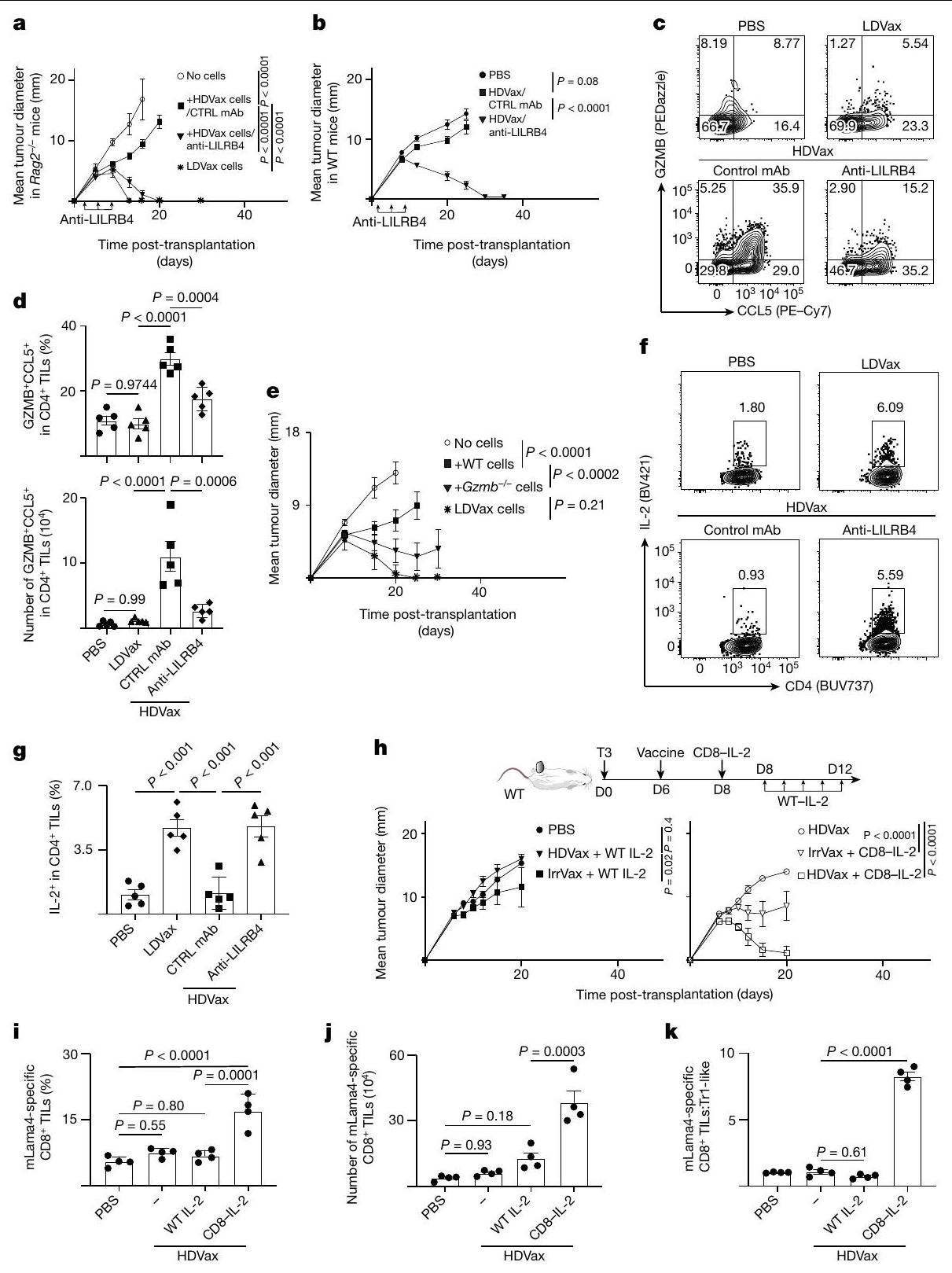
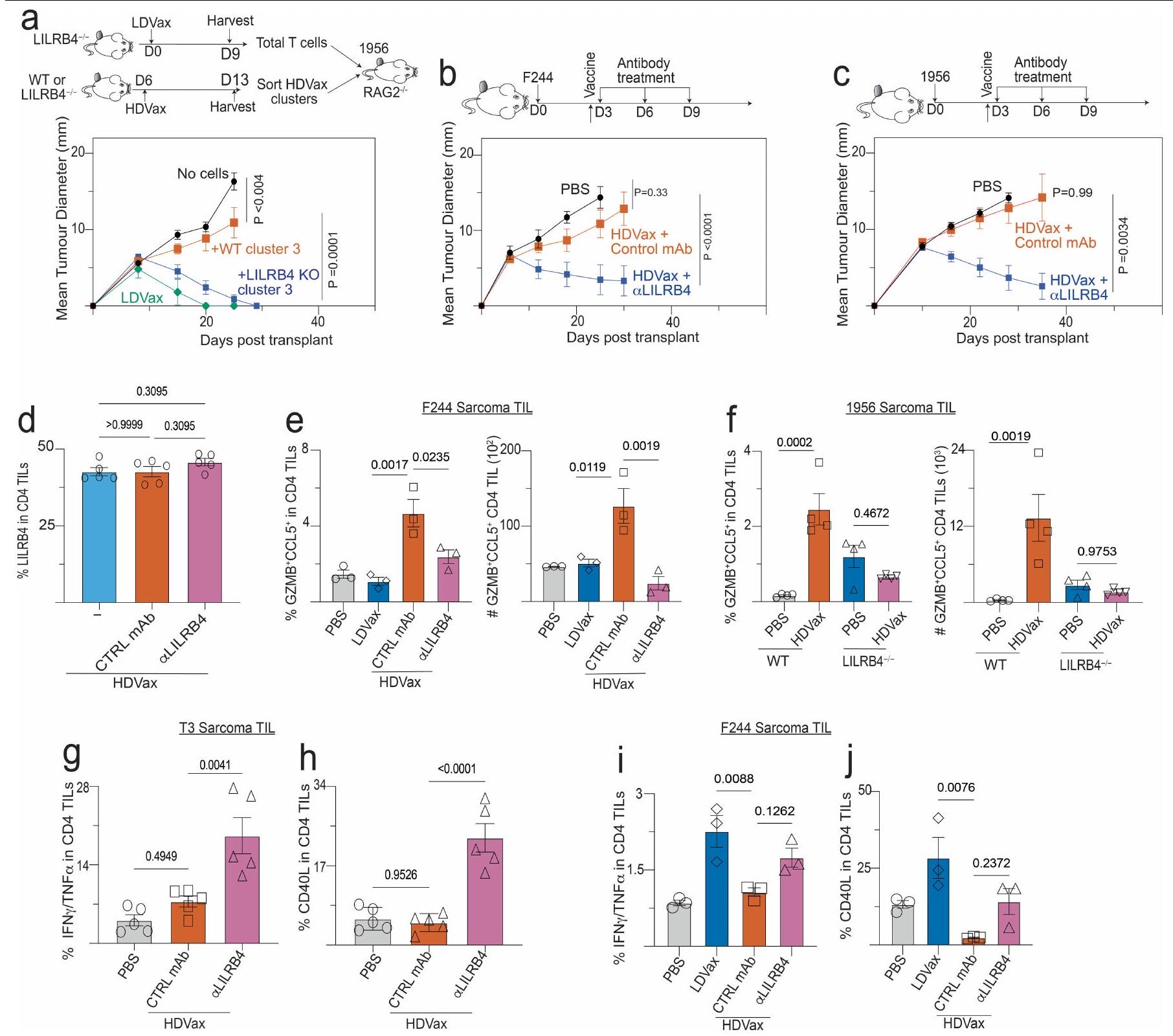
استنادًا إلى الاكتشاف بأن خلايا Tr1 المثبطة تعبر عن بروتين LILRB4 على سطحها، تساءلنا عما إذا كان جسم مضاد وحيد النسيلة الذي تم الإبلاغ عنه مؤخرًا والذي يحجب LILRB4يمكن أن تلغي الإجراءات القمعية لهذه الخلايا. كما هو متوقع، في الفئران Rag2-/، أدى نقل خلايا T الفعالة المستحثة بواسطة T3-LDVax إلى رفض T3، وخلط خلايا T الفعالة مع خلايا Tr1 المستحثة بواسطة T3-HDVax أعاق الرفض (الشكل 4a). بينما لم يعكس إضافة الأجسام المضادة الأحادية النسيلة الضابطة إلى الخليط الأخير تثبيط رفض الورم، فإن إضافة الأجسام المضادة المضادة لـ LILRB4 سهلت رفض الورم. وبالمثل، بينما أدى نقل خلايا المجموعة 3 المستحثة بواسطة HDVax من الفئران WT إلى تثبيط رفض الورم في هذا الاختبار، فإن نقلخلايا الكتلة 3 من Lilrb4لم تمنع الفئران رفض الورم (الشكل التوضيحي 6a). علاوة على ذلك، نمت أورام T3 بشكل متزايد في الفئران البرية المعالجة بـ T3-HDVax والأجسام المضادة الأحادية التحكم، ولكن تم رفضها في الفئران البرية التي تلقت مجموعة من T3-HDVax + مضاد LILRB4 (الشكل 4b). تم ملاحظة نتائج مماثلة باستخدام نماذج الساركومة F244 و1956 مع علاجات HDVax الخاصة بهما (الشكل التوضيحي 6b وc، على التوالي).
سألنا بعد ذلك عما إذا كان علاج مضاد LILRB4 قد أثار تغييرات في النمط الظاهري و/أو الوظيفي في خلايا Tr1 المستحثة بواسطة HDVax الخاصة بالورم. وقد أكدنا أن مضاد LILRB4 لم يقم بتقليل عدد خلايا CD4 التي تعبر عن LILRB4.تيلز في الجسم الحي (الشكل البياني الموسع 6d). أدى العلاج بهذا الجسم المضاد وحيد النسيلة إلى تقليل مستويات GZMB/CCL5 المعبر عنها لكل CD4تم حصاد خلايا T من الفئران الحاملة لورم T3 (الشكل 4c، d) أو F244 (الشكل 6e من البيانات الموسعة). تم تسجيل انخفاض مماثل في خلايا CD4 التي تعبر عن GZMB/CCL5.تمت ملاحظة TILs عند إجراء مقارنة بين فئران WT الحاملة للأورام من عام 1956 وفئران ذات تعطيل مستهدف لجين Lilrb4 (الشكل 6f من البيانات الموسعة). تشير هذه النتائج إلى وجود صلة بين التعبير الخلوي عن LILRB4 و GZMB.
لاختبار دور GZMB في العملية المثبطة المعتمدة على Tr1، قمنا بإجراء اختبار Rag2-‘ ACT باستخدام خلايا المجموعة 3 المشتقة من WT المعالج بـ T3-HDVax مقابل Gzmb.الفئران بعد التأكيد أولاً على مستويات مماثلة من تعبير LILRB4 في الخلايا المشتقة من كل سلالة. بينما أظهرت خلايا المجموعة 3 المستحثة بواسطة T3-HDVax من الفئران WT تثبيطًا قويًا لرفض الورم، فإن Gzmb المستحث بواسطة T3-HDVaxأظهرت خلايا المجموعة 3 قدرة مثبطة مخفضة (الشكل 4e).
ثم قمنا بتقييم قدرة مضاد LILRB4 على التأثير في تحفيز CD4 الأخرىمنتجات مشتقة من خلايا T التي تقدم المساعدة للاستجابات المضادة للورم. أعاد حجب LILRB4 إنتاج IL-2 في الخلايا المستحثة بواسطة HDVax (الشكل 4f، g) وزاد من تعبيرها عن IFN. TNF و CD40L في كل من نماذج الأورام T3 و F244 (الشكل 6g,j من البيانات الموسعة). بالتزامن مع هذه التغيرات، زاد مضاد LILRB4 من عدد خلايا CD8 الخاصة بـ mLama4.تمت ملاحظة خلايا TILs وانخفاض التعبير عن علامات الإرهاق PD1 وLAG3 وTIM3. تم تكرار نتائج مماثلة باستخدام أورام F244 وF244-HDVax (الشكل البياني الممتد 7a-e). وبالتالي، في الفئران، يتم تمييز خلايا Tr1-like المثبطة بتعبير LILRB4، وتُعكس قدرتها المثبطة إما عن طريق حجب الأجسام المضادة وحيدة النسيلة أو الحذف الجيني للبروتين الذي، بدوره، يقلل من تعبير GZMB ويزيد من تعبير IL-2 وجزيئات أخرى تسهل الفعالية المضادة للأورام.
CD8-IL-2 يتجاوز تثبيط HDVax
قدرة مضاد LILRB4 على استعادة إنتاج IL-2 في CD4 المستحث بواسطة HDVaxدفعتنا خلايا T إلى التساؤل عما إذا كان تكميل IL-2 سيعيد فعاليةوظيفة الخلايا ورفض الورم. على الرغم من أن IL-2 البري هو علاج معتمد لبعض أنواع السرطان، إلا أن استخدامه السريري محدود بسبب تأثيراته المتعددة السلبية وسمّيته العالية.. لذلك استخدمنا متغير IL-2 المستهدف بشكل خاص لخلايا T (CD8-IL-2) الذي يرتبط بشكل انتقائي بـالخلايا ولكن ليس إلىالخلايا، خلايا القاتل الطبيعي أو
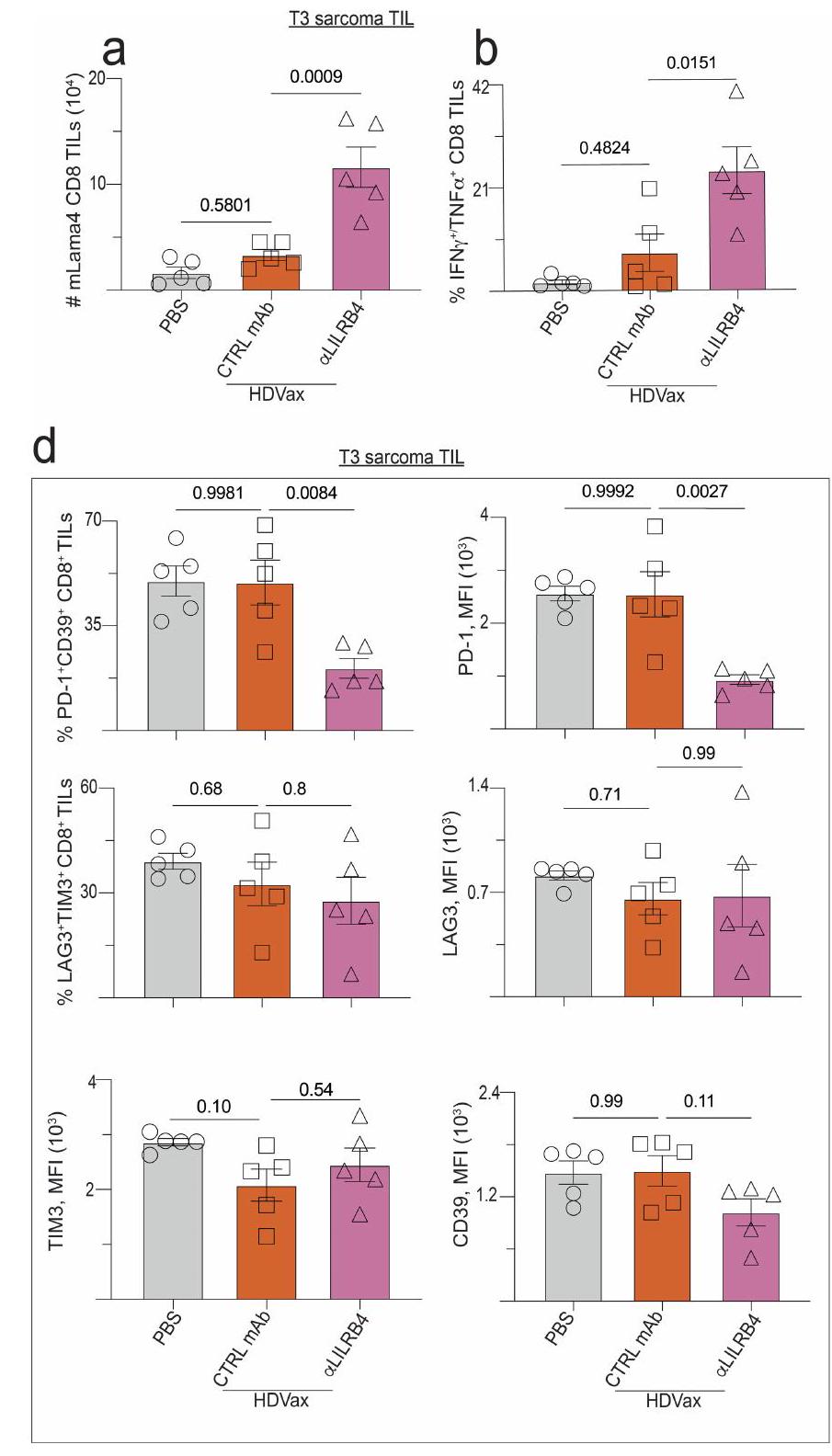
الشكل 4 | علاج مضاد لـ LILRB4 يعكس تحفيز HDVax لـ GZMB/CCL5 وكبح إنتاج IL-2. أ، نمو ورم T3 في فئران Rag2-/ التي تتلقى خلايا T من فئران تم علاجها بـ LDVax بالإضافة إلى CD4 المحدد لـ mItgb1 الذي تم تحفيزه بواسطة HDVax والذي لا يحتوي على CD25.خلايا T (حوالي ) تليها معالجة بالأجسام المضادة الأحادية النسيلة المضادة لـ LILRB4 أو التحكم (CTRL) ( (PBS)، 5 (LDVax و anti-LILB4) و 8 لمضاد التحكم mAb).b، نمو ورم T3 في الفئران WT المعالجة بـ HDVax وحده أو مع anti-LILRB4 ( (PBS) و 10 لمجموعات أخرى). ج، مخططات تدفق تمثيلية تظهر تعبير GZMB/CCL5 في CD4تيلز المحملة على CD4. د، التردد (أعلى) والأرقام (أسفل) ل CD4 الذي يعبر عن GZMB/CCL5الخلايا التائية المستحثة في الأورام من النوع T3 بعد 7 أيام من العلاج ). e، 1956 نمو الورم في فئران Rag2-/- التي تتلقى خلايا T من فئران LDVax WT وخلايا CD4 الخاصة بـ mItgb1 السلبية لـ CD25 المستحثة بواسطة HDVaxخلايا T (حوالي ) من WT أو Gzmb فئران (لـ PBS و LDVax، و 7 لمجموعات أخرى). ف، تظهر مخططات تدفق الخلايا التمثيلية تعبير IL-2 في CD4تكرار الخلايا التي تعبر عن IL-2 في CD4خلايا T في عدة فئران ). نمو ورم T3 في الفئران البرية المعالجة كما هو موضح ومفصل في الطرق ( ). تم علاج الفئران الحاملة لورم T3 كما هو موضح في اللوحة، وتكرار ( ) والأرقام ( ) من CD8 الخاص بـ mLama4 الخلايا التائية المستحثة، ونسبة خلايا CD8 المحددة لـ mLama4الخلايا الشبيهة بـ Tr1تم تقييم TILs في اليوم الثالث عشر ). البيانات في الألواح يتم التعبير عنها كمتوسط قطر الورم س.م. ويمثل ثلاثة واثنين واثنين وثلاثة تجارب، على التوالي؛ تم إجراء الإحصائيات باستخدام تحليل التباين الثنائي، مع تصحيح المقارنات المتعددة باستخدام اختبار المقارنة المتعددة لسيداك. البيانات في الألواح تظهر كمتوسطيمثل s.e.m. ثلاثة، ثلاثة، اثنان وثلاثة تجارب، على التوالي؛ تم إجراء الإحصائيات باستخدام تحليل التباين الأحادي، مع تصحيح المقارنات المتعددة باستخدام طريقة توكي. CD4 التقليديخلايا T. CD8-IL-2 يوسع بشكل انتقائي CD8خلايا T في الفئران والرئيسيات وتحافظ على وظيفة CTL دون التسبب في السمية. لا T3-HDVax ولا الجرعات العلاجية المنخفضة من CD8-IL-2 (أدى إلى رفض الورم في الفئران الحاملة لـ T3 كعلاجات أحادية. بالمقابل، أدى الجمع بين T3-HDVax وجرعات تحت علاجية من CD8-IL-2 إلى رفض ورم T3 (الشكل 4h). كان دمج IrrVax مع CD8-IL-2 غير فعال. كانت المعالجة بالجمع لم تؤدِ WT IL-2 و T3-HDVax أو Irr-HDVax إلى رفض الورم ولكنها تسببت في سمية كبيرة، كما يتضح من فقدان الوزن في الفئران المعالجة. تم ملاحظة نتائج مشابهة باستخدام نماذج الورم F244 و 1956 (الشكل 8a و b في البيانات الموسعة، على التوالي).
بعد العلاج بـ HDVax بالإضافة إلى CD8-IL-2، لوحظ زيادة بمقدار 2.5 مرة في التردد (الشكل 4i والشكل الإضافي 8c) والأعداد المطلقة (الشكل 4j) من خلايا CD8 الخاصة بـ mLama4.تمت ملاحظة TILs. بالإضافة إلى ذلك،
الشكل 5 | تتولد خلايا شبيهة بـ Tr1 نتيجة تقديم مستضد cDC2 المفضل بواسطة HDVax وتقتل cDC1 بطريقة تعتمد على GZMB. أ، بروتوكول الفرز وإنتاج IL-2 بواسطة هجين محدد لـ mItgb1 (، تمثل تجربتين). ب، تكرار تعبير NIR في cDC1 () بعد الحضانة مع تحت المجموعات المصنفة من CD4 المستحثة بواسطة HDVaxخلايا T المستخرجة من الفئران WT (المعالجة بـ HDVax بالإضافة إلى الأجسام المضادة التحكمية أو anti-LILRB4) أوفئران (، تمثل ثلاثة تجارب). ج، مخططات تدفق الخلايا التمثيلية تظهر cDC1 و cDC2 في أورام T3 التي تم تصنيفها مسبقًا علىوتكرار (أعلى) والأعداد (أسفل) من cDC1 في أورام T3 التي تم تقييمها بعد 5 أيام من التطعيم، تمثل ثلاثة تجارب). هـ، التردد (أعلى) والأعداد (أسفل) لـ LILRB4CD25فوكس بي 3CD4خلايا T في عام 1956 تم جمع TILs من WT أومحمل بالأورام فئران بعد التطعيم (، تمثل ثلاثة تجارب). ف، التردد (أعلى) والأعداد (أسفل) من cDC1 في أورام 1956 في WT والفئران التي تم تقييمها بعد 5 أيام من التطعيم، تمثل ثلاثة تجارب). ج، 1956 نمو الورم في WT وفئران تم تطعيمها بـ 1956 HDVax، LDVax أو IrrVax في اليومين 5 و 16. هـ، 1956 نمو الورم في WT والفئران المعالجة بـمن مضاد PD1 في الأيام 12 و15 و18 (، تمثل تجربتين). البيانات في الألواحتُعبر كمتوسطبيانات s.e.m. في اللوحاتيتم التعبير عنها كمتوسط قطر الورمتم إجراء الإحصائيات في اللوحة د باستخدام تحليل التباين الأحادي، مع تصحيح المقارنات المتعددة باستخدام طريقة توكي. الإحصائيات في اللوحاتتم تحديدها بواسطة تحليل التباين ثنائي الاتجاه، مع تصحيح المقارنات المتعددة باستخدام اختبار المقارنات المتعددة لسيداك.
كان علاج CD8-IL-2 مرتبطًا بزيادات كبيرة في نسبةتحويل TILs إلى خلايا شبيهة بـ Tr1 (الشكل 4k). تم ملاحظة نتائج مماثلة أيضًا في نموذج ورم F244 (الشكل التمديدي 8d-f). وبالتالي، استهداف ميوتيني IL-2 إلىتتجاوز خلايا T تثبيط HDVax من خلال زيادة تكرار السيتوتوكسية المحددة بمستضد.الخلايا، مما يؤدي إلى تراجع الورم.
يتطلب تحفيز خلايا Tr1 المثبطة وجود cDC2s/ وحيدات النواة
نظرنا في إمكانية أن تعكس النتائج المختلفة بين HDVax و LDVax اختلافات في خلايا تقديم المستضد (APCs) التي تقدم المItgb1 neoAg. استكشفنا هذه الإمكانية من خلال تطعيم الفئران السليمة إما بـ T3-LDVax أو T3-HDVax، وعزل مجموعات مختلفة من خلايا APC في نقاط زمنية مختلفة واختبار قدرتها على تقديم mItgb1 إلى هجين خلايا T محددة بـ mItgb1باستخدام T3-LDVax، لاحظنا أن كل من cDC1s و cDC2s قدمت النيوبيبتيد mItgb1 إلى نفس الدرجة تقريبًا (الشكل 9a من البيانات الموسعة). بالمقابل، باستخدام T3-HDVax، برزت مجموعة cDC2 كأهم خلايا تقديم المستضد (الشكل 5a). يتماشى هذا مع النتيجة، حيث أن HDVax وليس LDVax أثار استجابات قوية للأجسام المضادة تجاه ببتيد mItgb1 (الشكل 9b،c من البيانات الموسعة). كما هو متوقع، فإن خلايا Tr1 المستحثة بواسطة HDVax عبرت عن مستويات عالية من GZMB و perforin (الشكل 3c والشكل 9d من البيانات الموسعة)، مما يشير إلى أنها قد تظهر على الأقل بعض وظائفها المثبطة من خلال قتل خلايا تقديم المستضد التي تعبر عن MHC-II.نظرًا لأن خلايا T3 لا تعبر أبدًا عن MHC-II، فقد اعتبرنا إمكانية أن تكون الخلايا المستهدفة لـ Tr1 السامة هي cDC1 لأنها تقدم كل من مستضدات الورم MHC-I وMHC-II.لذلك، فهي بشكل خاص مهم للاستجابة المضادة للأورام. أظهرت اختبارات القتل في المختبر أن خلايا Tr1 المستحثة بواسطة T3-HDVax ولكن ليس خلايا CD4 غير Tr1قتلت خلايا T خلايا cDC1 المحملة بـ mltgb1 ولكنها لم تقتل خلايا cDC2 بطريقة تعتمد على GZMB (الشكل 5b والشكل الإضافي 9e، f). من الجدير بالذكر أن خلايا Tr1 التي تم جمعها من الفئران المعالجة بـ T3-HDVax في وجود مضاد LILRB4 أظهرت نشاط قتل في المختبر منخفض. بالإضافة إلى ذلك، تم تقليل تردد وعدد خلايا cDC1 المستعادة من الفئران الحاملة للأورام المعالجة بـ T3-HDVax بمقدار 4.5 مرة مقارنة بالفئران المعالجة بـ T3-LDVax أو PBS. تم استعادة استعادة CDC1 جزئيًا عندما تم دمج T3-HDVax مع علاج مضاد LILRB4 (الشكل 5c، d والشكل الإضافي 9g) أو عندما تم إعطاء HDVax لـ Lilrb4.الفئران (الشكل 9h من البيانات الموسعة). وبالمثل، فإن العلاج بمضاد CTLA4، الذي يستنفد خلايا Tr1 والخلايا (الشكل الإضافي 3j)، استعادة تعافي cDC1s في فئران T3-HDVax (الشكل الإضافي 9i). لم يُلاحظ أي انخفاض في استعادة cDC1 في الفئران المعالجة بـ T3-HDVax Gzmb.الفئران (الشكل 9j من البيانات الموسعة). وبالتالي، فإن اللقاحات التي تحتوي على جرعات عالية من neoAg MHC-II تفضل تقديم المستضد بشكل أساسي بواسطة cDC2s، مما يؤدي إلى تكوين خلايا Tr1 السامة التي تقتل cDC1s التي تقدم مستضدات الأورام بطريقة تعتمد على GZMB.
HDVax يحمي الفئران التي تفتقر إلى خلايا cDC2/الوحيدات
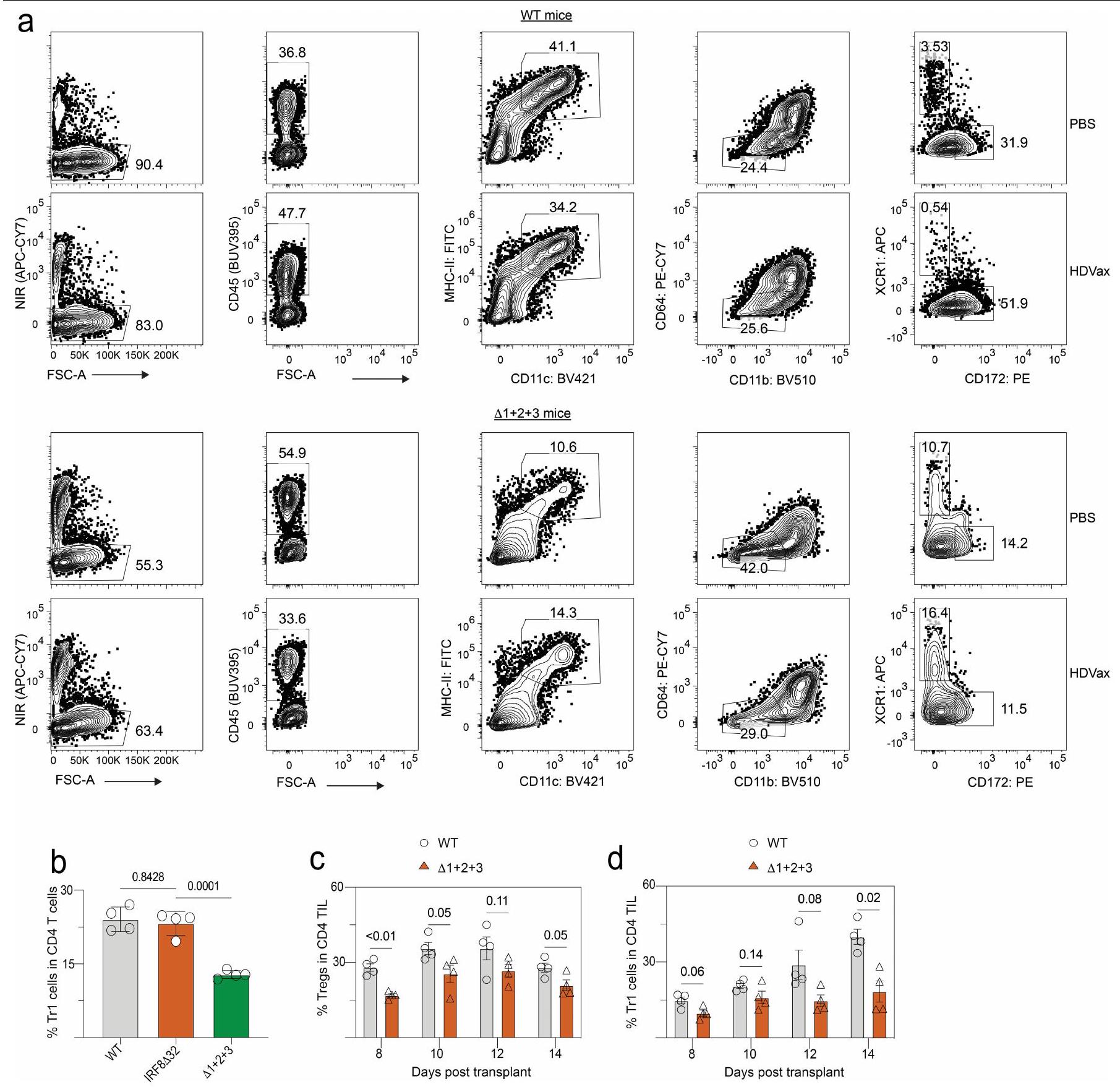
افترضنا أن الإزالة الجينية لخلايا cDC2s/الوحيدات في الفئران قد تقلل من إنتاج خلايا Tr1 وتلغي التأثير المثبط لـ HDVax. لذلك، قمنا بحقن 1956 خلية ساركوما في فئران C57BL/6 WT أو فئران C57BL/6 التي تفتقر إلى خلايا cDC2s/الوحيدات. ) ولكن لا يزال قادرًا على توليد على عكس الفئران WT، تم علاجها بـ 1956-HDVaxلم تُظهر الفئران زيادة في تكرار خلايا Tr1 (الشكل 5e) ولا انخفاضًا في استعادة cDC1 (الشكل 5f). كما هو متوقع، نمت أورام 1956 بشكل تدريجي سواء في الفئران غير المعالجة أو في الفئران المعالجة بـ Irr-HDVax من النوع البري.الفئران، وتم رفضها في كلا السلالتين عندما تم علاج الفئران الحاملة للأورام بـ 1956-LDVax. 1956-HDVax أدى إلى رفض الورم فيالفئران ولكن ليس في الفئران WT (الشكل 5g). من الجدير بالذكر أن تردد خلايا Tr1 كان مماثلاً في كل من الفئران WT والفئران التي تفتقر إلى cDC1s (IRF8فئران) (الشكل 10ب من البيانات الموسعة).
كما لاحظنا زيادة في خلايا Tr1 في الساركوما المتزايدة بشكل تدريجي حيث فقدت حساسيتها لمضاد PD1 (الشكل. )، قمنا بمقارنة مظهر هذه الخلايا في أورام عام 1956 التي تنمو في WT مقابل الفئران. تراكمت خلايا Tr1 في الأورام المتزايدة بشكل تدريجي 1956 في الفئران WT حيث أصبحت غير حساسة لمضاد PD1. بالمقابل، لم تتراكم خلايا Tr1 في أورام 1956 التي تنمو فيالفئران (الشكل 10c، d من البيانات الموسعة). أخيرًا، أدى مضاد PD1 إلى رفض أورام 1956 التي تم تأسيسها منذ 12 يومًا فيالفئران ولكن ليس في فئران WT (الشكل 5h)، مما يشير إلى أن الفعالية المعززة لمضاد PD1 في غياب cDC2s و/أو العدلات كانت متجسدة، على الأقل جزئيًا، من خلال الفشل في توليد خلايا Tr1 السامة للخلايا. تكشف هذه النتائج أنه تحت ظروف HDVax، تقوم cDC2s و/أو العدلات بتحفيز خلايا Tr1 CD4 السامة للخلايا.خلايا T التي تقتل أو تضر بسلامة/استقرار cDC1s، مما يؤدي إلى عيوب في تطوير/نضوج الخلايا السامة.استجابات خلايا T، مما يضعف التأثيرات المضادة للورم للمناعة.
نقاش
يظهر العمل المقدم هنا أن فعالية لقاحات البيبتيدات النيوأنتيجينية الخاصة بالأورام تعتمد بشكل كبير على جرعة النيوأنتيجين MHC-II الموجودة في اللقاح. لقد أظهرنا أن الجرعة الزائدة من النيوبيبتيد MHC-II المعبر عنه بواسطة الورم في اللقاح يمكن أن تعكس فعالية اللقاح تمامًا. هذه الملاحظة مهمة جدًا وفي وقتها، حيث يتم الآن السعي بنشاط نحو لقاحات السرطان العلاجية كعلاج مناعي شخصي للسرطان.لم نفهم بعد القواعد التي تحكم كميات مستضدات MHC-II الجديدة اللازمة لتحقيق فعالية لقاح مثالية، كما أننا لا نقترح أن يتم تحديد ذلك تجريبيًا لكل ورم من أورام المرضى. بدلاً من ذلك، نركز على تحديد طبيعة المثبطات. تم توليد خلايا T في ظل ظروف تطعيم غير فعالة ثم تم التحقق من استراتيجيات لتجاوز الآثار الضارة للجرعات العالية من MHC-II neoAg على الاستجابة المضادة للورم. باستخدام هذا النهج، أظهرنا أن تثبيط فعالية مضادات الورم بواسطة HDVax يرجع إلى تحفيز خلايا Tr1 المحددة لمستضدات الورم السيتوليتية التي تقتل cDC1s في المختبر وفي الجسم الحي. وبالتالي، يحدد عملنا وظيفة مثبطة حاسمة ل CD4 المعروف.مجموعة فرعية من خلايا T التي لم ترتبط عمومًا بالعلاج المناعي للسرطان أو هروب المناعة من السرطان. إن إثباتنا أن خلايا Tr1 تصبح قابلة للاكتشاف في الأورام التي تنمو بشكل متزايد مع مقاومتها للعلاج المضاد لـ PD1 يشير بقوة إلى أن الخلايا المستحثة بواسطة HDVax هي نظائر لتلك التي تظهر خلال نمو الورم المتقدم، وبالتالي، فهي مشاركون ذوو صلة فسيولوجية في قمع المناعة المرتبط بالسرطان.ثم قمنا بتحديد الآليات التي يتم من خلالها تحفيز خلايا Tr1 وأظهرنا ثلاث استراتيجيات بديلة لتجاوز نشاطها في الجسم الحي.
لقد أظهرنا أيضًا أن العلامات الظاهرية على خلايا Tr1 تختلف بناءً على المواقع التشريحية التي يتم عزلها منها. وبالتالي، بينما يتم تمييز خلايا Tr1-like المستحثة بواسطة HDVax في الطحال بشكل مختلف من خلال تعبير CD49b و LAG3 (المرجع 39)، فإن الخلايا في TILs تفتقر إلى التعبير التفاضلي عن هذه العلامات لكنها لا تزال تحتفظ بالقدرة على إنتاج IL-10، السيتوكين المميز لخلايا Tr1. بالمقابل، يحدد تعبير LILRB4 بوضوح خلايا Tr1-like في الفئران من كل من الطحال وبيئة الورم الدقيقة. ومع ذلك، بينما يتم ترميز LILRB4 في الفئران بواسطة جين Gb49b واحد، فإنه ينتمي إلى عائلة أكثر تعقيدًا في البشر تتكون من خمسة جينات مختلفة.ستكون الجهود المبذولة لتعريف النظير البشري هدفًا أساسيًا للعمل المستقبلي وقد توفر أداة لتحديد وجود مثل هذه الخلايا بسهولة لدى المرضى المصابين بالسرطان. هذا مهم لأن خلايا Tr1 المثبطة المستحثة بواسطة HDVax وخلايا CD4 المساعدة المستحثة بواسطة LDVax.تعبّر عن مستويات مشابهة من CD39، مما يشير إلى أن عوامل أخرى تتجاوز CD39 مطلوبة للتفريق بين CD4خلايا T التي تثبط مقابل تعزز رفض الورم.
أظهرت الأعمال الأخيرة من قبل آخرين أن بعض مستضدات الأورام، التي تُسمى مثبطات، يمكن أن تقمع فعالية اللقاحات المضادة للسرطان وanti-PD1 (المرجع 56)، وأن وظيفتها المثبطة كانت نتيجة لخصائصها الهيكلية الكامنة ولكن غير المحددة. تميز هذه التصنيف المثبطات عن مستضدات MHC-II الجديدة المقدمة في الدراسة الحالية، التي ترتبط خصائصها المنشطة مقابل المثبطة بجرعاتها. تثير دراستنا إمكانية أن أي مستضد جديد من MHC-II يمكن أن يكون شبيهاً بالمثبط عندما يتم إنتاجه أو إطلاقه بمستويات مرتفعة، مما يوسع تعريف المثبط. سيكون من الضروري أيضاً في المستقبل استكشاف ما إذا كانت تحفيز خلايا Tr1 يعتمد على نوع منصة اللقاح المستخدمة (على سبيل المثال، اللقاحات القائمة على الببتيد مقابل اللقاحات القائمة على الحمض النووي مقابل اللقاحات القائمة على الحمض النووي الريبي). ستتناول الدراسات المستقبلية هذه الأسئلة. ومع ذلك، فإن تحديد خلايا Tr1 ووظيفتها البيولوجية المثبطة للمناعة في السرطان وتقديم ثلاث استراتيجيات لتجاوز وظائفها المثبطة توفر أهدافاً جديدة محتملة لتحسين فعالية العلاجات المناعية للسرطان بشكل عام.
المحتوى عبر الإنترنت
أي طرق، مراجع إضافية، ملخصات تقارير Nature Portfolio، بيانات المصدر، بيانات موسعة، معلومات تكميلية، شكر وتقدير، معلومات مراجعة الأقران؛ تفاصيل مساهمات المؤلفين والمصالح المتنافسة؛ وبيانات توفر البيانات والرموز متاحة علىhttps://doi.org/10.1038/s41586-024-07752-y.
4. ختري، ر.، كوكس، ت.، ياسايكو، س. أ. ورامسديل، ف. دور أساسي لسكورفين في CD4CD25خلايا تنظيم T. نات. مناعه. 4، 337-342 (2003). 5. ثورنتون، أ. م. وشيفاش، إ. م. CD4تقوم خلايا T المناعية التنظيمية بتثبيط تنشيط خلايا T متعددة النسائل في المختبر عن طريق تثبيط إنتاج الإنترلوكين 2. J. Exp. Med. 188، 287-296 (1998). 6. إيشويلر، س. وآخرون. خلايا تنظيم T الجريبية داخل الورم تحد من فعالية علاج PD-1. نات. إيمونول. 22، 1052-1063 (2021). 7. زاباسودي، ر. وآخرون. مثبطات غير تقليدية CD4+Foxp3-PD-1خلايا T كعلامة حيوية لنشاط حجب نقاط التفتيش المناعية. خلية السرطان 34، 1017-1032.e7 (2018). 8. ألسباتش، إ. وآخرون. تشكل النيوانتجينات MHC-II مناعة الورم واستجابة للعلاج المناعي. ناتشر 574، 696-701 (2019). 9. أهريندس، ت. وآخرون. CD4يساعد الخلية على منح برنامج تأثير الخلايا التائية السامة للخلايا بما في ذلك تقليل مستقبلات التثبيط المساعدة وزيادة القدرة على التوغل في الأنسجة. المناعة 47، 848-861.e5 (2017). 10. بينيت، س. ر.، كاربوني، ف. ر.، كاراماليس، ف.، ميلر، ج. ف. وهيث، و. ر. تحفيز لـاستجابة الخلايا التائية السامة للخلايا عن طريق التحفيز المتبادل تتطلب التعرف المتبادلمساعدة الخلايا. ج. تجريب. ميد. 186، 65-70 (1997). 11. أوسندورب، ف.، مينغيدي، إ.، كامبس، م.، فيليوس، ر. وملييف، ج. س. متطلبات خلايا T المساعدة المحددة للتحفيز الأمثل للخلايا السامة.الخلايا اللمفاوية ضد الأورام السلبية لمركب التوافق النسيجي الرئيسي من الفئة الثانية. J. Exp. Med. 187، 693-702 (1998). 12. كلارنيت، ج. ب. وآخرون. CD8 المتفاعل مع FBLسامة للخلايا و CD4تتعرف الخلايا اللمفاوية التائية المساعدة على مستضدات مميزة مشفرة بواسطة فيروس لوكيميا الفئران فريد. ج. التجريبية. ميد. 169، 457-467 (1989). 13. دولينا، ج. س. وآخرون. مرتبطتطعيم الخلايا النيوأنتيجينية يتغلب على مقاومة حجب نقاط التفتيش المناعية ويمكّن من تراجع الورم. J. Clin. Invest. 133، e164258 (2023). 14. بوت، سي.، أبيتو، ل. وكوتشرو، ف. ك. خلايا T التنظيمية من النوع 1 (Tr1) في المناعة الذاتية. سيمين. إيمونول. 23، 202-208 (2011). 15. كوتش، ك. وآخرون. زيادة تكرار خلايا T التنظيمية من النوع 1 CD49b/LAG-3+ في الأفراد المصابين بفيروس نقص المناعة البشرية. أبحاث الإيدز. الفيروسات العكسية البشرية 31، 1238-1246 (2015). 16. مكغويرك، ب.، مككان، ج. & ميلز، ك. هـ. خلايا تنظيمية T محددة مسببة للأمراض يتم تحفيزها في الجهاز التنفسي بواسطة جزيء بكتيري يحفز إنتاج الإنترلوكين 10 بواسطة الخلايا الشجرية: استراتيجية جديدة للتملص من استجابات T المساعدة من النوع 1 الواقية بواسطة بوردتيلا السعال الديكي. ج. تجريبي. ميد. 195، 221-231 (2002). 17. بيرغمان، سي. وآخرون. خلايا تنظيم المناعة من النوع 1 في سرطان الخلايا الحرشفية للرأس والعنق: آليات القمع والتوسع في المرض المتقدم. أبحاث السرطان السريرية 14، 3706-3715 (2008). 18. غروكس، هـ. وآخرون. CD4مجموعة فرعية من خلايا T تثبط استجابات خلايا T المحددة للمستضد وتمنع التهاب القولون. ناتشر 389، 737-742 (1997). 19. غوبين، م. م. وآخرون. تستهدف علاجات المناعة ضد السرطان عن طريق حجب نقاط التفتيش المستضدات الطافرة المحددة للأورام. ناتشر 515، 577-581 (2014). 20. مينغ، و. وآخرون. تحسين اكتشاف نيوأنتيجين الورم بواسطة مطياف الكتلة بتقنية التنقل الأيوني ذات الموجات غير المتناظرة عالية المجال. أبحاث مناعة السرطان.https://doi.org/10.1158/2326-6066.CIR-23-0900 (2024). 21. غوبين، م. م. وآخرون. التحليل عالي الأبعاد يحدد إعادة تشكيل حجرة النخاع والعقد اللمفاوية خلال العلاج الناجح لسرطان نقاط التفتيش المناعية. خلية 175، 1014-1030.e19 (2018). 22. غابريشوفا، ل. وآخرون. c-Maf يتحكم في الاستجابات المناعية من خلال تنظيم شبكات الجينات الخاصة بالأمراض وكبح IL-2 في CD4خلايا T. مناعة طبيعية. 19، 497-507 (2018). 23. شارما، ن.، أتولاجبي، أ. ت.، جي، ز. وأليسون، ج. ب. LILRB4 يثبط المناعة في الأورام الصلبة ويعتبر هدفًا محتملاً للعلاج المناعي. ج. إكسب. ميد. 218، e20201811 (2021). 24. سلا، م. وآخرون. مستقبل مثبط جديد (ILT3) معبر عنه على وحيدات النوى، البلعميات، والخلايا الشجرية المعنية في معالجة المستضدات. ج. تجريبي. طب. 185، 1743-1751 (1997). 25. تشانغ، سي. سي. وآخرون. تحمّل خلايا الدندريت بواسطةالخلايا: الدور الحاسم لمستقبلات التثبيط ILT3 وILT4. نات. مناعه. 3، 237-243 (2002). 26. مانافالان، ج. س. وآخرون. التعبير العالي عن ILT3 وILT4 هو سمة عامة للخلايا التغصنية التوليرية. زراعة. المناعة 11، 245-258 (2003). 27. تشابوفال، س. ب. وآخرون. سيميورين 4A يثبت نمط الخلايا التائية التنظيمية البشرية عبر بلكسين B1. إيميونهورايزونز 3، 71-87 (2019). 28. ديلغوف، ج. م. وآخرون. يتم الحفاظ على استقرار ووظيفة خلايا T التنظيمية بواسطة محور نيوروبيلين-1-سيمفورين-4أ. ناتشر 501، 252-256 (2013). 29. جاجلياني، ن. وآخرون. تتحول خلايا Th17 إلى خلايا T تنظيمية خلال انتهاء الالتهاب. ناتشر 523، 221-225 (2015). 30. تشوي، ج. وآخرون. Bcl-6 هو عامل النسخ المحوري لخلايا المساعدة الجريبية T عبر دوائر مثبط المثبط. نات. إيمونول. 21، 777-789 (2020). 31. كروفورد، أ. وآخرون. الأساس الجزيئي والنسخي لـ CD4خلل وظيفة خلايا T خلال العدوى المزمنة. المناعة 40، 289-302 (2014). 32. فان دير فيكن، ج. وآخرون. تتبع وراثي يكشف عن وظيفة تعتمد على عامل النسخ Foxp3 ووظيفة مستقلة عن Foxp3 للنوع المحفز طرفياًالخلايا. المناعة 55، 1173-1184.e7 (2022). 33. زانغ، هـ. وآخرون. شبكة النسخ المدفوعة بواسطة IL-27 تحدد منظمات تعبير IL-10 عبر مجموعات خلايا T المساعدة. تقرير الخلية. 33، 108433 (2020). 34. هاينمان، سي. وآخرون. IL-27 و IL-12 يعارضان IL-23 المؤيد للالتهابات فيخلايا T عن طريق تحفيز Blimp1. نات. كوم. 5، 3770 (2014). 35. كوك، ل. وآخرون. الوظائف المثبطة والمُعَادِلة للأمعاء لخلايا تنظيم T من النوع 1 البشرية. أمراض الجهاز الهضمي 157، 1584-1598 (2019). 36. سليدزينسكا، أ. وآخرون. خلايا T التنظيمية تقيد اكتساب الوظيفة السامة للخلايا المعتمدة على الإنترلوكين-2 و blimp-1 بواسطة CD4خلايا T. المناعة 52، 151-166.e6 (2020). 37. سادي-فيلدمان، م. وآخرون. تحديد حالات خلايا T المرتبطة بالاستجابة للعلاج المناعي عبر نقاط التفتيش في الميلانوما. خلية 175، 998-1013.e20 (2018). 38. عوض، م. م. وآخرون. لقاح النيوانتجين الشخصي NEO-PV-O1 مع العلاج الكيميائي ومضاد PD-1 كعلاج خط أول لسرطان الرئة غير صغير الخلايا غير الحرشفي. خلية السرطان 40، 1010-1026.e11 (2022). 39. جاجلياني، ن. وآخرون. التعبير المشترك عن CD49b وLAG-3 يحدد خلايا T التنظيمية من النوع 1 في البشر والفئران. نات. ميد. 19، 739-746 (2013). 40. باتشيلا، ل. أ.، مادسن، ل. ت. ودينز، ج. إ. سمية وفائدة استراتيجيات الجرعات المختلفة للإنترلوكين-2 في الميلانوما النقيلي وسرطان الخلايا الكلوية. ج. ممارسات متقدمة. أونكول. 6، 212-221 (2015). 41. موينيهان، ك. د. وآخرون. IL-2 المستهدف إلىالخلايا تعزز استجابات فعالة قوية للخلايا التائية ومناعة مضادة للأورام قوية. اكتشاف السرطان.https://doi.org/10.1158/2159-8290.CD-23-1266 (2024). 42. ماغناني، سي. إف. وآخرون. قتل خلايا APCs النخاعية عبر HLA الفئة I، CD2 وCD226 يحدد آلية جديدة من القمع بواسطة خلايا Tr1 البشرية. المجلة الأوروبية للمناعة 41، 1652-1662 (2011). 43. فيريس، س.ت. وآخرون. خلايا cDC1 تهيئ وتُرخص بواسطة خلايا T CD4 + لتحفيز المناعة المضادة للأورام. ناتشر 584، 624-629 (2020). 44. هوانغ، إكس. وآخرون. الاستخدام المختلف لمُعززات المثبط النسخي زيب2 يميز بين تكوين الدم البالغ والجنيني. المناعة 54، 1417-1432.e7 (2021). 45. ليو، تي. تي. وآخرون. إلغاء تطوير cDC2 بواسطة طفرات ثلاثية داخل معزز Zeb2. ناتشر 607، 142-148 (2022). 46. دوراي، ف. وآخرون. التنشيط الخفي لمُعزز Irf8 يحكم تحديد مصير cDC1. نات. إيمونول. 20، 1161-1173 (2019). 47. أوت، ب. أ. وآخرون. لقاح نيوأنتيجين شخصي مثير للمناعة للمرضى المصابين بالميلانوما. ناتشر 547، 217-221 (2017). 48. ويبر، ج. س. وآخرون. العلاج الفردي للنيوانتيجين mRNA-4157 (V940) بالإضافة إلى بيمبروليزوماب مقابل العلاج الأحادي ببيمبروليزوماب في الميلانوما المستأصلة (KEYNOTE-942): دراسة عشوائية، المرحلة 2ب. لانسيت 403، 632-644 (2024). 49. شاهين، U. وآخرون. لقاحات RNA المخصصة تحفز مناعة علاجية متعددة التخصصات ضد السرطان. ناتشر 547، 222-226 (2017). 50. كارينو، ب. م. وآخرون. علاج السرطان المناعي. لقاح الخلايا الجذعية يزيد من نطاق وتنوع خلايا T الخاصة بالنيوانتجين الميلانيني. ساينس 348، 803-808 (2015). 51. ساكسانا، م.، فان دير بورغ، س. هـ.، ميليف، س. ج. م. & بهاردواج، ن. لقاحات السرطان العلاجية. نات. ريف. كانسر 21، 360-378 (2021). 52. Zheng، ل. وآخرون. المشهد الخلوي الفردي للسرطان الشامل لخلايا T المتسللة إلى الورم. ساينس 374، abe6474 (2021). 53. أرم، ج. ب.، نوانكوا، س. & أوستن، ك. ف. التعرف الجزيئي على عائلة جديدة من أعضاء عائلة الأجسام المضادة البشرية التي تمتلك أنماط تثبيط تعتمد على التيروزين لمستقبلات المناعة وتشابه مستقبل التثبيط الفأري gp49B1. مجلة المناعة 159، 2342-2349 (1997). 54. بالانكا، سي. سي. وآخرون. حجب PD-1 يستعيد نشاط المساعدين للخلايا المناعية المتسللة إلى الورم والتي تعاني من الإرهاق PD-1سي دي 39خلايا T CD4. JCI Insighthttps://doi.org/10.1172/jci.insight. 142513 (2021). 55. مارتينيز-غوميز، سي. وآخرون. PD- المتعبة المتداولةقرص مضغوطتعتبر خلايا CD4 T المساعدة محددة لمستضدات الورم وتتنبأ بالاستجابة لحجب محور PD-1/PD-L1. السرطاناتhttps://doi.org/10.3390/cancers14153679 (2022). 56. لام، هـ. وآخرون. طريقة اختيار مستضد تجريبية تحدد المستضدات الجديدة التي إما تحفز استجابات واسعة من خلايا T المضادة للورم أو تدفع نمو الورم. اكتشاف السرطان. 11، 696-713 (2021).
ملاحظة الناشر: تظل شركة سبرينغر ناتشر محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
الوصول المفتوح هذه المقالة مرخصة بموجب رخصة المشاع الإبداعي النسب 4.0 الدولية، التي تسمح بالاستخدام والمشاركة والتكيف والتوزيع وإعادة الإنتاج بأي وسيلة أو صيغة، طالما أنك تعطي الائتمان المناسب للمؤلفين الأصليين والمصدر، وتوفر رابطًا لرخصة المشاع الإبداعي، وتوضح ما إذا كانت هناك تغييرات قد أُجريت. الصور أو المواد الأخرى من طرف ثالث في هذه المقالة مشمولة في رخصة المشاع الإبداعي الخاصة بالمقالة، ما لم يُشار إلى خلاف ذلك في سطر الائتمان للمواد. إذا لم تكن المادة مشمولة في رخصة المشاع الإبداعي الخاصة بالمقالة وكان استخدامك المقصود غير مسموح به بموجب اللوائح القانونية أو يتجاوز الاستخدام المسموح به، فسيتعين عليك الحصول على إذن مباشرة من صاحب حقوق الطبع والنشر. لعرض نسخة من هذه الرخصة، قم بزيارةhttp://creativecommons.org/licenses/by/4.0/. (ج) المؤلف(ون) 2024
طرق
فئران تم شراء ذكور الفئران WT 129S6 (للتجارب التي تتضمن خلايا T3 و F244) وإناث الفئران C57BL/6 (للتجارب التي تتضمن خلايا 1956) من مزرعة تاكونيك ومؤسسة تشارلز ريفر، على التوالي.على خلفية C57BL/6 وفئران Rag2-/(على خلفيتي 129S6 و C57BL/6) تم تربيتها وإيواؤها في منشأتنا الخالية من مسببات الأمراض المحددة. نقص cDC2ونقص cDC1 (IRF8تم الحصول على الفئران من سلالة C57BL/6 من K. Murphy (جامعة واشنطن في سانت لويس). تم الحصول على فئران knockout لـ Lilrb4a (التي تشفر gp49b) من M. Colonna (جامعة واشنطن في سانت لويس) وتم إنتاجها بواسطة مركز هندسة الجينوم والخلايا الجذعية المستحثة (GEiC) ونواة الفئران المعدلة وراثيًا وknockout والحقن الدقيق، قسم علم الأمراض والمناعة، في جامعة واشنطن في سانت لويس. تم إدخال RNA دليلي واحد (cctttagttgcagctcatccata) وبروتينات Cas9 عن طريق التحفيز الكهربائي في الزيغوتات قبل نقلها إلى إناث فئران B6/J الحوامل زيفيًا. تم فحص المؤسسين الناتجين باستخدام تسلسل الجيل التالي (NGS). تم تربية مؤسس ذكر لديه حذف مكون من 21 نيوكليوتيد في الإكسون 4 مع إناث فئران B6/J التي تم الحصول عليها من مختبر جاكسون (JAX stock #000664) لعدة أجيال، وتم تزاوج الفئران الهجينة F1 لإنتاج knockout. تم تأكيد سلامة الجين المتجانس بشدة Lilrb4b بواسطة NGS. تم تأكيد غياب تعبير بروتين LILRB4A في وحيدات الدم والعدلات بواسطة قياس التدفق باستخدام الأجسام المضادة H1.1.استنساخ (BioLegend). تم الحصول على فئران eGFP لتتبع السلالة على خلفية C57BL/6 من J. Kipnis (جامعة واشنطن في سانت لويس). تم استخدام الفئران بين 8 و 10 أسابيع من العمر. تم إجراء جميع التجارب الحية في منشأتنا الخالية من مسببات الأمراض المحددة باستخدام إجراءات معتمدة من لجنة الدراسات الحيوانية المعتمدة من AAALAC في جامعة واشنطن في سانت لويس واتبعت جميع اللوائح الأخلاقية ذات الصلة.
زراعة الورم
T3 و F244 و 1956 هي خلايا ساركوما مستحثة بواسطة MCA تم إنشاؤها سابقًا في مختبرنا في ذكور فئران 129S6 (خطوط خلايا T3 و F244) وإناث فئران C57BL/6 (خط خلايا 1956). تم زراعة خلايا الورم في وسط RPMI (Hyclone) معززة بـ 10% من مصل العجل الجيني (Hyclone). كانت جميع خطوط الخلايا المستخدمة سلبية بالنسبة للميكوبلازما وعوامل العدوى الأخرى. من أجل حقن الورم، تم جمع خلايا الورم عن طريق التحلل بالإنزيم، وغسلها ثلاث مرات بمحلول PBS وإعادة تعليقها في محلول PBS بكثافةخلايا لكل مليلتر. ثم،تم حقنها تحت الجلد في الجوانب الخلفية لفئران متلقية متجانسة. تم تحديد نمو الورم في الفئران الفردية باستخدام الكالبرات وعبر التعبير عنه كمتوسط لقطرين عموديين. تم euthanizing الفئران عندما وصل قطر الورم إلى 20 مم في أي اتجاه.
بروتوكول NeoAgs والتطعيم
تم وصف T3 neoAgs سابقًاوتم إدراجها في الجدول التكميلي 1. تم استخدام ببتيد فيروس الورم الحليمي البشري (DKCLKFYSKISEYRHY CYSLYGTTL) كأنتيجين غير ذي صلة بالأورام الساركوما. تم شراء جميع الببتيدات من Peptide 2.0 أو Genscript مع نقاء محدد يزيد عنكانت مستويات الإندوتوكسين في جميع الببتيدات المختبرة أقل من (لينكو). تم خلط الجرعات المحددة من الببتيد مع حمض البوليمر الإينوسينيك-البوليمر السيتيدينيك المعقد مع البوليمر-ل-لايسين والكاربوكسي ميثيل السليلوز (البوليمر-ICLC؛ أونكوفير) المخفف في محلول فوسفات البفر. من أجل التطعيم العلاجي، تم حقن الفئران الحاملة للأورام عن طريق الوريد أو تحت الجلد باللقاحات.في اليومين 6 و 17 بعد زراعة الورم. في التجارب التي استخدمت مزيج التطعيم والعلاج المناعي، تم إعطاء اللقاحات عن طريق الوريد في اليومين 3 أو 6 بعد زراعة الورم. في تجارب الفرز،تم خلط مضاد CD40 مع الجرعة الثانية من اللقاح لزيادة إنتاج المستضد المحدد.خلايا T.
علاج الأجسام المضادة في الجسم الحي
للعلاج داخل الجسم الحي، تم شراء أجسام مضادة أحادية النسيلة خالية من LPS وخالية من مسببات الأمراض ضد PD1 (IgG2a من الجرذان، النسخة RMP1-14)، وضد CTLA4 (IgG2b من الفئران، النسخة 9D9) أو ضد 4-1BB (IgG2a من الجرذان، النسخة 3H3) من شركة Leinco Technologies. تم تطعيم الفئران الحاملة للأورام في اليوم الثالث بعد زراعة الورم ثم تم حقنها عن طريق البطن بـكل جسم مضاد في الأيام 3 و6 و9. لتقييم دور IL-10 في تثبيط HDVax، تم شراء أجسام مضادة أحادية النسيلة لـ IL-10 (IgGl من الجرذان، النسخة JES5-2A5) و IL-10R (IgG1 من الجرذان، النسخة 1b1.3A) من Bioxcell وتم حقنها مع HDVax. لعلاج حجب anti-IL-10، تم معالجة الفئران بـيوم واحد قبل زراعة الورم، تلتها معالجة كل 3 أيام طوال مدة التجربة. بالنسبة لحجب IL-10R، تم معالجة الفئران بجرعة 1 ملغ لكل فأر كل 7 أيام، بدءًا من يوم واحد قبل حقن الورم. تم توفير الأجسام المضادة الأحادية النسيلة المحايدة ولكن غير المستنفدة المحددة لـ LILRB4 (النسخة # 2F1.F9.E6) من قبل ن. شارما وج. أليسون (جامعة تكساس (مركز أندرسون للسرطان))، وتم معالجة الفئران بـفي الأيام 3 و 6 و 9 بعد تلقيح الورم.
تلطيخ الرباعي
تلطيخ التترايمر لخلايا CD8+ T الخاصة بـ mLama4 وخلايا CD4 الخاصة بـ mItgb1تم إجراء خلايا T كما هو موصوف سابقًا. خلايا TILs أو خلايا الطحال (مليون)تم تحضين محلول FACS مع ببتيد mLama4 المسمى بـ PE/APC-رباعيلمدة 20 دقيقة. ثم، مزيج مضاد الأجسام الرئيسي الذي يتكون من CD90.2 و CD4 و CD11b وصبغة حية/ميتة (NIR) فيتم إضافة محلول FACS إلى كل بئر وتم حضنه لمدة 30 دقيقة عندتم الحصول على التترايمرات من مختبر مراقبة المناعة في مركز بورسكي بجامعة واشنطن في مدرسة الطب في سانت لويس.
تم حصاد TILs بعد 7 أيام من التطعيم، وتم صبغها بـ mItgbl-I-Aتترايمر تم تحضيره بواسطة مختبر مراقبة المناعة في جامعة واشنطن في سانت لويس باستخدام ببتيد mItgb1 المرتبط تساهميًا بـ-سلسلة، وإجمالي CD4 المحدد بـ mItgb1تم فرز الخلايا (الشكل 2ج) أو الخلايا التي تشغل مجموعات مختلفة (الشكل 3ج) بواسطة تحليل تدفق الخلايا. تم فرز الخلايا (تم تحفيزها في وسط خالٍ من المصل بـخلايا الطحال المشعة (المعزولة من فئران غير محصنة) التي تم تنشيطها بـتم قياس إفراز العديد من السيتوكينات باستخدام تقنية التدفق بعد .h الحضانة لوحة السيتوكين المخصصة ProcartaPlex (تقنيات لوميكس) وفقًا لبروتوكول الشركة المصنعة.
لتحديد استجابات الأجسام المضادة للببتيد mItgb1، تم طلاء ألواح ELISA بـإما mItgb1 أو SLP غير ذي صلة فيبين عشية وضحاها. تم غسل الأطباق وحجبها بـمصل الماعز لمدة ساعتين في درجة حرارة الغرفة، ثم أضيفت تخفيفات مختلفة من المصل من فئران LDVax أو HDVax لـتم غسل الأطباق مرة أخرى وتخزينها مع جسم مضاد من الماعز مرتبط بالبيروكسيداز لإجمالي IgG الفأري (H + L) (115-035-003، جاكسون إيمونوريسيرش) أو مواد ثانوية محددة لنوع الأيزوتوب مختلفة مرتبطة بالبيروكسيداز من الفجل لمدة ساعتين في درجة حرارة الغرفة، تلتها إضافة متتابعة لمادة الركيزة تيترا ميثيل بنزيدين (TMB). تم إيقاف التفاعل عن طريق التحمض، وتم قياس الكثافة الضوئية لكل بئر عند 405 نانومتر.
تجارب نقل التبني
لإجراء تجارب نقل التبني إلى فئران Rag2-/، تم تطعيم فئران WT بلقاح LDVax، وبعد 9 أيام، تم نقل 5 ملايين خلية T إجمالاً إلى فئران Rag2-/- التي تم حقنها بـخلايا الورم قبل يوم واحد. لعزل CD4 المحدد بـ mItgb1تم جمع الخلايا من فئران HDVax، وTILs والطحال من الفئران المعالجة بـ HDVax، وتم إثراء الخلايا باستخدام CD4طقم عزل الخلايا من ميلتيني (130-117-043) ملون بتترايمر ميتجب1، وCD25-negative mItgb1-specific CD4تم تنقية خلايا T باستخدام تقنية تحليل التدفق الخلوي. نصف مليون من خلايا CD4 المحددة بـ mItgb1تم حقن خلايا T داخل الورم (في ) إلى فئران Rag2-‘ الحاملة لورم T3.
لترتيب ونقل الأفراد من المجموعات الفردية،تم إثراء الخلايا بواسطة اختيار الخرز الإيجابي وتم صبغها بـ mItgb1-I-Abتترايمرات، CD25، CD200، CD153 و CD39.تم فرز الخلايا بناءً على التعبير الإيجابي لـ CD25، وتم تعريف الخلايا غير المجموعة 3/5 من خلال التعبير السلبي لـ CD25 والتعبير الإيجابي لـ CD200 و/أو CD153. تم فرز خلايا المجموعة 3 بناءً على التعبير السلبي لـ CD25 وCD200 وCD153 والتعبير الإيجابي لـ CD39. تم استخدام عدد متساوٍ من الخلايا (الخلايا) التي تم فرزها من هذه المجموعات تم إعادة تعليقها فيتم حقن PBS داخل الورم. لتصنيف الخلايا التي تعبر عن LILRB4 و SEMA4، تم استخدام CD25 و CD200 و CD39 لتحديد الخلايا في المجموعة 3، وتم حقن الخلايا في فئران Rag2-/- الحاملة لورم T3 التي تلقت أيضًاخلايا T الكلية المستنفدة من الخلايا من فئران LDVax.
تلطيخ السيتوكينات داخل الخلوية وCD40L
خلايا الطحالتم حصادها من الفئران الساذجة وتم تعريضها للإشعاع (30 غيغا) وتم تنبيهها بـببتيد، وTILs ) تم إضافتها لاحقًا، وتم حضن تعليق الخلايا عند تم إضافة GolgiPlug (BD Biosciences) بعد ساعة واحدة وتم حضنه لمدة 4 ساعات أخرى. تم صبغ الخلايا لعلامات سطحية مختلفة، بما في ذلك علامة الحياة/الموت (NIR)، ثم تم اختراقها باستخدام مجموعة اختراق الخلايا الداخلية (BD Biosciences)، تلاها صبغ لـ IFN.TNF و CD40L (النسخة؛ SA047C3؛ 157010، BioLegend).
اختبار تقديم المستضد خارج الجسم
تم تطعيم فئران WT الساذجة عن طريق الوريد بلقاح HDVax الذي يتكون منمن mItgb1 SLP و بولي-ICLC (HDVax). في الوقت المحدد، تم جمع الطحال والعقد اللمفاوية وهضمها بوجود الكولاجيناز. تم فرز خلايا APCs باستخدام تقنية FACS إلى أربع مجموعات فرعية مختلفة: و.APCs مرتبة ( ) تم حضنها مع خلايا الهجين المحددة لـ mItgb1في ألواح ELISPOT المطلية مسبقًا بـ IL-2 (IMMUNOSPOT). بعد أربع وعشرين ساعة، تم تطوير الألواح وفقًا لبروتوكول الشركة المصنعة، وتم قياس بقع IL-2 باستخدام جهاز CTL ImmunoSpot S6.
اختبار قتل الخلايا الشجرية في المختبر
لعزل الخلايا الشجرية، تم جمع الطحال من الفئران البرية السليمة وتم تكسيره بواسطة هضم الكولاجيناز. CD11cتم إثراء الخلايا باستخدام مجموعة اختيار إيجابية (130-125-835، ميلتني بيوتيك). تم إثراء CD11cتم تحفيز الخلايا بـ mItgb1 SLP عند وتم حضنه مع CD4 المحدد للأورامتمت زراعة خلايا T بنسبة 1:1 لمدة 12 ساعة. تم غسل الخلايا مرتين بمحلول PBS، وتم صبغها بمؤشرات MHC-II وCD11c وXCR1 وCD172a وصبغة Zombie NIR لتحديد تردد NIR في cDC1s وcDC2s باستخدام تقنية تحليل التدفق الخلوي.
CD8-IL-2
تم توفير CD8-IL-2 من قبل Asher Biotherapeuticsتم إنتاج CD8-IL-2 من خلال دمج ميوتين IL-2 الذي لا يرتبط بـ IL-2R.ويظهر ارتباطًا مخفضًا بشكل ملحوظ مع IL-2Rتم ربط هذا المتغير بعد ذلك بمضاد أحادي التكافؤ مضاد للفأر CD8. تم تحقيق ربط IL-2 أحادي التكافؤ باستخدام تقنية زوج الشحن ثنائي الخصوصية، وتم إلغاء الربط من خلال تغييرمنطقة الربط للأجسام المضادة المضادة لـ CD8. تم التعبير عن CD8-IL-2 في خلايا HEK293 وتم تنقيته عبر كروماتوغرافيا الارتباط بالبروتين A، تليها كروماتوغرافيا تبادل الأيونات ثم كروماتوغرافيا فصل الحجم. الجرعات العلاجية ( ) من CD8-IL-2 أدى إلى نشاط مضاد للأورام بشكل كبير (حوالي معدل الاستجابة) ضد أورام T3 التي تم تأسيسها لمدة 8 أيام عندما أصبحت غير حساسة لعلاج مضاد PD1. في الدراسة الحالية، تم إعطاء CD8-IL-2 بجرعة دون العلاجمخفف في PBS وتم حقنه داخل الصفاق. تم شراء WT IL-2 (202-IL/CF) من (R&D Systems). تم علاج كل فأر بـ 25,000 وحدة دولية في PBS تم حقنها داخل الصفاق يوميًا لمدة 5 أيام.
تحليل تسلسل RNA أحادي الخلية
تجميع UMAP وفصل الخلايا الكلية والخلايا المحددة للمستضد. تم علاج الفئران الحاملة لورم T3 بـ HDVax أو LDVax أو PBS بعد 6 أيام من زراعة الورم. بعد سبعة أيام، تم إعداد تعليقات خلوية مفردة من TILs (مجمعة من سبعة فئران لكل مجموعة). إجمالي CD4تم إثراء خلايا T باستخدام CD4مجموعة اختيار إيجابي لخلايا T (ميلتيني). CD4 الغنيتم تقسيم خلايا T إلى قسمين. تم وسم أحد القسمين بـ TCR (TotalSeq-C0120 مضاد لمستقبلات TCR الفأرية أجسام مضادة سلسلة، 109259، BioLegend) واستخدمت كمصدر لإجمالي CD4خلايا T. تم وسم الجزء الآخر بـ CD90.2 (أجسام مضادة مضادة للفأر CD90.2 TotalSeq-C0075، 105353، BioLegend) واستخدم لعزل خلايا محددة لـ mItgb1 بناءً على mItgb1-I-A.تلطيخ التترا مير والفرز بالتدفق. الفئتين (الإجمالية والمحددة بمستضد CD4تم خلط الخلايا فينسبة وتم تقديمها إلى معهد الجينوم في جامعة واشنطن لإنشاء مكتبات 10 X باستخدامتم استخدام تسلسل RNA أحادي الخلية v 2. تم استخدام المحاذاة، وتعيين الرموز الشريطية، وعدّ المعرفات الجزيئية الفريدة باستخدام Cell Ranger (v6.1.1) لإعداد مصفوفات العد لمكتبة التعبير الجيني باستخدام الجينوم الفأري (GRCm38) كمرجع.تم تصفية رموز الباركود في جميع العينات التي تمثل خلايا منخفضة الجودة باستخدام استراتيجية الانعطاف القياسية المتاحة في Cell Ranger. للتحليل اللاحق، تم استخدام حزمة Seurat (الإصدار 4.0.4)؛ وتمت تصفية الجينات المعبر عنها في أقل من ثلاث خلايا من مصفوفات التعبير، وتمت إزالة الخلايا التي تحتوي على أقل من 200 جين معبر عنه. تم حساب نسبة الجينات الميتوكوندرية لجميع الخلايا، وتمت تصفية الخلايا التي كانت تحتوي على نسبة ميتوكوندرية أكثر من أعلى فترة ثقة لنسبة الميتوكوندرية المقاسة. تم تطبيع كل عينة باستخدام دالة SCTransform مع محتوى الميتوكوندريا كمتغير للتراجع عنه في تحليل انحدار خطي غير منتظم ثانٍ. لأغراض الدمج، تم تحديد الجينات المتغيرة عبر العينات بواسطة دالة SelectIntegrationFeatures مع عدد من الميزات يساوي 2000. ثم، تم إعداد الكائن للدمج (دالة PrepSCTIntegration)، وتم العثور على المراسي (دالة FindIntegrationAnchors) وتم دمج العينات في الكائن الكامل (دالة IntegrateData).تم استخدام تحليل المكونات الرئيسية (PCA) لتقليل الأبعاد، وتم استخدام أول 20 مكونًا رئيسيًا لاحقًا لتوليد تقليل الأبعاد باستخدام UMAP من خلال وظيفة RunUMAP. تم تنفيذ إجراء التجميع بواسطة FindNeighbors و FindClusters. مع مجموعة من الدقة (من 0.2 إلى 1.0، مع 0.2 كخطوة) وأول 20 مكونًا رئيسيًا كمدخلات. تم تطبيع بيانات العلامات المشتقة من الأجسام المضادة (ADT) بواسطة طريقة تحويل النسبة اللوغاريتمية المركزية، وتم قياسها وتحويلها إلى فضاء PCA الخاص بها. أخيرًا، تم تعريف الخلايا على أنها خلايا محددة لـ mItgb1 وخلايا إجمالية (تتكون في الغالب من خلايا غير محددة لـ mItgb1) بناءً على القيمة المقاسة للـ ADTs المقابلة. تم تنظيف كل كائن بشكل تكراري من الثنائيات وكذلك الخلايا ذات الجودة المنخفضة.
تحليلات بيانات TCR
تم محاذاة بيانات TCR إلى الجينوم المرجعي للفأر GRCm38 وتم عدها باستخدام Cell Ranger (الإصدار 6.1.1) سير عمل vdj.تم تحويل كائن Seurat إلى تنسيق h5ad، وتم دمج الخلايا التي اجتازت مراقبة الجودة لمستويات التعبير الجيني وTCR في الكائنات (خلايا محددة mItgb1 والخلايا الكلية). تم تنفيذ التحليلات اللاحقة، بما في ذلك تحليل توسع الكلونات وتقدير أزواج جينات VDJ المستخدمة، باستخدام مجموعة أدوات scirpy. تم حساب نسب الكلونات المشتركة والفريدة معتم عد الخلايا (العنقود 5). تم إجراء تحليلات TCR باستخدام حزمة Immunarch.
تسلسل RNA بكميات كبيرة
ثلاث مجموعاتتم تطعيم الفئران الحاملة لورم T3 بـ HDVax، وبعد 7 أيام تم قياس CD4تم إثراء خلايا T من TILs باستخدام CD4 الفأري من ميلتيني.طقم عزل خلايا T، مصبوغ بـ mItgb1-I-Aرباعي ثم تم فرزه إلى ثلاث نسخ من CD25 ( الخلايا)،(غير-الخلايا/غير المجمعة 3) و(العنقود 3). تم تحلل الخلايا، وتم استخراج الرنا المرسال باستخدام Nucleospin RNA Plus (740990.50، تاكارا). تم تسلسل الرنا بواسطة معهد جينوم جامعة واشنطن. باختصار، تم تحديد سلامة الرنا الكلي باستخدام جهاز تحليل البيولوجيا من أجيلنت. تم إعداد المكتبة باستخدام 10 نانوغرام من الرنا الكلي مع درجة رقم سلامة الرنا (RIN) أكبر من 8.0. تم إعداد الحمض النووي مزدوج الشريطة (ds-cDNA) باستخدام مجموعة SMARTer Ultra Low RNA لتسلسل إيلومينا (740990.50، تاكارا) وفقًا لبروتوكول الشركة المصنعة. تم تجزئة cDNA باستخدام جهاز سونيكاتور كوفاريس E220 باستخدام طاقة ذروة حادثة تبلغ 18، وعامل واجبتمت معالجة cDNA بحيث كانت نهاياتها مستقيمة، وتم إضافة قاعدة A إلى النهايات 3’، ثم تم ربط محولات تسلسل Illumina بالنهايات. تم تضخيم الأجزاء المربوطة لمدة 12-14 دورة باستخدام بادئات تحتوي على علامات فريدة مزدوجة. تم تسلسل الأجزاء على جهاز Illumina NovaSeq-6000 باستخدام قراءات مزدوجة الطرف تمتد 150 قاعدة. بالنسبة للتحليل، تم محاذاة البيانات الخام وعدها إلى الجينوم المرجعي GRCm 38.101 بواسطة سير عمل Dragen من خدمة GTAC@MGI. للتحليل اللاحق، تم اختيار 12,000 جين بأعلى عدد من العد عبر العينات. تم تطبيع جميع العينات بواسطة تحويل اللوغاريتم المنتظم (وظيفة rlogTransformation) وتحويل تثبيت التباين (وظيفة vst)، والتي تم إنجازها في حزمة DESeq2 (v1.30.1). تم تشغيل PCA بناءً على مخرجات vst، ولم يتم تحديد أي نقاط شاذة.تم تقليص قيم التغير النسبي لجميع المقارنات (CD25 مقابل CD39، وCD200 مقابل CD39) بواسطة دالة IfcShrink، وتم استخدام ناتجها لبناء مخطط البركان..
تحليل توقيع الجينات
لتقييم أهمية الإثراء للجينات ذات التعبير العالي ضمن المجموعة 3، حددنا أعلى 200 جين معبر عنه بشكل مختلف من مجموعة بيانات RNA-seq الكثيفة (الجدول التكميلي 2). بعد ذلك، قمنا بإجراء تحليل دقيق للتعبير الجيني المختلف باستخدام حزمة limma على بيانات العد المعيارية المستمدة من مجموعات بيانات متاحة للجمهور. على وجه التحديد، قمنا بإجراء المقارنات الزوجية التالية.
لـ CD4 السامة للخلاياتم تحليل بيانات GSE141540، وتم إجراء مقارنة لـ خلايا Tخلية-سياق مقابلتم إجراء الخلية. بالنسبة لتوقيع جين Tr1، تم الحصول على GSE158703 و GSE139990، و مقارنة Tr1-IL-10 ضد أو تم إجراء مقارنة بين Tr1 و naive، على التوالي.
لـ و تم الحصول على توقيع الجين، GSE68242 أو GSE140187، وتم إجراء مقارنة بين FOXP3 مقابل غيره أو WT T في العقد اللمفية.تم إجراء مقارنة بين العقد اللمفاوية غير المدربة. بالنسبة لخلايا CD4 المستنفدة التي تم تحفيزها بشكل مزمنتم تحليل بيانات GSE30431 الخاصة بخلايا T، وتم إجراء مقارنة بين CD4 D30 المزمن وCD4 D30 الحاد. استخدمنا GSEA لتحديد أهمية الثراء لتوقيع الجينات المرتبط بالعناقيد 3. هذه الطريقة تقيم ما إذا كانت توقيعات الجينات المحددة مسبقًا للعناقيد 3 مفرطة التمثيل إحصائيًا أو ناقصة التمثيل ضمن قوائم الجينات المعبر عنها بشكل مختلف المستمدة من مجموعات بيانات RNA-seq العامة المتاحة لكل CD4.نوع فرعي من خلايا T.
لتحديد تردد خلايا Tr1-like في المرضى المصابين بالسرطان، نقوم أولاً بتقسيم الخلايا التي تم التحكم في جودتها من كل مخطوطة إلىالخلايا. بعد ذلك، استخدمنا AUC ellلتحديد أي الخلايا تتطابق مع مجموعة جينات Tr1 البشرية. بعد ذلك، حددنا نسبة خلايا Tr1 ضمن خلايا CD4.تعداد خلايا T لكل مريض. قمنا باختبار الفروق في نسب خلايا Tr1 بين المستجيبين وغير المستجيبين باستخدام اختبار ستودنت.-اختبارات دون افتراض تساوي التباينات. هناك فرق كبير في نسبة خلايا Tr1 بين المستجيبين وغير المستجيبين من مجموعة بيانات سادي-فيلدمان وآخرون.، د.ف.لم يكن هناك فرق كبير في مجموعة بيانات عوض وآخرون. ).
تحليل GSEA لمسار IL-2
تم استخدام دالة FindMarkers للعثور على الجينات المحددة لكل من عينات LDVax و HDVax. المتوسطتم استخدام قيم التغير المطوي كمدخل في GSEA المنفذ في حزمة R fgsea (الإصدار 1.16.0). تم استخدام ثلاثة وأربعين جينًا مرتبطًا بمسار إشارة IL-2 لرسم غنى مجموعة الجينات.بالنسبة لمخططات الكمان، تم حساب التعبير الملخص لجينات إشارات IL-2 باستخدام اختبار sctransform (SCT) (فتحة البيانات)، وتم قياسه وتقسيمه على عدد الجينات في المسار.
الإحصائيات
تم استخدام برنامج GraphPad Prism (الإصدار 10.2.2) لإجراء جميع التحليلات الإحصائية. تم تحديد الدلالة باستخدام تحليل التباين الأحادي مع مقارنات متعددة تم تصحيحها بطريقة توكي ما لم يُذكر خلاف ذلك في أساطير الأشكال.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
تم إيداع بيانات تسلسل RNA الضخم وبيانات تسلسل RNA أحادي الخلية باستخدام رقم الوصول GSE268302؛ يمكن استخدام GSE268300 (تسلسل RNA الضخم) أو GSE268301 (تسلسل RNA أحادي الخلية) للوصول إلى البيانات الخام. تتوفر خطوط الأنابيب مع الشيفرة لمعالجة وتحليل بيانات تسلسل RNA الضخم وبيانات تسلسل RNA أحادي الخلية فيhttps://github.com/vdsukhov/neoantigen_vaccines_2024تم توفير بيانات المصدر مع هذه الورقة. 57. وانغ، ل. ل.، تشو، د. ت.، دوكون، أ. أ. ويوكوياما، و. م. التعبير القابل للتحفيز لمستقبل gp49B المثبط على خلايا NK. مجلة المناعة 164، 5215-5220 (2000). 58. زينغ، ج. إكس. وآخرون. التوصيف الرقمي المتوازي بشكل كبير للتعبير الجيني للخلايا الفردية. نات. كوميونيك. 8، 14049 (2017). 59. ستيوارت، ت. وآخرون. التكامل الشامل لبيانات الخلايا المفردة. خلية 177، 1888-1902.e21 (2019). 60. لوف، م. آي.، هوبر، و. & أندرس، س. تقدير معتدل لتغير الطي والتشتت لبيانات RNA-seq باستخدام DESeq2. جينوم بيو. 15، 550 (2014). 61. أيبار، س. وآخرون. SCENIC: استنتاج الشبكة التنظيمية على مستوى الخلية الواحدة والتجميع. نات. ميثودز 14، 1083-1086 (2017). 62. ستورم، ج. وآخرون. سكيري: امتداد لسكاني لتحليل بيانات تسلسل مستقبلات الخلايا التائية أحادية الخلية. المعلوماتية الحيوية 36، 4817-4818 (2020).
الشكر والتقدير نشكر جميع أعضاء مختبر شرايبر على المناقشات وتقديم الدعم الفني؛ ل. يانغ و د. بيندر من مختبر المناعة. (IML) الذي قدم التترايمرات لـ MHC-I و MHC-II neoAgs وأجرى تقييمات السيتوكين متعددة المعلمات؛ مركز هندسة الجينوم والخلايا الجذعية المستحثة (GEiC) في جامعة واشنطن في سانت لويس لخدمات التحقق من RNA الدليل؛ م. وايت لتوليد فئران Lilrb4aknockout؛ و ج. كيبنيس و ج. ج. دافانزو لتوفير فئران تتبع سلالة FOXP3. تم دعم هذا العمل من خلال منح لـ R.D.S. من المعهد الوطني للسرطان التابع للمعاهد الوطنية للصحة في الولايات المتحدة (RO1CA190700)، معهد باركر لعلاج السرطان المناعي، ومنحة بحثية ترجمة فريق أحلام تقارب مؤسسة سرطان البنكرياس Stand Up to Cancer-Lustgarten Foundation. Stand Up to Cancer هو برنامج تابع لمؤسسة صناعة الترفيه يديره الجمعية الأمريكية لأبحاث السرطان. K.M.M. مدعوم من المعاهد الوطنية للصحة (RO1Al150297، RO1CA248919، R21AI164142، RO1Al162643 و R21Al163421). M.C. مدعوم من مركز موارد أبحاث الأمراض الروماتيزمية في جامعة واشنطن (NIH/NIAMS P3OARO73752). H.S. تم دعمه من خلال منحة تدريب ما بعد الدكتوراه (T32CA009547) من المعهد الوطني للسرطان. M.F. تم دعمه من قبل وزارة العلوم والتعليم العالي في الاتحاد الروسي (الاتفاق رقم 075-15-2022-301). J.P.W. مدعوم من المعهد الوطني للسرطان التابع للمعاهد الوطنية للصحة (5K08CA245215). يتم دعم IML من قبل مركز أندرو م. وجين ن. بورسكي لعلم المناعة البشري وبرامج العلاج المناعي ومركز ألفين ج. سيتيمان الشامل للسرطان. الأخير مدعوم من منحة دعم مركز السرطان من المعهد الوطني للسرطان التابع للمعاهد الوطنية للصحة (P30CA91842).
مساهمات المؤلفين: قام H.S. و R.D.S. بتصميم التجارب وتفسير النتائج وكتابة المخطوطة. قام H.S. بإجراء التجارب وجمع البيانات. قام Y.T. و J.P.W. بتحديد neoAgs في أورام 1956 و F244 على التوالي. قام V.S. و M.F. بإجراء تحليلات scRNA-seq و bulk RNA-seq و GSEA تحت إشراف M.N.A. قام T.-T.L. و K.M.M. بإنشاء وتوفيرالفئران. قام D.K. و S.B. و M.C. بتوليد وتوفير الفئران المعدلة وراثيًا Lilrb4-knockout. قدم N.S. و J.P.A. الأجسام المضادة الأحادية النسيلة LILRB4 المحايدة. قدم C.D.A. و J.M.W. و Y.S. و H.K. المساعدة الفنية طوال الدراسة. أشرف K.C.F.S. على الدراسات التي أجريت في IML. قدم A.M.S. بوليمر-ICLC من الدرجة السريرية. قدم K.D.M. و Y.A.Y. و I.D. CD8-IL-2. شارك T.N.S. في تطوير CD8-IL-2. جميع المؤلفين ساهم في التطوير النهائي للمخطوطة ووافق على النتائج والاستنتاجات.
المصالح المتنافسة: R.D.S. هو أحد المؤسسين، وعضو في مجلس الاستشارات العلمية، ومساهم ومتلقٍ للحقوق من Asher Biotherapeutics، وهو عضو مدفوع في مجلس الاستشارات العلمية لشركات A2 Biotherapeutics وNGM Biopharmaceuticals وBioLegend وSensei Biotherapeutics وNeuvogen وBlueSphere Biopharmaceuticals. K.M.M. هو عضو في مجلس الاستشارات العلمية لشركة Harbour Biomed. J.P.A. يتلقى أتعاب استشارية من Achelois وAdaptive Biotechnologies وApricity وBioAtla وBioNTech وCandel Therapeutics وCodiak وDragonfly وEarli وEnable Medicine وHummingbird وImaginAb وJounce وLava Therapeutics وLytix وMarker وPBM Capital وPhenomic AI وPolaris Pharma وTime Bioventures وTrained Therapeutix وTwo Bear Capital وVenn Biosciences؛ ويمتلك أسهماً في Achelois وAdaptive Biotechnologies وApricity وBioAtla وBioNTech وCandel Therapeutics وCodiak وDragonfly وEarli وEnable Medicine وHummingbird وImaginAb وJounce وLava Therapeutics وLytix وMarker وPBM Capital وPhenomic AI وPolaris Pharma وTime Bioventures وTrained Therapeutix وTwo Bear Capital وVenn Biosciences. T.N.S. هو أحد المؤسسين، وعضو في مجلس الاستشارات العلمية، ومساهم في Asher Biotherapeutics وNeogene Therapeutics، وأحد المؤسسين ومساهم في Cell Control Bio، ومستشار ومساهم في Third Rock Ventures، وعضو في مجلس الاستشارات العلمية لشركات Allogene Therapeutics وCelsius وMerus وScenic Biotech. K.D.M. وY.A.Y. وI.D. هم موظفون بدوام كامل في Asher Biotherapeutics. R.B. وH.A.C. هما موظفان بدوام كامل في Natera, Inc. جميع المؤلفين الآخرين يعلنون عدم وجود مصالح متنافسة.
معلومات إضافية
معلومات إضافية النسخة الإلكترونية تحتوي على مواد إضافية متاحة فيhttps://doi.org/10.1038/s41586-024-07752-y. يجب توجيه المراسلات والطلبات للحصول على المواد إلى روبرت د. شرايبر. تشكر مجلة نيتشر أكسل كالييس والمراجعين الآخرين المجهولين على مساهمتهم في مراجعة الأقران لهذا العمل. معلومات إعادة الطباعة والتصاريح متاحة علىhttp://www.nature.com/reprints.
الشكل البياني الممتد 1|الملفات الظاهرية الناتجة عن HDVax مقابل LDVaxالخلايا. أ، مخططات تدفق تمثيلية تظهر صبغ التترايمر mitgbl في CD4تيلز. ب، التردد (يسار) والأعداد (يمين) من CD4 المحدد لـ mItgb1تم تقييم TILs في أورام T3 في اليوم 13، تمثل خمسة تجارب). ج، مخططات تدفق الخلايا التمثيلية تظهر صبغ CD25 و Foxp3 في CD4 المحدد بـ mItgb1تيلز. د، تردد CD25فوكس بي 3الخلايا في mItgb1-specificالخلايا في TILs (يسار) والطحال (يمين) من الفئران الحاملة لورم T3، تمثل خمسة تجارب). هـ، تردد CD25فوكس بي 3تريغس في CD4خلايا T في الأورام (يسار) والطحال (يمين) لفئران F244 الحاملة للأورام، تمثيل ثلاثة تجارب). ف، مخططات تدفق تمثيلية تظهر تكرار IFNTNFفي CD4تيلز.تكرار IFN-TNF-التعبير عن CD4تيلز في عدة فئران“، مستمدة من تجربتين). ح، مخططات تدفق تمثيلية تظهر تعبير CD40L في CD4 TILs. أنا، تردد تعبير CD40L في CD4تيلز الأورام T3 بعد التنشيط في المختبر، مجمعة من تجربتين). البيانات في جميع الألواح معروضة كمتوسطاتس.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع تصحيحات المقارنات المتعددة باستخدام طريقة توكي.
الشكل البياني الموسع 2 | توصيف الخلايا الشجرية و CD8TILs في الفئران المعالجة بـ HDVax و LDVax و PBS. أ، مخططات تدفق تمثيلية تظهر استراتيجية التصفية لـ cDC1 و cDC2. ب، تعبير CD86 و CD70 و MHC-II و CD40 على سطح الخلايا الشجرية (CD45CD90.2CD11cMHC-II) في أورام T3 التي تم جمعها بعد 5 أيام من التطعيم ( ، تمثل تجربتين). ج، أعداد cDC1 (في الأعلى) و cDC2 (في الأسفل) في أورام T3 التي تم تقييمها بعد خمسة أيام من التطعيم (تمثل ثلاثة تجارب). مخططات تدفق الخلايا التمثيلية التي تظهر استراتيجية التصفية لـ CD8 المحدد لـ mLama4تيلز في اليوم 13 ت3 تيلز. هـ-، تردد CD8 المحدد لـ mLama4تيلز التعبير عن e، TOX f، PD-1/CD39 و g، TIM-3/LAG3 (، تمثل تجربتين).تكرار الخلايا التي تعبر عن IFN-TNF-فيCD8 المحددالخلايا التائية بعد التنشيط في المختبر مع خلايا الطحال المنشطة بـ mLama4 SLP في وجود سدادة غولجي تليها صبغة داخل الخلايا (، مجمعة من ثلاثة تجارب). i، تردد CD8 المحدد بـ mLama4تظهر TILs TIM3 و TCF-1. البيانات في جميع اللوحات معبّرة كمتوسطاتس.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع تصحيحات المقارنات المتعددة باستخدام طريقة توكي.
الشكل 3 من البيانات الموسعة | الناتج عن HDVaxتفتقر خلايا T إلى توقيع جين Th1 وتعبر عن مستويات عالية من GZMB و CCL5. أ-ب، مخططات كمان تظهر مستويات التعبير عن عدة جينات متورطة فيالإرهاق أوتوقيع المؤثر Th1 في بيانات scRNAseq المحددة لـ mItgbl. ج، GSEA تظهر التعبير النسبي للجينات المشاركة في مسار إشارة IL-2. د، مخططات violing تظهر مستويات التعبير لعوامل النسخ المختلفة المشاركة في إشارة IL-2 في بيانات scRNAseq المحددة لـ mItgb1. هـ-و، التردد (يسار) والأعداد (يمين) من CD4 المعبر عن CCL5/GZMBخلايا T في، TILs و طحال الفئران الحاملة لورم T3 بعد 5-7 أيام من التطعيمات (TILs) و 5 (الطحال) وتمثل ثلاث تجارب). ج-ح، التردد (يسار) والأعداد (يمين) من
CD4 الذي يعبر عن CCL5/GZMBخلايا T في، TILs و طحال الفئران WT الحاملة لورم F244 بعد سبعة أيام من التطعيماتيمثلان تجربتين). i، تردد CTLA-4 في مجموعات مختلفة من CD4 الخاصة بـ mItgb1تيلز من فئران WT الحاملة لورم T3، تمثل ثلاثة تجارب).تكرار تعبير CCL5/GZMB في CD4تيلز من فئران WT الحاملة لورم T3 المعالجة بـ HDVax بالإضافة إلىPD-1 أوCTLA-4تمثل ثلاثة تجارب). تُعرض البيانات في جميع الألواح كمتوسطاتتم إجراء الإحصائيات باستخدام تحليل التباين الأحادي مع تصحيحات المقارنات المتعددة بطريقة توكي في e-h و j، وتم إجراء تحليل التباين الثنائي مع اختبار تصحيح المقارنات المتعددة بطريقة سيداك للإحصائيات في.
الشكل 4 من البيانات الموسعة | استراتيجية التصفية والفرز لتغنيص خلايا المجموعة 3 بشكل جزئي. أ، رسم بياني يوضح تعبير مجموعة متنوعة من الجينات متراكبًا علىCD4 المحدد mItgb1توزيع TILs UMAP. ب، مخططات تدفق الخلايا التمثيلية توضح استراتيجية الفرز لمجموعات مختلفة في CD4 المحدد بواسطة mItgb1TILs. ج، رسم بياني تمثيلي لتدفق الخلايا يوضح تعبير CD25 وCD39 وFoxp3 في مجموعات مختلفة بعد الفرز. د، نمو T3 في فئران RAG2-/- التي تلقت خلايا T الكلية من فئران WT المعالجة بـ LDVax. ) وحده أو زائد؛ خلايا المجموعة 3 المستحثة بواسطة LDVax؛ المستحثة بواسطة HDVax،
CD25-negative محدد mItgb1؛ أو متعدد النسائل (غير محدد mItgb1) CD4خلايا T ). البيانات معبّرة كمتوسط قطر الورم س.م.، تجربتين).تم حساب القيم باستخدام تحليل التباين ثنائي الاتجاه مصححًا باختبار المقارنات المتعددة لسيداك. على سبيل المثال، تكرار LILRB4 إيجابي مفرد، LILRB4/SEMA4a إيجابي مزدوج أو SEMA4a إيجابي مفرد يعبر عن CD4 المحدد بـ mItgb1.TILs (تمثل ثلاثة تجارب). يتم التعبير عن البيانات كمتوسطتم إجراء الإحصائيات باستخدام تحليل التباين الأحادي مع تصحيحات المقارنات المتعددة بطريقة توكي.
الشكل البياني الممتد 5| انظر الصفحة التالية للتعليق.
الشكل 5 من البيانات الموسعة | الخلايا التي تشكل العنقود-3 متميزة عن خلايا Tregs. أ، تحليل GSEA يقارن توقيع الجينات لخلايا العنقود-3 (تسلسل RNA الكلي) بتوقيع الجينات لمختلف CD4مجموعات خلايا T. تم استخدام درجة الثراء لتحديد-تحديد القيمة لكل مجموعة جينات بواسطة اختبار التبديل. ب، رسم بياني لتدفق الخلايا يظهر صبغ CD25 وFoxp3 في خلايا Tregs المفروزة والخلايا التي تشغل الكتلة-3 بعد 10 أيام من الحضانة في المختبر مع IL-2 فقط ( )، أو زائد IL-10 و أو . يمثل التجربة تجربتين. ج، أنماط تكرار مستقبلات الخلايا التائية في كل مجموعة من I- الناتجة عن HDVax-mItgb1 محدد CD4تم تحديد T بواسطة TCRseq ومقارنتها مع أنواع TCR في الخلايا التي تشغل الكتلة-3. د، تردد الخلايا التي تعبر عنفي Tregs (CD25فوكس بي 3) أو CD25 LILRB4CD4خلايا T بعد علاجات مختلفة في الطحال وTILs، تمثل تجربة واحدة). هـ، تردد الخلايا التي تعبر عن LAG3 و CD49b في الكتلة-3 أو غير الكتلة-3/5 من محدد mItgb1الخلايا بعد علاجات مختلفة في الطحال و TILsتمثل ثلاثة تجارب). البيانات معبّرة كمتوسط قطر الورمتم إجراء s.e.m. والإحصائيات بواسطة تحليل التباين الثنائي الاتجاه مصحح باستخدام طريقة سيداكس المتعددة اختبارات المقارنات. CD4 المعزولة حديثًاتم تحفيز خلايا T من مجموعات مختلفة بـmltgb1 SLP لمدة 72 ساعة، وإنتاج الأشكال النشطة من TGFتم تقييمه بواسطة Multiplex (نسخ بيولوجية)في الكتلة – 5 و 3 تكرارات في جميع الكتل الأخرى وكل القيم هي متوسط تكرارين تقنيين، تمثل تجربتين). الإحصائيات في و تمت باستخدام تحليل التباين الأحادي مع تصحيحات المقارنات المتعددة باستخدام طريقة توكي. ج، خريطة حرارية تظهر مستويات إنتاج السيتوكينات المختلفة (المشار إليها على المحور السيني للخريطة الحرارية). الأرقام تمثل متوسط ثلاث تكرارات بيولوجية مستقلة وتمثل تجربتين. ح، نمو ورم T3 في الفئران WT المعالجة بـ HDVax أو لقاحات غير ذات صلة في اليوم السادس و IL-10 (-1، 4، 7، 10 و 14) بعد حقن الورم T3 (، تمثل تجربتين). i، نمو ورم T3 في الفئران WT المعالجة بـ HDVax في اليوم الثالث و IL-10R (-1,5,11 و 18) بعد زراعة الورم ، تمثل تجربة واحدة). البيانات في و معبرًا عنه كمتوسط قطر الورمس.م.تم حساب القيم باستخدام تحليل التباين ثنائي الاتجاه مصححًا باختبار المقارنات المتعددة لسيداك.
الشكل البياني الموسع 6 | متعلق بالشكل 4:علاج LILRB4 يعكس تحفيز HDVax لـ GZMB/CCL5 وكبت إنتاج IL-2. أ، 1956 نمو في RAG2الفئران التي تتلقى LILRB4 الكليخلايا T من المعالجة بـ LDVax
ليلرب4فئران بالإضافة إلى CD4 السلبية CD25 المستحثة بواسطة HDVaxخلايا T ) من فئران WT أو LILRB4 فئران (لـ PBS و LDVax) 7 لمجموعات أخرى). ب، نمو ورم F244 في الفئران WT المعالجة بـ HDVax و LILRB4 كما هو موضح (، تمثل تجربتين). ج، نمو الورم في الفئران المعالجة بـ HDVax في عام 1956 و LILRB4 كما هو موضح (، تمثل تجربتين). د، تردد الخلايا التي تعبر عن LILRB4 في CD4ت3 تي آي إل بعد العلاج مع LILRB4 أو الأجسام المضادة التحكمية ( يمثل ثلاثة تجارب). e، تردد وأعداد CD4 التي تعبر عن GZMB/CCL5الخلايا التائية المستحثة في الفئران الحاملة لورم F244 (، تمثل تجربتين).تردد وأعداد CD4 التي تعبر عن GZMB/CCL5الخلايا التائية في WT و LILRB4فئران تحمل الأورام، تمثل تجربتين). ج-ح، تردد CD4تعبيرات TILs g، IFN-TNF- أو CD40L في T3 TILs بعد التنشيط في المختبر باستخدام ببتيد mItgb1 في وجود Golgi-plugتمثل تجربتين).تكرار CD4تعبيرات TILsIFN-TNF- أو CD40L في خلايا TILs F244 بعد التنشيط في المختبر باستخدام ببتيد mPlec في وجود Golgi-plug ). البيانات في (أ-ج) معبّرة عن متوسط قطر الورم س.م.تم حساب القيم باستخدام تحليل التباين ثنائي الاتجاه مع تصحيح اختبار المقارنات المتعددة لسيداك. البيانات في (د-ي) معبّرة كمتوسط.س.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع مقارنات متعددة تم تصحيحها باستخدام طريقة توكي.
الشكل 7 من البيانات الموسعة | حجب LILRB4 يعزز النمط الظاهري والوظائف الفعالة لـتيلز. أ، أرقام من ملما4CD8 المحددتم تقييم TILs في T3 TILs ). تكرارالخلايا التائية المستحثة تعبر عن IFNTNF-بعد التنشيط في المختبر باستخدام mLama4 SLP. ج، تردد CD8تعبير TILs عن IFN-TNF-بعد التنشيط في المختبر باستخدام mPex14 SLP ). تعبير عدة علامات إجهاد على خلايا CD8 المحددة بـ mLama4خلايا T
e ورم ساركوما F244
في TILs من الفئران الحاملة لورم T3 ). تمثل البيانات تجربتين مستقلتين وتظهر كمتوسط s.e.m.e، تعبير عن عدة علامات إجهاد في CD8 المحدد بـ mPex14 خلايا T في TILs F244 ). البيانات المعبر عنها تظهر كمتوسط س.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع مقارنات متعددة تم تصحيحها باستخدام طريقة توكي، والتي تم استخدامها للإحصائيات.
الشكل البياني الممتد 8 | متعلق بالشكل 4: علاج IL-2 الموجه نحو CD8 والمستهدف بشكل متزامن يعيد تأسيس فعالية HDVax المضادة للورم في نماذج الأورام F244 و1956. أ، نمو ورم F244 في الفئران الطبيعية المعالجة كما في الشكل 4 ح.، تمثل تجربتين). ب، نمو الورم في فئران C57BL6 WT المعالجة كما في الشكل 5i(تمثل تجربتين). البيانات في و تُعبر كمتوسطس.م.تم حساب القيم باستخدام تحليل التباين ثنائي الاتجاه مصححًا باختبار المقارنات المتعددة لسيداك. ج، تمثيلي رسم بياني لتدفق الخلايا يظهر تعبير mLama4+محددتم حصاد TILs من الفئران المعالجة، كما هو موضح في الشكل 5h. د-و، تم معالجة الفئران الحاملة للورم F244 كما هو موضح في الشكل 5h، والتردد،أعداد CD8 المحددة بـ mPex14تيلز، ونسبة CD8 المحددة لـ mPex14تم تقييم TILs إلى خلايا شبيهة بـ Tr1 في اليوم 13 من TILs. تُعبر البيانات عن المتوسطس.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع تصحيحات المقارنات المتعددة باستخدام طريقة توكي.
الشكل البياني الممتد 9 | متعلق بالشكل 5: خلايا شبيهة بـ Tr1 المستحثة بواسطة HDVax تقتل cDC1 بطريقة تعتمد على GZMB. أ، إنتاج IL-2 بواسطة خلايا الهجينة المخصصة لـ mItgb1 بعد الحضانة مع cDC1 و cDC2 المفصولة من خمسة فئران بعد يوم واحد من العلاج بـ T3- LDVax أو -HDVax ( وتمثل تجربتين). ب، تم تطعيم الفئران الحاملة لورم T3 بـ LDVax أو HDVax (جرعتان، بفارق 11 يومًا)، وتم جمع المصل بعد 7 أيام من آخر تطعيم، وتم تقييم مستوى الأجسام المضادة لـ mItgb1 SLP باستخدام ELISA (الطرق). ج، تم تحديد أنواع مختلفة من الأجسام المضادة الخاصة بـ mItgb1. البيانات في و تظهر كوسائل س.م. ويمثل ثلاثة تجارب مستقلة ( ). د، تردد التعبير عن البيرفورين (Prf1) في CD4 تيلز في مجموعات فرعية مختلفة من CD4تيلز (، تمثل تجربتين). هـ، تردد تعبير NIR (إشارة حية/ميتة) في cDC1 (“) (يسار) و cDC2 () (يمين) بعد الحضانة مع LDVax المحفز أو تحت مجموعات من HDVax المحفز CD4 المحدد لـ mItgb1خلايا T (الطرق). تُعرض البيانات كمتوسطاتالانحراف المعياري للمتوسط لثلاث تجارب. ف، تردد cDC2 (CD172aتعبير NIR بعد الحضانة مع تحت مجموعات فرز من CD4 المحدد بواسطة HDVax mItgb1تم جمع خلايا T من الفئران WT (المعالجة بـ HDVax بالإضافة إلى الأجسام المضادة الأحادية التحكم أو LILRB4) أو GZMB الفئران (مرتبط بالشكل 5ب). ج، التردد (يسار) والأعداد (يمين) من cDC1 (من CD45الخلايا) في F244 TILs المعالجة بـ HDVax بعد خمسة أيام من التطعيمات ( ). البيانات تمثل تجربتين مستقلتين وتظهر كمتوسط s.e.m. h، التردد (يسار) والأعداد (يمين) من cDC1 في TILs 1956 التي تم تقييمها بعد خمسة أيام من التطعيم في WT و LILRB4فئران ( ). أنا، تردد cDC1 في أورام T3 تم تقييمه بعد خمسة أيام من التطعيم والعلاج بجرعتين من PD-1 أو CTLA-4 في الفئران البرية ( ، تمثل تجربتين).تكرار cDC1 في TILs لعام 1956 تم تقييمه بعد خمسة أيام من التطعيم في WT و GZMBفئران ( ). البيانات في جميع اللوحات معروضة كمتوسطات س.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع تصحيحات المقارنات المتعددة باستخدام طريقة توكي.
الشكل البياني الممتد 10 | متعلق بالشكل 5: خلايا شبيهة بـ Tr1 الناتجة عن HDVax تنشأ من تقديم مستضدات cDC2 المفضل. أ، مخططات تدفق تمثيلية تظهر استراتيجية التصفية لمجموعات مختلفة من خلايا تقديم المستضد في WT وفئران تحمل أورام 1956. ب، تكرار خلايا شبيهة بـ Tr1 في طحال الفئران المعالجة بـ HDVax. (نقص cDC2) أو Irf8 فئران ناقصة cDC1تمثل تجربتين). البيانات هي
مُظهر كوسيلةس.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع تصحيحات المقارنات المتعددة بطريقة توكي. ج، تكرار خلايا Tregs و د، خلايا شبيهة بـ Tr1 في 1956 TIL التي تم جمعها في عدة أيام بعد حقن الورم.، تمثل تجربتين). يتم التعبير عن البيانات كمتوسطاتس.م.تم حساب القيم باستخدام تحليل التباين ثنائي الاتجاه مصححًا باختبار المقارنات المتعددة لسيداك.
محفظة الطبيعة
المؤلف (المؤلفون) المراسلون: روبرت شرايبر آخر تحديث بواسطة المؤلفين: 22 مايو 2024
ملخص التقرير
تسعى Nature Portfolio إلى تحسين إمكانية تكرار العمل الذي ننشره. يوفر هذا النموذج هيكلًا للاتساق والشفافية في التقرير. لمزيد من المعلومات حول سياسات Nature Portfolio، يرجى الاطلاع على سياسات التحرير وقائمة مراجعة سياسة التحرير.
الإحصائيات
لجميع التحليلات الإحصائية، تأكد من أن العناصر التالية موجودة في أسطورة الشكل، أسطورة الجدول، النص الرئيسي، أو قسم الطرق.
مؤكد X حجم العينة بالضبط ( لكل مجموعة/شرط تجريبي، معطاة كرقم منفصل ووحدة قياس بيان حول ما إذا كانت القياسات قد أُخذت من عينات متميزة أو ما إذا كانت نفس العينة قد تم قياسها عدة مرات اختبار(ات) الإحصاء المستخدمة وما إذا كانت أحادية الجانب أو ثنائية الجانب يجب أن تُوصف الاختبارات الشائعة فقط بالاسم؛ واصفًا التقنيات الأكثر تعقيدًا في قسم الطرق. وصف لجميع المتغيرات المشتركة التي تم اختبارها وصف لأي افتراضات أو تصحيحات، مثل اختبارات الطبيعية والتعديل للمقارنات المتعددة وصف كامل للمعلمات الإحصائية بما في ذلك الاتجاه المركزي (مثل المتوسطات) أو تقديرات أساسية أخرى (مثل معامل الانحدار) والتباين (مثل الانحراف المعياري) أو تقديرات عدم اليقين المرتبطة (مثل فترات الثقة) إكس لاختبار الفرضية الصفرية، إحصائية الاختبار (على سبيل المثال، ) مع فترات الثقة، أحجام التأثير، درجات الحرية وقيمة ملحوظة أعطِالقيم كقيم دقيقة كلما كان ذلك مناسبًا.
إكسلتحليل بايزي، معلومات حول اختيار القيم الأولية وإعدادات سلسلة ماركوف مونت كارلو Xللتصاميم الهرمية والمعقدة، تحديد المستوى المناسب للاختبارات والتقارير الكاملة عن النتائج X تقديرات أحجام التأثير (مثل حجم تأثير كوهين)بيرسون )، مما يشير إلى كيفية حسابها تحتوي مجموعتنا على الإنترنت حول الإحصائيات لعلماء الأحياء على مقالات تتناول العديد من النقاط المذكورة أعلاه.
البرمجيات والشيفرة
معلومات السياسة حول توفر كود الكمبيوتر جمع البيانات تم جمع بيانات تحليل تدفق الخلايا باستخدام برنامج BD FACSDIVA الإصدار 9.1. تم إجراء اختبار MultiPlex باستخدام Flex MAP 3D. تم جمع ELISPOT باستخدام جهاز وبرنامج CTL Immunospot S6 العالمي.
تحليل البيانات تم إجراء جميع التحليلات الإحصائية باستخدام برنامج GraphPad Prism الإصدار 10.2.2. تم استخدام cell ranger v6.1.1 لتحليل تسلسل RNA أحادي الخلية.
بالنسبة للمخطوطات التي تستخدم خوارزميات أو برامج مخصصة تكون مركزية في البحث ولكن لم يتم وصفها بعد في الأدبيات المنشورة، يجب أن تكون البرمجيات متاحة للمحررين والمراجعين. نحن نشجع بشدة على إيداع الشيفرة في مستودع مجتمعي (مثل GitHub). راجع إرشادات مجموعة Nature لتقديم الشيفرة والبرمجيات لمزيد من المعلومات.
بيانات
معلومات السياسة حول توفر البيانات يجب أن تتضمن جميع المخطوطات بيانًا حول توفر البيانات. يجب أن يوفر هذا البيان المعلومات التالية، حيثما ينطبق:
رموز الانضمام، معرفات فريدة، أو روابط ويب لمجموعات البيانات المتاحة للجمهور
وصف لأي قيود على توفر البيانات
بالنسبة لمجموعات البيانات السريرية أو بيانات الطرف الثالث، يرجى التأكد من أن البيان يتماشى مع سياستنا
جميع البيانات التي تدعم نتائج هذه الدراسة متاحة ضمن الورقة، أو ملفات البيانات التكميلية أو الموسعة، أو ملفات المصدر. أي معلومات إضافية تتعلق بالدراسة متاحة من المؤلف المراسل عند الطلب. جميع المواد متاحة من خلال اتفاقية نقل المواد.
البحث الذي يتضمن مشاركين بشريين، بياناتهم، أو مواد بيولوجية
معلومات السياسة حول الدراسات التي تشمل مشاركين بشريين أو بيانات بشرية. انظر أيضًا معلومات السياسة حول الجنس، الهوية/التقديم الجنسي، والتوجه الجنسي والعرق، والاثنية والعنصرية.
التقارير عن الجنس والنوع الاجتماعي غير متوفر التقارير عن العرق أو الإثنية أو غير متوفر اجتماعي ذو صلة تجمعات خصائص السكان غير متوفر التوظيف
غير متوفر
رقابة الأخلاقيات غير متوفر
يرجى ملاحظة أنه يجب أيضًا تقديم معلومات كاملة حول الموافقة على بروتوكول الدراسة في المخطوطة.
التقارير المتخصصة في المجال
يرجى اختيار الخيار أدناه الذي يناسب بحثك بشكل أفضل. إذا لم تكن متأكدًا، اقرأ الأقسام المناسبة قبل اتخاذ قرارك. علوم الحياة العلوم السلوكية والاجتماعية العلوم البيئية والتطورية والبيئية لنسخة مرجعية من الوثيقة مع جميع الأقسام، انظرnature.com/documents/nr-reporting-summary-flat.pdf
تصميم دراسة علوم الحياة
يجب على جميع الدراسات الإفصاح عن هذه النقاط حتى عندما يكون الإفصاح سلبياً.
حجم العينة
تم تحديد أحجام العينات بناءً على 25 عامًا من الخبرة مع هذه الأنواع من الأورام بشكل عام، وأكثر من 10 سنوات مع خطوط خلايا T3 و F244. (غوبين وآخرون 2014، ألباش 2019)
استثناءات البيانات
لم يتم استبعاد أي بيانات
التكرار
جميع التجارب كانت تحتوي على نسخ بيولوجية و/أو تقنية متعددة، كما هو موضح في أساطير الأشكال.
العشوائية
في الدراسات الحيوانية، تم توزيع الفئران عشوائيًا من دفعات كبيرة تم الحصول عليها من البائع إلى مجموعات تجريبية مختلفة بتوزيع متناسب مع العمر.
عمى
تمت حقن الورم بواسطة شخص واحد. تم إجراء العلاجات وقياس الورم بواسطة أشخاص مستقلين. لم يتم إجراء أي إخفاء في الدراسات المخبرية.
التقارير عن مواد وأنظمة وطرق محددة
نحتاج إلى معلومات من المؤلفين حول بعض أنواع المواد والأنظمة التجريبية والأساليب المستخدمة في العديد من الدراسات. هنا، يرجى الإشارة إلى ما إذا كانت كل مادة أو نظام أو طريقة مدرجة ذات صلة بدراستك. إذا لم تكن متأكدًا مما إذا كان عنصر القائمة ينطبق على بحثك، يرجى قراءة القسم المناسب قبل اختيار رد.
المواد والأنظمة التجريبية
طرق
غير متوفر
مشارك في الدراسة
غير متوفر
مشارك في الدراسة
خطوط خلايا حقيقية النواة
البحث ذو الاستخدام المزدوج الذي يثير القلق
الأجسام المضادة
الأجسام المضادة المستخدمة
تم شراء الأجسام المضادة للتدفق من: [BioLegend؛ IFN (XMG1.2؛ 1:100؛ رقم الكات 505808)، TNF (XMG1.2؛ 1:100؛ رقم الكات 505826)، CD200 (OX-90؛ 1:200؛ 123820)، PD-1 (29F.1A12؛ 1:200؛ رقم الكات 135228)، CCL5 (2E9/CCL5؛ 1:500؛ رقم الكات 149106)، GZMB (QA16A02؛ 1:50؛ رقم الكات 372216)، TIM-3 (RMT3-23؛ 1:200؛ رقم الكات 119723)، CD25 (PC61؛ 1:100؛ رقم الكات 102036)، CD154 (MR1؛ 1:100؛ رقم الكات 106506)، CD152 (UC10-4B9؛ 1:100؛ رقم الكات 106318)، CD4 (RM4-5: 1:500)، IL-2 (Jes6-5H4: 1:50؛ رقم الكات 503826)، LILRB4 (H1.1: 1:200؛ رقم الكات 144904)، CD11b (M1/70: 1:800؛ رقم الكات 101226)، XCR1 (ZET: 1:100؛ رقم الكات 148206)، MHC-II (M5/114.15.2: 1:1000؛ رقم الكات 107641)، CD11c (N418: 1:200؛ رقم الكات 117336)، CD172a (P84: 1:500؛ رقم الكات 144008)، (صبغة حيوية قابلة للإصلاح Zombie NIR؛ 1:500؛ رقم الكات 423106)، CD86 (GL-1؛ 1:200؛ رقم الكات 105042)، CD80 (16-10A1؛ 1:200؛ رقم الكات 104712)، CD40 (FGK45؛ 1:100؛ رقم الكات 157506)؛
eBioSciences: FOXP3 (Fjk-16a; 1:50; cat# 11-5773-82) و SEMA4a (5E3; 1:50; cat# 46-9753-41)]. تم استخدام مجموعة صبغ Foxp3/عامل النسخ (ThermoFischer; cat# 00-5523-00) لصبغ Foxp3 وبروتينات خلوية أخرى وفقًا لبروتوكول الشركة المصنعة.
التحقق
تم التحقق من جميع الأجسام المضادة الأولية ضد البروتينات المحددة من قبل الشركة المصنعة وفقًا لورقات البيانات المرتبطة بها وهي مدرجة على مواقعها الإلكترونية.
خطوط خلايا حقيقية النواة
معلومات السياسة حول خطوط الخلايا والجنس والنوع في البحث
مصدر(s) خط الخلية
تم إنتاج خطوط الخلايا T3 و F244 و 1956 من خلال الحقن تحت الجلد لـ MCA. تم الحصول على خط خلايا HEK293 من ATCC (CRL-1573).
المصادقة
تم التحقق من صحة خطوط الخلايا من خلال تسلسل الإكسوزومات الكاملة والتعبير عن بعض الجينات
تلوث الميكوبلازما
جميع خطوط الخلايا يتم اختبارها بانتظام كل شهر وكانت سلبية لعدوى الميكوبلازما.
الخطوط التي يتم التعرف عليها بشكل خاطئ بشكل شائع (انظر سجل ICLAC)
لم تتضمن هذه الدراسة أي خطوط خلوية يتم التعرف عليها بشكل خاطئ بشكل شائع.
الحيوانات وغيرها من الكائنات البحثية
معلومات السياسة حول الدراسات التي تشمل الحيوانات؛ إرشادات ARRIVE الموصى بها للإبلاغ عن أبحاث الحيوانات، والجنس والنوع في البحث
الحيوانات المخبرية
تم استخدام إناث من فئران C57BL/6J بعمر 6-8 أسابيع (رقم الكاتالوج 000664، مختبر جاكسون)، وفئران GZMB KO، وفئران LILRB4 KO، وفئران 129S6 الذكور المعوزة لـ Rag2 والنمط البري في التجارب. تم تربية الفئران في مجموعات من 3 إلى 5.على: 12 ساعة تشغيل/إيقاف دورة الظلام/الضوء. درجة حرارة غرفة الحيوانات المحيطة هيمتحكم فيه ضمنورطوبة الغرفة هيمتحكم فيه ضمن.
الحيوانات البرية
لم يتم استخدام حيوانات برية في هذه الدراسة.
التقارير عن الجنس
تم استخدام ذكور الفئران (129S6) لأورام T3 وF244، بينما تم استخدام إناث الفئران (C57BL6) لخط أورام 1956.
عينات تم جمعها من الميدان
لم يتم استخدام عينات تم جمعها من الميدان في هذه الدراسة.
رقابة الأخلاقيات
تمت جميع الدراسات الحيوانية بموافقة لجنة الدراسات الحيوانية المعتمدة من جمعية اعتماد رعاية الحيوانات المخبرية في جامعة واشنطن في سانت لويس.
يرجى ملاحظة أنه يجب أيضًا تقديم معلومات كاملة حول الموافقة على بروتوكول الدراسة في المخطوطة.
نباتات
مخزونات البذور
غير متوفر
أنماط جينية نباتية جديدة
غير متوفر
المصادقة
غير متوفر
تدفق الخلايا
المؤامرات
أكد أن:
توضح تسميات المحاور العلامة والفلوركروم المستخدم (مثل CD4-FITC). المقاييس على المحاور واضحة تمامًا. قم بتضمين الأرقام على المحاور فقط للرسم البياني في أسفل اليسار من المجموعة (المجموعة هي تحليل للعلامات المتطابقة). جميع الرسوم البيانية هي رسوم بيانية متساوية الارتفاع مع نقاط شاذة أو رسوم بيانية بالألوان الزائفة. تم توفير قيمة عددية لعدد الخلايا أو النسبة المئوية (مع الإحصائيات).
المنهجية
تحضير العينة
تم جمع الأورام، وتفكيكها يدويًا إلى قطع صغيرة وهضمها باستخدام الكولاجيناز 1A لمدة 45 دقيقة عند 37 درجة مئوية. ثم تم تصفية تعليق الخلايا من خلال مصفاة خلايا بفتحة 70 ميكرومتر. بعد ذلك، تم تحلل كريات الدم الحمراء. تم تصفية التعليق مرة أخرى من خلال مصفاة خلايا بفتحة 40 ميكرومتر وإعادة تعليقها في وسط RPMI كامل. بالنسبة لصبغات الخلايا داخل الخلوية، تم تثبيت الخلايا وتمريرها باستخدام مجموعة صبغ BD Foxp3.
آلة
برمجيات
وفرة تجمع الخلايا استراتيجية البوابة
تم استخدام BD FACS Symphony 3 و LSRFortessaX20 لجمع البيانات. تم استخدام ARIA-II و/أو Bigfoot للفرز.
تم استخدام برنامج BD FACSDIVA الإصدار 9.1 لجمع بيانات تحليل الخلايا باستخدام التدفق وللفرز، وتم تحليل البيانات باستخدام برنامج FlowJo الإصدار 10.10.
تم إظهار وفرة الخلايا من خلال استراتيجيات التصفية
تم تحديد الخلايا اللمفاوية الحية باستخدام CD90.2 و CD4. تم تحديد الخلايا المحددة بالمستضد باستخدام التترامر المناسب، وتم استبعاد Tregs باستخدام صبغة CD25 و Foxp3. تم قياس تعبيرات CD39 و CD200 و CD153 و LILRB4 و GZMB أو CCL5.
قم بتحديد هذا المربع لتأكيد أنه تم تقديم رقم يوضح استراتيجية البوابة في المعلومات التكميلية.
قسم علم الأمراض والمناعة، جامعة واشنطن كلية الطب، سانت لويس، ميزوري، الولايات المتحدة الأمريكية.مركز أندرو م. وجين م. بورسكي لعلم المناعة البشرية وبرامج العلاج المناعي، كلية الطب بجامعة واشنطن، سانت لويس، ميزوري، الولايات المتحدة الأمريكية.قسم الطب، قسم الأورام، جامعة واشنطن كلية الطب، سانت لويس، ميزوري، الولايات المتحدة الأمريكية.قسم المناعة، مركز أندرسون للسرطان بجامعة تكساس، هيوستن، تكساس، الولايات المتحدة الأمريكية.مركز أبحاث الطب الوطني ألمازوف، سانت بطرسبرغ، روسيا.أونكوفير إنك، واشنطن، دي سي، الولايات المتحدة الأمريكية.ناتيرا إنك، أوستن، تكساس، الولايات المتحدة الأمريكية.أشر بيوثيرابيوتيكس، جنوب سان فرانسيسكو، كاليفورنيا، الولايات المتحدة الأمريكية.معهد السرطان الهولندي، معهد أونكود، أمستردام، جامعة ليدن، ليدن، هولندا.معهد باركر لعلاج السرطان المناعي، سان فرانسيسكو، كاليفورنيا، الولايات المتحدة الأمريكية.البريد الإلكتروني: rdschreiber@wustl.edu
عواد، م. ونورث، ر. ج. التراجع المناعي لليمفوما الفأرية بعد العلاج بالأجسام المضادة المضادة لـ L3T4. نتيجة لإزالة L3T4خلايا T المثبطة من مضيف ينتج بشكل أساسي مناعة تعتمد على خلايا T Lyt-2+. J. Exp. Med. 168، 2193-2206 (1988).
هوري، س.، نومورا، ت. وساكاغوتشي، س. التحكم في تطوير خلايا T التنظيمية بواسطة عامل النسخ Foxp3. ساينس 299، 1057-1061 (2003).
فونتينو، ج. د.، غافين، م. أ. ورودنسكي، أ. ي. يبرمج Foxp3 تطوير ووظيفةالخلايا التائية التنظيمية. نات. مناعه. 4، 330-336 (2003).
Hussein Sultan , Yoshiko Takeuchi , Jeffrey P. Ward , Naveen Sharma , Tian-Tian Liu , Vladimir Sukhov , Maria Firulyova , Yuang Song , Samuel Ameh , Simone Brioschi , Darya Khantakova’, Cora D. Arthur , J. Michael White , Heather Kohlmiller , Andres M. Salazar , Robert Burns , Helio A. Costa , Kelly D. Moynihan , Yik Andy Yeung , Ivana Djuretic , Ton N. Schumacher , Kathleen C. F. Sheehan , Marco Colonna , James P. Allison , Kenneth M. Murphy , Maxim N. Artyomov & Robert D. Schreiber
Abstract
cells can either enhance or inhibit tumour immunity. Although regulatory T cells have long been known to impede antitumour responses , other cells have recently been implicated in inhibiting this response . Yet, the nature and function of the latter remain unclear. Here, using vaccines containing MHC class I (MHC-I) neoantigens (neoAgs) and different doses of tumour-derived MHC-II neoAgs, we discovered that whereas the inclusion of vaccines with low doses of MHC-II-restricted peptides (LDVax) promoted tumour rejection, vaccines containing high doses of the same MHC-II neoAgs (HDVax) inhibited rejection. Characterization of the inhibitory cells induced by HDVax identified them as type 1 regulatory T (Tr1) cells expressing IL-10, granzyme B, perforin, CCL5 and LILRB4. Tumour-specific Tr1 cells suppressed tumour rejection induced by anti-PD1, LDVax or adoptively transferred tumour-specific effector T cells. Mechanistically, HDVax-induced Tr1 cells selectively killed MHC-II tumour antigen-presenting type 1 conventional dendritic cells (cDC1s), leading to low numbers of cDC1s in tumours. We then documented modalities to overcome this inhibition, specifically via anti-LILRB4 blockade, using a CD8-directed IL-2 mutein, or targeted loss of cDC2/monocytes. Collectively, these data show that cytotoxic Tr1 cells, which maintain peripheral tolerance, also inhibit antitumour responses and thereby function to impede immune control of cancer.
A desirable outcome of cancer immunotherapy is the generation of tumour-specific cytolytic T cells (CTLs) capable of destroying tumours. Whereas induction of these effector cells is often promoted by ‘helper’ cells , this process can be inhibited by regulatory cells . Recent studies have suggested that conventional cells may not be the only immunosuppressive T cell population residing in progressively growing tumours , but establishing the origins, nature and function of the latter has been challenging. This has placed theoretical and practical constraints on cancer immunotherapy in general and cancer-specific neoAg vaccines in particular. The ability of CD4 Tr1 cells to maintain tolerance has been well established in autoimmunity and chronic infections and has been suggested to contribute to cancer immunosuppression . However, Tr1 cells have not been defined based on a master transcription factor but rather are identified as FOXP3-negative, IL-10-producing inhibitory cells . Here we use our well-characterized, syngeneic mouse sarcoma models to show that, whereas synthetic long peptide (SLP) neoAg vaccines containing tumour-specific MHC-I neoAgs and low inputs of tumour-specific MHC-II neoAgs efficiently promote tumour rejection, similar vaccines
expressing high quantities of the same MHC-II neoAgs unexpectedly induce immunosuppressive cells distinct from cells that inhibit tumour elimination. We characterize these cells as cytolytic Tr1 cells, provide insights into their development and mechanisms of action, and validate strategies to circumvent their inhibitory activity. We further show that a similar CD4 T cell population can be found in progressively growing tumours in mice as they become insensitive to anti-PD1 therapy and in human patients with cancer who respond poorly to immune checkpoint therapy (ICT) and cancer vaccines. We demonstrate that eliminating these cells or inhibiting their function renders cancer immunotherapy more effective. This study thus not only documents a new function for a known CD4 T cell subpopulation but also provides a strategy to circumvent their inhibitory effects on tumour rejection.
Complex MHC-II dosing profiles by peptide vaccines
We previously identified the unique major MHC-I and MHC-II neoAgs expressed in three of our methylcholanthrene (MCA) sarcoma lines
Fig. 1 | High MHC-II neoAg doses inhibit the antitumour efficacy of SLP vaccines and certain ICTs. a, Percent survival of T3 tumour-bearing mice after treatments with vaccines containing fixed ( 1.5 ng ) doses of mItgb1 SLP plus different doses of mLama4/mAlg8 SLP or IrrVax.b, Percent survival of T3 tumour-bearing mice after treatments with vaccines containing fixed doses of each plus different doses of mItgb1SLP.c, Percent survival of F244 tumour-bearing mice after treatment with vaccines containing fixed doses of mPex14 SLP ( per mouse) plus different doses of mPlec SLP. d, Percent survival of 1956 tumour-bearing mice after treatment with vaccines containing fixed doses of per mouse) plus different doses of mCs SLP or IrrVax. The numbers above each bar represent the number of mice that rejected the tumours over the total number of mice used.
(Supplementary Table1), showed that immune rejection of each tumour depends on and cell responses to the neoAgs uniquely expressed in each tumour line, and further showed that optimal rejection efficacy required the presence of both MHC-I and MHC-II neoAgs in tumours and vaccines . We were intrigued by the profound effects that cells had on cell-dependent protective immunity against MCA sarcomas that do not express MHC-II proteins, and therefore decided to study the interdependency between MHC-I and MHC-II neoAg responses in promoting antitumour efficacy. We approached this question by defining the MHC-I and MHC-II neoAg doses either alone or together that induced optimal antitumour efficacy. T3-specific SLP vaccines comprising only T3-specific MHC-I (G1254V mLama4 plus A506T mAlg8) or MHC-II (N710Y mItgb1) SLP plus poly-ICLC adjuvant or irrelevant vaccines containing human papillomavirus (HPV)-SLP (IrrVax) were ineffective in inducing T3 rejection when administered 6 days post-tumour cell inoculation (Fig. 1a,b). However, T3-specific vaccines containing MHC-I neoAgs at doses between 150 ng and together with constant low doses of mItgb1 SLP ( 1.5 ng ) induced tumour rejection in nearly all tumour-bearing mice (Fig. 1a,b). By contrast, when doses of T3-specific MHC-I neoAg SLPs were held constant at 150 ng and the dose of T3-specific MHC-II neoAg SLPs was varied, a bell-shaped dose-response curve was observed displaying maximum protection in 42 of 50 vaccinated mice at doses of 1.5 ng of -restricted mltgb1 SLP (hereafter referred to as LDVax; Fig. 1b). Vaccines containing the same dose of T3-specific MHC-I SLP plus higher doses of mItgb1 SLP,
for example, 150 ng , induced little or no antitumour efficacy (hereafter referred to as HDVax). Thus, high doses of tumour-specific MHC-IISLP render T3-specific vaccines ineffective.
Generalizability of HDVax ineffectiveness
To address whether the lack of efficacy of T3-specific HDVax was generalizable to other antigenically distinct tumours, we used vaccines containing the major MHC-I-restricted and MHC-II-restricted neoAgs for two other MCA sarcoma models: F244 (expressing an -restricted G122A mutation in peroxisomal membrane protein 14 (mPex14) and a D2944YI-A -restricted mutation in plectin-1 (mPlec)) and 1956 (expressing a -restricted M 50 I mutation in proteasome 26S subunit, non-ATPase 6 (mPsmd6) and an I-A -restricted R144S mutation in citrate synthase (mCs)) (Supplementary Table 1). As seen with T3-specific HDVax and T3 tumours, the same type of bell-shape curve for MHC-II neoAgs was observed with F244 and 1956 tumours and their specific HDVax (Fig. 1c,d).
HDVax inhibits antitumour efficacy of ICT
To discern whether HDVax actively inhibited immunotherapy, we assessed the effect of T3-HDVax on the efficacy of anti-PD1, anti-41BB or anti-CTLA4 as each of these antibodies, when administered as monotherapy, induced complete in vivo rejection of T3 tumours
Fig. 2 | HDVax inhibition is specific for the MHC-II neoAgs in the tumour. a, T3 and F244 tumour outgrowth in opposite contralateral flanks of male 129S6 WT mice vaccinated and treated with anti-PD1 ( , representative of two experiments). D0, day tumour outgrowth in mice receiving total T cells ( ) from LDVax-treated mice only or plus CD25-negative mltgb1-specific CD4 T cells sorted from HDVax-treated mice . Data in panels are shown as mean tumour diameter s.e.m., and statistics were done using two-way ANOVA, with multiple comparisons corrected using Sidak’s multiple comparison test.c-e, CD4 T cells were purified and stimulated
as shown. IL-2 production in the supernatant was assessed using MultiPlex ( , pooled from two experiments; d). The heatmap displays the production levels of different cytokines (indicated on the axis; e). The numbers in panel represent the mean of two independent biological replicates (representative of two experiments). Frequency ( ) and numbers ( ) of -specific CD8 TILs in T3 tumours harvested on day 13 ( , representative of three experiments) are also shown. Data in panels d,f,g are expressed as mean s.e.m.; statistics were done using one-way ANOVA, with multiple comparisons corrected with Tukey’s method.
(Fig. 1e). Combining ICT and LDVax did not alter the antitumour efficacy of the checkpoint antibodies. By contrast, T3-HDVax abrogated tumour rejection in all mice treated with anti-PD1 or anti-4-1BB, but did not ablate anti-CTLA4-induced tumour rejection. Similar results were obtained using the F244 and 1956 tumour models and their respective neoAg vaccines (Fig. 1f,g), demonstrating that HDVax treatment inhibits certain types of ICT. The latter observation was explained by subsequent experiments.
Antigen specificity of HDVax inhibition
To test the antigen specificity of HDVax inhibition, T3 sarcoma cells and antigenically distinct sex-matched syngeneic F244 sarcoma cells were injected into contralateral flanks of naive 129S6 wild-type (WT) mice. Three days later, mice were treated with anti-PD1 plus either irrelevant HPV-LDVax or HPV-HDVax, T3-LDVax or T3-HDVax, or F244-LDVax or
F244-HDVax, and tumour growth was monitored. Neither LDVax nor irrelevant HPV-HDVax treatment inhibited anti-PD1-induced rejection of T3 or F244 tumours (Fig. 2a, upper three panels and lower left panel). However, T3-HDVax treatment selectively inhibited anti-PD1-dependent rejection of T3 but not F244 tumours (Fig. 2a, bottom middle panel). Conversely, F244-HDVax treatment selectively inhibited anti-PD1-dependent rejection of F244 tumours but not T3 tumours (Fig. 2a, bottom right panel). Thus, the inhibitory action of HDVax is specifically linked to the MHC-II neoAg expressed by each tumour line.
HDVax-induced T cells inhibit tumour rejection
To formally test whether inhibition was due to induction of immunosuppressive cells, we established an adoptive T cell transfer (ACT) system in Rag2 mice (see Methods). ACT of cell-depleted T cells from LDVax-treated WT mice into T3 tumour-bearing, immunodeficient Rag2-/- mice was sufficient to induce T3 rejection (Fig. 2b). However, mixing HDVax-induced, cell-depleted mItgb1-specific CD4 T cells with LDVax-induced effector cells abrogated T3 tumour rejection. These data suggested that HDVax induces a cell population distinct from classical cells, which possesses potent immunosuppressive activity.
Characteristics of HDVax-induced CD4 cells
The frequency and absolute numbers of mItgb1-specific CD4 tumourinfiltrating lymphocytes (TILs) from HDVax-treated or LDVax-treated mice were similar (Extended Data Fig. 1a,b). Flow cytometry analyses revealed that the frequency and absolute numbers of cells in mltgb1-specific CD4 TILs or spleens decreased comparably following either HDVax or LDVax treatment compared with PBS-treated controls (Extended Data Fig. 1c,d). Similar findings were observed with F244 tumour-bearing mice treated with F244-HDVax or F244-LDVax (Extended Data Fig. 1e). Cytokine multiplex analyses revealed distinct phenotypes of CD4 TILs induced by LDVax versus HDVax. Whereas T3-LDVax-induced mItgb1-specific CD4 T cells produced IL-2 when stimulated with mItgb1-pulsed splenocytes, T3-HDVax-induced mItgb1-specific CD4 T cells failed to produce IL-2 (Fig. 2c,d). In addition, whereas T3-LDVax-induced CD4 T cells produced high levels of IFN , TNF and GM-CSF (Fig. 2e), T3-HDVax-induced CD4 T cells produced high levels of IL-10, TGF , IL-5 and IL-27. These results were recapitulated by intracellular staining analyses (Extended Data Fig.1f,g). Moreover, whereas mItgb1-specific CD4 TILs harvested from T3-LDVax-treated mice expressed increased levels of CD40L following antigen stimulation (LDVax: ; PBS: ), only a small fraction of antigen-stimulated T3-HDVax-induced mItgb1-specific CD4 TILs expressed CD40L (HDVax: ; Extended Data Fig. 1h,i).
Consistent with T3-LDVax-dependent elevated expression of CD40L on CD4 TILs, dendritic cells from the same mice displayed slightly higher but significantly enhanced expression of CD86 and CD70 than comparable cells from T3-HDVax-treated mice. No significant differences were observed in expression levels of CD40 on dendritic cells (Extended Data Fig. 2a,b). Of note, the number of cDC1s was reduced by 2.9 -fold in tumours from T3-bearing mice treated with T3-HDVax compared with PBS (Extended Data Fig. 2c). Only a minor reduction in the number of cDC2s was observed, and this was not statistically significant.
In agreement with the aberrant cytokine response and low recovery of cDC1s from T3-HDVax-treated mice, the frequencies and absolute numbers of mLama4-specific CD8 TILs were significantly lower than CD8 TILs harvested from T3-LDVax-treated mice (Fig. 2f,g and Extended Data Fig. 2d). Furthermore, mLama4-specific CD8 TILs from T3-HDVax-treated mice expressed high levels of exhaustion markers (PD1, CD39, TIM3, LAG3 and TOX) and produced low amounts of IFN and TNF compared with similar populations from T3-LDVax-treated tumour-bearing mice (Extended Data Fig. 2d-h). No differences were observed in the TIM3 TCF1 subpopulations (Extended Data Fig. 2i). These results revealed that whereas T3-LDVax generates tumour-specific CD4 T cells whose products promote help to antitumour responses, T3-HDVax induces tumour-specific CD4 T cells that dampen antitumour activity.
Identification of inhibitory CD4 TILs by scRNA-seq
To better define the HDVax-induced inhibitory CD4 T cells, we performed single-cell RNA sequencing (scRNA-seq) on mItgb1-specific CD4 TILs from mice treated with T3-HDVax, T3-LDVax or PBS. Unbiased uniform manifold approximation and projection (UMAP) clustering of mItgb1-specific CD4 TILs revealed 11 cellular clusters (cluster 0 to cluster C10) with distinct gene signatures (Fig. 3a,b). We found that
vaccination per se resulted in a threefold reduction in the frequencies of TILs populating cluster 0 (follicular helper cell cluster ( Bcl6 and Cxcr5 )) and cluster 5 ( cell cluster (Foxp3 and Cd25 )) in tumours from mice receiving either T3-HDVax or T3-LDVax compared with PBS. By contrast, the proportions of cells populating clusters 1-4, 6 and 8 were differentially affected by the MHC-II peptide dose used in the vaccine. Clusters 2 and 8 were more densely populated following T3-LDVax than following T3-HDVax or PBS treatment, whereas clusters 3, 4 and 6 were more densely populated following T3-HDVax treatment than following LDVax or PBS treatment. Whereas T3-HDVax induced only minor density increases in clusters 4 and 6 compared with T3-LDVax or PBS treatment, the cell density of cluster increased to the greatest extent and showed the largest differences between HDVax versus LDVax or PBS.
Additional analyses revealed that T3-HDVax-induced CD4 TILs expressed relatively high levels of and sir transcripts and relatively low levels of Lag3, Cd2OO, Havcr2 and Tigit transcripts compared with those induced by T3-LDVax or PBS (Extended Data Fig. 3a). CD4 TILs from T3-HDVax-treated mice also expressed lower mRNA levels for Ifng, Tnf, Cd28 and Cd4Ol and transcription factors associated with T helper cell polarization, for example, Bhlhe4O, than TILs from T3-LDVax-treated or PBS-treated mice (Extended Data Fig. 3b). As shown by Il2 gene set enrichment analysis (GSEA), HDVa-induced cells exhibited defects in the IL-2 signature (Extended Data Fig.3c), expressed low levels of transcription factors such as Stat5b, Rora and Nfatc, and showed elevated expression of the cMafgene that suppresses IL-2 production compared with corresponding cells following T3-LDVax treatment (Extended Data Fig. 3d).
To validate these observations, we assessed the levels of the corresponding proteins in tumour-specific CD4 TILs and splenocyte populations from mice treated with T3-HDVax, T3-LDVax or PBS. These analyses confirmed that mItgb1-specific CD4 TILs were present at fivefold higher frequencies and numbers in T3-HDVax-treated mice than in TILs from mice treated with T3-LDVax or PBS (Extended Data Fig. 3e,f). A 2-3-fold difference was also noted in splenocytes from mice receiving T3-HDVax compared with T3-LDVax or PBS. Similar results were observed using F244 tumour-bearing mice treated with F244-HDVax, F244-LDVax or PBS (Extended Data Fig. 3g,h). In both T3 and F244 tumour models, the respective HDVax-induced cluster 3 cells expressed higher levels of CTLA4 protein than cells in all other clusters, except cluster 5 (classical CD25 cells; Extended Data Fig. 3i). Treatment with anti-CTLA4 but not anti-PD1 eliminated cluster 3 and cluster 5 cells in TILs of HDVax-treated mice (Extended Data Fig. 3j). This finding explains why HDVax did not inhibit antitumour efficacy of anti-CTLA4, because the anti-CTLA4 monoclonal antibody (9D9) used in the study depletes both cells and the HDVax-induced inhibitory CD4 T cells (Fig. 1e,f).
Cluster 3 cell ACT inhibits tumour rejection
Although these results suggested that the inhibitory HDVax-induced cells of interest were localized to cluster 3, they did not identify surface proteins on these cells that would permit their isolation. We therefore established a sorting strategy in which we removed T cells from the tumour-specific CD4 T cell population that expressed genes of surface markers not found in cluster 3 cells (that is, Cd25, Cd2OO and Cd153), and positively sorted the remaining cells for CD39 expression (Extended Data Fig. 4a). This approach allowed us to generate three cellular pools: (1) partially purified cluster 3 cells (CD25 CD200 and CD39 ), (2) CD25 cluster cells (more than FOXP ), and (3) non-cluster cells (CD25 CD200 CD153 and ; Extended Data Fig. 4b,c). These cellular pools were then tested for their capacity to inhibit T3 rejection. T3 tumours were rejected in Rag2-/ mice receiving cell-depleted T3-LDVax-induced effector T cells either alone or in combination with HDVax-induced, T3-specific non-cluster
Fig. 3 | HDVax-induced cells are cytotoxic Tr1 cells that are necessary and sufficient for HDVax-mediated inhibition. a, UMAP shows individual clusters in I- -mItgb1-specific CD4 TILs under different conditions. b, Frequency of cells populating individual clusters. c, T3 tumour outgrowth in mice that received T cells from LDVax mice plus total CD25 CD4 cells (approximately ) or different subpopulations (approximately ) sorted from HDVax-treated mice ( ). d, Volcano plot of bulk RNA-seq comparing gene expression in cluster 3 cells relative to cells (left) and non-cluster cells (right). values were obtained after applying the DESeq 2 tool.e Representative flow cytometry plot showing LILRB4 and SEMA4a expression in HDVax-induced mItgb1-specific CD4 TILs in T3 tumours.f, T3 tumour outgrowth in Rag2-/- mice receiving T cells from LDVax mice and different subpopulations (approximately ) sorted from HDVax-treated mice based on LILRB4 and SEMA4a expression ( (PBS and LDVax) and for the other groups). , Frequency of cells (g) or LILRB4-expressing CD4 TILs (h) in T3 tumours harvested at multiple days post-tumour inoculation ( ). ND, not done. i, Percent of T3 tumour-bearing mice surviving after three doses of anti-PD1 started on different days ( or 12) post-tumour transplant, followed by two subsequent doses injected every 3 days. The numbers above each bar represent the number of mice that rejected the
3/5 cells (Fig. 3c). By contrast, mixtures of LDVax-induced effector T cells plus HDVax-induced T3-specific cluster 3 cells did not reject T3 tumours (Fig. 3c). Similar mixtures containing classical cluster
tumours over the total mice used pooled from two experiments. Data in panels c,f are expressed as mean tumour diameter s.e.m. and are representative of three and two experiments, respectively. Statistics were done using two-way ANOVA, with multiple comparisons corrected using Sidak’s multiple comparison test. Data in panels are expressed as mean s.e.m. representing two experiments; statistics were performed using one-way ANOVA, with multiple comparisons corrected with Tukey’s method. j, GSEA comparing the gene signature of cluster 3 cells (bulk RNA-seq) to the gene signature of different cell subsets. The red line shows the maximal deviation from zero of the curve, which is basically equal to the enrichment score in these cases. The enrichment score was used to identify the value identification for each gene set by permutation test. NES, normalized enrichment score.k, Box plot showing the frequency of Tr1-like cells between responders and non-responders within the Sade-Feldman et al. dataset (left) and durable clinical benefits (DCBs) or non-DCBs within the Awad et al. dataset (right). Statistics were performed using -tests without assuming equal variances. The values in the Sade-Feldman et al. dataset was , d.f. ; no significant difference in the Awad et al. dataset was found ( ). Data are shown as box plots extending from the 25th to 75th percentiles, with the median as the centre and the whiskers corresponding to the minimum and maximum values.
cells instead of cluster 3 cells only partially inhibited tumour rejection. As expected, mixtures of T3-LDVax-induced effector T cells plus either LDVax-induced T3-specific cluster 3 cells or polyclonal T cells from T3-HDVax-treated mice lacking mItgb1 specificity did not suppress T3 rejection (Extended Data Fig. 4d). Thus, tumour-specific HDVax-induced cells populating cluster 3 are both necessary and sufficient to mediate inhibition of antitumour efficacy.
LILRB4 is a marker of inhibitory cells
To better define the immunosuppressive cells populating cluster 3 and derive insights into their origins and mechanism of action, we subjected isolated HDVax-induced cluster 3, cluster 5 ( cells) and non-cluster cell populations to deep RNA-seq and compared their transcriptomes. Volcano plot analyses confirmed that cluster 3 cells expressed high levels of Gzmb and Ccl5 (Fig. 3d). This characteristic was not shared either by cells or non-cluster cells. T3-HDVax-induced cluster 3 cells also showed selective enrichment in the expression of two surface markers known to have immunosuppressive activities: Lilrb4 (encoding ILT3, CD85k or gp49b) and Sema4a . Flow cytometry analyses revealed that HDVax cluster 3 cells could be divided into four subpopulations based on single or dual expression of LILRB4 and SEMA4a proteins (Fig. 3e and Extended Data Fig. 4e-g). Adoptive transfer of HDVax-induced LILRB4 SEMA4a or LILRB4 SEMA4 mItgb1-specific CD4 T cells comparably ablated T3 tumour rejection in the Rag2-/ ACT assay (Fig. 3f). By contrast, adoptively transferred SEMA4a single-positive cells or double-negative cells did not inhibit tumour rejection.
As these results established surface markers that identified the inhibitory cells, we asked whether they were present in progressively growing tumours that became insensitive to anti-PD1. Indeed, the frequency of FOXP3_LILRB4 + TILs increased in progressively growing, untreated T3 tumours (Fig. 3g,h), and their appearance correlated with the onset of resistance to anti-PD1 (Fig. 3i). Similar increases in cells were not observed under these conditions. These results, together with the scRNA-seq data, reveal that LILRB4 expression identifies a non- cell suppressive CD4 T cell population induced by HDVax that also develops in untreated progressively growing tumours as they develop ICT insensitivity.
HDVax-induced cells are Tr1 cells
GSEA revealed that cluster 3 cells display a distinctive gene signature differentiating them from FOXP3+ cells cells and chronically stimulated exhausted T cells (Extended Data Fig. 5a). HDVax-induced inhibitory cells remained FOXP3 after 10 days of incubation with different cytokines in vitro (Extended Data Fig. 5b). In addition, HDVax-induced cells were not derived from cells as evidenced by lack of shared TCR clonotypes populating cluster 3 and cluster 5 and in vivo lineage-tracing studies using FOXP3 lineage-tracing mice (Extended Data Fig. 5c,d). Thus, cluster 3 cells were neither precursors of, nor differentiated from, cells. Conversely, the gene signature of cluster 3 cells significantly overlapped those reported for mouse and human Tr 1 cells , and cytotoxic cells (Fig. 3j). In addition, the proportion of Tr1-like cells was higher in patients displaying resistance to anti-PD1 therapy and personalized cancer vaccines (Fig.3k). Whereas expression of LAG3 and CD49b (sometimes used to define Tr1 cells ) were not different in TILs of HDVax-treated versus LDVax-treated mice, these markers were selectively expressed on mItgb1-specific CD4 T cells in the spleens of T3-HDVax-treated mice, but not in T3-LDVax-treated mice (Extended Data Fig. 5e). T3-HDVax-induced LILRB4 cluster 3 cells produced high amounts of IL-10 and low amounts of IL-2 (hallmarks of Tr1 cells), as well as low levels of TNF and GM-CSF compared with LILRB4 cluster 3 cells, cells or non-cluster cells (Extended Data Fig. 5 g ). TGF was produced mainly by non-cluster cells and cells (Extended Data Fig. 5f). IL-10 blockade using anti-IL-10 or anti-IL-10R did not rescue HDVax inhibition (Extended Data Fig. 5h,i), indicating that IL-10 was not sufficient to impede antitumour efficacy.
These data suggest that HDVax and progressively growing tumours that become insensitive to anti-PD1 therapy do so partly because of elaboration of inhibitory FOXP3- CD4 Tr1 cells.
Anti-LILRB4 reverses suppression by Tr1-like cells
On the basis of the finding that inhibitory Tr1 cells expressed LILRB4 protein on their surface, we asked whether a recently reported blocking LILRB4 monoclonal antibody could abrogate the suppressive actions of these cells. As expected, in Rag2-/ mice, transfer of T3-LDVax-induced effector T cells induced T3 rejection, and mixtures of effector T cells plus T3-HDVax-induced Tr1 cells inhibited rejection (Fig. 4a). Whereas addition of control monoclonal antibody to the latter mixture did not reverse inhibition of tumour rejection, addition of anti-LILRB4 facilitated tumour rejection. Similarly, whereas transfer of HDVax-induced cluster 3 cells from WT mice suppressed tumour rejection in this assay, transfer of cluster 3 cells from Lilrb4 mice did not inhibit tumour rejection (Extended Data Fig. 6a). Moreover, T3 tumours grew progressively in WT mice treated with T3-HDVax and control monoclonal antibody, but were rejected in WT mice receiving the combination of T3-HDVax + anti-LILRB4 (Fig. 4b). Similar results were observed using the F244 and 1956 sarcoma models with their respective HDVax treatments (Extended Data Fig. 6b,c, respectively).
We next asked whether anti-LILRB4 treatment provoked phenotypic and/or functional changes in tumour-specific HDVax-induced Tr1 cells. We confirmed that anti-LILRB4 did not deplete LILRB4-expressing CD4 TILs in vivo (Extended Data Fig. 6d). Treatment with this monoclonal antibody decreased levels of GZMB/CCL5 expressed per CD4 T cell harvested from T3 (Fig. 4c,d) or F244 tumour-bearing mice (Extended Data Fig. 6e). A similar reduction of GZMB/CCL5-expressing CD4 TILs was observed when a comparison was made between 1956 tumour-bearing WT mice and mice with a targeted disruption of the Lilrb4 gene (Extended Data Fig. 6f). These results implied a connection between cellular expression of LILRB4 and GZMB.
To test the role of GZMB in the Tr1-dependent inhibitory process, we performed the Rag2-‘ ACT assay using cluster 3 cells derived from T3-HDVax-treated WT versus Gzmb mice after first confirming similar levels of LILRB4 expression in cells derived from each strain. Whereas T3-HDVax-induced cluster 3 cells from WT mice displayed potent inhibition of tumour rejection, T3-HDVax-induced Gzmb cluster 3 cells displayed reduced inhibitory capacity (Fig. 4e).
We then assessed the capacity of anti-LILRB4 to affect the induction of other CD4 T cell-derived products that provide help to antitumour responses. LILRB4 blockade restored IL-2 production in HDVax-induced cells (Fig. 4f,g) and enhanced their expression of IFN TNF and CD40L in both T3 and F244 tumour models (Extended Data Fig. 6g,j). Concomitant with these changes, anti-LILRB4 increased the number of mLama4-specific CD8 TILs and decreased expression of the exhaustion markers PD1, LAG3 and TIM3. Similar findings were recapitulated using F244 tumours and F244-HDVax (Extended Data Fig. 7a-e). Thus, in mice, the inhibitory Tr1-like cells are marked by expression of LILRB4, and their inhibitory capacity is reversed either by monoclonal antibody blockade or genetic deletion of the protein that, in turn, reduces the expression of GZMB and increases expression of IL-2 and other molecules that facilitate antitumour efficacy.
CD8-IL-2 circumvents HDVax suppression
The ability of anti-LILRB4 to restore IL-2 production in HDVax-induced CD4 T cells prompted us to ask whether IL-2 supplementation would restore effector cell function and tumour rejection. Although WT IL-2 is an approved therapy for certain cancers, its clinical use is limited due to its pleiotropic effects and high toxicity . We therefore utilized a T cell cis-targeted IL-2 mutein (CD8-IL-2) that binds selectively to cells but not to cells, natural killer cells or
Fig. 4 | Anti-LILRB4 treatment reverses HDVax induction of GZMB/CCL5 and suppression of IL-2 production. a, T3 tumour outgrowth in Rag2-/ mice receiving T cells from LDVax-treated mice plus HDVax-induced, CD25-negative mItgb1-specific CD4 T cells (approximately ) followed by anti-LILRB4 or control (CTRL) monoclonal antibody (mAb) treatment ( (PBS), 5 (LDVax and anti-LILB4) and 8 for control mAb).b, T3 tumour outgrowth in WT mice treated with HDVax alone or plus anti-LILRB4 ( (PBS) and 10 for other groups).c, Representative flow cytometry plots showing GZMB/CCL5 expression in CD4 TILs pregated on CD4 . d, Frequency (top) and numbers (bottom) of GZMB/CCL5-expressing CD4 TILs in T3 tumours 7 days after treatments ( ). e, 1956 Tumour outgrowth in Rag2-/- mice receiving T cells from LDVax WT mice and HDVax-induced, CD25-negative mItgb1-specific CD4 T cells (approximately ) from WT or Gzmb mice
( for PBS and LDVax, and 7 for other groups). f, Representative flow cytometry plots show the IL-2 expression in CD4 TILs. g, Frequency of cells expressing IL-2 in CD4 T cells in multiple mice ( ). h, T3 tumour outgrowth in WT mice treated as shown and detailed in the Methods ( ). , T3 tumourbearing mice were treated as in panel , and the frequency ( ) and numbers ( ) of mLama4-specific CD8 TILs, and the ratio of mLama4-specific CD8 TILs to Tr1-like cells ( ) were assessed on day 13 TILs ( ). The data in panels are expressed as mean tumour diameter s.e.m. and represent three, two, two and three experiments, respectively; statistics were done using two-way ANOVA, with multiple comparisons corrected using Sidak’s multiple comparison test. Data in panels are shown as mean s.e.m. representing three, three, two and three experiments, respectively; statistics were done using one-way ANOVA, with multiple comparisons corrected with Tukey’s method.
conventional CD4 T cells . CD8-IL-2 selectively expands CD8 T cells in mice and primates and maintains CTL function without inducing toxicity . Neither T3-HDVax nor subtherapeutic doses of CD8-IL-2 ( ) induced tumour rejection in T3-bearing mice as monotherapies. By contrast, the combination of T3-HDVax plus subtherapeutic doses of CD8-IL-2 drove T3 tumour rejection (Fig. 4h). Combining IrrVax with CD8-IL-2 was ineffective. Treatment with the combination
of WT IL-2 and either T3-HDVax or Irr-HDVax did not induce tumour rejection but resulted in significant toxicity, evidenced by weight loss in treated mice. Similar findings were observed using the F244 and 1956 tumour models (Extended Data Fig. 8a,b, respectively).
Following treatment with HDVax plus CD8-IL-2, a 2.5 -fold increase in frequency (Fig. 4i and Extended Data Fig. 8c) and absolute numbers (Fig. 4j) of mLama4-specific CD8 TILs was observed. In addition,
Fig. 5 | HDVax-induced Tr1-like cells arise from preferential cDC2 antigen presentation and kill cDC1 in a GZMB-dependent manner. a, Sorting protocol and IL-2 production by mItgb1-specific hybridoma ( , representing two experiments). b, Frequency of NIR expression in cDC1 ( ) after incubation with sorted subpopulations of HDVax-induced CD4 T cells harvested from WT mice (treated with HDVax plus control mAb or anti-LILRB4) or mice ( , representing three experiments). c, Representative flow cytometry plots showing cDC1 and cDC2 in T3 tumours pregated on and , Frequency (top) and numbers (bottom) of cDC1 in T3 tumours assessed 5 days post-vaccination ( , representing three experiments). e, Frequency (top) and numbers (bottom) of LILRB4 CD25 FOXP3 CD4 T cells in 1956 TILs harvested from WT or tumour-bearing
mice after vaccination ( , representing three experiments). f, Frequency (top) and numbers (bottom) of cDC1 in 1956 tumours in WT and mice assessed 5 days post-vaccination ( , representing three experiments). g, 1956 Tumour outgrowth in WT and mice vaccinated with 1956 HDVax, LDVax or IrrVax on days 5 and 16. h, 1956 Tumour outgrowth in WT and mice treated with of anti-PD1 on days 12,15 and 18 ( , representing two experiments). Data in panels are expressed as mean s.e.m. Data in panels are expressed as mean tumour diameter s.e.m. Statistics in panel d were done using one-way ANOVA, with multiple comparisons corrected with Tukey’s method. Statistics in panels were determined by twoway ANOVA, with multiple comparisons corrected using Sidak’s multiple comparison test.
CD8-IL-2 treatment was associated with significant increases in the ratio of TILs to Tr1-like cells (Fig. 4k). Similar findings were also observed in the F244 tumour model (Extended Data Fig. 8d-f). Thus, targeting an IL-2 mutein to T cells circumvents HDVax inhibition by increasing the frequency of antigen-specific cytotoxic cells, leading to tumour regression.
Induction of inhibitory Tr1 cells requires cDC2s/ monocytes
We considered the possibility that the different outcomes between HDVax and LDVax might reflect differences in antigen-presenting cells (APCs) that present the mItgb1 neoAg. We explored this possibility by vaccinating naive mice with either T3-LDVax or T3-HDVax, isolating
different APC populations at different time points and testing their capacity to present mItgb1 to a mItgb1-specific T cell hybridoma . Using T3-LDVax, we observed that both cDC1s and cDC2s presented the mItgb1 neoepitope to approximately the same extent (Extended Data Fig. 9a). By contrast, using T3-HDVax, the cDC2 population emerged as the primary APCs (Fig. 5a). Consistent with this result, HDVax but not LDVax elicited robust antibody responses to the mItgb1 peptide (Extended Data Fig. 9b,c). As expected, HDVax-induced Tr1 cells expressed high levels of GZMB and perforin (Fig. 3c and Extended Data Fig. 9d), suggesting that they might manifest at least some of their inhibitory function by killing an MHC-II-expressing APC . As T3 cells never express MHC-II, we considered the possibility that the target cells for cytotoxic Tr1 might be cDC1 because they present both MHC-I and MHC-II tumour epitopes and are, therefore, particularly
important to the antitumour response. In vitro killing assays showed that T3-HDVax-induced Tr1 cells but not non-Tr1 CD4 T cells killed mltgb1-pulsed cDC1s but not cDC2s in a GZMB-dependent manner (Fig. 5b and Extended Data Fig. 9e,f). Of note, Tr1 cells harvested from T3-HDVax-treated mice in the presence of anti-LILRB4 showed reduced in vitro killing activity. In addition, the frequency and number of cDC1s recovered from T3-HDVax-treated tumour-bearing mice were reduced by 4.5 -fold compared with mice treated with T3-LDVax or PBS. CDC1 recovery was partially restored when T3-HDVax was combined with anti-LILRB4 treatment (Fig. 5c,d and Extended Data Fig. 9g) or when HDVax was administered to Lilrb4 mice (Extended Data Fig. 9h). Similarly, treatment with anti-CTLA4, which depletes Tr1 cells and cells (Extended Data Fig. 3j), restored recovery of cDC1s in T3-HDVax mice (Extended Data Fig. 9i). No reduction in cDC1 recovery was observed in T3-HDVax-treated Gzmb mice (Extended Data Fig. 9j). Thus, vaccines containing high doses of MHC-II neoAg favour antigen presentation predominantly by cDC2s, leading to the formation of cytotoxic Tr1 cells that kill tumour-antigen-presenting cDC1s in a GZMB-dependent manner.
HDVax protects mice lacking cDC2s/monocytes
We hypothesized that the genetic elimination of cDC2s/monocytes in mice might reduce the generation of Tr1 cells and ablate the suppressive effect of HDVax. We therefore injected 1956 sarcoma cells into either C57BL/6 WT mice or C57BL/6 mice lacking cDC2s/ monocytes ( ) but still capable of generating . In contrast to WT mice, 1956-HDVax-treated mice showed neither an increase in the frequency of Tr1 cells (Fig. 5e) nor a reduction in cDC1 recovery (Fig. 5f). As expected, 1956 tumours grew progressively in either untreated or Irr-HDVax-treated WT or mice, and were rejected in both strains when tumour-bearing mice were treated with 1956-LDVax. 1956-HDVax induced tumour rejection in mice but not in WT mice (Fig. 5g). Of note, the frequency of Tr1 cells was comparable in both WT mice and in mice lacking cDC1s (IRF8 mice) (Extended Data Fig. 10b).
As we observed increased Tr1 cells in progressively growing sarcomas as they lost sensitivity to anti-PD1 (Fig. ), we compared the appearance of these cells in 1956 tumours growing in WT versus mice. Tr1 cells accumulated in progressively growing 1956 tumours in WT mice as they became insensitive to anti-PD1. By contrast, Tr1 cells did not accumulate in 1956 tumours growing in mice (Extended Data Fig.10c,d). Finally, anti-PD1 induced the rejection of 12 -day-established 1956 tumours in mice but not in WT mice (Fig. 5h), suggesting that the enhanced efficacy of anti-PD1 in the absence of cDC2s and/or monocytes was manifested, at least in part, by the failure to generate cytotoxic Tr1 cells. These results reveal that under HDVax conditions, cDC2s and/or monocytes induce cytotoxic Tr1 CD4 T cells that kill or compromise the integrity/stability of cDC1s, leading to defects in the development/maturation of cytotoxic T cell responses, thereby compromising antitumour effects of immunity.
Discussion
The work presented here shows that the effectiveness of therapeutic tumour-specific neoAg peptide vaccines heavily depends on the dose of the MHC-II neoAg contained in the vaccine. We demonstrated that overdosing an MHC-II neoepitope expressed by the tumour in the vaccine can completely reverse vaccine efficacy. This observation is particularly timely and important as therapeutic cancer vaccines are now being actively pursued as personalized cancer immunotherapy . We do not yet understand the rules governing the amounts of MHC-II neoAgs needed for optimal vaccine effectiveness, nor are we proposing that this needs to be empirically determined for the tumour of each patient. Rather, we focus on defining the nature of the inhibitory T cell generated under ineffective vaccination conditions and then validate strategies to circumvent the deleterious effects of high doses of the MHC-II neoAg on the antitumour response. Using this approach, we showed that the inhibition of antitumour efficacy by HDVax is due to the induction of cytolytic tumour antigen-specific Tr1 cells that kill cDC1s in vitro and in vivo. Thus, our work identifies a critical inhibitory function of a known CD4 T cell subset that has not been generally associated with cancer immunotherapy or cancer immune escape. Our demonstration that Tr1 cells become detectable in progressively growing tumours as they become resistant to anti-PD1 argues strongly that the HDVax-induced cells are orthologues of those that arise during progressive tumour growth and, therefore, are physiologically relevant participants in cancer-related immunosuppression . We then defined the mechanisms by which Tr1 cells are induced and demonstrated three alternative strategies to circumvent their activity in vivo.
We also showed that the phenotypic markers on Tr1 cells differ based on the anatomical sites from which they are isolated. Thus, whereas HDVax-induced Tr1-like cells in the spleen are differentially marked by expression of CD49b and LAG3 (ref. 39), the cells in TILs lack differential expression of these markers but still retain the capacity to produce IL-10, the signature cytokine of Tr1 cells. By contrast, LILRB4 expression clearly identifies mouse Tr1-like cells from both the spleen and the tumour microenvironment. However, whereas LILRB4 in mice is encoded by a single Gb49b gene, it belongs to a more complex family in humans comprising five different genes . Efforts to define the human orthologue will be an essential goal of future work and could provide a tool to readily identify the presence of such cells in patients with cancer. This is important because HDVax-induced inhibitory Tr1 cells and effector LDVax-induced helper CD4 T express similar levels of CD39, suggesting that other factors beyond CD39 are required to differentiate between CD4 T cells that inhibit versus promote tumour rejection .
Recent work by others has demonstrated that certain tumour antigens, termed inhibigens, can suppress antitumour efficacy of cancer vaccines and anti-PD1 (ref. 56), and that their inhibitory function was due to their inherent but undefined structural characteristics. This classification differentiates inhibigens from the MHC-II neoAgs presented in the current study, whose activating versus inhibitory properties are linked to their dosing. Our study raises the possibility that any MHC-II neoAg can be inhibigen-like when produced or released at elevated levels, thus broadening the inhibigen definition. It will also be essential in the future to explore whether the induction of Tr1 cells depends on the type of vaccine platform used (for example, peptide-based versus DNA-based versus RNA-based vaccines). Future studies will address these questions. Nevertheless, identifying Tr1 cells and their immunosuppressive biological function in cancer and providing three strategies to circumvent their inhibitory functions provide potentially novel targets for improving the effectiveness of cancer immunotherapies in general.
Online content
Any methods, additional references, Nature Portfolio reporting summaries, source data, extended data, supplementary information, acknowledgements, peer review information; details of author contributions and competing interests; and statements of data and code availability are available at https://doi.org/10.1038/s41586-024-07752-y.
4. Khattri, R., Cox, T., Yasayko, S. A. & Ramsdell, F. An essential role for scurfin in CD4 CD25 T regulatory cells. Nat. Immunol. 4, 337-342 (2003).
5. Thornton, A. M. & Shevach, E. M. CD4 immunoregulatory T cells suppress polyclonal T cell activation in vitro by inhibiting interleukin 2 production. J. Exp. Med. 188, 287-296 (1998).
6. Eschweiler, S. et al. Intratumoral follicular regulatory T cells curtail anti-PD-1 treatment efficacy. Nat. Immunol. 22, 1052-1063 (2021).
7. Zappasodi, R. et al. Non-conventional inhibitory CD4+Foxp3-PD-1 T cells as a biomarker of immune checkpoint blockade activity. Cancer Cell 34, 1017-1032.e7 (2018).
8. Alspach, E. et al. MHC-II neoantigens shape tumour immunity and response to immunotherapy. Nature 574, 696-701 (2019).
9. Ahrends, T. et al. CD4 cell help confers a cytotoxic T cell effector program including coinhibitory receptor downregulation and increased tissue invasiveness. Immunity 47, 848-861.e5 (2017).
10. Bennett, S. R., Carbone, F. R., Karamalis, F., Miller, J. F. & Heath, W. R. Induction of a cytotoxic T lymphocyte response by cross-priming requires cognate cell help. J. Exp. Med. 186, 65-70 (1997).
11. Ossendorp, F., Mengedé, E., Camps, M., Filius, R. & Melief, C. J. Specific T helper cell requirement for optimal induction of cytotoxic lymphocytes against major histocompatibility complex class II negative tumors. J. Exp. Med. 187, 693-702 (1998).
12. Klarnet, J. P. et al. FBL-reactive CD8 cytotoxic and CD4 helper T lymphocytes recognize distinct Friend murine leukemia virus-encoded antigens. J. Exp. Med. 169, 457-467 (1989).
13. Dolina, J. S. et al. Linked cell neoantigen vaccination overcomes immune checkpoint blockade resistance and enables tumor regression. J. Clin. Invest. 133, e164258 (2023).
14. Pot, C., Apetoh, L. & Kuchroo, V. K. Type 1 regulatory T cells (Tr1) in autoimmunity. Semin. Immunol. 23, 202-208 (2011).
15. Koch, K. et al. Increased frequency of CD49b/LAG-3+ type 1 regulatory T cells in HIV-infected individuals. AIDS Res. Hum. Retroviruses 31, 1238-1246 (2015).
16. McGuirk, P., McCann, C. & Mills, K. H. Pathogen-specific T regulatory 1 cells induced in the respiratory tract by a bacterial molecule that stimulates interleukin 10 production by dendritic cells: a novel strategy for evasion of protective T helper type 1 responses by Bordetella pertussis. J. Exp. Med. 195, 221-231 (2002).
17. Bergmann, C. et al. T regulatory type 1 cells in squamous cell carcinoma of the head and neck: mechanisms of suppression and expansion in advanced disease. Clin. Cancer Res. 14, 3706-3715 (2008).
18. Groux, H. et al. A CD4 T-cell subset inhibits antigen-specific T-cell responses and prevents colitis. Nature 389, 737-742 (1997).
19. Gubin, M. M. et al. Checkpoint blockade cancer immunotherapy targets tumour-specific mutant antigens. Nature 515, 577-581 (2014).
20. Meng, W. et al. Improvement of tumor neoantigen detection by high field asymmetric waveform ion mobility mass spectrometry. Cancer Immunol. Res. https://doi.org/10.1158/ 2326-6066.CIR-23-0900 (2024).
21. Gubin, M. M. et al. High-dimensional analysis delineates myeloid and lymphoid compartment remodeling during successful immune-checkpoint cancer therapy. Cell 175, 1014-1030.e19 (2018).
22. Gabryšová, L. et al. c-Maf controls immune responses by regulating disease-specific gene networks and repressing IL-2 in CD4 T cells. Nat. Immunol. 19, 497-507 (2018).
23. Sharma, N., Atolagbe, O. T., Ge, Z. & Allison, J. P. LILRB4 suppresses immunity in solid tumors and is a potential target for immunotherapy. J. Exp. Med. 218, e20201811 (2021).
24. Cella, M. et al. A novel inhibitory receptor (ILT3) expressed on monocytes, macrophages, and dendritic cells involved in antigen processing. J. Exp. Med. 185, 1743-1751 (1997).
25. Chang, C. C. et al. Tolerization of dendritic cells by cells: the crucial role of inhibitory receptors ILT3 and ILT4. Nat. Immunol. 3, 237-243 (2002).
26. Manavalan, J. S. et al. High expression of ILT3 and ILT4 is a general feature of tolerogenic dendritic cells. Transpl. Immunol. 11, 245-258 (2003).
27. Chapoval, S. P. et al. Semaphorin 4A stabilizes human regulatory T cell phenotype via plexin B1. Immunohorizons 3, 71-87 (2019).
28. Delgoffe, G. M. et al. Stability and function of regulatory T cells is maintained by a neuropilin-1-semaphorin-4a axis. Nature 501, 252-256 (2013).
29. Gagliani, N. et al. Th17 cells transdifferentiate into regulatory T cells during resolution of inflammation. Nature 523, 221-225 (2015).
30. Choi, J. et al. Bcl-6 is the nexus transcription factor of T follicular helper cells via repressor-of-repressor circuits. Nat. Immunol. 21, 777-789 (2020).
31. Crawford, A. et al. Molecular and transcriptional basis of CD4 T cell dysfunction during chronic infection. Immunity 40, 289-302 (2014).
32. van der Veeken, J. et al. Genetic tracing reveals transcription factor Foxp3-dependent and Foxp3-independent functionality of peripherally induced cells. Immunity 55, 1173-1184.e7 (2022).
33. Zhang, H. et al. An IL-27-driven transcriptional network identifies regulators of IL-10 expression across T helper cell subsets. Cell Rep. 33, 108433 (2020).
34. Heinemann, C. et al. IL-27 and IL-12 oppose pro-inflammatory IL-23 in T cells by inducing Blimp1. Nat. Commun. 5, 3770 (2014).
35. Cook, L. et al. Suppressive and gut-reparative functions of human type 1 T regulatory cells. Gastroenterology 157, 1584-1598 (2019).
36. Sledzinska, A. et al. Regulatory T cells restrain interleukin-2- and blimp-1-dependent acquisition of cytotoxic function by CD4 T cells. Immunity 52, 151-166.e6 (2020).
37. Sade-Feldman, M. et al. Defining T cell states associated with response to checkpoint immunotherapy in melanoma. Cell 175, 998-1013.e20 (2018).
38. Awad, M. M. et al. Personalized neoantigen vaccine NEO-PV-O1 with chemotherapy and anti-PD-1 as first-line treatment for non-squamous non-small cell lung cancer. Cancer Cell 40, 1010-1026.e11 (2022).
39. Gagliani, N. et al. Coexpression of CD49b and LAG-3 identifies human and mouse T regulatory type 1 cells. Nat. Med. 19, 739-746 (2013).
40. Pachella, L. A., Madsen, L. T. & Dains, J. E. The toxicity and benefit of various dosing strategies for interleukin-2 in metastatic melanoma and renal cell carcinoma. J. Adv. Pract. Oncol. 6, 212-221 (2015).
41. Moynihan, K. D. et al. IL-2 targeted to cells promotes robust effector T cell responses and potent antitumor immunity. Cancer Discov. https://doi.org/10.1158/2159-8290.CD-23-1266 (2024).
42. Magnani, C. F. et al. Killing of myeloid APCs via HLA class I, CD2 and CD226 defines a novel mechanism of suppression by human Tr1 cells. Eur. J. Immunol. 41, 1652-1662 (2011).
43. Ferris, S.T. et al. cDC1 prime and are licensed by CD4 + T cells to induce anti-tumour immunity. Nature 584, 624-629 (2020).
44. Huang, X. et al. Differential usage of transcriptional repressor Zeb2 enhancers distinguishes adult and embryonic hematopoiesis. Immunity 54, 1417-1432.e7 (2021).
45. Liu, T. T. et al. Ablation of cDC2 development by triple mutations within the Zeb2 enhancer. Nature 607, 142-148 (2022).
46. Durai, V. et al. Cryptic activation of an Irf8 enhancer governs cDC1 fate specification. Nat. Immunol. 20, 1161-1173 (2019).
47. Ott, P. A. et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 547, 217-221 (2017).
48. Weber, J. S. et al. Individualised neoantigen therapy mRNA-4157 (V940) plus pembrolizumab versus pembrolizumab monotherapy in resected melanoma (KEYNOTE-942): a randomised, phase 2b study. Lancet 403, 632-644 (2024).
49. Sahin, U. et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature 547, 222-226 (2017).
50. Carreno, B. M. et al. Cancer immunotherapy. A dendritic cell vaccine increases the breadth and diversity of melanoma neoantigen-specific T cells. Science 348, 803-808 (2015).
51. Saxena, M., van der Burg, S. H., Melief, C. J. M. & Bhardwaj, N. Therapeutic cancer vaccines. Nat. Rev. Cancer 21, 360-378 (2021).
52. Zheng, L. et al. Pan-cancer single-cell landscape of tumor-infiltrating T cells. Science 374, abe6474 (2021).
53. Arm, J. P., Nwankwo, C. & Austen, K. F. Molecular identification of a novel family of human Ig superfamily members that possess immunoreceptor tyrosine-based inhibition motifs and homology to the mouse gp49B1 inhibitory receptor. J. Immunol. 159, 2342-2349 (1997).
54. Balanca, C. C. et al. PD-1 blockade restores helper activity of tumor-infiltrating, exhausted PD-1 CD39 CD4 T cells. JCI Insight https://doi.org/10.1172/jci.insight. 142513 (2021).
55. Martinez-Gomez, C. et al. Circulating exhausted PD- CD helper CD4 T cells are tumor-antigen-specific and predict response to PD-1/PD-L1 axis blockade. Cancers https://doi.org/10.3390/cancers14153679 (2022).
56. Lam, H. et al. An empirical antigen selection method identifies neoantigens that either elicit broad antitumor T-cell responses or drive tumor growth. Cancer Discov. 11, 696-713 (2021).
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
(c) The Author(s) 2024
Methods
Mice
WT male 129S6 mice (for experiments involving T3 and F244 cells) and female C57BL/6 mice (for experiments involving 1956 cells) were purchased from Taconic Farms and Charles River, respectively. on the C57BL/6 background and Rag2-/ mice (on the 129S6 and C57BL/6 backgrounds) were bred and housed in our specific pathogen-free facility. cDC2-deficient and cDC1-deficient (IRF8 mice on the C57BL/6 background were obtained from K. Murphy (Washington University in St. Louis). Lilrb4a (encoding gp49b)-knockout mice were obtained from M. Colonna (Washington University in St. Louis) and were generated by the Genome Engineering and iPSC Center (GEiC) and the Transgenic, Knockout, and Micro-Injection Core, Department of Pathology and Immunology, at Washington University in St. Louis. A single guide RNA (cctttagttgcagctcatccata) and Cas9 proteins were electroporated into zygotes before being transferred into pseudo-pregnant B6/J female mice. The resulting founders were screened using next-generation sequencing (NGS). A male founder with a 21 -nucleotide deletion in exon 4 was bred with B6/J female mice obtained from the Jackson Laboratory (JAX stock #000664) for one generation, and heterozygous F1 mice were intercrossed to generate knockouts. The integrity of the highly homologous gene Lilrb4b was confirmed by NGS. The absence of LILRB4A protein expression in blood monocytes and neutrophils was confirmed by flow cytometry using the H1.1 antibody clone (BioLegend). Lineage tracing eGFP mice on the C57BL/6 background were obtained from J. Kipnis (Washington University in St. Louis). Mice were used between 8 and 10 weeks of age. All in vivo experiments were performed in our specific pathogen-free facility using procedures approved by the AAALAC-accredited Animal Studies Committee of Washington University in St Louis and followed all relevant ethical regulations.
Tumour transplantation
T3, F244 and 1956 are MCA-induced sarcoma cells that were previously generated in our laboratory in male 129S6 (T3 and F244 cell lines) and female C57BL/6 mice (1956 cell line). Tumour cells were cultured in RPMI medium (Hyclone) supplemented with 10% FCS (Hyclone). All cell lines used were negative for mycoplasma and other infectious agents. For tumour inoculation, tumour cells were harvested by trypsinization, washed three times with PBS and resuspended in PBS at a density of cells per millilitre. Then, was injected subcutaneously into the rear flanks of syngeneic recipient mice. Tumour growth in individual mice was determined using callipers and expressed as the average of two perpendicular diameters. Mice were euthanized when the tumour diameter reached 20 mm in any direction.
NeoAgs and vaccination protocol
T3 neoAgs were previously described and are listed in Supplementary Table 1. The peptide of HPV (DKCLKFYSKISEYRHY CYSLYGTTL) was used as an irrelevant non-sarcomas antigen. All peptides were purchased from Peptide 2.0 or Genscript with a specified purity of over . Endotoxin levels in all peptides tested were below (Leinco). The indicated doses of peptide were mixed with of polyinosinic-polycytidylic acid complexed with poly-l-lysine and carboxymethylcellulose (poly-ICLC; Oncovir) diluted in PBS. For therapeutic vaccination, tumour-bearing mice were injected intravenously or subcutaneously with the vaccines on days 6 and 17 post-tumour transplant. In experiments using the combination of vaccination and ICT, vaccines were administered intravenously on days 3 or 6 post-tumour transplantation. In the sorting experiments, of anti-CD40 was mixed with the second dose of the vaccine to increase the yield of the antigen-specific T cells.
In vivo antibody treatment
For in vivo treatment, LPS-free and pathogen-free anti-PD1 (rat IgG2a, clone RMP1-14), anti-CTLA4 (mouse IgG2b, clone 9D9) or anti-4-1BB (rat IgG2a, clone 3H3) monoclonal antibodies were purchased from Leinco Technologies. Tumour-bearing mice were vaccinated on day 3 post-tumour implantation and then injected intraperitoneally with of each antibody on days 3,6 and 9. To assess the role of IL-10 in HDVax-induced inhibition, monoclonal antibodies to IL-10 (rat IgGl, clone JES5-2A5) and IL-10R (rat IgG1, clone 1b1.3A) were purchased from Bioxcell and injected with HDVax. For anti-IL-10 blockade, mice were treated with 1 day before tumour implantation, followed by treatment every 3 days for the duration of the experiment. For anti-IL-10R blockade, mice were treated with 1 mg per mouse every 7 days, starting 1 day before tumour injection. Neutralizing but not depleting hamster monoclonal antibody specific for LILRB4 (clone# 2F1.F9.E6) was provided by N. Sharma and J. Allison (University of Texas (MD Anderson Cancer Center)), and mice were treated with on days 3, 6 and 9 post-tumour inoculation.
Tetramer staining
Tetramer staining for mLama4-specific CD8+ T cells and mItgb1-specific CD4 T cells was performed as previously described . TILs or splenocytes ( million) in FACS buffer were incubated with PE/ APC-labelled mLama4 peptide- tetramer at for 20 min . Then, an antibody master mix consisting of CD90.2, CD4, CD11b and live/dead dye (NIR) in FACS buffer was added to each well and incubated for 30 min at . Tetramers were obtained from the Bursky Center Immunomonitoring Laboratory at Washington University School of Medicine in St. Louis.
Flow cytometry antibodies
Flow antibodies were: IFN (XMG1.2; 1:100; 505808), TNF (MP6-XT22; 1:100; 506323), CD200 (OX-90; 1:200; 123820), PD1 (29F.1A12; 1:200; 135228), CCL5 (2E9/CCL5; 1:500; 149106), GZMB (QA16A02; 1:50; 372216), TIM3 (RMT3-23; 1:200;119723), CD25 (PC61; 1:100; 102036), CD154 (MR1; 1:100; 106506), CD152 (UC10-4B9; 1:100; 106318), CD4 (RM4-5; 1:500), IL-2 (Jes6-5H4; 1:50; 503826), LILRB4 (H1.1; 1:200; 144904), CD11b (M1/70; 1:800; 101226), XCR1 (ZET; 1:100; 148206), MHC-II (M5/114.15.2; 1:1000; 107641), CD11c (N418; 1:200; 117336), CD172a (P84; 1:500; 144008), Zombie NIR fixable viability dye (1:500; 423106), CD86 (GL-1; 1:200; 105042), CD80 (16-10A1; 1:200; 104712) and CD40 (FGK45; 1:100; 157506) all from BioLegend; CD90.2 (53-2.1; 1:800; 565257), CD45 (HI30; 1:800; 563791), CD39 (Y23-1185; 1:400; 567105), CD153 (RM153; 1:200; 740751), CD70 (FR70; 1:100; 740741) and CD8 (53.6.7; 1:200; 564920) from BD Biosciences; or FOXP3 (Fjk-16a; 1:50; 11-5773-82) and SEMA4a (5E3; 1:50; 46-9753-41) from eBioSciences. Foxp3/Transcription factor Staining kit (00-5523-00, Thermo Fisher) was used to stain FOXP3 and other intracellular proteins according to the manufacturing protocol. BD FACSDIVA software V9.1 on Symphony 3, LSRFortessaX20 or ARIA-II was used for collecting the flow cytometry data and for sorting, respectively. Data were analysed using FlowJo software v10.10.
Multiplex assay and antibody ELISA
TILs were harvested 7 days post-vaccination, stained with mItgbl-I-A tetramer prepared by the Immunomonitoring Laboratory core at Washington University in St. Louis using mItgb1 peptide covalently attached to the -chain, and total mItgb1-specific CD4 cells (Fig. 2c) or cells populating different clusters (Fig. 3c) were sorted by flow cytometry. Sorted cells ( ) were stimulated in a serum-free medium with irradiated splenocytes (isolated from naive mice) pulsed with .h incubation, secretion of multiple cytokines was measured using a flow-based
customized ProcartaPlex cytokine panel (Luminex Technologies) following the manufacturer’s protocol.
To determine antibody responses to the mItgb1 peptide, ELISA plates were coated with of either mItgb1 or irrelevant SLP at overnight. Plates were washed and blocked with goat serum for 2 h at room temperature, and then different dilutions of serum from LDVax or HDVax mice were added for . Plates were washed again and incubated with peroxidase-conjugated goat antibody to total mouse IgG (H + L) (115-035-003, Jackson ImmunoResearch) or different isotype-specific secondary reagents conjugated with horseradish peroxidase for 2 h at room temperature, followed by sequential addition of tetramethylbenzidine substrate (TMB). The reaction was stopped by acidification, and the optical density of each well was measured at 405 nm .
Adoptive transfer experiments
For adoptive transfer experiments into Rag2-/ mice, WT mice were vaccinated with LDVax, and 9 days later, 5 million total T cells were transferred into Rag2-/- mice that were injected with tumour cells 1 day earlier. To isolate mItgb1-specific CD4 cells from HDVax mice, TILs and spleens were harvested from HDVax-treated mice, and cells were enriched using the CD4 cell isolation kit from Miltenyi (130-117-043) stained with mitgb1 tetramer, and CD25-negative mItgb1-specific CD4 T cells were purified by flow cytometry. Half a million mItgb1-specific CD4 T cells were injected intratumourally (in ) into T3 tumour-bearing Rag2-‘ mice.
For sorting and adoptive transfer of individual clusters, cells were enriched by positive bead selection and stained with mItgb1-I-Ab tetramers, CD25, CD200, CD153 and CD39. cells were sorted based on the positive expression of CD25, and non-cluster 3/5 cells were defined by the negative expression of CD25 and positive expression of CD200 and/or CD153. Cluster 3 cells were sorted based on the negative expression of CD25, CD200 and CD153 and the positive expression of CD39. An equal number of cells ( cells) sorted from these clusters were resuspended in PBS and were injected intratumourally. To sort for LILRB4-expressing and SEMA4-expressing cells, CD25, CD200 and CD39 were used to pregate on cluster 3 cells, and cells were injected into T3 tumour-bearing Rag2-/- mice that also received cell-depleted total T cells from LDVax mice.
Intracellular cytokine and CD40L staining
Splenocytes ( ) harvested from naive mice were irradiated ( 30 Gy) and pulsed with peptide, and TILs ( ) were subsequently added, and the cell suspension was incubated at . GolgiPlug (BD Biosciences) was added 1 h later and incubated for another 4 h . Cells were stained for different surface markers, including the live/dead marker (NIR), then permeabilized using the intracellular permeabilization kit (BD Biosciences), followed by staining for IFN , TNF and CD40L (clone; SA047C3; 157010, BioLegend).
Ex vivo antigen presentation assay
Naive WT mice were intravenously vaccinated with HDVax consisting of of mItgb1 SLP and poly-ICLC (HDVax). At the specified time point, spleens and lymph nodes were harvested and digested in the presence of collagenase. APCs were FACS sorted into four different subpopulations: and .Sorted APCs ( ) were incubated with mItgb1-specific hybridoma cells in IL-2 precoated ELISPOT plates (IMMUNOSPOT). Twenty-four hours later, plates were developed following the manufacturer’s protocol, and IL-2 spots were quantified using a CTL ImmunoSpot S6 machine.
In vitro dendritic cell killing assay
To isolate dendritic cells, spleens were harvested from naive WT mice and disrupted by collagenase digestion. CD11c cells were enriched
using a positive selection kit (130-125-835, Miltney Biotec). Enriched CD11c cells were pulsed with mItgb1 SLP at and incubated with tumour-specific CD4 T cells at a 1:1 ratio for 12 h . Cells were washed twice with PBS, stained with MHC-II, CD11c, XCR1, CD172a and Zombie NIR dye to determine the frequency of NIR in cDC1s and cDC2s using flow cytometry.
CD8-IL-2
CD8-IL-2 was provided by Asher Biotherapeutics . CD8-IL-2 was generated via fusion of an IL-2 mutein that does not bind to IL-2R and displays significantly reduced binding to IL-2R ү. This mutein was then coupled to a monovalent mouserized anti-mouse CD8 antibody. Monovalent IL-2 coupling was achieved using bispecific charge pair technology, and binding was abolished via mutating the binding region of an anti-CD8 antibody. CD8-IL-2 was expressed in HEK293 cells and purified via protein A affinity chromatography, followed by ion-exchange chromatography and then size-exclusion chromatography. Therapeutic doses ( ) of CD8-IL-2 induced significant antitumour activity (approximately response rate) against 8-day-established T3 tumours when they became insensitive to anti-PD1 therapy. For the current study, CD8-IL-2 was administered at a subtherapeutic dose of diluted in PBS and injected intraperitoneally. WT IL-2 (202-IL/CF) was purchased from (R&D Systems). Each mouse was treated with 25,000 IU in PBS injected intraperitoneally daily for 5 days.
scRNA-seq analysis
UMAP clustering and separation of total and antigen-specific cells. T3 tumour-bearing mice were treated with HDVax, LDVax or PBS 6 days post-tumour transplantation. Seven days later, single-cell suspensions were prepared from TILs (pooled from seven mice for each group). Total CD4 T cells were enriched using a CD4 T cell-positive selection kit (Miltenyi). Enriched CD4 T cells were split into two fractions. One fraction was labelled with TCR (TotalSeq-C0120 anti-mouse TCR chain antibody, 109259 , BioLegend) and used as a source of total CD4 T cells. The other fraction was labelled with CD90.2 (TotalSeq-C0075 anti-mouse CD90.2 antibody, 105353, BioLegend) and used to isolate mItgb1-specific cells based on mItgb1-I-A tetramer staining and flow sorting. The two fractions (total and antigen-specific CD4 cells) were mixed at a ratio and submitted to the Genome Institute at Washington University to generate 10 X libraries using v 2 singlecell RNA-seq. Alignment, barcode assignment and unique molecular identifier counting with Cell Ranger (v6.1.1) were used to prepare count matrices for the gene expression library using the mouse genome (GRCm38) as a reference . Barcodes in all samples representing low-quality cells were filtered out using the standard knee-inflection strategy available in Cell Ranger. For downstream analysis, the Seurat package (v4.0.4) was used; genes expressed in fewer than three cells were additionally filtered out from expression matrices, and cells that contained fewer than 200 expressed genes were removed. The mitochondrial gene fraction was calculated for all cells, and cells with a mitochondrial fraction more than the highest confidence interval for scaled mitochondrial percentage were filtered out. Each sample was normalized using the SCTransform function with mitochondrial content as a variable to regress out in a second non-regularized linear regression. For integration purposes, variable genes across the samples were identified by the SelectIntegrationFeatures function with a number of features equal to 2,000 . Then, the object was prepared for integration (PrepSCTIntegration function), the anchors were found (FindIntegrationAnchors function) and the samples were integrated into the whole object (IntegrateData function) . Principal component analysis (PCA) was used for dimensionality reduction, and the first 20 principal components were used further to generate UMAP dimensionality reduction by the RunUMAP function. The clustering procedure was performed by FindNeighbors and FindClusters
with a range of resolutions (from 0.2 to 1.0 , with 0.2 as a step) and the first 20 principal components as input. Antibody-derived tags (ADT) data were normalized by a centred log-ratio transformation method, scaled and transformed to its own PCA space. Finally, cells were defined as mItgb1-specific cells and total cells (predominantly mItgb1-nonspecific cells) based on the scaled value of corresponding ADTs. Each object was iteratively cleaned for doublets as well as low-quality cells.
TCR data analyses
TCR data were aligned to the reference mouse genome GRCm38 and counted using Cell Ranger (v6.1.1) vdj workflow . The Seurat object was converted to the h5ad format, and cells that passed quality control for gene expression and TCR levels were integrated into the objects (mItgb1-specific cells and total cells). Downstream analyses, including clonotype expansion analysis and estimation of used VDJ gene pairings, were executed using the scirpy toolkit. Proportions of shared and unique clonotypes with cells (cluster 5) were counted. TCR analyses were done using the Immunarch package.
Bulk RNA-seq
Three cohorts ( ) of T3 tumour-bearing mice were vaccinated with HDVax, and 7 days later CD4 T cells were enriched from TILs using the Miltenyi mouse CD4 T cells isolation kit, stained with the mItgb1-I-A tetramer and then sorted into three replicates of CD25 ( cells), (non- cells/non-cluster 3) and (cluster 3). Cells were lysed, and mRNA was extracted using Nucleospin RNA Plus (740990.50, TaKaRa). RNA was sequenced by the Washington University Genome Institute. In brief, total RNA integrity was determined using Agilent Bioanalyzer. Library preparation was performed with 10 ng of total RNA with a Bioanalyzer RNA integrity number (RIN) score greater than 8.0. double-stranded DNA (ds-cDNA) was prepared using the SMARTer Ultra Low RNA kit for Illumina sequencing (740990.50, TaKaRa) per the manufacturer’s protocol. cDNA was fragmented using a Covaris E220 sonicator using peak incident power of 18, duty factor of , and cycles per burst of 50 for 120 s. cDNA was blunt-ended, had an A base added to the 3′ ends, and then had Illumina sequencing adapters ligated to the ends. Ligated fragments were then amplified for 12-14 cycles using primers incorporating unique dual index tags. Fragments were sequenced on an Illumina NovaSeq-6000 using paired-end reads extending 150 bases. For analysis, raw data were aligned and counted to the GRCm 38.101 reference genome by the Dragen workflow of the GTAC@MGI service. For downstream analysis, 12,000 genes with the highest number of counts across the samples were selected. All samples were normalized by regularized log transformation (rlogTransformation function) and variance stabilizing transformation (vst function), which are accomplished in the DESeq2 (v1.30.1) package. PCA was run based on the vst output, and no outliers were identified. fold change values were shrunk for all comparisons (CD25 versus CD39, and CD200 versus CD39) by the IfcShrink function, and its output was used for volcano plot construction .
Gene signature analysis
To evaluate the significance of enrichment for highly expressed genes within cluster 3, we identified the top 200 differentially expressed genes from a bulk RNA-seq dataset (Supplementary Table 2). Subsequently, we conducted a rigorous analysis of differential gene expression using the limma package on normalized count data obtained from publicly available datasets. Specifically, we performed the following pairwise comparisons.
For the cytotoxic CD4 T cells, data from GSE141540 were analysed, and a comparison of cell-ctx versus cell was performed. For the Tr1 gene signature, GSE158703 and GSE139990 were obtained, and a
comparison of Tr1-IL-10 versus or Tr1 versus naive was performed, respectively.
For the and gene signature, GSE68242 or GSE140187 were obtained, and a comparison of FOXP3 versus other or lymph node WT T versus lymph node naive was performed, respectively. For the chronically stimulated exhausted CD4 T cells, data from GSE30431 were analysed, and a comparison of CD4 D30 chronic versus CD4 D30 acute was performed. We used GSEA to ascertain the significance of enrichment for the gene signature associated with cluster 3. This method evaluates whether cluster 3-predefined gene signatures are statistically overrepresented or underrepresented within the differentially expressed gene lists derived from the publicly available bulk RNA-seq datasets for each CD4 T cell subtype.
To determine the frequency of Tr1-like cells in patients with cancer, we first subset the quality-controlled cells from each manuscript down to cells. After this, we used AUC ell to determine which cells match the human Tr1 gene set. Following this, we determined the proportion of the Tr1 cells within the CD4 T cell population of each patient. We tested for differences in Tr1 cell proportions between responders and non-responders using Student’s -tests without assuming equal variances. There is a significant difference in the proportion of Tr1 cells between responders and non-responders from the Sade-Feldman et al. dataset , d.f. . There was no significant difference in the Awad et al. dataset ( ).
GSEA for the IL-2 pathway
The FindMarkers function was used to find genes specific to both LDVax and HDVax samples. The average fold change values were used as an input in GSEA implemented in the fgsea R package (v1.16.0). Forty-three genes associated with the IL-2 signalling pathway were used for the gene set enrichment plot . For violin plots, the summary expression of IL-2 signalling genes was calculated using the sctransform (SCT) assay (data slot), scaled and divided by the number of genes in the pathway.
Statistics
GraphPad Prism software (v10.2.2) was used to perform all statistical analyses. Significance was determined using one-way ANOVA with multiple comparisons corrected with Tukey’s method unless otherwise stated in the figure legends.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Bulk RNA-seq and scRNA-seq data were deposited using the accession number GSE268302; GSE268300 (bulk RNA-seq) or GSE268301 (scRNA-seq) can be used to access raw data. The pipelines with code to process and analyse the bulk RNA-seq and scRNA-seq data are available at https://github.com/vdsukhov/neoantigen_vaccines_2024. Source data are provided with this paper.
57. Wang, L. L., Chu, D. T., Dokun, A. O. & Yokoyama, W. M. Inducible expression of the gp49B inhibitory receptor on NK cells. J. Immunol. 164, 5215-5220 (2000).
58. Zheng, G. X. et al. Massively parallel digital transcriptional profiling of single cells. Nat. Commun. 8, 14049 (2017).
59. Stuart, T. et al. Comprehensive integration of single-cell data. Cell 177, 1888-1902.e21 (2019).
60. Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
61. Aibar, S. et al. SCENIC: single-cell regulatory network inference and clustering. Nat. Methods 14, 1083-1086 (2017).
62. Sturm, G. et al. Scirpy: a Scanpy extension for analyzing single-cell T-cell receptorsequencing data. Bioinformatics 36, 4817-4818 (2020).
Acknowledgements We thank all members of the Schreiber laboratory for the discussions and for providing technical support; L. Yang and D. Bender of the Immunomonitoring Laboratory
(IML) who provided tetramers for MHC-I and MHC-II neoAgs and performed the multiparameter cytokine assessments; the Genome Engineering and iPSC Center (GEiC) at Washington University in St. Louis for guide RNA validation services; M. White for generating the Lilrb4aknockout mice; and J. Kipnis and G. G. Davanzo for providing the FOXP3 lineage-tracing mice. This work was supported by grants to R.D.S. from the National Cancer Institute of the US National Institutes of Health (RO1CA190700), the Parker Institute for Cancer Immunotherapy, and a Stand Up to Cancer-Lustgarten Foundation Pancreatic Cancer Foundation Convergence Dream Team Translational Research Grant. Stand Up to Cancer is a program of the Entertainment Industry Foundation administered by the American Association for Cancer Research. K.M.M. is supported by the National Institutes of Health (RO1Al150297, RO1CA248919, R21AI164142, RO1Al162643 and R21Al163421). M.C. is supported by the Rheumatic Diseases Research Resource-based Center at Washington University (NIH/NIAMS P3OARO73752). H.S. was supported by a postdoctoral training grant (T32CA009547) from the National Cancer Institute. M.F. was supported by the Ministry of Science and Higher Education of the Russian Federation (agreement no. 075-15-2022-301). J.P.W. is supported by the National Cancer Institute of the National Institutes of Health (5K08CA245215). The IML is supported by the Andrew M. and Jane N. Bursky Center for Human Immunology and Immunotherapy Programs and the Alvin J. Siteman Comprehensive Cancer Center. The latter is supported by a National Cancer Institute of the National Institutes of Health Cancer Center support grant (P30CA91842).
Author contributions H.S. and R.D.S. conceived and designed the experiments, interpreted the results and wrote the manuscript. H.S. performed the experiments and collected the data. Y.T. and J.P.W. identified neoAgs in 1956 and F244 tumours, respectively. V.S. and M.F. performed analyses of scRNA-seq, bulk RNA-seq and GSEA data under the supervision of M.N.A. T.-T.L. and K.M.M. generated and provided mice. D.K., S.B. and M.C. generated and provided the Lilrb4-knockout mice. N.S. and J.P.A. provided the neutralizing LILRB4 monoclonal antibody. C.D.A., J.M.W., Y.S. and H.K. provided technical assistance throughout the study. K.C.F.S. oversaw the studies performed in the IML. A.M.S. provided clinical-grade poly-ICLC. K.D.M., Y.A.Y. and I.D. provided CD8-IL-2. T.N.S. participated in the development of CD8-IL-2. All authors
contributed to the final development of the manuscript and agreed with the results and conclusions.
Competing interests R.D.S. is a cofounder, scientific advisory board member, stockholder and royalty recipient of Asher Biotherapeutics, and is a paid scientific advisory board member for A2 Biotherapeutics, NGM Biopharmaceuticals, BioLegend, Sensei Biotherapeutics Neuvogen and BlueSphere Biopharmaceuticals. K.M.M. is on the Scientific Advisory Board for Harbour Biomed. J.P.A. receives consulting fees from Achelois, Adaptive Biotechnologies, Apricity, BioAtla, BioNTech, Candel Therapeutics, Codiak, Dragonfly, Earli, Enable Medicine, Hummingbird, ImaginAb, Jounce, Lava Therapeutics, Lytix, Marker, PBM Capital, Phenomic AI, Polaris Pharma, Time Bioventures, Trained Therapeutix, Two Bear Capital and Venn Biosciences; and has ownership of stocks for Achelois, Adaptive Biotechnologies, Apricity, BioAtla, BioNTech, Candel Therapeutics, Codiak, Dragonfly, Earli, Enable Medicine, Hummingbird, ImaginAb, Jounce, Lava Therapeutics, Lytix, Marker, PBM Capital, Phenomic AI, Polaris Pharma, Time Bioventures, Trained Therapeutix, Two Bear Capital and Venn Biosciences. T.N.S. is a cofounder, scientific advisory board member and stockholder of Asher Biotherapeutics and Neogene Therapeutics, cofounder and stockholder of Cell Control Bio, consultant and stockholder at Third Rock Ventures, and a scientific advisory board member for Allogene Therapeutics, Celsius, Merus and Scenic Biotech. K.D.M., Y.A.Y. and I.D. are full-time employees of Asher Biotherapeutics. R.B. and H.A.C. are full-time employees of Natera, Inc. All other authors declare no competing interests.
Additional information
Supplementary information The online version contains supplementary material available at https://doi.org/10.1038/s41586-024-07752-y.
Correspondence and requests for materials should be addressed to Robert D. Schreiber. Peer review information Nature thanks Axel Kallies and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Reprints and permissions information is available at http://www.nature.com/reprints.
Extended Data Fig. 1|Phenotypic profiles of HDVax versus LDVax-induced cells. a, representative flow cytometry plots showing mitgbl tetramer staining in CD4 TILs. b, frequency (left) and numbers (right) of mItgb1-specific CD4 TILs assessed in day 13 T3 tumours ( , representing five experiments). c, representative flow cytometry plots showing CD25 and Foxp3 staining in mItgb1-specific CD4 TILs. d, frequency of CD25 Foxp3 cells in mItgb1-specific cells in TILs (left) and spleens (right) of T3 tumour-bearing mice ( , representing five experiments). e, frequency of CD25 Foxp3 Tregs in CD4 T cells in tumours (left) and spleens (right) of F244 tumour-bearing mice ( ,
representing three experiments).f, representative flow cytometry plots showing the frequency of IFN TNF in CD4 TILs. , frequency of IFN- TNF- expressing CD4 TILs in multiple mice ( , pooled from two experiments). h, representative flow cytometry plots showing CD40L expression in CD4 TILs. i, frequency of CD40L expression in CD4 TILs of T3 tumours after in vitro activation ( , pooled from two experiments). Data in all panels are shown as means s.e.m. values were calculated using one-way ANOVA with multiple comparisons corrected with Tukey’s method.
Extended Data Fig. 2 | Characterization of dendritic cells and CD8 TILs in HDVax, LDVax and PBS-treated mice. a, representative flow cytometry plots showing the gating strategy for cDC1 and cDC2.b, expression of CD86, CD70, MHC-II and CD40 on the surface of dendritic cells (CD45 CD90.2 CD11c MHC-II ) in T3 tumours harvested 5 days after vaccination ( , represent two experiments). c, numbers of cDC1 (top) and cDC2 (bottom) in T3 tumours assessed five days post-vaccination ( , represent three experiments).
d, representative flow cytometry plots showing the gating strategy for mLama4specific CD8 TILs in day 13 T3 TILs. e-h, frequency of mLama4-specific CD8 TILs
expressing e, TOX f, PD-1/CD39 and g, TIM-3/LAG3 ( , represent two experiments). , frequency of cells expressing IFN- TNF- in specific CD8 TILs after in vitro activation with splenocytes pulsed with mLama4 SLP in the presence of Golgi-plug followed by intracellular staining ( , pooled from three experiments). i, frequency of mLama4-specific CD8 TILs showing TIM3 and TCF-1. Data in all panels are expressed as means s.e.m. values were calculated using one-way ANOVA with multiple comparisons corrected with Tukey’s method.
Extended Data Fig. 3 | HDVax-induced T cells are deficient in Th1 gene signature and express high levels of GZMB and CCL5. a-b, violin plots showing the expression levels of multiple genes involved in , exhaustion or , effector Th1 signature in mItgbl-specific scRNAseq data.c, GSEA showing relative expression of genes involved in the IL-2 signaling pathway. d, violing plots showing the expression levels of different transcription factors involved in IL-2 signaling in mItgb1-specific scRNAseq data. e-f, frequency (left) and numbers (right) of CCL5/GZMB expressing CD4 T cells in , TILs and , spleens of T3 tumour bearing WT mice 5-7 days post vaccinations ( (TILs) and 5 (spleen) and represent three experiments). g-h, frequency (left) and numbers (right) of
CCL5/GZMB expressing CD4 T cells in , TILs and , spleens of F244 tumourbearing WT mice seven days post vaccinations ( , represent two experiments). i, frequency of CTLA-4 in different clusters of mItgb1-specific CD4 TILs of T3 tumour-bearing WT mice ( , represent theee experiments). , frequency of CCL5/GZMB expressing CD4 TILs of T3 tumour-bearing WT mice treated with HDVax plus PD-1 or CTLA-4 ( , represent three experiments). Data in all panels are shown as means s.e.m. Statistics were performed using one-way ANOVA with multiple comparisons corrected with Tukey’s method in e-h and j, and two-way ANOVA corrected with sídák’s multiple comparisons test was performed for statistics in .
Extended Data Fig. 4 | Gating and sorting strategy to patially enrich for cluster-3 cells. a, feature plot showing the expression of various genes superimposed on mItgb1 specific CD4 TILs UMAP. b, representative flow cytometry plots showing the sorting strategy for different clusters in mItgb1-specific CD4 TILs. c, representative flow cytometry plot showing the expression of CD25, CD39 and Foxp3 in different clusters after sorting. d, T3 outgrowth in RAG2-/- mice receiving total T cells from LDVax-treated WT mice ( ) alone or plus; LDVax-induced cluster-3 cells; HDVax-induced,
CD25-negative mItgb1-specific; or polyclonal (mItgb1 non-specific) CD4 T cells ( ). Data expressed as mean tumour diameter s.e.m. ( , two experiments). values were calculated using Two-way ANOVA corrected with sídák’s multiple comparisons test. e-g, frequency of LILRB4 single positive, LILRB4/SEMA4a double positive or SEMA4a single positive expressing mItgb1-specific CD4 TILs ( , represent three experiments). Data are expressed as mean s.e.m. Statistics were performed using one-way ANOVA with multiple comparisons corrected with Tukey’s method.
Extended Data Fig. 5| See next page for caption.
Extended Data Fig. 5 | Cells populating cluster-3 are distinct from Tregs. a, GSEA comparing gene signature of cluster-3 cells (bulk-RNAseq) to gene signature of different CD4 T cell subsets. Enrichment Score was used to identify the -value identification for each gene set by permutation test.b, flow cytometry plot showing CD25 and Foxp3 staining in sorted Tregs and cells populating cluser-3 after 10 days of in vitro incubation with IL-2 alone ( ), or plus IL-10 and or . The experiment represents two experiments. c, TCR clonotypes in each cluster of HDVax-induced I- -mItgb1 specific CD4 T were determined by TCRseq and compared to the TCR clonotypes in cells populating cluster-3. d, frequency of cells expressing in Tregs (CD25 Foxp3 ) or CD25 LILRB4 CD4 T cells after different treatments in spleens and TILs ( , represent one experiment). e, frequency of cells expressing LAG3 and CD49b in cluster-3 or non-cluster-3/5 of mItgb1-specific cells after different treatments in spleens and TILs ( , represent three experimetns). Data expressed as mean tumour diameter s.e.m. and statistics were performed by two-way ANOVA corrected with sídák’s multiple
comparisons test.f-g, freshly isolated CD4 T cells from different clusters were stimulated with mltgb1 SLP for 72 hr , and production of active isoforms of TGF was assessed by Multiplex (biological replicates in cluster- 5 and 3 replicates in all other clusters and each values are mean of 2 technical replicates, represent two experiments). Statistics in and were performed using one-way ANOVA with multiple comparisons corrected with Tukey’s method. g, Heatmap showing levels of different cytokines production (indicated on the heatmap x -axis). Numbers represent the mean of three independent biological replicates and represent two experiments. h, T3 tumour outgrowth in WT mice treated with HDVax or irrelevant vaccines on day 6 and IL-10 (-1, 4, 7, 10 and 14) post-T3 tumour injection ( , representing two experiments). i, T3 tumour outgrowth in WT mice treated with HDVax on day 3 and IL-10R (-1,5,11 and 18) post tumour transplantation ( , representing one experiment). Data in and expressed as mean tumour diameter s.e.m. and values were calculated using two-way ANOVA corrected with sídák’s multiple comparisons test.
Extended Data Fig. 6 | related to Fig. 4: LILRB4 treatment reverses HDVax induction of GZMB/CCL5 and suppression of IL-2 production. a, 1956 outgrowth in RAG2 mice receiving total LILRB4 T cells from LDVax-treated
LILRB4 mice plus HDVax-induced, CD25-negative CD4 T cells ( ) from WT mice or LILRB4 mice ( for PBS and LDVax) 7 for other groups). b, F244 tumour outgrowth in WT mice treated with HDVax and LILRB4 as shown ( , represent two experiments). c, 1956 tumour outgrowth in mice treated with HDVax and LILRB4 as shown ( , represent two experiments). d, frequency of cells expressing LILRB4 in CD4 T3 TILs after treatment with LILRB4 or control mAb ( , represent three experimenrts). e, frequency and numbers of GZMB/CCL5 expressing CD4 TILs in F244 tumour-bearing mice
( , represent two experiments). , frequency and numbers of GZMB/CCL5 expressing CD4 TILs in WT and LILRB4 tumour-bearing mice ( , represent two experiments). g-h, frequency of CD4 TILs expressing g, IFN- TNF- or , CD40L in T3 TILs after in vitro activation with mItgb1 peptide in the presence of Golgi-plug ( , represent two experiments). , frequency of CD4 TILs expressing , IFN- TNF- or , CD40L in F244 TILs after in vitro activation with mPlec peptide in the presence of Golgi-plug ( ). Data in a-c expressed as mean tumour diameter s.e.m and values were calculated using Two-way ANOVA corrected with sídák’s multiple comparisons test. Data in d-j are expressed as mean s.e.m. and values were calculated using one-way ANOVA with multiple comparisons corrected with Tukey’s method.
Extended Data Fig. 7 | LILRB4 blockade enhances the phenotype and the effector functions of TILs. a, numbers of mLama4 specific CD8 TILs were assessed in T3 TILs ( ). , frequency of TILs expressing IFN TNF- after in vitro activation with mLama4 SLP. c, frequency of CD8 TILs expressing IFN- TNF- after in vitro activation with mPex14 SLP ( ).
d, expression of several exhaustion markers on mLama4-specific CD8 T cells
e
F244 sarcoma TIL
in TILs of T3 tumour-bearing mice ( ). Data represent two independent experiments and are shown as mean s.e.m.e, expression of several exhaustion markers in mPex14-specific CD8 T cells in F244 TILs ( ). Data expressed are shown as mean s.e.m. values were calculated using one-way ANOVA with multiple comparisons corrected with Tukey’s method, which was used for the statistics.
Extended Data Fig. 8 | related to Fig. 4: CD8-directed, cis-targeted IL-2 treatment re-establishes the anti-tumour efficacy of HDVax in F244 and 1956 tumour models. a, F244 tumour outgrowth in WT mice treated as in Fig. 4 h ( , represent two experiments). b, 1956 tumour growth in C57BL6 WT mice treated as in Fig. 5i( , represent two experiments). Data in and are expressed as mean s.e.m., and values were calculated using Two-way ANOVA corrected with sídák’s multiple comparisons test. c, representative
flow cytometry plot showing the expression of mLama4+ specific TILs in T3 TILs harvested from treated mice, as shown in Fig. 5h. d-f, F244 tumourbearing mice were treated as in Fig. 5 h , and , frequency, , numbers of mPex14specific CD8 TILs, and , the ratio of mPex14-specific CD8 TILs to Tr1-like cells were assessed on day 13 TILs. Data are expressed as mean s.e.m., and values were calculated using one-way ANOVA with multiple comparisons corrected with Tukey’s method.
Extended Data Fig. 9 | related to Fig. 5: HDVax-induced Tr1-like cells kill cDC1 in a GZMB-dependent manner. a, IL-2 production by mItgb1-specific hybridoma cells after incubation with cDC1 and cDC2 sorted from five mice one day after treatment with T3- LDVax or -HDVax ( and represent two experiments).b, T3 tumour-bearing mice were vaccinated with LDVax or HDVax ( 2 doses, 11 days apart), serum was collected 7 days after the last vaccination, and antibody titer to mItgb1 SLP was assessed using ELISA (Methods). c, different isotypes of mItgb1-specific antibodies were determined. Data in and are shown as means s.e.m. and represent three independent experiments ( ). d, frequency of perforin (Prf1) expressing CD4 TILs in different subpopulations of CD4 TILs ( , represent two experiments). e, frequency of NIR expression (live/dead signal) in cDC1 ( ) (left) and cDC2 ( ) (right) after incubation with sorted LDVax-induced or subpopulations of HDVax-induced mItgb1-specific CD4 T cells (Methods). Data are shown as means s.e.m. of three experiments.f, frequency of cDC2
( CD172a ) expressing NIR after incubation with sorted subpopulations of HDVax-induced mItgb1 specific CD4 T cells harvested from WT mice (treated with HDVax plus control mAb or LILRB4) or GZMB mice (related to Fig. 5b). g, frequency (left) and numbers (right) of cDC1 (of CD45 cells) in F244 TILs treated with HDVax five days post vaccinations ( ). Data are representative of two independent experiments and shown as mean s.e.m. h, frequency (left) and numbers (right) of cDC1 in 1956 TILs assessed five days post-vaccination in WT and LILRB4 mice ( ). i, frequency of cDC1 in T3 tumours assessed five days post-vaccination and treatment with 2 doses of PD-1 or CTLA-4 in WT mice ( , represent two experiments). , frequency of cDC1 in 1956 TILs assessed five days post-vaccination in WT and GZMB mice ( ). Data in all panels are shown as means s.e.m. and values were calculated using one-way ANOVA with multiple comparisons corrected with Tukey’s method.
Extended Data Fig. 10 | related to Fig. 5: HDVax-induced Tr1-like cells arise from preferential cDC2 antigen presentation. a, representative flow cytometry plots showing the gating strategy for different antigen-presenting cell subsets in WT and mice bearing 1956 tumours. b, frequency of Tr1-like cells in spleens of HDVax treated WT, (cDC2 deficient) or Irf8 (cDC1 deficient) mice ( , represent two experiments). Data are
shown as means s.e.m. and values were calculated using one-way ANOVA with multiple comparisons corrected with Tukey’s method. c, frequency of Tregs and d, Tr1-like cells in 1956 TIL harvested at multiple days post tumour inoculation ( , representing two experiments). Data are expressed as means s.e.m. and values were calculated using Two-way ANOVA corrected with sídák’s multiple comparisons test.
natureportfolio
Corresponding author(s):
Robert Schreiber
Last updated by author(s):
May 22, 2024
Reporting Summary
Nature Portfolio wishes to improve the reproducibility of the work that we publish. This form provides structure for consistency and transparency in reporting. For further information on Nature Portfolio policies, see our Editorial Policies and the Editorial Policy Checklist.
Statistics
For all statistical analyses, confirm that the following items are present in the figure legend, table legend, main text, or Methods section.
Confirmed X The exact sample size ( ) for each experimental group/condition, given as a discrete number and unit of measurement X A statement on whether measurements were taken from distinct samples or whether the same sample was measured repeatedly The statistical test(s) used AND whether they are one- or two-sided Only common tests should be described solely by name; describe more complex techniques in the Methods section. A description of all covariates tested A description of any assumptions or corrections, such as tests of normality and adjustment for multiple comparisons
A full description of the statistical parameters including central tendency (e.g. means) or other basic estimates (e.g. regression coefficient) AND variation (e.g. standard deviation) or associated estimates of uncertainty (e.g. confidence intervals) X
For null hypothesis testing, the test statistic (e.g. ) with confidence intervals, effect sizes, degrees of freedom and value noted Give values as exact values whenever suitable.
X For Bayesian analysis, information on the choice of priors and Markov chain Monte Carlo settings
X For hierarchical and complex designs, identification of the appropriate level for tests and full reporting of outcomes
X Estimates of effect sizes (e.g. Cohen’s , Pearson’s ), indicating how they were calculated
Our web collection on statistics for biologists contains articles on many of the points above.
Software and code
Policy information about availability of computer code
Data collection
Flow cytomtery data was collected using BD FACSDIVA software V9.1. MultiPlex assay was performed using Flex MAP 3D. ELISPOT was collecte using a CTL Immunospot S6 universal machine and software.
Data analysis
All statistical analysis was performed using GraphPad Prism software version 10.2.2. cell ranger v6.1.1 was used for scRNAseq
For manuscripts utilizing custom algorithms or software that are central to the research but not yet described in published literature, software must be made available to editors and reviewers. We strongly encourage code deposition in a community repository (e.g. GitHub). See the Nature Portfolio guidelines for submitting code & software for further information.
Data
Policy information about availability of data
All manuscripts must include a data availability statement. This statement should provide the following information, where applicable:
Accession codes, unique identifiers, or web links for publicly available datasets
A description of any restrictions on data availability
For clinical datasets or third party data, please ensure that the statement adheres to our policy
All data supporting the findings of this study are available within the paper, its Supplemental or Extended Data files, or Source files. Any additional information related to the study is available from the corresponding author upon request. All reagents are available through a Material Transfer Agreement.
Research involving human participants, their data, or biological material
Policy information about studies with human participants or human data. See also policy information about sex, gender (identity/presentation), and sexual orientation and race, ethnicity and racism.
Reporting on sex and gender
N/A
Reporting on race, ethnicity, or
N/A
other socially relevant
groupings
Population characteristics
N/A
Recruitment
N/A
Ethics oversight
N/A
Note that full information on the approval of the study protocol must also be provided in the manuscript.
Field-specific reporting
Please select the one below that is the best fit for your research. If you are not sure, read the appropriate sections before making your selection.
Life sciences
Behavioural & social sciences
Ecological, evolutionary & environmental sciences
For a reference copy of the document with all sections, see nature.com/documents/nr-reporting-summary-flat.pdf
Life sciences study design
All studies must disclose on these points even when the disclosure is negative.
Sample size
Sample sizes were determined based on 25 years experience with these types of tumors in general, and over 10 years with T3 and F244 cell lines. (Gubin, et al 2014, Alspach, 2019)
Data exclusions
No data were excluded
Replication
All experiments had multiple biological and/or technical replicates and are indicated the Figure legends.
Randomization
For animal studies, mice were randomly assigned from large batches obtained from the vendor to different experimental groups in an agematched distribution.
Blinding
Tumor injection was performed by one person. Treatments and tumor measurement were performed by independent people. No blinding was performed in vitro studies.
Reporting for specific materials, systems and methods
We require information from authors about some types of materials, experimental systems and methods used in many studies. Here, indicate whether each material, system or method listed is relevant to your study. If you are not sure if a list item applies to your research, read the appropriate section before selecting a response.
eBioSciences: FOXP3 (Fjk-16a; 1:50; cat# 11-5773-82) and SEMA4a (5E3; 1:50; cat# 46-9753-41)]. Foxp3/Transcription factor Staining kit (ThermoFischer; cat# 00-5523-00) was used to stain Foxp3 and other intracellular proteins according to the manufacturing protocol.
Validation
All primary antibodies were validated against indicated proteins by the manufacturer per their associated Data Sheets and are included on their websites.
Eukaryotic cell lines
Policy information about cell lines and Sex and Gender in Research
Cell line source(s)
T3, F244 and 1956 cell lines were T3, F244 and 1956 cell lines were generated via subcutaneous injection of MCAnerated via subcutaneous injection of MCA. HEK293 cell line were obtained from ATCC (CRL-1573).
Authentication
Cell lines were authenticated by whole exosome sequencing and expression of certain genes
Mycoplasma contamination
All cell lines are routinely tested each month and were negative for mycoplasma.
Commonly misidentified lines (See ICLAC register)
This study did not involve any commonly misidentified cell lines.
Animals and other research organisms
Policy information about studies involving animals; ARRIVE guidelines recommended for reporting animal research, and Sex and Gender in Research
Laboratory animals
Six-8-week-old female C57BL/6J mice (Cat # 000664, Jackson Laboratory), GZMB KO, LILRB4 KO, Rag2-deficient and wild type male 129S6 mice were used for experiments. Mice were housed in groups of 3 to 5 . Photoperiod on: 12 hr off dark/light cycle. Ambient animal room temperature is , controlled within and room humidity is , controlled within .
Wild animals
No wild animals were used in this study.
Reporting on sex
Both male mice (129S6) were used for T3 and F244 tumors and female mice (C57BL6) were used for 1956 tumor line.
Field-collected samples
No field collected samples were used in this study.
Ethics oversight
All animal studies were performed with the approval of the Association for the Accreditation of Laboratory Animal Careaccredited Animal Studies Committee of Washington University in St. Louis
Note that full information on the approval of the study protocol must also be provided in the manuscript.
Plants
Seed stocks
N/A
Novel plant genotypes
N/A
Authentication
N/A
Flow Cytometry
Plots
Confirm that:
The axis labels state the marker and fluorochrome used (e.g. CD4-FITC).
The axis scales are clearly visible. Include numbers along axes only for bottom left plot of group (a ‘group’ is an analysis of identical markers).
All plots are contour plots with outliers or pseudocolor plots.
A numerical value for number of cells or percentage (with statistics) is provided.
Methodology
Sample preparation
Tumours were harvested, manually disassociated into small pieces and digested with collagenase 1A for 45 minutes at 37 c. Single cell suspension was then filter through 70 um cell strainer. Subsequently, Red blood cells were lysed. Suspension were filtered again through 40 um cell strainer and resuspended into a complete RPMI media. For intracellular stains, cells were fixed and permeabilized using the BD Foxp3 staining kit
Instrument
Software
Cell population abundance
Gating strategy
BD FACS Symphony 3, LSRFortessaX20 were used to collect the data. ARIA-II and/or Bigfoot were used for sorting.
BD FACSDIVA software V9.1 was used for collecting the flow cytometry data and for sorting, Data were analyzed using FlowJo software version 10.10.
Cell abundance was shown through out gating strategies
Live Lymphocytes gated using CD90.2 and CD4. Antigen-specific cells were gated using the appropriate tetramer,Tregs were excluded using CD25 and Foxp3 staining. Expressions of CD39, CD200 CD153, LILRB4, GZMB or CCL5 were measured.
Tick this box to confirm that a figure exemplifying the gating strategy is provided in the Supplementary Information.
Department of Pathology and Immunology, Washington University School of Medicine, St. Louis, MO, USA. The Andrew M. and Jane M. Bursky Center for Human Immunology and Immunotherapy Programs, Washington University School of Medicine, St. Louis, MO, USA. Department of Medicine, Division of Oncology, Washington University School of Medicine, St. Louis, MO, USA. Department of Immunology, University of Texas MD Anderson Cancer Center, Houston, TX, USA. Almazov National Medical Research Centre, St. Petersburg, Russia. Oncovir Inc., Washington, DC, USA. Natera Inc., Austin, TX, USA. Asher Biotherapeutics, South San Francisco, CA, USA. Netherlands Cancer Institute, Oncode Institute, Amsterdam, Leiden University, Leiden, Netherlands. The Parker Institute for Cancer Immunotherapy, San Francisco, CA, USA. e-mail: rdschreiber@wustl.edu
Awwad, M. & North, R. J. Immunologically mediated regression of a murine lymphoma after treatment with anti-L3T4 antibody. A consequence of removing L3T4 suppressor T cells from a host generating predominantly Lyt-2+ T cell-mediated immunity. J. Exp. Med. 168, 2193-2206 (1988).
Hori, S., Nomura, T. & Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 299, 1057-1061 (2003).
Fontenot, J. D., Gavin, M. A. & Rudensky, A. Y. Foxp3 programs the development and function of regulatory T cells. Nat. Immunol. 4, 330-336 (2003).