تحتوي الآليات التي تتوسطها الميكروبات على أساس تراكم الكربون في التربة من خلال الزراعة المستدامة تحت ارتفاع درجات الحرارة لعقد من الزمن Microbially mediated mechanisms underlie soil carbon accrual by conservation agriculture under decade-long warming
قد يعيق زيادة الكربون العضوي في التربة (SOC) في الأراضي الزراعية من خلال التحول من الإدارة التقليدية إلى الإدارة المستدامة بسبب تحفيز التحلل الميكروبي تحت ارتفاع درجات الحرارة. هنا، نختبر التأثيرات التفاعلية للإدارة الزراعية وارتفاع درجات الحرارة على استمرارية SOC والآليات الميكروبية الأساسية في تجربة محكومة استمرت لعقد من الزمن على نظام زراعة القمح والذرة. زادت درجات الحرارة من محتوى SOC وسرعت من دوران المجتمع الفطري الزمني تحت الزراعة المستدامة (بدون حراثة، بقايا المحاصيل المقطعة)، ولكن ليس تحت الزراعة التقليدية (حراثة سنوية، إزالة بقايا المحاصيل). زادت كفاءة استخدام الكربون الميكروبي (CUE) والنمو بشكل خطي مع مرور الوقت، مع تأثيرات إيجابية أقوى لارتفاع درجات الحرارة بعد 5 سنوات تحت الزراعة المستدامة. وفقًا لنماذج المعادلات الهيكلية، نشأت هذه الزيادات من مدخلات الكربون الأكبر من المحاصيل، التي تحكمت بشكل غير مباشر في CUE الميكروبي عبر التغيرات في المجتمعات الفطرية. نتيجة لذلك، زادت الكتلة الميتة الفطرية من 28 إلى 53%، مما برز كأقوى مؤشر لمحتوى SOC. بشكل جماعي، تظهر نتائجنا كيف يمكن أن تتفاعل الإدارة والعوامل المناخية لتغيير تركيب المجتمع الميكروبي، وعلم وظائف الأعضاء والوظائف، وبالتالي، تشكيل وتراكم SOC في الأراضي الزراعية.
يعتبر الكربون العضوي في التربة (SOC) خاصية من خصائص النظام البيئي، حيث يقدم مجموعة واسعة من الفوائد لكل من المجتمع البشري والبيئة الطبيعية، بما في ذلك تنظيم المناخ العالمي. ومع ذلك، فإن حالة الزراعة، التي تغطي 38% من سطح الأرض، هي مسألة
قلق. لقد عانت مساحات كبيرة من الأراضي الزراعية من تدهور معتدل إلى شديد بسبب ممارسات الإدارة غير المناسبة, مما أدى إلى تقليل نصف إلى ثلثي إجمالي محتوى SOC مقارنة بالتربة الطبيعية أو غير المزروعة. معالجة هذه
المشكلة، هناك إمكانات كبيرة لتراكم SOC في الأراضي الزراعية من خلال اعتماد ممارسات الإدارة ‘الذكية مناخيًا’, والتي تعتبر حلاً طبيعياً رئيسياً للتخفيف من تغير المناخ وضمان الأمن الغذائي، وبالتالي تحقيق أهداف التنمية المستدامة. ومع ذلك، من الضروري الاعتراف بأن SOC عرضة للفقد بسبب تغير المناخ. لقد قدمت تجارب تسخين التربة ومجموعات البيانات العالمية أدلة على زيادة معدلات التحلل تحت ارتفاع درجات الحرارة. للأسف، الدراسات التي تحقق في التأثيرات التفاعلية للإدارة وارتفاع درجات الحرارة على تراكم SOC في الأراضي الزراعية نادرة للغاية. فهم التفاعلات بين الإدارة وارتفاع درجات الحرارة أمر حاسم لتحديد ممارسات الإدارة المناسبة التي تحتفظ بـ SOC وتزيده في مواجهة المناخ المتغير، ولتطوير استراتيجيات فعالة تزيد من مرونة الزراعة كعنصر حيوي من ‘الزراعة الذكية مناخيًا’.
لقد كانت الاضطرابات في التربة الناتجة عن الحراثة دافعًا رئيسيًا لفقدان SOC التاريخي، والذي يقدر بـ C في السنة على مستوى العالم. بشكل عام، يتراكم SOC بعد التحول من الحراثة المكثفة إلى الزراعة المستدامة. لقد تم اقتراح الزراعة المستدامة، التي تمثل عادةً من خلال الاحتفاظ ببقايا المحاصيل، وبدون حراثة أو حراثة مخفضة, كخيار مناسب لإعادة بناء مستويات SOC، والتي توفر أيضًا فوائد متنوعة لوظائف النظام البيئي المتعددة. يُقدّر أن من الأراضي الزراعية العالمية قد تم إدارتها بواسطة أساليب الزراعة المستدامة. ومع ذلك، يمكن أن تختلف استجابة SOC للتغيرات في ممارسات الحراثة بشكل كبير عبر مناطق مختلفة وعلى مر الزمن. نظرًا لأن درجات الحرارة المتزايدة من المتوقع أن تحفز التنفس الميكروبي، فإن التربة التي تحتوي على محتوى أعلى من الكربون العضوي تكون أكثر عرضة لفقدان الكربون تحت ظروف ارتفاع درجات الحرارة. يجب أن تحتوي التربة المدارة تحت الزراعة المستدامة، والتي تشمل زيادة الاحتفاظ ببقايا العضوية وتقليل أو عدم الحراثة, على مزيد من SOC. قد تحتوي أيضًا على نسب أكبر من تجمعات المواد العضوية الكيميائية المقاومة نسبيًا (مثل، الكتل الكبيرة أو المواد العضوية الجزيئية)، والتي تكون أكثر عرضة للفقد تحت ارتفاع درجات الحرارة. ومع ذلك، لم تكشف دراسة حضانة مختبرية قصيرة الأجل لمدة 3 أشهر عن أي فرق كبير في تمييع SOC بين الزراعة المستدامة والزراعة التقليدية تحت ظروف درجات حرارة مختلفة. المعلومات المتعلقة بحساسية واستمرارية SOC المعاد بناؤه من خلال إدارة الزراعة المستدامة تجاه ارتفاع درجات الحرارة على المدى الطويل محدودة جدًا.
تنبؤ التغيرات في SOC يعتمد بشكل كبير على تكيف الميكروبات، والذي يتضمن تعديلات فسيولوجية تعدل كفاءة استخدام الكربون الميكروبي (CUE) أي، النسبة المئوية من الكربون المخصص للنمو مقارنة بالخسائر التنفسية. إن CUE الميكروبي له آثار قوية على تخزين SOC في التربة، حيث يلعب الكربون العضوي المحول إلى الكتلة الحيوية الميكروبية والكتلة الميتة دورًا حاسمًا في استقرار SOC على المدى الطويل. تعتمد CUE الميكروبي على كل من العوامل غير الحية والحيوية, والتي تتأثر بارتفاع درجات الحرارة وممارسات الإدارة. يمكن أن يكون لارتفاع درجة الحرارة تأثيرات مباشرة وغير مباشرة على CUE الميكروبي. بشكل عام، يؤدي ارتفاع درجات الحرارة إلى تقليل CUE الميكروبي، حيث يتم إعادة تخصيص نسبة أكبر من الركيزة من النمو إلى الأيض الصيانة, مما يغير معدلات العمليات المدفوعة بالإنزيمات. يمكن أن يغير ارتفاع درجات الحرارة CUE بشكل غير مباشر عبر التغيرات في رطوبة التربة، وتوافر الركيزة أو تركيب و/أو بنية المجتمعات الميكروبية. قد يقلل ارتفاع درجات الحرارة من رطوبة التربة ويقلل من CUE الميكروبي لأن المزيد من الركيزة يتم تخصيصه للأيض غير المتجانس، وبالتالي أقل متاح للنمو. على العكس، قد يعزز ارتفاع درجات الحرارة نمو النبات، والإنتاجية ومدخلات الكربون في منطقة الجذور, مما يعزز نمو الميكروبات وتراكم الكتلة الميتة. يمكن أن تعدل المجموعات الميكروبية السائدة التي تتأثر بارتفاع درجات الحرارة CUE المجتمعي وبالتالي. يمكن أن يحول المحللون الميكروبيون الذين لديهم CUE أعلى الركائز إلى كتلة حيوية جديدة بشكل أكثر كفاءة من خلال زيادة النمو وتقليل التنفس لكل وحدة كربون تم امتصاصها بواسطة الميكروبات. وبالتالي، فإن زيادة CUE تنتج المزيد من الكتلة الميكروبية
والمنتجات الثانوية، أي، ‘الكتلة الميتة’، وكلاهما يساهم في تجمع SOC المستمر. عمومًا، تمتلك الفطريات CUE أعلى من البكتيريا, ولكن يمكن أن تتأثر سلبًا بالحراثة. قد تزيد الحساسية الأكبر للفطريات تجاه ارتفاع درجات الحرارة مقارنة بالبكتيريا من مساهمات بقايا الفطريات في الكتلة الميتة. ومع ذلك، قد تنخفض CUE إذا خصصت الفطريات الموارد الأيضية لزيادة إنتاج الإنزيمات الخارجية لاكتساب المغذيات تحت ارتفاع درجات الحرارة أو نقص المغذيات. يمكن أن يؤدي الاحتفاظ ببقايا المحاصيل في الزراعة المستدامة إلى زيادة CUE الميكروبي وتراكم الكتلة الميتة من خلال تخفيف نقص المغذيات، والحفاظ على رطوبة التربة، وتقليل مسارات تسرب الطاقة وإنتاج الإنزيمات الخارجية. ومع ذلك، فإن فهمنا للروابط المعقدة بين الخصائص الفسيولوجية والتمثيلية الميكروبية وتراكم SOC استجابةً للإدارة وارتفاع درجات الحرارة بدائي، خاصة عند النظر في مقاييس زمنية مختلفة.
هنا، نقدم الدراسة من تجربة زراعية طويلة الأجل تمتد لعقد من الزمن وتشمل نظامين إداريين متميزين (الزراعة المستدامة مقابل الزراعة التقليدية)مستويين من الاحترار (الاحترار مقابل البيئة المحيطة). كانت أهدافنا الرئيسية توضيح التأثيرات التفاعلية للاحتباس الحراري والإدارة على تراكم الكربون العضوي في التربة واستمراريته، بالإضافة إلى تحديد الخصائص الميكروبية الزمنية التي تدعم استجابتها على مدى 10 سنوات. على وجه التحديد، كان هدف دراستنا: (i) تقييم ما إذا كان الاحترار يؤثر بشكل مختلف على تراكم الكربون العضوي في الزراعة المستدامة (إعادة بقايا المحاصيل المقطوعة وعدم الحراثة) مقابل الزراعة التقليدية (إزالة بقايا المحاصيل والحراثة السنوية)؛ و (ii) دراسة التأثيرات التفاعلية للاحتباس الحراري والإدارة على تسلسل المجتمعات الميكروبية والتغيرات الزمنية في الصفات الفسيولوجية الميكروبية (مثل نمو الميكروبات، كفاءة استخدام الكربون، كربون الكتلة الميكروبية الميتة، والجينات الوظيفية الأيضية الميكروبية) وتقييم عواقبها على تشكيل وتراكم الكربون العضوي في التربة. افترضنا أنه، في ظل ظروف الاحترار المناخي، (i) تزيد الزراعة المستدامة من الكربون العضوي مباشرة من خلال زيادة المدخلات الكربونية المشتقة من النباتات وغير مباشرة من خلال زيادة توفر الركيزة لمجتمع الميكروبات في التربة؛ (ii) التكيف على مستوى المجتمع الميكروبي مع الاحترار وزيادة كفاءة نمو الميكروبات والوظائف الأيضية استجابة لتوفر ركيزة أكبر يزيد من مساهمة الكتلة الميكروبية الميتة في الكربون العضوي مع مرور الوقت تحت الزراعة المستدامة؛ (iii) زيادة الحجم وتسارع دوران مجتمع الفطريات يعزز تراكم الكتلة الميكروبية الميتة، مما يعزز تشكيل واستمرارية الكربون العضوي في التربة مع مرور الوقت تحت الزراعة المستدامة. لاختبار هذه الفرضيات، قمنا بقياس الكربون العضوي، والمجتمعات البكتيرية والفطرية باستخدام تسلسل الحمض النووي، والوظائف الميكروبية باستخدام الميتاجينوميات، والصفات الفسيولوجية الميكروبية (كفاءة استخدام الكربون والكتلة الميكروبية الميتة) باستخدام الركيزة المستقلة.تحليل العلامات الحيوية الميكروبية (السكريات الأمينية). في دراستنا، زادت درجة حرارة الأرض على المدى الطويل من الكربون العضوي في التربة تحت الزراعة المحافظة، ولكن ليس تحت الزراعة التقليدية. كانت هذه الاستجابة تحت الزراعة المحافظة مرتبطة بزيادة في الصفات الفسيولوجية الميكروبية الرئيسية، وكفاءة استخدام الكربون، والنمو وكتلة الفطريات الميتة، مع تسارع في دوران المجتمع الفطري وتباينه على مدى 10 سنوات. باستخدام تحليل نماذج المعادلات الهيكلية، نجد أن زيادة المدخلات الكربونية من المحاصيل قد سرعت من الخلافة الفطرية وعززت كفاءات نمو الميكروبات، مما أدى إلى زيادة تدريجية في مساهمات الكربون الميكروبي في تكوين وتراكم الكربون العضوي في التربة على مدى عقود تحت الزراعة المحافظة مع ارتفاع درجة الحرارة. تظهر أعمالنا أن إدارة الزراعة والعوامل المناخية يمكن أن تتفاعل لتغيير تركيب المجتمع الميكروبي، وفسيولوجيته، ووظائفه، وبالتالي، تكوين وتراكم الكربون العضوي في الأراضي الزراعية.
النتائج
زراعة الحفظ زادت من الكربون العضوي في التربة من خلال التوسط في تأثير الاحترار على خصائص التربة والنبات.
تم فرض تسخين حقول التجارب تحت الزراعة المحافظة أو التقليدية تجريبيًا لمدة 10 سنوات باستخدام الأشعة تحت الحمراء.
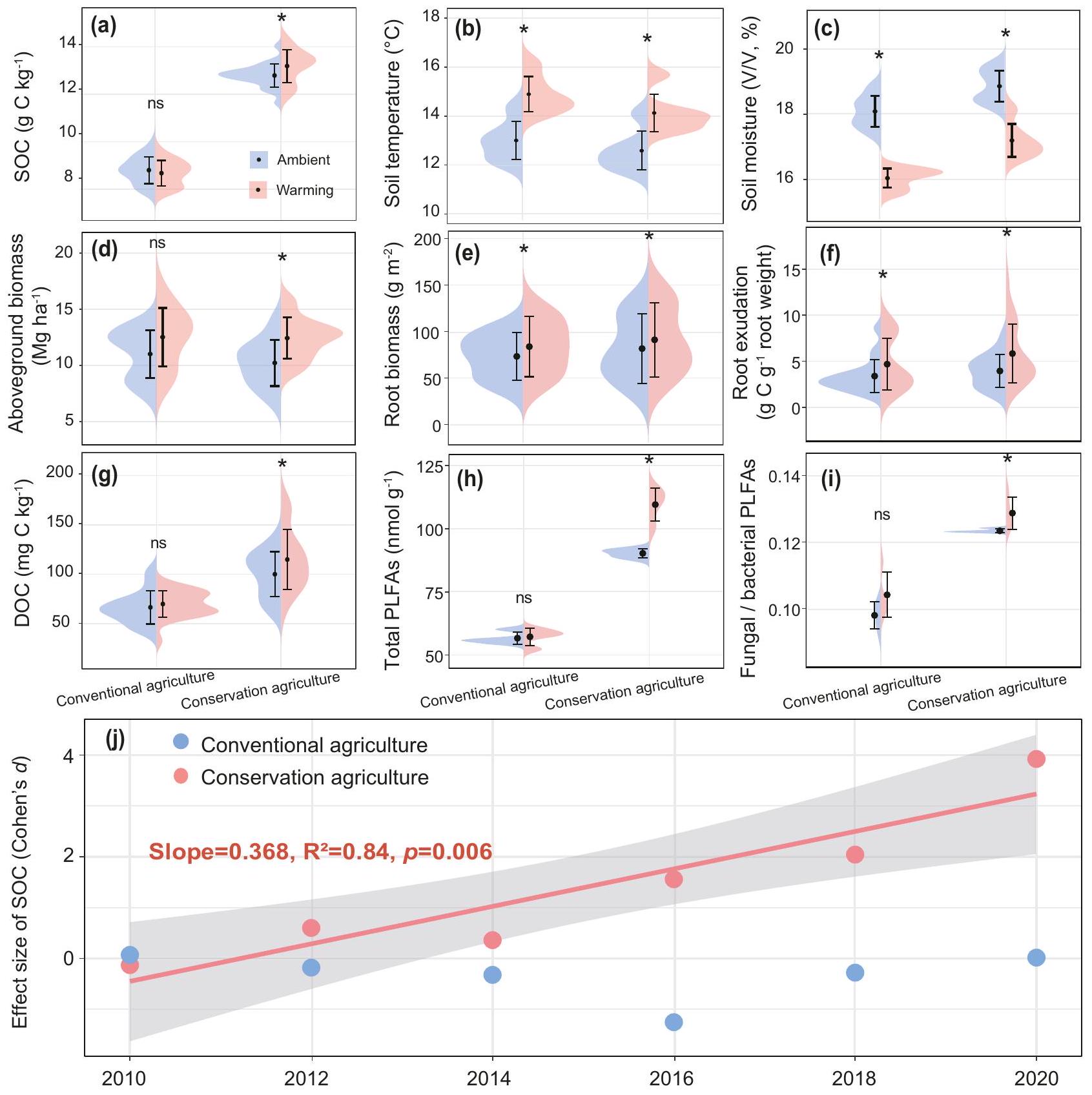
الشكل 1 | تأثير الاحترار على متغيرات النبات والتربة على مدى عشر سنوات اعتمادًا على أنظمة الإدارة. أ-ي متوسط محتوى الكربون العضوي في التربة، ودرجة حرارة التربة والرطوبة، والكتلة الحيوية فوق الأرض، وكتلة الجذور، وإفراز الجذور من الكربون، والكربون العضوي الذائب والكتلة الحيوية الميكروبية. تُعرض البيانات كرسوم بيانية على شكل كمان مع القيم المتوسطة.س.م. تم تحليل بيانات الكتلة الحيوية للجذور ومدخلات الكربون الناتجة عن إفرازات الجذور وبيانات الكتلة الحيوية الميكروبية استنادًا إلى أخذ عينات التربة في عام 2020 (عينات تربة مستقلة لكل معالجة). كانت المعلمات الأخرى بيانات متوسطة على ستة تواريخ أخذ عينات على مدى 10 سنوات (عينات التربة المستقلة لكل معالجة). تم إجراء التحليل الإحصائي باستخدام تحليل التباين ANOVA للقياسات المتكررة. جميع التقاريرتنتج القيم من اختبارات إحصائية ثنائية الجانب مع; غير دالة. تشير النجوم إلى الفروق الدالة في تأثير الاحترار لنظام الإدارة الفردي مقارنةً مع شرط عدم التسخين المتطابق. للحصول على القيم الإحصائية الدقيقة، انظر الجداول التكميلية 2.تحول في حجم تأثير الاحترار على الكربون العضوي في التربة على مر الزمن (2010-2020) للزراعة المحافظة والزراعة التقليدية. تم استخدام نموذج الانحدار الخطي مع اختبار ثنائي الجانب للتحليل الإحصائي، وتم استخدام R-squared المعدل. يتم الإشارة إلى العلاقات بخطوط صلبة وإحصائيات الملاءمة (الميل، و القيم) لكل ممارسة إدارية. الخط المتصل يمثل الانحدار الخطي المعنوي ( )، والتظليل الرمادي يشير إلى فترات الثقة. الزراعة المحافظة-الباردة الزراعة المحافظة بدون تسخين، الزراعة المحافظة-الساخنة الزراعة المحافظة مع تسخين، الزراعة التقليدية-الباردة الزراعة التقليدية بدون تسخين، الزراعة التقليدية-الساخنة الزراعة التقليدية مع تسخين. سخانات، تحافظ على درجة حرارة التربة عند فوق مستويات البيئة المحيطة (الشكل التوضيحي 1). قمنا أولاً بتحديد اتجاه التغيرات الزمنية في محتوى الكربون العضوي في التربة (SOC) وحققنا فيما إذا كانت ظاهرة الاحترار تؤثر بشكل مختلف على محتوى SOC تحت الزراعة المحافظة مقابل الزراعة التقليدية. على مدار 10 سنوات، زادت إدارة الزراعة المحافظة من محتوى SOC مقارنة بإدارة الزراعة التقليدية، بغض النظر عن معالجة الاحترار. كما زاد الاحترار من محتوى SOC. محتوى تحت الزراعة المستدامة بنسبة 3.1% مقارنةً بمعاملة عدم التسخين، ولكن ليس تحت الزراعة التقليدية.“; الشكل 1أ؛ الجداول التكميلية 1 و 2). بالإضافة إلى التأثيرات الفردية للإدارة والاحترار، كانت هناك تأثيرات تفاعلية للإدارةالتدفئة والإدارةتسخينعام على محتوى SOC (؛ الجدول التكميلي 1)، مما يشير إلى أن التأثير التحفيزي الإيجابي للاحتباس الحراري على محتوى الكربون العضوي في التربة تم التوسط فيه بواسطة الإدارة وتأثيرها زاد مع مرور الوقت. قمنا أيضًا بتقييم تأثير الاحترار على محتوى الكربون العضوي في التربة على مدى 10 سنوات باستخدام مقياس كوهين. زاد محتوى الكربون العضوي في التربة بشكل خطي مع مدة الاحترار تحت الزراعة المستدامة، وتسارع هذا الزيادة بعد السنة الخامسة. ; الشكل 1j). بالمقابل، لم يكن لتأثير الاحترار على محتوى الكربون العضوي في التربة تحت الزراعة التقليدية دلالة إحصائية (الشكل 1j؛ الجدول التكميلي 2). لذلك، تدعم نتائجنا جزئيًا الفرضية (i)، مما يشير إلى أن الزراعة المستدامة تحت ظروف أكثر دفئًا تزيد من محتوى الكربون العضوي في التربة.
عالجت الزراعة المستدامة، التي تتميز بتغطية مستمرة للتربة بفضلات المحاصيل دون حراثة، زيادة في الكربون العضوي في التربة، مما خفف من آثار الاحترار التجريبي على درجة حرارة التربة ومحتويات الرطوبة. على مدار فترة الدراسة التي استمرت 10 سنوات، قمنا بمراقبة درجة حرارة التربة ورطوبتها بشكل مستمر باستخدام أجهزة استشعار في الحقل. لاحظنا أن كلا العاملين قد تأثرا بالإدارة والاحترار التجريبي. كما هو متوقع، زاد الاحترار التجريبي من درجة حرارة التربة ولكنه قلل من رطوبة التربة تحت كل من الزراعة المستدامة والزراعة التقليدية.؛ الشكل 1ب، ج؛ الجداول التكميلية 1 و 2). ومع ذلك، كانت تأثيرات الاحترار متأثرة بالإدارة. كانت التربة المغطاة بفضلات المحاصيل تحت الزراعة المستدامة أكثر برودة ( مقابل ; “; الشكل 1ب) وأكثر رطوبة ( مقابل ; الشكل 1ج) مقارنة بالزراعة التقليدية.
قمنا أيضًا بتقييم مدخلات الكربون من النباتات باستخدام قياسات الوزن الجاف للكتلة الحيوية فوق الأرض وكتلة الجذور، وكربون إفرازات الجذور باستخدام طريقة تم تطويرها لجمع إفرازات الجذور في الموقع.زاد الاحترار من الكتلة الحيوية فوق الأرض تحت الزراعة المستدامة، ولكن ليس تحت الزراعة التقليدية.; الشكل 1d؛ الجدول التكميلي 2)، ربما بسبب التأثيرات الإيجابية للاحتفاظ بالمخلفات وعدم الحراثة على رطوبة التربة. زادت الاحترار من مدخلات الكربون النباتية تحت الأرض، بما في ذلك كتلة الجذور وكربون إفرازات الجذور، في كل من الزراعة المحافظة والزراعة التقليدية (؛ الشكل 1د، هـ، و؛ الجدول التكميلي 2). بشكل عام، زادت إجمالي مدخلات الكربون من الجذور بـتحت الاحترار مقارنة بالتحكم البيئي تحت الزراعة المستدامة; الشكل التوضيحي الإضافي 2)، مما يساهم في زيادة الكربون العضوي في التربة (; الشكل 1أ؛ الجدول التكميلي 2) والكربون العضوي المذاب (DOC) (؛ الشكل 1g؛ الجدول التكميلي 2) التركيزات.
أدى الاحترار إلى تحفيز كفاءة نمو الميكروبات وتراكم الكربون من الكتلة الحيوية الفطرية تحت الزراعة المستدامة
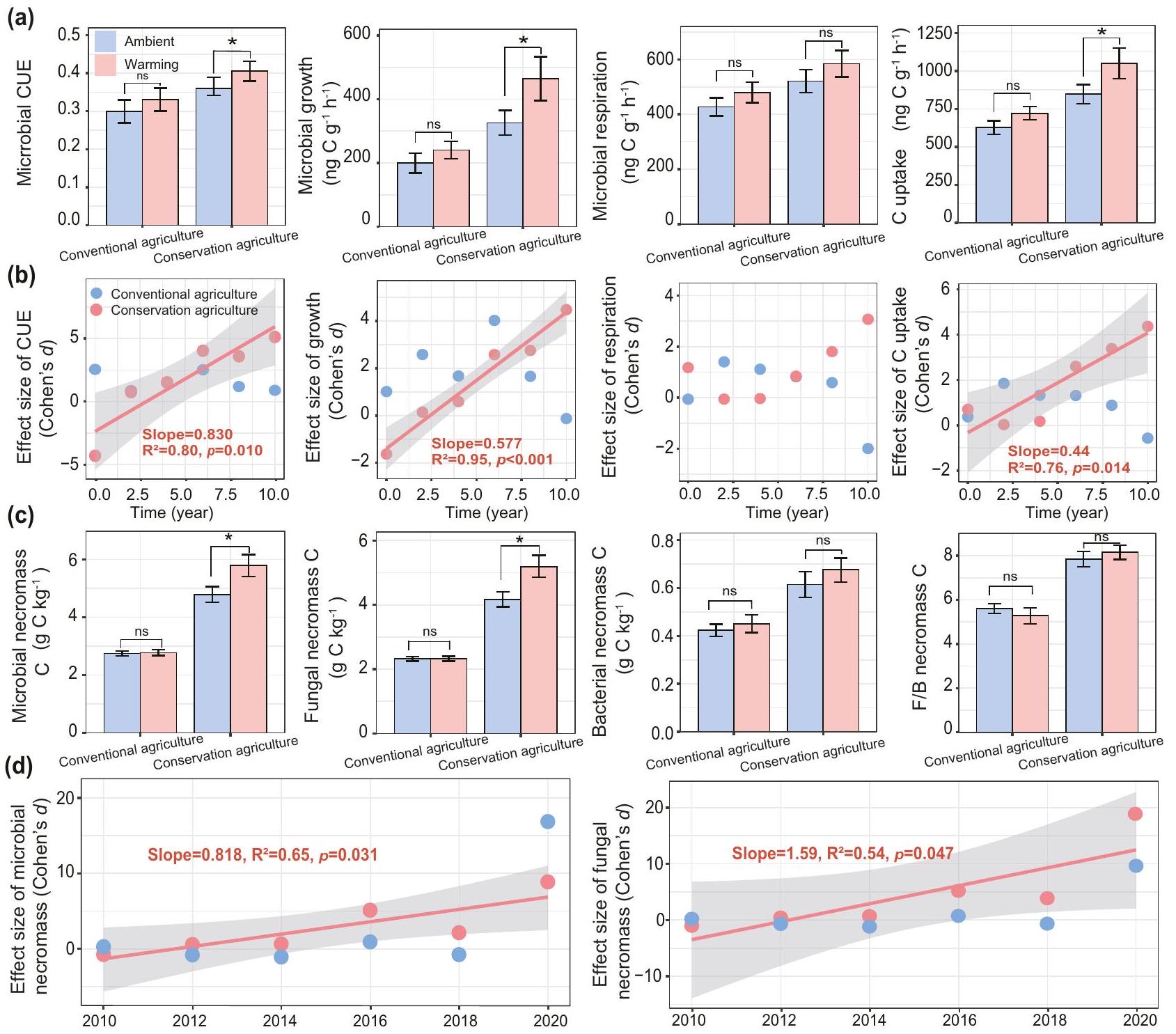
مثل محتوى SOC، قد تتغير آثار الإدارة والاحترار على كفاءة نمو الميكروبات على مدى 10 سنوات. لمعالجة ذلك، قمنا بقياس CUE لمجتمع الميكروبات في التربة باستخدام طريقة. على مدار جميع سنوات الدراسة، زاد الاحترار CUE الميكروبي بنسبة 12%، ونمو الميكروبات بـواستيعاب الكربون بـعند مقارنتها بالتحكم المحيط تحت الزراعة المستدامة (; الشكل 2أ؛ الجداول التكميلية 1 و 2)، مما يشير إلى تسريع دوران الميكروبات بما في ذلك التكاثر والنمو والموت. كانت التفاعلات بين الإدارة الاحترار والإدارة الاحترار السنة على CUE الميكروبي، والنمو، والتنفس واستيعاب الكربون (; الجدول التكميلية 1) تشير إلى أن آثار الاحترار كانت تعتمد على كل من الإدارة والوقت. قمنا بمزيد من الفحص للتغيرات الزمنية في تأثير الاحترار على كفاءة نمو الميكروبات تحت نظامي الإدارة على مدى فترة الدراسة التي استمرت 10 سنوات (الشكل 2ب). زادت آثار الاحترار على CUE الميكروبي تحت الزراعة المستدامة بشكل خطي مع مدة التجربة (الميل=0.83، ; الشكل 2ب).
لفهم هذه الآثار بشكل أفضل، قسمنا الدراسة إلى مراحل مبكرة (2010-2015) ومراحل لاحقة (2016-2020) لتقييم دور مدة الاحترار على اتجاهات الاستجابة وأحجامها (الشكل التكميلية 3). كان للاحتباس الحراري قصير وطويل الأجل آثار متناقضة على CUE تحت الزراعة المستدامة: لوحظ حجم تأثير سلبي لـ CUE خلال المرحلة المبكرة (2010-2015)، بينما كان حجم تأثير إيجابي واضح في المرحلة اللاحقة (2016-2020) (; الشكل التكميلية 3أ). وبالتالي، زاد الاحترار CUE الميكروبي بمقدار 1.1-1.5 مرة خاصة تحت الزراعة المستدامة خلال المرحلة اللاحقة (; الشكل التكميلية 3هـ). تبع تأثير الاحترار على نمو الميكروبات واستيعاب الكربون نمطًا مشابهًا لذلك الخاص بـ CUE، مع زيادة الفرق بين الاحترار وعدم الاحترار مع مرور الوقت في التربة تحت الزراعة المستدامة (الميل= 0.58 و 0.44، جميع ; الشكل 2ب؛ الشكل التكميلية 3ب، د). على العكس، لم تُلاحظ أي آثار تحت الزراعة التقليدية (الشكل 2ب؛ الشكل التكميلية 3). تشير الملاحظة بأن CUE الميكروبي، والنمو واستيعاب الكربون قد زادت في المرحلة اللاحقة من التجربة إلى توفر أكبر للركيزة، مما يتفق مع الفرضية (1) حيث زادت مدخلات الكربون النباتية ومحتوى SOC بشكل أسرع بعد السنة الخامسة (الشكل 1ي).
تم تأكيد تسريع دوران الميكروبات استجابةً للاحتباس الحراري تحت الزراعة المستدامة من خلال زيادة بنسبة 77% في إجمالي كربون النخر الميكروبي (المشار إليه بتركيز مؤشرات السكر الأميني ) مقارنة بالتربة غير المدفأة (; الشكل 2ج؛ الجداول التكميلية 2 و 3). رفع الاحترار مساهمة إجمالي النخر في SOC خلال المرحلة اللاحقة من التجربة (من 2016 إلى 2020؛ ; الشكل التكميلية 4)، مما يبرز الدور المهم لتأثير دفن مضخة الكربون الميكروبي في تشكيل SOC جديد. على وجه الخصوص، زاد كربون النخر الفطري بشكل خطي مع السنة تحت الزراعة المستدامة، مع كون حجم تأثير الاحترار أكبر بـ 19 مرة في 2020 مقارنة بـ 2010 (الميل =1.6، ; الشكل 2د؛ الجدول التكميلية 4). ومع ذلك، لم يكن هذا التأثير ذا دلالة تحت الزراعة التقليدية. لوحظ نمط زمني مشابه لكربون النخر الميكروبي الإجمالي (; الشكل 2د). كما زادت تركيز مؤشرات PLFA الميكروبية، التي تعمل كمؤشرات للكتلة الحيوية الميكروبية للمجموعات السائدة (الشكل 1ح؛ الجدول التكميلية 2)، مما يشير أيضًا إلى فوائد الإدارة المستدامة والاحترار للفطريات، كما يتضح من نسب PLFA الفطرية/البكتيرية الأكبر (الشكل 1ط؛ الجدول التكميلية 2). أدى حجم الكتلة الحيوية الفطرية الأكبر بالتالي إلى زيادة كربون النخر الفطري، مما ساهم بشكل كبير في إجمالي النخر مع مرور الوقت (; الشكل التكميلية 4) وزاد أيضًا بفعل الاحترار (بـ ; الشكل 2ج). وبالمثل، زاد الاحترار من مساهمة إجمالي النخر والنخر الفطري في SOC من إلى ، ومن إلى ، على التوالي، من 2010 إلى 2020 تحت الزراعة المستدامة (، الشكل التكميلية 4). تشير الزيادة في كفاءة نمو الميكروبات وإجمالي النخر تحت الزراعة المستدامة في التربة المدفأة، بالتوازي مع زيادة مدخلات الكربون النباتية وزيادة محتوى SOC، إلى أن المجتمع الميكروبي واستجاباته الفسيولوجية تكيفت مع ظروف الاحترار مع مرور الوقت، مما يدعم الفرضية (2).
سرع الاحترار دوران المجتمع الفطري الزمني وتباينه تحت الزراعة المستدامة
لفهم دور الميكروبيوم التربوي في دفع ديناميات SOC استجابةً للاحتباس الحراري والإدارة، قمنا بفحص كيف تباينت تنوع الميكروبات ودوران المجتمع على مدى 10 سنوات باستخدام تسلسل الحمض النووي. غيّر الاحترار كل من المجتمعات الميكروبية والميكروبات الفردية، اعتمادًا على الإدارة. تغيرت آثار الاحترار على الأنماط الفطرية مع السنة تحت الزراعة المستدامة (; الشكل التكميلية 5). في المرحلة اللاحقة من التجربة من 2016 إلى 2020، قلل الاحترار الأنماط الفطرية بنسبة 9.3-12% تحت الزراعة المستدامة، لكنه لم يكن له تأثير تحت الزراعة التقليدية (; الشكل التكميلية 5). تم تغيير تركيبات بكتيريا التربة والفطريات بواسطة الاحترار والإدارة والسنة، كما تم تصورها بواسطة تحليل المكونات الرئيسية (الشكل التكميلية 6؛ الجداول التكميلية 5). أكدت ثلاثة اختبارات إحصائية متعددة غير معلمية مكملة (Adonis،
الشكل 2 | آثار الاحترار على CUE الميكروبي في التربة، والنمو، والتنفس، واستيعاب الكربون والنخر على مدى عشر سنوات اعتمادًا على أنظمة الإدارة. أ متوسط CUE الميكروبي، والنمو، والتنفس واستيعاب الكربون في ستة تواريخ أخذ عينات على مدى 10 سنوات.تغيرات في حجم تأثير الاحترار على CUE الميكروبي، والنمو، والتنفس والاستيعاب مع مرور الوقت (2010-2020) للزراعة المستدامة والتقليدية.متوسط كربون النخر الميكروبي وتغير في آثار الاحترار على النخر مع مرور الوقت (2010-2020) اعتمادًا على أنظمة الإدارة. تشير الحروف إلى اختلافات ذات دلالة بين المدفأة والتحكم تحت نظامي الإدارة. تمثل الأعمدة المتوسط s.e.m. (عينات التربة المستقلة لكل معالجة). تم إجراء التحليل الإحصائي باستخدام ANOVA للقياسات المتكررة
التحليل. تشير الاختبارات الإحصائية ذات الجانبين إلى آثار ذات دلالة بواسطة ; ns، غير دالة. للحصول على القيم الإحصائية الدقيقة، انظر الجداول التكميلية 2 و 4. يتم الإشارة إلى العلاقات بخطوط صلبة وإحصائيات الملاءمة (الميل، والقيم) لكل ممارسة إدارية (تم عرض r -مربعة المعدلة والقيمة). يمثل الخط الصلب الانحدار الخطي الدال ()، ويشير التظليل الرمادي إلى فترات الثقة. Conserv-Amb الزراعة المستدامة بدون احترار، Conserv-Warm الزراعة المستدامة مع الاحترار، Conven-Amb الزراعة التقليدية بدون احترار، Conven-Warm الزراعة التقليدية مع الاحترار.
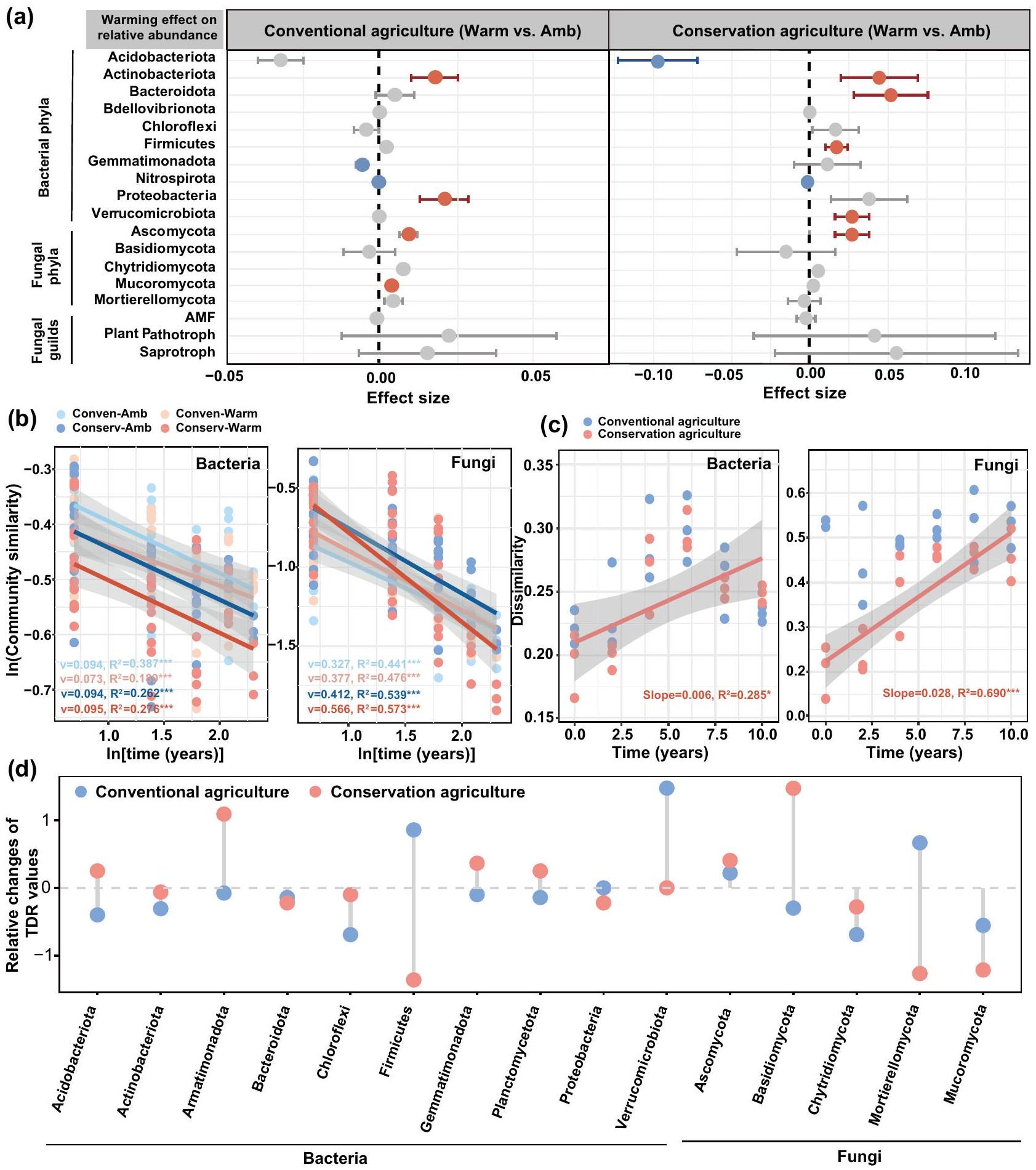
أكدت Anosim وMRPP أن المجتمعات البكتيرية والفطرية عبر جميع السنوات كانت مختلفة بشكل كبير بين المواقع المدفأة وغير المدفأة تحت كل من الزراعة المستدامة والتقليدية (; الجدول التكميلية 6). تحت الزراعة المستدامة، كان للاحتباس الحراري تأثير سلبي كبير على الوفرة النسبية لـ Acidobacteria (; الشكل 3أ)، بينما زاد من الوفرة النسبية لـ Actinobacteria وFirmicutes وBacteroidota وVerrucomicrobiota (إلى ; الشكل 3أ). زادت الوفرة النسبية لـ Proteobacteria، بما في ذلك Alphaproteobacteria، تحت الزراعة التقليدية مع الاحترار (; الشكل 3أ). كان للاحتباس الحراري تأثير إيجابي أكبر على الوفرة النسبية لـ Ascomycota تحت الزراعة المستدامة مقارنة بالزراعة التقليدية (مقابل ; الشكل 3أ). تحولت آثار الاحترار على الوفرة النسبية لـ Ascomycota من سلبية في المرحلة المبكرة (; الشكل التكميلية 7؛ من 2010 إلى 2015) إلى إيجابية في المرحلة اللاحقة (; من 2016 إلى 2020). قمنا بمزيد من التحقيق في مختلف النقابات الفطرية كما تم تصنيفها بواسطة FUNGuild. بينما زاد الاحترار بشكل طفيف من الوفرة النسبية لـ Saprotrophs تحت كلا نوعي الإدارة، كان التأثير أكثر وضوحًا تحت الزراعة المستدامة (مقابل 0.055؛ الشكل 3أ).
لتوضيح تأثيرات الاحترار على التغير الزمني في هيكل المجتمع الميكروبي، قمنا بتقييم علاقات تدهور الزمن (TDRs) للبكتيريا والفطريات. تمثل انحدارات قيم TDR معدلات التغير الزمني لمجتمعات الميكروبات في التربة. كانت معدلات التغير الزمني للفطريات أسرع من تلك الخاصة بالبكتيريا ( مقابل 0.72-0.97؛; الشكل 3ب). علاوة على ذلك، فإن TDR
الشكل 3 | الديناميات الزمنية للميكروبيوم التربة. أ أحجام التأثيرات للاحتباس الحراري على الوفرة النسبية لمجموعات الميكروبات الرئيسية بناءً على نماذج التأثيرات المختلطة الخطية. يتم تقديم البيانات كمتوسط s.e.m. لأحجام التأثير المقدرة. تعتمد الأهمية الإحصائية على اختبارات والد النوع الثاني X2 (ذو جانبين) ( عينات التربة المزدوجة لكل إدارة)؛ التغيرات غير المهمة تشير إليها النقاط الرمادية. علاقات تدهور الزمن لمجتمعات البكتيريا والفطريات لجميع العلاجات و(ج) التغيرات الزمنية في اختلافات المجتمع الميكروبي بين التربة المدفأة وتربة التحكم تحت الزراعة المحافظة والزراعة التقليدية. تم استخدام نموذج الانحدار الخطي مع اختبار ذو جانبين للتحليل الإحصائي، وتم استخدام R المعدل -مربع. يتم الإشارة إلى العلاقات بخطوط صلبة و
إحصائيات الملاءمة (الانحدارات، و القيم) لكل ممارسة إدارية. يمثل الخط الصلب الانحدار الخطي المهم ()، ويشير التظليل الرمادي إلى فترات الثقة 95%. د التغيرات النسبية الناتجة عن الاحترار في TDRs (v) بين السلالات التصنيفية تحت الزراعة المحافظة والزراعة التقليدية. يتم تقديم قيم TDR كتغيرات نسبية من (الاحترار-التحكم)/التحكم. Conserv-Amb: الزراعة المحافظة بدون احترار؛ Conserv-Warm: الزراعة المحافظة مع الاحترار؛ Conven-Amb: الزراعة التقليدية بدون احترار؛ Conven-Warm: الزراعة التقليدية مع الاحترار.
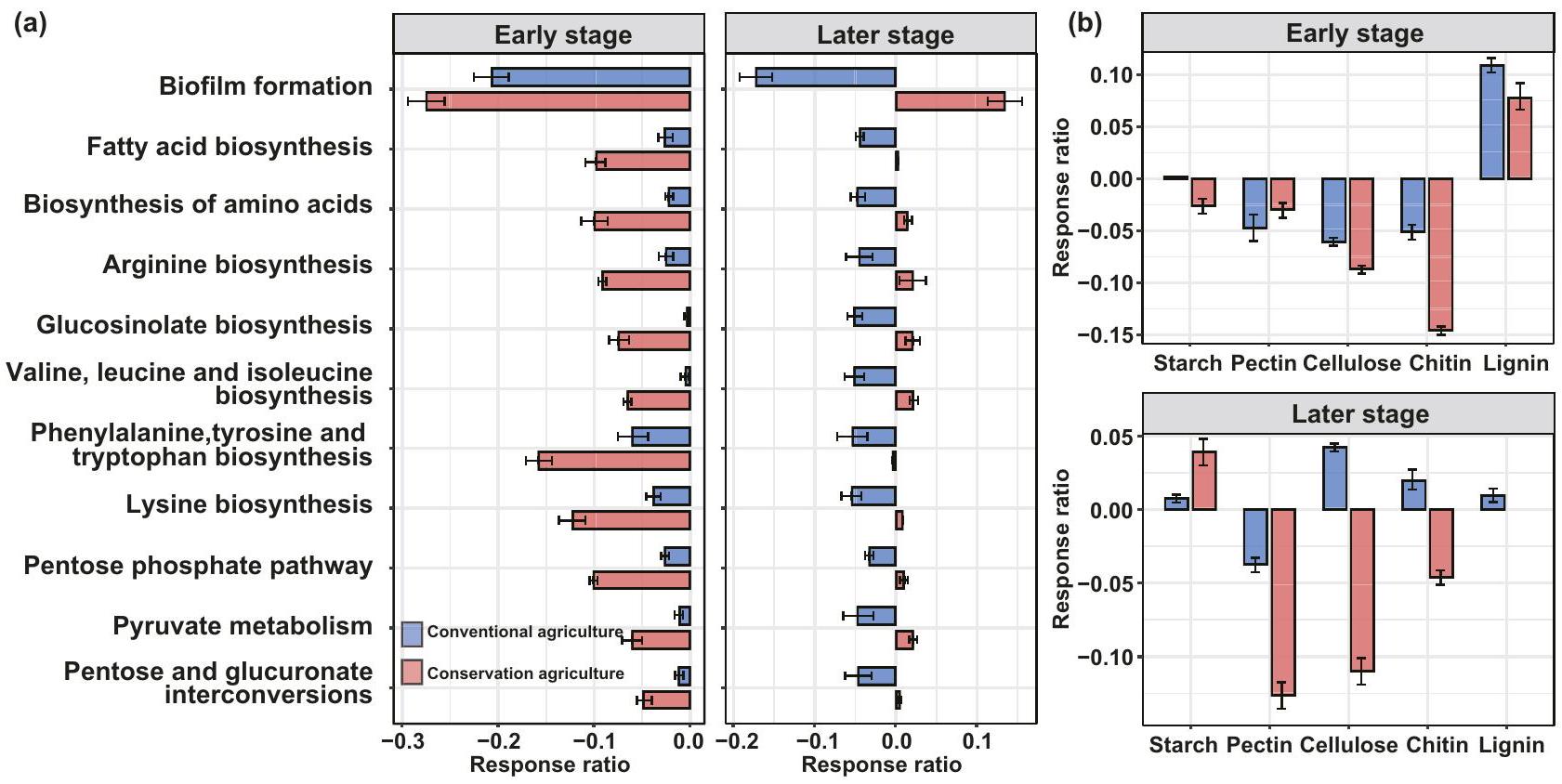
الشكل 4 | تأثير الاحترار على وظائف التمثيل الغذائي للكربون للميكروبات التربة اعتمادًا على الإدارة في المراحل المبكرة والمتأخرة. أ تأثير الاحترار على المسارات المتعلقة بنمو الميكروبات تحت نظامين إداريين بناءً على قاعدة بيانات KEGG. تم تقديم المسارات التي تغيرت بشكل كبير فقط بين التربة المدفأة وتربة التحكم تحت كل إدارة هنا. تأثير الاحترار على الوفرة النسبية لجينات التمثيل الغذائي للكربوهيدرات تحت نظامين
إداريين بناءً على الإنزيمات النشطة للكربوهيدرات (CAZy). تم تقديم الجينات التي تغيرت بشكل كبير فقط هنا. يتم تقديم البيانات كمتوسط s.e.m. لأحجام التأثير المقدرة ( عينات التربة المزدوجة لكل إدارة). ConservAmb: الزراعة المحافظة بدون احترار؛ Conserv-Warm: الزراعة المحافظة مع الاحترار؛ Conven-Amb: الزراعة التقليدية بدون احترار؛ Conven-Warm: الزراعة التقليدية مع الاحترار.
كانت الانحدارات أكثر حدة استجابةً للاحتباس الحراري تحت الزراعة المحافظة (اختبارات التبديل الزوجي؛; الشكل 3ب، الجدول التكميلي 4). لفحص تأثيرات الاحترار على تسلسل مجتمعات البكتيريا والفطريات بمرور الوقت، قمنا بتقييم الاختلافات في المجتمعات الميكروبية بين القطع المدفأة وغير المدفأة تحت هذين النظامين الإداريين على أساس سنوي. زادت الاختلافات في هياكل مجتمعات البكتيريا والفطريات بين الاحترار والتحكم البيئي بشكل خطي مع الوقت، ولكن فقط تحت الزراعة المحافظة (; الشكل 3ج). بالإضافة إلى ذلك، كانت الاختلافات المزدوجة في المجتمعات الميكروبية بين القطع المدفأة وقطع التحكم تحت الزراعة المحافظة أكثر وضوحًا بالنسبة للفطريات (الانحدار = 0.006؛ الشكل 3ج) مقارنة بالبكتيريا (الانحدار = 0.028؛ الشكل 3ب)، مما يشير إلى أن الاحترار كان له تأثير أكبر على التغيرات الزمنية للفطريات مقارنة بالبكتيريا تحت الزراعة المحافظة، مما يتفق مع الفرضية (iii) التي تفيد بأن التربة الأكثر دفئًا تحت الزراعة المحافظة تسارع من دوران مجتمع الفطريات.
كما لاحظنا أن مجموعات الفطريات ودوران المجتمع تغيرت استجابةً للاحتباس الحراري تحت الزراعة المحافظة، والتي كانت متوازية مع التغيرات في كفاءة نمو الميكروبات وتراكم الكتلة الميتة (الشكل 2). لاستكشاف هذه الديناميات بشكل أعمق، قمنا بتقييم TDRs لمجموعات مختلفة من البكتيريا والفطريات بين القطع المدفأة وغير المدفأة تحت كلا النوعين من الإدارة (الشكل 3ج). كانت TDRs للفطريات أعلى من TDRs للبكتيريا ( مقابل 0.04-0.89؛ الجدول التكميلي 7). اختلفت انحدارات TDR لمجتمعات Basidiomycota وAscomycota بين الاحترار وعدم الاحترار تحت الزراعة المحافظة (; الجداول التكملية 7 و8). تسارع الاحترار من معدلات التغير الزمني لـ Ascomycota وBasidiomycota تحت الزراعة المحافظة، ولكنه قلل من مجتمعات Firmicutes وPlanctomycetota (; الشكل 3ج؛ الجدول التكميلي 7). أظهر تحليل المؤشرات الحدية (TITAN) أيضًا أن Ascomycota وBasidiomycota شهدت زيادات كبيرة مع مرور الوقت (الشكل التكميلي 9). علاوة على ذلك، كان للاحتباس الحراري تأثيرات أكثر وضوحًا على TDRs للأنواع البكتيرية، بما في ذلك
Firmicutes وGemmatimonadota تحت الزراعة التقليدية (; الشكل 3ج؛ الجدول التكميلي 7).
اعتمدت تأثيرات الاحترار على وظائف الميكروبات المتعلقة بنمو الميكروبات على الإدارة
غيرت الإدارة والاحترار من مقادير مدخلات الكربون النباتي، بما في ذلك بقايا المحاصيل من الكتلة الحيوية فوق الأرض، وكتلة الجذور وإفرازات الجذور (الشكل 1). كان لهذه التغيرات تأثيرات متتالية على الكتلة الحيوية، والنشاط (الأشكال 1هـ، و، 2)، الهيكل والدوران (الشكل 3) لمجتمعات الميكروبات في التربة، وخاصة تكاثر الفطريات. استخدمنا أيضًا تحليل الميتاجينوم بالطلق لتحديد التغيرات في وظائف الميكروبات المتعلقة بالعمليات الخلوية والتمثيل الغذائي (الشكل 4أ). انخفضت الوفرة النسبية للمسارات المعنية في تخليق مكونات الخلايا الميكروبية استجابةً للاحتباس الحراري تحت كل من الزراعة المحافظة والزراعة التقليدية في المرحلة المبكرة (في 2010) (; الشكل 4أ). ومع ذلك، زادت هذه المسارات تحت الزراعة المحافظة مع الاحترار في المرحلة المتأخرة (في 2020)، خاصة لتكوين الأغشية الحيوية وتخليق الأحماض الأمينية، مما يشير إلى تحفيز محتمل لنشاط الجينات المرتبطة بإنتاج البروتين وإمكانات نمو الخلايا (; الشكل 4أ). للتعمق أكثر في العمليات الحرجة المرتبطة بهيكل غشاء الميكروبات، قمنا أيضًا بتحليل نسبة الاستجابة لـ KOs المتعلقة بتكوين الأغشية الحيوية، وتخليق الأحماض الدهنية وتخليق الأحماض الأمينية للاحتباس الحراري تحت كلا النوعين من الإدارة (; الشكل 4ب). على غرار التغيرات في المسارات الأيضية، أظهرت الوفرة النسبية للجينات الوظيفية المعلنة في تكوين الأغشية الحيوية أكبر استجابة إيجابية للاحتباس الحراري تحت الزراعة المحافظة في المرحلة المتأخرة، جنبًا إلى جنب مع الجينات المرتبطة بتخليق الأحماض الدهنية وتخليق الأحماض الأمينية (; الشكل التكميلي 8).
قدمت التغيرات في الجينات التي ترمز للإنزيمات النشطة للكربوهيدرات بناءً على قاعدة بيانات CAZy (الشكل 4ب) رؤى حول كيفية تأثير الإدارة والاحترار على مدخلات الكربون النباتي، بما في ذلك اللجنين، والسليلوز والبيكتين من جدران خلايا النبات، ومدخلات الميكروبات مثل الكيتين من الكتلة الحيوية الفطرية (المشار إليها بزيادة الجلوكوزامين الفطري؛ الشكل 2ج). زاد الاحترار من الوفرة النسبية لـ
gينات ترميز الإنزيمات اللجنينية بنسبة 7.8-11%، ولكنه قلل من تلك الخاصة بالجنيات المرتبطة بالبيكتين والسليلوز والكيتين تحت كل من الزراعة المحافظة والزراعة التقليدية في المرحلة المبكرة (; الشكل 4ب). تم العثور على بعض الاستجابات المميزة للاحتباس الحراري بين الممارسات الإدارية في المرحلة المتأخرة. تحت الزراعة المحافظة، زاد الاحترار من الوفرة النسبية للجينات التي ترمز لإنزيمات تحلل النشا، ولكنه قلل من الوفرة النسبية للجينات المرتبطة بتحلل البيكتين والسليلوز والكيتين (; الشكل 4ب). وهذا يشير إلى أن الزراعة المحافظة تميل إلى تفضيل الاحتفاظ بالكربون المقاوم (مثل اللجنين والكيتين) أكثر من الكربون القابل للتحلل (مثل النشا) تحت الاحترار، مما يتفق مع الفرضية (ii). على النقيض من ذلك، عزز الاحترار من الوفرة النسبية للجينات المرتبطة بتحلل النشا والسليلوز والكيتين واللجنين بنسبة، و، و، على التوالي، تحت الزراعة التقليدية في المرحلة المتأخرة (; الشكل 4ب).
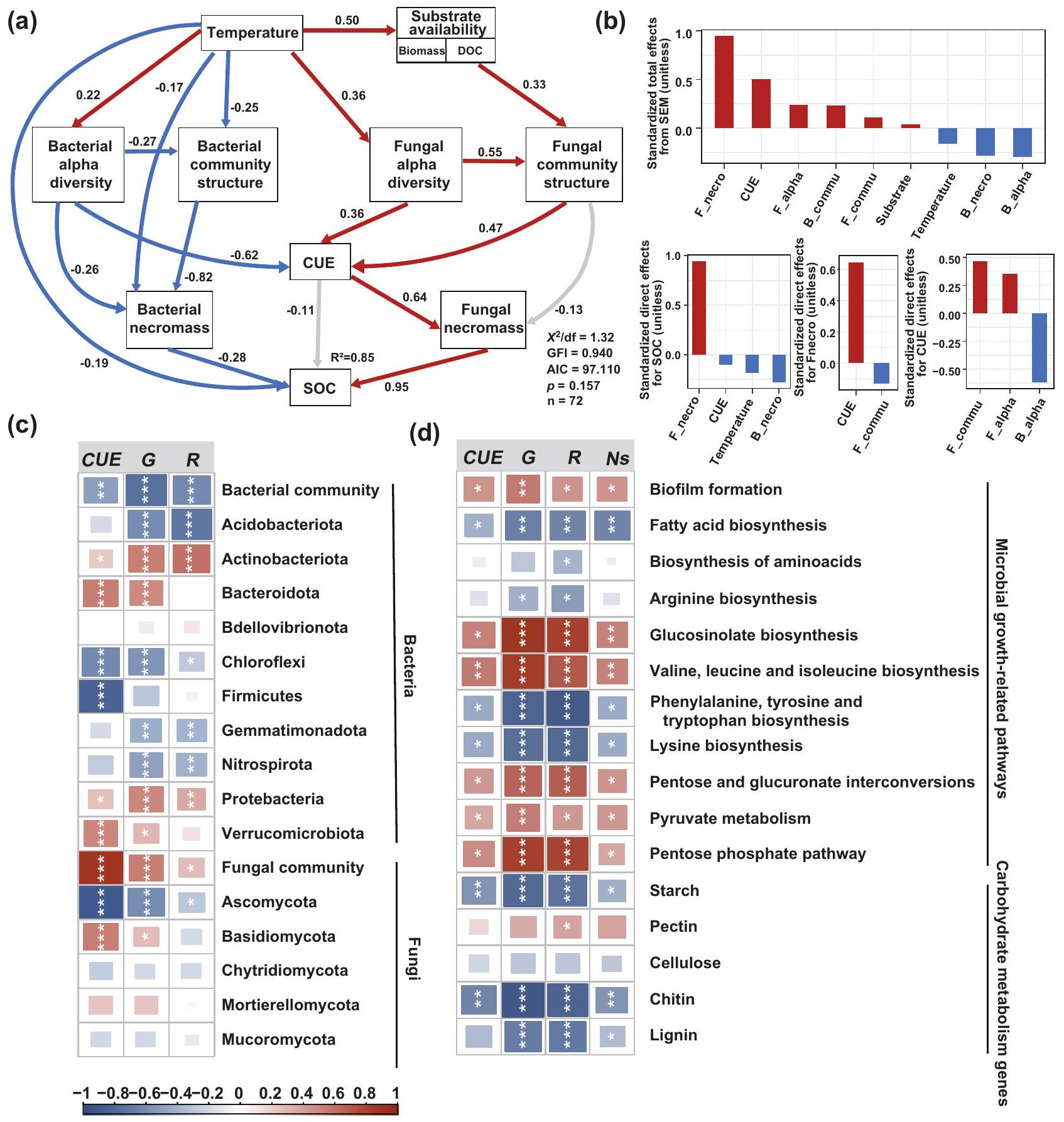
تراكم SOC كدالة لتوافر الركيزة الناتج عن الاحترار، المجتمع الميكروبي والخصائص الفسيولوجية
تم استخدام نمذجة المعادلات الهيكلية (SEM) لتحديد المكونات غير الحيوية (فسيولوجيا الميكروبات والتنوع) والحيوية (توافر الركيزة وعوامل المناخ المحلي) الناتجة عن الاحترار التي تؤثر على ديناميات SOC. كشفت نتائجنا عن الدور الحاسم لخصائص فسيولوجيا الميكروبات في التوسط في ديناميات SOC تحت الزراعة المحافظة. إجمالي من التغيرات في SOC تم حسابه من خلال تركيبات درجة حرارة التربة، توافر الركيزة (المشار إليه بكتلة النبات وDOC)، تنوع الميكروبات ألفا، تركيب المجتمع الميكروبي، CUE الميكروبي والكربون من النخر (الشكل 5a). من بين هذه العوامل، كان التأثير المباشر الإيجابي لكربون النخر الفطري على SOC هو الأكبر (الشكل 5b)، بينما كانت CUE الميكروبي مرتبطة بشكل رئيسي بالتأثيرات غير المباشرة (الشكل 5b). كان هذا متوافقًا مع الارتباط الإيجابي القوي بين كربون النخر الفطري وSOC (; الشكل التكميلي 9a). كان التأثير المباشر الإيجابي لـ CUE الميكروبي على كربون النخر الفطري هو الأكبر (معامل المسار ; الشكل 5b): زاد كربون النخر الفطري بمعدل أسرع مع نمو الميكروبات مقارنة بالتنفس، مما أدى إلى علاقة إيجابية كبيرة بين كربون النخر الفطري وCUE (; الشكل التكميلي 9b). exerted توافر الركيزة تأثيرًا قويًا على تركيب المجتمع الفطري من خلال التأثير المباشر. بشكل عام، أظهر تركيب المجتمع الفطري ارتباطات إيجابية مع نمو الميكروبات (; الشكل 5c؛ الجدول التكميلي 9)، مما أدى إلى تأثيرات مباشرة إيجابية قوية على CUE الميكروبي (معاملات المسار , الشكل 5a). أظهر الوفرة النسبية للفصيلة الفطرية السائدة Ascomycota ارتباطات سلبية مع CUE الميكروبي والنمو خلال المرحلة المبكرة، ولكن ارتباطات إيجابية في المرحلة اللاحقة (; الشكل التكميلي 9d). كانت الوفرة النسبية للفصيلة الفطرية Basidiomycota مرتبطة ارتباطات إيجابية مع CUE ونمو الميكروبات (; الشكل 5c؛ الجدول التكميلي 9). علاوة على ذلك، زادت CUE الميكروبي والنخر مع الوفرة النسبية للجينات المتعلقة بإمكانات نمو الميكروبات ولكنها انخفضت مع الجينات التي تشفر إنزيمات تحلل الكربوهيدرات (; الشكل 5d؛ الجدول التكميلي 10).
نقاش
تستكشف هذه الدراسة التأثيرات التفاعلية للإدارة والاحترار على تراكم SOC واستمراريته في الأراضي الزراعية على مدى عقد من الزمن. توفر نتائجنا أدلة تجريبية على أن الزراعة المحافظة يمكن أن تزيد من تراكم واستمرارية SOC تحت سيناريوهات الاحترار المناخي المتوقعة. يتم التحكم في ديناميات SOC من خلال التوازن بين إدخال الكربون من النباتات والتحلل عبر تنفس الميكروبات، وكلاهما يتم تعديله بواسطة الإدارة والعوامل المناخية . بشكل عام، من المتوقع أن يتسبب الاحترار بحد ذاته في فقدان SOC، خاصة في التربة ذات المحتويات العالية من الكربون العضوي . غالبًا ما تحتوي التربة المدارة تحت الزراعة المحافظة على مدى طويل على تجمعات كبيرة من الكربون العضوي غير المحمي (مثل المادة العضوية الجزيئية)، والتي قد تكون أكثر
عرضة للاحتباس الحراري . لم نلاحظ أن الاحترار أثر على محتوى SOC تحت الزراعة التقليدية (الشكل 1a)، ولكن تم زيادة الوفرة النسبية للجينات الميكروبية التي تشفر تحلل الكربون القابل للتحلل وغير القابل للتحلل بواسطة الاحترار، مما يشير إلى إمكانية فقدان SOC إضافي (الشكل 4b). على الرغم من أن الاحترار زاد من الكتلة الحيوية تحت الأرض وإفراز الجذور (الشكل 1e-f)، إلا أن هذه التغييرات لم تُترجم إلى زيادة في SOC تحت الزراعة التقليدية المدفأة. هنا، زاد محتوى SOC بشكل حصري تحت الزراعة المحافظة وتم تعزيزه بمقدار 10 سنوات من الاحترار (الشكل 1a، j)، والذي يمكن أن يُعزى جزئيًا إلى تغطية التربة المستمرة التي وسّطت تأثيرات الاحترار على درجة حرارة التربة ومنعت الجفاف المفرط. من المهم أن خصائص فسيولوجيا الميكروبات وتداول المجتمع قد تغيرت بعد السنة الخامسة من الاحترار تحت الزراعة المحافظة (الأشكال 2 و3؛ الأشكال التكملية 3 و4)، مما أدى إلى زيادة تراكم SOC (الشكل 1j). تشير هذه النتائج إلى أن دورة الكربون الميكروبية الأكثر كفاءة هي التي تكمن وراء تراكم SOC الملحوظ تحت الزراعة المحافظة استجابةً للاحتباس الحراري، خاصة بعد 5 سنوات.
CUE الميكروبي هو معلمة أيضية حاسمة تصف نسبة العمليات البنائية والهدمية . تشير CUE الميكروبي الأعلى في الزراعة المحافظة مع معالجة الاحترار، والتي تتميز بزيادة أكبر في نمو الميكروبات (33%) مقارنة بالتنفس (13%) على مدى 10 سنوات (الشكل 2a، b والشكل التكميلي 3)، إلى تعزيز عام في كفاءة نمو الميكروبات. يتضمن الزيادة في CUE الميكروبي تحت الزراعة المحافظة استجابات تكاليف الصيانة ومعدلات النمو للتغيرات الناتجة عن الاحترار في درجة حرارة التربة، الرطوبة، توافر الركيزة والمجتمعات الميكروبية في هذه الدراسة. exerted هيكل المجتمع الفطري أقوى تأثير إيجابي على CUE الميكروبي، على الرغم من أن العلاقة كانت متوسطة بشكل رئيسي بواسطة توافر الركيزة (الشكل 5a، b). تعتبر الكتلة الحيوية للجذور وإفراز الجذور أشكالًا رئيسية من إدخال الكربون إلى التربة، والتي تعمل كمصادر كربون مهمة للميكروبات التربة. مع الاحتفاظ بفضلات المحاصيل، زادت DOC، الكتلة الحيوية فوق الأرض، الكتلة الحيوية تحت الأرض للجذور ورسو الجذور (الشكل 1d-g) تحت الزراعة المحافظة على المدى الطويل مع الاحترار، مما يشير إلى إدخالات كربون نباتية أعلى (الشكل 1d-f) التي وفرت ركائز وفيرة لتكاثر الميكروبات. هذا فضل بشكل خاص نمو الفطريات، مما أدى إلى زيادة الكتلة الحيوية الفطرية (الشكل 1h) ونسب الفطريات/البكتيريا (الشكل 1i) تحت الإدارة المحافظة على المدى الطويل مع الاحترار. الفطريات حساسة للتغيرات في المناخ المحلي (مثل الرطوبة ودرجة الحرارة) والاضطرابات الخارجية . تعزز الزراعة المحافظة وفرة الفطريات من خلال زيادة توافر الركيزة، وتقليل اضطراب هيكل التربة، والحفاظ على الشبكات الهيفالية وتحسين توافر المياه . زادت CUE الميكروبي استجابةً للاحتباس الحراري بشكل خطي مع الوقت تحت الزراعة المحافظة، خاصة خلال المرحلة اللاحقة من 2016 إلى 2020 (الشكل 2b والشكل التكميلي 3). يتوافق هذا أيضًا مع زيادة في الوفرة النسبية للجينات الوظيفية الميكروبية المرتبطة بنمو الخلايا الميكروبية وإمكانات إنتاج البروتين (مثل تشكيل الأغشية الحيوية، تخليق الأحماض الدهنية والأحماض الأمينية؛ الشكل 4a)، مع عدد أقل من الجينات المتعلقة بتحلل الكربون غير القابل للتحلل تحت الزراعة المحافظة مع الاحترار خلال المرحلة اللاحقة (الشكل 4). يتفق هذا مع دراسة طويلة الأمد عن الاحترار في سهل العشب الطويل، والتي أفادت بتحول نحو تحلل الركائز الأكثر قابلية للتحلل بسبب زيادة إدخالات النبات . يتماشى هذا أيضًا مع الملاحظة بأن الميكروبات قد تقلل من إنتاج البروتين استجابةً لنضوب الركيزة الناتج عن الاحترار . على العكس من ذلك، زادت كفاءة استخدام الركائز الأكثر مقاومة عند درجات حرارة أعلى في التربة المعرضة للاحترار لمدة تقارب عقدين عند فوق المعدل الطبيعي في غابة هارفارد . بشكل عام، تؤكد نتائجنا على أهمية هيكل المجتمع الميكروبي وتوافر الركيزة في دفع استجابات العمليات الأيضية الميكروبية للاحتباس الحراري تحت الزراعة المحافظة.
تعد دراسات دوران المجتمع الميكروبي في التربة وخصائص الميكروبات الفسيولوجية على مر الزمن استجابةً لتغير المناخ أمرًا حيويًا لـ
الاستجابة للاحتباس الحراري في المراعي وغابة . تحتاج الفطريات إلى كربون أكبر من البكتيريا وعادة ما تكون الأولى في استخدام إدخالات الكربون النباتية تحت الأرض . وبالتالي، فإن النمو الفطري السريع وتغيرات المجتمع الفطري، المدفوعة بزيادة مدخلات الكربون من النباتات، هي عوامل رئيسية في التغيرات الزمنية في المجتمع تحت الزراعة المستدامة مع ارتفاع درجات الحرارة. كان لتوفر الركيزة الأكبر بسبب زيادة مدخلات الكربون من النباتات تأثير مباشر على تركيب المجتمع الفطري، كما تدعمه التغيرات الملحوظة في سلالة الفطريات (الشكل 5 أ-ج). من بين مختلف الفئات الفطرية، تم الإبلاغ عن زيادة Ascomycota سابقًا مع ارتفاع درجات الحرارة.، أوضح أكبر تباين في المجتمعات الفطرية الكلية (الشكل التوضيحي التكميلي 6). كفطريات غنية بالمغذيات، فإن Ascomycota عمومًا-استراتيجيون يستخدمون مصادر الكربون المتقلبة بكفاءة من أجل الأيض السريع والنمو والتكاثراستجابةً لزيادة توفر الركيزة، قد يكون لهذه الفطريات وقت جيل أقصر، مما يؤدي إلى زيادة عدد الأجيال في وحدة الزمن.لذلك، نفترض أن الاحترار لعب دورًا في دفع التباين الزمني للفطريات، كما يتضح من زيادة الفروق بين المواقع المدفأة وغير المدفأة تحت الزراعة المستدامة (الشكل 3ب). في المقابل، أوضحت البكتيريا الحمضية أكبر تباين لمجتمعات البكتيريا (الشكل التكميلي 6). يتميز هذا المجموعة بخصائص أوليغوتروفية، مما يعني أنها تزدهر في البيئات ذات المغذيات المنخفضة.وترتبط ارتباطًا وثيقًا بانخفاضات في التمعدن الصافي للنيتروجين بسبب انخفاض توفر الركيزة الناتج عن جفاف التربةلذلك، قد يُعزى التغير الزمني الأبطأ لمجتمعات البكتيريا إلى وجود نسبة كبيرة من البكتيريا غير النشطة (الخاملة أو بطيئة النمو) في التربة الأكثر دفئًا وجفافًا.تشير هذه النتائج إلى أن التعديلات الفسيولوجية الميكروبية التي اختارت سلالات ميكروبية كانت أكثر تكيفًا مع تغير الظروف الميكروكليماتية للتربة وتوافر الركيزة مع مرور الوقت.
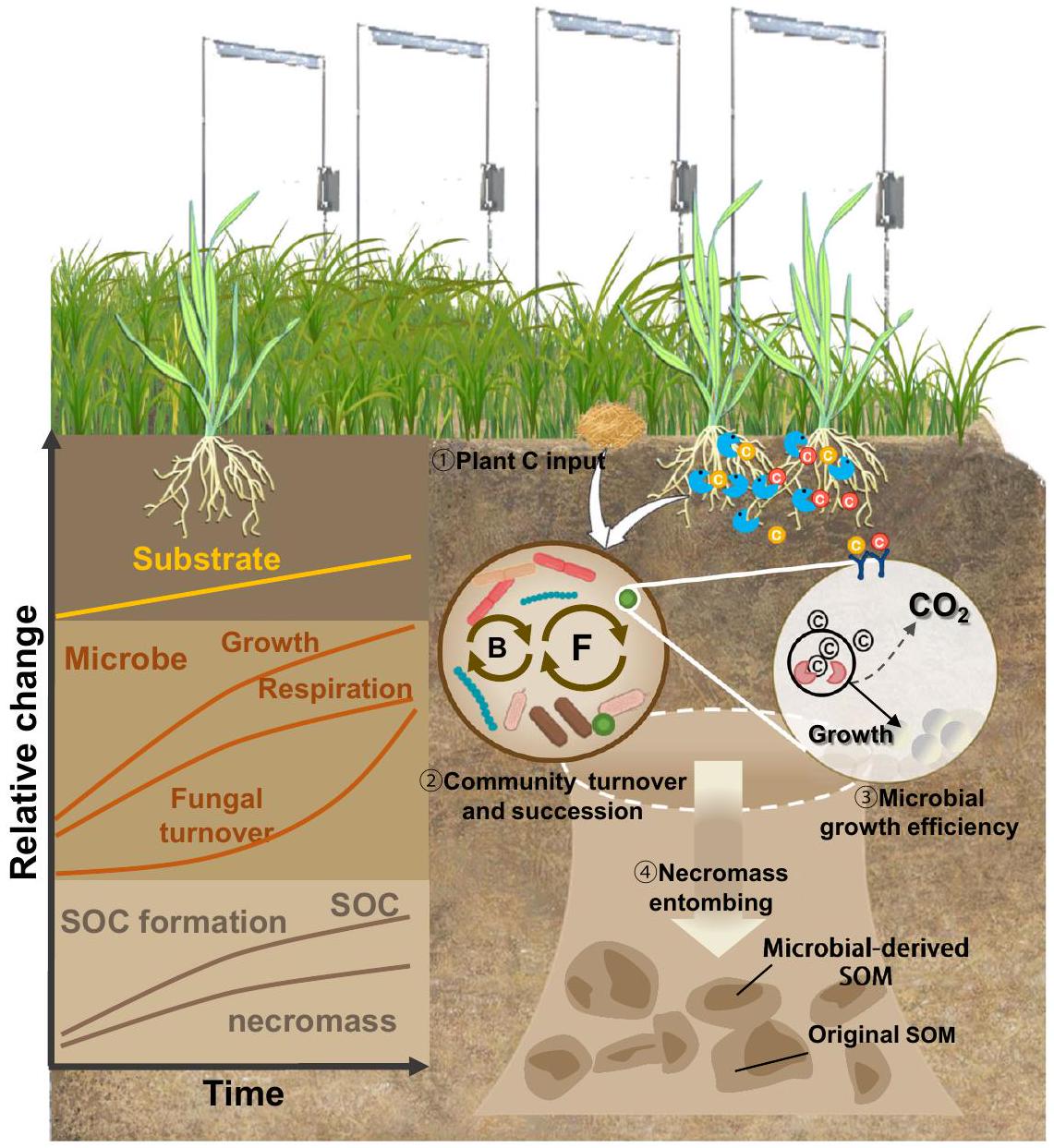
تؤكد النظرية المتطورة لتكوين المادة العضوية في التربة على إنتاج الكتلة الحيوية الميكروبية وتفاعلات مصفوفة المعادن كآليات رئيسية لتكوين واستقرار الكربون العضوي في التربة.تشكل الكتلة الميتة للميكروبات أكثر من 50% من الكربون العضوي في التربة، والتي تكون محمية إما بشكل مادي داخل كتل التربة أو كيميائيًا من خلال ارتباطها مع معادن التربة.زاد الكربون الناتج عن الكتلة الحيوية الفطرية مع ارتفاع درجات الحرارة ويمثل أكبر التغيرات في تراكم الكربون العضوي في التربة تحت الزراعة المستدامة: 36% من إجمالي الكربون العضوي في التربة كان مستمدًا من الكتلة الحيوية الميكروبية (السكريات الأمينية)، معمن أصل فطري (الأشكال 3ج، د و5أ). تدعم هذه النتائج الفكرة القائلة بأن الكتلة الميتة للفطريات هي محرك حاسم لتراكم الكربون العضوي المستقر.تراكم الكتلة الحيوية الميتة للميكروبات في التربة مدفوع بإدخال الكربون من النباتات ويتوسطه إنتاج الكتلة الحيوية الميتة للميكروبات واستقرارها.وبناءً عليه، كانت هناك علاقة إيجابية بين المجتمع الفطري وكفاءة استخدام الكائنات الدقيقة، التي كان لها أكبر تأثير إيجابي على الكتلة الميتة للفطريات (الشكل 5 أ، ب). تشير هذه النتائج إلى أن الاحترار زاد من كفاءة نمو الكائنات الدقيقة وتداول المجتمع الفطري بسبب زيادة مدخلات الكربون من النباتات، مما زاد في النهاية من إمدادات بقايا الكائنات الدقيقة التي تساهم في تشكيل الكربون العضوي في التربة. بالإضافة إلى ذلك، فإن اعتماد الزراعة المستدامة يقلل من اضطراب التربة ويعزز تكوين واستقرار الكتل، مما يوفر بيئة ملائمة لتراكم الكتلة الميتة المشتقة من الفطريات في الكتل الكبيرة.بشكل عام، تشير نتائجنا إلى أن الاحترار يعزز تشكيل وتراكم الكربون العضوي في التربة تحت الزراعة المستدامة من خلال زيادة كفاءة نمو الميكروبات عبر تسريع دوران مجتمع الفطريات، بالإضافة إلى زيادة تراكم الكربون من الكتلة الميتة للفطريات (الأشكال 2 و3). باختصار، قدمت هذه الدراسة أدلة تجريبية على الآليات التي تمكن من زيادة تراكم الكربون العضوي في التربة واستمراريته تحت الاحترار والزراعة المستدامة. كما هو موضح في الشكل 6، من خلال تنفيذ إدارة طويلة الأمد في الموقع.في تجربة حقلية للتدفئة، كشفنا عن تسارع تدريجي في تراكم الكربون العضوي في التربة استجابةً للاحترار المناخي تحت الزراعة المستدامة. تبرز أبحاثنا الأدوار المهمة لتغير مدخلات الكربون النباتي وظروف المناخ الصغير في دفع الاستجابات الميكروبية للاحتباس الحراري تحت الزراعة المستدامة. الزراعة. بشكل خاص، تحت الزراعة المستدامة على المدى الطويل مع ارتفاع درجات الحرارة، تسارعت زيادة مدخلات الكربون النباتي مما أدى إلى تسلسل الفطريات وتعزيز كفاءات نمو الميكروبات، مما أدى إلى زيادة تدريجية في مساهمات الكربون المشتق من الميكروبات في تشكيل وتراكم الكربون العضوي في التربة على مدى عقود. إذا كانت نتائجنا قابلة للتعميم على أنظمة ومناطق أخرى حيث لا يحد الماء من الإنتاجية (مثل المناطق المروية)، فإننا نقترح أن الإدارة التجديدية يمكن أن تعزز من تخزين الكربون الفعال تحت ارتفاع درجات الحرارة، مما يزيد من مرونة الزراعة كعنصر حيوي من “الزراعة الذكية مناخياً”. ومع ذلك، هناك حاجة إلى مزيد من البحث لفهم التأثيرات المشتركة للإدارة والمناخ على تراكم واستمرارية الكربون العضوي في التربة القابلة للزراعة عبر مناطق مختلفة ذات نطاق واسع من هطول الأمطار، مما يمكن أن يوجه ممارسات استخدام الأراضي الذكية مناخياً لتخزين الكربون بشكل فعال وتحسين إنتاج المحاصيل في مواجهة تغير المناخ.
طرق
موقع التجربة والتصميم
تم بدء تجربة ميدانية طويلة الأمد مع نظام القمح الشتوي مزدوج المحصول (Triticum aestivum L.) – الذرة الصيفية (Zea mays L.) في عام 2003 في محطة يوتشينغ الشاملة للتجارب في شمال الصين. )، الذي ينتمي إلى الأكاديمية الصينية للعلوم (CAS). تتمتع المنطقة بمناخ شبه جاف معتدل مع متوسط درجة حرارة سنوي قدره “، ومتوسط هطول الأمطار السنوي 561 مم. التربة لها قوام طيني سلتيني معرملطينطين، ومتوسط pH يبلغ 7.1. نوع التربة هو فلوسول كالكاريك وفقًا لنظام الفاو-يونسكو.
تم وضع أربعة علاجات في تصميم كتلة عشوائية كاملة مع أربع مكررات: الزراعة المستدامة مع وبدون تدفئة (Conserv-Amb، Conserv-Warm) والزراعة التقليدية مع وبدون تدفئة (Conven-Amb، Conven-Warm). تم تقديم التفاصيل الكاملة لتصميم التجربة والإدارة في المرجع 73. باختصار، تم إنشاء علاجات الزراعة التقليدية والزراعة المستدامة في عام 2003. تم تحديد مستويين من التدفئة (البيئة و ) تم فرضها على كل من الزراعة المحافظة والزراعة التقليدية منذ عام 2010. تم تعليق سخان بالأشعة تحت الحمراء (سخان الأشعة تحت الحمراء MSR-2420، Kalglo Electronics Inc، بيت لحم، بنسلفانيا) على ارتفاع حوالي 3 أمتار فوق الأرض في كل قطعة دافئة لتحقيق تسخين سطح التربة التي تم توقعها من قبل سيناريوهات غازات الدفيئة التابعة للهيئة الحكومية الدولية المعنية بتغير المناخ لمعدلات شمال الصين.
حجم كل قطعة كانتم زراعة القمح الشتوي في أوائل أكتوبر وتم حصاده في أوائل يونيو. تم زراعة الذرة الصيفية في يونيو وتم حصادها في نهاية سبتمبر. في معالجة الزراعة التقليدية، تمت إزالة المخلفات بعد حصاد الذرة. تم الزراعة باستخدام محراث دوار إلى عمقتم دمج باقي القش بالكامل في التربة. في معالجة الزراعة المستدامة، تم تقطيع جميع المخلفات إلى حوالي 5 سم في الطول واحتفظ بها على سطح التربة. كانت جميع إجراءات الإدارة الأخرى متطابقة لكلا نوعي الإدارة.
القياسات الميدانية، أخذ عينات التربة والتحليلات
تمت مراقبة درجة حرارة التربة على عمق 5 سم باستخدام ترمومتر PT 100. تم قياس رطوبة التربة الحجمية عندتم قياس العمق بواسطة حساسات رطوبة التربة FDS100 (شركة يونيزم تكنولوجيز، بكين). من 2010 إلى 2020، تم حصاد الكتلة الحيوية الحية فوق الأرض. ) في كل قطعة أرض في مايو من كل عام. تم تجفيف عينات الكتلة الحيوية للنباتات فوق الأرض في حتى تم الحصول على وزن ثابت. تم قياس الكتلة الحيوية للجذور وإفراز الكربون من الجذور من أكتوبر 2019 إلى مايو 2020 خلال مواسم نمو القمح الشتوي وفقًا للمرجع 52. (التفاصيل أدناه).
عينات التربةتم جمع عينات التربة (بعمق) كل عامين بعد حصاد القمح الشتوي من 2010 إلى 2020. تم أخذ خمسة أنوية من التربة السطحية بشكل عشوائي من كل قطعة أرض وتم دمجها كعينة واحدة لكل قطعة. ثم تم تمرير العينات المجمعة من التربة عبر منخل بفتحات 2 مم لإزالة الجذور المرئية والحصى. تم تقسيم العينة إلى 3 أجزاء فرعية-
الشكل 6 | رسم تخطيطي مفاهيمي يوضح تأثير الاحترار على تشكيل وتراكم الكربون العضوي في التربة والآليات الميكروبية الأساسية من خلال النباتاتالمدخلات، تسلسل المجتمع، تعديل الفسيولوجيا وإنتاج الكتلة الحيوية تحت الزراعة المستدامة على مدى عقود. تشمل العمليات الرئيسية (1) زيادة توفر الركيزة بسبب إدخال الكربون من النباتات (مثل بقايا المحاصيل من الكتلة الحيوية فوق الأرض، وكتلة الجذور وإفرازات الجذور) مع ارتفاع درجات الحرارة؛ (2-3) عملت العمليات السابقة كـ ‘محفز’ لدفع تسلسل المجتمع الميكروبي (الذي لوحظ من خلال معدلات التحجيم الزمني العالية وزيادة) تتابع متباين لمجتمع الفطريات) وتعديلات فيزيولوجية ميكروبية (مثل كفاءة استخدام الكربون العالية وكفاءة النمو). بالتوازي، تم ملاحظة الجينات الوظيفية المرتبطة بعملية الأيض الميكروبي المعدلة، حيث زادت الوفرة النسبية للجينات المعنية بإمكانات نمو الميكروبات وتفكيك الكربون القابل للتحلل، بينما انخفضت الجينات التي تشفر لتفكيك الكربون المقاوم؛ (4) تم تعزيز مضخة الكربون الميكروبية تدريجياً حيث تدخل الكتلة الحيوية الميكروبية (خاصة الكتلة الحيوية الفطرية) إلى خزانات الكربون وتدفع تشكيل الكربون الجديد وتراكمه. عينات للتحليل. تم تجفيف العينة الفرعية الأولى في الهواء عند درجة حرارة الغرفة، بينما تم تخزين العينة الفرعية الثانية في، وتم تخزين العينة الفرعية الثالثة في الفريزر عندتم قياس SOC والنيتروجين الكلي (TN) وDOC بعد جمع العينات في 2010 و2012 و2014 و2014 و2018 و2020. تم تحديد محتويات SOC وTN عن طريق احتراق التربة المجففة بالهواء باستخدام محلل العناصر Vario EL III (Elementar). تم قياس تركيز DOC في التربة الطازجة المخزنة فيوفقًا لطريقة جونز وويلت.
تم قياس CUE الميكروبي في عام 2021 في جميع التربة (التي تم أخذ عينات منها في 2010 و2012 و2014 و2016 و2018 و2020) التي تم تخزينها فياستخدامطريقة المتعقب بعد 7 أيام من التحضين المسبق (التفاصيل أدناه). تم إجراء تحليلات لتنوع البكتيريا والفطريات على ثلاثة من الأربع عينات المكررة التي تم تخزينها في في عام 2021. عينات التربة المخزنة (تم جمعها في عام 2010 و2020 وتم إخضاعها لمزيد من تسلسل الميتاجينوم. تم تحليل التربة الطازجة المأخوذة في عام 2020 لتكوين المجتمع الميكروبي باستخدام تحليل الأحماض الدهنية الفوسفوليبيدية (PLFA) وفقًا للمرجع 75؛ تم تقديم التغيرات في تكوين المجتمع الميكروبي كنسب مولارية (نسبة مولية %) لمؤشرات PLFA للبكتيريا أو الفطريات. تم استخراج السكريات الأمينية من التربة المجففة بالهواء وتحديدها وفقًا لطريقة زانغ وأميلونغ. (التفاصيل أدناه).
استخراج الحمض النووي وتسلسل الأمبليكون
تم استخراج الحمض النووي من 0.25 جرام من التربة باستخدام مجموعة عزل PowerSoil (مختبرات موبيو، كارلسباد، كاليفورنيا، الولايات المتحدة الأمريكية) وفقًا لتعليمات الشركة المصنعة. تم تقييم جودة الحمض النووي المنقى بناءً على “ و نسب الامتصاص التي تم الحصول عليها باستخدام مقياس الطيف الضوئي NanoDrop ND-1000 (شركة NanoDrop Technologies Inc.، ويلمنجتون، ديلاوير، الولايات المتحدة الأمريكية). تم تخزين الحمض النووي فيحتى تحليل التسلسل.
تم تضخيم منطقة V4-V5 من 16S rRNA ومنطقة الفاصل الداخلي (ITS) من rRNA لبناء ملفات تعريف المجتمع البكتيري والفطري، على التوالي، باستخدام التسلسل عالي الإنتاجية. تم استخدام مجموعات البرايمر العالمية، F515 (5″-GTGCCAGCMGCCGCGC3′) و R907 (-CCGTCAATTCMTTTRAGTTT-3′) تستهدف جينات الرنا الريباسي 16 S البكتيرية و gITS7F (-GTGARTCATCGARTCTTTG-3′) و ITS4R (-TCCTCCGCTTATTGATATGC-3) للـ ITS الفطرية، تم استخدامهاتم استخدام البرنامج الحراري التالي لتكبير جين 16 S rRNA: الت denaturation الأولي عندلمدة 3 دقائق، تليها 27 دورة من الت denaturing عند لمدة 30 ثانية، تلدين عند لمدة 30 ثانية وامتداد عند لمدة 45 ثانية، ثم تمديد واحد عند لمدة 10 دقائق وتنتهي عند تم إجراء تضخيم PCR لجين rRNA ITS2 على النحو التالي: الت denaturation الأولي عند لمدة 5 دقائق، تليها 30 دورة من الت denaturation عند لمدة 30 ثانية، تلدين عند لمدة 30 ثانية وامتداد عندلمدة 30 ثانية، وتمديد واحد عند لمدة 10 دقائق وتنتهي عند تم تجميع الأمبليكون المنقى بشكل متساوٍ وتم تسلسله من الطرفين على منصة Illumina Nova6000 (Illumina، سان دييغو، الولايات المتحدة الأمريكية) وفقًا للبروتوكولات القياسية من قبل شركة Majorbio Bio-Pharm Technology Co. Ltd. (شنغهاي، الصين).
تم إخضاع التسلسلات الخام لرقابة الجودة وفقًا للمعايير التالية: (ط) تم تقصير قراءات 300 نقطة أساس في أي موقع يحصل على متوسط درجة جودة منعلى نافذة منزلقة بطول 50 نقطة أساس، وتم التخلص من القراءات المقطوعة التي تقل عن 50 نقطة أساس، كما تم التخلص من القراءات التي تحتوي على أحرف غامضة؛ (ii) تم تجميع فقط التسلسلات المتداخلة التي تزيد عن 10 نقاط أساس وفقًا لتسلسلها المتداخل. كانت نسبة الخطأ القصوى في منطقة التداخل 0.2. وتم التخلص من القراءات التي لم يمكن تجميعها؛ (iii) تم تمييز العينات وفقًا للباركود والبرايمرات، وتم ضبط اتجاه التسلسل، مع مطابقة دقيقة للباركود، ووجود خطأين نوكليوتيديين في مطابقة البرايمر. تم اختيار الأنماط الفيلوجينية (أي، متغيرات تسلسل الأمبليكون، ASVs) باستخدام UNOISE3 مع المعلمات الافتراضية في USEARCH وتم التعرف عليها عند مستوى الهويةتم تحليل تصنيف كل تسلسل ممثل لـ ASV بواسطة خوارزمية RDP Classifier Bayesian مقابل قاعدة بيانات سيلفا.https://www.arb-silva.de/) وقاعدة بيانات UNITE (https://unite.ut.ee/باستخدام عتبة ثقة تبلغ 0.7. استخدمنا مجموعة فرعية مختارة عشوائيًا من 24,579 و 21,550 تسلسل لكل عينة لتحليل المجتمعات البكتيرية والفطرية لاحقًا.
تسلسل الميتاجينوم وتحليل البيانات
تم تجزئة مستخلص الحمض النووي إلى حجم متوسط يبلغ حوالي 400 قاعدة باستخدام جهاز Covaris M220 (شركة جين، الصين) لبناء مكتبة القراءة المزدوجة. تم بناء مكتبة القراءة المزدوجة باستخدام NEXTFLEX Rapid DNA-Seq (Bioo Scientific، أوستن، تكساس، الولايات المتحدة الأمريكية). تم ربط المحولات التي تحتوي على مجموعة كاملة من مواقع تزاوج بادئات التسلسل بنهاية القطع المستقيمة. تم إجراء تسلسل القراءة المزدوجة على جهاز Illumina NovaSeq (شركة إلومينا، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) في شركة Majorbio Bio-Pharm Technology Co.، Ltd. (شنغهاي، الصين).
تمت إزالة المحولات من قراءات إلومينا ذات الطرفين، وقراءات الجودة المنخفضة (الطولأو بقيمة جودةأو وجود N قواعد) تم إزالتها بواسطة fastp (https://github.com/OpenGene/fastp، الإصدار 0.20.0). تم تجميع بيانات الميتاجينوم باستخدام MEGAHIT (https://github.com/voutcn/megahit، الإصدار 1.1.2). القطع بطول تم اختيار bp كنتيجة للتجميع النهائي، ثم تم استخدام الكنتيجات لمزيد من توقع الجينات والتعليق. تم محاذاة التسلسلات التمثيلية للجينات غير المتكررة مع قاعدة بيانات KEGG. (http://www.diamondsearch.org/index.php، الإصدار 0.8.35) ضد قاعدة بيانات موسوعة كيوتو للجينات والجينومات (http:// www.genome.jp/keeg/تم إجراء توضيح إنزيمات نشطة للكربوهيدرات باستخدام hmmscan.http://hmmer.janelia.org/search/hmmscan) ضد قاعدة بيانات CAZy (http://www.cazy.org/“). استخدمنا طريقة RPM (القراءات لكل مليون) لتطبيع الوفرة النسبية للمسارات والجينات المعلنة ضمن قاعدة بيانات KEGG و CAZy..
كفاءة استخدام الكربون الميكروبي
تم قياس CUE الميكروبي باستخدامطريقة المتعقبعلى التربة التي تم تخزينها سابقًا فيوفقًا للمرجع 81. تم تعديل عينات التربة إلىسعة الماء في الحقل مع ماء ميلليبور (يتم تجديده وفقًا لفقدان الوزن كل 3 أيام) وتم تحضيره مسبقًا لمدة 7 أيام عندفي أنابيب مغطاة بأفلام ختم مثقبة لتجنب الإفراطتراكم. بعد التحضين المسبق، تم نقل 1 جرام من التربة المحضنة مسبقًا إلى أنبوبين من نوع فالكون سعة 2 مل (0.5 جرام من التربة في كل أنبوب). تم تعديل عينة واحدة بـماء غني بالأكسجين لتحقيق 30% من الذراتتم وسم الماء في التربة بعلامة O، وتم تعديل العينة الأخرى بنفس حجم ماء ميلليبور غير المسمى كتحكم في الوفرة الطبيعية. ثم تم حضانة جميع العينات لمدة 48 ساعة عندفي الظلام عندسعة مياه الحقل. الـتم قياس ما تم إنتاجه خلال هذه الفترة باستخدام جهاز كروماتوغرافيا الغاز Agilent 7890 A (Agilent Technologies، أيتو بالو، كاليفورنيا) مزود بكاشف TCD. تم استخراج الحمض النووي من التربة باستخدام
مجموعة عزل PowerSoil (مختبرات موبيو، كارلسباد، كاليفورنيا، الولايات المتحدة الأمريكية). تم استخراج الحمض النووي (تم تجفيفه في كبسولات فضية فيلمدة يومين. بعد ذلك،تم تحديد وفرة الكربون الكلي ومحتوى الأكسجين باستخدام محلل عنصري (FLASH 2000، Thermo Fisher Scientific، كامبريدج، المملكة المتحدة) متصل بنظام مطياف الكتلة لنسبة النظائر (IRMS) (واجهة ConFlo VI وMAT253 IRMS، Thermo Scientific، بريمن، ألمانيا) في معهد الزراعة شبه الاستوائية التابع للأكاديمية الصينية للعلوم، (تشانغشا، مقاطعة هونان، الصين).
تم حساب كفاءة استخدام الكربون الميكروبي بناءً على المعادلة التالية:
الكتلة الحيوية الميكروبية تحليل
تم استخدام الجلوكوزامين والغالكتوزامين وحمض الموراميك كعلامات حيوية لبقايا الميكروبات (“الكتلة الحيوية الميتة”). تم استخراجها وتحديدها وفقًا لطريقة زانغ وأميلونغ . تم تجفيف عينات التربة ( ) وتحليلها باستخدام 6 M HCl عند لمدة 8 ساعات بعد إضافة المايو-إنوزيتول (معيار داخلي). تم فصل المركبات المشتقة على جهاز كروماتوغرافيا الغاز المجهز بعمود HP-5 وتم تقديرها باستخدام كاشف تأين اللهب (Agilent 6890 A، Agilent Technologies، ليتلتون، كولورادو، الولايات المتحدة الأمريكية). تم حساب بقايا البكتيريا والفطريات (الكتلة الحيوية الميتة) C بناءً على المعادلات التالية:
بقايا الفطريات مليمول جلوكوزامين مليمول حمض موراميك
حيث 45 هو قيمة التحويل إلى بقايا البكتيريا؛ 179.2 هو الوزن الجزيئي للجلوكوزامين؛ و9 هو عامل التحويل من جلوكوزامين الفطريات إلى بقايا الفطريات. تم تقدير إجمالي بقايا الميكروبات كمجموع لبقايا الفطريات والبكتيريا.
مدخلات الكربون من الكتلة الحيوية للجذور والإفرازات
تم قياس الكتلة الحيوية للجذور ومدخلات الكربون من الإفرازات الجذرية من أكتوبر 2019 إلى مايو 2020 خلال نمو القمح الشتوي. خلال مراحل الإطالة والإزهار والنضج للقمح، جمعنا الإفرازات الجذرية من ستة نباتات فردية في أربعة قطع فرعية من كل معالجة. استخدمنا نظام قوارير ثقافي معدل تم تطويره خصيصًا لجمع الإفرازات الجذرية في الموقع . أولاً، تم اختيار جذور القمح بنفس الارتفاع وحالة النمو، وتم حفر الجذور النهائية بعناية من التربة السطحية ( ) يدويًا وغسلها بشكل مكثف لإزالة جزيئات التربة الملتصقة. ثانيًا، تم وضع الجذور السليمة في القوارير، ثم تم ملء القوارير بكرات زجاجية (بقطر حوالي 1 مم)، والتي وضعت لمحاكاة جزيئات التربة، وتم إغلاقها بسدادة مطاطية خاصة. ثالثًا، تم تغطية القوارير (بما في ذلك الضوابط التي تحتوي على كرات فقط) بورق الألمنيوم وإعادة دفنها في المنطقة المحفورة في التربة. بعد فترة توازن لمدة يوم واحد، تم تدفق كمية صغيرة من محلول مغذي خالي من الكربون ( ) عبر كل قارورة لإزالة الكربون القابل للذوبان. أخيرًا، تم جمع الإفرازات عن طريق تدفق القارورة ثلاث مرات مع 15 مل من محلول المغذيات الطازجة بعد فترة حضانة لمدة 24 ساعة. تم وضع ومعالجة القوارير الضابطة المملوءة فقط بالرمل السيليكا الخشن بنفس الطريقة في كل قطعة. تم تصفية حلول الفخ من خلال مرشحات حقن معقمة خلال من الجمع ثم تم تخزينها في حتى التحليل الإضافي. تم جمع الإفرازات لمدة ثلاثة أيام متتالية. تم تحليل إجمالي الكربون العضوي غير الجزيئي المتراكم في حلول الفخ باستخدام محلل TOC متعدد 3100 N/C (Analytik، يينا، ألمانيا).
تم حساب معدلات الإفراز الجذرية لكل معالجة عن طريق طرح تراكم الكربون داخل القوارير المملوءة بالجذور من الكربون
المكتشف في الضبط الخالي من الجذور ثم تم التعبير عنها كـ الكتلة الحيوية للجذور . قدرنا معدلات الإفراز الجذرية خلال فترة النمو الكاملة من خلال وزن معدلات الإفراز الجذرية في كل فترة نمو. حسبنا الإفرازات الجذرية السنوية من خلال ضرب الكتلة الحيوية الحية للجذور خلال فترة النمو بمعدل الإفراز الجذري الموزون وعدد الساعات لكل فترة نمو .
المعادلة التالية لتقييم إفراز الكربون خلال فترة نمو القمح الكاملة (c)
حيث هو فرق تركيز TOC بين محلول الإفراز الجذري والضبط الفارغ ( )؛ هو حجم محلول الإفراز الجذري (لتر)؛ هو وزن الجذر (غرام)؛ هو وقت الجمع (ساعات)؛ هو الكتلة الحيوية للجذر ( )؛ 24 هو 24 ساعة في اليوم؛ و يمثل فترة النمو.
التحليلات الإحصائية
تم استخدام نماذج مختلطة لتحليل التباين (ANOVAs) لفحص تأثيرات الإدارة، والتسخين، والسنة وتفاعلاتها على SOC، وتنوع البكتيريا والفطريات، والفيزيولوجيا الميكروبية والكتلة الحيوية الميتة. بالإضافة إلى ذلك، تم استخدام المقارنات المتعددة (LSD) لفحص الفروق في المعلمات المختلفة بين المعالجات. تم استخدام الانحدارات الخطية لاكتشاف الاتجاهات الخطية لـ SOC، والخصائص الفيزيولوجية الميكروبية وC الكتلة الحيوية الميتة مع الوقت تحت أربع معالجات. تم تحليل المنحدرات لتلك العلاقات الخطية ومقارنتها من خلال تحليل الانحدار المحوري القياسي باستخدام حزمة smatr في R v.3.2.1 . تم حساب قيمة كوهين كتقدير لحجم تأثير المعالجات المتعددة على استجابة قيم SOC، والفيزيولوجيا الميكروبية والكتلة الحيوية الميتة للتسخين تحت الزراعة المحافظة والزراعة التقليدية من خلال مقارنتها مع الضبط الخالي من التسخين؛ تشير القيم الإيجابية إلى أن المتغيرات الاستجابية في المعالجة لها قيمة أكبر من الضبط، والعكس صحيح . تم إجراء تحليلات حجم التأثير في البرنامج v.3.2.1 مع حزمة effsize.
تم استخدام تحليل المكونات الرئيسية لتقييم التغيرات في مجتمعات البكتيريا والفطريات. تم استخدام ثلاثة اختبارات إحصائية متعددة غير معلمية: تحليل التباين المتعدد غير المعلمي (ADONIS)، تحليل التشابه (ANOSIM) وإجراء التبديل متعدد الاستجابة (MRPP) لاختبار الفروق في مجتمعات الميكروبات في التربة تحت معالجات التسخين والضبط. تم إجراء هذه التحليلات في R v.3.2.1 مع حزمة vegan. تم تقييم تأثير المعالجات على الوفرة النسبية للأنواع البكتيرية والفطرية بناءً على نماذج التأثيرات المختلطة الخطية وفقًا لطرق المرجع 84. تعتمد الأهمية الإحصائية على اختبارات نوع والد II وجميع أحجام التأثير المقدرة تعتمد على المتغيرات الاستجابية المعاد قياسها. تم إجراء هذه التحليلات في v.3.2.1 مع حزمتي lme4 وcar.
تم تقييم علاقات تدهور الزمن (TDR) لمجتمعات البكتيريا والفطريات باستخدام الانحدار الخطي بين التشابهات اللوغاريتمية والمسافة الزمنية اللوغاريتمية. تم استخدام نهج النافذة المتحركة لتقييم تدهور الزمن في مجتمعات الميكروبات كما تم وصفه سابقًا ، وهو حاليًا النهج السائد لـ TDRs. في بيانات تنوع البكتيريا والفطريات التي تم مسحها كل سنتين على مدى 10 سنوات، تضمنت نافذة المجموعة 1 التشابه الثنائي العينات لعينة كانت تفصلها سنتان؛ نافذة المجموعة 2 هي التشابه الثنائي العينات لعينة تفصلها أربع سنوات، وهكذا. لذلك، في هذا السجل الذي يمتد لعشر سنوات، هناك 5 فترات زمنية مدتها سنتان، 4 فترات زمنية مدتها أربع سنوات، 3 فترات زمنية مدتها ست سنوات، 2 فترات زمنية مدتها ثماني سنوات، و1 فترة زمنية مدتها عشر سنوات لكل قطعة. بالنظر إلى تصميم القياسات المتكررة، تم حساب تحليل TDR فقط بالمقارنات الثنائية بين نقاط الزمن داخل كل قطعة (أي، 15 مقارنة ثنائية لكل قطعة وإجمالي 45 مقارنة ثنائية لكل معالجة). قمنا أيضًا بتقييم تأثير التسخين على تسلسل مجتمعات البكتيريا والفطريات في التربة باستخدام المسافة بين مجتمعات الميكروبات تحت التسخين و
الضبط في كل كتلة في كل عام للزراعة المحافظة والزراعة التقليدية، على التوالي وفقًا لطريقة المرجع 57.
تم حساب نسبة استجابة وظائف التمثيل الغذائي للكربون للميكروبات بين قطع التسخين وغير التسخين في الزراعة المحافظة والزراعة التقليدية وفقًا لكيرتس ووانغ . تم استخدام متوسطات مجموعة المعالجة ومجموعة الضبط لحساب نسبة الاستجابة باستخدام:
تم استخدام الطبيعي للاختبارات الإحصائية. إذا كانت و موزعة بشكل طبيعي وكلاهما أكبر من الصفر، فإن موزعة تقريبًا بشكل طبيعي بمتوسط يساوي نسبة الاستجابة الحقيقية.
تم تطبيق تحليل SEM للتحقيق في التأثيرات المباشرة وغير المباشرة للمناخ الميكروبي للتربة (درجة حرارة التربة والرطوبة)، والكتلة الحيوية فوق الأرض، وDOC والمتغيرات الميكروبية (تنوع الألفا الميكروبي، هيكل المجتمع، CUE الميكروبي والنيكرومس) على SOC. أولاً، اعتبرنا نموذجًا مفاهيميًا مفترضًا (الشكل التكميلي 10) الذي شمل جميع المسارات المعقولة. تم اختيار درجة الحرارة فقط لتحسين ملاءمة النموذج لأننا لاحظنا وجود ارتباط قوي بين درجة حرارة التربة والرطوبة (بيرسون). ). قمنا بإزالة المسارات غير المهمة بشكل متسلسل ما لم تكن المسارات ذات دلالة بيولوجية، أو أضفنا مسارات بناءً على الارتباطات المتبقية. تم استخدام 72 عينة تربة مستقلة لتشغيل نموذج المعادلات الهيكلية (SEM). تم تقييم جودة ملاءمة نتائج نموذج المعادلات الهيكلية من خلال اختبار كاي تربيع (X 2) ومؤشر جودة الملاءمة (GFI). عندما و GFI النموذج لديه ملاءمة جيدة. تم إجراء تحليل SEM باستخدام AMOS 23.0 (شركة تطوير AMOS).
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
تم إيداع تسلسلات الحمض النووي لجين 16 S rRNA و ITS amplicons في هذه الدراسة في المركز الوطني لمعلومات التكنولوجيا الحيوية (NCBI) تحت أرقام الوصول للمشاريع PRJNA903096 و PRJNA903090. تم إيداع تسلسلات الميتاجينوم الخام في هذه الدراسة في المركز الوطني لمعلومات التكنولوجيا الحيوية (NCBI) تحت رقم الوصول للمشروع PRJNA1007786. قاعدة بيانات سيلفا متاحة علىhttps://www.arb-silva.de/قاعدة بيانات UNITE متاحة علىhttps://unite.ut.ee/تم توفير بيانات المصدر في هذه الورقة. تم توفير بيانات المصدر مع هذه الورقة.
Schmidt, M. W. et al. Persistence of soil organic matter as an ecosystem property. Nature 478, 49-56 (2011).
Lehmann, J. & Kleber, M. The contentious nature of soil organic matter. Nature 528, 60-68 (2015).
Sanderman, J., Hengl, T. & Fiske, G. J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. USA 114, 9575-9580 (2017).
Lal, R., Follett, R. F., Stewart, B. A. & Kimble, J. M. Soil carbon sequestration to mitigate climate change and advance food security. Soil Sci. 172, 943-956 (2007).
Paustian, K. et al. Climate-smart soils. Nature 532, 49-57 (2016).
Bossio, D. et al. The role of soil carbon in natural climate solutions. Nat. Sustain. 3, 391-398 (2020).
Davidson, E. A. & Janssens, I. A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440, 165-173 (2006).
Crowther, T. W. et al. Quantifying global soil carbon losses in response to warming. Nature 540, 104-108 (2016).
Chappell, A., Baldock, J. & Sanderman, J. The global significance of omitting soil erosion from soil organic carbon cycling schemes. Nat. Clim. Change 6, 187-191 (2016).
Powlson, D. S. et al. Limited potential of no-till agriculture for climate change mitigation. Nat. Clim. Change 4, 678-683 (2014).
Kan, Z. R. et al. Mechanisms of soil organic carbon stability and its response to no-till: a global synthesis and perspective. Glob. Change Biol. 28, 693-710 (2022).
Pittelkow, C. M. et al. Productivity limits and potentials of the principles of conservation agriculture. Nature 517, 365-368 (2015).
Neufeldt, H., Kissinger, G. & Alcamo, J. No-till agriculture and climate change mitigation. Nat. Clim. Change 5, 488-489 (2015).
Wittwer, R. A. et al. Organic and conservation agriculture promote ecosystem multifunctionality. Sci. Adv. 7, eabg6995 (2021).
Qi, J.-Y. et al. Effects of long-term tillage regimes on the vertical distribution of soil iron/aluminum oxides and carbon decomposition in rice paddies. Sci. Total Environ. 776, 145797 (2021).
Karhu, K. et al. Temperature sensitivity of soil respiration rates enhanced by microbial community response. Nature 513, 81-84 (2014).
Lugato, E., Lavallee, J. M., Haddix, M. L., Panagos, P. & Cotrufo, M. F. Different climate sensitivity of particulate and mineral-associated soil organic matter. Nat. Geosci. 14, 295-300 (2021).
Kpemoua, T. P. et al. Are carbon-storing soils more sensitive to climate change? A laboratory evaluation for agricultural temperate soils. Soil Biol. Biochem. 183, 109043 (2023).
Allison, S. D., Wallenstein, M. D. & Bradford, M. A. Soil-carbon response to warming dependent on microbial physiology. Nat. Geosci. 3, 336-340 (2010).
Frey, S. D., Lee, J., Melillo, J. M. & Six, J. The temperature response of soil microbial efficiency and its feedback to climate. Nat. Clim. Change 3, 395-398 (2013).
Hagerty, S. B. et al. Accelerated microbial turnover but constant growth efficiency with warming in soil. Nat. Clim. Change 4, 903-906 (2014).
García-Palacios, P. et al. Evidence for large microbial-mediated losses of soil carbon under anthropogenic warming.Nat. Rev. Earth Environ. 2, 507-517 (2021).
Malik, A. A. et al. Land use driven change in soil ph affects microbial carbon cycling processes. Nat. Commun. 9, 3591 (2018).
Tao, F. et al. Microbial carbon use efficiency promotes global soil carbon storage. Nature 618, 981-985 (2023).
Cotrufo, M. F., Wallenstein, M. D., Boot, C. M., Denef, K. & Paul, E. The microbial efficiency-matrix stabilization (mems) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob. Change Biol. 19, 988-995 (2013).
Liang, C., Schimel, J. P. & Jastrow, J. D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2, 17105 (2017).
Domeignoz-Horta, L. A. et al. Microbial diversity drives carbon use efficiency in a model soil. Nat. Commun. 11, 3684 (2020).
Tian, W. et al. Thermal adaptation occurs in the respiration and growth of widely distributed bacteria. Glob. Change Biol. 28, 2820-2829 (2022).
Alster, C. J., Robinson, J. M., Arcus, V. L. & Schipper, L. A. Assessing thermal acclimation of soil microbial respiration using macromolecular rate theory. Biogeochemistry 158, 131-141 (2022).
Söllinger, A. et al. Down-regulation of the bacterial protein biosynthesis machinery in response to weeks, years, and decades of soil warming. Sci. Adv. 8, eabm3230 (2022).
Domeignoz-Horta, L. A. et al. Substrate availability and not thermal acclimation controls microbial temperature sensitivity response to long-term warming. Glob. Change Biol. 29, 1574-1590 (2023).
Li, J. et al. Reduced carbon use efficiency and increased microbial turnover with soil warming. Glob. Change Biol. 25, 900-910 (2019).
Wang, N. et al. Effects of climate warming on carbon fluxes in grasslands-a global meta-analysis. Glob. Change Biol. 25, 1839-1851 (2019).
Chang, R. et al. Soil organic carbon becomes newer under warming at a permafrost site on the tibetan plateau. Soil Biol. Biochem. 152, 108074 (2021).
Feng, X. & Wang, S. Plant influences on soil microbial carbon pump efficiency. Glob. Change Biol. 29, 3854-3856 (2023).
Dove, N. C., Torn, M. S., Hart, S. C. & Tas, N. Metabolic capabilities mute positive response to direct and indirect impacts of warming throughout the soil profile. Nat. Commun. 12, 2089 (2021).
Nottingham, A. T. et al. Microbial responses to warming enhance soil carbon loss following translocation across a tropical forest elevation gradient. Ecol. Lett. 22, 1889-1899 (2019).
Six, J., Frey, S. D., Thiet, R. K. & Batten, K. M. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 70, 555-569 (2006).
Buckeridge, K. M., Creamer, C. & Whitaker, J. Deconstructing the microbial necromass continuum to inform soil carbon sequestration. Funct. Ecol. 36, 1396-1410 (2022).
Malik, A. A. et al. Soil fungal: bacterial ratios are linked to altered carbon cycling. Front. Microbiol. 7, 1247 (2016).
Strickland, M. S. & Rousk, J. Considering fungal: bacterial dominance in soils-methods, controls, and ecosystem implications. Soil Biol. Biochem. 42, 1385-1395 (2010).
Yang, Y. et al. Fungi determine increased soil organic carbon more than bacteria through their necromass inputs in conservation tillage croplands. Soil Biol. Biochem. 167, 108587 (2022).
Jansson, J. K. & Hofmockel, K. S. Soil microbiomes and climate change. Nat. Rev. Microbiol. 18, 35-46 (2020).
Ding, X. L. et al. Warming increases microbial residue contribution to soil organic carbon in an alpine meadow. Soil Biol. Biochem. 135, 13-19 (2019).
Ullah, M. R., Carrillo, Y. & Dijkstra, F. A. Drought-induced and seasonal variation in carbon use efficiency is associated with fungi: bacteria ratio and enzyme production in a grassland ecosystem. Soil Biol. Biochem. 155, 108159 (2021).
Tian, J. et al. Microbial metabolic response to winter warming stabilizes soil carbon. Glob. Change Biol. 27, 2011-2028 (2021).
Sauvadet, M. et al. High carbon use efficiency and low priming effect promote soil c stabilization under reduced tillage. Soil Biol. Biochem. 123, 64-73 (2018).
Zhou, R. et al. Microbial necromass in cropland soils: a global metaanalysis of management effects. Glob. Change Biol. 29, 1998-2014 (2023).
Sinsabaugh, R. L., Manzoni, S., Moorhead, D. L. & Richter, A. Carbon use efficiency of microbial communities: Stoichiometry, methodology and modelling. Ecol. Lett. 16, 930-939 (2013).
Bradford, M. A. et al. Managing uncertainty in soil carbon feedbacks to climate change. Nat. Clim. Change 6, 751-758 (2016).
Zhang, Q., Qin, W., Feng, J. & Zhu, B. Responses of soil microbial carbon use efficiency to warming: review and prospects.Soil Ecol. Lett. 4, 307-318 (2022).
Phillips, R. P., Erlitz, Y., Bier, R. & Bernhardt, E. S. New approach for capturing soluble root exudates in forest soils. Funct. Ecol. 22, 990-999 (2008).
Yin, H. et al. Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming. Glob. Change Biol. 19, 2158-2167 (2013).
Spohn, M., Klaus, K., Wanek, W. & Richter, A. Microbial carbon use efficiency and biomass turnover times depending on soil depth-implications for carbon cycling. Soil Biol. Biochem. 96, 74-81 (2016).
Zheng, Q. et al. Growth explains microbial carbon use efficiency across soils differing in land use and geology. Soil Biol. Biochem. 128, 45-55 (2019).
Zhang, X. & Amelung, W. Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils. Soil Biol. Biochem. 28, 1201-1206 (1996).
Guo, X. et al. Climate warming leads to divergent succession of grassland microbial communities. Nat. Clim. Change 8, 813-818 (2018).
Paustian, K. et al. Quantifying carbon for agricultural soil management: From the current status toward a global soil information system. Carbon Manag. 10, 567-587 (2019).
Kivlin, S. N. & Hawkes, C. V. Spatial and temporal turnover of soil microbial communities is not linked to function in a primary tropical forest. Ecology 101, e02985 (2020).
Zhang, B. et al. Soil microbial community dynamics over a maize (zea mays l.) growing season under conventional- and no-tillage practices in a rainfed agroecosystem. Soil. Res. 124, 153-160 (2012).
Zhou, J. et al. Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Change 2, 106-110 (2011).
Averill, C., Cates, L. L., Dietze, M. C. & Bhatnagar, J. M. Spatial vs. Temporal controls over soil fungal community similarity at continental and global scales. ISME J. 13, 2082-2093 (2019).
Koch, B. J. et al. Estimating taxon-specific population dynamics in diverse microbial communities. Ecosphere 9, e02090 (2018).
Martinović, T. et al. Temporal turnover of the soil microbiome composition is guild-specific. Ecol. Lett. 24, 2726-2738 (2021).
Asemaninejad, A., Thorn, R. G., Branfireun, B. A. & Lindo, Z. Climate change favours specific fungal communities in boreal peatlands. Soil Biol. Biochem. 120, 28-36 (2018).
Ling, N., Wang, T. & Kuzyakov, Y. Microbiome of the rhizosphere: from structure to functions. Nat. Commun. 13, 836 (2022).
Deng, Y. et al. Elevated carbon dioxide accelerates the spatial turnover of soil microbial communities. Glob. Change Biol. 22, 957-964 (2015).
Fierer, N., Bradford, M. A. & Jackson, R. B. Toward an ecological classification of soil bacteria. Ecology 88, 1354-1364 (2007).
Lennon, J. T. & Jones, S. E. Microbial seed banks: the ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 9, 119-130 (2011).
Liang, C., Amelung, W., Lehmann, J. & Kastner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Glob. Change Biol. 25, 3578-3590 (2019).
Hannula, S. & Morriën, E. Will fungi solve the carbon dilemma? Geoderma 413, 115767 (2022).
Ding, X. L., Liang, C., Zhang, B., Yuan, Y. R. & Han, X. Z. Higher rates of manure application lead to greater accumulation of both fungal and bacterial residues in macroaggregates of a clay soil. Soil Biol. Biochem. 84, 137-146 (2015).
Hou, R., Ouyang, Z., Wilson, G. V., Li, Y. & Li, H. Response of carbon dioxide emissions to warming under no-till and conventional till systems. Soil Sci. Soc. Am. J. 78, 280-289 (2013).
Jones, D. L. & Willett, V. B. Experimental evaluation of methods to quantify dissolved organic nitrogen (don) and dissolved organic carbon (doc) in soil. Soil Biol. Biochem. 38, 991-999 (2006).
Frostegård, Å., Tunlid, A. & Bååth, E. Microbial biomass measured as total lipid phosphate in soils of different organic content. J. Microbiol. Methods 14, 151-163 (1991).
Yusoff, M. Z. M. et al. Influence of pretreated activated sludge for electricity generation in microbial fuel cell application. Bioresour. Technol. 145, 90-96 (2013).
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C. & Knight, R. Uchime improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194-2200 (2011).
Li, D., Liu, C.-M., Luo, R., Sadakane, K. & Lam, T.-W. Megahit: an ultrafast single-node solution for large and complex metagenomics assembly via succinct de bruijn graph. Bioinformatics 31, 1674-1676 (2015).
Buchfink, B., Xie, C. & Huson, D. H. Fast and sensitive protein alignment using diamond. Nat. Methods 12, 59-60 (2015).
Morse, N. et al. Plant-microbe interactions drive denitrification rates, dissolved nitrogen Removal, and the abundance of denitrification genes in stormwater control measures. Environ. Sci. Technol. 52, 9320-9329 (2018).
Schroeder, J., Kammann, L., Helfrich, M., Tebbe, C. C. & Poeplau, C. Impact of common sample pre-treatments on key soil microbial properties. Soil Biol. Biochem 160, 108321 (2018).
Shen, X., Yang, F., Xiao, C. & Zhou, Y. Increased contribution of root exudates to soil carbon input during grassland degradation. Soil Biol. Biochem. 146, 107817 (2020).
Warton, D. I., Wright, I. J., Falster, D. S. & Westoby, M. Bivariate linefitting methods for allometry. Biol. Rev. 81, 259-291 (2006).
Wu, L. et al. Reduction of microbial diversity in grassland soil is driven by long-term climate warming. Nat. Microbiol. 7, 1054-1062 (2022).
Curtis, P. S. & Wang, X. A meta-analysis of elevated effects on woody plant mass, form, and physiology. Oecologia 113, 299-313 (1998).
شكر وتقدير
تم دعم هذه الدراسة من قبل المؤسسة الوطنية للعلوم الطبيعية في الصين (أرقام المنح: 32071629، 32071607)، البرنامج الوطني الرئيسي للبحث والتطوير في الصين (2022YFD1901300)، برنامج تطوير المواهب 2115 في جامعة الزراعة الصينية وبرنامج البحث الاستراتيجي للأكاديميات المتقدمة في بكين والأكاديمية الصينية للعلوم (XDA28130301). كما تم دعم المخطوطة التي تم تعديلها بواسطة Y.K. من قبل برنامج القيادة الأكاديمية الاستراتيجية لجامعة RUDN.
مساهمات المؤلفين
ساهم جميع المؤلفين بمساهمات فكرية ومساعدة في هذه الدراسة وإعداد المخطوطة. قام J.T. و J.A.J.D. و F.Z. و M.F.C. و J.Z. بتصميم الفكرة الأصلية واستراتيجية التجربة. قام J.T. بإجراء التجارب في المختبر وجمع البيانات. قام R.H. بتنفيذ التجربة الميدانية وجمع البيانات. قام J.T. بتحليل البيانات بمساعدة Y.D. قام J.T. بصياغة المخطوطة بمساعدة J.A.J.D. و Y.Y. و I.P.H. و M.F.C. و Y.K. و J.Z. ساهم جميع المؤلفين في المقالة ووافقوا على النسخ المقدمة.
المصالح المتنافسة
يعلن المؤلفون عدم وجود مصالح متنافسة.
معلومات إضافية
معلومات إضافية النسخة الإلكترونية تحتوي على المواد التكميلية متاحة على https://doi.org/10.1038/s41467-023-44647-4. يجب توجيه المراسلات والطلبات للحصول على المواد إلى جينغ تيان، فوسو زانغ، م. فرانشيسكا كوتروفو أو جيزهونغ زو.
معلومات مراجعة الأقران تشكر مجلة Nature Communications المراجع(ين) المجهول(ين) على مساهمتهم في مراجعة هذا العمل. يتوفر ملف مراجعة الأقران.
المختبر الوطني الرئيسي لاستخدام وإدارة المغذيات، كلية الموارد والعلوم البيئية، جامعة الزراعة الصينية، 100193 بكين، جمهورية الصين الشعبية.الجغرافيا، كلية البيئة والعلوم والاقتصاد، جامعة إكستر، شارع رين، إكستر EX4 4RJ، المملكة المتحدة.مركز إدارة الكربون، كلية اسكتلندا الريفية، إدنبرة EH9 3JG، المملكة المتحدة.المختبر الرئيسي لمراقبة النظم البيئية ونمذجتها، معهد علوم الجغرافيا والبحوث في الموارد الطبيعية، الأكاديمية الصينية للعلوم (CAS)، 100101 بكين، جمهورية الصين الشعبية.مختبر المفاتيح لعلوم البيئة الحيوية، مركز أبحاث العلوم البيئية الإيكولوجية، الأكاديمية الصينية للعلوم، 100085 بكين، جمهورية الصين الشعبية.المختبر المشترك الرئيسي للدولة لمحاكاة البيئة ومراقبة التلوث، كلية البيئة، جامعة تسينغhua، بكين، جمهورية الصين الشعبية.قسم علوم التربة للأنظمة البيئية المعتدلة، جامعة غوتنغن، 37077 غوتنغن، ألمانيا.قسم علوم التربة والمحاصيل، جامعة ولاية كولورادو، فورت كولينز، كولورادو، الولايات المتحدة الأمريكية.معهد الجينوم البيئي، جامعة أوكلاهوما، نورمان، أوكلاهوما، الولايات المتحدة الأمريكية.كلية العلوم البيولوجية، جامعة أوكلاهوما، نورمان، أوكلاهوما، الولايات المتحدة الأمريكية.كلية الهندسة المدنية وعلوم البيئة، جامعة أوكلاهوما، نورمان، أوكلاهوما، الولايات المتحدة الأمريكية.كلية علوم الحاسوب، جامعة أوكلاهوما، نورمان، أوكلاهوما، الولايات المتحدة الأمريكية.علوم الأرض والبيئة، مختبر لورانس بيركلي الوطني، بيركلي، كاليفورنيا، الولايات المتحدة الأمريكية.ساهم هؤلاء المؤلفون بالتساوي: جينغ تيان، جينيفر أ. ج. دونغيت، رويزينغ هو، يي دينغ. البريد الإلكتروني:tianj@cau.edu.cn; zhangfs@cau.edu.cn; فرانشيسكا.كوتروفو@كولورادوستيت.إديو; jzhou@ou.edu
Increasing soil organic carbon (SOC) in croplands by switching from conventional to conservation management may be hampered by stimulated microbial decomposition under warming. Here, we test the interactive effects of agricultural management and warming on SOC persistence and underlying microbial mechanisms in a decade-long controlled experiment on a wheatmaize cropping system. Warming increased SOC content and accelerated fungal community temporal turnover under conservation agriculture (no tillage, chopped crop residue), but not under conventional agriculture (annual tillage, crop residue removed). Microbial carbon use efficiency (CUE) and growth increased linearly over time, with stronger positive warming effects after 5 years under conservation agriculture. According to structural equation models, these increases arose from greater carbon inputs from the crops, which indirectly controlled microbial CUE via changes in fungal communities. As a result, fungal necromass increased from 28 to 53%, emerging as the strongest predictor of SOC content. Collectively, our results demonstrate how management and climatic factors can interact to alter microbial community composition, physiology and functions and, in turn, SOC formation and accrual in croplands.
Soil organic carbon (SOC) stands as a property of ecosystem, offering a wide range of benefits to both human society and the natural environment, including global climate regulation . However, the state of agriculture, which covers 38% of the Earth’s land surface, is a matter of
concern. Large swathes of agricultural land have suffered from moderate to severe degradation due to inappropriate management practices , which has reduced one-half to two-thirds of total SOC content compared with natural or uncultivated soils . Addressing this
issue, there exists significant potential for SOC accrual in croplands through the adoption of ‘climate-smart’ management practices , which is considered a key natural solution for mitigating climate change and ensuring food security, and thereby achieving Sustainable Development Goals . However, it is essential to recognize that SOC is vulnerable to loss due to climate change. Both soil warming experiments and global datasets have provided evidence of increased rates of decomposition under warming . Unfortunately, studies investigating the interactive effects of management and warming on SOC accrual in croplands are extremely scarce. Understanding the interactions between management and warming is critical to identify suitable management practices that retain and augment SOC in face of a changing climate, and to develop effective strategies that increase agricultural resilience as a vital component of ‘climate-smart agriculture’.
Soil disturbance caused by tillage has been a primary driver of historical SOC loss, estimated at C year globally . In general, SOC accumulates after a shift from intensive tillage to conservation agriculture . Conservation agriculture, typically represented by crop residue retention, and no-tillage or reduced tillage , has been proposed as an appropriate option for rebuilding SOC levels, which also provides various benefits for ecosystem multifunctionality . It is estimated that of global arable land has been managed by conservation agriculture approaches . However, the response of SOC to alterations in tillage practices can differ significantly across different regions and over time . Since increasing temperatures are expected to stimulate microbial respiration, soils with higher organic carbon contents are more vulnerable to carbon loss under warming conditions . Soils managed under conservation agriculture, which involve increased retention of organic residue and reduced or zero tillage , should contain more SOC. They may also have greater proportions of relatively loosely protected chemically recalcitrant organic matter pools (e.g., macroaggregates or particulate organic matter), which are more vulnerable to loss under warming . However, a shortterm 3-month laboratory incubation study detected no significant difference in SOC mineralization between conservation and conventional agriculture under various temperature conditions . The information regarding the sensitivity and persistence of SOC rebuilt through conservation agriculture management to long-term climate warming is very limited.
Predicting changes in SOC is highly dependent on microbial acclimation, which involves physiological adjustments that modify microbial carbon use efficiency (CUE) i.e., the proportion of carbon allocated to growth relative to respiratory losses . Microbial CUE has strong implications for SOC storage in soils, as organic carbon converted into microbial biomass and necromass plays a crucial role in long-term SOC stabilization . Microbial CUE depends on both abiotic and biotic factors , which are affected by warming and management practices. Increased temperature can have both direct and indirect effects on microbial CUE. Generally, warming decreases microbial CUE, as a greater proportion of the substrate is reallocated from growth to maintenance metabolism , which alters rates of enzyme-driven processes . Warming can alter CUE indirectly via changes in soil moisture, substrate availability or the composition and/ or structure of microbial communities . Warming may decrease soil moisture and reduce microbial CUE because more substrate is allocated to dissimilatory metabolism, and hence less available for growth . In contrast, warming may enhance plant growth, productivity and rhizosphere carbon input , promoting microbial growth and necromass accumulation . The dominant microbial groups altered by warming could further modify community CUE and consequently . Microbial decomposers with higher CUE can convert substrates to new biomass more efficiently by increasing growth and reducing respiration per unit carbon taken up by microorganisms . Thus, increasing CUE yields more microbial
biomass and byproducts, i.e., ‘necromass’, both of which contribute to the persistent SOC pool . Fungi generally have a higher CUE than bacteria , but they can be negatively affected by tillage . The greater sensitivity of fungi to warming compared to bacteria may increase the contributions of fungal residues to necromass . However, CUE may decrease if fungi allocate metabolic resources to increase exoenzyme production for nutrient acquisition under warming or nutrient limitation . Crop residue retention in conservation agriculture can increase microbial CUE and necromass accumulation by alleviating nutrient limitation, preserving soil moisture, and reducing energy leakage pathways and exoenzyme production . However, our understanding of the intricate connections between microbial physiological and metabolic characteristics and SOC accrual in response to management and warming is primitive, particularly when considering different temporal scales .
Herein, we present the study from a long-term agricultural experiment that spans a decade and encompasses two distinct management systems (conservation versus conventional agriculture) two warming levels (warming versus ambient). Our primary objectives were to elucidate the interactive effects of warming and management on SOC accrual and persistence, as well as identify the temporal microbial attributes underpinning their responses over 10 years. Specifically, our study aimed to: (i) assess whether warming differentially affects SOC accrual under conservation agriculture (chopped crop residues returned and no tillage) versus conventional agriculture (crop residue removed and annual tillage); and (ii) examine the interactive effects of warming and management on the succession of microbial communities and temporal changes of microbial physiological traits (e.g., microbial growth, CUE, microbial necromass carbon, and microbial metabolic functional genes) and evaluate their consequences for SOC formation and accrual. We hypothesized that, under conditions of climate warming, (i) conservation agriculture increases SOC directly through increased plant-derived carbon inputs and indirectly via greater substrate availability to the soil microbial community; (ii) microbial community-level adaptation to warming and higher microbial growth efficiency and metabolic functions in response to larger substrate availability increases the contribution of microbial necromass to SOC over time under conservation agriculture; (iii) the increase in size and accelerated fungal community turnover boost fungal necromass accumulation, thereby promoting SOC formation and persistence over time under conservation agriculture. To test these hypotheses, we measured SOC, bacterial and fungal communities using DNA sequencing, microbial functions using metagenomics, and microbial physiological traits (CUE and microbial necromass) using substrate independent labeling and microbial biomarker (amino sugars) analysis. In our study, long-term warming increased SOC under conservation agriculture, but not under conventional agriculture. This response under conservation agriculture was related to increase in key microbial physiological traits, CUE, growth and fungal necromass, with accelerated fungal community turnover and divergence over the 10 years. Using structural equation models analysis, we find that an increase in carbon input from the crops accelerated fungal succession and enhanced microbial growth efficiencies, leading to a progressive increase of microbially-derived carbon contributions to SOC formation and accrual at decadal timescales under conservation agriculture with warming. Our work demonstrates that agricultural management and climatic factors can interact to alter microbial community composition, physiology and functions and, in turn, SOC formation and accrual in croplands.
Results
Conservation agriculture increased SOC by mediating the effect of warming on soil and plant properties
Warming the field plots under conservation or conventional agriculture was experimentally imposed for 10 years using infrared
Fig. 1 | Effect of warming on plant and soil variables over ten years depending on management systems. a-i Average SOC content, soil temperature and moisture, aboveground biomass, root biomass, root exudation C, DOC and microbial biomass. Data are presented as violin plots with mean values s.e.m. The root biomass and root exudation C input and microbial biomass data were analyzed based on soil sampling in 2020 ( independent soil samples per treatment). The other parameters were average data on six sampling dates over 10 years ( independent soil samples per treatment). Statistical analysis was performed using repeated measures ANOVA analysis. All reported values result from two-sided statistical tests with ; ns nonsignificant. Asterisks indicate significant differences in the warming effect of the individual management system as compared
with their matched no-warming condition. For exact statistical values, see Supplementary Tables 2. Shift in the effect size of warming on SOC over time (2010-2020) for conservation and conventional agriculture. Linear regression model with two-sided test was used for the statistical analysis, and adjusted R-squared was used. Relationships are denoted with solid lines and fit statistics (slopes, and values) for each management practice. The solid line represents the significant linear regression ( ), and the gray shading indicates the confidence intervals. Conserv-Amb conservation agriculture without warming, Conserv-Warm conservation agriculture with warming, Conven-Amb conventional agriculture without warming, Conven-Warm conventional agriculture with warming.
heaters, maintaining the soil temperature at above ambient levels (Supplementary Fig. 1). We first determined the direction of temporal changes in SOC and investigated whether warming differentially affected SOC content under conservation versus conventional agriculture. Over the course of 10 years, conservation management increased SOC content compared to conventional management, regardless of warming treatment. Warming further increased SOC
content under conservation agriculture by 3.1% as compared to the nowarming treatment, but not under conventional agriculture ( ; Fig. 1a; Supplementary Tables 1 and 2). Along with individual effects of management and warming, there were interactive effects of management warming and management warming year on SOC content ( ; Supplementary Table 1), suggesting that the positive stimulatory effect of warming on SOC content was mediated by
management and such effect increased with time. We further assessed the warming effect over 10 years on SOC content using Cohen’s index: SOC content increased linearly with the duration of warming under conservation agriculture, and this increase accelerated after the 5th year ( ; Fig. 1j). In contrast, the warming effect on SOC content under conventional agriculture was not significant (Fig. 1j; Supplementary Table 2). Therefore, our findings partially support the hypothesis (i), suggesting that conservation agriculture under warmer conditions increases SOC content.
The conservation agriculture treatment, characterized by continuous soil cover by crop residues with no tillage, increased SOC, which mitigated the effects of experimental warming on soil temperature and moisture contents. Over the 10 -year study period, we continuously monitored soil temperature and moisture using in-field sensors. We observed that both of these factors were changed by management and experimental warming. As expected, experimental warming increased soil temperature but decreased soil moisture under both conservation and conventional agriculture ( ; Fig. 1b, c; Supplementary Tables 1 and 2). However, the warming effects were modulated by management. Soils covered by crop residues under conservation agriculture were cooler ( vs. ; ; Fig. 1b) and wetter ( vs. ; Fig. 1c) compared with conventional agriculture.
We also evaluated plant carbon input using gravimetric measurements of aboveground biomass and root biomass, and root exudation carbon using a method developed for in situ collection of roots exudates . Warming increased aboveground biomass under conservation agriculture, but not under conventional agriculture ( ; Fig. 1d; Supplementary Table 2), probably due in part to the positive effects of residue retention and no tillage on soil moisture. Warming increased belowground plant carbon inputs, including root biomass and root exudate carbon, in both conservation and conventional agriculture ( ; Fig. 1d, e, f; Supplementary Table 2). Overall, total root carbon input increased by under warming compared with ambient control under conservation agriculture ( ; Supplementary Fig. 2), ultimately contributing to the increase in SOC ( ; Fig. 1a; Supplementary Table 2) and dissolved organic carbon (DOC) ( ; Fig. 1g; Supplementary Table 2) concentrations.
Warming stimulated microbial growth efficiency and fungal necromass carbon accumulation under conservation agriculture
Like SOC content, the effects of management and warming on microbial growth efficiency might change over 10 years. To address this, we measured soil microbial community CUE using the substrateindependent method . Across all years of the study, warming increased microbial CUE by 12%, microbial growth by and carbon uptake by when compared with the ambient control under conservation agriculture ( ; Fig. 2a; Supplementary Tables 1 and 2), indicating an acceleration of microbial turnover including proliferation, growth and death. The interactions between management warming and management warming year on microbial CUE, growth, respiration and carbon uptake ( ; Supplementary Table 1) indicated that warming effects were dependent on both management and time. We further examined the temporal changes of the warming effect on microbial growth efficiency under the two management systems over the 10 -year study period (Fig. 2b). The warming effects on microbial CUE under conservation agriculture increased linearly with experimental duration (Slope=0.83, ; Fig. 2b).
To better understand these effects, we divided the study into early (2010-2015) and later stages (2016-2020) to assess the role of warming duration on the response directions and magnitudes (Supplementary Fig. 3). Shorter and longer-term warming had contrasting
effects on CUE under conservation agriculture: a negative effect size of CUE was observed during the early stage (2010-2015), while a positive effect size was evident in the later stage (2016-2020) ( ; Supplementary Fig. 3a). Consequently, warming increased microbial CUE by 1.1-1.5 times particularly under conservation agriculture during the later stage ( ; Supplementary Fig. 3e). The warming effect on microbial growth and carbon uptake followed a similar pattern to that of CUE, with the difference between warming and no-warming increasing over time in soil under conservation agriculture (Slope= 0.58 and 0.44, all ; Fig. 2b; Supplementary Fig. 3b, d). Conversely, no effects were observed under conventional agriculture (Fig. 2b; Supplementary Fig. 3). The observation that microbial CUE, growth and carbon uptake were enhanced in the later stage of the experiment indicated greater substrate availability, which agrees with hypothesis (i) as plant carbon inputs and SOC content increased more rapidly after the 5th year (Fig. 1j).
The acceleration of microbial turnover in response to warming under conservation agriculture was substantiated by a 77% increase in total microbial necromass carbon (indicated by the concentration of amino sugar biomarkers ) compared with un-warmed soils ( ; Fig. 2c; Supplementary Tables 2 and 3). Warming elevated the contribution of total necromass to SOC during the later stage of the experiment (from 2016 to 2020; ; Supplementary Fig. 4), underscoring the important role of the entombing effect of the microbial carbon pump in the formation of new SOC. In particular, fungal necromass carbon increased linearly with year under conservation agriculture, with the effect size of warming being 19 times greater in 2020 than in 2010 (Slope =1.6, ; Fig. 2d; Supplementary Table 4). However, this effect was not significant under conventional agriculture. A similar temporal pattern was observed for total microbial necromass carbon ( ; Fig. 2d). An increase in the concentration of microbial PLFA biomarkers, which serve as indicators of microbial biomass of dominant groups (Fig. 1h; Supplementary Table 2), also indicated the benefits of conservation management and warming for fungi, as reflected in larger fungal/ bacterial PLFA ratios (Fig. 1i; Supplementary Table 2). The larger fungal biomass consequently led to larger fungal necromass, contributing significantly to total necromass over time ( ; Supplementary Fig. 4) and was further increased by warming (by ; Fig. 2c). Correspondingly, warming increased the contribution of total and fungal necromass to SOC from to , and to , respectively, from 2010 to 2020 under conservation agriculture ( , Supplementary Fig. 4). The increase in microbial growth efficiency and total necromass under conservation agriculture in warmed soils, in parallel with larger plant carbon input and increasing SOC content, suggests that the microbial community and its physiological responses adapted to warming conditions over time, which supports the hypothesis (ii).
Warming accelerated fungal community temporal turnover and its divergence under conservation agriculture
To understand the role of the soil microbiome in driving SOC dynamics in response to warming and management, we examined how microbial diversity and community turnover co-varied over the 10 years using DNA sequencing. Warming altered both microbial communities and individual microorganisms, depending on management. The effects of warming on fungal phylotypes changed with year under conservation agriculture ( ; Supplementary Fig. 5). In the later stage of the experiment from 2016 to 2020, warming decreased the fungal phylotypes by 9.3-12% under conservation agriculture, but had no effect under conventional agriculture ( ; Supplementary Fig. 5). The compositions of soil bacteria and fungi were altered by warming, management and year, as visualized by principal component analysis (Supplementary Fig. 6; Supplementary Tables 5). Three complementary non-parametric multivariate statistical tests (Adonis,
Fig. 2 | Effects of warming on soil microbial CUE, growth, respiration, C uptake and necromass over ten years depending on management systems. a Average microbial CUE, growth, respiration and C uptake on six sampling dates over 10 years. Shifts in the effect size of warming on microbial CUE, growth, respiration and uptake over time (2010-2020) for conservation and conventional agriculture. Average microbial necromass C and shift in the effects of warming on necromass over time (2010-2020) depending on the management systems. Letters indicate significant differences between warmed and control under two management systems. Bars represent mean s.e.m. ( independent soil samples per treatment). Statistical analysis was performed using repeated measures ANOVA
analysis. The two-sided statistical tests indicate significant effects by ; ns, nonsignificant. For exact statistical values, see Supplementary Tables 2 and 4. Relationships are denoted with solid lines and fit statistics (slopes, and values) for each management practice (adjusted r -squared and value are shown). The solid line represents the significant linear regression ( ), and the gray shading indicates the confidence intervals. Conserv-Amb conservation agriculture without warming, Conserv-Warm conservation agriculture with warming, Conven-Amb conventional agriculture without warming, Conven-Warm conventional agriculture with warming.
Anosim and MRPP) confirmed that bacterial and fungal communities across all years were significantly different between the warmed and unwarmed plots under both conservation and conventional agriculture ( ; Supplementary Table 6). Under conservation agriculture, warming had a large negative effect on the relative abundance of Acidobacteria ( ; Fig. 3a), while it increased the relative abundances of Actinobacteria, Firmicutes, Bacteroidota, and Verrucomicrobiota ( to ; Fig. 3a). The relative abundance of Proteobacteria, including Alphaproteobacteria, increased under conventional agriculture with warming ( ; Fig. 3a). Warming had a greater positive effect on the relative abundance of Ascomycota under conservation than conventional agriculture ( vs ; Fig. 3a). Warming effects on the
relative abundance of Ascomycota shifted from negative in the early stage ( ; Supplementary Fig. 7; from 2010 to 2015) to positive in the later stage ( ; from 2016 to 2020). We further investigated different fungal guilds as classified by FUNGuild. While warming marginally increased the relative abundance of Saprotrophs under both management types, the effect was more pronounced under conservation agriculture ( vs 0.055 ; Fig. 3a).
To elucidate the impacts of warming on the temporal turnover of microbial community structure, we assessed the time-decay relationships (TDRs) for bacteria and fungi. The slopes of the TDR values represent the temporal turnover rates of soil microbial communities . Temporal turnover rates of fungi were faster than those of bacteria ( VS 0.72-0.97; ; Fig. 3b). Moreover, the TDR
Fig. 3 | Temporal dynamics of the soil microbiome. a Effect sizes of warming on the relative abundance of major microbial groups based on linear mixed-effects models. Data are presented as mean s.e.m. of the estimated effect sizes. Statistical significance is based on Wald type II X2 tests (two-sided) ( paired soil samples per management); non-significant changes are denoted by gray dots. The time-decay relationships of bacterial and fungal communities for all treatments and (c) temporal changes in microbial community differences between warmed and control soils under conservation and conventional agriculture. Linear regression model with two-sided test was used for the statistical analysis, and adjusted R -squared was used. Relationships are denoted with solid lines and
fit statistics (slopes, and values) for each management practice. The solid line represents the significant linear regression ( ), and the gray shading indicates the 95% confidence intervals. d The warming-induced relative changes of TDRs (v) among phylogenetic lineages under conservation and conventional agriculture. The TDR values are presented as the relative changes of (warmingcontrol)/control. Conserv-Amb: conservation agriculture without warming; Conserv-Warm: conservation agriculture with warming; Conven-Amb: conventional agriculture without warming; Conven-Warm: conventional agriculture with warming.
Fig. 4 | Effect of warming on carbon metabolism-associated functions of soil microorganisms depending on management at early and later stages. a Effect of warming on microbial growth-related pathways under two management systems based on the KEGG database. Only significant changed pathways between warmed and control soils under each management were presented here. Effect of warming on the relative abundance of carbohydrate metabolism genes under two
management systems based on carbohydrate-active enzymes (CAZy). Only significant changed genes were presented here. Data are presented as mean s.e.m. of the estimated effect sizes ( paired soil samples per management). ConservAmb: conservation agriculture without warming; Conserv-Warm: conservation agriculture with warming; Conven-Amb: conventional agriculture without warming; Conven-Warm: conventional agriculture with warming.
slopes were steeper in response to warming under conservation agriculture (pairwise permutation tests; ; Fig. 3b, Supplementary Table 4). To further examine the warming effects on the succession of bacterial and fungal communities over time, we assessed the differences in microbial communities between paired warmed and unwarmed plots under these two managements on a yearly basis. The differences in bacterial and fungal community structures between warming and ambient control increased linearly with time, but only under conservation agriculture ( ; Fig. 3c). In addition, the paired differences in microbial communities between the warmed and control plots under conservation agriculture were more pronounced for fungi (slope = 0.006; Fig. 3c) than for bacteria (slope = 0.028; Fig. 3b), suggesting that warming had a more substantial impact on the temporal turnovers of fungi than bacteria under conservation agriculture, agreeing with the hypothesis (iii) that warmer soils under conservation agriculture accelerated fungal community turnover.
We also observed that fungal groups and community turnover changed in response to warming under conservation agriculture, which were in parallel with changes in microbial growth efficiency and necromass accumulation (Fig. 2). To further explore these dynamics, we evaluated the TDRs of various lineages of bacteria and fungi between warmed and unwarmed plots under both management types (Fig. 3c). The TDRs of fungal phyla were higher than bacterial phyla ( vs. 0.04-0.89; Supplementary Table 7). The TDR slopes for Basidiomycota and Ascomycota communities differed between warming and no-warming under conservation agriculture ( ; Supplementary Tables 7 and 8). Warming accelerated the temporal turnover rates of Ascomycota and Basidiomycota under conservation agriculture, but decreased Firmicutes and Planctomycetota communities ( ; Fig. 3c; Supplementary Table 7). The threshold indicator taxa analysis (TITAN) also showed that Ascomycota and Basidiomycota had large increases over time (Supplementary Fig. 9). Moreover, warming had more pronounced effects on the TDRs of bacterial taxa, including
Firmicutes and Gemmatimonadota under conventional agriculture ( ; Fig. 3c; Supplementary Table 7).
Warming effects on microbial functions related to microbial growth depended on management
Management and warming changed the magnitudes of plant carbon inputs, including crop residues from aboveground biomass, root biomass and root exudates (Fig. 1). These changes had cascading effects on the biomass, activity (Figs. 1h, i, 2), structure and turnover (Fig. 3) of the soil microbial communities, particularly the proliferation of fungi. We further used shotgun metagenomic analysis to determine the changes in microbial functions related to cellular processes and metabolism (Fig. 4a). The relative abundances of pathways involved in the biosynthesis of microbial cell constituents were decreased in response to warming under both conservation and conventional agriculture at the early stage (in 2010) ( ; Fig. 4a). However, these pathways increased under conservation agriculture with warming at the later stage (in 2020), especially for biofilm formation and amino acids biosynthesis, suggesting a potential stimulation of gene activity associated with protein production and cell growth potential ( ; Fig. 4a). To delve deeper into critical processes associated with microbial membrane structure, we further analyzed the response ratio of KOs related to biofilm formation, fatty acid biosynthesis and biosynthesis of amino acids to warming under both management types ( ; Fig. 4b). Similar to the variations in metabolic pathways, the relative abundances of functional genes annotated in biofilm formation showed the greatest positive response to warming under conservation agriculture at the later stage, along with genes associated with fatty acid biosynthesis and biosynthesis of amino acids ( ; Supplementary Fig. 8).
Changes in the genes encoding carbohydrate-active enzymes based on the CAZy database (Fig. 4b) provided insights into how management and warming affected plant carbon inputs, including lignin, cellulose and pectin from plant cell walls, and microbial inputs such as chitin from fungal biomass (indicated by the increase in fungal glucosamine; Fig. 2c). Warming increased the relative abundances of
genes encoding ligninolytic enzymes by 7.8-11%, but decreased those of genes encoding pectin, cellulose and chitin under both conservation and conventional agriculture at the early stage ( ; Fig. 4b). Some distinct responses to warming were found between the management practices at the later stage. Under conservation agriculture, warming increased the relative abundances of genes encoding starchdegrading enzymes by , yet decreased the relative abundances of genes associated with pectin, cellulose, and chitin degradation ( ; Fig. 4b). This suggested that conservation agriculture tends to favor more the retention of recalcitrant carbon (e.g., lignin and chitin) than labile carbon (e.g., starch) under warming, in agreement with the hypothesis (ii). In contrast, warming promoted the relative abundances of genes associated with starch, cellulose, chitin and lignin degradation by , and , respectively, under conventional agriculture in the later stage ( ; Fig. 4b).
SOC accumulation as a function of warming-induced substrate availability, microbial community and physiological traits
Structural equation modeling (SEM) was used to determine the warming-induced abiotic (microbial physiology and diversity) and biotic (substrate availability and microclimate factors) components that drive SOC dynamics. Our results revealed the crucial role of soil microbial physiological traits in mediating SOC dynamics under conservation agriculture. A total of of the SOC variations were accounted for by the combinations of soil temperature, substrate availability (indicated by plant biomass and DOC), microbial alpha diversity, microbial community composition, microbial CUE and necromass carbon (Fig. 5a). Among these factors, the positive direct effect of fungal necromass carbon on SOC was the greatest (Fig. 5b), while microbial CUE were mainly related to indirect effects (Fig. 5b). This corresponded to the strong positive correlation between fungal necromass carbon and SOC ( ; Supplementary Fig. 9a). The direct positive effect of microbial CUE on fungal necromass carbon was the largest (path coefficient ; Fig. 5b): fungal necromass carbon increased at a faster rate with microbial growth than respiration, leading to a significant positive relationship between fungal necromass carbon and CUE ( ; Supplementary Fig. 9b). Substrate availability exerted a strong selection on the fungal community composition through direct effect. Overall, fungal community composition showed positive associations with microbial growth ( ; Fig. 5c; Supplementary Table 9), leading to strong positive direct effects on microbial CUE (path coefficients , Fig. 5a). The relative abundance of dominant fungal phylum Ascomycota showed negative correlations with microbial CUE and growth during early stage, but positive correlations in the later stage ( ; Supplementary Fig. 9d). The relative abundance of fungal phylum Basidiomycota had positive associations with CUE and microbial growth ( ; Fig. 5c; Supplementary Table 9). Moreover, microbial CUE and necromass increased with the relative abundances of genes related to microbial growth potential but decreased with genes encoding carbohydrateactive enzyme degradation ( ; Fig. 5d; Supplementary Table 10).
Discussion
This study investigates the interactive effects of management and warming on SOC accrual and persistence in arable land over a decadelong timescale. Our findings provide empirical evidence that conservation agriculture can increase the accrual and persistence of SOC under predicted climate warming scenarios. SOC dynamics is controlled by the balance between carbon input from plants and decomposition via microbial respiration, both of which are modulated by management and climatic factors . Generally, warming per se is anticipated to cause SOC loss, particularly in soils with high organic carbon contents . Soils managed under long-term conservation agriculture often harbor substantial pools of unprotected organic carbon (e.g., particulate organic matter), which may be more
vulnerable to warming . We did not observe that warming affected the SOC content under conventional agriculture (Fig. 1a), but the relative abundances of microbial genes encoding labile and recalcitrant carbon degradation were increased by warming, suggesting the potential for further SOC loss (Fig. 4b). Though warming increased belowground biomass and root exudation (Fig. 1e-f), these changes were not translated to an increase in SOC under warmed conventional agriculture. Herein, SOC content increased exclusively under conservation agriculture and was further boosted by 10 years of warming (Fig. 1a, j), which can be partially attributed to continuous soil cover that mediated the effects of warming on soil temperature and prevented excessive drying. Importantly, microbial physiological traits and community turnover shifted after the 5th year of warming under conservation agriculture (Figs. 2 and 3; Supplementary Figs. 3 and 4), leading to higher SOC accumulation (Fig. 1j). These findings suggested that more efficient soil microbial carbon cycling underlies the observed SOC accumulation under conservation agriculture in response to warming, especially after 5 years.
Microbial CUE is a crucial metabolic parameter that describes the ratio of anabolic and catabolic processes . Higher microbial CUE in the conservation agriculture with warming treatment, characterized by a greater increase in microbial growth (33%) compared to respiration (13%) over 10 years (Fig. 2a, b and Supplementary Fig. 3), indicates an overall enhancement in microbial growth efficiency. The increase in microbial CUE under conservation agriculture incorporates the responses of maintenance costs and growth rates to warming-induced changes in soil temperature, moisture, substrate availability and microbial communities in this study. The fungal community structure exerted the strongest positive influence on microbial CUE, though the relationship was mainly mediated by substrate availability (Fig. 5a, b). Root biomass and root exudation are major forms of carbon input into soils, which serve as important carbon sources for soil microorganisms. Together with crop residue retention, DOC, aboveground biomass, belowground root biomass and rhizodeposition (Fig. 1d-g) increased under long-term conservation agriculture with warming, indicating higher plant carbon inputs (Fig.1d-f) that provided abundant substrates for microbial proliferation. This particularly favored fungal growth, leading to higher fungal biomass (Fig. 1h) and fungal/bacterial ratios (Fig. 1i) under long-term conservation management with warming. Fungi are sensitive to changes in local climate (e.g., moisture and temperature) and external disturbance . Conservation agriculture promotes fungal abundance by increasing substrate availability, reducing disturbance of soil structure, preserving hyphal networks and improving water availability . Microbial CUE in response to warming increased linearly with time under conservation agriculture, especially during the later stage from 2016 to 2020 (Fig. 2b and Supplementary Fig. 3). This also corresponded to an increase in the relative abundances of microbial functional genes associated with microbial cell growth and protein production potential (e.g., biofilm formation, fatty acid and amino acid biosynthesis; Fig. 4a), with fewer genes related to recalcitrant carbon decomposition under conservation agriculture with warming during the later stage (Fig. 4). This agrees with a long-term warming study in a tallgrass prairie, which reported a shift towards the decomposition of more labile substrates due to increased plant inputs . This is also consistent with the observation that microorganisms may downregulate their protein production machinery in response to warming-induced substrate depletion . Conversely, the utilization efficiency of more recalcitrant substrates was increased at higher temperatures in soils exposed to almost two decades of warming at above ambient at Harvard Forest . Altogether, our results underscore the importance of microbial community structure and substrate availability in driving the responses of microbial metabolic processes to warming under conservation agriculture.
Studies of soil microbial community turnover and microbial physiological traits over time in response to climate change are vital to
response to warming in grassland and a forest . Fungi have a greater carbon demand than bacteria and are usually the first to utilize belowground plant carbon inputs . Consequently, the rapid fungal growth and fungal community changes, driven by increased plant carbon input, are key drivers of temporal community turnovers under conservation agriculture with warming. Larger substrate availability due to increased plant carbon input had a direct effect on fungal community composition, as supported by the observed changes in fungal lineage (Fig. 5a-c). Among various fungal taxa, Ascomycota, previously reported to increase with warming , explained the largest variance in total fungal communities (Supplementary Fig. 6). As copiotrophic fungi, Ascomycota are generally -strategists that efficiently utilize labile carbon sources for rapid metabolism, growth and propagation . In response to increased substrate availability, these fungi may have a shorter generation time, leading to more generation per unit time . Therefore, we assume that warming played a role in driving fungal temporal divergence, as evidenced by increasing differences between warmed and un-warmed plots under conservation agriculture (Fig. 3b). In contrast, Acidobacteria explained the largest variance for bacterial communities (Supplementary Fig. 6). This group is characterized by oligotrophic traits, meaning they thrive in lownutrient environments and are closely associated with decreases in net nitrogen mineralization due to the reduced substrate availability caused by soil drying . Therefore, the slower temporal turnover of bacterial communities may be attributed to a large portion of inactive (dormant or slow-growing) bacteria in warmer and drier soils . These results suggest that microbial physiological adjustments that selected microbial lineages were better adapted to changing soil microclimatic conditions and substrate availability over time.
The evolving theory of soil organic matter formation emphasizes microbial necromass production and mineral-matrix interactions as dominant mechanisms for SOC formation and stabilization . Microbial necromass accounts for more than 50% of SOC, which is protected either physically within soil aggregates or chemically through its association with soil minerals . Fungal necromass carbon increased with warming and accounted for the largest variations in SOC accumulation under conservation agriculture: 36% of total SOC was derived from microbial necromass (amino sugars), with of fungal origin (Figs. 3c, d and 5a). These findings support the concept that fungal necromass is a critical driver of stable SOC accrual . Microbial necromass accumulation in soil is driven by plant carbon input and mediated by microbial necromass production and stabilization . Accordingly, the fungal community was positively correlated with microbial CUE, which had the largest positive effects on fungal necromass (Fig. 5a, b). These results suggested that warming increased microbial growth efficiency and fungal community turnover due to greater plant carbon input, which ultimately increased the supply of microbial residues contributing to SOC formation. In addition, the adoption of conservation agriculture reduces soil disturbance and promotes aggregate formation and stability, providing a conducive environment for the preferential accumulation of fungal-derived necromass in macroaggregates . Overall, our results suggested that warming enhances SOC formation and accumulation under conservation agriculture by increasing microbial growth efficiency via accelerating fungal community turnover, as well as via increased fungal necromass carbon accumulation (Figs. 2 and 3). In summary, this study provided empirical evidence for the mechanisms that enable the increase in SOC accrual and persistence under warming and conservation agriculture. As illustrated by Fig. 6, by carrying out a longterm in situ management warming field experiment, we revealed a gradual acceleration of SOC accrual in response to climate warming under conservation agriculture. Our research highlights the significant roles of altered plant carbon input and microclimate conditions in driving microbial responses to warming under conservation
agriculture. In particular, under long-term conservation agriculture with warming, the increase in plant carbon input accelerated fungal succession and enhanced microbial growth efficiencies, leading to a progressive increase of microbially-derived carbon contributions to SOC formation and accrual at decadal timescales. If our findings can be generalized to other systems and regions where water does not limit productivity (e.g., irrigated regions), then we propose that regenerative management can promote effective carbon sequestration under warming, which increases agricultural resilience as a vital component of ‘climate-smart agriculture’. However, more research is needed to understand the combined effects of management and climate on SOC accrual and persistence in arable soils across different regions with a wide precipitation range, which could guide climate-smart land use practices to sequester carbon effectively and optimize crop production in the face of a changing climate.
Methods
Experimental site and design
A long-term field experiment with a double-cropped winter wheat (Triticum aestivum L.)-summer maize (Zea mays L.) system was started in 2003 at the Yucheng Comprehensive Experiment Station in North China ( ), which belongs to the Chinese Academy of Science (CAS). The region has a temperate semi-arid climate with an annual mean temperature of , and annual mean precipitation of 561 mm . The soil has a silt loam texture with sand, silt, clay, and a mean pH of 7.1. The soil type is Calcaric Fluvisol according to the FAO-UNESCO system.
Four treatments were laid out in a randomized complete block design with four replicates: conservation agriculture with and without warming (Conserv-Amb, Conserv-Warm) and conventional agriculture with and without warming (Conven-Amb, Conven-Warm). Full details of the experimental design and management are provided by ref. 73. In brief, the conventional and conservation agriculture treatments were established in 2003. Two levels of warming (ambient and ) were imposed on both conservation and conventional agriculture since 2010. An infrared heater (MSR-2420 infrared heater, Kalglo Electronics Inc, Bethlehem, PA) was suspended approximately 3 m above the ground in each warmed plot to achieve a surface soil warming of , which was projected by IPCC greenhouse gas scenarios rates for northern China.
The size of each plot was . Winter wheat was seeded in early October and harvested in early June. The summer maize was grown in June and harvested at the end of September. In the conventional agriculture treatment, the residues were removed after the maize harvest. Cultivation with a rotary tiller to a depth of fully incorporated the remaining stubble into the soil. In the conservation agriculture treatment, all residues were chopped to approximately 5 cm in length and retained on the soil surface. All other management procedures were the same for both management types.
Field measurements, soil sampling and analyses
Soil temperature at 5 cm depth was monitored with PT 100 thermocouple. Volumetric soil moisture at depth was measured by FDS100 soil moisture sensors (Unism Technologies Incorporated, Beijing). From 2010 to 2020, the living aboveground biomass was harvested ( ) at each plot in May every year. The aboveground plant biomass samples were dried at until a constant weight was obtained. Measurement of root biomass and root exudation C was conducted from October 2019 to May 2020 during winter wheat growth seasons according to ref. 52. (details below).
Soil samples ( depth) were collected every 2 years after winter wheat harvest from 2010 to 2020. Five topsoil cores taken randomly from each plot were composited as one sample per plot. The composited soil samples were then passed through a 2 mm sieve to remove visible roots and gravel. The sample was divided into 3 sub-
Fig. 6 | A conceptual diagram illustrating the impact of warming on SOC formation and accrual and the underlying microbial mechanisms through plant inputs, community succession, physiology adjustment and necromass production under conservation agriculture over decadal timescales. The main processes include (1) substrate availability increased due to plant C input (e.g., crop residues from aboveground biomass, root biomass and root exudates) with warming; (2-3) the above processes worked as ‘trigger’ for driving microbial community succession (observed by high temporal scaling rates and increasingly
divergent succession for fungal community) and microbial physiological adjustments (e.g., high CUE and growth efficiency). In parallel, the altered microbial metabolism-associated functional genes were observed indicated by an increased relative abundance of genes involved in microbial growth potentials and labile C degradation, but decreased genes encoding recalcitrant C degradation; (4) gradually strengthened microbial carbon pump observed as microbial necromass (especially fungal necromass) enters the carbon pools and drive new carbon formation and accrual.
samples for analyses. The first sub-sample was air-dried at room temperature, the second sub-sample was stored at , and the third subsample was stored in a freezer at . SOC, total nitrogen (TN), and DOC were measured after sample collection in 2010, 2012, 2014, 2014, 2018 and 2020. The SOC and TN contents were determined by combustion of air-dried soils using a Vario EL III Elemental Analyzer (Elementar). DOC concentration was measured in fresh soils stored at following the method of Jones and Willett .
Microbial CUE was measured in 2021 in all soils (sampled in 2010, 2012, 2014, 2016, 2018 and 2020) that had been stored at using the tracer method after 7 days preincubation (details below). Analyses of bacterial and fungal diversity were performed on three of the four replicates samples that had been stored at in 2021 . The stored soil samples ( ) collected in 2010 and 2020 were further subjected to metagenomic sequencing. Fresh soils sampled in 2020 were analyzed for microbial community composition using phospholipid fatty acid (PLFA) analysis according to ref. 75; changes in microbial community composition were presented as molar percentages (mole %) of the PLFA biomarkers for bacteria or fungi. Amino sugars were extracted from air-dried soils and determined following the method of Zhang and Amelung (details below).
DNA extraction and amplicon sequencing
DNA was extracted from 0.25 g of soil using the PowerSoil Isolation kit (MoBio Laboratories, Carlsbad, CA, USA) according to the manufacturer’s instructions. The quality of the purified DNA was assessed based on the and absorbance ratios obtained, using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, USA). The DNA was stored at until sequencing analysis.
The V4-V5 region of 16S rRNA and the internal transcribed spacer (ITS) region of the rRNA were amplified to construct bacterial and fungal community profiles, respectively, using high-throughput sequencing. Universal primer sets, F515 (5″-GTGCCAGCMGCCGCGC3′) and R907 ( -CCGTCAATTCMTTTRAGTTT-3′) targeting the bacterial 16 S rRNA genes and gITS7F ( -GTGARTCATCGARTCTTTG-3′) and ITS4R ( -TCCTCCGCTTATTGATATGC-3) for fungal ITS, were used . The following thermal program was used for the amplification of the 16 S rRNA gene: initial denaturation at for 3 min , followed by 27 cycles of denaturing at for 30 s , annealing at for 30 s and extension at for 45 s , then a single extension at for 10 min and ending at . The PCR amplification of the ITS2 rRNA gene was performed as follows: initial denaturation at for 5 min , followed by 30 cycles of denaturing at for 30 s , annealing at
for 30 s and extension at for 30 s , and single extension at for 10 min and ending at . Purified amplicons were pooled in equimolar and paired-end sequenced on an Illumina Nova6000 platform (Illumina, San Diego, USA) according to the standard protocols by Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, China).
The raw sequences were subjected to quality control with the following criteria: (i) the 300 bp reads were truncated at any site receiving an average quality score of over a 50 bp sliding window, and the truncated reads shorter than 50 bp were discarded, and reads containing ambiguous characters were also discarded; (ii) only overlapping sequences longer than 10 bp were assembled according to their overlapped sequence. The maximum mismatch ratio of the overlap region was 0.2 . Reads that could not be assembled were discarded; (iii) samples were distinguished according to the barcode and primers, and the sequence direction was adjusted, exact barcode matching, and 2 nucleotide mismatches in primer matching. Phylotypes (i.e., Amplicon Sequencing Variants, ASVs) were picked using UNOISE3 with default parameters in USEARCH and were identified at the identity level . The taxonomy of each ASV representative sequence was analyzed by the RDP Classifier Bayesian algorithm against the Silva database (https://www.arb-silva.de/) and the UNITE database (https://unite.ut.ee/) using a confidence threshold of 0.7. We used a randomly selected subset of 24,579 and 21,550 sequences per sample for subsequent bacterial and fungal communities’ analysis.
Metagenomic sequencing and data analyses
DNA extract was fragmented to an average size of about 400 bp using Covaris M220 (Gene Company Limited, China) for paired-end library construction. The paired-end library was constructed using NEXTFLEX Rapid DNA-Seq (Bioo Scientific, Austin, TX, USA). Adapters containing the full complement of sequencing primer hybridization sites were ligated to the blunt end of fragments. Paired-end sequencing was performed on Illumina NovaSeq (Illumina Inc., San Diego, CA, USA) at Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China).
The paired-end Illumina reads were trimmed of adaptors, and lowquality reads (length or with a quality value or having N bases) were removed by fastp (https://github.com/OpenGene/fastp, version 0.20.0). Metagenomics data were assembled using MEGAHIT (https://github.com/voutcn/megahit, version 1.1.2). Contigs with a length bp were selected as the final assembling result, and then the contigs were used for further gene prediction and annotation. Representative sequences of non-redundant genes were aligned to the KEGG database (http://www.diamondsearch.org/index.php, version 0.8.35) against the Kyoto Encyclopedia of Genes and Genomes database (http:// www.genome.jp/keeg/). Carbohydrate-active enzyme annotation was conducted using hmmscan (http://hmmer.janelia.org/search/hmmscan) against the CAZy database (http://www.cazy.org/). We used the RPM (reads per million) method to normalize the relative abundance of pathways and genes annotated within the KEGG and CAZy database .
Microbial carbon use efficiency
Microbial CUE was measured using the tracer method on soils previously stored at according to ref. 81 . Soil samples were adjusted to field water capacity with Millipore water (replenished according to weight loss every 3 days) and pre-incubated for 7 days at in tubes covered by vented sealing films to avoid excessive accumulation. After the pre-incubation, 1 g pre-incubated soil was transferred to two 2 mL Falcon tubes ( 0.5 g soil in each tube). One sample was amended with O-enriched water to achieve 30 atom% O-labeled soil water, and the other was amended with the same volume of non-labeled Millipore water as a natural abundance control. All samples were then incubated for 48 h at in the dark at field water capacity. The produced during this time was measured using an Agilent 7890 A gas chromatograph (Agilent Technologies, Aito Palo, CA) equipped with a TCD detector. Soil DNA was extracted using the
PowerSoil Isolation kit (MoBio Laboratories, Carlsbad, CA, USA). The extracted DNA ( ) was dried in silver capsules at for 2 days. Subsequently, the abundance and total O content were determined using an elemental analyzer (FLASH 2000, Thermo Fisher Scientific, Cambridge, UK) coupled with an isotope ratio mass spectrometer (IRMS) system (ConFlo VI interface and MAT253 IRMS, Thermo Scientific, Bremen, Germany) at the Chinese Academy of Sciences Institute of Subtropical Agriculture, (Changsha, Hunan Province, China).
Microbial CUE was calculated based on the following equation:
Microbial necromass analysis
Glucosamine, galactosamine and muramic acid were used as biomarkers for microbial residues (“necromass”). They were extracted and determined following the method of Zhang and Amelung . Airdried soil samples ( ) were hydrolyzed with 6 M HCl at for 8 h after adding myo-inositol (internal standard). The derivatized compounds were separated on a gas chromatograph equipped with an HP-5 column and quantified using a flame ionization detector (Agilent 6890 A, Agilent Technologies, Littleton, CO, USA). Bacterial and fungal residue (necromass) C were calculated based on the following equations:
Fungal residue mmol glucosamine mmol muramic acid
where 45 is the conversion value to the bacterial residue; 179.2 is the molecular weight of glucosamine; and 9 is the conversion factor of fungal glucosamine to the fungal residue. The total microbial residue was estimated as the sum of fungal and bacterial residues.
Root biomass and exudation C input
Root biomass and root exudation C input were measured from October 2019 to May 2020 during winter wheat growth. During the elongation, flowering and maturation stages of wheat, we collected root exudates from six individual plants in four subplots of each treatment. We used a modified culture-based cuvette system developed especially for root exudates in situ field collection . Firstly, wheat roots with the same height and growth status were selected, and terminal fine roots were carefully excavated from the topsoil ( ) by hand and extensively washed to remove adhering soil particles. Secondly, the intact roots were placed into cuvettes, and then cuvettes were filled with glass beads (c. 1 mm diameter), which were placed to simulate soil particles, and sealed with a special rubber septum. Thirdly, the cuvettes (including the controls with beads only) were covered in foil and reburied in the excavated area in soil. After a one-day equilibration period, a small amount of fresh C-free nutrient solution ( ) was flushed through each cuvette to remove soluble C. Finally, the exudates were collected by flushing the cuvette three times with 15 ml of the fresh nutrient solution after the 24 -h incubation. The control cuvettes filled with only coarse silica sand were similarly placed and processed at each plot. The trap solutions were filtered through sterile syringe filters within of collection and then stored at until further analysis. Exudates were collected for three consecutive days. Total nonparticulate organic C accumulated in the trap solutions was analyzed using a Multi 3100 N/C TOC analyzer (Analytik, Jena, Germany).
Root exudation rates for each treatment were calculated by subtracting the C accumulation within the root-filled cuvettes from the C
detected in the root-free control and were then expressed as root biomass . We estimated the whole growth period root exudation rates by weighting root exudation rates in each growth period. We calculated yearly root exudates by multiplying the growth period living root biomass with the weighted root exudation rate and the number of hours per growth period .
The following equation to assess the whole growth period C exudation of wheat root (c)
where is the TOC concentration difference between root exudate solution and blank control ( ); is the volume of root exudate solution (L); is the root weight (g); is the collection time (h); is root biomass ( ); 24 is 24 h per day; and represents the growth period.
Statistical analyses
Mixed models for repeated measures analysis of variance (ANOVAs) were used to examine the effects of management, warming, year and their interactions on SOC, bacteria and fungi diversity, microbial physiology and necromass. In addition, multiple comparisons (LSD) were used to examine the differences in different parameters among treatments. Linear regressions were used to detect linear trends of SOC, microbial physiological traits and necromass C with time under four treatments. The slopes of those linear relationships were analyzed and compared by Standardized Major Axis regression analysis using the smatr package in R v.3.2.1 . Cohen’s was further calculated as an estimate of multiple-treatment effect sizes on SOC, microbial physiology and necromass values’ response to warming under conservation and conventional agriculture by comparing them against the no warming control; positive values indicate that the response variables in the treatment have a larger value than in the control, and vice versa . The effect size analyses were performed in the software v.3.2.1 with the package effsize.
Principal component analysis was used to assess changes in the bacterial and fungal communities. Three non-parametric multivariate statistical tests: nonparametric multivariate analysis of variance (ADONIS), analysis of similarity (ANOSIM) and multi-response permutation procedure (MRPP), were used to test the differences in soil microbial communities under warming and control treatments. These analyses were performed in R v.3.2.1 with the vegan package. The influence of treatments on the relative abundance of bacterial and fungal taxa was evaluated based on linear mixed-effect models following the methods of ref. 84. Statistical significance is based on Wald type II tests and all estimated effect sizes are based on rescaled response variables. These analyses were performed in v.3.2.1 with the lme4 and car packages.
The time-decay relationships (TDR) of bacterial and fungal communities were evaluated using the linear regression between logarithmic similarities and logarithmic temporal distance. The moving window approach was used to assess time decay in microbial communities as previously described , which is currently the dominant approach for TDRs. In our biennial surveyed bacterial and fungal diversity data of six sampling dates over 10 years, subset window 1 included the pairwise similarity of samples that were two years apart; subset window 2 is the pairwise similarity of samples four years apart, and so on. So, in this tenyear record, there are 5 two-year intervals, 4 four-year intervals, 3 sixyear intervals, 2 eight-year intervals, and 1 ten-year intervals for each plot. Considering the repeated measures design, TDR analysis counted only pairwise comparisons among time points within each plot (that is, 15 pairwise comparisons for each plot and a total of 45 pairwise comparisons for each treatment). We further evaluated the impact of warming on the succession of soil bacterial and fungal communities using the distance of microbial communities between warming and
control at each block in each year for conservation and conventional agriculture, respectively following the method of ref. 57.
The response ratio of carbon metabolism-associated functions of microorganisms between warming and un-warming plots at conservation and conventional agriculture was calculated according to Curtis and Wang . The means of the treatment and control group were used to compute a response ratio using:
The natural was used for statistical tests. If and are normally distributed and both are greater than zero, is approximately normally distributed with a mean equal to the true response ratio.
SEM analysis was applied to investigate the direct and indirect effects of soil microclimate (soil temperature and moisture), aboveground biomass, DOC and microbial variables (microbial alpha diversity, community structure, microbial CUE and necromass) on SOC. We first considered a hypothesized conceptual model (Supplementary Fig. 10) that included all reasonable pathways. Temperature was only selected to improve model fit because we observed strong collinearity between soil temperature and moisture (Pearson’s ). We sequentially eliminated non-significant pathways unless the pathways were biologically informative, or added pathways based on the residual correlations. A total of 72 independent soil samples were used to run the SEM. The overall goodness of fit of the SEM results was evaluated by the Chisquare test ( X 2 ) and the goodness fit index (GFI). When and GFI , the model has a good fit. The SEM analysis was performed using AMOS 23.0 (AMOS Development Corporation).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The DNA sequences of the 16 S rRNA gene and ITS amplicons in this study have been deposited in the National Center for Biotechnology Information (NCBI) under project accession numbers PRJNA903096 and PRJNA903090. Raw shotgun metagenomic sequences in this study have been deposited in the National Center for Biotechnology Information (NCBI) under project accession PRJNA1007786. Silva database is available at https://www.arb-silva.de/. UNITE database is available at https://unite.ut.ee/. Source data are provided in this paper. Source data are provided with this paper.
Schmidt, M. W. et al. Persistence of soil organic matter as an ecosystem property. Nature 478, 49-56 (2011).
Lehmann, J. & Kleber, M. The contentious nature of soil organic matter. Nature 528, 60-68 (2015).
Sanderman, J., Hengl, T. & Fiske, G. J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. USA 114, 9575-9580 (2017).
Lal, R., Follett, R. F., Stewart, B. A. & Kimble, J. M. Soil carbon sequestration to mitigate climate change and advance food security. Soil Sci. 172, 943-956 (2007).
Paustian, K. et al. Climate-smart soils. Nature 532, 49-57 (2016).
Bossio, D. et al. The role of soil carbon in natural climate solutions. Nat. Sustain. 3, 391-398 (2020).
Davidson, E. A. & Janssens, I. A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440, 165-173 (2006).
Crowther, T. W. et al. Quantifying global soil carbon losses in response to warming. Nature 540, 104-108 (2016).
Chappell, A., Baldock, J. & Sanderman, J. The global significance of omitting soil erosion from soil organic carbon cycling schemes. Nat. Clim. Change 6, 187-191 (2016).
Powlson, D. S. et al. Limited potential of no-till agriculture for climate change mitigation. Nat. Clim. Change 4, 678-683 (2014).
Kan, Z. R. et al. Mechanisms of soil organic carbon stability and its response to no-till: a global synthesis and perspective. Glob. Change Biol. 28, 693-710 (2022).
Pittelkow, C. M. et al. Productivity limits and potentials of the principles of conservation agriculture. Nature 517, 365-368 (2015).
Neufeldt, H., Kissinger, G. & Alcamo, J. No-till agriculture and climate change mitigation. Nat. Clim. Change 5, 488-489 (2015).
Wittwer, R. A. et al. Organic and conservation agriculture promote ecosystem multifunctionality. Sci. Adv. 7, eabg6995 (2021).
Qi, J.-Y. et al. Effects of long-term tillage regimes on the vertical distribution of soil iron/aluminum oxides and carbon decomposition in rice paddies. Sci. Total Environ. 776, 145797 (2021).
Karhu, K. et al. Temperature sensitivity of soil respiration rates enhanced by microbial community response. Nature 513, 81-84 (2014).
Lugato, E., Lavallee, J. M., Haddix, M. L., Panagos, P. & Cotrufo, M. F. Different climate sensitivity of particulate and mineral-associated soil organic matter. Nat. Geosci. 14, 295-300 (2021).
Kpemoua, T. P. et al. Are carbon-storing soils more sensitive to climate change? A laboratory evaluation for agricultural temperate soils. Soil Biol. Biochem. 183, 109043 (2023).
Allison, S. D., Wallenstein, M. D. & Bradford, M. A. Soil-carbon response to warming dependent on microbial physiology. Nat. Geosci. 3, 336-340 (2010).
Frey, S. D., Lee, J., Melillo, J. M. & Six, J. The temperature response of soil microbial efficiency and its feedback to climate. Nat. Clim. Change 3, 395-398 (2013).
Hagerty, S. B. et al. Accelerated microbial turnover but constant growth efficiency with warming in soil. Nat. Clim. Change 4, 903-906 (2014).
García-Palacios, P. et al. Evidence for large microbial-mediated losses of soil carbon under anthropogenic warming.Nat. Rev. Earth Environ. 2, 507-517 (2021).
Malik, A. A. et al. Land use driven change in soil ph affects microbial carbon cycling processes. Nat. Commun. 9, 3591 (2018).
Tao, F. et al. Microbial carbon use efficiency promotes global soil carbon storage. Nature 618, 981-985 (2023).
Cotrufo, M. F., Wallenstein, M. D., Boot, C. M., Denef, K. & Paul, E. The microbial efficiency-matrix stabilization (mems) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob. Change Biol. 19, 988-995 (2013).
Liang, C., Schimel, J. P. & Jastrow, J. D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2, 17105 (2017).
Domeignoz-Horta, L. A. et al. Microbial diversity drives carbon use efficiency in a model soil. Nat. Commun. 11, 3684 (2020).
Tian, W. et al. Thermal adaptation occurs in the respiration and growth of widely distributed bacteria. Glob. Change Biol. 28, 2820-2829 (2022).
Alster, C. J., Robinson, J. M., Arcus, V. L. & Schipper, L. A. Assessing thermal acclimation of soil microbial respiration using macromolecular rate theory. Biogeochemistry 158, 131-141 (2022).
Söllinger, A. et al. Down-regulation of the bacterial protein biosynthesis machinery in response to weeks, years, and decades of soil warming. Sci. Adv. 8, eabm3230 (2022).
Domeignoz-Horta, L. A. et al. Substrate availability and not thermal acclimation controls microbial temperature sensitivity response to long-term warming. Glob. Change Biol. 29, 1574-1590 (2023).
Li, J. et al. Reduced carbon use efficiency and increased microbial turnover with soil warming. Glob. Change Biol. 25, 900-910 (2019).
Wang, N. et al. Effects of climate warming on carbon fluxes in grasslands-a global meta-analysis. Glob. Change Biol. 25, 1839-1851 (2019).
Chang, R. et al. Soil organic carbon becomes newer under warming at a permafrost site on the tibetan plateau. Soil Biol. Biochem. 152, 108074 (2021).
Feng, X. & Wang, S. Plant influences on soil microbial carbon pump efficiency. Glob. Change Biol. 29, 3854-3856 (2023).
Dove, N. C., Torn, M. S., Hart, S. C. & Tas, N. Metabolic capabilities mute positive response to direct and indirect impacts of warming throughout the soil profile. Nat. Commun. 12, 2089 (2021).
Nottingham, A. T. et al. Microbial responses to warming enhance soil carbon loss following translocation across a tropical forest elevation gradient. Ecol. Lett. 22, 1889-1899 (2019).
Six, J., Frey, S. D., Thiet, R. K. & Batten, K. M. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 70, 555-569 (2006).
Buckeridge, K. M., Creamer, C. & Whitaker, J. Deconstructing the microbial necromass continuum to inform soil carbon sequestration. Funct. Ecol. 36, 1396-1410 (2022).
Malik, A. A. et al. Soil fungal: bacterial ratios are linked to altered carbon cycling. Front. Microbiol. 7, 1247 (2016).
Strickland, M. S. & Rousk, J. Considering fungal: bacterial dominance in soils-methods, controls, and ecosystem implications. Soil Biol. Biochem. 42, 1385-1395 (2010).
Yang, Y. et al. Fungi determine increased soil organic carbon more than bacteria through their necromass inputs in conservation tillage croplands. Soil Biol. Biochem. 167, 108587 (2022).
Jansson, J. K. & Hofmockel, K. S. Soil microbiomes and climate change. Nat. Rev. Microbiol. 18, 35-46 (2020).
Ding, X. L. et al. Warming increases microbial residue contribution to soil organic carbon in an alpine meadow. Soil Biol. Biochem. 135, 13-19 (2019).
Ullah, M. R., Carrillo, Y. & Dijkstra, F. A. Drought-induced and seasonal variation in carbon use efficiency is associated with fungi: bacteria ratio and enzyme production in a grassland ecosystem. Soil Biol. Biochem. 155, 108159 (2021).
Tian, J. et al. Microbial metabolic response to winter warming stabilizes soil carbon. Glob. Change Biol. 27, 2011-2028 (2021).
Sauvadet, M. et al. High carbon use efficiency and low priming effect promote soil c stabilization under reduced tillage. Soil Biol. Biochem. 123, 64-73 (2018).
Zhou, R. et al. Microbial necromass in cropland soils: a global metaanalysis of management effects. Glob. Change Biol. 29, 1998-2014 (2023).
Sinsabaugh, R. L., Manzoni, S., Moorhead, D. L. & Richter, A. Carbon use efficiency of microbial communities: Stoichiometry, methodology and modelling. Ecol. Lett. 16, 930-939 (2013).
Bradford, M. A. et al. Managing uncertainty in soil carbon feedbacks to climate change. Nat. Clim. Change 6, 751-758 (2016).
Zhang, Q., Qin, W., Feng, J. & Zhu, B. Responses of soil microbial carbon use efficiency to warming: review and prospects.Soil Ecol. Lett. 4, 307-318 (2022).
Phillips, R. P., Erlitz, Y., Bier, R. & Bernhardt, E. S. New approach for capturing soluble root exudates in forest soils. Funct. Ecol. 22, 990-999 (2008).
Yin, H. et al. Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming. Glob. Change Biol. 19, 2158-2167 (2013).
Spohn, M., Klaus, K., Wanek, W. & Richter, A. Microbial carbon use efficiency and biomass turnover times depending on soil depth-implications for carbon cycling. Soil Biol. Biochem. 96, 74-81 (2016).
Zheng, Q. et al. Growth explains microbial carbon use efficiency across soils differing in land use and geology. Soil Biol. Biochem. 128, 45-55 (2019).
Zhang, X. & Amelung, W. Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils. Soil Biol. Biochem. 28, 1201-1206 (1996).
Guo, X. et al. Climate warming leads to divergent succession of grassland microbial communities. Nat. Clim. Change 8, 813-818 (2018).
Paustian, K. et al. Quantifying carbon for agricultural soil management: From the current status toward a global soil information system. Carbon Manag. 10, 567-587 (2019).
Kivlin, S. N. & Hawkes, C. V. Spatial and temporal turnover of soil microbial communities is not linked to function in a primary tropical forest. Ecology 101, e02985 (2020).
Zhang, B. et al. Soil microbial community dynamics over a maize (zea mays l.) growing season under conventional- and no-tillage practices in a rainfed agroecosystem. Soil. Res. 124, 153-160 (2012).
Zhou, J. et al. Microbial mediation of carbon-cycle feedbacks to climate warming. Nat. Clim. Change 2, 106-110 (2011).
Averill, C., Cates, L. L., Dietze, M. C. & Bhatnagar, J. M. Spatial vs. Temporal controls over soil fungal community similarity at continental and global scales. ISME J. 13, 2082-2093 (2019).
Koch, B. J. et al. Estimating taxon-specific population dynamics in diverse microbial communities. Ecosphere 9, e02090 (2018).
Martinović, T. et al. Temporal turnover of the soil microbiome composition is guild-specific. Ecol. Lett. 24, 2726-2738 (2021).
Asemaninejad, A., Thorn, R. G., Branfireun, B. A. & Lindo, Z. Climate change favours specific fungal communities in boreal peatlands. Soil Biol. Biochem. 120, 28-36 (2018).
Ling, N., Wang, T. & Kuzyakov, Y. Microbiome of the rhizosphere: from structure to functions. Nat. Commun. 13, 836 (2022).
Deng, Y. et al. Elevated carbon dioxide accelerates the spatial turnover of soil microbial communities. Glob. Change Biol. 22, 957-964 (2015).
Fierer, N., Bradford, M. A. & Jackson, R. B. Toward an ecological classification of soil bacteria. Ecology 88, 1354-1364 (2007).
Lennon, J. T. & Jones, S. E. Microbial seed banks: the ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 9, 119-130 (2011).
Liang, C., Amelung, W., Lehmann, J. & Kastner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Glob. Change Biol. 25, 3578-3590 (2019).
Hannula, S. & Morriën, E. Will fungi solve the carbon dilemma? Geoderma 413, 115767 (2022).
Ding, X. L., Liang, C., Zhang, B., Yuan, Y. R. & Han, X. Z. Higher rates of manure application lead to greater accumulation of both fungal and bacterial residues in macroaggregates of a clay soil. Soil Biol. Biochem. 84, 137-146 (2015).
Hou, R., Ouyang, Z., Wilson, G. V., Li, Y. & Li, H. Response of carbon dioxide emissions to warming under no-till and conventional till systems. Soil Sci. Soc. Am. J. 78, 280-289 (2013).
Jones, D. L. & Willett, V. B. Experimental evaluation of methods to quantify dissolved organic nitrogen (don) and dissolved organic carbon (doc) in soil. Soil Biol. Biochem. 38, 991-999 (2006).
Frostegård, Å., Tunlid, A. & Bååth, E. Microbial biomass measured as total lipid phosphate in soils of different organic content. J. Microbiol. Methods 14, 151-163 (1991).
Yusoff, M. Z. M. et al. Influence of pretreated activated sludge for electricity generation in microbial fuel cell application. Bioresour. Technol. 145, 90-96 (2013).
Edgar, R. C., Haas, B. J., Clemente, J. C., Quince, C. & Knight, R. Uchime improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194-2200 (2011).
Li, D., Liu, C.-M., Luo, R., Sadakane, K. & Lam, T.-W. Megahit: an ultrafast single-node solution for large and complex metagenomics assembly via succinct de bruijn graph. Bioinformatics 31, 1674-1676 (2015).
Buchfink, B., Xie, C. & Huson, D. H. Fast and sensitive protein alignment using diamond. Nat. Methods 12, 59-60 (2015).
Morse, N. et al. Plant-microbe interactions drive denitrification rates, dissolved nitrogen Removal, and the abundance of denitrification genes in stormwater control measures. Environ. Sci. Technol. 52, 9320-9329 (2018).
Schroeder, J., Kammann, L., Helfrich, M., Tebbe, C. C. & Poeplau, C. Impact of common sample pre-treatments on key soil microbial properties. Soil Biol. Biochem 160, 108321 (2018).
Shen, X., Yang, F., Xiao, C. & Zhou, Y. Increased contribution of root exudates to soil carbon input during grassland degradation. Soil Biol. Biochem. 146, 107817 (2020).
Warton, D. I., Wright, I. J., Falster, D. S. & Westoby, M. Bivariate linefitting methods for allometry. Biol. Rev. 81, 259-291 (2006).
Wu, L. et al. Reduction of microbial diversity in grassland soil is driven by long-term climate warming. Nat. Microbiol. 7, 1054-1062 (2022).
Curtis, P. S. & Wang, X. A meta-analysis of elevated effects on woody plant mass, form, and physiology. Oecologia 113, 299-313 (1998).
Acknowledgements
This study was supported by the the National Natural Science Foundation of China (grant nos. 32071629, 32071607), National Key R&D Program of China (2022YFD1901300), 2115 Talent Development Program of China Agricultural University and Beijing Advanced Disciplines and Strategic Priority Research Program of the Chinese Academy of Sciences (XDA28130301). The manuscript revised by Y.K. was also supported by the RUDN University Strategic Academic Leadership Program.
Author contributions
All authors contributed intellectual input and assistance to this study and manuscript preparation. J.T., J.A.J.D., F.Z., M.F.C., and J.Z. designed the original concept and experiment strategy. J.T. performed the lab experiments and collected the data. R.H. carried out the field experiment and collected the data. J.T. analyzed the data with help of Y.D. J.T. drafted the manuscript with help from J.A.J.D., Y.Y., I.P.H., M.F.C., Y.K. and J.Z. All authors contributed to the article and approved the submitted versions.
Competing interests
The authors declare no competing interests.
Additional information
Supplementary information The online version contains
supplementary material available at https://doi.org/10.1038/s41467-023-44647-4.
Correspondence and requests for materials should be addressed to Jing Tian, Fusuo Zhang, M. Francesca Cotrufo or Jizhong Zhou.
Peer review information Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Reprints and permissions information is available at
State Key Laboratory of Nutrient Use and Management, College of Resources and Environmental Sciences, China Agricultural University, 100193 Beijing, PR China. Geography, Faculty of Environment, Science and Economy, University of Exeter, Rennes Drive, Exeter EX4 4RJ, UK. Carbon Management Centre, SRUC-Scotland’s Rural College, Edinburgh EH9 3JG, UK. Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences (CAS), 100101 Beijing, PR China. CAS Key Laboratory for Environmental Biotechnology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, 100085 Beijing, PR China. State Key Joint Laboratory of Environment Simulation and Pollution Control, School of Environment, Tsinghua University, Beijing, PR China. Department of Soil Science of Temperate Ecosystems, University of Göttingen, 37077 Göttingen, Germany. Department of Soil and Crop Science, Colorado State University, Fort Collins, CO, USA. Institute for Environmental Genomics, University of Oklahoma, Norman, OK, USA. School of Biological Sciences, University of Oklahoma, Norman, OK, USA. School of Civil Engineering and Environmental Sciences, University of Oklahoma, Norman, OK, USA. School of Computer Science, University of Oklahoma, Norman, OK, USA. Earth and Environmental Sciences, Lawrence Berkeley National Laboratory, Berkeley, CA, USA. These authors contributed equally: Jing Tian, Jennifer A. J. Dungait, Ruixing Hou, Ye Deng. e-mail: tianj@cau.edu.cn; zhangfs@cau.edu.cn; Francesca.Cotrufo@colostate.edu; jzhou@ou.edu