DOI: https://doi.org/10.1007/s42773-023-00296-w
تاريخ النشر: 2024-01-08
تطبيقات الفحم الحيوي والأسمدة العضوية تعزز وفرة الميكروبات الوظيفية في التربة وتعدد وظائف النظام الزراعي البيئي
الملخص
يتم دعم استخدام الفحم الحيوي والأسمدة العضوية على نطاق واسع للحفاظ على إنتاج المحاصيل والتنمية المستدامة للأنظمة الزراعية. ومع ذلك، لا يزال من غير الواضح كيف ينظم الفحم الحيوي والأسمدة العضوية بمفردهما أو معًا الميكروبيومات الوظيفية للتربة وعلاقتها بتعدد وظائف النظام البيئي (EMF). هنا، تم استخدام تجربة ميدانية طويلة الأمد (بدأت في عام 2013) تحتوي على خمسة علاجات للتسميد لاستكشاف تأثيرات تطبيقات الفحم الحيوي والأسمدة العضوية على EMF (استنادًا إلى 18 مؤشرًا وظيفيًا لإنتاج المحاصيل، وتوريد العناصر الغذائية للتربة، ودورات العناصر، والكتلة الحيوية الميكروبية) والميكروبيومات الوظيفية للتربة السائبة وتربة الجذور [مع تطبيع وفرة 64 جينًا مرتبطًا بدورات الكربون (C) والنيتروجين (N) والفوسفور (P) والكبريت (S)]. بالمقارنة مع التسميد الكيميائي الفردي، زادت مدخلات الفحم الحيوي والأسمدة العضوية بشكل كبير من معظم وظائف النظام البيئي الفردية، وبشكل خاص، زادت EMF بشكل كبير بنسبة 18.7-30.1٪؛ كما زادت تطبيقات الفحم الحيوي والأسمدة العضوية بشكل كبير من وفرة الأنواع الميكروبية الوظيفية للتربة المرتبطة بدورات C-N-P-S بدرجات متفاوتة. أظهرت التطبيق المشترك للفحم الحيوي والأسمدة العضوية تحسينًا أفضل في هذه المؤشرات مقارنة باستخدامها بشكل فردي. أظهرت معظم المجموعات الميكروبية الوظيفية في التربة، وخاصة الأنواع المعنية بتفكيك الكربون، والنيترة، وتقليل النترات، وتعدين الفوسفور العضوي، ودورة الكبريت، ارتباطات إيجابية كبيرة مع EMF عند مستويات عتبة مختلفة، والتي تم تنظيمها في النهاية بواسطة درجة حموضة التربة وتوافر العناصر الغذائية. تسلط هذه النتائج الضوء على الروابط القوية بين الميكروبيومات التربة ووظائف النظام الزراعي، بالإضافة إلى تقديم دعم علمي لإدراج الفحم الحيوي في الإنتاج الزراعي والخدمات مع التعديلات العضوية.
أهم النقاط
- أظهرت الأدلة الميدانية على مدى 8 سنوات تأثيرات الفحم الحيوي وروث الخنازير على الميكروبيوم الوظيفي للتربة ووظائف النظام البيئي.
- أدت مدخلات الفحم الحيوي وروث الخنازير إلى تحسين ملحوظ في معظم وظائف النظام البيئي الفردية وزادت EMF بنسبة 18.7-30.1%.
4. كانت الزيادة في القوة الدافعة الكهربائية مرتبطة بعمليات التربة المدفوعة بالميكروبات مثل تحلل الكربون، النترجة، وتعدين الفوسفور العضوي.
5. يمكن أن يؤدي إدخال الفحم الحيوي في إنتاج المحاصيل مع التعديلات العضوية إلى تعزيز وظائف وخدمات النظام البيئي الزراعي.
الملخص الرسومي
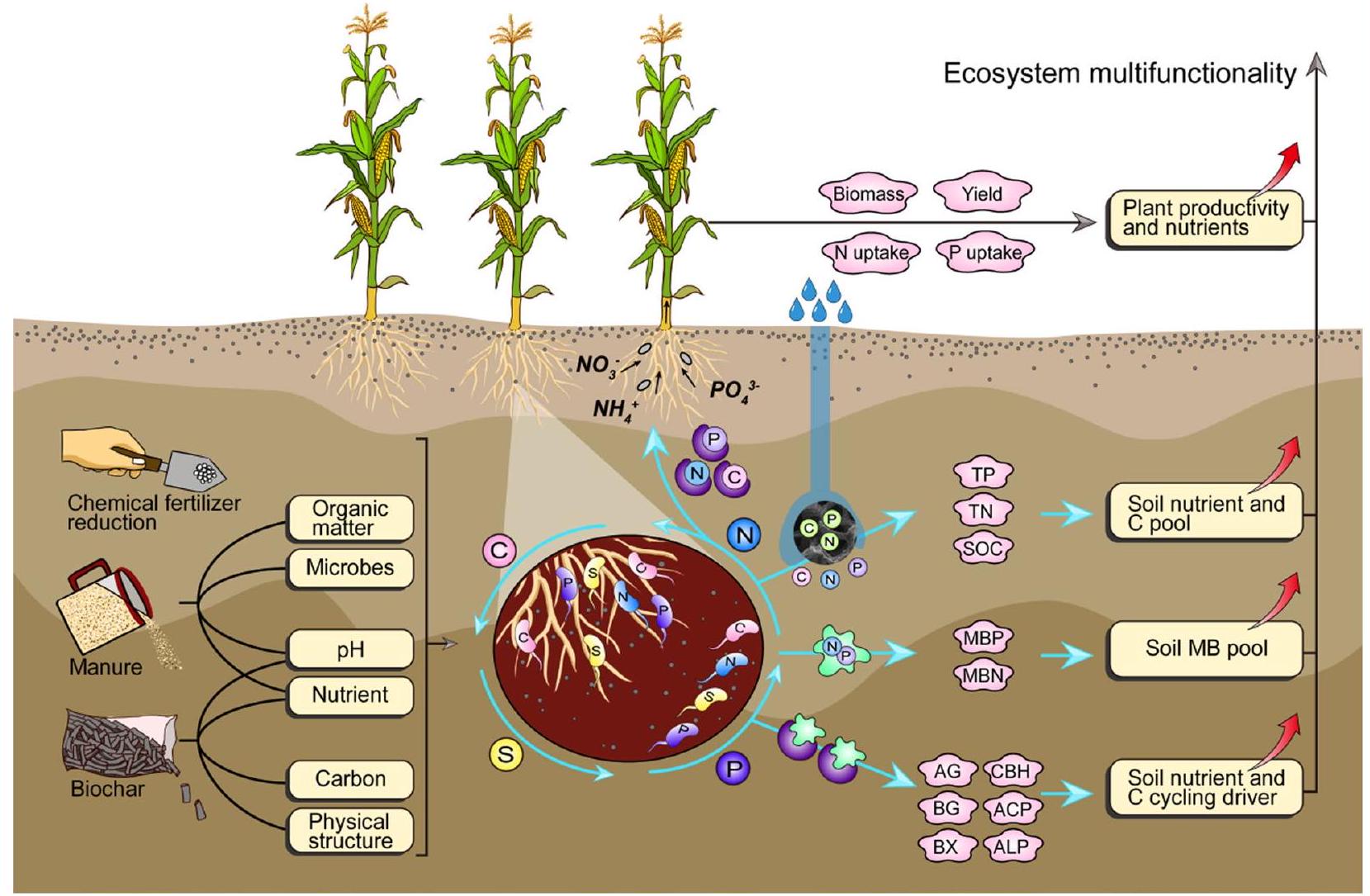
1 المقدمة
المغذيات المتاحة لتعزيز الأنشطة والعمليات الميكروبية في التربة، ثم الحفاظ على خدمات النظام البيئي أو تعزيزها (هان وآخرون 2022؛ وو وآخرون 2021). بالإضافة إلى الأسمدة العضوية، تم استخدام الفحم الحيوي على نطاق واسع كمعالج للتربة أو مادة عضوية في إنتاج المحاصيل وتحسين التربة في السنوات الأخيرة. يمكن أن يحسن الهيكل المسامي الغني، والمغذيات، والكربون (C) في الفحم الحيوي من بنية التربة وإمدادات المغذيات ويزيد من تخزين الكربون في التربة (ليو وآخرون 2023؛ باي وآخرون 2022). وبالتالي، قد تكون مكاسب خصوبة التربة وإنتاجية المحاصيل أكبر عند خلط الفحم الحيوي مع الأسمدة العضوية من
مع التطبيقات الفردية (Brtnicky et al. 2019؛ Bai et al. 2022؛ Liu et al. 2022a؛ Yan et al. 2023). ومع ذلك، فإن الدراسات الحالية حول تأثيرات تطبيقها المشترك على الوظائف المتعددة (المعروفة باسم “التعددية الوظائف”) للنظم البيئية الزراعية غير كافية.
تتعلق الخدمات أو الوظائف التي تقدمها النظم البيئية الزراعية ارتباطًا وثيقًا بتنوع ووظيفة المجتمعات الميكروبية في التربة (زابالا وآخرون 2021). تشارك الميكروبات التربة في عمليات وظيفية متنوعة مثل دورة المغذيات، وكبح الأمراض، والإنتاج الأولي، وبالتالي تلعب أنواع الميكروبات الخاصة في التربة دورًا حيويًا في EMF (سينغ وآخرون 2021؛ هان وآخرون 2022). على سبيل المثال، يمكن أن تدفع الأنواع الميكروبية المرتبطة بدورة الكربون تحلل المواد العضوية وتؤثر على تخزين الكربون في التربة، مما ينظم توازن الكربون في النظام البيئي (باسيلي-دولش وآخرون 2020). يمكن للبكتيريا المثبتة للنيتروجين أن تثبت النيتروجين (N) من الغلاف الجوي إلى الأمونيا لامتصاص النباتات (آسفار وآخرون 2021). كما تؤثر بعض الأنواع الوظيفية أيضًا على دورة النيتروجين في النظام البيئي من خلال تحفيز عمليات نترت التربة ونزع النيتروجين (داي وآخرون 2020). الفوسفور
قد تؤدي التغيرات في ممارسات الإدارة مثل استخدام الأراضي والتسميد إلى تغيير قدرة الكائنات الدقيقة في التربة على الحفاظ على الوظائف المتعددة للنظم البيئية فوق وتحت الأرض (Delgado-Baquerizo et al. 2016). قد يكون لأي فقدان في تنوع الكائنات الدقيقة في التربة تأثير سلبي على وظائف النظام البيئي (Delgado-Baquerizo et al. 2020). لذلك، يمكن أن يؤثر وضع مجموعات الكائنات الدقيقة الوظيفية الخاصة في التربة على EMF واستقراره (Luo et al. 2018, 2019). تُستخدم نشاط إنزيمات التربة، ومحتوى المواد العضوية والمواد الغذائية، وامتصاص النباتات للمواد الغذائية والإنتاجية، والكتلة الحيوية الميكروبية بشكل واسع كمؤشرات لتقييم وظائف وخدمات النظام البيئي (Delgado-Baquerizo et al. 2016؛ Luo et al. 2018؛ Zak et al. 2019). يمكن أن تؤثر الكائنات الدقيقة الوظيفية على عمليات كيميائية حيوية مختلفة في التربة، ويمكن أن تؤدي عمليات ميكروبية متنوعة تحدث في التربة بشكل مباشر أو غير مباشر.
تنظم البيئات الحيوية فوق الأرض (سينغ وآخرون 2021). لذلك، فإن تقييم كيفية ظهور التآزر أو التبادلات بين وظائف النظام البيئي مهم لفهم تنفيذ EMF (يوان وآخرون 2020). يتم قياس مؤشر EMF بشكل رئيسي من خلال طريقة القيمة المتوسطة وطريقة العتبة المتعددة، حيث يتم استخدام كلاهما معًا لتقييم تأثيرات تنوع التربة على EMF بشكل أفضل (لي وآخرون 2020؛ ليو وآخرون 2019ب). وبالتالي، يمكن أن يوضح التقييم متعدد الأبعاد بشكل فعال كيف تنظم إضافات الفحم الحيوي والأسمدة العضوية وظائف النظام البيئي وعلاقاتها مع الميكروبات الوظيفية الخاصة.
2 المواد والأساليب
2.1 تصميم التجربة وإدارة الحقل
2.2 جمع عينات التربة والنبات
تم أخذ عينات من التربة باستخدام طريقة النقاط الخمس وطريقة الاهتزاز. تم حفر خمس كتل من التربة تحتوي على أنظمة جذرية سليمة من كل قطعة أرض وإعادتها إلى المختبر على الفور. تم هز كميات كبيرة من التربة التي لا تحتوي على جذور بلطف أولاً، ثم تمت إزالة التربة الملتصقة بمحاذاة الجذر باستخدام سكين لتستخدم كتربة أساسية. وتمت إزالة التربة الملتصقة بمحاذاة الجذر باستخدام فرشاة لتستخدم كتربة ريزوسفير. تم خلط خمس عينات من التربة من كل قطعة أرض في جزء واحد ثم تم تخزينها للتحليل اللاحق.
2.3 تحديد الخصائص الفيزيائية والكيميائية للتربة
تم حساب محتوى (DOP) كـ TDP-
2.4 تقييم تعددية وظائف النظام البيئي
2.5 تقدير وفرة الجينات الوظيفية
2.6 تحليل البيانات
تم فك رموز المؤشرات الرئيسية لمؤشر EMF من خلال تحليل الغابات العشوائية الذي اقترحه بريمان (2001). أضافت خوارزميات الغابات العشوائية طبقة إضافية من العشوائية إلى تقنية التجميع، واستخدمت كل شجرة عينة مختلفة من البيانات. تم استخدام حزم “randomForest” و “rfPermute” لتنفيذ وتقدير أهمية مقاييس الأهمية لنموذج الغابات العشوائية من خلال تبديل المتغير الاستجابي. تم تقديم تفاصيل حول عملية التحليل في منشورات سابقة (Luo et al. 2019, 2020). تم تطوير نموذج المعادلات الهيكلية من خلال تحليل خصائص التربة وكثافات الميكروبات الوظيفية لتحديد المحركات الرئيسية والمسارات لمؤشر EMF. تم وضع نموذج أولي للارتباطات المحتملة المعروفة بين EMF و
تم تطوير السائقين المذكورين أعلاه على أساس استخراج البيانات باستخدام تحليل الغابة العشوائية. تم استخدام برنامج AMOS (SPSS AMOS 20.0.0) لبناء وتقييم النموذج. تم استخدام المقاييس التالية لتقييم جودة ملاءمة نموذجنا: جذر متوسط مربع خطأ الاقتراب (RMSEA)، قيمة كاي تربيع، إحصائية فيشر P، ومعيار معلومات أكايك (AIC). في النهاية، تم تحسين النموذج الأولي للحصول على النموذج الذي يلبي شرط جودة الملاءمة، وتم التحقق من دلالة المسار بشكل إضافي.
3 نتائج
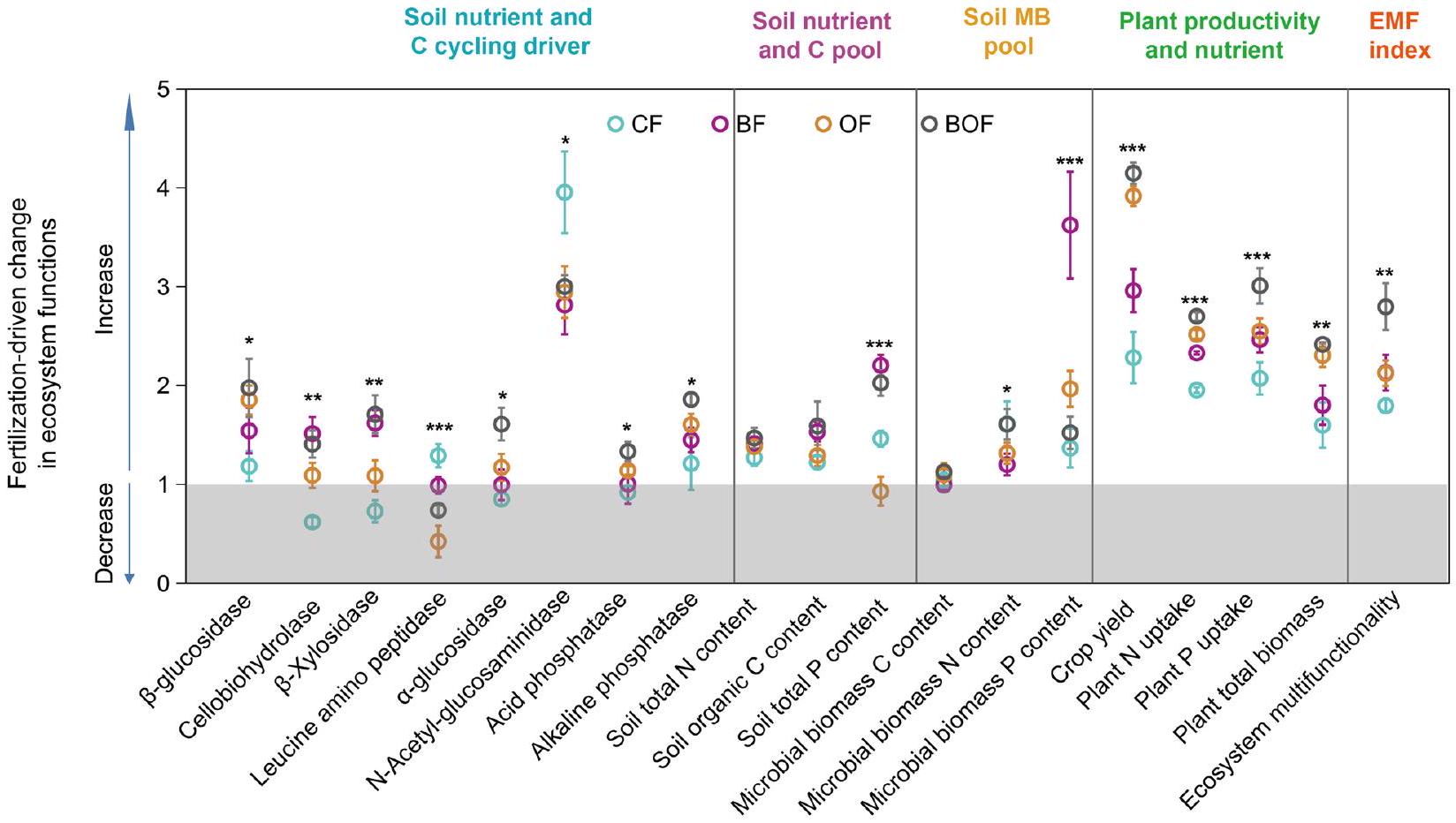
3.1 تأثيرات أنظمة التسميد على وظائف النظام البيئي
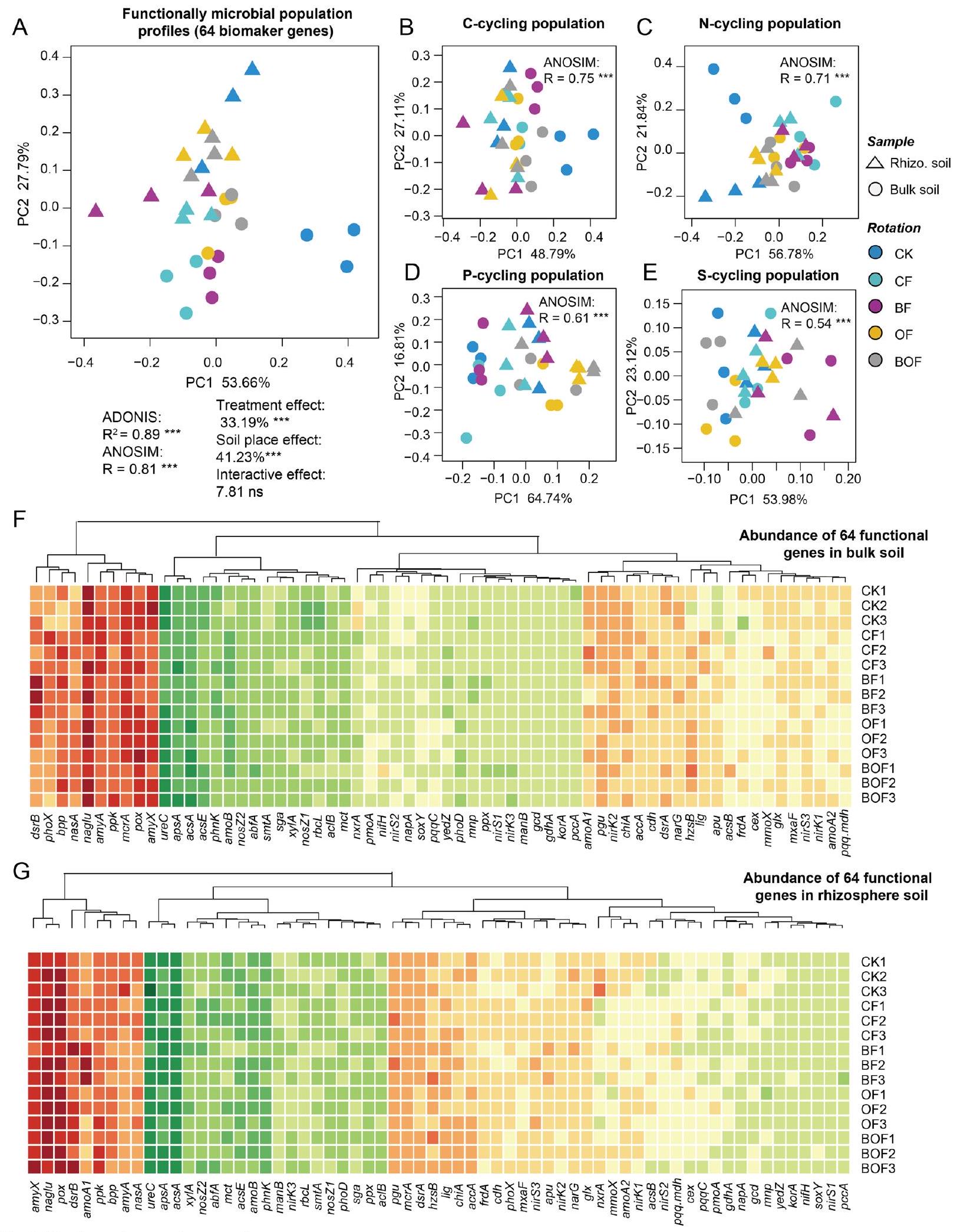
3.2 تأثيرات التعديلات العضوية على وفرة الميكروبات الوظيفية في التربة
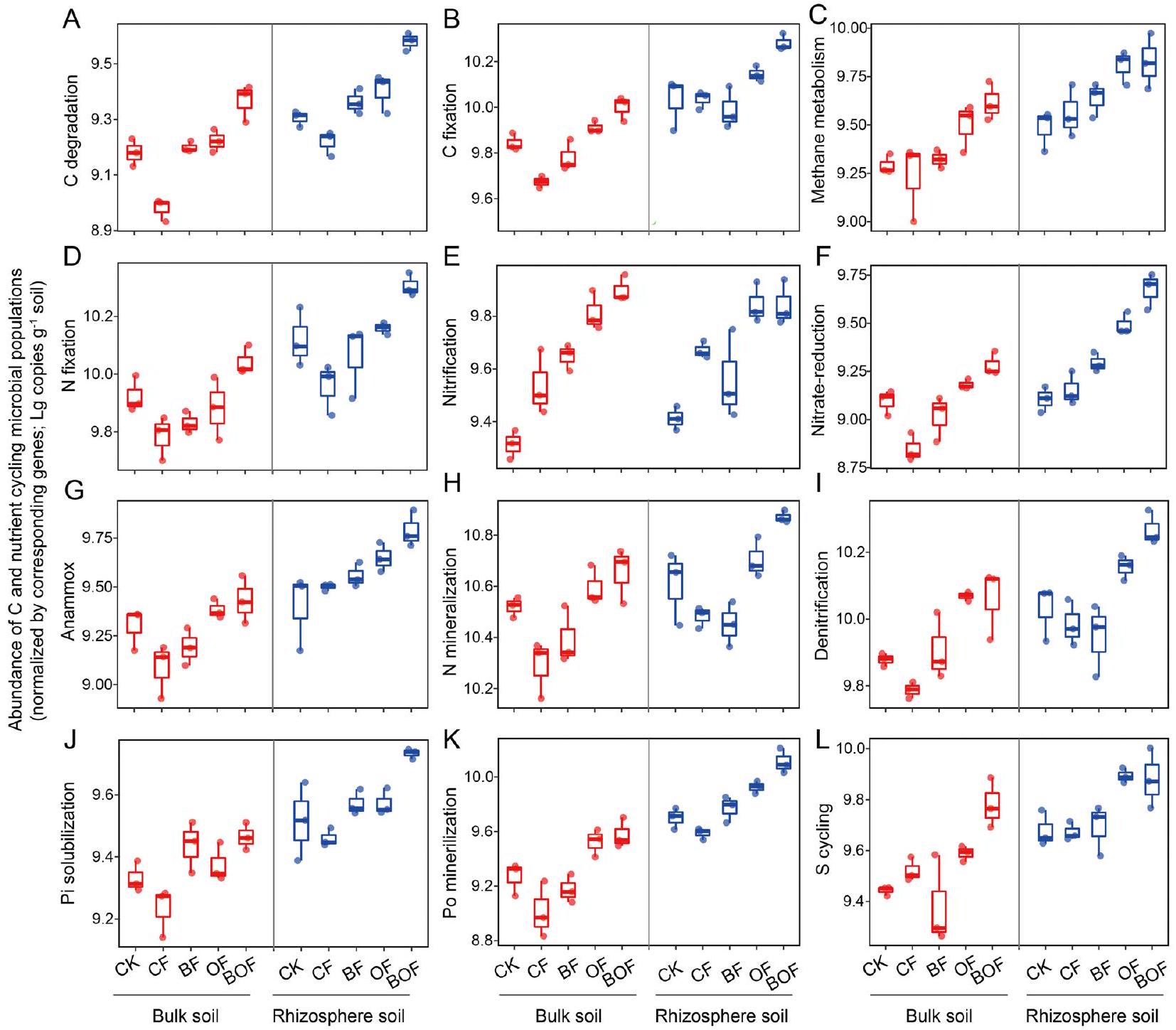
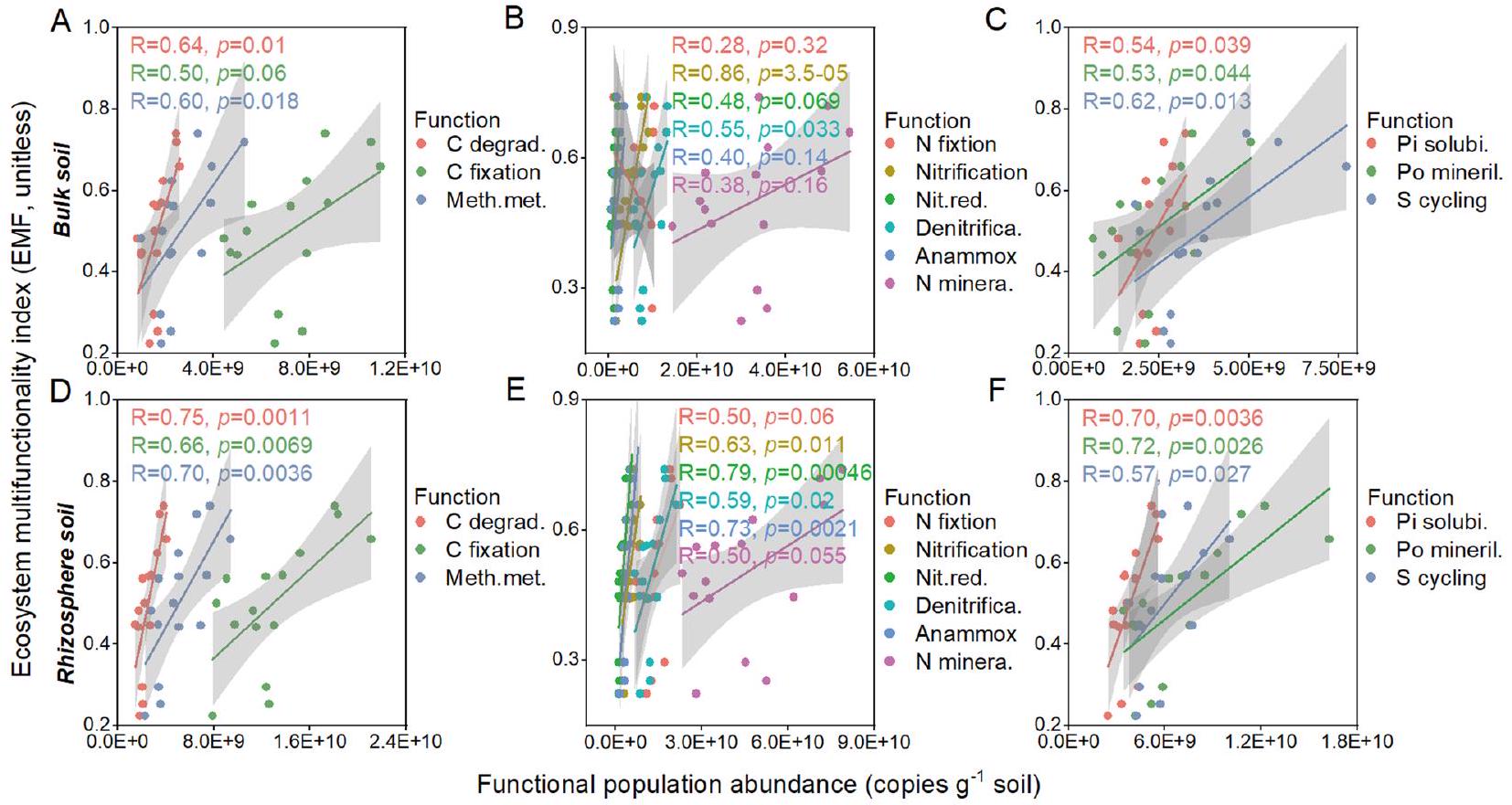
3.3 العلاقة بين وظائف النظام البيئي وكثافات الميكروبات الوظيفية
3.4 العلاقة بين القوة الدافعة الكهربائية وكثافة الميكروبات الوظيفية في التربة
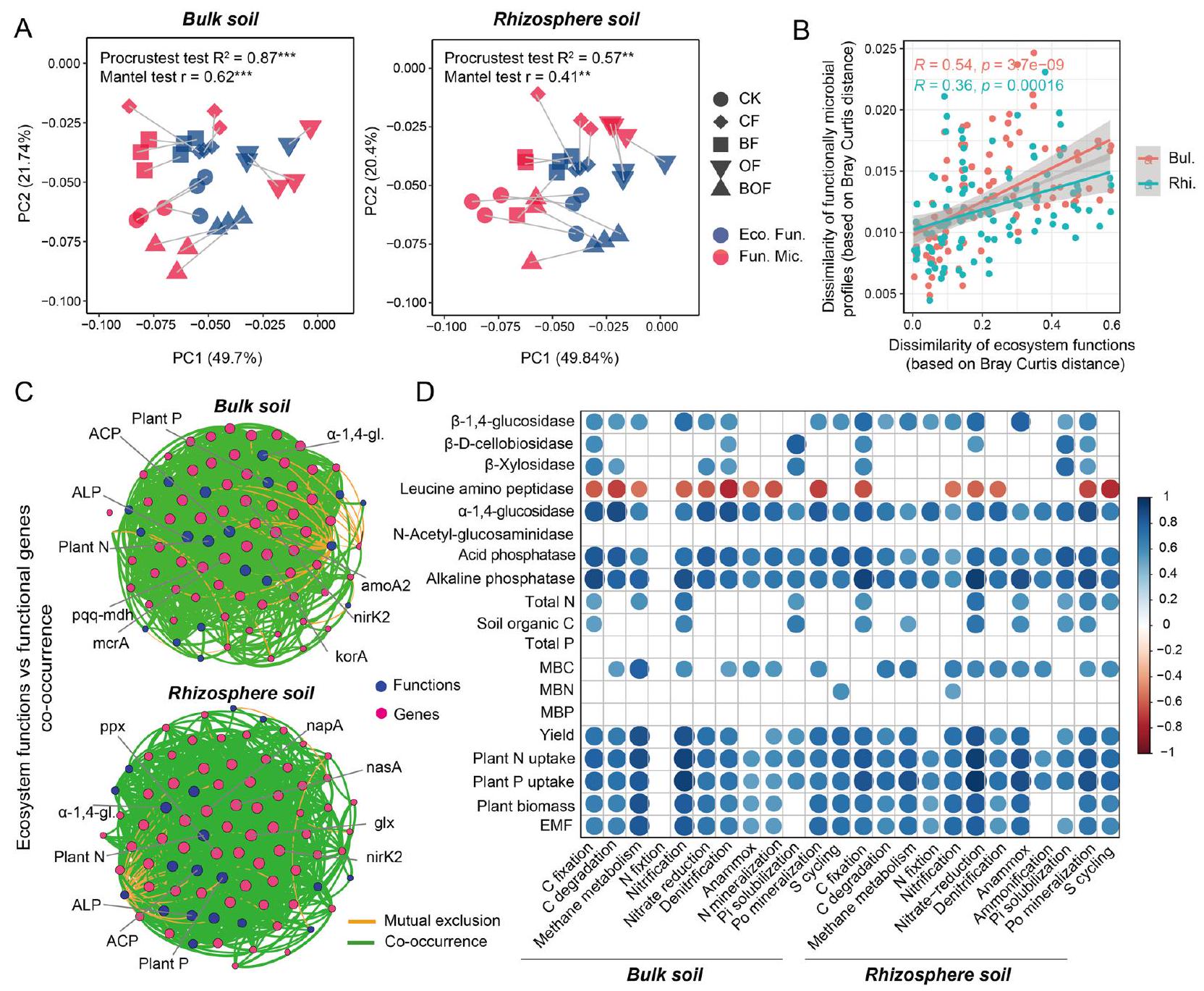
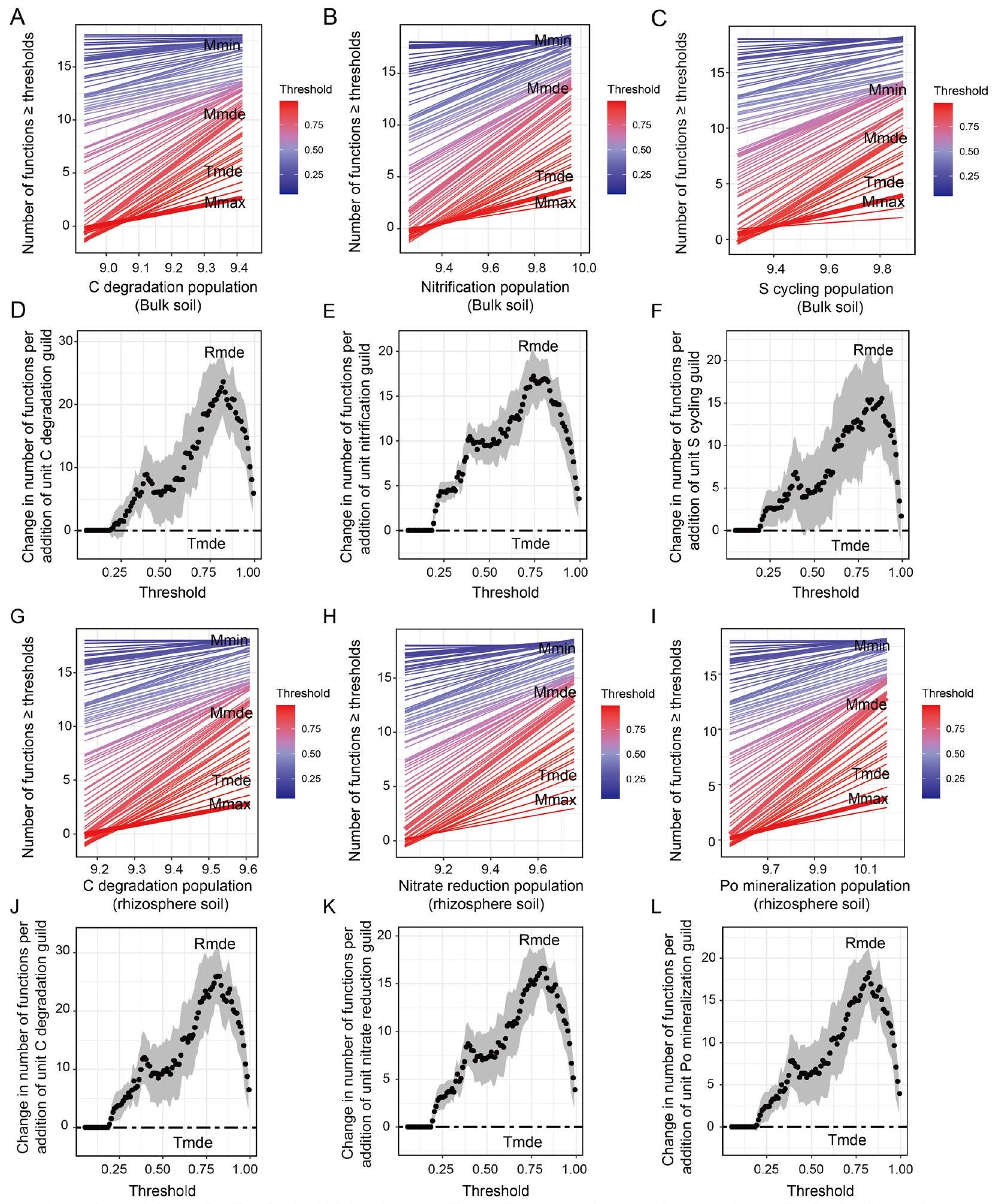
كانت وفرة الأنواع الميكروبية المرتبطة بتفكك الكربون، وتقليل النترات، وتحرير الفوسفور في تربة الجذور من المتنبئات المهمة للـ EMF.
العلاقات الانحدارية بين وفرة الميكروبات المتعلقة بتفكك الكربون، وتقليل النترات، وتعدين الفوسفات العضوي وEMF عند كل عتبة، كانت Tmin هي
3.5 تنظيم وفرة الميكروبات الوظيفية في التربة على EMF
أظهرت وفرة الأنواع الميكروبية في التربة المرتبطة بتدهور الكربون أكبر تأثير مباشر على الفطريات الميكورية، تليها توافر التربة.
4 المناقشة
4.1 تطبيقات الفحم الحيوي والأسمدة العضوية تحسن وظائف النظام البيئي
تأثير ترويجي أكبر على الحفاظ على توازن إمدادات المغذيات في التربة ومتطلبات المغذيات للنبات (كاترر وآخرون 2019). ومع ذلك، لا يمكن فصل تحلل المادة العضوية وتحولات المغذيات في التربة عن تنظيم إنزيمات التربة خارج الخلوية (دوتانيا وآخرون 2019).
كعلامات على وظائف النظام البيئي، تشارك أنشطة إنزيمات التربة في عمليات وظيفية متعددة في التربة (وانغ وآخرون 2021). الفحم الحيوي والأسمدة العضوية
يمكن أن تعزز التعديلات أنشطة إنزيمات التربة المرتبطة بدورات الكربون والمواد الغذائية (الشكل 1). يمكن أن يعزز الفحم الحيوي والأسمدة العضوية تخزين الكربون في التربة وتوافره الحيوي، مما يحفز نمو الميكروبات وتخليق الإنزيمات المرتبطة بتفكك الكربون (دومينشين وآخرون 2021؛ زو وآخرون 2020). وهذا ما تؤكده نتائجنا أيضًا (الشكل 1). يمكن أن يزيد الفحم الحيوي والمواد العضوية من درجة حموضة التربة ويحسن من تكيف بعض إنزيمات التربة مثل العضوية.
4.2 تعديلات الفحم الحيوي والأسمدة العضوية تنظم الميكروبات الوظيفية في التربة
يمكن أن يعزز أيضًا تكتل التربة، ويزيد من مسامية التربة، ويقدم المزيد من المواد الفعالة للميثانوتروفات (جونغ وآخرون 2022). وبالتالي، فإن التطبيق المشترك للبيوچار والأسمدة العضوية هو ممارسة إدارة واعدة تعزز احتجاز الكربون في التربة وتقلل من خطر انبعاثات الميثان في النظم البيئية الزراعية.
مرتبط ارتباطًا وثيقًا بـ pH التربة، وقد يكون زيادة pH التربة من خلال تطبيقات الفحم الحيوي والأسمدة العضوية أحد الأسباب وراء إثراء ميكروبات تحلل الفوسفور (Po-mineralization) وذوبان الفوسفور (Pisolubilization) (الملف الإضافي 1: الشكل S2؛ زينغ وآخرون 2019). معظم
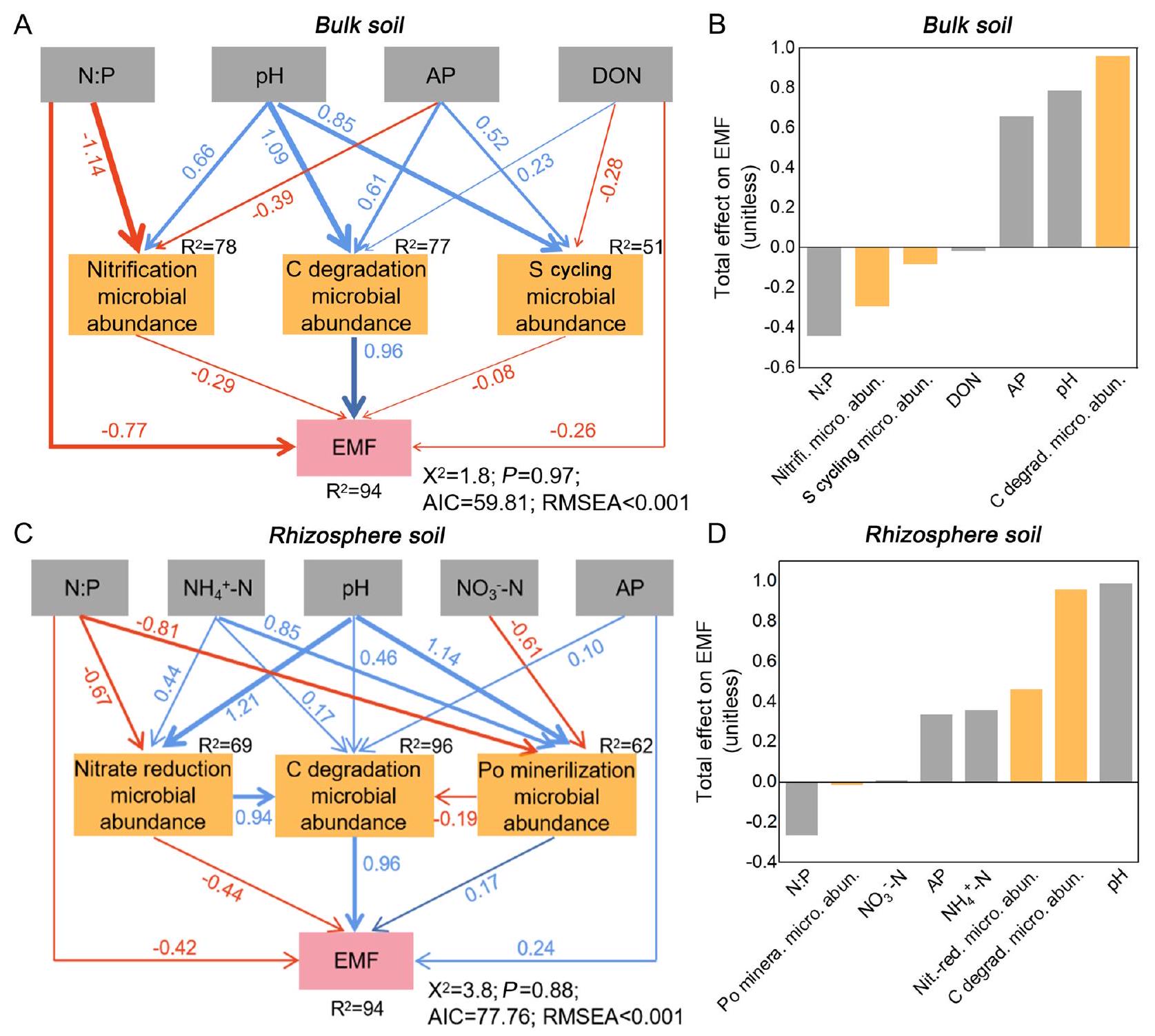
4.3 تنظيم مسارات الميكروبيومات الوظيفية للتربة على وظائف النظام البيئي
يمكن أن يؤثر الرقم الهيدروجيني للتربة والمواد الغذائية المتاحة بشكل مباشر على القوة الدافعة الكهربائية أو بشكل غير مباشر من خلال تنظيم الميكروبات الوظيفية الخاصة بالتربة (الشكل 7؛ ليو وآخرون 2018؛ لي وآخرون 2023). زيادة غير مستقرة
الميكروبات، مما يحفزها على تحلل المواد العضوية عن طريق إفراز سلسلة من الإنزيمات (تشو وآخرون 2020؛ إبراهيم وآخرون 2021). يتم إطلاق العناصر الغذائية في عملية تحلل المواد العضوية بواسطة ميكروبات التربة وتستخدم مرة أخرى من قبل المجتمع فوق الأرض، مما يحسن من امتصاص النباتات للعناصر الغذائية والكتلة الحيوية (دلغادو-باكيريزو وآخرون 2016). بالإضافة إلى ذلك، يمكن أن يؤدي توفر الكربون العالي في التربة إلى استقطاب ميكروبات معينة مرتبطة بتحولات النيتروجين والفوسفور، مما يعزز بدوره العلاقات الإيجابية للفطريات الميكورية الخارجية مع دورات العناصر الغذائية في التربة (رين وآخرون 2023). تؤدي زيادة درجة حموضة التربة إلى تعطيل الربط بين المكونات العضوية والطين، مما يؤثر على الميكروبات التربوية، وبالتالي يؤثر بشكل إيجابي على دورات الكربون والعناصر الغذائية (لو وآخرون 2018؛ وان وآخرون 2020). على سبيل المثال، يمكن أن تؤدي مدخلات الفحم الحيوي والأسمدة العضوية إلى زيادة درجة حموضة التربة لتعزيز نمو الميكروبات الفوسفورية ونشاط الفوسفاتاز، ثم تقوية تحلل الفوسفور العضوي. يؤدي زيادة تحلل الفوسفور إلى تحسين توفر الفوسفور في التربة وتعزيز امتصاص المحاصيل للفوسفور وإنتاجيتها (تشنغ وآخرون 2019؛ هو وآخرون 2023). وبالتالي، يمكن أن تؤدي تطبيقات الفحم الحيوي والأسمدة العضوية إلى تحسين المواطن الحية لمجموعات الميكروبات الوظيفية الخاصة من خلال التأثير على درجة حموضة التربة والعناصر الغذائية المتاحة، مما ينظم بشكل إيجابي الفطريات الميكورية الخارجية. ومع ذلك، فإن مجموعة واسعة ومتنوعة من الوظائف والخدمات عبر فئات النظام البيئي المتعددة، مثل غازات الدفيئة، وخصائص النباتات الوظيفية، وما إلى ذلك، تحتاج أيضًا إلى التحقيق من أجل تقديم مقياس تمثيلي لوظيفة النظام البيئي بشكل عام (غارلاند وآخرون 2021). في الوقت نفسه، يتطلب التقييم الشامل للفطريات الميكورية الخارجية تحت تطبيقات التعديلات العضوية جهدًا إضافيًا في الكشف عن تأثيرات منطقة الجذور التي تنظم العلاقات بين المجتمعات فوق الأرض وتحت الأرض.
5 الخاتمة
يمكن أن تعزز تطبيقات الفحم الحيوي والأسمدة العضوية، وخاصة تطبيقها المشترك، تكاثر الميكروبات الوظيفية في التربة من خلال تنظيم خصائص التربة، مما يحسن بعد ذلك وظائف النظام البيئي وخدماته.
معلومات إضافية
الملف الإضافي 1. طرق، أشكال وجداول إضافية: الجدول
شكر وتقدير
مساهمات المؤلفين
تمويل
توفر البيانات والمواد
الإعلانات
المصالح المتنافسة
تفاصيل المؤلف
ديسمبر 2023
نُشر على الإنترنت: 08 يناير 2024
References
Anantharaman K, Hausmann B, Jungbluth SP, Kantor RS, Lavy A, Warren LA, Rappé MS, Pester M, Loy A, Thomas BC, Banfield JF (2018) Expanded diversity of microbial groups that shape the dissimilatory sulfur cycle. ISME J 12(7):1715-1728
Azeem M, Hayat R, Hussain Q, Tahir MI, Imran M, Abbas Z, Sajid M, Latif A, Irfan M (2019) Effects of biochar and NPK on soil microbial biomass and enzyme activity during 2 years of application in the arid region. Arab J Geosci 12:1-13
Backer R, Rokem JS, Ilangumaran G, Lamont J, Praslickova D, Ricci E, Subramanian S, Smith DL (2018) Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front Plant Sci 9:1473
Bai SH, Omidvar N, Gallart M, Kämper W, Tahmasbian I, Farrar MB, Singh K, Zhou G, Muqadass B, Xu CY, Koech R, Li Y, Nguyen TTN, van Zwieten L (2022) Combined effects of biochar and fertilizer applications on yield: a review and meta-analysis. Sci Total Environ 808:152073
Bao SD (2000) Soil and Agro-Chemistry Analysis, 3rd ed. China Agric. Press, Beijing
Basile-Doelsch I, Balesdent J, Pellerin S (2020) Reviews and syntheses: the mechanisms underlying carbon storage in soil. Biogeosciences 17(21):5223-5242
Biederman LA, Harpole WS (2013) Biochar and its effects on plant productivity and nutrient cycling: a meta-analysis. GCB Bioenergy 5(2):202-214
Breiman L (2001) Random forest. Mach Learn 45:5-32
Brtnicky M, Dokulilova T, Holatko J, Pecina V, Kintl A, Latal O, Vyhnanek T, Prichystalova J, Datta R (2019) Long-term effects of biochar-based organic amendments on soil microbial parameters. Agronomy 9(11):747
Cao H, Ning L, Xun M, Feng F, Li P, Yue S, Song J, Zhang W, Yang H (2019) Biochar can increase nitrogen use efficiency of Malus hupehensis by modulating nitrate reduction of soil and root. Appl Soil Ecol 135:25-32
Chen W, Wang J, Chen X, Meng Z, Xu R, Duoji D, Zhang J, He J, Wang Z, Chen J, Liu K, Hu T, Zhang Y (2022) Soil microbial network complexity predicts ecosystem function along elevation gradients on the Tibetan Plateau. Soil Biol Biochem 172:108766
Dai Z, Yu M, Chen H, Zhao H, Huang Y, Su W, Xia F, Chang SX, Brookes PC, Dahlgren RA, Xu J (2020) Elevated temperature shifts soil N cycling from microbial immobilization to enhanced mineralization, nitrification and denitrification across global terrestrial ecosystems. Global Change Biol 26(9):5267-5276
Delgado-Baquerizo M, Maestre FT, Reich PB, Jeffries TC, Gaitan JJ, Encinar D, Gaitan JJ, Encinar D, Berdugo M, Campbell CD, Singh BK (2016) Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat Commun 7(1):10541
Delgado-Baquerizo M, Reich PB, Trivedi C, Eldridge DJ, Abades S, Alfaro FD, Bastida F, Berhe AA, Cutler NA, Gallardo A, García-Velázquez L, Hart CC, Hayes PE, He JZ, Hseu ZY, Hu HW, Kirchmair M, Neuhauser S, Pérez CA, Reed SC, Santos F, Sullivan BW, Trivedi P, Wang JT, Weber-Grullon L, Williams MA, Singh BK (2020) Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat Ecol Evol 4:210-220
Dominchin MF, Verdenelli RA, Berger MG, Aoki A, Meriles JM (2021) Impact of N-fertilization and peanut shell biochar on soil microbial community structure and enzyme activities in a Typic Haplustoll under different management practices. Eur J Soil Biol 104:103298
Dotaniya ML, Aparna K, Dotaniya CK, Singh M, Regar KL (2019) Role of soil enzymes in sustainable crop production. In: Kuddus M (ed) Enzymes in food biotechnology. Academic Press, Cambridge, pp 569-589
Doyle A, Weintraub MN, Schimel JP (2004) Persulfate digestion and simultaneous colorimetric analysis of carbon and nitrogen in soil extracts. Soil Sci Soc Am J 68:669-676
Galhardo CX, Masini JC (2000) Spectrophotometric determination of phosphate and silicate by sequential injection using molybdenum blue chemistry. Anal Chim Acta 417:191-200
Gao Y, Liang A, Zhang Y, Huang D, McLaughlin N, Zhang Y, Wang Y, Chen X, Zhang
Garland G, Banerjee S, Edlinger A, Miranda Oliveira E, Herzog C, Wittwer R, Philippot L, Maestre FT, van Der Heijden MG (2021) A closer look at
the functions behind ecosystem multifunctionality: a review. J Ecol 109(2):600-613
Giles M, Morley N, Baggs EM, Daniell TJ (2012) Soil nitrate reducing processesdrivers, mechanisms for spatial variation, and significance for nitrous oxide production. Front Microbiol 3:407
Gong H, Li J, Liu Z, Zhang Y, Hou R, Zhu O (2022) Mitigated greenhouse gas emissions in cropping systems by organic fertilizer and tillage management. Land 11(7):1026
Guo Z, Han J, Li J, Xu Y, Wang X (2019) Effects of long-term fertilization on soil organic carbon mineralization and microbial community structure. PLoS ONE 14(1):e0211163
Han Z, Xu P, Li Z, Lin H, Zhu C, Wang J, Zou J (2022) Microbial diversity and the abundance of keystone species drive the response of soil multifunctionality to organic substitution and biochar amendment in a tea plantation. GCB Bioenergy 14(4):481-495
Hossain ME, Shahrukh S, Hossain SA (2022) Chemical fertilizers and pesticides: impacts on soil degradation, groundwater, and human health in Bangladesh. Environmental degradation: challenges and strategies for mitigation. Springer International Publishing, Cham, pp 63-92
Hu W, Ran J, Dong L, Du Q, Ji M, Yao S, Sun Y, Gong C, Hou Q, Gong H, Chen R, Lu J, Xie S, Wang Z, Huang H, Li X, Xiong J, Xia R, Wei M, Zhao D, Zhang Y, Li J, Yang H, Wang X, Deng Y, Sun Y, Li H, Zhang L, Chu Q, Li X, Aqeel M, Manan A, Akram MA, Liu X, Li R, Li F, Hou C, Liu J, He JS, An L, Bardgett RD, Schmid B, Deng J (2021) Aridity-driven shift in biodiversity-soil multifunctionality relationships. Nat Commun 12(1):5350
Hu W, Zhang Y, Rong X, Fei J, Peng J, Luo G (2023) Coupling amendment of biochar and organic fertilizers increases maize yield and phosphorus uptake by regulating soil phosphatase activity and phosphorus-acquiring microbiota. Agr Ecosyst Environ 355:108582
Huang X, Wang C, Liu Q, Zhu Z, Lynn TM, Shen J, Whiteley AS, Kumaresan D, Tida GE, Wu J (2018) Abundance of microbial
Ibrahim MM, Zhang H, Guo L, Chen Y, Heiling M, Zhou B, Mao Y (2021) Biochar interaction with chemical fertilizer regulates soil organic carbon mineralization and the abundance of key C-cycling-related bacteria in rhizosphere soil. Eur J Soil Biol 106:103350
Jarvie HP, Withers PJA, Neal C (2002) Review of robust measurement of phosphorus in river water: sampling, storage, fractionation and sensitivity. Hydrol Earth Syst Sci 6:113-131
Jia R, Zhou J, Chu J, Shahbaz M, Yang Y, Jones DL, Zang H, Razavi BS, Zeng Z (2022) Insights into the associations between soil quality and ecosystem multifunctionality driven by fertilization management: a case study from the North China Plain. J Cleaner Prod 362:132265
Kätterer T, Roobroeck D, Andrén O, Kimutai G, Karltun E, Kirchmann H, Nybery G, Vanlauwe B, de Nowina KR (2019) Biochar addition persistently increased soil fertility and yields in maize-soybean rotations over 10 years in sub-humid regions of Kenya. Field Crop Res 235:18-26
Lazcano C, Zhu-Barker X, Decock C (2021) Effects of organic fertilizers on the soil microorganisms responsible for
Li Y, Chapman SJ, Nicol GW, Yao H (2018) Nitrification and nitrifiers in acidic soils. Soil Biol Biochem 116:290-301
Li S, Wang S, Fan M, Wu Y, Shangguan Z (2020) Interactions between biochar and nitrogen impact soil carbon mineralization and the microbial community. Soil Tillage Res 196:104437
Li M, Guo J, Ren T, Luo G, Shen Q, Lu J, Guo S, Ling N (2021) Crop rotation history constrains soil biodiversity and multifunctionality relationships. Agr Ecosyst Environ 319:107550
Li QM, Zhang D, Zhang JZ, Zhou ZJ, Pan Y, Yang ZH, Zhu JH, Liu YH, Zhang LF (2023) Crop rotations increased soil ecosystem multifunctionality by improving keystone taxa and soil properties in potatoes. Front Microbiol 14:1034761
Liu Y, Wang C, He N, Wen X, Gao Y, Li S, Niu S, Butterbach-Bahl K, Luo Y, Yu G (2017) A global synthesis of the rate and temperature sensitivity of soil nitrogen mineralization: latitudinal patterns and mechanisms. Global Change Biol 23(1):455-464
Liu H, Yang X, Liang C, Li Y, Qiao L, Ai Z, Xue S, Liu G (2019a) Interactive effects of microplastics and glyphosate on the dynamics of soil dissolved organic matter in a Chinese loess soil. CATENA 182:104177
Liu H, Du X, Li Y, Han X, Li B, Zhang X, Li Q, Liang W (2022a) Organic substitutions improve soil quality and maize yield through increasing soil microbial diversity. J Cleaner Prod 347:131323
Liu M, Linna C, Ma S, Ma Q, Song W, Shen M, Song L, Cui K, Zhou Y, Wang L (2022b) Biochar combined with organic and inorganic fertilizers promoted the rapeseed nutrient uptake and improved the purple soil quality. Front Nutr 9:997151
Liu Q, Meki K, Zheng H, Yuan Y, Shao M, Luo X, Li X, Jiang Z, Li F, Xing B (2023) Biochar application in remediating salt-affected soil to achieve carbon neutrality and abate climate change. Biochar 5(1):45
Luo G, Rensing C, Chen H, Liu M, Wang M, Guo S, Ling N, Shen Q (2018) Deciphering the associations between soil microbial diversity and ecosystem multifunctionality driven by long-term fertilization management. Funct Ecol 32(4):1103-1116
Luo G, Wang T, Li K, Li L, Zhang J, Guo S, Ling N, Shen Q (2019) Historical nitrogen deposition and straw addition facilitate the resistance of soil multifunctionality to drying-wetting cycles. Appl Environ Microb 85(8):e02251-e2318
Luo G, Xue C, Jiang Q, Xiao Y, Zhang F, Guo S (2020) Soil carbon, nitrogen, and phosphorus cycling microbial populations and their resistance to global change depend on soil C:N: P stoichiometry. mSystems 5(3):e00162-20
Maron PA, Sarr A, Kaisermann A, Lévêque J, Mathieu O, Guigue J, Karimi B, Bernard L, Dequiedt S, Terrat S, Chabbi A, Ranjard L (2018) High microbial diversity promotes soil ecosystem functioning. Appl Environ Microb 84(9):e02738-e2817
Neina D (2019) The role of soil pH in plant nutrition and soil remediation. Appl Environ Soil Sci 2019:1-9
Ren T, Liao J, Jin L, Delgado-Baquerizo M, Ruan H (2023) Application of biogasslurry and biochar improves soil multifunctionality in a poplar plantation during afforestation processes. Plant Soil. https://doi.org/10.1007/ s11104-023-05968-x
Saha B, Saha S, Roy PD, Padhan D, Pati S, Hazra GC (2018) Microbial transformation of sulphur: an approach to combat the sulphur deficiencies in agricultural soils. In: Meena VS (ed) Role of rhizospheric microbes in soil: Volume 2: nutrient management and crop improvement. Springer, Berlin, pp 77-97
Saifullah DS, Naeem A, Rengel Z, Naidu R (2018) Biochar application for the remediation of salt-affected soils: challenges and opportunities. Sci Total Environ 625:320-335
Singh VK, Rai S, Singh D, Upadhyay RS (2021) Application of soil microorganisms for agricultural and environmental sustainability: a review. In: Dubey SK, Verma SK (eds) Plant, soil and microbes in tropical ecosystems. Springer, Berlin, pp 151-175
Sun R, Guo X, Wang D, Chu H (2015) Effects of long-term application of chemical and organic fertilizers on the abundance of microbial communities involved in the nitrogen cycle. Appl Soil Ecol 95:171-178
Wan W, Tan J, Wang Y, Qin Y, He H, Wu H, Zuo W, He D (2020) Responses of the rhizosphere bacterial community in acidic crop soil to pH : changes in diversity, composition, interaction, and function. Sci Total Environ 700:134418
Wang J, Xie J, Li L, Luo Z, Zhang R, Wang L, Jiang Y (2021) The impact of fertilizer amendments on soil autotrophic bacteria and carbon emissions in maize field on the semiarid Loess Plateau. Front Microb 12:664120
Wang T, Duan Y, Liu G, Shang X, Liu L, Zhang K, Li J, Zou Z, Zhu X, Fang W (2022) Tea plantation intercropping green manure enhances soil functional microbial abundance and multifunctionality resistance to dryingrewetting cycles. Sci Total Environ 810:151282
Wang X, Ge H, Fang Y, Liu C, Eltohamy KM, Wang Z, Liang X (2023) Biochar reduces colloidal phosphorus in leachate by regulating phoD- and phoC-harboring microbial communities during drying/rewetting cycles. Biochar 5(1):1-15
Waqas MA, Li Y, Smith P, Wang X, Ashraf MN, Noor MA, Amou M, Shi S, Zhu Y, Li J, Wan Y, Qin X, Gao Q, Liu S (2020) The influence of nutrient management on soil organic carbon storage, crop production, and yield stability varies under different climates. J Clean Prod 268:121922
Wu J, Sha C, Wang M, Ye C, Li P, Huang S (2021) Effect of organic fertilizer on soil bacteria in maize fields. Land 10(3):328
Yan B, Zhang Y, Wang Y, Rong X, Peng J, Fei J, Luo G (2023) Biochar amendments combined with organic fertilizer improve maize productivity and mitigate nutrient loss by regulating the
Yuan Z, Ali A, Ruiz-Benito P, Jucker T, Mori AS, Wang S, Zhang X, Li H, Hao Z, Wang X, Loreau M (2020) Above-and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient. J Ecol 108(5):2012-2024
Zabala JA, Martínez-Paz JM, Alcon F (2021) A comprehensive approach for agroecosystem services and disservices valuation. Sci Total Environ 768:144859
Zak D, Stutter M, Jensen HS, Egemose S, Carstensen MV, Audet J, Strand JA, Feuerbach P, Hoffmann CC, Christen B, Knudsen M, Stockan J, Watson H, Heckrath G, Kronvang B (2019) An assessment of the multifunctionality of integrated buffer zones in Northwestern Europe. J Environ Qual 48(2):362-375
Zhang Y, Wang F, Zhang J, Zhu T, Lin C, Müller C, Cai Z (2015) Cattle manure and straw have contrasting effects on organic nitrogen mineralization pathways in a subtropical paddy soil. Acta Agr Scand B-S P 65(7):619-628
Zhang Y, Yan J, Rong X, Han Y, Yang Z, Hou K, Zhao H, Hu W (2021) Responses of maize yield, nitrogen and phosphorus runoff losses and soil properties to biochar and organic fertilizer application in a light-loamy fluvo-aquic soil. Agr Ecosyst Environ 314:107433
Zhang YJ, Gao W, Luan HA, Tang JW, Li RN, Li MY, Zhang HZ, Huang SW (2022) Effects of a decade of organic fertilizer substitution on vegetable yield and soil phosphorus pools, phosphatase activities, and the microbial community in a greenhouse vegetable production system. J Integr Agr 21(7):2119-2133
Zhao Q, Wang Y, Xu Z, Yun J, Yu Z (2022a) Unravelling how biochar and dung amendments determine the functional structure and community assembly related to methane metabolisms in grassland soils. Biochar 4(1):49
Zhao Y, Wang X, Yao G, Lin Z, Xu L, Jiang Y, Jin Z, Shan S, Ping L (2022b) Advances in the effects of biochar on microbial ecological function in soil and crop quality. Sustainability 14:10411
Zheng B, Zhu Y, Sardans J, Peñuelas J, Su J (2018) QMEC: a tool for highthroughput quantitative assessment of microbial functional potential in C, N, P, and S biogeochemical cycling. Sci China Life Sci 61(12):1451-1462
Zheng B, Zhang D, Wang Y, Hao X, Wadaan MAM, Hozzein WN, Peñuelas J, Zhu Y, Yang X (2019) Responses to soil pH gradients of inorganic phosphate solubilizing bacteria community. Sci Rep 9(1):25
Zhou W, He P, Li S, Lin B (2005) Mineralization of organic sulfur in paddy soils under flooded conditions and its availability to plants. Geoderma 125(1-2):85-93
Zhou G, Gao S, Lu Y, Liao Y, Nie J, Cao W (2020) Co-incorporation of green manure and rice straw improves rice production, soil chemical, biochemical and microbiological properties in a typical paddy field in southern China. Soil Tillage Res 197:104499
قدّم مخطوطتك إلى SpringerOpen
- تقديم مريح عبر الإنترنت
- مراجعة دقيقة من الأقران
- الوصول المفتوح: مقالات متاحة مجانًا على الإنترنت
- رؤية عالية داخل المجال
- الاحتفاظ بحقوق الطبع والنشر لمقالك
- المحرر المعالج: بين جاو.
*المراسلة:
يوبينغ زانغ
zhyp2003@126.com
شوان تشو
zhouxuan_123@126.com
جونغوين لو
gongwenluo@hunau.edu.cn
قائمة كاملة بمعلومات المؤلف متاحة في نهاية المقال - (انظر الشكل في الصفحة التالية.)
الشكل 2 عدم التشابه في ملفات وفرة الميكروبات الوظيفية بين خمسة علاجات استنادًا إلى اختبار ADONIS و ANOSIM (أ). PERMANOVA ثنائية الاتجاه تقارن التأثيرات الرئيسية والتفاعلية للعلاجات وأماكن التربة على ملفات وفرة الجينات الوظيفية (999 تبديل). استنادًا إلى النتائج، تم استكشاف عدم التشابه لأربعة تجمعات ميكروبية وظيفية محددة بشكل أكبر كالتربة الكلية وتربة الجذور بين خمسة علاجات (ب-هـ). النجوم (*)، و ) تشير إلى اختلافات كبيرة عند , و مستويات الاحتمال، على التوالي. تغييرات في وفرة 64 جينًا وظيفيًا في التربة السائبة ( ) وتربة الجذور ( ) بين خمسة علاجات. كلما كان اللون أكثر اخضرارًا، زادت قيمة المتغير. وعلى العكس، كلما كان اللون أكثر احمرارًا، قلت القيمة. CK بدون أسمدة، CF أسمدة غير عضوية، BF أسمدة غير عضوية وبيوچار، OF سماد عضوي N يحل محل من سماد N، BOF OF وبيوچار
DOI: https://doi.org/10.1007/s42773-023-00296-w
Publication Date: 2024-01-08
Biochar and organic fertilizer applications enhance soil functional microbial abundance and agroecosystem multifunctionality
Abstract
Biochar and organic fertilizer are widely supported to maintain crop production and sustainable development of agroecosystems. However, it is unclear how biochar and organic fertilizer alone or in combination regulate soil functional microbiomes and their relationships to ecosystem multifunctionality (EMF). Herein, a long-term (started in 2013) field experiment, containing five fertilization treatments, was employed to explore the effects of biochar and organic fertilizer applications on the EMF (based on 18 functional indicators of crop productivity, soil nutrient supply, element cycling, and microbial biomass) and the functional microbiomes of bulk soil and rhizosphere soil [normalizing the abundances of 64 genes related to carbon (C), nitrogen (N), phosphorus (P), and sulphur (S) cycles]. Compared with single-chemical fertilization, biochar and organic fertilizer inputs significantly enhanced most ecosystem-single functions and, in particular, the EMF significantly increased by 18.7-30.1%; biochar and organic fertilizer applications significantly increased the abundances of soil microbial functional taxa related to C-N-P-S cycles to varying degree. The combined application of biochar and organic fertilizer showed a better improvement in these indicators compared to using them individually. Most functional microbial populations in the soil, especially the taxa involved in C degradation, nitrification, nitrate-reduction, organic P mineralization, and S cycling showed significantly positive associations with the EMF at different threshold levels, which ultimately was regulated by soil pH and nutrient availability. These results highlight the strong links between soil microbiomes and agroecosystem functions, as well as providing scientific support for inclusion of biochar in agricultural production and services with organic amendments.
Highlights
- 8-year field evidence revealed impacts of biochar and pig manure on soil functional microbiome and ecosystem functions.
- Biochar and pig manure inputs notably enhanced most ecosystem-single functions and the EMF increased by 18.7-30.1%.
4. Increase in EMF was related to microbe-driven soil processes such as C degradation, nitrification, and Po mineralization.
5. Inclusion of biochar in crop production with organic amendments could enhance agro-ecosystem functions and services.
Graphical Abstract
1 Introduction
available nutrients to promote soil microbial activities and metabolisms, and then maintain or promote ecosystem services (Han et al. 2022; Wu et al. 2021). In addition to organic fertilizers, biochar has been widely used as a soil conditioner or organic material in crop production and soil improvement in recent years. Rich pore structure, nutrients, and carbon (C) of biochar can improve soil structure and nutrient supply and increase soil C storage (Liu et al. 2023; Bai et al. 2022). Thus, soil fertility and crop productivity gains may be greater with the mixing of biochar with organic fertilizer than
with single applications (Brtnicky et al. 2019; Bai et al. 2022; Liu et al. 2022a; Yan et al. 2023). However, current studies on the effects of their coupling application on the multiple functions (termed “multifunctionality”) of agroecosystems are insufficient.
The services or functions provided by agroecosystems are closely related to the diversity and function of microbial communities in soil (Zabala et al. 2021). Soil microorganisms participate in various functional processes such as nutrient cycling, disease suppression, primary production, and thus special microbial taxa in the soil play a vital role in EMF (Singh et al. 2021; Han et al. 2022). For example, the microbial taxa related to C cycle can drive organic matter degradation and affect soil C storage, thereby regulating ecosystem C balance (Basile-Doelsch et al. 2020). Nitrogen-fixing bacteria can fix nitrogen (N) from atmosphere to ammonia for plant absorption (Aasfar et al. 2021). Some functional taxa also affect ecosystem N cycle by triggering the processes of soil nitrification and denitrification (Dai et al. 2020). Phosphorus
Changes in management practices such as land use and fertilization may alter the ability of soil microorganisms to maintain the multiple-functions of above- and belowground ecosystems (Delgado-Baquerizo et al. 2016). Any loss of soil biodiversity may have a negative impact on ecosystem functioning (Delgado-Baquerizo et al. 2020). Therefore, the status of special functional microbial populations in soil can influence EMF and its stability (Luo et al. 2018, 2019). Soil enzyme activity, organic matter and nutrient content, plant nutrient uptake and productivity, and microbial biomass are broadly applied as the indicators for evaluating ecosystem functions and services (Delgado-Baquerizo et al. 2016; Luo et al. 2018; Zak et al. 2019). Functional microorganisms can influence various soil biochemical processes, and various microbial processes occurring in soil can directly or indirectly
regulate above-ground biomes (Singh et al. 2021). Thus, assessing how different ecosystem functions show synergies or trade-offs is important for understanding EMF implementation (Yuan et al. 2020). The index of EMF mainly be quantified through mean value method and multi-threshold method, both of which are used together to better assess the effects of soil biodiversity on the EMF (Li et al. 2020; Liu et al. 2019b). Thus, multi-faceted assessment can effectively clarify how the amendments of biochar and organic fertilizer regulate ecosystem functions and their relationships with special functional microbes.
2 Materials and methods
2.1 Experimental design and field management
2.2 Soil and plant sample collection
Soil was sampled using the five-point method and shake-down method. Five portions of soil mass with intact root systems were dug from each plot and brought back to laboratory immediately. Large patches of soil that did not contain roots were gently shaken off first, and then the soil adhering to the root perimeter removed with a knife was used as bulk soil. The soil adhering to the root perimeter was removed using a brush as rhizosphere soil. Five soil samples from each plot were mixed into one portion and then stored for subsequent analysis.
2.3 Soil physiochemical property determination
(DOP) content was calculated as TDP-
2.4 Ecosystem multifunctionality assessment
2.5 Functional gene abundance quantification
2.6 Data analysis
The main predictors of the EMF index were deciphered by random forest analysis proposed by Breiman (2001). Random forest algorithms added an additional layer of randomness to bagging, and each tree used a different bootstrap sample of the data. The “randomForest” and “rfPermute” packages were used to carry out and estimate the significance of importance metrics for a random forest model by permuting the response variable. Details on the analysis process have been given in former publications (Luo et al. 2019, 2020). Structural equation model was developed by analyzing soil properties and functional microbial abundances to identify the main drivers and pathways of the EMF index. A priori model of the known potential correlations between the EMF and
the above mentioned drivers was developed on the basis of data mining with random forest analysis. AMOS software (SPSS AMOS 20.0.0) was used for the construction and evaluation of the model. The following metrics were used to evaluate the goodness of fit of our model: the root mean squared error of approximation (RMSEA), chisquare value, Fisher’s P statistic, and the Akaike information criterion (AIC). Ultimately, the priori model was optimized to obtain the model that satisfied the goodness of fit condition, and the significance of the path was further verified.
3 Results
3.1 The impacts of fertilization regimes on ecosystem functions
3.2 The impacts of organic amendments on soil functional microbial abundances
3.3 Relationship between ecosystem functions and functional microbial abundances
3.4 Relationship between the EMF and soil functional microbial abundances
The abundances of the microbial taxa related to C degradation, nitrate-reduction, and Po-mineralization in the rhizosphere soil were the important predictors of the EMF (
the regression relationships between microbial abundance related to C degradation, nitrate-reduction, and Po-mineralization and the EMF at each threshold, the Tmin was
3.5 Regulation of soil functional microbial abundances on the EMF
The abundances of soil microbial taxa related to C degradation showed the largest direct effect on the EMF, followed by soil available
4 Discussion
4.1 Biochar and organic fertilizer applications improve ecosystem functions
a greater promotion effect on maintaining the balance of soil nutrient supply and plant nutrient requirement (Kätterer et al. 2019). However, organic matter decomposition and nutrient transformations in soil cannot be separated from the regulation of soil extracellular enzymes (Dotaniya et al. 2019).
As indicators of ecosystem functions, soil enzyme activities participate in multiple soil functional processes (Wang et al. 2021). Biochar and organic fertilizer
amendments could enhance the activities of soil enzymes correlated with C and nutrients cycles (Fig. 1). Biochar and organic fertilizer can enhance soil C storage and bio-availability, thereby stimulating microbial growth and synthesis of enzymes associated with C degradation (Dominchin et al. 2021; Zhou et al. 2020). This is also confirmed by our results (Fig. 1). Biochar and organic matter can increase soil pH and improve the adaptability of some soil enzymes such as organic
4.2 Biochar and organic fertilizer amendments regulate soil functional microbiomes
can also enhance soil agglomeration, increase soil porosity, and offer more active substances for methanotrophs (Gong et al. 2022). Thus, biochar and organic fertilizer co-application is a promising management practice enhancing soil C sequestration and reducing the risk of methane emissions in agroecosystems.
closely related to soil pH , and the increase of soil pH by biochar and organic fertilizer applications may be one of the reasons for enrichment of Po-mineralization and Pisolubilization microbes (Additional file 1: Fig. S2; Zheng et al. 2019). Most
4.3 Regulating pathways of soil functional microbiomes on ecosystem functions
Soil pH and available nutrients could directly influence the EMF or indirectly by regulating soil special functional microbes (Fig. 7; Luo et al. 2018; Li et al. 2023). Increased unstable
microbes, thereby stimulating them to degrade organic matter by secreting a series of enzymes (Zhou et al. 2020; Ibrahim et al. 2021). Nutrients in the process of organic matter degradation are released by soil microbes and utilized by above-ground community again, thus improving plant nutrient uptake and biomass (Delgado-Baquerizo et al. 2016). In addition, higher C availability in soil can enable the recruitment of specific microorganisms associated with N and P transformations, which in turn promotes the positive relationships of the EMF with soil nutrient cycles (Ren et al. 2023). Higher soil pH results in the disruption of the binding between organic components and clay, affecting soil microbiomes, and thus positively affecting C and nutrient cycles (Luo et al. 2018; Wan et al. 2020). For example, biochar and organic fertilizer inputs can increase soil pH to promote the growth of phosphorolytic microbes and phosphatase activity, and then strengthen organic P mineralization. Increased P mineralization further improves soil P availability and promotes crop P uptake and yield (Zheng et al. 2019; Hu et al. 2023). Thus, biochar and organic fertilizer applications can improve the living habitats of special functional microbial taxa by influencing soil pH and available nutrients, thereby positively regulating EMF. However, a wide and diverse range of functions and services across multiple ecosystem categories, e.g., greenhouse gases, plant functional traits, etc., also need to be investigated in order to provide a representative measure of overall ecosystem functioning (Garland et al. 2021). Meanwhile, comprehensive assessment of the EMF under organic amendment applications requires additional effort in disclosing rhizosphere effects that is regulate the associations between above- and below-ground communities.
5 Conclusion
biochar and organic fertilizer applications, especially their co-application, could promote the propagation of soil functional microbiomes by regulating soil attributes and then improve ecosystem functions and services.
Supplementary Information
Additional file 1. Supplementary methods, figures and tables: Table
Acknowledgements
Author contributions
Funding
Availability of data and materials
Declarations
Competing interests
Author details
December 2023
Published online: 08 January 2024
References
Anantharaman K, Hausmann B, Jungbluth SP, Kantor RS, Lavy A, Warren LA, Rappé MS, Pester M, Loy A, Thomas BC, Banfield JF (2018) Expanded diversity of microbial groups that shape the dissimilatory sulfur cycle. ISME J 12(7):1715-1728
Azeem M, Hayat R, Hussain Q, Tahir MI, Imran M, Abbas Z, Sajid M, Latif A, Irfan M (2019) Effects of biochar and NPK on soil microbial biomass and enzyme activity during 2 years of application in the arid region. Arab J Geosci 12:1-13
Backer R, Rokem JS, Ilangumaran G, Lamont J, Praslickova D, Ricci E, Subramanian S, Smith DL (2018) Plant growth-promoting rhizobacteria: context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front Plant Sci 9:1473
Bai SH, Omidvar N, Gallart M, Kämper W, Tahmasbian I, Farrar MB, Singh K, Zhou G, Muqadass B, Xu CY, Koech R, Li Y, Nguyen TTN, van Zwieten L (2022) Combined effects of biochar and fertilizer applications on yield: a review and meta-analysis. Sci Total Environ 808:152073
Bao SD (2000) Soil and Agro-Chemistry Analysis, 3rd ed. China Agric. Press, Beijing
Basile-Doelsch I, Balesdent J, Pellerin S (2020) Reviews and syntheses: the mechanisms underlying carbon storage in soil. Biogeosciences 17(21):5223-5242
Biederman LA, Harpole WS (2013) Biochar and its effects on plant productivity and nutrient cycling: a meta-analysis. GCB Bioenergy 5(2):202-214
Breiman L (2001) Random forest. Mach Learn 45:5-32
Brtnicky M, Dokulilova T, Holatko J, Pecina V, Kintl A, Latal O, Vyhnanek T, Prichystalova J, Datta R (2019) Long-term effects of biochar-based organic amendments on soil microbial parameters. Agronomy 9(11):747
Cao H, Ning L, Xun M, Feng F, Li P, Yue S, Song J, Zhang W, Yang H (2019) Biochar can increase nitrogen use efficiency of Malus hupehensis by modulating nitrate reduction of soil and root. Appl Soil Ecol 135:25-32
Chen W, Wang J, Chen X, Meng Z, Xu R, Duoji D, Zhang J, He J, Wang Z, Chen J, Liu K, Hu T, Zhang Y (2022) Soil microbial network complexity predicts ecosystem function along elevation gradients on the Tibetan Plateau. Soil Biol Biochem 172:108766
Dai Z, Yu M, Chen H, Zhao H, Huang Y, Su W, Xia F, Chang SX, Brookes PC, Dahlgren RA, Xu J (2020) Elevated temperature shifts soil N cycling from microbial immobilization to enhanced mineralization, nitrification and denitrification across global terrestrial ecosystems. Global Change Biol 26(9):5267-5276
Delgado-Baquerizo M, Maestre FT, Reich PB, Jeffries TC, Gaitan JJ, Encinar D, Gaitan JJ, Encinar D, Berdugo M, Campbell CD, Singh BK (2016) Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat Commun 7(1):10541
Delgado-Baquerizo M, Reich PB, Trivedi C, Eldridge DJ, Abades S, Alfaro FD, Bastida F, Berhe AA, Cutler NA, Gallardo A, García-Velázquez L, Hart CC, Hayes PE, He JZ, Hseu ZY, Hu HW, Kirchmair M, Neuhauser S, Pérez CA, Reed SC, Santos F, Sullivan BW, Trivedi P, Wang JT, Weber-Grullon L, Williams MA, Singh BK (2020) Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat Ecol Evol 4:210-220
Dominchin MF, Verdenelli RA, Berger MG, Aoki A, Meriles JM (2021) Impact of N-fertilization and peanut shell biochar on soil microbial community structure and enzyme activities in a Typic Haplustoll under different management practices. Eur J Soil Biol 104:103298
Dotaniya ML, Aparna K, Dotaniya CK, Singh M, Regar KL (2019) Role of soil enzymes in sustainable crop production. In: Kuddus M (ed) Enzymes in food biotechnology. Academic Press, Cambridge, pp 569-589
Doyle A, Weintraub MN, Schimel JP (2004) Persulfate digestion and simultaneous colorimetric analysis of carbon and nitrogen in soil extracts. Soil Sci Soc Am J 68:669-676
Galhardo CX, Masini JC (2000) Spectrophotometric determination of phosphate and silicate by sequential injection using molybdenum blue chemistry. Anal Chim Acta 417:191-200
Gao Y, Liang A, Zhang Y, Huang D, McLaughlin N, Zhang Y, Wang Y, Chen X, Zhang
Garland G, Banerjee S, Edlinger A, Miranda Oliveira E, Herzog C, Wittwer R, Philippot L, Maestre FT, van Der Heijden MG (2021) A closer look at
the functions behind ecosystem multifunctionality: a review. J Ecol 109(2):600-613
Giles M, Morley N, Baggs EM, Daniell TJ (2012) Soil nitrate reducing processesdrivers, mechanisms for spatial variation, and significance for nitrous oxide production. Front Microbiol 3:407
Gong H, Li J, Liu Z, Zhang Y, Hou R, Zhu O (2022) Mitigated greenhouse gas emissions in cropping systems by organic fertilizer and tillage management. Land 11(7):1026
Guo Z, Han J, Li J, Xu Y, Wang X (2019) Effects of long-term fertilization on soil organic carbon mineralization and microbial community structure. PLoS ONE 14(1):e0211163
Han Z, Xu P, Li Z, Lin H, Zhu C, Wang J, Zou J (2022) Microbial diversity and the abundance of keystone species drive the response of soil multifunctionality to organic substitution and biochar amendment in a tea plantation. GCB Bioenergy 14(4):481-495
Hossain ME, Shahrukh S, Hossain SA (2022) Chemical fertilizers and pesticides: impacts on soil degradation, groundwater, and human health in Bangladesh. Environmental degradation: challenges and strategies for mitigation. Springer International Publishing, Cham, pp 63-92
Hu W, Ran J, Dong L, Du Q, Ji M, Yao S, Sun Y, Gong C, Hou Q, Gong H, Chen R, Lu J, Xie S, Wang Z, Huang H, Li X, Xiong J, Xia R, Wei M, Zhao D, Zhang Y, Li J, Yang H, Wang X, Deng Y, Sun Y, Li H, Zhang L, Chu Q, Li X, Aqeel M, Manan A, Akram MA, Liu X, Li R, Li F, Hou C, Liu J, He JS, An L, Bardgett RD, Schmid B, Deng J (2021) Aridity-driven shift in biodiversity-soil multifunctionality relationships. Nat Commun 12(1):5350
Hu W, Zhang Y, Rong X, Fei J, Peng J, Luo G (2023) Coupling amendment of biochar and organic fertilizers increases maize yield and phosphorus uptake by regulating soil phosphatase activity and phosphorus-acquiring microbiota. Agr Ecosyst Environ 355:108582
Huang X, Wang C, Liu Q, Zhu Z, Lynn TM, Shen J, Whiteley AS, Kumaresan D, Tida GE, Wu J (2018) Abundance of microbial
Ibrahim MM, Zhang H, Guo L, Chen Y, Heiling M, Zhou B, Mao Y (2021) Biochar interaction with chemical fertilizer regulates soil organic carbon mineralization and the abundance of key C-cycling-related bacteria in rhizosphere soil. Eur J Soil Biol 106:103350
Jarvie HP, Withers PJA, Neal C (2002) Review of robust measurement of phosphorus in river water: sampling, storage, fractionation and sensitivity. Hydrol Earth Syst Sci 6:113-131
Jia R, Zhou J, Chu J, Shahbaz M, Yang Y, Jones DL, Zang H, Razavi BS, Zeng Z (2022) Insights into the associations between soil quality and ecosystem multifunctionality driven by fertilization management: a case study from the North China Plain. J Cleaner Prod 362:132265
Kätterer T, Roobroeck D, Andrén O, Kimutai G, Karltun E, Kirchmann H, Nybery G, Vanlauwe B, de Nowina KR (2019) Biochar addition persistently increased soil fertility and yields in maize-soybean rotations over 10 years in sub-humid regions of Kenya. Field Crop Res 235:18-26
Lazcano C, Zhu-Barker X, Decock C (2021) Effects of organic fertilizers on the soil microorganisms responsible for
Li Y, Chapman SJ, Nicol GW, Yao H (2018) Nitrification and nitrifiers in acidic soils. Soil Biol Biochem 116:290-301
Li S, Wang S, Fan M, Wu Y, Shangguan Z (2020) Interactions between biochar and nitrogen impact soil carbon mineralization and the microbial community. Soil Tillage Res 196:104437
Li M, Guo J, Ren T, Luo G, Shen Q, Lu J, Guo S, Ling N (2021) Crop rotation history constrains soil biodiversity and multifunctionality relationships. Agr Ecosyst Environ 319:107550
Li QM, Zhang D, Zhang JZ, Zhou ZJ, Pan Y, Yang ZH, Zhu JH, Liu YH, Zhang LF (2023) Crop rotations increased soil ecosystem multifunctionality by improving keystone taxa and soil properties in potatoes. Front Microbiol 14:1034761
Liu Y, Wang C, He N, Wen X, Gao Y, Li S, Niu S, Butterbach-Bahl K, Luo Y, Yu G (2017) A global synthesis of the rate and temperature sensitivity of soil nitrogen mineralization: latitudinal patterns and mechanisms. Global Change Biol 23(1):455-464
Liu H, Yang X, Liang C, Li Y, Qiao L, Ai Z, Xue S, Liu G (2019a) Interactive effects of microplastics and glyphosate on the dynamics of soil dissolved organic matter in a Chinese loess soil. CATENA 182:104177
Liu H, Du X, Li Y, Han X, Li B, Zhang X, Li Q, Liang W (2022a) Organic substitutions improve soil quality and maize yield through increasing soil microbial diversity. J Cleaner Prod 347:131323
Liu M, Linna C, Ma S, Ma Q, Song W, Shen M, Song L, Cui K, Zhou Y, Wang L (2022b) Biochar combined with organic and inorganic fertilizers promoted the rapeseed nutrient uptake and improved the purple soil quality. Front Nutr 9:997151
Liu Q, Meki K, Zheng H, Yuan Y, Shao M, Luo X, Li X, Jiang Z, Li F, Xing B (2023) Biochar application in remediating salt-affected soil to achieve carbon neutrality and abate climate change. Biochar 5(1):45
Luo G, Rensing C, Chen H, Liu M, Wang M, Guo S, Ling N, Shen Q (2018) Deciphering the associations between soil microbial diversity and ecosystem multifunctionality driven by long-term fertilization management. Funct Ecol 32(4):1103-1116
Luo G, Wang T, Li K, Li L, Zhang J, Guo S, Ling N, Shen Q (2019) Historical nitrogen deposition and straw addition facilitate the resistance of soil multifunctionality to drying-wetting cycles. Appl Environ Microb 85(8):e02251-e2318
Luo G, Xue C, Jiang Q, Xiao Y, Zhang F, Guo S (2020) Soil carbon, nitrogen, and phosphorus cycling microbial populations and their resistance to global change depend on soil C:N: P stoichiometry. mSystems 5(3):e00162-20
Maron PA, Sarr A, Kaisermann A, Lévêque J, Mathieu O, Guigue J, Karimi B, Bernard L, Dequiedt S, Terrat S, Chabbi A, Ranjard L (2018) High microbial diversity promotes soil ecosystem functioning. Appl Environ Microb 84(9):e02738-e2817
Neina D (2019) The role of soil pH in plant nutrition and soil remediation. Appl Environ Soil Sci 2019:1-9
Ren T, Liao J, Jin L, Delgado-Baquerizo M, Ruan H (2023) Application of biogasslurry and biochar improves soil multifunctionality in a poplar plantation during afforestation processes. Plant Soil. https://doi.org/10.1007/ s11104-023-05968-x
Saha B, Saha S, Roy PD, Padhan D, Pati S, Hazra GC (2018) Microbial transformation of sulphur: an approach to combat the sulphur deficiencies in agricultural soils. In: Meena VS (ed) Role of rhizospheric microbes in soil: Volume 2: nutrient management and crop improvement. Springer, Berlin, pp 77-97
Saifullah DS, Naeem A, Rengel Z, Naidu R (2018) Biochar application for the remediation of salt-affected soils: challenges and opportunities. Sci Total Environ 625:320-335
Singh VK, Rai S, Singh D, Upadhyay RS (2021) Application of soil microorganisms for agricultural and environmental sustainability: a review. In: Dubey SK, Verma SK (eds) Plant, soil and microbes in tropical ecosystems. Springer, Berlin, pp 151-175
Sun R, Guo X, Wang D, Chu H (2015) Effects of long-term application of chemical and organic fertilizers on the abundance of microbial communities involved in the nitrogen cycle. Appl Soil Ecol 95:171-178
Wan W, Tan J, Wang Y, Qin Y, He H, Wu H, Zuo W, He D (2020) Responses of the rhizosphere bacterial community in acidic crop soil to pH : changes in diversity, composition, interaction, and function. Sci Total Environ 700:134418
Wang J, Xie J, Li L, Luo Z, Zhang R, Wang L, Jiang Y (2021) The impact of fertilizer amendments on soil autotrophic bacteria and carbon emissions in maize field on the semiarid Loess Plateau. Front Microb 12:664120
Wang T, Duan Y, Liu G, Shang X, Liu L, Zhang K, Li J, Zou Z, Zhu X, Fang W (2022) Tea plantation intercropping green manure enhances soil functional microbial abundance and multifunctionality resistance to dryingrewetting cycles. Sci Total Environ 810:151282
Wang X, Ge H, Fang Y, Liu C, Eltohamy KM, Wang Z, Liang X (2023) Biochar reduces colloidal phosphorus in leachate by regulating phoD- and phoC-harboring microbial communities during drying/rewetting cycles. Biochar 5(1):1-15
Waqas MA, Li Y, Smith P, Wang X, Ashraf MN, Noor MA, Amou M, Shi S, Zhu Y, Li J, Wan Y, Qin X, Gao Q, Liu S (2020) The influence of nutrient management on soil organic carbon storage, crop production, and yield stability varies under different climates. J Clean Prod 268:121922
Wu J, Sha C, Wang M, Ye C, Li P, Huang S (2021) Effect of organic fertilizer on soil bacteria in maize fields. Land 10(3):328
Yan B, Zhang Y, Wang Y, Rong X, Peng J, Fei J, Luo G (2023) Biochar amendments combined with organic fertilizer improve maize productivity and mitigate nutrient loss by regulating the
Yuan Z, Ali A, Ruiz-Benito P, Jucker T, Mori AS, Wang S, Zhang X, Li H, Hao Z, Wang X, Loreau M (2020) Above-and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient. J Ecol 108(5):2012-2024
Zabala JA, Martínez-Paz JM, Alcon F (2021) A comprehensive approach for agroecosystem services and disservices valuation. Sci Total Environ 768:144859
Zak D, Stutter M, Jensen HS, Egemose S, Carstensen MV, Audet J, Strand JA, Feuerbach P, Hoffmann CC, Christen B, Knudsen M, Stockan J, Watson H, Heckrath G, Kronvang B (2019) An assessment of the multifunctionality of integrated buffer zones in Northwestern Europe. J Environ Qual 48(2):362-375
Zhang Y, Wang F, Zhang J, Zhu T, Lin C, Müller C, Cai Z (2015) Cattle manure and straw have contrasting effects on organic nitrogen mineralization pathways in a subtropical paddy soil. Acta Agr Scand B-S P 65(7):619-628
Zhang Y, Yan J, Rong X, Han Y, Yang Z, Hou K, Zhao H, Hu W (2021) Responses of maize yield, nitrogen and phosphorus runoff losses and soil properties to biochar and organic fertilizer application in a light-loamy fluvo-aquic soil. Agr Ecosyst Environ 314:107433
Zhang YJ, Gao W, Luan HA, Tang JW, Li RN, Li MY, Zhang HZ, Huang SW (2022) Effects of a decade of organic fertilizer substitution on vegetable yield and soil phosphorus pools, phosphatase activities, and the microbial community in a greenhouse vegetable production system. J Integr Agr 21(7):2119-2133
Zhao Q, Wang Y, Xu Z, Yun J, Yu Z (2022a) Unravelling how biochar and dung amendments determine the functional structure and community assembly related to methane metabolisms in grassland soils. Biochar 4(1):49
Zhao Y, Wang X, Yao G, Lin Z, Xu L, Jiang Y, Jin Z, Shan S, Ping L (2022b) Advances in the effects of biochar on microbial ecological function in soil and crop quality. Sustainability 14:10411
Zheng B, Zhu Y, Sardans J, Peñuelas J, Su J (2018) QMEC: a tool for highthroughput quantitative assessment of microbial functional potential in C, N, P, and S biogeochemical cycling. Sci China Life Sci 61(12):1451-1462
Zheng B, Zhang D, Wang Y, Hao X, Wadaan MAM, Hozzein WN, Peñuelas J, Zhu Y, Yang X (2019) Responses to soil pH gradients of inorganic phosphate solubilizing bacteria community. Sci Rep 9(1):25
Zhou W, He P, Li S, Lin B (2005) Mineralization of organic sulfur in paddy soils under flooded conditions and its availability to plants. Geoderma 125(1-2):85-93
Zhou G, Gao S, Lu Y, Liao Y, Nie J, Cao W (2020) Co-incorporation of green manure and rice straw improves rice production, soil chemical, biochemical and microbiological properties in a typical paddy field in southern China. Soil Tillage Res 197:104499
Submit your manuscript to a SpringerOpen
- Convenient online submission
- Rigorous peer review
- Open access: articles freely available online
- High visibility within the field
- Retaining the copyright to your article
- Handling editor: Bin Gao.
*Correspondence:
Yuping Zhang
zhyp2003@126.com
Xuan Zhou
zhouxuan_123@126.com
Gongwen Luo
gongwenluo@hunau.edu.cn
Full list of author information is available at the end of the article - (See figure on next page.)
Fig. 2 The dissimilarity of functional microbial abundance profiles between five treatments based on ADONIS and ANOSIM test (A). Two-way PERMANOVA comparing the main and interactive effects of the treatments and soil places on the functional gene abundance profiles (999 permutations). Based on the results, the dissimilarity of four specific functional microbial populations was further explored as the bulk soil and rhizosphere soil among five treatments (B-E). Asterisks (, and ) indicate significant differences at , and probability levels, respectively. Changes in abundance of 64 functional genes in the bulk soil ( ) and rhizosphere soil ( ) between five treatments. The greener the color, the greater the value of the variable. Conversely, the redder the color, the smaller the value. CK no fertilizers, CF inorganic fertilizers, BF inorganic fertilizers and biochar, OF organic fertilizer N replaces of fertilizer N , BOF OF and biochar