تقارب الميكروبيوم يمكّن بكتيريا الجذور المفرزة للحديد من تحسين تغذية الحديد وإنتاجية الفول السوداني المزروع بالتداخل مع الذرة Microbiome convergence enables siderophore-secreting-rhizobacteria to improve iron nutrition and yield of peanut intercropped with maize
يمكن أن يحسن الزراعة المختلطة من تغذية النباتات وكذلك من إنتاجية المحاصيل. ومع ذلك، لا يزال الآلية الدقيقة التي تعزز من تحسين اكتساب العناصر الغذائية والدور الذي قد تلعبه الميكروبيوم في منطقة الجذور في هذه العملية غير مفهومة بشكل جيد. هنا، نستخدم نظام الزراعة المختلطة بين الفول السوداني والذرة للتحقيق في دور الميكروبات المرتبطة بالجذور في تغذية الحديد في هذه المحاصيل، من خلال دمج تحليل الميكروبيوم، وعزل السلالات والمواد، والتحقق الوظيفي. نجد أن الزراعة المختلطة تزيد من تغذية الحديد في الفول السوداني ولكن ليس في نباتات الذرة، وأن تركيبة الميكروبات تتغير وتتقارب بين النباتين المختبرين في تجارب الزراعة المختلطة. نحدد سيفيروفور يفرزه بكتيريا الزائفة، وهو بايوفيردين، الذي يحسن من تغذية الحديد في تجارب البيوت الزجاجية والميدان. تشير نتائجنا إلى أن وجود بكتيريا الزائفة التي تفرز السيفيروفور في الفول السوداني والذرة المزروعة بشكل مختلط يلعب دورًا مهمًا في تغذية الحديد. يمكن استخدام هذه النتائج لتصور ممارسات الزراعة المختلطة المستقبلية التي تهدف إلى تحسين تغذية النباتات.
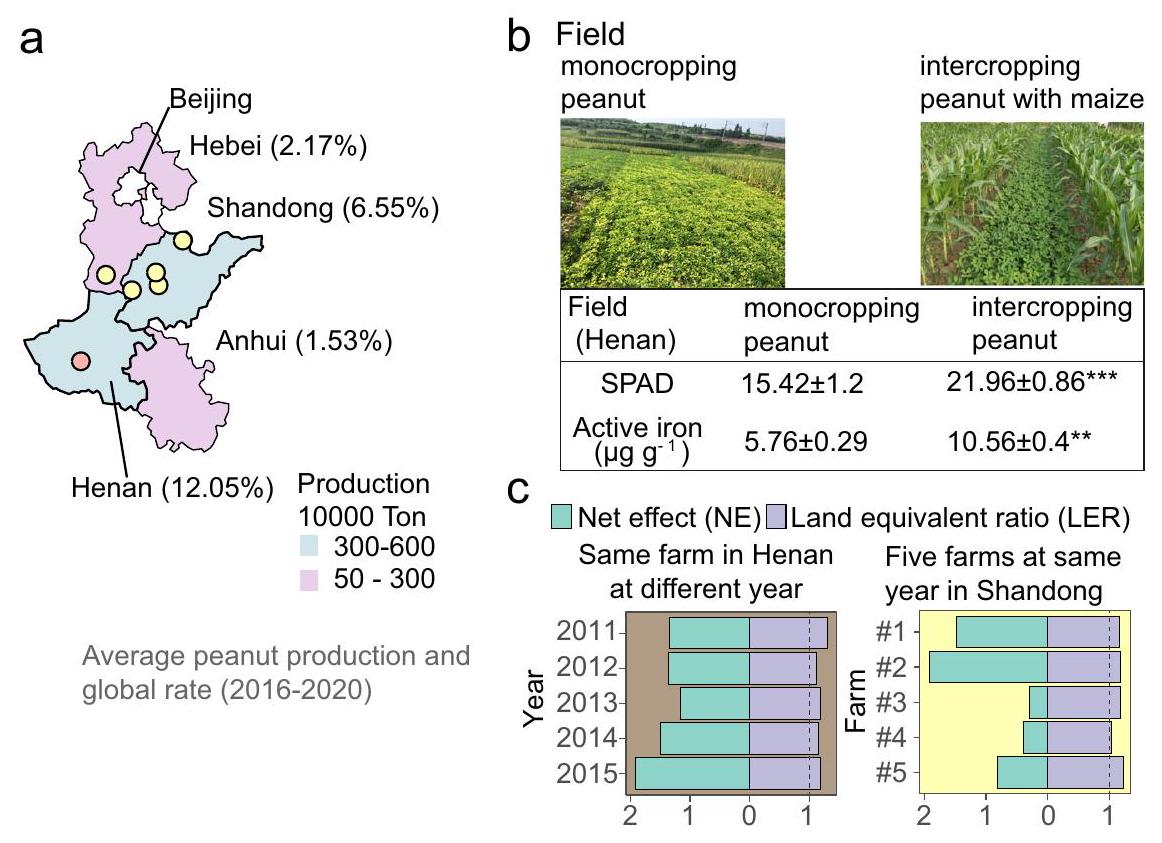
أحد أكبر التحديات التي تواجه البشرية هو كيفية إطعام عدد كبير من السكان بشكل مستدام.، خاصة في دول مثل الصين، التي تشكل حوالي خُمس سكان العالم ولكن لديها أراضٍ زراعية محدودة متاحة. إحدى الحلول لضمان الأمن الغذائي هي زراعة محاصيلين على الأقل في نفس الحقل في وقت واحد، لأن الزراعة المختلطة غالبًا ما تزيد من الإنتاجية وكفاءة استخدام الموارد ومكافحة الآفات والأمراض، والاستدامة البيئية.الفول السوداني (أراشيش زراعة الفول السوداني (Hypogaea L.) مع الذرة (Zea mays) شائعة في الصين لأنها أكثر فعالية وأكثر استدامة بيئيًا من زراعة الفول السوداني بشكل منفرد، خاصةً بالنسبة لمالكي الأراضي الصغيرة.الفول السوداني هو بقوليات زيتية مهمة. تنتج الصينوسهول شمال الصين تقريبًاإنتاج الفول السوداني العالمي (الشكل 1أ). ومع ذلك، فإن إنتاجية وجودة الفول السوداني تتأثر بشدة في هذه المنطقة بسبب نقص الحديد (Fe) الشديد السائد في التربة القلوية و
الشكل 1 | زراعة الذرة والفول السوداني معًا تحسن من تغذية الفول السوداني بالحديد وإنتاجه في التربة الكلسية. أ متوسط إنتاج الفول السوداني ومعدل الإنتاج العالمي لأربع مقاطعات من سهل شمال الصين وموقع التجارب الميدانية (المشار إليه بالنقاط).صور لزراعة المحاصيل الأحادية وزراعة المحاصيل المتداخلة للفول السوداني في مواقع الحقول ونوعين مرتبطين من ظاهرة اصفرار الأوراق الناتج عن نقص الحديد: قيمة تحليل التربة والنبات (قيمة SPAD التي تشير إلى محتوى الكلوروفيل) والحديد النشط (الذي يشير إلى مدى نقص الحديد) في الأوراق الشابة. القيم هي متوسطات. الانحراف المعياري (SD) عينات مستقلة بيولوجياً-قيمة لـ SPAD: ، -قيمة الحديد النشط: 0.002). تم تحويل توزيعات البيانات المنحرفة أو البيانات ذات تباين غير متجانس باستخدام خوارزمية BoxCox لتلبية افتراضات الاختبارات المعلمية. إذا لم تستوفِ البيانات الافتراضات بعد
تم استخدام اختبارات غير معلمية مثل اختبار ويلكوكسون. تم استخدام خوارزمية BH للتصحيح-قيم للمقارنات المتعددة. تشير النجوم إلى الفروق المهمة بين مجموعتين:; ; ، n.s غير دال. تم استخدام اختبارات ثنائية الجوانب لاختبار الفرضية البديلة. ج نسبة المعادل الأرضي (LER) (يتم تعريف LER على أنه الفرق بين العائد المرصود والعائد المتوقع، مما يشير إلى كفاءة استخدام الأراضي) والأثر الصافي (NE) يتم تعريف (NE) على أنه مجموع العوائد النسبية الجزئية لكل نوع، مما يشير إلى التأثير الصافي للزراعة المختلطة. تشير زراعة الفول السوداني/الذرة المختلطة إلى ميزة كبيرة في هنان خلال الفترة من 2011 إلى 2015 وفي خمسة مزارع من سهل شمال الصين في عام 2015 تحت ظروف الحقل. تم توفير بيانات المصدر. التربة الكلسية، مما يجعل الحديد غير قابل للذوبان وغير متاح حيوياًتُخفف هذه المشكلة من خلال الزراعة المختلطة مع الذرة، مما يحسن من تغذية الحديد وكفاءة التمثيل الضوئي للفول السوداني (الشكل 1ب) وبالتالي يزيد من الإنتاجية وكفاءة استخدام الأرض. (الشكل 1ج). يُعتقد أن هذه الزراعة المختلطة تقدم وسيلة فعالة ومستدامة لضمان تعزيز الحديد والزنك بيولوجيًا لمنع الجوع الخفي..
هنا، نستخدم زراعة الفول السوداني/الذرة كنظام نموذجي لتوضيح الآلية الدقيقة التي تؤدي إلى زيادة تغذية الحديد، مع التركيز بشكل خاص على الدور غير المستكشف بعد لميكروبيوم الجذور. من المعروف أن زراعة الفول السوداني/الذرة تحسن تغذية الحديد من خلال التسهيل المباشر تحت الأرض عبر إفرازات الجذور.تظهر النباتات العليا استجابتين جزيئيتين وفسيولوجيتين بديلتين ناتجتين عن نقص الحديد.الفول السوداني يتبع الاستراتيجية I (النباتات ثنائية الفلقة والنباتات أحادية الفلقة غير العشبية) ويقوم بتقليل الحديد (III) مباشرة إلى الحديد (II) تليها امتصاص الحديد (II).على النقيض من ذلك، يتبع الذرة الاستراتيجية الثانية (أحادية الفلقة العشبية) وتفرز الفيتوسيدروفورات من عائلة حمض الموجينيك (MAs) لتخلب الحديد غير القابل للذوبان Fe(III) تليها امتصاص معقد MAs-Fe(III).تتفوق الذرة على الفول السوداني في اكتساب الحديد في التربة القلوية لأن الاستراتيجية الثانية أقل حساسية للارتفاعتم اقتراح أن زراعة المحاصيل المتداخلة تفيد الفول السوداني لأنها تعزز إفراز حمض الديوكسي ميوجينك (DMA) من جذور الذرة، مما يحل المزيد من الحديد (III) في التربة، والذي يتم امتصاصه بعد ذلك كمعقد DMA-Fe(III) بواسطة نباتات الفول السوداني القريبة..
نموذج التفاعل الحالي بين النباتات لاستخدام الحديد لا يدمج تفاعلات الجذور مع الميكروبيوم. وهذا أمر مدهش لأنه من المعروف جيدًا أن ميكروبيوم منطقة الجذور حاسم لكل من لياقة النبات واكتساب الحديد.. على وجه التحديد، تفرز النباتات من الاستراتيجية الأولى والثانية إفرازات جذرية مختلفة في ظل ظروف نقص الحديد، لاستقطاب أنواع مختلفة من البكتيريا الجذرية التي تحسن تغذية النبات في الاكتساب العام والحديد بشكل خاصآلية شائعة لاكتساب الحديد في هذه البكتيريا المفيدة تعمل من خلال إفراز السايدروفورات لحل المواد غير القابلة للذوبانلذلك، نفترض أن زراعة الفول السوداني والذرة معًا يمكن أن تعدل الميكروبيوم في منطقة الجذور من خلال قرب جذور النباتات، مما قد يعزز خلط إفرازات الجذور، وأعضاء الميكروبيوم، والحديدات الخاصة بهم، وبالتالي تحسين فرص الحصول على الحديد.
لاختبار فرضيتنا، قمنا أولاً بفحص ما إذا كانت زيادة اكتساب الحديد في الزراعة المختلطة تعتمد على وجود ميكروبيوم جذري وظيفي. بعد ذلك، قمنا بدمج تحليل الميكروبيوم، وتوصيف السلالات الوظيفية، وتحديد المواد، وتجارب الدفيئة والميدان لتأسيس الروابط الميكانيكية بين أنظمة الزراعة، وشبكات تفاعل الجذور والميكروبيوم تحت الأرض، واكتساب الحديد. يسمح لنا تصميم دراستنا بتحديد الكائنات الدقيقة والمواد الأيضية الميكروبية التي تساهم في زيادة كفاءة استخدام المغذيات وإنتاجية المحاصيل من خلال الزراعة المختلطة.
النتائج
تساهم الميكروبيوم في منطقة الجذور في تحسين تغذية الحديد للفول السوداني المزروع بالتداخل مع الذرة
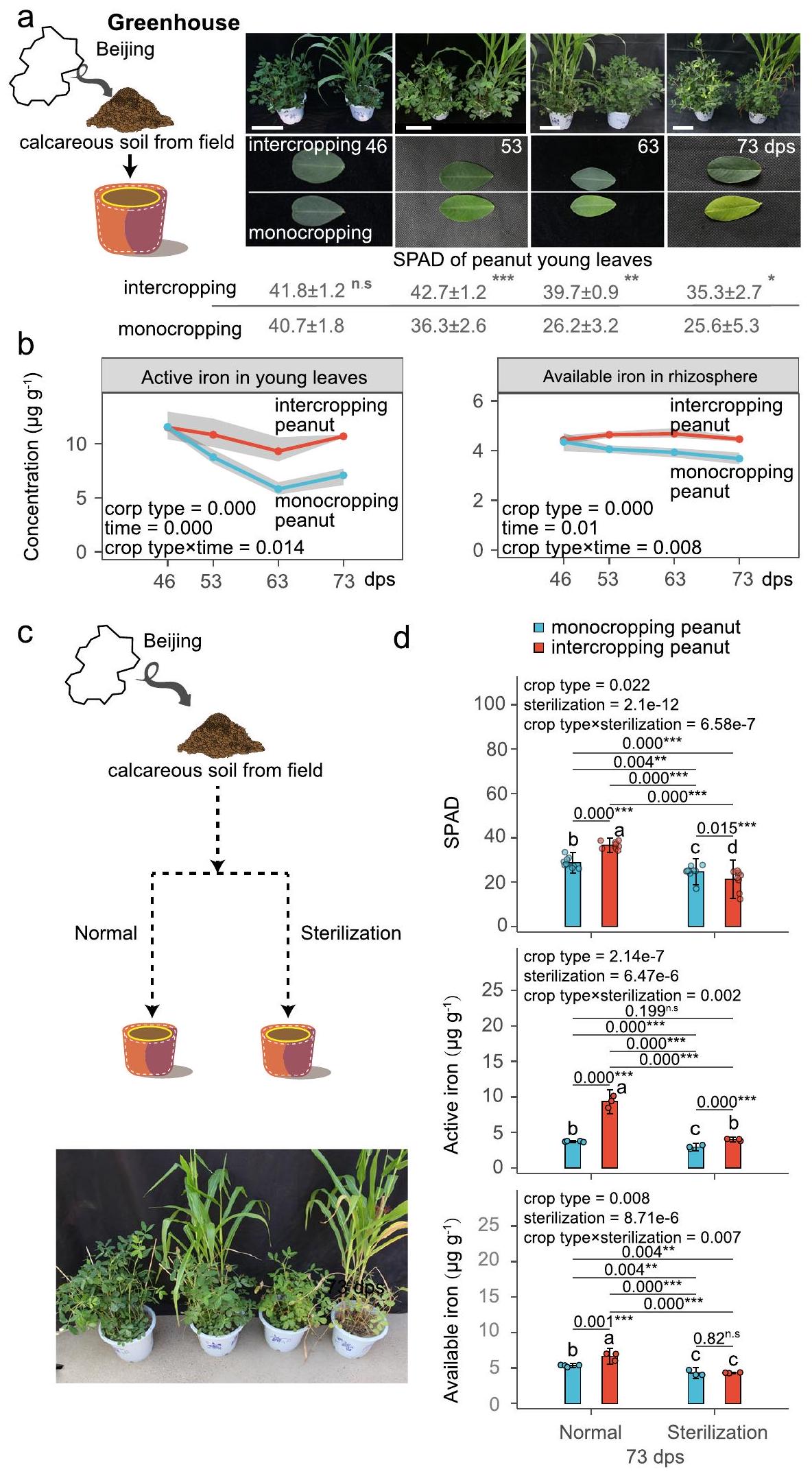
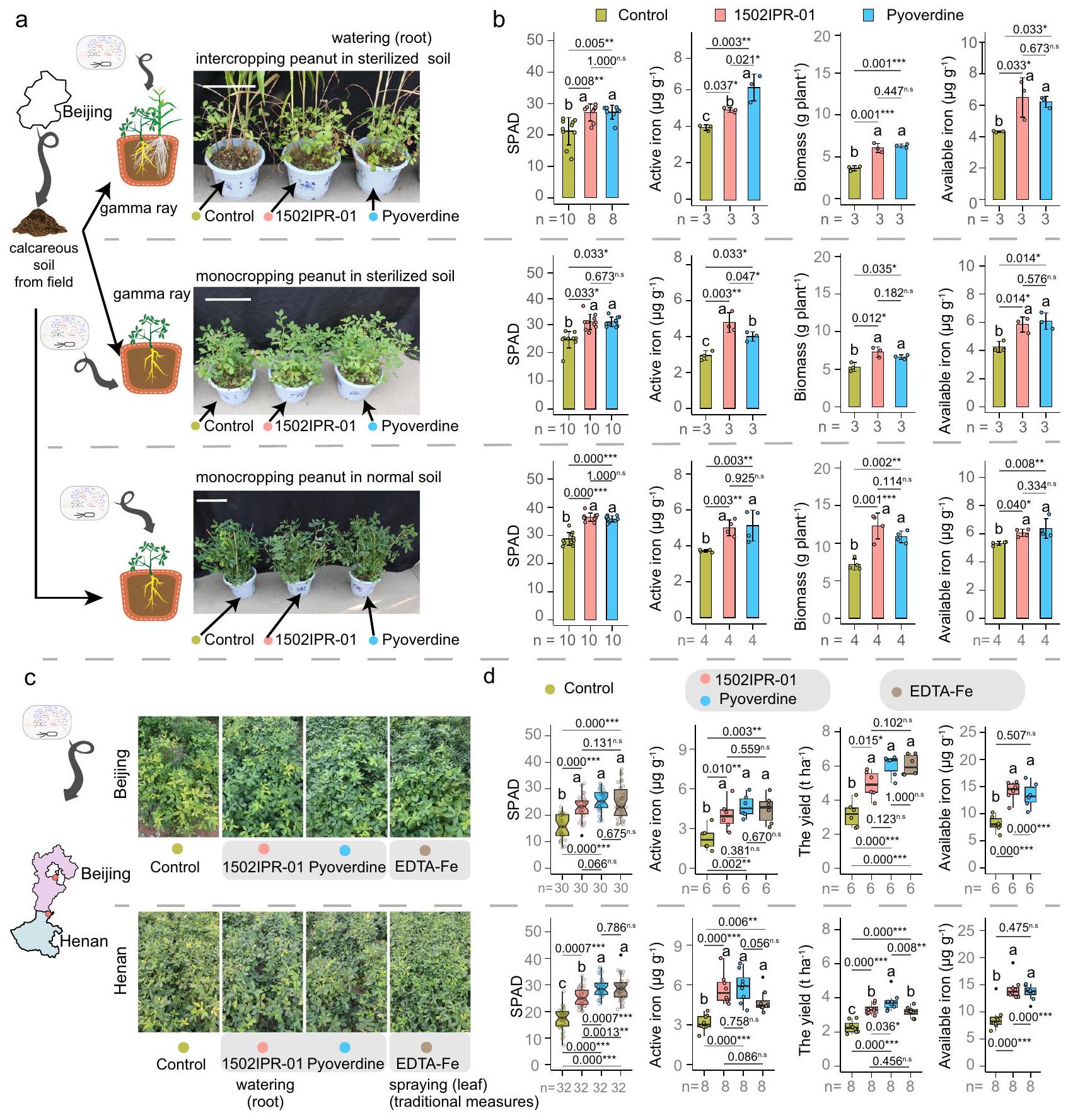
في التجربة الأولى، استكشفنا ما إذا كانت زراعة الفول السوداني والذرة معًا مفيدة في ظروف البيوت الزجاجية في التربة التي تعاني من نقص الحديد بشكل طبيعي. وجدنا أن زراعة الفول السوداني والذرة معًا قد حسنت بالفعل من تغذية الحديد في الفول السوداني (الشكل 2 أ، ب)، ولكن ليس في الذرة (الشكل التكميلي 1). على وجه التحديد، زادت زراعة الفول السوداني والذرة معًا من مستويات الكلوروفيل (قيمة تحليل التربة والنبات (SPAD)،إلى ) وتركيز الحديد النشط ( إلىمؤشرًا على مدى نقص الحديد) في الأوراق الشابة بدءًا من 53 يومًا بعد الزراعة (شكل 2 أ، ب).
مقارنة بزراعة الفول السوداني بشكل أحادي، كان هناك زيادة ملحوظة في تركيز الحديد المتاح في منطقة الجذور.أي، الحديد يتواجد بشكل يمكن أن تمتصه النبات) (الشكل 2ب). في المقابل، بدأت زراعة الفول السوداني الأحادي تظهر أعراض نقص الحديد في الأوراق الصغيرة بعد 53 يومًا من الزراعة، والتي تفاقمت تدريجياً مع مرور الوقت (الشكل 2أ، ب).
لاختبار ما إذا كان الميكروبيوم في منطقة الجذور متورطًا في تحسين تغذية الحديد الملحوظ في زراعة الفول السوداني بالتداخل، قمنا بتكرار تجاربنا في تربة عادية (غير معقمة) وتربة معقمة (الشكل 2c). بالنسبة لجميع المعايير الثلاثة المقاسة – SPAD، الحديد النشط في الأوراق والحديد المتاح في منطقة الجذور – كانت هناك تفاعلات إحصائية كبيرة بين معالجات التربة (المعقمة مقابل العادية).
الشكل 2 | الميكروبيوم في منطقة الجذور يساهم في تحسين تغذية الحديد للفول السوداني المزروع بالتداخل مع الذرة. أ) الظواهر وقيم تحليل التربة والنبات (SPAD) للنباتات والأوراق الصغيرة من الفول السوداني المزروع بشكل منفرد والفول السوداني المزروع بالتداخل.، و 73 يومًا بعد الزراعة (dps). الجدول يوضح قيم SPAD في أوراق الفول السوداني الصغيرة. القيم تظهر المتوسطات SD ( عينات مستقلة بيولوجياً و-القيم هيلـ 46 و 53 و 63 و 73 يوم بعد الزراعة). الشريط يشير إلى 32 سم. ب الحديد النشط في الأوراق الشابة والحديد المتاح (الذي يشير إلى كمية الحديد القابلة للاستخدام من قبل النباتات) في منطقة الجذور للفول السوداني المزروع في نظام الزراعة الأحادية والفول السوداني المزروع في نظام الزراعة المختلطة في الأواني. النقاط والمناطق المظللة تظهر القيم المتوسطة وفترة الثقة 95% من أربع تكرارات بيولوجية مستقلة، على التوالي. ج الشكل الظاهري للفول السوداني المزروع في نظام الزراعة الأحادية والفول السوداني المزروع في نظام الزراعة المختلطة في تربة جيرية معقمة وعادية عند 73 يوم بعد الزراعة. د قيم SPAD والحديد النشط في الأوراق الشابة والحديد المتاح في منطقة الجذور للفول السوداني عند 73 يوم بعد الزراعة المزروع في تربة عادية أو معقمة. الأشرطة مع أشرطة الخطأ تمثل المتوسط. وأنواع الزراعة المختلطة (زراعة الفول السوداني بالتداخل مقابل زراعة الفول السوداني الأحادية)، مما يوضح أن زراعة الفول السوداني بالتداخل كانت مفيدة فقط وزادت من توفر الحديد للفول السوداني في وجود ميكروبيوم الجذور الوظيفي (الشكل 2d).
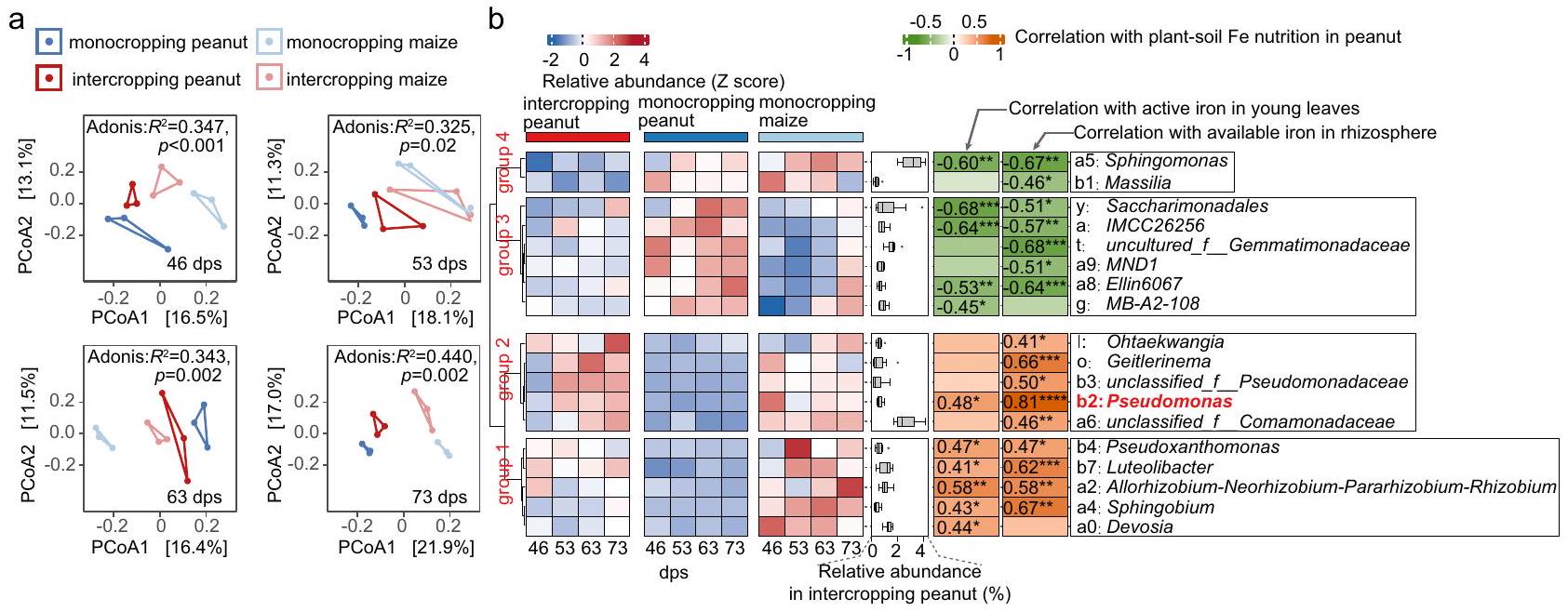
بكتيريا الزائفة – المعززة من الذرة إلى الفول السوداني – مرتبطة بتحسين تغذية الحديد تغيير زراعة المحاصيل يؤثر على ميكروبيوتا الجذور. نظرًا لأن الميكروبيوم الجذري ضروري لتحسين تغذية الحديد في زراعة الفول السوداني بالتداخل، افترضنا أن زراعة المحاصيل بالتداخل تحسن من اكتساب الحديد من خلال تعديل ميكروبيوم الفول السوداني. لاختبار فرضيتنا، استخدمنا تسلسل أمبليكون 16S rRNA لتوصيف الميكروبيوم الجذري لنباتات الفول السوداني والذرة من الزراعة الأحادية وزراعة المحاصيل بالتداخل في أربع نقاط زمنية ( و 73 يوم بعد الزراعة، الشكل التوضيحي 2). شكلت مجتمعات البكتيريا الجذرية أربعة تجمعات متميزة (زراعة الفول السوداني بشكل منفرد، زراعة الذرة بشكل منفرد، زراعة الفول السوداني بالتداخل، زراعة الذرة بالتداخل) في جميع المراحل وأظهرت أكبر تباين عند 73 يوم بعد الزراعة (الشكل 3أ). بينما كانت البكتيريا لم تتأثر تنوع المجتمعات البكتيرية في منطقة الجذور بالزراعة المختلطة (الشكل التكميلي 3أ)، لكننا لاحظنا أن الميكروبيومات الخاصة بالفول السوداني المزروع مع الذرة أكثر تشابهًا من الميكروبيومات الخاصة بالفول السوداني المزروع بمفرده والذرة المزروعة بمفردها (الشكل 3أ والشكل التكميلي 3ب). تشير هذه النمطية إلى أن أعضاء الميكروبيوم الخاص بالذرة والفول السوداني يتم تبادلهم، ربما من خلال الشبكة الجذرية المتداخلة بين الأنواع التي تتشكل مع الزراعة المختلطة (الشكل التكميلي 4). بشكل عام، تكشف نتائجنا أن الزراعة المختلطة تحفز كل من تغيير في تركيب الميكروبيوم وتقارب بين الميكروبيومات الخاصة بالفول السوداني والذرة.
البسودوموناس هو أقوى علامة حيوية مرتبطة بتحسين تغذية الحديد. لاكتشاف الأنواع البكتيرية الرئيسية المرتبطة بتحسين تغذية الحديد، استخدمنا تحليل التمييز الخطي لحجم التأثير (LefSe). حددنا 10 علامات حيوية على مستوى الجنس (درجات LDA المطلقة > 3.0 و ) التي كانت غنية في زراعة الفول السوداني بالتداخل مقارنة بزراعة الفول السوداني الأحادي (الشكل التكميلي 5) وكانت أكثر وفرة في زراعة الذرة الأحادية مقارنة بزراعة الفول السوداني الأحادي (الشكل 3ب). وبالتالي، فإن هذه الأنواع هي المرشحة الرئيسية لتكون غنية بالتبادل من الذرة إلى الفول السوداني في الزراعة بالتداخل. كما وجدنا أن وفرة تلك الأنواع العشر كانت مرتبطة إيجابياً إما بالحديد النشط في الأوراق الشابة، أو بتوافر الحديد في منطقة الجذور، أو كلا المقياسين (الشكل 3ب، الشكل التكميلي 6 الذي يظهر الارتباط الإيجابي بين مقياسي الحديد). بينما أظهرت خمسة أنواع (Pseudomonas، Pseudoxanthomonas، Luteolibacter، Allorhizobium-Neorhizobium-Para-rhizobium-Rhizobium، Sphingobium) ارتباطات إيجابية مع كلا مقياسي الحديد، كان لـ Pseudomonas أقوى ارتباط (معامل سبيرمانمع توفر الحديد في منطقة الجذور (الشكل 3ب والبيانات التكميلية 1). وبالتالي، تكشف تحليلاتنا عن خمسة أنواع رئيسية قد توجه تحسين تغذية الحديد في زراعة الفول السوداني بالتداخل، مع كون بكتيريا Pseudomonas هي المرشح الأول. الانحراف المعياري (SD) (عينات مستقلة بيولوجيًا لقيمة SPAD وللحديد النشط والحديد المتاح) والنقاط تمثل القيم الفردية. لـ، و نوع المحصول يشير إلى الزراعة المختلطة أو الزراعة الأحادية. بارامترية الطالب-تم استخدام اختبار t و ANOVA مع اختبار LSD كاختبار لاحق للبيانات التي كانت موزعة بشكل طبيعي ولديها تباينات متجانسة. تم تحويل توزيعات البيانات المنحرفة أو البيانات ذات تباين غير متجانس باستخدام خوارزمية BoxCox لتلبية افتراضات الاختبارات المعلمية. إذا لم تستوفِ البيانات الافتراضات بعد التحويل، تم استخدام اختبارات غير معلمية مثل اختبار ويلكوكسون، واختبار كروسكال-واليس مع اختبار دانييت T3 أو اختبار شيرير-راي-هار. تم استخدام خوارزمية BH للتصحيح.-قيم للمقارنات المتعددة. تشير النجوم إلى الفروق المهمة بين مجموعتين:; ; غير مهم. الحروف المختلفة تشير إلى اختلافات مهمة بين المجموعات. تم استخدام اختبارات ثنائية الجانب لاختبار الفرضية البديلة. تم توفير بيانات المصدر.
علاوة على ذلك، وجدنا أن الزراعة المختلطة تؤدي أيضًا إلى تغييرات في الميكروبيوم في منطقة جذور الذرة. على وجه الخصوص، حددنا ثلاثة أجناس (أزوتوباكتر، روبيلليمايكروبيوم وبرايوباكتير) التي كانت غنية في الذرة المزروعة بشكل مختلط مقارنة بالذرة المزروعة بشكل أحادي (الشكل التوضيحي 7).
بكتيريا البكتيريات الزائفة من الزراعة المختلطة تظهر قدرة عالية على إفراز السايدروفور
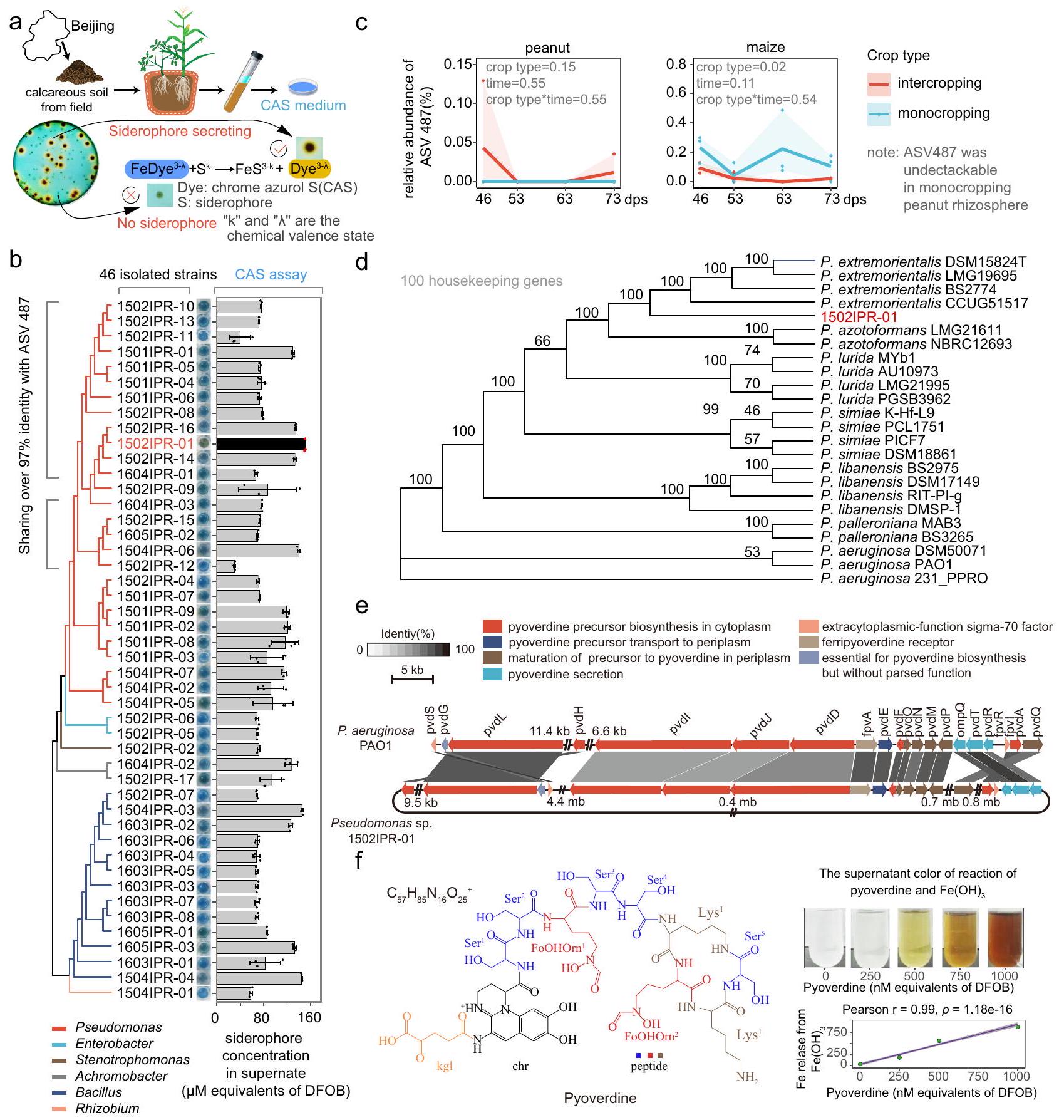
نظرًا للقدرة العالية للسايدروفورات على إذابة الحديد، افترضنا أن السايدروفورات التي تفرزها البكتيريا الجذرية في الزراعة المختلطة قد تكون مسؤولة عن زيادة توافر الحديد الحيوي في منطقة جذر الفول السوداني. لاختبار هذه الفرضية، قمنا بعزل بكتيريا جذرية تفرز السايدروفورات من زراعة الفول السوداني المختلطة باستخدام اختبار صفيحة الكروم أزول S (CAS) (الشكل 4أ). قمنا بعزل أكثر من 300 سلالة من البكتيريا الجذرية التي تفرز السايدروفورات، من بينها 46 سلالة كانت لديها نشاط عالٍ بشكل خاص في إفراز السايدروفورات (قطر الهالة على صفيحات CAS، الشكل 4ب). من بين أكبر المنتجين،كانت الأنواع Pseudomonas spp. (وفقًا لتسلسل 16S rRNA الخاص بها)، وكانت الغالبية العظمى من هذه السلالات (63.0%) تمتلك هويات تسلسلية تزيد عن 97% مع متغير تسلسل الأمبليكون التمثيلي (ASV487) الذي تم الحصول عليه بواسطة خوارزمية DADA2، والتي تلتقط مجموعة معينة من Pseudomonas spp. (الشكل 4ب والجدول التكميلي 1). من المهم أن ASV487 لم يكن قابلًا للاكتشاف في زراعة الفول السوداني الأحادي، ولكنه كان قابلًا للاكتشاف في زراعة الفول السوداني المختلط وكانت وفرة نسبته أعلى في زراعة الذرة الأحادية مقارنة بزراعة الذرة المختلطة (الشكل 4ج). بالإضافة إلى ذلك، تم عزل سلالات مشابهة لـ ASV487 في منطقة جذور الفول السوداني المختلط في مرحلة حاسمة من التحول الغذائي للحديد. ) وفي المرحلة التي كانت فيها فجوات تغذية الحديد بين زراعة المحاصيل الأحادية وزراعة الفول السوداني المتداخلة في أوسع حالاتها ( 73 يومًا بعد الزراعة ) (الجدول التكميلي 1). تشير هذه النتائج إلى أن ASV487 يمثل مجموعة من سلالات بكتيريا Pseudomonas التي تفرز كميات كبيرة من السايدروفور والتي تم إثراؤها بشكل متبادل من الذرة إلى الفول السوداني خلال الزراعة المتداخلة وترتبط بتحسين تغذية الحديد في الفول السوداني.
بعد ذلك، ركزنا على Pseudomonas sp. 1502IPR-01، وهو سلالة تعتبرمطابق لـ ASV487 وكان لديه أعلى قدرة على إفراز السايدروفور بين السلالات المعزولة (الشكل 4ب والشكل التكميلي 8). استخدمنا Pseudomonas sp. 1502IPR-01 كسلالة تمثيلية لاستكشاف دورها في تحسين تغذية الحديد في زراعة الفول السوداني بالتداخل. قمنا بتسلسل الجينوم الكامل لـ Pseudomonas sp. 1502 IPR-01 ووجدنا أن 1502IPR-01 مشابه لـ. extremorientalis (الشكل 4d والشكل التوضيحي 9). من خلال البحث في جينومه عبر تحليل المجموعات، اكتشفنا مجموعة تخليق وإفراز البيوفرادين التي تشترك في 51.4-84.1% من هوية تسلسل البروتين مع الجينات المتماثلة لـ. الأيروجينوزا PAO1، السلالة النموذجية لأبحاث البيوفرودين (الشكل 4e والجدول التكميلي 2).
لتأكيد أن Pseudomonas sp. 1502IPR-01 يفرز بالفعل البايفيردين كأهم سلاسل الحديد، قمنا بعزل المركب المخلبي للحديد (الشكل التكميلي 10) من السائل الفائق، بعد الزراعة في وسط محدود الحديد. كشفت تجربتنا أنقدرة السوبرناتانت على ربط الحديد كانت
الشكل 3 | الزراعة المختلطة تؤدي إلى تغييرات في تركيبة مجتمع البكتيريا الجذرية وتظهر أن بكتيريا Pseudomonas هي واحدة من الأنواع المرشحة الرئيسية المعنية بتحسين تغذية الحديد في الفول السوداني. أ تحليل المكونات الرئيسية غير المقيد (PCoA) باستخدام مسافات UniFrac (التي تقيس المسافة بين المجتمعات بناءً على السلالات التي تحتويها) لميكروبيومات البكتيريا الجذرية المجمعة من زراعة الفول السوداني الأحادي، وزراعة الفول السوداني المختلط، وزراعة الذرة المختلطة، وزراعة الذرة الأحادية. تظهر نتائج تحليل التباين المتعدد المتغيرات القائم على التباديل (PERMANOVA، المعروف أيضًا باسم Adonis). تم استخدام الوفرة النسبية في تحليل PcoA وAdonis.علامة جينية تميز بين زراعة الفول السوداني في نظام الزراعة الأحادية والزراعة المختلطة، تم تحديدها بواسطة تحليل التمييز الخطي وحجم التأثير، LefSe (LDA) و ). تظهر الكميات الديناميكية النسبية للأنواع البيولوجية المؤشر في زراعة الفول السوداني بالتداخل، وزراعة الفول السوداني الأحادي.
وزراعة الذرة كمعدل ز-score موحد للقيمة المتوسطة لثلاث عينات بيولوجية مستقلة، وتم تصويرها كخريطة حرارية تمتد من القيم المنخفضة (الأزرق) إلى القيم العالية (الأحمر) من z-scores. يتم عرض الوفرة النسبية لكل جنس باستخدام مخططات الصندوق: المركز = الوسيط، وحدود الصندوق = الربع الأول والثالث، والشعيرات = الحد الأدنى والحد الأقصى. الجدول يظهر معامل سبيرمان.الارتباطات بين الوفرة النسبية واثنين من المقاييس التي تصف حالة تغذية الحديد في النباتات، موضحة كخريطة حرارية تمتد من الارتباطات السلبية (الخضراء) إلى الارتباطات الإيجابية (الوردية). كانت تحليلات الارتباط قائمة على 24 عينة مستقلة. يتم الإشارة إلى الارتباطات المهمة بنجوم:; ; ; ن.س غير دالة. سبيرمان الدقيقتم إدراج القيم ذات المستويات المهمة. عدد العينات البيولوجية المستقلة لبيانات خريطة الحرارة هو ثلاثة. تم استخدام اختبارات ثنائية الجانب لاختبار الفرضيات البديلة. تم توفير بيانات المصدر. ناتج عن السايدروفورات (الجدول التكميلي 3). أسفر توضيح الهيكل والتحليل الكيميائي (الشكل 4f، الأشكال التكميلية 11-21 والجدولان التكامليان 4 و5، التفاصيل في قسم “الطرق”) عن صيغة كيميائية لـوتؤكد البنية الكيميائية أن السايدروفور الرئيسي الذي تفرزه بكتيريا Pseudomonas sp. 1502IPR-01 هو بايوفيردين (الشكل 4f).
بعد ذلك، قمنا بتقييم نشاط البايفيردين المكتشف لربط III) من غير القابل للحل مصدر في تعليق. III تشير عملية الخلب إلى تغيير في اللون من الشفاف إلى الأصفر إلى البني (الشكل 4f). وجدنا أن تركيز الحديد المخلب في المحلول زاد مع كمية متساوية من البايفيردين المضاف، مما يتطابق مع تغيير اللون لمركب التحكم بايفيردين-Fe(III) (الشكل 4f والشكل التكميلي 10c). باختصار، تحدد نتائجنا البايفيردين كالسيدروفور الرئيسي الذي تفرزه بكتيريا Pseudomonas sp. 1502IPR-01، وتظهر قدرته العالية على إذابة Fe(III) من المواد غير القابلة للذوبان.الذي يتوافق مع الشكل الرئيسي للحديد في التربة الكلسية.
بكتيريا Pseudomonas sp. 1502IPR-01 وبيوفيردينها تحسن من تغذية الفول السوداني بالحديد في تجارب الدفيئة والميدان
للتحقق تجريبيًا من أن بكتيريا الزائفة هي نوع أساسي يحسن من تغذية الحديد من خلال إفراز البايفيردين، قمنا بإجراء تجارب في البيوت الزجاجية والميدان. في تجارب البيوت الزجاجية، زرعنا الفول السوداني في زراعة أحادية وزراعة مختلطة في تربة جيرية معقمة إما بدون معالجة كتحكم، أو مع بكتيريا الزائفة sp. 1502IPR-01 أو بايفيردين خالي من الحديد كعلاجات. زادت كلا المعالجتين بشكل كبير من قيم SPAD، وتركيز الحديد النشط في الأوراق الشابة وفي منطقة الجذور، وحسنت من كتلة الفول السوداني في الزراعة الأحادية والزراعة المختلطة (الشكل 5أ، ب). لاحظنا نفس التحسينات الكبيرة عند تكرار التجربة في تربة غير معقمة عادية في زراعة الفول السوداني الأحادية (الشكل 5ب). وبالتالي، يمكن أن تعوض بكتيريا الزائفة sp. 1502IPR-01 ومعالجات البايفيردين عن نقص الحديد في الفول السوداني في غياب ميكروبيوم وظيفي في زراعة الفول السوداني بالتداخل وزراعة الفول السوداني الأحادي في التربة العادية. على النقيض من ذلك، لم تتحسن تغذية الحديد في الذرة عند معالجتها بـ Pseudomonas sp. 1502IPR-01 أو بايوفيردين (الشكل التوضيحي التكميلي 22). ومع ذلك، زادت الكتلة الحيوية للذرة بشكل كبير مع كلا العلاجين في التربة المعقمة والعادية في زراعة الذرة بالتداخل (الشكل التوضيحي التكميلي 22)، مما يشير إلى أن الذرة تستفيد من زراعة الفول السوداني بالتداخل بطرق أخرى غير تحسين تغذية الحديد.
بعد ذلك، بحثنا فيما إذا كانت تطبيقات Pseudomonas sp. 1502IPR-01 وpyoverdine الخالية من الحديد فعالة أيضًا كعلاجات في الميدان. مع زراعة الفول السوداني بالتداخل في التربة العادية، لم يكن لتطبيق Pseudomonas sp. 1502IPR-01 وpyoverdine الخالية من الحديد أي تأثير على تغذية الحديد والكتلة الحيوية (الشكل التوضيحي 23)، ربما لأن الزراعة بالتداخل وحدها تكفي للتخفيف من نقص الحديد في الفول السوداني. لذلك، ركزنا بشكل حصري على زراعة الفول السوداني بشكل أحادي وعالجنا النباتات إما بـ Pseudomonas sp. 1502IPR-01 أو pyoverdine (كلاهما تم تطبيقه عن طريق ري الجذور) في موقعين في سهل شمال الصين، الذي يتميز بالتربة الكلسية. في كلا الموقعين، زادت العلاجات بشكل كبير من قيم SPAD (زيادة: 45.9-67.6%)، تركيز الحديد النشط في الأوراق الشابة (78.2-107.5%)، توفر الحديد في منطقة الجذور (53.6-73.2%) وإنتاج المحاصيل (44.8-89.8%)، مقارنة بالنباتات الضابطة التي لم تتلق أي علاج (الشكل 5c، d). كما طبقنا علاجًا تقليديًا يستخدمه المزارعون المحليون، والذي يتضمن الرش الورقي بـ EDTA-Fe، وهو خالب للحديد صناعي. لاحظنا أن معالجة النباتات إما بـ Pseudomonas sp. 1502IPR-01 أو pyoverdine زادت من تغذية الحديد وإنتاج الفول السوداني إلى حد مشابه، وفي حالة واحدة حتى أكثر من علاج EDTA-Fe التقليدي (الشكل 5d). تشير هذه النتائج إلى أن pyoverdine المفرز أو المضاف من Pseudomonas sp. 1502IPR-01 يمكن أن يخلب Fe(III) بشكل فعال في التربة الكلسية في الميدان، وأن هذه الآلية مسؤولة عن تحسين تغذية الحديد وإنتاج المحاصيل في الفول السوداني. وبالتالي، يمكن اعتبار pyoverdine كسماد للحديد للزراعة المستدامة دون الحاجة إلى إمدادات خارجية من الحديد.
الشكل 4 | البكتيريا الجذرية المفرزة للسيدروفور الوظيفية وخصائص السيدروفور. أ رسم تخطيطي يوضح البروتوكول المستخدم لعزل
عزل Pseudomonas sp. 1502IPR-01 مع ممثلين آخرين من سلالات Pseudomonas spp. بناءً على 100 جين منزلي كعلامات فيلو جينية. تم بناء شجرة الإجماع من 1000 شجرة bootstrap. الأرقام على الفروع تظهر دعم bootstrap (%). e جينات إفراز وامتصاص تخليق البيوفيردين لـ Pseudomonas sp. 1502IPR-01. التسلسلات المشتركة بين Pseudomonas sp. 1502IPR-01 وPseudomonas aeruginosa PAO1 متصلة بواسطة شرائط تتدرج وفقًا لمستوى هويتها، من الأسود ( ) إلى الأبيض ( ). يشير لون السهم إلى وظيفة الجينات. البنية الكيميائية المفترضة للبايوفيردين من بكتيريا Pseudomonas sp. 1502IPR-01 وقدرتها على تشكيل معقد مع الحديد (III) من الرواسب. تُظهر الألواح تغير اللون في السائل العلوي الناتج عن البايفيردين والارتباط (معامل الارتباط بيرسون) بين إطلاق الحديد من وتركيز البيوفرادين في خليط التفاعل. تشير الخطوط الأرجوانية والمنطقة الرمادية إلى خط الانحدار الخطي وفترة الثقة 95%، على التوالي. تشير النقاط الخضراء إلى القيمة المتوسطة عبر ثلاث عينات مستقلة. تم استخدام اختبارات ثنائية الجانب لاختبار الفرضية البديلة. تم توفير بيانات المصدر.
الشكل 5 | بكتيريا Pseudomonas sp. 1502IPR-01 وموادها الجاذبة للحديد (بايوفيردين) تمنع الكلوروز الناتج عن نقص الحديد وتحسن نمو الفول السوداني وإنتاجه. أ الأنماط الظاهرية
لـالأعمدة وأشرطة الخطأ تمثل المتوسط الانحراف المعياري (SD) والنقاط تمثل القيم الفردية. من أجل تظهر خطوط المركز في مخطط الصندوق الوسيط، وتظهر حدود مخطط الصندوق الربع الأول Q1 والربع الثالث Q3، وتظهر الشعيرات 1.5 (Q3-Q1) أسفل وفوق Q1 وQ3، والنقطة السوداء تمثل القيم الشاذة والنقاط المفتوحة تمثل القيم الفردية، على التوالي. بالنسبة لـ و البيانات تتبع توزيعًا طبيعيًا وتجانس التباين، يتم استخدام تحليل التباين (ANOVA) مع اختبار LSD كاختبار لاحق في الاختبارات المعلمية. بالنسبة للبيانات المنحرفة أو البيانات ذات عدم تجانس التباين، يتم تحويل البيانات باستخدام خوارزمية BoxCox للاختبار المعلمي. يتم استخدام اختبار كروسكال مع اختبار Dunnett T3 إذا لم تستوفِ البيانات افتراضات الاختبار المعلمي بعد خوارزمية BoxCox. تشير الحروف المختلفة إلى اختلافات معنوية بين المجموعات. يتم إجراء تصحيحات للاختبارات المتعددة بواسطة خوارزمية BH. تشير النجوم إلى اختلافات معنوية بين مجموعتين:; ; غير دال إحصائيًا. بالنسبة لجميع المعايير،يمثل عدد العينات المستقلة بيولوجيًا. تم استخدام اختبارات ثنائية الجوانب لاختبار الفرضيات البديلة. تم توفير بيانات المصدر.
تكشف تجارب التحقق أن البايفيردين يؤثر مباشرة على استقلاب الحديد في النباتات
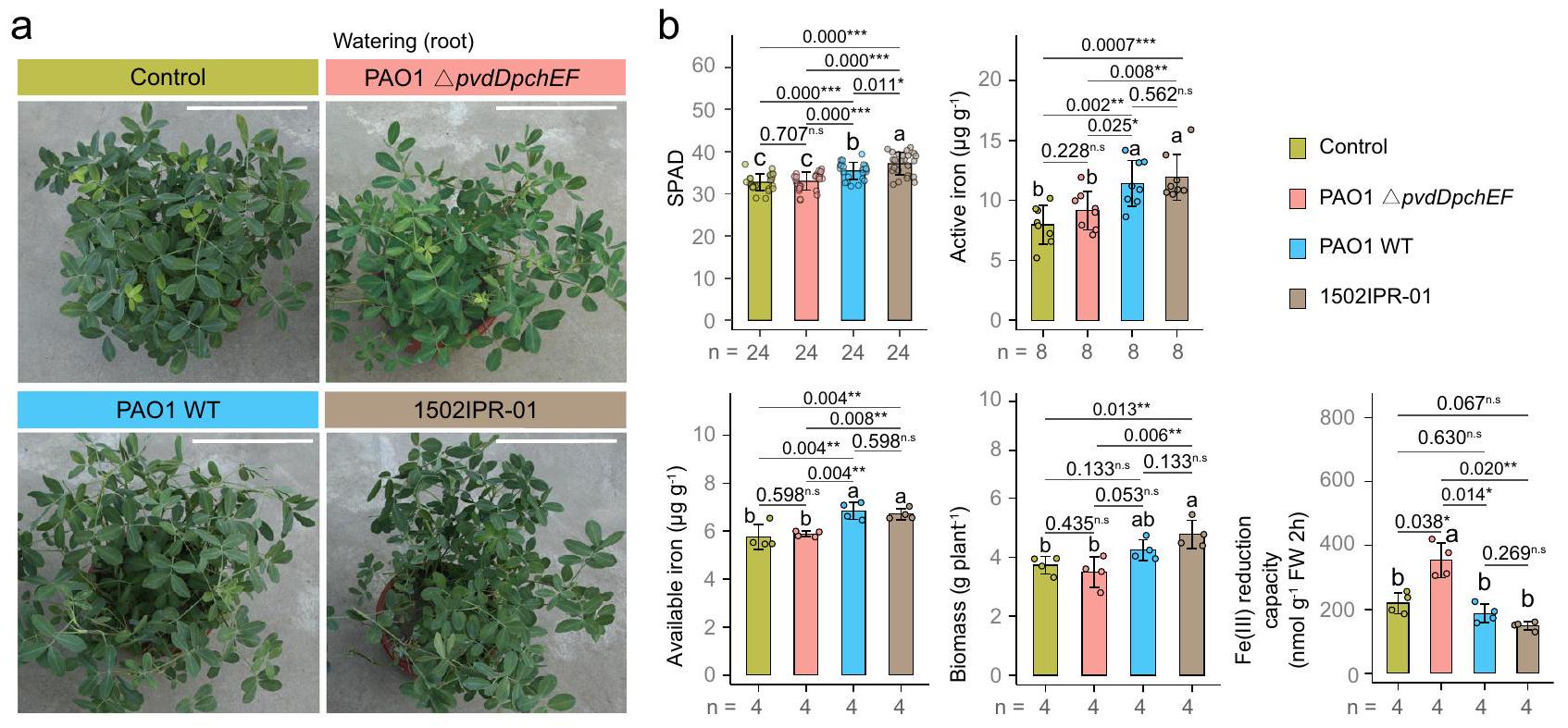
للتحقق من أن البايفيردين يساهم في تحسين تغذية الحديد في الفول السوداني، أجرينا تجارب مع طفرات محددة خالية من البايفيردين (الشكل 6أ). كل من السلالة المخبرية P. aeruginosa PAO1 والعزلة الطبيعية Pseudomonas sp. 1502IPR-01 حسنت بشكل كبير تغذية الحديد في الفول السوداني وزيادة الكتلة الحيوية في التربة العادية. على النقيض من ذلك، فإن المعالجة بالطافرة الخالية من السايدروفور PAO1 (الشكل 6ب) لم يكن له أي تأثيرات مفيدة على النباتات، مما يظهر أن البيوفيردين مطلوب لتحسين تغذية الفول السوداني بالحديد.
ثم سألنا عما إذا كانت إفرازات البيوفرادين تؤثر على نشاط أنظمة اكتساب الحديد الخاصة بالنباتات. في الواقع، لاحظنا أن نشاط مختزل الشوارد الحديدية (FCR) في الفول السوداني يرتفع بشكل ملحوظ في وجود الطفرة المفقودة للسايدروفور (الشكل 6ب)، ولكنه ينخفض في وجود سلالات النوع البري المنتجة للبيوفرادين وعند إضافة البيوفرادين (الشكل التكميلية 24). حدثت نفس الاستجابات في الذرة، التي خفضت من الفيتوسايدروفور DMA في وجود Pseudomonas sp. 1502IPR-01 أو البيوفرادين الخاص بها (الشكل التكميلية 24). تسلط هذه التجارب الضوء على أن البيوفرادين يؤثر بشكل مباشر على استقلاب الحديد في النباتات.
نقاش
الزراعة المختلطة مهمة للزراعة المستدامة وأمن الغذاء. ومع ذلك، فإن الآليات الدقيقة التي من خلالها يزيد الزراعة المختلطة من الغلة غالبًا ما تظل غير واضحة. في هذه الدراسة، نجد أن تحسين تغذية الحديد يتم بوساطة الميكروبيوم البكتيري في منطقة الجذور وينشأ من خلال تقارب الميكروبيوم بين نوعي النبات. على وجه التحديد، نجد أن الزراعة المختلطة تمكن البكتيريا المفيدة مثل Pseudomonas spp. من الانتقال من منطقة جذور الذرة إلى منطقة جذور الفول السوداني، حيث تزيد هذه البكتيريا من كمية الحديد المتاح حيوياً من خلال إفراز السايدروفور بايوفيردين. الحديد المرتبط بالبايوفيردين يزيد من تغذية الحديد في نباتات الفول السوداني ويعوض عن الكلوروز الناتج عن نقص الحديد في الأوراق (الأشكال 4-7). لقد ارتبطت السايدروفورات (مثل بايوفيردين) سابقاً بتأثيرات مفيدة للنباتات.، ودراستنا الآن تقدم أدلة تربط بين إنتاج البايفيردين البكتيري في النباتات وتحسين تغذية الحديد في النباتات (الشكل 6). هذه الرؤية تسد فجوة معرفية كبيرة في مجال البحث. من المهم أن نوضح أن التفاعل بين الميكروبات والنباتات الذي تم اكتشافه ضروري لتحسين تغذية الحديد، حيث أن الزراعة المختلطة في غياب ميكروبيوم وظيفي ليست مفيدة (الأشكال 2، 5). تظهر نتائجنا أيضًا كيف أن المعرفة التفصيلية بآليات التفاعل بين الممالك تساعد في تطوير الزراعة المستدامة بيئيًا وأمن الغذاء.
تشير خطتان من الأدلة إلى أن التفاعلات الميكروبية التي لوحظت في تجاربنا في البيوت الزجاجية تحدث في الحقل. أولاً، يمكن لجذور الفول السوداني والذرة أن تتصل بسهولة عبر مسافة لا تقل عن 60 سم في الحقل، مما يتطابق مع النتائج من تجارب الأواني لدينا.. علاوة على ذلك، تحدث أيضًا اتصالات جذرية عبر مسافات مشابهة بين الذرة/الفاصوليا الفولية والذرة/فول الصويا.. وبالتالي، من المحتمل أن يحدث تلاقي الميكروبيوم في منطقة الجذور بناءً على الروابط الجذرية بين الفول السوداني والذرة التي لوحظت في الدفيئة أيضًا في الحقل. ثانيًا، أظهرت تجاربنا الميدانية أن إضافة Pseudomonas sp. 1502IPR-01 أو البايفيردين الخاص بها قد حسنت من تغذية الفول السوداني بالحديد، مما يدل على أن نباتات الفول السوداني يمكن أن تستفيد من الحديد المتاح بواسطة Pseudomonas spp. ومن ثم، فإن تحريك الحديد بواسطة البايفيردين خلال تلاقي الميكروبيوم يمكن أن يكون عنصرًا رئيسيًا في تحسين تغذية الحديد وإنتاجية الفول السوداني المزروع مع الذرة.
عملنا يعزز الرأي القائل بأن بكتيريا الزائفة هي نوع أساسي يشارك في تغذية الحديد للنباتات. نوضح أن بكتيريا الزائفة sp. 1502IPR-01 وبيوفيردينها تحسن تغذية الحديد في الفول السوداني.
الشكل 6 | البايفيردين الذي تفرزه بكتيريا الزائفة هو أمر أساسي لتحسين تغذية الحديد في الفول السوداني والكتلة الحيوية. أ. أنماط الفول السوداني تحت ظروف التحكم وعند معالجتها بسلالة متحورة من بكتيريا الزائفة الزنجارية PAO1 بدون القدرة على إفراز البايفيردين (PAO1النوع البري من بكتيريا الزائفة الزنجارية PAO1 (PAO1 WT) مع القدرة على إفراز البيوفرودين، وبكتيريا الزائفة 1502IPR-01 مع القدرة على إفراز البيوفرودين (جميعها تطبيقات في منطقة الجذور).قيم تحليل التربة والنبات (SPAD)، الحديد النشط في الأوراق الشابة، الحديد المتاح في منطقة الجذور، الكتلة الحيوية وقدرة اختزال الحديد (III) للفول السوداني تحت ظروف التحكم وثلاثة علاجات (PAO1ApvdDpchEF، PAO1 WT و1502IPR-01) في ظروف الدفيئة.الأشرطة وأشرطة الخطأ تمثل المتوسطالانحراف المعياري (SD) والنقاط تمثل القيم الفردية. تحليل التباين (ANOVA) مع
تم استخدام اختبار LSD بعد التحليل عندما كانت البيانات تتبع التوزيع الطبيعي ولديها تباينات متجانسة. بالنسبة لمجموعات البيانات المنحرفة أو البيانات ذات التباينات غير المتجانسة، تم تطبيق خوارزمية تحويل BoxCox. عندما لم تستوفِ البيانات بعد افتراضات الاختبارات المعلمية، تم استخدام اختبار كروسكال-واليس مع اختبار دانييت T3. تشير الحروف المختلفة إلى فروق ذات دلالة إحصائية بين المجموعات. تم إجراء تصحيحات للاختبارات المتعددة بواسطة خوارزمية BH. تشير النجوم إلى فروق ذات دلالة إحصائية بين مجموعتين:، غير مهم. بالنسبة لجميع المعلمات، تمثل n عدد العينات المستقلة بيولوجياً. تم استخدام اختبارات ثنائية الجوانب لاختبار الفرضية البديلة. تم توفير بيانات المصدر.
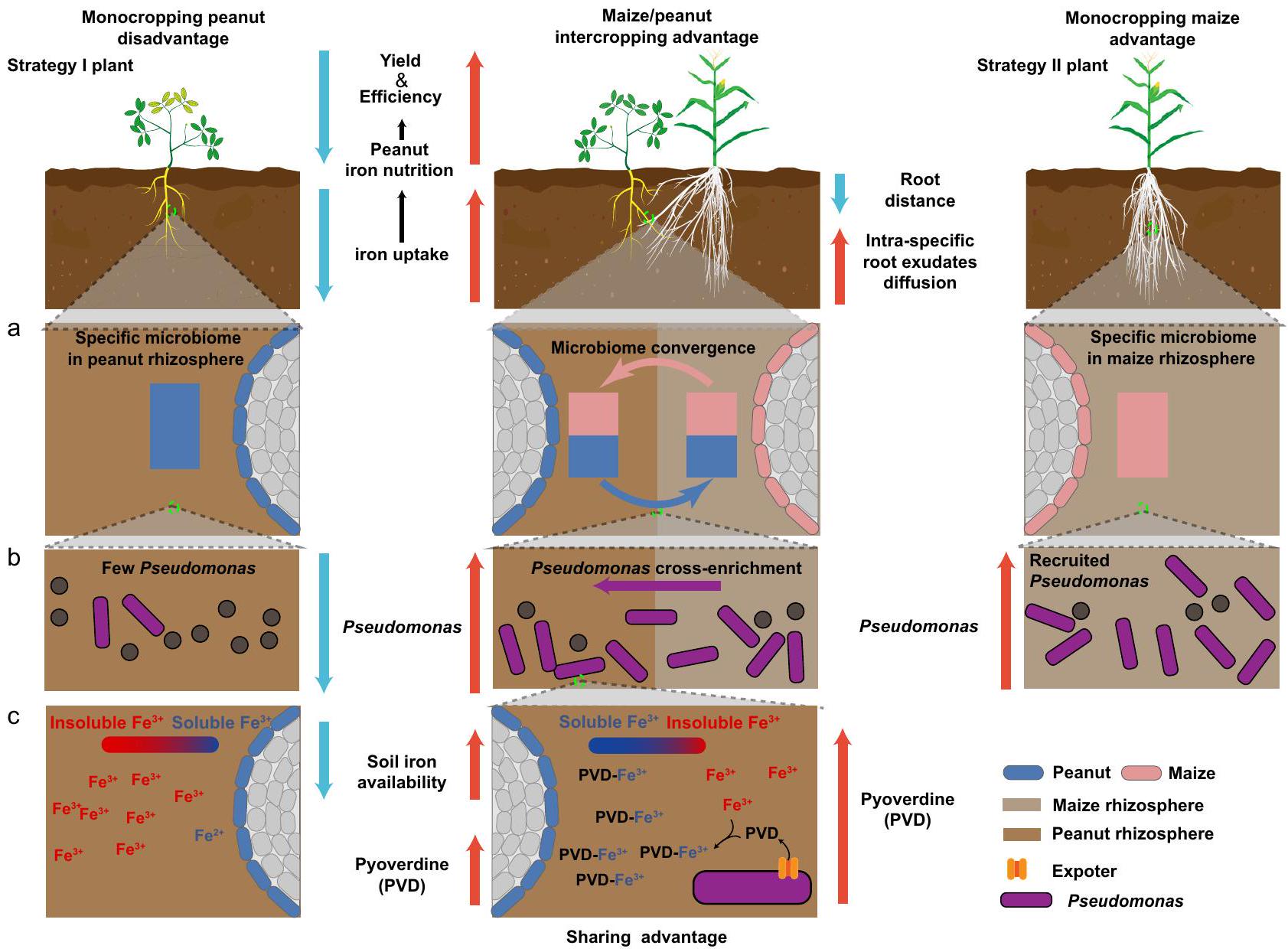
الشكل 7 | نموذج لكيفية إثراء زراعة الفول السوداني/الذرة للـ Pseudomonas spp. في منطقة الجذور لتحسين تغذية الحديد من خلال إفراز السايدروفور بايوفيردين. أ يحدث تلاقي الميكروبيوم في منطقة الجذور في زراعة الفول السوداني/الذرة من خلال تفاعلات الجذور (على مستوى الميكروبيوم الكامل). ب Pseudomonas يتبادل المحصولان الفائدة خلال الزراعة المختلطة من جذور الذرة إلى جذور الفول السوداني (ميكروب وظيفي). تقوم بكتيريا Pseudomonas بإفراز السايدروفور بايوفيردين، الذي يزيد من توفر الحديد في جذور الفول السوداني في الزراعة المختلطة (مستقلب ميكروبي وظيفي). التربة الكلسية في كل من ظروف البيوت الزجاجية والحقول من خلال تقارب الميكروبيوم بين الفول السوداني والذرة (الأشكال 3-6). وقد تم الإبلاغ عن نتائج مماثلة بشأن إثراء بكتيريا Pseudomonas في الفول السوداني المزروع مع الذرة في التربة الحمضية.ومع ذلك، من المهم أن نلاحظ أن تحليلنا كشف عن أعضاء إضافيين من الميكروبيوم في منطقة الجذور الذين كانوا مرتبطين أيضًا بتحسين تغذية الحديد، على سبيل المثال، أعضاء من أجناس Sphingobium وLuteolibacter. لذلك، سيكون من المهم إجراء أعمال تجريبية مع سلالات موصوفة من هذه الأنواع في المستقبل، لتقييم دورها بشكل أكثر دقة. من المحتمل أن تكون التأثيرات المفيدة الملحوظة في الزراعة المختلطة نتيجة لتأثير مجتمع بكتيري مع احتمال أن تكون Pseudomonas spp. هي الرائدة في هذه العملية (الأشكال 3 و4). إن نسب التأثيرات الإيجابية إلى نوع واحد فقط هو بالتأكيد ميزة عندما نهدف إلى تطوير علاجات سماد حيوي للتطبيقات الزراعية.
تشير التأثيرات الإيجابية للبايوفيردين على تغذية الحديد إلى أن هذه الجزيئات تلعب دورًا لم يتم وصفه بعد لمجتمعات الجذور في ظروف الحقل. كشفت الأبحاث السابقة أن بعض السايدروفورات البكتيرية المحددة، بما في ذلك البايوفيردين، يمكن أن تكون مفيدة للنباتات لأنها تقمع مسببات الأمراض النباتية في منطقة الجذور.الآلية المقترحة تنطوي على أن البايفيردينات (وغيرها من السايدروفورات) هي خالطات قوية للحديد، وبما أنها محددة ضريبياً، فإنها يمكن أن تحجز الحديد بعيداً عن مسببات الأمراض التي لا تمتلك المستقبلات المطابقة، وبالتالي تعمل كـ ‘أشياء سيئة عامة’.يمكن أن تؤدي المنافسة على الحديد إلى تعزيز السيطرة على الأمراض النباتية وحماية المحاصيل من خلال الميكروبيومات الطبيعية في منطقة الجذور.في الزراعة المختلطة، يبدو أن البايوفردين يعمل بشكل أساسي كـ ‘سلعة عامة’ تذيب الحديد للنبات وربما أيضًا لأعضاء الميكروبيوم في منطقة الجذور الذين يمتلكون مستقبلات متطابقة.من حيث المبدأ، يمكن أن تلعب البايفيردين دورًا مزدوجًا في الزراعة المختلطة من خلال قمع الأمراض النباتية (أثر سلبي عام) وتحسين تغذية الحديد للنباتات (أثر إيجابي عام). من المهم أن نلاحظ أن النباتات والميكروبات قد تتنافس أيضًا على الحديد. يتم إنتاج السايدروفورات مثل البايفيردين بشكل أساسي بواسطة البكتيريا لتلبية احتياجاتها الخاصة من الحديد. ولكن نظرًا لأن السايدروفورات قابلة للتشتت بشكل كبير، فقد تصبح متاحة وقابلة للاستخدام من قبل النباتات. (الأشكال 5-6). قد يؤدي فقدان السايدروفورات إلى النباتات إلى تعزيز إنتاج السايدروفورات البكتيرية كتعويض. وعلى العكس، قد تؤثر السايدروفورات الميكروبية أيضًا على أنظمة امتصاص الحديد في النباتات. على سبيل المثال، قد تقوم السايدروفورات الميكروبية بربط الحديد من معقدات السايدروفورات النباتية-Fe(III) أو Fe(III) المرتبط بمواد أخرى تخرجها الجذور، مما يجعل هذه الأنظمة أقل كفاءة. وبالتالي، لا يزال غير واضح ما إذا كان تحسين تغذية الحديد الملحوظ في الزراعة المختلطة يحدث لأن النباتات تستغل السايدروفورات البكتيرية بكفاءة أو ما إذا كان هناك تفاعل تكافلي بين الشريكين المتفاعلين.
تكشف دراستنا أن البيوفيردين يمكنه إذابة الحديد غير القابل للذوبان وجعله متاحًا حيويًا للنباتات (الأشكال 4f، 5، 6). وقد أفادت أعمال سابقة أيضًا بقدرة عدة أنواع من النباتات على الحصول على الحديد من البيوفيردين المحمّل بالحديد.. ومع ذلك، فإن الآلية البيوكيميائية الدقيقة لكيفية استرجاع النباتات للحديد من البايفيردين لا تزال غير واضحة. إحدى الخيارات هي أن النباتات تمتص المركب الكامل من Fe (III)-بايفيردين وتسترجع الحديد من خلال تحلل البايفيردين. البايفيردين يتكون العمود الفقري من سلسلة ببتيدية وبالتالي يمكن أن يتعرض للتحلل بواسطة البروتيازات. خيار آخر هو أن النباتات تحفز تقليل الحديد (III) إلى الحديد (II) من خلال مختزل الحديد الثلاثي (FCR) لتمتصه مباشرة.بينما تلعب FCR بالتأكيد دورًا، لاحظنا أن إضافة البايفيردين أدت إلى تقليل نشاط FCR في الفول السوداني وإنتاج DMA في الذرة، والتي تمثل الاستجابات العادية لنقص الحديد في هذين النوعين من النباتات. (الشكل التوضيحي 24). تشير هذه التغييرات إلى أن البايفيردين يصبح الطريق المفضل لاكتساب الحديد. تماشيًا مع هذه الفرضية، لاحظنا أن تقليص أنظمة امتصاص الحديد الخاصة بالنباتات يتزامن مع زيادة الكتلة الحيوية لكل من نباتات الفول السوداني والذرة (الشكل 5 والأشكال التوضيحية 22-24)، مما يشير إلى أن كلا النباتين يستفيدان ويوفران الموارد من خلال امتصاص الحديد المرتبط بالبايفيردين. هذه الصورة من دورة المغذيات التي تتوسطها تفاعلات النبات والميكروبيوم تتدمر في التربة المعقمة، مما يؤدي إلى انخفاض قيمة SPAD في الفول السوداني المزروع بالتداخل، حتى أقل من الفول السوداني المزروع بشكل أحادي (الأشكال 2d و5b). ومع ذلك، يمكن أن يزيد الزراعة المتداخلة من الحديد النشط في الأوراق الشابة للفول السوداني (الأشكال 2d و5b)، مما يشير إلى أن DMA الذي تفرزه الذرة يمكن استخدامه مباشرة من قبل الفول السوداني المزروع بالتداخل.في التربة المعقمة. لذلك، تشير نتائجنا إلى أن مصادر الحديد المتعددة تُستخدم خلال الزراعة المختلطة، بما في ذلك السايدروفورات من البكتيريا، وDMA من الذرة، وربما أيضًا الفلافينات أو الكومارينات من إفرازات جذور الفول السوداني، للتخفيف من نقص الحديد في النباتات. ومع ذلك، بسبب الفروق الكبيرة في قدرة ربط الحديد بين البيوفيردين وإفرازات الجذور.الاحتمال هو أن النباتات توازن استراتيجيات امتصاص الحديد بينها وبين الأنظمة المعتمدة على البيوفرودين في الزراعة المختلطة، ولكن هناك حاجة إلى مزيد من البحث لاختبار هذه الفرضية.
بينما ركزنا بشكل أساسي على كيفية تحسين تغذية الحديد في الفول السوداني عند زراعته مع الذرة، اكتشفنا أيضًا فوائد للذرة بطريقة تعتمد على الميكروبيوتا، مما يشير إلى أن زراعة المحاصيل المتداخلة تفيد كلا النوعين من النباتات. يمكن أن تشارك الذرة والفول السوداني في تفاعل ‘إعطاء-استقبال’ من خلال تقارب ميكروبيوماتها في الزراعة المتداخلة. تعتبر نباتات الفول السوداني من البقوليات وتستضيف بكتيريا مثبتة للنيتروجين في عقيدات جذورها.بحيث يمكن للفول السوداني تحسين اكتساب النيتروجين من الذرة. لقد وجدنا بالفعل أدلة تدعم هذه الفرضية. على وجه التحديد، اكتشفنا زيادة في عدد أعضاء جنس Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium وجنس Devosia من الذرة إلى الفول السوداني (الشكل 3ب). زيادة وفرة Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium المكونة للعُقَد وDevosia غير الريزوبية المسببة للعُقَد.يمكن أن تزيد البكتيريا من نشاط العقدة والتركيز على الفول السوداني، الذي يمكن أن يعزز بدوره اكتساب النيتروجين من قبل الذرة القريبة من خلال تقليل المنافسة على نيتروجين التربة والأسمدة بين النوعين النباتيين.علاوة على ذلك، وجدنا أن الأزوتوباكتر، وهو جنس بكتيري غير متعايش مفيد للنباتات ويقوم بتثبيت النيتروجين، قد تم تعزيزه بشكل متبادل من الفول السوداني إلى الذرة، مما قد يساهم أيضًا في تحسين تغذية النيتروجين في الذرة. تتماشى نتائجنا مع النتائج السابقة حول تحسين تغذية النيتروجين المعتمد على الميكروبيوتا.بشكل عام، تعتبر تركيبات البقوليات/الحشائش مرشحة واعدة للزراعة المختلطة، وبالتالي فهي موضوع بحث مكثف في علم التربة وتغذية النبات.
توافر الحديد الحيوي منخفض بشكل خاص في التربة القلوية الكلسية. هذه هي السبب الرئيسي الذي جعلنا نركز على الأعضاء البكتيرية من ميكروبيوم الجذور، لأن البكتيريا معروفة بإنتاج تنوع من السايدروفورات، حيث أن بعضها (مثل البايفيردين) له affinity عالية للحديد.. ومع ذلك، من المهم أن نلاحظ أن عوامل أخرى قد تلعب أيضًا دورًا في تحديد توفر الحديد وفوائد الزراعة المختلطة. على سبيل المثال، مغذيات التربة مثلو(الأشكال الرئيسية للنيتروجين في منطقة الجذور) يمكن أن تؤثر على الرقم الهيدروجيني وبالتالي توفر الحديد. التطبيق طويل الأمد لـيقلل من درجة حموضة منطقة الجذور، بينمايزيد من منطقة الجذور. ومع ذلك، في نظام الزراعة المختلطة لدينا، ظل الرقم الهيدروجيني مستقراً عبر مراحل المغذيات (الشكل التوضيحي التكميلي 6ب)، مما يوحي بأن دورة النيتروجين قد ليس لها تأثير مباشر على تحسين تغذية الحديد في نظامنا. قد يكون السبب في ذلك أن التربة الكلسية لديها قدرة قوية على تخزين الرقم الهيدروجيني.علاوة على ذلك، خصائص التربة، مثليمكن أن يؤثر الكربون العضوي، أو الكربون العضوي، على هيكل ووظيفة المجتمع الميكروبي، مما يعني أن الأنواع المفيدة مثل Pseudomonas قد لا تكون متساوية الوفرة في جميع التربة. أخيرًا، بينما ركزنا على البكتيريا، قد تكون أعضاء أخرى من الميكروبيوتا أيضًا متورطة في فوائد الزراعة المختلطة. من بين المرشحين الرئيسيين هي البكتيريا المثبتة للنيتروجين (كما وجدنا في دراستنا للذرة) والفطريات الميكورية الجذرية. سيكون فهم تأثيرات مثل هذه التفاعلات متعددة الممالك بين النباتات والبكتيريا والفطريات عبر تربة ذات خصائص مختلفة ومقاييس جغرافية للزراعة المختلطة هو التحدي في العمل المستقبلي.
تظهر نتائجنا أن الزراعة المختلطة في التربة الطبيعية كافية لتحسين تغذية الحديد في التربة الكلسية (الشكل التوضيحي 23). ومع ذلك، قد لا تكون الزراعة المختلطة ممكنة دائمًا. في تلك الحالات، توفر الرؤى من دراستنا علاجًا بسيطًا وقابلًا للتطبيق، يتكون من إضافة Pseudomonas sp. 1502IPR01 أو البايفيردين الخاص بها إلى منطقة الجذور. يتمتع علاجنا بعدة مزايا مقارنة باستخدام الأسمدة التقليدية للحديد مثل EDTA-Fe الاصطناعي الذي يتمتع بكفاءة منخفضة ومخاطر بيئية عالية في التربة.لا تنطبق هذه القضايا على علاج البيوفرادين لأنه يذيب الحديد مباشرة من المخزونات الطبيعية في التربة، والبيوفرادين نفسه قابل للتحلل البيولوجي.، و . extremorientalis (أقرب قريب لـ Pseudomonas sp. 1502IPR-01) مصنفة ككائن خطر حيوي من الفئة 1 غير ضار ومن غير المحتمل أن يسبب مرضًا في البشر. وبالتالي، يمكن اعتبار بكتيريا Pseudomonas sp. 1502IPR-01 والبايوفيردين الخاص بها كسماد حيوي صديق للبيئة. بالإضافة إلى ذلك، يمكن أن تعزز البايوفيردين أيضًا توافر النحاس الحيوي وبالتالي لديها القدرة على تحسين تغذية النباتات للعناصر الدقيقة الأخرى.. نظرًا لدوره المزدوج في تخصيب العناصر الدقيقة (خاصة الحديد) ومكافحة الأمراض النباتيةيمكن أن يقلل البايوفيردين في الوقت نفسه من استخدام الأسمدة الاصطناعية الدقيقة والمبيدات الحشرية، مما قد يمثل خطوة حاسمة نحو الزراعة المستدامة، وتحسين الأمن الغذائي، والحفاظ على البيئة.
باختصار، أوضحنا آلية كيف تستفيد النباتات من الزراعة المختلطة من خلال تقارب الميكروبيوم في منطقة الجذور. تبرز دراستنا أن فائدة الزراعة المختلطة من التيسير تحت الأرض تؤدي إلى وظيفة مكتسبة للفول السوداني من خلال إثراء متبادل لأعضاء الميكروبيوم في منطقة الجذور المستمدة من الذرة (الشكل 7). تقدم نتائجنا فرصًا تطبيقية لعلاج بروبيوتيك لتحسين صحة النبات وإنتاجية المحاصيل في الحالات التي لا تكون فيها الزراعة المختلطة ممكنة. بينما حددنا سلالة بروبيوتيك ومركبها المفيد الذي يتم إفرازه، هناك إمكانيات كبيرة لتحسين العملية من خلال الهندسة الحيوية. تقدم رؤانا الآلية حول تحسين تغذية الحديد وجهات نظر ويجب أن تثير اهتمامًا في تعزيز أنظمة الزراعة المختلطة الأخرى مع تركيبات نباتية أخرى لزيادة كل من الإنتاجية والاستدامة.
طرق
حساب NE و LER
لتحديد فوائد زراعة الفول السوداني/الذرة معًا، قمنا بحساب نسبة تكافؤ الأرض (LER) والأثر الصافي (NE) لهذا النظام الزراعي في مواقع حقول وسنوات مختلفة. تشير LER > 1 إلى أن الزراعة المختلطة لها ميزة على الزراعة الأحادية من حيث كفاءة استخدام الأرض. NE > 0 تشير إلى أن الزراعة المختلطة تحقق عائدًا مطلقًا أكبر من الزراعة الأحادية..
يتم تعريف NE على أنه الفرق بين العائد المرصود والعائد المتوقع.
أين و تمت ملاحظة غلات الأنواع 1 و 2 في الزراعة المختلطة، و و هي العوائد المتوقعة من النوعين، المحسوبة كحاصل ضرب عائد الزراعة الأحادية و حصة الأرض.
أين و هي العوائد لكل وحدة مساحة من الأنواع 1 و 2 في الزراعة الأحادية، و و هي حصة الأرض للنوعين 1 و 2 في الزراعة المختلطة. تم حساب حصة الأرض بناءً على كثافات الأنواع في الزراعة المختلطة وفي الزراعة الأحادية أو بناءً على عدد الصفوف أو ترتيبات النباتات..
يتم تعريف LER على أنه مجموع LER الجزئي (العوائد النسبية) لكل نوع (pLER1 و pLER2):
أين و هي العوائد (لكل وحدة من المساحة الإجمالية للزراعة المختلطة) للنوعين 1 و 2 في الزراعة المختلطة، و و هي عوائد الأنواع 1 و 2 في الزراعة الأحادية (نفس ما سبق).
تصميم تجريبي لزراعة الفول السوداني/الذرة في نظام الزراعة المختلطة تحت ظروف الأواني
قمنا بإجراء تجارب الزراعة المختلطة والزراعة الأحادية في الأواني. تم جمع تربة رملية جيرية نموذجية تعاني من نقص الحديد من عمق 0 إلى-عمق في قرية ليهوا، منطقة دا شينغ في بكين، الصينتم تحديد خصائص التربة قبل تطبيق الأسمدة وفقًا للبروتوكولات من الأدبيات السابقة.وتم إدراجها في الجدول التكميلي 6. احتوى كل وعاء على 8 كجم من التربة المخصبة بالأسمدة الأساسية [التركيب ( التربة): ن 100 (كـ )، ص 150 (كـ )، K 100 (كـ KCl )، Mg 50 (كـ ) وزنك 5 (كـ )].
تم استخدام الفول السوداني (Arachis hypogaea L. cv. Luhua14) والذرة (Zea mays L. cv. Zhengdan958). تم إعداد ثلاثة معالجات زراعية. تم زراعة ستة نباتات فول سوداني أو ثلاث نباتات ذرة في وعاء لمعالجات الزراعة الأحادية (زراعة أحادية للفول السوداني وزراعة أحادية للذرة). تم زراعة ثلاث نباتات فول سوداني مع ثلاث نباتات ذرة في وعاء لمعالجات الزراعة المختلطة. تم زراعة النباتات تحت ظروف الإضاءة الطبيعية معدرجة حرارة الهواءشدة الضوء والرطوبة النسبية. خلال التجربة، تم الحفاظ على محتوى الماء في التربة عند حواليسعة الحقل. تم حصاد عينات من نبات الفول السوداني لتقييم حالة التغذية في، و 73 يومًا بعد الزراعة (dps). في نفس الوقت، تم تدمير الأواني للحصول على تربة كاملة. تمت إزالة التربة السائبة عن طريق الاهتزاز برفق. تم فصل جذور الذرة المزروعة مع الفول السوداني المزروع بعناية عن بعضها البعض أثناء الاهتزاز. تم جمع تربة الجذور لتحليل الميكروبيوم وخصائص التربة عن طريق إزالة التربة الملتصقة بالجذور باستخدام فرشاة دقيقة معقمة (الشكل التوضيحي التكميلي 2). تم خلط نباتات الفول السوداني وتربة الجذور من نفس الأواني في تكرار واحد. كان لكل معالجة أربعة تكرارات في كل نقطة زمنية.
تصميم لتجربة تعقيم التربة في ظروف الأصص
لتقييم دور الميكروبيوم في منطقة الجذور في تحسين تغذية الحديد للفول السوداني في الزراعة المختلطة، تم زراعة الفول السوداني في زراعة أحادية وزراعة مختلطة مع الذرة في كل من التربة العادية والمعقمة. تم إعداد التربة كما هو موضح أعلاه. بالنسبة لعلاج تعقيم التربة، تم -مُشعّع بجرعة قصوى تبلغ 20 كيلوغراي (24 ساعة،شركة بكين للتكنولوجيا العالية في تطبيق تكنولوجيا الإشعاع جينهوي المحدودة). كانت أنواع النباتات وظروف النمو وإدارة المحاصيل كما في تصميم التجربة لزراعة الفول السوداني/الذرة في ظروف الأواني. في 73 يومًا بعد الزراعة، تم حصاد عينات من الفول السوداني لتقييم حالة التغذية بالحديد، وتم جمع التربة المحيطة بالجذور لقياس تركيز الحديد المتاح. الفول السوداني تم خلط النباتات والتربة المحيطة بالجذور من نفس الأصيص ومعاملتها كنسخة واحدة. كان لكل معالجة ثلاث أو أربع نسخ.
قياس المؤشرات المتعلقة بالحديد في النباتات والتربة
قيمة SPAD. تم قياس مستويات الكلوروفيل في الأوراق الشابة الطازجة (الأوراق العليا التي ظهرت بالكامل) من الفول السوداني باستخدام مقياس الكلوروفيل (كونيكا-مينولتا، أوساكا، اليابان). تم قياس كل ورقة في أربع تكرارات وتم حساب المتوسط مع تجنب الأوردة الرئيسية للورقة.
نشاط مختزل الحديد-الكلات (FCR) في نباتات الفول السوداني. وفقًا للأدبيات السابقةتم غسل نباتات الفول السوداني الطازجة ثم تم غمر الجذور في محلول مشبعحل لمدة 30 دقيقة. بعد الغسيل، تم نقل نباتات الفول السوداني إلى 100 مل من الكاشف الذي يتكون من – ثنائي بيريدين و 0.1 مليمول من الحديد (III)-EDTA، من أجل الحضانة، محمية من الضوء. امتصاصية المحلول عند 523 نانومترتم قياسه. تم حساب نشاط اختزال الحديد-الكلات باستخدام المعادلة (5) أدناه وتم استخدام الماء كعنصر تحكم:
أين هو امتصاصية المحلول تحت 523 نانومتر، و FW هو الوزن الطازج للجذر، و 8650 هو الامتصاصية المولية لـ – ثنائي بيريدين ( 100 هو حجم محلول حضانة النباتات.
قدرة إفراز حمض الديوكسي ميوجينك (DMA) من نباتات الذرة. DMA هو الفيتوسيدروفور الوحيد الذي تفرزه الذرة، لذا استخدمنا طريقة تحديد إفراز الفيتوسيدروفورلتحديد قدرة إفراز DMA. تم غسل نباتات الذرة الطازجة ثم تم غمر الجذور في محلول مشبعلمدة 30 دقيقة. بعد الغسل، تم تحويل نباتات الذرة إلى 200 مل من الماء من أجلالحضانة، محمية من الضوء. قبل الحضانة،تم إضافة ميكروبور (منتجات كاتادين، واليسيلين، سويسرا) لمنع التحلل الميكروبي للفيتوسيدروفورات. تم إضافة 10 مل من المحلول المحضن أعلاه إلى 0.5 مل منوتم خلطه لمدة 15 دقيقة، ثم تم إضافة 1 مل من محلول أسيتات الصوديوم (pH 7.0) وتم خلط المحلول لمدة 10 دقائق أخرى. لتقليل Fe (III) إلى Fe (II)، تم تصفية المحاليل إلى 0.25 مل من 6 M HCl ثمتم إضافة هيدروكسيلامين هيدروكلوريد. تم حضانة المحلول عندلمدة 30 دقيقة. ثم، يتم خلط المحلول معفيروزين و1 مل من محلول أسيتات الصوديوم 2.0 M (pH 4.7). بعدالحضانة، تم قياس امتصاص المحلول عند 562 نانومتر. تم حساب نشاط إفراز DMA باستخدام الصيغة (6) أدناه باستخدام الماء كتحكم:
حيثهو امتصاص المحلول تحت 562 نانومتر. FW هو الوزن الطازج للجذر.هو الامتصاص المولي (). 200 هو حجم محلول حضانة النبات. 1.35 هو نسبة حجم محلول التفاعل النهائي والأصلي.
الحديد النشط في الأوراق الشابة. تم غسل الأوراق الشابة الطازجة من الفول السوداني بالماء المقطر وجمعها لتقييم الحديد القابل للاستخراج بواسطة HCl (‘الحديد النشط’) وفقًا للأدبيات السابقةتم أخذ الأوراق الشابة وخلطها مع 10 مل من حمض الهيدروكلوريك 1 M. تم خلط السائل العلوي لمدة 5 ساعات عند 150 دورة في الدقيقة ثم تم تصفيته باستخدام ورق الترشيح. تم قياس تركيز الحديد بواسطة مطيافية الانبعاث الضوئي بالتحليل الطيفي المتصل بالتحليل الطيفي (ICP-OES) باستخدام نظام 7300DV (بيركن إلمر، وولثام، الولايات المتحدة الأمريكية). تم حساب الحديد النشط في الأوراق الشابة باستخدام الصيغة (7) أدناه وتم استخدام الماء منزوع الأيونات كـ تحكم:
حيثوCONCNهما تركيز الحديد () من ترشيح العينة والتحكم، على التوالي، وFW هو الوزن الطازج للأوراق الشابة. يتم تطبيق الضرب بـ 10 لحساب حجم حمض الهيدروكلوريك.
الحديد المتاح في منطقة الجذور
تم وزن 10 جرام من تربة سطح الجذور المجففة بالهواء وإضافتها إلى 20 مل من عامل استخراج حمض ثنائي إيثيلين ثلاثي الأمين (DTPA)، يحتوي على 0.005 M DTPA،و0.1 M ثلاثي إيثانول أمين. ثم، تم خلط المزيج لمدة ساعتين عند. عند. بعد ذلك، تم تصفية المحلول وتم قياس تركيز الحديد في الترشيح بواسطة ICP-OES. تم حساب الحديد المتاح في منطقة الجذور باستخدام الصيغة (8) أدناه وتم استخدام الماء منزوع الأيونات كتحكم:
الحديد المتاح في منطقة الجذورالوزن حيثوهما تركيز الحديد () من عينة التربة والتحكم، الوزن هو وزن عينة التربة. يتم تطبيق الضرب بـ 20 لحساب حجم عوامل استخراج DTPA.
استخراج الحمض النووي من التربة، تسلسل الأمبليكون 16S rRNA
تم إجراء تسلسل الأمبليكون 16S rRNA لفحص تأثير الزراعة المختلطة على تركيبات الميكروبيوم في منطقة الجذور. كما هو الحال مع أي دراسة تسلسل ميكروبيوم، من المهم مراعاة مصادر التلوث المحتملة. يمكننا تحديد مصدرين محتملين في مخطط جمع عينات الميكروبيوم في منطقة الجذور لدينا، وهما (أ) عدم فصل الجذور بشكل كامل و(ب) خلط التربة أثناء حصاد النباتات. بالنسبة لـ (أ)، قد يكون من الممكن أن جذور الفول السوداني والذرة لا يمكن تمييزها وفصلها بشكل موثوق عند جمعها. يمكننا دحض هذه الإمكانية لأن جذور الفول السوداني والذرة يمكن تمييزها بوضوح بناءً على لونها: جذور الذرة بيضاء، بينما جذور الفول السوداني بنية داكنة (الشكل التكميلي 25). هذه الاختلافات الواضحة سمحت لنا بفصل جذور النوعين بشكل كامل وموثوق. بالنسبة لـ (ب)، هناك احتمال أن جزيئات التربة السائبة المرتبطة أصلاً بجذر أحد الأنواع قد تنتقل إلى التربة المجمعة من النوع الآخر أثناء عملية الحصاد. لمنع هذا المصدر من التلوث، قمنا أولاً بهز جذور النبات المجمعة بقوة للتخلص من جميع جزيئات التربة السائبة التي ترتبط عادة بالتربة الرملية. بعد ذلك، جمعنا فقط تربة منطقة الجذور التي كانت مرتبطة بإحكام حول جذور الفول السوداني أو الذرة. هذان النقطتان تضمنان تقليل خطر التلوث إلى الحد الأدنى وأن التقارب الذي لاحظناه ينشأ من التفاعلات البيولوجية بين الجذور الدقيقة للنوعين النباتيين. تم عزل الحمض النووي من 500 ملغ من عينة التربة باستخدام مجموعة FastDNASpin Kit for Soil (MP Biomedicals، سولون، الولايات المتحدة الأمريكية) وفقًا لتعليمات الشركة المصنعة. تم تضخيم المنطقة المتغيرة V3-V4 من جين 16S rRNA البكتيري باستخدام البرايمر 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) و806R (5′-GGAC-TACHVGGGTWTCTAAT-3′) بواسطة جهاز ABI GeneAmpPCR thermocycler (Applied Biosystems، فوستر سيتي، الولايات المتحدة الأمريكية). كانت ظروف PCR كما يلي: 3 دقائق عندثم 27 دورة من كل 30 ثانية عندعندو30 ثانية عند. بعد هذه الدورات، كان هناك وقت إطالة لمدة 10 دقائق عند. تم تجميع الأمبليكون المنقى بشكل متساوي وتم تسلسله على منصة Illumina MiSeq PE300 (Illumina، سان دييغو، الولايات المتحدة الأمريكية) وفقًا للبروتوكولات القياسية من قبل شركة Majorbio Bio-Pharm Technology Co. Ltd. (شنغهاي، الصين).
تم معالجة تقدم التحليل العلوي لتسلسل الأمبليكون باستخدام QIIME2 v2021.8. باستخدام سير العمل DADA2 في
QIIME2، تم تقليم الرموز الشريطية، وبرايمرات PCR، والتسلسلات ذات الجودة المنخفضة، وتم بعد ذلك إزالة الضوضاء ودمج القراءات المتبقية لتوليد جدول ميزات متغيرات تسلسل الأمبليكون (ASV). تم استخدام مصنف بايزي الأصلي المدرب على المنطقة V3-V4 (338 F و806R) من تسلسل 16 S المرجعي (إصدار SILVA 138) لتعيين الهويات التصنيفية للتسلسلات التمثيلية. تم الاحتفاظ فقط بـ ASV التي تم تعيينها على الأقل إلى مستوى الشعبة. تم إزالة الميزات التي تم تحديدها على أنها كلوروبلاست أو ميتوكوندريا. تم تصفية الميزات التي كانت لديها وفرة إجمالية (مجمعة عبر جميع العينات) أقل من عشرة أو موجودة في أقل من ست عينات (عينات) تم تصفيتها. قمنا بتطبيع عمق التسلسل إلى 17,063 عدًا لكل عينة باستخدام التدرج مع أخذ عينات مرتبة (SRS)مع الحفاظ على الهيكل المجتمعي الأصلي من خلال تقليل أخطاء أخذ العينات. تم تصفية التسلسلات التمثيلية بناءً على جدول ASV المصفى، ثم استخدمت لتوليد شجرة تطورية تقريبية باستخدام FastTree 2.1.8 في QIIME2 مع المعلمات الافتراضية.
تم إجراء تحليل PCoA وحساب مسافة Bray-Curtis بواسطة حزمة R ‘phyloseq’ (الإصدار 1.34.0) بناءً على الوفرة النسبية لـ ASV. تم تحديد الميكروبات المختلفة بين المجموعات بواسطة تحليل التمييز الخطي (LDA) وحجم تأثير LDA (LEfSe) مع عتبة LDA تبلغ 3 وعتبة القيمة 0.05. تم إجراء تحليل LefSe على خادم مختبر هوتنهاور غالاكسي (http://huttenhower.sph.harvard.edu/galaxy). تم إجراء تحويل التنسيق من جدول ASV إلى جدول لـ Lefse بواسطة حزمة R ‘amplicon’ (الإصدار 1.14.2). كانت العلاقة بين وفرة العلامات البيولوجية لجنس الجذور النشطة واحتياجات الحديد للنباتات في التربة أيضًا تعتمد على الوفرة النسبية وتم حسابها باستخدام ارتباط رتبي غير معلمي (مقدم كـ Spearman rho) باستخدام حزمة R ‘phyloseq’ (الإصدار 2.2.5).
عزل الجذور البكتيرية المفرزة للفيتوسيدروفور
قمنا بعزل الجذور البكتيرية المفرزة للفيتوسيدروفور من منطقة الجذور للفول السوداني المزروع بشكل مختلط لتقييم الدور الذي تلعبه في تحسين تغذية الحديد للفول السوداني. تم تعديل طريقة العزل وفقًا للبروتوكولات من الأدبيات السابقة(الشكل 4أ) والتفاصيل موصوفة أدناه. باختصار، تم تحريك 1 جرام من عينة التربة من منطقة جذور الفول السوداني المزروع بشكل مختلط بشكل معتدل في 20 مل من محلول معقملمدة 10 دقائق أولاً. ثم تم نشر التعليق الميكروبي (مخفف بالتسلسل) على أطباق الأجار معمؤشر الكروم أزول S (CAS). احتوى مؤشر CAS على 1 مللي مول من CAS،بروميد الهكساديسيل تريميثيل الأمونيوم (HDTMA). تتكون المكونات الأخرى لوسيط أجار CAS من (): سكر، 2000؛ هيدروكسيليت حمض الكازين، , 241؛ أجار، 1800 و محلول عازل فوسفات ). تم ضبط الرقم الهيدروجيني للوسيط على 7.0. يشكل المركب الثلاثي CAS-Fe(III)-HTDMA صبغة زرقاء. تقوم السايدروفورات بإزالة Fe (III) من الصبغة عن طريق خلب Fe (III)، مما يؤدي إلى تغيير اللون من الأزرق إلى البرتقالي (في طبق الأجار) أو الأحمر (في المحلول) . تم حضانة الأطباق عند لمدة 48 ساعة في الظلام. تم اختيار المستعمرات الميكروبية مع أو بدون هالة برتقالية. ثم تم إعادة زراعة العزلات على أطباق LB لتنقية المستعمرات. كانت المجموعة النهائية تتكون من 324 عزلة من البكتيريا الجذرية التي تفرز السايدروفورات. تم زراعة جميع العزلات المنقاة في 1 مل من وسط LB السائل عند مع التحريك (خلاط دوار بسرعة 180 دورة في الدقيقة) قبل التجميد والتخزين عند في الجليسرول. يجب توجيه الطلبات للحصول على السلالات إلى zuoym@cau.edu.cn.
قياس قدرة إفراز السايدروفورات من البكتيريا الجذرية
لتحديد قدرة إفراز السايدروفورات لكل عزلة من البكتيريا الجذرية تحت ظروف نقص الحديد، تم إحياء عزلات البكتيريا الجذرية عن طريق نقل أحجام من مخزونها المجمد إلى 5 مل من وسط LB السائل وزراعتها لمدة 18 ساعة عند مع التحريك (خلاط دوار بسرعة 180 دورة في الدقيقة). ثم، تم نقل أحجام من الثقافات المعاد إحيائها إلى 5 مل من وسط حمض السكسينيك القياسي (SSM) بدون
حديد مضاف ( حمض السكسينيك، ). بعد الحضانة عند ، تم تخفيف الثقافات البكتيرية إلى . ثم، تم جمع السائل الطافي الخالي من الخلايا من الثقافة البكتيرية المخففة عن طريق الطرد المركزي ( عند ) والترشيح (باستخدام فلتر). تم قياس تركيز السايدروفورات في السائل الطافي باستخدام نسخة معدلة من اختبار CAS الكيميائي العالمي. باختصار، تم إعداد محلول اختبار CAS كما يلي: تم إذابة 21.9 ملغ من HDTMA في 25 مل من الماء مع التحريك المستمر على حرارة منخفضة. تم خلط 1.5 مل من (في 10 مليمول HCl) مع 7.5 مل من 2 مليمول CAS. تمت إضافة هذا المحلول ببطء إلى محلول HDTMA أثناء التحريك، وتم تكوين حجم الخليط إلى 100 مل باستخدام ماء منزوع الأيونات. تم خلط السائل الطافي الخالي من الخلايا أو ماء منزوع الأيونات كمرجع تحكم مع حجم متساوي من محلول اختبار CAS في طبق 96 بئر وتم حضانته لمدة أربعة عند درجة حرارة الغرفة (الشكل التكميلي 8). أزالت السايدروفورات Fe(III) من مركب Fe (III)-CAS-HDTMA الملون، مما أدى إلى تغيير اللون من الأزرق إلى الأحمر في محلول وسط CAS، مما خفض القياسات. ثم تم قياس للسوائل الطافية الخالية من الخلايا (A) ومراقبة ماء منزوع الأيونات (Ar) باستخدام قارئ الأطباق (SpectraMax M5، Molecular Devices، Sunnyvale، USA). يمكن تحديد قدرة إفراز السايدروفورات باستخدام الصيغة التالية: . تصف هذه الصيغة الانخفاض النسبي في قياس الناتج عن المركبات الخالبة للحديد التي تسترجع Fe (III) من مركب Fe (III)-CAS-HDTMA الملون. يظهر قيمة Su تغييرات أسية بدلاً من تغييرات خطية عندما تكون السايدروفورات مفرطة (). وبالتالي، يجب تخفيف السائل الطافي لمحلول تفاعل اختبار CAS بشكل تدريجي بواسطة 1/2 من محلول اختبار CAS لضمان . تم استخدام ديسفيريوكسامين B (DFOB)، وهو سايدروفور قابل للشراء، لإنشاء منحنى معايرة لتقدير تركيز السايدروفورات في السوائل الطافية للبكتيريا الجذرية. تم خلط تخفيفات متسلسلة (، و و) من معيار DFOB مع محلول اختبار CAS وتم حضانتها لمدة أربعة عند درجة حرارة الغرفة قبل التحديد. كانت قيمة Su لـ DFOB هي 0.71، وكانت معادلة منحنى المعايرة هي: مكافئ السايدروفور (). يتم التعبير عن النتائج كمكافئات
من DFOB. لحساب قدرة ربط الحديد للسايدروفورات البكتيرية، يعتبر 1 مليمول من DFOB مرتبطًا بمليمول واحد من Fe(III).
بناء الشجرة والتحليل الجيني لعزلات البكتيريا الجذرية التي تفرز السايدروفوراتقمنا بتسلسل جينات 16S rRNA لعزلات البكتيريا الجذرية التي تفرز السايدروفورات لتأكيد مجموعة جنسها وعلاقاتها التطورية. وفقًا لبروتوكول الشركة المصنعة، تم استخراج الحمض النووي الجينومي باستخدام مجموعة تنقية الحمض النووي الجينومي Wizard (Promega، Charbonnières، فرنسا). تم قياس الحمض النووي الجينومي المنقى باستخدام مقياس الفلورية TBS-380 (Turner BioSystems Inc.، Sunnyvale، USA). تم استخدام الحمض النووي عالي الجودة () لإجراء المزيد من الأبحاث. تم تضخيم جين 16S rRNA باستخدام بادئات 27 F (-AGAGTTTGATCMTGGCTCAG-3′) و1492 R (-TACGGY-TACCTTGTTACGACTT-3′) بواسطة تفاعل البوليميراز المتسلسل (PCR). احتوت تفاعلات PCR () على واحد من الحمض النووي البكتيري، و مزيج رئيسي، وواحد من كل من البادئات الأمامية والعكسية، و ماء منزوع الأيونات. تم تشغيل PCR كما يلي: البداية: إنكار عند لمدة 5 دقائق، 30 دورة من الإنكار عند لمدة 30 ثانية، التهجين عند لمدة 30 ثانية، التمديد عند لمدة دقيقة و30 ثانية، والتمديد النهائي عند
استخراج الحمض النووي الجينومي الكلي وتسلسل عزلة البكتيريا الجذرية
تم تسلسل جينوم عزلة Pseudomonas 1502IPR-01، التي كانت لديها أعلى قدرة على إفراز السايدروفورات في جميع عزلات البكتيريا الجذرية، باستخدام مزيج من منصات تسلسل PacBio RS II Single Molecule Real Time (SMRT) وIllumina. تم استخدام بيانات Illumina لتقييم تعقيد الجينوم.
للتسلسل باستخدام Illumina، تم استخدام على الأقل من الحمض النووي الجينومي في بناء مكتبة التسلسل. تم تقطيع عينات الحمض النووي إلى قطع بطول 400-500 نقطة باستخدام جهاز M220 Focused Acoustic Shearer (Covaris، Woburn، الولايات المتحدة الأمريكية) وفقًا لبروتوكول الشركة المصنعة. تم إعداد مكتبات تسلسل Illumina من القطع المقطعة باستخدام مجموعة NEXTflex Rapid DNA-Seq Kit (Bioo Scientific، أوستن، الولايات المتحدة الأمريكية). باختصار، تم إصلاح الأطراف الرئيسية 5′ أولاً وإضافة الفوسفات. بعد ذلك، تم إضافة ذيول A إلى الأطراف 3′ وربطها بمهايئات التسلسل. كانت الخطوة الثالثة هي إثراء المنتجات المرتبطة بالمهايئات باستخدام PCR. ثم تم استخدام المكتبات المعدة للتسلسل Illumina مزدوج النهاية () على آلة Illumina HiSeq X Ten (Illumina، سان دييغو، الولايات المتحدة الأمريكية).
للتسلسل باستخدام Pacific Biosciences، تم تدوير جزء من الحمض النووي في أنبوب Covaris g-TUBE (Covaris، Woburn، الولايات المتحدة الأمريكية) عند لمدة 60 ثانية باستخدام جهاز الطرد المركزي Eppendorf 5424 (Eppendorf، NY، الولايات المتحدة الأمريكية). ثم تم تنقية قطع الحمض النووي، وإصلاح الأطراف، وربطها بمهايئات تسلسل SMRTbell وفقًا لتوصيات الشركة المصنعة (Pacific Biosciences، Menlo Park، الولايات المتحدة الأمريكية). وفقًا لتوصيات الشركة المصنعة، تم تنقية مكتبة التسلسل الناتجة ثلاث مرات باستخدام أحجام من كرات Agencourt AMPure XP (Beckman Coulter Genomics، Danvers، الولايات المتحدة الأمريكية). بعد ذلك، تم إعداد مكتبة إدراج بحجم 10 كيلو بايت وتم تسلسلها على خلية SMRT واحدة باستخدام الطرق القياسية.
تجميع الجينوم
تم تجميع تسلسل الجينوم الكامل لـ Pseudomonas sp. 1502IPR-01 باستخدام كل من قراءات PacBio وIllumina. تم تحويل بيانات الصورة الأصلية إلى بيانات تسلسل عبر استدعاء القاعدة، والتي تعرف بالبيانات الخام، وتم حفظها كملف FASTQ. تحتوي تلك الملفات FASTQ على تسلسلات القراءة، ومعلومات الجودة مضمنة. تم تطبيق إحصائية لمعلومات الجودة لتقليم الجودة، حيث يمكن إزالة البيانات ذات الجودة المنخفضة لتشكيل بيانات نظيفة. ثم تم تجميع القراءات في contig باستخدام عملية تجميع الجينوم الهرمية (HGAP) وcanu، وهو برنامج يقوم بأداء تجميع دقيق وقابل للتوسع للقراءات الطويلة عبر وزن k -mer التكيفي وفصل التكرارات. تم التحقق من الخطوة الدائرية الأخيرة وإكمالها يدويًا، مما أدى إلى إنشاء جينوم كامل مع كروموسومات وبلازميدات سلسة. أخيرًا، تم إجراء تصحيح الأخطاء لنتائج تجميع PacBio باستخدام قراءات Illumina باستخدام Pilon. تم إجراء جميع التحليلات المذكورة أعلاه باستخدام منصة I-Sanger Cloud (www.i-sanger.com) من Shanghai Majorbio.
بناء شجرة النشوء والتطور لـ 1502IPR-01 وممثلي جنس الزائفة
لتأكيد العلاقات التطورية الدقيقة لـ 1502IPR-01 وممثلي أنواع Pseudomonas، تم إنشاء شجرة تطورية تعتمد على تحليل تسلسل متعدد المواقع باستخدام طريقة تم تأسيسها سابقًا.تم استخدام TBLASTN+ (الإصدار 2.10.0) بشكل محدد لتحديد واستخراج تسلسلات 100 جين من جينات المنزل المستخدمة في ديفيس وآخرون.تم محاذاة جميع الجينات المنزلية الـ 100 لـ 1502IPR-01 و23 نوعًا من بكتيريا Pseudomonas التي تم تسلسلها بالكامل باستخدام MAFFT (الإصدار 7.427). تم بناء شجرة النشوء والتطور باستخدام IQ-TREE2. تم تصور الأشجار باستخدام ggtree الإصدار 2.4.1 في R الإصدار 4.2.2. تم إدراج معلومات سلالات Pseudomonas spp. التي تم تسلسلها في الجدول التكميلية 7.
تحليل مجموعة جينات تخليق ونقل البيوفرادين
تم تحديد موضع جينات تخليق ونقل البيوفرادين في جينوم 1502IPR-01 باستخدام TBLASTN+ (الإصدار 2.10.0) مع تسلسلات البروتين من بكتيريا الزائفة الهوائية PAO1 المستخدمة كاستعلامات ضد تسلسلات البروتين المتوقعة من جينوم 1502IPR-01 كقواعد بيانات. تم إجراء مقارنات التسلسل باستخدام خوارزمية سميث-واترمان بواسطة برنامج SnapGene (الإصدار 6.0.2، GSL Biotech LLC، بوسطن، الولايات المتحدة الأمريكية) للحصول على هوية تسلسل الحمض النووي. تم إجراء التصوير باستخدام Adobe Illustrator CC، 2020 (أدوبي، سان خوسيه، الولايات المتحدة الأمريكية).
إنتاج وتنقية البيوفر دين
لإنتاج البيوفرادين، تم زراعة Pseudomonas sp. 1502IPR-01 في وسط SSM المذكور أعلاه. بعد الزراعة فيعند 180 دورة في الدقيقة لمدة 48 ساعة، تم عزل وسط الثقافة (10 لترات) من المواد الخلوية عن طريق الطرد المركزي فيلمدة 15 دقيقة ثم الترشيح باستخدام ورق الترشيح. تم تعديل مستخلصات الثقافة الخام إلى pH 5.5 عن طريق إضافة HCl وتم امتصاصها بواسطة XAD-4، وهو راتنج ماص بوليمري. بعد إزالة السائل العلوي، تم ترشيح الراتنج، ثم تم غسل الفraction المحتوي على البيوفرادين. 1:1 ( ” ). الـتم تبخير المحلول حتى الجفاف في فراغ. ثم تم إذابة الرواسب فيوخضعت لجهاز الكروماتوغرافيا السائلة عالية الأداء شبه التحضيرية من شيمادزو (شيمادزو، طوكيو، اليابان) فيبمعدل تدفق قدرهحجم الحقن كانتمت عملية الكروماتوغرافيا للعينات باستخدام نظام مذيبات منحمض ثلاثي فلورو الأسيتيك (TFA) في (المذيب A) و MeCN-MeOH 4:1 (، المذيب ب). تدرج ثنائي خطي, تم استخدامه. تم مراقبة الإيلاوات عند 380 نانومتر. تم جمع الفصيل الذي يحتوي على البيوفرادين عندثم تم تعديلها إلى pH 7.0 باستخدامهيدروكسيد الأمونيوم قبل التبخر. تم إزالة الملح في الكمية الناتجة خلال تحييد TFA بواسطة غشاء غسيل كلوي Spectra/Por بحد قطع الوزن الجزيئي (MWCO): 500-1000 دالتون (مختبر سبيكتروم، رانشو دومينغيز، الولايات المتحدة الأمريكية). ثم، نقاءتم تأكيد وجود البايفيردين المعزول بواسطة HPLC.
التعرف الهيكلي على البيوفيردين
مطيافية الكتلة. تم الحصول على طيف LC-HRMS على مطياف الكتلة Agilent 6530 Q-TOF المتصل بجهاز HPLC Agilent 1260 (Agilent Technologies GmbH، والدبرون، ألمانيا)؛ مطياف MS Thermo Orbitrap (Orbitrap Fusion Lumos، Thermo Fisher Scientific، سان خوسيه، الولايات المتحدة الأمريكية)؛ ESI-QQQ-MSMS (Xevo TQ-S micro، Waters، ميلفورد، الولايات المتحدة الأمريكية).
تم الحصول على بيانات الرنين المغناطيسي النووي باستخدام (يعمل بتردد 500.13 ميغاهرتز لـو125.75 ميغاهرتز لـ (بروكر بيوسبين GmbH، كارلسروه، ألمانيا). تم قياس جميع الأطياف عند 303 كلفن. تم استخدام إشارات المذيب المتبقية كمرجع للأطياف في و الأبعاد.
UV/Vis. تم تسجيل طيف الأشعة فوق البنفسجية بواسطة كاشف مصفوفة ثنائية الأبعاد SPD-M20A من شيمادزو (PDA، الكشف 190-800 نانومتر) (شيمادزو، كيوتو، اليابان) متصل بنظام HPLC من شيمادزو (LC-20AR، شيمادزو، اليابان) خلال عمليات الفصل شبه التحضيري HPLC في MeCN- يحتوي على 0.1% TFA في .
الكروماتوغرافيا السائلة عالية الأداء شبه التحضيرية. تم إجراء فصائل الكروماتوغرافيا السائلة عالية الأداء شبه التحضيرية على نظام كروماتوغرافيا سائلة عالية الأداء من شيمادزو (LC-20AR، شيمادزو، اليابان) مع عمود أتلانتس Prep OBD T3.واترز، الولايات المتحدة الأمريكية).
المواد الكيميائية. تم الحصول على الماء النقي للغاية من شركة واتسونز المحدودة (هونغ كونغ، الصين). تم شراء الأسيتونيتريل (درجة HPLC) وTFA (درجة HPLC) المستخدمة في UHPLC/Q-TOF-MS من شركة ميرك (دارمشتات، ألمانيا) وشركة رو ساينتيفيك إنك (نيوارك، الولايات المتحدة الأمريكية) على التوالي. تم شراء كربونات الصوديوم (درجة HPLC) من شركة سيغما-ألدريتش (سانت لويس، الولايات المتحدة الأمريكية). تم شراء غشاء الدياليز من شركة سبيكتروم لابوراتوريز إنك (رانشو دومينغيز، الولايات المتحدة الأمريكية).
بايوفيردين (الشكل التوضيحي 11أ)، الذي تم الحصول عليه على شكل مسحوق بلوري أصفر مائل إلى الأخضر، أظهر طيف الأشعة فوق البنفسجية/المرئية النموذجي للبايوفيردين: 387 نانومتر عند pH 7.0 وشريط مقسم عند 365 و407 نانومتر عند pH 3.0 و10.0. الصيغة الجزيئية لـتم تحديده بواسطة قمة الأيون الجزيئي الزائف عند(محسوب لـ1393.5872) في تحليل الكتلة الإيجابية HR-ESI-MS (الشكل التوضيحي 11b). أشار التحلل الكلي للبايوفيردين إلى سلسلة ببتيد تحتوي على ثلاثة أنواع من الأحماض الأمينية: السيرين (Ser)، والليسين (Lys) و5-N-formyl-5-N-hydroxyornithine (FoOHOrn) (الشكل التوضيحي 12).
لقد تم توثيق أن مطيافية الكتلة هي طريقة قوية لتوضيح البنية للبايوفيردينات.في هذا السياق، حصلنا على الأيونات المميزة التالية لبايوفيردين باستخدام طريقة تفكك الاصطدام عالي الطاقة (HCD) مع طاقات اصطدام متغيرة. يوجد أيون عند204.08، الأيون الأكثر تميزًا للكروموفور بايوفيردين (A1′-2، الشكل التكميلي 11c)، تم التعرف عليه بوضوح في جهاز قياس الكتلة Orbitrap (الشكل التكميلي 11d). بالقرب منه، كان هناك أيون وفير آخر عند، والذي يتوافق مع الكروموفور المتصل بسلسلة جانبية من الأميد الكربوكسيلي (A1′-1) (الشكل التكميلي 11c، d)، يمكن تفسيره كأيون شظية لفتح حلقة التتراهيدروبيريميدين للكروموفور بواسطة تفاعل ريترو-دييلز-ألدر (RDA). ثم، تم إثبات وجود حمض الكيتوجلوتاريك (kgl) لسلسلة البايفيردين الجانبية من خلال فقدان من A1 ( ) و B1 ( أيونات تعطي الاثنين المقابلين و الأيونات (الشكل التوضيحي التكميلي 11e)“. ومن ثم، تم اقتراح مسارات هيكل-التفكك من B1 إلى A1′-2 كما هو موضح في الشكل التوضيحي 11c. علاوة على ذلك، تمثل سلسلة الأيونات B من B8 ( ) إلى B1 ( ) مسموح بتحديد تسلسل الأحماض الأمينية لسلسلة الببتيد كالتالي: سير-سير-FoOHOrn-سير-سير-(لايس-FoOHOrn-لايس-سير) (الأشكال التكميلية 12-14). سلسلة أيونات B’ (فقدان من سلسلة kgl لأيونات B، الأشكال التكميلية 12-14) تشكل B8′ ( ) إلى أكدت تسلسل الببتيد. معًا، تم استنتاج التركيب الأساسي للبايوفيردين على أنه kgl-chr-Ser-Ser-FoOHOrn-Ser-Ser-(Lys-FoOHOrn-Lys-Ser)، وهو مطابق للبايوفيردين من Pseudomonas spp. CFML 95-275..
تم تأكيد هيكل البيوفيردين المستنتج من بيانات الطيف الكتلي (MS) بشكل إضافي من خلال بيانات الرنين المغناطيسي النووي (NMR). استنادًا إلى مجموعة من و تجارب الرنين المغناطيسي النووي الكربوني (C-NMR) ثنائي الأبعاد (COSY، NOESY، HSQC، وHMBC) (الأشكال التكميلية 15-20)، – و -تم تخصيص بيانات البيوفرادين في الجداول التكميلية 4 و 5. بالإضافة إلى ذلك، يتم عرض الارتباطات الرئيسية COSY و HMBC في الشكل التكميلية 21. أظهرت النتائج أن-كانت بيانات NMR للبايوفيردين شبه مطابقة لتلك الخاصة بالبايوفيردين المبلغ عنها. بينما -قيم الإزاحة الكيميائية لنمط الرنين المغناطيسي النووي للبايوفيردين قد تحولت، والتي قد تكون ناتجة عن درجات حرارة أو آلات اكتساب مختلفة. تتكون جزيئة البيوفرادين من ثلاثة أجزاء: (i) كروموفور من نوع ثنائي هيدروكينولين (chr): (1S)-5-أمينو-2،3-ثنائي هيدرو-8،9-ثنائي الهيدروكسي-1H-بيريميدو-[1،2-a]كينولون-1-حمض الكربوكسيليك، (ii) ببتيد يتكون من 9 أحماض أمينية: سير-سير-فوهورن-سير-سير-(ليس-فوهورن-ليس-سير) (سير: سيرين؛ فوهورن: 5-N-فورميل-5-N-هيدروكسي أورنيثين؛ ليس: ليسين)، و (iii) سلسلة جانبية (حمض الكيتوجلوتاريك، كجل ) مرتبط بذرة النيتروجين في الموضعمن الكروموفور.
قدرة البيوفرادين من بكتيريا Pseudomonas sp. 1502IPR-01 على إذابة الحديد من
هو الشكل الرئيسي غير القابل للذوبان من الحديد في التربة الكلسية. لذلك، قمنا بتحديد قدرة البايفيردين من Pseudomonas sp. 1502IPR-01 على إذابة الحديد من الشكل غير القابل للذوبان.الأسهم وفقًا للورقة السابقةلهذا الغرض،تم إضافة ما يعادل البايفيردين المنقى (المُحتسب بناءً على منحنى المعايرة DFOB) إلى 2 مل من محلول أسيتات الصوديوم 0.5 م. ثم، تم إضافة جميع المحاليل إلىتم تحضير المحلول عن طريق المعادلةمع NaOH إلىتعليقًا بحجم إجمالي قدره 5 مل. كانت المحاليل المذكورة أعلاه مُحضَّن فيلمدة خمس ساعات، وتم جمع السائل العلوي بعد الطرد المركزي فيفيلمدة 10 دقائق. أخيرًا، تم تحديد محتوى الحديد في السائل العلوي باستخدام ICP-OES بواسطة نظام 7300DV (بيركن إلمر، وولثام، الولايات المتحدة الأمريكية).
التحقق الوظيفي من بكتيريا Pseudomonas sp. 1502IPR-01 والحديدية الخاصة بها
لقد حددنا تأثير بكتيريا Pseudomonas sp. 1502IPR-01 والحديد الذي تنتجه على تحسين تغذية الحديد في الفول السوداني في كل من ظروف الدفيئة والحقول. تم إعداد التربة الرملية الكلسية العادية والمعقمة التي تعاني من نقص الحديد كما هو موضح أعلاه لتجربة الأصص. تم استخدام الفول السوداني (Arachis hypogaea L. cv. Luhua14) والذرة (Zea mays L. cv. Zhengdan958). تم إعداد أربع مجموعات علاجية: (1) زراعة الفول السوداني في تربة عادية؛ (2) زراعة الفول السوداني في تربة معقمة؛ (3) زراعة الفول السوداني والذرة معًا في تربة عادية؛ (4) زراعة الفول السوداني والذرة معًا في تربة معقمة. تم زراعة ستة نباتات فول سوداني في أصيص لعلاج زراعة الفول السوداني، وتم زراعة ثلاثة نباتات فول سوداني مع ثلاثة نباتات ذرة في أصيص لعلاج الزراعة المختلطة. احتوى كل أصيص على 8 كجم من التربة. ظلت بقية ظروف زراعة الدفيئة كما هو موضح أعلاه. عند الإزهار (50 يومًا بعد الزراعة)، وعند مرحلة تكوين القرون (64 يومًا بعد الزراعة)، وعند مرحلة انتفاخ الثمار (75 يومًا بعد الزراعة)، تم إضافة 600 مل منCFUمحلول سلالة 1502IPR-01نبات CFU ) أو 600 مل من بيوفر الدين المنقىنباتتم ري المحلول في منطقة الجذور كعلاج 1502IPR-01 وعلاج بايوفيردين. تم ري 600 مل من الماء في منطقة الجذور كتحكم. من الجدير بالذكر أننا طبقنامحلول بايوفيردين لعلاج منطقة الجذور للفول السوداني وفقًا للأدبيات السابقةوتجربتنا البحثية الخاصةتم خلط نباتات الفول السوداني في نفس الأصيص كنسخة مكررة. كان لكل معالجة ثلاث أو أربع نسخ مكررة. عند 84 يومًا بعد الزراعة، تم قياس قيم SPAD وتركيز الحديد النشط في الأوراق الشابة. تم جمع تربة منطقة الجذور لقياس تركيز الحديد المتاح.
تم إجراء التجربة الميدانية في قرية شيشانغإي،ن)، منطقة فانغشان في بكين، وقرية ليغو (، ن) في مدينة بويانغ، مقاطعة خنان، الصين في عام 2020. في بكين، كان صنف الفول السوداني هو لوهوا 14. كانت درجة الحرارة المتوسطة من مايو إلى سبتمبر هي، وكان معدل هطول الأمطار السنوي 603 مم. نوع التربة في بكين هو الكامبوسول الكلسي. كانت خاصية التربة pH 7.9، إجمالي النيتروجينمتاح P (أولسن-P)متاح ك ( ) متوفر في. في هنان، كان صنف الفول السوداني هو يوهوا 37. كانت درجة الحرارة المتوسطة من مايو إلى سبتمبر هي ، وكان معدل هطول الأمطار السنوي 604.1 ملم. نوع التربة في خنان هو التربة الفلوي-مائية. كانت خاصية التربة pH 7.7، إجماليمتاح P (أولسن- P)متاح ك ( ) متوفر فيتم تطبيق سماد مركب (N-P-K: 19-19-19) قبل الزراعة، وكانت إدارة الحقل وفقًا للممارسات المحلية. تم إعداد أربعة علاجات: (1) تم ري 150 مل من الماء في منطقة الجذور، ورش 16 مل من الماء على الأوراق الصغيرة لنبتة الفول السوداني كعلاج تحكم؛ (2) تم ري محلول سلالة 1502IPR-01 في منطقة الجذور، وتم رش 16 مل من الماء على الأوراق الصغيرة لنبات الفول السوداني كعلاج 1502IPR-01؛ (3)بيوفر الدين المنقىنباتتم ري الجذور بمقدار 16 مل من الماء على الأوراق الصغيرة لنبات الفول السوداني كعلاج بالبايوفيردين؛ (4) تم ري 150 مل من الماء في منطقة الجذور وتم رش محلول EDTA-FeNa على الأوراق الصغيرة لنبات الفول السوداني كعلاج EDTA-Fe. احتوت جميع محاليل الرش على 0.1% من Triton X-100 كعامل خافض للتوتر السطحي. تضمنت كل معالجة ست تكرارات في حقول في بكين وثماني تكرارات في حقول في بويانغ. كل تكرار في الحقل يتكون من 16 حفرة، وتم زراعة نباتين من الفول السوداني في حفرة واحدة. كانت المسافة بين النباتات 30 سم، والمسافة بين الصفوف 60 سم. عند الإزهار (53 يومًا بعد الزراعة في بكين وبويانغ)، وعند ظهور القرون (65 يومًا بعد الزراعة في بكين و68 يومًا بعد الزراعة في بويانغ)، وعند انتفاخ الثمار (79 يومًا بعد الزراعة في).
في بكين و84 نقطة في بويانغ) المرحلة، تم تكرار العلاجات المعنية. بعد أسبوعين من العلاج الثالث، تم قياس قيم SPAD وتركيز الحديد النشط في الأوراق الصغيرة. تم جمع تربة منطقة الجذور لقياس تركيز الحديد المتاح. ثم تم حصاد نباتات الفول السوداني لقياس المحصول.
تجربة وعاء مع طافح بيوفردين غير المتحول
لتعزيز النقطة التي تفيد بأن البيوفيردين يتم إنتاجه في التربة ويشارك في تغذية الحديد في الفول السوداني، قمنا بإجراء تجارب إضافية مع السلالة المخبرية.. الأيروجينوزا PAO1 وطفرتها المتجانسة المحددة الخالية من البيوفيردين PAO1. باو1 هو طفره مزدوجة من السايدروفور الأساسي بايوفيردين (من خلال حذف ) والسايدروفور الثانوي بايوشيلين (من خلال حذف و ) في P. aeruginosa PAO1. استخدمنا هذا الطافرة المزدوجة لأن البايوشيلين يتم تنظيمه بشكل زائد عندما يتم حذف البايوفيردين ويمكن أن يعوض عن تأثير البايوفيردين. لتقييم دور البايوفيردين، كان علينا استخدام الطافرة المزدوجة. تم إعداد التربة الرملية الكلسية العادية التي تفتقر إلى الحديد كما هو موضح أعلاه لتجربة الأصص، حيث تحتوي كل أصيص على 8 كجم من التربة. تم زراعة ستة نباتات فول سوداني (Arachis hypogaea L. cv. Luhua14) في أصيص لعلاج فول السوداني الأحادي. عند الإزهار (50 يوم بعد الزراعة)، مرحلة تحمل القرون (64 يوم بعد الزراعة)، ومرحلة انتفاخ الثمار (75 يوم بعد الزراعة)، تم إضافة 600 مل من ( نبات CFUنُسخ من Pseudomonas sp. 1502IPR-01، وP. aeruginosa PAO1 النوع البري، والطافرة الناقصة للسيدروفور PAO1تم ريها في منطقة الجذور. تم ري 600 مل من الماء في منطقة الجذور كتحكم. ظلت بقية ظروف زراعة الدفيئة متطابقة مع تلك الموصوفة أعلاه. تم خلط نباتات الفول السوداني التي تنمو في نفس الأصيص ومعاملتها كنسخة واحدة. كان لكل معالجة ثلاث أو أربع نسخ. في 84 يومًا بعد الزراعة، تم قياس قيم SPAD وتركيز الحديد النشط في الأوراق الشابة. تم جمع تربة منطقة الجذور لقياس تركيز الحديد المتاح.
تحليل البيانات
تم اختبار مجموعات البيانات للتوزيع الطبيعي باستخدام اختبار شابيرو-ويلك ولتباين التباين باستخدام اختبار ليفين. بالنسبة لمجموعات البيانات التي استوفت شروط التوزيع الطبيعي وتجانس التباين، تم استخدام اختبار الطالب البارامetric.تم استخدام اختبار t واختبار ANOVA مع اختبار LSD كاختبار لاحق. بالنسبة لمجموعات البيانات التي لم تستوفِ هذه الشروط، تم استخدام خوارزمية BoxCox لتحويل البيانات لتلبية افتراضات الاختبارات المعلمية. إذا لم يتم استيفاء هذه الشروط بعد التحويل، تم استخدام اختبارات ويلكوكسون غير المعلمية، واختبارات كروسكال-واليس مع اختبار دانييت T3 (للتصاميم ذات الاتجاه الواحد) أو اختبار شيرير-راي-هار (للتصاميم ذات الاتجاهين). تم استخدام خوارزمية BH للتصحيح.-القيم في المقارنات المتعددة. تم تحليل جميع الاختبارات المذكورة أعلاه، بالإضافة إلى PERMANOVA وتحليلات الارتباط، في R (الإصدار 4.2.2؛https://www.rproject.org) باستخدام حزم ‘stats’ (الإصدار 4.2.2)، ‘car’ (الإصدار 3.1-1)، ‘forecast’ (الإصدار 8.2.0)، ‘ggpubr’ (الإصدار 0.4.0)، ‘stats’ (الإصدار 4.2.2)، ‘agricolae’ (الإصدار 1.3-5)، ‘ggpubr’ (الإصدار 0.4.0)، ‘stats’ (الإصدار 4.2.2)، ‘PMCMRplus’ (الإصدار 1.9.6)، ‘rcompanion’ (الإصدار 2.4.21)، ‘vegan’ (الإصدار 2.5-7)، و ‘psych’ (الإصدار 2.2.5). بالنسبة لمجموعات البيانات التي لم تستوفِ توزيعًا طبيعيًا، تم حساب الارتباطات باستخدام ارتباط سبيرمان غير المعلمي (المقدم كـ سبيرمان رو). بالنسبة لمجموعات البيانات التي استوفت توزيعًا طبيعيًا، تم حساب الارتباطات باستخدام ارتباط بيرسون المعلمي (المقدم كـ بيرسون تم إنشاء خريطة الحرارة باستخدام حزمة ‘ComplexHeatmap’ (الإصدار 2.15.1) في R (الإصدار 4.2.2) بناءً على الوفرة النسبية للجنس. ثم تم تجميع الرسوم البيانية بما في ذلك المخططات المفاهيمية وتحسينها من الناحية الجمالية بواسطة Adobe Illustrator CC، 2020 (أدوبي، سان خوسيه، الولايات المتحدة الأمريكية). كانت جميع الاختبارات ثنائية الجانب. خريطة الحدود الإدارية للصين المستخدمة في الأشكال تم تنزيلها من منصة عامة: منصة بيانات مركز الموارد والبيئة السحابية.http://www.resdc.cn/).
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير Nature Portfolio المرتبط بهذه المقالة.
توفر البيانات
البيانات الخام لتسلسل 16 S rRNA المستخدمة في هذه الدراسة محفوظة في قاعدة بيانات NCBI تحت BioProject PRJNA788265. تسلسلات جينات 16S rRNA لـ 46 من البكتيريا الجذرية المعزولة محفوظة في GenBank برقم الوصول OL824679 – OL824724. تسلسل جينوم سلالة المرجع Pseudomonas sp. 1502IPR-01، الذي تم تحديده لهذه الدراسة، محفوظ في قاعدة بيانات NCBI تحت BioProject PRJNA788188. البيانات الخام التي تم إنشاؤها في هذه الدراسة متاحة في مستودع Figshare تحت رمز الوصول (https://doi.org/10.6084/m9.figshare. كما تم توفيرها أيضًا في ملف البيانات المصدر/البيانات التكميلية مع هذه الورقة. تم توفير بيانات المصدر مع هذه الورقة.
Li, C. et al. Syndromes of production in intercropping impact yield gains. Nat. Plants 6, 653-660 (2020).
Brooker, R. W. et al. Facilitation and biodiversity-ecosystem function relationships in crop production systems and their role in sustainable farming. J. Ecol. 109, 2054-2067 (2021).
Zuo, Y. & Zhang, F. Iron and zinc biofortification strategies in dicot plants by intercropping with gramineous species. A review. Agron. Sustain. Dev. 29, 63-71 (2009).
National Bureau of Statistics of China. China Statistical Yearbook (in Chinese) (China Statistics Press, 2021).
Dai, J. et al. From leguminosae/gramineae intercropping systems to see benefits of intercropping on iron nutrition. Front. Plant Sci. 10, 605 (2019).
Xiong, H. et al. Molecular evidence for phytosiderophore-induced improvement of iron nutrition of peanut intercropped with maize in calcareous soil. Plant Cell Environ. 36, 1888-1902 (2013).
Curie, C. & Mari, S. New routes for plant iron mining. N. Phytol. 214, 521-525 (2017).
Riaz, N. & Guerinot, M. L. All together now: regulation of the iron deficiency response. J. Exp. Bot. 72, 2045-2055 (2021).
Kobayashi, T., Nozoye, T. & Nishizawa, N. K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 133, 11-20 (2019).
Suzuki, M. et al. Development of a mugineic acid family phytosiderophore analog as an iron fertilizer. Nat. Commun. 12, 1558 (2021).
Lurthy, T., Pivato, B., Lemanceau, P. & Mazurier, S. Importance of the rhizosphere microbiota in iron biofortification of plants. Front. Plant Sci. 12, 744445 (2021).
Zhang, J. et al. NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 37, 676-684 (2019).
Voges, M., Bai, Y., Schulze-Lefert, P. & Sattely, E. S. Plant-derived coumarins shape the composition of an Arabidopsis synthetic root microbiome. Proc. Natl Acad. Sci. USA 116, 12558-12565 (2019).
Cadot, S. et al. Specific and conserved patterns of microbiotastructuring by maize benzoxazinoids in the field. Microbiome 9, 103 (2021).
Harbort, C. J. et al. Root-secreted coumarins and the microbiota interact to improve iron nutrition in Arabidopsis. Cell Host Microbe 28, 825-837 (2020).
Rajniak, J. et al. Biosynthesis of redox-active metabolites in response to iron deficiency in plants. Nat. Chem. Biol. 14, 442-450 (2018).
Ghosh, S. K., Bera, T. & Chakrabarty, A. M. Microbial siderophore – A boon to agricultural sciences. Biol. Control 144, 104214 (2020).
Sorrenti, G., Masiello, C. A. & Toselli, M. Biochar interferes with kiwifruit Fe-nutrition in calcareous soil. Geoderma 272, 10-19 (2016).
Maitra, S. et al. Intercropping-a low input agricultural strategy for food and environmental security. Agronomy 11, 343 (2021).
Shirley, M., Avoscan, L., Bernaud, E., Vansuyt, G. & Lemanceau, P. Comparison of iron acquisition from Fe-pyoverdine by strategy I and strategy II plants. Botany 89, 731-735 (2011).
Vansuyt, G., Robin, A., Briat, J. F., Curie, C. & Lemanceau, P. Iron acquisition from Fe-pyoverdine by Arabidopsis thaliana. Mol. PlantMicrobe Interact. 20, 441-447 (2007).
Soares, E. V. Perspective on the biotechnological production of bacterial siderophores and their use. Appl. Microbiol. Biotechnol. 106, 3985-4004 (2022).
Gu, S. et al. Competition for iron drives phytopathogen control by natural rhizosphere microbiomes. Nat. Microbiol. 5, 1002-1010 (2020).
Zuo, Y. M., Zhang, F. S., Li, X. L. & Cao, Y. P. Studies on the improvement in iron nutrition of peanut by intercropping with maize on a calcareous soil. Plant Soil 220, 13-25 (2000).
Li, L. et al. Root distribution and interactions between intercropped species. Oecologia 147, 280-290 (2006).
Gao, Y. et al. Distribution of roots and root length density in a maize/ soybean strip intercropping system. Agric. Water Manag. 98, 199-212 (2010).
Li, Q. S. et al. Belowground interactions impact the soil bacterial community, soil fertility, and crop yield in maize/peanut intercropping systems. Int. J. Mol. Sci. 19, 622 (2018).
Figueiredo, A. R. T., Özkaya, Ö., Kümmerli, R. & Kramer, J. Siderophores drive invasion dynamics in bacterial communities through their dual role as public good versus public bad. Ecol. Lett. 25, 138-150 (2022).
Kümmerli, R. Iron acquisition strategies in pseudomonads: mechanisms, ecology, and evolution. Biometals 36, 777-797 (2023).
Ahmed, E. & Holmstrom, S. J. Siderophores in environmental research: roles and applications. Micro. Biotechnol. 7, 196-208 (2014).
Shen, H. et al. AhDMT1, a transporter, is involved in improving iron nutrition and fixation in nodules of peanut intercropped with maize in calcareous soils. Planta 239, 1065-1077 (2014).
Dong, Q. et al. Maize and peanut intercropping improves the nitrogen accumulation and yield per plant of maize by promoting the secretion of flavonoids and abundance of Bradyrhizobium in rhizosphere. Front. Plant Sci. 13, 957336 (2022).
Martínez-Hidalgo, P. & Hirsch, A. M. The nodule microbiome: fixing rhizobia do not live alone. Phytobiomes J. 1, 70-82 (2017).
Li, B. et al. Root exudates drive interspecific facilitation by enhancing nodulation and fixation. Proc. Natl Acad. Sci. USA 113, 6496-6501 (2016).
Hu, H. Y. et al. Nitrogen fixation and crop productivity enhancements co-driven by intercrop root exudates and key rhizosphere bacteria. J. Appl. Ecol. 58, 2243-2255 (2021).
Vora, S. M., Joshi, P., Belwalkar, M. & Archana, G. Root exudates influence chemotaxis and colonization of diverse plant growth promoting rhizobacteria in the pigeon pea – maize intercropping system. Rhizosphere 18, 100331 (2021).
Wang, Z. et al. Forms of nitrogen inputs regulate the intensity of soil acidification. Glob. Change Biol. 29, 4044-4055 (2023).
Weng, Z. H., Li, G., Sale, P. W. G. & Tang, C. X. Application of calcium nitrate with phosphorus promotes rhizosphere alkalization in acid subsoil. Eur. J. Soil Sci. 73, e13153 (2022).
Ferreira, C. M. H. et al. Calcareous soil interactions of the iron(III) chelates of DPH and Azotochelin and its application on amending
iron chlorosis in soybean (Glycine max). Sci. Total Environ. 647, 1586-1593 (2019).
Havlin, J. L., Beaton, J. D., Tisdale, S. L. & Nelson, W. L. Soil Fertility and Fertilizers: An Introduction to Nutrient Management 7th edn (Pearson College Div, 2004).
Federal Ministry of Labour and Social Affairs (BMAS). Technical Rules for Biological Agents: Classification of Prokaryotes (Bacteria and Archaea) into Risk Groups (TRBA 466)-2015 (2015).
Cornu, J. Y. et al. Contrasting effects of pyoverdine on the phytoextraction of Cu and Cd in a calcareous soil. Chemosphere 103, 212-219 (2014).
Verma, S., Chakdar, H., Kumar, M., Varma, A. & Saxena, A. K. Microorganisms as a sustainable alternative to traditional biofortification of iron and zinc: status and prospect to combat hidden hunger. J. Soil Sci. Plant Nutr. 21, 1700-1717 (2021).
Li, C. J., Zhu, X. P. & Zhang, F. S. Role of shoot in regulation of iron deficiency responses in cucumber and bean plants. J. Plant Nutr. 23, 1809-1818 (2000).
Reichman, S. M. & Parker, D. R. Critical evaluation of three indirect assays for quantifying phytosiderophores released by the roots of Poaceae. Eur. J. Soil Sci. 58, 844-853 (2007).
Bolyen, E. et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852-857 (2019).
Beule, L. & Karlovsky, P. Improved normalization of species count data in ecology by scaling with ranked subsampling (SRS): application to microbial communities. PeerJ 8, e9593 (2020).
Schwyn, B. & Neilands, J. B. Universal chemical-assay for the detection and determination of siderophores. Anal. Biochem. 160, 47-56 (1987).
Davis, E. W., Okrent, R. A., Manning, V. A. & Trippe, K. M. Unexpected distribution of the 4-formylaminooxyvinylglycine (FVG) biosynthetic pathway in Pseudomonas and beyond. PLoS ONE 16, eO247348 (2021).
Budzikiewicz, H., Schafer, M., Fernandez, D. U., Matthijs, S. & Cornelis, P. Characterization of the chromophores of pyoverdins and related siderophores by electrospray tandem mass spectrometry. Biometals 20, 135-144 (2007).
Sultana, R. et al. A pyoverdin from Pseudomonas sp. CFML 95-275. Z. Naturforsch. C. 55, 857-865 (2000).
Jin, C. W., Li, G. X., Yu, X. H. & Zheng, S. J. Plant Fe status affects the composition of siderophore-secreting microbes in the rhizosphere. Ann. Bot. 105, 835-841 (2010).
Wang, N. Q. et al. Microbiome convergence enables siderophore-secreting-rhizobacteria to improve iron nutrition and yield of peanut intercropped with maize. Figshare. https://doi.org/10.6084/m9. figshare. 24648306 (2023).
Wang, N. Q. et al. Microbiome convergence enables siderophore-secreting-rhizobacteria to improve iron nutrition and yield of peanut intercropped with maize. wnq13579/Peanut-maize-intercroppingmicrobiome. Zenodo. https://doi.org/10.5281/zenodo. 10212602 (2023).
الشكر والتقدير
تم دعم هذا البحث ماليًا من قبل المؤسسة الوطنية للعلوم الطبيعية في الصين (رقم المنحة 32372810 لـ Y.-M. Z.، 32102469 لـ N.-Q. W.، و42325704 لـ Z. W.)، البرنامج الوطني الرئيسي للبحث والتطوير في الصين (رقم المنحة 2022YFD1901500/2022YFD1901501،
2023YFD1700203 لـ Y.-M. Z. و2021YFD1900100 لـ Z. W.)، ومؤسسة العلوم الوطنية السويسرية SNSF (رقم المنحة 31003A_182499 لـ R. K.). نشكر الدكتور البروفيسور يانغ باي من الأكاديمية الصينية للعلوم، والدكتور البروفيسور تشويهوا كونغ، والدكتور البروفيسور لينغ شو من جامعة الزراعة الصينية على المناقشات والاقتراحات القيمة.
مساهمات المؤلفين
Y.-M.Z.، Z. W.، N.-Q. W. وT.-Q. W. صمموا البحث. قام N.-Q. W. وT.-Q. W. بتنفيذ وتحليل معظم التجارب في المختبر والميدان، وQ.-F. L. K.-G. W.، Z.-C. D.، Z.-G. C.، W.-Q.، J.-D.، L.-N.، وJ.-Y. C. قاموا بتنفيذ وتحليل معظم التجارب في الدفيئة. قدمت Y. C. وM. W. ظروف المختبر ومدخلات فكرية لتحديد هيكل السايدروفور وكتبوا القسم “التعرف الهيكلي على البايفيردين” في الطرق. قدم R. K. وZ. W. مدخلات فكرية، وإرشادات لتحليل بيانات الميكروبيوم وساعدوا في تفسير البيانات. قدمت M. W. بعض المدخلات الفكرية لهذه المخطوطة، وإرشادات لتحليل بيانات الميكروبيوم والتصور. كتب N.-Q. W.، T.-Q. W.، F.-S. Z.، R. K.، Z. W.، وY.-M. Z. وراجعوا المخطوطة. ناقش جميع المؤلفين النتائج وعلقوا على المخطوطة. أشرف Y.-M. Z. على الدراسة.
كلية الموارد والعلوم البيئية، المختبر الوطني الرئيسي لاستخدام وإدارة المغذيات (SKL-NUM)، الأكاديمية الوطنية للتنمية الخضراء الزراعية، جامعة الزراعة الصينية، 100193 بكين، الصين. مختبر جيانغسو الرئيسي للبحث واستخدام الموارد النباتية، مركز أبحاث الهندسة في مقاطعة جيانغسو للزراعة البيئية والاستخدام العالي القيمة للمواد الطبية الصينية، معهد علم النبات، مقاطعة جيانغسو والأكاديمية الصينية للعلوم، 210014 نانجينغ، جيانغسو، الصين. قسم علم أمراض النبات، المختبر الرئيسي لمناعة النبات، جامعة نانجينغ الزراعية، نانجينغ 210095، الصين. مختبر جيانغسو الرئيسي لاستخدام النفايات العضوية الصلبة، المختبر الرئيسي للأسمدة العضوية في الصين، مركز الابتكار التعاوني في جيانغسو للنفايات العضوية الصلبة، مركز الهندسة التابع لوزارة التعليم للأسمدة الموفرة للموارد، جامعة نانجينغ الزراعية، نانجينغ 210095، الصين. قسم البيولوجيا الكمية، جامعة زيورخ، زيورخ، سويسرا. ساهم هؤلاء المؤلفون بالتساوي: نانكي وانغ، تيانكي وانغ، يوي تشين. البريد الإلكتروني: mwang@njau.edu.cn; weizhong@njau.edu.cn; zuoym@cau.edu.cn
Intercropping has the potential to improve plant nutrition as well as crop yield. However, the exact mechanism promoting improved nutrient acquisition and the role the rhizosphere microbiome may play in this process remains poorly understood. Here, we use a peanut/maize intercropping system to investigate the role of root-associated microbiota in iron nutrition in these crops, combining microbiome profiling, strain and substance isolation and functional validation. We find that intercropping increases iron nutrition in peanut but not in maize plants and that the microbiota composition changes and converges between the two plants tested in intercropping experiments. We identify a Pseudomonas secreted siderophore, pyoverdine, that improves iron nutrition in glasshouse and field experiments. Our results suggest that the presence of siderophore-secreting Pseudomonas in peanut and maize intercropped plays an important role in iron nutrition. These findings could be used to envision future intercropping practices aiming to improve plant nutrition.
One of humanity’s greatest challenges is how to sustainably feed a large population , especially in countries such as China, comprising almost a fifth of the world population but having limited available arable land. One solution to guarantee food security is to grow at least two crops simultaneously in the same field because intercropping often increases productivity, resource use efficiency, pest and pathology control, and ecological sustainability . Peanut (Arachis
hypogaea L .) /maize (Zea mays) intercropping is common in China as it is more effective and ecologically more sustainable than peanut monocropping, particularly for small land holders . Peanut is an important oilseed legume. China produces and Northern China Plain approximately of the global peanut production (Fig. 1a). However, peanut yield and quality are severely hampered in this area because of severe iron (Fe) deficiency prevailing in the alkaline and
Fig. 1 | Maize/peanut intercropping improves peanut iron nutrition and yield in calcareous soils. a Average peanut production and the global rate of four provinces from Northern China Plain and position of field experiments (indicated by dots). Pictures of monocropping and intercropping peanut in field sites and two associated phenotypes of iron-deficiency chlorosis: Soil and Plant Analyzer Development value (SPAD value indicating chlorophyll content) and active iron (indicating iron-deficiency extent) in young leaves. Values are means standard deviation (SD) ( biologically independent samples, -value for SPAD: , -value for active iron: 0.002 ). Skewed data distributions or data with heterogeneity of variances were transformed using the BoxCox algorithm to meet the assumptions of parametric tests. If data did not meet the assumptions after
transformation, non-parametric tests such as the Wilcoxon test were used. The BH algorithm was used to correct -values for multiple comparisons. Asterisks indicate significant differences between two groups: ; ; , n.s not significant. Two-sided tests were used for alternative hypothesis testing. c Land equivalent ratio (LER) (LER is defined as the difference between the observed and the expected yield, indicating the land-use efficiency) and net effect (NE) (NE is defined as the sum of partial relative yields per species, indicating the net effect of intercropping) of peanut/maize intercropping indicate significant advantage in Henan during 2011-2015 and in five farms from Northern China Plain in 2015 under field conditions. Source data are provided.
calcareous soils, making iron insoluble and bio-unavailable . This problem is alleviated through intercropping with maize, which improves the iron nutrition and photosynthetic efficiency of peanut (Fig. 1b) and thereby increases yield and land-use efficiency (Fig. 1c). This intercropping is thought to offer an effective and sustainable way to ensure iron and zinc biofortification to prevent hidden hunger .
Here, we use peanut/maize intercropping as a model system to elucidate the exact mechanism leading to increased iron nutrition, particularly focusing on the yet unexplored role of the rhizosphere microbiome. It is known that peanut/maize intercropping improves iron nutrition via direct belowground facilitation through root exudates . Higher plants show two alternative iron-deficiencyinduced molecular and physiological responses . Peanut follows strategy I (dicots and non-graminaceous monocots) and directly reduces Fe (III) to Fe (II) followed by Fe (II) absorption . In contrast, maize follows strategy II (graminaceous monocots) and secretes phytosiderophores of the mugineic acid family (MAs) to chelate insoluble Fe(III) followed by MAs-Fe(III) complex absorption . Maize outperforms peanut in iron acquisition in alkaline soils because strategy II is less sensitive to high . It was proposed that intercropping benefits peanut because it enhances deoxymugineic acid (DMA) secretion from maize root, which solubilizes more Fe (III) in the soil that is then absorbed as DMA-Fe(III) complex by nearby peanut plants .
This current plant-plant interaction model for iron utilization does not integrate root-microbiome interactions. This is surprising because it is well established that the rhizosphere microbiome is critical for both plant fitness and iron acquisition . Specifically, strategy I and II plants secrete different root exudates under iron-limited conditions, to recruit various rhizobacteria that improve plant nutrition in
general and iron acquisition in particular . A common mechanism of iron acquisition in these beneficial bacteria operates through the secretion of siderophores to dissolve insoluble . For these reasons, we hypothesize that peanut/maize intercropping could modulate the rhizosphere microbiome through the proximity of plant roots, which in turn could promote the mixing of root exudates, microbiome members, their siderophores and thus improve iron acquisition opportunities.
To test our hypothesis, we first examined whether increased iron acquisition in intercropping depends on the presence of a functional rhizosphere microbiome. Subsequently, we combined microbiome profiling, functional strain characterization, substance identification, and greenhouse and field experiments to establish the mechanistic links between cropping systems, belowground root-microbiome interaction networks and iron acquisition. Our study design allows us to identify microorganisms and microbial metabolites that contribute to the increase in nutrient use efficiency and crop yield by intercropping.
Results
The rhizosphere microbiome contributes to iron nutrition improvement of peanut intercropped with maize
In the first experiment, we explored whether peanut/maize intercropping is beneficial under greenhouse conditions in naturally ironlimited soils. We found that intercropping indeed improved iron nutrition in peanut (Fig. 2a, b), but not in maize (Supplementary Fig. 1). Specifically, intercropping increased chlorophyll levels (Soil Plant Analysis Development (SPAD) value, to ) and active iron concentration ( to , indicating Fe-deficiency extent) in young leaves from 53 days post sowing (dps) onwards (Fig. 2a, b).
Compared to monocropping peanut, there was a significant increase in the available iron concentration in the rhizosphere ( , i.e., iron prevails in a form that can be absorbed by the plant) (Fig. 2b). In contrast, monocropping peanut started to show iron-deficiency chlorosis in young leaves at 53 dps , which progressively worsened over time (Fig. 2a, b).
To test whether the rhizosphere microbiome is involved in the observed iron nutrition improvement in intercropping peanut, we repeated our experiments in normal (unsterilized) and sterilized soils (Fig. 2c). For all the three measured parameters – SPAD, active iron in leaves and available iron in the rhizosphere – there were significant statistical interactions between soil treatments (sterilized vs. normal)
Fig. 2 | Rhizosphere microbiome contributes to the iron nutrition improvement of peanut intercropped with maize. a Phenotypes and Soil Plant Analysis Development (SPAD) values of plants and young leaves from monocropping peanut and intercropping peanut at , and 73 days post sowing (dps). Table shows SPAD values in peanut young leaves. Values show means SD ( biologically independent samples and -values are for 46 , 53, 63 and 73 dps ). Bar indicates 32 cm . b Active iron in young leaves and available iron (indicating the amount of iron utilizable by plants) in the rhizosphere of monocropping peanut and intercropping peanut in pots. Points and shaded areas show mean values and 95% confidence interval from four independent biological replicates, respectively. c Phenotype of monocropping peanut and intercropping peanut grown in sterile and normal calcareous soil at 73 dps. d SPAD values and active iron in young leaves and available iron in the rhizosphere of peanut at 73 dps grown in either normal or sterile soil. Bars with error bars represent mean
and cropping types (intercropping peanut vs. monocropping peanut), demonstrating that intercropping peanut was only beneficial and increased iron availability for peanut in the presence of a functional rhizosphere microbiome (Fig. 2d).
Pseudomonas spp. – cross-enriched from maize to peanut – are associated with improved iron nutrition
Intercropping alters rhizosphere microbiota. Given that the rhizosphere microbiome is essential for improving iron nutrition in intercropping peanut, we hypothesized that intercropping improves iron acquisition via modification of the peanut microbiome. To test our hypothesis, we used 16 S rRNA amplicon sequencing to profile the rhizosphere microbiome of peanut and maize plants from monocropping and intercropping at four-time points ( and 73 dps , Supplementary Fig. 2). Rhizobacterial communities formed four distinct clusters (monocropping peanut, monocropping maize, intercropping peanut, intercropping maize) at all stages and showed the greatest divergence at 73 dps (Fig. 3a). While bacterial diversity in the rhizosphere bacterial communities was not affected by intercropping (Supplementary Fig. 3a), we observed that intercropping peanut and intercropping maize microbiomes are more similar than the monocropping peanut and monocropping maize microbiomes (Fig. 3a and Supplementary Fig. 3b). This pattern suggests that maize and peanut microbiome members are exchanged, possibly through the interwoven inter-species root network formed with intercropping (Supplementary Fig. 4). Altogether, our results reveal that intercropping induces both a change in microbiome composition and a convergence between the peanut and maize microbiomes.
Pseudomonas is the strongest biomarker closely associated with improved iron nutrition. To detect key bacterial taxa linked with improved iron nutrition, we employed the linear discriminant analysis effect size (LefSe). We identified 10 biomarkers at the genus level (absolute LDA-scores > 3.0 and ) that were enriched in intercropping peanut compared to monocropping peanut (Supplementary Fig. 5) and were more abundant in monocropping maize compared to monocropping peanut (Fig. 3b). Hence, these taxa are the top candidates for being cross-enriched from maize to peanut in intercropping. We further found that the abundance of those 10 genera positively correlated with either the active iron in young leaves, the iron availability in the rhizosphere, or both metrics (Fig. 3b, Supplementary Fig. 6 showing positive correlation of the two iron metrics). While five genera (Pseudomonas, Pseudoxanthomonas, Luteolibacter, Allorhizobium-Neorhizobium-Para-rhizobium-Rhizobium, Sphingobium) showed positive correlations with both iron metrics, Pseudomonas had the strongest association (Spearman’s ) with rhizosphere iron availability (Fig. 3b and Supplementary Data 1). Thus, our analysis reveals five keystone taxa possibly guiding iron nutrition improvement in intercropping peanut, with Pseudomonas being the top candidate.
standard deviation (SD) ( biologically independent samples for SPAD value and for active iron and available iron) and dots represent individual values. For , and crop type indicates intercropping or monocropping. Parametric Student’s -test and ANOVA with LSD as post-hoc test were used for data sets that were normally distributed and had homogeneous variances. Skewed data distributions or data with heterogeneity of variances were transformed using the BoxCox algorithm to meet the assumptions of parametric tests. If data did not meet the assumptions after transformation, non-parametric tests such as the Wilcoxon test, Kruskal-Wallis test with Dunnett T3 test or Scheirer-Ray-Hare test were used. The BH algorithm was used to correct -values for multiple comparisons. Asterisks indicate significant differences between two groups: ; ; , n.s not significant. Different letters indicate significant differences among groups. Two-sided tests were used for alternative hypothesis testing. Source data are provided.
Moreover, we found that intercropping also leads to microbiome shifts in the maize rhizosphere. Particularly, we identified three genera (Azotobacter, Rubellimicrobium and Bryobacter) that were enriched in intercropping maize compared to monocropping maize (Supplementary Fig. 7).
Pseudomonas rhizobacteria from intercropping show high siderophore-secreting ability
Due to the high capacity of siderophores to solubilize iron , we hypothesized that siderophores secreted by rhizobacteria in intercropping could be responsible for the enhanced iron bioavailability in the peanut rhizosphere. To test this hypothesis, we isolated siderophore-secreting rhizobacteria from intercropping peanut using the chrome azurol S (CAS) plate assay (Fig. 4a). We isolated over 300 siderophore-secreting rhizobacteria strains, among which 46 strains had particularly high siderophore-secreting activity (halo diameter on CAS plates , Fig. 4b). Among the top producers, were Pseudomonas spp. (according to their 16S rRNA sequence), and the majority of these strains (63.0%) had sequence identities > 97% with a representative amplicon sequence variant (ASV487) obtained by the DADA2 algorithm, capturing one particular clade of Pseudomonas spp. (Fig. 4b and Supplementary Table 1). Importantly, ASV487 was undetectable in monocropping peanut, but detectable in intercropping peanut and its relative abundance was higher in monocropping maize than intercropping maize (Fig. 4c). Additionally, ASV487-similar strains were isolated in the intercropping peanut rhizosphere at a key stage of the iron nutritional shift ( ) and at the stage where the iron nutrition gaps between monocropping and intercropping peanut were widest ( 73 dps ) (Supplementary Table 1). These results suggest that ASV487 represents a group of high siderophore-secreting Pseudomonas strains that were cross-enriched from maize to peanut during intercropping and are associated with improved iron nutrition in peanut.
Next, we focused on Pseudomonas sp. 1502IPR-01, a strain that is identical to ASV487 and had the highest siderophore-secreting ability among the isolated strains (Fig. 4b and Supplementary Fig. 8). We used Pseudomonas sp. 1502IPR-01 as a representative strain to experimentally explore its role in iron nutrition improvement in intercropping peanut. We sequenced the whole-genome of Pseudomonas sp. 1502 IPR-01 and found that 1502IPR-01 is similar to . extremorientalis (Fig. 4d and Supplementary Fig. 9). By searching its genome through cluster analysis, we detected the pyoverdine biosynthetic and secretion cluster sharing 51.4-84.1% protein sequence identity with homologous genes of . aeruginosa PAO1, the model strain for pyoverdine research (Fig. 4e and Supplementary Table 2).
To confirm that Pseudomonas sp. 1502IPR-01 indeed secretes pyoverdine as its primary siderophore, we isolated the iron-chelating compound (Supplementary Fig. 10) from the supernatant, after cultivation in the iron-limited medium. Our assay revealed that of the iron-chelating ability of supernatants was
Fig. 3 | Intercropping induces shifts in rhizobacteria community composition and reveals Pseudomonas as one of the top candidate taxa involved in intercropping peanut iron nutrition improvement. a Unconstrained principal coordinate analysis (PCoA) with UniFrac distances (measuring the distance between communities based on the lineages they contain) of rhizobacteria microbiomes collected from monocropping peanut, intercropping peanut, intercropping maize, and monocropping maize. The results of permutational multivariate analysis of variance (PERMANOVA, also known as Adonis) are shown. For PcoA and Adonis analysis, relative abundances were used. Genus biomarker discriminating monocropping and intercropping peanut, identified by the linear discriminant analysis effect size, LefSe (LDA and ). Shown are the relative dynamic abundances of the biomarker taxa in intercropping peanut, monocropping peanut
and monocropping maize as normalized z-score of mean value of three biologically independent samples and depicted as a heatmap spanning the continuum from low (blue) to high (red) z-scores. Relative abundances of each genus are shown with box plots: center = median, box boundaries = 1st and 3rd quartiles, whiskers = minima and maxima. The table shows Spearman correlations between the relative abundances and two metrics describing the plants’ iron nutrition state, indicated as heatmap spanning the continuum from negative (green) to positive (pink) correlations. The correlation analyses were based on 24 independent samples. Significant correlations are indicated with asterisks: ; ; ; n.s not significant. Exact Spearman values with significant levels are listed. The number of biologically independent samples for the heatmap date is three. Twosided tests were used for alternative hypothesis testing. Source data are provided.
attributable to siderophores (Supplementary Table 3). Structure elucidation and chemical analysis (Fig. 4f, Supplementary Figs. 11-21 and Supplementary Tables 4 and 5, details in “Methods” section) yielded a chemical formula of , and the chemical structure confirms that the primary siderophore secreted by Pseudomonas sp. 1502IPR-01 is a pyoverdine (Fig. 4f).
Next, we assessed the activity of the detected pyoverdine to bind III) from an insoluble source in suspension. III chelation is indicated by a color change from clear to yellow to brown (Fig. 4f). We found that the chelated iron concentration in the solution increased with an equimolar amount of the added pyoverdine, matching the color change of the pyoverdine-Fe(III) control complex (Fig. 4 f and Supplementary Fig. 10c). In sum, our results identify pyoverdine as the primary siderophore secreted by Pseudomonas sp. 1502IPR-01, and show its high capacity to dissolve Fe(III) from insoluble , which corresponds to the main form of iron in calcareous soil .
Pseudomonas sp. 1502IPR-01 and its pyoverdine improve peanut iron nutrition in greenhouse and field experiments
To experimentally validate that Pseudomonas is a keystone taxon improving iron nutrition through pyoverdine secretion, we conducted greenhouse and field experiments. For the greenhouse experiments, we grew peanut in monocropping and intercropping peanut in sterilized calcareous soil either without treatment as control, or with Pseudomonas sp. 1502IPR-01 or its iron-free pyoverdine as treatments. Both treatments significantly increased SPAD values, active iron concentration in young leaves and in the rhizosphere, and improved peanut biomass in monocropping and intercropping peanut (Fig. 5a, b). We observed the same significant improvements when repeating the experiment in normal non-sterile soil in monocropping peanut (Fig. 5b). Thus, Pseudomonas sp. 1502IPR-01 and pyoverdine treatments can compensate for iron deficiency in peanut in the
absence of a functional microbiome in intercropping and monocropping peanut in normal soil. In contrast, the iron nutrition of maize did not improve when treated with Pseudomonas sp. 1502IPR-01 or pyoverdine (Supplementary Fig. 22). However, maize biomass significantly increased with both treatments in sterilized and normal soil in intercropping maize (Supplementary Fig. 22), suggesting that maize benefits from intercropping with peanuts by other means than improved iron nutrition.
Next, we investigated whether the application of Pseudomonas sp. 1502IPR-01 and its iron-free pyoverdine are also effective treatments in the field. With intercropping peanut in normal soils, the application of Pseudomonas sp. 1502IPR-01 and its iron-free pyoverdine had no effect on iron nutrition and biomass (Supplementary Fig. 23), probably because intercropping itself suffices to mitigate iron deficiency in peanut. Hence, we focused exclusively on monocropping peanut and treated plants with either Pseudomonas sp. 1502IPR-01 or pyoverdine (both applied via root watering) at two field sites in North China Plain, characterized by calcareous soils. At both sites, treatments significantly increased SPAD values (increase: 45.9-67.6%), active iron concentration in young leaves (78.2-107.5%), iron availability in the rhizosphere (53.6-73.2%) and crop yield (44.8-89.8%), compared to the control plants that received no treatment (Fig. 5c, d). We further applied a traditional treatment used by local farmers, which involved foliar spraying with EDTA-Fe, a synthetic iron chelator. We observed that treating plants with either Pseudomonas sp. 1502IPR-01 or its pyoverdine increased peanut iron nutrition and yield to a similar extent and in one case even more than the traditional EDTA-Fe treatment (Fig. 5d). These results indicate that secreted or added pyoverdine of Pseudomonas sp. 1502IPR-01 could effectively chelate Fe(III) in calcareous soils in the field, and this mechanism is responsible for improved peanut iron nutrition and crop yield. Thus, pyoverdine could be considered as an iron fertilizer for sustainable agriculture without the need for external iron supply.
Fig. 4 | Functional siderophore-secreting rhizobacteria and siderophore characterization. a Schematic diagram showing the protocol used to isolate
isolate Pseudomonas sp. 1502IPR-01 with other representatives of Pseudomonas spp. strains based on 100 housekeeping genes as phylogenetic markers. The consensus tree is constructed from 1000 bootstrap trees. Numbers on branches show bootstrap supports (%). e The pyoverdine biosynthesis secretion and uptake genes of Pseudomonas sp. 1502IPR-01. Sequences shared between Pseudomonas sp. 1502IPR-01 and Pseudomonas aeruginosa PAO1 are connected by bands that are shaded according to their identity level, from black ( ) to white ( ). The arrow color indicates the function of the genes. The putative chemical structure of pyoverdine from Pseudomonas sp. 1502IPR-01 and its capacity to chelate Fe(III) from precipitated . The panels show the color change of the supernatant induced by the pyoverdine and the correlation (Pearson correlation coefficient) between the iron release from and the pyoverdine concentration in the reaction mix. The purple line and the gray area indicate a linear regression line and its 95% confidence interval, respectively. The green points indicate mean value across three independent samples. Two-sided tests were used for alternative hypothesis testing. Source data are provided.
Fig. 5 | Pseudomonas sp. 1502IPR-01 and its siderophore (pyoverdine) prevent iron-deficiency chlorosis and improve peanut growth and yield. a Phenotypes
For bars and error bars represent the mean standard deviation (SD) and dots represent individual values. For boxplot center lines show the median, boxplot bounds show the first quartile Q1 and the third quartile Q3, and whiskers show 1.5 (Q3-Q1) below and above Q1 and Q3, and black dot represent outliers and open dots represent individual values, respectively. For and data obeyed normal distribution and homogeneity of variance, ANOVA with LSD as post-hoc test is used in parametric test. For skewness data or data with heterogeneity of variance, data is transformed using BoxCox algorithm for parametric test. Kruskal test with Dunnett T3 test is used if data did not meet the assumptions of parametric test after BoxCox algorithm. Different letters indicate significant differences among groups. Multiple testing corrections are performed by BH algorithm. Asterisks indicate significant differences between two groups: ; ; , n.s not significant. For all parameters, represents the number of biologically independent samples. Two-sided tests were used for alternative hypothesis testing. Source data are provided.
Validation experiments reveal that pyoverdine directly affects plant iron metabolism
To functionally validate that pyoverdine contributes to improved peanut iron nutrition, we conducted experiments with a defined pyoverdine null mutant (Fig. 6a). Both the lab strain P. aeruginosa PAO1 and the natural isolate Pseudomonas sp. 1502IPR-01 significantly improved monocropping peanut iron nutrition and biomass in normal soil. In stark contrast, treatment with the siderophore-deficient mutant PAO1 (Fig. 6b) had no beneficial effects on plants, showing that pyoverdine is required to improve peanut iron nutrition.
We then asked whether pyoverdine secretion feeds back on the activity of the plants’ own iron acquisition systems. Indeed, we observed that the ferric-chelate reductase (FCR) activity of peanut is significantly upregulated in the presence of the siderophore-deficient mutant (Fig. 6b), but down-regulated in the presence of pyoverdineproducing wildtype strains and when pyoverdine is added (Supplementary Fig. 24). The same responses occurred in maize, which downregulated its phytosiderophore DMA in the presence of Pseudomonas sp. 1502IPR-01 or its pyoverdine (Supplementary Fig. 24). These experiments highlight that pyoverdine directly affects the iron metabolism of plants.
Discussion
Intercropping is important for sustainable agriculture and food security . However, the precise mechanisms by which intercropping increases yield often remain unclear. In this study, we find that improved iron nutrition is mediated by the bacterial rhizosphere microbiome and arises through microbiome convergence between the two plant species. Specifically, we find that intercropping enables beneficial bacteria such as Pseudomonas spp. to translocate from the maize to the peanut rhizosphere, where these bacteria increase the
amount of bioavailable iron through the secretion of their siderophore pyoverdine. The iron chelated to pyoverdine increases iron nutrition in peanut plants and negates iron-deficiency chlorosis in leaves (Figs. 4-7). Siderophores (e.g., pyoverdine) have previously been associated with beneficial effects for plants , and our study now provides evidence linking bacterial pyoverdine production in planta to improved plant iron nutrition (Fig. 6). This insight closes a major knowledge gap in the research field. Crucially, we show that the uncovered microbe-plant interaction is essential to improve iron nutrition, as intercropping in the absence of a functional microbiome is not beneficial (Figs. 2, 5). Our results further show how detailed knowledge of inter-kingdom interaction mechanisms helps to develop ecologically sustainable agriculture and food security.
Two lines of evidence suggest that the microbiome interactions observed in our greenhouse experiments occur in the field. First, the peanut and maize roots can easily connect across at least 60 cm in the field, matching the results from our pots experiments . Furthermore, root connections across similar distances also occur between maize/ faba bean and maize/soybean . Thus, the rhizosphere microbiome convergence based on root connections between peanut and maize observed in the greenhouse likely also occurs in the field. Second, our field experiments themselves demonstrated that the addition of Pseudomonas sp. 1502IPR-01 or its pyoverdine improved peanut iron nutrition, showing that peanut plants can benefit from iron made available by Pseudomonas spp. Hence, pyoverdine-mediated iron mobilization during microbiome convergence could be a major component of improved iron nutrition and yield of peanut intercropped with maize.
Our work strengthens the view that Pseudomonas is a keystone taxon involved in plant iron nutrition. We show that Pseudomonas sp. 1502IPR-01 and its pyoverdine improve peanut iron nutrition in
Fig. 6 | Pyoverdine secreted by Pseudomonas spp. is essential to improve peanut iron nutrition and biomass. a Phenotypes of peanut plants under control conditions and when treated with a Pseudomonas aeruginosa PAO1 mutant without pyoverdine secreting ability (PAO1 ), Pseudomonas aeruginosa PAO1 wild type (PAO1 WT) with pyoverdine secretion ability, and Pseudomonas sp. 1502IPR-01 with pyoverdine secretion ability (all rhizosphere application). Soil Plant Analysis Development (SPAD) values, active iron in young leaves, available iron in the rhizosphere, biomass and Fe (III) reduction capacity of peanut under control conditions and the three treatments (PAO1ApvdDpchEF, PAO1 WT and 1502IPR-01) in greenhouse conditions. For bars and error bars represent the mean standard deviation (SD) and dots represent individual values. ANOVA with
LSD post-hoc test was used when data followed the normal distribution and had homogenous variances. For skewed data sets or data with heterogeneous variances, the BoxCox transformation algorithm was applied. When the data still did not meet the assumptions of parametric tests, Kruskal-Wallis test with Dunnett T3 test was used. Different letters indicate significant differences between groups. Multiple testing corrections were performed by the BH algorithm. Asterisks indicate significant differences between two groups: , not significant. For all parameters, n represents the number of biologically independent samples. Two-sided tests were used for alternative hypothesis testing. Source data are provided.
Fig. 7 | Model of how peanut/maize intercropping enriches for Pseudomonas spp. in the rhizosphere to improve iron nutrition via secreting siderophore pyoverdine. a Rhizosphere microbiome convergence occurs in peanut/maize intercropping through root interactions (whole microbiome level). b Pseudomonas
cross-enriches during intercropping from the maize to the peanut rhizosphere (functional microbe). c Pseudomonas secretes the siderophore pyoverdine, which increases the iron availability in intercropping peanut rhizospheres (functional microbial metabolite).
calcareous soils in both greenhouse and field conditions through microbiome convergence between peanut and maize (Figs. 3-6). Similar findings regarding the enrichment of Pseudomonas in peanut intercropped with maize have been reported in acidic soil . However, important to note is that our analysis revealed additional members of the rhizosphere microbiome that were also associated with iron nutrition improvement, for example, members of the genera Sphingobium and Luteolibacter. Therefore, it would be important to conduct experimental work with characterized strains of these taxa in the future, to assess their role more closely. The observed beneficial effects in intercropping are likely a bacterial community effect with Pseudomonas spp. possibly taking the lead in the process (Figs. 3 and 4). Attributing positive effects to a single species is clearly advantageous when aiming at developing bio-fertilizer treatments for agricultural applications.
The positive effects of pyoverdines on iron nutrition suggest that these molecules take on a yet undescribed role for rhizosphere communities under field conditions. Previous research revealed that specific bacterial siderophores, including pyoverdines, can be beneficial for plants because they suppress plant pathogens in the rhizosphere . The proposed mechanism entails that pyoverdines (and other siderophores) are strong iron chelators, and because they are taxonspecific that can lock iron away from pathogens that do not possess the matching receptors, hence acting as ‘public bads . Competition for iron can therefore drive phytopathogen control and crop protection by natural rhizosphere microbiomes . In intercropping,
pyoverdine seems to primarily serve as a ‘public good’ that solubilizes iron for the plant and possibly also for rhizosphere microbiome members possessing matching receptors . In principle, pyoverdine could exert a double role in intercropping by suppressing phytopathogens (public bad effect) and by improving iron nutrition of plants (public good effect). Important to note is that plants and microbes may also compete for iron. Siderophores like pyoverdines are primarily produced by bacteria to satisfy their own need for iron. But given that siderophores are highly diffusible they might become available and utilizable for plants (Figs. 5-6). The loss of siderophores to plants could exert positive feedback on bacterial siderophore production as compensation. Conversely, microbial siderophores may also feedback on plant iron-uptake systems. For example, microbial siderophores may chelate iron from phytosiderophore-Fe(III) complexes or Fe(III) chelated by other root exudates, rendering these uptake systems less efficient. Thus, it remains unclear whether the observed iron nutrition improvement in intercropping occurs because plants efficiently exploit bacterial siderophores or whether there is a mutualistic interaction between the two interacting partners.
Our study reveals that pyoverdine can dissolve insoluble iron and make it bioavailable for plants (Figs. 4f, 5, 6). Earlier work has also reported the ability of several plant species to obtain iron from ironloaded pyoverdines . However, the exact biochemical mechanism of how plants retrieve iron from pyoverdines remains unclear. One option is that plants take up the entire Fe (III)-pyoverdine complex and retrieve iron through the degradation of pyoverdine. The pyoverdine
backbone consists of a peptide chain and could thus be degraded by proteases. Another option is that plants induce the reduction of Fe (III) to Fe (II) through their ferric-chelate reductase (FCR) to then directly absorb . While FCR certainly plays a role, we observed that pyoverdine supplementation led to the down-regulated FCR activity in peanuts and DMA production in maize, which represent the regular iron-deficiency responses of these two plant species (Supplementary Fig. 24). These changes indicate that pyoverdine becomes the preferred route of iron acquisition. In line with this hypothesis, we observed that the down-scaling of the plants’ own iron-uptake systems goes along with biomass increases for both peanut and maize plants (Fig. 5 and Supplementary Figs. 22-24), suggesting that both plants benefit and save resources by taking up iron chelated to pyoverdines. This form of nutrient cycling mediated by plant-microbiome interactions is destroyed in sterilized soil, resulting in a decreased SPAD value in intercropping peanut, even lower than in monocropping peanut (Figs. 2d and 5b). However, intercropping can still increase the active iron in young leaves of peanut (Figs. 2d and 5b), suggesting that DMA secreted by maize can directly be utilized by intercropping peanut in sterilized soil. Therefore, our findings indicate that multiple iron sources are tapped during intercropping, including siderophores from bacteria, DMA from maize and possibly also flavins or coumarins from peanut root exudates, to mitigate the plants’ iron-deficiency. However, due to the large differences in iron-chelating affinity between pyoverdine and root exudates , the possibility is that plants balance the iron-uptake strategies between themselves systems and pyoverdinebased ones in intercropping, but future research is needed to test this hypothesis.
While we primarily focused on how iron nutrition is improved in peanut when intercropped with maize, we also detected benefits for maize in a microbiota-dependent manner, suggesting that intercropping benefits both plant species. Maize and peanut could engage in a ‘giving-receiving feedback’ interaction through the convergence of their microbiomes in intercropping. Peanut plants are legumes and host nitrogen-fixing bacteria in their root nodules , such that peanut could improve nitrogen acquisition of maize . We indeed found evidence for this hypothesis. Specifically, we detected an enrichment of members of the Allorhizobium-Neorhizobium-PararhizobiumRhizobium genus and Devosia genus from maize to peanut (Fig. 3b). Increased abundance of nodule-forming Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium and nonrhizobial nodule-inducing Devosia bacteria could increase nodule activity and fixation in peanut, which in return could enhance nitrogen acquisition by nearby maize through reduced competition for soil and fertilizer nitrogen between the two plant species . Furthermore, we found Azotobacter, a typical plant beneficial non-symbiotic nitrogen-fixing bacterial genus, to be crossenriched from peanut to maize, possibly also contributing to improved nitrogen nutrition in maize. Our results are in line with previous findings on microbiota-dependent improvement of nitrogen nutrition . Overall, legume/gramineae combinations are promising candidates for intercropping and are therefore the subject of intensive research in soil science and plant nutrition.
Bioavailability of iron is particularly low in alkaline calcareous soils. That is the main reason why we focused on bacterial members of the rhizosphere microbiome, because bacteria are known for producing a diversity of siderophores, with some of them (e.g., pyoverdine) having a high affinity for iron . However, important to note is that other factors might also play a role in determining iron availability and benefits of intercropping. For instance, soil nutrients like and (the main forms of nitrogen in the rhizosphere) can impact pH and therefore iron availability. Long-term application of decreases rhizosphere pH , while increases rhizosphere . However, in our intercropping system, the pH remained stable across nutrient stages (Supplementary Fig. 6b), implying that the nitrogen cycle may
have no direct effect on iron nutrition improvement in our system. The reason may be that calcareous soils have strong pH buffering capacity . Moreover, soil properties, like , or organic carbon, can potentially affect microbial community structure and function, meaning that beneficial taxa like Pseudomonas might not be equally abundant in all soils. Finally, while we focused on bacteria other members of the microbiota might also be involved in intercropping benefits. Top candidates are the nitrogen-fixing rhizobium (as found in our study for maize) and arbuscular mycorrhizal fungi. Understanding the effects of such multi-kingdom plant-bacteria-fungi interactions across soils with different properties and geographical scales for intercropping will be the challenge of future work.
Our results show that intercropping in natural soils is sufficient to improve iron nutrition in calcareous soils (Supplementary Fig. 23). However, intercropping might not always be possible. For those situations, the insights from our study provide a simple practicable treatment, which consists of the addition of Pseudomonas sp. 1502IPR01 or its pyoverdine to the rhizosphere. Our treatment has several advantages over the use of traditional iron fertilizers such as synthetic EDTA-Fe with low efficiency and high environmental risk in soil . These issues do not apply to the pyoverdine treatment because it directly solubilizes iron from natural stocks in soil, pyoverdine itself is biodegradable , and . extremorientalis (closest relative of Pseudomonas sp. 1502IPR-01) is classified as a harmless biosafety risk 1 organism that is unlikely to cause disease in humans . Hence, Pseudomonas sp. 1502IPR-01 and its pyoverdine can be considered ecosystem-friendly biofertilizers. Additionally, pyoverdines can also enhance copper bioavailability and thus have the capacity to improve plant nutrition for other microelements . Given its dual role in microelements (especially iron) fertilization and phytopathogen control , pyoverdine could simultaneously reduce the use of synthetic microelements-fertilizers and pesticides, which could present a critical step forward toward sustainable agriculture, improved food security and environmental conservation.
In summary, we elucidated a mechanism of how intercropping benefits plants through rhizosphere microbiome convergence. Our study highlights that the benefit of intercropping from belowground facilitation leads to a gained function for peanut through a crossenrichment of rhizosphere microbiome members obtained from maize (Fig. 7). Our findings offer applied opportunities for a probiotic treatment to improve plant health and crop yield in situations where intercropping is not possible. While we have identified a probiotic strain and its secreted beneficial compound, there is great potential to improve the process through bioengineering. Our mechanistic insights on iron nutrition improvement offer perspectives and should trigger interest in promoting other intercropping systems with other plant combinations to increase both yield and sustainability.
Methods
NE and LER calculation
To quantify the benefits of peanut/maize intercropping, we calculated the land equivalency ratio (LER) and the net effect (NE) of this intercropping system for various field sites and years. LER > 1 indicates that intercropping has an advantage over monocropping in land-use efficiency. NE>0 indicates that intercropping has more absolute yield than monocropping .
The NE is defined as the difference between the observed yield and the expected yield.
where and are observed yields of species 1 and 2 in the intercropping, and and are expected yields of the two species, calculated as the product of the monocropping yield and the
land share .
where and are yields per unit area of species 1 and 2 in monocropping, and and are the land shares of species 1 and 2 in intercropping. The land share was calculated based on the densities of a species in the intercropping and in the monocropping or on the basis of the number of rows or plant arrangements .
The LER is defined as the sum of partial LERs (relative yields) per species (pLER1 and pLER2):
where and are the yields (per unit of the total area of the intercropping) of species 1 and 2 in intercropping, and and are the yields of species 1 and 2 in monocropping (same as above) .
Experimental design for peanut/maize intercropping under pot conditions
We conducted intercropping and monocropping experiments in pots. A typical iron-deficient calcareous sandy soil was collected from 0 to -depth in Lihua village, Daxing district of Beijing, China . Soil properties before fertilizer application were determined according to protocols from the previous literature and listed in Supplementary Table 6. Each pot contained 8 kg of soil amended with basal fertilizers [composition ( soil): N 100 (as ), P 150 (as ), K 100 (as KCl ), Mg 50 (as ) and Zn 5 (as )].
Peanut (Arachis hypogaea L. cv. Luhua14) and maize (Zea mays L. cv. Zhengdan958) were used. Three cropping treatments were set up. Six peanut plants or three maize plants were grown in a pot for the monocropping treatments (monocropping peanut and monocropping maize). Three peanut plants with three maize plants were grown in a pot for the intercropping treatments. The plants were grown under natural light conditions with air temperature, light intensity and relative humidity. During the experiment, soil water content was kept at approximately of the field capacity. Plant samples of peanut were harvested to assess the nutrition status at , and 73 days post sowing (dps). At the same time, pots were destroyed to acquire complete soil. Bulk soil was removed by gently shaking. The roots of intercropping maize and intercropping peanut were carefully separated from each other when shaking. Rhizosphere soil was collected for microbiome and soil properties analysis by removing the soil closely adhered to the roots using a sterilized fine brush (Supplementary Fig. 2). The peanut plants and rhizosphere soils from the same pots were mixed into one replicate. Each treatment had four replicates at each time point.
Design for soil sterilization experiment in pot conditions
To assess the role of the rhizosphere microbiome in intercropping peanut iron nutrition improvement, monocropping peanut and peanut/maize intercropping were grown in both normal and sterilized soil. Soil was prepared as described above. For soil sterilization treatment, the soil was -irradiated with a maximum dose of 20 kilograys ( 24 h , , Beijing Atomic High-tech Jinhui Radiation Technology Application Co. Ltd.). Plant species, growth conditions, and crop management were the same as in the experimental design for peanut/maize intercropping in pot conditions. At 73 dps , plant samples of peanut were harvested to assess the iron nutrition status, and rhizosphere soil was collected to measure available iron concentration. The peanut
plants and rhizosphere soil from the same pot were mixed and treated as one replicate. Each treatment had three or four replicates.
Measuring iron-related indicators in plants and soil
SPAD value. Chlorophyll levels in fresh young leaves (top fully emerged leaves) from peanut was measured using a chlorophyll meter (Konica-Minolta, Osaka, Japan). Each leaf was measured in four replications and averaged while avoiding the main leaf veins.
Ferric-chelate reductase (FCR) activity of peanut plants. According to previous literature , the fresh peanut plants were washed and then the roots were immersed in saturated solution for 30 min . After washing, peanut plants were transferred to a 100 Ml reagent consisting of – bipyridine and 0.1 Mm Fe (III)-EDTA, for a incubation, protected from light. The absorbance of the solution at 523 nm was measured. Ferric-chelate reductase activity was calculated with the formula (5) below and water was used as a control:
where is the absorbance of the solution under 523 nm , FW is the fresh weight of the root, and 8650 is the molar absorptivity of – bipyridine ( ). 100 is the volume of plant incubation solution.
Deoxymugineic acid (DMA) secretion capacity of maize plants. DMA is only phytosiderphore secreted by maize, so we used phytosiderphore secretion-determination method to determine the DMA secretion capacity. The fresh maize plants were washed and then the roots were immersed in saturated solution for 30 min . After washing, maize plants were transformed to 200 mL water for a incubation, protected from light. Before incubation, micropur (Katadyn Products Inc. Wallisellen, Switzerland) was added to prevent microbial degradation of phytosiderphores. 10 mL aboveincubated solution was added 0.5 mL of and shaken for 15 min , then 1 mL sodium acetate buffer ( pH 7.0 ) was added and the solution was shaken for a further 10 min . To reduce Fe (III) to Fe (II), the solutions were filtered into 0.25 ml of 6 M HCl and then hydroxylamine hydrochloride was added. The solution was incubated at for 30 min . Then, the solution is mixed with ferrozine and 1 mL 2.0 M sodium acetate buffer ( pH 4.7 ). After incubation, the absorbance of the solution at 562 nm was measured. DMA secretion activity was calculated with the formula (6) below by using water as a control:
where is the absorbance of the solution under 562 nm . FW is the fresh weight of the root. is the molar absorptivity ( ). 200 is the volume of plant incubation solution. 1.35 is the volume ratio of the final and original reaction solution.
Active iron in young leaves. Fresh young leaves from peanut were washed with distilled water and collected to assess HCl -extractable iron (‘active iron’) according to the previous literature of young leaves was taken and mixed with 10 mL of 1 M hydrochloric acid. The supernatant was shaken for 5 h at 150 r.p.m. and then filtered with filter paper. The iron concentration was measured by inductively coupled plasma-optical emission spectrometry (ICP-OES) using a 7300DV system (Perkin Elmer, Waltham, USA). Active iron in young leaves was calculated with the formula (7) below and deionized water was used as
a control:
where and CONCN are the Fe concentration ( ) of filtrate of sample and control, respectively, and FW is the fresh weight of the young leaves. Multiplication with 10 is applied to account for the volume of hydrochloric acid.
Available iron in the rhizosphere
10 g of air-dried rhizoplane soil was weighed and added to 20 mL of diethylenetriaminepentaacetic acid (DTPA) extracting agent, containing 0.005 M DTPA, and 0.1 M triethanolamine. Then, the mixture was shaken for 2 h at . at . Subsequently, the solution was filtered and the iron concentration in the filtrate was measured by ICP-OES. Available iron in the rhizosphere was calculated with the formula (8) below and deionized water was used as a control:
Available iron in the rhizosphere weight
where and are Fe concentration ( ) of the soil sample and control, weight is the weight of the soil sample. Multiplication with 20 is applied to account for the volume of DTPAextracting agents.
Soil DNA extraction, 16S rRNA amplicon sequencing
16 S rRNA amplicon sequencing was performed to examine the impact of intercropping on rhizosphere microbiome compositions. As with any microbiome sequencing study, it is important to consider potential contamination sources. We can identify two such potential sources in our rhizosphere microbiome sample collection scheme, which are (a) incomplete root separation and (b) soil mixing during plant harvest. For (a), it could be that peanut and maize roots cannot fully reliably be distinguished and separated when collecting them. We can refute this possibility because the roots of peanuts and maize can be clearly distinguished based on their color: roots of maize are white, while roots of peanut are dark brown (Supplementary Fig. 25). This distinct difference allowed us the completely and reliably separate the roots of the two species. For (b), there is the possibility that loose soil particles originally associated with the root of one of the species are transferred to the soil collected from the other species during the harvesting process. To prevent this source of contamination, we first vigorously shook the roots of the collected plant to get rid of all loose soil particles that are typically associated with the sandy soil. Subsequently, we only collected the rhizosphere soil that was tightly attached around the peanut or maize roots. These two points ensure to reduce the contamination risk to a minimum and that the convergence we observed stems from the biological interactions between the fine roots of the two plant species. DNA was isolated from 500 mg of soil sample using the FastDNA Spin Kit for Soil (MP Biomedicals, Solon, USA) according to the manufacturer’s instructions. The hypervariable region V3-V4 of the bacterial 16S rRNA gene was amplified with primer 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGAC-TACHVGGGTWTCTAAT-3′) by an ABI GeneAmp PCR thermocycler (Applied Biosystems, Foster City, USA). PCR conditions were as follows: 3 min at , then 27 cycles of each 30 s at at and 30 s at . After these cycles, there was a 10 -min elongation time at . Purified amplicons were pooled in equimolar and paired-end sequenced on an Illumina MiSeq PE300 platform (Illumina, San Diego, USA) according to the standard protocols by Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, China).
The upstream analysis progress of the amplicon sequence was processed using QIIME2 v2021.8 . Using the DADA2 workflow in
QIIME2, barcodes, PCR primers and low-quality sequences were trimmed, and the remaining reads were subsequently denoised and merged to generate the amplicon sequence variants (ASV) feature table. A taxonomy native Bayes classifier trained on the V3-V4 region ( 338 F and 806R) of the reference 16 S sequence (SILVA release 138 ) was used to assign taxonomic identities to the representative sequences. Only ASV that were at least annotated to the phylum level were retained. Features identified as chloroplast or mitochondrial were removed. Features with a total abundance (summed across all samples) of less than ten or present in less than six samples ( samples) were filtered. We normalized the sequencing depth to 17,063 counts per sample using scaling with ranked subsampling (SRS) , preserving the original community structure by minimizing subsampling errors. Representative sequences were filtered based on the filtered ASV table, then used to generate an approximately-maximum-likelihood phylogenetic tree using FastTree 2.1.8 in QIIME2 with default parameters.
PCoA analysis and Bray-Curtis distance calculation were performed by R ‘phyloseq’ package (version 1.34.0) based on the relative abundance of ASV. Differential microbiota between groups was identified by linear discriminant analysis (LDA) and LDA effect size (LEfSe) with an LDA threshold of 3 and -value threshold of 0.05. LefSe analysis was performed on the Huttenhower lab Galaxy server (http:// huttenhower.sph.harvard.edu/galaxy). The format conversion from ASV table to table for Lefse was performed by R ‘amplicon’ package (version 1.14.2). The correlation between rhizobacteria genus biomark’s relative abundance and soil plant iron nutrition was also based on relative abundance and calculated using non-parametric Spearman’s rank correlation (presented as Spearman rho) using R ‘phyloseq’ package (version 2.2.5).
Isolation of siderophore-secreting rhizobacteria
We isolated siderophore-secreting-rhizobacteria from the rhizosphere of intercropping peanut to assess the role they play in intercropping peanut iron nutrition improvement. The isolation method was modified according to protocols from the previous literature (Fig. 4a) and details are described below. Briefly, 1 g of the soil sample from intercropping peanut rhizosphere was moderately agitated in 20 mL of sterile solution for 10 min first. Then the microbial suspension (serially diluted) was spread on agar plates with chrome azurol S (CAS) indicator. CAS indicator contained 1 mM CAS, hexadecyltrimethylammonium bromide (HDTMA). Other components of the CAS agar medium consisted of ( ): sucrose, 2000; casein acid hydrolysate, , 241; agar, 1800 and phosphate buffer solution ). The pH of the medium was adjusted to 7.0. The ternary complex CAS-Fe(III)-HTDMA forms a blue dye. Siderophores remove Fe (III) from the dye by chelating Fe (III), which induces a color change from blue to orange (in agar plate) or red (in solution) . The plates were incubated at for 48 h in the dark. The microbial colonies with or without an orange halo were picked. The isolates were then re-streaked on LB plates for colony purification. The final collection consisted of 324 siderophoresecreting rhizobacteria isolates. All purified isolates were cultured in 1 mL liquid LB medium at with shaking (rotary shaker at 180 r.p.m.) before freezing and storing at in glycerol. Requests for strains should be addressed to zuoym@cau.edu.cn.
Measuring the siderophore-secreting capacity of rhizobacteria
To quantify the siderophore-secreting capacity of each rhizobacterium isolate under iron-limited conditions, the rhizobacteria isolates were revived by transferring volumes of their freezer stocks into 5 mL liquid LB medium and cultured 18 h at with shaking (rotary shaker at 180 r.p.m.). Then, the volumes of revived cultures were transferred into 5 mL liquid standard succinic medium (SSM) without
added iron ( succinic acid, ). After incubation at , the bacterial cultures were diluted to . Then, cellfree supernatant from diluted bacterial culture was harvested by centrifugation ( at ) and filtration (using a filter). Siderophore concentration in the supernatant was assayed using a modified version of the universal chemical CAS assay. Briefly, CAS assay solution was prepared as follows: a 21.9 mg quantity of HDTMA was dissolved in 25 mL water while constantly stirring over low heat. 1.5 mL of (in 10 mM HCl ) was mixed with 7.5 mL of 2 mM CAS. This solution was slowly added to the HDTMA solution while stirring, and the mixture volume was made up to 100 mL with deionized water. Cell-free supernatant or deionized water as a control reference was mixed with an equal volume of CAS assay solution in a 96 -well plate and incubated for four at room temperature (Supplementary Fig. 8). Siderophores removed the Fe(III) from pigmented Fe (III)-CAS-HDTMA complex, inducing a color change from blue to red in the CAS medium solution, lowering the measurements. The of the cell-free supernatants (A) and deionized water controls (Ar) was then measured using a plate reader (SpectraMax M5, Molecular Devices, Sunnyvale, USA). Siderophoresecreting capacity can be quantified using the following formula: . This formula quantifies the relative reduction in the measurement induced by iron-chelating compounds that retrieve Fe (III) from the pigmented Fe (III)-CAS-HDTMA complex. The Su-value exhibits exponential instead of linear changes when the siderophore is excessive ( ). Thus, the supernatant for the CAS assay reaction solution should be gradient diluted by 1/2 CAS assay solution to ensure . Desferrioxamine B (DFOB), a purchasable siderophore, was used to establish a calibration curve to estimate the concentration of siderophores in the supernatants of the rhizobacteria. Serial dilutions ( , and ) of the DFOB standard were mixed with CAS assay solution and incubated for four at room temperature before determination. The Su-value of DFOB was 0.71, and the calibration curve equation was: siderophore equivalent ( ). Results are expressed as equivalents of DFOB. To calculate the iron-binding capacity of bacterial siderophores, one mmol of DFOB is considered to bind one mmol Fe(III).
Tree construction and phylogenetic analysis of rhizobacterial siderophore-secreting isolates
We sequenced the 16S rRNA genes of the siderophore-secreting rhizobacterial isolates to confirm their genus-group and evolutionary relationships. According to the manufacturer’s protocol, the genomic DNA was extracted using Wizard Genomic DNA Purification Kit (Promega, Charbonnières, France). Purified genomic DNA was quantified by a TBS-380 fluorometer (Turner BioSystems Inc., Sunnyvale, USA). High-quality DNA ( ) was used to do further research. The 16 S rRNA gene was amplified using 27 F ( -AGAGTTTGATCMTGGCTCAG-3′) and 1492 R ( -TACGGY-TACCTTGTTACGACTT-3′) primers by the polymerase chain reaction (PCR). The PCR reactions ( ) contained one bacterial DNA, master mix, one each of the forward and reverse primers, and deionized water. The PCR was run as follows: initial: denaturation at for 5 min , 30 cycles of denaturation at for 30 s , annealing at for 30 s , extension at for 1 min 30 s , and the final extension at for 10 min . The PCR products were sequenced by Beijing Genomics Institution Co., Ltd (Beijing, China).
The 16S rRNA gene sequences were trimmed at both ends to obtain maximum overlap using the software DNAStar (version 7.1.0, DNASTAR Inc., Madison, USA). The 16 S rRNA gene sequences and homologous sequence similarity were identified using the NCBI database (https://www.ncbi.nlm.nih.gov/). We constructed an
approximately-maximum-likelihood phylogenetic tree using FastTree 2.1.8 in QIIME2 with default parameters. Trees were visualized using ggtree v2.4.1 in R v4.2.2.
Total genomic DNA extraction and sequencing of rhizobacteria isolate
The genome of Pseudomonas 1502IPR-01 isolate, which had the highest siderophore-secreting capacity in all rhizobacteria isolates, was sequenced using a combination of PacBio RS II Single Molecule Real Time (SMRT) and Illumina sequencing platforms. The Illumina data were used to evaluate the complexity of the genome.
For Illumina sequencing, at least genomic DNA was used in sequencing library construction. DNA samples were sheared into 400-500 bp fragments using a M220 Focused Acoustic Shearer (Covaris, Woburn, USA) following the manufacturer’s protocol. Illumina sequencing libraries were prepared from the sheared fragments using the NEXTflex Rapid DNA-Seq Kit (Bioo Scientific, Austin, USA). Briefly speaking, 5′ prime ends were first end-repaired and phosphorylated. Next, the 3 ‘ ends were A-tailed and ligated to sequencing adapters. The third step was to enrich the adapters-ligated products using PCR. The prepared libraries were then used for paired-end Illumina sequencing ( ) on an Illumina HiSeq X Ten machine (Illumina, San Diego, USA).
For Pacific Biosciences sequencing, an aliquot of DNA was spun in a Covaris g-TUBE (Covaris, Woburn, USA) at for 60 s using an Eppendorf 5424 centrifuge (Eppendorf, NY, USA). DNA fragments were then purified, end-repaired, and ligated with SMRTbell sequencing adapters following the manufacturer’s recommendations (Pacific Biosciences, Menlo Park, USA). Following the manufacturer’s recommendations, the resulting sequencing library was purified three times using volumes of Agencourt AMPure XP beads (Beckman Coulter Genomics, Danvers, USA). Next, a-10 kb insert library was prepared and sequenced on one SMRT cell using standard methods.
Genome assembly
The complete genome sequence of Pseudomonas sp. 1502IPR-01 was assembled using both the PacBio and Illumina reads. The original image data was transferred into sequence data via base calling, defined as raw data, and saved as a FASTQ file. Those FASTQ files contain read sequences, and quality information is included. A statistic of quality information was applied for quality trimming, by which the low-quality data can be removed to form clean data. The reads were then assembled into a contig using a hierarchical genome assembly process (HGAP) and canu, a software that performs scalable and accurate longread assembly via adaptive k -mer weighting and repeat separation. The last circular step was checked and finished manually, generating a complete genome with seamless chromosomes and plasmids. Finally, error correction of the PacBio assembly results was performed using the Illumina reads using Pilon. All of the above analyses were performed using I-Sanger Cloud Platform (www.i-sanger.com) from Shanghai Majorbio.
Phylogenetic tree construction of the 1502IPR-01 and representatives of Pseudomonas spp
To confirm the exact evolutionary relationships of 1502IPR-01 and representatives of Pseudomonas spp., a multi-locus sequence analysisbased phylogenetic tree was generated using a previously established method . Specifically, TBLASTN+ (v. 2.10.0) was used to identify and extract sequences for 100 housekeeping genes used in Davis et al. . All 100 housekeeping genes of 1502IPR-01 and 23 completely sequenced Pseudomonas spp. were aligned using MAFFT (v.7.427). Phylogenetic tree construction was done using IQ-TREE2. Trees were visualized using ggtree v2.4.1 in R v4.2.2. Information of used sequenced Pseudomonas spp. strains were listed in Supplementary Table 7.
Pyoverdine biosynthesis and transport gene cluster analysis
The pyoverdine biosynthesis and transport genes locus in 1502IPR-01 genome were identified using TBLASTN+ (v. 2.10.0) with the protein sequences from Pseudomonas aeruginosa PAO1 used as queries against the predicted protein sequences from 1502IPR-01 genome as the databases. Sequence comparisons were conducted using SmithWaterman algorithm by SnapGene software (version 6.0.2, GSL Biotech LLC, Boston, USA) to obtain DNA sequence identity. Visualization was performed by Adobe Illustrator CC, 2020 (Adobe, San Jose, USA).
Pyoverdine production and purification
For pyoverdine production, Pseudomonas sp. 1502IPR-01 was cultured in an above-mentioned SSM medium. After culturing at at 180 r.p.m for 48 h , the culture media ( 10 L ) was isolated from the cell materials by centrifugation at for 15 min and filtration with filter paper. The crude culture extracts were adjusted to pH 5.5 by the addition of HCl and adsorbed by XAD-4, a polymeric adsorbent resin. After removal of supernatant, the resin was then filtered, and the pyoverdine-containing fraction was subsequently eluted with 1:1 ( ). The elute was evaporated to dryness in a vacuum. Then the residues were dissolved in and subjected to the Shimadzu semi-preparative HPLC (Shimadzu, Tokyo, Japan) at with a flow rate of . The injection volume was . Samples were chromatographed using a solvent system of trifluoroacetic acid (TFA) in (solvent A) and MeCN-MeOH 4:1 ( , solvent B ). A linear binary gradient , -was used. The eluate was monitored at 380 nm . The faction containing pyoverdine was collected at and then adjusted to pH 7.0 with ammonium hydroxide before evaporation. The salt in the fraction generated during the neutralization of TFA was removed by a Spectra/ Por Dialysis membrane with a molecular weight cut-off (MWCO): 500-1000 Da (Spectrum Lab., Rancho Dominguez, USA). Then, the purity of isolated pyoverdine was confirmed by HPLC.
Structural identification of pyoverdine
Mass spectrometry. LC-HRMS spectra were obtained on an Agilent 6530 Q-TOF mass spectrometer coupled to an Agilent 1260 HPLC (Agilent Technologies GmbH, Waldbronn, Germany); Thermo Orbitrap MS (Orbitrap Fusion Lumos, Thermo Fisher Scientific, San Jose, USA); ESI-QQQ-MSMS (Xevo TQ-S micro, Waters, Milford, USA).
NMR data were obtained using (operating at 500.13 MHz for and 125.75 MHz for ) (Bruker Biospin GmbH, Karlsruhe, Germany). All spectra were measured at 303 K . The residual solvent signals were used for referencing spectra in the and dimensions.
UV/Vis. The UV spectra were recorded by a SPD-M20A Shimadzu photodiode array detector (PDA, detection 190-800 nm) (Shimadzu, Kyoto, Japan) connected to a Shimadzu HPLC system (LC-20AR, Shimadzu, Japan) during semi-preparative HPLC separations in MeCN- containing 0.1% TFA in .
Semi-preparative HPLC. Semi-preparative HPLC separations were conducted on a Shimadzu HPLC system (LC-20AR, Shimadzu, Japan) with an Atlantis Prep OBD T3 column ( , Waters, USA).
Chemicals. Ultrapure water was acquired from Watsons Ltd. (Hong Kong, China). Acetonitrile (HPLC grade) and TFA (HPLC grade) used for UHPLC/Q-TOF-MS were purchased from Merck (Darmstadt, Germany) and Roe Scientific Inc. (Newark, USA), respectively. Sodium carbonate (HPLC grade) was purchased from Sigma-Aldrich (St. Louis, USA). The dialysis membrane was purchased from Spectrum Laboratories, Inc. (Rancho Dominquez, USA).
Pyoverdine (Supplementary Fig. 11a), obtained as a yellowishgreen crystalline powder, showed the typical UV/Vis spectra of pyoverdines: 387 nm at pH 7.0 and a split band of 365 and 407 nm at pH 3.0 and 10.0. The molecular formula of was determined by the pseudo-molecular ion peak at (calculated for , 1393.5872) in positive HR-ESI-MS (Supplementary Fig. 11b). Total hydrolysis of pyoverdine indicated a peptide chain containing the three types of amino acids: serine (Ser), lysine (Lys) and 5-N-formyl-5-N-hydroxyornithine (FoOHOrn) (Supplementary Fig. 12).
It has been well documented that mass spectrometry is a powerful method for structural elucidation of pyoverdines . Herein, we obtained the following characteristic fragment ions of pyoverdine by using an orbitrap higher-energy collisional dissociation (HCD) approach with varying collision energies. An ion at 204.08, the most characteristic ion for the pyoverdine chromophore (A1′-2, Supplementary Fig. 11c), was clearly recognized in the orbitrap MS (Supplementary Fig. 11d). Nearby, another abundant ion at , corresponding to the chromophore connected with a carboxylic amide side chain (A1′-1) (Supplementary Fig. 11c, d), could be explained as the fragment ion of the opening of the tetrahydropyrimidine ring of the chromophore by retro-Diels-Alder (RDA) reaction. Then, the side chain ketoglutaric acid (kgl) of pyoverdine was evidenced by the loss of from the A1 ( ) and B1 ( ) ions yielding the two corresponding and ions (Supplementary Fig. 11e) . Hence, the structure-fragmentation pathways from B1 to A1′-2 were proposed as shown in Supplementary Fig. 11c. Furthermore, the series of B-ions from B8 ( ) to B1 ( ) allowed to determine of the amino acid sequence of the peptide chain as Ser-Ser-FoOHOrn-Ser-Ser-(Lys-FoOHOrn-Lys-Ser) (Supplementary Figs. 12-14). The series of B’-ions (lossing of from the kgl chain of the B-ions, Supplementary Figs. 12-14) form B8′ ( ) to corroborated the peptide sequence. Taken together, the primary structure of pyoverdine was deduced as kgl-chr-Ser-Ser-FoOHOrn-Ser-Ser-(Lys-FoOHOrn-Lys-Ser), which is identical to the pyoverdine from Pseudomonas spp. CFML 95-275 .
The structure of pyoverdine deduced from MS data was further confirmed by the NMR data. On the basis of a combination of and C-NMR) and 2D (COSY, NOESY, HSQC, and HMBC) NMR experiments (Supplementary Figs. 15-20), the – and -data of pyoverdine were assigned in Supplementary Tables 4 and 5. In addition, the key COSY and HMBC correlations are shown in Supplementary Fig. 21. The results showed that -NMR data of pyoverdine was almost identical to that of the reported pyoverdine . While the -NMR chemical shift values of pyoverdine shifted , which could be caused by the different acquisition temperatures or machines. The pyoverdine molecule consists of three parts: (i) a dihydroquinoline-type chromophore (chr): (1S)-5-amino-2,-3-dihydro-8,9-dihydroxy-1H-pyrimido-[1,2-a]quinolone-1-carboxylic acid, (ii) a peptide comprising 9 amino acids: Ser-Ser-FoOHOrn-Ser-Ser-(Lys-FoOHOrn-Lys-Ser) (Ser: serine; FoOHOrn: 5-N-formyl-5-N-hydroxyornithine; Lys: lysine), and (iii) a side chain ( -ketoglutaric acid, kgl ) bound to the nitrogen atom at position of the chromophore.
The capacity of pyoverdine of Pseudomonas sp. 1502IPR-01 to solubilize iron from
is the main insoluble form of iron in calcareous soil. Therefore, we determined the capacity of the pyoverdine of Pseudomonas sp. 1502IPR-01 to solubilize iron from insoluble stocks according to previous paper . For this purpose, equivalent of purified pyoverdine (calculated based on the DFOB calibration curve) was added into 2 mL 0.5 M sodium acetate solution. Then, all solutions were added to solution was prepared by neutralizing with NaOH to ) suspensions to a total volume of 5 mL . The above solutions were
incubated at for five hours, and the supernatant was collected after centrifugation at at for 10 min . Finally, the iron content in the supernatant was determined by ICP-OES using a 7300DV system (Perkin Elmer, Waltham, USA).
Functional verification of Pseudomonas sp. 1502IPR-01 and its siderophore
We identified the effect of Pseudomonas sp. 1502IPR-01 and its siderophore on peanut iron nutrition improvement in both greenhouse and field conditions. The normal and sterilized iron-deficient calcareous sandy soil was prepared as described above for pot experiment. Peanut (Arachis hypogaea L. cv. Luhua14) and maize (Zea mays L. cv. Zhengdan958) were used. Four treatment groups were set up: (1) monocropping peanut grown in normal soil; (2) monocropping peanut grown in sterilized soil; (3) peanut/maize intercropping grown in normal soil; (4) peanut/maize intercropping grown in sterilized soil. Six monocropping peanut plants were grown in a pot for the monocropping peanut treatment, and three peanut plants with three maize plants were grown in a pot for the intercropping treatment. Each pot contained 8 kg of soil. The rest of the greenhouse culture conditions remained identical as described above. At anthesis ( 50 dps ), pod bearing ( 64 dps ), and fruit-swelling stage ( 75 dps ), 600 mL of CFU 1502IPR-01 strain solution ( CFU plant ) or 600 mL of purified pyoverdine ( plant ) solution were irrigated into the rhizosphere as 1502IPR-01 and pyoverdine treatment. The 600 mL of water was irrigated into the rhizosphere as control. It is worth noting that we applied pyoverdine solution to treat the rhizosphere of peanut according to previous literature and our own research experience . The peanut plants in the same pot were mixed as a replicate. Each treatment had three or four replicates. At 84 dps , SPAD values and active iron concentration in young leaves were measured. Rhizosphere soil was collected to measure the available iron concentration.
The field experiment was conducted in Xichang village ( E, N), Fangshan district of Beijing, and Ligu village ( , N) in Puyang city, Henan province, China in 2020. In Beijing, the peanut cultivar was Luhua14. The average temperature from May to September was , and the annual precipitation was 603 mm . The type of soil in Beijing is calcic cambisol. The soil property was pH 7.9, total N , available P (Olsen-P) , available K ( ) , available Fe . In Henan, the peanut cultivar was Yuhua37. The average temperature from May to September was , and the annual precipitation was 604.1 mm . The type of soil in Henan is fluvo-aquic soil. The soil property was pH 7.7 , total , available P (Olsen- P ) , available K ( ) , available Fe compound fertilizer (N-P-K: 19-19-19) was applied before sowing, and field management was according to local practices. Four treatments were set up: (1) 150 mL water was irrigated in the rhizosphere, and 16 mL water was sprayed on young leaves for one peanut plant as a control treatment; (2) 1502IPR-01 strain solution was irrigated in rhizosphere, and 16 mL water was sprayed on young leaves for one peanut plant as 1502IPR-01 treatment; (3) purified pyoverdine ( plant ) was irrigated into rhizosphere, and 16 mL water was sprayed on young leaves for one peanut plant as pyoverdine treatment; (4) 150 mL water was irrigated in the rhizosphere and EDTA-FeNa solution was sprayed on young leaves for one peanut plant as EDTA-Fe treatment. All spraying solutions contained 0.1% Triton X-100 as surfactant. Each treatment consisted of six field plot replications in Beijing and eight field plot replications in Puyang. Each field plot replication consisted of 16 holes, and two peanut plants were grown in one hole. The plant spacing was 30 cm , and row spacing was 60 cm . At anthesis ( 53 dps in Beijing and Puyang), pod bearing ( 65 dps in Beijing and 68 dps in Puyang), and fruit-swelling ( 79 dps in
Beijing and 84 dps in Puyang) stage, the respective treatments were repeated. Two weeks after the third treatment, SPAD values and active iron concentration in young leaves were measured. Rhizosphere soil was collected to measure the available iron concentration. Then, peanut plants were harvested to measure yield.
Pot experiment with pyoverdine null mutant
To further strengthen the point that pyoverdine is produced in soil and involved in peanut iron nutrition, we conducted additional experiments with the laboratory strain . aeruginosa PAO1 and its defined isogenic pyoverdine null mutant PAO1 . PAO1 is a double knockout mutant of the primary siderophore pyoverdine (through the deletion of ) and the secondary siderophore pyochelin (through the deletion of and ) in P. aeruginosa PAO1. We used this double knockout mutant because pyochelin is upregulated when pyoverdine is knocked out and can compensate for the effect of pyoverdine. To assess the role of pyoverdine, we thus needed to use the double knockout mutant. The normal iron-deficient calcareous sandy soil was prepared as described above for the pot experiment with each pot containing 8 kg of soil. Six monocropping peanut plants (Arachis hypogaea L. cv. Luhua14) were grown in a pot for the monocropping peanut treatment. At anthesis ( 50 dps ), pod bearing ( 64 dps ), and fruit-swelling stage ( 75 dps ), 600 mL of ( CFU plant ), solutions of Pseudomonas sp. 1502IPR-01, P. aeruginosa PAO1 wildtype, and the siderophoredeficient mutant PAO1 were irrigated into the rhizoplane. 600 mL of water was irrigated into the rhizosphere as control. The rest of the greenhouse culture conditions remained identical to the ones described above. The peanut plants growing in the same pot were mixed and treated as one replicate. Each treatment had three or four replicates. At 84 dps , SPAD values and active iron concentration in young leaves were measured. Rhizosphere soil was collected to measure the available iron concentration.
Data analysis
Data sets were tested for normal distribution with the Shapiro-Wilk test and for heterogeneity of variance using Levene’s test. For data sets that met the conditions of normal distribution and homogeneity of variance, parametric Student’s -test and ANOVA with LSD as post-hoc test were used. For data sets that did not meet these conditions, the BoxCox algorithm was used for data transformation to meet the assumptions of parametric tests. If these conditions were still not met after transformation, non-parametric Wilcoxon tests, Kruskal-Wallis tests with Dunnett T3 test (for one-way designs) or Scheirer-Ray-Hare test (for two-way designs) were used. The BH algorithm was used to correct -values in multiple comparisons. All the above tests, as well as PERMANOVA and correlation analyses, were analyzed in R (version 4.2.2; https://www.rproject.org) using the ‘stats’ (version 4.2.2), ‘car’ (version 3.1-1), ‘forecast’ (version 8.2.0), ‘ggpubr’ (version 0.4.0), ‘stats’ (version 4.2.2), ‘agricolae’ (version 1.3-5), ‘ggpubr’ (version 0.4.0), ‘stats’ (version 4.2.2), ‘PMCMRplus’ (version 1.9.6), ‘rcompanion’ (version 2.4.21), ‘vegan’ (version 2.5-7), and ‘psych’ (version 2.2.5) packages. For data sets that did not meet normal distribution, correlations were calculated using non-parametric Spearman’s rank correlation (presented as Spearman rho). For data sets that met normal distribution, correlations were calculated using parametric Pearson correlation (presented as Pearson’s ). The heatmap was generated using ‘ComplexHeatmap’ (version 2.15.1) package in R (version 4.2.2) based on the relative abundance of the genus. Then, plots including conceptual diagrams were assembled and aesthetically perfected by Adobe Illustrator CC, 2020 (Adobe, San Jose, USA). All tests were two-sided. The China administrative border map used in the figures was downloaded from a public platform: resource and environment center data cloud platform (http://www.resdc.cn/).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Raw data of 16 S rRNA amplicon sequencing used in this study are deposited in the NCBI database under BioProject PRJNA788265. 16S rRNA gene sequences of 46 isolated rhizobacteria are deposited on GenBank with accession OL824679 – OL824724. The Pseudomonas sp. 1502IPR-01 reference strain genome sequence, determined for this study, is deposited at the NCBI database under BioProject PRJNA788188. The raw data generated in this study are available in the Figshare repository under accession code (https://doi.org/10.6084/m9.figshare. and also are provided in the Source Data file/Supplementary Data with this paper. Source data are provided with this paper.
Li, C. et al. Syndromes of production in intercropping impact yield gains. Nat. Plants 6, 653-660 (2020).
Brooker, R. W. et al. Facilitation and biodiversity-ecosystem function relationships in crop production systems and their role in sustainable farming. J. Ecol. 109, 2054-2067 (2021).
Zuo, Y. & Zhang, F. Iron and zinc biofortification strategies in dicot plants by intercropping with gramineous species. A review. Agron. Sustain. Dev. 29, 63-71 (2009).
National Bureau of Statistics of China. China Statistical Yearbook (in Chinese) (China Statistics Press, 2021).
Dai, J. et al. From leguminosae/gramineae intercropping systems to see benefits of intercropping on iron nutrition. Front. Plant Sci. 10, 605 (2019).
Xiong, H. et al. Molecular evidence for phytosiderophore-induced improvement of iron nutrition of peanut intercropped with maize in calcareous soil. Plant Cell Environ. 36, 1888-1902 (2013).
Curie, C. & Mari, S. New routes for plant iron mining. N. Phytol. 214, 521-525 (2017).
Riaz, N. & Guerinot, M. L. All together now: regulation of the iron deficiency response. J. Exp. Bot. 72, 2045-2055 (2021).
Kobayashi, T., Nozoye, T. & Nishizawa, N. K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 133, 11-20 (2019).
Suzuki, M. et al. Development of a mugineic acid family phytosiderophore analog as an iron fertilizer. Nat. Commun. 12, 1558 (2021).
Lurthy, T., Pivato, B., Lemanceau, P. & Mazurier, S. Importance of the rhizosphere microbiota in iron biofortification of plants. Front. Plant Sci. 12, 744445 (2021).
Zhang, J. et al. NRT1.1B is associated with root microbiota composition and nitrogen use in field-grown rice. Nat. Biotechnol. 37, 676-684 (2019).
Voges, M., Bai, Y., Schulze-Lefert, P. & Sattely, E. S. Plant-derived coumarins shape the composition of an Arabidopsis synthetic root microbiome. Proc. Natl Acad. Sci. USA 116, 12558-12565 (2019).
Cadot, S. et al. Specific and conserved patterns of microbiotastructuring by maize benzoxazinoids in the field. Microbiome 9, 103 (2021).
Harbort, C. J. et al. Root-secreted coumarins and the microbiota interact to improve iron nutrition in Arabidopsis. Cell Host Microbe 28, 825-837 (2020).
Rajniak, J. et al. Biosynthesis of redox-active metabolites in response to iron deficiency in plants. Nat. Chem. Biol. 14, 442-450 (2018).
Ghosh, S. K., Bera, T. & Chakrabarty, A. M. Microbial siderophore – A boon to agricultural sciences. Biol. Control 144, 104214 (2020).
Sorrenti, G., Masiello, C. A. & Toselli, M. Biochar interferes with kiwifruit Fe-nutrition in calcareous soil. Geoderma 272, 10-19 (2016).
Maitra, S. et al. Intercropping-a low input agricultural strategy for food and environmental security. Agronomy 11, 343 (2021).
Shirley, M., Avoscan, L., Bernaud, E., Vansuyt, G. & Lemanceau, P. Comparison of iron acquisition from Fe-pyoverdine by strategy I and strategy II plants. Botany 89, 731-735 (2011).
Vansuyt, G., Robin, A., Briat, J. F., Curie, C. & Lemanceau, P. Iron acquisition from Fe-pyoverdine by Arabidopsis thaliana. Mol. PlantMicrobe Interact. 20, 441-447 (2007).
Soares, E. V. Perspective on the biotechnological production of bacterial siderophores and their use. Appl. Microbiol. Biotechnol. 106, 3985-4004 (2022).
Gu, S. et al. Competition for iron drives phytopathogen control by natural rhizosphere microbiomes. Nat. Microbiol. 5, 1002-1010 (2020).
Zuo, Y. M., Zhang, F. S., Li, X. L. & Cao, Y. P. Studies on the improvement in iron nutrition of peanut by intercropping with maize on a calcareous soil. Plant Soil 220, 13-25 (2000).
Li, L. et al. Root distribution and interactions between intercropped species. Oecologia 147, 280-290 (2006).
Gao, Y. et al. Distribution of roots and root length density in a maize/ soybean strip intercropping system. Agric. Water Manag. 98, 199-212 (2010).
Li, Q. S. et al. Belowground interactions impact the soil bacterial community, soil fertility, and crop yield in maize/peanut intercropping systems. Int. J. Mol. Sci. 19, 622 (2018).
Figueiredo, A. R. T., Özkaya, Ö., Kümmerli, R. & Kramer, J. Siderophores drive invasion dynamics in bacterial communities through their dual role as public good versus public bad. Ecol. Lett. 25, 138-150 (2022).
Kümmerli, R. Iron acquisition strategies in pseudomonads: mechanisms, ecology, and evolution. Biometals 36, 777-797 (2023).
Ahmed, E. & Holmstrom, S. J. Siderophores in environmental research: roles and applications. Micro. Biotechnol. 7, 196-208 (2014).
Shen, H. et al. AhDMT1, a transporter, is involved in improving iron nutrition and fixation in nodules of peanut intercropped with maize in calcareous soils. Planta 239, 1065-1077 (2014).
Dong, Q. et al. Maize and peanut intercropping improves the nitrogen accumulation and yield per plant of maize by promoting the secretion of flavonoids and abundance of Bradyrhizobium in rhizosphere. Front. Plant Sci. 13, 957336 (2022).
Martínez-Hidalgo, P. & Hirsch, A. M. The nodule microbiome: fixing rhizobia do not live alone. Phytobiomes J. 1, 70-82 (2017).
Li, B. et al. Root exudates drive interspecific facilitation by enhancing nodulation and fixation. Proc. Natl Acad. Sci. USA 113, 6496-6501 (2016).
Hu, H. Y. et al. Nitrogen fixation and crop productivity enhancements co-driven by intercrop root exudates and key rhizosphere bacteria. J. Appl. Ecol. 58, 2243-2255 (2021).
Vora, S. M., Joshi, P., Belwalkar, M. & Archana, G. Root exudates influence chemotaxis and colonization of diverse plant growth promoting rhizobacteria in the pigeon pea – maize intercropping system. Rhizosphere 18, 100331 (2021).
Wang, Z. et al. Forms of nitrogen inputs regulate the intensity of soil acidification. Glob. Change Biol. 29, 4044-4055 (2023).
Weng, Z. H., Li, G., Sale, P. W. G. & Tang, C. X. Application of calcium nitrate with phosphorus promotes rhizosphere alkalization in acid subsoil. Eur. J. Soil Sci. 73, e13153 (2022).
Ferreira, C. M. H. et al. Calcareous soil interactions of the iron(III) chelates of DPH and Azotochelin and its application on amending
iron chlorosis in soybean (Glycine max). Sci. Total Environ. 647, 1586-1593 (2019).
Havlin, J. L., Beaton, J. D., Tisdale, S. L. & Nelson, W. L. Soil Fertility and Fertilizers: An Introduction to Nutrient Management 7th edn (Pearson College Div, 2004).
Federal Ministry of Labour and Social Affairs (BMAS). Technical Rules for Biological Agents: Classification of Prokaryotes (Bacteria and Archaea) into Risk Groups (TRBA 466)-2015 (2015).
Cornu, J. Y. et al. Contrasting effects of pyoverdine on the phytoextraction of Cu and Cd in a calcareous soil. Chemosphere 103, 212-219 (2014).
Verma, S., Chakdar, H., Kumar, M., Varma, A. & Saxena, A. K. Microorganisms as a sustainable alternative to traditional biofortification of iron and zinc: status and prospect to combat hidden hunger. J. Soil Sci. Plant Nutr. 21, 1700-1717 (2021).
Li, C. J., Zhu, X. P. & Zhang, F. S. Role of shoot in regulation of iron deficiency responses in cucumber and bean plants. J. Plant Nutr. 23, 1809-1818 (2000).
Reichman, S. M. & Parker, D. R. Critical evaluation of three indirect assays for quantifying phytosiderophores released by the roots of Poaceae. Eur. J. Soil Sci. 58, 844-853 (2007).
Bolyen, E. et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852-857 (2019).
Beule, L. & Karlovsky, P. Improved normalization of species count data in ecology by scaling with ranked subsampling (SRS): application to microbial communities. PeerJ 8, e9593 (2020).
Schwyn, B. & Neilands, J. B. Universal chemical-assay for the detection and determination of siderophores. Anal. Biochem. 160, 47-56 (1987).
Davis, E. W., Okrent, R. A., Manning, V. A. & Trippe, K. M. Unexpected distribution of the 4-formylaminooxyvinylglycine (FVG) biosynthetic pathway in Pseudomonas and beyond. PLoS ONE 16, eO247348 (2021).
Budzikiewicz, H., Schafer, M., Fernandez, D. U., Matthijs, S. & Cornelis, P. Characterization of the chromophores of pyoverdins and related siderophores by electrospray tandem mass spectrometry. Biometals 20, 135-144 (2007).
Sultana, R. et al. A pyoverdin from Pseudomonas sp. CFML 95-275. Z. Naturforsch. C. 55, 857-865 (2000).
Jin, C. W., Li, G. X., Yu, X. H. & Zheng, S. J. Plant Fe status affects the composition of siderophore-secreting microbes in the rhizosphere. Ann. Bot. 105, 835-841 (2010).
Wang, N. Q. et al. Microbiome convergence enables siderophore-secreting-rhizobacteria to improve iron nutrition and yield of peanut intercropped with maize. Figshare. https://doi.org/10.6084/m9. figshare. 24648306 (2023).
Wang, N. Q. et al. Microbiome convergence enables siderophore-secreting-rhizobacteria to improve iron nutrition and yield of peanut intercropped with maize. wnq13579/Peanut-maize-intercroppingmicrobiome. Zenodo. https://doi.org/10.5281/zenodo. 10212602 (2023).
Acknowledgements
This research was financially supported by the National Natural Science Foundation of China (grant no. 32372810 to Y.-M. Z., 32102469 to N.-Q. W., and 42325704 to Z. W.), the National Key Research and Development Program of China (grant no. 2022YFD1901500/2022YFD1901501,
2023YFD1700203 to Y.-M. Z. and 2021YFD1900100 to Z. W.), and the Swiss National Science Foundation SNSF (grant no. 31003A_182499 to R. K.). We thank Dr. Prof. Yang Bai from the Chinese Academy of Sciences, Dr. Prof. Chuihua Kong, and Dr. Prof. Ling Xu from China Agricultural University for valuable discussions and suggestions.
Author contributions
Y.-M.Z., Z. W., N.-Q. W. and T.-Q. W. designed the research. N.-Q. W. and T.-Q. W. performed and analyzed most of the experiments in the laboratory and field, and Q.-F. L. K.-G. W., Z.-C. D., Z.-G. C., W.-Q., J.-D., L.-N., and J.-Y. C. performed and analyzed most of the experiments in the greenhouse. Y. C. and M. W. provided lab conditions and intellectual input for identifying the siderophore structure and wrote the section “Structural identification of pyoverdine” in methods. R. K. and Z. W. provided intellectual input, and guidance to microbiome data analysis and helped to interpret data. M. W. provided some intellectual input for this manuscript, and guidance to microbiome data analysis and visualization. N.-Q. W., T.-Q. W., F.-S. Z., R. K., Z. W., and Y.-M. Z. wrote and revised the manuscript. All of the authors discussed the results and commented on the manuscript. Y.-M. Z. supervised the study.
Correspondence and requests for materials should be addressed to Ming Wang, Zhong Wei or Yuanmei Zuo.
Peer review information Nature Communications thanks Pierre Cornelis, and Eiko Kuramae for their contribution to the peer review of this work. A peer review file is available.
College of Resources and Environmental Sciences, State Key Laboratory of Nutrient Use and Management (SKL-NUM), National Academy of Agriculture Green Development, China Agricultural University, 100193 Beijing, China. Jiangsu Key Laboratory for the Research and Utilization of Plant Resources, Jiangsu Province Engineering Research Center of Eco-cultivation and High-value Utilization of Chinese Medicinal Materials, Institute of Botany, Jiangsu Province and Chinese Academy of Sciences, 210014 Nanjing, Jiangsu, China. Department of Plant Pathology, The Key Laboratory of Plant Immunity, Nanjing Agricultural University, Nanjing 210095, China. Jiangsu provincial key lab for solid organic waste utilization, Key lab of organic-based fertilizers of China, Jiangsu Collaborative Innovation Center for Solid Organic Wastes, Educational Ministry Engineering Center of Resource-saving fertilizers, Nanjing Agricultural University, Nanjing 210095, China. Department of Quantitative Biomedicine, University of Zurich, Zurich, Switzerland. These authors contributed equally: Nanqi Wang, Tianqi Wang, Yu Chen. e-mail: mwang@njau.edu.cn; weizhong@njau.edu.cn; zuoym@cau.edu.cn