تمنع الأحماض الجاليكية والجليكيرهيزين تلف الكبد الناتج عن الأزيثروميسين في الجرذان من خلال التخفيف من الإجهاد التأكسدي والالتهاب Gallic and glycyrrhetinic acids prevent azithromycin-induced liver damage in rats by mitigating oxidative stress and inflammation
تمنع الأحماض الجاليكية والجليكيرهيزين تلف الكبد الناتج عن الأزيثروميسين في الجرذان من خلال التخفيف من الإجهاد التأكسدي والالتهاب
مروة أحمد محمد عليأسماء إبراهيم مطوقعلاء الدين أحمد حمزةعمرو أمين و جيهان حسين هيبا
لقد أظهرت المنتجات الطبيعية مثل حمض الجاليك (GA)، وهو مركب فينولي، وحمض الجلسيرهيزين (GLA)، وهو تربتيني ثلاثي الحلقات، خصائص مضادة للأكسدة ومضادة للالتهابات وحامية للكبد. تهدف هذه الدراسة إلى التحقيق في التأثيرات الوقائية لـ GA و GLA وتركيبهما واستكشاف آلياتها الأساسية ضد تلف الكبد الحاد الناتج عن الأزيثروميسين (AZM) في الجرذان. تم استخدام سبع مجموعات من الجرذان الذكور من نوع ويستار، وهي مجموعة التحكم، GA، GLA، AZM، AZM + GA، AZM + GLA، و AZM + GA + GLA. عالجنا الجرذان لمدة 21 يومًا، حيث تم إعطاء GA و GLA فيأسبوع واحد قبل AZMمستويات السيروم من الأسبارتات أمينوترانسفيراز، الألانين أمينوترانسفيراز، والفوسفاتاز القلوي في الجرذان المعالجة بـ AZM والمُعالجة مسبقًا بـ GA وGLA والعلاج المشترك كانت أقل بكثير من تلك في مجموعة AZM. أظهرت المستويات المرتفعة من الجلوتاثيون، الكاتالاز، والسوبر أكسيد ديسموتاز في الجرذان الوقائية أن GA وGLA وتركيباتهما قللت بشكل ملحوظ من مستويات المالونديالديهايد المستحثة بواسطة AZM، وهو علامة على الإجهاد التأكسدي، مما حسن النظام المضاد للأكسدة في تلك المجموعات. علاوة على ذلك، قللت GA وGLA وتركيباتهما من السيتوكينات المؤيدة للالتهابات، مثل عامل نخر الورم-وعامل النسخ النووي كابا بيتا (NF-kB)، مما يعكس الالتهاب الكبدي. كما اكتشفنا أن AZM خفض من مستوى عامل النسخ المرتبط بالخلية الحمراء 2 (Nrf2)، وأن مستوياته الطبيعية تم استعادتها بعد العلاج بـ GA و GLA ومزيجهما، مما يشير إلى أن هناك مسار إشارة Nrf2 يساهم في الوقاية من تلف الكبد الناتج عن AZM. في الختام، حمت GA و GLA ومزيجهما الفئران من إصابة الكبد الناتجة عن AZM من خلال قدرتها المثبتة على خفض الإجهاد التأكسدي والالتهاب عن طريق منع خفض مستوى Nrf2 وزيادة مستوى NF-kB. بعد تحديد فعاليتها وسلامتها في البيئات السريرية في الدراسات المستقبلية، يمكن أن تصبح GA و GLA أدوية علاجية مفيدة للتخفيف من السمية الكبدية الناتجة عن AZM.
الكلمات الرئيسية: أزيثروميسين، حمض الغاليك، أحماض الجلسيرهيزين، تلف الكبد، Nrf2، NF-kB يمكن أن تؤدي إصابة الكبد الناتجة عن الأدوية (DILI)، على الرغم من كونها غير شائعة، إلى عواقب تهدد الحياة مثل ارتفاع إنزيمات الكبد، وفشل الكبد الحاد، وحتى الوفاة.الأزيثرومايسين (AZM)، وهو مضاد حيوي من فئة الماكروليد، يعد من بين العشرة الأوائل من الأدوية التي تسبب تلف الكبد الناتج عن الأدوية (DILI).. إنه يعالج بشكل فعال العدوى البولية التناسلية، والأمراض المعوية مثل التيفوئيد، والحالات التنفسية مثل الالتهاب الرئوي، مما يدل على نشاطه المضاد للبكتيريا واسع الطيف ضد كل من البكتيريا إيجابية الجرام وسلبية الجرام. علاوة على ذلك، أظهر AZM إمكانيات علاجية في إدارة الشفاء لمجموعة واسعة من الأمراض، بما في ذلك العدوى الفيروسية مثل COVID-19 4،5،6.
لقد أثبتت الأبحاث العلمية بلا شك وجود تأثيرين سلبيين معترف بهما لإدارة AZM: السمية الكبدية والسمية القلبية.. لي وآخرونأجروا دراسة شاملة عن السمية الكبدية في الحيوانات، وإليسون وبلاكويلتمت ملاحظته في البشر. أكدت الدراسات السريرية أن AZM يمكن أن يكون له آثار سلبية خطيرة على كل من البالغين والأطفال، مثل ركود الصفراء، واليرقان، واضطراب وظائف الكبد.لقد وثقت الأبحاث حالات التهاب الكبد الصفراوي الحاد الذي تطور خلال الأسابيع الثلاثة الأولى من بدء تناول دواء AZM.وفقًا لمارتينيز وآخرونيظهر تلف الكبد عادةً في غضون أسبوع إلى ثلاثة أسابيع بعد
بدء العلاج. بينما يسبب AZM غالبًا تلفًا خفيفًا في الكبد، هناك حالات يمكن أن تؤدي إلى تلف كبير في الكبد، مما يتطلب زراعة كبد وقد يؤدي إلى فشل كبدي مفاجئ.إن التعرف على هذه المخاطر المحتملة والحفاظ على موقف دقيق تجاه الكشف عن أي مؤشرات على تلف الكبد لدى المرضى الذين يتلقون علاج AZM هو أمر في غاية الأهمية. نحن لا نفهم تمامًا كيف يؤثر AZM على الأنسجة الكبدية، لكننا نفترض أن الإجهاد التأكسدي والالتهاب يلعبان دورًا كبيرًا.
حالياً، هناك خيارات علاجية ووقائية محدودة لأمراض الكبد، على الرغم من انتشارها العالمي ومساهمتها الكبيرة في معدل الوفيات في جميع أنحاء العالم. يدرك الناس على نطاق واسع أن الإجهاد التأكسدي والالتهاب هما عاملان رئيسيان في تطور أمراض الكبد.يولي الباحثون والمهنيون في مجال الرعاية الصحية اهتمامًا متزايدًا بالمركبات الطبيعية من البوليفينول لتقليل الآثار الضارة لمختلف الأدوية والملوثات البيئية.مؤخراً، حظيت هذه المركبات باهتمام كبير بفضل خصائصها الدوائية المتنوعة وإمكاناتها العلاجية.حمض الجاليك (GA)، مركب فينولي طبيعي يتمتع بخصائص مضادة للأكسدة قوية، يتميز بوزنه الجزيئي المنخفض نسبيًا ويعرف كيميائيًا باسم حمض 3،4،5-ثلاثي الهيدروكسي بنزويك.تمتلك GA عدة خصائص حيوية وكيميائية، مثل التأثيرات المضادة للالتهابات، والمضادة للبكتيريا، والمضادة للأكسدة، والحامية للقلب، والمضادة للفيروسات، والمضادة للسرطان.. هذا المركب الفينولي الطبيعي موجود في مجموعة متنوعة من المصادر النباتية بما في ذلك قشور التفاح، والجوز، والمانجو، والعنب، والشاي الأخضر. يظهر GA خصائص واقية للكبد بشكل ملحوظ.مظهراً فعاليته في مكافحة عوامل تلف الكبد المختلفة بما في ذلك الكربون رباعي الكلور (CCL4)نيتروسو ثنائي إيثيل أمين، قيادةبيروكسيد التيرت-بيوتيلبيسفينولالإيثانولثيوأسيتاميدوالكادميومتعمل الأنشطة البيولوجية لـ GA، وخاصة خصائصه المضادة للأكسدة، على حماية خلايا وأنسجة الإنسان من الأضرار الناتجة عن الإجهاد التأكسدي.أنواع الجذور الحلوة، المعروفة عادةً باسم العرقسوس، هي المصدر الرئيسي للجليكوسيد ثلاثي التربين الخماسي.جذر العرقسوس هو المصدر الرئيسي لحمض الجلسيرهيزيك (GLA)، وهو تريتربين بيولوجي نشط خماسي الحلقات.يُنتج العرقسوس نوعين من متماكبات حمض الجلسيرهيزيك:حمض الجلسيرهيزين وحمض 18a-جلسيرهيزينتظهر هذه المركبات مجموعة من الخصائص الصيدلانية بما في ذلك الأنشطة المضادة للفيروسات، والمضادة للالتهابات، والمضادة للتليف، والأنشطة المضادة للأكسدة.أظهرت الدراسات الحديثة أن GLA يمكن أن تحمي خلايا الكبد من الضرر الناتج عن سموم مختلفة مثل CCL4.2-أسيتيل أمينوفلورينسيكلوفوسفاميدميثوتريكساتوإيزوثيوسيانات ألفا-نافثيلتشير النتائج إلى أن GLA تمتلك خصائص واقية للكبد ويمكن أن تخفف من تلف الكبد الناجم عن مواد كيميائية ضارة متنوعة.
إيرانشاهى وآخرونأظهرت أن عامل النووية المرتبط بالحديد 2 (Nrf2) يلعب دورًا حاسمًا في حماية الكبد من السمية الناتجة عن الأدوية. كشفت دراستهم أن Nrf2 ينظم تعبير الجينات المعتمدة على عناصر الاستجابة لمضادات الأكسدة (ARE)، والتي تعتبر ضرورية للحفاظ على مرونة الخلايا ضد الإجهاد التأكسدي. يؤدي تنشيط Nrf2 إلى تحفيز نسخ الإنزيمات المضادة للأكسدة والواقية للخلايا، بما في ذلك SOD، الكاتالاز، هيم أوكسجيناز 1، NAD(P) H، كينون أوكسيدوريدوكتاز 1، وNAD(P).وبذلك تعزز آليات الدفاع في الخلية.
لقد أظهرت العديد من المنتجات الطبيعية خصائص واقية للكبد.في هذه الدراسة، ركزنا على فئتين رئيسيتين من المركبات النشطة بيولوجيًا: الأحماض الفينولية، مثل حمض الجاليك، والمركبات التريترينية، مثل حمض الجلوتاميك.من الجدير بالذكر أن تنشيط Nrf2 يبدو أنه آلية رئيسية وراء التأثيرات الواقية للكبد لهذه المركبات الطبيعية.لقد أظهرت المركبات المستمدة من العرقسوس مثل GLA، عند استخدامها بالاشتراك مع عوامل علاجية أخرى، أنها تعزز حماية الكبد ضد إصابة الكبد الناتجة عن الأدوية.وبناءً عليه، تهدف هذه الدراسة إلى تقييم التأثيرات الوقائية لـ AZM و GA و GLA وتركيباتها في التخفيف من السمية الكبدية الحادة في ذكور الجرذان ويستار. تشير النتائج الحالية إلى أن GA و GLA وإعطاؤهما معًا يحميان الكبد من خلال تعديل الإجهاد التأكسدي ومسارات الالتهاب.
المواد والأساليب
المواد الكيميائية
تم الحصول على حمض الثيوباربيتيوريك، والبيوروجالول، وحمض 18β1-غليسيريتينيك (رقم الكاتالوج G10105) وحمض الغاليك (رقم الكاتالوج G7384) من شركة سيغما-ألدريتش (سانت لويس، ميزوري، الولايات المتحدة الأمريكية). تم شراء AZM (Xithrone، 500 ملغ، 211515) من شركة أمون، الواقعة في مدينة العبور، القليوبية، مصر. وقد اختارت الاستفسارات أعلى درجة تحليلية لجميع المواد الأخرى.
الحيوانات
شملت الدراسة عينة من 42 من ذكور الجرذان الألبينو من نوع ويستار، بأوزان تتراوح بين 180 إلى 220 جرام. تم الحصول على هذه الجرذان من بيت الحيوانات التجريبية في جامعة النهضة، بني سويف. تم تقسيم الجرذان إلى 7 مجموعات تجريبية باستخدام التوزيع العشوائي. لحساب حجم التأثير، استخدمنا بيانات منشورة سابقًا من بولوس وآخرون.، بقوة 0.9 وعتبة دلالة 0.05. قبل بدء الإجراءات التجريبية، تم منح الفئران أسبوعًا للتكيف مع بيئة المختبر. تم الاحتفاظ بالمواضيع في حاويات بلاستيكية مع رطوبة منظمة (درجة الحرارة )، ودورة ضوء وظلام مدتها 12 ساعة لكل منهما. تم إعطاء الجرذان نظام غذائي تجاري قياسي وكان لديها وصول غير محدود إلى الماء.
إعلان الأخلاقيات
تم إجراء تجارب على الحيوانات وفقًا للمعايير الدولية لرعاية واستخدام الحيوانات المخبرية (لجنة المجلس الوطني للبحوث (الولايات المتحدة) لتحديث الدليل لرعاية واستخدام الحيوانات المخبرية، الطبعة الثامنة. واشنطن (دي سي): مطبعة الأكاديميات الوطنية (الولايات المتحدة)؛ 2011). تم اعتماد جميع بروتوكولات التجارب من قبل لجنة أخلاقيات البحث لرعاية واستخدام الحيوانات المخبرية في جامعة المنيا، كلية الصيدلة (رقم الموافقة الأخلاقية: 2301004). بالإضافة إلى ذلك، تم الإبلاغ عن جميع بروتوكولات التجارب وفقًا للإرشادات واللوائح ذات الصلة المبلغ عنها في إرشادات ARRIVE 2.0.https://arriveguidelines.org).
تصميم تجربة
تضمن التجربة توزيع الفئران عشوائيًا على مجموعات مختلفة. شملت الدراسة سبع مجموعات، كل منها تحتوي على ستة فئران. تلقت مجموعة التحكم من الفئران محلولًا من الماء وقطرات توين 80. يوم ) لمدة 21 يومًا. تلقت الفئران في مجموعة GA جرعة يومية من GA عن طريق الفم ( يوم، مذاب فيماء يحتوي على قطرات توين 80) لمدة 21 يومًا. تلقت الفئران في مجموعة GLA GLA عن طريق الفم ( يوم ) في محلول من الماء وقطرات توين 80 لمدة 21 يومًا. تم إعطاء مجموعة AZM محلولًا من الماء مختلطًا بقطرات توين 80 عن طريق الفم ( يوم ) لمدة 21 يومًا. بدءًا من اليوم الثامن، تم إعطاء هذه الفئران أيضًا AZM عن طريق الفم بجرعة من يوم في محلول من الماء وقطرات توين 80 لمدة 14 يومًا. تم إعطاء الفئران في مجموعة GA + AZM GA عن طريق الفم (اليوم) لمدة 21 يومًا. في اليوم الثامن، تلقت الفئران AZM عن طريق الفم يوم ، في محلول من الماء وقطرات توين 80) لمدة 14 يومًا. تم إعطاء مجموعة الفئران المعروفة باسم GLA + AZM GLA عن طريق الفم بجرعة من وزن الجسم لمدة 21 يومًا. في اليوم الثامن، تلقوا إدارة فموية لـ AZM بجرعة منيوم في محلول من الماء وقطرات توين 80 لمدة 14 يومًا. تم إعطاء مجموعة الفئران المسمّاة GA + GLA + AZM كل من GA وGLA عن طريق الفم لمدة 21 يومًا، بجرعة منوزن الجسم لكل مادة. بدءًا من اليوم الثامن، قمنا بإعطاء AZM عن طريق الفم بجرعة يومية منلمدة 14 يومًا.
تم اختيار جرعات GLA و GA وفقًا لأبحاث سابقة أظهرت تأثيراتها الواقية للكبد ضد مواد مختلفة.بالإضافة إلى ذلك، تم إنشاء نموذج إصابة الكبد الناتج عن AZM وفقًا للتجارب التجريبية الأولية وكذلك كما تم وصفه سابقًا بواسطة بولوس وآخرون..
جمع العينات والتحضيرات
تم تخدير الفئران باستخدام الكيتامين/زيلازين24 ساعة بعد العلاج النهائي. ثم تم أخذ عينات من الدم من الضفيرة الوردية خلف العين. تم euthanize الفئران من خلال خلع العنق بعد استخراج الدم. بعد ذلك، تم استخراج الكبد على الفور، وغسله بمحلول ملحي عادي بارد، ووزنه. لإجراء دراسات نسيجية، تم غمر قطعة من نسيج الكبد على الفور فيمحلول الفورمالين المخزن. تم تجانس أنسجة الكبد باستخدام محلول تريس-هيدروكلوريك بنسبة (نسيج الكبد للتخزين) بتركيز 150 مللي مولار ودرجة حموضة 7.4. تم تقسيم الهوموجينات إلى قوارير وتخزينها عند درجة حرارة . باستخدام العوازل المناسبة، تم إعداد تخفيفات من الأخيرة لاختبار كميات GSH، ومنتج LP MDA، والبروتينات الكلية، ونشاط SOD و CAT. تم استخدام أنابيب الطرد المركزي لجمع الدم، الذي تم بعد ذلك إخضاعه للطرد المركزي لمدة 20 دقيقة بسرعة 3000 دورة في الدقيقة في جهاز طرد مركزي مبرد تم ضبطه على درجة حرارة تم تنفيذ هذه العملية لاستخراج المصل. تم استخدام عينات المصل لقياس مستويات ALT و AST و ALP والألبومين.
الاختبارات الكيميائية الحيوية والأنسجة المرضية
تقييم مؤشرات سمية الكبد
تم تقييم مؤشرات إصابة الكبد من خلال قياس مستويات الألانين أمينوترانسفيراز (ALT) والأسبارتات أمينوترانسفيراز (AST) والفوسفاتاز القلوي (ALP) والألبومين. تم استخدام مجموعات تجارية من BIOMED في العاشر من رمضان، القاهرة، مصر، وفقًا لتعليمات الشركة المصنعة.
تحليل الأنسجة باستخدام صبغة الهيماتوكسيلين والإيوزين
وفقًا للإجراء الموضح في كتاب بانكروفتتم قص العينات المحفوظة وتنظيفها وتجفيفها باستخدام محاليل كحولية تتزايد قوتها، ومعالجتها بالزيلين لإزالة الشوائب، وتغليفها بالشمع، ثم قطعها إلى شرائح قياسميكرومترات في السمك. تم صبغ المقاطع بعد ذلك باستخدام طريقة الهيماتوكسيلين والإيوزين (H&E). تم إجراء التقييمات النسيجية من خلال تحليل خمس مناطق متميزة من كل عينة كبد بتكبير عشوائي 200x. كانت التقييمات مركزة حول تحديد التغيرات النسيجية، بما في ذلك التنكس المائي، النخر الموضعي، الاحتقان الوعائي، وت infiltrations الخلوية الالتهابية. تم تقييم حجم هذه التغيرات باستخدام طريقة تصنيف تم استخدامها في تحقيق سابق أجراه إبراهيم فؤاد وأحمد.. كان نظام الدرجات يتراوح بين 0 و 3، وكان لكل قيمة دلالة محددة. تم تحديد الدرجات كنسبة مئوية لقياس مدى التغيرات النسيجية المرضية المكتشفة. كانت الدرجة 0 (صفر) تشير إلى نتيجة سلبية، بينما كانت الدرجات، و تشير إلى تعديلات خفيفة ومتوسطة وشديدة، على التوالي.
قياس تركيز المالونديالديهايد (MDA)
اختبار حمض الثيوباربيتيوريك (TBA) الذي وصفه بويج وأوستتم استخدامه لقياس تركيز المالونديالديهايد (MDA) في مستخلصات أنسجة الكبد، حيث يتفاعل MDA مع TBA في وسط حمضي مكونًا مركبًا ملونًا باللون الوردي يتم قياسه طيفيًا عند 535 نانومتر. تم إجراء الاختبار بالطريقة التالية: أولاً، يتم تحضير كاشف (TBA-TCA-HCl) (الكروموجين) عن طريق إذابة 15 جرام من حمض ثلاثي كلور الأسيتيك TCA ( النهائي) و 375 ملغ من TBA ( النهائي) في ، تليها إضافة 2.08 مل من المركز النهائية). ثم، تم تسخين المزيج إلى في حمام مائي (لمساعدة ذوبان TBA)، ثم يتم إكمال الحجم إلى 100 مل بالماء المقطر، وتم تغطيته أثناء التحضير لمنع تغيير التركيز. تم إنشاء التفاعل عن طريق خلط 0.5 مل من مستحلب الكبد، 0.5 مل من الماء المقطر و 2 مل من كاشف TBA/TCA/HCl. تم تعريض المحلول لمعالجة حرارية في حمام مائي لمدة 15 دقيقة. بعد عملية التبريد، يُسمح بالتبريد والطرد المركزي بسرعة 5000 دورة في الدقيقة لمدة 10 دقائق لعزل طبقة السائل الملون. تم قياس امتصاص المنتج الوردي عند 535 باستخدام مطياف الأشعة فوق البنفسجية والمرئية، مع عينة فارغة للمقارنة. تم تحديد تركيز MDA في نسيج الكبد كـ البروتين. من خلال استخدام هذه الطريقة، تم تقييم تركيز MDA في مستخلصات أنسجة الكبد، مما يعد مؤشراً على مستويات تأكسد الدهون.
قياس نشاط سوبر أكسيد ديسموتاز (SOD)
تم تحديد مستوى نشاط سوبر أكسيد ديسموتاز (SOD) في مستحلب الكبد باستخدام الطريقة الموضحة من قبل ناندي وتشاتيرجي.. تعتمد هذه التقنية على قدرة SOD على إعاقة الأكسدة التلقائية للبايروجالول في ظل ظروف قلوية. فيما يلي نظرة عامة مختصرة على العملية: تم إضافة عينة الكبد إلى 2 مل من محلول يحتوي على 20 مليمول من عازلة Tris-HCL (تحتوي على 1 مليمول EDTA عند درجة حموضة 8.2). تم بدء التفاعل عن طريق إدخال محلول بايروجالول بتركيز 15 مليمول تم إنتاجه مؤخرًا. تم قياس الامتصاص عند طول موجي 420 نانومتر باستمرار لمدة دقيقتين، مع الحصول على قياسات خلال الفترة الزمنية من 1 إلى 3 دقائق. تم تحديد النتائج بوحدات هوموجينات الأنسجة لكل ملليجرام من البروتين. يتم تحديد وحدة واحدة من نشاط SOD على أنها الكمية من الإنزيم اللازمة لتقليل الأكسدة الذاتية للبايروجالول بواسطة . تم التعبير عن النتائج بوحدات هوموجينات الأنسجة لكل ملليجرام من البروتين.
قياس محتوى الجلوتاثيون ونشاط الكاتالاز
تم قياس محتوى الجلوتاثيون (GSH) ونشاط الكاتالاز في هوموجينات الكبد باستخدام مجموعات تشخيصية حيوية تم الحصول عليها من 29 شارع التحرير، الدقي، الجيزة، مصر. كانت المجموعات المستخدمة هي رقم الكات. GR 2511 للجلوتاثيون (GSH) ورقم الكات. CA 2517 للكاتالاز. تم إجراء القياسات وفقًا لتعليمات الشركة المصنعة.
تم استخدام كاشف Biuret باستخدام مجموعة Spectrum-diagnostics التي تم الحصول عليها من منطقة الصناعات في مدينة العبور، القاهرة، مصر، لتحديد محتوى البروتين الكلي في أنسجة الكبد. كانت المجموعة المستخدمة هي رقم الكات. 310,001. تم أخذ القياسات باستخدام مطياف UV/VIS من نوع PerkinElmer Lambda 25 وتم الإبلاغ عنها كقراءات امتصاص.
تحليل Western blot
تم سحق الأنسجة إلى خليط متجانس باستخدام محلول تحلل Tris الذي يتكون من Triton X-100، و50 مليمول من Tris pH 7.4. بالإضافة إلى ذلك، تم تضمين خليط مثبطات البروتياز (Biospes، الصين). تم إجراء عملية الهوموجينيزه لمدة 30 دقيقة عند درجة حرارة . تم التخلص من الأنسجة المتبقية باستخدام الطرد المركزي بسرعة 10,000 دورة في الدقيقة لمدة 10 دقائق عند درجة حرارة . تم قياس تركيز البروتين بعد ذلك باستخدام مجموعة اختبار بروتين BCA (Thermo Fisher Scientific، الولايات المتحدة الأمريكية) وفقًا للتعليمات المقدمة من الشركة المصنعة. بعد ذلك، تم إيداع كميات متساوية من البروتين (60 ملليجرام من البروتين الكلي في كل حارة) التي تم فصلها باستخدام هلام بولي أكريلاميد SDS بتركيز 10% على غشاء PVDF (Millipore، Merk، الولايات المتحدة الأمريكية) . تم حجب الأغشية بمحلول يحتوي على حليب خالي من الدسم في عازلة TBST لمدة ساعة واحدة عند درجة حرارة الغرفة. بعد ذلك، خضعت الأغشية لعملية غسيل ثم تم تعريضها لحضانة ليلية عند درجة حرارة مع الأجسام المضادة الأولية: مضاد Nrf2 (Biospes، الصين؛ تخفيف 1:200؛ CAT# YPA1865)، مضاد NF-B p65 (Cell Signaling Technologies؛ تخفيف 1:1000، CAT# 8242)، أو مضاد الأكتين (Cell Signaling Technologies؛ تخفيف 1:1000، CAT# 7074). بعد إجراء جولة أخرى من الغسيل، تم تعريض الأغشية للأجسام المضادة الثانوية المرتبطة بالفوسفاتاز القلوي المناسبة (1:5000، Novus Biologicals، LLC، Littleton، CO، الولايات المتحدة الأمريكية) لمدة ساعة واحدة. تم ملاحظة الأشرطة باستخدام مجموعة كشف الركيزة BCIP/NBT من Genemed Biotechnologies، Inc.، الموجودة في كاليفورنيا، الولايات المتحدة الأمريكية. كان الأكتين بمثابة مرجع داخلي لتحميل البروتين، وتمت مقارنة الأشرطة الناتجة مع الأكتين باستخدام برنامج Fiji ImageJ (2.16.0) http://imagej.net/software/fiji/ (NIH، الولايات المتحدة الأمريكية).
تقدير السيتوكين المسبب للالتهابات، TNF-
تم تقدير مستويات TNF- في الكبد باستخدام مجموعات ELISA التي تم الحصول عليها من Elabscience (CAT# E-EL-R0019، 14780 Memorial Drive، Suite 108، هيوستن، تكساس، 77079، الولايات المتحدة الأمريكية)، وفقًا لتعليمات الشركة المصنعة.
التحليل الإحصائي
تم إجراء الدراسة الإحصائية باستخدام برنامج SPSS من IBM، (الإصدار 25.0، شيكاغو، إلينوي، الولايات المتحدة الأمريكية). تم إنشاء الرسوم البيانية باستخدام برنامج GraphPad Prism 9 (سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية). تم استخدام المتوسط ± الخطأ المعياري (SEM) لتمثيل البيانات. تم تقييم التوزيع الطبيعي للبيانات باستخدام اختبار Shapiro-Wilk. تم إجراء تحليل التباين الأحادي (ANOVA)، تلاه تحليل Tukey’s post-hoc. تعتبر قيمة P فرقًا ذا دلالة إحصائية.
النتائج
حسنت GA وGLA ومزيجها من زيادة وزن الجسم ووزن الكبد في الجرذان المعالجة بـ AZM
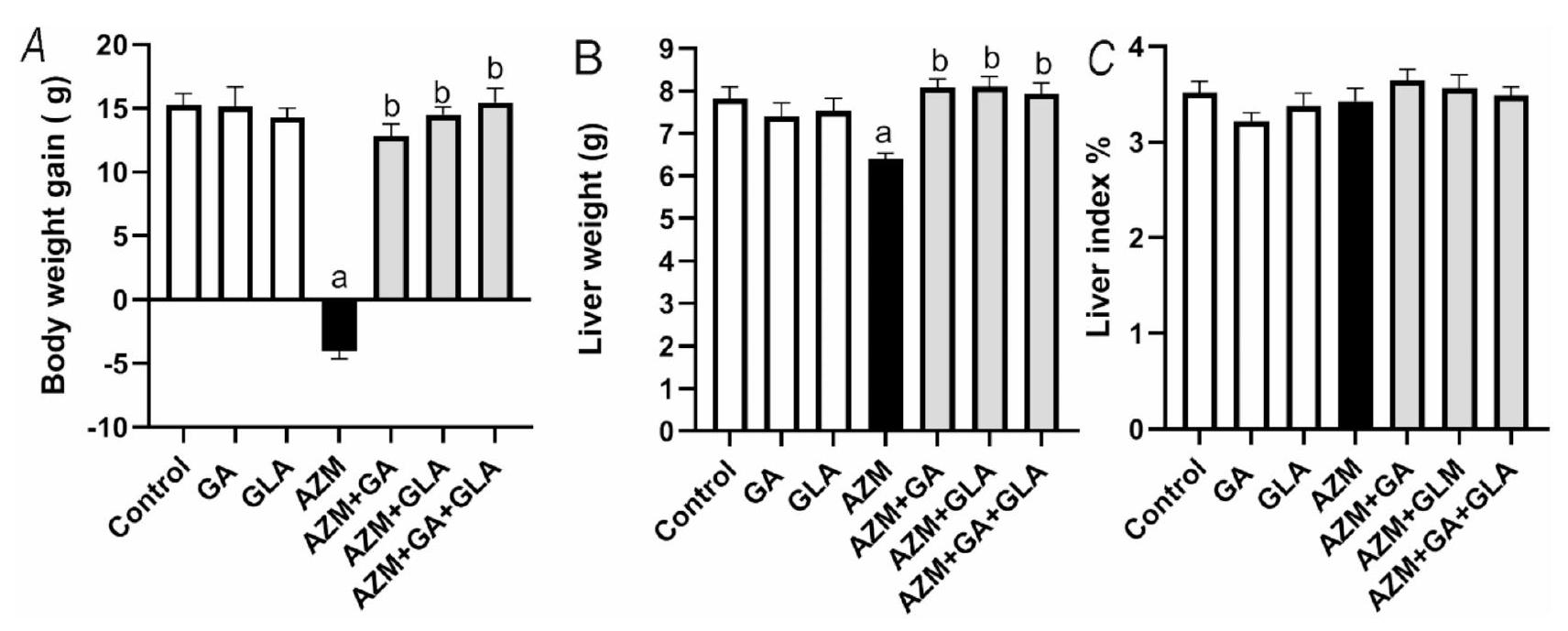
تظهر الشكل 1 زيادة وزن الجسم ووزن الكبد وقيم مؤشر الكبد للمجموعات التجريبية. أدى إعطاء AZM للجرذان إلى تقليل كبير في زيادة وزن الجسم ووزن الكبد ( )، ولكن لم يتم الكشف عن أي تغييرات في مؤشر الكبد. ومع ذلك، أدى إعطاء AZM بالتزامن مع GA وGLA ومزيجها إلى زيادة كبيرة في مؤشرات الجرذان، مما يدل على تعزيز فعال جدًا لهذه المؤشرات.
خففت GA وGLA ومزيجها من إصابة الكبد في الجرذان المعالجة بـ AZM
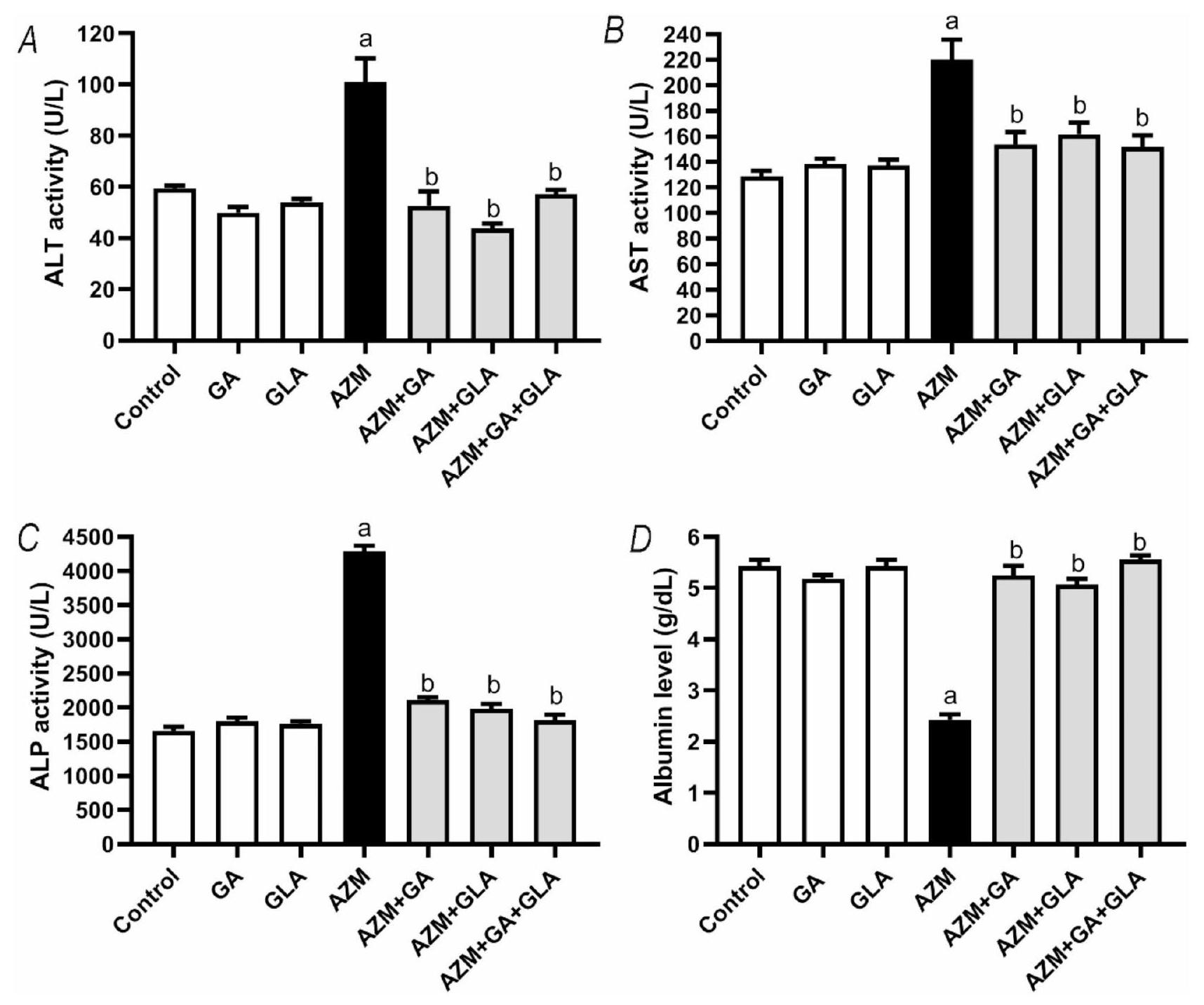
بعد تلقي AZM، زادت مستويات ALT وAST وALP في مصل الدم للجرذان بشكل كبير. على وجه التحديد، كان هناك زيادة تقريبية بنسبة في ALT، وزيادة بنسبة في AST، وزيادة بنسبة في ALP عند مقارنتها بمجموعة التحكم من الجرذان. انخفضت مستويات الألبومين في المصل بشكل كبير بنسبة مقارنة بمجموعة التحكم ( ) (الشكل 2). أظهرت مجموعة AZM + GA انخفاضًا كبيرًا في نشاط ALT وAST وALP في المصل مقارنة بمجموعة ASM، بنسبة و على التوالي. بالمقارنة مع مجموعة AZM، أظهرت مجموعة AZM + GLA مستويات مصل أقل بشكل كبير من ALT وAST وALP ( و على التوالي). علاوة على ذلك، لاحظنا انخفاضًا كبيرًا في قيم ALT وAST وALP في المصل في مجموعة AZM + GA + GLA. عند مقارنتها بمجموعة AZM، انخفضت مستويات ALT وAST وALP بنسبة و على التوالي. كانت مستويات الألبومين في المصل في مجموعة AZM + GA،
الشكل 1. تأثير GA وGLA ومزيجها على (A) زيادة وزن الجسم، (B) وزن الكبد، و(C) مؤشر الكبد في الجرذان المعالجة بـ AZM. تم تقديم النتائج كمتوسط ± SEM ( ). تم استخدام تحليل ANOVA الأحادي، تلاه تحليل Tukey’s post-hoc مع عتبة دلالة إحصائية قدرها . ; تختلف بشكل كبير عن مجموعة التحكم؛ ; تختلف بشكل كبير عن مجموعة AZM. حيث تشير GA وGLA وAZM إلى حمض الجاليك وحمض الجليسرهيتيك وكذلك الأزيثرومايسين على التوالي.
كانت مجموعات AZM + GLA وAZM + GA + GLA أعلى بكثير من مجموعة AZM ( و على التوالي) (الشكل 2D). توضح هذه البيانات أن مجموعة AZM + GA + GLA هي الأكثر حماية ضد تلف الكبد الناتج عن AZM.
خففت GA وGLA ومزيجها من التغيرات النسيجية في كبد الجرذان المعالجة بـ AZM
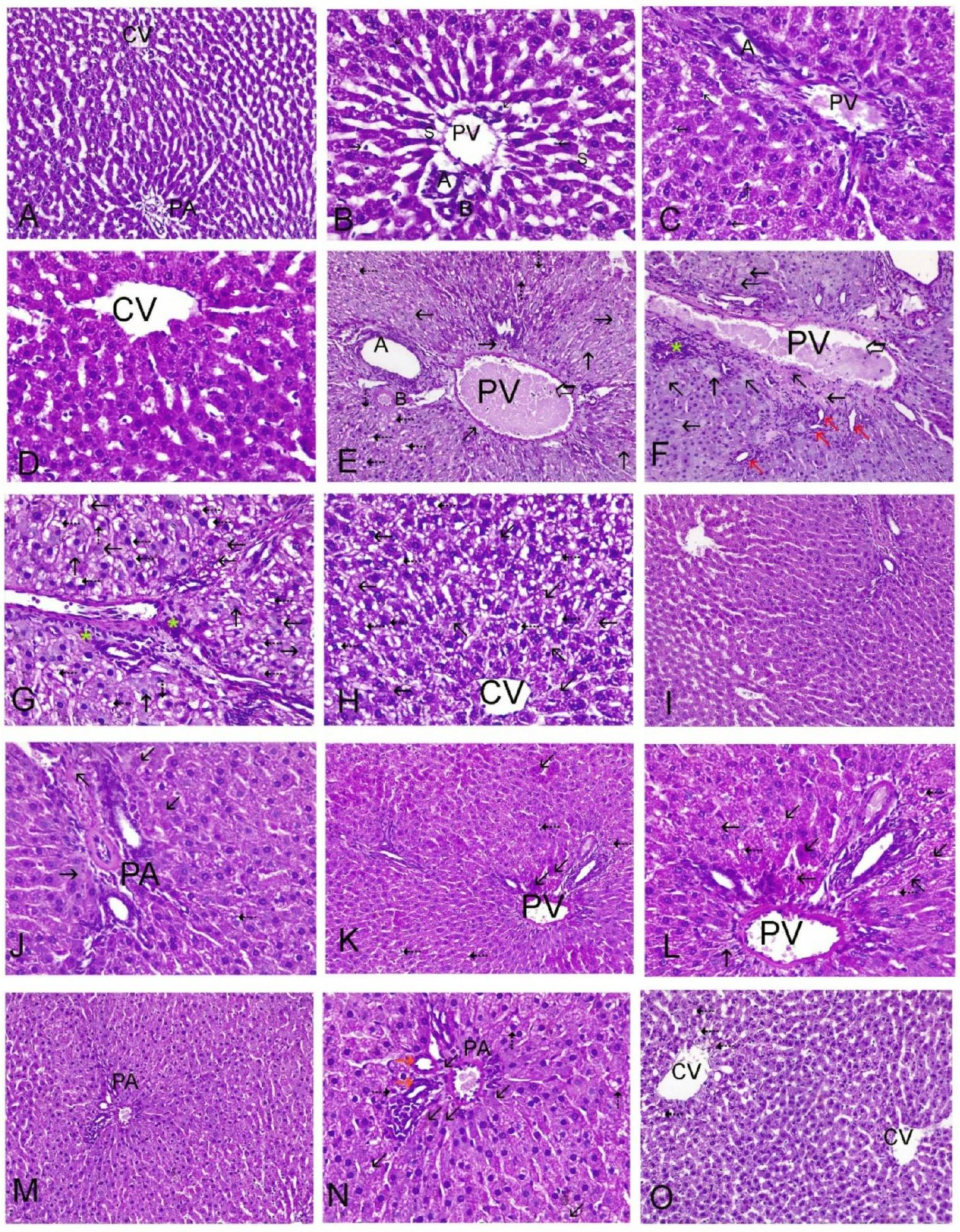
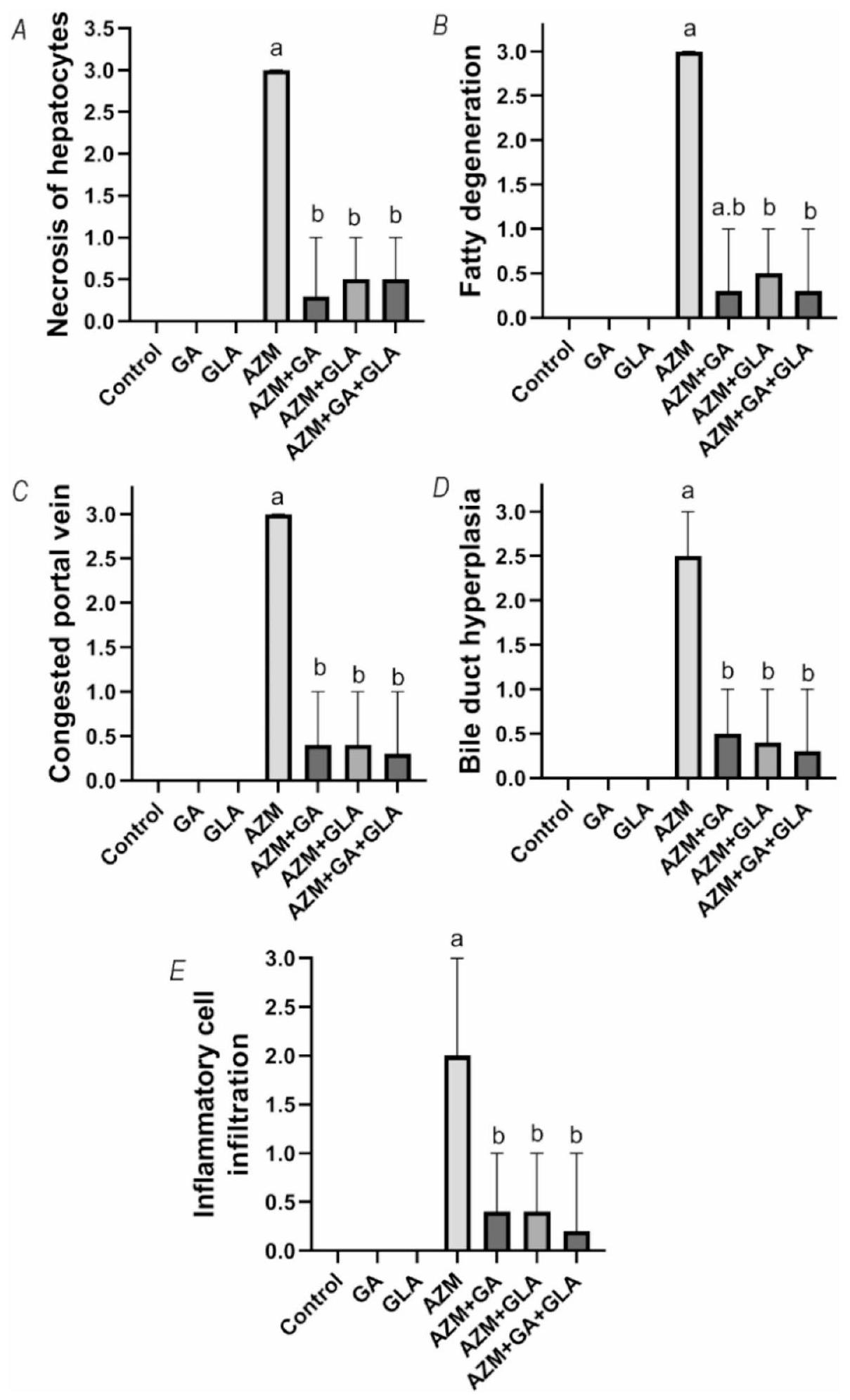
أكدت الدراسة النسيجية المرضية لأنسجة الكبد نتائج الاختبارات الكيميائية الحيوية. كانت مجموعات التحكم وGA وGLA جميعها تحتوي على هياكل كبدية نموذجية، كما هو موضح في الشكل 3A&B لمجموعة التحكم، وفي الشكل 3C&D لمجموعات GA وGLA. تضمنت الهيكل الكبدي حبال كبدية كلاسيكية، وأوردة مركزية (CV)، ومنطقة بوابة (PA) مع شعيرات دموية سليمة. أظهرت أكباد الجرذان التي تلقت AZM نخرًا شديدًا وتحلل دهني. عرضت كل من المناطق البوابة (الشكل 3E وF وG) والمناطق المركزية (H) سيتوبلازمًا متجوفًا ونخرًا. أظهرت مقاطع AZM قنوات صفراوية متكاثرة، وتسلل خلايا التهابية، واحتقان كبير في الوريد البابي. يظهر الشكل 3I&J نخرًا خفيفًا وتجويفًا في المنطقة البوابة بعد علاج GA وAZM. أظهرت مقاطع كبد الجرذان المعالجة بـ AZM وGLA نخرًا خلويًا بؤريًا، والتهابًا خفيفًا، وسيتوبلازمًا متجوفًا في المنطقة البوابة (الشكل 3K & L). احتفظت خلايا الكبد في المناطق البوابة والمناطق الوريدية المركزية لمقاطع كبد الجرذان المعالجة بـ AZM وGA مع GLA بسلامتها الهيكلية. ومع ذلك، احتوت بعض الخلايا الكبدية على تجويفات وبؤر صغيرة من النخر، بالإضافة إلى قنوات صفراوية متكاثرة بشكل خفيف (الشكل 3M وN وO). يظهر الشكل 4 أن علاج AZM زاد بشكل كبير من درجة النسيجية الكبدية مقارنة بالمجموعة الطبيعية ( ). ومع ذلك، في مجموعات و كانت درجة النسيجية الكبدية أقل بكثير من علاج AZM.
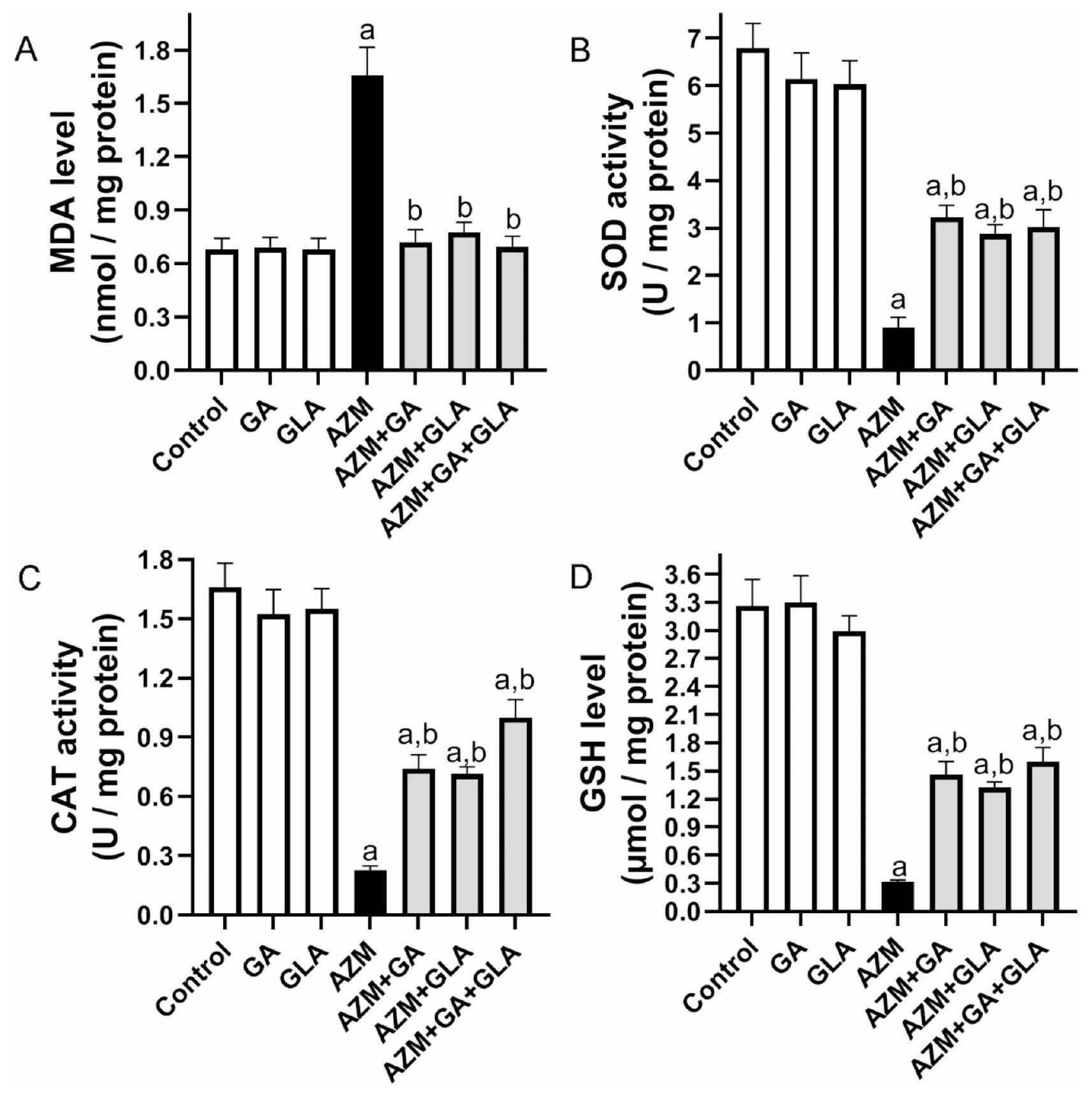
خففت GA وGLA ومزيجها من الإجهاد التأكسدي في أكباد الجرذان المعالجة بـ AZM. في مجموعة AZM، ارتفع مقدار MDA في الكبد بنسبة ، لكن نشاط SOD وCAT انخفض بنسبة وعلى التوالي، مقارنة بمجموعة التحكم (الشكل 5). مقارنة بمجموعة AZM، كانت مجموعات AZM + GA وAZM + GLA وAZM + GA + GLA تحتوي على كميات أقل بشكل ملحوظ من MDA الكبدي.من، و على التوالي. ومع ذلك، مقارنةً بمجموعة AZM، كانت هذه المجموعات تتمتع بنشاط أعلى بكثير من SOD الكبدي بواسطة، و بالإضافة إلى زيادة نشاط CAT بواسطة، و ، على التوالي (الشكل 5). من ناحية أخرى، انخفض محتوى الجلوتاثيون في كبد مجموعة AZM بـ مقارنةً بمجموعة التحكم. الـ ، وكانت مستويات الجلوتاثيون في الكبد في مجموعات AZM + GA + GLA أعلى بكثير من مجموعة AZM (367.26، 322.46%، وتظهر هذه البيانات أن مجموعة AZM + GA + GLA هي الأكثر حماية ضد تلف الكبد الناتج عن AZM.
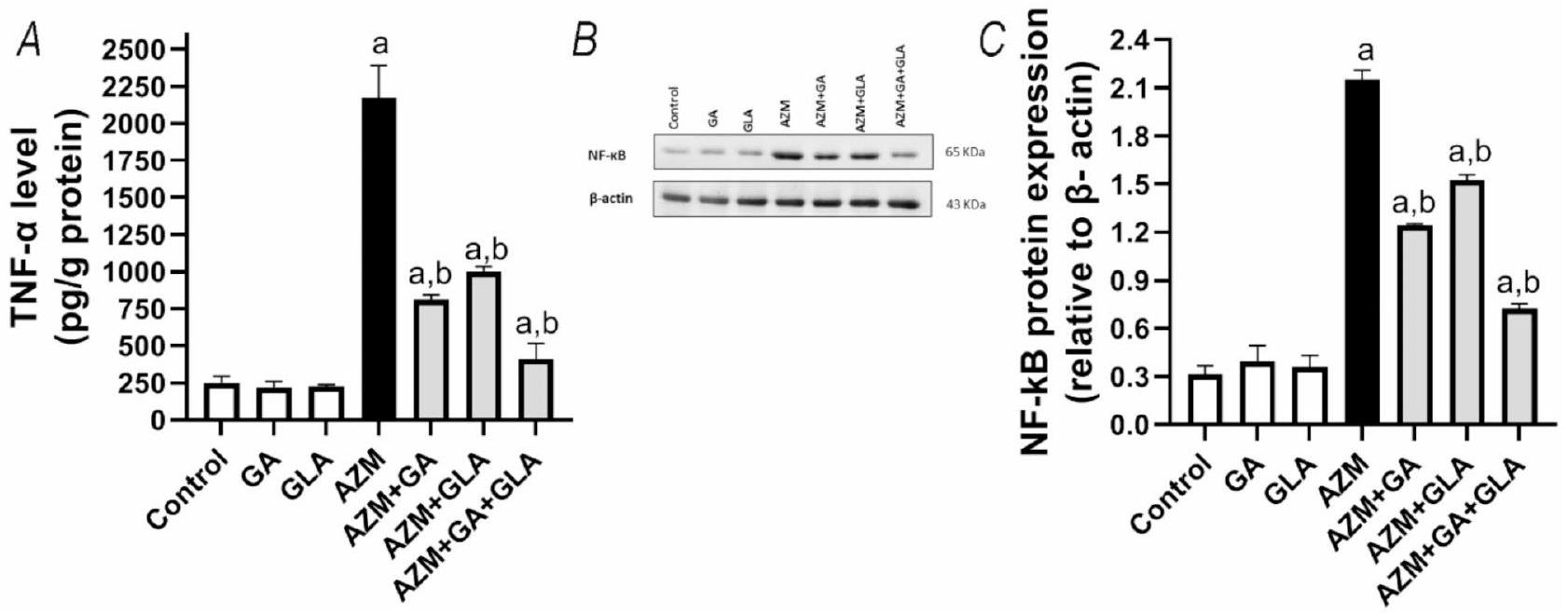
GA و GLA وتركيبهما قللا من الالتهاب في كبد الفئران المعالجة بـ AZM
تم تقييم التأثيرات المضادة للالتهابات لـ GA و GLA في الجرذان المعالجة بـ AZM من خلال تقييم علامات الالتهاب مثل TNF- وزاد العلاج بـ AZM من مستويات TNF- وبروتينات NF-kB في الكبد بواسطة و على التوالي، مقارنة بمجموعة التحكم (قيمة p ). من ناحية أخرى، TNF- الكبدي وكانت مستويات تعبير NF-kB قد انخفضت بشكل ملحوظ بعد معالجة الحيوانات بـ GA و GLA و GA+GLA بحوالي و و ، و و ، على التوالي، مقارنةً بـ AZM عند قيمة pتظهر هذه البيانات أن مجموعة AZM + GA + GLA هي الأكثر حماية ضد الالتهاب الكبدي الناتج عن AZM.
الشكل 2. تأثير GA وGLA وتركيبهما على سمية الكبد الناتجة عن AZM في الجرذان. تشمل المؤشرات (A) نشاط ALT في المصل، (B) نشاط AST في المصل، (C) نشاط ALP في المصل، و(D) مستوى الألبومين في المصل. يتم عرض البيانات كمتوسطSEMقمنا أولاً بإجراء تحليل ANOVA الإحصائي، تلاه تحليل توكي بعد الاختبار.تشير إلى فرق كبير عن مجموعة التحكم، بينما بتشير إلى فرق كبير عن مجموعة AZM.
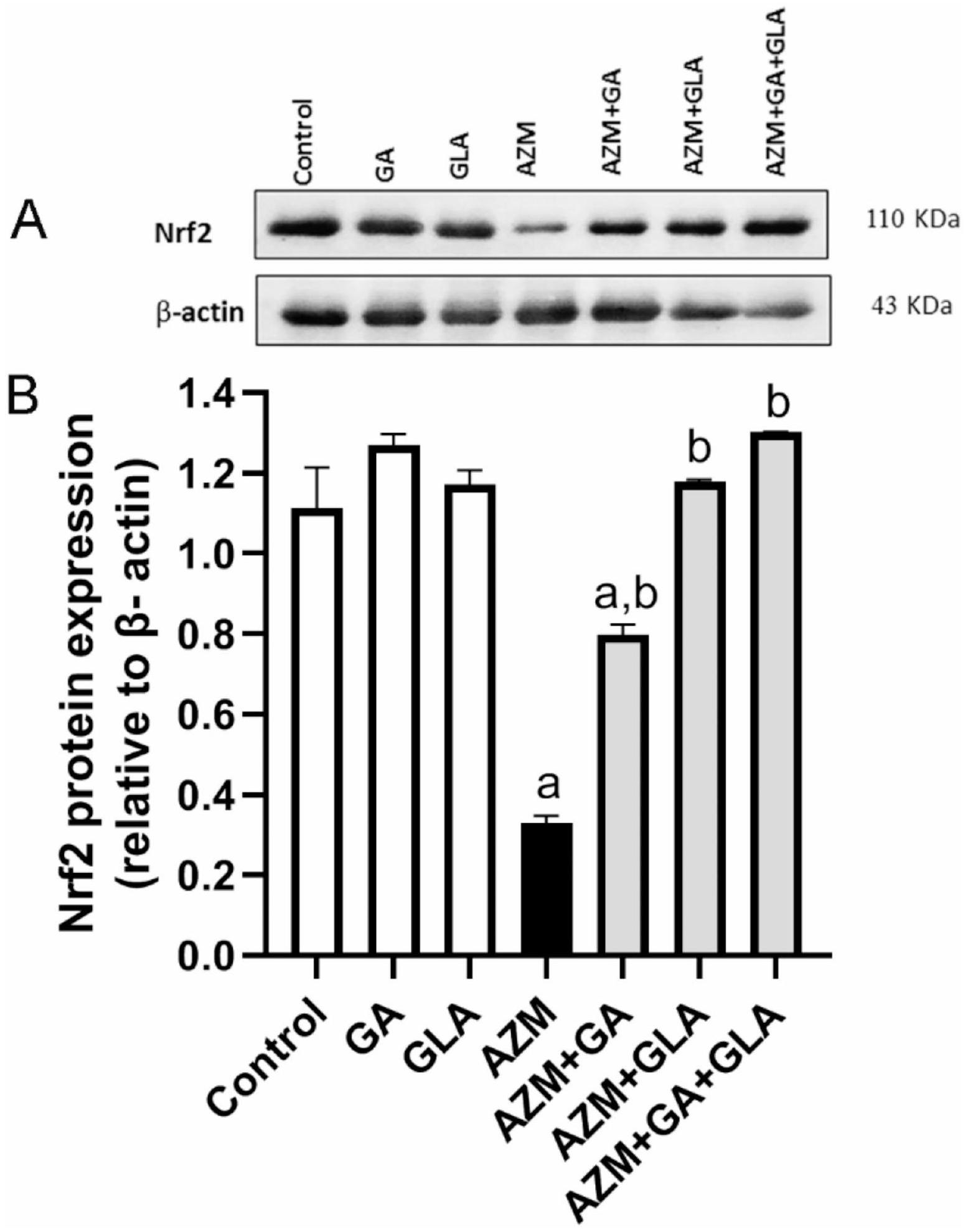
GA و GLA وتركيبهما قللا من تثبيط تعبير Nrf2 في كبد الفئران المعالجة بـ AZM
أظهر تحليل Western blot أن كمية بروتين Nrf2 في كبد الجرذان التي تم إعطاؤها AZM كانتأخفضمقارنةً بمجموعة التحكم (الشكل 7B). ومع ذلك، بعد المعالجة المسبقة بـ GA أو GLA أو GA + GLA، زادت مستويات تعبير Nrf2 في كبد الفئران المعالجة بـ AZM بشكل كبير بحوالي، ، و ، على التوالي، مقارنةً بمجموعة AZM. (الأشكال 7A وB).
نقاش
تُعد إصابة الكبد الحادة الناتجة عن الأدوية قضية صحية عالمية رئيسية، وغالبًا ما تؤدي إلى فشل الكبد، والحاجة إلى زراعة الأعضاء، وحتى الوفاة. لمعالجة هذه القضية الحرجة، هناك حاجة ملحة لتطوير وتنفيذ تدخلات علاجية فعالة. تُعتبر إصابة الأنسجة الناتجة عن الأكسدة والالتهاب آليتين رئيسيتين تدفعان آلية حدوث إصابة الكبد الحادة.لذلك، فإن المركبات الجديدة التي تتمتع بخصائص مضادة للأكسدة ومضادة للالتهابات تحمل وعدًا في الحماية من خلل وظائف الكبد. في الدراسة الحالية، قمنا بالتحقيق في الفوائد الصحية المحتملة لـ GA و GLA ومزيجهما في التخفيف من سمية الكبد الحادة الناتجة عن AZM في الفئران. أظهرت نتائجنا أن هذه المركبات خففت بشكل فعال من الإجهاد التأكسدي، وقللت الالتهاب، ومنعت الأضرار اللاحقة في أنسجة الكبد.
بعض الأدوية، وخاصة AZM، لديها خطر مرتفع للإصابة الحادة بالكبد.. في دراستنا، أظهرت الفئران المعالجة بـ AZM وحده فقدان الوزن وانخفاض وزن الكبد، مما يدل على السمية الجهازية واضطراب الأيض. بالمقابل، أدى العلاج المشترك مع GA أو GLA أو تركيبهما إلى زيادة في وزن الجسم ووزن الكبد، مما يشير إلى أن هذه المركبات تعزز من عملية الأيض في الكبد وعمليات إزالة السموم. وقد أفادت دراسات سابقة أن مضاد حيوي يُستخدم بشكل روتيني، مثل AZM، غالبًا ما يسبب إصابة حادة في الكبد.أظهرت الدراسات الحيوانية أن الضرر الحاد الذي يسببه AZM للكبد مرتبط بارتفاع مؤشرات الكبد الدائرة، وضعف وظيفة الكبد، وتطور تغييرات هيستوباثولوجية تشمل التنكس الكبدي، والتغيرات الدهنية، والاحتقان الجيبي، وتسلل الخلايا الالتهابية.. وبالمثل، اكتشفنا أن معالجة الفئران من نوع ويستار بـ AZM بجرعة من تسبب في إصابة شديدة في الكبد كما يتضح من ارتفاع مستويات ALT وAST وALP. في نموذجنا التجريبي، كانت مستويات المؤشرات الحيوية للكبد المرتفعة مرتبطة بوجود تشوهات شكلية كبيرة، بما في ذلك النخر الشديد، والتنكس الدهني، وتكاثر القنوات الصفراوية، وتسلل الخلايا الالتهابية. تتماشى هذه التغيرات الملحوظة في تلف الكبد وبنيته مع النتائج من الدراسات السابقة.تسبق نخر الخلايا الكبدية والتنكس تمزق الغشاء، مما يؤدي إلى إطلاق المؤشرات الحيوية الكبدية في مجرى الدم وارتفاع مستوياتها في الدم لاحقًا.وبالتالي، في ظل ظروف الضغط الكبدي، تعتبر مستويات إنزيمات الكبد المرتفعة في المصل مؤشراً حساساً على زيادة نفاذية غشاء الخلايا الكبدية وتلف الخلايا.بالإضافة إلى تلف الكبد، أظهرت الفئران المعالجة بـ AZM خللاً في وظائف الكبد، كما يتضح من انخفاض مستويات الألبومين في المصل. تظهر نتائجنا أن GA وGLA وتركيباتهما توفر حماية ضد إصابة الكبد الناتجة عن AZM. أكدت التحليلات النسيجية هذه التأثيرات الوقائية، كما يتضح من الحفاظ على مستويات إنزيمات الكبد الطبيعية (ALT وAST وALP) والألبومين، إلى جانب الحفاظ على سلامة نسيج الكبد. كشفت دراسات مختلفة أن GA قدمت تأثيرات واقية للكبد ضد الأضرار الناتجة عن الهيدروبيروكسيد التيرت-بيوتيلي.بيسفينولالإيثانولوثيوأسيتاميد. وبالمثل، تم إظهار أن GLA يحمي خلايا الكبد من السموم مثل الكربون تتراكلورايد، 2-أسيتيل أمينوفلورين، سيكلوفوسفاميد، ميثوتريكسات، وألفا-نافثيل إيزوثيوسياناتفي هذه الدراسة، تظهر مجموعة AZM + GA + GLA أعلى مستوى من الحماية ضد إصابة الكبد الناتجة عن AZM، كما يتضح من نسبة الحماية الملاحظة.
يلعب الإجهاد التأكسدي ونظام الدفاع المضاد للأكسدة دورًا حاسمًا في إصابة الكبد الحادة الناتجة عن الأدويةتحت الظروف العادية، يقوم نظام الدفاع المضاد للأكسدة – الذي يتكون من SOD و CAT و GSH – بشكل فعال بتحييد الجذور الحرة الناتجة عن استقلاب المواد السامة والأدوية.يحدث الإجهاد التأكسدي عندما يكون هناك عدم توازن بين الجذور الحرة ومضادات الأكسدة.لقد أظهرت العديد من التحقيقات أن AZM يحفز إنتاج الجذور الحرة للأكسجين (ROS)، مما يؤدي إلى زيادة مستويات منتجات تأكسد الدهون (MDA) وتقليل مستويات مضادات الأكسدة، بما في ذلك CAT وSOD وGSH في قلوب وكبد الحيوانات المعالجة.كشفت نتائجنا أن AZM زاد من الإجهاد التأكسدي، كما يتضح من ارتفاع مستويات MDA بشكل ملحوظ. علاوة على ذلك، لاحظنا انخفاضًا مستمرًا في نشاط CAT وSOD، بالإضافة إلى انخفاض تركيزات GSH، مما يتماشى مع النتائج التي أبلغ عنها سينغ وآخرون.أظهرت الدراسة الحالية أن GA وGLA وتركيباتهما زادت بشكل فعال من مستويات مضادات الأكسدة (GSH وCAT وSOD) وقللت من منتجات تأكسد الدهون (MDA) في أنسجة كبد الجرذان المعالجة بـمن AZM، مع تسليط الضوء على خصائصها المحتملة كمضادات للأكسدة. على سبيل المثال، لقد أظهر GA أنه يحمي الكبد من الأضرار التأكسدية الناتجة عن مواد كيميائية مثل الرصاص.ثيوأسيتاميدورابع كلوريد الكربون. وبالمثل، أظهرت GLA القدرة على التخلص من أنواع الأكسجين التفاعلية وتخفيف الإجهاد التأكسدي في نماذج إصابة الكبد التي تتضمن حمض الفالبرويكسيكلوفوسفاميدو ميثوتريكساتأكدت نسبة الحماية أن مجموعة AZM أظهرت أعلى مستوى من الحماية ضد تلف الكبد الناتج عن AZM.
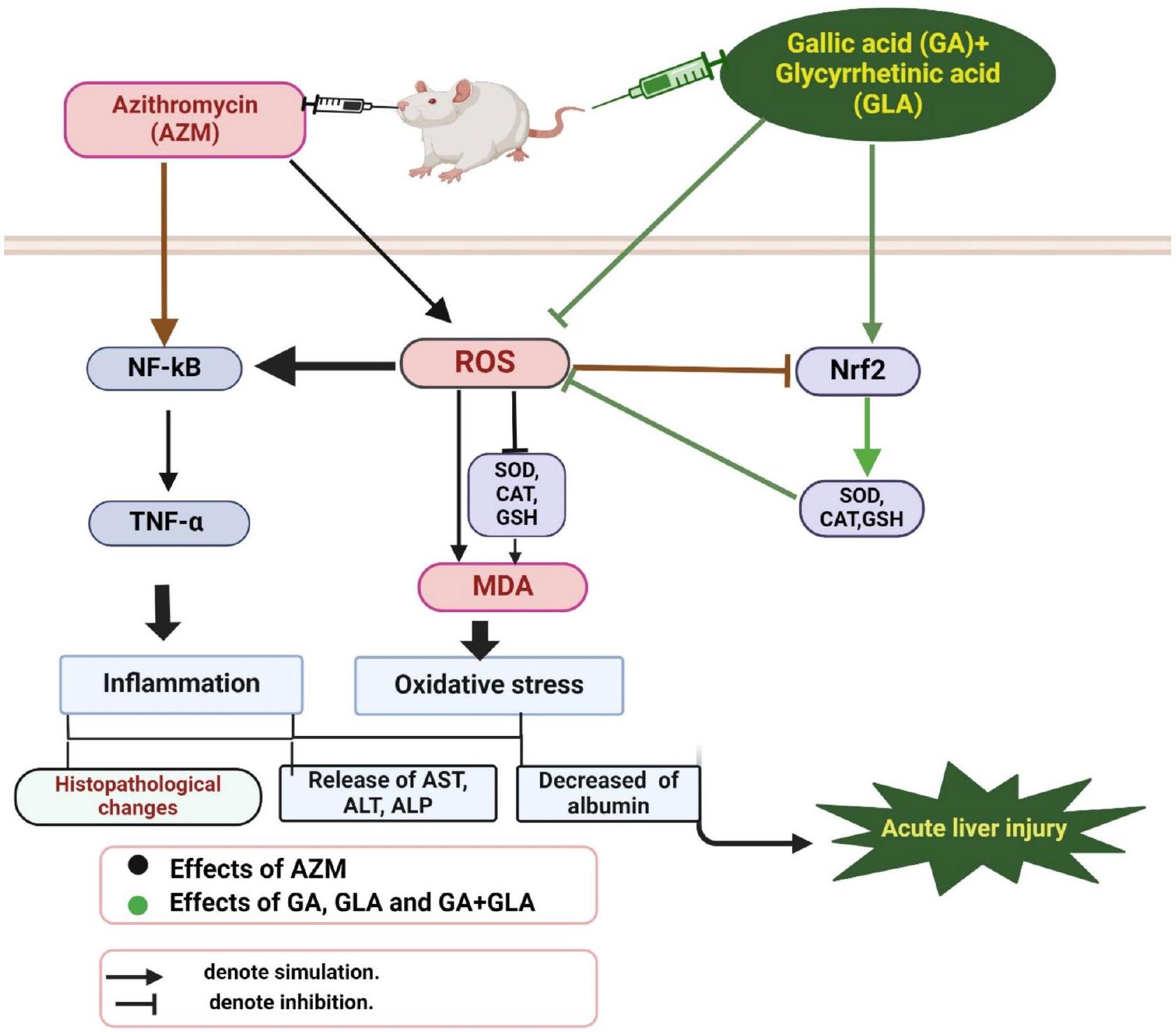
تحليل المسار الذي يتوسطه Nrf2 يوفر رؤى حول الآليات الكامنة وراء القدرات المضادة للأكسدة لـ GA و GLA. كلا من GA و GLA يزيدان من تنظيم Nrf2، مما يؤدي إلى تعزيز التعبير والنشاط لإنزيمات الدفاع المضادة للأكسدة. هذا يقلل من مستوى المؤكسدات التفاعلية ويخفف من الأضرار الخلوية المرتبطة بها. العلاج المسبق للفئران بـ GA أو GLA أو تركيبهما عكس الانخفاض الكبير في تعبير بروتين Nrf2 في نسيج الكبد الذي تسببت فيه AZM. وبالتالي، فإن زيادة تنظيم Nrf2 تعزز إنتاج إنزيمات الدفاع المضادة للأكسدة، مما يقلل من المؤكسدات التفاعلية ويقلل من إصابة الخلايا. تحت الضغط التأكسدي، ينتقل Nrf2 إلى النواة ويرتبط بعنصر الاستجابة المضادة للأكسدة (ARE)، مما يعزز التعبير عن الإنزيمات المشاركة في تخليق SOD و CAT و الجلوتاثيون.أظهر الباحثون أن GA ينظم مسار إشارة Nrf2، مما يوفر الحماية ضد تلف الكبد الناتج عن بيروكسيد التيرت-بيوتيل والإيثانول.وبالمثل، فإن تأثيرات GLA الواقية للكبد ضد إصابة الكبد الناتجة عن السيكلوفوسفاميد والميثوتريكسات تتم من خلال استجابة مضادة للأكسدة تعتمد على Nrf2.استنادًا إلى نسبة الحماية، أظهرت مجموعة AZM + GA + GLA أعلى مستوى من الحماية ضد تلف الكبد التأكسدي الناتج عن AZM وانخفاض تعبير بروتين Nrf2.
يلعب زيادة إنتاج أنواع الأكسجين التفاعلية دورًا رئيسيًا في بدء مسار الإشارات المؤيدة للالتهابات. تقوم هذه المسارات بتنشيط NF-kB، مما يؤدي إلى إنتاج وسطاء مؤيدين للالتهابات والتهاب الكبد اللاحق. ليو وآخرون.أظهرت أن الإجهاد التأكسدي يعزز نشاط NF-kB، مما يؤدي إلى إطلاق السيتوكينات المؤيدة للالتهابات مثل TNF-أظهرت الدراسات أن مواد مثل حمض الفالبرويك، 2-أسيتيل أمينوفلورين، ميثوتريكسات، وثيوأسيتيمايد تزيد من مستويات NF-kB والسيتوكينات المؤيدة للالتهابات، مما يساهم في تلف الكبد الحاد.في دراستنا، أدى العلاج المسبق بـ GA و GLA في الفئران المعالجة بـ AZM إلى تقليل مستويات NF-kB و TNF- بشكل ملحوظ.المستويات، مؤكدةً تأثيراتها الواقية ضد تلف الكبد الناتج عن AZM، ومسلطةً الضوء على خصائصها المضادة للالتهابات المحددة للكبد. وقد استكشفت الدراسات السابقة التأثيرات المضادة للالتهابات لكل من GA وGLA في نماذج مختلفة من إصابة الكبد. على سبيل المثال، أدى علاج GLA إلى تقليل مستويات TNF-a وNF-kB في نموذج الفئران لالتهاب الكبد الحاد الناتج عن السيكلوفوسفاميد والميثوتريكسات.. بالمثل، محمد وحافظأُبلغ في عام 2023 أن علاج GA قلل من TNF-NF-kB، ووسائط التهابية أخرى في إصابة الكبد الناتجة عن الثيوأسيطاميد.
علاوة على ذلك، كشفت هذه الدراسة عن علاقة عكسية كبيرة بين Nrf2 وNF-kB في كبد الفئران المعالجة بـ AZM. وُجد أن تنشيط Nrf2 كان حاسمًا للتأثيرات المضادة للالتهابات لـ GA وGLA. يمكن لـ Nrf2 أن يثبط مباشرة إشارة NF-kB وإنتاج السيتوكينات المؤيدة للالتهابات، مما يمارس السيطرة التنظيمية على سلسلة الالتهابات. وقد أبرزت الدراسات الحية الدور المحوري لـ Nrf2 في إدارة الأمراض الالتهابية عبر أنظمة مختلفة.مسار إشارة Nrf2 يثبط نشاط السيتوكينات ووسائط الالتهاب الأخرى، مما يؤثر على تعبير NF-kB وشبكات أخرى. المشاركة في تنظيم الالتهابات. يدعم هذا المفهوم العديد من الدراسات، بما في ذلك تلك التي أجراها جين وآخرون.محمود وآخرونكي وآخرون، ويونس وآخرونالتي تُظهر أن بعض المركبات النباتية ذات الخصائص المضادة للالتهابات يمكن أن تعزز بشكل فعال Nrf2 وتقلل من NF-kB. بشكل جماعي، قد تساعد GA وGLA وتركيبهما في التخفيف من تلف الكبد الناتج عن AZM من خلال تقليل الإجهاد التأكسدي والالتهاب من خلال تعزيز تعبير Nrf2.
الاستنتاجات
في الدراسة الحالية، حمى GA و GLA الفئران من تلف الكبد الناتج عن AZM. يرتبط هذا الحماية الكبدية ارتباطًا وثيقًا بزيادة تنظيم Nrf2، وهو عامل نسخ أساسي في أنظمة الدفاع الخلوية، والذي يخفف من الإجهاد التأكسدي والالتهاب الناتج عن AZM. من خلال تقليل الإجهاد التأكسدي، يمنع GA و GLA تراكم ROS الذي يمكن أن يتلف خلايا الكبد. بالإضافة إلى ذلك، تساعد خصائصهما المضادة للالتهابات
4 الشكل 3. التحقيقات النسجية المرضية لشرائح الكبد الملونة بصبغة H&E. كانت مجموعات التحكم (A&B) ومجموعات GA (C) ومجموعات GLA (D) جميعها تحتوي على هياكل كبدية طبيعية. (A) تحتوي فصوص الكبد على حواجز بين الفصوص بارزة مع خلايا كبدية متعددة الأضلاع تمتد من الشريان البابي (PA) والوريد المركزي (CV). ). (B&C) تتكون منطقة البوابة (PA) من الشريان الكبدي (A) وقناة الصفراء (B) وفروع الوريد البابي. تحتوي الخلايا الكبدية على سيتوبلازم إيوزينوفيلي ونوى حويصلية عندما تكون مرئية. تصطف الخلايا البطانية المسطحة وخلايا كوبفر (الأسهم السوداء) على المساحات الجيبية (S) بين الخلايا الكبدية ( ). (E) مجموعة AZM لديها وريد بورتال متوسع (سهم سميك)، والعديد من المناطق النخرية (أسهم سوداء)، وتفريغ الخلايا الكبدية، والتنكس الدهني (5). (F) أظهرت مجموعة AZM تضخم القنوات الصفراوية (سهم أحمر)، وريد بورتال متوسع (سهم سميك)، ونخر مع تسرب خلايا التهابية.. (G&H) مجموعة AZM يظهر نخر شديد (أسهم سوداء)، وتفريغ الخلايا الكبدية، وتDegenerations دهنية (5)، وت infiltrations خلايا التهابية (*). (I&J) تشير البنية النسيجية لمجموعة AZM + GA إلى استعادة الهيكل الطبيعي للكبد مع بعض التفريغ، والتDegenerations دهنية (5) لخلايا الكبد. )، ونخر طفيف (أسهم سوداء). (K&L)، تُظهر البنية النسيجية لمجموعة AZM + GLA بقع نخرية في المناطق البابية (أسهم سوداء) الهيكل النسيجي لمجموعة AZM + GA + GLA طبيعي، مع بؤر نادرة من النخر (الأسهم السوداء) والتجويف (5) في المناطق البابية (M & N، ) ومنطقة الوريد المركزي (O، )، بالإضافة إلى تضخم القناة الصفراوية (السهم الأحمر). ومع ذلك، فإن الآليات الدقيقة التي من خلالها تعدل GA وGLA تعبير Nrf2 وتؤثر على تأثيراتها الحامية للكبد تتطلب مزيدًا من التحقيق (الشكل 8).
تحتاج التحقيقات الإضافية إلى تقييم فعاليتها وسلامتها في الحالات السريرية.
الشكل 4. الدرجات النسجية المرضية لـ (A) نخر الخلايا الكبدية، (B) التنكس الدهني، (C) احتقان الوريد البابي، (D) تضخم القناة الصفراوية، و (E) تسرب الخلايا الالتهابية، القيم معبّر عنها كوسيطفترة الثقة لخمس حيوانات في كل مجموعة. تم تحديد الأهمية من خلال اختبار كروسكال-واليس H غير المعلمي تلاه المقارنة الزوجية.; فرق كبير عن مجموعة التحكم؛ ؛ اختلاف كبير عن مجموعة AZM.
الشكل 5. تأثير GA وGLA وتركيبهما على الإجهاد التأكسدي الكبدي في الجرذان المعالجة بـ AZM. تشمل علامات الأكسدة (A) مستوى MDA، (B) نشاط SOD، (C) نشاط CAT، و(D) مستوى GSH. يتم عرض البيانات كمتوسط ± SEM.قمنا أولاً بإجراء تحليل ANOVA الإحصائي، تلاه تحليل توكي بعد الاختبار. قيمة Pيشير إلى فرق كبير عن مجموعة التحكم، بينماتشير إلى فرق كبير عن مجموعة AZM.
الشكل 6. تأثير GA و GLA وتركيبهما على الالتهاب الكبدي في الجرذان المعالجة بـ AZM. (A) مستوى تعبير TNF-تم تحديده في الكبد بواسطة ELISA. (ب) تم تحديد مستوى التعبير عن NF-kB و B-actin في الكبد بواسطة تقنية Western blotting. تم استخدام B-actin كتحكم داخلي. تم تقديم البقع الأصلية في الشكل التكميلية S1 و S2 (ج) التحليل الكمي لبروتين NF-kB. يتم تقديم البيانات كمتوسط ± SEM من ثلاث تجارب مستقلة، تم تنفيذها ثلاث مرات. أ تختلف بشكل ملحوظ عن التحكم عند قيمة p; ب اختلاف كبير عن AZM عند قيمة p.
الشكل 7. (أ) تم تحديد مستوى التعبير عن Nrf2 وB-actin في الكبد بواسطة تقنية الويسترن بلوت. تم استخدام B-actin كتحكم داخلي. تم تقديم الصور الأصلية في الشكل التكميلي S3 وS4 (ب) التحليل الكمي لبروتينات Nrf2. تم تقديم البيانات كمتوسط ± SEM لثلاث تجارب مستقلة، تم تنفيذها ثلاث مرات. أ تختلف بشكل ملحوظ عن التحكم عند-قيمة; ب يختلف بشكل كبير عن AZM عند -قيمة .
الشكل 8. الآلية المقترحة لعمل حمض الجاليك وحمض الجلسيرهيزين. يحمي حمض الجاليك وحمض الجلسيرهيزين الفئران من إصابة الكبد الحادة الناتجة عن AZM من خلال تعديل مسارات الإشارات Nrf2 و NF-kB. هذه التعديلات على مسارات الإشارات Nrf2 و NF-kB تقلل من الإجهاد التأكسدي والالتهاب الناجمين عن AZM. تم إنشاؤه في BioRender، BioRender. https://BioRender.com/q891287.
توفر البيانات
بيان توفر البيانات. تتوفر مجموعات البيانات المستخدمة و/أو التي تم تحليلها خلال الدراسة الحالية من المؤلف المراسل عند الطلب المعقول.
تاريخ الاستلام: 8 ديسمبر 2024؛ تاريخ القبول: 4 مارس 2025
تم النشر عبر الإنترنت: 20 مارس 2025
References
Björnsson, H. K. & Björnsson, E. S. Drug-induced liver injury: Pathogenesis, epidemiology, clinical features, and practical management. Eur. J. Intern. Med. 97, 26-31. https://doi.org/10.1016/j.jim.2021.10.035 (2022).
Clinton, J. W. et al. Drug-Induced liver injury: Highlights and controversies in the recent literature. Drug Saf. 44, 1125-1149. https://doi.org/10.1007/s40264-021-01109-4 (2021).
Jin, C. et al. Treatment responses to Azithromycin and Ciprofloxacin in uncomplicated Salmonella Typhi infection: A comparison of clinical and Microbiological data from a controlled human infection model. PLoS Negl. Trop. Dis. 13, e0007955 (2019).
Bal, A. M. Macrolide antibiotics. (2022).
Zhai, G., Li, M., Wang, Y. & Wu, J. Drug-induced liver disturbance during the treatment of COVID-19. Front. Pharmacol. 12, 719308 (2021).
Firth, A. & Prathapan, P. Azithromycin: The first broad-spectrum therapeutic. Eur. J. Med. Chem. 207, 112739 (2020).
Atli, O., Ilgin, S., Altuntas, H. & Burukoglu, D. Evaluation of Azithromycin induced cardiotoxicity in rats. Int. J. Clin. Exp. Med. 8, 3681 (2015).
Hamza, R. Z. et al. Ascorbic acid ameliorates cardiac and hepatic toxicity induced by azithromycin-etoricoxib drug interaction. Curr. Issues Mol. Biol. 44, 2529-2541(2022).
Li, S. et al. Establishment of a new animal model of azithromycin-induced liver injury and study the molecular pathological change during the process. Hum. Exp. Toxicol. 35, 511-525 (2016).
Ellison, C. A. & Blackwell, S. B. Acute hepatocellular injury associated with Azithromycin. J. Pharm. Pract. 34, 493-496 (2021).
Danica, J. et al. Vanishing bile duct syndrome associated with Azithromycin in a 62-year-old man. Basic. Clin. Pharmacol. Toxicol. 106, 62-65 (2010).
Martinez, M. A. et al. Clinical and histologic features of azithromycin-induced liver injury. Clin. Gastroenterol. Hepatol. 13, 369376 (2015).
Park, H. J., Seo, K. I. & Choi, Y. I. Liver transplantation for azithromycin-induced severe liver injury. Korean J. Transplantation 34, 279-285 (2020).
Choi, Y. I., Moon, H. H., Jo, J. H. & Shin, D. H. Liver transplantation for azithromycin-induced severe liver injury: A case report. Korean J. Transplantation 35, 130 (2021).
Reyes-Gordillo, K., Shah, R. & Muriel, P. Oxidative stress and inflammation in hepatic diseases: Current and future therapy. Oxid. Med. Cell. Longev. 2017 (2017).
Villanueva-Paz, M. et al. Oxidative stress in drug-induced liver injury (DILI): From mechanisms to biomarkers for use in clinical practice. Antioxidants 10, 390 (2021).
Zhang, M. M., Qiao, Y., Ang, E. L. & Zhao, H. Using natural products for drug discovery: The impact of the genomics era. Expert Opin. Drug Discov. 12, 475-487 (2017).
Govea-Salas, M. et al. In the Liver317-322 (Elsevier, 2018).
Aglan, H. A., Ahmed, H. H., El-Toumy, S. A. & Mahmoud, N. S. Gallic acid against hepatocellular carcinoma: An integrated scheme of the potential mechanisms of action from in vivo study. Tumor Biol. 39, 1010428317699127 (2017).
Gao, J., Hu, J., Hu, D. & Yang, X. A role of Gallic acid in oxidative damage diseases: A comprehensive review. Nat. Prod. Commun. 14, 1934578X19874174 (2019).
Jadon, A., Bhadauria, M. & Shukla, S. Protective effect of Terminalia Belerica Roxb. And Gallic acid against carbon tetrachloride induced damage in albino rats. J. Ethnopharmacol. 109, 214-218 (2007).
Latief, U., Husain, H., Mukherjee, D. & Ahmad, R. Hepatoprotective efficacy of Gallic acid during Nitrosodiethylamine-induced liver inflammation in Wistar rats. J. Basic. Appl. Zool. 76, 31-41 (2016).
Reckziegel, P. et al. Antioxidant protection of Gallic acid against toxicity induced by Pb in blood, liver and kidney of rats. Toxicol. Rep. 3, 351-356 (2016).
Feng, R. B., Wang, Y., He, C., Yang, Y. & Wan, J. B. Gallic acid, a natural polyphenol, protects against tert-butyl hydroperoxideinduced hepatotoxicity by activating ERK-Nrf2-Keap1-mediated antioxidative response. Food Chem. Toxicol. 119, 479-488 (2018).
Dutta, M. & Paul, G. Gallic acid protects rat liver mitochondria ex vivo from bisphenol A induced oxidative stress mediated damages. Toxicol. Rep. 6, 578-589 (2019).
Zhou, Y. et al. Gallic acid protects against ethanol-induced hepatocyte necroptosis via an NRF2-dependent mechanism. Toxicol. Vitro. 57, 226-232 (2019).
Hussein, R. M., Anwar, M. M., Farghaly, H. S. & Kandeil, M. A. Gallic acid and ferulic acid protect the liver from thioacetamideinduced fibrosis in rats via differential expression of miR-21, miR-30 and miR-200 and impact on TGF- /Smad3 signaling. Chemico-Biol. Interact. 324, 109098 (2020).
Sundaresan, S. et al. Gallic acid attenuates cadmium mediated cardiac hypertrophic remodelling through upregulation of Nrf2 and PECAM-1signalling in rats. Environ. Toxicol. Pharmacol. 87, 103701 (2021).
Li, X., Sun, R. & Liu, R. Natural products in licorice for the therapy of liver diseases: Progress and future opportunities. Pharmacol. Res. 144, 210-226 (2019).
Shinu, P. et al. Pharmacological features of -glycyrrhetinic acid: A pentacyclic triterpenoid of therapeutic potential. Plants 12, 1086 (2023).
Chen, S., Zou, L., Li, L. & Wu, T. The protective effect of glycyrrhetinic acid on carbon tetrachloride-induced chronic liver fibrosis in mice via upregulation of Nrf2. PLoS One 8, e53662 (2013).
Yang, Y. et al. Protective effects of hepatocyte-specific glycyrrhetic derivatives against carbon tetrachloride-induced liver damage in mice. Bioorg. Chem. 72, 42-50. https://doi.org/10.1016/j.bioorg.2017.03.009 (2017).
Hasan, S. et al. glycyrrhetinic acid alleviates 2 -acetylaminofluorene-induced hepatotoxicity in Wistar rats: Role in hyperproliferation, inflammation and oxidative stress. Hum. Exp. Toxicol. 34, 628-641 (2015).
Mahmoud, A. M. & Al Dera, H. S. 18β-Glycyrrhetinic acid exerts protective effects against cyclophosphamide-induced hepatotoxicity: Potential role of PPAR and Nrf2 upregulation. Genes Nutr. 10, 41. https://doi.org/10.1007/s12263-015-0491-1 (2015).
Mahmoud, A. M., Hussein, O. E., Hozayen, W. G. & Abd El-Twab, S. M. Methotrexate hepatotoxicity is associated with oxidative stress, and down-regulation of PPAR and Nrf2: Protective effect of -Glycyrrhetinic acid. Chem. Biol. Interact. 270, 59-72. https://doi.org/10.1016/j.cbi.2017.04.009 (2017).
Iranshahy, M., Iranshahi, M., Abtahi, S. R. & Karimi, G. The role of nuclear factor erythroid 2-related factor 2 in hepatoprotective activity of natural products: A review. Food Chem. Toxicol. 120, 261-276. https://doi.org/10.1016/j.fct.2018.07.024 (2018).
Paulose, P. et al. Ameliorative effect of vitamin E on Azithromycin induced biochemical and histological changes in liver tissue of rats. Adv. Anim. Vet. Sci. 4, 85-91 (2016).
Ojeaburu, S. & Oriakhi, K. Hepatoprotective, antioxidant and, anti-inflammatory potentials of Gallic acid in carbon tetrachlorideinduced hepatic damage in Wistar rats. Toxicol. Rep. 8, 177-185 (2021).
Bancroft, J. D. & Gamble, M. Theory and Practice of Histological Techniques (Elsevier health sciences, 2008).
Ibrahim Fouad, G. & Ahmed, K. A. Curcumin ameliorates doxorubicin-induced cardiotoxicity and hepatotoxicity via suppressing oxidative stress and modulating iNOS, NF- , and TNF- in rats. Cardiovasc. Toxicol. 22, 152-166 (2022).
Buege, J. A. & Aust, S. D. In Methods in Enzymology 52302-310 (Elsevier, 1978).
Nandi, A. & Chatterjee, I. Assay of superoxide dismutase activity in animal tissues. J. Biosci. 13, 305-315 (1988).
Towbin, H., Staehelin, T. & Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. 76, 4350-4354 (1979).
Usadadia, S., Patel, J., Vihol, P. & Urkude, A. Protective effect of Quercetin on Azithromycin induced hepatotoxicity and nephrotoxicityin Wistar rats. Int. J. Curr. Microbiol. Appl. Sci. 9, 2926-2934 (2020).
Plaa, G. & Zimmerson, H. Evaluation of hepatotoxicity: physiological and biochemical measures of hepatic function. Compr. Toxicol. 9, 97-109 (1997).
Wang, H. et al. Glycyrrhizin and glycyrrhetinic acid inhibits alpha-naphthyl isothiocyanate-induced liver injury and bile acid cycle disruption. Toxicology 386, 133-142 (2017).
Liguori, I. et al. Oxidative stress, aging, and diseases. Clin. Interv Aging, 757-772 (2018).
Jomova, K. et al. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: Chronic diseases and aging. Arch. Toxicol. 97, 2499-2574 (2023).
Singh, H., Prakash, A., Kalia, A. & Majeed, A. B. A. Synergistic hepatoprotective potential of ethanolic extract of Solanum xanthocarpum and Juniperus communis against Paracetamol and Azithromycin induced liver injury in rats. J. Traditional Complement. Med. 6, 370-376 (2016).
Jin, J. et al. Role of Nrf2 activation and NF- Inhibition in valproic acid induced hepatotoxicity and in diammonium glycyrrhizinate induced protection in mice. Food Chem. Toxicol. 73, 95-104 (2014).
Liu, T., Zhang, L., Joo, D. & Sun, S. C. NF-кB signaling in inflammation. Signal. Transduct. Target. Therapy. 2, 1-9 (2017).
Mohamed, E. K. & Hafez, D. M. Gallic acid and Metformin co-administration reduce oxidative stress, apoptosis and inflammation via Fas/caspase-3 and NF- signaling pathways in thioacetamide-induced acute hepatic encephalopathy in rats. BMC Complement. Med. Ther. 23, 265 (2023).
Ahmed, S. M. U., Luo, L., Namani, A., Wang, X. J. & Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Et Biophys. Acta (BBA)-Molecular Basis Disease 1863, 585-597 (2017).
Qi, W. et al. Cardamonin protects against doxorubicin-induced cardiotoxicity in mice by restraining oxidative stress and inflammation associated with Nrf2 signaling. Biomed. Pharmacother 122, 109547 (2020).
Younis, N. S., Elsewedy, H. S., Shehata, T. M. & Mohamed, M. E. Geraniol averts methotrexate-induced acute kidney injury via keap1/Nrf2/HO-1 and MAPK/NF- B pathways. Curr. Issues Mol. Biol. 43, 1741-1755 (2021).
مساهمات المؤلفين
تم تصميم الدراسة بواسطة G.H. و A.H. تم إجراء التجارب والتحليل الإحصائي بواسطة M.A. و A.I. و G.H. تم توفير الموارد والمساعدة المنهجية بواسطة G.H. و A.H. و M.A. و A.I. و A.A. تم كتابة المسودة الأصلية للمخطوطة بواسطة A.H. و G.H. و M.A. و A.I. و A.A. ثم قام جميع المؤلفين بتحرير وقبول النسخة المحدثة من المقال. كل مؤلف قام بمراجعة النسخة المنشورة من المخطوطة ومنح موافقته.
التمويل
تم دعم AA من قبل منحة البذور رقم (24010901156) من جامعة الشارقة.
الإعلانات
المصالح المتنافسة
يعلن المؤلفون عدم وجود مصالح متنافسة.
معلومات إضافية
معلومات إضافية. تحتوي النسخة عبر الإنترنت على مواد إضافية متاحة على https://doi.org/1 0.1038/s41598-025-93120-3.
يجب توجيه المراسلات والطلبات للحصول على المواد إلى A.A.H. أو A.A.
معلومات إعادة الطبع والتصاريح متاحة على www.nature.com/reprints.
ملاحظة الناشر. تظل Springer Nature محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
إدارة الصحة بأبو قرقاص، مديرية الصحة بالمنيا، وزارة الصحة، المنيا 61611، مصر. قسم علم الأدوية والسموم، كلية الصيدلة، جامعة المنيا، المنيا، مصر. قسم البيولوجيا، الهيئة الوطنية لمراقبة الأدوية والبحوث، الجيزة 12611، مصر. كلية الطب، جامعة الشارقة، الشارقة 27272، الإمارات العربية المتحدة. البريد الإلكتروني: alaa17mm@gmail.com; a.amin@sharjah.ac.ae
Gallic and glycyrrhetinic acids prevent azithromycin-induced liver damage in rats by mitigating oxidative stress and inflammation
Marwa Ahmed Mohamed Ali , Asmaa Ibrahim Matouk , Alaaeldin Ahmed Hamza , Amr Amin & Gehan Hussein Heeba
Natural products like gallic acid (GA), a phenolic compound, and glycyrrhetinic acid (GLA), a pentacyclic triterpene, have been shown to exhibit antioxidant, ant-inflammatory, and hepatoprotective properties. This study aims to investigate the protective effects of GA, GLA and their combination and to explore their underlying mechanisms against acute liver damage induced by azithromycin (AZM) in rats. Seven groups of male Wistar rats were used namely control, GA, GLA, AZM, AZM + GA, AZM + GLA, and AZM + GA + GLA groups. We treated the rats for 21 days, administering GA and GLA at one week prior to AZM ( ). Serum levels of aspartate aminotransferase, alanine aminotransferase, and alkaline phosphatase in rats AZM-induced and pre-treated with of GA, GLA, and combination therapy were much lower than those in the AZM group Elevated levels of glutathione, catalase, and superoxide dismutase in preventive rats demonstrated that GA, GLA, and their combinations notably reduced AZM-induced malondialdehyde levels, an oxidative stress marker, improving the antioxidant system in those groups. Furthermore, GA, GLA, and their combinations reduced pro-inflammatory cytokines, such as tumor necrosis factor- and nuclear factor kappa beta (NF-kB), thus reversing hepatic inflammation. We also discovered that AZM down-regulated nuclear factor erythroid 2-related factor 2 (Nrf2), and that its normal levels were restored post treatments with GA, GLA, and their combination suggesting a Nrf2 signaling pathway-mediated prevention of AZM-induced liver damage. In conclusion, GA, GLA, and their combination protected rats against AZMinduced liver injury through their demonstrated ability to lower oxidative stress and inflammation by preventing downregulation of Nrf2 and upregulation of NF-kB. After determining their efficacy and safety in clinical settings in future studies, GA and GLA could potentially make useful therapeutic drugs to mitigate AZM-induced hepatotoxicity.
Keywords Azithromycin, Gallic acid, Glycyrrhetinic acids, Liver damage, Nrf2, NF-kB
Drug-induced liver injury (DILI), while uncommon, can lead to potentially life-threatening consequences including elevated liver enzymes, acute liver failure, and even mortality . Azithromycin (AZM), a macrolide antibiotic, is among the top ten medications causing DILI . It effectively treats genitourinary infections, enteric diseases such as typhoid, and respiratory conditions like pneumonia , indicating its broad-spectrum antibacterial activity against both gram-positive and gram-negative bacteria. Moreover, AZM has demonstrated therapeutic potential in manging cure for a wide range of illnesses, including viral infections such as COVID-19 4,5,6.
Scientific research has undoubtedly established two recognized adverse effects of AZM administration: hepatotoxicity and cardiotoxicity . Li et al. conducted an extensive study on hepatotoxicity in animals, and Ellison and Blackwell observed it in humans. Clinical studies have confirmed that AZM can have serious negative effects on both adults and children, such as cholestasis, jaundice, and liver dysfunction . Research has documented cases of acute cholestatic hepatitis developed within the first three weeks of starting AZM medication . According to Martinez et al. , liver damage typically appears within one to three weeks after
starting medication. While AZM often causes mild liver damage, there are cases when it can result in significant liver damage, requiring a liver transplant and potentially leading to abrupt liver failure . Recognizing these potential risks and maintaining a diligent attitude towards detecting any indications of liver damage in patients receiving AZM treatment, is of utmost importance. We do not fully understand how AZM affects hepatic tissue, but we hypothesize that oxidative stress and inflammation play a significant role.
Currently, there are limited therapeutic and preventive options for liver disease, despite its global prevalence and significant contribution to mortality rate worldwide. People widely recognize oxidative stress and inflammation as major factors in liver disease development . Researchers and healthcare professionals are increasingly focusing on naturally occurring polyphenol compounds to reduce the harmful effects of various drugs and environmental pollutants . Recently, these compounds have gained significant attention thanks to their diverse pharmacological properties and therapeutic potential . Gallic acid (GA), a naturally occurring phenolic compound with potent antioxidant properties, is characterized by its relatively low molecular weight and is chemically known as 3,4,5-trihydroxybenzoic acid . GA has several biochemical and pharmacological characteristics, such as anti-inflammatory, antibacterial, antioxidant, cardioprotective, antiviral, and anticancer effects . This natural phenolic compound is found in a variety of plant-based sources including apple peels, walnuts, mangoes, grapes, and green tea. GA exhibits significant hepatoprotective properties , demonstrating its efficacy in combating different liver-damaging agents including carbon tetrachloride (CCL4) , nitroso diethylamine , lead , tert-butyl hydroperoxide , bisphenol , ethanol , thioacetamide , and cadmium . GA’s biological activities, particularly its antioxidant properties, protect human cells and tissues from oxidative stress-induced damage . Glycyrrhiza species, commonly known as licorice, is the primary source of pentacyclic triterpene glycoside . The root of licorice is the main source of glycyrrhetinic acid (GLA), a bioactive pentacyclic triterpene . Licorice yields two isomers of glycyrrhetinic acid: -glycyrrhetinic acid and 18a-glycyrrhetinic acid . These compounds exhibit a range of pharmacological characteristics including antiviral, anti-inflammatory, antifibrotic, and antioxidant activities . Recent studies have shown that GLA can protect liver cells from damage induced by various toxins such as CCL4 , 2-acetylaminofluorene , cyclophosphamide , methotrexate , and alpha-naphthyl isothiocyanate . The findings indicate that GLA possesses hepatoprotective characteristics and can mitigate liver damage induced by various harmful chemicals.
Iranshahy et al. showed that nuclear factor Erythroid 2-Related Factor 2 (Nrf2) plays a critical role in protecting the liver from drug-induced toxicity. Their study revealed that Nrf2 regulates the expression of genes dependent on antioxidant response elements (ARE), which are essential for maintaining cellular resilience against oxidative stress. The activation of Nrf2 triggers the transcription of antioxidant and cytoprotective enzymes, including SOD, catalase, heme oxygenase 1, NAD(P) H, quinone oxidoreductase 1, and NAD(P) , thereby enhancing the cell’s defense mechanisms.
Many natural products have demonstrated hepatoprotective properties . In this study, we focused on two main classes of bioactive compounds: Phenolic acids, such as GA, and triterpenoid compounds, such as GLA . Notably, Nrf2 activation appears to be a key mechanism underlying the hepatoprotective effects of these natural compounds . Licorice-derived compounds like GLA, when used in combination with other therapeutic agents, have been shown to enhance liver protection against DILI . Accordingly, this study aims to evaluate the protective effects of AZM, GA, GLA, and their combinations in mitigating acute hepatotoxicity in male Wistar rats. The present findings suggested that GA, GLA, and their co-administration protect the liver by modulating oxidative stress and inflammatory pathways.
Materials and methods
Chemicals
Thiobarbituric acid, pyrogallol, and 18β1-glycyrrhetinic acid (CAT# G10105) and gallic acid (CAT# G7384) were obtained from Sigma-Aldrich (St. Louis, MO, USA). The AZM (Xithrone, 500 mg , 211515) was acquired from Amoun, located in El Obour City, Qalyubia, Egypt. The inquiry selected the highest analytical grade for all other substances.
Animals
The study included a sample of 42 male albino Wistar rats, with weights ranging from 180 to 220 g . These rats were obtained from the experimental animal house at Nahda University, Beni Suef. The rats were put into 7 experimental groups using random assignment. To calculate the effect size, we utilized previously published data from Paulose et al. , with a power of 0.9 and a significance threshold of 0.05 . Before commencing the experimental procedures, the rats were provided with one week to adapt to the laboratory setting. The subjects were kept in plastic enclosures with regulated humidity ( ), temperature ( ), and a light-dark cycle of 12 h each. The rats were given a standard commercial diet and had unrestricted access to water.
Ethics declaration
Animal experiments were conducted in compliance with international standards for the care and utilization of laboratory animals (National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals, 8th edition. Washington (DC): National Academies Press (US); 2011). All experimental protocols were approved by the Research Ethics Committee for the care and use of laboratory animals at Minia University, Faculty of Pharmacy (ethical approval number: 2301004). In addition, all experimental protocols were reported following the relevant guidelines and regulations reported in ARRIVE guidelines 2.0. (https://arriveguidelines.org).
An experiment’s design
The experiment involved randomly assigning rats to different groups. The study involved seven groups, each containing six rats. The control group of rats received a solution of water and tween 80 drops ( day ) for 21 days. The rats in the GA group received a daily dosage of GA orally ( day, dissolved in water containing Tween 80 droplets) for 21 days. Rats in the GLA group received GLA orally ( day ) in a solution of water and Tween 80 drops for 21 days. The AZM group was administered a solution of water mixed with Tween 80 drops orally ( day ) for 21 days. Starting on the eighth day, these rats were also given AZM orally at a dose of day in a solution of water and Tween 80 drops for 14 days. The rats in the GA + AZM group were administered oral GA ( day ) for 21 days. On the eighth day, the rats received oral AZM day , in a solution of water and Tween 80 drops) for a duration of 14 days. The group of rats known as GLA + AZM were administered GLA orally at a dosage of body weight for 21 days. On the eighth day, they received oral administration of AZM at a dosage of day in a solution of water and Tween 80 drops for 14 days. The group of rats labeled as GA + GLA + AZM were administered both GA and GLA orally for 21 days, with a dosage of body weight for each substance. Starting on the eighth day, we administered AZM orally at a daily dose of for 14 days.
The doses of GLA and GA were chosen according to prior research that has shown their hepatoprotective effects against different substances . In addition, a liver injury model generated by AZM was established according to pilot experimental trials as well a as to previously described by Paulose et al. .
Sample collections and preparations
The rats were anaesthetized using a ketamine/xylazine ( , i.p.) 24 h after the final treatment. Blood samples were then taken from the retro-orbital plexus. The rats were euthanized through cervical dislocation following blood extraction. Subsequently, the liver was promptly extracted, rinsed with cold normal saline, and weighed. To conduct histological studies, a section of liver tissue was promptly immersed in a buffered formalin solution. The liver tissues were homogenized using a Tris-HCL buffer at a ratio of (liver tissue to buffer) with a concentration of 150 mM and a pH of 7.4. The homogenate was partitioned into vials and stored at a temperature of . Using appropriate buffers, dilutions were prepared from the latter to test the amounts of GSH, LP product MDA, total proteins, and SOD and CAT activity. Centrifuge tubes were used to collect blood, which was then subjected to centrifugation for 20 min at a speed of 3000 revolutions per minute in a refrigerated centrifuge set at a temperature of . This process was carried out to extract serum. Serum samples were utilized for the measurement of the levels of ALT, AST, ALP and albumin.
Biochemical and histopathological assays
Assessment of liver toxicity biomarkers
Assessment of liver injury biomarkers was conducted by measuring the levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), and albumin. Commercial kits from BIOMED in Al Asher Men Ramadan, Cairo, Egypt, were used, following the manufacturer’s instructions.
Histopathological analysis using haematoxylin and eosin staining
According to the procedure outlined in Bancroft’s methodology, the preserved samples were clipped, cleaned, dried using progressively stronger alcohol solutions, treated with xylene to remove impurities, encased in paraffin, and then cut into sections measuring micrometres in thickness. The sections were subsequently stained using the hematoxylin and eosin (H&E) method. Histological evaluations were conducted by analyzing five distinct regions of each liver sample at a random 200x magnification. The evaluations were centred around the identification of histological alterations, including hydropic degeneration, localized necrosis, vascular congestion, and inflammatory cell infiltrations. The magnitude of these alterations was assessed using a grading method employed in a prior investigation conducted by Ibrahim Fouad and Ahmed . The grading system had a range of values from 0 to 3 , and each value had a specific connotation. The grades were determined as a percentage to measure the extent of the detected histopathological alterations. Grades 0 (zero) signified a negative outcome, whereas Grades , and denoted mild, moderate, and severe modifications, respectively.
Measurement of malondialdehyde (MDA) concentration
Thiobarbituric acid (TBA) assay described by Buege and Aust was employed to measure the concentration of malondialdehyde (MDA) in the liver tissue homogenates, where MDA reacts with TBA in acidic medium forming a pink colored adduct measured spectrophotometrically at 535 nm . The test was carried out in the following manner: First, prepare (TBA-TCA-HCl reagent (chromagen)) by dissolving 15 g of trichloroacetic acid TCA ( final) and 375 mg of TBA ( final) in , followed by addition of 2.08 ml of concentrated final). Then, the mixture was heated to in a water bath (to help dissolution of TBA), then the volume completed to 100 ml with distilled , and covered during prepartion to prevent change in concentration. The reaction was created by mixing 0.5 mL of liver homogenate, 0.5 ml of distilled water and 2 ml of TBA/TCA/ HCl reagent. The solution was subjected to thermal treatment in a water bath for 15 min . Following the chilling process, allow cooling and centrifugation at a speed of 5000 revolutions per minute for 10 min to isolate the colored supernatant layer. The pink product’s absorbance was quantified at 535 using a UVvisible spectrophotometer, with a blank sample for comparison. The concentration of MDA in the liver tissue was quantified as protein. By employing this procedure, the concentration of MDA was assessed in the liver tissue homogenates, serving as an indicator of lipid peroxidation levels.
Measurement of superoxide dismutase (SOD) activity
The level of superoxide dismutase (SOD) activity in liver homogenate was determined using the method outlined by Nandi and Chatterjee . This technique relies on the capacity of SOD to hinder the spontaneous oxidation of pyrogallol under alkaline conditions. Below is a concise overview of the process: of the liver sample was added to 2 mL of a solution containing 20 mmol Tris-HCL buffer (containing 1 Mm EDTA at a pH of 8.2). The reaction was started by introducing of a recently generated 15 mM pyrogallol solution. The absorbance at a wavelength of 420 nm was continually measured for a duration of 2 min , with measurements obtained during the time interval of 1 to 3 min . The results were quantified as tissue homogenate units per milligram of protein. A single unit of SOD activity is determined as the quantity of enzyme needed to reduce the auto-oxidation of pyrogallol by . The results were expressed as tissue homogenate units per milligram of protein.
Measurement of glutathione content and catalase activity
The glutathione content (GSH) and catalase activity in the liver homogenate were measured using Bio diagnostic kits obtained from 29 Tahreer St., Dokki, Giza, Egypt. The kits utilized were Cat. No. GR 2511 for glutathione (GSH) and Cat. No. CA 2517 for catalase. The measurements were carried out according to the manufacturer’s instructions.
Biuret reagent using Spetcrum-diagnostics kit obtained from Obour city industrial area, Cairo, Egypt, was employed to determine the total protein content of the liver tissues. The kit utilized was Cat. No. 310,001 . Measurements were taken using a PerkinElmer Lambda 25 UV/VIS spectrophotometer and reported as absorbance readings.
Western blot analysis
The tissues were crushed into a uniform mixture using Tris lysis solution that consisted of Triton X-100, and 50 mM Tris pH 7.4. Additionally, Protease Inhibitor Cocktail (Biospes, China) was included. The process of homogenization was conducted for 30 min at a temperature of . The residual tissue was eliminated using centrifugation at a speed of 10,000 revolutions per minute for 10 min at a temperature of . The protein concentration was subsequently measured using a BCA Protein Assay Kit (Thermo Fisher Scientific, USA) by the instructions provided by the manufacturer. Subsequently, equal quantities of protein ( 60 mg of total protein in each lane) that had undergone separation using 10% SDS-polyacrylamide gel electrophoresis were deposited onto a PVDF membrane (Millipore, Merk, USA) . The membranes were obstructed with a solution containing non-fat milk in TBST buffer for 1 h at ambient temperature. Subsequently, the membranes underwent a washing process and were then subjected to overnight incubation at a temperature of with the primary antibodies: anti-Nrf2 (Biospes, China; dilution 1:200; CAT# YPA1865), anti-NF-B p65 (Cell Signaling Technologies; dilution 1:1000, CAT# 8242), or anti-actin (Cell Signaling Technologies; dilution 1:1000, CAT# 7074). After undergoing another round of washing, the membranes were then exposed to the suitable alkaline phosphatase-conjugated secondary antibody (1:5000, Novus Biologicals, LLC, Littleton, CO, USA) for 1 h . The bands were observed using the BCIP/NBT substrate detection Kit from Genemed Biotechnologies, Inc., located in California, USA. Actin served as an internal reference for protein loading, and the resulting bands were compared to actin using Fiji ImageJ (2.16.0) software, http://imagej.net/software/fiji/ (NIH, USA).
Quantification of Proinflammatory cytokine, TNF-
The levels of TNF- in the liver were quantified using ELISA kits obtained from Elabscience (CAT# E-EL-R0019, 14780 Memorial Drive, Suite 108, Houston, Texas, 77079, USA), according to the manufacturer’s instructions.
Statistical analysis
The statistical study was conducted using IBM’s SPSS software, (version 25.0, Chicago, IL, USA). The graphs were made using GraphPad Prism 9 software (San Diego, CA, USA). The mean ± standard error means (SEM) were employed to represent the data. The data’s normal distribution was evaluated using the Shapiro-Wilk test. A one-way analysis of variance (ANOVA) was performed, followed by Tukey’s post-hoc analysis. P value is considered a significant difference.
Results
GA, GLA, and their combination improved body weight gain and liver weight in AZM-treated rats
Figure 1 shows the experimental groups’ body weight gain, liver weight, and liver index values. Administration of AZM to rats significantly reduced body weight gain and liver weight ( ), but no changes was detected in the liver index. However, administering AZM concurrently with GA, GLA, and their combination significantly increased the rats’ indices, indicating a very efficient intensification of these indicators.
GA, GLA, and their combination attenuated liver injury in AZM-treated rats
After receiving AZM, the rats’ blood serum levels of ALT, AST, and ALP significantly increased. Specifically, there was an approximately increase in ALT, a increase in AST, and a increase in ALP when compared to the control group of rats. Serum albumin levels decreased significantly by compared to the control group ( ) (Fig. 2). The AZM + GA group demonstrated a significant decrease in serum ALT, AST, and ALP activity compared to the ASM group, by , and , respectively. In comparison to the AZM group, the AZM + GLA group exhibited significantly lower serum levels of ALT, AST, and ALP ( , and , respectively). Furthermore, we observed a significant reduction in serum ALT, AST, and ALP values in the AZM + GA + GLA group. When compared to the AZM group, the levels of ALT, AST, and ALP dropped by , and , respectively. Serum albumin levels in the AZM + GA,
Fig. 1. Effect of GA, GLA, and their combination on (A) body weight gain, (B) liver weight, and (C) liver index in AZM-treated rats. The results are presented as the mean ± SEM ( ). A one-way ANOVA was utilized, followed by Tukey’s post-hoc analysis with a significant threshold of . ; significantly different from the control group; ; significantly different from the AZM group. Where GA, GLA, and AZM denotes gallic acid, glycerihetenic acid, as well as azithromycin respectively.
AZM + GLA, and AZM + GA + GLA groups were considerably higher than in the AZM group ( , and , respectively) (Fig. 2D). This data demonstrates that the AZM + GA + GLA group is the most protected against AZM-induced liver damage.
GA, GLA, and their combination alleviated the histological changes in the livers of AZMtreated rats
The histopathological study of liver tissues confirmed the biochemical assay results. The control, GA, and GLA groups all had typical hepatic structures, as shown in Fig. 3A&B for the control group, and in Fig. 3C&D for the GA and GLA groups. The hepatic structure included classic hepatic cords, centrally positioned veins (CV), and a portal region (PA) with intact blood sinusoids. The livers of rats given AZM showed severe necrosis and fatty degeneration. Both the portal (Fig. 3E, F, and G) and central areas (H) displayed vacuolated cytoplasm and necrosis. The AZM sections showed proliferating bile ducts, inflammatory cell infiltration, and significant portal vein congestion. Figure 3I&J shows mild necrosis and vacuolization in the portal area following GA and AZM treatment. Rat liver sections treated with AZM and GLA showed focal cellular necrosis, mild inflammation, and vacuolated cytoplasm in the portal region (Fig. 3K & L). The hepatic cells in the portal and central venous areas of rat liver slices treated with AZM and GA with GLA retained their structural integrity. However, a few hepatocytes contained vacuoles and minor foci of necrosis, as well as mildly proliferating bile ducts (Fig. 3M, N, and O ). Figure 4 shows that AZM treatment significantly increased the liver histopathology score compared to the normal group ( ). However, in the , and groups, the liver histopathology score was significantly lower than in the AZM treatment.
GA, GLA, and their combination attenuated oxidative stress in the livers of AZM-treated rats In the AZM group, the amount of MDA in the liver rose by , but SOD and CAT activity dropped by and , respectively, compared to the control group (Fig. 5). Compared to the AZM group, the AZM + GA, AZM + GLA, and AZM + GA + GLA groups had significantly lower hepatic MDA contents of , and , respectively. However, compared to the AZM group, these groups had considerably higher hepatic SOD activity by , and , as well as higher CAT activity by , and , respectively (Fig. 5). On the other hand, the AZM group’s liver GSH content decreased by compared to the control group. The , and AZM + GA + GLA groups had considerably higher hepatic GSH levels than the AZM group (367.26, 322.46%, and , respectively). This data demonstrates that the AZM + GA + GLA group is the most protected against AZM-induced liver damage.
GA, GLA, and their combination reduced inflammation in the livers of AZM-treated rats
GA and GLA’s anti-inflammatory effects in rats treated with AZM were evaluated by assessing inflammatory markers like TNF- and NF- kB (Fig. 6). The treatment with AZM raised the levels of TNF- and NF- kB proteins in the liver by and , respectively, compared to the control group ( p -value ). On the other hand, hepatic TNF- and NF-kB expression levels were significantly decreased after treatment of animals with GA, GLA, and GA+GLA by about and and , and and , respectively, compared to the AZM at p-value . This data demonstrates that the AZM + GA + GLA group is the most protected against AZM-induced hepatic inflammation.
Fig. 2. Effect of GA, GLA, and their combination on AZM-induced liver toxicity in rats. The markers include (A) serum ALT activity, (B) serum AST activity, (C) serum ALP activity, and (D) serum albumin level. The data is displayed as the mean SEM . We performed the ANOVA statistical analysis first, followed by Tukey’s post-hoc analysis. A indicates a significant difference from the control group, while b indicates a significant difference from the AZM group.
GA, GLA, and their combination attenuated the suppression of Nrf2 expression in the livers of AZM-treated rats
Western blot analysis revealed that the amount of Nrf2 protein in the livers of rats that were given AZM was lower than in the control group (Fig. 7B). However, after pretreatment with GA, GLA, or GA + GLA, the Nrf2 expression levels in the liver of AZM-treated rats significantly increased by approximately , , and , respectively, in comparison to the AZM group. (Figs. 7A and B).
Discussion
Drug-induced acute liver injury is a major global health issue, often resulting in liver failure, the need for transplantation, and even death. To address this critical issue, the development and implementation of effective therapeutic interventions are urgently needed. Oxidative tissue injury and inflammation are key mechanisms driving the pathogenesis of acute liver injury . Therefore, novel compounds with antioxidant and anti-inflammatory properties hold promise in protecting against liver dysfunction. In the present study, we investigated the potential health benefits of GA, GLA, and their combination in mitigating AZM-induced acute liver toxicity in rats. Our findings demonstrated that these compounds effectively alleviated oxidative stress, reduced inflammation, and prevented subsequent liver tissue damage.
Some drugs, particularly AZM, have a high risk of acute liver injury . In our study, rats treated with AZM alone exhibited weight loss and reduced liver weight, indicative of systemic toxicity and metabolic disruption. In contrast, co-treatment with GA, GLA, or their combination led to increased body and liver weights, suggesting
that these compounds enhance liver metabolism and detoxification processes. Previous studies have reported that a routinely used antibiotic, like AZM often cause acute liver injury . Animal studies have demonstrated that AZM-induced acute liver damage is associated with elevated circulating liver biomarkers, impaired liver function, and the development of histopathological alterations including hepatic degeneration, fatty changes, sinusoidal congestion, and inflammatory cell infiltration . Similarly, we discovered that treating Wistar rats AZM at a dose of caused severe liver injury reflected in high levels of ALT, AST, and ALP. In our experimental model, elevated liver biomarkers were correlated with significant morphological abnormalities, including severe necrosis, fatty degeneration, proliferating bile ducts, and inflammatory cell infiltration. These observed changes in liver damage and structure align with findings from previous studies . Hepatocyte necrosis and degeneration precede membrane disruption, resulting in the release of hepatic biomarkers into the bloodstream and their subsequent elevation in blood levels . Thus, under hepatic stress conditions, elevated serum levels of hepatic enzyme markers serve as a sensitive indicator of increased hepatocyte membrane permeability and cellular damage . In addition to hepatic damage, AZM-treated rats showed hepatic dysfunction, as evidenced by a reduced serum albumin levels. Our results demonstrate that GA, GLA, and their combinations confer protection against AZM-induced liver injury. Histological analysis confirmed these protective effects, as evidenced by the maintenance of normal liver enzyme levels (ALT, AST, and ALP) and albumin, alongside the preservation of liver tissue integrity. Different studies have revealed that GA provided hepatoprotective effects against damage induced by tert-butyl hydroperoxide , bisphenol , ethanol , and thioacetamide . Similarly, GLA has been shown to protect liver cells from toxins such as carbon tetrachloride, 2 -acetylaminofluorene, cyclophosphamide, methotrexate, and alpha-naphthyl isothiocyanate . In this study, the AZM + GA + GLA group exhibits the highest level of protection against AZM-induced liver injury, as evidenced by the percentage of protection observed.
Oxidative stress and the antioxidant defense system play a critical role in drug-induced acute liver injury . Under normal conditions, the antioxidant defense system – comprising SOD, CAT, and GSH – effectiviely neutralizes free radicals generated during the metabolism of toxic substances and medications . Oxidative stress arises when there is an imbalance between free radicals and antioxidants are out of equilibrium . Numerous investigations have demonstrated that AZM induces the production of ROS, leading to increased levels of lipid peroxidation products (MDA) and a reduction in antioxidant levels, including CAT, SOD, and GSH in the hearts and livers of treated animals . Our findings revealed that AZM increased oxidative stress, as evidenced by markedly elevated MDA levels. Furthermore, we observed a consistent reduction in the activity of CAT and SOD, as well as decreased GSH concentrations, aligning with the results reported by Singh et al. . The present study revealed that GA, GLA, and their combinations effectively increased antioxidant levels (GSH, CAT, and SOD) and reduced lipid peroxidation products (MDA) in liver tissue of rats treated with of AZM, highlighting their potentialantioxidant properties. For instance, GA has been shown to protect the liver against oxidative damage induced by chemicals like lead , thioacetamide , and carbon tetrachloride . Similarly, GLA demonstrated the ability to scavenge ROS and mitigate oxidative stress in liver injury models involving valproic acid , cyclophosphamide , and methotrexate . The percentage of protection further confirmed that the AZM group exhibited the highest protection against AZM-induced liver damage.
Analyzing the Nrf2-mediated pathway provides insights into the mechanisms underlying the antioxidant capabilities of GA and GLA. Both GA and GLA upregulate Nrf2, leading to enhanced expression and activity of antioxidant defense enzymes. This reduces the level of reactive oxidants and mitigate their associated cellular damage. Pretreatment of rats with GA, GLA, or their combination reversed the significant decline in Nrf2 protein expression in liver tissue induced by AZM. Consequently, upregulating Nrf2 enhances the production of antioxidant defense enzymes, thereby reducing reactive oxidants and minimizing cellular injury. Under oxidative stress, Nrf2 translocates to the nucleus and binds to the antioxidant response element (ARE), promoting the expression of enzymes involved in the synthesis of SOD, CAT, and glutathione . Researchers have demonstrated that GA modulates the Nrf2 signaling pathway, offering protection against liver damage induced by tert-butyl hydroperoxide and ethanol . Similarly, GLA’s hepatoprotective effects against cyclophosphamide- and methotrexate-induced liver injury are mediated through the Nrf2-dependent antioxidant response . Based on the percentage of protection, the AZM + GA + GLA group exhibited the highest level of protection against AZM-induced oxidative liver oxidative damage and the decline in Nrf2 protein expression.
Increased ROS generation play a major role in initiating pro-inflammatory signaling pathway. These pathways activate NF-kB, leading to the production of proinflammatory mediators and subsequent liver inflammation. Liu et al. demonstrated that oxidative stress enhances NF- kB activity, resulting in the release of proinflammatory cytokines such as TNF- . Studies have shown that substances like Valproic acid, 2-acetylaminofluorene, methotrexate, and thioacetamide increase NF-kB and proinflammatory cytokine levels, contributing to acute liver damage . In our study, pre-treatment with GA and GLA in AZM-treated rats significantly reduced levels of NF-kB and TNF- levels, confirming their protective effects against AZM-induced liver damage and highlighting their liver-specific anti-inflammatory properties. Previous studies have explored the antiinflammatory effects of GA and GLA in various liver injury models. For instance, GLA treatment reduced TNF-a and NF-kB levels in rat model of cyclophosphamide- and methotrexate-induced acute hepatitis . Similarly, Mohamed and Hafez reported in 2023 that GA treatment reduced TNF- , NF-kB, and other inflammatory mediators in thioacetamide-induced liver injury.
Furthermore, this study revealed a significant inverse relationship between Nrf2 and NF-kB in the livers of AZM-treated rats. The activation of Nrf2 was found to be crucial for the anti-inflammatory effects of GA and GLA. Nrf2 can directly inhibit NF-kB signaling and the production of pro-inflammatory cytokines, thereby exerting regulatory control over the inflammatory cascade. In vivo studies have highlighted the pivotal role of Nrf2 in managing inflammatory diseases across various systems . The Nrf2 signaling pathway suppresses the activity of cytokines and other inflammatory mediators, influencing NF-kB expression and other networks
involved in inflammation regulations . This concept is supported by several studies, including those by Jin et al. , Mahmoud et al. , Qi et al. , and Younis et al. , which demonstrate that certain phytochemicals with anti-inflammatory properties can effectively up-regulate Nrf2 and downregulate NF-kB. Collectively, GA and GLA and their combination may help mitigate AZM-induced liver damage by reducing the oxidative stress and inflammation through the upregulation of Nrf2 expression.
Conclusions
In the present study, GA and GLA protected rats from AZM-induced liver damage. This hepatoprotection is closely associated with the upregulation of Nrf2, a transcription factor integral to cellular defence systems, which mitigates oxidative stress and inflammation caused by AZM. By reducing oxidative stress, GA and GLA prevent the accumulation of ROS that can damage liver cells. Additionally, their anti-inflammatory properties help
4Fig. 3. Histopathological investigations of liver sections stained with H&E. Control groups (A&B), GA groups (C), and GLA groups (D) all had normal liver structures. (A) The liver’s lobules have prominent interlobular septa with polygonal hepatocytes extending from the PA and CV ( ). (B&C) The portal region (PA) is comprised of the hepatic artery (A), the bile duct (B), and portal vein branches. Hepatocytes have eosinophilic cytoplasm and vesicular nuclei when they are visible. Flat endothelial cells and Kupffer cells (black arrows) line the sinusoidal spaces (S) between hepatocytes ( ). (E) The AZM group has a dilated portal vein (thick arrow), numerous necrotic areas (black arrows), hepatocyte vacuolization, and fatty degeneration (5). (F) The AZM group showed bile duct hyperplasia (red arrow), dilated portal vein (thick arrow), and necrosis with inflammatory cell infiltrations . (G&H) The AZM group exhibits severe necrosis (black arrows), hepatocyte vacuolization, fatty degenerations (5), and inflammatory cell infiltrations (*). (I&J) The histological structure of the AZM + GA group indicates a recovery to normal liver architecture along with some vacuolization, fatty degenerations (5) of the liver cells ( ), and minimal necrosis (black arrows). (K&L), The AZM + GLA group’s histological structure shows necrotic patches in the portal areas (black arrows) . The histological structure of the AZM + GA + GLA group is normal, with rare foci of necrosis (black arrows) and vacuolization (5) in the portal areas (M & N, ) and central vein area (O, ), as well as bile duct hyperplasia (red arrow).
counteract the inflammatory response triggered by AZM. However, the precise mechanisms though which GA and GLA modulate Nrf2 expression and exert their hepatoprotective effects require further investigation (Fig. 8).
Additional investigations are necessary to evaluate their efficacy and safety in clinical situations.
Fig. 4. Histopathological scores for (A) necrosis of hepatocytes, (B) fatty degeneration, (C) congested portal vein, (D) bile duct hyperplasia, and (E) inflammatory cell infiltrations, Values expressed as median confidence interval for five animals in each group. Significance was determined by the non-parametric Kruskal-Wallis H test flowed by pairwise comparison:. ; significantly difference from the control group; ; significantly difference from the AZM group.
Fig. 5. Effect of GA, GLA, and their combination on the hepatic oxidative stress in AZM-treated rats. The oxidative markers include (A) MDA level, (B) SOD activity, (C) CAT activity, and (D) GSH level. The data is displayed as the mean ± SEM . We performed the ANOVA statistical analysis first, followed by Tukey’s post-hoc analysis. a P value indicates a significant difference from the control group, while indicates a significant difference from the AZM group.
Fig. 6. Effect of GA, GLA and their combination on the hepatic inflammation in AZM-treated rats. (A) The expression level of TNF- in the liver was determined by ELISA. (B) The expression level of NF- kB and B-actin in the liver was determined by western blotting. B-actin was used as an internal control. Original blots are presented in supplementary figure S1 and S2 (C) Quantitative analysis of the Nf-kB protein. Data are presented as the mean ± SEM of three independent experiments, performed in triplicate. a Significantly different from the control at a p -value ; b Significantly different from AZM at a p -value .
Fig. 7. (A) The expression level of Nrf2 and B-actin in the liver was determined by western blotting. B-actin was used as an internal control. Original blots are presented in supplementary figure S3 and S4 (B) Quantitative analysis of the Nrf2 proteins. Data are presented as the mean ± SEM of three independent experiments, performed in triplicate. a Significantly different from the control at a -value ; b Significantly different from AZM at a -value .
Fig. 8. The proposed mechanism of gallic acid and glycyrrhetinic acid action. Gallic acid and glycyrrhetinic acid protect rats from AZM-induced acute liver injury by modifying the Nrf2 and NF-kB signaling pathways. These modifying the Nrf2 and NF-kB signaling pathways reduce oxidative stress and inflammation caused by AZM. Created in BioRender, BioRender. https://BioRender.com/q891287.
Data availability
Data Availability StatementThe datasets used and/or analysed during the current study available from the corresponding author on reasonable request.
Received: 8 December 2024; Accepted: 4 March 2025
Published online: 20 March 2025
References
Björnsson, H. K. & Björnsson, E. S. Drug-induced liver injury: Pathogenesis, epidemiology, clinical features, and practical management. Eur. J. Intern. Med. 97, 26-31. https://doi.org/10.1016/j.jim.2021.10.035 (2022).
Clinton, J. W. et al. Drug-Induced liver injury: Highlights and controversies in the recent literature. Drug Saf. 44, 1125-1149. https://doi.org/10.1007/s40264-021-01109-4 (2021).
Jin, C. et al. Treatment responses to Azithromycin and Ciprofloxacin in uncomplicated Salmonella Typhi infection: A comparison of clinical and Microbiological data from a controlled human infection model. PLoS Negl. Trop. Dis. 13, e0007955 (2019).
Bal, A. M. Macrolide antibiotics. (2022).
Zhai, G., Li, M., Wang, Y. & Wu, J. Drug-induced liver disturbance during the treatment of COVID-19. Front. Pharmacol. 12, 719308 (2021).
Firth, A. & Prathapan, P. Azithromycin: The first broad-spectrum therapeutic. Eur. J. Med. Chem. 207, 112739 (2020).
Atli, O., Ilgin, S., Altuntas, H. & Burukoglu, D. Evaluation of Azithromycin induced cardiotoxicity in rats. Int. J. Clin. Exp. Med. 8, 3681 (2015).
Hamza, R. Z. et al. Ascorbic acid ameliorates cardiac and hepatic toxicity induced by azithromycin-etoricoxib drug interaction. Curr. Issues Mol. Biol. 44, 2529-2541(2022).
Li, S. et al. Establishment of a new animal model of azithromycin-induced liver injury and study the molecular pathological change during the process. Hum. Exp. Toxicol. 35, 511-525 (2016).
Ellison, C. A. & Blackwell, S. B. Acute hepatocellular injury associated with Azithromycin. J. Pharm. Pract. 34, 493-496 (2021).
Danica, J. et al. Vanishing bile duct syndrome associated with Azithromycin in a 62-year-old man. Basic. Clin. Pharmacol. Toxicol. 106, 62-65 (2010).
Martinez, M. A. et al. Clinical and histologic features of azithromycin-induced liver injury. Clin. Gastroenterol. Hepatol. 13, 369376 (2015).
Park, H. J., Seo, K. I. & Choi, Y. I. Liver transplantation for azithromycin-induced severe liver injury. Korean J. Transplantation 34, 279-285 (2020).
Choi, Y. I., Moon, H. H., Jo, J. H. & Shin, D. H. Liver transplantation for azithromycin-induced severe liver injury: A case report. Korean J. Transplantation 35, 130 (2021).
Reyes-Gordillo, K., Shah, R. & Muriel, P. Oxidative stress and inflammation in hepatic diseases: Current and future therapy. Oxid. Med. Cell. Longev. 2017 (2017).
Villanueva-Paz, M. et al. Oxidative stress in drug-induced liver injury (DILI): From mechanisms to biomarkers for use in clinical practice. Antioxidants 10, 390 (2021).
Zhang, M. M., Qiao, Y., Ang, E. L. & Zhao, H. Using natural products for drug discovery: The impact of the genomics era. Expert Opin. Drug Discov. 12, 475-487 (2017).
Govea-Salas, M. et al. In the Liver317-322 (Elsevier, 2018).
Aglan, H. A., Ahmed, H. H., El-Toumy, S. A. & Mahmoud, N. S. Gallic acid against hepatocellular carcinoma: An integrated scheme of the potential mechanisms of action from in vivo study. Tumor Biol. 39, 1010428317699127 (2017).
Gao, J., Hu, J., Hu, D. & Yang, X. A role of Gallic acid in oxidative damage diseases: A comprehensive review. Nat. Prod. Commun. 14, 1934578X19874174 (2019).
Jadon, A., Bhadauria, M. & Shukla, S. Protective effect of Terminalia Belerica Roxb. And Gallic acid against carbon tetrachloride induced damage in albino rats. J. Ethnopharmacol. 109, 214-218 (2007).
Latief, U., Husain, H., Mukherjee, D. & Ahmad, R. Hepatoprotective efficacy of Gallic acid during Nitrosodiethylamine-induced liver inflammation in Wistar rats. J. Basic. Appl. Zool. 76, 31-41 (2016).
Reckziegel, P. et al. Antioxidant protection of Gallic acid against toxicity induced by Pb in blood, liver and kidney of rats. Toxicol. Rep. 3, 351-356 (2016).
Feng, R. B., Wang, Y., He, C., Yang, Y. & Wan, J. B. Gallic acid, a natural polyphenol, protects against tert-butyl hydroperoxideinduced hepatotoxicity by activating ERK-Nrf2-Keap1-mediated antioxidative response. Food Chem. Toxicol. 119, 479-488 (2018).
Dutta, M. & Paul, G. Gallic acid protects rat liver mitochondria ex vivo from bisphenol A induced oxidative stress mediated damages. Toxicol. Rep. 6, 578-589 (2019).
Zhou, Y. et al. Gallic acid protects against ethanol-induced hepatocyte necroptosis via an NRF2-dependent mechanism. Toxicol. Vitro. 57, 226-232 (2019).
Hussein, R. M., Anwar, M. M., Farghaly, H. S. & Kandeil, M. A. Gallic acid and ferulic acid protect the liver from thioacetamideinduced fibrosis in rats via differential expression of miR-21, miR-30 and miR-200 and impact on TGF- /Smad3 signaling. Chemico-Biol. Interact. 324, 109098 (2020).
Sundaresan, S. et al. Gallic acid attenuates cadmium mediated cardiac hypertrophic remodelling through upregulation of Nrf2 and PECAM-1signalling in rats. Environ. Toxicol. Pharmacol. 87, 103701 (2021).
Li, X., Sun, R. & Liu, R. Natural products in licorice for the therapy of liver diseases: Progress and future opportunities. Pharmacol. Res. 144, 210-226 (2019).
Shinu, P. et al. Pharmacological features of -glycyrrhetinic acid: A pentacyclic triterpenoid of therapeutic potential. Plants 12, 1086 (2023).
Chen, S., Zou, L., Li, L. & Wu, T. The protective effect of glycyrrhetinic acid on carbon tetrachloride-induced chronic liver fibrosis in mice via upregulation of Nrf2. PLoS One 8, e53662 (2013).
Yang, Y. et al. Protective effects of hepatocyte-specific glycyrrhetic derivatives against carbon tetrachloride-induced liver damage in mice. Bioorg. Chem. 72, 42-50. https://doi.org/10.1016/j.bioorg.2017.03.009 (2017).
Hasan, S. et al. glycyrrhetinic acid alleviates 2 -acetylaminofluorene-induced hepatotoxicity in Wistar rats: Role in hyperproliferation, inflammation and oxidative stress. Hum. Exp. Toxicol. 34, 628-641 (2015).
Mahmoud, A. M. & Al Dera, H. S. 18β-Glycyrrhetinic acid exerts protective effects against cyclophosphamide-induced hepatotoxicity: Potential role of PPAR and Nrf2 upregulation. Genes Nutr. 10, 41. https://doi.org/10.1007/s12263-015-0491-1 (2015).
Mahmoud, A. M., Hussein, O. E., Hozayen, W. G. & Abd El-Twab, S. M. Methotrexate hepatotoxicity is associated with oxidative stress, and down-regulation of PPAR and Nrf2: Protective effect of -Glycyrrhetinic acid. Chem. Biol. Interact. 270, 59-72. https://doi.org/10.1016/j.cbi.2017.04.009 (2017).
Iranshahy, M., Iranshahi, M., Abtahi, S. R. & Karimi, G. The role of nuclear factor erythroid 2-related factor 2 in hepatoprotective activity of natural products: A review. Food Chem. Toxicol. 120, 261-276. https://doi.org/10.1016/j.fct.2018.07.024 (2018).
Paulose, P. et al. Ameliorative effect of vitamin E on Azithromycin induced biochemical and histological changes in liver tissue of rats. Adv. Anim. Vet. Sci. 4, 85-91 (2016).
Ojeaburu, S. & Oriakhi, K. Hepatoprotective, antioxidant and, anti-inflammatory potentials of Gallic acid in carbon tetrachlorideinduced hepatic damage in Wistar rats. Toxicol. Rep. 8, 177-185 (2021).
Bancroft, J. D. & Gamble, M. Theory and Practice of Histological Techniques (Elsevier health sciences, 2008).
Ibrahim Fouad, G. & Ahmed, K. A. Curcumin ameliorates doxorubicin-induced cardiotoxicity and hepatotoxicity via suppressing oxidative stress and modulating iNOS, NF- , and TNF- in rats. Cardiovasc. Toxicol. 22, 152-166 (2022).
Buege, J. A. & Aust, S. D. In Methods in Enzymology 52302-310 (Elsevier, 1978).
Nandi, A. & Chatterjee, I. Assay of superoxide dismutase activity in animal tissues. J. Biosci. 13, 305-315 (1988).
Towbin, H., Staehelin, T. & Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. 76, 4350-4354 (1979).
Usadadia, S., Patel, J., Vihol, P. & Urkude, A. Protective effect of Quercetin on Azithromycin induced hepatotoxicity and nephrotoxicityin Wistar rats. Int. J. Curr. Microbiol. Appl. Sci. 9, 2926-2934 (2020).
Plaa, G. & Zimmerson, H. Evaluation of hepatotoxicity: physiological and biochemical measures of hepatic function. Compr. Toxicol. 9, 97-109 (1997).
Wang, H. et al. Glycyrrhizin and glycyrrhetinic acid inhibits alpha-naphthyl isothiocyanate-induced liver injury and bile acid cycle disruption. Toxicology 386, 133-142 (2017).
Liguori, I. et al. Oxidative stress, aging, and diseases. Clin. Interv Aging, 757-772 (2018).
Jomova, K. et al. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: Chronic diseases and aging. Arch. Toxicol. 97, 2499-2574 (2023).
Singh, H., Prakash, A., Kalia, A. & Majeed, A. B. A. Synergistic hepatoprotective potential of ethanolic extract of Solanum xanthocarpum and Juniperus communis against Paracetamol and Azithromycin induced liver injury in rats. J. Traditional Complement. Med. 6, 370-376 (2016).
Jin, J. et al. Role of Nrf2 activation and NF- Inhibition in valproic acid induced hepatotoxicity and in diammonium glycyrrhizinate induced protection in mice. Food Chem. Toxicol. 73, 95-104 (2014).
Liu, T., Zhang, L., Joo, D. & Sun, S. C. NF-кB signaling in inflammation. Signal. Transduct. Target. Therapy. 2, 1-9 (2017).
Mohamed, E. K. & Hafez, D. M. Gallic acid and Metformin co-administration reduce oxidative stress, apoptosis and inflammation via Fas/caspase-3 and NF- signaling pathways in thioacetamide-induced acute hepatic encephalopathy in rats. BMC Complement. Med. Ther. 23, 265 (2023).
Ahmed, S. M. U., Luo, L., Namani, A., Wang, X. J. & Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Et Biophys. Acta (BBA)-Molecular Basis Disease 1863, 585-597 (2017).
Qi, W. et al. Cardamonin protects against doxorubicin-induced cardiotoxicity in mice by restraining oxidative stress and inflammation associated with Nrf2 signaling. Biomed. Pharmacother 122, 109547 (2020).
Younis, N. S., Elsewedy, H. S., Shehata, T. M. & Mohamed, M. E. Geraniol averts methotrexate-induced acute kidney injury via keap1/Nrf2/HO-1 and MAPK/NF- B pathways. Curr. Issues Mol. Biol. 43, 1741-1755 (2021).
Author contributions
Study design was done by G.H., and A.H. The experiments and statistical analysis were carried out by M.A. A.I., and G.H. Resources and methodological assistance were provided by G.H., A.H., M.A. A.I., and A.A The original draft of the manuscript was written by A.H., G.H., M.A. A.I., and A.A All authors then edited and accepted the updated version of the article. Each author has reviewed the published version of the manuscript and given their approval.
Funding
AA was supported by the University of Sharjah’s Seed Grant No. (24010901156).
Declarations
Competing interests
The authors declare no competing interests.
Additional information
Supplementary Information The online version contains supplementary material available at https://doi.org/1 0.1038/s41598-025-93120-3.
Correspondence and requests for materials should be addressed to A.A.H. or A.A.
Reprints and permissions information is available at www.nature.com/reprints.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abu Qurqas Health Administration, Minia Directorate of Health, Ministry of Health, El-Minia 61611, Egypt. Department of Pharmacology and Toxicology, Faculty of Pharmacy, Minia University, El-Minia, Egypt. Biology Department, National Organization for Drug Control and Research, Giza 12611, Egypt. College of Medicine, University of Sharjah, Sharjah 27272, UAE. email: alaa17mm@gmail.com; a.amin@sharjah.ac.ae