الحديد يحفز أكسدة الدهون في الأغشية البيولوجية ويعزز شكلًا من أشكال موت الخلايا يسمى الفيروبتوز.تحديد مكان حدوث هذه الكيمياء في الخلية يمكن أن يُفيد في تصميم أدوية قادرة على تحفيز أو تثبيط الفيروبتوز في سياقات مرضية مختلفة. وقد كشفت الأساليب الجينية عن مثبطات الفيروبتوز.؛ بالمقابل، يمكن للجزيئات الصغيرة أن توفر تحكمًا مكانيًا وزمنيًا في الكيمياء التي تعمل.. هنا نوضح أن مثبط الفيروبتوزيس ليبروكستاتين-1 يمارس تأثيرات واقية للخلايا من خلال تعطيل الحديد في الليزوزومات. كما نوضح أن المحفز للفيروبتوزيس RSL3 يبدأ أكسدة الدهون الغشائية في الليزوزومات. قمنا بتصميم منشط جزيئي صغير للحديد الليزوزومي – فنتوميسين-1 – لتحفيز التحلل الأكسيدي للفوسفوليبيدات وفي النهاية الفيروبتوزيس. فنتوميسين-1 قادر على قتل CD44 الغني بالحديد.خلايا الساركوما الأولية والأدينوكارسينوما القناة البنكرياسية، التي يمكن أن تعزز الانتقال وتغذي تحمل الأدوية. في مثل هذه الخلايا، ينظم الحديد تكيف الخلايا.بينما يمنح القابلية للفيروبتوسيستكتسب خلايا الساركوما المعرضة لجرعات غير مميتة من الفينتوميسين-1 حالة مقاومة للفيروبتوز تتميز بتقليل علامات الميزانشيم وتنشيط استجابة تلف الغشاء. يمكن لهذا المحلل للفوسفوليبيد القضاء على خلايا السرطان المقاومة للأدوية في المختبر ويقلل من نمو الأورام داخل العقد اللمفاوية في نموذج فأر لانتشار سرطان الثدي. معًا، تُظهر هذه النتائج أن التحكم في تفاعل الحديد يوفر فوائد علاجية، وتؤسس الحديد الليزوزومي كهدف قابل للعلاج وتبرز قيمة استهداف حالات الخلايا..
يتفاعل الحديد مع البيروكسيدات لإنتاج جذر حُر مركزي بالأكسجين. في الخلية، يمكن أن يبدأ مثل هذا العملية وينشر سلسلة من التفاعلات الجذرية التي تؤدي إلى منتجات عضوية مؤكسدة، وهي كيمياء تُعرف بشكل عام بتفاعل فنتون.الدهون الكيميائية التفاعلية في الأغشية البيولوجية هي ركائز مثالية لمثل هذه التفاعلات وقد تم الإشارة إليها في عملية الفيروبتوسيس.يمكن أن يؤدي تراكم الفوسفوليبيدات التالفة في النهاية إلى فقدان سلامة الغشاء مع تغير وظائف العضيات وإطلاق أنواع مختلفة من الكيانات الكيميائية في الخلية، والتي يمكن أن تسبب مزيدًا من العيوب الخلوية وموت الخلايا. تتضمن الفيروبتوزيس عضيات مميزة.بما في ذلك البيروكسيوماتالميتوكوندرياالشبكة الإندوبلازمية (ER)والليزوزومات. ومع ذلك، فإنه من غير الواضح حاليًا ما إذا كانت العضيات الفردية تساهم في الفيروبتوز من خلال إعاقة إشارات الخلايا، أو الأيض، أو تخليق جزيئات حيوية معينة، أو ما إذا كانت الدهون الغشائية
تعتبر هذه الحُجرات أيضًا ركائز مباشرة للأكسدة المعتمدة على الحديد التي تؤدي إلى موت الخلايا. وقد أظهرت الدراسات أن خلايا السرطان تحتاج إلى ليسوزومات وظيفية لتخضع لعملية الفيروبتوز.. تسلط هذه النتيجة الضوء على أهمية استيراد الحديد الخلوي وتدعم الفكرة القائلة بأن الحديد له دور في تعزيز موت الخلايا. إن تثبيط الحركة الدوائية للحديد من الجسيمات الحالة يعزز أكسدة الدهون في أغشية الجسيمات الحالة.، مما يشير إلى أن مجموعة الحديد النشطة في هذا العضية يمكن أن تحفز الفيروبتوز. يمكن أن تحفز المركبات الاصطناعية من الإندوبيروكسيدات، وهي بدائل للدهون المؤكسدة التي تميل إلى التراكم في الشبكة الإندوبلازمية، الفيروبتوز، مما يقترح أن تأكسد الدهون يمكن أن ينتشر من خلال هذه العضية.ومع ذلك، فإنه غير واضح حاليًا أين يتم بدء أكسدة الدهون الغشائية بواسطة الحديد في الخلية قبل أن تنتشر عبر الدهون الغشائية القريبة الأخرى.
الحديد الليزوزومي يبدأ عملية الفيروبتوز
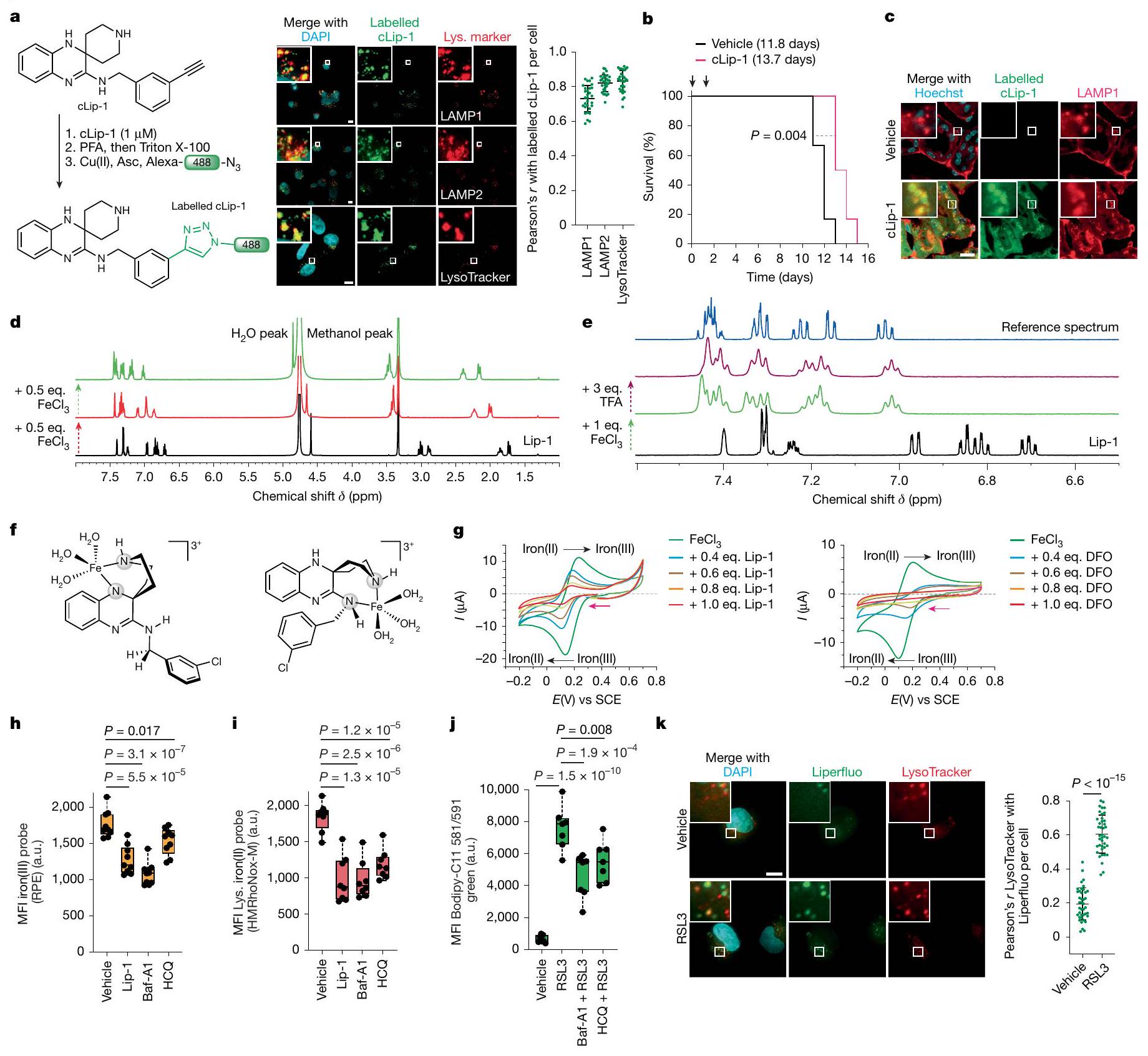
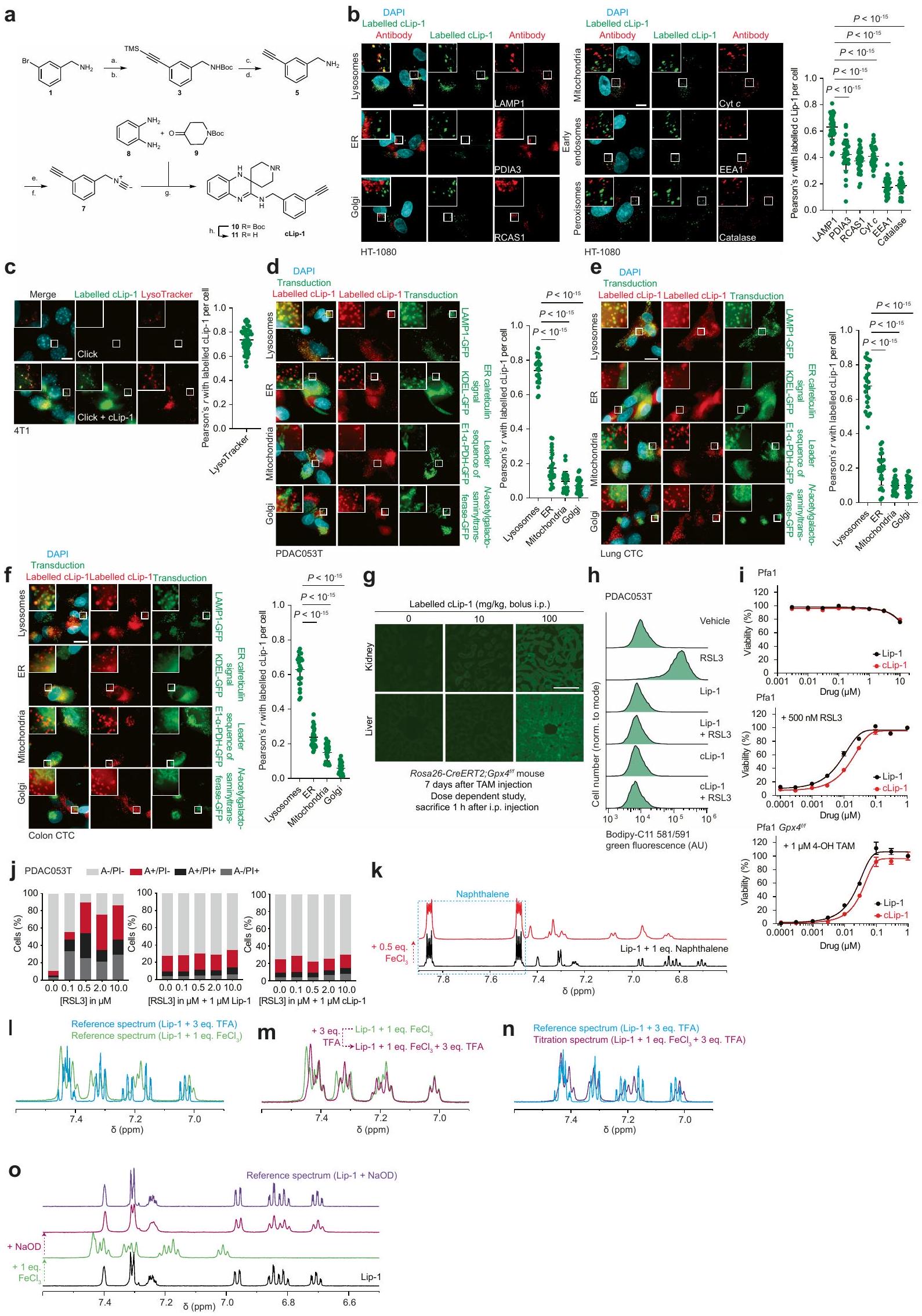
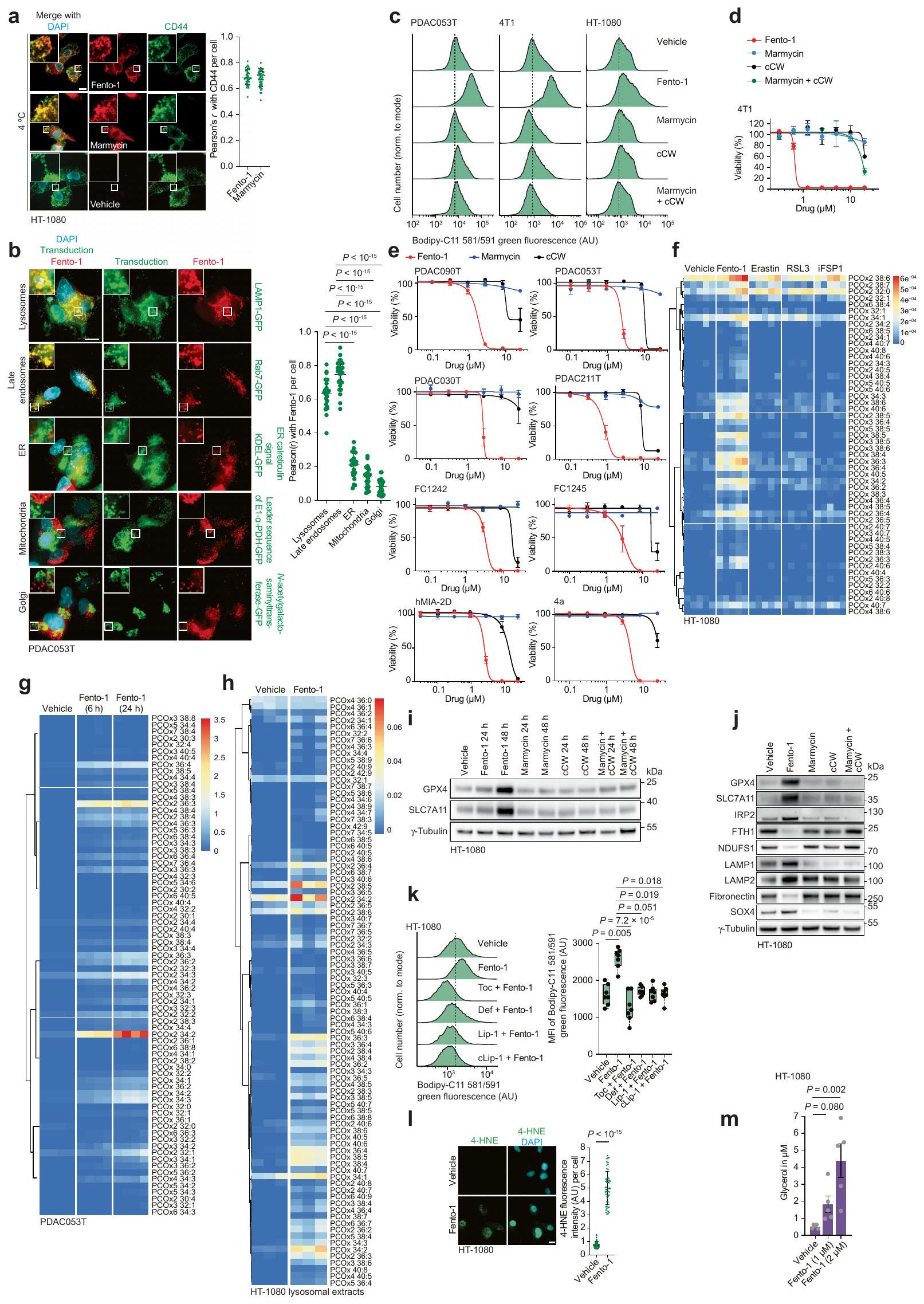
قمنا بتحديد المواقع تحت الخلوية لعمل ليبروكستاتين-1 (Lip-1)، وهو جزيء صغير يحمي الخلايا من الموت الناتج عن تعطيل مثبطات الفيروبتوز.قمنا بتسمية نظير يحتوي على الألكاين قابل للنقر من Lip-1 (المسمى cLip-1) باستخدام كيمياء النقر داخل الخلايا وأجرينا مجهرية الفلورسنت.في خطوط الخلايا المختلفة. كشفت النتائج عن تراكم الليزوزومات في خلايا الساركوما الليفية HT-1080، وفي خلايا سرطان القناة البنكرياسية البشرية الأولية (PDAC)، وفي خلايا الأورام الدائرة في الرئة والقولون، وفي خلايا سرطان الثدي في الفئران 4T1 (الشكل 1 أ والشكل الممتد 1 أ-و). من الجدير بالذكر أن cLip-1 المعلم تم اكتشافه فقط في الحجرة الإندوليزوزومية، حتى عند تركيز أعلى من الجرعة الفعالة. تدعم هذه النتيجة الفكرة القائلة بأن Lip-1 يمارس نشاطه المضاد للفيروبوتوز في هذه العضية. في الجسم الحي، زاد cLip-1 من بقاء الفئران التي تم تحفيز الفشل الكلوي الحاد فيها من خلال الحذف الجيني لـالذي يشفر مثبط الفيروبتوزيس بيروكسيداز الجلوتاثيون (الشكل 1ب). أظهر وسم cLip-1 في أنسجة الفئران تراكمه في الكلى، حيث تزامن مع علامة ليفوسوم في الأنابيب القريبة من الكلى (الشكل 1ج والشكل التمديدي 1ز). في المختبر، منع cLip-1 أكسدة الدهون الغشائية وحمى الخلايا من آثار النقص الجيني في Gpx4 أو التثبيط الدوائي لـ GPX4 باستخدام RSL3، مما يعيد تجسيد النشاط البيولوجي لـ Lip-1 (الشكل التمديدي 1ح-ي). هذه البيانات تؤكد أن cLip-1 هو بديل مناسب لـ Lip-1 للتحقيق في الفيروبتوز.
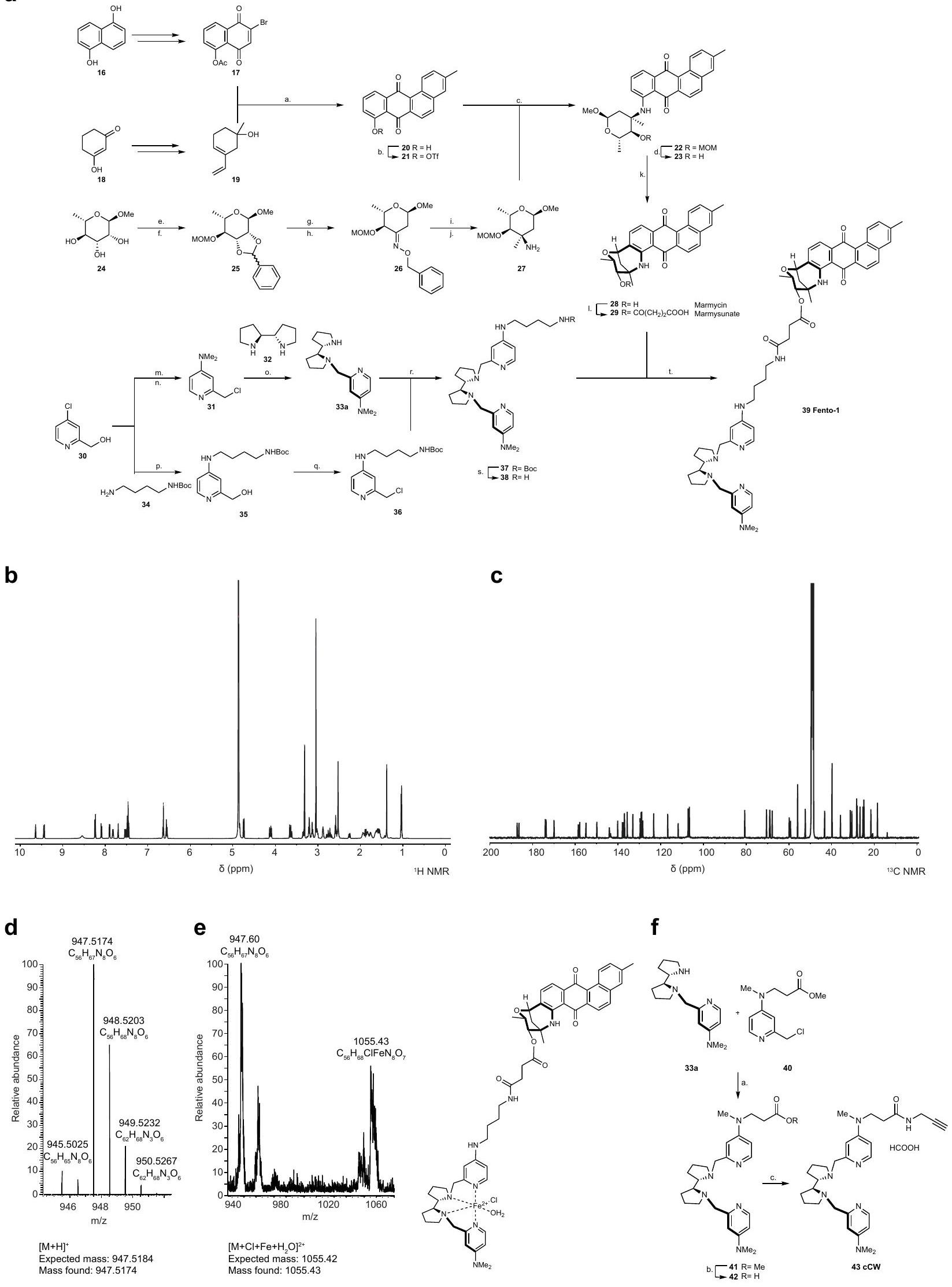
الكلاتير الحديدي الضيق ديفيروكسامين (DFO) يحمي الخلايا من الموت الخلوي الناتج عن الإراستين.بالمقابل، يظهر Lip-1 خصائص مضادة للأكسدة قادرة على التقاط الجذور الحرة (RTA) في المختبر.. ومع ذلك، على عكس معظم RTAs التي تعيق الفيروبتوز، مثللا يتبع ليب-1 الاتجاه العام للعديد من مثبطات الفيروبتوز، حيث أن نشاط RTA مرتبط ارتباطًا وثيقًا بتثبيط الفيروبتوز.. تشير هذه النتيجة إلى أن خصائص أخرى لـ Lip-1 تساهم في تأثيراته المضادة للفيروبتوز. في خلايا السرطان، يمكن أن يتم امتصاص الحديد عن طريق الإندوسيتوز.وينظم انتقالات حالة الخلية وتكاثر الخلايا. لأن cLip-1 يتراكم في الليزوزومات ويحتوي على ذرات نيتروجين قابلة للتفاعل مع الحديد، قمنا بالتحقيق في إمكانية أن خصائص تثبيط الفيروبتوز قد تنتج عن تعطيل الحديد في هذه العضية. أظهرت مطيافية الرنين المغناطيسي النووي (NMR) أن Lip-1 يتفاعل مع الحديد(III) بنسب 1:1 و 2:1 (الشكل 1d). أدى تيترا Lip-1 مع كلوريد الحديد(III) إلى توسيع الإشارة مع تحول في الأسفل لإشارات البروتونات المجاورة لذرات النيتروجين في Lip-1، مما يدل على حدوث تنسيق مع أزواج النيتروجين الوحيدة مع الحمض لويس. بالمقابل، ظلت إشارات البروتونات للنفتالين، الذي تم استخدامه كتحكم داخلي لعدم احتوائه على ذرات غير متجانسة، غير متأثرة (الشكل 1k من البيانات الموسعة). علاوة على ذلك، أدى إضافة حمض برونستيد إلى Lip-1 إلى تحولات مميزة في الأسفل لإشارات البروتونات، مما يدل على بروتنة ذرات النيتروجين (الشكل 1e). من الجدير بالذكر أن توسيع خط إشارات البروتونات لوحظ فقط في وجود المعدن البارامغناطيسي (الشكل 11-n من البيانات الموسعة). علاوة على ذلك، لم تؤدي إضافة حمض ثلاثي فلورو الأسيتيك (TFA) إلى محلول من الحديد(III) و Lip-1 إلى إزاحة الحديد بالكامل من Lip-1، كما هو موضح بـ تحولات كيميائية أعادت تلخيص تأثيرات الأحماض لويس وبرونستيد. تشير هذه البيانات إلى أن ارتباط Lip-1 بالحديد يمكن أن يحدث في ظروف حمضية، مثل تلك الموجودة في الليزوزومات. أدت إضافة هيدروكسيد الصوديوم إلى استعادة التحولات الكيميائية لـ Lip-1 غير المرتبط، وهو نتيجة تُظهر الطبيعة القابلة للعكس لتفاعلات Lip-1 مع الحديد (الشكل التمديدي 1o). أنتجت نمذجة جزيئية باستخدام حساب تحويل فورييه المنفصل عدة أوضاع ارتباط محتملة بين الحديد وLip-1، بما في ذلك معقدات الحديد الرباعية الجزيئية الثنائية التي تشمل وظائف البيبيريدين، والأميدين، والأنيلين لـ Lip-1 (الشكل 1f والمعلومات التكميلية).
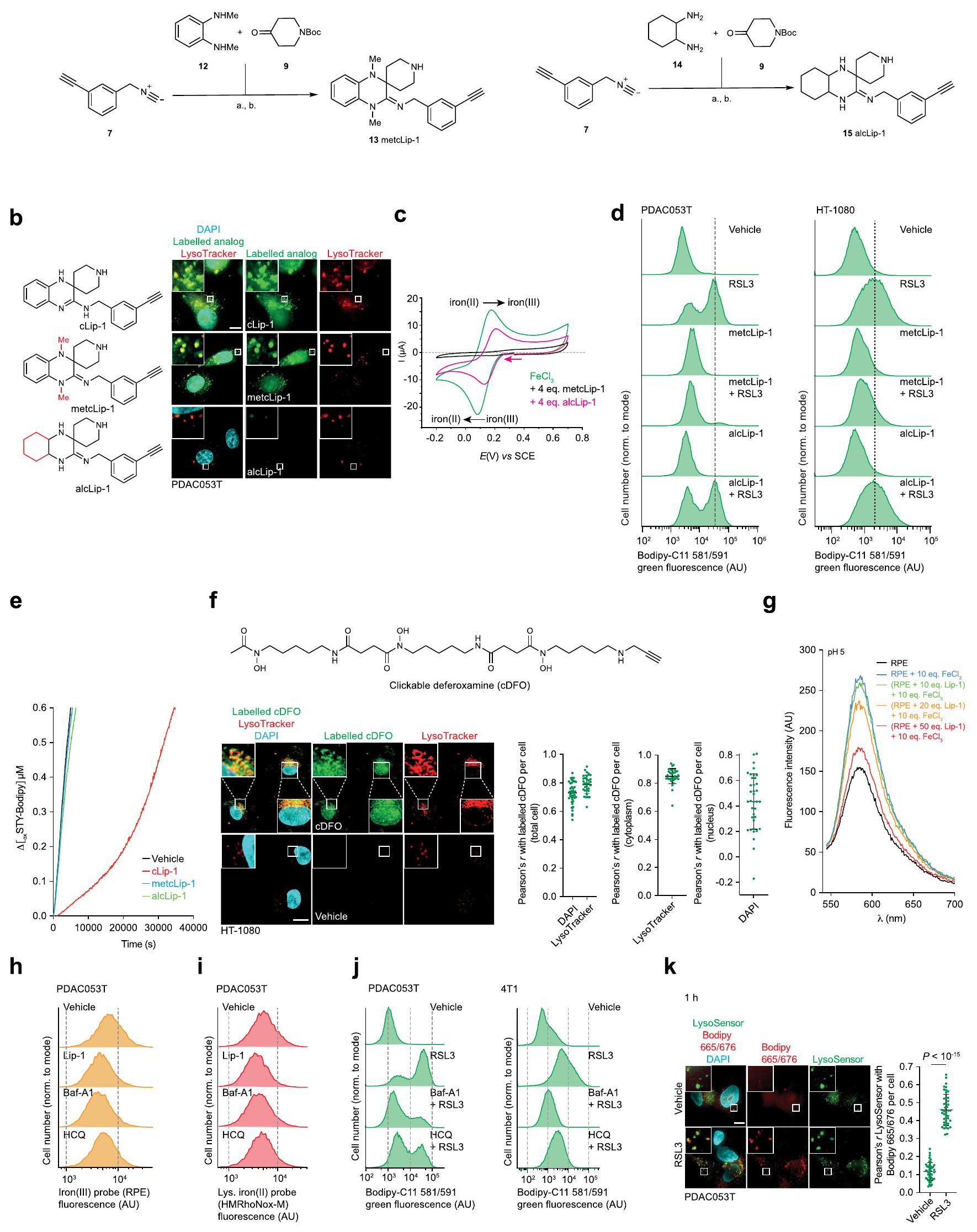
الجهد الكهربائي الدوري، الذي يُستخدم لتحديد الجهد الاختزالي للمعادن في المحلول“، أشار إلى أن Lip-1 و DFO يغيران خصائص الأكسدة والاختزال للحديد (الشكل 1g). كضوابط، قمنا بدراسة اثنين من النظائر الاصطناعية الأخرى لـ Lip-1: metcLip-1 و alcLip-1. في metcLip-1، تم ميثلة الأمينات العطرية، مما أثر سلبًا على نشاط RTA. بالمقابل، alcLip-1 هو نظير أليفاتي يحتوي على أمينات لها ثابت تفكك حمضي سالب لوغاريتمي أعلى. ) القيمة مقارنة بنظيراتها العطرية. كان من المتوقع أن يكون هذا النظير مؤينًا بالكامل عند درجة الحموضة المحايدة وبالتالي يظهر ميلًا أقل لدخول الخلايا والتراكم في الليزوزومات (الشكل 2a من البيانات الموسعة). أظهر وسم metcLip-1 في الخلايا نمط تلوين مشابه لذلك الخاص بـ cLip-1 المسمى، مما يدل على التوضع المشترك مع علامة ليزوزومية. بالمقابل، أظهر alcLip-1 المسمى فلوريسنس ضعيف (الشكل 2b من البيانات الموسعة). أشارت نتائج الفولتمترية الدورية أيضًا إلى أن metcLip-1 غيرت الجهد الكهروكيميائي للحديد بشكل أكبر من alcLip-1 (الشكل 2c من البيانات الموسعة). وبشكل متسق، حمى metcLip-1 الخلايا من أكسدة الدهون الغشائية الناتجة عن RSL3 بشكل أقوى من alcLip-1 (الشكل 2d من البيانات الموسعة)، ولم يظهر أي من metcLip-1 أو alcLip-1 خصائص RTA في المختبر (الشكل 2e من البيانات الموسعة). تم وسم مشتق يحتوي على الألكاين من DFO (cDFO) في الخلاياكما كشفت عن إشارات ليفوسومية، مما قدم دعماً للنشاط المؤيد للفيروبتوز في هذه العضية (الشكل 2f من البيانات الموسعة). كما وُجدت إشارات cDFO في نواة الخلية، مما يفسر جزئياً سمية هذا الخالب للحديد ونمطه الظاهري المميز مقارنةً بجزيئات صغيرة أخرى تتفاعل مع الحديد، بما في ذلك Lip-1.
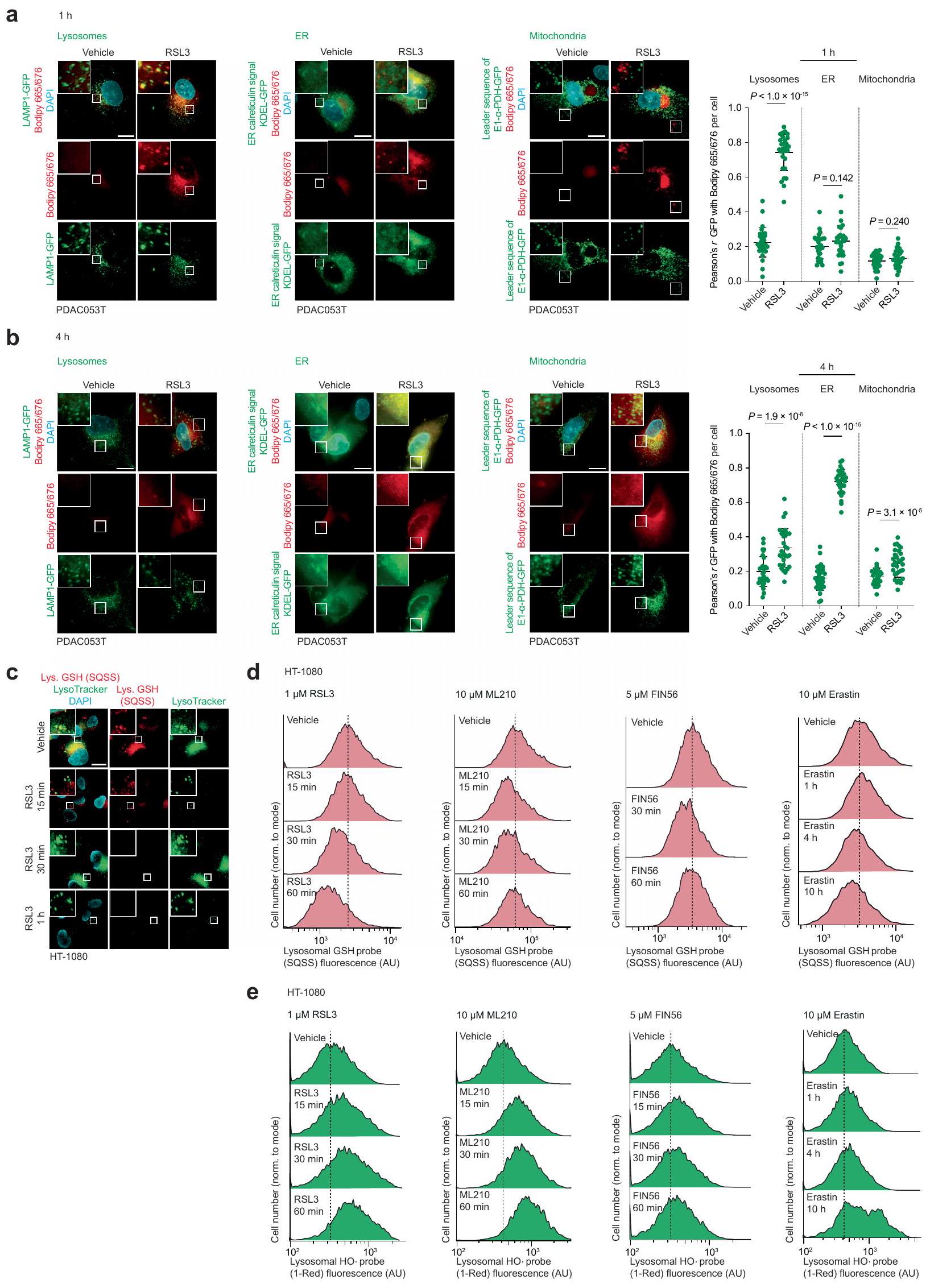
في نظام خالٍ من الخلايا، قلل Lip-1 من الفلورية لمسبار فلوري محدد للحديد (III) في وجود الحديد (III). (الشكل 2g من البيانات الموسعة). أدى علاج الخلايا بالهيدروكسي كلوروكين أو بافيلوميسين-A1، اللذان يزيدان من درجة حموضة الليسوزوم ويمنعان تفريغ الحديد (III) من حامليه الداخليين، إلى تقليل مخزونات الحديد (III) الخلوي القابل للتخلب والحديد (II) الليسوزومي التفاعلي، كما تم قياسه باستخدام مجسات فلورية محددة.. هذه العلاجات أيضًا حمت الخلايا من أكسدة الدهون الغشائية الناتجة عن RSL3 في خلايا السرطان (الشكل 1h-j والبيانات الموسعة الشكل.تم الحصول على نتائج مماثلة مع Lip-1، والتي كانت متسقة مع التفاعلات المباشرة لهذه المركب مع الحديد الليزوزومي (الشكل 1h-j وبيانات ممتدة الشكل 2h-j). بعد العلاج بـ RSL3 لمدة ساعة واحدة، تم الكشف عن أكسدة الدهون الغشائية بشكل رئيسي في الليزوزومات، كما تم تقييمه من خلال مستويات فلورية من Liperfluo وBodipy 665/676 (الشكل 1k وبيانات ممتدة الأشكال 2k و3a). بالمقابل، أدى العلاج بـ RSL3 لمدة 4 ساعات إلى نمط تلوين للدهون الغشائية المؤكسدة التي تزامنت بشكل رئيسي مع علامة بيولوجية ملونة فلوريسنتياً للشبكة الإندوبلازمية (بيانات ممتدة الشكل 3b). تشير بياناتنا إلى أن بدء تفاعل السلسلة الجذرية يحدث مبكراً في الحجرة الإندوليزوزومية، حيث يمكن العثور على الحديد النشط في حالة الأكسدة.يمكن أن تنتشر هذه التفاعل السلسلي الجذري بعد ذلك إلى الدهون الغشائية للعضيات الأخرى في الجوار، بما في ذلك الشبكة الإندوبلازمية. دعمًا لهذه الفرضية، أدى علاج الخلايا بمحفزات الفيروبتوسيس المعروفة إلى استنفاد الجلوتاثيون وزيادة مستويات الجذور الحرة المرتكزة على الأكسجين في الليزوزومات (الشكل 3c-e من البيانات الموسعة). معًا، تشير هذه البيانات إلى أن الحديد الليزوزومي له دور مركزي في تحفيز الفيروبتوسيس.
تنشيط الحديد يعزز الفيروبتوز
خلايا السرطان التي اكتسبت حالة خلوية مقاومة للأدوية (DTP) تكون عرضة للفيروبتوز.تتميز خلايا سرطان DTP
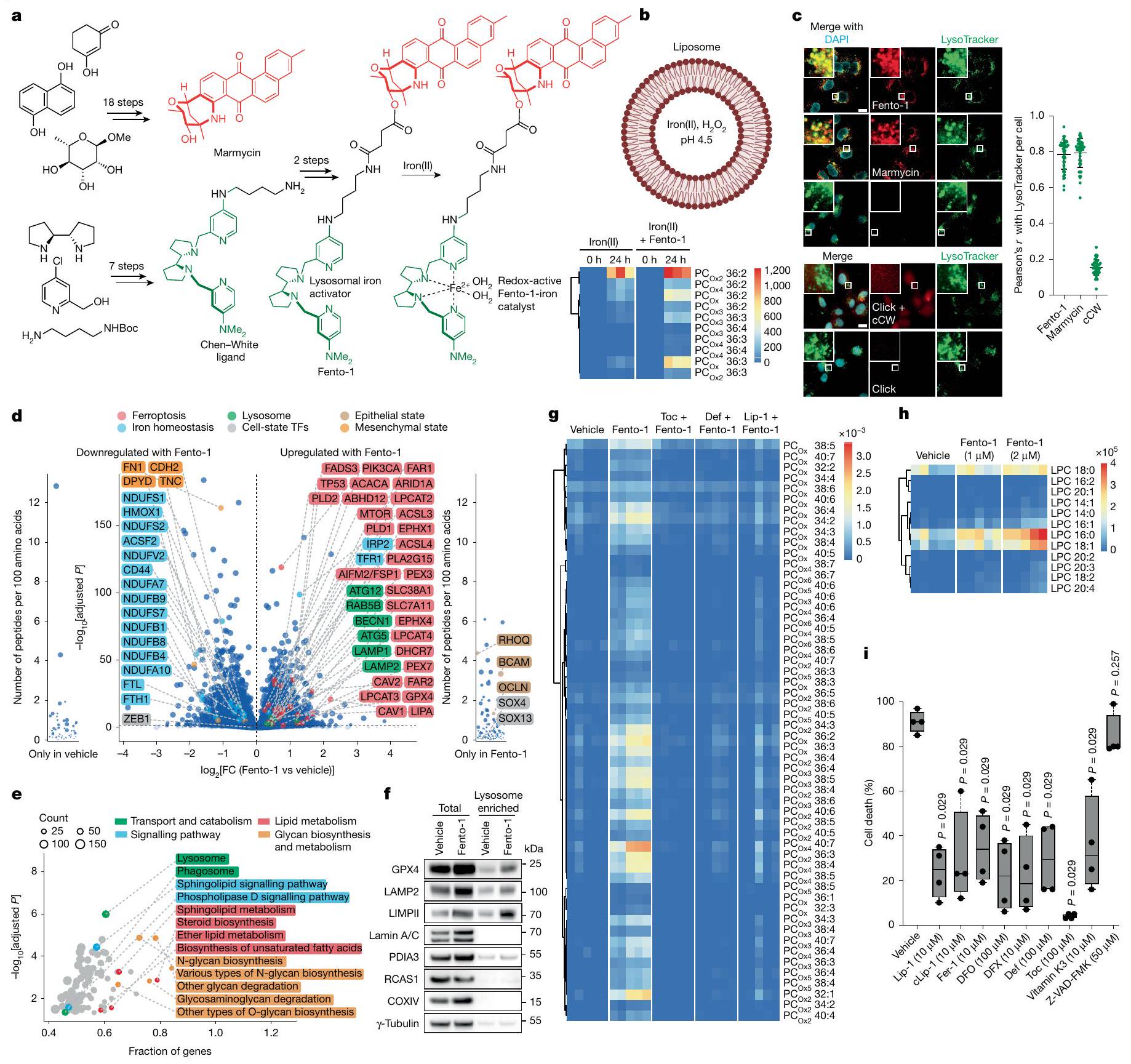
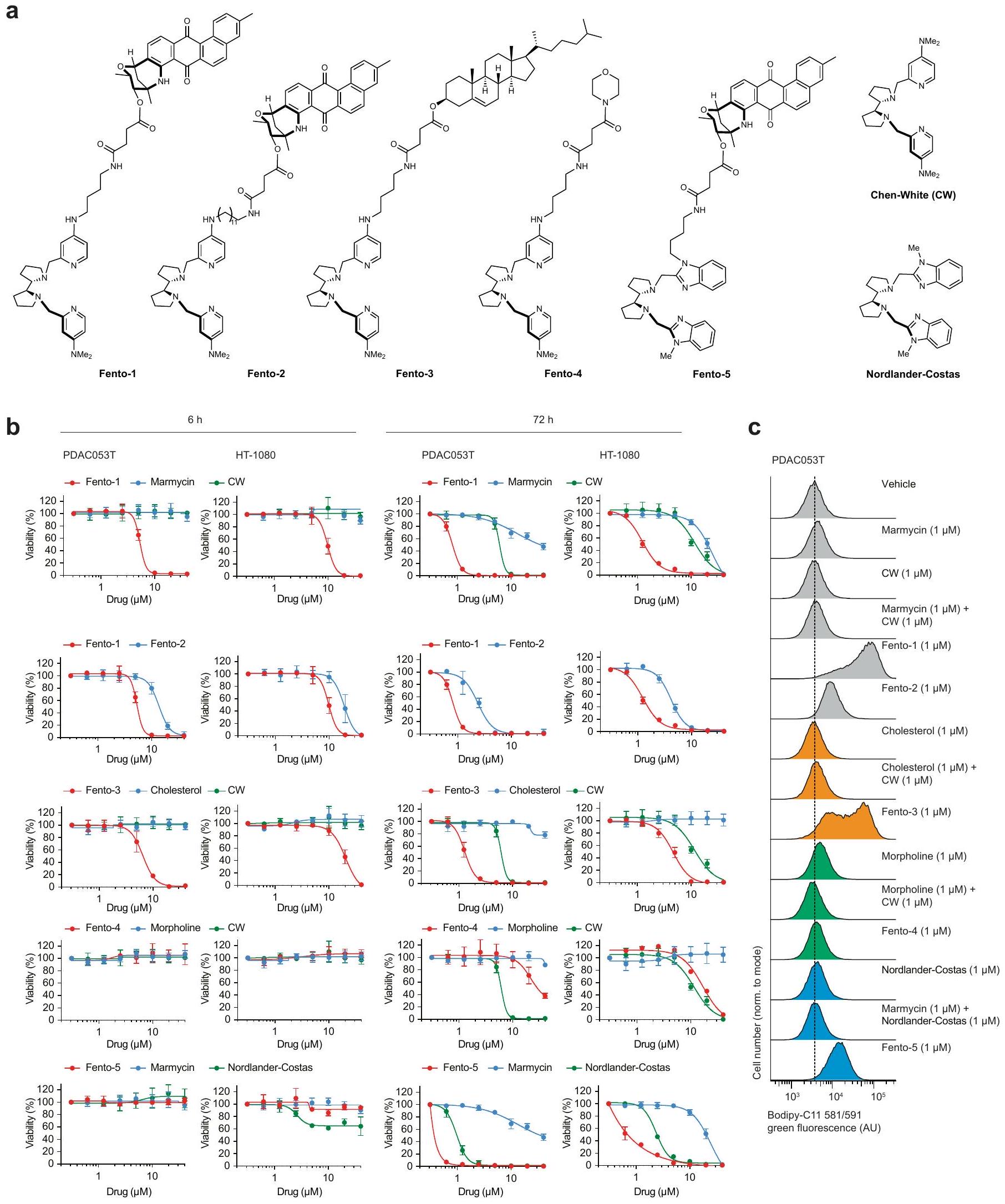
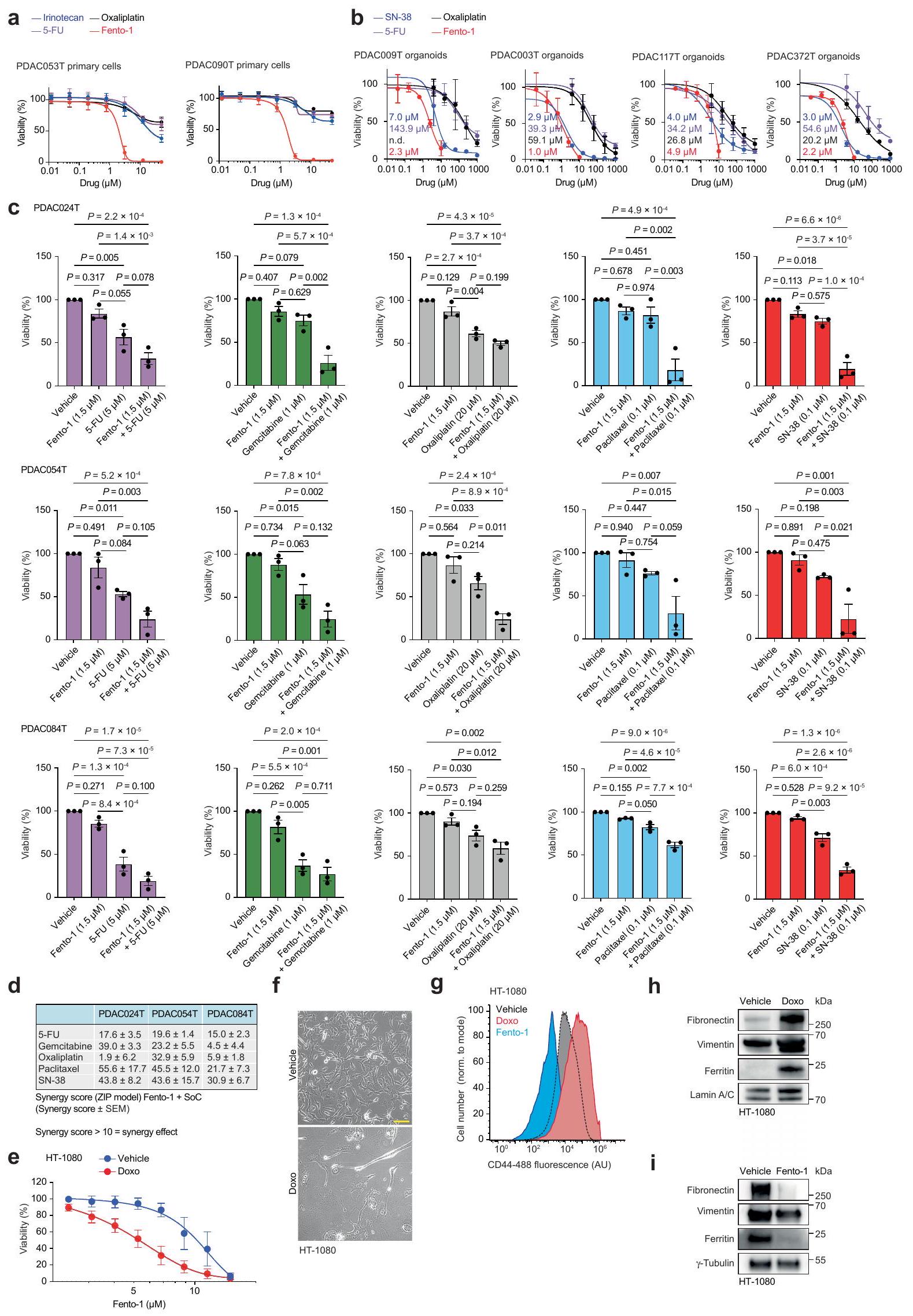
الشكل 1 | الحديد الليزوزومي يحفز أكسدة الدهون الغشائية. أ، مخطط (يسار)، صور (وسط) وقياس (يمين) لتوسيم cLip-1في الخلايا.تجارب مستقلة في خلايا HT-1080. البيانات هي المتوسط ± الانحراف المعياري. Asc، أسكوربات؛ Lys.، ليسوزوم؛ PFA، بارافورمالدهيد. ب، منحنيات بقاء كابلان-ماير لـ Rosa26-CreERT2؛ Gpx4الفئران المعالجة بـ cLip-1 لكل كيلوجرام في اليوم عن طريق الحقن داخل البطن؛ الفئران لكل مجموعة). اختبار مانتل-كوكس لوج-رانك. الأسهم تشير إلى توقيت حقنتي التاموكسيفين. متوسط أيام البقاء على قيد الحياة موضح. ج، صور فلورية لـ cLip-1 المعلم في الأنابيب القريبة الكلوية لـ Rosa26-CreERT2;Gpx4فأر بعد 7 أيام من علاج التاموكسيفين. الصور تمثلفئران. د،طيف الرنين المغناطيسي النووي لـ Lip-1 المعياري مع (أحمر، 2:1 ليب-1 إلى الحديد؛ أخضر، 1:1 ليب-1 إلى الحديد). هـ، طيف الرنين المغناطيسي النووي لـ Lip-1 المضاف إليهثم TFA. الطيف المرجعي الأزرق هو Lip-1 + 3 معادلة TFA.f، أوضاع الربط المحتملة لـ Lip-1 والحديد المستمدة من النمذجة الجزيئية.g، الفولتمترية الدورية لـحل مع Lip-1 (يسار) أو DFO (يمين) (إمكانات الاختزال موضحة بالأسهم الوردية). SCE، إلكترود الكالوميل المشبع. ح-ك، البيانات خاصة بخلايا HT-1080. ح، تحليل تدفق الخلايا. تمت معالجته بالمركبات المحددة لمدة 15 دقيقة ثم بمسبار الحديد (III) RPE لـتجارب مستقلة. وحدة عشوائية؛ MFI، متوسط شدة الفلورة. i، قياس التدفق الخلوي للخلايا المعالجة بالمركبات المحددة لمدة 15 دقيقة ثم مع مجس الحديد (II) HMRhoNox-M لمدة 15 دقيقة.تجارب مستقلة. قياس تدفق الخلايا المعالجة بـ RSL3 (1 ساعة) ومع بافيلوميسين-A1 (Baf-A1) أو هيدروكسي كلوروكوين (HCQ) ساعتين قبل.تجارب مستقلة. تحليل التباين أحادي الاتجاه (ANOVA). ك، تصوير الفلورسنت لـ Liperfluo في خلايا تم علاجها بـ RSL3. تجارب مستقلة. غير متطابقة ذات جانبين-اختبار. البيانات هي المتوسط ± الانحراف المعياري، طيف NMR المسجل عند 310 كلفن في الميثانول-. ل و ، القيم من نوع غير متزاوج ذو جانبين-اختبار مقارنة مع المركبة. تم استخدام التركيزات التالية: h، i، Lip-1باف-أ1تظهر مخططات الصندوق نطاق الربع، مع خطوط مركزية تشير إلى الوسيطات وشعيرات تشير إلى القيم الدنيا والقصوى. شريط القياس،. عن طريق التعبير العالي لبروتين امتصاص الحديد CD44، وهو بروتين سكري موجود في غشاء الخلية يتم إدخاله عن طريق البلعمة ويستخدم كعلامة على خصائص الخلايا السرطانية والانتقال.. وبالتالي، تظهر هذه الخلايا زيادة امتصاص الحديد، مما يؤدي إلى زيادة مستويات الحديد الخلوية التي بدورها تعزز نشاط الديميثيلاز المعتمد على الحديد. هذه العملية تنشط برامج محددة على مستوى الإبيجينيتك والترجمة الجينية التي تكمن وراء انتقالات حالة الخلية واكتساب نمط ظاهرة DTPوبالتالي، ظهر الحديد الليزوزومي كهدف قابل للعلاج لتعزيز الفيروبتوزيس بطريقة تعتمد على حالة الخلية.مع وضع ذلك في الاعتبار ومع العلم أن الحديد الليزوزومي يمكن أن يحفز أكسدة الدهون الغشائيةقمنا بتصميم جزيء صغير لاستهداف الدهون في الغشاء البلازمي وتنشيط الحديد (II) في الليزوزومات عند الابتلاع الخلوي. مثل هذا المركب سيستغل الدهون التفاعلية القريبة في الغشاء كركائز للأكسدة. لهذا الغرض، قمنا بتخليق مركب ثنائي الوظيفة يتكون من المنتج الطبيعي الفلوري الليبوفيلك مارمايسين والليغاند الاصطناعي تشين-وايت، الذي أطلقنا عليه اسم فنتومايسين-1 (Fento-1) (الشكل 2أ والشكل التمديدي 4أ-هـ). يتجمع مارمايسين عند الغشاء البلازمي ويتم امتصاصه عن طريق الابتلاع الخلوي.بينما يُستخدم ligand Chen-White عادةً في التخليق الكيميائي لأكسدة الركائز العضوية عن طريق تنشيط الحديد (II)في وجود بيروكسيد الهيدروجين وتحت ظروف مائية حمضية خفيفة، مثل تلك الموجودة في الليزوزومات، يُعتقد أن محفز الحديد تشين-وايت يشكل نوعًا تفاعليًا من الحديد-أوكسي.. مثل الجذور الحرة الهيدروكسيلية والهيدروبيروكسيلية، يمكن لهذا الوسط أن يستخلص ذرة هيدروجين من الركائز العضوية، بما في ذلك المشبعة، لإنتاج جذور كربونية تفاعلية ومنتجات أكسدة لاحقة. لذلك، فإن فنتو-1 يشبه جزيئًا صغيرًا ثنائي الوظيفة قادرًا على تحفيز القرب من جديد بين محفز والأحماض النووية أو البروتينات لتعزيز التحلل.اقترحنا أنه من خلال استغلال الكمية الوفيرة من الحديد (II) التفاعلي في خلايا سرطان DTPستشكل جزيء ثنائي الوظيفة هذا محفزًا نشطًا في الخلية لتعزيز التحلل التأكسدي للدهون الغشائية الليزوزومية وفي النهاية تحفيز الفيروبتوز.
في نظام خالٍ من الخلايا، عزز Fento-1 الأكسدة الناتجة عن الحديد للفوسفوليبيد غير المشبع الذي يشكل الحويصلات تحت ظروف تجريبية مشابهة لتلك الموجودة في الليزوزومات. أي، درجة حموضة حمضية، وجود بيروكسيد الهيدروجين وملح الحديد (II) القابل للذوبان في الماء (الشكل 2ب والجدول التكميلي 1). من الجدير بالذكر أن الحديد عزز أكسدة طبقة الفوسفوليبيد في وجود بيروكسيد الهيدروجين دون إنزيمات، مما يدعم بالتالي فكرة أن مثل هذه العملية تحدث في الخلية. كشفت الفلورية الذاتية لـ Fento-1 عن موضعه عند الغشاء البلازمي عندما تم إجراء التجارب في درجة حرارة منخفضة لتقليل تدفق الإدخال الخلوي (الشكل التمديدي 5أ)، مما يعيد تجسيد الخصائص الضوئية الفيزيائية للمنتج الطبيعي الأم. بالمقابل، عند درجة حرارة فسيولوجية، والتي تمكن حدوث الإدخال الخلوي، تم اكتشاف Fento-1 وmarmycin في الحجرة الداخلية الليزوزومية (الشكل 2ج). كشفت الوسم الكيميائي لرباط Chen-White المحتوي على الألكاين القابل للنقر (cCW) عن تلطيخ ضعيف على مستوى الخلايا (الشكل 2ج والشكل التمديدي 4ف)، مما يشير إلى أن الرباط غير المرتبط لا يتراكم بشكل محدد في الحجرة الداخلية الليزوزومية. علاوة على ذلك، لم يستهدف Fento-1 بشكل رئيسي عضيات أخرى (الشكل التمديدي 5ب). هذا الجزيء الصغير أدى إلى أكسدة الفوسفوليبيدات الغشائية وتقليل حيوية الخلايا في عدة خطوط خلايا سرطانية بشرية وفئران وخلايا أولية، بينما كانت آثار cCW وmarmycin هامشية (الشكل التمديدي 5ج-هـ). كشفت علم الدهون القائم على مطيافية الكتلة (MS) أن Fento-1 أدى إلى أكسدة الفوسفوليبيدات في عدة خطوط خلايا إلى حد أكبر من المحفزات المعروفة للفيروبتوز تحت هذه الظروف (الشكل التمديدي 5و، ز، المعلومات التكميلية والجدول التكميلي 2 و3). أكدت تحليلات الليزوزومات المعزولة من خلايا HT-1080 المعالجة بـ Fento-1 لمدة ساعة واحدة أكسدة الفوسفوليبيدات المرتبطة في هذه النقطة الزمنية المبكرة (الشكل التمديدي 5ح، المعلومات التكميلية والجدول التكميلي 4). بما يتماشى مع أكسدة الفوسفوليبيدات الغشائية، زادت الجرعات دون القاتلة من Fento-1 من مستويات مثبطات الفيروبتوز في خلايا HT-1080، بما في ذلك GPX4، بروتين مثبط الفيروبتوز 1 (FSP1) وناقل السيستين-جلوتامات (المكون من SLC7A11 وSLC3A2) (الشكل 2د والشكل التمديدي 5ط). كشفت البروتيوميات الكمية أن الخلايا المعالجة بجرعات دون القاتلة من Fento-1 لمدة 48 ساعة عززت تنظيم الفيروبتوز.وبروتينات مرتبطة بالليزوزوم، بما في ذلك مثبطات الفيروبتوز والاغشية المرتبطة بالليزوزوم البروتينات (LAMPs)، على التوالي، وهي نتيجة تدعم دور الليزوزومات في تحفيز الفيروبتوز (الشكل 2d، e والجدولين التكميلين 5 و6). كما تم تمييز الخلايا من خلال انخفاض تنظيم البروتينات المرتبطة بتوازن الحديد، بما في ذلك بروتين امتصاص الحديد CD44 وبروتين تخزين الحديد الفيريتين. كما أظهرت هذه الخلايا مستويات مرتفعة من مستقبل ترانسفيرين 1 (TFR1) وبروتين ربط عنصر الاستجابة للحديد 2 (IRP2)، والتي تعتبر نموذجية لنقص الحديد الخلوي.جرعات تحت القاتلة من Fento-1 عززت تقليل التعبير عن علامات الميزانشيم، والتي تم ربطها باكتساب حالات خلايا السرطان المقاومة للعلاجات القياسية.. أخيرًا، أظهرت هذه الخلايا زيادة في تنظيم عوامل النسخ الأخرى المعنية في تنظيم حالات الخلايا، مثل SOX4 و SOX13، بالإضافة إلى علامات الظهارة. تشير هذه البيانات إلى أن تأثيرات علاج Fento-1 على خلايا HT-1080، التي هي من أصل ميزانشيمي، تعود إلى اكتساب حالة خلوية مميزة تشبه تلك الخاصة بالخلايا الظهارية التي تكون عادة حساسة للعلاجات المضادة للتكاثر القياسية. تشير هذه النتيجة أيضًا إلى أن خلايا السرطان يمكن أن تتكيف مع التعرض لـ Fento-1 وتطور حالة خلوية تتحمل الفيروبتوز. أي أن الخلايا المعالجة بجرعة منخفضة من Fento-1 تظهر بقاء مستدام ونمط ظاهري مميز بسبب التكيف الخلوي بدلاً من الاختيار الكلوني.تحليلات Western blot أكدت المزيد من بيانات البروتيوم (الشكل 5j من البيانات الموسعة). علاوة على ذلك، تم الكشف عن مستويات مرتفعة من GPX4 في الفractions الغنية بالليسوزوم المعزولة من خلايا HT-1080 المعالجة بـ Fento-1 (الشكل 2f).
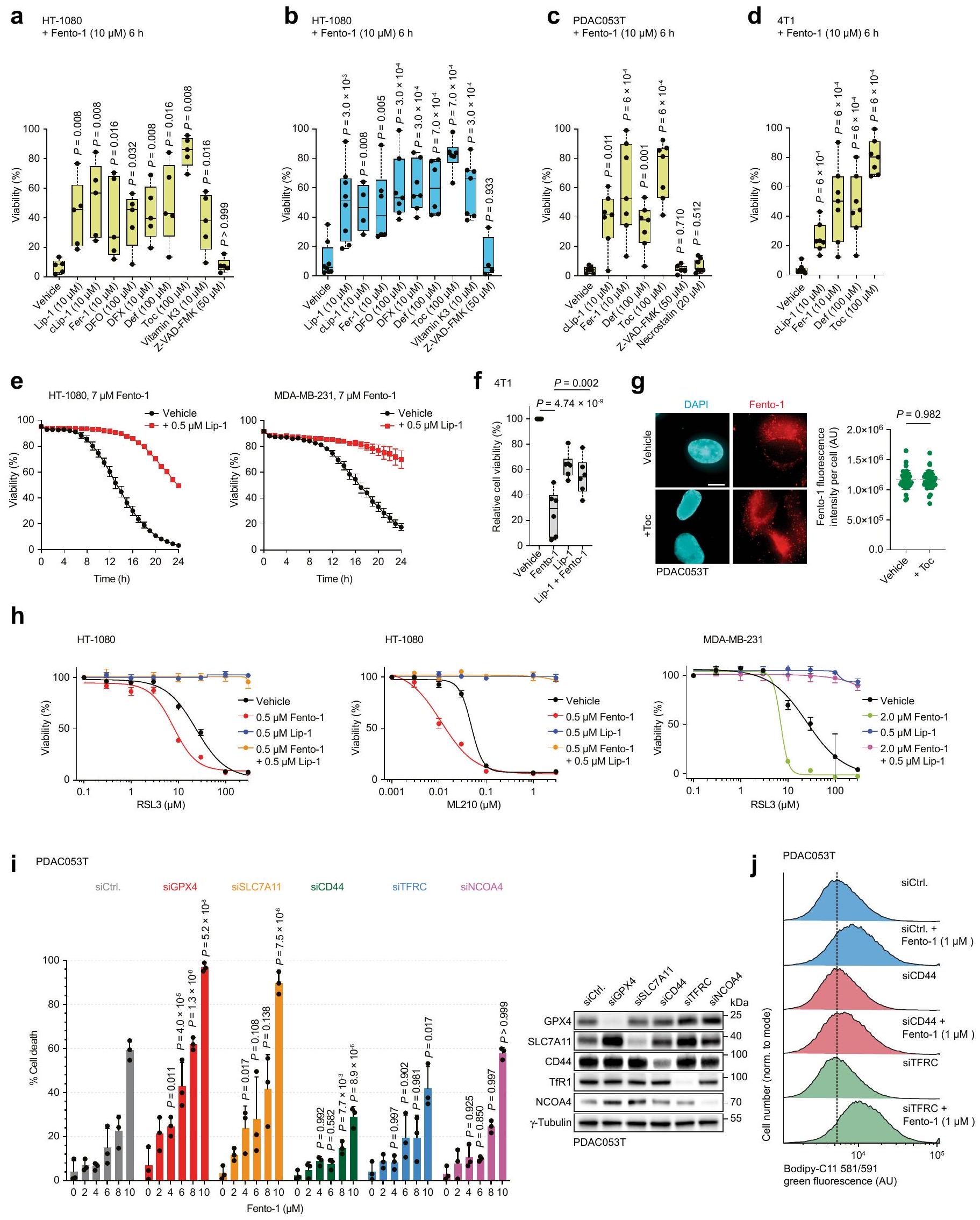
تم تقليل أكسدة الفوسفوليبيدات الغشائية التي تسببها Fento-1 بواسطة RTA المحبة للدهون.-توكوفيرول، الخالب للحديد ديفيريبيرون وLip-1 (الشكل 2g، الشكل البياني الموسع 5k، المعلومات التكميلية والجدول التكميلية 7). علاوة على ذلك، أدى العلاج بـ Fento-1 إلى إنتاج 4-هيدروكسي نونينال (4-HNE) (الشكل البياني الموسع 51)، وهو نتيجة تشير إلى الأكسدة وانهيار الفوسفوليبيدات التي تحتوي على أحماض دهنية غير مشبعة.من الجدير بالذكر أن 4 -HNE يمكن أن يسبب تلفًا خلويًا، وإنتاجه هو علامة مميزة للفيروبتوز.أدى العلاج بـ Fento-1 أيضًا إلى زيادة مستويات الليسوفوسفوليبيدات والجليسرول، مما يشير بشكل أكبر إلى تحلل الفوسفوليبيدات المؤكسدة (الشكل 2h، الشكل الإضافي 5m، المعلومات التكميلية والجدول التكميلية 8). تشير هذه النتيجة، جنبًا إلى جنب مع زيادة تنظيم عملية الأيض الدهني بعد العلاج بـ Fento-1 (الشكل 2e)، إلى أن الدهون المؤكسدة في الأغشية تحفز استجابة تلف الأغشية.تمت معاكسة موت الخلايا الذي تم تحفيزه بواسطة Fento-1 بواسطة مثبطات الفيروبتوز المعروفة، والتي شملت خافضات الحديد وRTAs، ولكن ليس بواسطة مثبطات الاستماتة أو النخر المبرمج (الشكل 2i والشكل الإضافي 6a-f). ومن الجدير بالذكر أن Fento-1 أظهر سمية متبقية لم تتمكن مثبطات الفيروبتوز من التغلب عليها بالكامل، مما يعكس قوة Fento-1.-توكوفيرول لم يمنع استهداف الليزوزوم مع Fento-1، مما يدل على أن هذا المثبط للفيروبتوزيس يمارس نشاطه الوقائي من خلال العمل كـ RTA في الدهون الغشائية وليس من خلال التنافس على التفاعل عند الغشاء البلازمي (الشكل 6g من البيانات الموسعة). كان Fento-1 قاتلاً اصطناعياً بعد المعالجة المشتركة مع مثبطات GPX4، وتم إنقاذ حيوية الخلايا بواسطة Lip-1 (الشكل 6h من البيانات الموسعة)، وهو نتيجة تؤكد النشاط المؤيد للفيروبتوزيس لـ Fento-1. أخيرًا، أدى تقليل مثبطات الفيروبتوزيس إلى زيادة حساسية الخلايا تجاه Fento-1، في حين أن تقليل Cd44 منح تأثيرًا وقائيًا (الشكل 6i من البيانات الموسعة). وبشكل متسق، فإن تقليل Cd44 حمى الخلايا جزئيًا من أكسدة الدهون الغشائية، في حين أن التثبيط الجيني لمسار امتصاص الحديد التقليدي الذي يشمل TFR1 كان أقل فعالية (الشكل 6j من البيانات الموسعة). تؤكد هذه النتيجة دور CD44 كعامل رئيسي في امتصاص الحديد في الحالة الميزنشيمية لخلايا السرطان.. بعد ذلك استكشفنا نشاط المتغيرات الهيكلية لـ Fento-1 (الشكل 7a من البيانات الموسعة). أظهر Fento-2، الذي يحتوي على رابط أطول بين المارمايسين و ligand تشين-وايت، فعالية أقل ضد خلايا PDAC والساركوما مقارنة بـ Fento-1. تدعم هذه النتيجة الفكرة القائلة بأن Fento-1 يمارس نشاطه من خلال تعزيز القرب بين محفز الحديد التفاعلي والدهون الغشائية (الشكل 7b,c من البيانات الموسعة).
الشكل 2 | تحفيز الفيروبتوز بواسطة مُحلل الفوسفوليبيد المعتمد على الحديد والاختيارية للليزوزوم. أ، التركيب الكيميائي لـ Fento-1 وتكوين المحفز الحديدي النشط بالأكسدة. ب، أعلى، توضيح لليبوزوم المحمّل بالحديد (II) و. القاع، علم الدهون للأغشية الدهنية المؤكسدة من DOPC مع الحديد (II) وFento-1. n = 3 تجارب مستقلة. ج-ي، البيانات تخص خلايا HT-1080. ج، تصوير الفلورسنت لـ Fento-1، مارمايسين وcCW المعلم.في الخلايا المعالجة عند لمدة ساعة واحدة. شريط القياس، تجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري، البروتيوميات الكمية للخلايا المعالجة بـ Fento-1. الخط العمودي المتقطع يشير إلى التعديلتجارب مستقلة. FC، تغيير الطي؛ TFs، عوامل النسخ. e، تحليل إثراء KEGG للبروتينات المرتفعة في الخلايا المعالجة بـ Fento-1تجارب مستقلة. تحليل البروتينات باستخدام تقنية الويسترن بلوت من مستخلصات إجمالية وغنية بالليزوزومات من خلايا تم معالجتها بـ Fento-1 (48 ساعة). تمثيلي لـتجارب مستقلة.-التوبولين هو عنصر تحكم في معالجة العينة. ج، علم الدهون للأفوسفوليبيدات المؤكسدة في الخلايا المعالجة بـ Fento-1 والمثبطات (أضيفت قبل ساعتين من Fento-1). تجارب مستقلة. خريطة الحرارة الكاملة موجودة في المعلومات التكميلية. ه، دهنات الليسوفوسفوليبيدات في الخلايا المعالجة بـ Fento-1 (24 ساعة).تجارب مستقلة. أنا، إطلاق اللاكتات ديهيدروجيناز (LDH) من الخلايا المعالجة بفينتو-لمدة 6 ساعاتومع إضافة المثبطات قبل ساعتين.تجارب مستقلة. تم مقارنة اختبار مان-ويتني ذو الجانبين مع المركبة. تُظهر الرسوم البيانية الصندوقية نطاق الربع، مع وجود خطوط المركز التي تشير إلى الوسيطات والشعيرات التي تمثل القيم الدنيا والقصوى. ديف، ديفيريبيرون؛ DFX، ديفيراسيروك؛ Fer-1، فيروستاتين-1؛ توك،-توكوفيرول. تم استخدام التركيزات التالية ما لم يُذكر خلاف ذلك: فينتو-1توك، تعريف وشفاه-تظهر الدهون الفوسفوليبيدية الفوسفatidylcholine (PC) في الشكل الرئيسي. يشير الحرف السفلي Oxn إلى إضافةالأكسجين. يتم عرض الفوسفatidylethanolamine (PE) والفوسفatidylinositol (PI) والفوسفatidylserine (PS) في المعلومات التكميلية. يتم عرض الدهون lysophosphatidylcholine (LPC) في الشكل الرئيسي. يتم عرض LPE و LPI و LPS في المعلومات التكميلية. بالنسبة لـ d و e، نموذج خطي، ذو جانبين.-اختبار على تغيير الطي.تم التعديل باستخدام معدل الاكتشاف الخاطئ لبيجاميني-هوشبرغ. تم إنشاء التوضيح في (ب) باستخدام BioRender (https:// biorender.com).
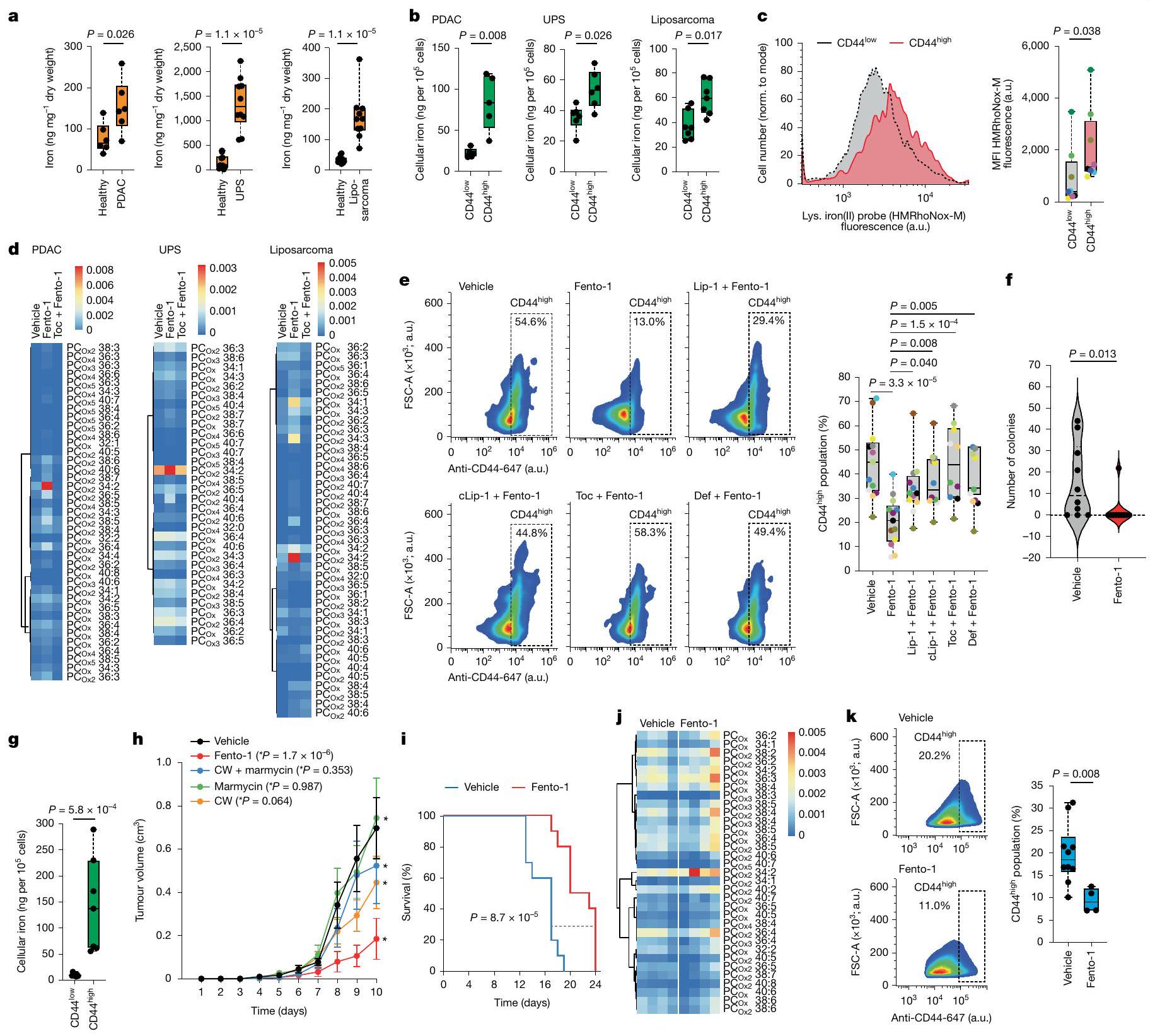
الشكل 3 | تنشيط الحديد الليزوزومي يستهدف CD44خلايا السرطان وتقلل من نمو الورم. أ، ICP-MS للأنسجة البشرية السليمة والسرطانية. PDAC،قطع لكل نسيج؛ UPS والليبو ساركوما،قطع لكل نسيج. ب، ICP-MS لخلايا سرطان الورم البشري المفككة (CD45CD31فاب“). PDAC،; يو بي إس، ورم دهنيالتكرارات لكل ورم. ج، قياس التدفق HMRhoNox-M لخلايا سرطان البنكرياس البشرية المفصولة (CD45CD31فاب). مرضى. د، علم الدهون للفوسفوليبيدات المؤكسدة لخلايا الورم البشرية المفصولة المعالجة بـ Fento-1(1 ) و Toc (قبل ساعتين). e، تحليل تدفق CD44 لخلية سرطان البنكرياس البشرية المفصولة (CD45CD31فابعولجت بـ Fento-ومثبطات (قبل ساعتين).المرضى. تحليل التباين الأحادي. اختبار تشكيل المستعمرات لخلايا سرطان DTP SUM159 بعد علاج الدوكسوروبيسين ( )، ثم فينتو-1 ( ) أو DMSO ( ) علاج لمدة 72 ساعة. البيانات هي المتوسط ± الانحراف المعياري، ICP-MS لخلايا الورم 4T1 المفروزة (CD45). نسخ.حجم الورم في حاملي ورم 4T1 فئران.فئران لكل حالة. تحليل التباين ثنائي الاتجاه.القيم (اليوم 10) مقارنةً بالمركبة. البيانات هي المتوسط ± الانحراف المعياري. CW، ligand تشين-وايت. i، الحد الأقصى لقطر الورم كنقطة نهاية البقاء للفئران الحاملة للورم. اختبار مانتل-كوكس لرتبة اللوغاريتم. j، دهنات الفوسفوليبيدات المؤكسدة في أورام الفئران 4T1 (علاج لمدة 15 يومًا).فئران لكل حالة. ك، تحليل تدفق CD44 لخلايا سرطان الورم المفصولة (CD45) من فئران تحمل ورم 4T1 المعالجة بـ Fento-1لكل حيوان كل يومين). المركبة،فأران، فينتو-الفئران. تم استخدام التركيزات التالية ما لم يُذكر خلاف ذلك:، ، “ و cLip-1 . ل و القيم مأخوذة من اختبارات مان-ويتني ذات الجانبين. لـ و كل نقطة ملونة تمثل ورمًا لمريض مختلف في اللوحة المعطاة. خرائط الحرارة الكاملة لـ و مقدمة في المعلومات التكميلية. تُظهر الرسوم البيانية الصندوقية نطاق الربع، مع خطوط مركزية تشير إلى الوسيطات والشعيرات تمثل القيم الدنيا والقصوى.
كما استبدلنا المارمايسين بالكوليسترول، وهو منتج طبيعي غير قطبي يمكنه التداخل بين الفوسفوليبيدات في الغشاء البلازمي.ولها دور في الحالة الميزانشيمية لخلايا السرطان، ربما عند الغشاء البلازمي. أدت هذه التعديلات إلى النظير غير الفلوري Fento-3، الذي أظهر قوة مشابهة لتلك الخاصة بـ
فينتو-1. معًا، توضح هذه النظائر الطبيعة المتنوعة لهذه الاستراتيجية (الشكل البياني الموسع 7 أ-ج). كتحكم، أدى استبدال الهيكل غير القطبي بموحد أكثر قطبية، مثل المورفولين، إلى إنتاج فينتو-4، الذي كان غير نشط بيولوجيًا (الشكل البياني الموسع 7 أ-ج). أخيرًا، أدى استبدال ligand تشين-وايت بـ ligand أكثر تعقيدًا هيكليًا
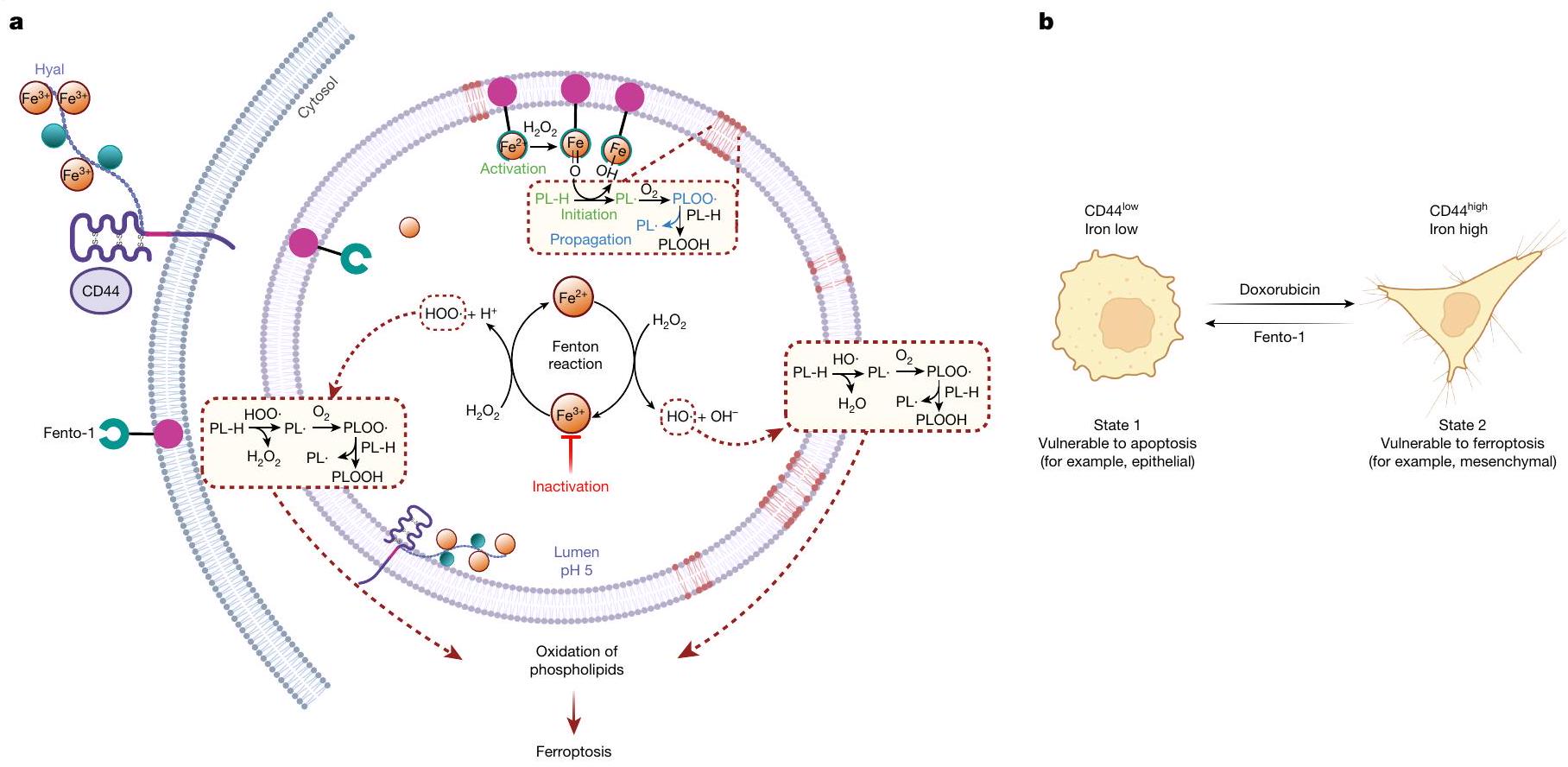
الشكل 4 | الحديد في الفيروبتوزيس وانتقال حالة الخلية. أ، يتم امتصاص الحديد عن طريق الإندوسيتوز. يحفز الحديد الليزوزومي إنتاج الجذور الحرة المرتكزة على الأكسجين من الهيدروبيروكسيدات في ظل ظروف حمضية. يمكن أن تقوم هذه الجذور بسحب هيدروجين من الفوسفوليبيدات التفاعلية لإنتاج جذور حرة مرتكزة على الكربون، مما يؤدي إلى منتجات الأكسدة والفيروبتوزيس. تعطيل
الحديد الليزوزومي يحمي الخلايا من كيمياء الحديد-الأكسدة. تنشيط الحديد الليزوزومي يؤدي إلى أكسدة الدهون الغشائية والفيروبتوز. هيال، هيالورونات ب، انتقالات حالة الخلية التي تسببها أدوية الرعاية القياسية والأدوية المؤيدة للفيروبتوز. الرسوم التوضيحية في و تم إنشاؤها باستخدام BioRender (https://biorender.com).
نورلاندير-كوستاسالليغاند المنشط للحديد أنتج Fento-5، الذي أظهر خصائص سامة للخلايا وأكسدة الدهون الغشائية مشابهة لتلك الخاصة بـ Fento-1، على الرغم من استجابة موت الخلايا المتأخرة. تعكس هذه النتيجة التفاعل المميز لمحفز الحديد تجاه الركائز العضوية. (الشكل 7a-c من البيانات الموسعة). معًا، تشير هذه البيانات إلى أن التنشيط الدوائي للحديد الليزوزومي يمكن أن يحفز الفيروبتوز.
تنشيط الحديد يقلل من نمو الورم
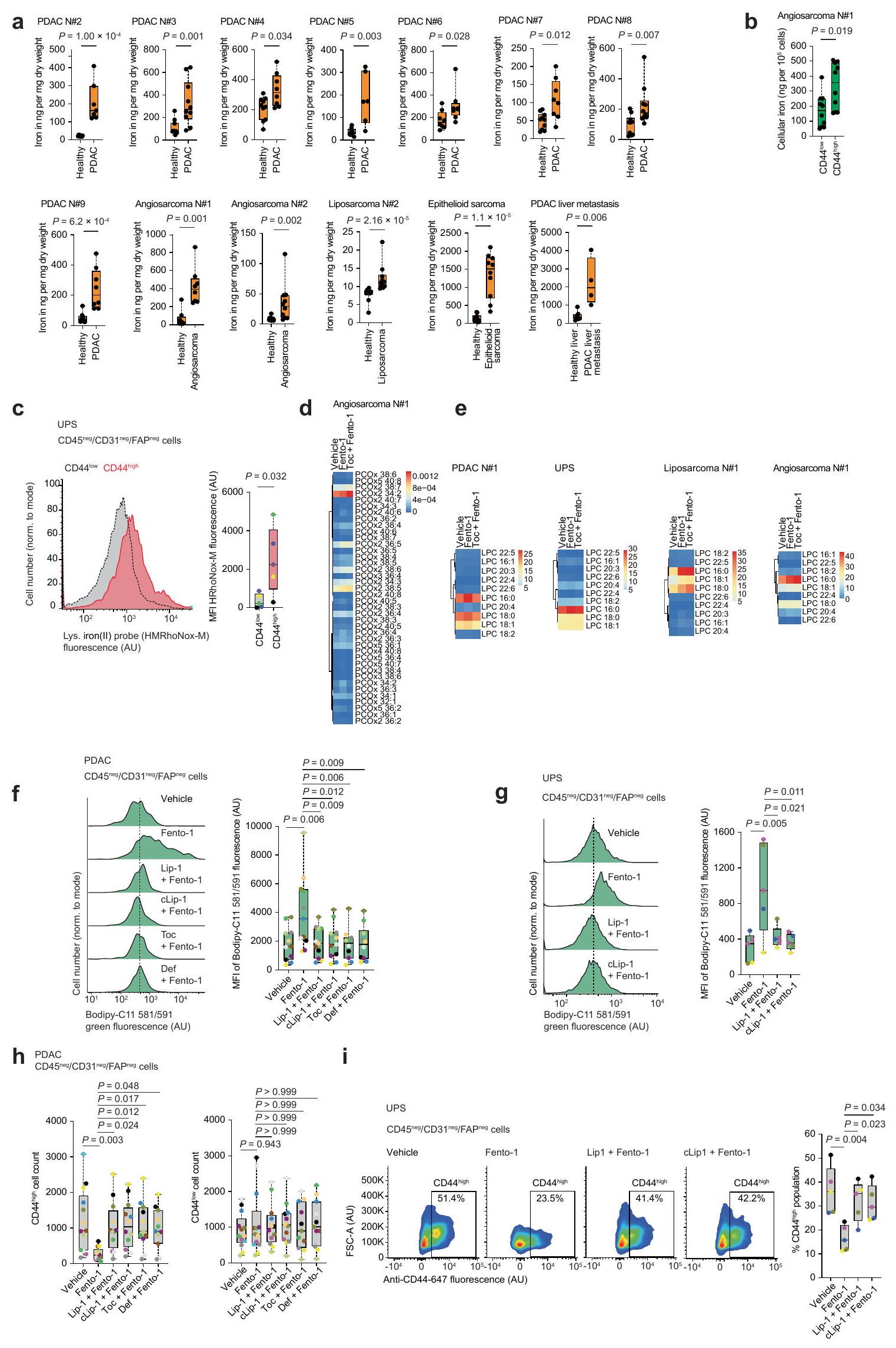
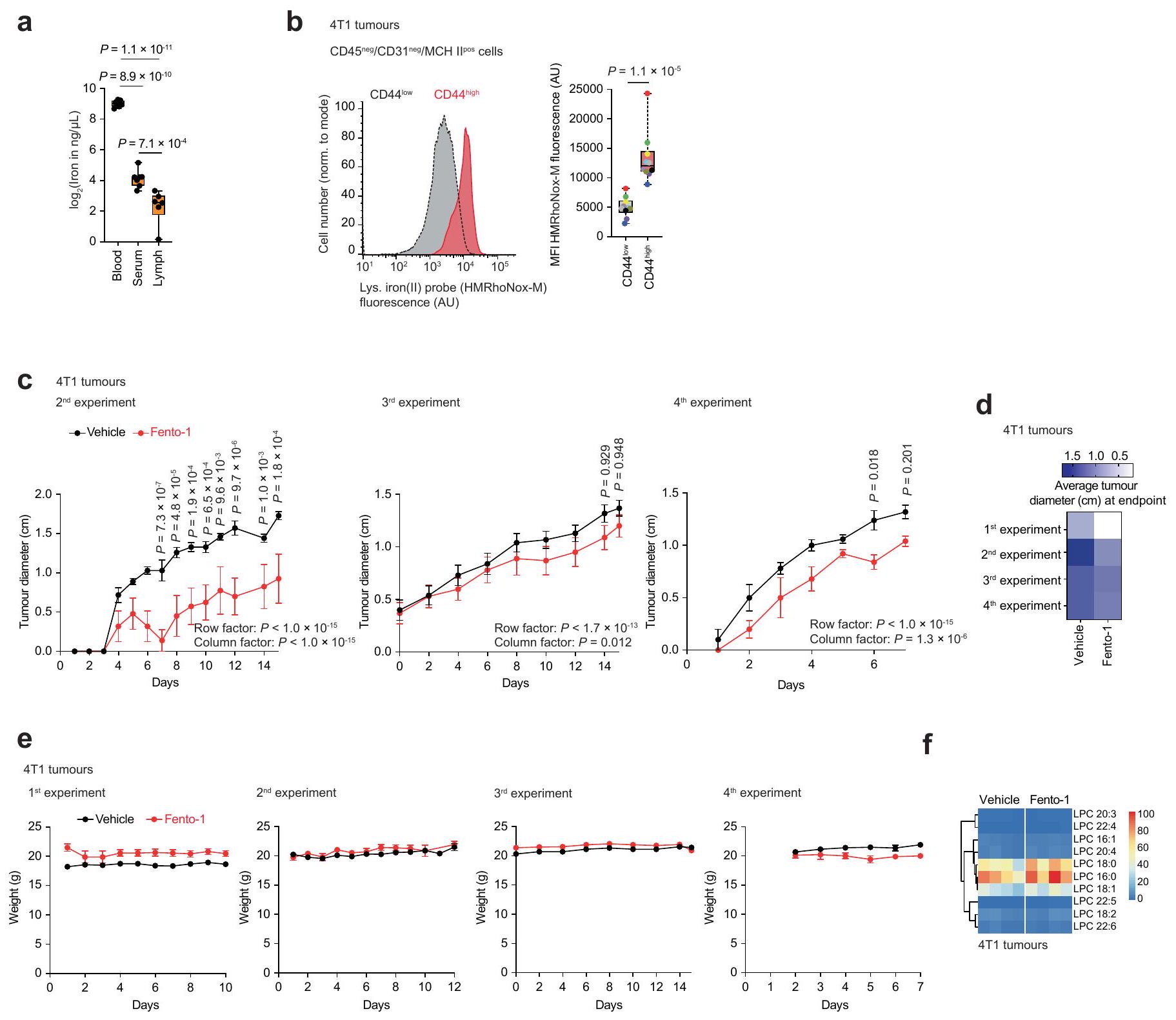
بعد ذلك، قمنا بالتحقيق في تأثير تنشيط الحديد الليزوزومي في نماذج ذات صلة بالمرض. لهذا الغرض، قمنا بتحديد محتوى الحديد في أنسجة الأورام الأولية لأنواع سرطانية مختلفة، بما في ذلك سرطان البنكرياس القنوي البشري، وأنواع مختلفة من الساركوما البشرية، ونموذج فأر لانتقال سرطان الثدي العفوي. تم اختيار هذه الأنسجة السرطانية لطبيعتها المقاومة للعلاجات القياسية وقدرتها على تشكيل النقائل، مما يساهم في نتائج سريرية سيئة. علاوة على ذلك، فقد أظهرت هذه المؤشرات أنها عرضة للفيروبتوز..
كان محتوى الحديد الكلي أعلى في أنسجة السرطان من سرطان البنكرياس القنوي (PDAC) وساركوما غير المتمايزة (UPS) وساركوما الأوعية الدموية وساركوما الدهون وساركوما الظهارة مقارنةً بالأنسجة غير السرطانية المجاورة، كما تم قياسه بواسطة مطيافية الكتلة المتصلة بالتحريض (ICP-MS) (الشكل 3أ والشكل التمديدي 8أ). علاوة على ذلك، كان الحمل الخلوي للحديد أعلى في تحت مجموعات خلايا السرطان ذات التعبير العالي عن CD44 (الشكل 3ب والشكل التمديدي 8ب). أظهرت خلايا من أنسجة PDAC وساركوما البشرية الأولية المفرومة حديثًا مستويات أعلى من الحديد(II) الليزوزومي النشط في حالة الأكسدة في CD44.تحت مجموعات من خلايا السرطان أكثر من CD44 الخاص بهمالنظائر (الشكل 3c والشكل الإضافي 8c). هذه النتيجة تتماشى مع النتائج السابقة التي تظهر أن CD44 يساهم في ابتلاع الحديد في خلايا السرطان التي اكتسبت نمط DTP.. في الخلايا المستخلصة من الأورام الأولية البشرية المفككة حديثًا، أدى تحفيز Fento-1 إلى الأكسدة وتحلل الدهون للفوسفوليبيدات الغشائية، وهو تأثير تم معاداته بواسطة مثبطات الفيروبتوز (الشكل 3d الشكل التمديدي 8d-g، المعلومات التكميلية و
الجداول التكميلية 9 و 10). كما قلل فنتو-1 من عدد CD44الخلايا في سرطان البنكرياس الغدي البشري المنفصل وUPS، وقد تم عكس هذا التأثير بواسطة مثبطات الفيروبتوز (الشكل 3e والشكل الممتد 8h، i).
في خلايا PDAC الأولية والأورغانويدات المشتقة من PDAC البشري، قلل Fento-1 من حيوية الخلايا بشكل أقوى من الأدوية القياسية (الشكل التمديدي 9a، b) وتعاون مع هذه الأدوية في عدة خلايا سرطان أولية من PDAC البشري (الشكل التمديدي 9c، d). في اختبار تشكيل المستعمرات، قضى Fento-1 على DTP لخلايا سرطان الثدي الثلاثي السلبية SUM159 التي تعبر عن CD44 والتي نجت من العلاج بالدكسوروبسين (الشكل 3f). تماشيًا مع ذلك، كان Fento-1 أكثر فعالية ضد خلايا HT-1080 التي تم علاجها بجرعات منخفضة من الدكسوروبسين لفترة طويلة من الزمن (الشكل التمديدي 9e). في ظل هذه الظروف، اعتمدت خلايا HT-1080 شكلًا مميزًا وتميزت بزيادة مستويات CD44 والفيريتين مع زيادة علامات الميزانشيم (الشكل التمديدي 9f-h). تشير هذه البيانات إلى أن الخلايا المعرضة للدكسوروبسين تزيد من CD44 لتعزيز امتصاص الحديد وتعتمد نمطًا ميزانشيميًا أكثر للهروب من الحالة الحساسة للدكسوروبسين. تؤدي هذه التكيفات إلى زيادة القابلية للفيروبتوز والحساسية لـ Fento-1. على العكس، أظهرت خلايا HT-1080 المعرضة لجرعات منخفضة من Fento-1 مستويات منخفضة من CD44 والفيريتين مع انخفاض علامات الميزانشيم، مما يشير إلى أن هذه الخلايا تتكيف مع استهداف الحديد الليزوزومي (الشكل 2d والشكل التمديدي 9g، i).
تعتبر آفات سرطان العقد اللمفاوية الإقليمية مؤشرات على النقائل البعيدة والوفيات، وتكون الخلايا المنتشرة محمية من الفيروبتوز في اللمف.أظهرت القياسات في الفئران أن مستويات الحديد الكلية في سائل اللمف كانت أقل من تلك الموجودة في الدم والمصل (الشكل 10a من البيانات الموسعة). وبالتالي، قمنا بتقييم تأثير Fento-1 على بقاء خلايا السرطان مباشرة في الأوعية اللمفاوية. لهذا الغرض، استخدمنا نموذج الفئران Balb/c المناعي 4T1 لسرطان الثدي الثلاثي السلبي العفوي. أظهرت الكمية من الحديد في الخلايا المعزولة من أورام 4T1 داخل العقد اللمفاوية حمولة حديد أعلى في CD44.خلايا السرطان أكثر من CD44المجموعات الفرعية (الشكل 3g). هذه
CD44أظهرت الخلايا أيضًا نشاطًا أعلى في اختزال الحديد (II) في الجسيمات الحالّة (الشكل 10b من البيانات الموسعة). عندما تم علاج الفئران التي تحتوي على أورام 4T1 داخل العقد اللمفاوية بـ Fento-1 (إدارة داخل اللمف كل يومين)، أظهرت الفئران انخفاضًا في نمو الورم، وزيادة في نقاط البقاء المستندة إلى حجم الورم، وعدم وجود آثار سلبية على وزن الجسم (الشكل 3h,i والشكل 10c-e من البيانات الموسعة). أظهرت التحليلات الإضافية للأورام المتبقية أن Fento-1 أدى إلى أكسدة الدهون الغشائية وإنتاج الليسوفوسفوليبيدات، مما أثر بشكل تفضيلي على CD44.على CD44تحت مجموعات خلايا السرطان (الشكل 3j، k، الشكل البياني الموسع 10f، المعلومات التكميلية والجداول التكميلية 11 و12). معًا، تُظهر هذه البيانات أن Fento-1 يحفز الموت الخلوي الحديدي في الجسم الحي من خلال استغلال الوفرة الأعلى من الحديد(II) الليزوزومي في CD44.تحت مجموعات خلايا السرطان.
نقاش
الفيروبتوز هو شكل من أشكال موت الخلايا الناتج عن الأكسدة غير المنضبطة للدهون الغشائية. حيث وكيف يتم بدء ذلك في الخلية لا يزال غير واضح. يمكن أن يتفاعل الحديد مع الهيدروبيروكسيدات لبدء أو نشر تفاعل سلسلة جذرية بشكل مستقل عن الإنزيمات. تعتبر الحويصلات الليزوزومية منظمًا حاسمًا لتوازن الحديد الخلوي.وطبيعتها الحمضية، جنبًا إلى جنب مع وجود الحديد التفاعلي وبيروكسيد الهيدروجين، توفر بيئة كيميائية مثالية لتحفيز أكسدة الفوسفوليبيدات الغشائية (الشكل 4أ). تقوم مجموعات فرعية محددة من خلايا السرطان بزيادة تنظيم بروتين امتصاص الحديد CD44 لتعزيز إزالة الميثيل المؤكسدة للعلامات الكروماتينية القمعية وفتح التعبير عن الجينات التي تنظم انتقالات حالة الخلية المرتبطة بتقدم السرطان.. وبالتالي، على الرغم من أن مستويات الحديد المرتفعة تمكّن هذه الخلايا من اكتساب ملف DTP، إلا أن هذه السمة تمنح أيضًا بشكل حتمي قابلية للتعرض للفيروبتوسيس. (الشكل 4ب). إن حقيقة أن CD44 يميز أيضًا التطور، والتجديد الذاتي، وشفاء الجروح، وتنشيط خلايا المناعة تشير إلى أن الحديد الليزوزومي يساهم أيضًا في تحفيز الفيروبتوز في هذه الإعدادات البيولوجية. إن تعزيز النشاط الأكسدي للحديد الليزوزومي من خلال التدخلات الجينية للقضاء على خلايا السرطان يمثل تحديًا. هنا قمنا بتطوير مُحلل للفوسفوليبيد يستفيد من الحمل العالي للحديد في مجموعات فرعية معينة من خلايا السرطان لتحفيز الفيروبتوز عن طريق تنشيط الحديد الليزوزومي (II) (الشكل 4أ). أدت جرعات غير مميتة من هذا المحلل إلى اكتساب حالة خلوية مقاومة للفيروبتوز، والتي أعادت جزئيًا ميزات الخلايا الظهارية (الشكل 4ب). تسلط هذه النتيجة الضوء على قيمة دمج الأدوية المحددة لحالة الخلية لعلاج السرطان. يمكن أن يتسبب دواء السرطان دوكسوروبيسين في إحداث ضرر للحمض النووي الجيني في الخلايا المتكاثرة، مما يؤدي، إذا لم يتم إصلاحه، إلى تحفيز الموت الخلوي المبرمج.على العكس من ذلك، فإن حالة الخلايا المقاومة للفيروبتوز، التي تتميز بزيادة تنظيم مثبطات الفيروبتوز، والهيكليين الإيبوكسيديين، والفوسفوليباز، وزيادة في عملية الأيض الدهني (الشكل 2د)، يمكن أن تنشأ نتيجة لقدرة الخلايا على إصلاح الأغشية التالفة بكفاءة. تظهر هذه الدراسة أن ضبط النشاط الأكسدي للحديد الليزوزومي يوفر تحكمًا في أكسدة الدهون الغشائية، مما يدعم فكرة أن الحديد الليزوزومي هو محفز للفيروبتوز. يمكن الآن استخدام هذه المعرفة في تصميم معدلات الفيروبتوز. يتمتع Fento-1 بنوع كيميائي مميز للتحقيق في الفيروبتوز ويوفر إطارًا مفاهيميًا لاستهداف خلايا سرطان DTP..
المحتوى عبر الإنترنت
أي طرق، مراجع إضافية، ملخصات تقارير Nature Portfolio، بيانات المصدر، بيانات موسعة، معلومات تكميلية، شكر وتقدير، معلومات مراجعة الأقران؛ تفاصيل مساهمات المؤلفين والمصالح المتنافسة؛ وبيانات توفر البيانات والرموز متاحة علىhttps://doi.org/10.1038/s41586-025-08974-4.
2. فريدمان أنجيلي، ج. ب. وآخرون. تعطيل منظم الفيروبتوز Gpx4 يؤدي إلى فشل كلوي حاد في الفئران. نات. خلية بيولوجيا. 16، 1180-1191 (2014). 3. بيرسوكير، ك. وآخرون. إن إنزيم CoQ الأكسيدوريدوكتاز FSP1 يعمل بالتوازي مع GPX4 لمنع الفيروبتوز. ناتشر 575، 688-692 (2019). 4. دول، س. وآخرون. FSP1 هو مثبط للفيروبتوز غير المعتمد على الجلوتاثيون. ناتشر 575، 693-698 (2019). 5. يانغ، و. س. و ستوكويل، ب. ر. تحديد الفحص القاتل الاصطناعي للمركبات التي تنشط موت الخلايا غير المبرمج المعتمد على الحديد في خلايا السرطان الحاملة لـ RAS المسرطنة. كيم. بيول. 15، 234-245 (2008). 6. مولر، س. وآخرون. CD44 ينظم اللدونة الجينية عن طريق التوسط في ابتلاع الحديد. نات. كيم. 12، 929-938 (2020). 7. سولير، س. وآخرون. مسار إشارة النحاس القابل للعلاج الذي يحفز الالتهاب. ناتشر 617، 386-394 (2023). 8. ماي، ت. ت. وآخرون. سالينوميسين يقتل خلايا السرطان الجذعية عن طريق احتجاز الحديد في الليزوزومات. نات. كيم. 9، 1025-1033 (2017). 9. رودريغيز، ر.، شرايبر، س. ل. وكونراد، م. خلايا السرطان المستمرة: إدمان الحديد والضعف تجاه الفيروبتوز. مول. سيل 82، 728-740 (2022). 10. فرانكا، ج. س. وآخرون. التكيف الخلوي مع علاج السرطان على طول استمرارية المقاومة. ناتشر 631، 876-883 (2024). 11. فينتون، هـ. ج. هـ. أكسدة حمض الطرطريك في وجود الحديد. مجلة الجمعية الكيميائية 65، 899-910 (1894). 12. سيلر، أ. وآخرون. إن إنزيم الجلوتاثيون بيروكسيداز 4 يستشعر ويترجم الإجهاد التأكسدي إلى موت الخلايا المعتمد على 12/15 ليبكسجيناز والمُدار بواسطة AIF. ميتابوليزم الخلايا. 8، 237-248 (2008). 13. تشين، إكس، كانغ، ر، كرويمر، ج. وتانغ، د. تنظيم محدد للعضيات للفيروبتوسيس. موت الخلايا والتمايز. 28، 2843-2856 (2021). 14. زو، ي. وآخرون. مرونة الدهون الأثيرية تعزز القابلية للإصابة بالفيروبتوز وتجنبها. الطبيعة 585، 603-608 (2020). 15. قاو، م. وآخرون. دور الميتوكوندريا في الفيروبتوز. مول. خلية 73، 354-363 (2019). 16. ديكسون، س. ج. وآخرون. التثبيط الدوائي لتبادل السيستين-جلوتامات يسبب إجهاد الشبكة الإندوبلازمية والفيروبتوز. eLife 3، eO2523 (2014). 17. توري، س. وآخرون. دور أساسي للليسوزومات الوظيفية في الفيروبتوزيس لخلايا السرطان. مجلة الكيمياء الحيوية 473، 769-777 (2016). 18. أنطوشاك، م. وآخرون. بروتينات حساسة للحديد تحفز الفيروبتوز النشط في خلايا سرطان البنكرياس المقاومة للعلاج. ج. أم. كيم. سوس. 144، 11536-11545 (2022). 19. فون كروزنشتيرن، أ. ن. وآخرون. تحديد المواقع الأساسية لعملية أكسدة الدهون في الفيروبتوز. نات. كيم. بيول. 19، 719-730 (2023). 20. جيانغ، إكس، ستوكويل، ب. ر. وكونراد، م. فيروبتوسيس: الآليات، البيولوجيا والدور في المرض. نات. ريف. مول. سيل. بيول. 22، 266-282 (2021). 21. كانيكي، ت.، مولر، س. ورودريغيز، ر. تصور الجزيئات الصغيرة النشطة بيولوجيًا في الخلايا باستخدام كيمياء النقر. نات. ريف. كيم. 2، 202-215 (2018). 22. زيلكا، أ. وآخرون. حول آلية الحماية الخلوية بواسطة فيروستاتين-1 وليبروكستاتين-1 ودور أكسدة الدهون في موت الخلايا الناتج عن الفيروبتوز. ACS Cent. Sci. 3، 232-243 (2017). 23. شاه، ر.، فارمر، ل. أ.، زيلكا، أ.، فان كيسل، أ. ت. م. و برات، د. أ. ما وراء DPPH: استخدام التثبيط المعتمد على الفلورية للتنبؤ بإنقاذ موت الخلايا التأكسدي. كيمياء الخلايا. بيولوجيا 26، 1594-1607 (2019). 24. شونبرغ، د. ل. وآخرون. تميز النقل التفضيلي للحديد خلايا الجذعية الشبيهة بالورم الدبقي. خلية السرطان 28، 441-455 (2015). 25. باسولي، د. وآخرون. إدمان الحديد: هدف علاجي جديد في سرطان المبيض. أونكوجين 36، 4089-4099 (2017). 26. Elgrishi، ن. وآخرون. دليل عملي للمبتدئين في الفولتمترية الدورية. مجلة التعليم الكيميائي 95، 197-206 (2018). 27. وي، ي.، أيدين، ز.، تشانغ، ي.، ليو، ز. وغو، م. جهاز استشعار فلوري متحول للتصويرفي خلايا عصبية حية بدقة تحت خلوية. ChemBioChem 13، 1569-1573 (2012). 28. نوا، م.، هيراياما، ت.، أوكودا، ك. وناجاساوا، هـ. فئة جديدة من مجسات الفلورسنت الانتقائية لـ Fe(II) ذات التباين العالي تعتمد على هياكل حلزونية لتصور الحديد القابل للتغيير داخل الخلايا الذي يتم توصيله بواسطة الترانسفيرين. الكيمياء العضوية والجزيئية 12، 6590-6597 (2014). 29. فيسواناثان، ف. س. وآخرون. اعتماد حالة مقاومة العلاج لخلايا السرطان على مسار بيروكسيداز الدهون. ناتشر 547، 453-457 (2017). 30. هانغاور، م. ج. وآخرون. خلايا السرطان المقاومة للعقاقير عرضة لتثبيط GPX4. ناتشر 551، 247-250 (2017). 31. غونترت، يو. وآخرون. نوع جديد من الجليكوبروتين CD44 يمنح الخلايا السرطانية في الجرذان القدرة على الانتقال. خلية 65، 13-24 (1991). 32. كانيكي، ت. وآخرون. تخليق المارمايسين A والتحقيق في نشاطه الخلوي. نات. كيم. 7، 744-751 (2015). 33. تشين، م. س. ووايت، م. س. تفاعل أكسدة C-H الأليفاتي الانتقائي بشكل متوقع لتخليق الجزيئات المعقدة. ساينس 318، 783-787 (2007). 34. هيرتزبيرغ، ر. ب. وديرفان، ب. ب. انقسام الحمض النووي الحلزوني المزدوج بواسطة ميثيديم-بروبيل-EDTA-حديد(II). مجلة الجمعية الكيميائية الأمريكية 104، 313-315 (1982). 35. شرايبر، س. ل. صعود اللصقات الجزيئية. خلية 184، 3-9 (2021). 36. تشيو، ب. وآخرون. الفوسفوليبيدات ذات ذيول الأحماض الدهنية المتعددة غير المشبعة تعزز الفيروبتوز. خلية 187، 1177-1190 (2024). 37. رودريغيز، ر. وآخرون. إشارات أيونات المعادن في الطب الحيوي. مراجعة الكيمياء 125، 660-744 (2025). 38. مارين، ج. س.، داوسون، س. ج. وداوسون، م. أ. آليات المقاومة العلاجية غير الوراثية في السرطان. نات. ريف. كانسر 20، 743-756 (2020). 39. ين، هـ.، شو، ل. وبورتر، ن. أ. أكسدة الدهون بواسطة الجذور الحرة: الآليات والتحليل. مراجعة الكيمياء 111، 5944-5972 (2011). 40. داي، إ. وآخرون. دليل حول النظام البيئي الجزيئي الذي ينظم الفيروبتوسيس. نات. بيولوجيا الخلايا 26، 1447-1457 (2024). 41. بامبلونا، ر. الفوسفوليبيدات الغشائية، الضرر الناتج عن الأكسدة الدهنية والنزاهة الجزيئية: دور سببي في الشيخوخة وطول العمر. بيوكيمي. بيوفيز. أكتا 1777، 1249-1262 (2008). 42. سيزجين، إ.، ليفنتال، إ.، مايور، س. وإيغيلينغ، س. لغز تنظيم الغشاء: التركيب، التنظيم وأدوار طوفان الدهون. نات. ريف. مول. سيل. بيول. 18، 361-374 (2017). 43. عبد الله، ن.، فينسنت، س. ت. وكور، م. رؤى آلية توضح دور الكوليسترول في الانتقال الظهاري الميزانشيمي ومقاومة الأدوية في السرطان. فرونت. خلية تطوير. بيولوجيا. 9، 728325 (2021). 44. ميترا، م. وآخرون. أكسدة الإيبوكسيد عالية الانتقائية الإنزيمية للأوليفينات بواسطةتم تحفيزه بواسطة محفز غير هيمي Fe(II) من ligand رباعي الأسنان غير متماثل. دالتون ترانس. 48، 6123-6131 (2019).
مقالة
فورستنر، أ. تحفيز الحديد في التخليق العضوي: تقييم نقدي لما يتطلبه جعل هذا المعدن الأساسي بطلاً متعدد المهام. ACS Cent. Sci. 2، 778-789 (2016).
بادجلي، م. أ. وآخرون. نقص السيستين يحفز موت الخلايا بالحديد في أورام البنكرياس في الفئران. ساينس 368، 85-89 (2020).
أوبلاكير، ج. م. وآخرون. اللمف يحمي خلايا الميلانوما المنتشرة من الفيروبتوسيس. ناتشر 585، 113-118 (2020).
ريزولو، ف.، مور، س.، فانغيلوي، ب. وأغوستينيس، ب. الليزوزوم كمنظم رئيسي لتمثيل الحديد. اتجاهات العلوم البيوكيميائية 46، 960-975 (2021).
رودريغيز، ر. وآخرون. الضرر الناتج عن الجزيئات الصغيرة في الحمض النووي يحدد هياكل بديلة للحمض النووي في الجينات البشرية. نات. كيم. بيو. 8، 301-310 (2012).
ملاحظة الناشر: تظل شركة سبرينغر ناتشر محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
الوصول المفتوح هذه المقالة مرخصة بموجب رخصة المشاع الإبداعي النسبية-غير التجارية-بدون مشتقات 4.0 الدولية، التي تسمح بأي استخدام غير تجاري، ومشاركة، وتوزيع، وإعادة إنتاج في أي وسيلة أو صيغة، طالما أنك تعطي الائتمان المناسب للمؤلفين الأصليين والمصدر، وتوفر رابطًا لرخصة المشاع الإبداعي، وتوضح إذا قمت بتعديل المادة المرخصة. ليس لديك إذن بموجب هذه الرخصة لمشاركة المواد المعدلة المشتقة من هذه المقالة أو أجزاء منها. الصور أو المواد الأخرى من طرف ثالث في هذه المقالة مشمولة في رخصة المشاع الإبداعي الخاصة بالمقالة، ما لم يُشار إلى خلاف ذلك في سطر الائتمان للمادة. إذا لم تكن المادة مشمولة في رخصة المشاع الإبداعي الخاصة بالمقالة واستخدامك المقصود غير مسموح به بموجب اللوائح القانونية أو يتجاوز الاستخدام المسموح به، ستحتاج إلى الحصول على إذن مباشرة من صاحب حقوق الطبع والنشر. لعرض نسخة من هذه الرخصة، قم بزيارة http:// creativecommons.org/licenses/by-nc-nd/4.0/. (ج) المؤلف(ون) 2025
طرق
الأخلاق
تم الحصول على عينات جديدة من الأورام عن طريق الجراحة من مرضى في مراكز رعاية بول بروسي ومعهد كوري. قدم جميع المرضى موافقة خطية مستنيرة لاستخدام عينات الأورام. تمت الموافقة على الدراسة من قبل اللجان التنظيمية المؤسسية (رقم 587 و DATA190160). تم إجراء جميع تجارب cLip-1 في الجسم الحي وفقًا لقانون الرفق بالحيوانات الألماني وتمت الموافقة عليها من قبل اللجنة المؤسسية لتجارب الحيوانات وحكومة بافاريا العليا (رقم ROB-55.2-2532.Vet_02-18-13). امتثلت جميع تجارب حقن العقد اللمفاوية في الفئران لجميع اللوائح الأخلاقية ذات الصلة وتمت وفقًا للبروتوكولات المعتمدة من قبل لجنة رعاية واستخدام الحيوانات المؤسسية في مدرسة هارفارد تي. إتش. تشان للصحة العامة (البروتوكول IS00003460). بالنسبة لجمع اللمف والدم من الفئران، تم إجراء تجارب الحيوانات وفقًا لإرشادات المجتمع الأوروبي لرعاية واستخدام الحيوانات. تم إجراء تجارب الحيوانات بما يتماشى مع الإرشادات الفرنسية للتعامل مع الحيوانات وتمت الموافقة عليها من قبل اللجان الأخلاقية المحلية (رقم الاتفاقية). ).
التركيب الكيميائي
تم شراء المواد الأولية بأعلى جودة تجارية واستخدمت دون مزيد من التنقية ما لم يُذكر خلاف ذلك. تم الحصول على المذيبات اللامائية عن طريق تمرير المذيبات المنزوعة الغاز عبر مصافي جزيئية وأعمدة الألومينا المنشطة. تم مراقبة التفاعلات بواسطة كروماتوغرافيا الطبقة الرقيقة (TLC) باستخدام ألواح الألمنيوم المغلفة بهلام السيليكا أو أكسيد الألمنيوم المحايد من ميرك.تمت رؤية ألواح TLC بواسطة الأشعة فوق البنفسجية أو عن طريق المعالجة بمحلول نينهدين، أو موليبدات الأمونيوم السيري، أو برمنغنات البوتاسيوم والتسخين. تم تنقية نواتج التفاعل بواسطة كروماتوغرافيا العمود السريع على هلام السيليكا 60 (230-400 شبكة، ماشيراي ناغل) أو أكسيد الألمنيوم (نشط محايد، سيغما-ألدريش) باستخدام نظام CombiFlash NextGen و HPLC التحضيري Gradient رباعي 2545 المزود بكاشف مصفوفة الفوتوديود 2998 (واترز) المجهز بعمود عكسي (XBridge BEH C18 OBD prep col).تم إجراء مطيافية الرنين المغناطيسي النووي باستخدام أجهزة Bruker 400 أو 500 ميغاهرتز. تم تشغيل الطيف في الميثانول-ثنائي ميثيل سلفوكسيد-كلوريد الميثيلين-أو كلوروفورم-عند 298 كلفن أو 310 كلفن كما هو موضح.التحولات الكيميائيةيتم التعبير عنها بوحدات ppm باستخدام المذيب غير الديوتيري كمعيار داخلي، وتحدد ثوابت الاقتران J بوحدات Hz. يتم استخدام الاختصارات التالية: bs، قمة عريضة؛ s، قمة مفردة؛ d، قمة مزدوجة؛ dd، قمة مزدوجة من مزدوجات؛ ddd، قمة مزدوجة من مزدوجة من مزدوجات؛ dt، قمة مزدوجة من ثلاثيات؛ dq، قمة مزدوجة من رباعيات؛ q، قمة رباعية؛ t، قمة ثلاثية؛ td، قمة ثلاثية من مزدوجات؛ quint.، قمة خماسية؛ و m، مجموعة.التحولات الكيميائيةيتم التعبير عنها بوحدات ppm باستخدام المذيب غير الديوتيري كمعيار داخلي. تم تحديد نقاء المركبات النهائية ليكون >98% بواسطة UPLC-MS. تم تسجيل طيف الكتلة منخفض الدقة باستخدام جهاز Waters Acquity H-class مزود بكاشف مصفوفة ثنائية الأبعاد وكاشف SQ (UPLC-MS) مزود بعمود عكسي الطور (Acquity UPLC BEH C18).تم تسجيل طيف الكتلة عالي الدقة على جهاز Thermo Scientific Q-Exactive Plus المزود بجهاز Robotic TriVersa NanoMate Advion. يتم تفصيل إجراءات تخليق الجزيئات الصغيرة في المعلومات التكميلية.
تجارب المعايرة باستخدام الرنين المغناطيسي النووي
تم تسجيل طيف NMR على جهاز مطياف Bruker بتردد 500 ميجاهرتز عند 310 كلفن باستخدام برنامج Bruker Topspin (الإصدار 4.1.4) وتم تحليله باستخدام برنامج MestRenova (الإصدار 5.0.1-35756) أو برنامج Bruker Topspin (الإصدار 4.1.4)، والانزياحات الكيميائيةيتم التعبير عنها بوحدات جزء في المليون (ppm) باستخدام إشارات المذيبات غير الديوتيرية المتبقية كمعيار داخلي. تم تحضير جميع المحاليل في الميثانول-. لتسجيلات طيف الرنين المغناطيسي النووي لـ Lip-1 المضاف إليهقبل إضافة TFA أو الصوديوم ديتروكسيدمعادلة محلول لـ (ألفا أيسر، 12357، ، 145 مليمول) أضيفت إلى محلول Lip-1 (سيغما-ألدريتش، SML1414، 1 ملغ في ). ثم، تم تحضير محلول من TFA (سيغما-ألدريتش، T6508، الدفعة STBG1988V، ) أو محلول من ديوتيريد الصوديوم (Eurisotop، D076Y، الدفعة R2621، ) تم إضافته. لتسجيلات طيف الرنين المغناطيسي النووي لـ Lip-1 والنفتالين المعاير بـمعادلة محلول لـتمت إضافته إلى محلول Lip-1 (1 ملغ فيونفتالين (سيغما-ألدريتش،بعد كل إضافة، تم تحريك المحلول لبضع ثوان وسُجلت طيفيات الرنين المغناطيسي النووي.
الجهد الدوري
الجهد الدوريتم إجراء التجارب باستخدام خلية ذات ثلاثة أقطاب. تم استخدام قطب زئبقي مشبع كمرجع، وتم اختيار قطب كربون زجاجي ثابت بقطر 3 مم كقطب عمل، وسلك من البلاتين كقطب مضاد. تم تسجيل جميع الفولتموجرامات الدورية في درجة حرارة الغرفة مع-autolab III من ميتروهيم باستخدام برنامج نوفا (الإصدار 2.1) بمعدل مسحتم استخدام الأسيتونيتريل والميثانول من درجة HPLC للتسجيلات. لجميع التجارب،في الأسيتونيتريلتم استخدام محلول مخزون. تم استخدام 10 مل من محلول مخزونتم تحضيره بـمنمحلول في ماء الميلي Q و 9.5 مل من الأسيتونيتريل. ثم،تم إضافة (0.2 معادل) من محلول مخزون بتركيز 40 مللي مولار من DFO، Lip-1، metcLip-1 (محلي الصنع) أو alcLip-1 (محلي الصنع، مذاب في الأسيتونيتريل أو الميثانول) إلى 1 مل منتم الوصول إلى 1.0 مكافئ من المحلول. بعد كل إضافة، تم تحريك المحلول لبضع ثوانٍ وتم تسجيل الفولتموجرامات.
النمذجة الجزيئية
تم تحسين جميع الهياكل باستخدام مجموعة برامج Gaussian 16 (المرجع 50) على مستوى BHLYP/SVP لجميع الذرات (حالة دوران سداسية لمركبات الحديد(III)) (Gaussian 16، المراجعة C.01). تم حساب التصحيح الحراري للطاقة الحرة لجيبس عند 310.15 كلفن. تم إجراء نقاط مفردة على مستوى UMP2/SVP باستخدام نموذج الإذابة SMD (الميثانول). النتائج المقدمة هيفي.
الفلوريمترية لمسبار الحديد (II) RPE
تم تخفيف محلول RPE (محلي، 10 مللي مول) في DMSO بمحلول من الأسيتونيتريل-مخزن الأسيتاتللوصول إلى تركيز منفي لوحة مكونة من 96 بئرًا، ثلاث حلول من RPE10 مللي مول و 10 أو 20 أو 50 معادلة من Lip-1 (داخلي 1، 2 أومحلول مخزون بتركيز 10 مللي مولار في الميثانول، على التوالي) تم تحضيرها وتم خلط المحاليل عدة مرات. ثم، 10 معادلات. ( محلول مخزون في محلول أسيتونيتريل-أسيتات ( )) تم إضافته إلى كل حل وخلط مرة أخرى. تم تسجيل الأطياف باستخدام جهاز الفلورية (Tecan spark 10 M ) ( باستخدام برنامج SoftMax Pro 7.1 GxP. تم استخدام الأسيتونيتريل والميثانول من درجة UPLC بالإضافة إلى ماء الميلي Q.
اختبار التثبيط المعتمد على الفلورية للأكسدة الذاتية (FENIX)
تم شحن أنبوب الطرد المركزي المخروطي (15 مل) بحويصلات دهنية أحادية الطبقة من فوسفاتيديل كولين البيض (0.4 مل من تعليق 20 مليمول في PBS، بمتوسط قطر 100 نانومتر)، 134 مل من محلول 12 مليمول من (E)-1,2-bis((2-methyldecan-2-yl)oxy)diazene (Cayman، 32742) في الإيثانول وPBS (7.46 مل من 12 مليمول عند pH 7.4) وتم خلطه بواسطة جهاز الخلط. STY-Bodipy (تم إضافة محلول بتركيز 1.74 مليمول في DMSO، ثم تم خلط التعليق بواسطة دوامة مرة واحدة. تم نقل خليط الليبوسوم-المبادر-الصبغة إلى خزان، واستخدامتم تحميل لوحة 96 بئر (سوداء، نونك) بالمزيج باستخدام بيبتيد متعدد القنوات لكل بئر). مركبات الاختبار ( من محلول بتركيز 0.24 مليمول في الأسيتونيتريل) تم إضافته لرفع الحجم النهائي إلىتم خلط محتويات الآبار يدويًا باستخدام ماصة متعددة القنوات ثم تم خلطها في قارئ الميكرو بلايت لمدة دقيقة واحدة عند، بعد ذلك تم مراقبة أكسدة STY-Bodipy بواسطة الفلورية (تم جمع البيانات باستخدام برنامج Agilent BioTek Gen5 (الإصدار 3.08.01) بين عشية وضحاها.
الأجسام المضادة
“تم توضيح الأجسام المضادة على النحو التالي: WB، البقعة الغربية؛ FCy، قياس التدفق الخلوي؛ FI، تصوير الفلورية؛ Hu، مستخدمة لعينات بشرية؛ Ms، مستخدمة لعينات فئران. يتم الإشارة إلى التخفيفات. أي تحقق من الأجسام المضادة من قبل الشركة المصنعة موضح ويمكن العثور عليه على مواقع الشركات المصنعة. استراتيجيات تحققنا من الأجسام المضادة من خلال تقنيات خفض التعبير (KD) و/أو حذف الجينات (KO) للأجسام المضادة ذات الصلة موضحة. تم استخدام الأجسام المضادة الأولية التالية: AIFM2/FSP1 (Merck، MABC1638، النسخة 6D8-11، الدفعة Q3745998، WB، 1:500، Hu، تم التحقق من KD داخليًا)؛ الكاتالاز (Cell Signaling، 12980T، النسخة D4P7B، الدفعة 3، FI، 1:200، Hu)؛ CD3-BV510 (BioLegend، 317332، النسخة OKT3، الدفعة B263750، FCy، 1:100، Hu)؛ CD31-PE-Cy7 (BioLegend، 303118، النسخة WM59، الدفعة B276836، FCy، 1:100، Hu)؛ CD31-BV605 (BioLegend، 303122، النسخة WM59، الدفعة 331683، FCy، 1:100، Hu)؛ CD31-BV605 (BioLegend، 102427، النسخة 390، الدفعة B375532، FCy، 1:100، Ms)؛ CD44 (Abcam، ab189524، النسخة EPR18668، الدفعة 1014086-32، WB، 1:30,000، Hu، تم التحقق من KO وKD داخليًا)؛ CD44-AF647 (Novus Biologicals، NB500-481AF488، النسخة MEM-263، الدفعة P158343، FCy، 1:100، Hu)؛ CD44-AF647 (Novus Biologicals، NB500-481AF647، النسخة MEM-263، الدفعة D145771، FCy، 1:100، Hu)؛ CD44-AF647 (BioLegend، 103018 (إشارة الخلية، 12963S، النسخة “، الدفعة 2، FI، 1:200، Hu)؛ EEA1 (Abcam، ab70521، النسخة 1G11، الدفعة GR315680-1، FI، 1:200، Hu، تم التحقق منها بواسطة المناعية الخلوية/ المناعية الفلورية من قبل الشركة المصنعة)؛ FAP-AF700 (R&D Systems، FAB3715N، النسخة 427819، الدفعة AEVI020011، FCy، 1:100، Hu)؛ FAP-AF750 (Bio-Techne، FAB3715S-100UG، النسخة 427819، الدفعة 1718688، FCy، 1:100، Hu)؛ الفيريتين (Abcam، ab75973، النسخة EPR300AY، الدفعة 10136442-29، WB، 1:1,000، Hu، تم التحقق منها بواسطة WB من قبل الشركة المصنعة)؛ الفيبرو نكتين (Abcam، ab45688، النسخة F14، الدفعة 1016266-35، WB، 1:1,000، Hu)؛ FTH1 (Santa Cruz Biotechnology، sc-376594، النسخة B-12، الدفعة G2622، WB، 1:200، Hu)؛ GPX4 (Abcam، ab125066، النسخة EPNCIR144، الدفعة GR3369574-4، WB، 1:2,000، Hu، تم التحقق من KO من قبل الشركة المصنعة، تم التحقق من KD داخلياً)؛ 4-HNE (Abcam، ab48506، النسخة HNEJ-2، الدفعة 1062274-2، FI، 1:200، Hu)؛ IRP2 (Novus Biologicals، NB100-1798، الدفعة D-4، WB، 1:1,000، Hu)؛ لامين A/C (Cell Signaling، 2032S، الدفعة 6، WB، 1:1,000، Hu)؛ LAMP1 (Cell Signaling، 9091S، النسخة D2D11، الدفعة 7، FI، 1:200، Hu)؛ LAMP1 (Santa Cruz Biotechnology، sc-20011، النسخة H4A3، FI، 1:100، Ms)؛ LAMP1 (Abcam، ab24170، الدفعة GR3235630-1، WB، 1:1,000، Hu)؛ LAMP2 (Abcam، ab25631، النسخة H4B4، الدفعة 1011336-1، FI، 1:200، Hu)؛ LAMP2 (Thermo Fisher Scientific، MA1-205، النسخة H4B4، الدفعة YG377512، WB، 1:1,000، Hu)؛ LIMPII (Proteintech، 27102-1-AP، WB، 1:1,000، Hu)؛ NCOA4 (Abcam، ab86707، الدفعة GR3244520-13، WB، 1:10,000، Hu، تم التحقق من KD داخلياً)؛ NDUFS1 (Ab-توبولين (سيغما-ألدريتش، T5326، النسخة GTU-88، الدفعة 0000140390، WB، 1:1,000، Hu، تم التحقق منها من قبل الشركة المصنعة)؛ وفيمنتين (سيل سيجنالينغ، 5741S، النسخة D21H3، الدفعة 8، WB، 1:1,000، Hu). تم استخدام الأجسام المضادة الثانوية التالية: أليكسا فلور 647 مضاد للفأر (أبكام، ab150115، وسم الأنسجة، 1:500، Ms)؛ أليكسا فلور 647 مضاد للفأر (إنفيتروجين، A21237، الدفعة 1485202، FI، 1:1,000،
تم الحصول على خلايا HT-1080، MDA-MB-231 وخلايا 4T1 من ATCC. تم زراعة خلايا الأورام البشرية والفأر المنفصلة وخلايا 4T1 في RPMI1640 معززة بـ GlutaMAX (Gibco، 61870010) وFBS (Eurobio Scientific, CVFSVF00-01). تم زراعة خلايا HT-1080 في وسط دلبوكو المعدل (DMEM)-GlutaMAX (Gibco, 61965059) معززة بـ 10% FBS (Gibco, 10270-106) والبنسلين-ستربتوميسين (BioWhittaker/Lonza, DE17-602E). تم زراعة خلايا MDA-MB-231 في DMEM-GlutaMAX معززة بالبيروفات (Thermo Fisher Scientific, 31966021) ومعززة بـ 10% FBS والبنسلين-ستربتوميسين (Gibco, 15140148). خلايا FC1242 و FC1245 من سرطان البنكرياس الفأري، وخلايا 4a وخلايا البنكرياس البشرية hMIA-2D كانت هدية من مختبر توفيسون (مختبر كولد سبرينغ هاربر) وتم زراعتها في DMEM-GlutaMAX معززة بـ 10% FBS والبنسلين-ستربتوميسين. تم زراعة خلايا PDAC024T و PDAC030T و PDAC053T و PDAC054T و PDAC084T و PDAC090T و PDAC211T البشرية الأولية في وسط قنوي خالٍ من المصل DMEM/F12 (Gibco, 10565018) معززة بـ 0.61 جرام في 500 مل من النيكوتيناميد (Sigma-Aldrich, 3376)، و2.50 جرام في 500 مل من الجلوكوز (Sigma-Aldrich, G6152)، و1:200 ITS+ (Corning, 354352)، و1:20 Nu-serum IV (Corning, 355104).سم الكوليراديكساميثازون (سيغما-ألدريتش، D4902)،-ثلاثي يودو-L-ثيرونين (سيغما-ألدريتش، T6397) وبنسلين-ستربتوميسين). مستخلص الغدة النخامية البقرية الطازج (جيبكو، ) و تم إضافة EGF البشري المعاد تركيبه الخالي من الحيوانات (Thermo Fisher Scientific، AF-100-15-1MG) إلى وسط جديد عند تقسيم أو زراعة الخلايا. تم زراعة خلايا SUM159 في Ham F12، GlutaMAX (Gibco، 31765035)، NEAA (Gibco، 11140050)، مضاد حيوي-مضاد للفطريات (Thermo Fisher Scientific، 15240062)، الأنسولين (Humalog، Cip: 3400934142680)، الهيدروكورتيزون (Sigma-Aldrich، H0888-10G) و5% FBS (Thermo Fisher Scientific، A5256701). تم زراعة خلايا الأورام الدائرية الأولية من الرئة (Celprogen، 36107-34CTC، الدفعة 219411، الجنس: أنثى) وخلايا الأورام الدائرية الأولية من القولون (Celprogen، 36112-39CTC، الدفعة 20188، الجنس: أنثى) باستخدام وسط الخلايا الجذعية الكامل (Celprogen، M36102-29PS) حتى المرور الثالث. تم زراعة خلايا السرطان الدائرية في قوارير T75 من ECM الخلايا الجذعية (Celprogen، E77002-07-T75). تم زراعة خلايا Pfa1 باستخدام DMEM عالي الجلوكوز وFBS، 2 مللي مول من L-glutamine و 1% من البنسلين-ستربتوميسين.
فصل عينات الأورام البشرية والفأرية
تم جمع عينات الأورام من المرضى بعد الجراحة. كانت عينات الأورام من مرضى يعانون من سرطان البنكرياس القنوي، ساركوما الأنسجة الرخوة غير المحددة، ليبوساركوما، أنجيو ساركوما، ساركوما الظهارية أو نقائل سرطان البنكرياس إلى الكبد. تم فصل الأورام باستخدام مجموعة فصل الأورام البشرية (ميلتيني، 130-095929) وفقًا لبروتوكول الشركة المصنعة. باختصار، تم قطع الأورام إلى قطع صغيرة ( )، وضعت في خليط الإنزيمات في RPMI وتم تفكيكها باستخدام جهاز التفكيك gentleMACS Octo مع السخانات (Miltenyi) باستخدام البرنامج المناسب gentleMACS (37C_h_TDK). لكل عينة ورم، تم استخدام 9.4 مل من وسط RPMI مع تركيز الإنزيم المناسب وفقًا لبروتوكول الشركة المصنعة. تم تفكيك عينات الأورام من الفئران باستخدام مجموعة تفكيك أورام الفئران (Miltenyi، 130-096-730) وفقًا لبروتوكول الشركة المصنعة. تم قطع الأورام إلى قطع صغيرة ( )، وضعت في خليط الإنزيمات في RPMI وتم تفكيكها باستخدام جهاز التفكيك gentleMACS Octo مع السخانات (Miltenyi) مع البرنامج المناسب gentleMACS (37C_m_TDK). بعد ذلك، تم تطبيق تعليق الورم المفكك على مصفاة MACS SmartStrainer. (ميلتيني). تم تخفيف العينات بـتمت معالجة العينة بمحلول PBS وتم الطرد المركزي عند 300 جرام. تم إعادة تعليق راسب الخلايا في RPMI. (مع 10% من مصل فتي وبيتا-لاكتام المضاد الحيوي)، وتم عد الخلايا باستخدام عداد خلايا آلي (Entek). يتم الإشارة إلى إجمالي عدد الخلايا المفصولة على أنه خلايا ورمية مفصولة، بينما يتم الإشارة إلى المجموعة الفرعية التي تتوافق مع خلايا السرطان من الأورام من خلال الاختيار السلبي (انظر قسم ‘تدفق الخلايا’) على أنها خلايا سرطان ورمية مفصولة.
إنشاء ثقافات خلايا أولية مشتقة من زراعة الأنسجة الغريبة
تم اشتقاق هذه النماذج في الأصل من نماذج زراعة الأنسجة المستمدة من المرضى (PDX). تم معالجة قطع PDX المخصصة لزراعة الخلايا في غرفة أمان حيوي. بعد تقطيعها إلى قطع صغيرة، تم معالجتها بالإنزيم الكولاجيناز من النوع الخامس (Sigma-Aldrich، C9263) والتربسين EDTA (Gibco، 25200-056) وتم تعليقها في DMEM المدعوم بـالبنسلين-ستربتوميسين وFBS. بعد الطرد المركزي، تم إعادة تعليق الخلايا في وسط قنوي خالٍ من المصل تم تكييفه من بروتوكولات سابقة.فيفيالحاضنة. تم تخزين الخلايا المعززة في النيتروجين السائل. تم فصل الخلايا عن المضادات الحيوية لأكثر من 48 ساعة قبل الاختبار. تم استخدام هذا البروتوكول لإنشاء الخلايا المعينة باسم PDAC024T و PDAC030T و PDAC053T و PDAC054T و PDAC084T و PDAC090T و PDAC211T.
إنشاء عضيات البنكرياس المشتقة من زراعة الأنسجة الغريبة
تم اشتقاق نماذج الأعضاء البنكرياسية المستمدة من زراعة الأنسجة الغريبة (XDPO) في الأصل من نماذج PDX. تم تقسيم زراعات الأنسجة إلى عدة قطع صغيرة ومعالجتها في غرفة أمان حيوي. بعد تقطيعها إلى قطع صغيرة، تم معالجتها باستخدام مجموعة تفكيك الأورام البشرية (Miltenyi، 130-095-929). تم هضم الكريات غير المهضومة باستخدام الأكتوز (Thermo Fisher Scientific، A1110501) عندلمدة 30 دقيقة. تم نقل معلق نسيج البنكرياس إلىمنخل الأنسجة ثم وضع في لوحة 12 بئر مغطاة بـماتريجيل (كورنينج، 354230). تم زراعة العينات في وسط تغذية الأعضاء البنكرياسية، والذي يتكون من DMEM/F12 المتقدم معزز بـ 10 مللي مول من HEPES (ثيرمو فيشر ساينتيفيك، 15630056)،غلوتا ماكس (ثيرمو فيشر ساينتيفيك، 35050087)، بنسلين-ستربتوميسين،FGF10 البشري المعاد تركيبه الخالي من الحيوانات (ثيرمو فيشر ساينتيفيك،EGF البشري المعاد تركيبه الخالي من الحيوانات (ثيرمو فيشر ساينتيفيك، AF-100-15-1MG)نوجين بشري مؤتلف (بايو-تيكن، 6057-NG)، وسط مُشَشَّد بـ WNT3Aوسط مشروط بـ RSPO1 (10 نانومتر من الجاسترين البشري 1 (سيغما-ألدريتش، SCP0152)، 10 ملليمول من النيكوتيناميد (سيغما-ألدريتش، NO636)،-أسيتيل سيستين (سيغما-ألدريتش، A9165)، (ثيرمو فيشر ساينتيفيك، 17504001)، 500 نانومتر A83-01 (توكريس، 2939/10) وY27632 (Tocris, 1254/1). تم حضن الأطباق فيفيالحاضنة، وتم تغيير الوسط كلأيام. تم استخدام هذه الإجراءات لتوليد XDPOs PDAC009T و PDAC003T و PDAC117T و PDAC372T.
توصيف حساسية الكيمياء لـ XDPOs
لتحليل حساسية الأدوية الكيميائية، تم زرع XDPOs في أطباق 96 بئرًا ثم تعرضت لتركيزات متزايدة من الأدوية. تم قياس حيوية الخلايا بعد 72 ساعة من العلاج باستخدام CellTiter-Glo 3D (بروماجا، G9683). تم حساب أوقات تضاعف حيوية XDPO في ظروف العلاج بالمركب على اليومين 0 و3. كانت نسبة اليوم 3 إلى اليوم 0 تعادل معدل التكاثر (RR) للخلايا بعد 72 ساعة. تم حساب أوقات التضاعف باستخدام الصيغةتم قياس قيم الفلورية واللمعان باستخدام جهاز قراءة الألواح Tristar LB941 (تقنيات بيرثولد). تم إجراء كل تجربة على الأقل ثلاث مرات مع ثلاثة مكررات على الأقل.
اختبارات موت الخلايا والحيوية
اختبار موت الخلايا باستخدام Annexin-V ويوديد البروبيديوم. تم زراعة خلايا PDAC053T في أطباق 6 آبار بكثافةخلايا لكل بئر. RSL3 (سيغما-ألدريتش، SML2234، 0.1، 0.5، 2 و ) تم إضافته مع Lip-1 ، cLip-1الكليب-1أو ميتكليب-1في اليوم التالي. بعد 24 ساعة، تم استعادة الوسط. وتمت معالجة الخلايا بالتريبسين. تم جمع الخلايا، وتكويرها مع الوسط المستعاد وغسلها بـPBS. التالي،منتم إضافة محلول ربط الأنكسين- V الذي يحتوي على الأنكسين- V واليوديد البروبيوم وفقًا لبروتوكول الشركة المصنعة (مجموعة تحليل التدفق للأنكسين- V، ثيرمو فيشر ساينتيفيك، V13242).محلول PBS يحتوي على FBS و EDTA تمت إضافة ) لقياس التدفق الخلوي. تم إجراء قياس التدفق الخلوي باستخدام جهاز قياس التدفق AttuneTM NxT وتم تحليل البيانات باستخدام FlowJo.
اختبار موت الخلايا باستخدام Annexin-V و Sytox الأزرق. تم زراعة خلايا PDAC053T في أطباق 6 آبار بكثافةخلايا لكل بئر. في اليوم التالي، تم نقل الخلايا باستخدام siRNA كما هو موضح في قسم ‘تداخل RNA’. تم استبدال الوسط بعد 6 ساعات من النقل. بعد ثلاثة أيام من النقل، تم معالجة الخلايا بالتركيزات المحددة من Fento-1 لمدة 6 ساعات. تم تحليل موت الخلايا باستخدام Annexin-V AF488 (Thermo Fisher Scientific، A13201) و Sytox blue (Thermo Fisher Scientific، S34857). تم إجراء تحليل تدفق الخلايا باستخدام جهاز تحليل تدفق AttuneTM NxT وتم تحليل البيانات باستخدام FlowJo.
اختبار LDH. تم قياس إفراز LDH باستخدام مجموعة كشف السمية الخلوية (سيغما-ألدريتش، 11644793001) وفقًا لبروتوكول الشركة المصنعة في صفيحة 96 بئر. تم تقييم حيوية الخلايا باستخدام مجموعة CellTiterGlo 2.0 (بروماجا، G9241) أو CellTiter blue (بروماجا، G8081) وفقًا لبروتوكول الشركة المصنعة في صفيحة 96 بئر. باختصار، تم زراعة 4000 خلية (HT-1080، PDAC053T أو 4T1) لكل بئر في صفائح 96 بئر ذات قاع شفاف ومظلم (غرينر، 655090، دفعة E23063EG) قبل 24 ساعة من التجربة. ثم تم معالجة الخلايا مسبقًا لمدة ساعتين بـ, كليب-1 فيروستاتين-1 (SML0583،DFO (سيغما-ألدريتش، D9533،ديفرسيروك (كيميان كيميكل،ديفيريبيرون (سيغما-ألدريتش، Y0001976، ) ، -توكوفيرول (فيتامين K3 (سيغما-ألدريتش، M5625-25G،Z-VAD-FMK (إنزو لايف ساينسز، ALX-260-020-M005، ) أو نيكروستاتين-1 (سيغما-ألدريتش، N9037، . بعد ذلك، فنتو-1تمت الإضافة. بالنسبة لقياسات الجرعة والاستجابة، تم معالجة الخلايا بكميات متفاوتة من الجزيء المعني لمدة الوقت المحدد. تم معالجة العينات كما هو موضح في بروتوكولات الشركة المصنعة وتم تسجيل البيانات على جهاز قراءة الألواح SpectraMax ID3 (Molecular Devices). بالنسبة لقياسات بقاء الخلايا وفقًا لمعايير الرعاية، تم زراعة الخلايا بمعدل 2000 خلية لكل بئر قبل 24 ساعة من التجربة. تم حضانة الخلايا مع تخفيفات متسلسلة من Fento-1، إيرينوتيكان (Sigma-Aldrich، I1406)، 5-فلورويوراسيل (5-FU؛ Alfa Aesar، A13456-06) أو أوكساليبلاتين (Bio-Techne، 2623) لمدة 72 ساعة.
اختبار MTT. تم زرع الخلايا في صفيحة 96 بئرًا، وتم حضنها لمدة 24 ساعة ثم تم معالجتها مسبقًا بـ Lip-1. لمدة 10 دقائق قبل العلاج مع التحكم في المركبة أو Fento-1 لمدة 24 ساعة. بعد 24 ساعة، تمت إزالة الوسط بعناية ووسط خالٍ من المصل وتم إضافة محلول MTT (Cayman Chemical، 21795) لكل بئر. تم حضن اللوحة فيلمدة 3 ساعات. بعد الحضانة، تمت إزالة المحلول وتم إضافة DMSO لكل بئر. تم تغطية اللوحة وهزها على هزاز مداري لمدة 15 دقيقة قبل قراءة الامتصاص على جهاز قراءة اللوحات (OD ). لعلاج RSL3، تم زراعة خلايا Pfa1 في أطباق 96 بئر (2000 خلية لكل بئر) وتم زراعتها طوال الليل. في اليوم التالي، تم معالجة الخلايا بـ RSL3 والجزيئات الصغيرة المشار إليها في التخفيف التسلسلي. بالنسبة لعلاج 4-OH-TAM، تم زراعة خلايا Pfa1 في أطباق 96 بئر (500 خلية لكل بئر) مع 4-OH-TAM ( ) وسلسلة تخفيف من المركبات المحددة. بعد 24 ساعة (لـ RSL3) أو 72 ساعة (لـ 4-OH TAM) من الحضانة، تم تقييم حيوية الخلايا باستخدام الريزازورين كمؤشر لحيوية الخلايا. تم قياس شدة الفلورية عند باستخدام جهاز قراءة الميكرو بلايت SpectraMax iD5 مع برنامج SoftMax Pro الإصدار 7 (Molecular Devices) بعد 4 ساعات من الحضانة في وسط زراعة الخلايا القياسي الذي يحتوي علىريزازورين.
اختبار ألامار الأزرق. تم تقييم تأثير الجرعات غير القاتلة من Fento-1 مع RSL3 أو ML210 على بقاء الخلايا باستخدام اختبار ألامار الأزرق. لتحديد بقاء الخلايا، تم استخدام 3,000 من خلايا HT-1080 أو MDA-MB-231. تم زراعة الخلايا في لوحة 96 بئر باستخدام وسط قياسي (10% مصل بقري فتي).البنسلين-ستربتوميسين) حوالي 20 ساعة قبل العلاج. تم معالجة الخلايا مسبقًا بـفينتو- 1 و/أولمدة ساعة واحدة. بعد 48 ساعة من علاج RSL3 أو ML210 (سيغما-ألدريتش، SML0521-5MG)، تم إجراء اختبار حيوية الخلايا. تم تحضير محلول ألامار الأزرق عن طريق إذابة 0.5 جرام من ملح الصوديوم ريسازورين (سيغما-ألدريتش، 263-718-5) في 100 مل من PBS معقم وتم ترشيحه بشكل معقم من خلالفلتر. بعد ساعتين من الحضانة، تم تقييم الحيوية عن طريق قياس الفلورية باستخدامفلتر الإثارة وفلتر الانبعاث على قارئ الميكرو بلايت Spark (تيكان).
تم قياس السمية الخلوية لـ Fento-1 باستخدام جهاز IncuCyte. تم جمع حركيات موت الخلايا باستخدام منصة التصوير الحيوي IncuCyte (Essen). تم زراعة الخلايا في أطباق 96 بئرًا (3,000 خلية لكل بئر) قبل يوم واحد من العلاج. تم علاج الخلايا بـ Fento-1 بالاشتراك مع Lip-1.في وسط FluoroBrite DMEM (Thermo Fisher Scientific A1896701). تم التقاط أربع صور لكل بئر كل ساعة، وتم تحليلها ومتوسط البيانات. تم قياس موت الخلايا بناءً على دمج DRAQ7.تم جمع البيانات كعدد للخلايا الإيجابية لـ DRAQ7 لكل عدد إجمالي من الخلايا في كل حالة.
توليد خلايا سرطان HT-1080 DTP
حولتم زراعة خلايا HT-1080 في قوارير T75 قبل 24 ساعة من إضافة دوكسوروبيسين (Clinisciences، HY-15142، 25 نانومول). تم غسل القوارير بـتم استبدال محلول PBS ووسيط يحتوي على دوكسوروبيسين (25 نانومول) كلأيام. بعد 30 يومًا من العلاج بالدوكسوروبيسين، تم زراعة 4000 خلية في أطباق 96 بئرًا لكل بئر لتقييم حيوية الخلايا بعد العلاج مع Fento-1.
تحليل بقاء الخلايا المستنسخة
حولتم زراعة خلايا SUM159 في أطباق سعة 10 سم في وسط نمو كامل. تم معالجة الخلايا المزروعة بالدoxorubicin عند التركيز المثبط نصف الأقصى. ) التركيز ( 150 نانومولار ). بعد 72 ساعة ، تم غسل الخلايا ومعالجتها بـ Fento- ) أو دي إم إس أو بدون دواء. بعد 72 ساعة، تم غسل الخلايا، واستُبدل العلاج بوسط نمو كامل. بعد 10-15 يومًا، تم غسل الخلايا، وتثبيتها، وصبغها بحمض الأسيتيك والميثانول (1:7) وكوماسي الأزرق 1 ساعة في درجة حرارة الغرفة. عدد المستعمرات معتم عد الخلايا باستخدام عداد المستعمرات SCAN1200 (إنترساينس).
تحليل تآزر الأدوية
تم زراعة الخلايا في أطباق 96 بئر (5000 خلية لكل بئر). بعد 12 ساعة، تم معالجة الخلايا بـ 5-FU (Selleckchem، S1209،جمسيتابين (سيلك كيم، S1714،أوكساليبلاتين (سيلك كيم، S1224،بكليتاكسيل (Selleckchem، S1150، ) أو SN-38 (Selleckchem، S4908، بمفرده أو بالاشتراك مع Fento-1 لمدة 72 ساعة. تم تقدير حيوية الخلايا بعد إضافة PrestoBlue (Life Technologies، A13261) لمدة ساعتين وفقًا لتعليمات الشركة المصنعة. لتحديد وجود تأثير تآزري على تكاثر الخلايا أو حيوية الخلايا بين أدوية الرعاية القياسية وFento-1، استخدمنا برنامج SynergyFinder. لتحليل هذا التأثير، تم استخدام درجة ZIP كنموذج لحساب التآزر..
علاج cLip-1 في الجسم الحي
تم الاحتفاظ بالفئران تحت ظروف قياسية مع الماء والطعام بحرية وفي بيئة خاضعة للرقابة.الرطوبة، دورة ضوء-ظلام لمدة 12 ساعة). بالنسبة لدراسات الحيوانات، تم توزيع فئران C57BL6/J عشوائيًا في أقفاص منفصلة. تم استخدام الفئران التي تتراوح أعمارها بين 12-24 أسبوعًا والمتطابقة في الجنس لجميع التجارب. لدراسة مجموعة البقاء، Rosa26-creERT2;Gpx4تم علاج الفئران عن طريق الحقن داخل البطن بالتاموكسيفين (سيغما-ألدريتش، T2859، 2 ملغ في اليوم مذاب في Myglyol 812) في اليومين 0 و 1 من أجل الحذف لـفي الجسم كله باستثناء الدماغ. من اليوم الثاني، cLip-1 (محلي الصنع، 10 ملغ لكل كغ يوميًا مذاب فيبي بي إس تم إعطاء 20% PEG400 و5% Solutol HS15) أو المركب كحقن داخل الصفاق للفئران يوميًا حتى انتهاء دراسة البقاء. بالنسبة لتحليلات الكيمياء النسيجية، Rosa26-creERT2;Gpx4تم استخدام الفئران المعالجة بحقن التاموكسيفين داخل الصفاق (2 ملغ يوميًا في اليومين 0 و1). في اليوم السابع، cLip-1 (محلي الصنع، 10 وتم حقن المركب (أو المركبة) داخل الصفاق في الفئران. بعد ساعة من الحقن، تم euthanized الفئران، وتم معالجة عينات الكلى والكبد كما هو موضح في قسم ‘توسيم الجزيئات الصغيرة باستخدام كيمياء النقر’.
عزل الدم والمصل والسائل اللمفاوي من الفئران
تم شراء فئران Balb/c (إناث بعمر 25 أسبوعًا) من شركة تشارلز ريفر وتم إيواؤها في منشأة الحيوانات الأساسية في CRCM وتم توزيعها عشوائيًا في أقفاص منفصلة. تم استخدام الفئران التي تتراوح أعمارها بين 12-24 أسبوعًا والمتطابقة في الجنس لجميع التجارب. تم إيواء الفئران في ظروف معقمة مع توفير الطعام والماء المعقمين بحرية، وتم الحفاظ عليها على دورة ضوء-ظلام مدتها 12 ساعة.الرطوبة. لم تتعرض الفئران لأي إجراءات قبل جمع اللمف والدم والمصل.
لجمع عينة اللمف، تم إعطاء البوبرينورفين (Buprecare)، وهو مسكن، عن طريق الحقن داخل الصفاق قبل 30 دقيقة من بدء التجربة.تم euthanizing الفئران عن طريق الحقن داخل البطن بمزيج من الكيتامين والزلازين (الكيتامين“إيمالجين” وزيلزين (رومبوم)؛ بعد إجراء الشقوق الجلدية والبطنية، فإن اللمف في جذع اللمف المعويتم جمعها باستخدام شعيرة زجاجية. تم وضع عينات اللمف في أنابيب تجميد، وتم تجميدها عندوتم تخزينه في.
لجمع عينات الدم والمصل، بعد جمع اللمف، قمنا بإجراء ثقب قلبي نهائي (إبرة 23 G مع إبرة) عن طريق فتح الصدر لجمع حجم كبير من الدم دون مضادات التخثر. بعد ذلك،عينة الدم وضعت في قنينة، وتم تجميدها عندوتم تخزينه فيتم ترك بقية عينة الدم لتقف في درجة حرارة الغرفة لمدة 30 دقيقة، ثم تم الطرد المركزي عندلمدة 15 دقيقة. تم جمع الطور العلوي (المصل) في قنينة، وتم تجميده عندوتم تخزينه في.
البروتيوميات الكمية
تحضير العينة. تم زراعة خلايا HT-1080 فيأطباق دائرية فيالتقاء ومعالجته بـ Fento-تم جمع مستخلصات الخلايا الكاملة ثم تم الطرد المركزي عند 500 جرام لمدة 5 دقائق فيمغسولمع الثلج الباردتمت معالجة PBS وتحليله باستخدام محلول التحلل (8 م يوريا، 200 مللي مولار بيكربونات الصوديوم وخلطة مثبطات البروتياز الكاملة (روش، 000000011697498001)). تم تحقيق التحلل عن طريق الحضانة على عجلة دوارة في. لمدة 45 دقيقة في تمت عملية الطرد المركزي للمستخلصات عندلمدة 20 دقيقة عند، وتم استخدام السائل العلوي الذي يحتوي على البروتين الكلي للتحليل البروتيني الكمي العالمي. باختصار، تم تقليل إجمالي بروتين lysate الخلايا عن طريق الحضانة مع 5 مليمول من ديثيوثريتال عندلمدة 30 دقيقة ثم تم ألكلة باستخدام يودوأستاميد ) في درجة حرارة الغرفة في الظلام. تم إضافة التربسين/ليس سي (بروماجا) بنسبة 1:100 (وزن/وزن) من الإنزيم إلى الركيزة. تم إجراء الهضم طوال الليل في ثم تم تحميل العينات على StageTips C18 المصنوعة خصيصًا (AttractSPE Disk Bio C18-100.47.20، Affinisep) لإزالة الأملاح. تم استرجاع الببتيدات باستخدام نسبة 40:60 من الأسيتونيتريل وتم تركيز حمض الفورميك تحت الفراغ حتى الجفاف باستخدام جهاز SpeedVac. تم إعادة تكوين الببتيدات في 0.3% TFA قبل استخدام الكروماتوغرافيا السائلة مع مطياف الكتلة الثنائي (LC-MS/MS).
تحليلات LC-MS/MS. تم إجراء LC باستخدام نظام Vanquish Neo LC (ثيرمو ساينتيفيك) متصل بمطياف الكتلة Orbitrap Astral، مع واجهة بواسطة مصدر أيون Nanospray Flex (ثيرمو ساينتيفيك). تم حقن الببتيدات على عمود C18 (قطر داخلي مدوبل نانو فايبر بيب ماب نيو، ثيرمو فيشر العلمية) التي تخضع للتنظيم فيومفصولة بتدرج خطي منالمخزن A ( و “حمض الفورميك” إلىمخزنأسيتونيتريل وحمض الفورميك) بمعدل تدفق قدرهأكثر من 104 دقيقة. تم تشغيل الجهاز في وضع الاكتساب غير المعتمد على البيانات (DIA). تحليل الطيف الكتلي الكامل تم تسجيل المسح على محلل الكتلة Orbitrap في وضع المركز لمدىبدقة 240,000، مع ضبط هدف التحكم التلقائي في الكسب (AGC) علىووقت حقن أقصى قدره 5 مللي ثانية. تم إجراء DIAs في محلل أسترا في وضع المركز لمدى الكتلة منبعرض نافذة يبلغ 2 دا (بدون تداخل)، ووقت حقن أقصى قدره 3 مللي ثانية وهدف ضبط تلقائي عادي يبلغ 500% بعد التفتت باستخدام التفكك بالاصطدام عالي الطاقة (25% طاقة اصطدام عادية).
معالجة البيانات. لأغراض التعريف، تم البحث عن البيانات مقابل قاعدة بيانات يوني بروت (UP000005640) الخاصة بالبشر (Homo sapiens) باستخدام برنامج سبكترو نوت (v.18.7 أو v.19؛ بيوجنوسيس) من خلال تحليل directDIA + باستخدام إعدادات البحث الافتراضية. تم تعيين خصوصية الإنزيم إلى التربسين، وتم السماح بحد أقصى من انقضاضين مفقودين. تم تعيين الكارباميدوميثيل كتعديل ثابت، وتم تعيين الأسيتيل على الطرف الأميني والأكسدة للميثيونين كتعديلات متغيرة. تم معالجة الملفات الناتجة بشكل إضافي باستخدام myProMS (v.3.10؛https://github.com/bioinfo-pf-curie/مبروكلتحديد كمية البروتين، تم السماح باستخدام XICs من الببتيدات البروتيوتيبية المشتركة بين الظروف المقارنة (مطابقة TopN) مع الانقسامات المفقودة والكارباميدوميثيليشن. تم تطبيق التطبيع الوسيط والنطاق على مستوى الببتيد على الإشارة الكلية لتصحيح XICs لكل تكرار بيولوجي مستقل. ). لتقييم أهمية التغير في وفرة البروتين، تم إجراء نموذج خطي (معدل على الببتيدات والتكرارات البيولوجية)، وتم استخدام اختبار ذو جانبين تم تطبيق اختبار على التغير النسبي المقدر بواسطة النموذج.تم تعديل القيم بعد ذلك باستخدام إجراء معدل الاكتشاف الخاطئ لبنجاميني-هوشبرغ. تم إيداع بيانات البروتيوميات الخام الخاصة بـ MS في اتحاد بروتيوم إكس من خلال مستودع شريك PRIDE..
نماذج انتشار الورم في العقد اللمفاوية في الفئران والإدارة الحية لـ Fento-1
تم زرع خلايا سرطان الثدي من الفئران (4T1) في فئران أنثوية من نوع Balb/c تتراوح أعمارها بين 6-8 أسابيع (متجانسة مع نموذج 4T1). تم الاحتفاظ بالفئران في منشأة الحيوانات الأساسية بمدرسة هارفارد الطبية وتم توزيعها عشوائيًا في أقفاص منفصلة. تم الاحتفاظ بالفئران في ظروف معقمة مع توفير الطعام والماء المعقمين بحرية، وتم الحفاظ عليها على دورة ضوء-ظلام مدتها 12 ساعة.الرطوبة. لأداء الحقن في العقد اللمفاوية، تم تتبع الأوعية اللمفاوية أولاً عن طريق حقن صبغة إيفانز الزرقاء 2% (سيغما-ألدريتش، E2129) في دواسة القدم قبل 5 دقائق من إجراء الحقن داخل العقد. بعد حقن صبغة إيفانز الزرقاء، تم تخدير الفئران باستخدام الإيزوفلوران وقطعة صغيرة (تم إجراء شق في منطقة العقدة اللمفاوية البقعية اليمنى. تم تحديد موقع العقدة اللمفاوية بناءً على صبغة إيفانز الزرقاء، وتم تثبيتها بالملاقط، وخلايا معلقة فيتم حقن PBS بحجمإلى العقدة اللمفاوية الفخذية باستخدام حقنة هاملتون 27 ج. تم تأكيد الحقن في العقدة اللمفاوية من خلال تورم مرئي للعقدة اللمفاوية. تم إغلاق الشق باستخدام لاصق جراحي (لاصق الأنسجة 3 م فيت بوند، 1469 SB) وتم مراقبة الفئران عن كثب بحثًا عن علامات الألم أو الضيق. بمجرد أن أصبحت الأورام ملحوظة في على الأقلالفئران (حوالي أسبوع بعد الحقن)،فينتو-1تم تسليم (لكل حيوان) أو مركبة داخل العقدة اللمفاوية الحاملة للورم كل يومين حتى نقطة النهاية التجريبية. تم قياس أقطار الأورام داخل العقدة 3 مرات في الأسبوع باستخدام الكالبر حتى وصل أي ورم في مجموعة الفئران إلى 2.0 سم في أكبر قطر له، وهو ما كان نقطة النهاية التجريبية المحددة مسبقًا لهذه التجارب. في جميع التجارب، لم يتم تجاوز هذا الحد الأقصى المسموح به لقطر الورم. في تلك المرحلة، تم euthanized جميع الفئران في المجموعة، وفقًا للبروتوكولات المعتمدة، لتحليل قطر الورم داخل العقدة، وكتلة الورم ومعلمات أخرى. عند نقطة النهاية التجريبية، تم جمع بيانات حول تكوين الورم بطريقة غير معروفة لهوية العينة أو العلاج. لتحديد منحنيات البقاء، تم تطبيق طرق معدل البقاء المعتمدة على حجم الورم، مع 1.5 سم كقيمة مقطوعة مقبولة أخلاقيًا لتحديد الفعالية العلاجية في دراسات الفئرانتم قياس أحجام الأورام بعددين، الطول ( المحور) والعرض ( المحور)، مع المحور يُعرف تقليديًا بأنه أطول محور للورم. يتم تمثيل أحجام الأورام كقطر الورم (سم) أو حجم الورم ( )، كما تم استنتاجه بواسطة الطول × العرض ، كما هو موضح لكل تجربة حية. تم تجميد عينات الورم فيدي إم إس أو في FBSحتى ) لتحليلات خلوية لاحقة. لم تُستخدم تقنيات عشوائية رسمية. ومع ذلك، تم تخصيص الحيوانات عشوائيًا لمجموعات العلاج وتم معالجة العينات بترتيب عشوائي. في جميع التجارب، تم إبقاء الفئران على نظام غذائي عادي وتم إطعامها بحرية.
تصوير الفلورسنت
تم زراعة الخلايا على شرائح زجاجية وتم معالجتها كما هو موضح. بوديبي 665/676 (ثيرمو فيشر ساينتيفيك، B3932،LysoTracker Deep Red (ثيرمو فيشر ساينتيفيك، L12492، ), DND-189 ليزو سينسور (ثيرمو فيشر ساينتيفيك، L7535، ليبرفلو (دوجيندو، ) و SQSS (داخلي، ) أُضيفت إلى الخلايا الحية قبل التثبيت. لدراسات التداخل، تم معالجة الخلايا بـ Fento- ) أو مارمايسين (داخلي، ) عند درجة الحرارة المحددة بعد 16 ساعة من النقل. بعد 16 ساعة من النقل، تم معالجة الخلايا التي تم نقلها بواسطة BacMam بـ RSL3لمدة 15 دقيقة أو لمدة 3 ساعات و15 دقيقة ثم تم معالجتها بـ Bodipy 665/676 لمدة 45 دقيقة لتحديد النقاط الزمنية النهائية وهي 1 ساعة و4 ساعات. من أجل التداخل المكاني لدهون البيروكسيد مع الليزوزومات، تم معالجة الخلايا بـ RSL3.وإما بوديبي 665/676 وDND-189 لايسوسينسور أو ليبرفلور وليسوتراكر ديب ريد لمدة ساعة واحدة. لتقييم شدة الفلورسنت لفينتو-1 في وجودتمت معالجة الخلايا مسبقًا بـ -توكوفيرول-توكوفيرول (سيغما-ألدريتش، PHR1031، ) ثم مع Fento-1 ( ). ثم تم غسل الخلايا 3 مرات بـ PBS، ثابت معبارافورمالدهيد فيPBS لمدة 12 دقيقة ثم تم غسلها 3 مرات بـPBS. لتلوين الأجسام المضادة، تم اختراق الخلايا بـترايتون X-100 فيPBS لمدة 5 دقائق وغسل 3 مرات بـPBS. بعد ذلك، تم حجب الخلايا في 2% BSA (يوروميدكس، 04-100-812-E)، 0.2% توين-20 وPBS (محلول ملحي فوسفاتي) لمدة 20 دقيقة في درجة حرارة الغرفة. تم حضن الخلايا مع الأجسام المضادة المناسبة في محلول الحجب لمدة ساعة واحدة في درجة حرارة الغرفة، وغسلت 3 مرات معتمت معالجة PBS وتم حضنه مع الأجسام المضادة الثانوية لمدة ساعة واحدة. أخيرًا، تم غسل الشرائح ثلاث مرات بـتم تثبيت الخلايا المعالجة بـ Bodipy 665/676 وLiperfluo باستخدام مواد كيميائية باردة ووضعها فيعلى الفور بعد التركيب على شرائح الزجاج وتم تصويرها. تم الحصول على صور الفلورسنت باستخدام ميكروسكوب ديلتا فيجن في الوقت الحقيقي (أبلايد بريسيجن)، أو ميكروسكوب ثاندر (لايكا) أو ميكروسكوب أبوتوم وايس مع برنامج تصوير ديلتا فيجن، أو برنامج اكتساب الصور ثاندر لايكا، أو برنامج تصوير أبوتوم زيس، على التوالي. أو تم استخدام الهدف NA للاستحواذات، وتم الحصول على جميع الصور كـتم فك تشويش الصور باستخدام SoftWorx (نسبة محافظة، 15 تكرار، Applied Precision) ومعالجتها باستخدام Fijiتم التقاط الصور بالأبيض والأسود وتم تطبيق التلوين باستخدام فيجي. يتم عرض شدة الفلورية بوحدات عشوائية ولا يمكن مقارنتها بين الألواح المختلفة. تم حساب تقدير التداخل باستخدام فيجي 2.0.0-rc-69/1.52n. تم الكشف عن النوى باستخدام فلوروسينس DAPI أو Hoechst كما هو موضح.
المجهر الضوئي المضيء والصور الرقمية
تم الحصول على صور المجال الساطع باستخدام ميكروسكوب CKX41 (أوليمبوس) وبرنامج تصوير cellSens Entry (أوليمبوس).
توسيم الجزيئات الصغيرة باستخدام كيمياء النقر
تسمية النقر داخل الخلايا. تم معالجة الخلايا على الشرائح الزجاجية كما هو موضح باستخدام cDFO (داخلي، ), cLip-1 (داخلي، أو 2 ساعة)، ميتكليب-1 (داخلي، ), alcLip-1 (داخلي، ) أو cCW ثم تم تثبيتها وتمريرها كما هو موضح في قسم ‘تصوير الفلورسنت’. تم إضافة Lyso Tracker Deep Red إلى الخلايا الحية لـ
45 دقيقة قبل التثبيت. تم تحضير خليط تفاعل النقر باستخدام مجموعة تصوير Click-iT EdU (Invitrogen, C10337) وفقًا لبروتوكول الشركة المصنعة. في تجربة نموذجية، قمنا بخلطمنعازل تفاعل Click-iT معحلألكسا-فلور-أزيدمضافات عازلة للتفاعل (أسكوربات الصوديوم) وماء نقي للغاية للوصول إلى حجم نهائي منتم تحضين الشرائح الزجاجية مع خليط تفاعل النقر في الظلام عند درجة حرارة الغرفة لمدة 30 دقيقة، ثم تم غسلها 3 مرات بـتم إجراء تصوير المناعة الفلورية كما هو موضح في قسم ‘تصوير الفلورسنت’.
انقر على وضع العلامات في الأنسجة. تم تثبيت عينات أنسجة الكلى والكبد المجمعة من الفئران المعالجة بـ cLip-1 فيبارافورمالدهيد فيPBS طوال الليل فيتم تحضين الأنسجة المثبتة في السكروز في PBS لمدة 30 دقيقة ثم تم حضنه في السكروز في PBS فيلمدة 4 ساعات، تليها التضمين في مركب تثبيت OCT (TissueTek، ساكورا) على ثلج جاف وتخزينه فيتم قطع الأنسجة المجمدة إلى-شرائح سميكة باستخدام جهاز Cryostat Microm HM 560 (ثيرمو فيشر ساينتيفيك) عندتم تثبيت مقاطع الأنسجة بعد ذلك باستخدام 1% من البارافورمالدهيد فيPBS لمدة 10 دقائق ثم تم حضنه معأسيتون لمدة 10 دقائق عندتم تحضين الأقسام في محلول الحجب (PBS يحتوي على BSA و تريتونيك X-100) لمدة 30 دقيقة. تم وضع علامات على العينات باستخدام تفاعل النقر كما هو موضح أعلاه. للتلوين المشترك مع علامة ليفية، تم حضانة العينات المعلّمة بتفاعل النقر مع مضاد LAMP1 أو الأجسام المضادة المخففة فيPBS يحتوي على مصل الماعز العادي طوال الليل في . في اليوم التالي، تم حضانة الأقسام مع الأجسام المضادة الثانوية فيPBS يحتوي على 1% BSA وتم استخدام Triton X-100 لمدة ساعتين في درجة حرارة الغرفة. تم تصور النوى باستخدام صبغة Hoechst 33342، وتم تثبيت الشرائح في Aqua/ PolyMount (Polysciences). تم التقاط الصور باستخدام ميكروسكوب فلوري (DS-Qi2، نيكون) أو ميكروسكوب ضوئي متماسك (LSM880، كارل زيس) ومجموعات الفلاتر المناسبة المقابلة للفلوروفورات.
التحليل الغربي
تم معالجة الخلايا كما هو موضح ثم تم غسلها بـPBS. تم إذابة البروتينات فيمحلول لايملي المحتوي على بنزونايز (VWR، 70664-3، 1:100). تم حضن المستخلصات فيلمدة ساعة واحدة، تم تسخينه عندلمدة 10 دقائق وتم قياسها باستخدام مقياس الطيف الضوئي NanoDrop 2000 (ثيرمو فيشر ساينتيفيك). تم فصل مستخلصات البروتين بواسطة كهربائية الهلام SDS-PAGE (نظام إنفيتروجين شور لوك وNu-PAGE)هلامات بيس-تريس مسبقة الصنع). في تجربة نموذجية،تم تحميل إجمالي مستخلص البروتين لكل حارة فيمحلول لايملي يحتوي على بروموفينول أزرق. تم تشغيل علامة حجم على كل هلام:PageRuler (ثيرمو فيشر ساينتيفيك، 26616) أوPageRuler Plus (ثيرمو فيشر ساينتيفيك، 26620) ومحلول لايملي. ثم تم نقل البروتينات إلى أغشية نيتروسليلوز (أميرشام بروترانباستخدام خلية نقل كهربائية شبه جافة Trans-Blot SD (بايو راد) باستخدامحافظة نقل NuPage (Invitrogen, NP00061) معالميثانول. تم حجب الأغشية باستخدام 5% من مسحوق الحليب الخالي من الدسم (ريجيلاي) فيتوين-20تمت معالجة الأنسجة بمحلول PBS لمدة 20 دقيقة. تم قطع الأغشية بحجم العلامة المناسب لتمكين اختبار عدة أجسام مضادة على نفس الغشاء. ثم تم اختبار البقع بالأجسام المضادة الأولية ذات الصلة فيبساتويين-20PBS أو في 5% من مسحوق الحليب الخالي من الدسم في 0.1% توين-20 وPBS فيبين عشية وضحاها مع حركة لطيفة في كيس بلاستيكي شفاف مختوم باليد. تم غسل الأغشية بـتوين-20PBS ثلاث مرات وتم حضنه مع أجسام مضادة ثانوية مرتبطة بالبيروكسيداز من الفجل (مختبرات جاكسون) في 5% من مسحوق الحليب الخالي من الدسم (ريجيلاي)تويين-20PBS لمدة ساعة واحدة في درجة حرارة الغرفة وغسل 3 مرات معتوين-20تم الكشف عن المستضدات باستخدام مجموعات الكشف الكيميائي الضوئي SuperSignal West Pico PLUS (Thermo Fisher Scientific، 34580) وSuperSignal West Femto (Thermo Fisher Scientific، 34096). تم تسجيل الإشارات باستخدام نظام تصوير Fusion Solo S (Vilber) مع برنامج FusionCapt Advance. تم معالجة الصور باستخدام برنامج Fiji 2.0.0-rc-69/1.52n. تم توفير مسحات كاملة من البقع في المعلومات التكميلية.
تحويل
تم زراعة خلايا PDAC053T في أطباق ذات 4 آبار بكثافةخلايا لكل بئر 24 ساعة قبل التجربة في 1 مل من وسط زراعة الخلايا. ثم تم نقل الخلايا باستخدام CellLight Lysosomes-GFP، BacMam 2.0 (ثيرمو فيشر ساينتيفيك، C10596)، CellLight ER-GFP، BacMam 2.0 (ثيرمو فيشر ساينتيفيك، C10590) أو CellLight Mitochondria-GFP، BacMam 2.0 (ثيرمو فيشر ساينتيفيك، C10600) وفقًا لتعليمات الشركة المصنعة. باختصار،تم إضافة الكاشف إلى الوسط وخلطه. تم حضانة الخلايا لمدة 16 ساعة.
تداخل RNA
تم زراعة الخلايا قبل 24 ساعة من التجارب. تم نقل الخلايا باستخدام siRNA المحدد باستخدام jetPRIME (Polyplus، 114-15) وفقًا لبروتوكول الشركة المصنعة بتركيز 100 نانومتر من siRNA. باختصار، تم زرع خلايا PDAC053T في أطباق 6 آبار بكثافةتم زراعة الخلايا في كل بئر وتم نقلها بعد 24 ساعة. تم استبدال الوسط بعد 6 ساعات. تم إجراء التحليل بعد 72 ساعة من النقل. تم تصميم siRNAs المناسبة بواسطة دارماكون لتقليل التعبير المستهدف للجينات. تم توفير تسلسلات siRNA في المعلومات التكميلية.
تدفق الخلايا
تم غسل الخلايا بماء مثلجPBS. لتلوين الأجسام المضادة، تم حضانة الخلايا مع حجب Fc (Human TruStain FcX، BioLegend، 422302، ) لمدة 15 دقيقة، ثم تم حضنه مع الأجسام المضادة في درجة حرارة منخفضةFBSPBS، 2 مللي مول EDTA لمدة 20 دقيقة عندثم تم غسلها بـPBS وإعادة تعليقها فيFBS،PBS و 2 مللي مولار EDTA قبل التحليل باستخدام جهاز تحليل الخلايا BD LSR Fortessa X-20 مع برنامج FACS DIVA (الإصدار 9.0.1).
تدفق الخلايا باستخدام مجسات الحديد. تم زراعة خلايا PDAC053T و HT-1080 في أطباق 6 آبار بكثافةخلايا لكل بئر. في اليوم التالي، ليب-1الهيدروكسي كلوروكين (سيغما-ألدريتش، ) أو بافيلوميسين A1 (سيغما-ألدريتش، B1793، 75 نانومتر ) أضيف إلى خلايا PDAC053T لمدة ساعة وإلى خلايا HT-1080 لمدة 30 دقيقة. بالنسبة لخلايا PDAC053T، بعد 30 دقيقة من العلاج بالمركبات، RPE داخلي، ) أو HMRhoNox- داخلي،تمت إضافة المجسات لمدة 30 دقيقة. بالنسبة لخلايا HT-1080، بعد 15 دقيقة من المعالجة بالمركبات، RPEداخلي، ) أو HMRhoNox- داخلي،تم إضافة المجسات وتركها لمدة 15 دقيقة. بعد الحضانة مع مجسات الحديد، تمت إزالة الوسط وتم غسل الخلايا بـتم جمع الخلايا، وتكويرها، وغسلها بـ PBS قبل التربسين.PBS وأخيرًامنمحلول PBS يحتوي على FBS و EDTA تمت الإضافة. تم تسجيل البيانات على جهاز BD LSR Fortessa X-20.
تدفق الخلايا باستخدام بوديبي-C11 581/591. تم زراعة خلايا PDAC053T في أطباق 6 آبار بكثافةخلايا لكل بئر. في اليوم التالي، تم معالجة الخلايا بـ Lip-1، cLip-1 (داخلي، ، ميتكليب-1 (داخلي، ), alcLip-1 (داخلي، ) و BodipyC11 581/591 ( 200 نانومتر ) قبل إضافة RSL3 ( 100 نانومتر ). بعد ساعة واحدة ، تمت إزالة الوسط وتم غسل الخلايا بـ تم جمع الخلايا مرتين قبل معالجة التربسين. تم جمع الخلايا، وتكويرها، وغسلها بـPBS وأخيرًامنمحلول PBS يحتوي علىFBS و EDTAتمت الإضافة لتحليل تدفق الخلايا. تم زراعة خلايا PDAC053T و HT-1080 و 4T1 في أطباق 6 آبار بكثافة خلايا لكل بئر. في اليوم التالي، تم معالجة الخلايا بالبافيلوميسين A1 (75 نانومتر) والهيدروكسي كلوروكين ( ) لمدة ساعتين قبل إضافة RSL3 (200 نانومتر لـ PDAC053T و HT-1080، 500 نانومتر لـ 4T1). بعد ساعة، تم معالجة الخلايا بـ Bodipy-C11لمدة ساعة إضافية. تم إزالة الوسط وتم غسل الخلايا بـ تم جمع الخلايا مرتين قبل معالجة التربسين. تم جمع الخلايا، وتكويرها، وغسلها بـPBS وأخيرًامنمحلول PBS يحتوي علىFBS و EDTAتمت الإضافة لقياس التدفق الخلوي. تم تسجيل البيانات على جهاز قياس التدفق الخلوي AttuneTM NxT (ثيرمو فيشر ساينتيفيك) باستخدام Attune NxT (الإصدار 4.2.0).
تحليلات الجلوتاثيون الليزوزومي والجذور الحرة الهيدروكسيلية الليزوزومية. تم حضن الخلايا مع SQSS (داخلي، ) أو 1-أحمر (داخلي، تم تحضين خلايا HT-1080 مع RSL3ML210 ، سيغما-ألدريتش، SML0521)، FIN56 سيغما-ألدريتش، SML1740)، بيوثيونين سلفوكسيمين (سيغما-ألدريتش، B2515) أو إراستين (، سيغما-ألدريتش، 329600) للنقاط الزمنية المحددة. تم تسجيل البيانات على جهاز تحليل التدفق AttuneTM NxT (ثيرمو فيشر ساينتيفيك).
تحليلات محتوى الحديد في الليزوزومات. عادةً،تم حضن خلايا الأنسجة البشرية المفصولة في وسط (RPMI 1610، 10% FBS وبيتا-لاكتام) مع HMRhoNox-داخلي،تم تحليل محتوى الحديد الليزوزومي في عينات الأورام البشرية والأنسجة السليمة المجاورة باستخدام لوحة الأجسام المضادة والصبغات التالية: DAPI (CD3-BV510 (BioLegend, 317332)، CD31-PE-Cy7 (BioLegend, 303118)، CD44-AF647 (Novus Biologicals, NB500481AF647)، CD45-BV785 (BioLegend, 304048)، CD163-PerCP/Cyanine5.5 (BioLegend, 326512)، FAP-AF700 (R&D Systems, FAB3715N) و TFR1-APC-AF750 (Beckman Coulter, A89313). كانت خلايا السرطان الحية تتوافق مع DAPIتم تسجيل البيانات على جهاز تحليل الخلايا BD LSRFortessa X-20 أو AttuneTM NxT (ثيرمو فيشر ساينتيفيك).
تحليلات CD44 في عينات الأورام المعالجة بـ Fento-1. عادةً،تم تحضين خلايا الورم المنفصلة في وسط (RPMI1610، FBS والبنسلين-ستربتوميسين) مع Fento-1 (داخلي، ، 24 ساعة). تم معالجة الخلايا مسبقًا بـ Lip-1، cLip-1 (داخلي، ) ، -توكوفيرولأو ديفيريبيرونلمدة ساعتين.تم الاحتفاظ بالتوكوفيرول نقيًا تحت جو خامل، وتم إعداد محلول مخزون جديد طوال الدراسة قبل كل تجربة. تم استخدام لوحة الأجسام المضادة والصبغة التالية لتحليل تدفق الخلايا اللاحق: سايتوكس الأزرق (ثيرمو فيشر ساينتيفيك، S34857،CD31-BV605 (BioLegend, 303122)، CD45-BV510 (BioLegend, 368526، الدفعة B373428)، CD44-AF647 (Novus Biologicals، NB500-481AF647)، TFR1-PE (BioLegend، 334106) و FAP-AF750 (Novus Biologicals، FAB3715S-100UG). كانت خلايا الورم الحية تتوافق مع Sytox blue.الخلايا. تم تسجيل البيانات على جهاز تحليل تدفق الخلايا AttuneTM NxT (ثيرمو فيشر ساينتيفيك). لتحليلات تدفق الخلايا لمستويات CD44 في أورام الفئران 4T1، عادةً،تم استخدام خلايا الورم المفصولة لكل حالة. تم صبغ الخلايا المفصولة حديثًا باستخدام الأجسام المضادة ولوحة الصبغة التالية: Sytox blue (ثيرمو فيشر ساينتيفيك، S34857،، CD31-BV605 (BioLegend، 102427)، CD44-AF647 (BioLegend، 103018)، CD45-BV510 (BioLegend، 103138) و MHCII-APC/Cyanine7 (BioLegend، 107628). كانت خلايا السرطان الحية تتوافق مع Sytox blueتم تسجيل البيانات على جهاز تحليل الخلايا AttuneTM NxT (ثيرمو فيشر ساينتيفيك). تم تحليل جميع البيانات باستخدام FlowJo (الإصدار 10.10.0).
فكس
تم فرز خلايا بشرية باستخدام الأجسام المضادة التالية: CD31-PE-Cy7 (BioLegend، 303118)، CD44-AF647 (Novus Biologicals، NB500-481AF647) و CD45-BV785 (BioLegend، 304048). كانت الخلايا المفروزة تت corresponded إلىالخلايا وCD44الخلايا، التي تم عزلها على جهاز FACSAria Fusion (BD) باستخدام FACS DIVA (الإصدار 9.0.1). تم إجراء تجارب ICP-MS باستخدام CD44 و CD44 خلايا الورم كما هو موصوف في قسم ‘ICP-MS’. تم فرز خلايا الفئران باستخدام الأجسام المضادة التالية: CD44-AF647 (BioLegend، 103018) و MHCII-APC/Cyanine7 (BioLegend، 107628). كانت الخلايا المفروزة تت corresponded إلىالخلايا والخلايا. تم طرد الخلايا المفروزة في جهاز الطرد المركزي عندوتم معالجة كريات الخلايا للتطبيقات اللاحقة.
ICP-MS
تم تنظيف قوارير الزجاج المزودة بسدادات تفلون باستخدام حمض النيتريك (VWR، سوبر بور، 1.00441.0250)، مغسول بماء نقي للغاية (سيغما-ألدريتش، 1012620500) وجففت. تم جمع الخلايا وغسلها مرتين بـتم بعد ذلك عد الخلايا باستخدام عداد خلايا آلي (Entek) ونقلها فيPBS أو ماء نقي للغاية إلى قوارير الزجاج النظيفة. نفس الحجم منتم نقل مياه PBS أو المياه فائقة النقاء إلى قوارير منفصلة لطرح الخلفية، على الأقل في نسختين لكل تجربة. بالنسبة لعينات الأنسجة، تم أخذ قطعة صغيرة بحجم حواليتم نقل العينة إلى قنينة نظيفة وموزونة مسبقًا. تم تجفيف العينات بالتجميد باستخدام مجفف بالتجميد (كريست، 2-4 LDplus). تم وزن القناني التي تحتوي على عينات الأنسجة لاحقًا لتحديد الوزن الجاف للأنسجة. ثم تم خلط العينات بحمض النيتريك.وتم تسخينه عندبينما كانت العينات في نفس القوارير الزجاجية المغلقة بغطاء يحمل غشاء تيفلون. ثم تم تبريد العينات إلى درجة حرارة الغرفة وتخفيفها بماء نقي للغاية إلى تركيز نهائي قدره 0.475 N من حمض النيتريك ونقلها إلى قوارير طرد مركزي خالية من المعادن (VWR، 89049-172) لإجراء تحليلات MS لاحقة. تم قياس كميات المعادن باستخدام جهاز Agilent 7900 ICP-QMS في وضع الدقة المنخفضة، مع الأخذ في الاعتبار توزيع النظائر الطبيعية. تم إدخال العينات باستخدام جهاز ميكرونيبولايزر (MicroMist، ) من خلال غرفة رش سكوت. تم قياس النظائر باستخدام واجهة تفاعل تصادمي مع غاز الهيليوم ( ) لإزالة التداخلات متعددة الذرات. تم حقن معايير داخلية من السكنديم والإنديوم بعد الخلط المباشر مع العينات للتحكم في غياب انحراف الإشارة وتأثيرات المصفوفة. تم قياس مزيج من المعايير المعتمدة عند تركيزات تتراوح بين تلك الخاصة بالعينات لتحويل قياسات العد إلى تركيزات في المحلول. تم تطبيع القيم مقابل عدد الخلايا أو الوزن الجاف للأنسجة.
تحضير الليبوسومات ودراسات أكسدة الدهون
تم إعداد الهياكل الليبوزومية باستخدام طريقة ترطيب فيلم الدهون التقليدية. باختصار،محلول مخزون (كلوروفورم ( -cis) PC (DOPC، أفانتي بولار ليبيدز) تم إذابته فيكلوروفورم وتم نقله إلى دورق دائري القاع. تم إزالة المذيب العضوي تحت ضغط منخفض في جهاز التبخر الدوار لمدة 15 دقيقة عند. في حمام مائي عند . بعد ذلك، تم تجفيف الفيلم الدهني باستخدام مضخة تفريغ طوال الليل. تم ترطيب العينة بـ 1 مل من محلول أسيتات الصوديوم 0.1 مليمول (رقم الهيدروجيني 4.5) وتم خلطها في جهاز الخلط كل 5 دقائق لمدة 20 دقيقة. تم تمرير المستحلب عبر غشائين من البولي كربونات (حجم المسام مرات. للتجربة الضابطة، تم إضافة محلول الليبوسوم إلى أنبوب إيبندورف وتسخينه عندمع القلق في. ثم، محلول مائي من ثلاثي فوسفات الحديد (II) (1.4 ملغ في 1.5 مل) وتم إضافة 0.1 مليمول من محلول الأسيتات (pH 4.5). عندمحلول مائي منتم إضافة (1 مل). بالنسبة لتجربة Fento-1،تم إضافة محلول الليبوسوم إلى أنبوب إيبندورف وتم تسخينه عندفي. ثم، محلول بتركيز 1 مللي مولار من Fento-1 في DMSO وتم إضافة 1.4 ملغ من محلول مائي لثلاثي فوسفات الحديد (II) في 1.5 مل.محلول مائي منتم إضافة (1 مل). تم تسجيل أكسدة DOPC باستخدام مطياف الكتلة QExactive (Thermo Fisher Scientific) المزود بمصدر أيون TriVersa NanoMate (Advion Biosciences) كما هو موضح في قسم ‘الليبيدوميات المعتمدة على مطياف الكتلة’. تم حقن العينات في، 7 ساعات و 24 ساعة من أوقات التفاعل.
عزل الفصائل الغنية بالليزوزومات
تم معالجة خلايا HT-1080 بـ Fento-1 (10لمدة ساعة واحدة، وتم عزل الفractions الغنية بالليسوزومات باستخدام مجموعة عزل الليسوزومات (Abcam، ab234047) وفقًا لبروتوكول الشركة المصنعة. باختصار،تم غسل الخلايا وطردها مركزيًا عند 600 جرام لمدة 10 دقائق وتم إزالة السائل العلوي. تم إعادة تعليق الخلايا في محلول عزل الليزوزومات، وتم خلطها باستخدام جهاز الخلط الدوار وتم الاحتفاظ بها على الثلج لمدة دقيقتين. تم الحصول على تكسير كامل للخلايا باستخدام جهاز خلط دوانس. بعد إضافة محلول تعزيز الليزوزومات، تم طرد المستحلب مركزيًا عند 500 جرام لمدة 10 دقائق فيتم إضافة السائل العلوي إلى قمة تدرج الكثافة غير المتواصل وتم إجراء الطرد المركزي الفائق عندلمدة ساعتين عندتم الأداء.
كانت الكمية الغنية بالليزوزومات موجودة في أعلى 10% من حجم التدرج. بالنسبة لعلم الدهون القائم على مطيافية الكتلة، تم خلط الكمية الليزوزومية مع حجمين منPBS، تم دوامه وطرده مركزيًا عند لمدة 30 دقيقة عند . التالي، تم إضافة 150 مللي مول من بيكربونات الصوديوم إلى الرواسب وتم تجميد العينة بسرعة في النيتروجين السائل. تم إجراء تحليلات الدهون على هذه العينات المجمدة بسرعة. بالنسبة لتحليلات الوسترن بلوت، تم قياس محتوى البروتين في السائل الفائق الغني بالليزوزومات باستخدام جهاز قياس الفلورية Qbit 1 (ثيرمو فيشر ساينتيفيك) ومجموعة قياس البروتين (ثيرمو فيشر ساينتيفيك، Q33212). تم تحميل كميات متساوية من البروتين الكلي من مستخلصات الخلايا الكلية والمستخلصات الغنية بالليزوزومات للمقارنة في تحليلات الوسترن بلوت.
الليبيدوميكس القائم على مطيافية الكتلة
للمقارنة بين المحفزات المسببة للفيروبتوز، تم معالجة خلايا HT-1080 بـ Fento-1 (داخل المنزل،إيراستين )، RSL3 ( 100 نانومتر ) أو iFSP1 ( ، سيغما-ألدريتش، SML2749) لمدة 24 ساعة. من أجل المعالجة المشتركة مع مثبطات الفيروبتوسيس، تم معالجة خلايا HT-1080 مسبقًا بـ-توكوفيرولديفيريبيرون أو لمدة ساعتين، ثم مع Fento-1لمدة 24 ساعة. تم معالجة عينات الأورام البشرية المنفصلة مسبقًا بـ-توكوفيرول ثم معفينتو-1 لمدة 24 ساعة. تم معالجة خلايا PDAC053T بـفينتو-1 لمدة 6 ساعات أو 24 ساعة. تم عد عينات الأورام المشتقة من الفئران ومعالجتها مباشرة بعد التفكك وتمت معالجتها مسبقًا بـ-توكوفيروللمدة ساعتين ثم مع Fento-1تم غسل الخلايا بعد ذلك بـتم غسل الخلايا بمحلول PBS ثم بمحلول يحتوي على 150 مليمول من بيكربونات الأمونيوم. بعد ذلك، تم إعادة تعليق الخلايا في 150 مليمول من بيكربونات الأمونيوم وتم الطرد المركزي عند 300 جرام لمدة 5 دقائق. تم إزالة السائل العلوي وأعيد تعليق الخلايا في 1 مل من 150 مليمول من بيكربونات الأمونيوم. تم طرد المحاليل مركزيًا عند 12,000 دورة في الدقيقة لمدة 10 دقائق وتم إزالة السائل العلوي. بعد ذلك،تم إضافة 150 مللي مول من بيكربونات الصوديوم إلى الرواسب وتم تجميد العينات بسرعة في النيتروجين السائل. تم إجراء تحليلات الدهون في نفس اليوم لجميع النسخ التقنية والبيولوجية لمجموعة بيانات معينة. بالنسبة لتحليلات الدهون،تم إضافة مستخلص الخلايا إلىمزيج الدهون القياسي الداخلي يحتوي على 300 بيكومول من الفوسفatidylcholine (PC) 17:0-17:0، 50 بيكومول من الفوسفatidylethanolamine (PE) 17:0-17:0، 50 بيكومول من الفوسفatidylinositol (PI) 16:0-16:0، 50 بيكومول من الفوسفatidylserine (PS) 17:0-17:0، 30 بيكومول من الفوسفatidic acid (PA) 17:0-17:0، 30 بيكومول من الفوسفatidylglycerol (PG)، 30 بيكومول من lysophosphatidylcholine (LPC) 12:0، 30 بيكومول من lysophosphatidylethanolamine (LPE) 17:1، 30 بيكومول من lysophosphatidylserine (LPS) 17:1 و30 بيكومول من lysophosphatidic acid (LPA) 17:0 وتم إخضاعها لاستخراج الدهون فيكما تم وصفه سابقًاباختصار، تم استخراج العينة باستخدام 1 مل من الكلوروفورم والميثانول (10:1) لمدة ساعتين عندمع اهتزاز قوي في جهاز الخلط الحراري (1000 دورة في الدقيقة). تم جمع المرحلة العضوية السفلية وتجفيفها في جهاز تركيز الفراغ SpeedVac. تم إعادة استخراج المرحلة المائية المتبقية باستخدام 1 مل من الكلوروفورم والميثانول.لمدة ساعة واحدة عند نفس درجة الحرارة وظروف الاهتزاز. تم جمع الطور العضوي السفلي وتبخيره في جهاز تركيز الفراغ SpeedVac. تم إذابة مستخلصات الدهون فيخليط التسريب يتكون من 7.5 مللي مولار من أسيتات الأمونيوم المذاب في البروبانول والكلوروفورم والميثانول ). لتجارب الليبوسوم في المختبر، من كل خليط تفاعل تم أخذه في نقاط زمنية مختلفة تم إضافته إلىخليط التسريب يتكون من 7.5 مللي مولار من أسيتات الأمونيوم المذاب في البروبانول والكلوروفورم والميثانول ) تحتوي على 300 بيكومول PC 17:0-17:0. تم تحليل العينات عن طريق الحقن المباشر في مطياف الكتلة QExactive (Thermo Fisher Scientific) المزود بمصدر أيوني TriVersa NanoMate (Advion Biosciences). باختصار، تم حقن العينة بضغط الغاز والفولتية المحددة على 1.25 psi و 0.95 kV، على التوالي. PC والأكسدةتم الكشف عنها في المستخلص بنسبة 10:1 بواسطة FTMS في وضع الأيونات الموجبة كأدكتات بروتونية من خلال المسح، في مع تفعيل الكتلة المقفلة في خلفية مشتركةلمدة 30 ثانية. كل مسح هو متوسط 2 ميكروسكان، وتم ضبط AGC علىوتم تعيين الحد الأقصى لوقت حقن الأيونات (IT) إلى 50 مللي ثانية. PE، PE مؤكسد ( ) وتم الكشف عن LPE كأدوات مضافة مزيلة للبروتون، وتم الكشف عن LPC كأدوات مضافة للأسيتات في المستخلص بنسبة 10:1 بواسطة FTMS في وضع الأيون السالب من خلال المسح، في مع تفعيل الكتلة المقفلة في خلفية مشتركة لمدة 30 ثانية. كل مسح هو متوسط 2 ميكروسكان، تم ضبط AGC على وكان الحد الأقصى للتيار الأيوني محددًا بـ 50 مللي ثانية. PI، PI المؤكسد ( )، PS، PS المؤكسد (PSتم الكشف عن الليسوفوسفوليديلينوسيتول (LPI) والليسوفوسفوليديلسيرين (LPS) في المستخلص بنسبة 2:1، بواسطة تقنية FTMS في وضع الأيونات السالبة كأيونات مزاحة البروتون من خلال المسح.، في مع تنشيط الكتلة المقفلة في خلفية مشتركة ( ) لمدة 30 ثانية. كل مسح هو متوسط 2 ميكروسكان، تم ضبط AGC على وتم تعيين الحد الأقصى لأيون IT إلى 50 مللي ثانية. توضح التعليقات التوضيحية للفوسفوليبيدات المؤكسدة فقط عدد ذرات الأكسجين الإضافية والروابط المزدوجة الموجودة في كتلة معينة، ويمكن أن تشير بشكل محتمل إلى عدة هياكل مختلفة بنفس الكتلة الأحادية النظائر. تم الحصول على جميع البيانات في وضع المركز. تم تحليل جميع بيانات الليبيدوميات باستخدام برنامج تحديد الليبيد LipidXplorer (الإصدار 1.2.8؛http://genomebiology.com/2011/12/1/R8تم ضبط التسامح لـ MS والتحديد على 2 جزء في المليون. تم تطبيع البيانات مقابل المعايير الداخلية والإدخال الكلي لأنواع الدهون المعنية. تم تجميع بيانات الدهون باستخدام مسافة الارتباط ورابط وورد، وتم تمثيلها كخرائط حرارية للقيم المطبوعة.
تحديد كمية الجلسرين
تم معالجة خلايا HT-1080 بـ Fento-1 (داخلي، 1 أوتم قياس الجليسرول باستخدام اختبار Glycerol-glo (Promega, J3150) وفقًا لبروتوكول الشركة المصنعة. باختصار، تم إجراء الاختبار في لوحة 96 بئر وتم زراعة 4000 خلية لكل بئر قبل 24 ساعة من التجربة. تم إعداد منحنى قياسي لكل تجربة بيولوجية وتم إجراء ثلاث تكرارات فنية لكل حالة ولكل تكرار بيولوجي. تم تسجيل إشارات اللمعان باستخدام جهاز قراءة الألواح SpectraMax ID3 (Molecular Devices). تم تصدير البيانات وتحليلها باستخدام Excel (Microsoft) وبرنامج Prism.
برمجيات للرسوم التوضيحية
تم إنشاء الرسوم التوضيحية باستخدام Fiji 2.0.0-rc-69/1.52n و Prism 10.0.3 و Adobe Illustrator 26.0.2. تم إنشاء الرسوم التوضيحية في الأشكال 2b و 4 باستخدام BioRender.https://biorender.com).
الإحصائيات وإمكانية التكرار
تُعرض النتائج كمتوسط ± الانحراف المعياري أو الخطأ المعياري كما هو موضح. بالنسبة لمخططات الصندوق، تمثل الصناديق النطاق الربعي الوسيط، وتشير الشعيرات إلى القيم الدنيا والقصوى. تم استخدام برنامج Prism (الإصدار 10.0.3) للحساب.القيم باستخدام اختبار مان ويتني ذو الجانبين، غير المقترن ذو الجانبين-اختبار، اختبار كروسكال-واليس مع اختبار دان بعده، تحليل التباين أحادي الاتجاه، تحليل التباين ثنائي الاتجاه أو اختبار مانتل-كوكس لوج-رانك كما هو موضح. تم استخدام برنامج بريزم (الإصدار 10.0.3) لإنشاء تمثيلات رسومية للقياسات ما لم يُذكر خلاف ذلك. أحجام العينات ( ) موضحة في أساطير الأشكال ولم تكن محددة مسبقًا. الحد الأدنى من تم إجراء تجارب مستقلة كمعيار، وتم زيادة أحجام العينات في التجارب الأكثر تعقيدًا لضمان القابلية للتكرار. بالضبطالقيم موضحة في الأرقام.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
تم تقديم بيانات الليبيدوميكس في الجداول التكميلية 1-4 و 7-12. تم إيداع بيانات البروتيوميك الكمية الخام في اتحاد بروتيوم إكس من خلال PRIDE.مستودع الشريك مع معرف مجموعة البيانات PXD054449. يتم توفير البيانات المصدر مع هذه الورقة. 50. فريش، م. ج. وآخرون. غاوسي 09، الإصدار ب. 01 (غاوسي، 2010). 51. شرايبر، ف. س. وآخرون. النمو الناجح وتوصيف خلايا القناة البنكرياسية للفئران: الخصائص الوظيفية لـ Ki-RASجين الأورام. أمراض الجهاز الهضمي 127، 250-260 (2004). 52. ياداف، ب.، وينربرغ، ك.، أيتوكاليو، ت. وتانغ، ج. البحث عن تآزر الأدوية في مشاهد استجابة الجرعة المعقدة باستخدام نموذج قوة التفاعل. مجلة الحوسبة والهندسة الحيوية 13، 504-513 (2015). 53. أيوناك، م. تقنية واحدة، نهجان، ونتائج: قسطرة القناة الصدرية في الحيوانات المخبرية الصغيرة. الجراحة المجهرية 23، 239-245 (2003). 54. بوليه، ب.، كاربنتييه، س. وباريلو، إ. myProMS، خادم ويب لإدارة والتحقق من بيانات البروتيوميات المستندة إلى مطيافية الكتلة. بروتيوميات 7، 2553-2556 (2007). 55. بيريز-ريفرول، ي. وآخرون. موارد قاعدة بيانات PRIDE في عام 2022: مركز لأدلة البروتيوميات المعتمدة على مطيافية الكتلة. أبحاث الأحماض النووية. 50، D543-D552 (2022). 56. ووركمان، ب. وآخرون. إرشادات لرعاية واستخدام الحيوانات في أبحاث السرطان. المجلة البريطانية للسرطان 102، 1555-1577 (2010). 57. سامبايو، ج. ل. وآخرون. دهن الغشاء لخط خلوى ظهاري. وقائع الأكاديمية الوطنية للعلوم في الولايات المتحدة الأمريكية 108، 1903-1907 (2011).
الشكر والتقدير R.R. مدعوم من المجلس الأوروبي للبحث تحت برنامج الأبحاث والابتكار الخاص بالاتحاد الأوروبي (الاتفاقية رقم 647973)، Ligue Contre le Cancer (فريق معتمد)، Fondation pour la Recherche Médicale، Fondation Charles Defforey-Institut de France، Klaus Grohe Foundation، Fondation Bettencourt Schueller، Région Ile-de-France، Institut National du Cancer (PLBIO-2022-099) وAgence Nationale de la Recherche (ANR-23-CE44-0030 وANR-23-IAHU-0006). V.G. يشكر UPSaclay وCNRS على الدعم المالي. تم منح هذا العمل الوصول إلى موارد HPC من CINES بموجب التخصيص 2020-A0070810977 الذي تم إجراؤه بواسطة GENCI. حصلت M.C. على تمويل من Deutsche Forschungsgemeinschaft (DFG) (CO 291/7-1، 428858739 وبرنامج الأولوية SPP 2306 (CO 291/9-1، 461385412؛ CO 291/10-1، 461507177؛ CO 291/9-2، CO 291/10-2، CO 291/14-1) وCRC 353 (CO 291/11-1؛ 471011418)، وزارة التعليم والبحث الفيدرالية الألمانية (BMBF) برنامج VIP+ NEUROPROTEKT (O3VPO4260)، برنامج BMBF FERROPATH (01EJ2205B) والمجلس الأوروبي للبحث تحت برنامج الأبحاث والابتكار الخاص بالاتحاد الأوروبي (الاتفاقية رقم 884754). J.P.F.A. يعترف بالدعم المؤسسي من جامعة وورزبورغ والدعم الإضافي من Deutsche Forschungsgemeinschaft (DFG)، FR 3746/3-1، FR 3746/5-1، FR 3746/6-1 وCRC205 (INST 269/886-1)، الاتحاد الأوروبي (ERC-Consolidator، DeciFERR، منحة Horizon Europe ERC رقم 101126134) ومؤسسة Deutsche Jose Carreras Leukämie Stiftung (DJCLS 01R/2022). J.M.U. مدعوم من منحة الباحث الشاب من Breast Cancer Alliance ومركز لودفيغ في هارفارد. A.C. مدعوم من زمالة أبحاث بيولوجيا السرطان من Landry. تم تمويل A.C. وC.S.F. من قبل NIH NIDDK (T32DK128781). E.C.-J. وC. Ginestier مدعومون من Ligue Contre le Cancer (فريق معتمد). D.A.P. مدعوم من العلوم الطبيعية و
مجلس أبحاث الهندسة في كندا (RGPIN-2022-05058). يتم دعم منصة ICP-MS في معهد فيزياء الكرة الأرضية في باريس من قبل البرنامج متعدد التخصصات IPGP PARI ومنطقة إيل دو فرانس (اتفاقية منحة SESAME رقم 12015908). مرافق تصوير الخلايا والأنسجة (PICT-IBiSA) في معهد كوري هي جزء من البنية التحتية الوطنية للتصوير الحيوي في فرنسا (ANR-10-INBS-04 و ANR-24-INBS-0005 FBI BIOGEN)، وتدعم منصة MS من قبل منطقة إيل دو فرانس (NºEX061034) و ITMO Cancer من Aviesan و INCa المدعومة من الأموال التي تديرها INSERM (رقم 21CQ016-00)، ومنصة السيتومتر هي جزء من معهد كوري. نشكر S. Hidalgo على المساعدة في تجارب ICP-MS؛ وR.R. يشكر S. Balasubramanian و S.L. Schreiber و J.-M. Lehn و E.M. Carreira على الدعم.
“مساهمات المؤلفين: قام R.R. بتصميم الدراسة وتوجيه البحث. قام R.R. وT.C. وL.B. وS.M. وJ.M.U. بتصميم التجارب. قام T.C. وL.B. وL.C. وA.V. وC. Gaillet بإجراء التخليق الكيميائي، وتحليل الطيف بالرنين المغناطيسي النووي، وقياس الكتلة. قام L.B. وL.G. بإجراء الفولتمترية الدائرية. قام A.U. بقياس نشاط RTA في المختبر. قام V.G. بإجراء النمذجة الجزيئية. قام S.M. وL.B. وS.S. وF.S. وA.P.-B. وL.K.T. بإجراء عمليات النقل، والتحويل، وتصوير الفلورية، وقياس تدفق الخلايا، وتحليل البروتينات الغربية، وعزل الليسوزومات، واختبارات موت الخلايا، واختبارات بقاء الخلايا. قام S.M. بتحضير العينات لتحليل ICP-MS. قام J.L.S. بإجراء تحليل الدهون. قام F.S. وB.L. وD.L. بإجراء تحليل البروتينات الكمي. قام L.S. وN.S. بإجراء التحليلات الحيوية. قام E.D.K. وC.B. وP.S.-C. وM.E. وN.D. وJ.L.I. وF.P.F. وJ.Z. بإجراء اختبارات تشكيل المستعمرات، والفتك الاصطناعي، واختبارات العلاج المركب. قام A.S.C. وG.P. وD.T. وS.B. باختيار عينات الأنسجة السليمة والورمية أثناء العملية. قام S.M. وS.S. بإجراء تحليلات لعينات الأنسجة الورمية البشرية. قام E.M. وB.P. بتقييم cLip-1 في الجسم الحي. قام A.C. وM.S. وC.S.F. وK.J.S. بإجراء علاجات داخل اللمفاوية لفئران تحمل ورم 4T1. ساهم جميع المؤلفين في تحليل البيانات. كتب R.R. وT.C. وS.M. المقال. علق E.C-.J. وC. Ginestier وP.H. وS.W. وD
المصالح المتنافسة قدم معهد كوري طلب براءة اختراع (PCT/EP2025/O51999) يتعلق بمحللات الدهون لتحفيز الفيروبتوز في السرطان. يحمل م.ك. و ب.ب. براءات اختراع لبعض المركبات الموصوفة هنا وهما مؤسسان مشاركان لشركة ROSCUE Therapeutics. ب.ر.س. هو مخترع في براءات اختراع وطلبات براءات تتعلق بالفيروبتوز؛ شارك في تأسيس ويعمل كمستشار لشركة ProJenX و Exarta Therapeutics؛ يمتلك أسهماً في شركة Sonata Therapeutics؛ ويعمل كمستشار لشركة Weatherwax Biotechnologies و Akin Gump Strauss Hauer & Feld. جميع المؤلفين الآخرين يعلنون عدم وجود مصالح متنافسة.
معلومات إضافية
معلومات إضافية النسخة الإلكترونية تحتوي على مواد إضافية متاحة فيhttps://doi.org/10.1038/s41586-025-08974-4. يجب توجيه المراسلات والطلبات للحصول على المواد إلى رافائيل رودريغيز. معلومات مراجعة الأقران تشكر مجلة نيتشر المراجعين المجهولين على مساهمتهم في مراجعة هذا العمل. تقارير مراجعي الأقران متاحة. معلومات إعادة الطباعة والتصاريح متاحة علىhttp://www.nature.com/reprints.
الشكل البياني الممتد 1|انظر الصفحة التالية للتعليق.
الشكل البياني الممتد 1 | تعطيل الحديد الليزوزومي يمنع الفيروبتوز (الجزء 1). أ، التركيب الكيميائي لـ cLip-1. ب، تصوير الفلورسنت لـ cLip-1 المعلم.وعلامات محددة للعضيات الخلوية. قضبان القياس،. تجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري. ج، تصوير الفلورسنت لـ cLip-1 المعلم.و LysoTracker. شريط القياس،تجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري، تصوير الفلورسنت لـ cLip-1 المعلم.وخلايا تم نقلها بواسطة BacMam مع علامات عضيات خلوية موسومة بـ GFP. مقياس الرسم،تجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري. ج، تصوير الفلورسنت لـ cLip-1 المعلم في أنسجة الكلى القريبة والكبد من Rosa26-CreERT2;Gpx4فأر تم معالجته بـ cLip-1 و يوم؛ تم التضحية بعد ساعة واحدة من الحقن داخل الصفاق). شريط القياس،.ه، بوديبي-C11581/591تدفق تحليل الخلايا المعالجة بـ Lip-1 و RSL3 لمدة 1 ساعة. i، قابلية الخلايا (الأعلى، بعد 72 ساعة من الحضانة؛ الوسط، الحضانة لمدة 24 ساعة والعلاج المشترك مع RSL3; القاع، الحضانة لمدة 72 ساعة والمعالجة المشتركة مع 4-OH TAM لإلغاء التأثير. البيانات هي المتوسط ± الانحراف المعياري. صبغة Annexin-V/Propidium Iodide (A/PI) في الخلايا المعالجة لمدة 24 ساعة. RSL3: تجارب مستقلة؛ RSL3+Lip-1:تجارب مستقلة؛ RSL3 + cLip-1:تجارب مستقلة. ك،طيف الرنين المغناطيسي النووي لـ Lip-1 والنفتالين المعاير بـ. طيف الرنين المغناطيسي النووي لـ Lip-1 المعياري معثم TFA.o،طيف الرنين المغناطيسي النووي لـ Lip-1 المضاف إليهثم ديترواكسيد الصوديوم (NaOD). k-o، طيف NMR المسجل عند 310 كلفن في الميثانول-. ب، د-ف، تحليل التباين الأحادي. وحدة عشوائية. ن
الشكل البياني الممتد 2 | انظر الصفحة التالية للتعليق.
الشكل البياني الموسع 2 | تعطيل الحديد الليزوزومي يمنع الفيروبتوز (الجزء 2). أ، التخليق الكيميائي لـ metcLip-1 و alcLip-1. ب، تصوير الفلورسنت لـ cLip-1 المعلم.ميتكلب-1 و alcLip-1 مقياس. ج، الفولتمترية الدورية لـ حل (إمكانات الاختزال)السهم الوردي). البيانات المسجلة في وجود metcLip-1 أو alcLip-1. د، تحليل تدفق الخلايا باستخدام Bodipy-C11581/591 للخلايا المعالجة بـ metcLip-1الكليب-1و RSL3لمدة ساعة واحدة. ممثل لـتجارب مستقلة لـ PDAC053T ولـ HT-1080. e، اختبار FENIX لـ cLip-1 و metcLip-1 و alcLip-1. f، التركيب الجزيئي لـ cDFO وتصوير الفلورسنت لـ cDFO المعلم.مقياستجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري. ج، طيف انبعاث الفلورسنت لـ RPE عند pH5 في وجودو Lip-1.h، تحليل تدفق الخلايا باستخدام تقنية الفلورية للخلايا المعالجة لمدة 30 دقيقة ثم المعالجة المشتركة مع المجس لـممثل لـتجارب مستقلة. أنا، تحليل تدفق HMRhoNox-M للخلايا المعالجة لمدة 30 دقيقة ثم المعالجة المشتركة مع المجس لـممثل لـتجارب مستقلة. ج، قياس تدفق الخلايا باستخدام Bodipy-C11581/591 للخلايا المعالجة بـ RSL3 (200 نانومتر لـ PDAC053T، 500 نانومتر لـ 4T1، 1 ساعة) و Baf-A1 أو HCQ ( ) تم استخدامه قبل ساعتين. ممثل عن تجارب مستقلة. ك، تصوير الفلورسنت لبوديبي 665/676 في خلايا تم علاجها بـ RSL3مقياستجارب مستقلة. غير متطابقة ذات جانبين-اختبار. البيانات هي المتوسط ± الانحراف المعياري. التركيزات المستخدمة ما لم يُذكر خلاف ذلك: ليب-1باف-أ1.
الشكل البياني الممتد 3 | انظر الصفحة التالية للتعليق.
الشكل البياني الممتد 3 | المحفزات المؤدية إلى الفيروبتوزيس تبدأ أكسدة الدهون في الليزوزومات. أ، تصوير فلوري لمؤشرات العضيات الخلوية المعلمة بـ GFP في خلايا تم نقلها بواسطة BacMam ومعالجة بـ RSL3.شريط القياس،تجارب مستقلة. البيانات هي المتوسطتصوير الفلورية لبوديبي 665/676 وخلايا تم نقلها بواسطة باك مام مع علامات عضيات الخلايا المعلّمة بـ GFP المعالجة بـ RSL3مقاييس الرسم تجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري. ج، تصوير الفلورية لمسبار الجلوتاثيون في الليزوزومات (SQSS) وعلامة ليزوزومية في الخلايا المعالجة بـ RSL3.مقياسممثل لـتجارب مستقلة. د، قياس التدفق الخلوي باستخدام SQSS للخلايا المعالجة بمحفزات الفيروبتوز. هـ، قياس التدفق الخلوي 1-ريد للخلايا المعالجة بمحفزات الفيروبتوز. أ، ب، اختبار غير متزاوج ذو جانبين.-اختبار. ن
الشكل 4 من البيانات الموسعة | التركيب الكيميائي والبيانات الطيفية لفينتو-1. أ. التخليق الكيميائي لفينتو-1.bطيف الرنين المغناطيسي النووي لفينتو-1.cطيف الرنين المغناطيسي النووي لفينتو-1.d. طيف الكتلة عالي الدقة (HRMS) لفينتو-1.
طيف الكتلة منخفض الدقة (LRMS) لمركب فينتو-1-حديد.كيميائي تركيب cCW.طيف NMR المسجل عند 298 كلفن في الميثانول-. كامل الوصف، معلومات إضافية. طيف الكتلة منخفض الدقة (LRMS) لمركب فنتو-1-حديد. التركيب الكيميائي لـ cCW.b، c، طيف NMR المسجل عند 298 كلفن في الميثانول-. الوصف الكامل، المعلومات التكميلية.
الشكل 5 من البيانات الموسعة | انظر الصفحة التالية للتعليق.
مقالة
الشكل 5 من البيانات الموسعة | Fento-1 يحفز أكسدة الفوسفوليبيدات وموت الخلايا عن طريق تنشيط الحديد الليزوزومي (الجزء 1). أ، تصوير فلوري لـ Fento-1 وmarmycin وعلامة سطح الخلية CD44 في الخلايا المعالجة عند لمدة ساعة واحدة. شريط القياس، تجارب مستقلة. البيانات هي المتوسطتصوير الفلورية لـ Fento-1 في خلايا تم نقلها بواسطة BacMam مع علامات عضيات خلوية موسومة بـ GFP. مقياس الرسم،تجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري، تحليل تدفق الخلايا باستخدام Bodipy-C11581/591 للخلايا المعالجة بـ Fento-1 و marmycin و cCW بمفردها أو مجتمعة لمدة 6 ساعات. تمثيلي لـتجارب مستقلة. د، قابلية الخلايا المعالجة لمدة 6 ساعات. البيانات هي المتوسط ± الانحراف المعياري.تجارب مستقلة. هـ، قابلية الخلايا المعالجة لمدة 72 ساعة. البيانات هي متوسطس.م.تجارب مستقلة. علم الدهون للفوسفوليبيدات المؤكسدة في الخلايا المعالجة بـ Fento-1 أو المحفزات المؤدية إلى الفيروبتوز لمدة 24 ساعة.تجارب مستقلة. ج، ليبيدوميكس للفوسفوليبيدات المؤكسدة في الخلايا المعالجة بـ Fento-1. ن = 4 تجارب مستقلة. خريطة حرارية كاملة، معلومات إضافية. ح، ليبيدوميكس للفوسفوليبيدات المؤكسدة من مستخلصات الليزوزومات في الخلايا المعالجة بـ Fento-تجارب مستقلة. أنا، تحليل البروتينات باستخدام تقنية الويسترن بلوت من خلايا تم معالجتها بـ Fento-1، مارمايسين، cCW أو مارمايسين-توبولين هو عنصر تحكم في تحميل العينة. يمثلتجارب مستقلة. ج، تحليل البروتينات باستخدام تقنية الويسترن بلوت من خلايا تم معالجتها بـ Fento-1، مارمايسين، cCW أو مارمايسين. -توبولين هو عنصر تحكم في معالجة العينة. يمثلتجارب مستقلة مع نتائج مشابهة. ك، تحليل تدفق الخلايا باستخدام Bodipy-C11581/591 للخلايا المعالجة بـ Fento-ومثبطات استخدمت قبل ساعتين.تجارب مستقلة. مخططات الصندوق: نطاق الربيع الداخلي، خطوط المركزالوسائط والشعيراتالقيم الدنيا والقصوى. أنا، تصوير الفلورية لـ 4-HNE المعالج بـ Fento-1 (1 ساعة). اختبار غير متزاوج ذو جانبين-اختبار.تجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري. شريط المقياس،تحديد كمية الجليسرول في الخلايا المعالجة بـ Fento-1البيانات هي متوسطس.م.تجارب مستقلة. التركيزات المستخدمة ما لم يُذكر خلاف ذلك: فينتو-1مارميسين, cCW إيراستينRSL3، و cLip-1 . ك، م، اختبار كروسكال-واليس مع اختبار دان. تم عرض الدهون الفوسفورية (PC) في الأشكال الرئيسية. الفوسفatidylethanolamine (PE)، الفوسفatidylinositol (PI)، الفوسفatidylserine (PS)، معلومات إضافية.
الشكل البياني الممتد 6 | انظر الصفحة التالية للتعليق.
مقالة
الشكل 6 من البيانات الموسعة | Fento-1 يحفز أكسدة الفوسفوليبيدات وموت الخلايا عن طريق تنشيط الحديد الليزوزومي (الجزء 2). أ، توهج CellTiterGlo للخلايا المعالجة بـ Fento-ومثبطات تم استخدامها قبل ساعتين.تجارب مستقلة. ب، فلوروسين ريسوروفين للخلايا المعالجة بـ Fento-1ومثبطات استخدمت قبل ساعتين. المركب، ليب-1:تجارب مستقلة، كليب-1 و Z-VAD-FMK:تجارب مستقلة، فير-1، ديف وتوك:تجارب مستقلة، DFO، DFX وفيتامين K3:تجارب مستقلة. ج، د، توهج CellTiterGlo للخلايا المعالجة بـ Fento-1ومثبطات استخدمت قبل ساعتين.تجارب مستقلة. قياس حيوية الخلايا باستخدام إنكوسايت. تمثيلي لـتجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري للخطأ، قابلية بقاء الخلايا المعالجة بـ Fento-1أو ليب-1أو فينتو-1 وليب-1 تحليل التباين أحادي الاتجاه.التكرارات الفنية. تصوير الفلورسنت لـ Fento-1. Fento-1 و سابقشريط القياس،تجارب مستقلة. غير متطابقة ذات جانبين-اختبار. البيانات هي المتوسط ± الانحراف المعياري. ح، حيوية الخلايا المعالجة لـتجارب مستقلة. البيانات هي المتوسط ± الانحراف المعياري. اليسار: موت الخلايا في الخلايا التي تم تقليلها بواسطة قياس Annexin-V/Sytox الأزرق في الخلايا المعالجة بـ Fento-1 لـتجارب مستقلة. تحليل التباين ثنائي الاتجاه. البيانات هي المتوسط ± الانحراف المعياري. اليمين: تحليل البروتينات بواسطة تقنية الويسترن بلوت في خلايا تم تقليل التعبير فيها.-توبولين هو عنصر تحكم في معالجة العينة. ج، تدفق السيتومترية لبوديبى-C11581/591 للخلايا التي تم تقليلها بعد المعالجة لمدة 6 ساعات. مخططات الصندوق: النطاق بين الربعين، والخطوط المركزية = الوسيطات والشعيرات.القيم الدنيا والقصوى. أ-د، اختبار مان-ويتني ذو الجانبين مقارنةً بالعقار الوهمي.
الشكل البياني الممتد 7| نظائر الفينتوميسين. أ، الهياكل الجزيئية. ب، حيوية الخلايا المعالجة لمدة 6 ساعات أو 72 ساعة. البيانات هي المتوسط ± الانحراف المعياري.تجارب مستقلة. تدفق السيتومترية للخلايا المعالجة لمدة 6 ساعات. تمثيلي لـتجارب مستقلة.
الشكل البياني الممتد 8 | انظر الصفحة التالية للتعليق.
الشكل 8 من البيانات الموسعة | Fento-1 يحفز الفيروبتوز في CD44خلايا السرطان الغنية بالحديد (الجزء 1). أ، ICP-MS للأنسجة الصحية والسرطانية من المرضى. PDAC #2: صحيقطع نسيج، سرطان البنكرياس القنويقطع نسيج؛ PDAC #3: صحيقطع نسيج، سرطان البنكرياس القنويقطع نسيج؛ PDAC #4: صحيقطع نسيج، سرطان البنكرياس القنويقطع نسيج؛ PDAC #5: صحيقطع نسيج، سرطان البنكرياس القنويقطع نسيج؛ PDAC #6: صحيقطع نسيج، سرطان البنكرياس القنويقطع نسيج؛ PDAC #7: صحيقطع نسيج، سرطان البنكرياس القنويقطع نسيج؛ PDAC #8: صحيقطع الأنسجة،قطع نسيجية؛ PDAC #9: صحيقطع نسيج، سرطان البنكرياس القنويقطع نسيجية؛ ورم وعائي #1: صحيقطع نسيج، ورم وعائيقطع نسيجية؛ ورم وعائي #2: صحيقطع نسيج، ورم وعائي ساركوميقطع نسيج؛ ليفوساركوما #2: صحيقطع نسيج، ليفوساركوماقطع نسيج؛ ساركوما ظهارية: صحيقطع نسيج، ساركوما ظهاريةقطع نسيج؛ نقائل كبدية من سرطان البنكرياس: صحيقطع نسيج، نقائل كبدية لسرطان البنكرياسقطع الأنسجة. تحليل الطيف الكتلي بالتحليل الطيفي للكتلة المتزامنة للخلايا السرطانية البشرية المفككة.نسخ. تدفق الخلايا السرطانية من UPS البشري باستخدام تقنية HMRhoNox-M.المرضى. د، علم الدهون للفوسفوليبيدات المؤكسدة في خلايا الورم البشرية المفصولة المعالجة بـ Fento-1واستخدم توك قبل ساعتين. علم الدهون من الليسوفوسفوليبيدات في خلايا الأورام البشرية المفصولة المعالجة بـ Fento-1واستخدم توك قبل ساعتين. ف، تحليل تدفق الخلايا لبوديبى-C11581/591 لخلايا سرطان البنكرياس البشرية المفصولة التي تم علاجها بفينتو-1 (24 ساعة) والمثبطات المستخدمة قبل ساعتين.المرضى.g، تحليل تدفق الخلايا باستخدام Bodipy-C11581/591 لخلايا سرطان UPS المفصولة المعالجة بـ Fento-1ومثبطات استخدمت قبل ساعتين.المرضى.h، عد خلايا الفلوسيتومترية لخلايا PDAC البشرية المنفصلة CD44خلايا السرطان المعالجة بـ Fento-1ومثبطات تم استخدامها قبل ساعتين.المرضى. أنا، تحليل تدفق الخلايا لـ CD44 لخلايا سرطان UPS المفصولة المعالجة بـ Fento-ومثبطات تم استخدامها قبل ساعتين.المرضى. التركيزات المستخدمة ما لم يُذكر خلاف ذلك: فينتو-1، تعريفليپ-1 و cLip-1 اختبار f-i، تحليل التباين الأحادي. ج، f-i، كل نقطة ملونة تمثل ورمًا لمريض مميز في لوحة معينة. مخططات الصندوق: نطاق الربع، خطوط المركزالوسيطات والشعيراتالقيم الدنيا والقصوى.
الشكل البياني الموسع 9 | انظر الصفحة التالية للتعليق.
الشكل 9 من البيانات الموسعة | Fento-1 يحفز الفيروبتوز في CD44خلايا السرطان الغنية بالحديد (الجزء 2). أ، حيوية الخلايا المعالجة بـ Fento-1 أو العلاج الكيميائي القياسي لمدة 72 ساعة. البيانات هي متوسطس.م.تجارب مستقلة. ب، قابلية البقاء للأورغانويد المستمد من PDAC المعالج بـ Fento-1 أو العلاج الكيميائي القياسي لمدة 72 ساعة. البيانات هي المتوسط ± الانحراف المعياري.-القيم موضحة. PDAC009T: أوكساليبلاتين، 5-FU وSN-38تجارب مستقلة، فينتو-1تجارب مستقلة. PDAC003T: أوكزالبلاتينتجارب مستقلة، 5-FU و SN38تجارب مستقلة، فينتو-تجارب مستقلة. PDAC117T: أوكساليبلاتين، 5-FU و SN-38تجارب مستقلة، فينتو-تجارب مستقلة. PDAC372T: أوكساليبلاتين، 5-FU و SN-38تجارب مستقلة، فينتو-تجارب مستقلة. ج، قابلية الخلايا المعالجة بالعلاج الكيميائي القياسي وجرعات غير مميتة من Fento-1.1-way ANOVA. البيانات هي المتوسط ± الانحراف المعياري. n = 3 تجارب مستقلة. د، درجة التآزر لـ العلاج الكيميائي القياسي وFento-1 في خلايا PDAC الأولية البشرية. e، حيوية الخلايا المعالجة بـ Fento-1 (72 ساعة) في خلايا غير معالجة أو خلايا تم معالجتها مسبقًا بـ Doxoلمدة 30 يومًا، ثم تم العلاج بـ Fento-1 لمدة 72 ساعة. البيانات هي متوسط ± الانحراف المعياري.تجارب مستقلة. صور مجال ساطع للخلايا المعالجة بـ Doxo (أيام). مقياس الرسم،ممثل لـتجارب مستقلة مع نتائج مشابهة. على سبيل المثال، تحليل تدفق الخلايا CD44 للخلايا المعالجة بدوكسو (25 نانومول) أو فينتو-لمدة 10 أيام. ممثل لـتجارب مستقلة. ه، تحليل الغربي لمؤشرات الخلايا الجذعية وتوازن الحديد في الخلايا المعالجة بدوكسو (25 نانومول) لمدة 10 أيام. لامين A/C هو عنصر تحكم في تحميل العينة. تمثيلي لـتجارب مستقلة مع نتائج مشابهة. تحليل الغربي لمؤشرات الخلايا الجذعية وتوازن الحديد في الخلايا المعالجة بـ Fento-لمدة 10 أيام.-توبولين هو عنصر تحكم في تحميل العينة. يمثلتجارب مستقلة مع نتائج مشابهة. دوكسو، دوكسوروبيسين.
الشكل 10 من البيانات الموسعة | Fento-1 يحفز الفيروبتوز في CD44خلايا السرطان الغنية بالحديد (الجزء 3). أ، ICP-MS من الدم، المصل والسائل اللمفاوي من الفئران. الدم والمصل:فئران؛ لمففالفئران. تحليل التباين الأحادي الاتجاه. ب، قياس التدفق HMRhoNox-M لخلايا سرطان الورم المشتتة من الفئران.الفئران. كل نقطة ملونة تمثل ورمًا لفأر مميز. اختبار مان ويتني ثنائي الجانب. ج، قطر الورم في الفئران المعالجة بـ Fento-1لكل حيوان كل يومين). تحليل التباين ثنائي الاتجاه. البيانات هي المتوسط ± الانحراف المعياري.تجربة: مركبةفئران، فينتو-فئران؛تجربة:فئران لكل مجموعة؛تجربة:فئران لكل مجموعة. د، قطر الورم في الفئران الحاملة للورم المعالجة بـ Fento-1مركب لكل حيوان كل يومين).تجارب مستقلة.تجربة:فئران لـ كل مجموعة؛سيارة التجربة:فئران، فينتو-فئران؛تجربة:فئران لكل مجموعة؛تجربة:الفئران لكل مجموعة. وزن جسم الفئران المعالجة كما هو موضح في الشكل 3h والشكل الإضافي 10c.1تجربة:فئران لكل مجموعة؛تجربة: مركبةفئران، فينتو-فئران؛تجربة:فئران لكل مجموعة؛تجربة:فئران لكل مجموعة. البيانات هي المتوسط ± الانحراف المعياري. علم الدهونيات للدهون الفوسفورية في أورام الفئران الحاملة للأورام المعالجة بـ Fento-1 كما في الشكل 3j.أورام 4T1. تم عرض الدهون LPC. LPE، LPS، معلومات إضافية. مخططات الصندوق: النطاق الربعي، خطوط المركزالوسائط والشعيراتالقيم الدنيا والقصوى.
محفظة الطبيعة
المؤلف (المؤلفون) المراسلون: رافائيل رودريغيز آخر تحديث من المؤلف(ين): 7 مارس 2025
ملخص التقرير
تسعى Nature Portfolio إلى تحسين إمكانية تكرار العمل الذي ننشره. يوفر هذا النموذج هيكلًا للاتساق والشفافية في التقرير. لمزيد من المعلومات حول سياسات Nature Portfolio، يرجى الاطلاع على سياسات التحرير وقائمة التحقق من سياسة التحرير.
الإحصائيات
لجميع التحليلات الإحصائية، تأكد من أن العناصر التالية موجودة في أسطورة الشكل، أسطورة الجدول، النص الرئيسي، أو قسم الطرق. غير متوفر □ X □ □ □ □
وصف لجميع المتغيرات المشتركة التي تم اختبارها □ □ □ □ تم التأكيد
اختبار(ات) الإحصاء المستخدمة وما إذا كانت أحادية الجانب أو ثنائية الجانبالقيم كقيم دقيقة كلما كان ذلك مناسبًا. □ □
حجم العينة بالضبطلكل مجموعة/شرط تجريبي، معطاة كرقم منفصل ووحدة قياس بيان حول ما إذا كانت القياسات قد أُخذت من عينات متميزة أو ما إذا كانت نفس العينة قد تم قياسها عدة مرات
يجب أن تُوصف الاختبارات الشائعة فقط بالاسم؛ واصفًا التقنيات الأكثر تعقيدًا في قسم الطرق.
وصف لأي افتراضات أو تصحيحات، مثل اختبارات الطبيعية والتعديل للمقارنات المتعددة وصف كامل للمعلمات الإحصائية بما في ذلك الاتجاه المركزي (مثل المتوسطات) أو تقديرات أساسية أخرى (مثل معامل الانحدار) وَ التباين (مثل الانحراف المعياري) أو تقديرات مرتبطة بعدم اليقين (مثل فترات الثقة)
لاختبار الفرضية الصفرية، إحصائية الاختبار (على سبيل المثال، ) مع فترات الثقة، أحجام التأثير، درجات الحرية والقيمة ملحوظة
لتحليل بايزي، معلومات حول اختيار القيم الأولية وإعدادات سلسلة ماركوف مونت كارلو للتصاميم الهرمية والمعقدة، تحديد المستوى المناسب للاختبارات والتقارير الكاملة عن النتائج تقديرات أحجام التأثير (مثل حجم تأثير كوهين)مؤشر بيرسون (r)، مما يدل على كيفية حسابها
البرمجيات والشيفرة
معلومات السياسة حول توفر كود الكمبيوتر
تعديل Attune NxT باستخدام برنامج Attune NxT الإصدار 4.2.0 FACSAria Fusion (BD) باستخدام برنامج FACS DIVA الإصدار 9.0.1 جهاز تحليل الخلايا BD LSR Fortessa X-20 باستخدام برنامج FACS DIVA الإصدار 9.0.1 البلوت الغربي: برنامج FusionCapt Advance
التصوير الفلوري: برنامج تصوير ديلتافيجن، برنامج استحواذ الصور ثاندر لايكا أو برنامج تصوير أبوتوم زيس تصوير برايتفيلد: برنامج تصوير cellSens Entry (أوليمبوس)
قارئ الألواح: قارئ الألواح SpectraMax ID3 (Molecular Devices)، تم تصديره كملفات Excel قارئ الميكرو بلايت SpectraMax iD5 مع برنامج SoftMax Pro v7 (Molecular Devices)
الرنين المغناطيسي النووي: برنامج Bruker Topspin 4.1.4 محفظة الطبيعة | ملخص التقرير أبريل 202
الجهد الدوري نوفا سوفتوير الفلوريمترية: سوفت ماكس برو 7.1 جي إكس بي
النمذجة الجزيئية: غاوسي 16، الإصدار C. 01
الليبيدوميكس: ليبيد إكسبلورر الإصدار 1.2.8 (http://genomebiology.com/2011/12/1/R8) اختبار FENIX: أجilent BioTek Gen5 الإصدار 3.08.01 تحليل البيانات البرمجيات: مايكروسوفت إكسل و PRISM 10.0.3 ما لم يُذكر خلاف ذلك
الرنين المغناطيسي النووي: ميست رينوفا v.5.0.1-35756 برنامج Bruker Topspin 4.1.4 تآزر الأدوية: برنامج SynergyFinder بالنسبة للمخطوطات التي تستخدم خوارزميات أو برامج مخصصة تكون مركزية في البحث ولكن لم يتم وصفها بعد في الأدبيات المنشورة، يجب أن تكون البرمجيات متاحة للمحررين والمراجعين. نحن نشجع بشدة على إيداع الشيفرة في مستودع مجتمعي (مثل GitHub). راجع إرشادات مجموعة Nature لتقديم الشيفرة والبرمجيات لمزيد من المعلومات.
بيانات
معلومات السياسة حول توفر البيانات يجب أن تتضمن جميع المخطوطات بيانًا حول توفر البيانات. يجب أن يوفر هذا البيان المعلومات التالية، حيثما ينطبق:
رموز الانضمام، معرفات فريدة، أو روابط ويب لمجموعات البيانات المتاحة للجمهور
وصف لأي قيود على توفر البيانات
بالنسبة لمجموعات البيانات السريرية أو بيانات الطرف الثالث، يرجى التأكد من أن البيان يتماشى مع سياستنا
تم تقديم بيانات الليبيدوميكس في الجداول التكميلية. تم إيداع بيانات البروتيوميات الكمية الخام في اتحاد بروتيوم إكس عبر مستودع شريك PRIDE مع معرف مجموعة البيانات PXD054449.مراجع_pxd054449@ebi.ac.uk; كلمة المرور: o1gh7kRYdTEP). لأغراض التعريف، تم البحث عن البيانات مقابل قاعدة بيانات يوني بروت (UP000005640) الخاصة بالإنسان العاقل باستخدام برنامج سبكترو نوت الإصدار 18.7 أو 19 (بيوجنوسيس).
البحث الذي يتضمن مشاركين بشريين، بياناتهم، أو مواد بيولوجية
معلومات السياسة حول الدراسات التي تشمل مشاركين بشريين أو بيانات بشرية. انظر أيضًا معلومات السياسة حول الجنس، الهوية/التقديم الجنسي، والتوجه الجنسي والعرق، والاثنية والعنصرية.
التقارير عن الجنس والنوع الاجتماعي
التقارير عن العرق أو الإثنية أو
لم يتم إجراء أي تقارير عن الجنس أو الجنس لعينات خزعة الأورام البشرية. تم جمع عينات الأورام بشكل مجهول لحماية المرضى والبيانات وفقًا لإرشادات المستشفى والقانون الفرنسي ولم يتم تحليلها حسب الجنس.
كانت عينات الأورام مجهولة الهوية ولم يتم الإبلاغ عن العرق أو الإثنية أو أي مجموعات اجتماعية ذات صلة وفقًا للقانون الفرنسي. اجتماعي ذو صلة تجمعات خصائص السكان
التوظيف
رقابة الأخلاقيات
تم الحصول على عينات مجهولة من سرطان القناة البنكرياسية الغدي (PDAC)، الساركوما غير المتمايزة متعددة الأشكال (UPS)، الساركوما الدهنية، الساركوما الوعائية، الساركوما الظهارية، و metastasis الكبدية لسرطان القناة البنكرياسية.
لا اختيار للمشاركين
تم الحصول على عينات جديدة من الأورام عن طريق الجراحة من مرضى في مراكز رعاية بول بروسي ومعهد كوري. قدم جميع المرضى موافقة خطية مستنيرة لاستخدام عينات الأورام. تمت الموافقة على الدراسة من قبل اللجان التنظيمية المؤسسية (رقم 587 و DATA190160).
يرجى ملاحظة أنه يجب أيضًا تقديم معلومات كاملة حول الموافقة على بروتوكول الدراسة في المخطوطة.
التقارير الخاصة بالمجال
يرجى اختيار الخيار أدناه الذي يناسب بحثك بشكل أفضل. إذا لم تكن متأكدًا، اقرأ الأقسام المناسبة قبل اتخاذ قرارك. علوم الحياة العلوم السلوكية والاجتماعية □ العلوم البيئية والتطورية والبيئية لنسخة مرجعية من الوثيقة مع جميع الأقسام، انظرnature.com/documents/nr-reporting-summary-flat.pdf
تصميم دراسة العلوم الحياتية
يجب على جميع الدراسات الإفصاح عن هذه النقاط حتى عندما يكون الإفصاح سلبياً.
حجم العينة
لم يتم إجراء حساب حجم العينة، ولكن الحد الأدنى منتم اختيارها لتجارب علم الأحياء الخلوية. بالنسبة للتجارب الحية، كانت مجموعات الفئران إما 5 أو 10. لم يتم إجراء حساب لحجم العينة لعينات الأورام البشرية، وتمت التحليلات بناءً على توفر هذه العينات النادرة بعد الجراحة، اعتمادًا على توفرها من المرضى.
استبعاد البيانات
لم يتم استبعاد أي بيانات من التحليلات.
استنساخ
تم إجراء التجارب ثلاث مرات على الأقل ما لم يُذكر خلاف ذلك. كانت عملية التكرار ناجحة وجميع البيانات معروضة في المخطوطة. جميع النسخ المكررة لجميع التجارب مذكورة في التعليقات التوضيحية و/أو التحليل الإحصائي، مع توفر البيانات كملفات مصدر.
العشوائية
للتجارب الحية، تم تخصيص الفئران عشوائيًا إلى مجموعات تجريبية. بالنسبة لعينات الورم، تم تجميعها حسب نوع الورم (PDAC، UPS، إلخ) ولم تكن عشوائية. كانت المواد متاحة من المستشفى وتم جمعها بطريقة مجهولة للتحليل، باستثناء الإشارة إلى نوع الورم.
مُعَمي
تمت إخفاء تخصيص المجموعات عن المحققين أثناء جمع البيانات لتجارب ICP-MS و lipidomics و proteomics. لم تكن تحليلات البيانات مخفية.
التقارير عن مواد وأنظمة وطرق محددة
نحتاج إلى معلومات من المؤلفين حول بعض أنواع المواد والأنظمة التجريبية والأساليب المستخدمة في العديد من الدراسات. هنا، يرجى الإشارة إلى ما إذا كانت كل مادة أو نظام أو طريقة مدرجة ذات صلة بدراستك. إذا لم تكن متأكدًا مما إذا كان عنصر القائمة ينطبق على بحثك، يرجى قراءة القسم المناسب قبل اختيار رد.
المواد والأنظمة التجريبية
طرق
غير متوفر
مشارك في الدراسة
غير متوفر
مشارك في الدراسة
□
– الأجسام المضادة
إكس
□
□
– خطوط خلايا حقيقية النواة
□
– قياس التدفق الخلوي
–
□
V
□
□
【 الحيوانات وغيرها من الكائنات الحية
إكس
□
–
□
إكس
□
الأجسام المضادة
الأجسام المضادة المستخدمة
تم توضيح الأجسام المضادة أدناه على النحو التالي. WB، التحليل الغربي؛ FCy، قياس التدفق الخلوي؛ FI، تصوير الفلورية. Hu، مستخدم لعينات بشرية. Ms، مستخدم لعينات فئران. يتم الإشارة إلى التخفيفات. أي تحقق من الأجسام المضادة من قبل الشركة المصنعة موضح ويمكن العثور عليه في مواقع الشركات المصنعة. استراتيجيات التحقق من الأجسام المضادة لدينا من خلال تقنيات تقليل التعبير (KD) و/أو حذف الجينات (KO) للأجسام المضادة ذات الصلة موضحة.
الأجسام المضادة الأولية: AIFM2/FSP1 (ميرك، MABC1638، النسخة 6D8-11، الدفعة Q3745998، WB، 1:500، Hu، تم التحقق من صحة KD داخلياً)، الكاتالاز
” CD31-PE-Cy7 (BioLegend, 303118, النسخة WM59، الدفعة B276836، FCy، 1:100، إنسان)، CD31-BV605 (BioLegend, 303122، النسخة WM59، الدفعة 331683، FCy، 1:100، إنسان)، CD31-BV605 (BioLegend, 102427، النسخة 390، الدفعة B375532، FCy، 1:100، فأر)، CD44 (Abcam، ab189524، النسخة EPR18668، الدفعة 1014086-32، WB، 1:30000، إنسان، تم التحقق من KO و KD داخليًا)، CD44-AF647 (Novus Biologicals، NB500-481AF488، النسخة MEM-263، الدفعة P158343، FCy، 1:100، إنسان)، CD44-AF647 (Novus Biologicals، NB500-481AF647، النسخة MEM-263، الدفعة D145771، FCy، 1:100، إنسان)، CD44-AF647 (BioLegend، 103018، النسخة IM7، الدفعة B317762، FCy، 1:100، فأر)، CD45-BV785 (BioLegend، 304048، النسخة HI30، الدفعة B339809، FCy، 1:100، إنسان)، CD45-BV510 (BioLegend، 368526، النسخة 2D1، الدفعة B373428، FCy، 1:100، إنسان)، CD45-BV510 (BioLegend، 103138، النسخة 30-F11، الدفعة B386738، FCy، 1:100، فأر)، CD163-PerCP/Cyanine5.5 (BioLegend، 326512، النسخة RM3/1، الدفعة B291202، FCy، 1:100، إنسان)، COXIV (Abcam، ab16056، الدفعة GR3206555-1، WB، 1:1000، إنسان)، Cytochrome c (Cell Signaling، 12963S، النسخة 6H2.B4، الدفعة 2، FI، 1:200، إنسان)، EEA1 (Abcam، ab70521، النسخة 1G11، الدفعة GR315680-1، FI، 1:200، إنسان، تم التحقق منه بواسطة ICC/IF من قبل الشركة المصنعة)، FAP-AF700 (R&D Systems، FAB3715N، النسخة 427819، الدفعة AEVIO20011، FCy، 1:100، إنسان)، FAP-AF750 (Bio-Techne، FAB3715S-100UG، النسخة 427819، الدفعة 1718688، FCy، 1:100، إنسان)، Ferritin (Abcam، ab75973، النسخة EPR300AY، الدفعة 10136442-29، WB، 1:1000، إنسان، تم التحقق منه بواسطة WB من قبل الشركة المصنعة)، Fibronectin (Abcam، ab45688، النسخة F14، الدفعة 1016266-35، WB، 1:1000، إنسان)، FTH1 (Santa Cruz Biotechnology، sc-376594، النسخة B-12، الدفعة G2622، WB، 1:200، إنسان)، GPX4 (Abcam، ab125066، النسخة EPNCIR144، الدفعة GR3369574-4، WB، 1:2000، إنسان، تم التحقق من KO من قبل الشركة المصنعة، KD تم التحقق منه داخليًا)، 4-Hydroxynonenal (4-HNE) (Abcam، ab48506، النسخة HNEJ-2، الدفعة 1062274-2، FI، 1:200، إنسان)، IRP2 (Novus Biologicals، NB100-1798، الدفعة D-4، WB، 1:1000، إنسان)، Lamin A/C (Cell Signaling، 2032S، الدفعة 6، WB، 1:1000، إنسان)، LAMP1 (Cell Signaling، 9091S، النسخة D2D11، الدفعة 7، FI، 1:200، إنسان)، LAMP1 (Santa Cruz Biotechnology، sc-20011، النسخة H4A3، FI، 1:100، فأر)، LAMP1 (Abcam، ab24170، الدفعة GR3235630-1، WB، 1:1000، إنسان)، LAMP2 (Abcam، ab25631، النسخة H4B4، الدفعة 1011336-1، FI، 1:200، إنسان)، LAMP2 (Thermo Fisher Scientific، MA1-205، النسخة
التحقق أي تحقق من الأجسام المضادة من قبل الشركة المصنعة موضح ويمكن العثور عليه في مواقع الشركات المصنعة. استراتيجيات التحقق من الأجسام المضادة لدينا (KD) و/أو (KO) التي تم تنفيذها في مختبرنا للأجسام المضادة ذات الصلة موضحة أعلاه وتم تنفيذها والتحقق منها في المختبر باستخدام طرق CRISPR أو siRNA ذات الصلة (المشار إليها على أنها داخلية في القائمة أعلاه). معلومات تحقق الأجسام المضادة من قبل الشركات المصنعة موضحة في القائمة أعلاه. كل هذه المعلومات مفصلة أيضًا في قسم الطرق.
خطوط خلايا حقيقية النواة
معلومات السياسة حول خطوط الخلايا والجنس والنوع في البحث
مصدر(s) خط الخلايا
المصادقة
تلوث الميكوبلازما
الخطوط التي يتم التعرف عليها بشكل خاطئ بشكل شائع (انظر سجل ICLAC)
تم الحصول على خلايا HT-1080 و MDA-MB-231 وخلايا 4T1 من ATCC. كانت خلايا FC1242 و FC1245 لسرطان البنكرياس الفأري، وخلايا 4a، وخلايا البنكرياس البشرية hMIA-2D هدية من مختبر توفيسون (CSHL). تم الحصول على خلايا البنكرياس البشرية الأولية PDAC024T و PDAC030T و PDAC053T و PDAC054T و PDAC084T و PDAC090T و PDAC211T كما هو موضح في فقرة “إنشاء ثقافات خلايا البنكرياس المستمدة من زراعة الأنسجة”. تم الحصول على الأعضاء العضوية المستمدة من زراعة الأنسجة (XDPO) PDAC009T و PDAC003T و PDAC117T و PDAC372T كما هو موضح في فقرة “إنشاء الأعضاء العضوية المستمدة من زراعة الأنسجة”. كانت خلايا الورم المنتشرة في الرئة الأساسية من شركة سيلبروجن. كانت خلايا الورم المنتشرة من القولون الأولي من شركة سيلبروجن. تم إنتاج خلايا Pfa1 في مختبر كونراد (Seiler A et al., 2008). كانت خلايا SUM159 هدية كريمة من الدكتور س. إيثير (مركز كارمانوس للسرطان، ديترويت، ميشيغان، الولايات المتحدة الأمريكية). تم الاعتماد على المصادقة من قبل المصدر التجاري حيثما كان ذلك مناسبًا (ATCC، Celprogen). لم يتم إجراء أي تحقق من خلايا سرطان البنكرياس المشتقة من الإنسان / الأعضاء العضوية. تم إنتاج هذه الخلايا من مصادر أولية مباشرة في مختبر لوفانا. تم إنتاج خلايا Pfa1 والمشتقات في مختبر كونراد (Seiler A et al., 2008) وكانت خلايا SUM159 هدية كريمة من الدكتور S. Ethier (مركز كارمانوس للسرطان، ديترويت، ميشيغان، الولايات المتحدة الأمريكية). لم يتم التحقق من هذه الخلايا، وكذلك الخلايا المهداه بسخاء من مختبر توفيسون (انظر أعلاه).
تم اختبار الخلايا بانتظام سلبياً لوجود تلوث بالميكوبلازما. لم يتم استخدام أي خطوط خلوية يتم التعرف عليها بشكل خاطئ بشكل شائع في هذه الدراسة.
الحيوانات وغيرها من الكائنات البحثية
معلومات السياسة حول الدراسات التي تشمل الحيوانات؛ إرشادات ARRIVE الموصى بها للإبلاغ عن أبحاث الحيوانات، والجنس والنوع في البحث
الحيوانات المخبرية
علاج cLip-1 في الجسم الحي: تم الاحتفاظ بالفئران تحت ظروف قياسية مع الماء والطعام بحرية وفي بيئة خاضعة للرقابةالرطوبة، دورة ضوء/ظلام لمدة 12 ساعة). بالنسبة للدراسات الحيوانية، تم توزيع فئران Rosa26-CreERT2;Gpx4f/f (C57BL6/J) عشوائيًا في أقفاص منفصلة. تم استخدام فئران متطابقة في الجنس تتراوح أعمارها بين 12 إلى 24 أسبوعًا في جميع التجارب.
عزل الدم والمصل والسائل اللمفاوي من الفئران: تم شراء فئران Balb/C (إناث بالغة تبلغ من العمر 25 أسبوعًا) من شركة تشارلز ريفر وتم تربيتها في منشأة الحيوانات الأساسية في CRCM وتم توزيعها عشوائيًا في أقفاص منفصلة. تم الاحتفاظ بالفئران في ظروف معقمة مع توفير الطعام والماء المعقمين بحرية، وتم الحفاظ عليها على دورة ضوء/ظلام مدتها 12 ساعة.الرطوبة.
نماذج انتشار الأورام في العقد اللمفاوية الفأرية وعلاج Fento-1/Marmycin/CW: تم زرع خلايا سرطان الثدي الفأرية (4T1) في إناث من فصيلة Balb/c تتراوح أعمارها بين 6 إلى 8 أسابيع (متجانسة مع نموذج 4T1). تم تربية الفئران في منشأة الحيوانات الأساسية بمدرسة هارفارد الطبية وتم توزيعها عشوائيًا في أقفاص منفصلة. تم الاحتفاظ بالفئران في ظروف معقمة مع توفير طعام وماء معقمين بحرية، وتم الحفاظ عليها على دورة ضوء/ظلام مدتها 12 ساعة.الرطوبة.
الحيوانات البرية
التقارير عن الجنس عينات تم جمعها من الميدان رقابة الأخلاقيات
لم تتضمن الدراسة حيوانات برية. كما هو موضح في الطرق.
لم تتضمن الدراسة عينات تم جمعها من الميدان.
تم إجراء جميع تجارب cLip-1 في الجسم الحي وفقًا لقانون الرفق بالحيوانات الألماني وتمت الموافقة عليها من قبل اللجنة المؤسسية لتجارب الحيوانات وحكومة بافاريا العليا (رقم الموافقة ROB-55.2-2532.Vet_02-18-13). جميع تجارب حقن العقد اللمفاوية في الفئران امتثلت لجميع اللوائح الأخلاقية ذات الصلة وتمت وفقًا للبروتوكولات المعتمدة من قبل لجنة رعاية واستخدام الحيوانات المؤسسية في مدرسة هارفارد تي. إتش. تشان للصحة العامة (البروتوكول ISOOOO3460). بالنسبة لجمع اللمف والدم من الفئران، تم إجراء تجارب الحيوانات وفقًا للإرشادات الأوروبية في رعاية واستخدام الحيوانات. تم إجراء تجارب الحيوانات بما يتماشى مع الإرشادات الفرنسية للتعامل مع الحيوانات وتمت الموافقة عليها من قبل اللجنة الأخلاقية المحلية (رقم الاتفاق 16487-2018082108541206 v3).
يرجى ملاحظة أنه يجب أيضًا تقديم معلومات كاملة حول الموافقة على بروتوكول الدراسة في المخطوطة.
النباتات
مخزونات البذور
تقرير عن مصدر جميع مخزونات البذور أو المواد النباتية الأخرى المستخدمة. إذا كان ذلك مناسبًا، يرجى ذكر مركز مخزون البذور ورقم الفهرس. إذا تم جمع عينات نباتية من الحقل، يرجى وصف موقع الجمع، التاريخ وإجراءات أخذ العينات.
أنماط جينية نباتية جديدة وصف الطرق التي تم من خلالها إنتاج جميع الأنماط الجينية النباتية الجديدة. يشمل ذلك تلك التي تم إنشاؤها من خلال الأساليب الوراثية، وتحرير الجينات، والطفرات المعتمدة على المواد الكيميائية/الإشعاع، والتهجين. بالنسبة للسلالات الوراثية، وصف طريقة التحويل، وعدد السلالات المستقلة التي تم تحليلها، والجيل الذي تم إجراء التجارب عليه. بالنسبة للسلالات المعدلة جينياً، وصف المحرر المستخدم، والتسلسل الداخلي المستهدف للتعديل، وتسلسل RNA الدليل المستهدف (إذا كان ذلك مناسباً) وكيف تم تطبيق المحرر.
المصادقة
تدفق الخلايا
المؤامرات
أكد أن:
توضح تسميات المحاور العلامة والفلوكروم المستخدم (مثل CD4-FITC). المقاييس على المحاور مرئية بوضوح. قم بتضمين الأرقام على المحاور فقط للرسم البياني في أسفل اليسار من المجموعة (المجموعة هي تحليل للعلامات المتطابقة). جميع الرسوم البيانية هي رسوم بيانية متساوية الارتفاع مع نقاط شاذة أو رسوم بيانية بالألوان الزائفة. تم توفير قيمة عددية لعدد الخلايا أو النسبة المئوية (مع الإحصائيات).
المنهجية
تحضير العينة
تدفق الخلايا:
تم غسل الخلايا بماء مثلجPBS. لتلوين الأجسام المضادة، تم حضانة الخلايا مع Fc block (Human TruStain FcX، Biolegend، 422302، 1:20) لمدة 15 دقيقة، ثم تم حضانتها مع الأجسام المضادة في درجة حرارة منخفضة.FBSPBS، 2 مللي مولار EDTA لمدة 20 دقيقة عند 4ثم تم غسله بـPBS وإعادة تعليقها فيFBS،PBS، 2 مللي مولار EDTA قبل التحليل باستخدام BD LSR Fortessa
جهاز قياس التدفق X-20. قياس التدفق الخلوي باستخدام مجسات الحديد: تم زراعة خلايا PDAC053T و HT-1080 في أطباق 6 آبار بكثافة خلايا/بئر. في اليوم التالي، ليب-1 الهيدروكسي كلوروكين (HCQ، سيغما-ألدريتش، )، تم إضافة بافيلوميسين A1 (Baf-A1، سيغما-ألدريتش، B1793، 75 نانومتر) إلى PDAC053T لمدة ساعة وإلى HT-1080 لمدة 30 دقيقة. في PDAC053T، بعد 30 دقيقة من العلاج بالمركبات، RPE (داخلي، ) أو HMRhoNox-M (داخلي، تمت إضافة المجسات لمدة 30 دقيقة. في HT-1080، بعد 15 دقيقة من العلاج بالمركبات، RPE (داخلي، ) أو HMRhoNox-M (داخلي، تم إضافة المجسات وتركها لمدة 15 دقيقة. بعد الحضانة مع المجسات الحديدية، تمت إزالة الوسط وتم غسل الخلايا بـتمت معالجة PBS قبل التربسين. تم جمع الخلايا، وتكويرها، وغسلها بـPBS وأخيرًامنمحلول PBS يحتوي على 10% FBS و EDTA ) تم إضافته. تم تسجيل البيانات على جهاز BD LSR Fortessa X-20. تم استخدام تقنية تحليل تدفق الخلايا مع Bodipy-C11 581/591: تم زراعة خلايا PDAC053T في أطباق 6 آبار بكثافة الخلايا/بئر. في اليوم التالي، تم معالجة الخلايا بـ Lip-1، cLip-1 (داخلي، ، ميتكليب-1 (داخلي، ), alcLip-1 (في المنزل، 10 وبوديبي-C11قبل إضافة RSL3بعد ساعة واحدة، تم إزالة الوسط وتم غسل الخلايا بـتمت معالجة PBS مرتين قبل التربسين. تم جمع الخلايا، وتكويرها، وغسلها بـPBS وأخيرًامنمحلول PBS يحتوي على 10% FBS و EDTA )، تم إضافته لتحليل تدفق الخلايا. تم زراعة خلايا PDAC053T و HT-1080 و 4T1 في أطباق 6 آبار بكثافة الخلايا/بئر. في اليوم التالي، تم معالجة الخلايا بـ Baf-A1 (75 نانومتر) و HCQلمدة ساعتين قبل إضافة RSL3 (200 نانومتر لـ PDAC053T و HT-1080، 500 نانومتر لـ 4T1). ثم بعد ساعة، تم معالجة الخلايا بـ Bodipy-C11.لمدة ساعة إضافية. تم إزالة الوسط وتم غسل الخلايا بـتمت معالجة PBS مرتين قبل التربسين. تم جمع الخلايا، وتكويرها، وغسلها بـPBS وأخيرًامنمحلول PBS يحتوي على FBS و EDTA )، تم إضافته لتحليل تدفق الخلايا. تم تسجيل البيانات على جهاز تحليل تدفق الخلايا AttuneTM NxT (ثيرمو فيشر ساينتيفيك). تم إجراء تحليلات تدفق الخلايا لمستويات GSH في الجسيمات الحالة والجذور الحرة الهيدroxيلية في الجسيمات الحالة: تم حضانة الخلايا مع SQSS (داخلية، ) أو 1-أحمر (في المنزل، تم تحضين خلايا HT-1080 مع RSL3 ، سيغما-ألدريتش، SML0521)، FIN56 (سيغما-ألدريش، SML1740)، بيوثيونين سلفوكسيمين (BSO،سيغما-ألدريتش، B2515) أو إيراستين، سيغما-ألدريتش، 329600) للفترات الزمنية المحددة. تم تسجيل البيانات على جهاز تحليل الخلايا بالتدفق AttuneTM NxT (ثيرمو فيشر ساينتيفيك). تحليلات الخلايا بالتدفق لمحتوى الحديد في الليزوزومات: عادةًتم حضن خلايا الأنسجة البشرية المفصولة في وسط (RPMI 1610، 10% FBS، بنسلين/ستربتوميسين) مع HMRhoNox-M (داخلي، 1تم تحليل محتوى الحديد الليزوزومي في عينات الأورام البشرية والأنسجة السليمة المجاورة باستخدام لوحة الأجسام المضادة والصبغات التالية: DAPI ( ), CD3-BV510 (BioLegend, 317332)، CD31-PE-Cy7 (BioLegend, 303118)، CD44AF647 (Novus Biologicals, NB500-481AF647)، CD45-BV785 (BioLegend, 304048)، CD163-PerCP/Cyanine5.5 (BioLegend, 326512)، FAP-AF700 (R&D Systems, FAB3715N)، TfR1-APC-AF750 (Beckman Coulter, A89313). كانت خلايا الورم الحية تت correspond إلى خلايا DAPIneg/CD45neg/CD31neg/FAPneg. تم تسجيل البيانات على جهاز BD LSRFortessa X-20. تحليلات تدفق الخلايا لـ CD44 في عينات الورم المعالجة بـ Fento-1: عادةً تم تحضين خلايا الورم المفصولة في وسط (RPMI 1610، 10% FBS، بنسلين/ستربتوميسين) مع Fento-1 (داخل المنزل،تم معالجة الخلايا مسبقًا بـ Lip-1، cLip-1 (داخلي، )، فهرس أو تعريف لمدة ساعتين. تم الحفاظ على Toc نقياً تحت جو خامل وتم إعداد محلول مخزون جديد طوال الدراسة قبل كل تجربة. تم استخدام لوحة الأجسام المضادة والصبغة التالية للتحليل التدفق اللاحق: SYTOX الأزرق (ثيرمو فيشر ساينتيفيك، S34857، ), CD31-BV605 (BioLegend, 303122)، CD45BV510 (BioLegend, 368526، الدفعة B373428)، CD44-AF647 (Novus Biologicals، NB500-481AF647)، TfR1-PE (BioLegend، 334106)، FAP-AF750 (Novus Biologicals، FAB3715S-100UG). كانت خلايا الورم الحية تت correspond إلى خلايا SYTOX زرقاء/ CD45 سلبية/ CD31 سلبية/ FAP سلبية. تم تسجيل البيانات على جهاز تحليل التدفق الخلوي AttuneTM NxT (Thermo Fisher Scientific). لتحليلات التدفق الخلوي لمستويات CD44 في أورام الفئران 4T1، عادةً تم استخدام خلايا الورم المفصولة لكل حالة. تم صبغ الخلايا المفصولة حديثًا باستخدام الأجسام المضادة ولوحة الصبغة التالية: SYTOX الأزرق (ثيرمو فيشر ساينتيفيك، S34857، 1CD31-BV605 (BioLegend, 102427)، CD44-AF647 (BioLegend, 103018)، CD45-BV510 (BioLegend, 103138)، MHCII-APC/ Cyanine7 (BioLegend, 107628). كانت خلايا الورم الحية تت corresponded إلى خلايا SYTOX blueneg/CD45neg/CD31neg/MCH IIpos. تم تسجيل البيانات على جهاز تحليل التدفق AttuneTM NxT (Thermo Fisher Scientific). تم تحليل جميع البيانات باستخدام برنامج FlowJo الإصدار 10.10.0.
فرز الخلايا المعتمد على الفلورية: تم فرز خلايا بشرية باستخدام الأجسام المضادة التالية: CD31-PE-Cy7 (BioLegend, 303118)، CD44-AF647 (Novus Biologicals, NB500-481AF647)، CD45-BV785 (BioLegend, 304048). كانت الخلايا المفروزة تت correspond إلى خلايا CD45neg/CD31neg/CD44pos وخلايا CD45neg/CD31neg/CD44neg وتم عزلها على جهاز FACSAria Fusion (BD). تم إجراء تجارب ICP-MS على خلايا الأورام CD44high و CD44low كما هو موضح في قسم ICP-MS. تم فرز خلايا الفئران باستخدام الأجسام المضادة التالية: CD44-AF647 (Biolegend, 103018)، MHCII-APC/Cyanine7 (BioLegend, 107628). كانت الخلايا المفروزة تت correspond إلى خلايا MHCIIpos/CD44high وخلايا MHCIIpos/CD44low. تم طرد الخلايا المفروزة في جهاز الطرد المركزي عندوتم معالجة كريات الخلايا للتطبيقات اللاحقة.
أجهزة تحليل التدفق الخلوي: BD LSRFortessa X-20، Thermo Fisher Attune NxT و BD FACSAria Fusion sorter.
برنامج FlowJo الإصدار 10.10.0
فرز الخلايا يسمح بالحصول على نقاء مرضٍ
كانت خلايا الورم البشري الحية تت correspond إلى خلايا DAPIneg/CD45neg/CD31neg/FAPneg. وكانت خلايا الورم الفأري الحية تت correspond إلى خلايا SYTOX blueneg/CD45neg/CD31neg/MCH IIpos. كانت الخلايا البشرية المفروزة تت correspond إلى خلايا CD45neg/CD31neg/CD44pos وخلايا CD45neg/CD31neg/CD44neg. وكانت الخلايا الفأرية المفروزة تت correspond إلى خلايا MHCllpos/CD44pos وخلايا MHCllpos/CD44neg.
قم بتحديد هذا المربع لتأكيد أنه تم تقديم رقم يوضح استراتيجية البوابة في المعلومات التكميلية.
معهد كوري، CNRS، INSERM، جامعة PSL للبحث، باريس، فرنسا.قسم الأيض الجزيئي، مدرسة هارفارد تي. إتش. تشان للصحة العامة، بوسطن، ماساتشوستس، الولايات المتحدة الأمريكية.قسم الوراثة، معهد كوري، باريس، فرنسا.جامعة باريس ساكلاي، UVSQ، مونتيني-لي-بريتونيو، فرنسا.معهد الأيض وموت الخلايا، مركز هيلمهولتز ميونيخ، نيوهيربرغ، ألمانيا.معهد كوري، INSERM، معهد باريس للتكنولوجيا، جامعة PSL للبحث، باريس، فرنسا.معهد كوري، جامعة PSL للبحث، باريس، فرنسا.المدرسة العليا للأساتذة، المركز الوطني للبحث العلمي، جامعة PSL للبحث، باريس، فرنسا.CRCM، CNRS، INSERM، معهد باولي-كالمات، جامعة إكس-مارسيليا، مارسيليا، فرنسا.CRCM، CNRS، INSERM، معهد باولي-كالمت، جامعة إكس-مارسيليا، الحديقة العلمية والتكنولوجية في ليميني، مارسيليا، فرنسا.قسم الجهاز الهضمي وعلم الأورام الطبية، جامعة باريس ساكلاي ومستشفى بول بروسا (APHP سود)، فيليجويف، فرنسا.قسم الأورام الجراحية، معهد كوري، باريس، فرنسا.قسم الأورام الطبية، معهد كوري، إنسرم، جامعة بي إس إل للبحث، باريس، فرنسا.معهد الكيمياء الجزيئية ومواد أورساي، CNRS، جامعة باريس ساكلاي، أورساي، فرنسا.قسم الكيمياء والعلوم الجزيئية الحيوية، جامعة أوتاوا، أوتاوا، أونتاريو، كندا. مركز رودولف فيرشو، مركز التصوير الحيوي التكاملي والترجماتي، جامعة يوليوس ماكسيميليان في وورزبورغ، وورزبورغ، ألمانيا.قسم الكيمياء، جامعة كولومبيا، نيويورك، نيويورك، الولايات المتحدة الأمريكية.قسم العلوم البيولوجية، جامعة كولومبيا، نيويورك، نيويورك، الولايات المتحدة الأمريكية.قسم علم الأمراض وعلم الأحياء الخلوية، كلية فاجيلوس للأطباء والجراحين، جامعة كولومبيا، نيويورك، نيويورك، الولايات المتحدة الأمريكية.علم الأحياء الأكسدة والاختزال الانتقالي، مدرسة العلوم الطبيعية بجامعة تكنولوجيا ميونيخ، غارشينغ، ألمانيا.ساهم هؤلاء المؤلفون بالتساوي: تاتيانا كانيكي، ليروي بارون، سيباستيان مولر.البريد الإلكتروني: raphael.rodriguez@curie.fr
Tatiana Cañeque , Leeroy Baron , Sebastian Müller , Alanis Carmona , Ludovic Colombeau , Antoine Versini , Stéphanie Solier , Christine Gaillet , Fabien Sindikubwabo , Julio L. Sampaio , Marie Sabatier , Eikan Mishima , Armel Picard-Bernes¹, Laurène Syx , Nicolas Servant , Bérangère Lombard , Damarys Loew , Jiashuo Zheng , Bettina Proneth , Leishemba K. Thoidingjam , Laurence Grimaud , Cameron S. Fraser , Krystina J. Szylo , Emma Der Kazarian , Caroline Bonnet , Emmanuelle Charafe-Jauffret , Christophe Ginestier , Patricia Santofimia-Castaño , Matias Estaras , Nelson Dusetti , Juan Lucio Iovanna , Antonio Sa Cunha , Gabriella Pittau , Pascal Hammel , Dimitri Tzanis , Sylvie Bonvalot , Sarah Watson , Vincent Gandon , Aditya Upadhyay , Derek A. Pratt , Florêncio Porto Freitas , José Pedro Friedmann Angeli , Brent R. Stockwell , Marcus Conrad , Jessalyn M. Ubellacker & Raphaël Rodriguez
Iron catalyses the oxidation of lipids in biological membranes and promotes a form of cell death called ferroptosis . Defining where this chemistry occurs in the cell can inform the design of drugs capable of inducing or inhibiting ferroptosis in various diseaserelevant settings. Genetic approaches have revealed suppressors of ferroptosis ; by contrast, small molecules can provide spatiotemporal control of the chemistry at work . Here we show that the ferroptosis inhibitor liproxstatin-1 exerts cytoprotective effects by inactivating iron in lysosomes. We also show that the ferroptosis inducer RSL3 initiates membrane lipid oxidation in lysosomes. We designed a small-molecule activator of lysosomal iron-fentomycin-1-to induce the oxidative degradation of phospholipids and ultimately ferroptosis. Fentomycin-1 is able to kill iron-rich CD44 primary sarcoma and pancreatic ductal adenocarcinoma cells, which can promote metastasis and fuel drug tolerance. In such cells, iron regulates cell adaptation while conferring vulnerability to ferroptosis . Sarcoma cells exposed to sublethal doses of fentomycin-1 acquire a ferroptosis-resistant cell state characterized by the downregulation of mesenchymal markers and the activation of a membranedamage response. This phospholipid degrader can eradicate drug-tolerant persister cancer cells in vitro and reduces intranodal tumour growth in a mouse model of breast cancer metastasis. Together, these results show that control of iron reactivity confers therapeutic benefits, establish lysosomal iron as a druggable target and highlight the value of targeting cell states .
Iron reacts with peroxides to produce oxygen-centred radicals. In the cell, such a process can initiate and propagate a radical chain reaction that leads to oxidized organic products, a chemistry broadly defined as the Fenton reaction . Chemically reactive lipids in biological membranes are ideal substrates for such reactions and have been implicated in ferroptosis . The accumulation of damaged phospholipids can eventually cause the loss of membrane integrity together with altered
organelle functions and the release of various types of chemical entities in the cell, which can induce further cellular defects and cell death. Ferroptosis involves distinct organelles , including peroxisomes , mitochondria , the endoplasmic reticulum (ER) and lysosomes . However, it is currently unclear whether individual organelles contribute to ferroptosis through impaired cell signalling, metabolism and biosynthesis of specific biomolecules or whether membrane lipids
of these compartments are also direct substrates for iron-mediated oxidation that leads to cell death. Cancer cells have been shown to require functional lysosomes to undergo ferroptosis . This finding highlights the importance of cellular iron import and lends support for the idea that iron has a role in promoting cell death. Pharmacological inhibition of iron trafficking from lysosomes promotes the oxidation of lysosomal membrane lipids , which indicates that a redox-active pool of iron in this organelle can trigger ferroptosis. Synthetic endoperoxides, surrogates of peroxidized lipids with a propensity to accumulate in the ER, can induce ferroptosis, which suggests that lipid peroxidation can propagate through this organelle . However, it is currently unclear where iron-mediated oxidation of membrane lipids is first initiated in the cell before propagating through other proximal membrane lipids.
Lysosomal iron initiates ferroptosis
We set out to identify the subcellular sites of action of liproxstatin-1 (Lip-1), a small molecule that protects cells against death induced through the inactivation of ferroptosis suppressors . We labelled a clickable alkyne-containing analogue of Lip-1 (named cLip-1) using in-cell click chemistry and performed fluorescence microscopy in various cell lines. The results revealed lysosomal accumulation in HT-1080 fibrosarcoma cells, in primary cells of human pancreatic ductal adenocarcinoma (PDAC), in lung and colon circulating tumour cells and in 4T1 mouse breast cancer cells (Fig. 1a and Extended Data Fig. 1a-f). Notably, labelled cLip-1 was detected only in the endolysosomal compartment, even at a concentration higher than the effective dose. This result supports the notion that Lip-1 exerts its anti-ferroptotic activity in this organelle. In vivo, cLip-1 increased the survival of mice in which acute renal failure was induced through genetic deletion of , which encodes the ferroptosis suppressor glutathione peroxidase (Fig. 1b). Labelling cLip-1 in mouse tissues revealed its accumulation in the kidney, colocalizing with a lysosomal marker in renal proximal tubules (Fig. 1c and Extended Data Fig. 1g). In vitro, cLip-1 prevented the oxidation of membrane lipids and protected cells against the effects of genetic depletion of Gpx4 or pharmacological inhibition of GPX4 with RSL3, thereby recapitulating the biological activity of Lip-1 (Extended Data Fig. 1h-j). These data validate cLip-1 as a suitable surrogate of Lip-1 to investigate ferroptosis.
The tight iron chelator deferoxamine (DFO) protects cells against erastin-induced ferroptosis .By contrast, Lip-1exhibits radical-trapping antioxidant (RTA) properties in vitro . However, unlike most RTAs that inhibit ferroptosis, such as -tocopherol, Lip-1 does not follow the general trend of many ferroptosis suppressors, whereby RTA activity is closely correlated with ferroptosis inhibition . This result suggests that other properties of Lip-1 contribute to its anti-ferroptotic effects. In cancer cells, iron can be internalized by endocytosis and regulates cell-state transitions and cell proliferation . Because cLip-1 accumulates in lysosomes and contains nitrogen atoms susceptible to interactions with iron, we investigated the possibility that its ferroptosis inhibition properties can result from iron inactivation in this organelle. Nuclear magnetic resonance (NMR) spectroscopy indicated that Lip-1 interacts with iron(III) with stoichiometries of 1:1 and 2:1(Fig.1d). Titrating Lip-1 with iron(III) chloride induced signal broadening together with a shift downfield of signals of the protons adjacent to nitrogen atoms of Lip-1, which indicated the occurrence of coordination with the nitrogen lone pairs with the Lewis acid. By contrast, the proton signals of naphthalene, which was used as an internal control as it lacks heteroatoms, remained unaffected (Extended Data Fig. 1k). Moreover, adding a Brønsted acid to Lip-1induced distinct shifts downfield of the proton signals, which indicated protonation of the nitrogen atoms (Fig.1e). Notably, line broadening of the proton signals was observed only in the presence of the paramagnetic metal (Extended Data Fig. 11-n). Furthermore, the addition of trifluoroacetic acid (TFA) to a solution of iron(III) and Lip-1 did not fully displace iron from Lip-1, as shown by
chemical shifts that recapitulated the effects of Lewis and Brønsted acids. These data indicate that the binding of Lip-1 to iron can take place under acidic conditions, such as those found in lysosomes. The addition of sodium hydroxide restored the chemical shifts of unbound Lip-1, a result that demonstrates the reversible nature of Lip-1 interactions with iron (Extended Data Fig. 1o). Molecular modelling using discrete Fourier transform computation produced several possible binding modes between iron and Lip-1, including bimolecular tetrahedral iron complexes involving the piperidine, amidine and aniline functions of Lip-1 (Fig. 1f and Supplementary Information).
Cyclic voltammetry, which is used to quantify the redox potential of metals in solution , indicated that Lip-1 and DFO alter the redox properties of iron (Fig. 1g). As controls, we investigated two other synthetic analogues of Lip-1: metcLip-1 and alcLip-1. In metcLip-1, aromatic amines are methylated, which adversely affected RTA activity. By contrast, alcLip-1is an aliphatic analogue with amines that have a higher negative logarithmic acid dissociation constant ( ) value compared with their aromatic counterparts. This analogue was expected to be fully protonated at neutral pH and therefore to exhibit a reduced propensity to enter cells and to accumulate in lysosomes (Extended Data Fig. 2a). Labelling of metcLip-1 in cells revealed a similar staining pattern to that of labelled cLip-1, showing colocalization with a lysosomal marker. By contrast, labelled alcLip-1 showed weak fluorescence (Extended Data Fig. 2b). Cyclic voltammetry results further indicated that metcLip-1 altered the redox potential of iron to a greater extent than alcLip-1 (Extended Data Fig. 2c). Consistently, metcLip-1 protected cells against RSL3-induced oxidation of membrane lipids more potently than alcLip-1 (Extended Data Fig. 2d), and neither metcLip-1 nor alcLip-1 exhibited RTA properties in vitro (Extended Data Fig. 2e). In-cell labelling of an alkyne-containing derivative of DFO (cDFO) also revealed lysosomal signals, which provided support for the pro-ferroptotic activity of iron in this organelle (Extended Data Fig. 2f). Signals of cDFO were also found in the cell nucleus, which partially explains the toxicity of this iron chelator and its distinct associated phenotype compared with other iron-interacting small molecules, including Lip-1.
In a cell-free system, Lip-1 reduced the fluorescence of an iron(III)specific fluorescent probe in the presence of iron(III) (Extended Data Fig. 2g). Treatment of cells with hydroxychloroquine or bafilomycin-A1, which increase the lysosomal pH and prevent iron(III) unloading from its endocytic carriers, led to reduced pools of chelatable cellular iron(III) and reactive lysosomal iron(II), as measured using specific fluorescent probes . These treatments also protected cells against RSL3-induced oxidation of membrane lipids in cancer cells (Fig. 1h-j and Extended Data Fig. ). Comparable results were obtained with Lip-1, which was consistent with the direct interactions of this compound with lysosomal iron (Fig. 1h-jand Extended Data Fig. 2h-j). After treatment with RSL3 for 1 h , oxidation of membrane lipids was predominantly detected in lysosomes, as assessed through Liperfluo and Bodipy 665/676 fluorescence levels (Fig. 1k and Extended Data Figs. 2k and 3a). By contrast, treatment with RSL3 for 4 h led to a staining pattern of oxidized membrane lipids that colocalized mainly with a fluorescently labelled biological marker of the ER (Extended Data Fig. 3b). Our data suggest that initiation of the radical chain reaction takes place early in the endolysosomal compartment, where redox-active iron can be found . This radical chain reaction can then propagate to membrane lipids of other organelles in the vicinity, including the ER. In support of this hypothesis, treatment of cells with well-established ferroptosis inducers led to a depletion of glutathione and increased levels of oxygen-centred radicals in lysosomes (Extended Data Fig. 3c-e). Together, these data indicate that lysosomal iron has a central role in the induction of ferroptosis.
Iron activation promotes ferroptosis
Cancer cells that have acquired a drug-tolerant persister (DTP) cell state are vulnerable to ferroptosis . DTP cancer cells are characterized
Fig. 1 | Lysosomal iron triggers the oxidation of membrane lipids. a, Schematic (left), images (middle) and quantification (right) of labelling cLip-1 in cells. independent experiments in HT-1080 cells. Data are the mean ± s.d. Asc, ascorbate; Lys., lysosome; PFA, paraformaldehyde. b, Kaplan-Meier survival curves of Rosa26-CreERT2;Gpx4 mice treated with cLip-1 per kg per day by intraperitoneal injection; mice per group). Mantel-Cox log-rank test. Arrows indicate the timing of the two tamoxifen injections. Average days of survival are indicated. c, Fluorescence images of labelled cLip-1 in renal proximal tubules of a Rosa26-CreERT2;Gpx4 mouse 7 days after tamoxifen treatment. Images representative of mice. d, NMR spectra of Lip-1 titrated with (red, 2:1 Lip-1 to iron; green, 1:1 Lip-1 to iron). e, NMR spectra of Lip-1 titrated with and then TFA. The blue reference spectrum is Lip-1 + 3 eq. TFA.f, Putative binding modes of Lip-1 and iron obtained from molecular modelling.g, Cyclic voltammetry of a solution with Lip-1 (left) or DFO (right) (reduction potentials indicated by pink arrows). SCE, saturated calomel electrode.h-k, Data are for HT-1080 cells.h, Flow cytometry of cells
treated with the indicated compounds for 15 min and then with the iron(III) probe RPE for independent experiments. a.u., arbitrary unit; MFI, mean fluorescence intensity. i, Flow cytometry of cells treated with the indicated compounds for 15 min and then with the iron(II) probe HMRhoNox-M for 15 min . independent experiments.j, Bodipy-C11581/591flow cytometry of cells treated with RSL3 ( 1 h ) and with bafilomycin-A1 (Baf-A1) or hydroxychloroquine (HCQ) 2 h before. independent experiments. One-way analysis of variance (ANOVA). k, Fluorescence imaging of Liperfluo in cells treated with RSL3 . independent experiments. Two-sided unpaired -test. Data are the mean ± s.d.d,e, NMR spectra recorded at 310 K in methanol- . For and , values are from two-sided unpaired -test compared with vehicle. The following concentrations were used: h, i, Lip-1 , Baf-A1 . Box plots show the interquartile range, with centre lines indicating the medians and whiskers indicating the minimum and maximum values. Scale bar, .
by high expression of the iron-uptake protein CD44, a cell membrane glycoprotein that is internalized by endocytosis and is used as a marker of cancer cell stemness and metastasis . Thus, these cells exhibit
upregulated iron uptake, which leads to increased cellular iron levels that in turn promote the activity of iron-dependent demethylases . This process activates specific epigenetic and transcriptional programs
that underlie cell-state transitions and the acquisition of a DTP phenotype . Thus, lysosomal iron has emerged as a druggable target to promote ferroptosis in a cell-state-dependent manner . With this in mind and the knowledge that lysosomal iron can trigger the oxidation of membrane lipids , we designed a small molecule to target lipids at the plasma membrane and to activate iron(II) in lysosomes upon endocytosis. Such a compound would exploit nearby reactive membrane lipids as substrates for oxidation. To this end, we synthesized a bifunctional compound composed of the fluorescent lipophilic natural product marmycin and the synthetic Chen-White ligand, which we named fentomycin-1 (Fento-1) (Fig. 2a and Extended Data Fig. 4a-e). Marmycin accumulates at the plasma membrane and is internalized by endocytosis , whereas the Chen-White ligand is commonly used in chemical synthesis to oxidize organic substrates by activating iron(II) . In the presence of hydrogen peroxide and under mild acidic aqueous conditions, such as those found in lysosomes, the Chen-White iron catalyst is thought to form a reactive iron-oxo species . Like hydroxyl and hydroperoxyl radicals, this intermediate can abstract a hydrogen atom from organic substrates, including saturated ones, to produce reactive carbon-centred radicals and subsequently oxidation products. Fento-1 is therefore reminiscent of a bifunctional small molecule that is able to induce proximity de novo between a catalyst and nucleic acids or proteins to promote degradation . We proposed that by exploiting the abundant amount of reactive iron(II) in DTP cancer cells , such a bifunctional molecule would form an active catalyst in the cell to promote the oxidative degradation of lysosomal membrane lipids and ultimately trigger ferroptosis.
In a cell-free system, Fento-1 enhanced the iron-induced oxidation of a liposome-forming unsaturated phospholipid under experimental conditions comparable to that found in lysosomes. That is, acidic pH , the presence of hydrogen peroxide and a water-soluble iron(II) salt (Fig. 2b and Supplementary Table 1). Notably, iron promoted oxidation of the phospholipid bilayer in the presence of hydrogen peroxide without enzymes, which therefore supports the idea that such a process occurs in the cell. The intrinsic fluorescence of Fento-1 revealed its localization at the plasma membrane when experiments were conducted at a low temperature to reduce endocytic flux (Extended Data Fig. 5a), thereby recapitulating the photophysical properties of the parental natural product. By contrast, at a physiological temperature, which enables endocytosis to take place, Fento-1 and marmycin were detected in the endolysosomal compartment (Fig. 2c). Chemical labelling of a clickable alkyne-containing Chen-White ligand (cCW) revealed weak pan-cellular staining (Fig. 2c and Extended Data Fig. 4f), which suggested that the untethered ligand does not specifically accumulate in the endolysosomal compartment. Moreover, Fento-1 did not predominantly target other organelles (Extended Data Fig. 5b). This small molecule induced the oxidation of membrane phospholipids and reduced cell viability in several human and mouse cancer cell lines and primary cells, whereas the effects of cCW and marmycin were marginal (Extended Data Fig. 5c-e). Mass-spectrometry (MS)-based lipidomics revealed that Fento-1 induced the oxidation of phospholipids in several cell lines to a greater extent than well-established ferroptosis inducers under these conditions (Extended Data Fig. 5f,g, Supplementary Information and Supplementary Tables 2 and 3). Analyses of lysosomes isolated from HT-1080 cells treated with Fento-1 for 1 h confirmed the oxidation of associated phospholipids at this early time point (Extended Data Fig. 5h, Supplementary Information and Supplementary Table 4). Consistent with the oxidation of membrane phospholipids, sublethal doses of Fento-1 increased levels of ferroptosis suppressors in HT-1080 cells, including GPX4, ferroptosis suppressor protein 1 (FSP1) and the cystine-glutamate antiporter (composed of SLC7A11 and SLC3A2) (Fig. 2d and Extended Data Fig. 5i). Quantitative proteomics revealed that cells treated with sublethal doses of Fento-1 for 48 h promoted the upregulation of ferroptosis and lysosome-associated proteins, including ferroptosis suppressors and lysosome-associated membrane
proteins (LAMPs), respectively, a result that provides support for the role of lysosomes in ferroptosis induction (Fig. 2d,e and Supplementary Tables 5 and 6). Cells were also characterized by the downregulation of iron homeostasis-associated proteins, including the iron-uptake protein CD44 and the iron-storage protein ferritin. These cells also exhibited increased levels of transferrin receptor 1(TFR1) and iron-responsive element-binding protein 2 (IRP2), which are characteristic of cellular iron depletion . Sublethal doses of Fento-1 promoted the downregulation of mesenchymal markers, which have been linked to the acquisition of cancer cell states refractory to standard-of-care treatments . Last, these cells exhibited upregulation of other transcription factors involved in the regulation of cell states, such as SOX4 and SOX13, as well as epithelial markers. These data indicate that the effects of Fento-1 treatment on HT-1080 cells, which are of mesenchymal origin, are due to the acquisition of a distinct cell state that resembles that of epithelial cells typically sensitive to standard antiproliferative treatments. This result also suggests that cancer cells can adapt to Fento-1 exposure and develop a ferroptosis-tolerant cell state. That is, cells treated with a low dose of Fento-1 exhibit sustained viability and a distinct phenotype due to cellular adaptation rather than clonal selection . Western blot analyses further corroborated the proteomic data (Extended Data Fig. 5j). Furthermore, increased levels of GPX4 were detected in lysosome-enriched fractions isolated from HT-1080 cells treated with Fento-1 (Fig. 2f).
Oxidation of membrane phospholipids induced by Fento- 1 was reduced by the lipophilic RTA -tocopherol, the iron chelator deferiprone and Lip-1 (Fig. 2g, Extended Data Fig. 5k, Supplementary Information and Supplementary Table 7). Furthermore, treatment with Fento-1 induced the production of 4-hydroxynonenal (4-HNE) (Extended Data Fig. 51), a result indicative of peroxidation and the breakdown of phospholipids that contain polyunsaturated fatty acids . Notably, 4 -HNE can cause cellular damage, and its production is a hallmark of ferroptosis . Treatment with Fento-1 also led to increased levels of lysophospholipids and glycerol, further indicating the degradation of oxidized phospholipids (Fig. 2h, Extended Data Fig. 5m, Supplementary Information and Supplementary Table 8). This result, together with the upregulation of lipid metabolism after treatment with Fento-1 (Fig. 2e), suggests that oxidized membrane lipids trigger a membrane-damage response . Cell death induced by Fento- 1 was antagonized by well-established ferroptosis inhibitors, which included iron chelators and RTAs, but not by apoptosis or necroptosis inhibitors (Fig.2i and Extended Data Fig. 6a-f). Notably, Fento-1 exhibited a residual toxicity that ferroptosis inhibitors could not fully overcome, which reflects the potency of Fento-1. -Tocopherol did not prevent lysosome targeting with Fento-1, which indicated that this ferroptosis inhibitor exerts its protective activity by acting as a RTA in membrane lipids and not by competing for interaction at the plasma membrane (Extended Data Fig. 6g). Fento-1 was synthetically lethal after cotreatment with GPX4 inhibitors, and cell viability was rescued by Lip-1 (Extended Data Fig. 6h), a result that further confirmed the pro-ferroptotic activity of Fento-1. Finally, knocking down ferroptosis suppressors sensitized cells to Fento-1, whereas knocking down Cd44 conferred a protective effect (Extended Data Fig. 6i). Consistently, knocking down Cd44 partially protected cells against oxidation of membrane lipids, whereas genetic suppression of the canonical iron endocytosis pathway involving TFR1 was less effective (Extended Data Fig. 6j). This result confirms the role of CD44 as the main effector of iron uptake in the mesenchymal state of cancer cells .
We next explored the activity of structural variants of Fento-1 (Extended Data Fig. 7a). Fento-2, which contains a longer linker between marmycin and the Chen-White ligand, exhibited a lower potency against PDAC and sarcoma cells than Fento-1. This result supports the notion that Fento-1 exerts its activity by promoting proximity between a reactive iron catalyst and membrane lipids (Extended Data Fig. 7b,c).
Fig. 2 | Induction of ferroptosis by a lysosome-selective iron-dependent phospholipid degrader. a, Chemical synthesis of Fento-1 and redox-active iron catalyst formation. b, Top, illustration of a liposome loaded with iron(II) and . Bottom, lipidomics of oxidized DOPC-forming liposomes with iron(II) and Fento-1.n = 3 independent experiments. c-i, Data are for HT-1080 cells. c, Fluorescence imaging of Fento-1, marmycin and labelled cCW in cells treated at for 1 h . Scale bar, independent experiments. Data are the mean ± s.d.d, Quantitative proteomics of cells treated with Fento-1 . The dashed vertical line indicates adjusted independent experiments. FC, fold change; TFs, transcription factors. e, KEGG enrichment analysis of upregulated proteins in cells treated with Fento-1 independent experiments.f, Western blot of proteins from total and lysosome-enriched extracts from cells treated with Fento-1(48 h). Representative of independent experiments. -Tubulin is a sampleprocessing control.g, Lipidomics of oxidized phospholipids in cells treated with Fento-1 and inhibitors (added 2 h before Fento-1). independent experiments. Full heat map is in Supplementary Information.h, Lipidomics of
lysophospholipids in cells treated with Fento-1( 24 h ). independent experiments. i, Lactate dehydrogenase (LDH) release from cells treated with Fento- for 6 h and with inhibitors added 2 h before. independent experiments. Two-sided Mann-Whitney test compared to vehicle. The box plots show the interquartile range, with the centre lines indicating the medians and the whiskers the minimum and maximum values. Def, deferiprone; DFX, deferasirox; Fer-1, ferrostatin-1; Toc, -tocopherol. The following concentrations were used unless stated otherwise: Fento-1 , Toc , Def and Lip- . Phosphatidylcholine (PC) lipids are displayed in the main figure. Subscript Oxn denotes the addition of oxygens. Phosphatidylethanolamine (PE), phosphatidylinositol (PI) and phosphatidylserine (PS) are shown in the Supplementary Information. Lysophosphatidylcholine (LPC) lipids are displayed in the main figure. LPE, LPI and LPS are shown in the Supplementary Information. For d and e, linear model, two-sided -test on fold change. adjusted with Benjamini-Hochberg false-discovery rate. Illustration in b was created using BioRender (https:// biorender.com).
Fig. 3 | Activation of lysosomal iron targets CD44 cancer cells and reduces tumour growth. a, ICP-MS of human healthy and cancer tissues. PDAC, pieces per tissue; UPS and liposarcoma, pieces per tissue. b, ICP-MS of dissociated human tumour cancer cells (CD45 CD31 FAP ). PDAC, ; UPS, ; liposarcoma, replicates per tumour.c, HMRhoNox-M flow cytometry of dissociated human PDAC cancer cells (CD45 CD31 FAP ). patients. d, Lipidomics of oxidized phospholipids of dissociated human tumour cells treated with Fento-1(1 ) and Toc (2 h before). e, CD44 flow cytometry of dissociated human PDAC cancer cells (CD45 CD31 FAP ) treated with Fento- and inhibitors ( 2 h before). patients. One-way ANOVA.f, Colony-formation assay of DTP SUM159 cancer cells after doxorubicin treatment ( ), then Fento-1 ( ) or DMSO ( ) treatment for 72 h . Data are the mean ± s.d.g, ICP-MS of sorted 4T1 tumour cells (CD45 ). replicates. , Tumour volume in 4T1 tumour-bearing
mice. mice per condition. Two-way ANOVA. values (day 10) compared to vehicle. Data are the mean ± s.e.m. CW, Chen-White ligand. i, Maximum tumour diameter as the survival end point of tumour-bearing mice. Mantel-Cox log-rank test.j, Lipidomics of oxidized phospholipids in mouse 4T1 tumours (treatment for 15 days). mice per condition. k, CD44 flow cytometry of dissociated tumour cancer cells (CD45 ) from 4T1 tumour-bearing mice treated with Fento-1 per animal every other day). Vehicle, mice,Fento- mice. The following concentrations were used unless stated otherwise: , , and cLip-1 . For and values are from two-sided Mann-Whitney tests. For and , each coloured dot represents a tumour of a distinct patient per given panel. Full heat maps for and are provided in the Supplementary Information. Box plots show the interquartile range, with centre lines indicating the medians and whiskers the minimum and maximum values.
We also replaced marmycin with cholesterol, an apolar natural product that can intercalate between phospholipids at the plasma membrane and has a role in the mesenchymal state of cancer cells , potentially at the plasma membrane. This modification led to the non-fluorescent analogue Fento-3, which exhibited a potency comparable to that of
Fento-1. Together, these analogues illustrate the versatile nature of this strategy (Extended Data Fig. 7a-c). As a control, replacing the apolar scaffold with a more polar moiety, such as morpholine, produced Fento4, which was biologically inactive (Extended Data Fig. 7a-c). Finally, replacing the Chen-White ligand with the structurally more hindered
Fig. 4 | Iron in ferroptosis and cell-state transitioning. a, Iron is internalized by endocytosis. Lysosomal iron catalyses the production of oxygen-centred radicals from hydroperoxides under acidic conditions. These radicals can abstract a hydrogen from reactive phospholipids to produce carbon-centred radicals, which leads to oxidation products and ferroptosis. Inactivation of
lysosomal iron protects cells against iron-redox chemistry. Activation of lysosomal iron triggers the oxidation of membrane lipids and ferroptosis. Hyal, hyaluronate b, Cell-state transitions induced by standard-of-care and pro-ferroptotic drugs. Illustrations in and were created using BioRender (https://biorender.com).
Nordlander-Costas iron-activating ligand produced Fento-5, which exhibited similar cytotoxic and membrane-lipid-oxidation properties to those of Fento-1, albeit with a delayed cell-death response. This result reflects the distinct reactivity of the iron catalyst towards organic substrates (Extended Data Fig. 7a-c). Together, these data indicate that pharmacological activation of lysosomal iron can trigger ferroptosis.
Iron activation reduces tumour growth
Next, we investigated the effect of lysosomal iron activation in diseaserelevant models. To this end, we quantified the iron content of primary tumour tissues of distinct cancer types, including human PDAC, various human sarcoma subtypes and a mouse model of spontaneous breast cancer metastasis. These cancer tissues were chosen for their refractory nature to standard-of-care treatments and their capacity to form metastases, which contribute to poor clinical outcomes. Furthermore, these indications have been shown to be vulnerable to ferroptosis .
Total iron content was higher in cancer tissues of PDAC, undifferentiated pleomorphic sarcoma (UPS), angiosarcoma, liposarcoma and an epithelioid sarcoma compared with adjacent non-cancerous tissues, as measured by inductively coupled plasma mass spectrometry (ICP-MS) (Fig. 3a and Extended Data Fig. 8a). Moreover, the cellular iron load was higher in subpopulations of cancer cells with high CD44 expression (Fig. 3b and Extended Data Fig. 8b). Cells from freshly dissociated human primary PDAC and sarcoma tissues showed higher levels of redox-active lysosomal iron(II) in the CD44 subpopulations of cancer cells than their CD44 counterparts (Fig. 3c and Extended Data Fig. 8c). This result is consistent with previous findings showing that CD44 mediates iron endocytosis in cancer cells that have acquired a DTP phenotype . In cells obtained from freshly dissociated human primary tumours, Fento-1induced oxidation and lipolysis of membrane phospholipids, an effect that was antagonized by ferroptosis inhibitors (Fig. 3d Extended Data Fig. 8d-g, Supplementary Information and
Supplementary Tables 9 and 10). Fento-1 also reduced the number of CD44 cells in dissociated human PDAC and UPS, and this effect was antagonized by ferroptosis inhibitors (Fig. 3e and Extended Data Fig. 8h,i).
In primary PDAC cells and human PDAC-derived organoids, Fento-1 decreased cell viability more potently than standard-of-care drugs (Extended Data Fig. 9a,b) and synergized with these drugs in several primary cancer cells of human PDAC (Extended Data Fig. 9c,d). In a colony-formation assay, Fento-1 eradicated DTP of triple-negative breast cancer SUM159 cells overexpressing CD44 that survived treatment with doxorubicin (Fig. 3f). In line with this, Fento-1 was more potent against HT-1080 cells that were treated with low doses of doxorubicin for a prolonged period of time (Extended Data Fig. 9e). Under these conditions, HT-1080 cells adopted a distinct morphology and were characterized by increased levels of CD44 and ferritin together with upregulated mesenchymal markers (Extended Data Fig. 9f-h). These data suggest that cells exposed to doxorubicin increase CD44 to promote iron uptake and adopt a more mesenchymal phenotype to escape the doxorubicin-sensitive state. This adaptation leads to higher vulnerability to ferroptosis and sensitivity to Fento-1. Conversely, HT-1080 cells exposed to low doses of Fento-1 exhibited reduced CD44 and ferritin levels together with downregulated mesenchymal markers, which indicated that these cells adapt to lysosomal iron targeting (Fig. 2d and Extended Data Fig. 9g, i).
Regional lymph-node cancer lesions are predictors of distant metastases and mortality, and metastasizing cells are protected against ferroptosis in the lymph . Measurements in mice showed that total iron levels in lymph fluid were lower than in blood and serum (Extended Data Fig. 10a). Thus, we evaluated the effect of Fento-1 on the viability of cancer cells directly in the lymphatics. To this end, we used the 4T1 immunocompetent mouse model of spontaneous triple-negative breast cancer metastases in Balb/c mice. Quantification of iron in cells isolated from intranodal 4T1 tumours showed a higher iron load in CD44 cancer cells than in CD44 subpopulations (Fig. 3g). These
CD44 cells also exhibited a higher iron(II) redox activity in lysosomes (Extended Data Fig. 10b). When mice with intranodal 4T1 tumours were treated with Fento-1(intralymphatic administration every other day), the mice showed reduced tumour growth, increased tumour-size-based survival end points and no adverse effects on body weight (Fig. 3h,i and Extended Data Fig.10c-e). Further analyses of residual tumours indicated that Fento-1 induced oxidation of membrane lipids and the production of lysophospholipids, which affected preferentially CD44 over CD44 cancer-cell subpopulations (Fig. 3j,k, Extended Data Fig. 10f, Supplementary Information and Supplementary Tables 11 and 12). Together, these data show that Fento-1 induces ferroptosis in vivo by exploiting the higher abundance of lysosomal iron(II) in CD44 cancer-cell subpopulations.
Discussion
Ferroptosis is a form of cell death resulting from the uncontrolled oxidation of membrane lipids. Where and how it is initiated in the cell remained unclear. Iron can react with hydroperoxides to initiate or propagate a radical chain reaction independently of enzymes. The lysosomal compartment is a crucial regulator of cellular iron homeostasis , and its acidic nature, together with the presence of reactive iron and hydrogen peroxide, provide an ideal chemical environment to catalyse the oxidation of membrane phospholipids (Fig. 4a). Specific cancer-cell subpopulations upregulate the iron-uptake protein CD44 to promote oxidative demethylation of repressive chromatin marks and to unlock the expression of genes that regulate cell-state transitions involved in cancer progression . Thus, although higher iron levels enable these cells to acquire a DTP profile, this characteristic also inexorably confers vulnerability to ferroptosis (Fig. 4b). The fact that CD44 also marks development, self-renewal, wound healing and immune-cell activation suggests that lysosomal iron also contributes to the induction of ferroptosis in these biological settings. Enhancing the redox activity of lysosomal iron through genetic interventions to eradicate cancer cells is challenging. Here we developed a phospholipid degrader that takes advantage of the higher iron load of specific cancer-cell subpopulations to induce ferroptosis by activating lysosomal iron(II) (Fig. 4a). Sublethal doses of this degrader triggered the acquisition of a ferroptosis-resistant cell state, which partially recapitulated features of epithelial cells (Fig.4b). This result highlights the value of combining cell-state-specific drugs to treat cancer. The anticancer drug doxorubicin can induce damage to genomic DNA in proliferating cells, which, if unrepaired, triggers apoptosis . Conversely, the ferroptosis-resistant cell state, which is characterized by the upregulation of ferroptosis suppressors, epoxide hydrolases, phospholipases and by increased lipid metabolism (Fig. 2d), can arise as a result of cells being able to efficiently repair damaged membranes.
This study shows that fine-tuning the redox activity of lysosomal iron provides control over membrane lipid oxidation, thereby supporting the idea that lysosomal iron is a trigger of ferroptosis. This knowledge can now inform the design of ferroptosis modulators. Fento-1 exhibits a distinct chemotype to investigate ferroptosis and provides a conceptual framework to target DTP cancer cells .
Online content
Any methods, additional references, Nature Portfolio reporting summaries, source data, extended data, supplementary information, acknowledgements, peer review information; details of author contributions and competing interests; and statements of data and code availability are available at https://doi.org/10.1038/s41586-025-08974-4.
2. Friedmann Angeli, J. P. et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat. Cell Biol. 16, 1180-1191 (2014).
3. Bersuker, K. et al. The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis. Nature 575, 688-692 (2019).
4. Doll, S. et al. FSP1 is a glutathione-independent ferroptosis suppressor. Nature 575, 693-698 (2019).
5. Yang, W. S. & Stockwell, B. R. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells. Chem. Biol. 15, 234-245 (2008).
6. Müller, S. et al. CD44 regulates epigenetic plasticity by mediating iron endocytosis. Nat. Chem. 12, 929-938 (2020).
7. Solier, S. et al. A druggable copper-signalling pathway that drives inflammation. Nature 617, 386-394 (2023).
8. Mai, T. T. et al. Salinomycin kills cancer stem cells by sequestering iron in lysosomes. Nat. Chem. 9, 1025-1033 (2017).
9. Rodriguez, R., Schreiber, S. L. & Conrad, M. Persister cancer cells: iron addiction and vulnerability to ferroptosis. Mol. Cell 82, 728-740 (2022).
10. Franca, G. S. et al. Cellular adaptation to cancer therapy along a resistance continuum. Nature 631, 876-883 (2024).
11. Fenton, H. J. H. Oxidation of tartaric acid in presence of iron. J. Chem. Soc. 65, 899-910 (1894).
12. Seiler, A. et al. Glutathione peroxidase 4 senses and translates oxidative stress into 12/15lipoxygenase dependent- and AIF-mediated cell death. Cell Metab. 8, 237-248 (2008).
13. Chen, X., Kang, R., Kroemer, G. & Tang, D. Organelle-specific regulation of ferroptosis. Cell Death Differ. 28, 2843-2856 (2021).
14. Zou, Y. et al. Plasticity of ether lipids promotes ferroptosis susceptibility and evasion. Nature 585, 603-608 (2020).
15. Gao, M. et al. Role of mitochondria in ferroptosis. Mol. Cell 73, 354-363 (2019).
16. Dixon, S. J. et al. Pharmacological inhibition of cystine-glutamate exchange induces endoplasmic reticulum stress and ferroptosis. eLife 3, eO2523 (2014).
17. Torii, S. et al. An essential role for functional lysosomes in ferroptosis of cancer cells. Biochem. J. 473, 769-777 (2016).
18. Antoszczak, M. et al. Iron-sensitive prodrugs that trigger active ferroptosis in drug-tolerant pancreatic cancer cells. J. Am. Chem. Soc. 144, 11536-11545 (2022).
19. von Krusenstiern, A. N. et al. Identification of essential sites of lipid peroxidation in ferroptosis. Nat. Chem. Biol. 19, 719-730 (2023).
20. Jiang, X., Stockwell, B. R. & Conrad, M. Ferroptosis: mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 22, 266-282 (2021).
21. Cañeque, T., Müller, S. & Rodriguez, R. Visualizing biologically active small molecules in cells using click chemistry. Nat. Rev. Chem. 2, 202-215 (2018).
22. Zilka, O. et al. On the mechanism of cytoprotection by ferrostatin-1 and liproxstatin-1 and the role of lipid peroxidation in ferroptotic cell death. ACS Cent. Sci. 3, 232-243 (2017).
23. Shah, R., Farmer, L. A., Zilka, O., Van Kessel, A. T. M. & Pratt, D. A. Beyond DPPH: use of fluorescence-enabled inhibited autoxidation to predict oxidative cell death rescue. Cell Chem. Biol. 26, 1594-1607 (2019).
24. Schonberg, D. L. et al. Preferential iron trafficking characterizes glioblastoma stem-like cells. Cancer Cell 28, 441-455 (2015).
25. Basuli, D. et al. Iron addiction: a novel therapeutic target in ovarian cancer. Oncogene 36, 4089-4099 (2017).
26. Elgrishi, N. et al. A practical beginner’s guide to cyclic voltammetry. J. Chem. Ed. 95, 197-206 (2018).
27. Wei, Y., Aydin, Z., Zhang, Y., Liu, Z. & Guo, M. A turn-on fluorescent sensor for imaging labile in live neuronal cells at subcellular resolution. ChemBioChem 13, 1569-1573 (2012).
28. Niwa, M., Hirayama, T., Okuda, K. & Nagasawa, H. A new class of high-contrast Fe(II) selective fluorescent probes based on spirocyclized scaffolds for visualization of intracellular labile iron delivered by transferrin. Org. Biomol. Chem. 12, 6590-6597 (2014).
29. Viswanathan, V. S. et al. Dependency of a therapy-resistant state of cancer cells on a lipid peroxidase pathway. Nature 547, 453-457 (2017).
30. Hangauer, M. J. et al. Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition. Nature 551, 247-250 (2017).
31. Günthert, U. et al. A new variant of glycoprotein CD44 confers metastatic potential to rat carcinoma cells. Cell 65, 13-24 (1991).
32. Cañeque, T. et al. Synthesis of marmycin A and investigation into its cellular activity. Nat. Chem. 7, 744-751 (2015).
33. Chen, M. S. & White, M. C. A predictably selective aliphatic C-H oxidation reaction for complex molecule synthesis. Science 318, 783-787 (2007).
34. Hertzberg, R. P. & Dervan, P. B. Cleavage of double helical DNA by methidium-propyl-EDTA-iron(II). J. Am. Chem. Soc. 104, 313-315 (1982).
35. Schreiber, S. L. The rise of molecular glues. Cell 184, 3-9 (2021).
36. Qiu, B. et al. Phospholipids with two polyunsaturated fatty acyl tails promote ferroptosis. Cell 187, 1177-1190 (2024).
37. Rodriguez, R. et al. Metal ion signaling in biomedicine. Chem. Rev. 125, 660-744 (2025).
38. Marine, J. C., Dawson, S. J. & Dawson, M. A. Non-genetic mechanisms of therapeutic resistance in cancer. Nat. Rev. Cancer 20, 743-756 (2020).
39. Yin, H., Xu, L. & Porter, N. A. Free radical lipid peroxidation: mechanisms and analysis. Chem. Rev. 111, 5944-5972 (2011).
40. Dai, E. et al. A guideline on the molecular ecosystem regulating ferroptosis. Nat. Cell Biol. 26, 1447-1457 (2024).
41. Pamplona, R. Membrane phospholipids, lipoxidative damage and molecular integrity: a causal role in aging and longevity. Biochim. Biophys. Acta 1777, 1249-1262 (2008).
42. Sezgin, E., Levental, I., Mayor, S. & Eggeling, C. The mystery of membrane organization: composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 18, 361-374 (2017).
43. Abdulla, N., Vincent, C. T. & Kaur, M. Mechanistic insights delineating the role of cholesterol in epithelial mesenchymal transition and drug resistance in cancer. Front. Cell Dev. Biol. 9, 728325 (2021).
44. Mitra, M. et al. Highly enantioselective epoxidation of olefins by catalyzed by a non-heme Fe(II) catalyst of a chiral tetradentate ligand. Dalton Trans. 48, 6123-6131(2019).
Article
Fürstner, A. Iron catalysis in organic synthesis: a critical assessment of what it takes to make this base metal a multitasking champion. ACS Cent. Sci. 2, 778-789 (2016).
Badgley, M. A. et al. Cysteine depletion induces pancreatic tumor ferroptosis in mice. Science 368, 85-89 (2020).
Ubellacker, J. M. et al. Lymph protects metastasizing melanoma cells from ferroptosis. Nature 585, 113-118 (2020).
Rizzollo, F., More, S., Vangheluwe, P. & Agostinis, P. The lysosome as a master regulator of iron metabolism. Trends Biochem. Sci. 46, 960-975 (2021).
Rodriguez, R. et al. Small-molecule-induced DNA damage identifies alternative DNA structures in human genes. Nat. Chem. Biol. 8, 301-310 (2012).
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http:// creativecommons.org/licenses/by-nc-nd/4.0/.
(c) The Author(s) 2025
Methods
Ethics
Fresh tumour samples were obtained by surgery from patients at Paul Brousse and Institut Curie care centres. All patients provided written informed consent for use of tumour samples. The study was approved by institutional regulatory boards (no. 587 and DATA190160). All cLip-1 in vivo experiments were performed in compliance with the German Animal Welfare Law and were approved by the Institutional Committee on Animal Experimentation and the Government of Upper Bavaria (no. ROB-55.2-2532.Vet_02-18-13). All intranodal injection mouse experiments complied with all relevant ethical regulations and were performed according to protocols approved by the Institutional Animal Care and Use Committee at Harvard T. H. Chan School of Public Health (protocol IS00003460). For mouse lymph and blood collection, animal experiments were performed in accordance with the European Community guidelines for the care and use of animals. Animal experiments were performed in agreement with the French guidelines for animal handling and approved by local ethics committees (agreement no. ).
Chemical synthesis
Starting materials were purchased at the highest commercial quality and used without further purification unless otherwise stated. Anhydrous solvents were obtained by passing the degassed solvents through molecular sieves and activated alumina columns. Reactions were monitored by thin layer chromatography (TLC) using aluminium plates coated with silica gel or neutral aluminium oxide from Merck ( ). TLC plates were visualized by UV or by treatment with a ninhydrin, ceric ammonium molybdate or potassium permanganate solution and heating. Reaction products were purified by flash column chromatography on silica gel 60 (230-400 mesh, Macherey Nagel) or aluminium oxide (activated neutral, Sigma-Aldrich) using a CombiFlash NextGen System and a preparative HPLC Quaternary Gradient 2545 equipped with a photodiode array detector 2998 (Waters) fitted with a reverse-phase column (XBridge BEH C18 OBD prep col ). NMR spectroscopy was performed using Bruker 400 or 500 MHz instruments. Spectra were run in methanol- , dimethylsulfoxide- , methylene chloride- or chloroform- at 298 K or 310 K as indicated. chemical shifts are expressed in ppm using the residual non-deuterated solvent as an internal standard, and the coupling constants J are specified in Hz. The following abbreviations are used: bs, broad singlet; s, singlet; d, doublet; dd, doublet of doublets; ddd, doublet of doublet of doublets; dt, doublet of triplets; dq, doublet of quadruplets; q, quadruplet; t, triplet; td, triplet of doublets, quint., quintet; and m, multiplet. chemical shifts are expressed in ppm using the residual non-deuterated solvent as an internal standard. The purity of the final compounds was determined to be >98% by UPLC-MS. Low-resolution mass spectra were recorded using a Waters Acquity H-class equipped with a photodiode array detector and a SQ Detector 2 (UPLC-MS) fitted with a reverse-phase column (Acquity UPLC BEH C181.7 ). High-resolution MS spectra were recorded on a Thermo Scientific Q-Exactive Plus equipped with a Robotic TriVersa NanoMate Advion. Procedures for the synthesis of small molecules are detailed in the Supplementary Information.
NMR titration experiments
NMR spectra were recorded on a Bruker 500 MHz spectrometer at 310 K using Bruker Topspin (v.4.1.4) software and analysed using MestRenova (v.5.0.1-35756) or Bruker Topspin (v.4.1.4) software, and chemical shifts are expressed in ppm using the residual non-deuterated solvent signals as an internal standard. All the solutions were prepared in methanol- . For recordings of NMR spectra of Lip-1 titrated with before the addition of TFA or sodium
deuteroxide, eq. of a solution of (Alfa Aesar, 12357, , 145 mM ) was added to a solution of Lip-1 (Sigma-Aldrich, SML1414, 1 mg in ). Then, a solution of TFA (Sigma-Aldrich, T6508, lot STBG1988V, ) or a solution of sodium deuteroxide (Eurisotop, D076Y, lot R2621, ) was added. For recordings of NMR spectra of Lip-1 and naphthalene titrated with eq. of a solution of was added over a solution of Lip-1 (1 mg in and naphthalene (Sigma-Aldrich, ). After each addition, the solution was stirred for few seconds and NMR spectra were recorded.
Cyclic voltammetry
Cyclic voltammetry experiments were performed with a threeelectrode cell. A saturated calomel electrode was used as reference, a steady glassy carbon electrode of diameter 3 mm was selected as the working electrode and a platinum wire as the counter-electrode. All cyclic voltammograms were recorded at room temperature with a -autolab III from Metrohm using Nova software (v.2.1) with a scan rate of . HPLC-grade acetonitrile and methanol were used for recordings. For all experiments, in acetonitrile ( stock solution) was used. A 10 ml of a stock solution of was prepared with of solution in milliQ water and 9.5 ml acetonitrile. Then, ( 0.2 eq.) of a 40 mM stock solution of DFO, Lip-1, metcLip-1 (in-house) or alcLip-1 (in-house, solubilized in acetonitrile or methanol) was added to 1 ml of solution until 1.0 eq. was reached. After each addition, the solution was stirred for few seconds and voltammograms were recorded.
Molecular modelling
All structures were optimized using the Gaussian 16 (ref. 50) set of programs at the BHLYP/SVP level for all atoms (sextet spin state for iron(III) complexes) (Gaussian 16, revision C.01). Thermal correction to the Gibbs free energy was computed at 310.15 K . Single points at the UMP2/SVP level were performed using the SMD solvation model (methanol). The results presented are in .
Fluorimetry of the iron(II) probe RPE
A solution of RPE (in-house, 10 mM ) in DMSO was diluted with a solution of acetonitrile-acetate buffer ( ) to reach a concentration of . In a 96-well plate, three solutions of RPE , 10 mM ) and 10, 20 or 50 eq. of Lip-1(in-house 1, 2 or of 10 mM stock solution in methanol, respectively) were prepared and the solutions were mixed several times. Then, 10 eq. ( of stock solution in acetonitrile-acetate buffer ( )) was added to each solution and mixed again. Spectra were recorded with a fluorimeter (Tecan spark 10 M ) ( ) using SoftMax Pro 7.1 GxP software. UPLC-grade acetonitrile and methanol as well as milliQ water were used.
A conical centrifuge tube ( 15 ml ) was charged with unilamellar liposomes of egg phosphatidylcholine ( 0.4 ml of a 20 mM suspension in PBS, 100 nm average diameter), 134 ml of a 12 mM solution of (E)-1,2-bis((2-methyldecan-2-yl)oxy)diazene (Cayman, 32742) in ethanol and PBS ( 7.46 ml of 12 mM at pH 7.4 ) and vortexed. STY-Bodipy ( of a 1.74 mM solution in DMSO) was then added and the suspension vortexed once. The liposome-initiator-dye mixture was transferred to a reservoir, and using a multichannel pipette, a 96 -well plate (black, Nunc) was charged with the mixture ( per well). Test compounds ( of a 0.24 mM solution in acetonitrile) were added to bring the final volume to . The contents of the wells were manually mixed using a multichannel pipette and then mixed in a microplate reader for 1 min at , after which the oxidation of STY-Bodipy was monitored by fluorescence ( ) overnight. Data were collected using Agilent BioTek Gen5 software (v.3.08.01).
Antibodies
Antibodies are annotated as follows: WB, western blot; FCy, flow cytometry; FI, fluorescence imaging; Hu, used for human samples; Ms, used for mouse samples. Dilutions are indicated. Any antibody validation by the manufacturer is indicated and can be found on the manufacturers’ websites. Our antibody validation knockdown (KD) and/or knockout (KO) strategies for relevant antibodies are indicated. The following primary antibodies were used: AIFM2/FSP1 (Merck, MABC1638, clone 6D8-11, lot Q3745998, WB, 1:500, Hu, KD validated in-house); catalase (Cell Signaling, 12980T, clone D4P7B, lot 3, FI, 1:200, Hu); CD3-BV510 (BioLegend, 317332, clone OKT3, lot B263750, FCy, 1:100, Hu); CD31-PE-Cy7 (BioLegend, 303118, clone WM59, lot B276836, FCy, 1:100, Hu); CD31-BV605 (BioLegend, 303122, clone WM59, lot 331683, FCy, 1:100, Hu); CD31-BV605 (BioLegend, 102427, clone390, lot B375532, FCy, 1:100, Ms); CD44 (Abcam, ab189524, clone EPR18668, lot 1014086-32, WB, 1:30,000, Hu, KO and KD validated in-house); CD44-AF647 (Novus Biologicals, NB500-481AF488, clone MEM-263, lot P158343, FCy, 1:100, Hu); CD44-AF647 (Novus Biologicals, NB500-481AF647, clone MEM-263, lot D145771, FCy, 1:100, Hu); CD44-AF647 (BioLegend, 103018, clone IM7, lot B317762, FCy, 1:100, Ms); CD45-BV785 (BioLegend, 304048, clone HI30, lot B339809, FCy, 1:100, Hu); CD45-BV510 (BioLegend, 368526, clone 2D1, lot B373428, FCy, 1:100, Hu); CD45-BV510 (BioLegend, 103138, clone 30-F11, lot B386738, FCy, 1:100, Ms); CD163-PerCP/Cyanine5.5(BioLegend, 326512, clone RM3/1, lot B291202, FCy, 1:100, Hu); COXIV (Abcam, ab16056, lot GR3206555-1, WB, 1:1,000, Hu); cytochrome (Cell Signaling, 12963S, clone , lot 2, FI, 1:200, Hu); EEA1 (Abcam, ab70521, clone 1G11, lot GR315680-1, FI, 1:200, Hu, validated by immunocytochemistry/ immunofluorescence by manufacturer); FAP-AF700 (R&D Systems, FAB3715N, clone 427819, lot AEVI020011, FCy, 1:100, Hu); FAP-AF750 (Bio-Techne, FAB3715S-100UG, clone 427819, lot 1718688, FCy, 1:100, Hu); ferritin (Abcam, ab75973, clone EPR300AY, lot 10136442-29, WB, 1:1,000, Hu, validated by WB by manufacturer); fibronectin (Abcam, ab45688, clone F14, lot 1016266-35, WB, 1:1,000, Hu); FTH1 (Santa Cruz Biotechnology, sc-376594, clone B-12, lot G2622, WB, 1:200, Hu); GPX4 (Abcam, ab125066, clone EPNCIR144, lot GR3369574-4, WB, 1:2,000, Hu, KO validated by manufacturer, KD validated in-house); 4-HNE (Abcam, ab48506, clone HNEJ-2, lot 1062274-2, FI, 1:200, Hu); IRP2 (Novus Biologicals, NB100-1798, lot D-4, WB, 1:1,000, Hu); Lamin A/C (Cell Signaling, 2032S, lot 6, WB, 1:1,000, Hu); LAMP1 (Cell Signaling, 9091S, clone D2D11, lot 7, FI, 1:200, Hu); LAMP1 (Santa Cruz Biotechnology, sc-20011, clone H4A3, FI, 1:100, Ms); LAMP1 (Abcam, ab24170, lot GR3235630-1, WB, 1:1,000, Hu); LAMP2 (Abcam, ab25631, clone H4B4, lot 1011336-1, FI, 1:200, Hu); LAMP2 (Thermo Fisher Scientific, MA1-205, clone H4B4, lot YG377512, WB, 1:1,000, Hu); LIMPII (Proteintech, 27102-1-AP, WB, 1:1,000, Hu); NCOA4 (Abcam, ab86707, lot GR3244520-13, WB, 1:10,000, Hu, KD validated in-house); NDUFS1 (Abcam, ab157221, clone EPR11522(B), lot YJ110907DS, WB, 1:1,000, Hu); PDIA3 (Sigma-Aldrich, AMAB90988, clone CL2444, lot 02879, FI, 1:200, Hu; WB, 1:1,000, Hu); MHCII-APC/Cyanine7 (BioLegend, 107628, clone M5/114.12.2, lot B370049, FCy, 1:100, Ms); RCAS1 (Cell Signaling, 12290S, clone D2B6N, lot 1, FI, 1:200, Hu, WB, 1:1,000, Hu); SLC7A11/ xCT (Cell Signaling, 12691S, clone D2M7A, lot 5, WB, 1:1,000, Hu, KD validated in-house); SOX4 (Santa Cruz Biotechnology, sc-518016, clone B-7, lot G2023, WB, 1:200, Hu); TFR1 (Life Technologies, 13-6800, clone H68.4, lot VJ313549, WB, 1:1,000, Hu, KO and KD validated in-house); TFR1-APC-AF750 (Beckman Coulter, A89313, clone YDJ1.2.2, lot 200060, FCy, 1:100, Hu); TFR1-PE (BioLegend, 334106, clone CY1G4, lot B364886, FCy, 1:100, Hu); -tubulin (Sigma-Aldrich, T5326, clone GTU-88, lot 0000140390, WB, 1:1,000, Hu, validated by manufacturer); and vimentin (Cell Signaling, 5741S, clone D21H3, lot 8, WB, 1:1,000, Hu). The following secondary antibodies were used: Alexa Fluor 647 anti-mouse (Abcam, ab150115, tissue labelling, 1:500, Ms); Alexa Fluor 647 anti-mouse (Invitrogen, A21237, lot 1485202, FI,1:1,000,
HT-1080 cells, MDA-MB-231 and 4T1 cells were obtained from the ATCC. Dissociated human and mouse tumour cells and 4T1 cells were cultured in RPMI1640 supplemented with GlutaMAX (Gibco, 61870010) and FBS (Eurobio Scientific, CVFSVF00-01). HT-1080 cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM)-GlutaMAX (Gibco, 61965059) supplemented with 10% FBS (Gibco, 10270-106) and penicillin-streptomycin (BioWhittaker/Lonza, DE17-602E). MDA-MB-231 cells were cultured in DMEM-GlutaMAX supplemented with pyruvate (Thermo Fisher Scientific, 31966021) and supplemented with 10% FBS and penicillin-streptomycin (Gibco, 15140148). FC1242 and FC1245 mouse pancreatic cancer cells, 4 a cells and human pancreatic hMIA-2D cells were a gift from the Tuveson laboratory (Cold Spring Harbor Laboratory) and were cultured in DMEM-GlutaMAX supplemented with 10% FBS and penicillin-streptomycin. Primary human pancreatic PDAC024T, PDAC030T, PDAC053T, PDAC054T, PDAC084T, PDAC090T and PDAC211T cells were grown in serum-free ductal medium DMEM/F12 (Gibco, 10565018) supplemented with 0.61 g in 500 ml nicotinamide (Sigma-Aldrich, 3376), 2.50 g in 500 ml glucose (Sigma-Aldrich, G6152), 1:200 ITS+ (Corning, 354352), 1:20 Nu-serum IV (Corning, 355104 ), cholera toxin, dexamethasone (Sigma-Aldrich, D4902), -triiodo-L-thyronine (Sigma-Aldrich, T6397) and penicillin-streptomycin). Fresh bovine pituitary extract (Gibco, ) and animal-free recombinant human EGF (Thermo Fisher Scientific, AF-100-15-1MG) were added to new medium when cells were split or plated. SUM159 cells were cultured in Ham F12, GlutaMAX (Gibco, 31765035), NEAA (Gibco, 11140050), antibiotic-antimycotic (Thermo Fisher Scientific, 15240062), insulin (Humalog, Cip: 3400934142680), hydrocortisone (Sigma-Aldrich, H0888-10G) and 5% FBS (Thermo Fisher Scientific, A5256701). Primary lung circulating tumour cells (Celprogen, 36107-34CTC, lot 219411, sex: female) and primary colon circulating tumour cells (Celprogen, 36112-39CTC, lot 20188, sex: female) were grown using stem cell complete medium (Celprogen, M36102-29PS) until the third passage. Circulating cancer cells were grown in stem cell ECM T75-flasks (Celprogen, E77002-07-T75). Pfa1 cells were cultured using DMEM high glucose and FBS, 2 mM L -glutamine and 1% penicillin-streptomycin.
Dissociation of human and mouse tumour samples
Tumour samples were collected from patients after surgery. Tumour samples were from patients with PDAC, UPS, liposarcoma, angiosarcoma, epithelioid sarcoma or PDAC liver metastasis. Tumours were dissociated using a human tumour dissociation kit (Miltenyi, 130-095929) according to the manufacturer’s protocol. In brief, tumours were cut into small pieces ( ), put into the enzyme mix in RPMI and dissociated using gentleMACS Octo Dissociator with Heaters (Miltenyi) with the appropriate gentleMACS program (37C_h_TDK). Per tumour sample, 9.4 ml RPMI medium was used with the corresponding enzyme concentration according to the manufacturer’s protocol. Mouse tumour samples were dissociated using a mouse tumour dissociation kit (Miltenyi, 130-096-730) according to the manufacturer’s protocol. Tumours were cut into small pieces ( ), put into the enzyme mix in RPMI and dissociated using gentleMACS Octo Dissociator with Heaters (Miltenyi) with the appropriate gentleMACS program (37C_m_TDK). Subsequently, the dissociated tumour suspension was applied to a MACS SmartStrainer (Miltenyi). Samples were diluted with PBS and centrifuged at 300 g . The cell pellet was resuspended in RPMI
(with 10% FBS and penicillin-streptomycin), and cells were counted using an automated cell counter (Entek). The total population of dissociated cells is denoted as dissociated tumour cells, whereas the subpopulation corresponding to cancer cells from tumours by negative selection (see the section ‘Flow cytometry’) is denoted as dissociated tumour cancer cells.
Establishment of xenograft-derived primary cell cultures
These models were originally derived from patient-derived xenograft (PDX) models. The PDX fragments designated for cell culture were processed in a biosafety chamber. After mincing to small pieces, they were treated with collagenase type V (Sigma-Aldrich, C9263) and trypsinEDTA (Gibco, 25200-056) and suspended in DMEM supplemented with penicillin-streptomycin and FBS. After centrifugation, cells were resuspended in serum-free ductal medium adapted from previous protocols at in a incubator. Amplified cells were stored in liquid nitrogen. Cells were weaned from antibiotics for more than 48 h before testing. This protocol was used to establish the cells designated as PDAC024T, PDAC030T, PDAC053T, PDAC054T, PDAC084T, PDAC090T and PDAC211T.
Establishment of xenograft-derived pancreatic organoids
Xenograft-derived pancreatic organoid (XDPO) models were originally derived from PDX models. Xenografts were split into several small pieces and processed in a biosafety chamber. After mincing to small pieces, they were treated using a human tumour dissociation kit (Miltenyi, 130-095-929). Undigested pellets were digested with accutase (Thermo Fisher Scientific, A1110501) at for 30 min . The pancreatic tissue slurry was transferred to a tissue strainer and then placed into a 12 -well plate coated with Matrigel (Corning, 354230). The samples were cultured in pancreatic organoid feeding medium, which consisted of advanced DMEM/F12 supplemented with 10 mM HEPES (Thermo Fisher Scientific, 15630056), GlutaMAX (Thermo Fisher Scientific, 35050087), penicillin-streptomycin, animal-free recombinant human FGF10 (Thermo Fisher Scientific, animal-free recombinant human EGF (Thermo Fisher Scientific, AF-100-15-1MG), recombinant human noggin (Bio-Techne, 6057-NG), WNT3A-conditioned medium , RSPO1-conditioned medium ( ), 10 nM human gastrin 1 (Sigma-Aldrich, SCP0152), 10 mM nicotinamide (Sigma-Aldrich, NO636), -acetylcysteine (Sigma-Aldrich, A9165), (Thermo Fisher Scientific, 17504001), 500 nM A83-01 (Tocris, 2939/10) and Y27632(Tocris, 1254/1). The plates were incubated at in a incubator, and the medium was changed every days. This procedure was used to generate the XDPOs PDAC009T, PDAC003T, PDAC117T and PDAC372T.
Chemosensitivity profiling of XDPOs
For chemosensitivity profiling, XDPOs were plated into 96-well plates and then subjected to incrementally increasing concentrations of drugs. Cell viability was measured 72 h after treatment using CellTiter-Glo 3D (Promega, G9683). Doubling times of XDPO viability for vehicle-treated conditions were calculated on days 0 and 3 . The ratio of day 3 over day 0 corresponded to the replication rate (RR) of the cells at 72 h . Doubling times were calculated using the formula RR . Fluorescence and luminescence values were quantified using a Tristar LB941 plate reader (Berthold Technologies). Each experiment was performed at least three times with at least three replicates.
Cell death and viability assays
Cell death assay with Annexin-V and propidium iodide. PDAC053T cells were seeded in 6-well plates at a density of cells per well. RSL3 (Sigma-Aldrich, SML2234, 0.1, 0.5, 2 and ) was added together with Lip-1 , cLip-1 , alcLip-1 or metcLip-1 on the following day. After 24 h , the medium was recovered
and cells were trypsinized. Cells were collected, pelleted along with the recovered medium and washed with PBS. Next, of Annexin-V binding buffer containing Annexin-V and propidium iodide was added according to the manufacturer’s protocol (Annexin-V flow cytometry kit, Thermo Fisher Scientific, V13242). PBS buffer containing FBS and EDTA ( ) was added for flow cytometry. Flow cytometry was performed using an AttuneTM NxT flow cytometer and data were analysed using FlowJo.
Cell death assay with Annexin-V and Sytox blue. PDAC053T cells were seeded in 6 -well plates at a density of cells per well. The following day, cells were transfected with siRNA as detailed in the section ‘RNA interference’. Medium was replaced 6 h after transfection. Three days after transfection, cells were treated with the indicated concentrations of Fento-1 for 6 h . Cell death was analysed using Annexin-V AF488 (Thermo Fisher Scientific, A13201) and Sytox blue (Thermo Fisher Scientific, S34857). Flow cytometry was performed using an AttuneTM NxT flow cytometer and data were analysed using FlowJo.
LDH assay. LDH release was measured using a Cytotoxicity detection kit (Sigma-Aldrich, 11644793001) according to the manufacturer’s protocol in a 96-well plate. Cell viability was assessed using a CellTiterGlo 2.0 (Promega, G9241) or CellTiter blue (Promega, G8081) kit according to the manufacturer’s protocol in a 96 -well plate. In brief, 4,000 cells (HT-1080, PDAC053T or 4T1) were seeded per well in clear-bottom and darkened 96-well plates (Greiner, 655090, lot E23063EG) 24 h before the experiment. Cells were then pretreated for 2 h with , cLip-1 , ferrostatin-1 (SML0583, ), DFO (Sigma-Aldrich, D9533, ), deferasirox (Cayman chemical, ), deferiprone (Sigma-Aldrich, Y0001976, ), -tocopherol ( ), vitamin K3 (Sigma-Aldrich, M5625-25G, ), Z-VAD-FMK (Enzo Life Sciences, ALX-260-020-M005, ) or necrostatin-1 (Sigma-Aldrich, N9037, . Subsequently, Fento-1 was added. For doseresponse measurements, cells were treated with varying amounts of the respective molecule for the time indicated. Samples were processed as detailed in the manufacturer’s protocols and data were recorded on a SpectraMax ID3 plate reader (Molecular Devices). For standard-of-care cell-viability measurements, cells were plated at 2,000 cells per well 24 h before the experiment. Cells were incubated with serial dilutions of Fento-1, irinotecan (Sigma-Aldrich, I1406), 5-fluorouracil (5-FU;Alfa Aesar, A13456-06) or oxaliplatin (Bio-Techne, 2623) for 72 h .
MTT assay. Cells were plated in a 96 -well plate, incubated for 24 h and then pretreated with Lip-1 for 10 min before treatment with vehicle control or Fento-1 for 24 h . After 24 h , the medium was carefully removed and serum-free medium and MTT solution (Cayman Chemical, 21795) were added per well. The plate was incubated at for 3 h . After incubation, the solution was removed and DMSO was added per well. The plate was covered and shaken on an orbital shaker for 15 min before reading the absorbance on a plate reader (OD ). For RSL3 treatment, Pfa1 cells were seeded in 96-well plates (2,000 cells per well) and cultured overnight. The following day, cells were treated with RSL3 and the indicated small molecules in serial dilution. For 4-OH-TAM treatment, Pfa1 cells were seeded in 96-well plates ( 500 cells per well) with 4-OH-TAM ( ) and a dilution series of the indicated compounds. After 24 h (for RSL3) or 72 h (for 4-OH TAM) incubation, cell viability was assessed using resazurin as a cell-viability indicator. Fluorescence intensity was measured at using a SpectraMax iD5 microplate reader with SoftMax Pro v. 7 software (Molecular Devices) after 4 h of incubation in standard cell culture medium containing resazurin.
Alamar blue assay. The effect of sublethal doses of Fento-1 together with RSL3 or ML210 on cell viability was assessed using the Alamar blue assay. To determine cell viability, 3,000 HT-1080 or MDA-MB-231
cells were seeded in a 96-well plate with standard medium (10% FBS and penicillin-streptomycin) around 20 h before treatment. Cells were pretreated with Fento- 1 and/or for 1 h . After 48 h of RSL3 or ML210 (Sigma-Aldrich, SML0521-5MG) treatment, a cell-viability assay was performed. Alamar blue solution was made by dissolving 0.5 g resazurin sodium salt (Sigma-Aldrich, 263-718-5) in 100 ml sterile PBS and sterile filtrated through a filter. After 2 h of incubation, viability was assessed by measuring the fluorescence using a excitation filter and a emission filter on a Spark microplate reader (Tecan).
IncuCyte measurements of the cytotoxicity of Fento-1. Kinetics of cell death were collected using an IncuCyte bioimaging platform (Essen). Cells were seeded in 96 -well plates (3,000 cells per well) 1 day before treatment. Cells were treated with Fento-1 in combination with Lip-1 in FluoroBrite DMEM medium (Thermo Fisher Scientific A1896701). Four images per well were captured every hour, analysed and data were averaged. Cell death was measured on the basis of the incorporation of DRAQ7 ( , Cell Signaling, 7406S). Data were collected as count of DRAQ7-positive cells per total number of cells in each condition.
Generation of HT-1080 DTP cancer cells
About HT-1080 cells were plated in T75 flasks 24 h before adding doxorubicin (Clinisciences, HY-15142, 25 nM ). Flasks were washed with PBS buffer, and medium containing doxorubicin ( 25 nM ) was replaced every days. After 30 days of treatment with doxorubicin, 4,000 cells were plated in 96-well plates per well to assess cell viability after treatment with Fento-1.
Clonogenic survival analysis
About SUM159 cells were plated in 10 cm dishes in complete growth medium. Seeded cells were treated with doxorubicin at the half maximal inhibitory concentration ( ) concentration ( 150 nM ). After 72 h , cells were washed and treated with Fento- ) or DMSO without drug. After 72 h , cells were washed, and the treatment was replaced with complete growth medium. After 10-15 days, cells were washed, fixed and stained with acetic acid and methanol (1:7) and Coomassie blue 1 h at room temperature. The number of colonies with cells was counted using a SCAN1200 colony counter (Interscience).
Drug synergy analysis
Cells were plated in 96-well plates (5,000 cells per well). After 12 h , cells were treated with 5-FU (Selleckchem, S1209, ), gemcitabine (Selleckchem, S1714, ), oxaliplatin (Selleckchem, S1224, ), paclitaxel (Selleckchem, S1150, ) or SN-38 (Selleckchem, S4908, alone or in combination with Fento-1 for 72 h . Cell viability was estimated after the addition of PrestoBlue (Life Technologies, A13261) for 2 h following the manufacturer’s instructions. To determine the existence of a synergistic effect on cell proliferation or cell viability between standard-of-care drugs and Fento-1, we used the software SynergyFinder. For the analysis of this effect, the ZIP score was used as a model for calculating synergy .
cLip-1 treatment in vivo
Mice were kept under standard conditions with water and food ad libitum and in a controlled environment ( humidity, 12 h light-dark cycle). For animal studies, C57BL6/J mice were randomized into separate cages. Mice aged 12-24 weeks and sex-matched were used for all experiments. For the survival cohort study, Rosa26-creERT2;Gpx4 mice were intraperitoneally treated with tamoxifen (Sigma-Aldrich, T2859, 2 mg per day dissolved in Myglyol 812) on days 0 and 1 for deletion of in the whole body except in the brain . From day 2, cLip-1 (in-house, 10 mg per kg per day dissolved in PBS
containing 20% PEG400 and 5% Solutol HS15) or vehicle was intraperitoneally administered to the mice each day until the completion of the survival study. For histochemistry analyses, Rosa26-creERT2;Gpx4 mice treated with intraperitoneal tamoxifen injection ( 2 mg per day on days 0 and 1 ) were used. On day 7 , cLip -1 (in-house, 10 and ) or vehicle was intraperitoneally injected into the mice. At 1 h after the injection, mice were euthanized, and the kidney and liver samples were processed as described in the section ‘Small-molecule labelling using click chemistry’.
Isolation of blood, serum and lymphatic fluid from mice
Balb/c mice (25-week-old females) were purchased from Charles River and housed at the CRCM animal core facility and randomized into separate cages. Mice aged 12-24 weeks and sex-matched were used for all experiments. Mice were housed under sterile conditions with sterilized food and water provided ad libitum and maintained on a 12 h light-dark cycle and humidity. Mice were not subjected to any procedures before collection of lymph, blood and serum.
For lymph sample collection, 30 min before the beginning of the experiment, buprenorphine (Buprecare), an analgesic, was administered by intraperitoneal injection ( ). Mice were euthanized by intraperitoneal injection of a ketamine-xylazine combination (ketamine (Imalgène) and xylazine (Rompum); ). After cutaneous and peritoneal incisions, the lymph in the intestinal lymph trunk was collected using a glass capillary. Lymph samples were placed in cryotubes, frozen at and stored at .
For blood and serum sample collection, after lymph collection, we performed a terminal cardiac puncture ( 23 G needle with a syringe) by thoracotomy to collect a large volume of blood without anticoagulants. Next, of the blood sample was placed in a vial, frozen at and stored at . The rest of the blood sample was left to stand at room temperature for 30 min , then centrifuged at for 15 min . The supernatant (serum) was collected in a vial, frozen at and stored at .
Quantitative proteomics
Sample preparation. HT-1080 cells were grown in round dishes at confluence and treated with Fento- . Whole-cell extracts were collected and then centrifuged at 500 g for 5 min at , washed with ice-cold PBS and lysed using lysis buffer ( 8 M urea, 200 mM sodium bicarbonate and complete protease inhibitor cocktail (Roche, 000000011697498001)). Lysis was achieved by incubating on a rotating wheel at . for 45 min at . Lysates were centrifuged at for 20 min at , and the supernatant that contained total protein was used for quantitative global proteomic analysis. In brief, total protein cell lysate was reduced by incubating with 5 mM dithiothreitol at for 30 min and then alkylated with iodoacetamide ( ) at room temperature in the dark. Trypsin/LysC (Promega) was added at 1:100 (w/w) enzyme to substrate. Digestion was performed overnight at . Samples were then loaded onto custom-made C18 StageTips (AttractSPE Disk Bio C18-100.47.20, Affinisep) for desalting. Peptides were eluted using a ratio of 40:60 acetonitrile and formic acid and vacuum concentrated to dryness with a SpeedVac. Peptides were reconstituted in 0.3% TFA before liquid chromatography-tandem mass spectrometry (LC-MS/MS).
LC-MS/MS analyses. LC was performed with a Vanquish Neo LC system (Thermo Scientific) coupled to an Orbitrap Astral mass spectrometer, interfaced by a Nanospray Flex ion source (Thermo Scientific). Peptides were injected onto a C18 column ( m inner diameter double nanoViper PepMap Neo, , Thermo Scientific) regulated at and separated with a linear gradient from buffer A ( and formic acid) to buffer acetonitrile and formic acid) at a flow rate of over 104 min . The instrument was operated in data-independent acquisition (DIA) mode. MS full
scans were recorded on the Orbitrap mass analyser in centroid mode for ranges with a resolution of 240,000 , a normalized automatic gain control (AGC) target set at and a maximum injection time of 5 ms . The DIAs in the Astral analyser were performed in centroid mode for a mass range of with a window width of 2 Da (without overlap), a maximum injection time of 3 ms and a normalized AGC target of 500% after fragmentation using higher-energy collisional dissociation (25% normalized collision energy).
Data processing. For identification, the data were searched against the Homo sapiens (UP000005640) UniProt database using Spectronaut (v.18.7 or v.19; Biognosys) by directDIA + analysis using default search settings. Enzyme specificity was set to trypsin, and a maximum of two missed cleavages were allowed. Carbamidomethyl was set as a fixed modification, and amino-terminal acetylation and oxidation of methionine were set as variable modifications. The resulting files were further processed using myProMS(v.3.10;https://github.com/bioinfo-pf-curie/ myproms) . For protein quantification, XICs from proteotypic peptides shared between compared conditions (TopN matching) with missed cleavages and carbamidomethylation were allowed. Median and scale normalization at the peptide level was applied on the total signal to correct the XICs for each biologically independent replicate ( ). To evaluate the significance of the change in protein abundance, a linear model (adjusted on peptides and biological replicates) was performed, and a two-sided -test was applied on the fold change estimated by the model. The values were then adjusted using the Benjamini-Hochberg false-discovery rate procedure. The MS proteomics raw data have been deposited to the ProteomeXchange Consortium through the PRIDE partner repository .
Intranodal mouse metastasis models and in vivo administration of Fento-1
Mouse breast cancer cells (4T1) were transplanted into 6-8-week-old female Balb/c mice (syngeneic with the 4T1 model). Mice were housed at the Harvard Medical School animal core facility and randomized into separate cages. Mice were housed under sterile conditions with sterilized food and water provided ad libitum and maintained on a 12 h light-dark cycle and humidity. To perform injections into lymph nodes, the lymphatics were first traced by injecting 2% Evans Blue dye (Sigma-Aldrich, E2129) into the foot pedal 5 min before performing intranodal injections. After injecting Evans Blue dye, the mice were anaesthetized using isoflurane and a small ( ) incision was made in the region of the right popliteal lymph node. The lymph node was located on the basis of Evans Blue staining, immobilized with forceps, and cells suspended in PBS were injected in a volume of into the popliteal lymph node using a 27 G Hamilton syringe. Injection into the lymph node was confirmed by visible swelling of the lymph node. The incision was closed using surgical glue ( 3 M VetBond Tissue Adhesive, 1469 SB ) and the mice were closely monitored for signs of pain or distress. Once tumours were palpable in at least of the mice (approximately 1 week after injection), Fento-1 per animal) or vehicle was delivered intralymphatically into the tumour-bearing lymph node every other day until the experimental end point. Intranodal tumour diameters were measured 3 times per week with calipers until any tumour in the mouse cohort reached 2.0 cm in its largest diameter, which was the predetermined experimental end point for these experiments. For all experiments, this maximum permitted tumour diameter was not exceeded. At that point, all mice in the cohort were euthanized, per approved protocols, for analyses of intranodal tumour diameter, tumour mass and other parameters. At the experimental endpoint, data on tumour formation were collected in a manner blinded to sample identity or treatment. For survival-curve determinations, tumour-sized-based survival rate methods were applied, with 1.5 cm as the recommended predetermined survival rate per ethically acceptable cut-off values for determining
therapeutic efficacy in mouse studies . Tumour sizes were measured in two dimensions, length ( axis) and width ( axis), with the axis conventionally defined as the longest axis of the tumour. Tumour sizes are represented as tumour diameter (cm) or tumour volume ( ), as extrapolated by length × width , as indicated for each in vivo experiment. Tumour samples were frozen in DMSO in FBS ( until ) for subsequent cellular analyses. No formal randomization techniques were used. However, animals were allocated randomly to treatment groups and specimens were processed in an arbitrary order. For all experiments, mice were kept on a normal chow diet and fed ad libitum.
Fluorescence imaging
Cells were plated on coverslips and treated as indicated. Bodipy 665/676 (Thermo Fisher Scientific, B3932, ), LysoTracker Deep Red (Thermo Fisher Scientific, L12492, ), DND-189 LysoSensor (Thermo Fisher Scientific, L7535, ), Liperfluo (Dojindo, ) and SQSS (in-house, ) were added to live cells before fixation. For colocalization studies, cells were treated with Fento- ) or marmycin (in-house, ) at the indicated temperature 16 h after transduction. At 16 h after transduction the BacMam-transduced cells were treated with RSL3 for 15 min or for 3 h 15 min and then treated with Bodipy 665/676 for 45 min to give the final time points of 1 h and 4 h . For colocalization of lipid peroxides with lysosomes, cells were treated with RSL3 and either Bodipy 665/676 and DND-189 LysoSensor or Liperfluo and LysoTracker Deep Red for 1 h . To assess the fluorescence intensity of Fento- 1 in the presence of -tocopherol, cells were pretreated with -tocopherol (Sigma-Aldrich, PHR1031, ) and then with Fento-1 ( ). Cells were then washed 3 times with PBS, fixed with paraformaldehyde in PBS for 12 min and then washed 3 times with PBS. For antibody staining, cells were permeabilized with Triton X-100 in PBS for 5 min and washed 3 times with PBS. Subsequently, cells were blocked in 2% BSA (Euromedex, 04-100-812-E), 0.2% Tween-20 and PBS (blocking buffer) for 20 min at room temperature. Cells were incubated with the relevant antibody in blocking buffer for 1 h at room temperature, washed 3 times with PBS and were incubated with secondary antibodies for 1 h . Finally, coverslips were washed 3 times with PBS and mounted using Vectashield containing DAPI (Vector Laboratories, H-1200-10). Bodipy 665/676 and Liperfluo-treated cells were fixed using ice-cold reagents and placed at immediately after mounting on coverslips and imaged. Fluorescence images were acquired using a Deltavision real-time microscope (Applied Precision), a thunder microscope (Leica) or an Apotome Weiss microscope with Deltavision imaging software, Thunder Leica image acquisition software or Apotome Zeiss Imaging software, respectively. or NA objective was used for acquisitions, and all images were acquired as stacks. Images were deconvoluted using SoftWorx (ratio conservative, 15 iterations, Applied Precision) and processed with Fiji . Images were taken in black and white and colouring was applied with Fiji. Fluorescence intensity is displayed as arbitrary units and is not comparable between different panels. Colocalization quantification was calculated using Fiji 2.0.0-rc-69/1.52n. Nuclei were detected using DAPI or Hoechst fluorescence as indicated.
Bright-field microscopy and digital photographs
Bright-field images were acquired using a CKX41 microscope (Olympus) and cellSens Entry imaging software (Olympus).
Small-molecule labelling using click chemistry
In-cell click labelling. Cells on coverslips were treated as indicated with cDFO (in-house, ), cLip-1 (in-house, or 2 h ), metcLip-1 (in-house, ), alcLip-1 (in-house, ) or cCW , then fixed and permeabilized as indicated in the section ‘Fluorescence imaging’. Lyso Tracker Deep Red was added to live cells for
45 min before fixation. The click reaction cocktail was prepared using a Click-iT EdU Imaging kit (Invitrogen, C10337) according to the manufacturer’s protocol. In a typical experiment, we mixed of Click-iT reaction buffer with solution, Alexa-Fluor-azide, reaction buffer additive (sodium ascorbate) and ultrapure water to reach a final volume of . Coverslips were incubated with the click reaction cocktail in the dark at room temperature for 30 min , then washed 3 times with PBS. Immunofluorescence imaging was then performed as described in the section ‘Fluorescence imaging’.
Click labelling in tissues. Kidney and liver tissue samples collected from cLip-1-treated mice were fixed in paraformaldehyde in PBS overnight at . Fixed tissues were incubated in sucrose in PBS for 30 min and then incubated in sucrose in PBS at for 4 h , followed by embedding in OCT mounting compound (TissueTek, Sakura) on dry ice and stored at . Frozen tissues were cut into -thick sections using a Cryostat Microm HM 560 (Thermo Fisher Scientific) at . Tissue sections were post-fixed with 1% paraformaldehyde in PBS for 10 min and subsequently incubated with acetone for 10 min at . Sections were incubated in blocking solution ( PBS containing BSA and Triton X-100) for 30 min . The specimens were labelled using a click reaction as described above. For costaining with a lysosomal marker, the click-labelled specimens were incubated with anti-LAMP1 or antibody diluted in PBS containing normal goat serum overnight at . The next day, sections were incubated with secondary antibody in PBS containing 1% BSA and Triton X-100 for 2 h at room temperature. Nuclei were visualized with Hoechst 33342 staining, and slides were mounted in Aqua/ PolyMount (Polysciences). Images were acquired using a fluorescence microscope (DS-Qi2, Nikon) or a confocal microscope (LSM880, Carl Zeiss) and the corresponding appropriate filter sets for fluorophores.
Western blotting
Cells were treated as indicated and then washed with PBS. Proteins were solubilized in Laemmli buffer containing benzonase (VWR, 70664-3, 1:100). Extracts were incubated at for 1 h , heated at for 10 min and quantified using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific). Protein lysates were resolved by SDS-PAGE electrophoresis (Invitrogen sure-lock system and Nu-PAGE Bis-Tris precast gels). In a typical experiment, of total protein extract were loaded per lane in Laemmli buffer containing bromophenol blue. On each gel, a size marker was run: PageRuler (Thermo Fisher Scientific, 26616) or PageRuler plus (Thermo Fisher Scientific, 26620) and Laemmli buffer. Proteins were then transferred onto nitrocellulose membranes (Amersham Protran ) using a Trans-Blot SD semi-dry electrophoretic transfer cell (Bio-Rad) using NuPage transfer buffer (Invitrogen, NP00061) with methanol. Membranes were blocked with 5% non-fat skimmed milk powder (Régilait) in Tween- 20 and PBS for 20 min . Membranes were cut at the appropriate marker size to enable the probing of several antibodies on the same membrane. Blots were then probed with the relevant primary antibodies in BSA, Tween-20 and PBS or in 5% non-fat skimmed milk powder in 0.1% Tween-20 and PBS at overnight with gentle motion in a hand-sealed transparent plastic bag. Membranes were washed with Tween- 20 and PBS 3 times and incubated with horseradish-peroxidase-conjugated secondary antibodies (Jackson Laboratories) in 5% non-fat skimmed milk powder (Régilait) in Tween-20 and PBS for 1 h at room temperature and washed 3 times with Tween- 20 and PBS. Antigens were detected using SuperSignal West Pico PLUS (Thermo Fisher Scientific, 34580) and SuperSignal West Femto (Thermo Fisher Scientific, 34096) chemiluminescent detection kits. Signals were recorded using a Fusion Solo S Imaging System (Vilber) with FusionCapt Advance software. Images were processed using Fiji 2.0.0-rc-69/1.52n software. Full scans of blots are provided in the Supplementary Information.
Transduction
PDAC053T cells were seeded in 4-well plates at a density of cells per well 24 h before the experiment in 1 ml cell culture medium. Cells were then transduced with CellLight Lysosomes-GFP, BacMam 2.0 (Thermo Fisher Scientific, C10596), CellLight ER-GFP, BacMam 2.0 (Thermo Fisher Scientific, C10590) or CellLight Mitochondria-GFP, BacMam 2.0 (Thermo Fisher Scientific, C10600) according to the manufacturer’s instructions. In brief, reagent was added to the medium and mixed. Cells were incubated for 16 h .
RNA interference
Cells were plated 24 h before the experiments. Cells were transfected with the specified siRNA using jetPRIME (Polyplus, 114-15) according to the manufacturer’s protocol with 100 nM siRNA. In brief, PDAC053T cells were seeded in 6-well plates at a density of cells per well and transfected 24 h later. The medium was replaced after 6 h . Analysis was performed 72 h after transfection. Suitable siRNAs were designed by Dharmacon for specific downregulation of target genes. siRNA sequences are provided in the Supplementary Information.
Flow cytometry
Cells were washed with ice-cold PBS. For antibody staining, cells were incubated with Fc block (Human TruStain FcX, BioLegend, 422302, ) for 15 min , then incubated with antibodies in ice-cold FBS, PBS, 2 mM EDTA for 20 min at and then washed with PBS and resuspended in FBS, PBS and 2 mM EDTA before analysis using a BD LSR Fortessa X-20 flow cytometer with FACS DIVA software (v.9.0.1).
Flow cytometry with iron probes. PDAC053T and HT-1080 cells were seeded in 6-well plates at a density of cells per well. On the next day, Lip-1 , hydroxychloroquine (Sigma-Aldrich, ) or bafilomycin A1 (Sigma-Aldrich, B1793, 75 nM ) was added to PDAC053T cells for 1 h and to HT-1080 cells for 30 min . For PDAC053T cells, after 30 min of treatment with compounds, RPE (in-house, ) or HMRhoNox- (in-house, ) probes were added for 30 min . For HT-1080 cells, after 15 min of treatment with compounds, RPE (in-house, ) or HMRhoNox- (in-house, probes were added and left for 15 min . After incubation with the iron probes, the medium was removed and cells were washed with PBS once before trypsinization. Cells were collected, pelleted, washed with PBS and finally of PBS buffer containing FBS and EDTA ( ) was added. Data were recorded on a BD LSR Fortessa X-20.
Flow cytometry with Bodipy-C11 581/591. PDAC053T cells were seeded in 6 -well plates at the density of cells per well. On the next day, cells were treated with Lip-1 , cLip-1 (in-house, ), metclip-1 (in-house, ), alcLip-1 (in-house, ) and BodipyC11 581/591 ( 200 nM ) before adding RSL3 ( 100 nM ). After 1 h , the medium was removed and cells were washed with PBS twice before trypsinization. Cells were collected, pelleted, washed with PBS and finally of PBS buffer containing FBS and EDTA was added for flow cytometry. PDAC053T, HT-1080 and 4T1 cells were seeded in 6-well plates at a density of cells per well. On the next day, cells were treated with bafilomycin A1 ( 75 nM ) and hydroxychloroquine ( ) for 2 h before adding RSL3 ( 200 nM for PDAC053T and HT-1080, 500 nM for 4T1). After 1 h , cells were treated with Bodipy-C11 for an additional 1 h . The medium was removed and cells were washed with PBS twice before trypsinization. Cells were collected, pelleted, washed with PBS and finally of PBS buffer containing FBS and EDTA , was added for flow cytometry. Data were recorded on an AttuneTM NxT flow cytometer (Thermo Fisher Scientific) using Attune NxT (v.4.2.0).
Analyses of lysosomal glutathione and lysosomal hydroxy radicals. Cells were incubated with SQSS (in-house, ) or 1-Red (in-house, ). HT-1080 cells were incubated with RSL3 , ML210 , Sigma-Aldrich, SML0521), FIN56 , Sigma-Aldrich, SML1740), buthionine sulfoximine ( , Sigma-Aldrich, B2515) or erastin ( , Sigma-Aldrich, 329600) for the indicated time points. Data were recorded on an AttuneTM NxT flow cytometer (Thermo Fisher Scientific).
Analyses of lysosomal iron content. Typically, dissociated human tissue cells were incubated in medium (RPMI 1610,10% FBS and penicillin-streptomycin) with HMRhoNox- (in-house, ). Lysosomal iron content of human tumour samples and healthy adjacent tissues was analysed with the following antibody and stain panel: DAPI ( ), CD3-BV510 (BioLegend, 317332), CD31-PE-Cy7 (BioLegend, 303118), CD44-AF647 (Novus Biologicals, NB500481AF647), CD45-BV785 (BioLegend, 304048), CD163-PerCP/Cyanine5.5 (BioLegend, 326512), FAP-AF700 (R&D Systems, FAB3715N) and TFR1-APC-AF750 (Beckman Coulter, A89313). The live tumour cancer cells corresponded to DAPI cells. Data were recorded on a BD LSRFortessa X-20 or AttuneTM NxT flow cytometer (Thermo Fisher Scientific).
Analyses of CD44 in Fento-1-treated tumour samples. Typically, dissociated tumour cells were incubated in medium (RPMI1610, FBS and penicillin-streptomycin) with Fento-1 (in-house, , 24 h ). Cells were pretreated with Lip-1 , cLip-1 (in-house, ), -tocopherol or deferiprone for 2 h . -Tocopherol was kept pure under inert atmosphere, and a fresh stock solution was prepared throughout the study before each experiment. The following antibody and stain panel was used for subsequent flow cytometry analysis: Sytox blue (Thermo Fisher Scientific, S34857, ), CD31-BV605 (BioLegend, 303122), CD45-BV510 (BioLegend, 368526, lot B373428), CD44-AF647 (Novus Biologicals, NB500-481AF647), TFR1-PE (BioLegend, 334106) and FAP-AF750 (Novus Biologicals, FAB3715S-100UG). The live tumour cells corresponded to Sytox blue cells. Data were recorded on an AttuneTM NxT flow cytometer (Thermo Fisher Scientific). For flow cytometry analyses of CD44 levels in mouse 4T1 tumours, typically, dissociated tumour cells were used per condition. Freshly dissociated cells were stained using the following antibody and stain panel: Sytox blue (Thermo Fisher Scientific, S34857, , CD31-BV605 (BioLegend, 102427), CD44-AF647 (BioLegend, 103018), CD45-BV510 (BioLegend, 103138) and MHCII-APC/Cyanine7 (BioLegend, 107628). The live tumour cancer cells corresponded to Sytox blue cells. Data were recorded on an AttuneTM NxT flow cytometer (Thermo Fisher Scientific). All data were analysed using FlowJo (v.10.10.0).
FACS
Sorting of human cells was performed using the following antibodies: CD31-PE-Cy7 (BioLegend, 303118), CD44-AF647 (Novus Biologicals, NB500-481AF647) and CD45-BV785 (BioLegend, 304048). The sorted cells corresponded to cells and CD44 cells, which were isolated on a FACSAria Fusion (BD) using FACS DIVA (v.9.0.1). ICP-MS experiments were conducted using CD44 and CD44 tumour cells as described in the section ‘ICP-MS’. Sorting of mouse cells was performed using the following antibodies: CD44-AF647 (BioLegend, 103018) and MHCII-APC/Cyanine7 (BioLegend, 107628). The sorted cells corresponded to cells and cells. Sorted cells were centrifuged at and cell pellets were processed for subsequent applications.
ICP-MS
Glass vials equipped with Teflon septa were cleaned with nitric acid (VWR, Suprapur, 1.00441.0250), washed with ultrapure water
(Sigma-Aldrich, 1012620500) and dried. Cells were collected and washed twice with PBS. Cells were then counted using an automated cell counter (Entek) and transferred in PBS or ultrapure water to the cleaned glass vials. The same volume of PBS or ultrapure water was transferred into separate vials for background subtraction, at least in duplicate per experiment. For tissue samples, a small piece of about was transferred into a clean preweighed vial. Samples were lyophilized using a freeze dryer (Christ, 2-4 LDplus). Vials with tissue samples were subsequently weighed to determine the tissue dry weight. Samples were then mixed with nitric acid and heated at overnight in the same glass vials closed with a lid carrying a Teflon septum. Samples were then cooled to room temperature and diluted with ultrapure water to a final concentration of 0.475 N nitric acid and transferred to metal-free centrifuge vials (VWR, 89049-172) for subsequent MS analyses. Amounts of metal were measured using an Agilent 7900 ICP-QMS in low-resolution mode, taking natural isotope distribution into account. Sample introduction was achieved with a micronebulizer (MicroMist, ) through a Scott spray chamber. Isotopes were measured using a collision-reaction interface with helium gas ( ) to remove polyatomic interferences. Scandium and indium internal standards were injected after inline mixing with the samples to control the absence of signal drift and matrix effects. A mix of certified standards was measured at concentrations spanning those of the samples to convert count measurements to concentrations in the solution. Values were normalized against cell number or tissue dry weight.
Liposome preparation and lipid oxidation studies
Liposomal structures were prepared using the traditional lipid film hydration method. In brief, of a stock solution ( chloroform) of ( -cis) PC (DOPC, Avanti Polar Lipids) was dissolved in chloroform and transferred into a round-bottom flask. The organic solvent was removed under reduced pressure in a rotary evaporator for 15 min at . in a water bath at . Afterwards, the lipid film was dried with a vacuum pump overnight. The sample was hydrated with 1 ml of 0.1 mM sodium acetate buffer ( pH 4.5 ) and vortexed every 5 min for 20 min . Liposomes were extruded by passing the suspension through 2 polycarbonate membranes (pore size times. For the control experiment, of the liposome solution was added into an Eppendorf tube and heated at with agitation at . Then, of an aqueous solution of iron(II) triflate ( 1.4 mg in 1.5 ml ) and of 0.1 mM acetate buffer ( pH 4.5 ) were added. At of an aqueous solution of in 1 ml ) was added. For the Fento-1 experiment, of the liposome solution was added to an Eppendorf tube and heated at at . Then, of a 1 mM solution of Fento- 1 in DMSO and of an aqueous solution of iron(II) triflate ( 1.4 mg in 1.5 ml ) were added. At of an aqueous solution of in 1 ml ) was added. DOPC oxidation was recorded with a QExactive mass spectrometer (Thermo Fisher Scientific) equipped with a TriVersa NanoMate ion source (Advion Biosciences) as detailed in the section ‘MS-based lipidomics’. Samples were injected at , 7 h and 24 h reaction times.
Isolation of lysosome-enriched fractions
HT-1080 cells were treated with Fento-1(10 for 1 h, and lysosomeenriched fractions were isolated using a Lysosome Isolation kit (Abcam, ab234047) according to the manufacturer’s protocol. In brief, cells were washed and centrifuged at 600 g for 10 min and the supernatant was removed. Cells were resuspended in lysosome isolation buffer, vortexed and incubated on ice for 2 min. Complete cell disruption was obtained using a dounce homogenizer. After adding lysosome enrichment buffer, the homogenate was centrifuged at 500 g for 10 min at . The supernatant was added to the top of a discontinuous gradient density and an ultracentrifugation at for 2 h at was performed.
The lysosome-enriched fraction was present in the top 10% of the gradient volume. For MS-based lipidomics, the lysosomal fraction was mixed with 2 volumes of PBS, vortexed and centrifuged at for 30 min at . Next, of 150 mM sodium bicarbonate was added to the pellet and the sample was flash-frozen in liquid nitrogen. Lipidomic analyses were performed on these flash-frozen samples. For western blot analyses, the protein content of the lysosomal-enriched gradient supernatant was quantified using a Qbit 1 fluorometer (Thermo Fisher Scientific) and a protein quantification kit (Thermo Fisher Scientific, Q33212). Equal total protein amounts of total cell extracts and lysosome-enriched extracts were loaded for comparison for western blot analyses.
MS-based lipidomics
For comparison of ferroptosis inducers, HT-1080 cells were treated with Fento-1 (in house, ), erastin ( ), RSL3 ( 100 nM ) or iFSP1 ( , Sigma-Aldrich, SML2749) for 24 h . For cotreatment with ferroptosis inhibitors, HT-1080 cells were pretreated with -tocopherol , deferiprone or for 2 h , and then with Fento-1 for 24 h . Dissociated human tumour samples were pretreated with -tocopherol and then with Fento-1 for 24 h . PDAC053T cells were treated with Fento-1 for 6 h or 24 h . Dissociated mouse tumour samples were counted and processed directly after dissociation and pretreated with -tocopherol for 2 h and then with Fento-1 . Cells were subsequently washed with PBS and then with 150 mM ammonium bicarbonate. Cells were then resuspended in 150 mM ammonium bicarbonate and centrifuged at 300 g for 5 min . The supernatant was removed and cells were resuspended in 1 ml of 150 mM ammonium bicarbonate. The solutions were centrifuged at 12,000 r.p.m. for 10 min and the supernatant was removed. Next, of 150 mM sodium bicarbonate was added to the pellet and samples were flash-frozen in liquid nitrogen. Lipidomic analyses were performed on the same day for all technical and biological replicates for a given dataset. For lipidomic analyses, cell lysate was spiked with l internal standard lipid mixture containing 300 pmol phosphatidylcholine (PC) 17:0-17:0, 50 pmol phosphatidylethanolamine (PE) 17:0-17:0, 50 pmol phosphatidylinositol (PI) 16:0-16:0, 50 pmol phosphatidylserine (PS) 17:0-17:0, 30 pmol phosphatidic acid (PA) 17:0-17:0,30 pmol phosphatidylglycerol (PG), 30 pmol lysophosphatidylcholine (LPC) 12:0, 30 pmol lysophosphatidylethanolamine (LPE) 17:1,30 pmol lysophosphatidylserine (LPS) 17:1 and 30 pmol lysophosphatidic acid (LPA) 17:0 and subjected to lipid extraction at as previously described . In brief, the sample was extracted with 1 ml chloroform and methanol (10:1) for 2 h at with vigorous shaking in a thermomixer (1,000 r.p.m.). The lower organic phase was collected and dried in a SpeedVac vacuum concentrator. The remaining aqueous phase was re-extracted with 1 ml chloroform and methanol for 1 h at the same temperature and shaking conditions. The lower organic phase was collected and evaporated in a SpeedVac vacuum concentrator.Lipid extracts were dissolved in infusion mixture consisting of 7.5 mM ammonium acetate dissolved in propanol, chloroform and methanol ( ). For the in vitro liposome experiments, of each reaction mixture taken at different time points was added to infusion mixture consisting of 7.5 mM ammonium acetate dissolved in propanol, chloroform and methanol ( ) containing 300 pmol PC 17:0-17:0. Samples were analysed by direct infusion in a QExactive mass spectrometer (Thermo Fisher Scientific) equipped with a TriVersa NanoMate ion source (Advion Biosciences). In brief, of sample was infused with the gas pressure and voltage set to 1.25 psi and 0.95 kV , respectively. PC and oxidized were detected in the 10:1 extract by positive-ion mode FTMS as protonated adducts by scanning , at with lock mass activated at a common background for 30 s . Every scan is the average of 2 microscans, and the AGC was set to and the maximum ion injection time (IT) was set to 50 ms . PE, oxidized PE
( ) and LPE were detected as deprotonated adducts and LPC were detected as acetate adducts in the 10:1 extract by negative-ion mode FTMS by scanning , at with lock mass activated at a common background for 30 s . Every scan is the average of 2 microscans, AGC was set to and the maximum ion IT was set to 50 ms . PI, oxidized PI ( ), PS, oxidized PS (PS ), lysophosphatidylinositol (LPI) and lysophosphatidylserine (LPS) were detected in the 2:1 extract, by negative-ion mode FTMS as deprotonated ions by scanning , at with lock mass activated at a common background ( ) for 30 s . Every scan is the average of 2 microscans, the AGC was set to and the maximum ion IT was set to 50 ms . Annotation of the oxidized phospholipids only details the number of additional oxygen atoms and double bonds present in a given mass and it can potentially refer to several different structures with the same monoisotopic mass. All data were acquired in centroid mode. All lipidomic data were analysed with the lipid identification software LipidXplorer (v.1.2.8;http://genomebiology.com/2011/12/1/R8). Tolerance for MS and identification was set to 2 ppm. Data were normalized against internal standards and total input to the respective lipid species. Lipidomic data were clustered using a correlation distance and Ward linkage, and represented as heat maps of normalized values.
Glycerol quantification
HT-1080 cells were treated with Fento-1(in-house, 1 or ). Glycerol was quantified using a Glycerol-glo assay (Promega,J3150) according to the manufacturer’s protocol. In brief, the assay was performed in a 96 -well plate and 4,000 cells were plated per well 24 h before the experiment. A standard curve was prepared for each biological experiment and three technical replicates were performed for each condition and each biological replicate. Luminescence signals were recorded using a SpectraMax ID3 plate reader (Molecular Devices). Data were exported and analysed using Excel (Microsoft) and Prism software.
Software for illustrations
Illustrations were created using Fiji 2.0.0-rc-69/1.52n, Prism 10.0.3 and Adobe Illustrator 26.0.2. Illustrations in Figs. 2b and 4 were created using BioRender (https://biorender.com).
Statistical and reproducibility
Results are presented as the mean ± s.e.m. or s.d. as indicated. For box plots, boxes represent the interquartile range and median, and whiskers indicate the minimum and maximum values. Prism (v.10.0.3) software was used to calculate values using a two-sided MannWhitney test, two-sided unpaired -test, Kruskal-Wallis test with Dunn’s post-test, one-way ANOVA, two-way ANOVA or Mantel-Cox log-rank test as indicated. Prism (v.10.0.3) software was used to generate graphical representations of quantifications unless stated otherwise. Sample sizes ( ) are indicated in the figure legends and were not predetermined. A minimum of independent experiments were performed as a standard, and sample sizes were increased in more complex experiments to ensure reproducibility. Exact values are indicated in the figures.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Lipidomics data are presented in the Supplementary Tables 1-4 and 7-12. Quantitative proteomic raw data have been deposited to the ProteomeXchange Consortium through the PRIDE partner repository with the dataset identifier PXD054449. Source data are provided with this paper.
50. Frisch, M. J. et al. Gaussian 09, Revision B. 01 (Gaussian, 2010).
51. Schreiber, F. S. et al. Successful growth and characterization of mouse pancreatic ductal cells: functional properties of the Ki-RAS oncogene. Gastroenterology 127, 250-260 (2004).
52. Yadav, B., Wennerberg, K., Aittokallio, T. & Tang, J. Searching for drug synergy in complex dose-response landscapes using an interaction potency model. Comput. Struct. Biotechnol. J. 13, 504-513 (2015).
53. Ionac, M. One technique, two approaches, and results: thoracic duct cannulation in small laboratory animals. Microsurgery 23, 239-245 (2003).
54. Poullet, P., Carpentier, S. & Barillot, E. myProMS, a web server for management and validation of mass spectrometry-based proteomic data. Proteomics 7, 2553-2556 (2007).
55. Perez-Riverol, Y. et al. The PRIDE database resources in 2022: a hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 50, D543-D552 (2022).
56. Workman, P. et al. Guidelines for the welfare and use of animals in cancer research. Br. J. Cancer 102, 1555-1577 (2010).
57. Sampaio, J. L. et al. Membrane lipidome of an epithelial cell line. Proc. Natl Acad. Sci. USA 108, 1903-1907 (2011).
Acknowledgements R.R. is supported by the European Research Council under the European Union’s Horizon 2020 research and innovation programme (grant agreement no. 647973), Ligue Contre le Cancer (Equipe Labellisée), Fondation pour la Recherche Médicale, Fondation Charles Defforey-Institut de France, Klaus Grohe Foudation, Fondation Bettencourt Schueller, Région Ile-de-France, Institut National du Cancer (PLBIO-2022-099) and Agence Nationale de la Recherche (ANR-23-CE44-0030 and ANR-23-IAHU-0006). V.G. thanks the UPSaclay and CNRS for financial support. This work was granted access to the HPC resources of CINES under the allocation 2020-A0070810977 made by GENCI. M.C. received funding from the Deutsche Forschungsgemeinschaft (DFG) (CO 291/7-1, 428858739 and the Priority Program SPP 2306 (CO 291/9-1, 461385412; CO 291/10-1, 461507177; CO 291/9-2, CO 291/10-2, CO 291/14-1) and CRC 353 (CO 291/11-1; 471011418), the German Federal Ministry of Education and Research (BMBF) VIP+ programme NEUROPROTEKT (O3VPO4260), the BMBF programme FERROPATH (01EJ2205B) and the European Research Council under the European Union’s Horizon 2020 research and innovation programme (grant agreement no. 884754). J.P.F.A. acknowledges institutional support of the University of Würzburg and additional support from the Deutsche Forschungsgemeinschaft (DFG), FR 3746/3-1, FR 3746/5-1, FR 3746/6-1 and CRC205 (INST 269/886-1), the European Union (ERC-Consolidator, DeciFERR, Horizon Europe ERC grant no. 101126134) and the Deutsche Jose Carreras Leukämie Stiftung (DJCLS 01R/2022). J.M.U. is supported by the Breast Cancer Alliance Young Investigator Grant and The Ludwig Center at Harvard. A.C. is supported by the Landry Cancer Biology Research Fellowship. A.C. and C.S.F. were funded by NIH NIDDK (T32DK128781). E.C.-J. and C. Ginestier are supported by Ligue Contre le Cancer (Equipe Labellisée). D.A.P. is supported by the Natural Sciences and
Engineering Research Council of Canada (RGPIN-2022-05058). The ICP-MS platform at Institut de Physique du Globe de Paris is supported by IPGP multidisciplinary program PARI and Région Ile-de-France (SESAME grant agreement no. 12015908). The cell and tissue imaging facilities (PICT-IBiSA) of Institut Curie are part of the France Bioimaging National Infrastructure (ANR-10-INBS-04 and ANR-24-INBS-0005 FBI BIOGEN), the MS platform is supported by Région Ile-de-France (NºEX061034) and ITMO Cancer of Aviesan and INCa supported by funds administered by INSERM (no. 21CQ016-00), and the cytometry platform is part of the Institut Curie. We thank S. Hidalgo for assistance with ICP-MS experiments; and R.R. thanks S.Balasubramanian, S.L.Schreiber, J.-M.Lehn and E.M. Carreira for support.
Author contributions R.R. conceptualized the study and directed the research. R.R., T.C., L.B., S.M. and J.M.U. designed the experiments. T.C., L.B., L.C., A.V. and C. Gaillet performed chemical synthesis, NMR spectroscopy and MS. L.B. and L.G. performed cyclic voltammetry. A.U. measured RTA activity in vitro. V.G. performed molecular modelling. S.M., L.B., S.S., F.S., A.P.-B. and L.K.T. performed transductions, transfections, fluorescence imaging, flow cytometry, western blotting, lysosome isolation, cell death and cell viability assays. S.M. prepared samples for ICP-MS. J.L.S. performed lipidomics. F.S., B.L. and D.L. performed quantitative proteomics. L.S. and N.S. performed bioinformatics. E.D.K., C.B., P.S.-C., M.E., N.D., J.L.I., F.P.F. and J.Z. performed colony-formation, synthetic lethality and combination therapy assays. A.S.C., G.P., D.T. and S.B. selected intraoperative healthy and tumour tissue samples. S.M. and S.S. performed analyses of human tumour tissues. E.M. and B.P. performed evaluation of cLip-1 in vivo. A.C., M.S., C.S.F. and K.J.S. performed intralymphatic treatments of 4T1 tumour-bearing mice. All the authors contributed to data analyses. R.R., T.C. and S.M. wrote the article. E.C-.J., C. Ginestier, P.H., S.W., D.A.P., J.P.F.A., B.R.S., M.C. and J.M.U. commented on the manuscript.
Competing interests Institut Curie filed a patent application (PCT/EP2025/O51999) related to lipid degraders to induce ferroptosis in cancer. M.C. and B.P. hold patents for some of the compounds described herein and are co-founders of ROSCUE Therapeutics. B.R.S. is an inventor on patents and patent applications involving ferroptosis; co-founded and serves as a consultant to ProJenX and Exarta Therapeutics; holds equity in Sonata Therapeutics; and serves as a consultant to Weatherwax Biotechnologies and Akin Gump Strauss Hauer & Feld. All the other authors declare no competing interests.
Additional information
Supplementary information The online version contains supplementary material available at https://doi.org/10.1038/s41586-025-08974-4.
Correspondence and requests for materials should be addressed to Raphaël Rodriguez. Peer review information Nature thanks the anonymous reviewers for their contribution to the peer review of this work. Peer reviewer reports are available.
Reprints and permissions information is available at http://www.nature.com/reprints.
Extended Data Fig.1|See next page for caption.
Extended Data Fig. 1 | Inactivation of lysosomal iron inhibits ferroptosis (Part 1). a, Chemical synthesis of cLip-1.b, Fluorescence imaging of labelled cLip-1 and specific markers of cellular organelles. Scale bars, . independent experiments. Data are mean ± s.d. c, Fluorescence imaging of labelled cLip-1 and LysoTracker. Scale bar, independent experiments. Data are mean ± s.d.d-f, Fluorescence imaging of labelled cLip-1 and BacMam transduced cells of GFP-labelled cell organelle markers. Scale bar, independent experiments. Data are mean ± s.d. g, Fluorescence imaging of labelled cLip-1 in proximal kidney and liver tissues of a Rosa26-CreERT2;Gpx4 mouse treated with cLip-1 and day; sacrificed 1 h after i.p.injection). Scale bar, .h, Bodipy-C11581/591flow
cytometry of cells treated with Lip-1 and RSL3 for 1 h .i, Viability of cells (top, after 72 h of incubation; middle, incubation for 24 h and cotreatment with RSL3 ; bottom, incubation for 72 h and cotreatment with 4-OH TAM to knock out . Data are mean ± s.d. j, Annexin-V/Propidium Iodide (A/PI) staining in cells treated for 24 h. RSL3: independent experiments; RSL3+Lip-1: independent experiments; RSL3 + cLip-1: independent experiments. k, NMR spectra of Lip-1 and naphthalene titrated with . NMR spectra of Lip-1 titrated with then TFA.o, NMR spectra of Lip-1 titrated with and then sodium deuteroxide (NaOD). k-o, NMR spectra recorded at 310 K in methanol- . b, d-f, 1-way ANOVA. AU, arbitrary unit.
a
Extended Data Fig. 2 | See next page for caption.
Extended Data Fig. 2 |Inactivation of lysosomal iron inhibits ferroptosis
(Part 2). a, Chemical syntheses of metcLip-1 and alcLip-1.b, Fluorescence imaging of labelled cLip-1 , metcLip-1 and alcLip-1 . Scale bar, . c, Cyclic voltammetry of a solution (reduction potentials pink arrow). Data recorded in the presence of metcLip-1 or alcLip-1. d, Bodipy-C11581/591 flow cytometry of cells treated with metcLip-1 , alcLip-1 and RSL3 for 1 h . Representative of independent experiments for PDAC053T and for HT-1080. e, FENIX assay of cLip-1, metcLip-1 and alcLip-1.f, Molecular structure of cDFO and fluorescence imaging of labelled cDFO . Scale bar, independent experiments. Data are mean ± s.d. g, Fluorescence emission spectra of RPE at
pH5 in the presence of and Lip-1.h, RPE flow cytometry of cells treated for 30 min and then cotreated with the probe for . Representative of independent experiments. i, HMRhoNox-M flow cytometry of cells treated for 30 min and then cotreated with the probe for . Representative of independent experiments. j, Bodipy-C11581/591 flow cytometry of cells treated with RSL3 (200 nM for PDAC053T, 500 nM for 4T1, 1 h ) and Baf-A1 or HCQ ( ) used 2 h prior. Representative of independent experiments. k, Fluorescence imaging of Bodipy 665/676 in cells treated with RSL3 . Scale bar, independent experiments. Two-sided unpaired -test. Data are mean ± s.d. Concentrations used unless stated otherwise: Lip-1 , Baf-A1 .
Extended Data Fig. 3 | See next page for caption.
Extended Data Fig. 3 | Ferroptosis inducers initiate lipid peroxidation in lysosomes. a, Fluorescence imaging of Bodipy 665/676 and BacMam transduced cells of GFP-labelled cell organelle markers treated with RSL3 . Scale bars, independent experiments. Data are mean s.d.b, Fluorescence imaging of Bodipy 665/676 and BacMam transduced cells of GFP-labelled cell organelle markers treated with RSL3 . Scale bars, independent experiments. Data are mean ± s.d. c, Fluorescence imaging of the lysosomal GSH probe (SQSS) and a lysosomal marker in cells treated with RSL3 . Scale bar, . Representative of independent experiments.d, SQSS flow cytometry of cells treated with ferroptosis inducers. e,1-Red flow cytometry of cells treated with ferroptosis inducers.a,b, Two-sided unpaired -test.
a
Extended Data Fig. 4 | Chemical synthesis and spectral data of Fento-1.
a, Chemical synthesis of Fento-1.b, NMR spectrum of Fento-1.c, NMR spectrum of Fento-1.d. High-resolution mass spectrum (HRMS) of Fento-1.
, Low-resolution mass spectrum (LRMS) of a Fento-1-iron complex. , Chemical
synthesis of cCW. , NMR spectra recorded at 298 K in methanol- . Full
description, Supplementary Information.
e, Low-resolution mass spectrum (LRMS) of a Fento-1-iron complex. f, Chemical synthesis of cCW.b, c, NMR spectra recorded at 298 K in methanol- . Full description, Supplementary Information.
Extended Data Fig.5|See next page for caption.
Article
Extended Data Fig. 5| Fento-1 induces the oxidation of phospholipids and cell death by activating lysosomal iron (Part 1). a, Fluorescence imaging of Fento-1, marmycin and the cell surface marker CD44 in cells treated at for 1 h . Scale bar, independent experiments. Data are mean . b, Fluorescence imaging of Fento-1 in BacMam transduced cells of GFP-labelled cell organelle markers. Scale bar, independent experiments. Data are mean ± s.d.c, Bodipy-C11581/591 flow cytometry of cells treated with Fento-1, marmycin and cCW alone or in combination for 6 h . Representative of independent experiments.d, Viability of cells treated for 6 h. Data are mean ± s.e.m. independent experiments. e, Viability of cells treated for 72 h . Data are mean s.e.m. independent experiments.f, Lipidomics of oxidised phospholipids in cells treated with Fento-1 or ferroptosis inducers for 24 h. independent experiments.g, Lipidomics of oxidised phospholipids in cells treated with Fento-1.n = 4 independent experiments. Full heatmap, Supplementary Information. h, Lipidomics of oxidised phospholipids of lysosomal extracts in cells treated with Fento- independent experiments.i, Western blot of proteins from cells treated with Fento-1,
marmycin, cCW or marmycin -tubulin is a sample loading control. Representative of independent experiments. j, Western blot of proteins from cells treated with Fento-1, marmycin, cCW or marmycin . -tubulin is a sample processing control. Representative of independent experiments with similar results. k, Bodipy-C11581/591flow cytometry of cells treated with Fento- and inhibitors used 2 h prior. independent experiments. Box plots: interquartile range, centre lines medians and whiskers the minimum and maximum values. I, Fluorescence imaging of 4-HNE treated with Fento-1(1 h). Two-sided unpaired -test. independent experiments. Data are mean ± s.d. Scale bar, , Quantification of glycerol in cells treated with Fento-1 . Data are mean s.e.m. independent experiments. Concentrations used unless stated otherwise: Fento-1 , marmycin , cCW , erastin , RSL3 , and cLip-1 . k, m, Kruskal-Wallis test with Dunn’s post-test. Phosphatidylcholine (PC) lipids displayed in the main figures. Phosphatidylethanolamine (PE), phosphatidylinositol (PI), phosphatidylserine(PS), Supplementary Information.
Extended Data Fig. 6 | See next page for caption.
Article
Extended Data Fig. 6 | Fento-1 induces the oxidation of phospholipids and cell death by activating lysosomal iron (Part 2). a, CellTiterGlo luminescence of cells treated with Fento- and inhibitors used 2 h prior. independent experiments.b, Resorufin fluorescence of cells treated with Fento-1 and inhibitors used 2 h prior. Vehicle, Lip-1: independent experiments, clip-1 and Z-VAD-FMK: independent experiments, Fer-1, Def and Toc: independent experiments, DFO, DFX and Vitamin K3: independent experiments.c, d, CellTiterGlo luminescence of cells treated with Fento-1 and inhibitors used 2 h prior. independent experiments. e, Incucyte cell viability measurement. Representative of independent experiments. Data are mean ± s.e.m.f, Viability of cells treated with Fento-1 or Lip-1 or Fento-1 and Lip-1
prior).1-way ANOVA. technical replicates.g, Fluorescence imaging of Fento-1. Fento-1 and prior .Scale bar, independent experiments. Two-sided unpaired -test. Data are mean ± s.d. h, Viability of cells treated for independent experiments. Data are mean ± s.d.i, Left:Cell death in knock-down cells measured by Annexin-V/Sytox blue in cells treated with Fento-1 for independent experiments. 2-way ANOVA. Data are mean ± s.d. Right: Western blot of proteins in knock-down cells. -tubulin is a sample processing control.j, Bodipy-C11581/591 flow cytometry of knock-down cells treated for 6 h . Box plots: interquartile range, centre lines= medians and whiskers the minimum and maximum values. a-d, Two-sided Mann-Whitney test compared to vehicle.
Extended Data Fig. 7| Fentomycin analogues. a, Molecular structures.b, Viability of cells treated for 6 h or 72 h . Data are mean ± s.d. independent experiments.
c, Bodipy-C11581/591flow cytometry of cells treated for 6 h . Representative of independent experiments.
Extended Data Fig. 8 |See next page for caption.
Extended Data Fig. 8 | Fento-1 induces ferroptosis in CD44 iron-rich cancer cells (Part 1). a, ICP-MS of healthy and cancer tissues from patients. PDAC #2: healthy tissue pieces, PDAC tissue pieces; PDAC #3: healthy tissue pieces, PDAC tissue pieces; PDAC #4: healthy tissue pieces, PDAC tissue pieces; PDAC #5: healthy tissue pieces, PDAC tissue pieces; PDAC #6: healthy tissue pieces, PDAC tissue pieces; PDAC #7: healthy tissue pieces, PDAC tissue pieces; PDAC #8: healthy tissue pieces, tissue pieces; PDAC #9: healthy tissue pieces, PDAC tissue pieces; angiosarcoma #1: healthy tissue pieces, angiosarcoma tissue pieces; angiosarcoma #2: healthy tissue pieces, angiosarcoma tissue pieces;liposarcoma #2: healthy tissue pieces, liposarcoma tissue pieces; epithelioid sarcoma: healthy tissue pieces, epithelioid sarcoma tissue pieces; PDAC liver metastasis: healthy tissue pieces, PDAC liver met astasis tissue pieces.b, ICP-MS of dissociated human tumour cancer cells. replicates.
c, HMRhoNox-M flow cytometry of cancer cells from human UPS. patients.
d, Lipidomics of oxidised phospholipids in dissociated human tumour cells treated with Fento-1 and Toc used 2 h prior. e, Lipidomics of lysophospholipids in dissociated human tumour cells treated with Fento-1 and Toc used 2 h prior. f, Bodipy-C11581/591flow cytometry of dissociated human PDAC cancer cells treated with Fento-1 ( 24 h ) and inhibitors used 2 h prior. patients.g, Bodipy-C11581/591flow cytometry of dissociated UPS cancer cells treated with Fento-1 and inhibitors used 2 h prior. patients.h, Flow cytometry cell counting of dissociated human PDAC CD44 cancer cells treated with Fento-1 and inhibitors used 2 h prior. patients. i, CD44 flow cytometry of dissociated UPS cancer cells treated with Fento- and inhibitors used 2 h prior. patients. Concentrations used unless stated otherwise: Fento-1 , , Def , Lip-1 and cLip-1 test.f-i, 1-way ANOVA. c, f-i, each coloured dot represents a tumour of a distinct patient for a given panel. Box plots: interquartile range, centre lines medians and whiskers the minimum and maximum values.
Extended Data Fig. 9 | See next page for caption.
Extended Data Fig. 9 | Fento-1 induces ferroptosis in CD44 iron-rich cancer cells (Part 2). a, Viability of cells treated with Fento-1 or standard-of-care chemotherapy for 72 h . Data are mean s.e.m. independent experiments. b, Viability of PDAC-derived organoids treated with Fento-1 or standard-of-care chemotherapy for 72 h. Data are mean ± s.e.m. IC -values are indicated. PDAC009T: Oxaliplatin, 5-FU and SN-38 independent experiments, Fento-1 independent experiments. PDAC003T: Oxaliplatin independent experiments, 5-FU and SN38 independent experiments, Fento- independent experiments. PDAC117T: Oxaliplatin, 5-FU and SN-38 independent experiments, Fento- independent experiments. PDAC372T: Oxaliplatin, 5-FU and SN-38 independent experiments, Fento- independent experiments.c, Viability of cells treated with standard-of-care chemotherapy and sublethal doses of Fento-1 .1-way ANOVA. Data are mean ± s.e.m. n = 3 independent experiments. d, Synergy score of
standard-of-care chemotherapy and Fento-1 in human primary PDAC cells. e, Viability of cells treated with Fento-1(72 h) in naive cells or cells pretreated with Doxo for 30 days, then treated with Fento- 1 for 72 h . Data are mean ± s.e.m. independent experiments.f, Bright field images of cells treated with Doxo ( days). Scale bar, . Representative of independent experiments with similar results.g, CD44 flow cytometry of cells treated with Doxo ( 25 nM ) or Fento- for 10 days. Representative of independent experiments. h, Western blot of mesenchymal and iron homeostasis markers in cells treated with Doxo ( 25 nM ) for 10 days. Lamin A/C is a sample loading control. Representative of independent experiments with similar results.i, Western blot of mesenchymal and iron homeostasis markers in cells treated with Fento- for 10 days. -tubulin is a sample loading control. Representative of independent experiments with similar results. Doxo, doxorubicin.
Extended Data Fig. 10 | Fento-1 induces ferroptosis in CD44 iron-rich cancer cells (Part 3). a, ICP-MS of blood, serum and lymphatic fluid from mice. Blood and serum: mice; Lymph: mice.1-way ANOVA.b, HMRhoNox-M flow cytometry of dissociated mouse tumour cancer cells. mice. Each coloured dot represents a tumour of a distinct mouse. Two-sided MannWhitney test.c, Tumour diameter in mice treated with Fento-1 per animal every-other-day). 2-way ANOVA. Data are mean ± s.e.m. experiment: vehicle mice, Fento- mice; experiment: mice for each group; experiment: mice for each group. d, Tumour diameter in tumour-bearing mice treated with Fento-1 compound per animal every-other-day). independent experiments. experiment: mice for
each group; experiment vehicle: mice, Fento- mice; experiment: mice for each group; experiment: mice for each group.e, Mice body weight of mice treated as in Fig. 3h and Extended Data Fig.10c.1 experiment: mice for each group; experiment: vehicle mice, Fento- mice; experiment: mice for each group; experiment: mice for each group. Data are mean ± s.e.m.f, Lipidomics of lysophospholipids in tumours of tumour-bearing mice treated with Fento-1 as in Fig. 3j. 4T1 tumours. LPC lipids displayed. LPE, LPS, Supplementary Information.Box plots: interquartile range, centre lines medians and whiskers the minimum and maximum values.
natureportfolio
Corresponding author(s): Raphaël RODRIGUEZ
Last updated by author(s): Mar 7, 2025
Reporting Summary
Nature Portfolio wishes to improve the reproducibility of the work that we publish. This form provides structure for consistency and transparency in reporting. For further information on Nature Portfolio policies, see our Editorial Policies and the Editorial Policy Checklist.
Statistics
For all statistical analyses, confirm that the following items are present in the figure legend, table legend, main text, or Methods section.
n/a
□ X
□
□
□
□
A description of all covariates tested
□
□
□
□
Confirmed
The statistical test(s) used AND whether they are one- or two-sided Give values as exact values whenever suitable.
□
□
The exact sample size for each experimental group/condition, given as a discrete number and unit of measurement
A statement on whether measurements were taken from distinct samples or whether the same sample was measured repeatedly
Only common tests should be described solely by name; describe more complex techniques in the Methods section.
A description of any assumptions or corrections, such as tests of normality and adjustment for multiple comparisons
A full description of the statistical parameters including central tendency (e.g. means) or other basic estimates (e.g. regression coefficient) AND variation (e.g. standard deviation) or associated estimates of uncertainty (e.g. confidence intervals)
For null hypothesis testing, the test statistic (e.g. ) with confidence intervals, effect sizes, degrees of freedom and value noted
For Bayesian analysis, information on the choice of priors and Markov chain Monte Carlo settings
For hierarchical and complex designs, identification of the appropriate level for tests and full reporting of outcomes
Estimates of effect sizes (e.g. Cohen’s , Pearson’s r), indicating how they were calculated
Software and code
Policy information about availability of computer code
Attune NxT using Attune NxT Sovtware v4.2.0
FACSAria Fusion (BD) using FACS DIVA software v9.0.1
BD LSR Fortessa X-20 flow cytometer using FACS DIVA software v9.0.1
Western blot:
FusionCapt Advance Software
Plate reader:
SpectraMax ID3 plate reader (Molecular Devices), exported as Excel files
SpectraMax iD5 microplate reader with SoftMax Pro v7 software (Molecular Devices)
NMR:
Bruker Topspin 4.1.4 software
nature portfolio | reporting summary
April 202
Cyclic Voltammetry:
Nova software
Fluorimetry:
SoftMax Pro 7.1 GxP
Molecular modeling:
Gaussian 16, Revision C. 01
Lipidomics:
LipidXplorer v 1.2.8 (http://genomebiology.com/2011/12/1/R8)
FENIX assay:
Agilent BioTek Gen5 v. 3.08.01
Data analysis
Softwares:
Microsoft Excel and PRISM 10.0.3 unless stated otherwise
NMR:
MestRenova v.5.0.1-35756
Bruker Topspin 4.1.4 software
Drug synergy:
SynergyFinder software
For manuscripts utilizing custom algorithms or software that are central to the research but not yet described in published literature, software must be made available to editors and reviewers. We strongly encourage code deposition in a community repository (e.g. GitHub). See the Nature Portfolio guidelines for submitting code & software for further information.
Data
Policy information about availability of data
All manuscripts must include a data availability statement. This statement should provide the following information, where applicable:
Accession codes, unique identifiers, or web links for publicly available datasets
A description of any restrictions on data availability
For clinical datasets or third party data, please ensure that the statement adheres to our policy
Lipidomics data are presented in Supplementary Tables. Quantitative proteomics raw data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD054449 (reviewer_pxd054449@ebi.ac.uk; password: o1gh7kRYdTEP). For identification, the data were searched against the Homo sapiens (UP000005640) Uniprot database using Spectronaut v18.7 or 19 (Biognosys).
Research involving human participants, their data, or biological material
Policy information about studies with human participants or human data. See also policy information about sex, gender (identity/presentation), and sexual orientation and race, ethnicity and racism.
Reporting on sex and gender
Reporting on race, ethnicity, or
No gender or sex reporting was performed for biopsies of human tumours. Tumour samples collected were anonimised for patient and data protection purposes in accordance with hospital guidelines and the French law and not analysed by sex.
Tumour samples were anonymous and no reporting of race, ethnicity or other socially relevant groupings were performed in accordance with French law.
other socially relevant
groupings
Population characteristics
Recruitment
Ethics oversight
Pancreatic ductal adenocarcinoma (PDAC), undifferentiated pleomorphic sarcoma (UPS), liposarcoma, angiosarcoma, epitheloid sarcoma, PDAC liver metastasis were obtained as anonymous samples.
No selection of the participants
Fresh tumour samples were obtained by surgery from patients at Paul Brousse and Institut Curie care centres. All patients provided written informed consent for use of tumour samples. The study was approved by institutional regulatory boards (no. 587 and DATA190160).
Note that full information on the approval of the study protocol must also be provided in the manuscript.
Field-specific reporting
Please select the one below that is the best fit for your research. If you are not sure, read the appropriate sections before making your selection.
Life sciences
Behavioural & social sciences □ Ecological, evolutionary & environmental sciences
For a reference copy of the document with all sections, see nature.com/documents/nr-reporting-summary-flat.pdf
Life sciences study design
All studies must disclose on these points even when the disclosure is negative.
Sample size
No sample size calculation was performed, but a minimum of chosen for cell biology experiments. For in vivo experiments, mouse cohort groups were either 5 or 10 . No sample size calculation was performed for human tumour samples, and analyses were performed on availability of these rare samples post-surgery, depending on availability from the patients.
Data exclusions
No data were excluded from the analyses.
Replication
Experiments were performed at least three times unless stated otherwise. Replication was successful and all data are displayed in the manuscript. All replicates of all experiments noted in the captions and/ or the statistical analysis, with data available as source files.
Randomization
For in vivo experiments, mice were allocated randomly into experimental groups. For tumour samples, these were grouped by tumour type (PDAC, UPS etc.) and not randomized. Material was available from the hospital and collected in an anonymous fashion for the analysis, other than the indication of the type of tumour.
Blinding
Investigators were blinded to group allocation during data collection for ICP-MS, lipidomics and proteomics experiments. Data analyses were not blinded.
Reporting for specific materials, systems and methods
We require information from authors about some types of materials, experimental systems and methods used in many studies. Here, indicate whether each material, system or method listed is relevant to your study. If you are not sure if a list item applies to your research, read the appropriate section before selecting a response.
Materials & experimental systems
Methods
n/a
Involved in the study
n/a
Involved in the study
□
– Antibodies
X
□
□
– Eukaryotic cell lines
□
– Flow cytometry
–
□
V
□
□
【 Animals and other organisms
X
□
–
□
X
□
Antibodies
Antibodies used
Antibodies are annotated below as follows. WB, Western blot; FCy, flow cytometry; FI, fluorescence imaging. Hu, used for human samples. Ms, used for mouse samples. Dilutions are indicated. Any antibody validation by manufacturer is indicated and can be found on the manufacturers’ websites. Our antibody validation knockdown (KD) and/or knockout (KO) strategies for relevant antibodies are indicated.
Validation
Any antibody validation by the manufacturer is indicated and can be found on the manufacturers’ websites. Our antibody validation knockdown (KD) and/or knockout (KO) strategies performed in our laboratory for relevant antibodies are indicated above an were performed and verified in the laboratory with relevant CRISPR or siRNA approaches (denoted as in-house in the list above). Information of antibody validation by manufacturers are indicated in thee list above. All this information is also detailed in the methods section.
Eukaryotic cell lines
Policy information about cell lines and Sex and Gender in Research
Cell line source(s)
Authentication
Mycoplasma contamination
Commonly misidentified lines (See ICLAC register)
HT-1080 cells, MDA-MB-231 and 4T1 cells were obtained from the ATCC. FC1242 and FC1245 murine pancreatic cancer cells, 4a cells and human pancreatic hMIA-2D cells were a gift from the Tuveson laboratory (CSHL).
Primary human pancreatic PDAC024T, PDAC030T, PDAC053T, PDAC054T, PDAC084T, PDAC090T and PDAC211T cells were obtained as described in the “Establishment of xenograft derived pancreatic cell cultures” paragraph.
Xenograft derived pancreatic organoids (XDPO) PDAC009T, PDAC003T, PDAC117T and PDAC372T were obtained as described in the “Establishment of xenograft derived pancreatic organoids” paragraph.
Primary lung circulating tumour cells were from Celprogen.
Primary colon circulating tumour cells were from Celprogen.
Pfa1 cells were generated in the Conrad laboratory (Seiler A et al., 2008).
SUM159 cells were a genrous gift from Dr. S.Ethier (Karmanos Cancer Center, Detroit, MI, USA).
Authentication was relied on by the commercial source where applicable (ATCC, Celprogen).
No authentication of human derived pancreatic cancer cells/ organoids was performed. These cells were generated from primary sources directly in the lovanna lab. Pfa1 cells and derivatives were generated in the Conrad lab (Seiler A et al., 2008) and SUM159 cells were a generous gift from Dr. S.Ethier (Karmanos Cancer Center, Detroit, MI, USA). These cells, and the generously gifted cells from the Tuveson laboratory (see above) were not further authenticated.
Cells were regularly tested negative for mycoplasma contamination.
No commonly misidentified cell lines were used in this study.
Animals and other research organisms
Policy information about studies involving animals; ARRIVE guidelines recommended for reporting animal research, and Sex and Gender in Research
Laboratory animals
cLip-1 treatment in vivo:
Mice were kept under standard conditions with water and food ad libitum and in a controlled environment humidity, 12 h light/dark cycle). For animal studies, Rosa26-CreERT2;Gpx4f/f (C57BL6/J) mice were randomised into separate cages. 12 to 24 -week-old sex-matched mice were used for all experiments.
Isolation of blood, serum and lymphatic fluid from mice:
Balb/C mice (25-week-old adult female mice) were purchased from Charles River and grown at the CRCM animal core facility and randomised into separate cages. Mice were housed under sterile conditions with sterilised food and water provided ad libitum and maintained on a 12 h light/dark cycle and humidity.
Intranodal murine metastasis models and Fento-1/Marmycin/CW treatment:
Murine breast cancer cells (4T1) were transplanted into 6 to 8 -week-old female Balb/c mice (syngeneic with the 4T1 model). Mice were grown at the Harvard medical school animal core facility and randomised into separate cages. Mice were housed under sterile conditions with sterilised food and water provided ad libitum and maintained on a 12 h light/dark cycle and humidity.
Wild animals
Reporting on sex
Field-collected samples
Ethics oversight
The study did not involve wild animals.
As detailed in the Methods.
The study did not involve samples collected from the field.
All cLip-1 in vivo experiments were performed in compliance with the German Animal Welfare Law and have been approved by the Institutional Committee on Animal Experimentation and the Government of Upper Bavaria (approved no. ROB-55.2-2532.Vet_02-18-13). All intranodal injection mouse experiments complied with all relevant ethical regulations and were performed according to protocols approved by the Institutional Animal Care and Use Committee at Harvard T.H. Chan School of Public Health (protocol ISOOOO3460). For mouse lymph and blood collection, animal experiments were performed in accordance with the European Community guiding in the care and use of animals. Animal experiments were performed in agreement with the French Guidelines for animal handling and approved by local ethics committee (Agreement no. 16487-2018082108541206 v3).
Note that full information on the approval of the study protocol must also be provided in the manuscript.
Plants
Seed stocks
Report on the source of all seed stocks or other plant material used. If applicable, state the seed stock centre and catalogue number. If plant specimens were collected from the field, describe the collection location, date and sampling procedures.
Novel plant genotypes
Describe the methods by which all novel plant genotypes were produced. This includes those generated by transgenic approaches, gene editing, chemical/radiation-based mutagenesis and hybridization. For transgenic lines, describe the transformation method, the number of independent lines analyzed and the generation upon which experiments were performed. For gene-edited lines, describe the editor used, the endogenous sequence targeted for editing, the targeting guide RNA sequence (if applicable) and how the editor was applied.
Authentication
Flow Cytometry
Plots
Confirm that:
The axis labels state the marker and fluorochrome used (e.g. CD4-FITC).
The axis scales are clearly visible. Include numbers along axes only for bottom left plot of group (a ‘group’ is an analysis of identical markers).
All plots are contour plots with outliers or pseudocolor plots.
A numerical value for number of cells or percentage (with statistics) is provided.
Methodology
Sample preparation
Flow cytometry:
Cells were washed with ice-cold PBS. For antibody staining, cells were incubated with Fc block (Human TruStain FcX, Biolegend, 422302, 1:20) for 15 min , then incubated with antibodies in ice cold FBS, PBS, 2 mM EDTA for 20 min at 4 and then washed with PBS and resuspended in FBS, PBS, 2 mM EDTA before analysis using a BD LSR Fortessa
X-20 flow cytometer. Flow cytometry with iron probes: PDAC053T and HT-1080 cells were seeded in 6 -well plates at the density of cells/well. On the following day, Lip-1 , hydroxychloroquine (HCQ, Sigma-Aldrich, ), Bafilomycin A1 (Baf-A1, Sigma-Aldrich, B1793, 75 nM ) were added to PDAC053T for 1 h and in HT-1080 for 30 min . In PDAC053T, after 30 min of treatment with compounds, RPE (in-house, ) or HMRhoNox-M (in-house, ) probes were added for 30 min . In HT-1080, after 15 min of treatment with compounds, RPE (in-house, ) or HMRhoNox-M (inhouse, ) probes were added and left for 15 min . After incubation with iron probes, the media was removed and cells were washed with PBS once before trypsinization. Cells were harvested, pelleted, washed with PBS and finally of PBS buffer containing 10% FBS and EDTA ( ) was added. Data were recorded on a BD LSR Fortessa X-20. Flow cytometry with Bodipy-C11 581/591: PDAC053T cells were seeded in 6 -well plates at the density of cells/well. On the next day, cells were treated with Lip-1 , cLip-1 (in house, ), metclip-1 (in house, ), alcLip-1 (in house, 10 and Bodipy-C11 before adding RSL3 . After 1 h , the media was removed and cells were washed with PBS twice before trypsinization. Cells were harvested, pelleted, washed with PBS and finally of PBS buffer containing 10% FBS and EDTA ( ), was added for flow cytometry. PDAC053T, HT-1080 and 4T1 cells were seeded in 6 -well plates at the density of cells/well. On the next day, cells were treated with Baf-A1 ( 75 nM ) and HCQ for 2 h before adding RSL3 ( 200 nM for PDAC053T and HT-1080, 500 nM for 4T1). Then after 1 h , cells were treated with Bodipy-C11 for an additional hour. The media was removed and cells were washed with PBS twice before trypsinization. Cells were harvested, pelleted, washed with PBS and finally of PBS buffer containing FBS and EDTA ( ), was added for flow cytometry. Data were recorded on an AttuneTM NxT Flow Cytometer (Thermo Fisher Scientific). Flow cytometry analyses of lysosomal GSH and lysosomal hydroxyl radicals: Cells were incubated with SQSS (in house, ) or 1-Red (in house, ). HT-1080 cells were incubated with RSL3 , Sigma-Aldrich, SML0521), FIN56 ( , Sigma-Adrich, SML1740), Buthionine sulfoximine (BSO, , Sigma-Aldrich, B2515) or erastin , Sigma-Aldrich, 329600) for the indicated time points. Data were recorded on an AttuneTM NxT Flow Cytometer (Thermo Fisher Scientific). Flow cytometry analyses of lysosomal iron content: Typically dissociated human tissue cells were incubated in media (RPMI 1610, 10% FBS, penicillin/streptomycin) with HMRhoNox-M (in-house, 1 . Lysosomal iron content of human tumour samples and healthy adjacent tissues was analysed with the following antibody and stain panel: DAPI ( ), CD3-BV510 (BioLegend, 317332), CD31-PE-Cy7 (BioLegend, 303118), CD44AF647 (Novus Biologicals, NB500-481AF647), CD45-BV785 (BioLegend, 304048), CD163-PerCP/Cyanine5.5 (BioLegend, 326512), FAP-AF700 (R&D Systems, FAB3715N), TfR1-APC-AF750 (Beckman Coulter, A89313). The live tumour cells corresponded to DAPIneg/CD45neg/CD31neg/FAPneg cells. Data were recorded on a BD LSRFortessa X-20. Flow cytometry analyses of CD44 in Fento-1 treated tumour samples: Typically dissociated tumour cells were incubated in media (RPMI 1610, 10% FBS, penicillin/streptomycin) with Fento-1 (in house, ). Cells were pre-treated with Lip-1 , cLip-1 (in house, ), Toc or Def for 2 h . Toc was kept pure under inert atmosphere and a fresh stock solution was prepared throughout the study before each experiment. The following antibody and stain panel was used for subsequent flow analysis: SYTOX blue (Thermo Fisher Scientific, S34857, ), CD31-BV605 (BioLegend, 303122), CD45BV510 (BioLegend, 368526, lot B373428), CD44-AF647 (Novus Biologicals, NB500-481AF647), TfR1-PE (BioLegend, 334106), FAP-AF750 (Novus Biologicals, FAB3715S-100UG). The live tumour cells corresponded to SYTOX blueneg/CD45neg/CD31neg/ FAPneg cells. Data were recorded on an AttuneTM NxT Flow Cytometer (Thermo Fisher Scientific). For flow cytometry analyses of CD44 levels in mouse 4T1 tumours, typically dissociated tumour cells were used per condition. Freshly dissociated cells were stained using the following antibody and stain panel: SYTOX blue (Thermo Fisher Scientific, S34857, 1 , CD31-BV605 (BioLegend, 102427), CD44-AF647 (BioLegend, 103018), CD45-BV510 (BioLegend, 103138), MHCII-APC/ Cyanine7 (BioLegend, 107628). The live tumour cells corresponded to SYTOX blueneg/CD45neg/CD31neg/MCH IIpos cells. Data were recorded on an AttuneTM NxT Flow Cytometer (Thermo Fisher Scientific). All data were analysed with FlowJo software v. 10.10.0.
Fluorescence-activated cell sorting:
Sorting of human cells was performed using the following antibodies: CD31-PE-Cy7 (BioLegend, 303118), CD44-AF647 (Novus Biologicals, NB500-481AF647), CD45-BV785 (BioLegend, 304048). The sorted cells corresponded to CD45neg/CD31neg/ CD44pos cells and CD45neg/CD31neg/CD44neg cells and were isolated on a FACSAria Fusion (BD). ICP-MS experiments were conducted in CD44high and CD44low tumour cells as described in the ICP-MS section. Sorting of mouse cells was performed using the following antibodies: CD44-AF647 (Biolegend, 103018), MHCII-APC/Cyanine7 (BioLegend, 107628). The sorted cells corresponded to MHCIIpos/CD44high cells and MHCIIpos/CD44low cells. Sorted cells were centrifuged at and cell pellets were processed for subsequent applications.
Cell sorting allows to obtain a satisfactory purity
The live human tumour cells corresponded to DAPIneg/CD45neg/CD31neg/FAPneg cells The live murine tumour cells corresponded to SYTOX blueneg/CD45neg/CD31neg/MCH IIpos cells. The sorted human cells corresponded to CD45neg/CD31neg/CD44pos cells and CD45neg/CD31neg/CD44neg cells. The sorted murine cells corresponded to MHCllpos/CD44pos cells and MHCllpos/CD44neg cells.
Tick this box to confirm that a figure exemplifying the gating strategy is provided in the Supplementary Information.
Institut Curie, CNRS, INSERM, PSL Research University, Paris, France. Department of Molecular Metabolism, Harvard T. H. Chan School of Public Health, Boston, MA, USA. Department of Genetics, Institut Curie, Paris, France. Paris Saclay University, UVSQ, Montigny-le-Bretonneux, France. Institute of Metabolism and Cell Death, Helmholtz Zentrum München, Neuherberg, Germany. Institut Curie, INSERM, Mines Paris Tech, PSL Research University, Paris, France. Institut Curie, PSL Research University, Paris, France. Ecole Normale Supérieure, CNRS, PSL Research University, Paris, France. CRCM, CNRS, INSERM, Institut Paoli-Calmettes, Aix-Marseille University, Marseille, France. CRCM, CNRS, INSERM, Institut Paoli-Calmettes, Aix-Marseille University, Parc Scientifique et Technologique de Luminy, Marseille, France. Department of Digestive and Medical Oncology, Paris Saclay University and Paul-Brousse Hospital (APHP Sud), Villejuif, France. Department of Surgical Oncology, Institut Curie, Paris, France. Department of Medical Oncology, Institut Curie, INSERM, PSL Research University, Paris, France. Institut de Chimie Moléculaire et des Matériaux d’Orsay, CNRS, Paris Saclay University, Orsay, France. Department of Chemistry and Biomolecular Sciences, University of Ottawa, Ottawa, Ontario, Canada. Rudolf-Virchow-Zentrum, Center for Integrative and Translational Bioimaging, Julius-Maximilians-Universität Würzburg, Würzburg, Germany. Department of Chemistry, Columbia University, New York, NY, USA. Department of Biological Sciences, Columbia University, New York, NY, USA. Department of Pathology and Cell Biology, Vagelos College of Physicians & Surgeons, Columbia University, New York, NY, USA. Translational Redox Biology, TUM Natural School of Sciences, Technical University of Munich, Garching, Germany. These authors contributed equally: Tatiana Cañeque, Leeroy Baron, Sebastian Müller. e-mail: raphael.rodriguez@curie.fr
Dixon, S. J. & Olzmann, J. A. The cell biology of ferroptosis. Nat. Rev. Mol. Cell Biol. 25, 424-442 (2024).