DOI: https://doi.org/10.1038/s41467-023-44464-9
PMID: https://pubmed.ncbi.nlm.nih.gov/38172570
تاريخ النشر: 2024-01-03
تنويع دورة المحاصيل يزيد من إنتاج الغذاء، يقلل من انبعاثات غازات الدفيئة الصافية ويحسن صحة التربة
تم القبول: 14 ديسمبر 2023
نُشر على الإنترنت: 03 يناير 2024
(أ) التحقق من التحديثات
الملخص
تواجه الإنتاجية الغذائية العالمية تحديات في تحقيق التوازن بين الحاجة إلى زيادة الغلات والاستدامة البيئية. تقدم هذه الدراسة تجربة ميدانية لمدة ست سنوات في سهل شمال الصين، توضح فوائد تنويع الزراعة الأحادية التقليدية للحبوب (القمح والذرة) مع المحاصيل النقدية (البطاطا الحلوة) والبقوليات (الفول السوداني وفول الصويا). تزيد الدورات المتنوعة من الغلة المعادلة بنسبة تصل إلى
لقد أصدرت الأنظمة أيضًا كميات كبيرة من غازات الدفيئة (GHG) وتسببت في تدهور البيئة
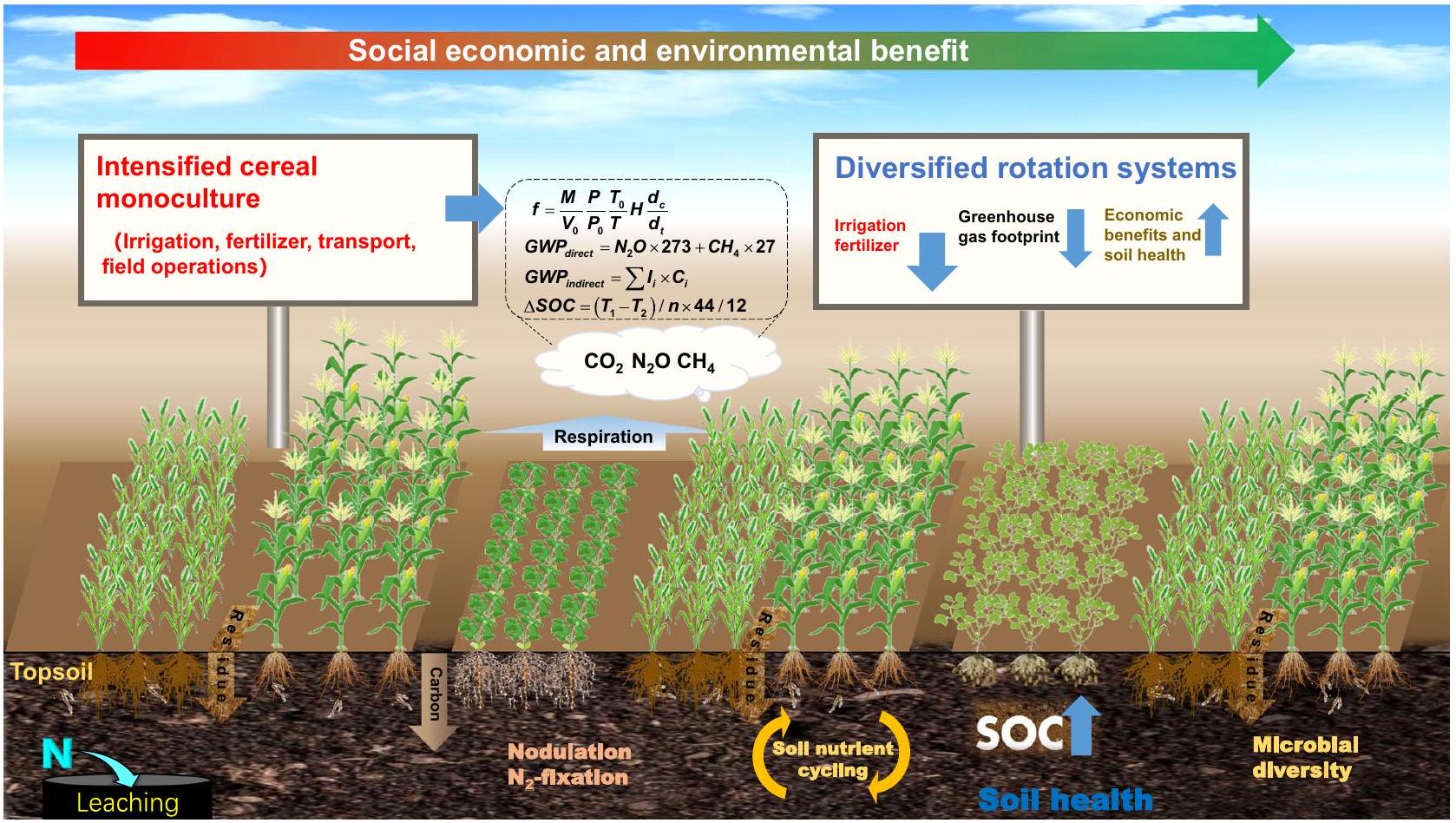
الفوسفور المتاح (AP)، الكربون في الكتلة الحيوية الميكروبية (MBC)، النيتروجين في الكتلة الحيوية الميكروبية (MBN)، وتركيب وتنوع المجتمع الميكروبي. هذه القياسات مكنتنا من اختبار الفرضيات التالية: (أ) أن نظام المحاصيل النقدية المتنوع يزيد من صافي دخل المزارعين دون jeopardizing غلات المحاصيل، (ب) أن الأنظمة المتنوعة من البقوليات تقلل من انبعاثات غازات الدفيئة على مستوى الحقل، و (ج) أن دمج الدورات المتنوعة يزيد من إنتاج الغذاء، ويقلل من انبعاثات غازات الدفيئة، ويعود بالنفع على صحة التربة (الشكل 1). تظهر نتائجنا أن (أ) النتائج الإرشادية من الأنظمة المتنوعة التي تم تصميمها واختبارها والتحقق من صحتها حديثًا يمكن أن توجه سهل شمال الصين في إنشاء نظام أكثر استدامة للحفاظ على أو زيادة إنتاج الحبوب والبروتين مع تقليل الأضرار على البيئة ونظم التربة البيئية، و (ب) أن النتائج من مثل هذه المنطقة المنتجة للغذاء بشكل مكثف قد توفر دليلاً للدول/المناطق ذات البيئات الزراعية المماثلة لتتبعها على نطاق أوسع.
النتائج
تزيد دورات المحاصيل المتنوعة من إنتاجية النظام البيئي
يمكن لمحاصيل البقوليات الحفاظ على غلات المحاصيل، وزيادة دخل المزارعين، وتقليل انبعاثات غازات الدفيئة بسبب العملية البيولوجية
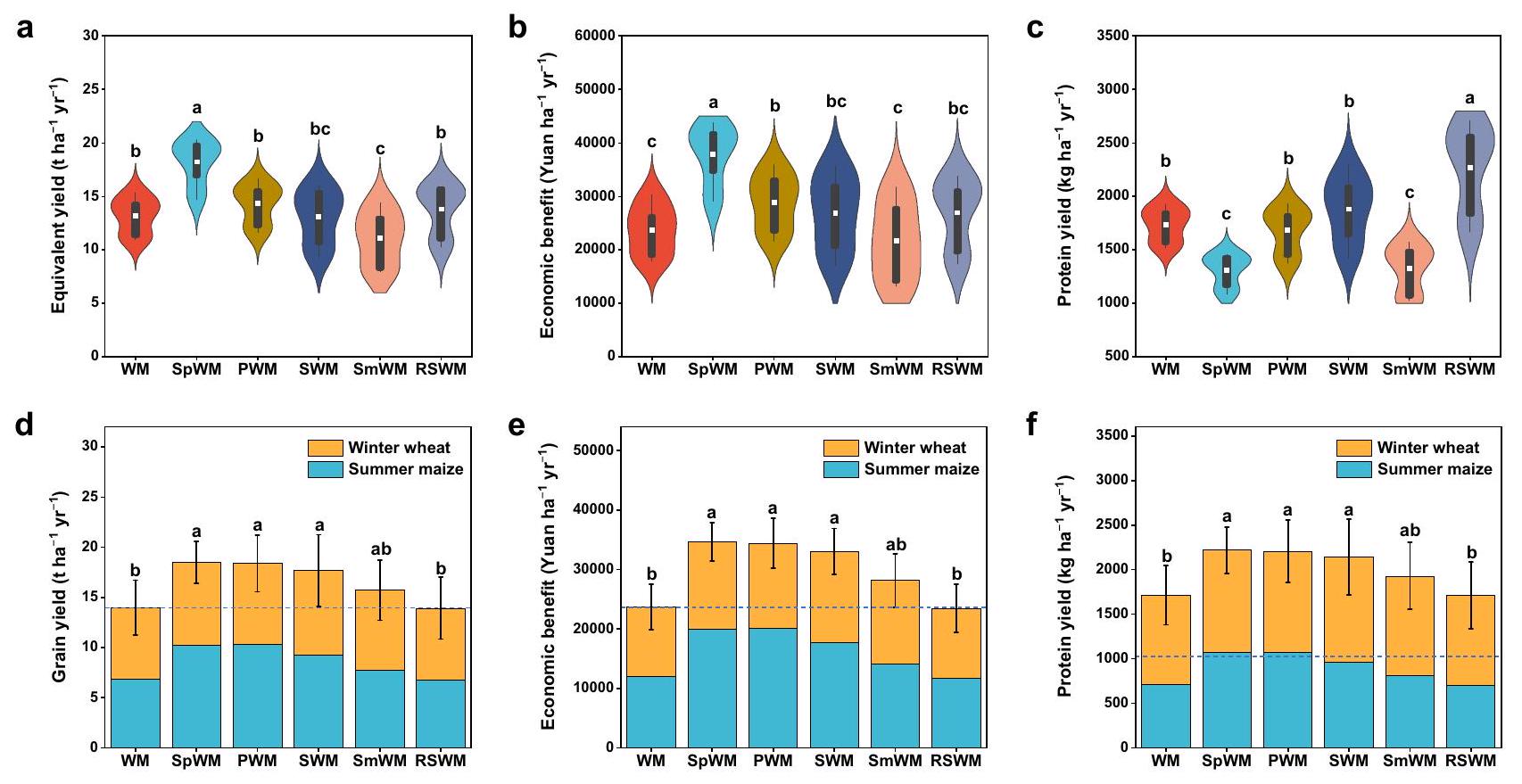
تفاوت العائد بين أنظمة الدوران، مع دوران الجاودار (Lolium multiflorum L.)-الدخن (Sorghum bicolor L.) (RSWM) وفول الصويا
تقلل دورات المحاصيل المتنوعة من انبعاثات غازات الدفيئة الصافية
التحكم الذي كان في
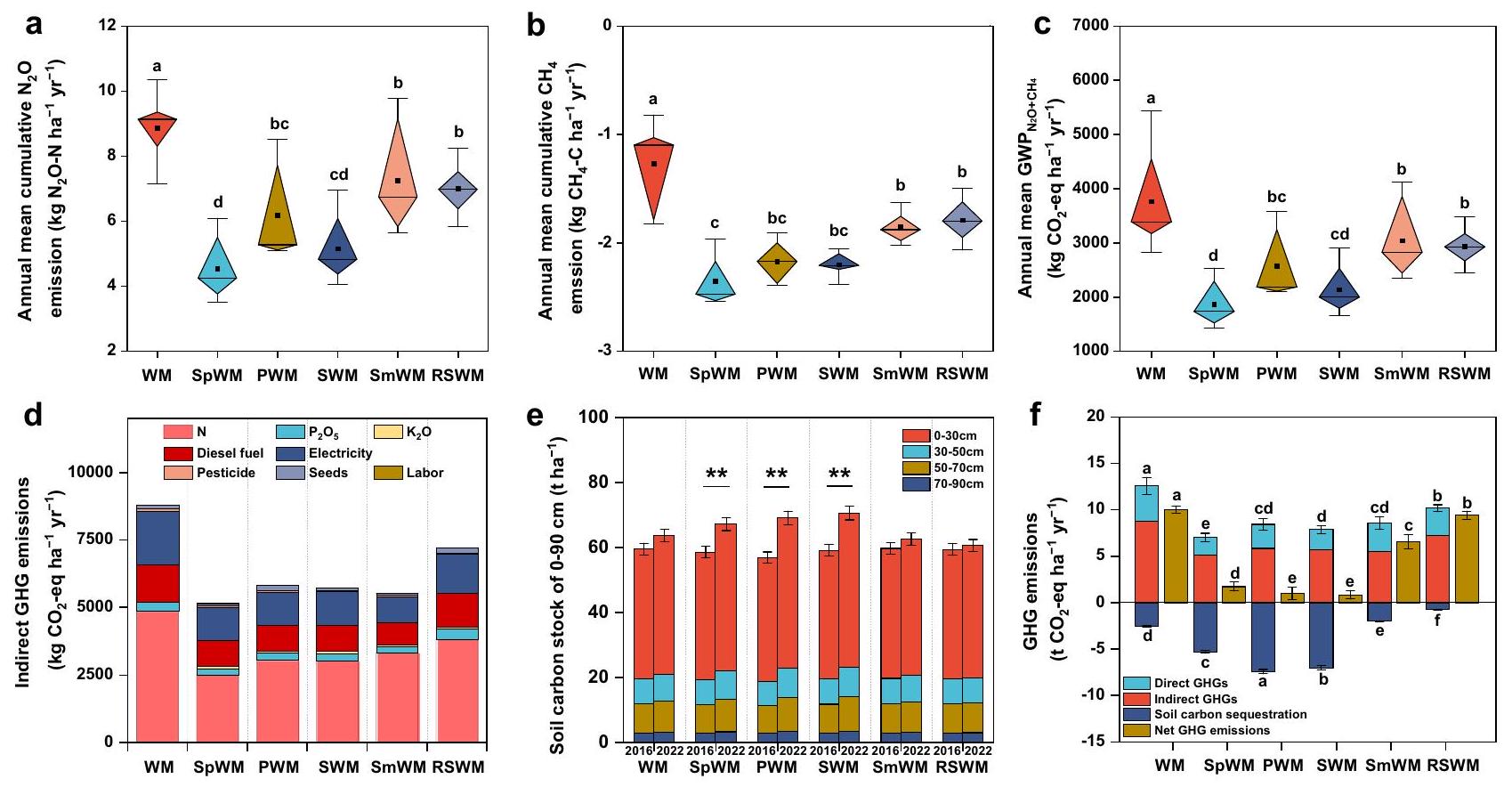
بين الدورات في
انبعاثات بواسطة
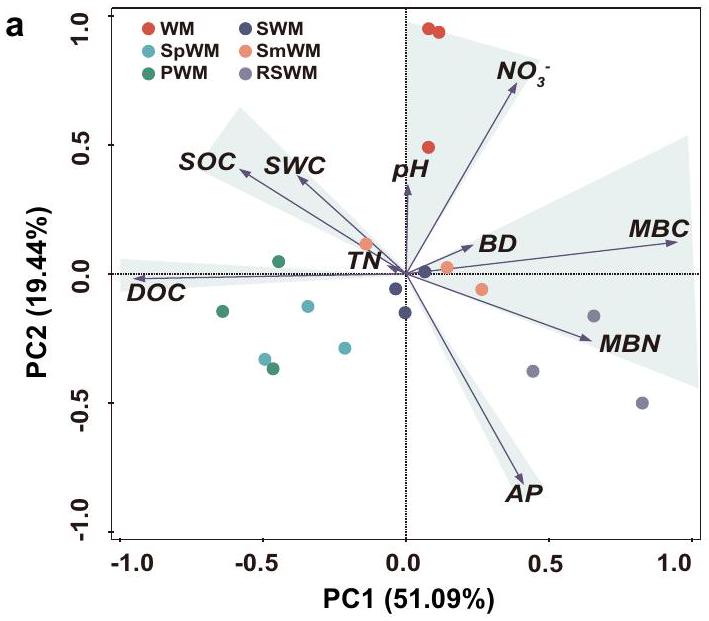
تساهم دورات المحاصيل المتنوعة في تعزيز صحة التربة وتنوع الميكروبات
دُخن
معقد، لكن كان من الواضح أن تركيبة المجتمعات البكتيرية والفطرية قد تغيرت من 2016 إلى 2022. كانت الدورات مع البطاطا الحلوة والفول السوداني وفول الصويا لها تركيبات مجتمعية بكتيرية مميزة عن الأنظمة الثلاثة الأخرى (الشكل 5e)، في حين أن الدورات المتنوعة الخمس المدروسة كانت لها تركيبات مجتمعية فطرية مختلفة بشكل ملحوظ عن التحكم في القمح والذرة (الشكل 5f). أثرت 6 سنوات من التغذية الدورية على الخصائص الفيزيائية والكيميائية للتربة بشكل تفاعلي مع المجتمعات الميكروبية في التربة. كانت المجتمعات البكتيرية للدورات مع البطاطا الحلوة والفول السوداني وفول الصويا مرتبطة ارتباطًا وثيقًا بالكربون العضوي في التربة والفوسفات المتاح، في حين أن تلك الخاصة بالقمح والذرة الربيعية…
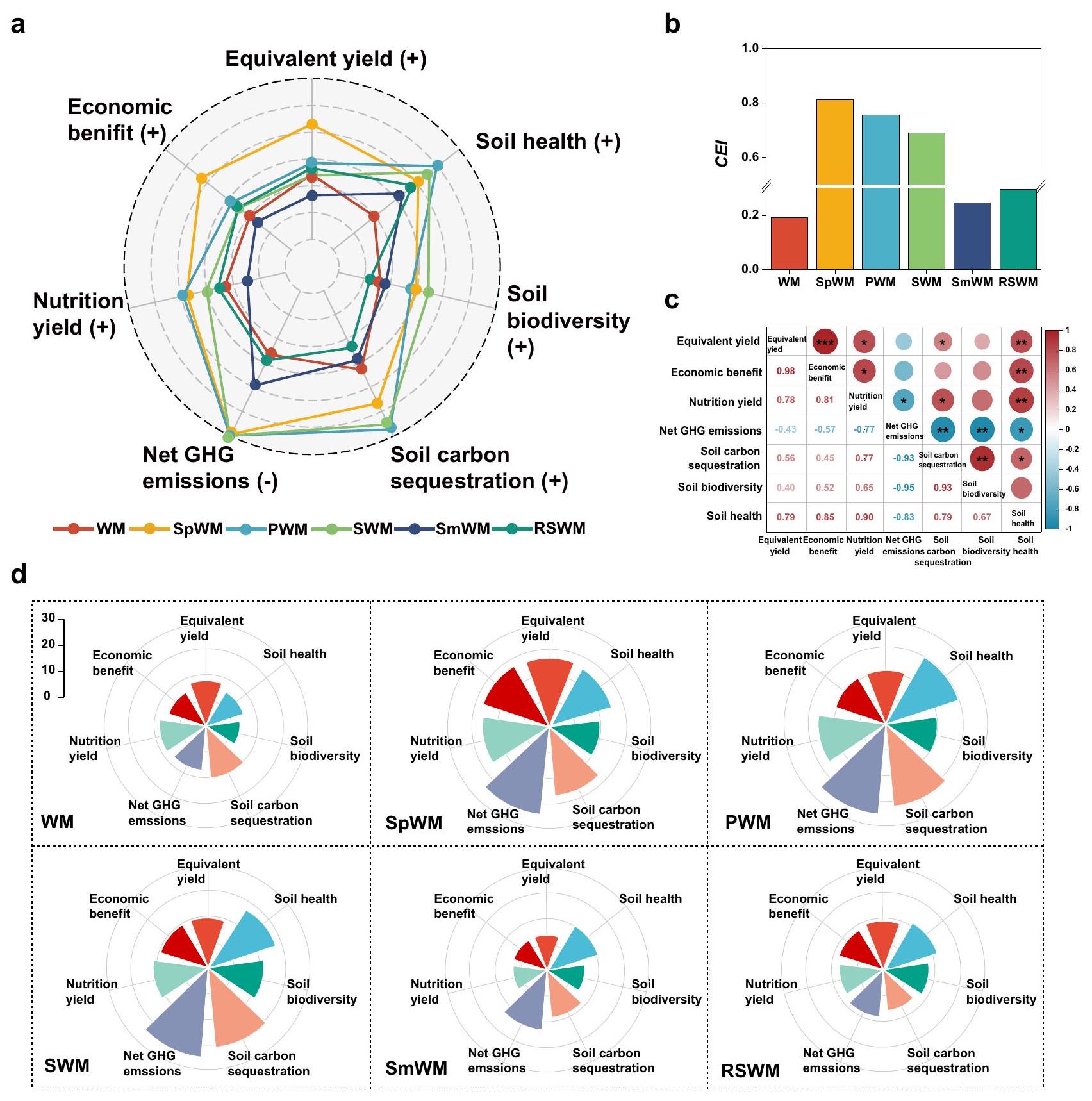
تقييمات متعددة الوظائف لدورات المحاصيل المتنوعة
نقاش
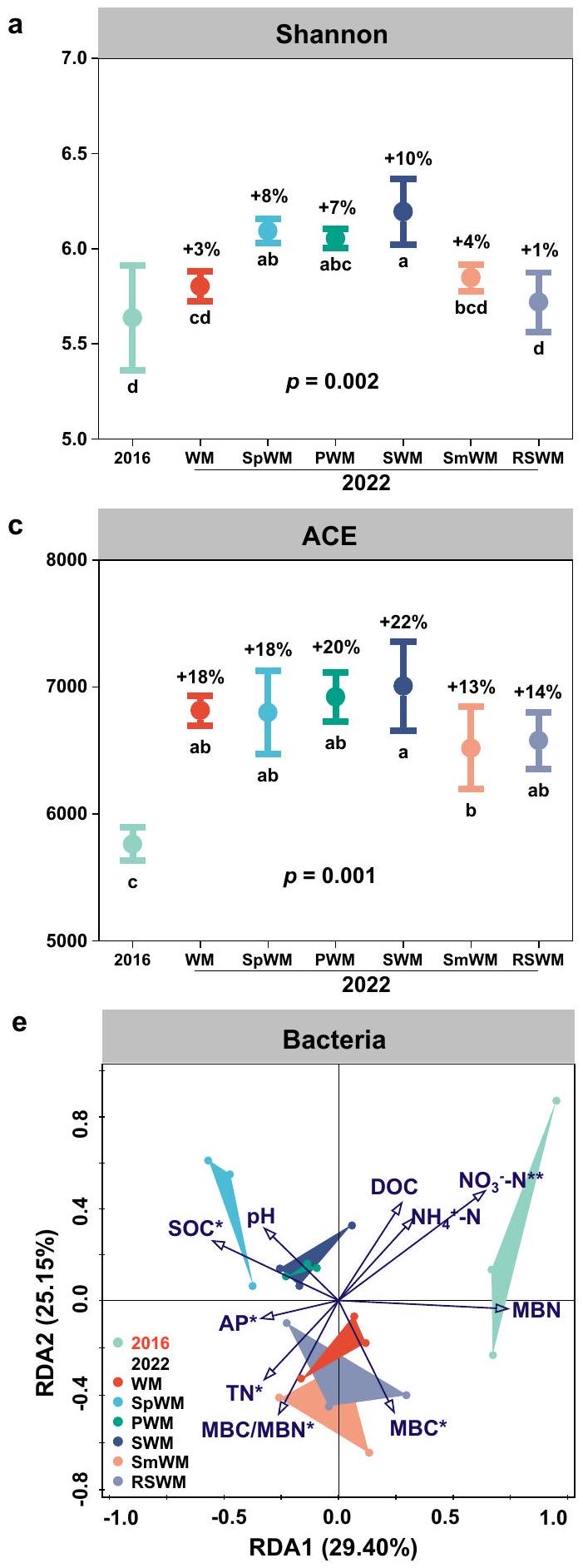
تشير إلى خصائص التربة بما في ذلك الكربون العضوي في التربة (SOC) ، الكربون العضوي المذاب (DOC) ، النيتروجين الكلي (TN) ،
مخزون الكربون في التربة في طبقة 0-90 سم من أكتوبر 2016 إلى أكتوبر 2022؛ تشير انبعاثات غازات الدفيئة الصافية إلى ميزانيات غازات الدفيئة الصافية بما في ذلك الانبعاثات المباشرة وغير المباشرة لغازات الدفيئة مطروحًا منها تخزين الكربون في التربة؛ يتم تمثيل تنوع التربة بواسطة مؤشر شانون لمجتمعات البكتيريا والفطريات؛ يتم حساب درجة صحة التربة من 10 مؤشرات باستخدام مصفوفة تقييم صحة التربة في كورنيل (CSHA) مجتمعة مع تحليل المكونات الرئيسية (PCA).
فوائد
لقد زادت البطاطا والفول السوداني وفول الصويا بشكل كبير من كربون التربة بعد 6 سنوات من تغذية الدورة الزراعية مقارنة بنظام زراعة القمح الشتوي والذرة الصيفية التقليدي. لقد أظهر إدخال البطاطا الحلوة والبقوليات في هذه الدورات الزراعية تحفيز نمو الميكروبات وزيادة كفاءة استخدام الكربون.
مصدر للمنتجات الميكروبية التي تؤثر على الكربون العضوي المستقر بقدرة عالية على استقرار الكربون
زيادة تنوع المجتمع الفطري مقارنة بالقمح الشتوي والذرة الصيفية، مما يشكل ملفات مغذيات التربة مع ركائز كربونية مختلفة أثرت على تنوع الميكروبات. علاوة على ذلك، فإن دورات المحاصيل المتنوعة قد تقلل من تلوث التربة بالمواد الكيميائية الزراعية بسبب انخفاض استخدام الأسمدة النيتروجينية الاصطناعية والمبيدات الحشرية، مما يفضل البيئات الميكروبية في التربة.
طرق
العائد المكافئ والفائدة الاقتصادية
تدفق الغاز وانبعاثات غازات الدفيئة الصافية
الخصائص الفيزيائية والكيميائية للتربة، تنوع الميكروبات وتقييم صحة التربة
تم تحديد العينات باستخدام محلل TOC متعدد 3100 N/C (Analytik Jena، ألمانيا). تم استخدام بروتوكول بريمنر القياسي المعدل لاستخراج التربة
الإنتروبيا-توبسيس لتقييم الوظائف المتعددة لدورات المحاصيل المتنوعة
(1) تطبيع المؤشرات:
(2) احسب قيمة الانتروبيا
التحليل الإحصائي
عدم اليقين المرتبط بالأحداث الجوية المتطرفة وتغير المناخ.
ملخص التقرير
توفر البيانات
توفر الشيفرة
References
- Webb, P. et al. The urgency of food system transformation is now irrefutable. Nat. Food 1, 584-585 (2020).
- Gerten, D. et al. Feeding ten billion people is possible within four terrestrial planetary boundaries. Nat. Sustain. 3, 200-208 (2020).
- Beerling, D. J. et al. Potential for large-scale
removal via enhanced rock weathering with croplands. Nature 583, 242-248 (2020). - Cui, Z. L. et al. Pursuing sustainable productivity with millions of smallholder farmers. Nature 555, 363-366 (2018).
- Garnett, T. et al. Sustainable intensification in agriculture: Premises and policies. Science 341, 33-34 (2013).
- Gu, B. J. et al. Cost-effective mitigation of nitrogen pollution from global croplands. Nature 613, 77-84 (2023).
- National Bureau of Statistics of China. Agriculture. China Statistical Yearbook – 2021. (China Statistics Press, Beijing, China, 2021).
- Liu, G., Zhang, F. & Deng, X. Z. Half of the greenhouse gas emissions from China’s food system occur during food production. Commun. Earth Environ. 4, 161 (2023).
- Friedlingstein, P. et al. Global carbon budget 2021. Earth Syst. Sci. Data 14, 1917-2005 (2022).
- Cumming, G. S. et al. Implications of agricultural transitions and urbanization for ecosystem services. Nature 515, 50-57 (2014).
- Marris, E. A world without soil: The past, present, and precarious future of the earth beneath our feet. Nature 601, 503-504 (2022).
- Xu, H. G. et al. Ensuring effective implementation of the post-2020 global biodiversity targets. Nat. Ecol. Evol. 5, 411-418 (2021).
- Wang, B. et al. Sources of uncertainty for wheat yield projections under future climate are site-specific. Nat. Food 1, 720-728 (2020).
- Shi, Y. et al. Building social resilience in North Korea can mitigate the impacts of climate change on food security. Nat. Food 3, 499-511 (2022).
- Suweis, S., Carr, J. A., Maritan, A., Rinaldo, A. & D’Odorico, P. Resilience and reactivity of global food security. Proc. Natl Acad. Sci. USA 112, 6902-6907 (2015).
- Tilman, D. et al. Future threats to biodiversity and pathways to their prevention. Nature 546, 73-81 (2017).
- Chai, Q. et al. Integrated farming with intercropping increases food production while reducing environmental footprint. Proc. Natl Acad. Sci. USA 118, e2106382118 (2021).
- Xie, W. et al. Crop switching can enhance environmental sustainability and farmer incomes in China. Nature 616, 300-305 (2023).
- Guo, Y. X. et al. Air quality, nitrogen use efficiency and food security in China are improved by cost-effective agricultural nitrogen management. Nat. Food 1, 648-658 (2020).
- Lucas, E., Guo, M. & Guillén-Gosálbez, G. Low-carbon diets can reduce global ecological and health costs. Nat. Food 4, 394-406 (2023).
- Ray, D. K., Ramankutty, N., Mueller, N. D., West, P. C. & Foley, J. A. Recent patterns of crop yield growth and stagnation. Nat. Commun. 3, 1293 (2012).
- Ranganathan, J., Daniels, R. J. R., Chandran, M. D. S., Ehrlich, P. R. & Daily, G. C. Sustaining biodiversity in ancient tropical countryside. Proc. Natl Acad. Sci. USA 105, 17852-17854 (2008).
- Crippa, M. et al. Food systems are responsible for a third of global anthropogenic GHG emissions. Nat. Food 2, 198-209 (2021).
- Yang, X. L. et al. Diversified crop rotations enhance groundwater and economic sustainability of food production. Food Energy Secur. 10, e311 (2021).
- Liu, Z. J. et al. Patterns and causes of winter wheat and summer maize rotation area change over the North China Plain. Environ. Res. Lett. 17, 044056 (2022).
- FAO. FAOSTAT. (2023). https://www.fao.org/faostat/en/#data/EM/ visualize, last access: 19 Jul 2023.
- Bowles, T. M. et al. Long-term evidence shows that crop-rotation diversification increases agricultural resilience to adverse growing conditions in North America. One Earth 2, 284-293 (2020).
- Li, J. X. et al. Diversifying crop rotation improves system robustness. Agron. Sustain. Dev. 39, 38 (2019).
- Giller, K. E. The food security conundrum of sub-Saharan Africa. Glob. Food Secur. 26, 100431 (2020).
- Hijbeek, R. et al. Do organic inputs matter – a meta-analysis of additional yield effects for arable crops in Europe. Plant Soil 411, 293-303 (2017).
- Mhlanga, B., Ercoli, L., Pellegrino, E., Onofri, A. & Thierfelder, C. The crucial role of mulch to enhance the stability and resilience of cropping systems in southern Africa. Agron. Sustain. Dev. 41, 29 (2021).
- MacLaren, C. et al. Long-term evidence for ecological intensification as a pathway to sustainable agriculture. Nat. Sustain. 5, 770-779 (2022).
- Lin, B. B. Resilience in agriculture through crop diversification: adaptive management for environmental change. BioScience 61, 183-193 (2011).
- Robertson, G. P., Paul, E. A. & Harwood, R. R. Greenhouse gases in intensive agriculture: contributions of individual gases to the radiative forcing of the atmosphere. Science 289, 1922-1925 (2000).
- Powlson, D. S. et al. Limited potential of no-till agriculture for climate change mitigation. Nat. Clim. Change 4, 678-683 (2014).
- Liu, K., Bandara, M., Hamel, C., Knight, J. D. & Gan, Y. T. Intensifying crop rotations with pulse crops enhances system productivity and soil organic carbon in semi-arid environments. Field Crops Res. 248, 107657 (2020).
- Chenu, C. et al. Increasing organic stocks in agricultural soils: knowledge gaps and potential innovations. Soil Tillage Res. 188, 41-52 (2019).
- Drinkwater, L. E., Wagoner, P. & Sarrantonio, M. Legume-based cropping systems have reduced carbon and nitrogen losses. Nature 396, 262-265 (1998).
- Montgomery, D. R. & Biklé, A. Soil health and nutrient density: beyond organic vs. conventional farming. Front. Sustain. Food Syst. 5, 417 (2021).
- Guinet, M., Nicolardot, B. & Voisin, A.-S. Nitrogen benefits of ten legume pre-crops for wheat assessed by field measurements and modelling. Eur. J. Agron. 120, 126151 (2020).
- Abdalla, M. et al. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity. Glob. Change Biol. 25, 2530-2543 (2019).
- Olesen, J. E. et al. Challenges of accounting nitrous oxide emissions from agricultural crop residues. Glob. Change Biol. 00, 1-10 (2023).
- Gan, Y. T. et al. Improving farming practices reduces the carbon footprint of spring wheat production. Nat. Commun. 5, 5012 (2014).
- Ma, B. L., Liang, B. C., Biswas, D. K., Morrison, M. J. & McLaughlin, N.
. The carbon footprint of maize production as affected by nitrogen fertilizer and maize-legume rotations. Nutr. Cycl. Agroecosyst. 94, 15-31 (2012). - Karlsson, H., Ahlgren, S., Strid, I. & Hansson, P.-A. Faba beans for biorefinery feedstock or feed? Greenhouse gas and energy balances of different applications. Agric. Syst. 141, 138-148 (2015).
- Plaza-Bonilla, D., Nolot, J.-M., Raffaillac, D. & Justes, E. Innovative cropping systems to reduce N inputs and maintain wheat yields by inserting grain legumes and cover crops in southwestern France. Eur. J. Agron. 82, 331-341 (2017).
- Roesch-McNally, G. E., Arbuckle, J. G. & Tyndall, J. C. Barriers to implementing climate resilient agricultural strategies: The case of crop diversification in the U.S. Corn Belt. Glob. Environ. Change 48, 206-215 (2018).
- Shah, K. K., Modi, B., Pandey, H. P., Subedi, A. & Shrestha, J. Diversified crop rotation: an approach for sustainable agriculture production. Adv. Agric. 2021, 1-9 (2021).
- Liang, J. P. & Shi, W. J. Cotton/halophytes intercropping decreases salt accumulation and improves soil physicochemical properties and crop productivity in saline-alkali soils under mulched drip irrigation: a three-year field experiment. Field Crops Res. 262, 108027 (2021).
- Hazra, K. K. et al. Diversification of maize-wheat cropping system with legumes and integrated nutrient management increases soil aggregation and carbon sequestration. Geoderma 353, 308-319 (2019).
- Mooshammer, M. et al. Microbial feedbacks on soil organic matter dynamics underlying the legacy effect of diversified cropping systems. Soil Biol. Biochem. 167, 108584 (2022).
- Dias, T., Dukes, A. & Antunes, P. M. Accounting for soil biotic effects on soil health and crop productivity in the design of crop rotations. J. Sci. Food Agric. 95, 447-454 (2015).
- Garbeva, P., Veen, J. A. V. & Elsas, J. D. V. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 42, 243-270 (2004).
- Selim, M. M. A review of advantages, disadvantages and challenges of crop rotations. Egypt. J. Agron. 41, 1-10 (2019).
- Garland, G. et al. Crop cover is more important than rotational diversity for soil multifunctionality and cereal yields in European cropping systems. Nat. Food 2, 28-37 (2021).
- Yang, Y. H. et al. Optimizing crop rotation increases soil carbon and reduces GHG emissions without sacrificing yields. Agric. Ecosyst. Environ. 342, 108220 (2023).
- Jarecki, M. et al. Long-term trends in corn yields and soil carbon under diversified crop rotations. J. Environ. Qual. 47, 635-643 (2018).
- Baldwin-Kordick, R. et al. Comprehensive impacts of diversified cropping on soil health and sustainability. Agroecol. Sustain. Food Syst. 46, 331-363 (2022).
- Chahal, I., Hooker, D. C., Deen, B., Janovicek, K. & Van Eerd, L. L. Long-term effects of crop rotation, tillage, and fertilizer nitrogen on soil health indicators and crop productivity in a temperate climate. Soil Tillage Res. 213, 105121 (2021).
- Fang, Q., Zhang, X. Y., Chen, S. Y., Shao, L. & Sun, H. Selecting traits to increase winter wheat yield under climate change in the North China Plain. Field Crops Res. 207, 30-41 (2017).
- da Silva, P. C. G. et al. No-tillage and crop rotation increase crop yields and nitrogen stocks in sandy soils under agroclimatic risk. Field Crops Res. 258, 107947 (2020).
- Hu, Z. Y. et al. Italian ryegrass-rice rotation system for biomass production and cadmium removal from contaminated paddy fields. J. Soils Sediment. 20, 874-882 (2020).
- Song, D. G. et al. Influence of planting distance and density on the yield and photosynthetic traits of sweet potato (Ipomoea balatas L.) under an intercropping system with walnut (Juglans regia) saplings. Soil Tillage Res. 196, 104484 (2020).
- Zhao, J. H. et al. The impacts of soil tillage combined with plastic film management practices on soil quality, carbon footprint, and peanut yield. Eur. J. Agron. 148, 126881 (2023).
- Liu, X. B. et al. Yield response of continuous soybean to one-season crop disturbance in a previous continuous soybean field in Northeast China. Field Crops Res. 138, 52-56 (2012).
- Alam, M. K., Bell, R. W. & Biswas, W. K. Decreasing the carbon footprint of an intensive rice-based cropping system using conservation agriculture on the Eastern Gangetic Plains. J. Clean. Prod. 218, 259-272 (2019).
- Wang, S. Q. et al. Diversified crop rotations reduce groundwater use and enhance system resilience. Agric. Water Manag. 276, 108067 (2023).
- Groot, J. C. J. & Yang, X. L. Trade-offs in the design of sustainable cropping systems at a regional level: a case study on the North China Plain. FASE 9, 14 (2022).
- Zheng, X. H. et al. Quantification of
fluxes from soil-plant systems may be biased by the applied gas chromatograph methodology. Plant Soil 311, 211-234 (2008). - Huang, J. X., Chen, Y. Q., Sui, P. & Gao, W. S. Estimation of net greenhouse gas balance using crop- and soil-based approaches: Two case studies. Sci. Total Environ. 456-457, 299-306 (2013).
- IPCC. Climate change 2021: The physical science basis. Contribution of working group i to the sixth assessment report of the intergovernmental panel on climate change. (ed. Masson-Delmotte, V. P. Z. A.) (Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2021). https://www.ipcc.ch/report/sixth-assessment-report-working-group-i/.
- Mclean, E. O. Soil pH and Lime Requirement. in Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties, 9.2.2, Second Edition (ed. Page, A. L.) 199-224 (American Society of Agronomy, Inc. and Soil Science Society of America, Inc., Madison, Wisconsin, USA, 1982).
- Bremner, J. M. & Shaw, K. Denitrification in soil. I. Methods. Investig. J. Agric. Sci. 51, 22-39 (1958).
- Ladd, J. N. & Amato, M. Relationship between microbial biomass carbon in soils and absorbance ( 260 nm ) of extracts of fumigated soils. Soil Biol. Biochem. 21, 457-459 (1989).
- Bremner, J. M. Inorganic forms of nitrogen. in Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties (ed. Norman, A. G.) 1179-1237 (American Society of Agronomy Madison, 1965).
- Olsen, S. R. & Sommers, L. E. Phosphorus. in Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties, 9.2.2, 2nd Edition (ed. Page, A. L.) 403-430 (American Society of Agronomy, Inc. and Soil Science Society of America, Inc., 1982).
- Harris, D., Horwáth, W. R. & van Kessel, C. Acid fumigation of soils to remove carbonates prior to total organic carbon or CARBON-13 isotopic analysis. Soil Sci. Soc. Am. J. 65, 1853-1856 (2001).
- Grossman, R. B. & Reinsch, T. G. 2.1 Bulk density and linear extensibility. in Methods of Soil Analysis: Part 4 Physical Methods (eds. Dane, J. H. & Topp, G. C.) 201-228 (Soil Science Society of America, Inc., 2002).
- Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335-336 (2010).
- Edgar, R. C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996-998 (2013).
- Gugino, B. K. et al. Cornell Soil Health Assessment Training Manual, Edition 1.2, (Cornell University Press, Cornell University, 2007).
- Idowu, O. J. et al. Farmer-oriented assessment of soil quality using field, laboratory, and VNIR spectroscopy methods. Plant Soil 307, 243-253 (2008).
- Moebius-Clune, B. N. et al. Comprehensive Assessment of Soil Health – The Cornell Framework, Edition 3.2, (Cornell University Press, Cornell University, 2016).
- Chang, T. T., Feng, G., Paul, V., Adeli, A. & Brooks, J. P. Chapter Three – Soil health assessment methods: Progress, applications and comparison. in Adv. Agron., Vol. 172 (ed. Sparks, D. L.) 129-210 (Academic Press, 2022).
- Congreves, K. A., Hayes, A., Verhallen, E. A. & Van Eerd, L. L. Longterm impact of tillage and crop rotation on soil health at four temperate agroecosystems. Soil Tillage Res. 152, 17-28 (2015).
- Wang, H. D. et al. Multi-objective optimization of water and fertilizer management for potato production in sandy areas of northern China based on TOPSIS. Field Crops Res. 240, 55-68 (2019).
- Chen, Y., Zhu, M. K., Lu, J. L., Zhou, Q. & Ma, W. B. Evaluation of ecological city and analysis of obstacle factors under the background of high-quality development: Taking cities in the Yellow River Basin as examples. Ecol. Indic. 118, 106771 (2020).
- Liu, X. G., Peng, Y. L., Yang, Q. L., Wang, X. K. & Cui, N. B. Determining optimal deficit irrigation and fertilization to increase mango yield, quality, and WUE in a dry hot environment based on TOPSIS. Agric. Water Manag. 245, 106650 (2021).
- Delgado, A. & Romero, I. Environmental conflict analysis using an integrated grey clustering and entropy-weight method: A case study of a mining project in Peru. Environ. Modell. Softw. 77, 108-121 (2016).
- Chen, C. W., Wang, J. H., Wang, J. C. & Shen, Z. H. Developing indicators for sustainable campuses in Taiwan using fuzzy Delphi method and analytic hierarchy process. J. Clean. Prod. 193, 661-671 (2018).
- R Core Team. R: A language and environment for statistical computing. in Computing (R foundation for statistical computing, Vienna, Austria, 2014).
شكر وتقدير
مساهمات المؤلفين
المصالح المتنافسة
معلومات إضافية
المواد التكميلية متاحة على
https://doi.org/10.1038/s41467-023-44464-9.
http://www.nature.com/reprints
ملاحظة الناشر: تظل شركة سبرينغر ناتشر محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
© المؤلفون 2024
المختبر الوطني الرئيسي للاستخدام الفعال لموارد المياه الزراعية، بكين 100083، الصين. كلية موارد المياه والهندسة المدنية، جامعة الزراعة الصينية، بكين 100083، الصين. مدرسة الزراعة والغابات الاستوائية، جامعة هاينان، هايكو 570228، الصين. كلية علوم الحياة والبيئة، جامعة ونتشو، ونتشو، تشجيانغ 325035، الصين. الـ مجموعة BC للتربة، مرتفعات تالوس، كيلونا، كولومبيا البريطانية، كندا. المختبر الوطني الرئيسي للتربة والزراعة المستدامة، معهد علوم التربة، الأكاديمية الصينية للعلوم، نانجينغ 210008، الصين. المختبر الرئيسي لموارد المياه الزراعية، مركز أبحاث الموارد الزراعية، معهد الوراثة وعلم الأحياء التنموي، الأكاديمية الصينية للعلوم، شيجياتشوانغ 050022، الصين. قسم الهندسة البيولوجية والبيئية، قاعة رايلي-روب، جامعة كورنيل، إيثاكا، نيويورك 14853، الولايات المتحدة الأمريكية. معهد الزراعة بجامعة أستراليا الغربية، جامعة أستراليا الغربية، بيرث، WA 6001، أستراليا. لاند-كرافت، قسم الزراعة البيئية، جامعة آرهوس، آرهوس، الدنمارك. معهد الأرصاد الجوية وبحوث المناخ، بحوث البيئة الجوية (IMK-IFU)، معهد كارلسروه للتكنولوجيا (KIT)، غارميش-بارتنكيرشن، ألمانيا. البريد الإلكتروني:yangxiaolin429@cau.edu.cn; دوتايسينغ@cau.edu.cn; juxt@cau.edu.cn; gary.gan@wzu.edu.cn
DOI: https://doi.org/10.1038/s41467-023-44464-9
PMID: https://pubmed.ncbi.nlm.nih.gov/38172570
Publication Date: 2024-01-03
Diversifying crop rotation increases food production, reduces net greenhouse gas emissions and improves soil health
Accepted: 14 December 2023
Published online: 03 January 2024
(A) Check for updates
Abstract
Global food production faces challenges in balancing the need for increased yields with environmental sustainability. This study presents a six-year field experiment in the North China Plain, demonstrating the benefits of diversifying traditional cereal monoculture (wheat-maize) with cash crops (sweet potato) and legumes (peanut and soybean). The diversified rotations increase equivalent yield by up to
systems have also emitted great amounts of greenhouse gases (GHG) and caused environmental degradation
available phosphorus (AP), microbial biomass carbon (MBC), microbial biomass nitrogen (MBN), and microbial community composition and diversity. These measurements enabled us to test the hypotheses that (a) cash-crop diversified system increases farmers’ net income without jeopardizing crop yields, (b) legume diversified systems reduce field scale GHG emissions, and (c) integrating diversified rotations increases food production, reduces GHG emissions, and benefits soil health (Fig. 1). Our results demonstrate that (a) instructive findings from newly designed, tested, and validated diversified systems could guide the North China Plain in establishing a more sustainable system to maintain or increase grain and protein production with reducing the damage to the environment and soil ecosystems, and (b) the results from such a representatively intensive food producing region may provide a guide for the countries/regions with similar agricultural environments to follow on an expanded scale.
Results
Diversified crop rotations increase ecosystem productivity
legume crops can maintain crop yields, increase farmers’ income, and reduce GHG emissions due to the biological
yield varied among the rotation systems, with ryegrass (Lolium multiflorum L.)-sorghum (Sorghum bicolor L.) rotation (RSWM) and soybean
Diversified crop rotations reduce net GHG emissions
control which was at
between the rotations at
emissions by
Diversified crop rotations enhance soil health and microbial diversity
sorghum
complex, but it was evident that bacterial and fungal community composition changed from 2016 to 2022. The rotations with sweet potato, peanut, and soybean had distinct bacterial community compositions from the other three systems (Fig. 5e), whereas the five studied diversified rotations had fungal community compositions distinctly different from the wheat-maize control (Fig. 5f). The 6 years of rotational nourishment affected soil physiochemical properties interactively with soil microbial communities. The rotations with sweet potato, peanut, and soybean bacterial communities were closely associated with SOC and AP, whereas those of wheat-maize, spring maize
Multiple functions assessments of diversified crop rotations
Discussion
indicating soil properties including soil organic carbon (SOC), dissolved organic carbon (DOC), total nitrogen (TN),
soil carbon stock in the 0-90 cm layer from October 2016 to October 2022; Net GHG emissions denotes the net GHG budgets including direct and indirect GHG emissions minus soil carbon sequestration; Soil biodiversity is represented by the Shannon Index of bacterial and fungal communities; Soil health score is calculated from 10 indicators using the overall Cornell Soil Health Assessment (CSHA) matrix combined with PCA
benefits
potato, peanut and soybean significantly increased soil C after 6 years of rotation nourishment compared to the traditional winter wheat-summer maize double-cropping system. Introducing sweet potatoes and legumes into these rotations has been shown to stimulate microbial growth, increased C use efficiency
source of microbial products that affect stable SOC with a high capacity for C stabilization
increased fungal community diversity compared to winter wheat-summer maize, shaping soil nutrient profiles with different carbon substrates that impacted microbial diversity. Moreover, diversified crop rotations potentially reduce soil agrochemical contamination due to decreased application of synthetic N fertilizer and herbicides, favoring soil microenvironments
Methods
Equivalent yield and economic benefit
Gas flux and net GHG emissions
Soil physiochemical properties, microbial diversity and soil health scoring
samples were determined using a Multi 3100 N/C TOC analyzer (Analytik Jena, Germany). A modified Bremner’s standard protocol was used to extract soil
Entropy-TOPSIS for multiple function assessment of diversified crop rotations
(1) Normalize the indicators:
(2) Calculate the entropy value
Statistical analysis
uncertainty associated with extreme weather events and climate change.
Reporting summary
Data availability
Code availability
References
- Webb, P. et al. The urgency of food system transformation is now irrefutable. Nat. Food 1, 584-585 (2020).
- Gerten, D. et al. Feeding ten billion people is possible within four terrestrial planetary boundaries. Nat. Sustain. 3, 200-208 (2020).
- Beerling, D. J. et al. Potential for large-scale
removal via enhanced rock weathering with croplands. Nature 583, 242-248 (2020). - Cui, Z. L. et al. Pursuing sustainable productivity with millions of smallholder farmers. Nature 555, 363-366 (2018).
- Garnett, T. et al. Sustainable intensification in agriculture: Premises and policies. Science 341, 33-34 (2013).
- Gu, B. J. et al. Cost-effective mitigation of nitrogen pollution from global croplands. Nature 613, 77-84 (2023).
- National Bureau of Statistics of China. Agriculture. China Statistical Yearbook – 2021. (China Statistics Press, Beijing, China, 2021).
- Liu, G., Zhang, F. & Deng, X. Z. Half of the greenhouse gas emissions from China’s food system occur during food production. Commun. Earth Environ. 4, 161 (2023).
- Friedlingstein, P. et al. Global carbon budget 2021. Earth Syst. Sci. Data 14, 1917-2005 (2022).
- Cumming, G. S. et al. Implications of agricultural transitions and urbanization for ecosystem services. Nature 515, 50-57 (2014).
- Marris, E. A world without soil: The past, present, and precarious future of the earth beneath our feet. Nature 601, 503-504 (2022).
- Xu, H. G. et al. Ensuring effective implementation of the post-2020 global biodiversity targets. Nat. Ecol. Evol. 5, 411-418 (2021).
- Wang, B. et al. Sources of uncertainty for wheat yield projections under future climate are site-specific. Nat. Food 1, 720-728 (2020).
- Shi, Y. et al. Building social resilience in North Korea can mitigate the impacts of climate change on food security. Nat. Food 3, 499-511 (2022).
- Suweis, S., Carr, J. A., Maritan, A., Rinaldo, A. & D’Odorico, P. Resilience and reactivity of global food security. Proc. Natl Acad. Sci. USA 112, 6902-6907 (2015).
- Tilman, D. et al. Future threats to biodiversity and pathways to their prevention. Nature 546, 73-81 (2017).
- Chai, Q. et al. Integrated farming with intercropping increases food production while reducing environmental footprint. Proc. Natl Acad. Sci. USA 118, e2106382118 (2021).
- Xie, W. et al. Crop switching can enhance environmental sustainability and farmer incomes in China. Nature 616, 300-305 (2023).
- Guo, Y. X. et al. Air quality, nitrogen use efficiency and food security in China are improved by cost-effective agricultural nitrogen management. Nat. Food 1, 648-658 (2020).
- Lucas, E., Guo, M. & Guillén-Gosálbez, G. Low-carbon diets can reduce global ecological and health costs. Nat. Food 4, 394-406 (2023).
- Ray, D. K., Ramankutty, N., Mueller, N. D., West, P. C. & Foley, J. A. Recent patterns of crop yield growth and stagnation. Nat. Commun. 3, 1293 (2012).
- Ranganathan, J., Daniels, R. J. R., Chandran, M. D. S., Ehrlich, P. R. & Daily, G. C. Sustaining biodiversity in ancient tropical countryside. Proc. Natl Acad. Sci. USA 105, 17852-17854 (2008).
- Crippa, M. et al. Food systems are responsible for a third of global anthropogenic GHG emissions. Nat. Food 2, 198-209 (2021).
- Yang, X. L. et al. Diversified crop rotations enhance groundwater and economic sustainability of food production. Food Energy Secur. 10, e311 (2021).
- Liu, Z. J. et al. Patterns and causes of winter wheat and summer maize rotation area change over the North China Plain. Environ. Res. Lett. 17, 044056 (2022).
- FAO. FAOSTAT. (2023). https://www.fao.org/faostat/en/#data/EM/ visualize, last access: 19 Jul 2023.
- Bowles, T. M. et al. Long-term evidence shows that crop-rotation diversification increases agricultural resilience to adverse growing conditions in North America. One Earth 2, 284-293 (2020).
- Li, J. X. et al. Diversifying crop rotation improves system robustness. Agron. Sustain. Dev. 39, 38 (2019).
- Giller, K. E. The food security conundrum of sub-Saharan Africa. Glob. Food Secur. 26, 100431 (2020).
- Hijbeek, R. et al. Do organic inputs matter – a meta-analysis of additional yield effects for arable crops in Europe. Plant Soil 411, 293-303 (2017).
- Mhlanga, B., Ercoli, L., Pellegrino, E., Onofri, A. & Thierfelder, C. The crucial role of mulch to enhance the stability and resilience of cropping systems in southern Africa. Agron. Sustain. Dev. 41, 29 (2021).
- MacLaren, C. et al. Long-term evidence for ecological intensification as a pathway to sustainable agriculture. Nat. Sustain. 5, 770-779 (2022).
- Lin, B. B. Resilience in agriculture through crop diversification: adaptive management for environmental change. BioScience 61, 183-193 (2011).
- Robertson, G. P., Paul, E. A. & Harwood, R. R. Greenhouse gases in intensive agriculture: contributions of individual gases to the radiative forcing of the atmosphere. Science 289, 1922-1925 (2000).
- Powlson, D. S. et al. Limited potential of no-till agriculture for climate change mitigation. Nat. Clim. Change 4, 678-683 (2014).
- Liu, K., Bandara, M., Hamel, C., Knight, J. D. & Gan, Y. T. Intensifying crop rotations with pulse crops enhances system productivity and soil organic carbon in semi-arid environments. Field Crops Res. 248, 107657 (2020).
- Chenu, C. et al. Increasing organic stocks in agricultural soils: knowledge gaps and potential innovations. Soil Tillage Res. 188, 41-52 (2019).
- Drinkwater, L. E., Wagoner, P. & Sarrantonio, M. Legume-based cropping systems have reduced carbon and nitrogen losses. Nature 396, 262-265 (1998).
- Montgomery, D. R. & Biklé, A. Soil health and nutrient density: beyond organic vs. conventional farming. Front. Sustain. Food Syst. 5, 417 (2021).
- Guinet, M., Nicolardot, B. & Voisin, A.-S. Nitrogen benefits of ten legume pre-crops for wheat assessed by field measurements and modelling. Eur. J. Agron. 120, 126151 (2020).
- Abdalla, M. et al. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity. Glob. Change Biol. 25, 2530-2543 (2019).
- Olesen, J. E. et al. Challenges of accounting nitrous oxide emissions from agricultural crop residues. Glob. Change Biol. 00, 1-10 (2023).
- Gan, Y. T. et al. Improving farming practices reduces the carbon footprint of spring wheat production. Nat. Commun. 5, 5012 (2014).
- Ma, B. L., Liang, B. C., Biswas, D. K., Morrison, M. J. & McLaughlin, N.
. The carbon footprint of maize production as affected by nitrogen fertilizer and maize-legume rotations. Nutr. Cycl. Agroecosyst. 94, 15-31 (2012). - Karlsson, H., Ahlgren, S., Strid, I. & Hansson, P.-A. Faba beans for biorefinery feedstock or feed? Greenhouse gas and energy balances of different applications. Agric. Syst. 141, 138-148 (2015).
- Plaza-Bonilla, D., Nolot, J.-M., Raffaillac, D. & Justes, E. Innovative cropping systems to reduce N inputs and maintain wheat yields by inserting grain legumes and cover crops in southwestern France. Eur. J. Agron. 82, 331-341 (2017).
- Roesch-McNally, G. E., Arbuckle, J. G. & Tyndall, J. C. Barriers to implementing climate resilient agricultural strategies: The case of crop diversification in the U.S. Corn Belt. Glob. Environ. Change 48, 206-215 (2018).
- Shah, K. K., Modi, B., Pandey, H. P., Subedi, A. & Shrestha, J. Diversified crop rotation: an approach for sustainable agriculture production. Adv. Agric. 2021, 1-9 (2021).
- Liang, J. P. & Shi, W. J. Cotton/halophytes intercropping decreases salt accumulation and improves soil physicochemical properties and crop productivity in saline-alkali soils under mulched drip irrigation: a three-year field experiment. Field Crops Res. 262, 108027 (2021).
- Hazra, K. K. et al. Diversification of maize-wheat cropping system with legumes and integrated nutrient management increases soil aggregation and carbon sequestration. Geoderma 353, 308-319 (2019).
- Mooshammer, M. et al. Microbial feedbacks on soil organic matter dynamics underlying the legacy effect of diversified cropping systems. Soil Biol. Biochem. 167, 108584 (2022).
- Dias, T., Dukes, A. & Antunes, P. M. Accounting for soil biotic effects on soil health and crop productivity in the design of crop rotations. J. Sci. Food Agric. 95, 447-454 (2015).
- Garbeva, P., Veen, J. A. V. & Elsas, J. D. V. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 42, 243-270 (2004).
- Selim, M. M. A review of advantages, disadvantages and challenges of crop rotations. Egypt. J. Agron. 41, 1-10 (2019).
- Garland, G. et al. Crop cover is more important than rotational diversity for soil multifunctionality and cereal yields in European cropping systems. Nat. Food 2, 28-37 (2021).
- Yang, Y. H. et al. Optimizing crop rotation increases soil carbon and reduces GHG emissions without sacrificing yields. Agric. Ecosyst. Environ. 342, 108220 (2023).
- Jarecki, M. et al. Long-term trends in corn yields and soil carbon under diversified crop rotations. J. Environ. Qual. 47, 635-643 (2018).
- Baldwin-Kordick, R. et al. Comprehensive impacts of diversified cropping on soil health and sustainability. Agroecol. Sustain. Food Syst. 46, 331-363 (2022).
- Chahal, I., Hooker, D. C., Deen, B., Janovicek, K. & Van Eerd, L. L. Long-term effects of crop rotation, tillage, and fertilizer nitrogen on soil health indicators and crop productivity in a temperate climate. Soil Tillage Res. 213, 105121 (2021).
- Fang, Q., Zhang, X. Y., Chen, S. Y., Shao, L. & Sun, H. Selecting traits to increase winter wheat yield under climate change in the North China Plain. Field Crops Res. 207, 30-41 (2017).
- da Silva, P. C. G. et al. No-tillage and crop rotation increase crop yields and nitrogen stocks in sandy soils under agroclimatic risk. Field Crops Res. 258, 107947 (2020).
- Hu, Z. Y. et al. Italian ryegrass-rice rotation system for biomass production and cadmium removal from contaminated paddy fields. J. Soils Sediment. 20, 874-882 (2020).
- Song, D. G. et al. Influence of planting distance and density on the yield and photosynthetic traits of sweet potato (Ipomoea balatas L.) under an intercropping system with walnut (Juglans regia) saplings. Soil Tillage Res. 196, 104484 (2020).
- Zhao, J. H. et al. The impacts of soil tillage combined with plastic film management practices on soil quality, carbon footprint, and peanut yield. Eur. J. Agron. 148, 126881 (2023).
- Liu, X. B. et al. Yield response of continuous soybean to one-season crop disturbance in a previous continuous soybean field in Northeast China. Field Crops Res. 138, 52-56 (2012).
- Alam, M. K., Bell, R. W. & Biswas, W. K. Decreasing the carbon footprint of an intensive rice-based cropping system using conservation agriculture on the Eastern Gangetic Plains. J. Clean. Prod. 218, 259-272 (2019).
- Wang, S. Q. et al. Diversified crop rotations reduce groundwater use and enhance system resilience. Agric. Water Manag. 276, 108067 (2023).
- Groot, J. C. J. & Yang, X. L. Trade-offs in the design of sustainable cropping systems at a regional level: a case study on the North China Plain. FASE 9, 14 (2022).
- Zheng, X. H. et al. Quantification of
fluxes from soil-plant systems may be biased by the applied gas chromatograph methodology. Plant Soil 311, 211-234 (2008). - Huang, J. X., Chen, Y. Q., Sui, P. & Gao, W. S. Estimation of net greenhouse gas balance using crop- and soil-based approaches: Two case studies. Sci. Total Environ. 456-457, 299-306 (2013).
- IPCC. Climate change 2021: The physical science basis. Contribution of working group i to the sixth assessment report of the intergovernmental panel on climate change. (ed. Masson-Delmotte, V. P. Z. A.) (Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2021). https://www.ipcc.ch/report/sixth-assessment-report-working-group-i/.
- Mclean, E. O. Soil pH and Lime Requirement. in Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties, 9.2.2, Second Edition (ed. Page, A. L.) 199-224 (American Society of Agronomy, Inc. and Soil Science Society of America, Inc., Madison, Wisconsin, USA, 1982).
- Bremner, J. M. & Shaw, K. Denitrification in soil. I. Methods. Investig. J. Agric. Sci. 51, 22-39 (1958).
- Ladd, J. N. & Amato, M. Relationship between microbial biomass carbon in soils and absorbance ( 260 nm ) of extracts of fumigated soils. Soil Biol. Biochem. 21, 457-459 (1989).
- Bremner, J. M. Inorganic forms of nitrogen. in Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties (ed. Norman, A. G.) 1179-1237 (American Society of Agronomy Madison, 1965).
- Olsen, S. R. & Sommers, L. E. Phosphorus. in Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties, 9.2.2, 2nd Edition (ed. Page, A. L.) 403-430 (American Society of Agronomy, Inc. and Soil Science Society of America, Inc., 1982).
- Harris, D., Horwáth, W. R. & van Kessel, C. Acid fumigation of soils to remove carbonates prior to total organic carbon or CARBON-13 isotopic analysis. Soil Sci. Soc. Am. J. 65, 1853-1856 (2001).
- Grossman, R. B. & Reinsch, T. G. 2.1 Bulk density and linear extensibility. in Methods of Soil Analysis: Part 4 Physical Methods (eds. Dane, J. H. & Topp, G. C.) 201-228 (Soil Science Society of America, Inc., 2002).
- Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335-336 (2010).
- Edgar, R. C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996-998 (2013).
- Gugino, B. K. et al. Cornell Soil Health Assessment Training Manual, Edition 1.2, (Cornell University Press, Cornell University, 2007).
- Idowu, O. J. et al. Farmer-oriented assessment of soil quality using field, laboratory, and VNIR spectroscopy methods. Plant Soil 307, 243-253 (2008).
- Moebius-Clune, B. N. et al. Comprehensive Assessment of Soil Health – The Cornell Framework, Edition 3.2, (Cornell University Press, Cornell University, 2016).
- Chang, T. T., Feng, G., Paul, V., Adeli, A. & Brooks, J. P. Chapter Three – Soil health assessment methods: Progress, applications and comparison. in Adv. Agron., Vol. 172 (ed. Sparks, D. L.) 129-210 (Academic Press, 2022).
- Congreves, K. A., Hayes, A., Verhallen, E. A. & Van Eerd, L. L. Longterm impact of tillage and crop rotation on soil health at four temperate agroecosystems. Soil Tillage Res. 152, 17-28 (2015).
- Wang, H. D. et al. Multi-objective optimization of water and fertilizer management for potato production in sandy areas of northern China based on TOPSIS. Field Crops Res. 240, 55-68 (2019).
- Chen, Y., Zhu, M. K., Lu, J. L., Zhou, Q. & Ma, W. B. Evaluation of ecological city and analysis of obstacle factors under the background of high-quality development: Taking cities in the Yellow River Basin as examples. Ecol. Indic. 118, 106771 (2020).
- Liu, X. G., Peng, Y. L., Yang, Q. L., Wang, X. K. & Cui, N. B. Determining optimal deficit irrigation and fertilization to increase mango yield, quality, and WUE in a dry hot environment based on TOPSIS. Agric. Water Manag. 245, 106650 (2021).
- Delgado, A. & Romero, I. Environmental conflict analysis using an integrated grey clustering and entropy-weight method: A case study of a mining project in Peru. Environ. Modell. Softw. 77, 108-121 (2016).
- Chen, C. W., Wang, J. H., Wang, J. C. & Shen, Z. H. Developing indicators for sustainable campuses in Taiwan using fuzzy Delphi method and analytic hierarchy process. J. Clean. Prod. 193, 661-671 (2018).
- R Core Team. R: A language and environment for statistical computing. in Computing (R foundation for statistical computing, Vienna, Austria, 2014).
Acknowledgements
Author contributions
Competing interests
Additional information
supplementary material available at
https://doi.org/10.1038/s41467-023-44464-9.
http://www.nature.com/reprints
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
© The Author(s) 2024
State Key Laboratory of Efficient Utilization of Agricultural Water Resources, Beijing 100083, China. College of Water Resources & Civil Engineering, China Agricultural University, Beijing 100083, China. School of Tropical Agriculture and Forestry, Hainan University, Haikou 570228, China. College of Life and Environmental Sciences, Wenzhou University, Wenzhou, Zhejiang 325035, China. The BC-Soil Group, Tallus Heights, Kelowna, BC, Canada. State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China. Key Laboratory of Agricultural Water Resources, Centre for Agricultural Resources Research, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Shijiazhuang 050022, China. Department of Biological and Environmental Engineering, Riley-Robb Hall, Cornell University, Ithaca, NY 14853, USA. The UWA Institute of Agriculture, The University of Western Australia, Perth, WA 6001, Australia. Land-CRAFT, Department of Agroecology, Aarhus University, Aarhus, Denmark. Institute of Meteorology and Climate Research, Atmospheric Environmental Research (IMK-IFU), Karlsruhe Institute of Technology (KIT), Garmisch Partenkirchen, Germany. e-mail: yangxiaolin429@cau.edu.cn; dutaisheng@cau.edu.cn; juxt@cau.edu.cn; gary.gan@wzu.edu.cn