لقد أظهرت جزيئات الدهون النانوية القابلة للتأين (LNP) التي مكنت من نجاح لقاحات الحمض النووي الرسول (mRNA) أنها تحفز المناعة في غياب الحمض النووي الرسول. ومع ذلك، فإن الآليات التي من خلالها تنشط هذه الجزيئات خلايا المناعة الفطرية لا تزال غير مفهومة تمامًا. باستخدام خط خلايا وحيدة النواة، قمنا بمقارنة قدرة ثلاثة تركيبات من LNP على تنشيط عوامل النسخ مثل عامل نواة كابا ب (NF-kB) وعامل تنظيم الإنترفيرون (IRF). أظهرت المقارنة في خطوط الخلايا المعطلة دورًا لمستقبلات المناعة الفطرية (TLR) 4 في بدء هذه السلسلة من الإشارات ومساهمة مكون الدهون القابلة للتأين. كان التنشيط الناتج عن LNPs الفارغة مشابهًا لذلك الناتج عن LNPs التي تحتوي على الحمض النووي الرسول، مما يشير إلى أن LNPs قد توفر الجزء الأكبر من التحفيز الفطري لمنصة لقاح الحمض النووي الرسول. توضح نتائجنا أن الدهون القابلة للتأين داخل LNPs تشير عبر TLR4 لتنشيط NF-кB وIRF، مما يحدد آلية للتنشيط الفطري يمكن تحسينها لتصميم المحفزات.
تم تطوير الجسيمات النانوية لاستخدامها في توصيل العلاجات ومولدات اللقاح. تم استخدام تركيبات متميزة من الجسيمات النانوية الدهنية (LNP) للقاحين BNT162b2 وmRNA-1273 لتوصيل mRNA المعدل بـ N1-methylpseudouridine الذي يشفر بروتين السنبلة لفيروس SARS-CoV-2.على الرغم من إثبات المناعية، إلا أن المتانة محدودةعلاوة على ذلك، فإن سبب ملف الآثار الجانبية الملحوظة غير معروف.تقدم هذه النتائج فرصة لتعزيز كل من التحمل والفعالية. ومع ذلك، يتطلب تحقيق هذا التحسين فهمًا أعمق للآليات الوظيفية للقاحات mRNA-LNP. كيفية بدء هذه اللقاحات للاستجابات المناعية غير مفهومة تمامًا. تحتوي اللقاحات عادةً على مادة مساعدة تحفز الاستجابة المناعية، وبعضها يستحث هذه الاستجابة من خلال إشارات مستقبلات المناعة الشبيهة بالمسار (TLR).قد يشير مكون mRNA من خلال TLR7 و8، اللذان يتعرفان على RNA أحادي السلسلة (ssRNA). بالإضافة إلى ذلك، قد يؤدي RNA مزدوج السلسلة (dsRNA)، وهو ناتج ثانوي لعملية إنتاج mRNA، إلى تحفيز الإشارات من خلال TLR3 أو RIG-I أو MDA-5. ومع ذلك، فإن mRNA المدمج في هذه اللقاحات قد خضع لتعديلات تهدف إلى تقليل التعرف على ssRNA وتم تنقيته لتقليل وجود dsRNA.. من المثير للاهتمام أن الدراسات التي أجريت في المختبر وفي الجسم الحي أظهرت أن الفراغ
يمكن لـ LNP تنشيط الجهاز المناعي الفطريومع ذلك، فإن الآليات وراء التحفيز المناعي الذي تسببه جزيئات الدهون النانوية غير مفهومة جيدًا.
تختلف جزيئات الدهون النانوية في التركيب، لكنها مصممة بشكل أساسي مع دهون مشحونة أو قابلة للتأين ودهون مساعدة.يتكون LNP من لقاح BNT162b2 mRNA من الدهون الكاتيونية القابلة للتأين ALC-0315، والكوليسترول، وDSPC (1،2-ديستيارويل-sn-غليسيرول-3-فوسفوكولين)، وALC-0159، وهو دهون PEGylated.بالمقارنة، تستخدم LNP من mRNA-1273 SM102 كدهون قابلة للتأين وPEG-DMG كدهون PEGylated.الدهون القابلة للإيون في حالة حيادية عند وقت الإعطاء لكنها تصبح مشحونة إيجابياً في الإندوسوم الحمضي، مما يسهل اندماج جزيئات الدهون النانوية وإطلاق الرنا المرسال إلى السيتوبلازم.الكوليسترول والدهون المساعدة المغلفة بـ PEG تمنع التكتل أو تعزز كفاءة النقل الجيني. تؤثر المكونات أيضًا على الحجم، حيث أن جزيئات الدهون النانوية (LNPs) للقاحين BNT162b2 و mRNA-1273 هي و على التواليمراجعة لصيغ LNP وجدت أنأثارت استجابة مناعية، مع predominating تنشيط المكمل والفطرة.يمكن اكتشاف بعض الجسيمات النانوية الكاتيونية بواسطة TLR4 ومركب الالتهاب NLRP3.. بالإضافة إلى ذلك، يمكن للأجسام المضادة المتولدة ضد الدهون الم PEGylated تنشيط مسار التكملليس من الواضح ما إذا كانت جزيئات LNPs القابلة للتأين الفارغة ستنشط جهاز المناعة.
من خلال مسارات مشابهة للصيغ الكاتيونية حيث تحتوي اللقاحات على دهون كاتيونية قابلة للتأين متميزة قد تؤثر بشكل مختلف على الامتصاص والتحلل..
مؤخراً، تم إثبات أن LNP، في غياب mRNA، له تأثير مناعي محفز في نموذج فئري.أدى إزالة الدهون القابلة للأيون في هذه الجسيمات النانوية الدهنية إلى تقليل إنتاج السيتوكينات، مما يشير إلى أن الدهون القابلة للأيون تلعب دورًا مهمًا في تعزيز المناعة.وبالمثل، فإن تحفيز خلايا الدندريتية المشتقة من وحيدات النوى البشرية باستخدام جزيئات نانوية قابلة للتأين يؤدي إلى إنتاج السيتوكينات مثل IL-1 وIL-6 وIL-12 وIFNα وIFN.حتى الآن، ومع ذلك، فإن الآليات التي من خلالها تنشط الجسيمات النانوية الفارغة الجهاز المناعي الفطري ليست مفهومة جيدًا.
لاحظ العلامة وآخرون أن فعالية التحفيز لجزيئات النانو الدهنية القابلة للأيون في نموذج فئري كانت تعتمد جزئيًا على MyD88، مما يشير إلى دور لمستقبلات TLRs أو مستقبلات IL-1.يمكن لمستقبلات TLRs تنشيط العامل النوويوعوامل النسخ لعوامل تنظيم interferon (IRF) من خلال جزيئات التكيف MyD88 و TRIFتعتبر هذه العوامل النسخية ضرورية للاستجابة الفطرية حيث تنظم إنتاج السيتوكينات، والكيموكينات، والإنترفيرونات من النوع الأول (IFNs).. ومع ذلك، لم يجد آخرون أي دليل على مشاركة TLR في الاستجابة المناعية الفطرية للجزيئات النانوية الدهنية في نماذج الفئران.
في هذه الدراسة، سعينا للتحقيق في قدرة LNPs القابلة للتأين الفارغة على تنشيط خلايا المناعة الفطرية البشرية. لذلك، قمنا بمقارنة تنشيط المناعة الفطرية الذي تسببه ثلاثة LNPs باستخدام خط خلايا أحادية النواة البشرية. وقد حددنا أن LNPs القابلة للتأين تنشط NF- بشكل مختلف. و IRF. تظهر نتائجنا أن الإشارات تتم بشكل أساسي من خلال TLR4. ومن المثير للاهتمام أن NF-وكانت استجابات IRF تجاه LNP BNT162b2 الفارغة مشابهة من حيث الحجم لـ mRNA-LNP، مما يشير إلى أن LNP القابلة للتأين قد تكون مسؤولة بشكل أساسي عن تنشيط الاستجابة المناعية الفطرية خلال التطعيم بـ mRNA.
النتائج
تثير الجسيمات النانوية القابلة للأيون NF-кB واستجابات IRF من خلايا THP1
تعد الخلايا الوحيدة من الخلايا المناعية الفطرية الرئيسية التي تستجيب بسرعة للمحفزات المرتبطة بالعدوى أو الضرر. يرتبط تنشيط ووظيفة الخلايا الوحيدة بعامل النسخ NF-نشاط وتفعيل مسار IRFلتحديد ما إذا كانت جزيئات LNP القابلة للتأين الفارغة يمكن أن تنشط الخلايا الوحيدة، استخدمنا خط خلايا شبيهة بالوحيدات THP-1 مع تقرير الفوسفاتاز القلوي لـ NF-تنشيط ومراسل لوسيفيراز لتنشيط IRF. قمنا بمقارنة التنشيط الناتج عن LNPs القابلة للتأين المختلفة مع R848 (منبه TLR7/8) وMPLA (منبه TLR4). كانت LNPs الفارغة المستخدمة هي LNP-1 وLNP-ALC315 (تركيبة BNT162b2) وLNP-SM102 (تركيبة mRNA-1273) (الجدول التكميلي 1). يختلف LNP-SM102 في هيكل الدهون القابلة للتأين عن LNP-ALC315 وكذلك في الدهون الم PEGylated (الشكل التكميلي 1) (الجدول التكميلي 1).
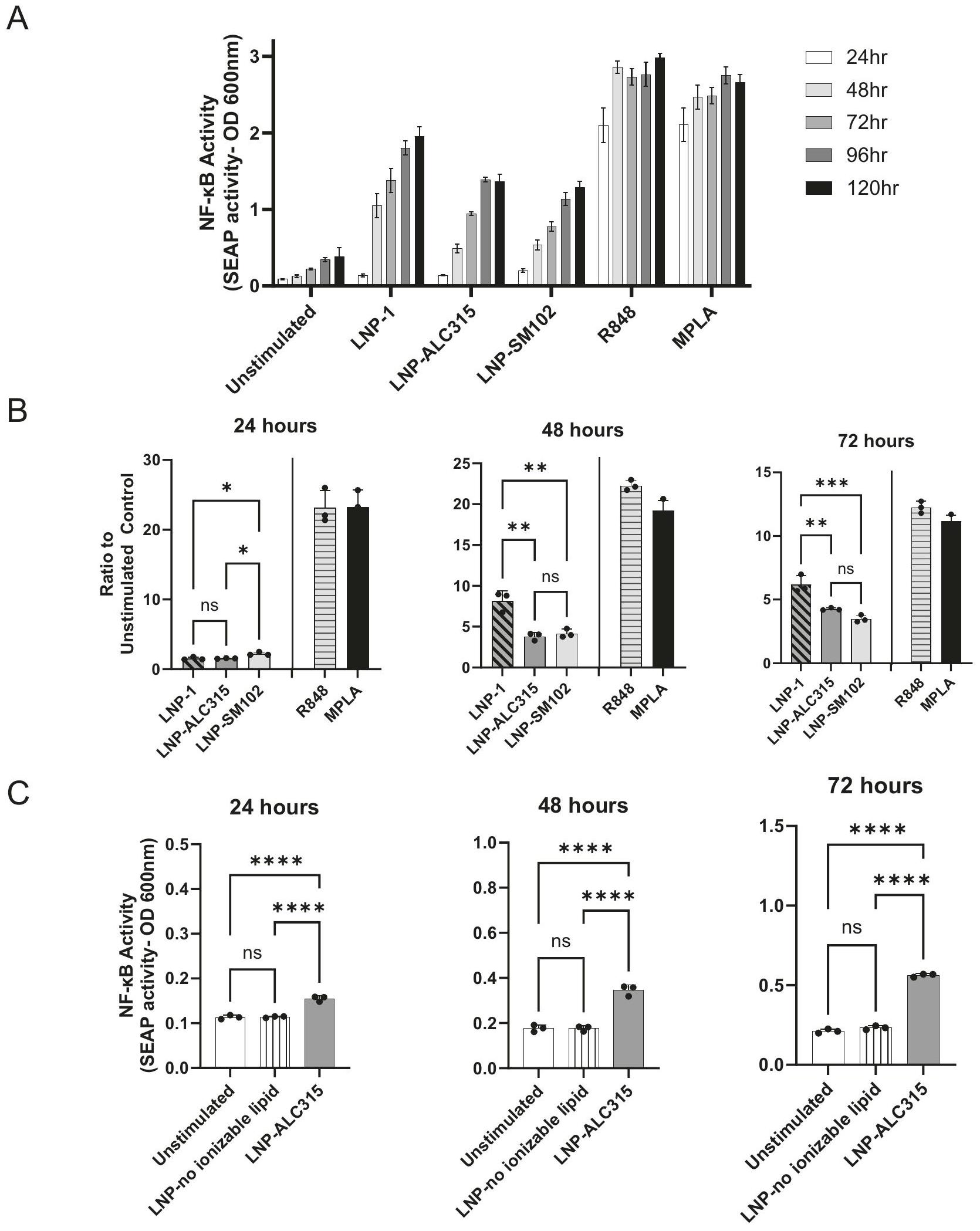
تحفيز خلايا THP-1 أدى إلى NF-كشف المراسل بعد 24 ساعة، مع زيادة NF-التفعيل عند 48 ساعة الذي زاد بشكل طفيف أو استقر حتى 120 ساعة (الشكل 1A). بالنسبة لـ LNP-ALC315، فإن NF-كانت الاستجابة أعلى بمقدار 2 مرة مقارنة بالتحكم غير المحفز عند 24 ساعة ثم 4 مرات بعد 48 ساعة. كانت الاستجابة لـ LNP-1 بمقدار 6-7 مرات عند 48 و72 ساعة. كانت الاستجابة لـ LNP-1 بمقدار 6-7 مرات عند 48 و72 ساعة. NF-كانت الاستجابة الناتجة عن LNP-SM102 مشابهة من حيث الحجم لتلك الناتجة عن LNP-ALC315 (الشكل 1). زادت حركية إنتاج المراسل لجميع LNPs على مدار 24 ساعة مع بلوغ الذروة في 48 و72 ساعة (الشكل التوضيحي 3A). بالمقارنة، أظهرت جميع المحفزات TLR استجابات قوية خلال 24 ساعة. كما لاحظنا أن NF-كانت عملية نقل الإشارة تعتمد على الجرعة وبلغت ذروتها عند أعلى جرعة بغض النظر عن LNP (الشكل التكميلي 2A). أظهرت جميع الظروف تغييرات مماثلة في البقاء على قيد الحياة مع مرور الوقت في الثقافة، وأظهرت R848 و MPLA أعلى نسبة من موت الخلايا (الشكل التكميلي 3C). نظرًا لأن انخفاض البقاء على قيد الحياة يُلاحظ في جميع الظروف، هناك احتمال أن تساهم منتجات موت الخلايا في التنشيط المرصود، خاصةً مع بدء موت الخلايا حول بدء NF- الكشف. في يؤدي إلى أعلى مستويات موت الخلايا المرتبطة بزيادة NF-التفعيل مقارنةً بصيغ LNP الأخرى، بينما تحفيز LNP-SM102 يؤدي إلى مستويات مشابهة من الموت مع الحد الأدنى من NF-التفعيل في هذه النقطة الزمنية.
للتحقيق في سبب التنشيط الفطري، استخدمنا LNP يفتقر إلى مكون الدهون القابلة للتأين لتحديد المساهمة في التحفيز. يتكون هذا LNP من الكوليسترول وDSPC وALC-0159، وهو نفس التركيب لـ LNP-ALC315 باستثناء عدم وجود الدهون القابلة للتأين (الجدول التكميلي 1).تم فقدان الاستجابة في غياب مكون الدهون القابلة للأيون (الشكل 1C) مما يشير إلى المساهمة الأساسية للدهون القابلة للأيون فيتفعيل.
كما قمنا بالتحقيق في تنشيط مسارات IRF استجابةً لـ LNPs القابلة للتأين. قمنا بقياس زيادة بمقدار 3 و 1.7 مرة عند 48 و 72 ساعة، على التوالي، في تقرير IRF استجابةً لـ LNP-ALC315 مقارنةً بالتحكم (الشكل 2A، B). على عكس ما حدث مع NF-كان لدى LNP-1 استجابة IRF مشابهة لـ LNP-ALC315 بينما أثار LNP-SM102 استجابة أكبر بشكل ملحوظ (الشكل 2A، B). أظهر المحفز TLR R848 أعلى زيادة في استجابة IRF، والتي كانت أعلى بشكل ملحوظ من LNPs الفارغة ومحفز TLR MPLA (الشكل التوضيحي 3B). كما لاحظنا استجابة IRF تعتمد على الجرعة لكل LNP والتي وصلت إلى مستوى ثابت عند (الشكل التوضيحي 2B). بالإضافة إلى ذلك، قمنا بتقييم مساهمة الدهون القابلة للتأين وأظهرنا أن LNP التي تفتقر إلى الدهون القابلة للتأين لم تكن قادرة على تحفيز تنشيط IRF (الشكل 2C). وهذا يشير إلى أن استجابة IRF، مثل NF-يتم تحفيزه بشكل أساسي بواسطة مكون الدهون القابلة للأيون.
بشكل عام، تظهر بياناتنا أن جزيئات LNP القابلة للتأين الفارغة تنشط بشكل أساسيو، إلى حد أقل، عوامل النسخ IRF. تختلف شدة هذه الاستجابات بين LNPs القابلة للتأين، ولكن، من المهم، أنها فقدت في غياب المكون القابل للتأين.
لا يرتبط إدخال LNP بتنشيط عوامل النسخ
تؤدي سلاسل الإشارات، بمجرد بدءها، عادةً إلى NF-وفسفرة IRF في فترة زمنية قصيرة، حتى بسرعة تصل إلىتفعيل NF-وطرق IRF بواسطة LNPs القابلة للتأين بلغت ذروتها عندبعد التحفيز، الذي تأخر مقارنة بالتفعيل بواسطة المحفزات TLR. لمعالجة حركيات الاستجابة الأبطأ، أكدنا أولاً أنها كانت متسقة عند جرعات مختلفة (الشكل التوضيحي 2). ثم افترضنا أن الإندوسيتوز قد يكون مطلوبًا لبدء تفعيل LNPs. باستخدام LNP موسوم بدي أو فلوريسنت، بحثنا في حركيات الامتصاص بواسطة خلايا THP-1. تم تصور الامتصاص خلال ساعتين، وبلغ ذروته في الكثافة عند 48 ساعة (الشكل التوضيحي 4A، B). مع الجرعات الأقل، ينخفض الامتصاص بشكل أسرع عند 72 ساعة. من المحتمل أن يكون الانخفاض ناتجًا عن استهلاك DiO-LNP في الوسط بينما تتكرر الخلايا، مما يجعل الخلايا المكررة حديثًا غير قادرة على امتصاص DiO-LNP (الشكل التوضيحي 4A، B). تظهر هذه النتائج امتصاصًا مستمرًا لـ LNP بواسطة خلايا THP-1 على مر الزمن.
نظرًا لأن هذه البيانات تشير إلى عتبة للاستهلاك، استخدمنا مثبطات الإندوسيتوز لتحديد ما إذا كانت هذه العملية ضرورية بالفعل لتنشيط الخلايا. عالجنا خلايا THP-1 بالدينازور، السيتوكالاسين D، الكلوروكين، أو الميثيل-السيكلودكسترين قبل ساعتين من التحفيز وكل 24 ساعة بعد ذلك من أجل تثبيط مسارات مختلفة من الإندوسيتوز.مثبط الانغماس المعتمد على الكلاثرين (CME) الكلوروكين قلل من استجابات IRF (الشكل التوضيحي 4C). الدينازور أيضًا يثبط CME لكنه لم يؤثر على تنشيط LNP (الشكل التوضيحي 4C). وبالتالي، فإن الانخفاض في استجابة IRF الذي لوحظ مع الكلوروكين هو على الأرجح نتيجة لتأثير غير مستهدف. لم يكن موت الخلايا عاملًا حيث كانت قابلية الحياة لكل حالة ضمنمن التحكم غير المعالج طوال الوقت. لم تؤثر المثبطات الأخرى أو تزيد من الاستجابة. تم ملاحظة تقليل طفيف في امتصاص LNP المسمى بـ DiO بعد 24 ساعة ولم يتم قياس أي تثبيط بعد 48 ساعة (الشكل التوضيحي 5). قامت المثبطات بتقليلواستجابات IRF لمراقبات MPLA و R848 في النقاط الزمنية المبكرة (الشكل التوضيحي 4D). بشكل عام، لم نلاحظ أي تأثير كبير لمثبطات الإندوسيتوز على NF- الذي يتم وساطته بواسطة LNP.وتفعيل IRF. ومع ذلك، نظرًا للحد من التثبيط المرئي، لا يمكننا أن نستنتج بشكل قاطع أن التفعيل لا يتطلب دخولًا إلى الحويصلات.
تحفيز باستخدام جزيئات نانوية دهنية قابلة للأيون يثير زيادة في إشارات مستقبلات المناعة الشبيهة بالجراثيم
تظهر بياناتنا أن خط خلايا المونوسيت يمكن تنشيطه بواسطة LNP القابلة للتأين الفارغة لتحفيز NF-ومسارات IRF، التي تحفز السيتوكينات
الشكل 1 | NF-إشارات النقل في خط خلايا التقرير THP-1. تم معالجة خلايا التقرير THP-1-Dual بـ LNP-1 ، LNP-ALC315 ، LNPSM102 R848مPLA LNP بدون دهون قابلة للتأين، أو الوسائط فقط. تم استخدام كاشف الفوسفاتاز القلوي لقياس NF-تم قياس تراكم المستجيب في السائل الفائق فوق الرواسبتمت مقارنة إنتاج تقرير تغيير الطي بين LNPs تمت مقارنة إنتاج المراسل استجابةً لـ LNP بدون دهون قابلة للتأين مع LNP-ALC315 المشابهة في التركيب. ) (C). يتم تمثيل البيانات كمتوسط ± SEM. تم تقييم الأهمية باستخدام تحليل التباين الأحادي مع اختبار دانييت لمقارنات متعددة.، ، ، .
الشكل 2 | نقل إشارات IRF في خط خلايا التقرير THP-1. تم معالجة خلايا التقرير THP-1-Dual بـ LNP-1 ، LNP-ALC315 ، LNPSM102 R848حركة الشعبية لتحرير أنغولا LNP بدون دهون قابلة للتأين، أو الوسائط فقط. تم استخدام تقرير لوكفيراز لقياس استجابة IRF. تم قياس تراكم التقرير في السائل فوق الطرد خلال 120 ساعة. (تم مقارنة إنتاج تقرير تغيير الطي بين LNPs ) (B). تم مقارنة إنتاج المراسل استجابةً لـ LNP بدون دهون قابلة للتأين مع LNP-ALC315 المشابهة في التركيب ( ) (C). تم تمثيل البيانات كمتوسط ± SEM. تم تقييم الأهمية باستخدام تحليل التباين الأحادي مع اختبار دانييت للمقارنات المتعددة..
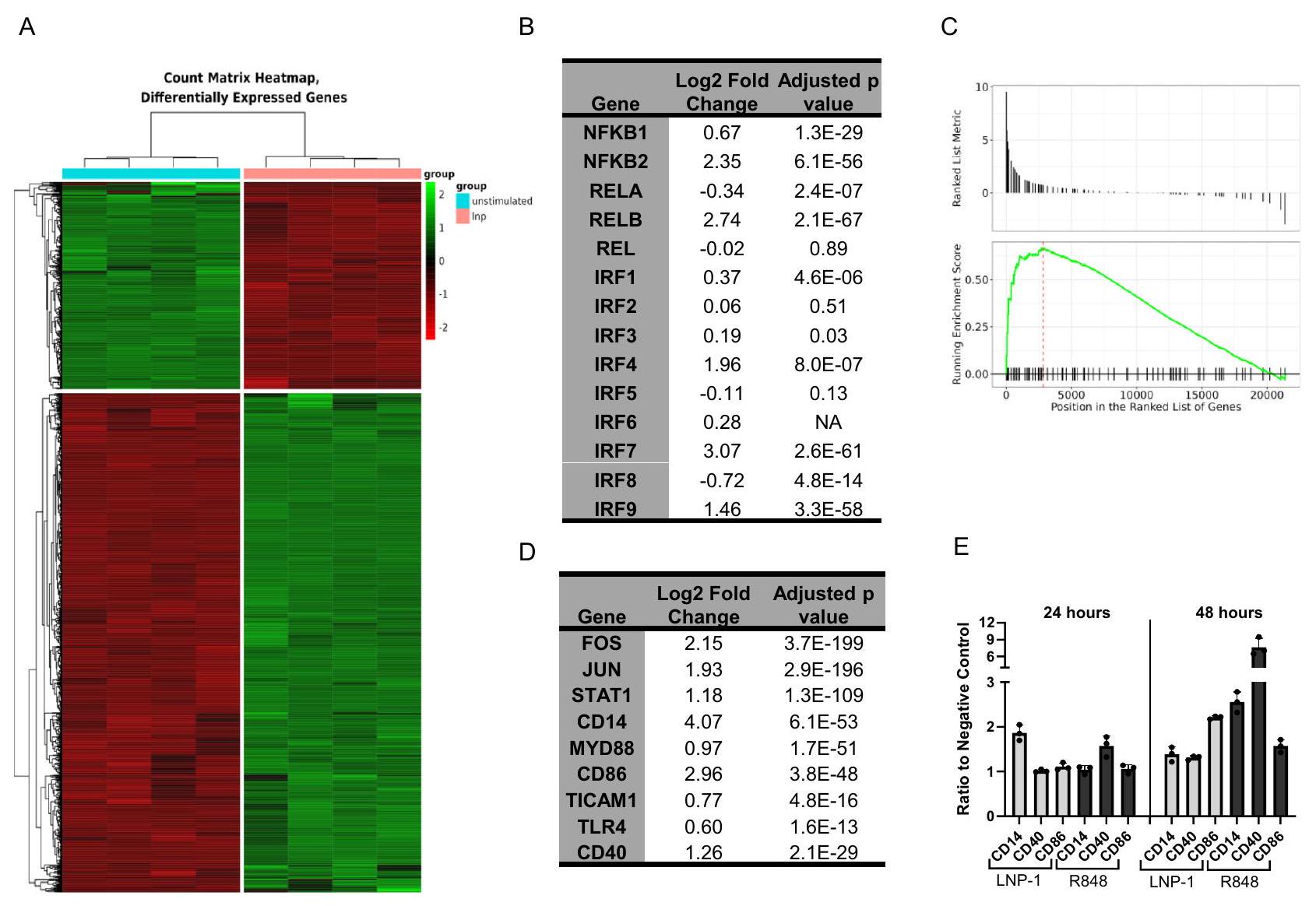
الشكل 3 | استجابة THP-1 النسخية لـ LNP-1. تم معالجة خلايا THP-1-Dual reporter بـ LNP-1أو الوسائط وحدها قبل عزل RNA والتسلسل. تم تسلسل العينات بأربعة مكررات. خريطة حرارية لـ 10,585 جينًا معبرًا عنه بشكل مختلف عند مقارنة خلايا THP-1 غير المنشطة بخلايا THP-1 المنشطة بـ LNP-1 من إجمالي 57,773 جينًا تم اكتشافها (A). جدول NF-جينات الوحدة الفرعية وIRF مع تغيير اللوغاريتم الثنائي المقابل لها والتعديلقيم (ب). تحليل إثراء مجموعة الجينات (GSEA) لمجموعة الجينات KEGG hsa04620 مستقبلات Toll-like مسار الإشارة (C). جدول الجينات المختارة من مسار إشارة مستقبلات Toll-like hsa04620 (D). تم تحفيز خلايا THP-1 باستخدام LNP-1أو الوسائط فقط، وتم قياس تعبير علامات السطح بعد 24 أو 48 ساعة بواسطة تحليل تدفق الخلايا ( ). تُظهر البيانات نسبة شدة الفلورسنت المتوسطة (MFI) للحالة المحفزة إلى التحكم السلبي غير المحفز وتمثل كمتوسط ± SEM (E). تم حساب قيم p المعدلة بواسطة إجراء بنجاميني-هوشبرغ. وملفات التهابيةلتعريف cascades الإشارة المعنية في تنشيط LNP، قمنا بتحليل الاستجابة النسخية لخلايا THP1 عند تحفيز LNP. اخترنا LNP-1 كنموذج لأنه أثار أقوى تنشيط من بين LNPs المختبرة. عند مقارنة الخلايا المحفزة بـ LNP-1 بالخلايا غير المحفزة بعد 24 ساعة، حددنا 10,585 جينًا معبرًا عنه بشكل مختلف (DEGs) من أصل 57,773 تم قياسها. استخدمنا معيار قطع صارم بمعدل اكتشاف خاطئ قدره 0.01 وقطع تغيير لوغاريتمي بمقدار 2 (LFC) فوق 1 وتحت -1 لتضييق تحليل المسار (الشكل 3A). خمسة جينات تشفر NF-تم قياس الوحدات الفرعية ومن بينها كانت NFKB1 و NFKB2 و RELB مرتفعة بشكل ملحوظ. وقد استوفى اثنان منها معايير DEG، بينما كانت NFKB1 تحت العتبة بقليل. أظهرت تقييمات عوامل تنظيم الإنترفيرون أن IRF3 و IRF4 و IRF7 و IRF9 كانت مرتفعة. يبدأ تنشيط NF-кB transcription لمزيد من NF-الوحدات الفرعية بينما يبدأ IRF حلقة تغذية راجعة إيجابية من خلال إشارات IFN. وبالتالي، فإن التعبير المتزايد عن هذه النسخ يتماشى مع نتائج اختبار المراسل. من بين IRFs التي تم تنظيمها لأعلى، فقط IRF4 و IRF7 استوفيا معاييرنا لـ DEG (الشكل 3B). أظهر IRF3 زيادة صغيرة ولكنها ملحوظة مع LFC قدره 0.19. جين مراسل اللوكيفيراز في نموذج THP-1 لدينا تحت سيطرة محفز ISG54 الحد الأدنى بالتزامن مع خمسة عناصر استجابة محفزة بواسطة IFN. وبالتالي، قد تساهم جميع IRFs التي تم تنظيمها لأعلى في استجابة المراسل المرئية (الشكل 2A-C).
باستخدام قائمة DEGs، حددنا مجموعات الجينات والمسارات في قاعدة بيانات علم الجينات (GO) و hallmark و KEGG لفهم الآليات التي قد تكون مرتبطة بتنشيط استجابات المناعة بواسطة LNP القابلة للتأين. من قاعدة بيانات GO، حددنا 791 مجموعة جينية
تحتوي 70 منها على أكثر من 50 DEG (الشكل التوضيحي 6A). كما حددنا ثلاثة عشر وخمسة مسارات منظمة بشكل مختلف باستخدام قواعد بيانات hallmark و KEGG، على التوالي (الشكل التوضيحي 6B، C). من بين المسارات المحددة، كانت العديد مرتبطة بتنشيط المناعة، بما في ذلك موت الخلايا، إنتاج الإنترفيرون، السيتوكينات المنظمة بواسطة NF-кB، وإشارات TLR (الشكل التوضيحي 6A-C). إن زيادة تنظيم إشارات الإنترفيرون والسيتوكينات المنظمة بواسطة NF-кB تتماشى مع نتائجنا باستخدام نظام مراسل THP-1.
بعد ذلك استخدمنا تحليل إثراء مجموعة الجينات (GSEA)، الذي يأخذ في الاعتبار جميع الجينات بدلاً من DEGs فقط (الشكل 3C). حدد هذا التحليل زيادة تنظيم مسار إشارات TLR، مما يشير إلى آلية التنشيط. ضمن هذا المسار، تم تنظيم الجينات الرئيسية المعنية بتنشيط خلايا تقديم المستضد وتفاعل خلايا T لأعلى (الشكل 3D). أكدنا هذه النتائج على مستوى البروتين من خلال قياس التعبير السطحي لـ CD14 و CD40 و CD86 على خلايا THP-1 بعد التحفيز بـ LNP-1 (الشكل 3D، E). كما هو موضح بواسطة قياس التدفق الخلوي، أدى LNP-1 إلى زيادة تقريبية بمقدار 2 ضعف في CD14 و CD86 عند مستويات التعبير القصوى وزيادة بمقدار 1.3 في CD40 (الشكل 3E). تتوافق هذه البيانات مع LFCs الأعلى لـ CD14 و CD86، 4.07 و 2.96 على التوالي، مع CD40 الذي يظهر LFC قدره 1.26. بالإضافة إلى NF- و IRF، يتم تنشيط عوامل نسخ أخرى استجابةً لـ LNP-1. تم تنظيم JUN و FOS اللذان يشفران AP-1 لأعلى، مما يشير إلى التداخل بين مسارات الإشارة (الشكل التوضيحي 6A-C، الشكل 3D).
الشكل 4 | اعتماد نقل إشارات NF-кB و IRF بواسطة LNP على TLR4 وجزيئات التكيف. تم حضانة خطوط خلايا THP-1-Dual reporter لمدة 72 ساعة مع LNP-1 , LNP-ALC315 , LNP-SM102 , MPLA , TNFa( , R848 , IFNα( أو وسائل الإعلام فقط. تمت مقارنة استجابة NF- و IRF بين خطوط خلايا النوع البري (WT) وخطوط خلايا TLR4 knockout (A) وبين WT وخطوط خلايا MyD88 أو TRIF knockout
(B). تم حضانة خلايا THP-1 WT مع المحفزات في وسائل الإعلام التي تحتوي على مصل (FBS) أو بدون مصل (C). تم تقييم استجابات NF- kB و IRF وتقريرها كنسبة من إنتاج المراسل للحالة المحفزة إلى التحكم في وسائل الإعلام فقط ( ). تم تمثيل البيانات كمتوسط ± SEM. تم تقييم الأهمية بواسطة -اختبار (A) أو ANOVA أحادي الاتجاه مع اختبار المقارنات المتعددة لدونيت (A، B) أو اختبار الطالب -اختبار .
يبدأ TLR4 الإشارات من خلال MyD88 استجابةً لـ LNPs القابلة للتأين
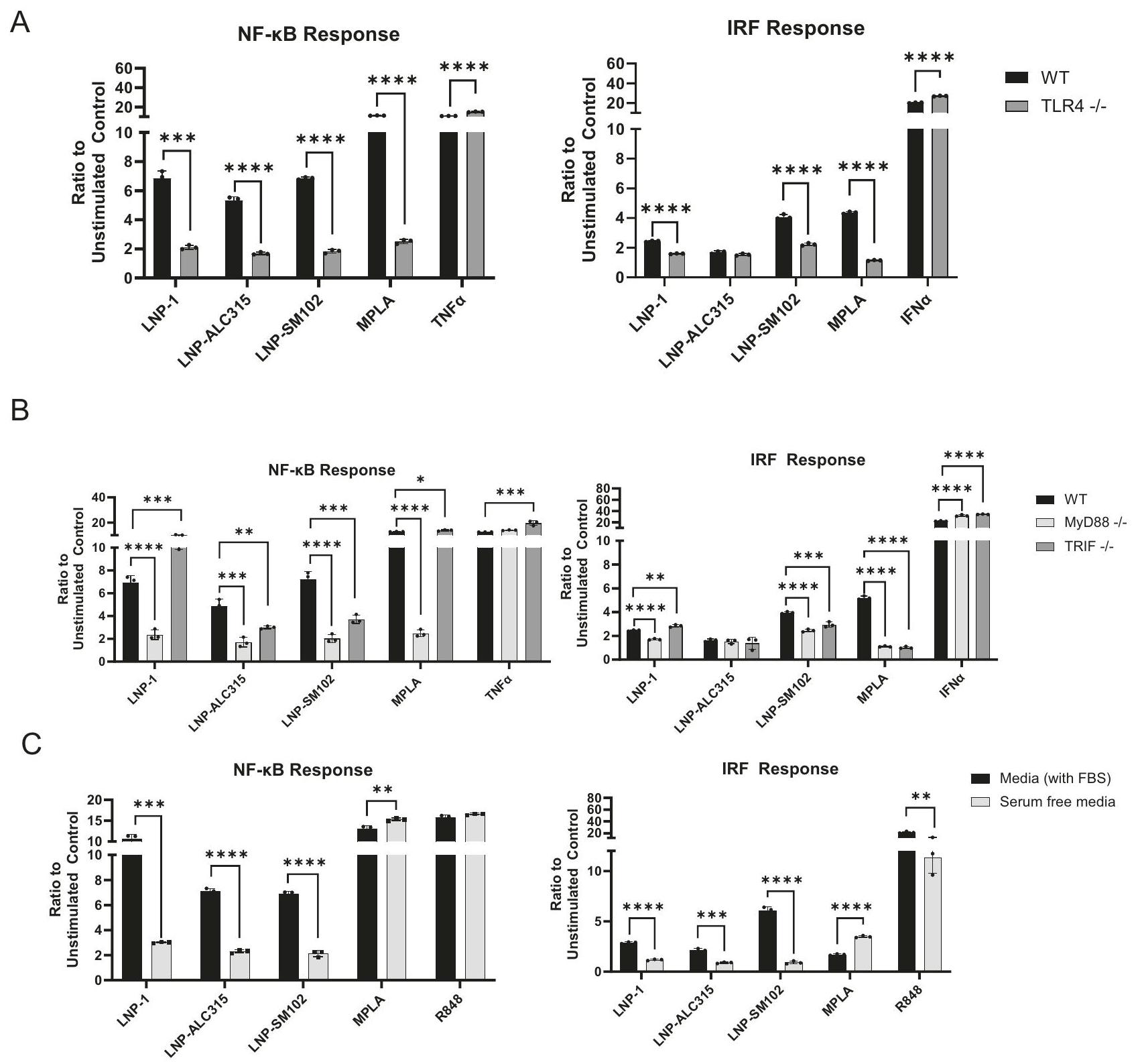
استنادًا إلى زيادة تنظيم مسار إشارات TLR في مجموعة بيانات النسخ الجينية لدينا والنتائج السابقة حول LNP القابلة للتأين والموجبة، افترضنا أن TLR4 يلعب دورًا رئيسيًا في تنشيط LNP القابلة للتأين للوحيدات . استخدمنا مراسل THP-1 مع جين TLR4 الذي تم تعطيله لتحديد ما إذا كانت زيادة تنظيم NF- و IRF في THP-1 مدفوعة بواسطة مسار الإشارة هذا (الشكل التوضيحي 7). عند مقارنة THP-1 مع TLR4-KO، قمنا بقياس انخفاض ملحوظ في استجابات NF-кB و IRF لجميع LNPs القابلة للتأين (الشكل 4A) (الشكل التوضيحي 8A). ومع ذلك، لم تؤدي خسارة TLR4 إلى إلغاء نشاط NF- أو IRF تمامًا (الشكل 4A) (الشكل التوضيحي 8A). تشير هذه النتائج إلى أن TLR4 قد يكون المستقبل الرئيسي المسؤول عن تنشيط خلايا THP-1 بواسطة LNP القابلة للتأين، ولكن قد تساهم مسارات أخرى. لضمان
دقة بياناتنا، اختبرنا LNP الخاص بنا لوجود الإندوتوكسين، الذي يمكن أن يحفز أيضًا TLR4، ووجدنا أنه كان أقل من حد الكشف في الاختبار.
بالإضافة إلى ذلك، يمكن أن يتم تحفيز TLR4 بواسطة موت الخلايا . هناك احتمال أن منتجات موت الخلايا التي يتم تحفيزها بواسطة LNP قد تبدأ إشارات TLR4 التي تم اكتشافها في هذه الدراسة. ومع ذلك، أظهرت دراسة حديثة أن الميزات الهيكلية للدهون القابلة للتأين من تركيبات LNP المختلفة تشارك TLR4 مما يؤكد نتائجنا مع TLR4-KO وإزالة الدهون القابلة للتأين. تدعم هذه النتائج معًا حدوث إشارات TLR4 بشكل منفصل عن موت الخلايا .
لتقييم المزيد من مشاركة TLR4 في استشعار LNPs القابلة للتأين، بحثنا في بروتينات التكيف MyD88 و TRIF، التي تشارك في سلسلة الإشارات (الشكل التوضيحي 7). أظهر تحليل بيانات النسخ الجينية زيادة تنظيم جينات MyD88 و TICAM1، الأخيرة تشفر TRIF، استجابةً لـ LNP-1 (الشكل 3D). تم تقليل كل من NF- و IRF
التنشيط بواسطة جميع LNPs القابلة للتأين في خلايا THP-1 التي تفتقر إلى MyD88 بعد 72 ساعة (الشكل 4B) (الشكل التوضيحي 8B). في المقابل، أظهرت خلايا THP-1 التي تفتقر إلى TRIF زيادة طفيفة في تنشيط NF- بواسطة LNP-1. ومع ذلك، أثارت LNPALC315 و LNP-SM102 استجابة مخفضة في TRIF-KO، على الرغم من أنها ليست بنفس الدرجة كما في MyD88-KO (الشكل 4B) (الشكل التوضيحي 8B). تم رؤية نتائج مماثلة مع استجابة IRF في TRIF-KO. توضح هذه النتائج أنه على الرغم من أن جميع LNPs الثلاثة يتم التعرف عليها بواسطة TLR4، إلا أن لديها اعتمادًا متنوعًا على بروتينات التكيف على الرغم من أن الإشارات كانت بشكل أساسي عبر مسار TLR4/MyD88.
لتحقيق المزيد من التحقيق في المسارات البديلة وتقييم دور TLR4 في استشعار LNPs القابلة للتأين، قمنا بتقييم تنشيط NF- و IRF باستخدام خط خلايا A549 بنفس تركيبات المراسل. خط خلايا A549 مشتق من خلايا الظهارة القاعدية الهوائية التي لا تعبر عن TLR4 . لم يتم ملاحظة أي استجابة NF- أو IRF لأي LNP (الشكل التوضيحي 9) مما يشير إلى الدور الرئيسي لـ TLR4 في استشعار LNPs وفشل في تحديد مسارات بديلة.
بعد ذلك، افترضنا أن المصل يسهل تفاعل LNP-TLR4 وامتصاصه، كما تم إظهاره مع LPS . لاختبار ما إذا كان المصل مطلوبًا لاستجابات LNP، استخدمنا ظروفًا خالية من المصل وأظهرنا انخفاضًا كبيرًا في كل من تنشيط NF- و IRF (الشكل 4C، الشكل التوضيحي 8C). أظهرت الثقافات الخالية من المصل انخفاضًا في التكاثر وتم استخدام عينات خالية من المصل غير المحفزة للتطبيع. توضح هذه النتائج أن LNPs القابلة للتأين تعتمد على وجود المصل لتنشيط THP-1. لتحديد ما إذا كان تفاعل TLR4 يحدث أيضًا في الإندوسوم، بحثنا في التوضع المشترك لـ LNP المسمى DiO مع TLR4 في الإندوسومات المبكرة، كما تم تمييزها بواسطة Rab5a، والتي لم يتم ملاحظتها (الشكل التوضيحي 8D). تشير هذه البيانات بشكل أكبر إلى أن تفاعل TLR4-LNP يحدث عند سطح الخلية (الشكل التوضيحي 7).
إضافة mRNA لا تعزز استجابات NF-кB أو IRF التي تحفزها LNP
بعد ذلك سعينا لتحديد دور mRNA في التنشيط الفطري الذي تحفزه لقاح BNT162b2. على الرغم من إجراء تعديلات على مكون mRNA لـ BNT162b2 لتقليل قدرته على التحفيز المناعي , افترضنا أن لقاح mRNA BNT162b2 سيؤدي إلى زيادة تنشيط NF- و IRF مقارنةً بـ LNP وحده. بشكل مفاجئ، أظهرت بياناتنا أن الاستجابة لـ mRNA-LNP كانت مشابهة أو أقل من LNP الفارغ المقابل عند معالجة خلايا مراسل THP-1 (الشكل 5A، B). تم إظهار أن R848 يثير تنشيط NF و IRF بشكل قوي (الشكل 1A، الشكل 2A)، مما يدل على استجابة TLR7/8 وظيفية. وبالتالي، لم يقم مكون mRNA بزيادة إشارات NF- و IRF ولم يحفز مسارات إضافية في غياب TLR4 أو MyD88 أو TRIF (الشكل 5A، B). علاوة على ذلك، كان تعبير CD86 على THP-1s مشابهًا بين mRNA-LNPs وخلايا LNP الفارغة المعالجة (الشكل 5C). بالإضافة إلى ذلك، لم تؤدي mRNA-LNP و LNP الفارغ إلى إثارة NFأو استجابات IRF في خلايا A549 (الشكل التوضيحي 9). تشير هذه البيانات إلى أن LNPs القابلة للتأين قادرة تمامًا على تنشيط نموذج أحادي النواة للمناعة الفطرية في غياب mRNA وأن إضافة mRNA لم تؤدِ إلى زيادة الاستجابة.
نقاش
إن ظهور لقاحات mRNA المرخصة يمثل تحولًا جذريًا في استراتيجيات التطعيم، حيث يوفر منصة تحفز استجابات مناعية تكيفية قوية ويمكن إنتاجها بسرعة.. من المثير للاهتمام أن هذه اللقاحات تفتقر إلى مادة مساعدة محددة، مما يشير إلى وجود خصائص مساعدة داخلية. على الرغم من أن الآلية الدقيقة لا تزال غير مفهومة تمامًا، إلا أن كل من مكون RNA و LNP متورطان في تحفيز المناعة الفطرية.فهم آلية فعالية المحفزات في لقاحات mRNA قد يوفر رؤى لتحسين المناعية و/أو ردود الفعل السلبية للقاحات المستقبلية. لذلك، كان هدف هذه الدراسة هو تحديد المسارات المناعية الفطرية التي يتم تنشيطها بواسطة جزيئات LNP القابلة للتأين الفارغة، بما في ذلك تلك المستخدمة في لقاحات BNT162b2 وmRNA-1273.
باستخدام خط خلايا وحيدة النواة THP-1 مع أنظمة تقارير، وجدنا أن الثلاثة LNPs القابلة للتأين تحفز NF-وتفعيل IRF بدرجات متفاوتة. كان التفعيل يعتمد إلى حد كبير على TLR4 من خلال MyD88 محول، مع اعتماد TRIF الحد الأدنى. دعم تحليل التعبير الجيني لخلايا THP-1 المنشطة بواسطة LNP1 هذه النتائج، حيث أظهر زيادة في تنظيم إشارات TLR. لا يوجد NF-أو حدث استجابة IRF في غياب مكون الدهون القابلة للأيون، مما يشير إلى أنه المحرك لتنشيط المناعة الفطرية ويقترح أن الاختلافات الهيكلية في الدهون القابلة للأيون تؤثر على حجم الإشارة. بالإضافة إلى ذلك، أشارت بياناتنا إلى أن الإشارة كانت تعتمد بشكل أساسي على TLR4-MyD88، ولكن من المحتمل أن يتم تحفيز مسارات إضافية بواسطة LNPs القابلة للأيون. علاوة على ذلك، كانت التنشيط يتطلب المصل، ولكن ليس الإندوسيتوز، مما يدعم دور TLR4 المعبر عنه على السطح.
من الجدير بالذكر أننا لاحظنا أن الجسيمات النانوية المحتوية على mRNA لم تزيد من IRF أو NF-الإشارات في خلايا THP-1 مقارنةً بـ LNPs الفارغة، مما يشير إلى أن LNP هو المعزز الرئيسي في لقاح BNT162b2 mRNA. بينما قد يتم تفعيل مسارات أخرى، فإن المسار السائد المستخدم هو عبر TLR4/MyD88 على سطح الخلية.
تفاوتت الاستجابات عبر LNPs. أثار LNP-SM102 (تركيبة mRNA-1273) أكبر استجابة IRF، بينما كان LNP-1 الأقوى في NF-استجابة، وأظهرت LNP-ALC315 (صيغة BNT162b2) NF مشابهةالتفعيل لـ LNP-SM102 وتفعيل IRF مشابه لـ LNP-1. لم يُلاحظ أي استجابة بدون مكون الدهون القابلة للتأين، مما يؤكد دوره كمحرك لـ NF-وتفعيل IRF. على الرغم من أننا لم نختبر ذلك مع LNP-SM102، من المحتمل أن يكون الدهون القابلة للأيون هي المحرك، لكن لا يمكننا استبعاد دور الدهون المPegylated. نفترض أن الاختلافات الهيكلية قد تؤثر على قوة NF-وتفعيل IRF، كما هو موضح مع اختلافات LPS عبر الأنواع البكتيرية في إشارات TLR4وبالتالي، فإن تصميم الدهون القابلة للتأين هو وسيلة محتملة يمكن من خلالها تعديل جزيئات الدهون النانوية لتعزيز فعالية اللقاح أو تقليل ردود الفعل السلبية.
تدعم نتائجنا حول تنشيط المناعة الفطرية بواسطة LNP ما أظهره كونورز وزملاؤه، الذين أثبتوا أن تجمعات الخلايا الشجرية والوحيدة من PBMCs المحفزة بـ LNP-1 تظهر تنشيط TBK1 و IRF7 بين 6 و 24 ساعة.. ينشط TBK1 كل من IRF3 وIRF7، وبما أن تقرير IRF لدينا يتم إنتاجه عند تنشيط أي IRF، فقد يكون IRF7 هو مصدر إنتاج التقرير الذي تم رؤيته في THP-بينما يتم تنشيط IRF3 downstream من TLRs، مثل TLR4لم تُظهر بياناتنا زيادة قوية في تنظيم IRF3 ولا اعتماد TRIF على LNP-1. ومع ذلك، تم تنظيم IRF4 وIRF7 بشكل مرتفع، مما يشير إلى أنهما يساهمان في الاستجابة لـ LNP-1. نظرًا لأن LNP-ALC315 وLNP-SM102 أظهرت بعض الاعتماد على TRIF، قد يلعب IRF3 دورًا لهذه LNPs. معًا، تُظهر بياناتنا أن LNPs القابلة للأيونات الفارغة يمكن أن تحفز كل من NF-واستجابات IRF، على الرغم من أن تفعيل IRF النهائي يتطلب مزيدًا من التحقيق.
باستخدام خطوط نقص THP-1، وجدنا أن تنشيط الخلايا كان يعتمد بشكل أساسي على إشارة TLR4-MyD88. دعمًا لنتائجنا، فإن LNP مع الدهون الأيونية MC3 يثير ردود فعل سلبية في الفئران من خلال مسار TLR4-MyD88، بغض النظر عن TRIF.يمكن اكتشاف بعض الجسيمات النانوية الكاتيونية بواسطة TLR4، وبالتالي فهي قادرة على تحفيز الاستجابات المناعية الفطرية.يبدو أن الشحنة وبنية مكونات الدهون تساهم في قدراتها التحفيزية. وقد تم إثبات أن التغيرات في مجموعات الرأس للدهون الكاتيونية تعدل إشارات TLR4.علاوة على ذلك، أظهرت دراسة حديثة أهمية هياكل مجموعات الرأس الأمينية في تفاعلات TLR4 مع الدهون القابلة للتأين المختلفة عن تلك التي تم التحقيق فيها في هذه الدراسة.بالإضافة إلى ذلك، يمكن أن تعزز التغيرات الهيكلية في طول سلسلة الهيدروكربون وكذلك تعديلات مجموعة الرأس أو تلغي تنظيم جزيئات التكامل المساعدة على خلايا الدندريت في الفئران.. وبالتالي، يمكن أن تؤدي التغيرات الهيكلية إلى تغيير قوة الإشارات. ومع ذلك، على الرغم من الاختلافات الهيكلية في الدهون، يبدو أن TLR4-MyD88 هو مسار شائع لتنشيط المناعة الفطرية. علاوة على ذلك، لا يوجد NF-تم الكشف عن استجابات IRF في خلايا A549، التي تفتقر إلى TLR4، مما يدعم بشكل أكبر دور TLR4 في التنشيط بواسطة LNPs القابلة للتأين.
تظهر بياناتنا بشكل أكبر أن بروتينات المصل من المحتمل أن تكون متورطة، حيث كان المصل مطلوبًا لتفعيل LNP بواسطة LNPs المختبرة. قد تساهم هذه البروتينات في تشكيل الغلاف البروتيني المحيط بـ LNP، مما يسهل امتصاصه بواسطة المستقبلات، أو قد تشمل بروتين ربط LPS (LBP)، الذي يعد ضروريًا لتفاعل LPS مع TLR4.. ومع ذلك، هناك حاجة إلى مزيد من التحقيق لتوضيح الآليات المحددة المعنية. تعتبر الغلاف البروتيني مهمًا بشكل خاص لأن تركيبه يعتمد على البيئة المحلية لـ.
الشكل 5 | الاستجابة لـ LNPs مقارنة بـ mRNA-LNP في THP-1. تم تحضين خلايا THP-1-Dual reporter مع LNP-1 ، LNP-ALC315 ، LNPSM102 mRNA-LNP-1mRNA-LNPALC315مPLATNFaإنترفيرون ألفاR848، أو وسائل الإعلام وحدها لمدة 72 ساعة. NF-تمت مقارنة استجابة NF-kB وIRF بين خطوط الخلايا من النوع البري (WT) وخطوط الخلايا المعطلة لـ TLR4 (A) وبين WT وخطوط الخلايا المعطلة لـ MyD88 أو TRIF (B). تم تقييم استجابات NF-kB وIRF وتقريرها كـ
نتيجة لذلك، قد يؤدي الإيصال العضلي للجزيئات النانوية الدهنية إلى تكوين غلاف بروتيني يختلف عن ذلك الذي يتم الحصول عليه من الجزيئات النانوية الدهنية التي يتم إيصالها عن طريق الوريد. لذلك، قد يتطلب فهم دور الغلاف البروتيني في تنشيط المناعة الفطرية بواسطة الجزيئات النانوية الدهنية تقييمًا في سياقات بيولوجية محددة.
حيث لا الـ NF-لم يتم إلغاء استجابة IRF تمامًا مع غياب TLR4 أو ملحقاته، ونشتبه في أن هناك مسارات إضافية تساهم في اكتشاف LNPs القابلة للتأين. ومن الجدير بالذكر أن نموذج الفأر المعطل لـ TLR4 الذي تم استخدامه في الدراسة التي أجراها لي وآخرون لم يظهر فقدان الاستجابة للقاح.أظهر آخرون أن LNP قابل للتأين مختلف يمكن أن يثير ردود فعل مناعية في الفئران من خلال TLR4، مما يشير إلى أن الاختلافات في قوة التنشيط والنمذجة قد تؤثر على النتائج.. نظرًا لأن بياناتنا تشير إلى أن تنشيط LNP ليس معتمدًا تمامًا على إشارات TLR4، فقد توفر مسارات أخرى تحفيزًا كافيًا في غياب TLR4. بالإضافة إلى ذلك، فإن إشارات وتعبير TLR في البشر والفئران ليست متسقة تمامًا وقد تؤدي إلى نتائج متضاربة..
أظهر تحليل النسخ للاستجابات الخلوية لـ LNP-1 زيادة في تنظيم العديد من المسارات المتعلقة بالالتهاب، وموت الخلايا، والإجهاد، مما قد يوفر رؤى حول طرق بديلة للتعرف على LNP القابلة للأيون. وقد تم إثبات أن مواد التحفيز الأخرى تستخدم تلك الآليات لتنشيط المناعة.على سبيل المثال، المحفزات القائمة على الألومنيوم والصابونين تسبب إجهادًا خلويًا. علاوة على ذلك، فإن اللقاحات التي تحفز موت الخلايا قد زادت من الفعاليةنظرًا لأن الجسيمات النانوية الكاتيونية قد لوحظ أنها تبدأ موت الخلايا الالتهابية، فإن هذه طريقة محتملة لتنشيط المناعة للجسيمات النانوية القابلة للتأين وتتطلب مزيدًا من التحقيق..
قد تكون الآليات التي من خلالها تنشط LNPs القابلة للتأين خلايا THP-1 مرتبطة أيضًا بالتأخير في الحركيات الملحوظة. خلال تحقيقنا في حركيات استجابة THP-1 لـ LNPs المختلفة، حدث ذروة الإشارة عند 48 ساعة. على الرغم من أن كونورز وآخرين وجدوا أن إشارة LNP-1 كانت الأعلى عند 24 ساعة، إلا أنهم لم يحققوا في النقاط الزمنية اللاحقة.نظرًا لأن MPLA (منبه TLR4) و R848 (منبه TLR7/8) أثارا استجابات قوية لـ NF-кB خلال 24 ساعة، توقعنا رؤية استجابة أكثر وضوحًا للجزيئات النانوية الدهنية في هذه النقطة الزمنية. قد يكون أحد التفسيرات لتأخر تفعيل الاستجابة الملحوظ هو أن البلعمة الخلوية مطلوبة. نظرًا لأن الخلايا تنمو في وسط يحتوي على مصل، ستتكون غلاف بروتيني حول الجزيئات النانوية الدهنية.. هذه البنية ستعيق بشكل ستيري اكتشاف TLR4 حتى يحدث مستوى معين من الانهيار أثناء الإندوسيتوز. لذلك، اعتبرنا أن الديناميات قد تعكس هذا المتطلب. باستخدام LNP المعلم بـ DiO، أظهرنا من خلال قياس التدفق والميكروسكوب المناعي الفلوري أن خلايا THP-1 تأخذ LNP خلال ساعتين وتستمر في ذلك على مدى 72 ساعة المقاسة. حيث كان هذا يشير إلى عتبة من الامتصاص لتنشيط الوساطة بواسطة LNP، استخدمنا مثبطات كيميائية مختلفة لحجب الامتصاص. فقط الكلوروكين أدى إلى تقليل استجابة IRF. على الرغم من أن المثبطات أضعفت الاستجابة للضوابط في النقاط الزمنية المبكرة، إلا أنها قللت فقط بشكل طفيف من امتصاص LNP خلال أول 24 ساعة. وبالتالي، لا يزال من الممكن أن يحدث الإشارة من الإندوسوم. ومع ذلك، فإن TLR4 يستخدم بشكل أساسي MyD88 من سطح الخلية ويتحول إلى الاعتماد على TRIF بعد الإندوسيتوز.نظرًا لأننا لاحظنا بشكل أساسي اعتماد MyD88 في LNPs المختبرة، فإن هذا يدعم تنشيط TLR4 على سطح الخلية. هناك حاجة إلى مزيد من التحقيق لتحديد الآليات الكامنة وراء حركيات تنشيط LNP القابلة للتأين.
قمنا بتوسيع دراستنا للتحقيق في مساهمة مكون mRNA مقارنةً بـ LNP القابل للتأين بمفرده. على الرغم من أنه تم إجراء تعديلات على RNA لجعله أقل تحفيزًا للمناعة، لا يزال هناك احتمال لاكتشاف هذا RNA بطريقة ما بواسطة TLR7/. بالإضافة إلى ذلك، يمكن اكتشاف أي dsRNA متبقي بعد تنقية mRNA بواسطة TLR3 أو PRRs السيتوسولية مثل MDA5. ومع ذلك، تظهر بياناتنا أنه في خلايا THP-1، كانت استجابات NF-кB وIRF لـ mRNA-LNPs المختبرة مشابهة للاستجابات تجاه LNP الفارغة المقابلة لها. وبالتالي، لم يوفر المكون RNA أي فائدة مناعية في هذا النظام النموذجي. تتناقض بياناتنا مع النتائج التي تشير إلى أن MDA5 هو مستقبل مناعي فطري رئيسي للتطعيم بـ mRNA، حيث أن الفئران التي تم حذف MDA5 منها أظهرت انخفاضًا ملحوظًا في الاستجابات المناعية الفطرية والتكيفية بعد التطعيم بـ BNT162b2.. اللقاح المستخدم في تلك الدراسة كان BNT162b2 الذي تم إنشاؤه وفقًا لمعايير GMP، في حين أن LNPs المحتوية على mRNA لدينا تم إنشاؤها باستخدام mRNA منقى بالسيليلوز، وبالتالي يمكن أن تكون مسؤول عن النتائج المختلفة. علاوة على ذلك، قد يأتي تنشيط MDA5 في الجسم الحي من منتجات dsRNA الناتجة عن إجهاد الخلايا بدلاً من بقايا dsRNA الناتجة عن تخليق mRNA..
بشكل عام، تظهر بياناتنا أن ثلاثة من جزيئات الدهون النانوية ذات التغيرات الهيكلية في الدهون القابلة للأيون، يتم اكتشافها بواسطة TLR4 وتفعيل NF- وعوامل النسخ IRF في خلايا المناعة الفطرية. من المعروف أن إشارات TLR تؤدي إلى زيادة تنظيم الجزيئات المعنية في التواصل بين خلايا T وإنتاج السيتوكينات، وقد تم استخدام المحفزات TLR كمواد مساعدة في التجارب السريرية واللقاحات المرخصة من أجل هذا التنشيط.تشير بياناتنا النسخية التي تظهر زيادة واسعة في تنظيم الجزيئات المشاركة في مسارات تنشيط الخلايا إلى أن LNPs القابلة للتأين قد توفر تحفيزًا مناعيًّا إضافيًّا من خلال مسارات لم يتم تأكيدها بعد. قد يسمح التحقيق في الآلية وراء تفاعل TLR4 بإمكانية تعديل التركيب الدهني لتحقيق توازن بين التحمل وقوة التحفيز. بالإضافة إلى ذلك، تشير بياناتنا إلى أن LNP، أكثر من RNA، هو المحرك الرئيسي لتنشيط أحادية النواة THP-1 الناتج عن لقاح mRNA. لذلك، يمكن أن تركز الجهود على فهم كيفية إنشاء LNPs لاستهداف مسارات الإشارة المحددة بدلاً من المزيد من تعديل mRNA للمساعدة في توجيه تصميم اللقاحات والمساعدات المناعية من أجل لقاحات mRNA أكثر أمانًا وفعالية، وربما أكثر ديمومة.
طرق
خطوط الخلايا وزراعتها
THP-1-Dوالتم الحصول على خط خلايا وحيدة النواة البشرية المزدوجة المراسل (InvivoGen، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) لاستخدامه في تجارب المراسل، والميكروسكوبية، وقياس التدفق الخلوي. يعبر هذا الخط الخلوي المعلق بشكل مستقر عن مراسل الفوسفاتاز القلوي المفرز (SEAP) القابل للتحفيز بواسطة NF-ومراسل لوسيفيراز لوسيا سري قابل للتحفيز بواسطة تنشيط مسار IRF. تم الحصول أيضًا على طفرات لجين MyD88 وTRIF وTLR4 في خلفية المراسل المزدوج من InvivoGen. تم زراعة خلايا THP-1 في RPMI 1640 مع 2 مللي مول من L-glutamine (Cytiva، مارلبورو، ماساتشوستس، الولايات المتحدة الأمريكية) مضافًا إليها 25 مللي مول من HEPES (Thermo Fisher، وولثام، ماساتشوستس، الولايات المتحدة الأمريكية)،FBS المعطل حرارياً، وPen-Strep ). تم الحفاظ على الخلايا في معاختيار المضادات الحيوية، بلاستيسيدين ( ) وزيوسين ( ) (إنفيفوجين، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية)، تم تطبيقها في كل تمريرة أخرى وفقًا لتوصيات الشركة المصنعة.
A549-Dوالتم الحصول على خط خلايا الظهارة التنفسية البشرية المزدوجة المراسل (InvivoGen، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) لاستخدامه في تجارب المراسل. تم زراعة هذه الخلايا في DMEM مع 2 مللي مول من L-glutamine (Thermo Fisher، وولثام، ماساتشوستس، الولايات المتحدة الأمريكية) معززة بـمصل فتي (FBS) المعطل حرارياً وPen-Strep ). تم الحفاظ على الخلايا في مع.
تحضير mRNA وتشكيل جزيئات الدهون النانوية
تم تحسين كودون بروتين السنبلة المتغير أوميكرون SARS-CoV-2 المعدل بالديبروتين (S2P) واستنساخه في بلازميد قالب النسخ في المختبر الذي يحتوي على محفز T7، و تمتلك مناطق UTR وذيل بولي (A) بطول 100 نيوكليوتيد. تم تخليق mRNA وتغطيته أثناء النسخ باستخدام مجموعة النسخ Megascript (Thermo Fisher، رقم الكاتالوج AMB 1334) ونظام CleanCapTM ثنائي النوكليوتيد (Trilink Biotechnologies)، وتم ترسيبه وتنقيته باستخدام طريقة كروماتوغرافيا قائمة على السليلوز معدلة. تم تقييم الطول وسلامة mRNA باستخدام نظام Agilent Bioanalyzer 2100. تم تأكيد إزالة ملوثات RNA مزدوج الشريطة (dsRNA) باستخدام طريقة النقاط، وتم قياس مستويات الإندوتوكسين باستخدام اختبار Genscript ToxiSensor الكروموجيني. ). تم تخزين mRNA S2P مجمدًا ( ) في في ماء خالٍ من النوكلياز والمواد المسببة للحمى حتى الاستخدام. تم احتواء mRNA في جزيئات الدهون النانوية كما هو موصوف سابقًا (51). تم خلط مكونات الدهون LNP في محلول إيثانولي، ثم تم خلطها بسرعة مع مرحلة مائية تحتوي على mRNA، وتمت عملية الغسيل والتركيز إلىوتم تخزينه فيحتى يتم الاستخدام (الجدول التكميلي 1، الشكل التكميلي 1).
كان LNP-ALC315، الذي يتبع تركيبة BNT162b2، مكونًا من الدهون الأيونية ALC-0315، والكوليسترول، وDSPC، ودهون PEG ALC-0159. كان LNP-SM102، الذي يتبع تركيبة mRNA-1273، يتكون من الدهون الأيونية SM-102، والكوليسترول، وDSPC، وPEGDMG PEG-lipid. تم تصميم تركيبة LNP ثالثة، تُعرف باسم LNP-1، وهي ملكية لشركة Acuitas Therapeutics. يتم وصف الدهون الملكية وتركيبة LNP في براءة الاختراع الأمريكية US10,221,127. تتكون LNP-1 من دهون أيونية ملكية، وDSPC، وكوليسترول، وPEG-lipid. تم أيضًا وسم LNP-1 بعلامة الفلورسنت DiO وتم تسميتها DiO-LNP. كانت التركيبة الرابعة التي تم إعدادها هي LNP، المعروفة باسم LNP-no ionizable lipid، والتي تحتوي فقط على كوليسترول، وDSPC، وALC-0159 PEG-lipid. تم تصنيف LNPs من حيث حجمها الهيدروديناميكي، ومؤشر التوزيع المتعدد (PDI)، باستخدام تشتت الضوء الديناميكي (DLS)، وتم قياس كفاءة الاحتواء والتركيز باستخدام اختبار RiboGreen RNA (Invitrogen، Cat# R11490). كان حجم LNP-ALC315 الفارغ المستخدم 54.07 نانومتر مع PDI قدره 0.28 وإمكانات زيتا قدرها -4.447. كان حجم LNP-SM102 الفارغ المستخدم 51.590 نانومتر مع PDI قدره 0.1605 وإمكانات زيتا قدرها -4.417. كانت LNP بدون الدهون الأيونية أكبر بحجم 80.26 نانومتر مع PDI قدره 0.05 وإمكانات زيتا قدرها -8.548. تم تصنيع mRNA-LNPs باستخدام LNP-1 و LNP-ALC315، وتم تسميتهما mRNA-LNP-1 و mRNA-LNP-ALC315. كان لدى mRNA-LNP-ALC315 كفاءة احتواء قدرهاالقطر، 0.1090 PDI، وإمكانات زيتا -11.55. يتم إعطاء تركيزات LNP كمعادل لـ mRNA لتمكين المقارنة مع mRNA-LNP كتركيز لـ mRNA.
اختبارات المراسل
لكل من NF-واختبارات تقارير IRF باستخدام خط خلايا THP-1، تم إعادة تعليق الخلايا في RPMI 1640 مع 2 مللي مول من L-glutamine (Cytiva، مارلبورو، ماساتشوستس، الولايات المتحدة الأمريكية)،مصل بقري مفعّل بالحرارة، وPen-Strep ) بتركيز خلاياوتم توزيع 1 مل لكل بئر من صفيحة 24 بئر. تشمل المواد الكيميائية المستخدمة بشكل فردي لعلاج خلايا THP-1 LNP-1 و LNP-ALC315 و LNP-SM102 و LNP-بدون دهون قابلة للتأين و mRNA-LNP-1 و mRNA-LNP-ALC315 و VacciGrade R848 (InvivoGen، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) و MPLA الاصطناعية (InvivoGen، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) و IFNa (Miltenyi، غايثربيرغ، ماريلاند، الولايات المتحدة الأمريكية) و LPS (Sigma-Aldrich، سانت لويس، ميزوري، الولايات المتحدة الأمريكية) و TNFa (Thermo Fisher، وولثام، ماساتشوستس، الولايات المتحدة الأمريكية). تم توفير LNPs الفارغة و mRNA-LNPs بسخاء من مختبر العلّامة وجامعة بنسلفانيا التي تم تصنيعها كما هو موضح أعلاه..
لتحليل الإندوسيتوز، تم ترك الخلايا دون معالجة أو معالجتها بمثبطات الإندوسيتوز الدينازور.فيشر ساينتيفيك، هامبتون، نيوهامبشير، الولايات المتحدة الأمريكية)، سيتوكالاسين D (فيشر ساينتيفيك، هامبتون، نيوهامبشير، الولايات المتحدة الأمريكية)، الكلوروكين (فشر ساينتيفيك، هامبتون، نيوهامبشير، الولايات المتحدة الأمريكية) أو ميثيل-سيكلوديكسترينتمت معالجة خلايا THP-1 لمدة ساعتين قبل إضافة المحفزات وكل 24 ساعة بعد ذلك. بعد إعادة تعليق العامل بشكل كامل مع خلايا THP-1،تمت إزالة من السائل العلوي في النقاط الزمنية المحددة لقياس جزيئات التقرير. تم أخذ عدّ الخلايا قبل إزالة السائل العلوي في كل يوم لضمان منحنيات نمو مشابهة. بالنسبة لـ NF-اختبار المراسلتم خلط السائل العلوي معمن محلول كوانتي-بلو (InvivoGen، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) في لوحة 96 بئر وتم حضنه فيمعلمدة 24 ساعة. تم قراءة الامتصاص عند 600 نانومتر باستخدام جهاز قراءة الألواح GlowMax من بروماجا (بروماجا، ماديسون، ويسكونسن، الولايات المتحدة الأمريكية). بالنسبة لاختبار تقرير IRF،تم خلط السائل العلوي معتم استخدام محلول Quanti-Luc (InvivoGen، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) وتم قراءة اللمعان على الفور عند دمج 0.1 ثانية باستخدام جهاز قراءة الألواح GlowMax من Promega. تم إجراء نسخ بيولوجية وتقنية لكل حالة في كل اختبار.
لكل من NF-واختبارات تقرير IRF باستخدام خط خلايا A549،تم زراعة الخلايا في كل بئر من أطباق 48 بئر بحجم نهائي من وتم حضنه في معتم استبدال السائل العلوي بوسائط جديدة وتم إضافة التحفيز. تم حضن الخلايا لمدة الفترة الزمنية المحددة وتم إزالة السائل العلوي لاختبار التقرير كما هو موضح أعلاه.
المجهر الفلوري
تم حضانة الخلايا مع أو بدون LNP الموسوم بـ DiO (DiO-LNP) بتركيز خلايافيمعفي صفيحة 24 بئر.
تم توفير DiO-LNP بسخاء من قبل مختبر العلامة وجامعة بنسلفانيا كما هو موضح أعلاه.بعد الحضانة، تم جمع الخلايا، وغسلها، وإعادة تعليقها في PBS. تم السماح للخلايا بالاستقرار على شريحة زجاجية في قاع أطباق 6 آبار لمدة 30 دقيقة. تم إزالة السائل الفائق برفق، وثبتت الخلايا بـمحلول بارافورمالدهيد لمدة 15 دقيقة. بعد التثبيت، تم غسل الخلايا بمحلول PBS لمدة دقيقتين مرتين. للتلوين داخل الخلايا، تم نفاذية الخلايا باستخدام السابونين أثناء التثبيت، وتم حجبها بـألبومين مصل البقر (BSA) (ميلتيني، غيثرزبرغ، ماريلاند، الولايات المتحدة الأمريكية) لمدة ساعة، وتم صبغها بأجسام مضادة مضادة لـ TLR4 (antiCD284) (رقم الكاتالوج 14-9917-82) (ثيرمو فيشر، وولثام، ماريلاند، الولايات المتحدة الأمريكية) وأجسام مضادة مضادة لـ Rab5a (رقم الكاتالوج 11947-1-AP) (ثيرمو فيشر، وولثام، ماريلاند، الولايات المتحدة الأمريكية) لمدة ساعة. ثم تم غسل الخلايا بمزيد من BSA وصبغها بأجسام مضادة ثانوية؛ جسم مضاد مضاد للفأر IgG ألكسا فلور 555 (رقم الكاتالوج 4409S) (تكنولوجيا الإشارة الخلوية، دانفرز، ماريلاند، الولايات المتحدة الأمريكية) وجسم مضاد مضاد للأرنب IgG ألكسا فلور 647 (رقم الكاتالوج 4414S) (تكنولوجيا الإشارة الخلوية، دانفرز، ماريلاند، الولايات المتحدة الأمريكية) لمدة ساعة. ثم، تم صبغ الخلايا بـتم استخدام DAPI (BioLegend، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) لمدة 10 دقائق في الظلام ثم تم غسلها. تم تثبيت الشرائح باستخدام مادة ProLong Gold المضادة للتلاشي (Invitrogen، وولثام، ماساتشوستس، الولايات المتحدة الأمريكية) وتم تصويرها باستخدام المجهر الضوئي المتماسك Zeiss700 (Zeiss، يينا، ألمانيا) باستخدام برنامج ZEN 2.3 SP1.
تدفق الخلايا
خلايا THP-1 بتركيزخلاياتم حضنها مع DiOLNP LNP-1 LNP-ALC315 LNP-SM102 mRNA-LNP-1mRNA-LNP-ALC315فاسيغريد R848 ( ) (إنفيفوجين، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية)، أو الوسط فقط في معلتحقيق في علامات السطح، تم غسل الخلايا بمحلول PBS وتم صبغها بـ Human TruStain FcX (BioLegend، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) عندلمدة 5 دقائق لمنع الارتباط غير المحدد تليها صبغ السطح بالأجسام المضادة المدرجة في الجدول التكميلي 2 فيلمدة 20 دقيقة. تم تثبيت الخلايا باستخداممحلول بارافورمالدهيد لمدة 15 دقيقة. لتقييم حركية امتصاص DiO-LNP، تم غسل الخلايا وتثبيتها على الفور. تم تشغيل الخلايا على جهاز قياس التدفق الطيفي Cytek Aurora وتحليلها باستخدام FlowJo v10.
تسلسل
خلايا THP-1 بتركيزخلاياتم حضنها مع أو بدون LNP-1 فيلمدة 24 ساعة عندقبل استخراج RNA باستخدام مجموعة Qiagen RNeasy Plus Mini Kit (Qiagen، هيلدن، ألمانيا). تم إعداد كل حالة في أربعة مكررات مع 1 مليون خلية لكل عينة. تم قياس RNA باستخدام مجموعة Qubit RNA Broad Range Assay Kit (Thermo Fisher، وولثام، ماساتشوستس، الولايات المتحدة الأمريكية). بالنسبة لجميع العينات، كان إجمالي كمية RNAتم استخدامه كمدخل لإعداد المكتبة باستخدام مجموعة إعداد مكتبة RNA الكلية TruSeq Stranded (Illumina، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية) باستخدام IDT لـ Illumina – TruSeq RNA UD Indexes. تم قياس مكتبات التسلسل بواسطة PCR في الوقت الحقيقي باستخدام مجموعة قياس مكتبة KAPA الكاملة (روش، بازل، سويسرا) وتم تقييمها من حيث توزيع الحجم وغياب المحولات الحرة وثنائيات المحولات على محلل الشظايا. تم تجميع مكتبات التسلسل وقياسها بواسطة PCR في الوقت الحقيقي كما هو موضح أعلاه، وتم تجميعها وتسلسلها على جهاز Illumina NovaSeq 6000 باستخدام مجموعة المواد الكيميائية v1.5 (200 دورة) مع معلمات التشغيل التي تنتج قراءات مزدوجة النهاية بطول 100 نقطة أساسية.
تحليل التسلسل
تمت معالجة ملفات Fastq التي تم الحصول عليها من عملية التسلسل في البداية من خلال خط أنابيب nf-core rnaseq عبر NextFlow باستخدام إعدادات الخط الموصى بها.. تتضمن سلسلة العمليات FASTQC وTrim Galore وSortMeRNA وSTAR وfeatureCounts وStringTie وSalmon وQualimap وdupRadar وPreseq وRSeQC. تم محاذاة القراءات إلى جينوم المرجع GRCh37. تم إجراء التحليل اللاحق باستخدام R 4.3.2 مع Jupyter Notebooks وحزمة Tidyverse (v2.0.0). استخدمنا DESeq2 (v1.42.0) لتحديد الجينات المعبر عنها بشكل مختلف (DEGs) بين العينات غير المحفزة وعينات LNP-1 المحفزة مع 4 عينات في كل مجموعة. تم استخدام إجراء بنجاميني-هوشبرغ لتصحيح الاختبارات المتعددة. بالإضافة إلى ذلك، قمنا بتنفيذ معدل اكتشاف خاطئ قدره 0.01 وحد قطع لتغير اللوغاريتم فوق 1 أو أقل من -1. ثم، قمنا بتنفيذ تقليص تغير اللوغاريتم باستخدام apeglm (v1.24.0) وتم مطابقة معرفات الجينات مع رموز الجينات لإنشاء جدول للجينات المعبر عنها بشكل مختلف (DEGs). تم تمرير جدول DEGs عبر مجموعات Human MSigDB (الإصدار 7.5.1): مجموعات الجينات الرئيسية (H)، KEGG (C2: مجموعات الجينات المنسقة)، وGO (C5: مجموعات الجينات الخاصة بعلم الأحياء – مجموعات الجينات الخاصة بعلم الأحياء الجيني). تم إجراء تحليل إثراء مجموعة الجينات (GSEA) باستخدام حزمة Bioconducter clusterProfiler (الإصدار 4.8.3).
التحليل الإحصائي
تم إجراء التحليلات الإحصائية باستخدام برنامج GraphPad PRISM (الإصدار 10، شركة GraphPad Software Inc.، بوسطن، ماساتشوستس، الولايات المتحدة الأمريكية) باستخدام تحليل التباين الأحادي (ANOVA) متبوعًا باختبار دنّيت للمقارنات المتعددة مع مجموعة تحكم واحدة أو باستخدام اختبار توكي للمقارنات المتعددة بين جميع المجموعات. يتم التعبير عن النتائج كمتوسطات مع خطأ معياري للمتوسط (SEM). القيم لـاعتُبرت ذات دلالة إحصائية. بالنسبة لبيانات النسخ الجيني، تم تنفيذ إجراء بنجاميني-هوشبرغ أثناء تحديد الجينات المعبر عنها بشكل مختلف وتحليل إثراء المسارات لتصحيح المقارنات المتعددة. تم تعديلتم الإبلاغ عن قيم p لهذه النتائج. تم تحديد الأهمية لتحليل المسار الجيني (GSEA) من خلال مقارنة درجة الإثراء بتوزيع فارغ من خلال اختبار التبديل.
توفر البيانات
ستكون البيانات متاحة عند الطلب. بيانات التسلسل متاحة في أرشيف قراءة التسلسل NCBI تحت رقم المرجع PRJNA1196969.
تاريخ الاستلام: 2 مايو 2024؛ تاريخ القبول: 24 مارس 2025؛
تم النشر على الإنترنت: 17 أبريل 2025
References
Verbeke, R., Lentacker, I., De Smedt, S.C. & Dewitte, H. The dawn of mRNA vaccines: The COVID-19 case. J. Controlled Release 333, 511-520 (2021).
O’Shaughnessy, J. A. (ed U.S. Food & Drug Administration) 1-23 (2022).
Patalon, T., et al. Waning effectiveness of the third dose of the BNT162b2 mRNA COVID-19 vaccine. Nat. Commun. 13, 3203 (2022).
Arunachalam, P. S. et al. Durability of immune responses to mRNA booster vaccination against COVID-19. J. Clin. Invest. org/10.1172/JCI167955 (2023).
Ishii, T., et al. Waning cellular immune responses and predictive factors in maintaining cellular immunity against SARS-CoV-2 six months after BNT162b2 mRNA vaccination. Sci. Rep. 13, 9607 (2023).
Baden, L. R. et al. Efficacy and Safety of the mRNA-1273 SARS-CoV2 Vaccine. N. Engl. J. Med. 384, 403-416 (2021).
Corbett, K. S. et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 586, 567-571 (2020).
Puranik, A., et al. Durability analysis of the highly effective mRNA-1273 vaccine against COVID-19. PNAS Nexus 1, pgac058 (2022).
Polack, F.P. et al. Safety and Efficacy of the BNT162b2 mRNA Covid19 Vaccine. N. Engl. J. Med. 383, 2603-2615 (2020).
Coggins, S.A., et al. Adverse Effects and Antibody Titers in Response to the BNT162b2 mRNA COVID-19 Vaccine in a Prospective Study of Healthcare Workers. Open Forum Infect. Dis. 9, ofab575 (2022).
Li, Y. & Chen, X. CpG 1018 Is an Effective Adjuvant for Influenza Nucleoprotein. Vaccines (Basel) 11 https://doi.org/10.3390/ vaccines11030649 (2023).
Pulendran, B., P, S. A. & O’Hagan, D. T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 20, 454-4-75 (2021).
Andries, O. et al. N(1)-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J. Control Release 217, 337-344 (2015).
Freund, I., Eigenbrod, T., Helm, M. & Dalpke, A.H. RNA Modifications Modulate Activation of Innate Toll-Like Receptors. Genes10,92 (2019).
Kariko, K., Buckstein, M., Ni, H. & Weissman, D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity23, 165-175 (2005).
Baiersdorfer, M. et al. A Facile Method for the Removal of dsRNA Contaminant from In Vitro-Transcribed mRNA. Mol. Ther. Nucleic Acids 15, 26-35 (2019).
Connors, J. et al. Lipid nanoparticles (LNP) induce activation and maturation of antigen presenting cells in young and aged individuals. Commun. Biol. 6, 188 (2023).
Alameh, M.G. et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity 54, 2877-2892 e2877 (2021).
Tahtinen, S. et al. IL-1 and IL-1ra are key regulators of the inflammatory response to RNA vaccines. Nat. Immunol. 23, 532-542 (2022).
Sun, D. & Lu, Z.R. Structure and Function of Cationic and Ionizable Lipids for Nucleic Acid Delivery. Pharm. Res. 40, 27-46 (2023).
Litvinova, V.R., Rudometov, A.P., Karpenko, L.I. & Ilyichev, A.A. mRNA Vaccine Platform: mRNA Production and Delivery. Russ. J. Bioorg. Chem. 49, 220-235 (2023).
Zhang, L. et al. Effect of mRNA-LNP components of two globallymarketed COVID-19 vaccines on efficacy and stability. NPJ Vaccines 8, 156 (2023).
Hald Albertsen, C., et al. The role of lipid components in lipid nanoparticles for vaccines and gene therapy. Adv. Drug Deliv. Rev. 188, 114416 (2022).
Pham, C.T. et al. Variable antibody-dependent activation of complement by functionalized phospholipid nanoparticle surfaces. J. Biol. Chem. 286, 123-130 (2011).
Tanaka, T. et al. DiC14-amidine cationic liposomes stimulate myeloid dendritic cells through Toll-like receptor 4. Eur. J. Immunol. 38, 1351-1357 (2008).
Halamoda-Kenzaoui, B. & Bremer-Hoffmann, S. Main trends of immune effects triggered by nanomedicines in preclinical studies. Int J. Nanomed. 13, 5419-5431 (2018).
Lonez, C., Vandenbranden, M. & Ruysschaert, J.M. Cationic lipids activate intracellular signaling pathways. Adv. Drug Deliv. Rev. 64, 1749-1758 (2012).
de Groot, A. M. et al. Immunogenicity Testing of Lipidoids In Vitro and In Silico: Modulating Lipidoid-Mediated TLR4 Activation by Nanoparticle Design. Mol. Ther. Nucleic Acids 11, 159-169 (2018).
Lonez, C. et al. Critical residues involved in Toll -like receptor 4 activation by cationic lipid nanocarriers are not located at the lipopolysaccharide-binding interface. Cell Mol. Life Sci. 72, 3971-3982 (2015).
Forster III, J., Nandi, D. & Kulkarni, A. mRNA-carrying lipid nanoparticles that induce lysosomal rupture activate NLRP3 inflammasome and reduce mRNA transfection efficiency. Biomater. Sci. 10, 5566-5582 (2022).
Kedmi, R., Ben-Arie, N. & Peer, D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials 31, 6867-6875 (2010).
Ju, Y. et al. Anti-PEG Antibodies Boosted in Humans by SARS-CoV-2 Lipid Nanoparticle mRNA Vaccine. ACS Nano 16, 11769-11780 (2022).
Dey, A. K. et al. Tuning the Immunostimulation Properties of Cationic Lipid Nanocarriers for Nucleic Acid Delivery. Front Immunol. 12, 722411 (2021).
Deguine, J & Barton, GM MyD88: a central player in innate immune signaling. F1000Prime Rep. 6, 97 (2014).
Kawasaki, T. & Kawai, T. Toll-like receptor signaling pathways. Fronteirs Immunol. 5, 461 (2014).
Li, C. et al. Mechanisms of innate and adaptive immunity to the PfizerBioNTech BNT162b2 vaccine. Nat. Immunol. 23, 543-555 (2022).
Billack, B. Macrophage activation: role of toll-like receptors, nitric oxide, and nuclear factor kappa B.Am. J. Pharm. Educ. 70, 102 (2006).
Ernst, O., Vayttaden, S.J. & Fraser, I.D.C. Measurement of NF-kappaB Activation in TLR-Activated Macrophages. Methods Mol. Biol. 1714, 67-78 (2018).
Peters, K.L., Smith, H.L., Stark, G.R. & Sen, G.C. IRF-3-dependent, NFkappa B- and JNK-independent activation of the 561 and IFN-beta genes in response to double-stranded RNA. Proc. Natl. Acad. Sci. USA 99, 6322-6327 (2002).
Rennick, J.J., Johnston, A.P.R. & Parton, R.G. Key principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. Nat. Nanotechnol. 16, 266-276 (2021).
Mussbacher, M., Derler, M., Basilio, J. & Schmid, J.A. NF-kappaB in monocytes and macrophages – an inflammatory master regulator in multitalented immune cells. Front Immunol. 14, 1134661 (2023).
Mussbacher, M. et al. Cell Type-Specific Roles of NF-kappaB Linking Inflammation and Thrombosis. Front Immunol. 10, 85 (2019).
Ma, F. et al. Positive feedback regulation of type I interferon by the interferon-stimulated gene STING. EMBO Rep. 16, 202-212 (2015).
Korzun, T. et al. Lipid Nanoparticles Elicit Reactogenicity and Sickness Behavior in Mice Via Toll-Like Receptor 4 and Myeloid Differentiation Protein 88 Axis. ACS Nano https://doi.org/10.1021/ acsnano.4c05088 (2024).
Lee, K.M. & Seong, S.Y. Partial role of TLR4 as a receptor responding to damage-associated molecular pattern. Immunol. Lett. 125, 31-39 (2009).
Chaudhary, N. et al. Amine headgroups in ionizable lipids drive immune responses to lipid nanoparticles by binding to the receptors TLR4 and CD1d. Nat. Biomed. Eng. 8, 1483-1498 (2024).
Tsutsumi-Ishii, Y. & Nagaoka, I. Modulation of human beta-defensin-2 transcription in pulmonary epithelial cells by lipopolysaccharidestimulated mononuclear phagocytes via proinflammatory cytokine production. J. Immunol. 170, 4226-4236 (2003).
Ciesielska, A., Matyjek, M. & Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol. Life Sci. 78, 1233-1261 (2021).
Maeshima, N. & Fernandez, RC Recognition of lipid A variants by the TLR4-MD-2 receptor complex. Front Cell Infect. Microbiol 3, 3 (2013).
Park, B.S. et al. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature 458, 1191-1195 (2009).
Carter, D., et al. A structure-function approach to optimizing TLR4 ligands for human vaccines. Clin. Transl. Immunol. 5, e108 (2016).
Farlik, M. et al. Contribution of a TANK-binding kinase 1-interferon (IFN) regulatory factor 7 pathway to IFN-gamma-induced gene expression. Mol. Cell Biol. 32, 1032-1043 (2012).
Fitzgerald, K.A. et al. IKKepsilon and TBK1 are essential components of the IRF3 signaling pathway. Nat. Immunol. 4, 491-496 (2003).
Uematsu, S. & Akira, S. Toll-like receptors and Type I interferons. J. Biol. Chem. 282, 15319-15323 (2007).
Ng, S.L. et al. IkappaB kinase epsilon (IKK(epsilon)) regulates the balance between type I and type II interferon responses. Proc. Natl. Acad. Sci. USA 108, 21170-21175 (2011).
Fitzgerald, K.A. et al. LPS-TLR4 signaling to IRF-3/7 and NF-kappaB involves the toll adapters TRAM and TRIF. J. Exp. Med 198, 1043-1055 (2003).
Vangasseri, D.P. et al. Immunostimulation of dendritic cells by cationic liposomes. Mol. Membr. Biol. 23, 385-395 (2006).
Ngo, W. et al. Identifying cell receptors for the nanoparticle protein corona using genome screens. Nat. Chem. Biol. 18, 1023-1031 (2022).
Liu, J. et al. A five-amino-acid motif in the undefined region of the TLR8 ectodomain is required for species-specific ligand recognition. Mol. Immunol. 47, 1083-1090 (2010).
Vaure, C. & Liu, Y. A comparative review of toll-like receptor 4 expression and functionality in different animal species. Front Immunol. 5, 316 (2014).
Schroder, K. et al. Conservation and divergence in Toll-like receptor 4-regulated gene expression in primary human versus mouse macrophages. Proc. Natl. Acad. Sci. USA 109, E944-E953 (2012).
Schneberger, D., Caldwell, S., Kanthan, R. & Singh, B. Expression of Tolllike receptor 9 in mouse and human lungs. J. Anat. 222, 495-503 (2013).
Wang, Y., Rahman, D. & Lehner, T. A comparative study of stressmediated immunological functions with the adjuvanticity of alum. . Biol. Chem. 287, 17152-17160 (2012).
Welsby, I. et al. Lysosome-Dependent Activation of Human Dendritic Cells by the Vaccine Adjuvant QS-21. Front Immunol. 7, 663 (2016).
Ho, N. I. et al. Saponin-based adjuvants enhance antigen crosspresentation in human CD11c(+) CD1c(+) CD5(-) CD163(+) conventional type 2 dendritic cells. J. Immunother Cancer 11 https:// doi.org/10.1136/jitc-2023-007082 (2023).
Furman, D. et al. Apoptosis and other immune biomarkers predict influenza vaccine responsiveness. Mol. Syst. Biol. 9, 659 (2013).
Palchetti, S. et al. The protein corona of circulating PEGylated liposomes. Biochim Biophys. Acta 1858, 189-196 (2016).
Wang, Y. et al. Inhibition of clathrin/dynamin-dependent internalization interferes with LPS-mediated TRAM-TRIF-dependent signaling pathway. Cell Immunol. 274, 121-129 (2012).
Vig, S., et al. ER stress promotes mitochondrial DNA mediated type-1 interferon response in beta-cells and interleukin-8 driven neutrophil chemotaxis. Front Endocrinol. (Lausanne) 13, 991632 (2022).
Chen, Y.G. & Hur, S. Cellular origins of dsRNA, their recognition and consequences. Nat. Rev. Mol. Cell Biol. 23, 286-301 (2022).
Dhir, A. et al. Mitochondrial double-stranded RNA triggers antiviral signalling in humans. Nature 560, 238-242 (2018).
Toussi, D.N. & Massari, P. Immune Adjuvant Effect of Molecularlydefined Toll-Like Receptor Ligands. Vaccines (Basel) 2, 323-353 (2014).
Ewels, P.A. et al. The nf-core framework for community-curated bioinformatics pipelines. Nat. Biotechnol. 38, 276-278 (2020).
nf-core/rnaseq v. 3.12.0 (Zenodo, 2023).
Zhu, A., Ibrahim, J.G. & Love, M.I. Heavy-tailed prior distributions for sequence count data: removing the noise and preserving large differences. Bioinformatics 35, 2084-2092 (2019).
الشكر والتقدير
نشكر كاتيرينا لوند وسينتايهو جيبريوهانس في مركز الأدوات البيولوجية بجامعة USU على المساعدة في قياس التدفق؛ دينيس مكداينيل في مركز الأدوات البيولوجية بجامعة USU على المساعدة في المجهر؛ وأندرو فرانك في مبادرة المعلوماتية الحيوية للطلاب على المساعدة في تحليل بيانات النسخ. تم دعم هذا العمل من خلال تمويل من منحة RAMP من قسم طب الأطفال وأموال اتحادية من برنامج الدفاع الصحي (HU000120200067، HU00012120094) وفرع الرعاية الصحية للتطعيم (HU00012120104) من وكالة الدفاع الصحي، وزارة الدفاع الأمريكية. لم يكن للرعاة أي تدخل في تصميم الدراسة، أو جمع البيانات، أو تحليل البيانات، أو تفسير البيانات، أو كتابة التقرير، أو في قرار تقديم المقال للنشر. الآراء والادعاءات المعبر عنها هنا هي آراء المؤلفين ولا ينبغي تفسيرها على أنها تعكس وجهات نظر USUHS، أو القوات الجوية الأمريكية، أو الجيش الأمريكي، أو البحرية الأمريكية، أو الجيش الأمريكي بشكل عام، أو وزارة الدفاع الأمريكية. ينص العنوان 17 U.S.C. 105 على أن ‘حماية حقوق الطبع والنشر بموجب هذا العنوان غير متاحة لأي عمل من أعمال حكومة الولايات المتحدة.’ يحدد العنوان 17 U.S.C. 101 “عمل حكومة الولايات المتحدة” على أنه عمل أعده عضو في الخدمة العسكرية أو موظف من حكومة الولايات المتحدة كجزء من واجبات ذلك الشخص الرسمية.
مساهمات المؤلفين
A.Z. و A.M.W.M. تصوروا وصمموا الدراسة، وحللوا، وفسروا البيانات. M.G.A. قدمت LNPs وساهمت في تصميم الدراسة. H.S. قامت بتخليق LNPs ولقاحات mRNA. E.M. ساهمت في تصميم الدراسة. A.Z. أجرت التجارب. G.S. و C.D. قاموا بإجراء تسلسل RNA
. Z.L. ساعدت في تحليل البيانات. A.Z. و A.M.W.M. تصوروا وكتبوا المخطوطة. Z.L. و M.G.A. و E.M. ساعدوا في كتابة ومراجعة المخطوطة.
المصالح المتنافسة
يعلن المؤلفون أن البحث تم إجراؤه في غياب أي علاقات تجارية أو مالية يمكن أن تُفسر على أنها صراع محتمل للمصالح. الآراء والادعاءات المعبر عنها هنا هي آراء المؤلفين ولا تعكس السياسة الرسمية أو موقف جامعة الخدمات الموحدة للعلوم الصحية أو وزارة الدفاع، أو مؤسسة هنري م. جاكسون لتقدم الطب العسكري، Inc. (HJF). الإشارة إلى أسماء العلامات التجارية أو المنتجات التجارية أو المنظمات لا تعني تأييدًا من حكومة الولايات المتحدة.
ملاحظة الناشر تظل Springer Nature محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
الوصول المفتوح هذه المقالة مرخصة بموجب رخصة المشاع الإبداعي للاستخدام والمشاركة والتكيف والتوزيع وإعادة الإنتاج في أي وسيلة أو تنسيق، طالما أنك تعطي الائتمان المناسب للمؤلفين الأصليين والمصدر، وتوفر رابطًا لرخصة المشاع الإبداعي، وتوضح ما إذا كانت هناك تغييرات قد أُجريت. الصور أو المواد الأخرى من طرف ثالث في هذه المقالة مشمولة في رخصة المشاع الإبداعي للمقالة، ما لم يُشار إلى خلاف ذلك في سطر الائتمان للمادة. إذا لم تكن المادة مشمولة في رخصة المشاع الإبداعي للمقالة واستخدامك المقصود غير مسموح به بموجب اللوائح القانونية أو يتجاوز الاستخدام المسموح به، ستحتاج إلى الحصول على إذن مباشرة من صاحب حقوق الطبع والنشر. لعرض نسخة من هذه الرخصة، قم بزيارةhttp://creativecommons.org/licenses/by/4.0/.
هذا عمل حكومي أمريكي وليس تحت حماية حقوق الطبع والنشر في الولايات المتحدة؛ قد تنطبق حماية حقوق الطبع والنشر الأجنبية 2025
1قسم طب الأطفال، جامعة الخدمات الموحدة للعلوم الصحية، بيثيسدا، MD، الولايات المتحدة.مؤسسة هنري م جاكسون لتقدم الطب العسكري، بيثيسدا، MD، الولايات المتحدة.قسم التشريح وعلم وظائف الأعضاء وعلم الوراثة، جامعة الخدمات الموحدة للعلوم الصحية، بيثيسدا، MD، الولايات المتحدة.قسم علم الأمراض وطب المختبرات، مستشفى الأطفال في فيلادلفيا، فيلادلفيا، PA، الولايات المتحدة.قسم علم الأمراض وطب المختبرات، جامعة بنسلفانيا، فيلادلفيا، PA، الولايات المتحدة.معهد بن للابتكار في RNA، جامعة بنسلفانيا، فيلادلفيا، PA، الولايات المتحدة.قسم علم الأحياء الدقيقة وعلم المناعة، جامعة الخدمات الموحدة للعلوم الصحية، بيثيسدا، PA، الولايات المتحدة.
utmb
الصحة
⟶ البريد الإلكتروني: allison.malloy@usuhs.edu
معهد سيالي لعلوم
اللقاحات
lonizable lipid nanoparticles of mRNA vaccines elicit NF- KB and IRF responses through toll-like receptor 4
Amanda E. Zelkoski , Zhongyan Lu , Gauthaman Sukumar , Clifton Dalgard , Hooda Said , Mohamad-Gabriel Alameh , Edward Mitre & Allison M. W. Malloy
Ionizable lipid nanoparticles (LNP) that have enabled the success of messenger RNA (mRNA) vaccines have been shown to be immunostimulatory in the absence of mRNA. However, the mechanisms through which they activate innate immune cells is incompletely understood. Using a monocyte cell line, we compared the ability of three LNP formulations to activate transcription factors Nuclear Factor-kappa B (NF-kB) and Interferon Regulatory Factor (IRF). Comparison of signaling in knockout cell lines illustrated a role for Toll-like receptor (TLR) 4 in initiation of this signaling cascade and the contribution of the ionizable lipid component. Activation induced by empty LNPs was similar to that induced by LNPs containing mRNA, indicating that LNPs may provide the majority of innate stimulation for the mRNA vaccine platform. Our findings demonstrate that ionizable lipids within LNPs signal through TLR4 to activate NF-кB and IRF, identifying a mechanism for innate activation that can be optimized for adjuvant design.
Nanoparticles have been developed for use in the delivery of therapeutics and vaccine antigen. Distinct lipid nanoparticle (LNP) formulations were used for the BNT162b2 and mRNA-1273 vaccines to deliver a N1-methylpseudouridine-modified mRNA encoding SARS-CoV-2 spike . Despite demonstrated immunogenicity, durability is limited . Furthermore, the cause of the observed side effect profile is unknown . These findings present an opportunity for enhancing both tolerability and efficacy. However, achieving this improvement requires a deeper understanding of the functional mechanisms of mRNA-LNP vaccines. How these vaccines initiate immune responses is incompletely understood. Vaccines typically contain an adjuvant that stimulates the immune response, and some elicit this response through Toll-like receptor (TLR) signaling . The mRNA component may signal through TLR7 and 8, which recognize singlestranded RNA (ssRNA). Additionally, double-stranded RNA (dsRNA), a byproduct of the mRNA production process, may elicit signaling through TLR3, RIG-I, or MDA-5. However, the mRNA incorporated into these vaccines have undergone modifications aimed at diminishing ssRNA recognition and been purified to reduce dsRNA presence . Interestingly, both in vitro and in vivo studies have demonstrated that the empty
LNP can activate the innate immune system . However, the mechanisms behind LNP-mediated immunostimulation are poorly understood.
LNPs vary in composition, but are primarily designed with a charged or ionizable lipid and helper lipids . The LNP of the BNT162b2 mRNA vaccine is comprised of the ALC-0315 ionizable cationic lipid, cholesterol, DSPC (1,2-distearoyl-sn-glycero-3-phosphocholine), and ALC-0159, a PEGylated lipid . In comparison, the LNP of mRNA-1273 uses SM102 as the ionizable lipid and PEG-DMG as the PEGylated lipid . The ionizable lipids are neutral at time of administration but become positively charged in the acidic endosome, which facilitates LNP fusion and mRNA release into the cytoplasm . Cholesterol and PEGylated helper lipids prevent aggregation or enhance transfection efficiency . The components also impact size, with the LNPs of the BNT162b2 and mRNA-1273 vaccines being and , respectively . A review of LNP formulations found that elicited an immune response, with complement and innate activation predominating . Some cationic LNPs can be detected by TLR4 and the NLRP3 inflammasome . Additionally, antibodies generated to the PEGylated lipid can activate the complement pathway . It is unclear whether empty ionizable LNPs would activate the immune system
through similar pathways as cationic formulations since the vaccines incorporate distinct ionizable cationic lipids that may differentially influence uptake and degradation .
Recently, LNP, in the absence of mRNA, was demonstrated to be immunostimulatory in a murine model . Removal of the ionizable lipid in this LNP resulted in a reduction in cytokine production, suggesting the ionizable lipid plays an important role in adjuvanticity . Similarly, stimulation of human monocyte-derived dendritic cells with ionizable LNPs elicits production of cytokines such as IL-1, IL-6, IL-12, IFNα, and IFN . To date, however, the mechanisms by which empty LNPs activate the innate immune system are not well understood.
Alameh et al. observed that the adjuvanticity of an empty ionizable LNP in a murine model was partially reliant on MyD88, suggesting a role for TLRs or IL-1 receptors . TLRs can activate nuclear factor and interferon regulatory factor (IRF) transcription factors through MyD88 and TRIF adaptor molecules . These transcription factors are essential to the innate response as they regulate the production of cytokines, chemokines, and type I interferons (IFNs) . However, others have found no evidence for TLR involvement in the innate immune response to LNPs in mouse models .
In this study, we sought to investigate the ability of empty ionizable LNPs to activate human innate immune cells. Therefore, we compared innate immune activation induced by three LNPs using a human monocyte cell line. We determined that ionizable LNPs differentially activate NF- and IRF. Our findings show signaling is predominantly mediated through TLR4. Interestingly, NF- and IRF responses to the empty BNT162b2 LNP were similar in magnitude to mRNA-LNP, indicating that the ionizable LNP may be primarily responsible for activating the innate immune response during mRNA vaccination.
Results
The ionizable LNPs elicits NF-кB and IRF responses from THP1 cells
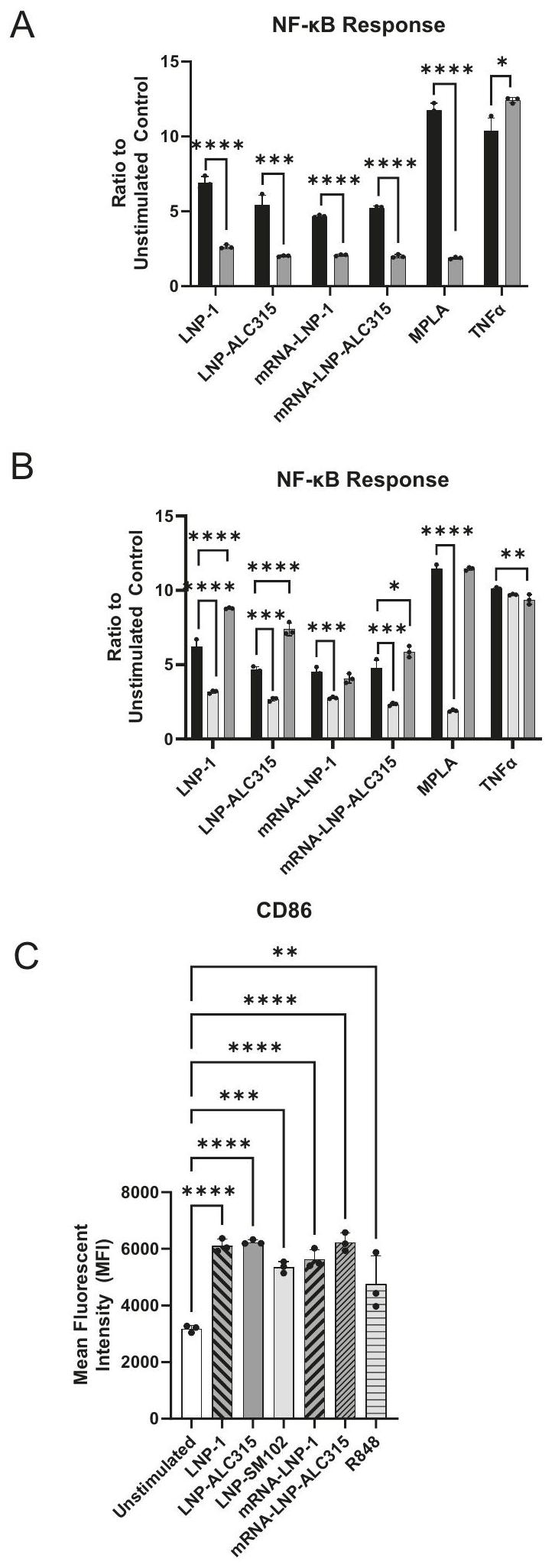
Monocytes are key innate immune cells that rapidly respond to pathogen or damage-associated stimuli. Activation and function of monocytes is associated with NF- activity and IRF pathway activation . To determine if empty ionizable LNPs could activate monocytes, we utilized a THP-1 monocyte-like cell line with an alkaline phosphatase reporter for NF- activation and a luciferase reporter for IRF activation. We compared activation induced by different ionizable LNPs to R848 (TLR7/8 agonist), and MPLA (TLR4 agonist). The empty LNPs used were LNP-1, LNP-ALC315 (BNT162b2 formulation), and LNP-SM102 (mRNA-1273 formulation) (Supplementary Table 1). LNP-SM102 differs in ionizable lipid structure from LNP-ALC315 as well as in PEGylated lipid (Supplementary Fig. 1) (Supplementary Table 1).
Stimulation of THP-1s resulted in NF- reporter detection at 24 h , with increased NF- activation at 48 h that minimally increased or plateaued through 120 h (Fig. 1A). For LNP-ALC315, the NF- response was 2 -fold above the unstimulated control at 24 h and then 4 -fold past 48 h . The response to LNP-1 was 6-7-fold at 48 and 72 h . The response to LNP-1 was 6-7-fold at 48 and 72 h . NF- activation induced by LNP-SM102 was similar in magnitude to that of LNP-ALC315 (Fig. 1). The kinetics of reporter production to all LNPs increased over 24 h with activation peaking at 48 and 72 h (Supplementary Fig. 3A). In comparison, all TLR agonists exhibited strong responses within 24 h . We also observed that NF- signal transduction was dose dependent and plateaued at the highest dose regardless of the LNP (Supplementary Fig. 2A). All conditions exhibited similar changes in viability over time cultured, and R848 and MPLA exhibited the highest percentage of cell death (Supplementary Fig. 3C). As decrease of viability is seen in all conditions, there is the potential for cell death products to contribute to activation seen, particularly as cell death begins around the initiation of NF- detection. At induces the greatest levels of cell death corresponding with the heightened NF- activation compared to the other LNP formulations, while LNP-SM102 stimulation induces similar levels of death with minimal NF- activation at this time point.
To investigate the cause of the innate activation, we utilized a LNP that lacks the ionizable lipid component to determine the contribution to stimulation. This LNP is comprised of cholesterol, DSPC, and ALC-0159, the same composition as LNP-ALC315 except devoid of ionizable lipid (Supplementary Table 1). The response was lost in the absence of the ionizable lipid component (Fig. 1C) indicating the primary contribution of the ionizable lipid to the activation.
We also investigated the activation of IRF pathways in response to the ionizable LNPs. We measured a 3 and 1.7 -fold increase at 48 and 72 h , respectively, in the IRF reporter in response to LNP-ALC315 in comparison to the control (Fig. 2A, B). Unlike with NF- , LNP-1 had a similar IRF response to LNP-ALC315 while LNP-SM102 elicited a significantly greater response (Fig. 2A, B). The TLR agonist R848 exhibited the highest increase in IRF response, which was significantly higher than the empty LNPs and TLR agonist MPLA (Supplementary Fig. 3B). We also observed a dose dependent IRF response to each LNP that plateaued at (Supplementary Fig. 2B). Additionally, we assessed the ionizable lipid’s contribution and showed that an LNP lacking ionizable lipid was unable to elicit IRF activation (Fig. 2C). This indicates that the IRF response, like NF- , is primarily driven by the ionizable lipid component.
Overall, our data show empty ionizable LNPs predominantly activate and, to a lesser extent, IRF transcription factors. The magnitude of these responses differs between the ionizable LNPs, but, importantly, was lost in the absence of the ionizable component.
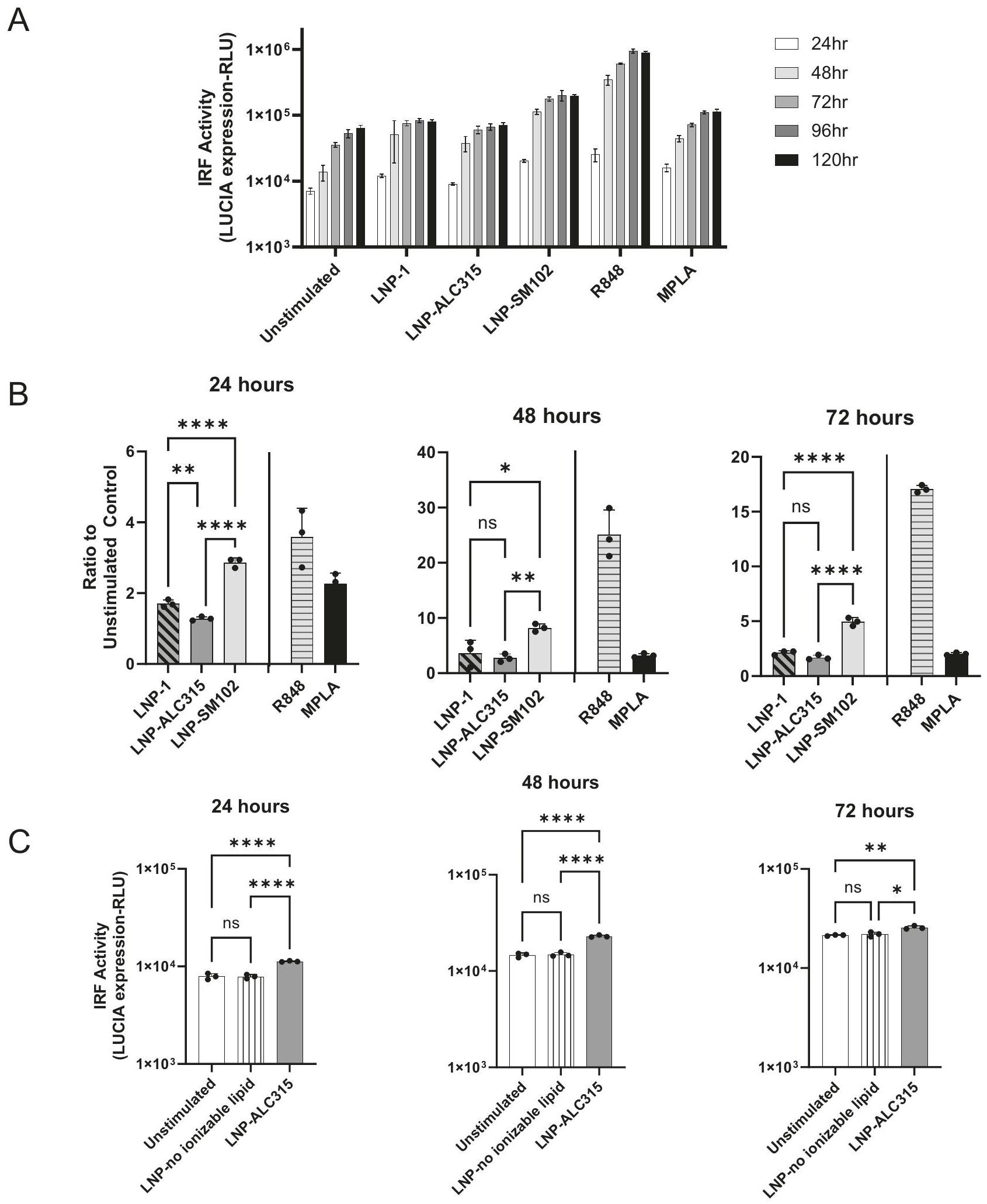
LNP endocytosis is not associated with transcription factor activation
Signaling cascades, once initiated, typically result in NF- and IRF phosphorylation in a short time period, even as quickly as . The activation of the NF- and IRF pathways by ionizable LNPs peaked at after stimulation, which was delayed compared to activation by the TLR agonists. To address the slower response kinetics, we first confirmed it was consistent at different doses (Supplementary Fig. 2). We then hypothesized that endocytosis may be required for LNPs to initiate activation. Using a fluorescent DiO-tagged LNP, we investigated the kinetics of uptake by the THP-1 cells. Uptake was visualized within 2 h , peaking in intensity at 48 h (Supplementary Fig. 4A, B). With lower doses, uptake declines more rapidly at 72 h . The decline is likely due to consumption of DiO-LNP in the media as cells replicate, leaving newly replicated cells unable to uptake DiO-LNP (Supplementary Fig. 4A, B). These findings demonstrate a continuous uptake of LNP by THP-1 cells over time.
As this data is suggestive of a threshold of uptake, we utilized endocytosis inhibitors to determine if this process was indeed critical for cellular activation. We treated THP-1 cells with dynasore, cytochalasin D, chloroquine, or methyl- cyclodextrin 2 h prior to stimulation and every 24 h subsequently in order to inhibit different pathways of endocytosis . The clathrin-mediated endocytosis (CME) inhibitor chloroquine decreased the IRF responses (Supplementary Fig. 4C). Dynasore also inhibits CME but did not impact LNP-mediated activation (Supplementary Fig. 4C). Thus, the decrease in IRF response seen with chloroquine is potentially the result of an off-target effect. Cell death was not a factor as viability for each condition was within of the untreated control throughout. The other inhibitors did not impact or heightened response. Minimal reduction in uptake of DiOtagged LNP was observed at 24 h and no inhibition was measured at 48 h (Supplementary Fig. 5). The inhibitors reduced and IRF responses to MPLA and R848 controls at early time points (Supplementary Fig. 4D). Overall, we observed no significant impact of endocytosis inhibitors on LNP-mediated NF- and IRF activation. However, given the limited inhibition visualized, we cannot definitively conclude that activation does not require endosomal entry.
Stimulation with ionizable LNPs elicit upregulation of toll-like receptor signaling
Our data demonstrate that a monocyte cell line can be activated by empty ionizable LNP to induce NF- and IRF pathways, which induce cytokines
Fig. 1 | NF- signaling transduction in THP-1 reporter cell line. THP-1-Dual reporter cells were treated with LNP-1 , LNP-ALC315 , LNPSM102 , R848 , MPLA , LNP without ionizable lipid , or media alone. An alkaline phosphatase reporter was used to measure the NF- response. Reporter accumulation was measured in the supernatant over . Fold change reporter production was compared between LNPs . Production of reporter in response to LNP without ionizable lipid was compared to the compositionally similar LNP-ALC315 ( ) (C). Data are represented as mean ± SEM. Significance was assessed using a one-way ANOVA with Dunnett’s test for multiple comparisons. , , , .
Fig. 2 | IRF signaling transduction in THP-1 reporter cell line. THP-1-Dual reporter cells were treated with LNP-1 , LNP-ALC315 , LNPSM102 , R848 , MPLA , LNP without ionizable lipid , or media alone. A luciferase reporter was used to measure the IRF response. Reporter accumulation was measured in the supernatant over 120 h
( ) (A). Fold change reporter production was compared between LNPs ( ) (B). Production of reporter in response to LNP without ionizable lipid was compared to the compositionally similar LNP-ALC315 ( ) (C). Data are represented as mean ± SEM. Significance was assessed using a one-way ANOVA with Dunnett’s test for multiple comparisons. .
Fig. 3 | THP-1 transcriptional response to LNP-1. THP-1-Dual reporter cells were treated with LNP-1 or media alone prior to RNA isolation and sequencing. Samples were sequenced in quadruplicate. Heatmap of 10,585 differentially expressed genes upon comparison of unstimulated to LNP-1-stimulated THP-1 cells out of a total of 57,773 genes detected (A). Table of NF- subunit and IRF genes with their corresponding log2-fold change and adjusted values (B). Gene set enrichment analysis (GSEA) for KEGG gene set hsa04620Toll-like receptor
signaling pathway (C). Table of selected genes from hsa04620Toll-like receptor signaling pathway (D). THP-1 cells were stimulated with LNP-1 or media alone, and surface marker expression was measured after 24 or 48 hours by flow cytometry ( ). Data are shown as the ratio of mean fluorescence intensity (MFI) of the stimulated condition to the unstimulated negative control and represented as mean ± SEM (E). Adjusted p values were calculated by the BenjaminiHochberg procedure.
and inflammatory profiles . To define signaling cascades involved in the LNP-mediated activation, we analyzed the transcriptional response of THP1 cells upon LNP stimulation. We chose LNP-1 as the model as it elicited the strongest activation of the tested LNPs. Comparing LNP-1 stimulated to unstimulated cells at 24 h , we identified 10,585 differential expressed genes (DEGs) out of 57,773 measured. We used a stringent cutoff criterion of a 0.01 false discovery rate and log 2-fold change (LFC) cutoff above 1 and below -1 to narrow down pathway analysis (Fig. 3A). Five genes encoding NF- subunits were measured and among those NFKB1, NFKB2 and RELB were significantly upregulated. Two met the DEG criteria, with NFKB1 just under the threshold. Evaluation of interferon regulatory factors demonstrated that IRF3, IRF4, IRF7, and IRF9 were upregulated. NF-кB activation initiates transcription of additional NF- subunits while IRF initiates a positive feedback loop through IFN signaling . Thus, heightened expression of these transcripts is consistent with the reporter assay findings. Of the upregulated IRFs, only IRF4 and IRF7 met our criteria for DEG (Fig. 3B). IRF3 exhibited a small, but significant upregulation with a LFC of 0.19 . The luciferase reporter gene in our THP-1 model is under the control of an ISG54 minimal promoter in conjunction with five IFNstimulated response elements. Thus, all upregulated IRFs may contribute to the reporter response seen (Fig. 2A-C).
Using the list of DEGs, we identified gene sets and pathways in the gene ontology (GO), hallmark, and KEGG databases to understand the mechanisms that might be linked to ionizable LNP activation of immune responses. From the GO database, we identified 791 gene sets
with 70 containing more than 50 DEGs (Supplementary Fig. 6A). We also identified thirteen and five differentially regulated pathways with the hallmark and KEGG databases, respectively (Supplementary Fig. 6B, C). Of the pathways identified, many were associated with immune activation, including cell death, interferon production, NF-кB-regulated cytokines, and TLR signaling (Supplementary Fig. 6A-C). The upregulation of interferon and NF-кB-regulated cytokine signaling pathways is consistent with our findings using the THP-1 reporter system.
We next utilized gene set enrichment analysis (GSEA), which considers all genes rather than only the DEGs (Fig. 3C). This analysis identified upregulation of the TLR Signaling pathway, indicating a mechanism of activation. Within this pathway, key genes involved in antigen presenting cell activation and T cell engagement were upregulated (Fig. 3D). We confirmed these findings at the protein level by measuring surface expression of CD14, CD40, and CD86 on THP-1 cells after stimulation with LNP-1 (Fig. 3D, E). As seen by flow cytometry, LNP-1 induced approximately 2-fold increases in CD14 and CD86 at peak expression levels and a 1.3 -fold increase in CD40 (Fig. 3E). This data corresponds to the higher LFCs of CD14 and CD86, 4.07 and 2.96 respectively, with CD40 exhibiting an LFC of 1.26. In addition to NF- and IRF, other transcription factors are activated in response to LNP-1. JUN and FOS which encode AP-1 are both upregulated, suggestive of the crosstalk between signaling pathways (Supplementary Fig. 6A-C, Fig. 3D).
Fig. 4 | LNP-mediated NF-кB and IRF signaling transduction dependence on TLR4 and adaptor molecules. THP-1-Dual reporter cell lines were incubated for 72 h with LNP-1 , LNP-ALC315 , LNP-SM102 , MPLA , TNFa( , R848 , IFNα( or media alone. NF- and IRF response were compared between wild-type (WT) and TLR4 knockout cell lines (A) and between the WT and MyD88 or TRIF knockout cell lines
(B). THP-1 WT cells were incubated with stimuli in media containing serum (FBS) or without serum (C). The NF- kB and IRF responses were assessed and reported as a ratio of the reporter production of the stimulated condition to the media alone control ( ). Data are represented as mean ± SEM. Significance was assessed by -test (A) or one-way ANOVA with Dunnett’s multiple comparisons test (A,B) or student’s -test .
TLR4 initiates signaling through MyD88 in response to ionizable LNPs
Based on the upregulation of the TLR Signaling pathway in our transcriptomics data set and prior findings on cationic and one ionizable LNP, we hypothesized that TLR4 plays a key role in the ionizable LNP activation of monocytes . We used the THP-1 reporter with the TLR4 gene knocked out to determine if THP-1 upregulation of NF- and IRF were driven by this signaling pathway (Supplementary Fig. 7). Upon comparing the THP-1 with the TLR4-KO, we measured a marked reduction in NF-кB and IRF responses to all ionizable LNPs (Fig. 4A) (Supplementary Fig. 8A). The loss of TLR4, however, did not completely abrogate the NF- or IRF activity (Fig. 4A) (Supplementary Fig. 8A). These findings suggest that TLR4 may be the principal receptor responsible for ionizable LNP-mediated activation of THP-1 cells, but other pathways may contribute. To ensure the
accuracy of our data, we tested our LNP for presence of endotoxin, which can also trigger TLR4, and determined it was below the assay detection limit.
Additionally, TLR4 activation can be triggered by cell death . There is a potential for LNP-mediated products of cell death to initiate TLR4 signaling detected in this study. However, a recent study demonstrated structural features of ionizable lipids of different LNP formulations engage TLR4 corroborating our findings with TLR4-KO and removal of the ionizable lipid. Together these finding support TLR4 signaling occurring separate from cell death .
To further evaluate TLR4 involvement in sensing ionizable LNPs, we investigated adaptor proteins MyD88 and TRIF, which participate in the signaling cascade (Supplementary Fig. 7). Analysis of the transcriptomic data showed upregulation of MyD88 and TICAM1 genes, the latter encoding TRIF, in response to LNP-1 (Fig. 3D). Both NF- and IRF
activation by all ionizable LNPs was reduced in MyD88 deficient THP-1 cells after 72 h (Fig. 4B) (Supplementary Fig. 8B). In contrast, TRIF deficient THP-1 cells showed a slight increase in NF- activation by LNP-1. LNPALC315 and LNP-SM102, however, elicited a reduced response in the TRIF-KO, albeit not to the same degree as with the MyD88-KO (Fig. 4B) (Supplementary Fig. 8B). Similar results were seen with the IRF response in the TRIF-KO. These results demonstrate that although all three LNPs are recognized by TLR4, they have varying reliance on the adaptor proteins though signaling was predominantly through the TLR4/MyD88 pathway.
To further investigate alternative pathways and evaluate the role of TLR4 in sensing the ionizable LNPs, we evaluated NF- and IRF activation using the A549 cell line with the same reporter constructs. The A549 cell line is derived from alveolar basal epithelial cells that do not express TLR4 . No NF- or IRF response to any LNP was observed (Supplementary Fig. 9) indicating the key role of TLR4 in sensing LNPs and failed to identify alternative pathways.
Next, we hypothesized that serum, facilitates LNP-TLR4 interaction and uptake, as has been shown with LPS . To test if serum is required for LNP-mediated responses, we used serum-free conditions and demonstrated a significant decrease in both NF- and IRF activation (Fig. 4C, Supplementary Fig. 8C). Serum-free cultures exhibited reduced proliferation and unstimulated serum-free samples were used for normalization. These findings demonstrate ionizable LNPs rely on the presence of serum for THP-1 activation. To determine if TLR4 interaction also occurred in the endosome, we investigated colocalization of DiO-tagged LNP with TLR4 in early endosomes, as marked by Rab5a, which was not observed (Supplementary Fig. 8D). This data further suggests TLR4-LNP interaction occurs at the cell surface (Supplementary Fig. 7).
Addition of mRNA does not enhance NF-кB or IRF responses induced by LNP
We next sought to determine the role of mRNA in innate activation induced by the BNT162b2 vaccine. Although modifications were made to the BNT162b2 mRNA component to reduce its immunostimulatory capacity , we hypothesized that BNT162b2 mRNA vaccine would have heightened NF- and IRF activation compared to LNP alone. Surprisingly, our data showed that the response to mRNA-LNP was comparable to or lower than the corresponding empty LNP upon treatment of THP-1 reporter cells (Fig. 5A, B). R848 was previously shown to elicit a robust NF and IRF activation (Fig. 1A, Fig. 2A), demonstrating a functional TLR7/8 response. Thus, the mRNA component did not augment the NF- and IRF signaling and did not stimulate additional pathways in the absence of TLR4, MyD88, or TRIF (Fig. 5A, B). Furthermore, CD86 expression on THP-1s was similar between mRNA-LNPs and corresponding empty LNP treated cells (Fig. 5C). Additionally, mRNA-LNP and empty LNP did not elicit NF or IRF responses in A549s (Supplementary Fig. 9). These data suggest that ionizable LNPs are fully competent to activate a monocyte model of innate immunity in the absence of mRNA and that the addition of mRNA did not increase the response.
Discussion
The advent of licensed mRNA vaccines marks a paradigm shift in immunization strategies, providing a platform which elicits robust adaptive immune responses and can be produced rapidly . Interestingly, these vaccines lack a defined adjuvant, indicating intrinsic adjuvanticity. Though the precise mechanism remains incompletely understood, both the RNA component and LNP are implicated in induction of innate immunity . Understanding the mechanism of mRNA vaccine adjuvanticity may provide insights for improving immunogenicity and/or reactogenicity of future vaccines. Therefore, the goal of this study was to better define the innate immune pathways activated by the empty ionizable LNPs, including those used in the BNT162b2 and mRNA-1273 mRNA vaccines.
Using a THP-1 monocyte cell line with reporter systems, we found that the three ionizable LNPs induce NF- and IRF activation to varying degrees. Activation was largely dependent on TLR4 through the MyD88
adaptor, with minimal TRIF dependence. Transcriptional profiling of LNP1 stimulated THP-1 cells supported these findings, showing upregulation of TLR signaling. No NF- or IRF response occurred in the absence of the ionizable lipid component, indicating it is the driver of innate immune activation and suggesting structural differences in the ionizable lipids influenced signaling magnitude. Additionally, our data indicated signaling was primarily TLR4-MyD88 dependent, but additional pathways are likely induced by ionizable LNPs. Furthermore, activation required serum, but not endocytosis, supporting the role of surface expressed TLR4.
Notably, we observed that mRNA-containing LNPs did not increase IRF or NF- signaling in THP- 1 cells compared to their empty LNPs, suggesting the LNP is the primary adjuvant in the BNT162b2 mRNA vaccine. While other pathways may be engaged, the predominant pathway utilized is via TLR4/MyD88 at the cell surface.
Responses varied across LNPs. LNP-SM102 (mRNA-1273 formulation) elicited the greatest IRF response, LNP-1 the strongest NF- response, and LNP-ALC315 (BNT162b2 formulation) showed similar NF activation to LNP-SM102 and IRF activation similar to LNP-1. No response was observed without the ionizable lipid component, confirming its role as the driver of NF- and IRF activation. Although we did not test this with LNP-SM102, the ionizable lipid is likely the driver but we cannot rule out a role for its pegylated lipid. We posit that structural differences may influence strength of NF- and IRF activation, as seen with LPS variations across bacterial species in TLR4 signaling . Thus, ionizable lipid design is a potential manner in which LNPs can be modified to enhance adjuvanticity or diminish reactogenicity.
Our findings on LNP-mediated innate immune activation are supported by Connors et al, who have demonstrated that dendritic cell and monocyte populations from PBMCs stimulated with LNP-1 exhibit TBK1 and IRF7 activation between 6 and 24 hours . TBK1 activates both IRF3 and IRF7, and as our IRF reporter is produced upon activation of any IRF, IRF7 could be the source of reporter production seen in the THP- . While, IRF3 is activated downstream of TLRs, such as TLR4 , our data did not show strong upregulation of IRF3 nor TRIF-dependence for LNP-1. However, IRF4 and IRF7 were both upregulated, suggesting they contribute to the response to LNP-1. As LNP-ALC315 and LNP-SM102 exhibited some TRIF dependence, IRF3 may play a role for these LNPs. Together, our data demonstrate that empty ionizable LNPs can elicit both NF- and IRF responses, albeit the definitive IRF activation requires further investigation.
Using THP-1 knockout lines, we found that cellular activation was predominantly dependent on TLR4-MyD88 signaling. Supporting our findings, an LNP with the MC3 ionizable lipid elicits reactogenicity in mice through the TLR4-MyD88 pathway, independent of TRIF . Certain cationic LNPs can be detected by TLR4, and thus are able to stimulate innate immune responses . Charge and structure of the lipid components appear to contribute to their stimulatory capabilities. Variations in head groups of cationic lipids have been demonstrated to modulate TLR4 signaling . Moreover, a recent study highlighted the significance of amine head group structures in TLR4 interactions with ionizable lipids distinct from those investigated in this study . Additionally, structural changes to hydrocarbon chain length as well as head group modifications could enhance or abrogate costimulatory molecule upregulation on murine dendritic cells . Thus, structural variations can alter strength of signaling. However, despite structural differences in lipids, TLR4-MyD88 appears to be a common pathway for innate immune activation. Furthermore, no NF- and IRF responses were detected in A549s, which lack TLR4, further supporting TLR4 involvement in activation by ionizable LNPs.
Our data further demonstrates that serum proteins, are likely involved as serum was required for LNP-mediated activation for LNPs tested. These proteins may contribute to the formation of the protein corona surrounding the LNP, facilitating receptor-mediated uptake, or they may include the LPS-binding protein (LBP), which is essential for LPS engagement with TLR4 . However, further investigation is needed to clarify the specific mechanisms involved. The protein corona is particularly significant because its composition depends on the local environment of the .
Fig. 5 | Response to LNPs compared to mRNA-LNP in THP-1. THP-1-Dual reporter cells were incubated with LNP-1 , LNP-ALC315 , LNPSM102 , mRNA-LNP-1 , mRNA-LNPALC315 , MPLA , TNFa , IFNα , R848 , or media alone for 72 h . NF- and IRF response were compared between wild-type (WT) and TLR4 knockout cell lines (A) and between the WT and MyD88 or TRIF knockout cell lines (B). NF-kB and IRF responses were assessed and reported as a
Consequently, intramuscular delivery of LNPs could result in a protein corona distinct from that of intravenously delivered LNPs. Therefore, understanding the role of the protein corona in LNP-mediated innate immune activation may require evaluation in specific biological contexts.
As neither the NF- nor IRF response was completely abrogated with absence of TLR4 or its adaptors, we suspect additional pathways contribute to detection of ionizable LNPs. Notably, a TLR4 knockout mouse model utilized in the study by Li et al. did not exhibit loss of vaccine responsiveness . Others have shown a different ionizable LNP can elicit reactogenicity in mice through TLR4, suggesting that variations in activation strength and modeling may influence results . Since our data indicates that LNP activation is not entirely dependent on TLR4 signaling, other pathways may provide sufficient stimulation in the absence of TLR4. Additionally, human and murine TLR signaling and expression are not fully consistent and may result in conflicting findings .
Transcriptional profiling of cellular responses to LNP-1 showed upregulation of several pathways related to inflammation, cell death, and stress, which could provide insight into alternative methods of ionizable LNP recognition. Other adjuvants have been demonstrated to utilize those mechanisms for immune activation . For instance, alum and saponinbased adjuvants induce cell stress . Furthermore, vaccines that induce cell death have enhanced effectiveness . As cationic LNPs have been observed to initiate inflammatory cell death, this is a plausible method for ionizable LNP immune activation and requires further investigation .
The mechanisms by which ionizable LNPs activate THP-1 cells may also be linked to the delay in kinetics observed. During our investigation of the kinetics of the THP-1 response to the different LNPs, the peak of signaling occurred at 48 h . Although Connors et al. found that signaling of LNP-1 was highest at 24 h , they did not investigate later timepoints . As MPLA (TLR4 agonist) and R848 (TLR7/8 agonist) elicited strong NF-кB responses within 24 h , we expected to see a more pronounced response to the LNPs at this time point. One explanation for the late onset activation observed may be that endocytosis is required. Since the cells are grown in media containing serum, a protein corona will form around the LNP . This structure would sterically impede TLR4 detection until some level of breakdown occurs during endocytosis. Thus, we considered that the kinetics may be reflective of this requirement. Using a DiO-tagged LNP, we showed with flow cytometry and immunofluorescence microscopy that THP-1 cells take up LNP within 2 h and continuously over the 72 h measured. As this was suggestive of a threshold of uptake for LNP-mediated activation, we used different chemical inhibitors to block uptake. Only chloroquine elicited reduction in IRF response. Although the inhibitors impaired response to controls at early time points, they only minutely reduced LNP uptake for the first 24 -hour period. Thus, it remains possible that signaling from the endosome occurs. However, TLR4 primarily utilizes MyD88 from the cell surface and switches to the TRIF-dependence after endocytosis . Since we predominantly observed MyD88-dependence in the LNPs tested, this supports cell surface activation of TLR4. Additional investigation is needed to determine the mechanisms underlying the kinetics of ionizable LNPmediated activation.
We took our study further to investigate the contribution of the mRNA component compared to the ionizable LNP alone. Although modifications have been made to the RNA to render it less immunostimulatory, there is still potential for detection of this RNA in some capacity by TLR7/ . Additionally, any dsRNA remaining after mRNA purification may be detected by TLR3 or cytosolic PRRs such as MDA5. However, our data show that in the THP-1s, the NF-кB and IRF responses to mRNA-LNPs tested were similar to the responses to their corresponding empty LNP. Thus, the RNA component provided no immunostimulatory benefit in this model system. Our data contrast with findings suggesting that MDA5 is a key innate immune receptor for mRNA vaccination, with MDA5 knockout mice having markedly decreased innate and adaptive immune responses after vaccination with BNT162b2 . The vaccine used in that study was BNT162b2 created with GMP standards whereas our mRNA-containing LNPs were created with cellulose-purified mRNA, and thus could be
responsible for the different results. Furthermore, in vivo MDA5 activation could come from dsRNA products of cell stress rather than dsRNA remnants from mRNA synthesis .
Overall, our data show that three LNPs with structural variations in the ionizable lipid, are detected by TLR4 and activate NF- and IRF transcription factors in innate immune cells. TLR signaling is known to cause up-regulation of molecules involved in T-cell crosstalk and production of cytokines and TLR agonists have been utilized as adjuvants in clinical trials and licensed vaccines for such activation . Our transcriptional data showing broad upregulation of molecules involved in cell activation pathways suggests that ionizable LNPs may be providing additional immune stimulation through, as of yet, unconfirmed pathways. Investigation into the mechanism behind TLR4 engagement may allow for the ability to adjust the lipid composition to balance tolerability with strength of stimulation. Additionally, our data indicates that the LNP, more so than RNA, is the predominant driver of mRNA vaccine-induced activation of THP- 1 monocytes. Therefore, efforts could be focused on understanding how to create LNPs to target specific signaling pathways rather than further modification of mRNA to help guide vaccine and adjuvant design for safer, more effective, and potentially more durable mRNA vaccines.
Methods
Cell lines and culture
THP-1-Dual a dual reporter human monocyte cell line (InvivoGen, San Diego, CA, USA) was obtained for use in reporter assays, microscopy, and flow cytometry. This suspension cell line stably expresses a secreted alkaline phosphatase (SEAP) reporter inducible by NF- and a secreted lucia luciferase reporter inducible by IRF pathway activation. Knockouts for MyD88, TRIF, and TLR4 in the dual-reporter background were also obtained from InvivoGen. THP-1 cells were grown in RPMI 1640 with 2 mM L-glutamine (Cytiva, Marlborough, MA, USA) supplemented with 25 mM HEPES (Thermo Fisher, Waltham, MA, USA), heatinactivated FBS, and Pen-Strep ( ). Cells were maintained at with . Selection antibiotics, Blasticidin ( ) and Zeocin ( ) (InvivoGen, San Diego, CA, USA), were applied every other passage per manufacturer recommendation.
A549-Dual a dual reporter human respiratory epithelial cell line (InvivoGen, San Diego, CA, USA) was obtained for use in reporter assays. These cells were grown in DMEM with 2 mM L-glutamine (Thermo Fisher, Waltham, MA, USA) supplemented with heat-inactivated FBS and Pen-Strep ( ). Cells were maintained at with .
mRNA preparation and lipid nanoparticle formulation
The diproline-modified SARS-CoV-2 Omicron variant spike glycoprotein (S2P) was codon optimized and cloned into an in vitro transcription template plasmid containing a T7 promoter, and UTR regions, and a 100 nucleotide poly(A) tail. mRNA was synthesized and cotranscriptionally capped using the Megascript Transcription kit (Thermo Fisher, Cat# AMB 1334) and the CleanCapTM dinucleotide system (Trilink Biotechnologies), precipitated, and purified using a modified cellulose-based chromatography method. Length and mRNA integrity were assessed using the Agilent Bioanalyzer 2100 system. Removal of double stranded RNA (dsRNA) contaminants was confirmed using dot blot, and endotoxin levels were measured using the Genscript ToxiSensor chromogenic assay ( ). The S2P mRNA was stored frozen ( ) at in nuclease and pyrogen free water until use. mRNA was encapsulated into lipid nanoparticles as previously described (51). LNP lipid components were mixed in an ethanolic solution, and rapidly mixed with an aqueous phase containing the mRNA, dialyzed, and concentrated to , and stored at until used (Supplementary Table 1, Supplementary Fig. 1).
LNP-ALC315, following the BNT162b2 formulation, was comprised of the ALC-0315 ionizable lipid, cholesterol, DSPC, and the ALC-0159 PEG-lipid. LNP-SM102, following the mRNA-1273 formulation, was
comprised of the SM-102 ionizable lipid, cholesterol, DSPC, and PEGDMG PEG-lipid. A third LNP formulation, designated as LNP-1, is proprietary to Acuitas Therapeutics. The proprietary lipid and LNP composition are described in US patent US10,221,127. LNP-1 is comprised of the a proprietary ionizable lipid, DSPC, cholesterol, and a PEG-lipid. LNP-1 was also tagged with the DiO fluorescent marker and designated as DiO-LNP. The fourth formulation made was an LNP, designated as LNP-no ionizable lipid, containing only cholesterol, DSPC, and the ALC-0159 PEG-lipid. LNPs were characterized for their hydrodynamic size, and polydispersity index (PDI), using dynamic light scattering (DLS), and the encapsulation efficiency and concentration measured using the RiboGreen RNA assay (Invitrogen, Cat# R11490). The empty LNP-ALC315 used was 54.07 nm with a PDI of 0.28 and zeta potential of -4.447. The empty LNP-SM102 used was 51.590 nm with a PDI of 0.1605 and zeta potential of -4.417. The LNP without the ionizable lipid was larger at 80.26 nm with a PDI of 0.05 and zeta potential of -8.548.
mRNA-LNPs were made with LNP-1 and LNP-ALC315, designated as mRNA-LNP-1 and mRNA-LNP-ALC315. The mRNA-LNP-ALC315 had an encapsulation efficiency of diameter, 0.1090 PDI, and zeta potential of -11.55 . LNP concentrations are given as the mRNA equivalent to enable comparison with the mRNA-LNP as the concentration of mRNA.
Reporter assays
For both NF- and IRF reporter assays using the THP-1 cell line, cells were resuspended in RPMI 1640 with 2 mM L-glutamine (Cytiva, Marlborough, MA, USA), heat-inactivated FBS, and Pen-Strep ( ) at a concentration of cells and 1 ml aliquoted per well of a 24-well plate. Reagents used individually to treat THP-1 cells included the LNP-1, LNP-ALC315, LNP-SM102, LNP-no ionizable lipid, mRNA-LNP-1, mRNA-LNP-ALC315, VacciGrade R848 (InvivoGen, San Diego, CA, USA), synthetic MPLA (InvivoGen, San Diego, CA, USA), IFNa (Miltenyi, Gaithersburg, MD, USA), LPS (Sigma-Aldrich, St. Louis, MO, USA), and TNFa (Thermo Fisher, Waltham, MA, USA). Empty LNPs and mRNA-LNPs were generously provided by the Alameh lab and University of Pennsylvania synthesized as described above .
For analysis of endocytosis, cells were either left untreated or treated with the endocytosis inhibitors dynasore , Fisher Scientific, Hampton, NH, USA), cytochalasin D ( , Fisher Scientific, Hampton, NH, USA), chloroquine ( , Fisher Scientific, Hampton, NH, USA) or methyl- cyclodextrin , Thermo Fisher, Waltham, MA, USA). THP-1s were treated two hours before addition of stimuli and every 24 h afterward. After thorough resuspension of reagent with THP-1, of supernatant was removed at the indicated time points for measurement of reporter molecules. Cell counts were taken prior to removal of supernatant on each day to ensure similar growth curves. For the NF- reporter assay, of supernatant was mixed with of Quanti-Blue Solution (InvivoGen, San Diego, CA, USA) in a 96-well plate and incubated at with for 24 h . The absorbance at 600 nm was read using the Promega GlowMax plate reader (Promega, Madison, WI, USA). For the IRF reporter assay, of supernatant was mixed with of Quanti-Luc Solution (InvivoGen, San Diego, CA, USA) and the luminescence was immediately read at 0.1 s integration using the Promega GlowMax plate reader. Biological and technical replicates were performed for each condition in each assay.
For both NF- and IRF reporter assays using the A549 cell line, cells were seeded in each well of a 48 well plate with a final volume of and incubated at with for 24 h . Supernatant was replaced with fresh media and stimulation was added. Cells were incubated for the designated time period and supernatant was removed for reporter assay as described above.
Fluorescent microscopy
Cells were incubated with or without DiO-tagged LNP (DiO-LNP) at a concentration of cells at with in 24 well plate.
DiO-LNP was generously provided by the Alameh lab and the University of Pennsylvania synthesized as described above . After incubation, cells were pelleted, washed, and resuspended in PBS. Cells were allowed to settle onto a coverslip at the bottom of 6 -well plates for 30 min . Supernatant was gently removed, and cells were fixed with a paraformaldehyde solution for 15 min . After fixation, cells were washed with PBS for two minutes twice. For intracellular staining, cells were permeabilized with saponin during fixation, blocked with bovine serum albumin (BSA) (Miltenyi, Gaithersburg, MD, USA) for 1 h , and stained with anti-TLR4 antibody (antiCD284) (Cat. No. 14-9917-82) (Thermo Fisher, Waltham, MA, USA) and anti-Rab5a antibody (Cat. No. 11947-1-AP) (Thermo Fisher, Waltham, MA, USA) for 1 h . Cells were subsequently washed with additional BSA and stained with secondary antibodies; anti-mouse IgG Alexa Fluor555 (Cat No. 4409S) (Cell Signaling Technology, Danvers, MA, USA) and anti-rabbit IgG Alexa Fluor647 (Cat No. 4414S) (Cell Signaling Technology, Danvers, MA, USA) for 1 h . Then, cells were stained with DAPI (BioLegend, San Diego, CA, USA) for 10 min in the dark and subsequently washed. Coverslips were mounted using ProLong Gold antifade reagent (Invitrogen, Waltham, MA, USA) and imaged on the Zeiss700 confocal microscope (Zeiss, Jena, Germany) using the ZEN 2.3 SP1 software.
Flow cytometry
THP-1 cells at a concentration of cells were incubated with DiOLNP , LNP-1 , LNP-ALC315 , LNP-SM102 , mRNA-LNP-1 , mRNA-LNP-ALC315 , VacciGrade R848 ( ) (InvivoGen, San Diego, CA, USA), or media alone at with . For surface marker investigation, cells were washed with PBS and stained with Human TruStain FcX (BioLegend, San Diego, CA, USA) at for 5 min to prevent nonspecific binding followed by surface staining with antibodies listed in Supplementary Table 2 at for 20 min . Cells were fixed with a paraformaldehyde solution for 15 min . For assessing kinetics of DiO-LNP uptake, cells were immediately washed and fixed. Cells were run on the Cytek Aurora Spectral Flow Cytometer and analyzed using FlowJo v10.
Sequencing
THP-1 cells at a concentration of cells were incubated with or without LNP-1 at for 24 h at prior to RNA extraction with Qiagen RNeasy Plus Mini Kit (Qiagen, Hilden, Germany). Each condition was set up in quadruplicate with 1 million cells per sample. RNA was quantified using Qubit RNA Broad Range Assay Kit (Thermo Fisher, Waltham, MA, USA). For all samples, a total RNA amount of was used as input for library preparation using the TruSeq Stranded total RNA Library Preparation Kit (Illumina, San Diego, CA, USA) using IDT for Illumina – TruSeq RNA UD Indexes. Sequencing libraries were quantified by real-time PCR using the KAPA Library Quantification Complete kit (Roche, Basel, Switzerland) and assessed for size distribution and absence of free adapters and adapter dimers on a Fragment Analyzer. Sequencing libraries were pooled and quantified by real-time PCR as above, clustered and sequenced on an Illumina NovaSeq 6000 using a Reagent Kit v1.5 (200 cycles) with run parameters generating paired-end reads at 100 bp length.
Sequencing analysis
Fastq files obtained from the sequencing run were initially processed through the nf-core rnaseq pipeline via NextFlow using recommended pipeline settings . The pipeline includes FASTQC, Trim Galore, SortMeRNA, STAR, featureCounts, StringTie, Salmon, Qualimap, dupRadar, Preseq, and RSeQC. Reads were aligned to the GRCh37 reference genome. Subsequent analysis was performed with R 4.3.2 using Jupyter Notebooks and the Tidyverse (v2.0.0) package. We used DESeq2 (v1.42.0) to identify differentially expressed genes (DEGs) between unstimulated and LNP-1stimulated samples with 4 samples in each group. The Benjamini-Hochberg procedure was used for multiple testing correction. Additionally, we implemented a false discovery rate of 0.01 and a cutoff of a logfold change above 1 or below -1 . Then, we performed logfold change shrinkage using
apeglm (v1.24.0) and matched gene IDs to gene symbols to create a table of DEGs. The table of DEGs was run through Human MSigDB Collections (v7.5.1): hallmark gene sets (H), KEGG (C2: curated gene sets), and the GO (C5: ontology gene sets – GO: Gene Ontology gene sets). Gene Set Enrichment Analysis (GSEA) was performed using the Bioconducter clusterProfiler (v4.8.3) package.
Statistical analysis
Statistical analyses were performed using GraphPad PRISM (v10, GraphPad Software Inc., Boston, MA, USA) using a one-way ANOVA followed by Dunnet’s test for multiple comparisons to a single control or by Tukey’s test for multiple comparisons between all groups. Results are expressed as means with standard error of the mean (SEM). Values of were considered statistically significant. For transcriptomics data, the Benjamini-Hochberg procedure was implemented during determination of DEGs and pathway enrichment analysis to correct for multiple comparisons. The adjusted -values were reported for these results. Significance for GSEA was determined by comparing the enrichment score to a null distribution through permutation testing.
Data availability
Data will be made available upon request. Sequencing data is available in the NCBI Sequence Read Archive under reference number PRJNA1196969.
Received: 2 May 2024; Accepted: 24 March 2025;
Published online: 17 April 2025
References
Verbeke, R., Lentacker, I., De Smedt, S.C. & Dewitte, H. The dawn of mRNA vaccines: The COVID-19 case. J. Controlled Release 333, 511-520 (2021).
O’Shaughnessy, J. A. (ed U.S. Food & Drug Administration) 1-23 (2022).
Patalon, T., et al. Waning effectiveness of the third dose of the BNT162b2 mRNA COVID-19 vaccine. Nat. Commun. 13, 3203 (2022).
Arunachalam, P. S. et al. Durability of immune responses to mRNA booster vaccination against COVID-19. J. Clin. Invest. org/10.1172/JCI167955 (2023).
Ishii, T., et al. Waning cellular immune responses and predictive factors in maintaining cellular immunity against SARS-CoV-2 six months after BNT162b2 mRNA vaccination. Sci. Rep. 13, 9607 (2023).
Baden, L. R. et al. Efficacy and Safety of the mRNA-1273 SARS-CoV2 Vaccine. N. Engl. J. Med. 384, 403-416 (2021).
Corbett, K. S. et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 586, 567-571 (2020).
Puranik, A., et al. Durability analysis of the highly effective mRNA-1273 vaccine against COVID-19. PNAS Nexus 1, pgac058 (2022).
Polack, F.P. et al. Safety and Efficacy of the BNT162b2 mRNA Covid19 Vaccine. N. Engl. J. Med. 383, 2603-2615 (2020).
Coggins, S.A., et al. Adverse Effects and Antibody Titers in Response to the BNT162b2 mRNA COVID-19 Vaccine in a Prospective Study of Healthcare Workers. Open Forum Infect. Dis. 9, ofab575 (2022).
Li, Y. & Chen, X. CpG 1018 Is an Effective Adjuvant for Influenza Nucleoprotein. Vaccines (Basel) 11 https://doi.org/10.3390/ vaccines11030649 (2023).
Pulendran, B., P, S. A. & O’Hagan, D. T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 20, 454-4-75 (2021).
Andries, O. et al. N(1)-methylpseudouridine-incorporated mRNA outperforms pseudouridine-incorporated mRNA by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. J. Control Release 217, 337-344 (2015).
Freund, I., Eigenbrod, T., Helm, M. & Dalpke, A.H. RNA Modifications Modulate Activation of Innate Toll-Like Receptors. Genes10,92 (2019).
Kariko, K., Buckstein, M., Ni, H. & Weissman, D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity23, 165-175 (2005).
Baiersdorfer, M. et al. A Facile Method for the Removal of dsRNA Contaminant from In Vitro-Transcribed mRNA. Mol. Ther. Nucleic Acids 15, 26-35 (2019).
Connors, J. et al. Lipid nanoparticles (LNP) induce activation and maturation of antigen presenting cells in young and aged individuals. Commun. Biol. 6, 188 (2023).
Alameh, M.G. et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity 54, 2877-2892 e2877 (2021).
Tahtinen, S. et al. IL-1 and IL-1ra are key regulators of the inflammatory response to RNA vaccines. Nat. Immunol. 23, 532-542 (2022).
Sun, D. & Lu, Z.R. Structure and Function of Cationic and Ionizable Lipids for Nucleic Acid Delivery. Pharm. Res. 40, 27-46 (2023).
Litvinova, V.R., Rudometov, A.P., Karpenko, L.I. & Ilyichev, A.A. mRNA Vaccine Platform: mRNA Production and Delivery. Russ. J. Bioorg. Chem. 49, 220-235 (2023).
Zhang, L. et al. Effect of mRNA-LNP components of two globallymarketed COVID-19 vaccines on efficacy and stability. NPJ Vaccines 8, 156 (2023).
Hald Albertsen, C., et al. The role of lipid components in lipid nanoparticles for vaccines and gene therapy. Adv. Drug Deliv. Rev. 188, 114416 (2022).
Pham, C.T. et al. Variable antibody-dependent activation of complement by functionalized phospholipid nanoparticle surfaces. J. Biol. Chem. 286, 123-130 (2011).
Tanaka, T. et al. DiC14-amidine cationic liposomes stimulate myeloid dendritic cells through Toll-like receptor 4. Eur. J. Immunol. 38, 1351-1357 (2008).
Halamoda-Kenzaoui, B. & Bremer-Hoffmann, S. Main trends of immune effects triggered by nanomedicines in preclinical studies. Int J. Nanomed. 13, 5419-5431 (2018).
Lonez, C., Vandenbranden, M. & Ruysschaert, J.M. Cationic lipids activate intracellular signaling pathways. Adv. Drug Deliv. Rev. 64, 1749-1758 (2012).
de Groot, A. M. et al. Immunogenicity Testing of Lipidoids In Vitro and In Silico: Modulating Lipidoid-Mediated TLR4 Activation by Nanoparticle Design. Mol. Ther. Nucleic Acids 11, 159-169 (2018).
Lonez, C. et al. Critical residues involved in Toll -like receptor 4 activation by cationic lipid nanocarriers are not located at the lipopolysaccharide-binding interface. Cell Mol. Life Sci. 72, 3971-3982 (2015).
Forster III, J., Nandi, D. & Kulkarni, A. mRNA-carrying lipid nanoparticles that induce lysosomal rupture activate NLRP3 inflammasome and reduce mRNA transfection efficiency. Biomater. Sci. 10, 5566-5582 (2022).
Kedmi, R., Ben-Arie, N. & Peer, D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials 31, 6867-6875 (2010).
Ju, Y. et al. Anti-PEG Antibodies Boosted in Humans by SARS-CoV-2 Lipid Nanoparticle mRNA Vaccine. ACS Nano 16, 11769-11780 (2022).
Dey, A. K. et al. Tuning the Immunostimulation Properties of Cationic Lipid Nanocarriers for Nucleic Acid Delivery. Front Immunol. 12, 722411 (2021).
Deguine, J & Barton, GM MyD88: a central player in innate immune signaling. F1000Prime Rep. 6, 97 (2014).
Kawasaki, T. & Kawai, T. Toll-like receptor signaling pathways. Fronteirs Immunol. 5, 461 (2014).
Li, C. et al. Mechanisms of innate and adaptive immunity to the PfizerBioNTech BNT162b2 vaccine. Nat. Immunol. 23, 543-555 (2022).
Billack, B. Macrophage activation: role of toll-like receptors, nitric oxide, and nuclear factor kappa B.Am. J. Pharm. Educ. 70, 102 (2006).
Ernst, O., Vayttaden, S.J. & Fraser, I.D.C. Measurement of NF-kappaB Activation in TLR-Activated Macrophages. Methods Mol. Biol. 1714, 67-78 (2018).
Peters, K.L., Smith, H.L., Stark, G.R. & Sen, G.C. IRF-3-dependent, NFkappa B- and JNK-independent activation of the 561 and IFN-beta genes in response to double-stranded RNA. Proc. Natl. Acad. Sci. USA 99, 6322-6327 (2002).
Rennick, J.J., Johnston, A.P.R. & Parton, R.G. Key principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. Nat. Nanotechnol. 16, 266-276 (2021).
Mussbacher, M., Derler, M., Basilio, J. & Schmid, J.A. NF-kappaB in monocytes and macrophages – an inflammatory master regulator in multitalented immune cells. Front Immunol. 14, 1134661 (2023).
Mussbacher, M. et al. Cell Type-Specific Roles of NF-kappaB Linking Inflammation and Thrombosis. Front Immunol. 10, 85 (2019).
Ma, F. et al. Positive feedback regulation of type I interferon by the interferon-stimulated gene STING. EMBO Rep. 16, 202-212 (2015).
Korzun, T. et al. Lipid Nanoparticles Elicit Reactogenicity and Sickness Behavior in Mice Via Toll-Like Receptor 4 and Myeloid Differentiation Protein 88 Axis. ACS Nano https://doi.org/10.1021/ acsnano.4c05088 (2024).
Lee, K.M. & Seong, S.Y. Partial role of TLR4 as a receptor responding to damage-associated molecular pattern. Immunol. Lett. 125, 31-39 (2009).
Chaudhary, N. et al. Amine headgroups in ionizable lipids drive immune responses to lipid nanoparticles by binding to the receptors TLR4 and CD1d. Nat. Biomed. Eng. 8, 1483-1498 (2024).
Tsutsumi-Ishii, Y. & Nagaoka, I. Modulation of human beta-defensin-2 transcription in pulmonary epithelial cells by lipopolysaccharidestimulated mononuclear phagocytes via proinflammatory cytokine production. J. Immunol. 170, 4226-4236 (2003).
Ciesielska, A., Matyjek, M. & Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol. Life Sci. 78, 1233-1261 (2021).
Maeshima, N. & Fernandez, RC Recognition of lipid A variants by the TLR4-MD-2 receptor complex. Front Cell Infect. Microbiol 3, 3 (2013).
Park, B.S. et al. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature 458, 1191-1195 (2009).
Carter, D., et al. A structure-function approach to optimizing TLR4 ligands for human vaccines. Clin. Transl. Immunol. 5, e108 (2016).
Farlik, M. et al. Contribution of a TANK-binding kinase 1-interferon (IFN) regulatory factor 7 pathway to IFN-gamma-induced gene expression. Mol. Cell Biol. 32, 1032-1043 (2012).
Fitzgerald, K.A. et al. IKKepsilon and TBK1 are essential components of the IRF3 signaling pathway. Nat. Immunol. 4, 491-496 (2003).
Uematsu, S. & Akira, S. Toll-like receptors and Type I interferons. J. Biol. Chem. 282, 15319-15323 (2007).
Ng, S.L. et al. IkappaB kinase epsilon (IKK(epsilon)) regulates the balance between type I and type II interferon responses. Proc. Natl. Acad. Sci. USA 108, 21170-21175 (2011).
Fitzgerald, K.A. et al. LPS-TLR4 signaling to IRF-3/7 and NF-kappaB involves the toll adapters TRAM and TRIF. J. Exp. Med 198, 1043-1055 (2003).
Vangasseri, D.P. et al. Immunostimulation of dendritic cells by cationic liposomes. Mol. Membr. Biol. 23, 385-395 (2006).
Ngo, W. et al. Identifying cell receptors for the nanoparticle protein corona using genome screens. Nat. Chem. Biol. 18, 1023-1031 (2022).
Liu, J. et al. A five-amino-acid motif in the undefined region of the TLR8 ectodomain is required for species-specific ligand recognition. Mol. Immunol. 47, 1083-1090 (2010).
Vaure, C. & Liu, Y. A comparative review of toll-like receptor 4 expression and functionality in different animal species. Front Immunol. 5, 316 (2014).
Schroder, K. et al. Conservation and divergence in Toll-like receptor 4-regulated gene expression in primary human versus mouse macrophages. Proc. Natl. Acad. Sci. USA 109, E944-E953 (2012).
Schneberger, D., Caldwell, S., Kanthan, R. & Singh, B. Expression of Tolllike receptor 9 in mouse and human lungs. J. Anat. 222, 495-503 (2013).
Wang, Y., Rahman, D. & Lehner, T. A comparative study of stressmediated immunological functions with the adjuvanticity of alum. . Biol. Chem. 287, 17152-17160 (2012).
Welsby, I. et al. Lysosome-Dependent Activation of Human Dendritic Cells by the Vaccine Adjuvant QS-21. Front Immunol. 7, 663 (2016).
Ho, N. I. et al. Saponin-based adjuvants enhance antigen crosspresentation in human CD11c(+) CD1c(+) CD5(-) CD163(+) conventional type 2 dendritic cells. J. Immunother Cancer 11 https:// doi.org/10.1136/jitc-2023-007082 (2023).
Furman, D. et al. Apoptosis and other immune biomarkers predict influenza vaccine responsiveness. Mol. Syst. Biol. 9, 659 (2013).
Palchetti, S. et al. The protein corona of circulating PEGylated liposomes. Biochim Biophys. Acta 1858, 189-196 (2016).
Wang, Y. et al. Inhibition of clathrin/dynamin-dependent internalization interferes with LPS-mediated TRAM-TRIF-dependent signaling pathway. Cell Immunol. 274, 121-129 (2012).
Vig, S., et al. ER stress promotes mitochondrial DNA mediated type-1 interferon response in beta-cells and interleukin-8 driven neutrophil chemotaxis. Front Endocrinol. (Lausanne) 13, 991632 (2022).
Chen, Y.G. & Hur, S. Cellular origins of dsRNA, their recognition and consequences. Nat. Rev. Mol. Cell Biol. 23, 286-301 (2022).
Dhir, A. et al. Mitochondrial double-stranded RNA triggers antiviral signalling in humans. Nature 560, 238-242 (2018).
Toussi, D.N. & Massari, P. Immune Adjuvant Effect of Molecularlydefined Toll-Like Receptor Ligands. Vaccines (Basel) 2, 323-353 (2014).
Ewels, P.A. et al. The nf-core framework for community-curated bioinformatics pipelines. Nat. Biotechnol. 38, 276-278 (2020).
nf-core/rnaseq v. 3.12.0 (Zenodo, 2023).
Zhu, A., Ibrahim, J.G. & Love, M.I. Heavy-tailed prior distributions for sequence count data: removing the noise and preserving large differences. Bioinformatics 35, 2084-2092 (2019).
Acknowledgements
We thank Kateryna Lund and Sintayehu Gebreyohannes at the USU Biomedical Instrument Core (BIC) for flow cytometry assistance; Dennis McDaniel at the USU BIC for microscopy assistance; and Andrew Frank at the Student Bioinformatics Initiative for assistance with transcriptomic data analysis. This work was supported by funding through the Department of Pediatrics RAMP grant and federal funds from the Defense Health Program (HU00012020067, HU00012120094) and the Immunization Healthcare Branch (HU00012120104) of the Defense Health Agency, United States Department of Defense. The sponsors had no involvement in the study design, the collection of data, the analysis of data, the interpretation of data, the writing of the report, or in the decision to submit the article for publication. The opinions and assertions expressed herein are those of the authors and are not to be construed as reflecting the views of USUHS, the U.S. Air Force, the U.S. Army, U.S. Navy, the U.S. military at large, or the U.S. Department of Defense. Title 17 U.S.C. 105 provides that ‘Copyright protection under this title is not available for any work of the United States Government.’ Title 17 U.S.C. 101 defines a “United States Government work” as a work prepared by a military service member or employee of the United States Government as part of that person’s official duties.
Author contributions
A.Z. and A.M.W.M. conceived and designed the study, analyzed, and interpreted data. M.G.A. provided the LNPs and contributed to study design. H.S. synthesized LNPs and mRNA-vaccines. E.M. contributed to study design. A.Z. performed experiments. G.S. and C.D. performed RNA
sequencing. Z.L. assisted in data analysis. A.Z. and A.M.W.M. conceived and wrote the manuscript. Z.L., M.G.A., and E.M. assisted in writing and reviewing the manuscript.
Competing interests
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The opinions and assertions expressed herein are those of the authors and do not reflect the official policy or position of the Uniformed Services University of the Health Sciences or the Department of Defense, or the Henry M. Jackson Foundation for the Advancement of Military Medicine, Inc. (HJF). Mention of trade names, commercial products, or organizations does not imply endorsement by the U.S. Government.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply 2025
1Department of Pediatrics, Uniformed Services University of Health Sciences, Bethesda, MD, USA. Henry M Jackson Foundation for the Advancement of Military Medicine, Bethesda, MD, USA. Department of Anatomy, Physiology & Genetics, Uniformed Services University of Health Sciences, Bethesda, MD, USA. Department of Pathology and Laboratory Medicine, Children’s Hospital of Philadelphia, Philadelphia, PA, USA. Department of Pathology and Laboratory Medicine, University of Pennsylvania, Philadelphia, PA, USA. Penn Institute for RNA Innovation, University of Pennsylvania, Philadelphia, PA, USA. Department of Microbiology and Immunology, Uniformed Services University of Health Sciences, Bethesda, PA, USA.
utmb
Health
⟶ e-mail: allison.malloy@usuhs.edu
Sealy Institute for
Vaccine Sciences