DOI: https://doi.org/10.1038/s41467-024-45115-3
PMID: https://pubmed.ncbi.nlm.nih.gov/38280872
تاريخ النشر: 2024-01-27
علاقات ستوكيومترية طويلة الأمد متسقة بين المغذيات والكلوروفيل-أ عبر البحيرات الضحلة
تاريخ القبول: 15 يناير 2024
تاريخ النشر على الإنترنت: 27 يناير 2024
الملخص
تتعرض النظم البيئية المائية للتهديد بسبب الإثراء الناتج عن تلوث العناصر الغذائية. في البحيرات، يتسبب الإثراء في مجموعة من الآثار الضارة، مثل ازدهار الطحالب الضارة، وموت الأسماك، وزيادة انبعاثات الميثان. ومع ذلك، فإن الاستجابات الخاصة بالبحيرات للتغيرات في العناصر الغذائية متغيرة للغاية، مما يعقد إدارة الإثراء. قد تكون هذه الاستجابات الخاصة بالبحيرات ناتجة عن محركات عشوائية قصيرة الأمد تطغى على العلاقات الطويلة الأمد المستقلة عن البحيرات بين العوالق النباتية والعناصر الغذائية. هنا، نوضح أن هناك علاقات ستيوكيومترية قوية طويلة الأمد بين العناصر الغذائية والكلوروفيل أ
الخصائص
التغيرات الدقيقة
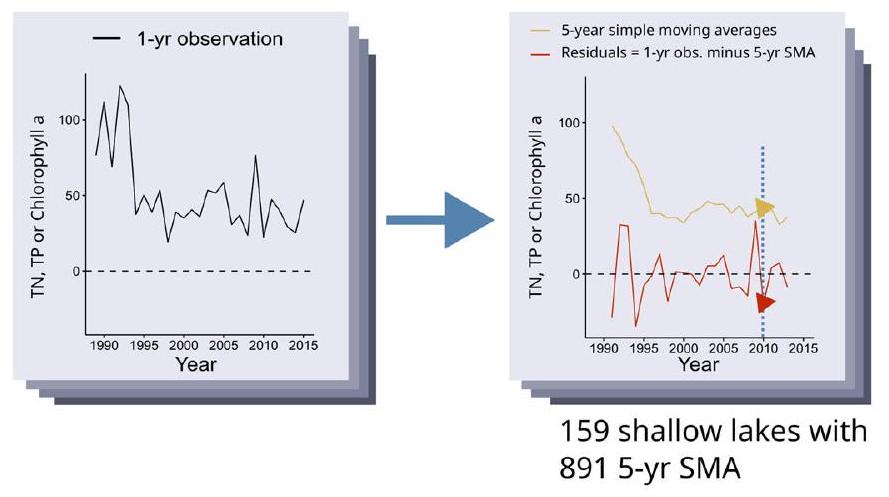
تقسيم إلى نوافذ نطاق TN:TP
SMA لمدة 5 سنوات، العناصر الغذائية الكلية مقابل Chla على طول TN:TP
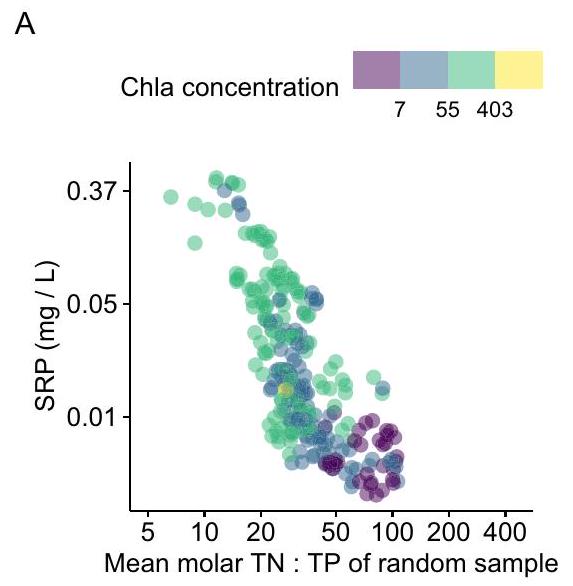
متغير ستيوكيومتري، حيث يمكن أن تتراكم العناصر الغذائية المذابة عندما يكون امتصاص العوالق النباتية لعنصر غذائي مذاب مقيدًا بتوافر عنصر غذائي آخر (الشكل 1). في هذه الحالة، قد يتراكم العنصر الغذائي في الفائض النسبي في عمود الماء في شكل مذاب وتفاعلي
النتائج والمناقشة
المغذيات – روابط الكلوروفيل أ للمتوسطات طويلة الأجل
عند
في البحيرات، تم الحفاظ على البروتينات المذابة بشكل انتقائي على مدى عقود إلى قرون، ربما بسبب التحلل البكتيري والضوئي
في مسار نزع النيتروجين-النيتروجين العضوي المذاب، مع تراكم النيتروجين العضوي المذاب المقاوم، على الأرجح البروتينات المذابة، في الغالبية العظمى من البحيرات (الشكل 3B). يشبه هذا المسار التخزيني المترابط مفهوم مضخة الكربون الميكروبية في المحيطات
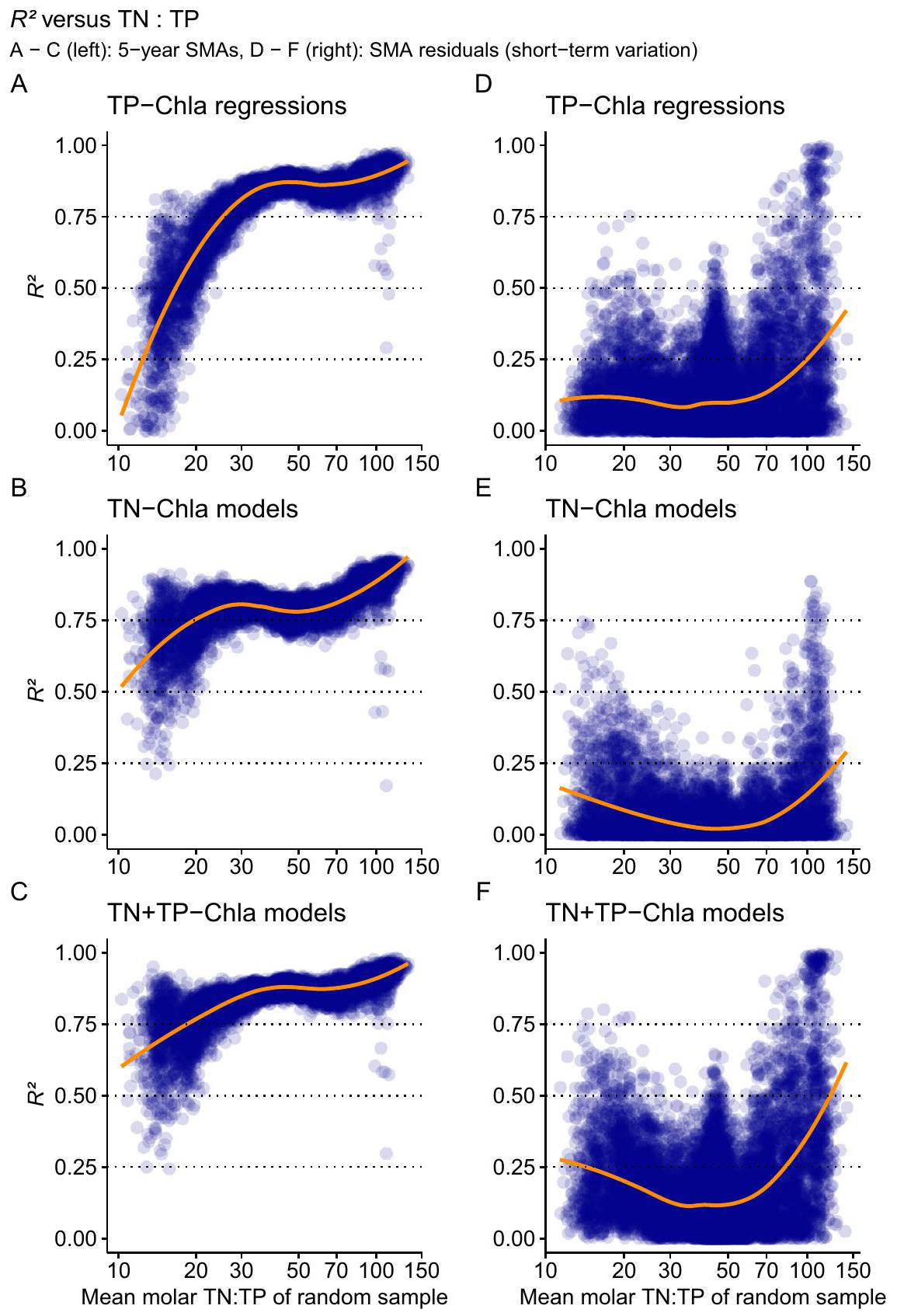
نماذج المغذيات الفردية والإضافية مقابل TN:TP
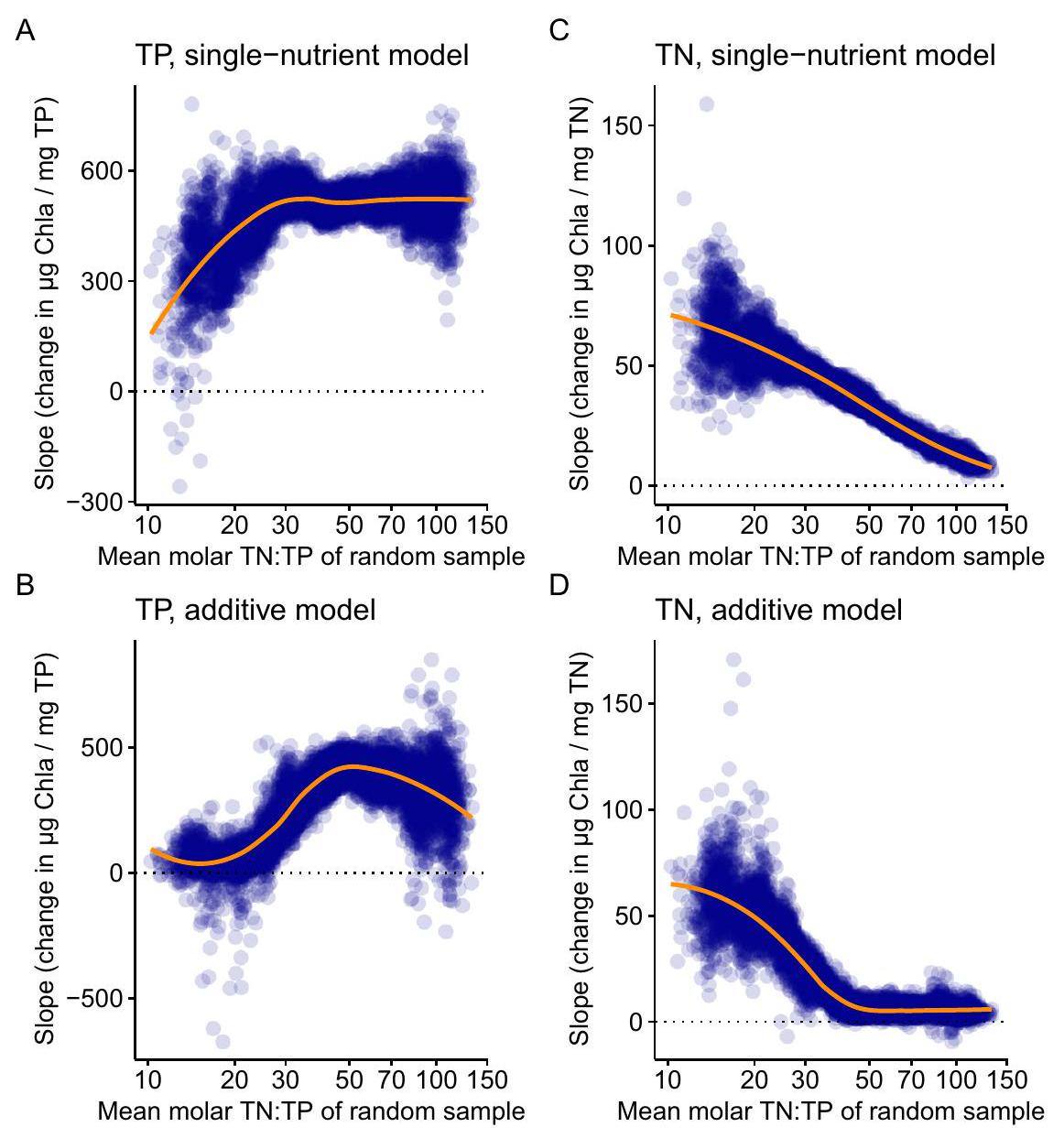
النماذج الخطية العامة مع توزيع غاما لـ Chla لمدة 5 سنوات. كلما كانت النقاط أقل شفافية، زادت الحلول المتداخلة التي تم العثور عليها للمنحدرات بواسطة إجراء bootstrap (مما يدل على خطأ المنحدرات). الخط البرتقالي هو متوسط الاستجابة بناءً على دالة LOESS.
كشفت عن أهمية نسبية أعلى لنزع النيتروجين مقارنة بتثبيت النيتروجين الجوي في الغالبية العظمى من البحيرات المدروسة
مقارنة روابط المغذيات-الكلوروفيل أ في البيانات طويلة الأمد وقصيرة الأمد
التحكم في المغذيات للتغذية الزائدة على نطاق النظام البيئي هو عملية تدريجية
كل ملاحظة TN وTP. لذلك، تتضمن اللوحة A 278 ملاحظة من 37 بحيرة ضحلة، وتحتوي اللوحة B على 620 ملاحظة من 138 بحيرة ضحلة. تم تحويل المحاور إلى اللوغاريتمات لأن نسب TN:TP اتبعت توزيع لوغاريتمي طبيعي، ولجعل تركيزات SRP ونترات-N الصغيرة مرئية.
المراقبة متعددة السنوات أو التجارب. استثناء ملحوظ هو دراسة حديثة أجراها كوسر وآخرون.
استخدام العلاقات الاستوكيومترية طويلة الأجل في إدارة البحيرات وحوض المياه.
تسمح للمديرين بإجراء توقعات طويلة الأجل لنجاح إدارة الإثراء بناءً على نسب TN:TP وتركيزاتها.
طرق
جمع البيانات
حساب متوسطات موسم النمو والمتوسطات المتحركة البسيطة
الفصل إلى نطاقات مختلفة من نسب TN:TP
تراوحت النوافذ من
الإحصائيات وإعادة أخذ العينات باستخدام طريقة البوتستراب
البرمجيات والحزم
ملخص التقرير
توفر البيانات
توفر الكود
References
- Jeppesen, E. et al. Lake responses to reduced nutrient loading an analysis of contemporary long-term data from 35 case studies. Freshw. Biol. 50, 1747-1771 (2005).
- Paerl, H. W. et al. It takes two to Tango: When and where dual nutrient (N & P) reductions are needed to protect lakes and downstream ecosystems. Environ. Sci. Technol. 50, 10805-10813 (2016).
- Søndergaard, M., Lauridsen, T. L., Johansson, L. S. & Jeppesen, E. Nitrogen or phosphorus limitation in lakes and its impact on phytoplankton biomass and submerged macrophyte cover. Hydrobiologia 795, 35-48 (2017).
- Wurtsbaugh, W. A., Paerl, H. W. & Dodds, W. K. Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum. WIREs Water 6, e1373 (2019).
- Guildford, S. J. & Hecky, R. E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? Limnol. Oceanogr. 45, 1213-1223 (2000).
- Moon, D. L., Scott, J. T. & Johnson, T. R. Stoichiometric imbalances complicate prediction of phytoplankton biomass in U.S. Lakes: Implications for nutrient criteria. Limnol. Oceanogr. 66, 2967-2978 (2021).
- Lewis, W. M. & Wurtsbaugh, W. A. Control of Lacustrine phytoplankton by nutrients: erosion of the phosphorus paradigm. Int. Rev. Hydrobiol. 93, 446-465 (2008).
- Dolman, A. M. & Wiedner, C. Predicting phytoplankton biomass and estimating critical N:P ratios with piecewise models that conform to Liebig’s law of the minimum. Freshw. Biol. 60, 686-697 (2015).
- McCarthy, M. J., Gardner, W. S., Lehmann, M. F. & Bird, D. F. Implications of water column ammonium uptake and regeneration for the nitrogen budget in temperate, eutrophic Missisquoi Bay, Lake Champlain (Canada/USA). Hydrobiologia 718, 173-188 (2013).
- McCarthy, M. J., Gardner, W. S., Lehmann, M. F., Guindon, A. & Bird, D. F. Benthic nitrogen regeneration, fixation, and denitrification in a temperate, eutrophic lake: Effects on the nitrogen budget and cyanobacteria blooms. Limnol. Oceanogr. 61, 1406-1423 (2016).
- Strayer, D. et al. Long-term ecological studies: An illustrated account of their design, operation, and importance to ecology. Occas. Publ. Inst. Ecosyst. Sci. 2, 1-36 (1986).
- Carpenter, S. R. & Leavitt, P. R. Temporal variation in a paleolimnological record arising from a trophic cascade. Ecology 72, 277-285 (1991).
- Kasprzak, P., Padisák, J., Koschel, R., Krienitz, L. & Gervais, F. Chlorophyll a concentration across a trophic gradient of lakes: An estimator of phytoplankton biomass? Limnologica 38, 327-338 (2008).
- Droop, M. R. Vitamin B12 and Marine Ecology. IV. The kinetics of uptake, growth and inhibition in Monochrysis Lutheri. J. Mar. Biol. Assoc. U. Kingd. 48, 689-733 (1968).
- Droop, M. R. The nutrient status of algal cells in continuous culture. J. Mar. Biol. Assoc. U. Kingd. 54, 825-855 (1974).
- Shatwell, T. & Köhler, J. Decreased nitrogen loading controls summer cyanobacterial blooms without promoting nitrogen-fixing taxa: Long-term response of a shallow lake. Limnol. Oceanogr. 64, S166-S178 (2019).
- Glibert, P. M. et al. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 61, 165-197 (2016).
- Trommer, G., Poxleitner, M. & Stibor, H. Responses of lake phytoplankton communities to changing inorganic nitrogen supply forms. Aquat. Sci. 82, 22 (2020).
- Reed, M. H. et al. Effects of filtration timing and pore size on measured nutrient concentrations in environmental water samples. Limnol. Oceanogr.: Methods 21, 1-12 (2023).
- Massé, S., Botrel, M., Walsh, D. A. & Maranger, R. Annual nitrification dynamics in a seasonally ice-covered lake. PLOS ONE 14, eO213748 (2019).
- Small, G. E. et al. Rates and controls of nitrification in a large oligotrophic lake. Limnol. Oceanogr. 58, 276-286 (2013).
- Messager, M. L., Lehner, B., Grill, G., Nedeva, I. & Schmitt, O. Estimating the volume and age of water stored in global lakes using a geo-statistical approach. Nat. Commun. 7, 13603 (2016).
- Kirillin, G. & Shatwell, T. Generalized scaling of seasonal thermal stratification in lakes. Earth-Sci. Rev. 161, 179-190 (2016).
- Scott, J. T., McCarthy, M. J. & Paerl, H. W. Nitrogen transformations differentially affect nutrient-limited primary production in lakes of varying trophic state. Limnol. Oceanogr. Lett. 4, 96-104 (2019).
- Westphal, K., Musolff, A., Graeber, D. & Borchardt, D. Controls of point and diffuse sources lowered riverine nutrient concentrations asynchronously, thereby warping molar N:P ratios. Environ. Res. Lett. 15, 104009 (2020).
- Graeber, D. et al. Bioavailable DOC: Reactive nutrient ratios control heterotrophic nutrient assimilationAn experimental proof of the macronutrient-access hypothesis. Biogeochemistry 155, 1-20 (2021).
- Wachholz, A. et al. Stoichiometry on the edge – Humans induce strong imbalances of reactive C:N:P ratios in streams. Environ. Res. Lett. 18, 044016 (2023).
- Elser, J. J. et al. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135-1142 (2007).
- Seitzinger, S. et al. Denitrification across landscapes and waterscapes: a synthesis. Ecol. Appl. 16, 2064-2090 (2006).
- Holmroos, H., Hietanen, S., Niemistö, J. & Horppila, J. Sediment resuspension and denitrification affect the nitrogen to phosphorus ratio of shallow lake waters. Fundam. Appl. Limnol. 180, 193-205 (2012).
- Filstrup, C. T. & Downing, J. A. Relationship of chlorophyll to phosphorus and nitrogen in nutrient-rich lakes. Inland Waters 7, 385-400 (2017).
- Klausmeier, C. A., Litchman, E., Daufresne, T. & Levin, S. A. Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton. Nature 429, 171-174 (2004).
- Godwin, C. M. & Cotner, J. B. What intrinsic and extrinsic factors explain the stoichiometric diversity of aquatic heterotrophic bacteria? ISME J. 12, 598-609 (2018).
- Goldberg, S. J. et al. Refractory dissolved organic nitrogen accumulation in high-elevation lakes. Nat. Commun. 6, 6347 (2015).
- Elser, J. J. et al. Sustained stoichiometric imbalance and its ecological consequences in a large oligotrophic lake. Proc. Natl Acad. Sci. 119, e2202268119 (2022).
- Jiao, N. et al. Microbial production of recalcitrant dissolved organic matter: Long-term carbon storage in the global ocean. Nat. Rev. Microbiol. 8, 593-599 (2010).
- Zhu, X., Jackson, R. D., DeLucia, E. H., Tiedje, J. M. & Liang, C. The soil microbial carbon pump: From conceptual insights to empirical assessments. Glob. Change Biol. 26, 6032-6039 (2020).
- Taylor, P. G. & Townsend, A. R. Stoichiometric control of organic carbon-nitrate relationships from soils to the sea. Nature 464, 1178-1181 (2010).
- Hansen, A. T., Dolph, C. L. & Finlay, J. C. Do wetlands enhance downstream denitrification in agricultural landscapes? Ecosphere 7, e01516 (2016).
- Schindler, D. W., Carpenter, S. R., Chapra, S. C., Hecky, R. E. & Orihel, D. M. Reducing phosphorus to curb lake eutrophication is a success. Environ. Sci. Technol. 50, 8923-8929 (2016).
- Scott, J. T. & McCarthy, M. J. Nitrogen fixation may not balance the nitrogen pool in lakes over timescales relevant to eutrophication management. Limnol. Oceanogr. 55, 1265-1270 (2010).
- Trautmann, N. M., McCulloch, C. E. & Oglesby, R. T. Statistical determination of data requirements for assessment of lake restoration programs. Can. J. Fish. Aquat. Sci. 39, 607-610 (1982).
- Tilman, D., May, R. M., Lehman, C. L. & Nowak, M. A. Habitat destruction and the extinction debt. Nature 371, 65-66 (1994).
- Weber, T. S. & Deutsch, C. Ocean nutrient ratios governed by plankton biogeography. Nature 467, 550-554 (2010).
- Jeppesen, E. et al. Structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 45, 201-218 (2000).
- Boedecker, A. R., Niewinski, D. N., Newell, S. E., Chaffin, J. D. & McCarthy, M. J. Evaluating sediments as an ecosystem service in western Lake Erie via quantification of nutrient cycling pathways and selected gene abundances. J. Gt. Lakes Res. 46, 920-932 (2020).
- Hoffman, D. K., McCarthy, M. J., Boedecker, A. R., Myers, J. A. & Newell, S. E. The role of internal nitrogen loading in supporting non-N-fixing harmful cyanobacterial blooms in the water column of a large eutrophic lake. Limnol. Oceanogr. 67, 2028-2041 (2022).
- Koussoroplis, A.-M., Schälicke, S., Raatz, M., Bach, M. & Wacker, A. Feeding in the frequency domain: Coarser-grained environments increase consumer sensitivity to resource variability, covariance and phase. Ecol. Lett. 22, 1104-1114 (2019).
- Cusser, S., Helms, J. IV, Bahlai, C. A. & Haddad, N. M. How long do population level field experiments need to be? Utilising data from the 40-year-old LTER network. Ecol. Lett. 24, 1103-1111 (2021).
- EPA, U. Preventing Eutrophication: Scientific support for dual nutrient criteria. (2015).
- Westphal, K. et al. Multi-decadal trajectories of phosphorus loading, export, and instream retention along a catchment gradient. Sci. Total Environ. 667, 769-779 (2019).
- Kronvang, B. et al. Effects of policy measures implemented in Denmark on nitrogen pollution of the aquatic environment. Environ. Sci. Policy 11, 144-152 (2008).
- Hansen, A. T., Dolph, C. L., Foufoula-Georgiou, E. & Finlay, J. C. Contribution of wetlands to nitrate removal at the watershed scale. Nat. Geosci. 11, 127-132 (2018).
- Zak, D. et al. An Assessment of the multifunctionality of integrated buffer zones in Northwestern Europe. J. Environ. Qual. 48, 362-375 (2019).
- Filazzola, A. et al. A database of chlorophyll and water chemistry in freshwater lakes. Sci. Data 7, 310 (2020).
- R. Core Team. R: A Language and Environment for Statistical Computing. (R Foundation for Statistical Computing, 2023).
- QGIS Development Team. QGIS geographic information system. (2022).
- Soranno, P. A. et al. LAGOS-NE: A multi-scaled geospatial and temporal database of lake ecological context and water quality for thousands of US lakes. GigaScience 6, gix101 (2017).
- Isles, P. D. F. The misuse of ratios in ecological stoichiometry. Ecology 101, e03153 (2020).
- Ren, S. et al. Nonparametric bootstrapping for hierarchical data. J. Appl. Stat. 37, 1487-1498 (2010).
- Kruschke, J. Doing Bayesian Data Analysis: A Tutorial with R, JAGS, and Stan. (Academic Press, 2014).
- Davidson, T. A. et al. Bimodality and alternative equilibria do not help explain long-term patterns in shallow lake chlorophyll-a. Nat. Commun. 14, 398 (2023).
الشكر والتقدير
مساهمات المؤلفين
التمويل
المصالح المتنافسة
معلومات إضافية
المواد التكميلية المتاحة على
https://doi.org/10.1038/s41467-024-45115-3.
© المؤلف(ون) 2024
قسم تحليل النظام البيئي المائي، مركز هيلمهولتز للبحث البيئي – UFZ، ماجدبورغ، ألمانيا. كرسي علم الأحياء المائية ومصايد الأسماك، جامعة إستونيا للعلوم الحياتية، تارту، إستونيا. قسم بحوث البحيرات، مركز هيلمهولتز للبحث البيئي – UFZ، ماجدبورغ، ألمانيا. قسم علوم البيئة، و WATEC، C.F. Møllers Allé 3، جامعة آرهوس، آرهوس، الدنمارك. مركز التعليم والبحث الصيني الدنماركي، بكين، الصين. مختبر علم المياه، قسم العلوم البيولوجية ومركز أبحاث وتنفيذ النظام البيئي، جامعة الشرق الأوسط التقنية، أنقرة، تركيا. معهد العلوم البحرية، جامعة الشرق الأوسط التقنية، مرسين، تركيا. معهد التحكم البيئي والتلوث لبحيرات الهضبة، مدرسة العلوم البيئية والبيئية، جامعة يونان، كونمينغ، الصين. البريد الإلكتروني: daniel.graeber@ufz.de
DOI: https://doi.org/10.1038/s41467-024-45115-3
PMID: https://pubmed.ncbi.nlm.nih.gov/38280872
Publication Date: 2024-01-27
Consistent stoichiometric long-term relationships between nutrients and chlorophylla across shallow lakes
Accepted: 15 January 2024
Published online: 27 January 2024
Abstract
Aquatic ecosystems are threatened by eutrophication from nutrient pollution. In lakes, eutrophication causes a plethora of deleterious effects, such as harmful algal blooms, fish kills and increased methane emissions. However, lake-specific responses to nutrient changes are highly variable, complicating eutrophication management. These lake-specific responses could result from short-term stochastic drivers overshadowing lake-independent, long-term relationships between phytoplankton and nutrients. Here, we show that strong stoichiometric long-term relationships exist between nutrients and chlorophyll
characteristics
variability obscuring subtle changes
Split into TN:TP range windows
5-yr SMA, total nutrients versus Chla along TN:TP
stoichiometric variant, where dissolved nutrients can accumulate when phytoplankton assimilation of a dissolved nutrient is constrained by the availability of another nutrient (Fig. 1). In this case, the nutrient in relative excess may accumulate in the water column in dissolved, reactive form
Results and discussion
Nutrient – chlorophyll a links for long-term averages
at
lakes, dissolved proteins were selectively preserved over decadal to centennial timescales potentially due to bacterial and photochemical degradation
in a coupled denitrification-dissolved organic N pathway, with accumulation of refractory dissolved organic N , likely dissolved proteins, in the majority of lakes (Fig. 3B). Such a coupled storage pathway resembles the microbial carbon pump concept in oceans
Single-nutrient and additive models versus TN : TP
generalised linear models with gamma distribution of Chla for 5-year SMAs. The less transparent the points, the more overlapping solutions were found for the slopes by the bootstrap procedure (indicating the error of the slopes). The orange line is the mean response based on a LOESS function.
revealed relatively higher importance of denitrification over atmospheric N fixation in the majority of studied lakes
Comparison of nutrient-chlorophyll a links in long-term and short-term data
nutrient control of eutrophication at the ecosystem scale is a gradual process
every TN and TP observation. Therefore, panel A includes 278 observations from 37 shallow lakes, and panel B includes 620 observations from 138 shallow lakes. The axes have been log-transformed since TN:TP ratios followed a log-normal distribution, and to make small SRP and nitrate-N concentrations visible.
multi-year monitoring or experiments. One notable exception is a recent study by Cusser et al.
Usage of long-term stoichiometric relationships in lake and catchment management
allow managers to make long-term predictions of eutrophication management success based on TN:TP ratios and their concentrations.
Methods
Data collection
Calculation of growing season means and simple moving averages
Separation into different TN:TP ratio ranges
windows ranged from
Statistics and bootstrap resampling
Software and packages
Reporting summary
Data availability
Code availability
References
- Jeppesen, E. et al. Lake responses to reduced nutrient loading an analysis of contemporary long-term data from 35 case studies. Freshw. Biol. 50, 1747-1771 (2005).
- Paerl, H. W. et al. It takes two to Tango: When and where dual nutrient (N & P) reductions are needed to protect lakes and downstream ecosystems. Environ. Sci. Technol. 50, 10805-10813 (2016).
- Søndergaard, M., Lauridsen, T. L., Johansson, L. S. & Jeppesen, E. Nitrogen or phosphorus limitation in lakes and its impact on phytoplankton biomass and submerged macrophyte cover. Hydrobiologia 795, 35-48 (2017).
- Wurtsbaugh, W. A., Paerl, H. W. & Dodds, W. K. Nutrients, eutrophication and harmful algal blooms along the freshwater to marine continuum. WIREs Water 6, e1373 (2019).
- Guildford, S. J. & Hecky, R. E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? Limnol. Oceanogr. 45, 1213-1223 (2000).
- Moon, D. L., Scott, J. T. & Johnson, T. R. Stoichiometric imbalances complicate prediction of phytoplankton biomass in U.S. Lakes: Implications for nutrient criteria. Limnol. Oceanogr. 66, 2967-2978 (2021).
- Lewis, W. M. & Wurtsbaugh, W. A. Control of Lacustrine phytoplankton by nutrients: erosion of the phosphorus paradigm. Int. Rev. Hydrobiol. 93, 446-465 (2008).
- Dolman, A. M. & Wiedner, C. Predicting phytoplankton biomass and estimating critical N:P ratios with piecewise models that conform to Liebig’s law of the minimum. Freshw. Biol. 60, 686-697 (2015).
- McCarthy, M. J., Gardner, W. S., Lehmann, M. F. & Bird, D. F. Implications of water column ammonium uptake and regeneration for the nitrogen budget in temperate, eutrophic Missisquoi Bay, Lake Champlain (Canada/USA). Hydrobiologia 718, 173-188 (2013).
- McCarthy, M. J., Gardner, W. S., Lehmann, M. F., Guindon, A. & Bird, D. F. Benthic nitrogen regeneration, fixation, and denitrification in a temperate, eutrophic lake: Effects on the nitrogen budget and cyanobacteria blooms. Limnol. Oceanogr. 61, 1406-1423 (2016).
- Strayer, D. et al. Long-term ecological studies: An illustrated account of their design, operation, and importance to ecology. Occas. Publ. Inst. Ecosyst. Sci. 2, 1-36 (1986).
- Carpenter, S. R. & Leavitt, P. R. Temporal variation in a paleolimnological record arising from a trophic cascade. Ecology 72, 277-285 (1991).
- Kasprzak, P., Padisák, J., Koschel, R., Krienitz, L. & Gervais, F. Chlorophyll a concentration across a trophic gradient of lakes: An estimator of phytoplankton biomass? Limnologica 38, 327-338 (2008).
- Droop, M. R. Vitamin B12 and Marine Ecology. IV. The kinetics of uptake, growth and inhibition in Monochrysis Lutheri. J. Mar. Biol. Assoc. U. Kingd. 48, 689-733 (1968).
- Droop, M. R. The nutrient status of algal cells in continuous culture. J. Mar. Biol. Assoc. U. Kingd. 54, 825-855 (1974).
- Shatwell, T. & Köhler, J. Decreased nitrogen loading controls summer cyanobacterial blooms without promoting nitrogen-fixing taxa: Long-term response of a shallow lake. Limnol. Oceanogr. 64, S166-S178 (2019).
- Glibert, P. M. et al. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr. 61, 165-197 (2016).
- Trommer, G., Poxleitner, M. & Stibor, H. Responses of lake phytoplankton communities to changing inorganic nitrogen supply forms. Aquat. Sci. 82, 22 (2020).
- Reed, M. H. et al. Effects of filtration timing and pore size on measured nutrient concentrations in environmental water samples. Limnol. Oceanogr.: Methods 21, 1-12 (2023).
- Massé, S., Botrel, M., Walsh, D. A. & Maranger, R. Annual nitrification dynamics in a seasonally ice-covered lake. PLOS ONE 14, eO213748 (2019).
- Small, G. E. et al. Rates and controls of nitrification in a large oligotrophic lake. Limnol. Oceanogr. 58, 276-286 (2013).
- Messager, M. L., Lehner, B., Grill, G., Nedeva, I. & Schmitt, O. Estimating the volume and age of water stored in global lakes using a geo-statistical approach. Nat. Commun. 7, 13603 (2016).
- Kirillin, G. & Shatwell, T. Generalized scaling of seasonal thermal stratification in lakes. Earth-Sci. Rev. 161, 179-190 (2016).
- Scott, J. T., McCarthy, M. J. & Paerl, H. W. Nitrogen transformations differentially affect nutrient-limited primary production in lakes of varying trophic state. Limnol. Oceanogr. Lett. 4, 96-104 (2019).
- Westphal, K., Musolff, A., Graeber, D. & Borchardt, D. Controls of point and diffuse sources lowered riverine nutrient concentrations asynchronously, thereby warping molar N:P ratios. Environ. Res. Lett. 15, 104009 (2020).
- Graeber, D. et al. Bioavailable DOC: Reactive nutrient ratios control heterotrophic nutrient assimilationAn experimental proof of the macronutrient-access hypothesis. Biogeochemistry 155, 1-20 (2021).
- Wachholz, A. et al. Stoichiometry on the edge – Humans induce strong imbalances of reactive C:N:P ratios in streams. Environ. Res. Lett. 18, 044016 (2023).
- Elser, J. J. et al. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135-1142 (2007).
- Seitzinger, S. et al. Denitrification across landscapes and waterscapes: a synthesis. Ecol. Appl. 16, 2064-2090 (2006).
- Holmroos, H., Hietanen, S., Niemistö, J. & Horppila, J. Sediment resuspension and denitrification affect the nitrogen to phosphorus ratio of shallow lake waters. Fundam. Appl. Limnol. 180, 193-205 (2012).
- Filstrup, C. T. & Downing, J. A. Relationship of chlorophyll to phosphorus and nitrogen in nutrient-rich lakes. Inland Waters 7, 385-400 (2017).
- Klausmeier, C. A., Litchman, E., Daufresne, T. & Levin, S. A. Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton. Nature 429, 171-174 (2004).
- Godwin, C. M. & Cotner, J. B. What intrinsic and extrinsic factors explain the stoichiometric diversity of aquatic heterotrophic bacteria? ISME J. 12, 598-609 (2018).
- Goldberg, S. J. et al. Refractory dissolved organic nitrogen accumulation in high-elevation lakes. Nat. Commun. 6, 6347 (2015).
- Elser, J. J. et al. Sustained stoichiometric imbalance and its ecological consequences in a large oligotrophic lake. Proc. Natl Acad. Sci. 119, e2202268119 (2022).
- Jiao, N. et al. Microbial production of recalcitrant dissolved organic matter: Long-term carbon storage in the global ocean. Nat. Rev. Microbiol. 8, 593-599 (2010).
- Zhu, X., Jackson, R. D., DeLucia, E. H., Tiedje, J. M. & Liang, C. The soil microbial carbon pump: From conceptual insights to empirical assessments. Glob. Change Biol. 26, 6032-6039 (2020).
- Taylor, P. G. & Townsend, A. R. Stoichiometric control of organic carbon-nitrate relationships from soils to the sea. Nature 464, 1178-1181 (2010).
- Hansen, A. T., Dolph, C. L. & Finlay, J. C. Do wetlands enhance downstream denitrification in agricultural landscapes? Ecosphere 7, e01516 (2016).
- Schindler, D. W., Carpenter, S. R., Chapra, S. C., Hecky, R. E. & Orihel, D. M. Reducing phosphorus to curb lake eutrophication is a success. Environ. Sci. Technol. 50, 8923-8929 (2016).
- Scott, J. T. & McCarthy, M. J. Nitrogen fixation may not balance the nitrogen pool in lakes over timescales relevant to eutrophication management. Limnol. Oceanogr. 55, 1265-1270 (2010).
- Trautmann, N. M., McCulloch, C. E. & Oglesby, R. T. Statistical determination of data requirements for assessment of lake restoration programs. Can. J. Fish. Aquat. Sci. 39, 607-610 (1982).
- Tilman, D., May, R. M., Lehman, C. L. & Nowak, M. A. Habitat destruction and the extinction debt. Nature 371, 65-66 (1994).
- Weber, T. S. & Deutsch, C. Ocean nutrient ratios governed by plankton biogeography. Nature 467, 550-554 (2010).
- Jeppesen, E. et al. Structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 45, 201-218 (2000).
- Boedecker, A. R., Niewinski, D. N., Newell, S. E., Chaffin, J. D. & McCarthy, M. J. Evaluating sediments as an ecosystem service in western Lake Erie via quantification of nutrient cycling pathways and selected gene abundances. J. Gt. Lakes Res. 46, 920-932 (2020).
- Hoffman, D. K., McCarthy, M. J., Boedecker, A. R., Myers, J. A. & Newell, S. E. The role of internal nitrogen loading in supporting non-N-fixing harmful cyanobacterial blooms in the water column of a large eutrophic lake. Limnol. Oceanogr. 67, 2028-2041 (2022).
- Koussoroplis, A.-M., Schälicke, S., Raatz, M., Bach, M. & Wacker, A. Feeding in the frequency domain: Coarser-grained environments increase consumer sensitivity to resource variability, covariance and phase. Ecol. Lett. 22, 1104-1114 (2019).
- Cusser, S., Helms, J. IV, Bahlai, C. A. & Haddad, N. M. How long do population level field experiments need to be? Utilising data from the 40-year-old LTER network. Ecol. Lett. 24, 1103-1111 (2021).
- EPA, U. Preventing Eutrophication: Scientific support for dual nutrient criteria. (2015).
- Westphal, K. et al. Multi-decadal trajectories of phosphorus loading, export, and instream retention along a catchment gradient. Sci. Total Environ. 667, 769-779 (2019).
- Kronvang, B. et al. Effects of policy measures implemented in Denmark on nitrogen pollution of the aquatic environment. Environ. Sci. Policy 11, 144-152 (2008).
- Hansen, A. T., Dolph, C. L., Foufoula-Georgiou, E. & Finlay, J. C. Contribution of wetlands to nitrate removal at the watershed scale. Nat. Geosci. 11, 127-132 (2018).
- Zak, D. et al. An Assessment of the multifunctionality of integrated buffer zones in Northwestern Europe. J. Environ. Qual. 48, 362-375 (2019).
- Filazzola, A. et al. A database of chlorophyll and water chemistry in freshwater lakes. Sci. Data 7, 310 (2020).
- R. Core Team. R: A Language and Environment for Statistical Computing. (R Foundation for Statistical Computing, 2023).
- QGIS Development Team. QGIS geographic information system. (2022).
- Soranno, P. A. et al. LAGOS-NE: A multi-scaled geospatial and temporal database of lake ecological context and water quality for thousands of US lakes. GigaScience 6, gix101 (2017).
- Isles, P. D. F. The misuse of ratios in ecological stoichiometry. Ecology 101, e03153 (2020).
- Ren, S. et al. Nonparametric bootstrapping for hierarchical data. J. Appl. Stat. 37, 1487-1498 (2010).
- Kruschke, J. Doing Bayesian Data Analysis: A Tutorial with R, JAGS, and Stan. (Academic Press, 2014).
- Davidson, T. A. et al. Bimodality and alternative equilibria do not help explain long-term patterns in shallow lake chlorophyll-a. Nat. Commun. 14, 398 (2023).
Acknowledgements
Author contributions
Funding
Competing interests
Additional information
supplementary material available at
https://doi.org/10.1038/s41467-024-45115-3.
© The Author(s) 2024
Department Aquatic Ecosystem Analysis, Helmholtz-Centre for Environmental Research – UFZ, Magdeburg, Germany. Chair of Hydrobiology & Fisheries, Estonian University of Life Sciences, Tartu, Estonia. Department Lake Research, Helmholtz-Centre for Environmental Research – UFZ, Magdeburg, Germany. Department of Ecoscience, and WATEC, C.F. Møllers Allé 3, Aarhus University, Aarhus, Denmark. Sino-Danish Education and Research Centre, Beijing, China. Limnology Laboratory, Department of Biological Sciences and Centre for Ecosystem Research and Implementation, Middle East Technical University, Ankara, Turkey. Institute of Marine Sciences, Middle East Technical University, Mersin, Turkey. Institute for Ecological and Pollution Control of Plateau Lakes, School of Ecology and Environmental Science, Yunnan University, Kunming, China. e-mail: daniel.graeber@ufz.de