إنتاج حمض الديوكسيكوليك بواسطة أنواع ميكروبية منخفضة الوفرة مرتبط باضطراب في استقلاب الجلوكوز Production of deoxycholic acid by low-abundant microbial species is associated with impaired glucose metabolism
يُقترح أن التغيرات في تركيبة ميكروبيوتا الأمعاء تساهم في الأمراض القلبية الأيضية، جزئيًا من خلال إنتاج جزيئات نشطة حيويًا. يتم إنتاج بعض المستقلبات بواسطة أنواع بكتيرية نادرة جدًا، والتي تم تجاهلها إلى حد كبير بسبب حدود الكشف. ومع ذلك، يمكن أن تصل تركيزات المستقلبات المنتجة ميكروبيًا من هذه الأنواع إلى مستويات عالية ولها تأثير كبير على فسيولوجيا المضيف. لاستكشاف هذا المفهوم، ركزنا على إنتاج الأحماض الصفراوية الثانوية بواسطةالبكتيريا المزالة للهيدروكسيل وأظهرت أن إضافة بكتيريا نادرة جداً إلى مجتمع يمكن أن تغير الناتج الأيضي بشكل كبير. نوضح أن Clostridium scindens تحول حمض الكوليك إلى حمض الديوكسيكوليك (DCA) بكفاءة عالية على الرغم من وفرة. إن مستوى C. scindens منخفض، لكنه لا يزال قابلاً للاكتشاف بواسطة PCR القطرات الرقمية. كما نوضح أن استعمار الفئران الإناث الخالية من الجراثيم بمجتمع يحتوي على C. scindens يحفز إنتاج DCA ويؤثر على استقلاب المضيف. أخيرًا، نوضح أن DCA يرتبط بضعف استقلاب الجلوكوز وسوء ملف الدهون لدى الأفراد المصابين بداء السكري من النوع 2، مما يوحي بأن هذه المسار الأيضي قد يساهم في تطور الأمراض القلبية الأيضية.
تزداد الأمراض القلبية الأيضية مثل السمنة ومرض السكري من النوع الثاني (T2D) على مستوى العالم وهناك حاجة ملحة لعلاجات جديدة. التغيرات في ميكروبيوم الأمعاء مرتبطة بالأمراض الأيضية.، لكن كيف تساهم هذه التغيرات في المرض لا يزال غير كامل فُهم. أحد التحديات الرئيسية في أبحاث الميكروبيوتا هو فهم كيفية عمل أنواع البكتيريا المختلفة في المجاميع لإنتاج جزيئات نشطة حيوياً. يتم إنتاج بعض المستقلبات الميكروبية بواسطة أنواع بكتيرية نادرة جداً، ومع ذلك تصل المستقلبات إلى مستويات عالية جداً.
تركيزات في الأمعاء والدورة الدموية حيث يمكن أن يكون لها تأثير كبير على فسيولوجيا المضيفمثال على هذه المستقلبات هو الأحماض الصفراوية الثانوية، التي تُعتبر من بين أكثر الجزيئات المشتقة من الميكروبات وفرة، على الرغم من أن عددًا قليلاً من الأنواع البكتيرية المعروفة فقط قادرة على إنتاجها. تعمل الأحماض الصفراوية كإشارات عبر مستقبلات النواة ومُستقبلات البروتين G المرتبطة بالمضيف، ويمكن أن تؤثر على العديد من المسارات الأيضية المختلفة.تتمتع أحماض الصفراء المختلفة بميول متفاوتة لمستقبلاتها؛ وبالتالي يمكن أن تعدل ميكروبات الأمعاء الإشارات عبر مستقبلات المضيف من خلال تغيير تركيبة أحماض الصفراء.تقوم ميكروبات الأمعاء بتعديل مجموعة الأحماض الصفراوية من خلال استقلاب الأحماض الصفراوية الأولية إلى أحماض صفراوية ثانوية عبر إزالة الهيدروكسيل، وإزالة الهيدروجين، والإيبيميريزيشن.أكثر الأحماض الصفراوية الثانوية انتشارًا في البشر هما حمض الديوكسيكوليك (DCA) وحمض الليثوكوليك (LCA)، اللذان يتم إنتاجهما بواسطةالبكتيريا المزالة للهيدروكسيل من حمض الكوليك (CA) وحمض الشينوديوكسيكوليك (CDCA)، على التواليميكروبيإزالة الهيدروكسيل هي عملية معقدة متعددة الخطوات، تم الكشف عنها بالكامل مؤخرًا وأظهرت أنها تتطلب إنزيمات مشفرة بواسطة جين الباي.عدد الأنواع البكتيرية التي تمتلك الجين الكامل bai-operon محدود وقد تم التعرف عليها في Eubacterium ومجموعة Clostridium XIVa وXI.. على الرغم من، تكون البكتيريا المزالة للهيدروكسيل عادةً موجودة بكميات منخفضة في أمعاء الإنسان، لكن إمكانياتها الأيضية عالية، وأكثر من 95% من مجموعة أحماض الصفراء في الأمعاء الغليظة تتكون من أحماض صفراء مزالة للهيدروكسيل.. علاوة على ذلك، بالإضافة إلى حمض الديوكسيكوليك وحمض الليثوكوليك، هناك مجموعة واسعة من المستقلبات الحمضية الصفراوية التي تنتجها الميكروبات مثل الأحماض الصفراوية الإيزو، والإيبي، والأوكزو، والتي قد تكون مهمة بيولوجياً ولكن لم يتم دراستها بشكل جيد.
في هذه الدراسة، نستكشف ما إذا كان إضافة نوع بكتيري ذو وفرة منخفضة إلى مجتمع ما يمكن أن يكون له تأثير كبير على الناتج الأيضي. اخترنا التركيز على الأحماض الصفراوية لأنها تشارك في العديد من العمليات الفسيولوجية وتعتبر الأحماض الصفراوية الثانوية من بين أكثر المستقلبات المشتقة من الميكروبات وفرة. قمنا بالتحقيق في إنتاج DCA بواسطة نوع بكتيري ذو وفرة منخفضة، Clostridium scindens، سواء في المختبر أو في الجسم الحي، وأظهرنا وجود صلة بين DCA وخلل في الأيض لدى المضيف.
النتائج
توليد مجتمع مبسط بقدرة على استقلاب أحماض الصفراء
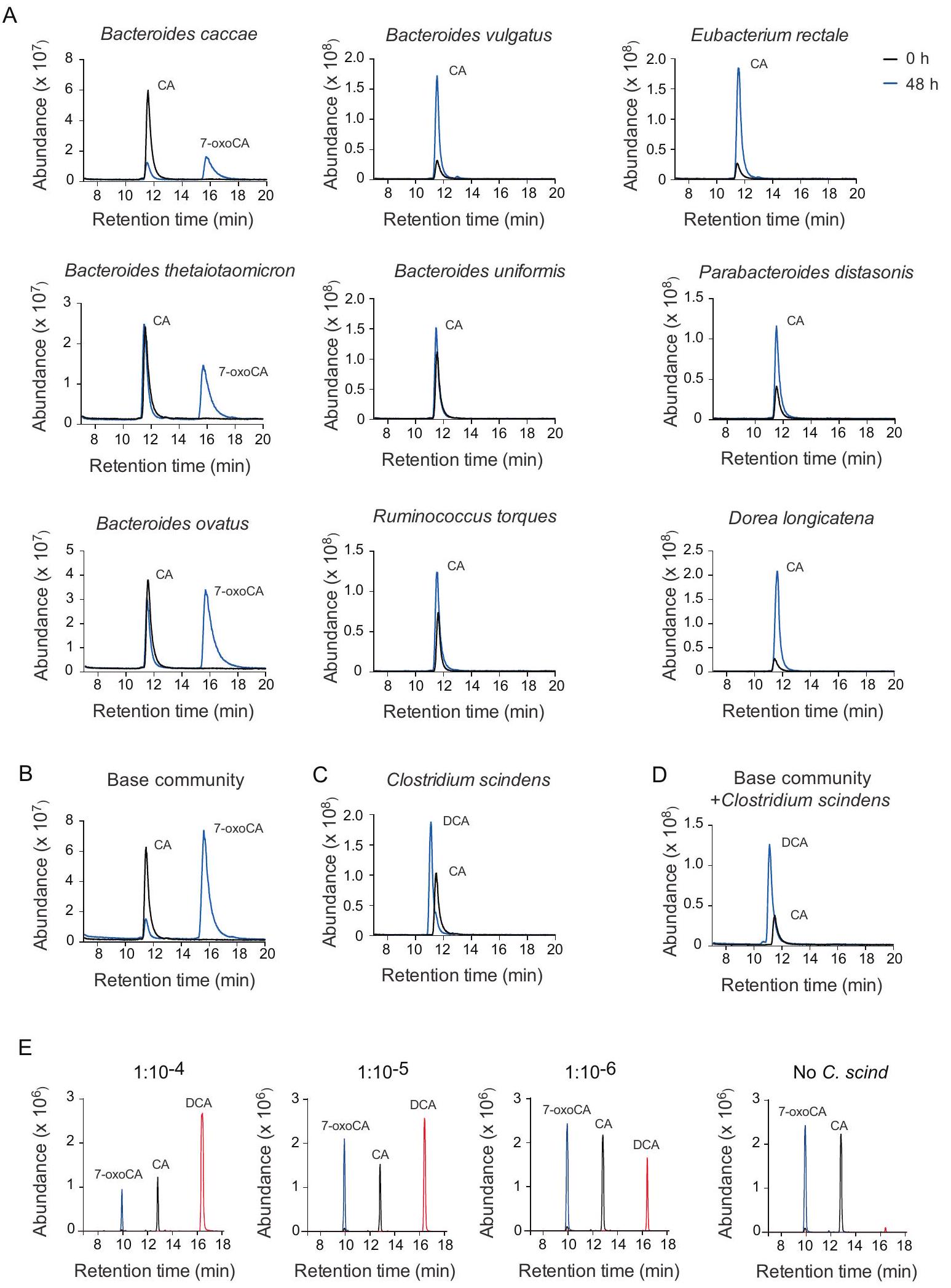
للتحقيق فيما إذا كان بإمكاننا إنتاج نظام لتقييم وظيفة البكتيريا ذات الوفرة المنخفضة، قمنا أولاً بإنشاء نظام في المختبر حيث…كانت الأنواع البكتيرية التي تقوم بإزالة الهيدروكسيل موجودة بكميات منخفضة، لدراسة تكوين DCA من CA. أولاً، قمنا بإنشاء مجتمع اصطناعي ‘أساسي’، استنادًا إلى الأعمال السابقة.يتكون من أنواع بكتيرية تفتقر إلى جينات الباي. تم زراعة البكتيريا التسع المختارة بشكل فردي في وجود حمض الكوليك لتقييم قدرتها على استقلاب الأحماض الصفراوية. أنتج ثلاثة منها (Bacteroides caccae وBacteroides thetaiotaomicron وBacteroides ovatus) 7-أوكسي حمض الكوليك، بينما لم ينتج أي من أعضاء المجتمع الأساسي حمض الديوكسيكوليك (الشكل 1A). وبالمثل، لم تنتج البكتيريا التسع حمض الديوكسيكوليك في المختبر عندما تم زراعتها معًا (الشكل 1B). على النقيض من ذلك، C. scindens، المعروف بـبكتيريا إزالة الهيدروكسيلتم تحويل CA إلى DCA سواء عند زراعتهما بمفردهما (الشكل 1C) أو مع المجتمع الأساسي (الشكل 1D).
أخيرًا، لمحاكاة الوضع في الأمعاء البشرية، قمنا بإجراء تخفيفات متسلسلة لتقليل وفرة. سكيندينس في المجتمع المبسط. لقياس مستوى . scindens في مجتمعاتنا المخففة استخدمنا تسلسل الميتاجينوم باستخدام تقنية تسلسل الشوتغن مع PCR القائم على القطرات الرقمية، مع مجموعات محددة من البرايمر-بروب. . scindens و B. caccae، عضو مجتمع ذو وفرة عالية تم اكتشافه في بيانات الميتاجينوم الشوتغن واستخدم كمرجع (الجدول التكميلي 1). ثم قمنا بتقييم قدرة C. scindens على إنتاج DCA في التخفيفات المختلفة ووجدنا أنه في التخفيفات التي تتراوح منإلى. يمكن أن تتحول scindens لا يزالمن CA إلى DCA في 24 ساعة، تم إثبات أنه في بيئة مجتمعية صناعية خاضعة للرقابة، يمكن أن يكون للكائن الحي ذو الوفرة المنخفضة إنتاجية أيضية عالية جداً (الشكل 1E).
إنتاج DCA من C. scindens ينظم الفسيولوجيا وعمليات الأيض للغلوكوز في الفئران المستعمرة
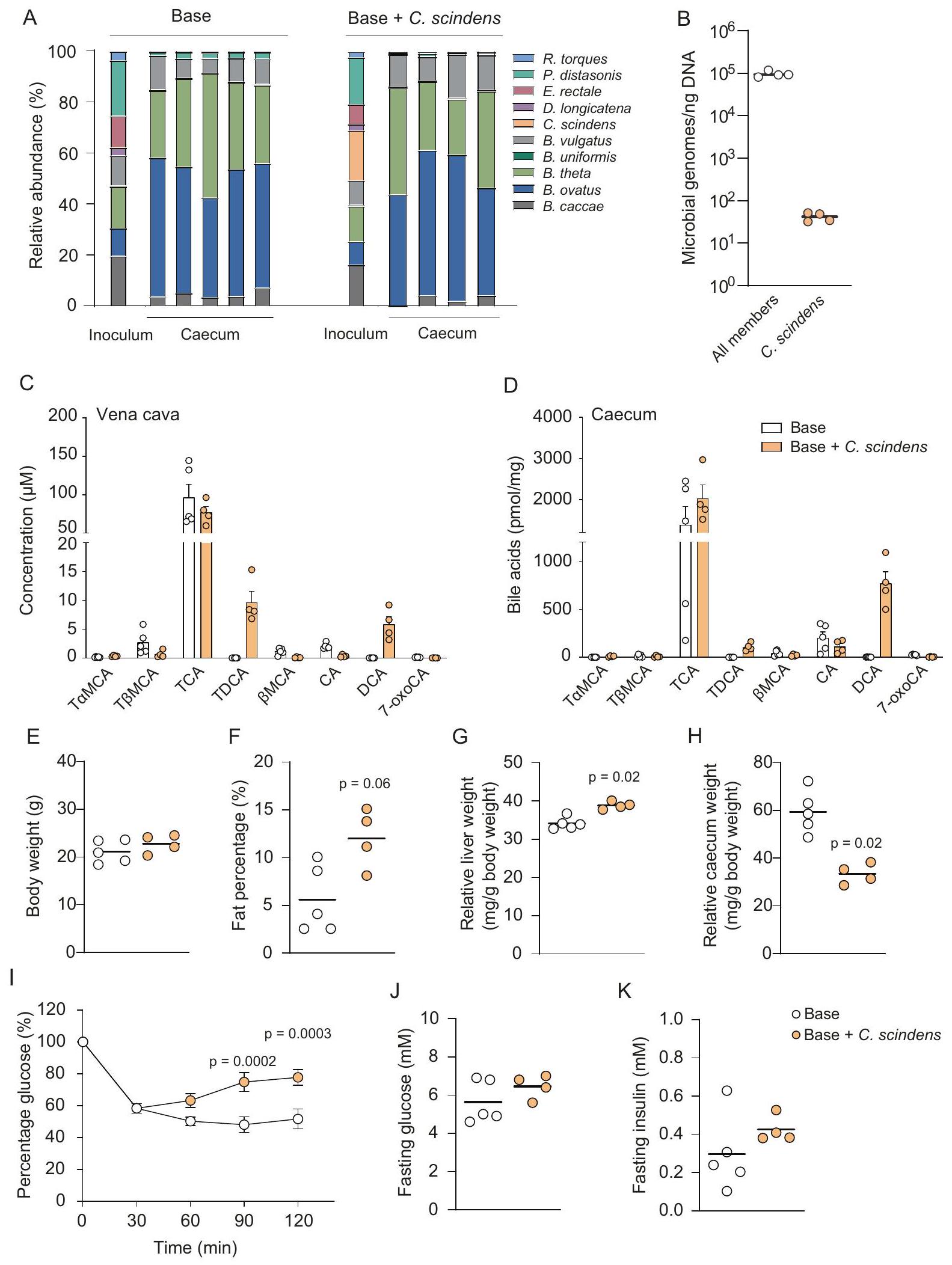
بعد ذلك، تحققنا مما إذا كانت قاعدة المجتمع. تم إنتاج DCA في الجسم الحي بواسطة scindens من خلال استعمار الفئران الخالية من الجراثيم لمدة أسبوعين. نظرًا لأن الفئران الخالية من الجراثيم لديها مستويات منخفضة من ، وهو سلف DCA، قمنا بإطعام الفئران نظامًا غذائيًا يحتوي علىعند الانتهاء، قمنا بتحليل تركيب الميكروبات في اللقاح وفي الأمعاء الغليظة من خلال إجراء تسلسل الميتاجينوم الشامل (الشكل 2A). لاحظنا أن ميكروبات الأمعاء الغليظة كانت مهيمنة بواسطة أنواع البكتيرويدس وأن الوفرة النسبية لـ. كان مستوى scindens أقل من حد الكمية القابلة للقياس. لذلك، قمنا بإجراء PCR بالقطرات الرقمية ووجدنا أن نسبة كانت نسخ جينوم C. scindens إلى إجمالي المجتمع 1:14,397 (الشكل 2B). من المهم أن C. scindens لم يتم اكتشافها في أي من الفئران التي استعمرت بالمجتمع الأساسي، مما يثبت أننا نستطيع إنشاء مجتمعات تختلف في نوع واحد قليل الوفرة في الجسم الحي.
ثم قمنا بتحليل الأحماض الصفراوية في مصل الدم من الوريد الأجوف ومحتوى القولون من الفئران المستعمرة بالمجتمع الأساسي وأظهرنا أن هذه الفئران يمكنها فك ارتباط حمض التوروموريتشوليك (. ) و تورو- CA (TCA) إلى و CA على التوالي، لكنهم لم يقوموا بتمثيلها بشكل أكبر إلى أحماض صفراوية ثانوية (الشكل 2C، D). على النقيض من ذلك، الفئران التي استعمرتها المجتمع الأساسي أظهرت scindens إنتاجًا كبيرًا من DCA على الرغم من انخفاض وفرتها. سكيندينس ( من جميع أعضاء المجتمع). تراوحت مستويات DCA منفي المصل وفي الأعور من الفئران المستعمرة مع المجتمع الأساسي + C. scindens، بينما كان DCA غير قابل للاكتشاف في مصل الفئران المستعمرة مع المجتمع الأساسي ولم يتم العثور إلا على آثار في الأعور (الشكل 2C، D). لمقارنة مستويات DCA في فئراننا المستعمرة مع المستويات الفسيولوجية، قمنا أيضًا بقياس DCA في مصل وأعور الفئران التي تم تربيتها تقليديًا على علف عادي أو نظام غذائي يحتوي على 1% CA. كانت مستويات DCA في مصل الفئران التي تم تربيتها تقليديًا على نظام غذائي يحتوي على CA مرتفعة بنفس القدر كما في الفئران المستعمرة مع المجتمع الأساسي.. ولكن مستويات DCA أعلى في الأعور (الشكل S1A). وبما يتماشى مع النتائج السابقة، أدى النظام الغذائي CA إلى ارتفاع مستويات السيروم من الألانين ترانسفيراز (ALT) والأسبارتات أمينوترانسفيراز (AST)، مما يشير إلى تأثير سام على الكبد لـ. ومع ذلك، لم تكن هناك اختلافات بين الفئران المستعمرة بالمجتمع الأساسي والمجتمع الأساسي + C. scindens (الشكل S1B). ومن الجدير بالذكر أن الفئران المستعمرة بـ C. scindens أظهرت انخفاضًا في التعبير الجيني الكبدي لجين Cyp7a1 بينما لم يكن هناك فرق ذو دلالة إحصائية في تعبير Cyp8b1 بين المجموعتين (الشكل S1C).
علاوة على ذلك، تم استعمار الفئران بمجتمع أساسيكان وزن جسم C. scindens مشابهًا لوزن الفئران المستعمرة بالمجتمع الأساسي (الشكل 2E). ومع ذلك، كانت الفئران المستعمرة بالمجتمع الأساسي + C. scindens تعاني من زيادة في الدهون (الشكل 2F) ووزن الكبد (الشكل 2G)، ولكن وزن الأمعاء الغليظة كان أقل (الشكل 2H)، مقارنة بالفئران المستعمرة بالمجتمع الأساسي. تماشيًا مع زيادة الدهون، كانت الدهون الثلاثية في الكبد أيضًا مرتفعة في الفئران المستعمرة بالمجتمع الأساسي + C. scindens وارتبطت بوزن الكبد النسبي (الشكل S1D). أظهرت تحليل الأحماض الدهنية قصيرة السلسلة في الأمعاء الغليظة مستويات أعلى من الزبدات في الفئران المستعمرة بالمجتمع الأساسي + C. scindens، والتي قد تكون بسبب زيادة تخمير الكربوهيدرات وقد تفسر جزئيًا الانخفاض في وزن الأمعاء الغليظة النسبي (الشكل S1E).
في نهاية التجربة، قمنا بإجراء اختبار تحمل الأنسولين داخل الصفاق ولاحظنا أن الفئران المستعمرة بمجتمع أساسي + C. scindens كانت تعاني من ضعف في تحمل الأنسولين، مما يشير إلى أن استقلاب الجلوكوز قد يتغير (الشكل 2I) على الرغم من عدم وجود اختلافات في مستوى الجلوكوز في الدم أثناء الصيام أو الأنسولين أثناء الصيام (الشكل 2J، K). مجتمعة، تظهر أنظمتنا في المختبر وفي الجسم الحي أن إضافة نوع بكتيري ذو وفرة منخفضة إلى مجتمع يمكن أن يكون له تأثير كبير على الناتج الأيضي الميكروبي، مما يمكن أن يغير بدوره استقلاب المضيف.
يتمتع DCA بزيادة في الأفراد المصابين بداء السكري من النوع الثاني ويرتبط بتدهور الخصائص الأيضية.
للتحقيق فيما إذا كانت التغيرات في مستويات DCA قد تكون مرتبطة بالأمراض الأيضية البشرية، اخترنا 100 فرد لديهم تحمل طبيعي للجلوكوز (NGT) و100 فرد من مرضى السكري من النوع الثاني غير المعالج (T2D) من مجموعتي IGT Microbiota وSCAPIS وقمنا بتحليل ملفات الأحماض الصفراوية لديهم. كانت مجموعتا NGT وT2D متطابقتين من حيث الجنس والعمر والوزن.
مؤشر كتلة الجسم، واستخدام الستاتينات، ومثبطات مضخة البروتون، ولم يكن أي من مجموعة السكري من النوع الثاني يتناول أدوية السكري (الجدول 1).
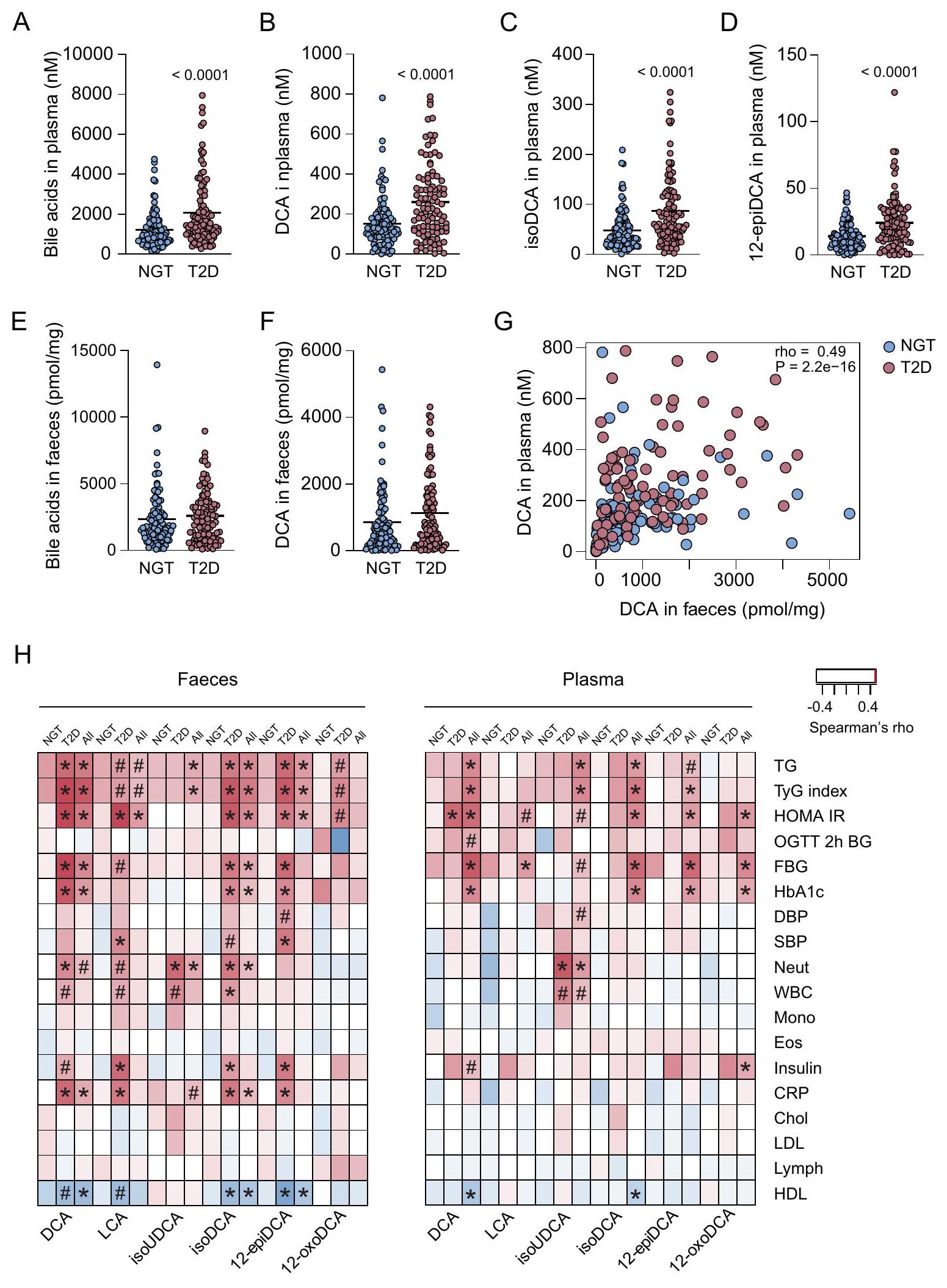
أظهر تحليل أحماض الصفراء في البلازما أن مستويات أحماض الصفراء الكلية كانت مرتفعة في الأفراد المصابين بداء السكري من النوع الثاني مقارنةً بالضوابط ذات مستويات الجلوكوز الطبيعية. في مجموعة السكري من النوع 2 مقابل في مجموعة NGT،; الشكل 3A والبيانات التكميلية 1). أظهر تحليل الأحماض الصفراوية الفردية أن الأحماض الصفراوية الثانوية الهيدروكسيليت 12، مثل،
الشكل 1 | التقييم في المختبر لتكوين DCA بواسطة الأنواع البكتيرية المدرجة في المجتمعات المبسطة. A-D تقييم تكوين DCA بعد 48 ساعة من الثقافة في وسط ميغا يحتوي علىCA بواسطة أعضاء المجتمع الأساسي Bacteroides caccae ATCC 43185، Bacteroides vulgatus ATCC 8482، Eubacterium rectale ATCC 33656، Bacteroides thetaiotaomicron VPI-5482، Bacteroides uniformis ATCC 8492، Parabacteroides distasonis ATCC 8503، Bacteroides ovatus ATCC 8483، Ruminococcus torques ATCC 27756 و Dorea longicatena DSM 13814 المزروعة بشكل فردي (A)، أعضاء المجتمع الأساسي المزروعون معًا (B)، Clostridium تم زراعة Clostridium scindens ATCC 35704 بشكل فردي (C) وفي مجتمع أساسي + Clostridium scindens معًا (D). E تقييم إنتاج DCA بعد 24 ساعة في تخفيفات متسلسلة من Clostridium scindens في المجتمع الأساسي المضاف إليه CA من محلول مخزون بتركيز 100 مليمول لتر إلى تركيز نهائي منتم قياس وفرة CA وDCA و7-oxoCA بواسطة GC-MS في (A-D) وبواسطة UPLC-Q-TOF-MS في (E). CA، حمض الكوليك؛ DCA، حمض الديوكسيكوليك؛ 7-oxoCA، حمض 7-أوكسيكوليك. الخط الأسود يشير إلى وفرة الأحماض الصفراوية عند 0 ساعة والخط الأزرق يشير إلى وفرة الأحماض الصفراوية عند; الخط الأزرق يشير إلى 7-أوكسيCA؛ الخط الأسود يشير إلى CA والخط الأحمر يشير إلى DCA (E).
كانت الأحماض الصفراوية DCA و isoDCA و 12-epiDCA هي الأكثر زيادة بشكل ملحوظ في الأفراد الذين يعانون من السكري من النوع 2 (الشكل 3B-D والشكل التوضيحي 2A) كما زادت 8 أحماض صفراوية إضافية أيضًا (الشكل التوضيحي 2A والبيانات التكميلية 1).
بعد ذلك، قمنا بتحليل تركيبة الأحماض الصفراوية في البراز ولكن لم نلاحظ أي اختلافات ملحوظة في مستويات الأحماض الصفراوية الكلية (الشكل 3E) أو في أي من أنواع الأحماض الصفراوية التي تم تحليلها (الشكل التوضيحي 2B والبيانات التكميلية 2). ومع ذلك، كانت DCA هي الحمض الصفراوي الأكثر وفرة وكانت تميل إلى الزيادة في براز الأفراد المصابين بداء السكري من النوع الثاني.قبل التعديل لاختبار متعدد) (الشكل 3F والبيانات التكميلية 2) ومرتبط بمستويات DCA في البلازما (الشكل 3G).
للتحقيق في العلاقات بين DCA والمعايير الفسيولوجية المتعلقة باضطراب استقلاب الجلوكوز، قمنا بربط المعايير السريرية (الجدول 1) مع الأحماض الصفراوية في البراز والبلازما. على الرغم من أننا لم نلاحظ اختلافات كبيرة في DCA البرازي بين مجموعتي NGT وT2D، إلا أننا لاحظنا علاقات كبيرة بين DCA البرازي والمعايير المرتبطة باضطراب استقلاب الجلوكوز مثل HOMA-IR، سكر الدم الصائم (FBG)، HbA1c، والأنسولين (الشكل 3H والبيانات التكميلية 2). بالإضافة إلى ذلك، ارتبط DCA إيجابيًا مع الدهون الثلاثية (TG) وسلبيًا مع البروتين الدهني عالي الكثافة (HDL)، مما يتماشى مع تدهور ملف الدهون. تم ملاحظة العلاقات عندما تم تحليل جميع الأفراد معًا وأيضًا عندما تم تحليل الأفراد المصابين بـ T2D بشكل منفصل (الشكل 3H). على النقيض من ذلك، لم نلاحظ أي علاقات كبيرة عندما تم تحليل مجموعة NGT بشكل منفصل. كما ارتبط DCA إيجابيًا مع بروتين C-reactive (CRP) وكريات الدم البيضاء (مثل العدلات) مما يشير إلى أنه مرتبط بالالتهاب. أظهر DCA البلازمي نمط ارتباط مشابه لـ DCA البرازي، وكانت أقوى علاقة موجودة مع HOMA-IR، سواء في المجموعة الكلية أو عندما تم تحليل مجموعة T2D بشكل منفصل (الشكل 3H والبيانات التكميلية 1). باختصار، يرتبط DCA، في كل من البراز والبلازما، باضطراب استقلاب الجلوكوز وتدهور ملف الدهون.
نقاش
في هذه الدراسة، أظهرنا أن نوعًا من البكتيريا ذو وفرة منخفضة، قد يتم تجاهله بسبب قيود الكشف، يمكن أن يكون له تأثير كبير على إنتاج المستقلبات الحيوية النشطة.
أظهرنا أن C. scindens، على الرغم من وجوده بكميات منخفضة، يمكن أن ينتج كميات كبيرة من DCA ويؤثر على تحمل الأنسولين في الفئران المستعمرة، مما يشير إلى أن DCA قد يساهم في ضعف استقلاب الجلوكوز. علاوة على ذلك، أظهرنا أن DCA، في البلازما والبراز، يرتبط بضعف استقلاب الجلوكوز وسوء ملف الدهون في الأفراد المصابين بداء السكري من النوع 2. تسلط نتائجنا الضوء على أن الأنواع البكتيرية ذات الوفرة المنخفضة قد يكون لها تأثيرات عميقة على مستويات المستقلبات في الأمعاء والدورة الدموية ويجب أن تحظى بمزيد من الاهتمام. إحدى التحديات في دراسة هذه الكائنات في الميتاجينومات هي أنها قد تكون قريبة من حد الكشف وبالتالي تفتقر التحليلات القوية. وبناءً عليه، يمكن توسيع هذا المفهوم ليشمل مستقلبات أخرى تنتجها الأنواع ذات الوفرة المنخفضة ويؤكد على أهمية دراسة مستقلبات الميكروبات المعوية في الدورة الدموية للكائنات المضيفة. تحدٍ آخر هو أن تحويل الأحماض الصفراوية معقد للغاية وأن مجموعة متنوعة من العوامل البيئية، مثل الأدوية والنظام الغذائي ودرجة الحموضة والجو الغازي ووجود العوامل المساعدة الإنزيمية يمكن أن تؤثر على التفاعلات بين الأحماض الصفراوية والبكتيريا في الأمعاء.. هذه المعلمات تعتمد على التداخل بين يمكن أن تختلف البكتيريا المختلفة في مجتمع ما ووظيفة نوع بكتيري محدد عندما يتم زراعته بمفرده مقارنةً عندما يتم زراعته في تجمع مع بكتيريا أخرى. يمكن أن تكون هناك أيضًا اختلافات في تحويل الأحماض الصفراوية بواسطة البكتيريا في المختبر وفي الجسم الحي.التي تؤكد على أهمية الدراسات الشاملة حول بكتيريا استقلاب الأحماض الصفراوية.
تظهر نتائجنا بوضوح أن الوفرة المنخفضة جداً لـالبكتيريا التي تزيل الهيدروكسيل كافية لإنتاج كميات كبيرة من DCA وهذا يدعم الفكرة أنقد تكون إزالة الهيدروكسيل حصرية لعدد قليل جداً من البكتيريا في أمعاء الإنسان. ومع ذلك، كانت مستويات DCA في الأعور أقل من المستويات في الفئران التي تربى تقليدياً، والتي تحتوي على ميكروبيوتا كاملة، ولا يمكننا استبعاد وجود أنواع بكتيرية أخرى غير محددة في أمعاء الإنسان تساهم في تكوين DCA، مثل Clostridium hylemonae.وما زالت هناك حاجة إلى مزيد من الدراسات لتوفير رؤية موسعة لبكتيريا استقلاب الأحماض الصفراوية. قد تكون هناك أيضًا تأثيرات تآزرية بين C. scindens وبكتيريا أخرى في الأمعاء، مقارنةً بما أظهرناه سابقًا بين Faecalibacterium prausnitzii وDesulfovibrio piger.. في تلك الدراسة، تم استعمار الفئران بشكل مشترك مع زادت بكتيريا البيجر من وفرة وقدرة الأيض لـ. براوسنيتسيا وسيكون من المهم توضيح آليات مماثلة لـ. سكندنس وبكتيريا أخرى تنتج DCA.
لقد أثارت أحماض الصفراء اهتمامًا كبيرًا كجزيئات إشارة على مدى السنوات الماضية، وهنا نوضح أن تحمل الأنسولين يتأثر في الفئران المستعمرة بـ C. scindens المنتجة لـ DCA. وهذا يتماشى مع دراسة سابقة حيث أظهر زابورسكا وآخرون أن مكملات DCA في الفئران تؤدي إلى ضعف في توازن الجلوكوز (كلا من اختبارات تحمل الأنسولين والجلوكوز) وزيادة في إجهاد الشبكة الإندوبلازمية الكبدية وانخفاض في إشارة الأنسولين الكبدية.في دراسة أخرى، أظهر يوشيموتو وزملاؤه أن إنتاج DCA قد زاد في الفئران التي تعاني من السمنة الناتجة عن النظام الغذائي أو الوراثة، وأن DCA ساهم في تطور السرطان في الفئران السمنة.على عكس دراسة يوشيموتو، استخدمنا نظام غذائي عادي مدعوم بـ CA ولم تكن فئراننا بدينة. ومع ذلك، وجدنا زيادة في الدهون ودهون الكبد في فئراننا المنتجة لـ DCA، مما يدعم وجود صلة بين DCA وتراكم الدهون على الرغم من أن الآليات قد تكون مختلفة.
تم الإشارة أيضًا إلى أن تخليق أحماض الصفراء مرتبط بالتغيرات الأيضية وقد لاحظنا انخفاضًا في تعبير Cyp7a1 في كبد الفئران التي استعمرت بمجتمع أساسي.. scindens ولكن لا يوجد فرق في تعبير Cyp8b1. ومع ذلك، فإن مساهمة الأحماض الصفراوية التي تم تصنيعها داخليًا في نموذج الفأر لدينا ربما تكون ضئيلة نظرًا لأن الفئران تم إعطاؤها جرعة عالية جدًا من CA الخارجي من خلال النظام الغذائي.
لقد أظهرنا أيضًا أن مستويات DCA مرتفعة في بلازما الأفراد المصابين بداء السكري من النوع الثاني وأن التغير في ملف الأحماض الصفراوية ليس نتيجة لأدوية السكري، حيث إن جميع الأفراد المصابين بداء السكري من النوع الثاني تم تشخيصهم حديثًا ولم يتلقوا علاجًا لداء السكري. من غير الواضح حاليًا لماذا كانت تركيزات DCA في البراز غير مختلفة بشكل ملحوظ، ولكن أحد التفسيرات المحتملة يمكن أن يكون أننا حصلنا فقط على عينات عشوائية بدلاً منعينات، أو أن هناك اختلافات في امتصاص أحماض الصفراء.
أظهرت الدراسات السابقة نتائج متضاربة فيما يتعلق بملفات أحماض الصفراء في البلازما وعمليات الأيض للغلوكوز.أظهر هاوسلر وآخرون أن زيادة نسبةلاأحماض الصفراء مرتبطة بمقاومة الأنسولينبينما وجدت دراسة أخرى عدم وجود علاقة بين نسبة و HOMA-IR أو خطر ظهور مرض السكري الجديدقد يكون أحد الأسباب المحتملة لهذه الفجوة هو وجود اختلافات في التحويل الميكروبي داخلأنواع أحماض الصفراء، أي تحويل حمض الكوليك إلى حمض الديوكسيكوليك و/أو iso-DCA و12epiDCA، ومن ثملاقد تكون النسبة مقياسًا بدائيًا وتحتاج إلى مزيد من التحليل. وجدنا علاقات قوية. بين DCA والمعايير المتعلقة بتمثيل الجلوكوز والدهون ونتائجنا السريرية تشير إلى أن DCA قد يكون علامة حيوية للالتهاب وأمراض القلب الأيضية. ومع ذلك، هناك حاجة إلى مزيد من الدراسات لتحديد العلاقة السببية بين DCA وأمراض التمثيل الغذائي وقد تكشف الدراسات المستقبلية ما إذا كان يمكن استخدام DCA لتحديد الأفراد المعرضين لخطر تطوير السكري من النوع الثاني.
الشكل 2 | استعمار الفئران الخالية من الجراثيم بمجموعات بكتيرية. أ وفرة نسبية للبكتيريا في اللقاح ومحتويات الأعور للفئران المستعمرة بمجتمع أساسي أو مجتمع أساسي + C. scindens تم تقييمها بواسطة تسلسل الميتاجينوم بالأسلوب العشوائي. ب نسخ الجينوم الميكروبي لكل نانوجرام من الحمض النووي البكتيري في محتوى الأعور للفئران المستعمرة بمجتمع أساسي.. تم تقييم C. scindens بواسطة PCR القطرات الرقمية. تم قياس أعضاء المجتمع الأساسي باستخدام مجموعات البرايمر-بروب لجين tilS، وتم قياس C. scindens باستخدام مجموعات البرايمر-بروب لجين baiCD (انظر الجدول التكميلي 1). ج، د مستويات الأحماض الصفراوية في الوريد الأجوف (ج) والقولون (د) للفئران المستعمرة بالمجتمع الأساسي أو المجتمع الأساسي + C. scindens تم قياسها بواسطة UPLC-MS/MS. هـ-ح المعايير الفسيولوجية للفئران بما في ذلك وزن الجسم (هـ)، السمنة (و)، الوزن النسبي للكبد ( ) ووزن الأعور النسبي ( تم قياسه بعد أسبوعين من الاستعمار. تم إجراء اختبار تحمل الأنسولين عن طريق حقن الأنسولين داخل الصفاق. الجلوكوز تم قياسه عندبعد الحقن، يتم تقديم البيانات كنسبة مئوية من الجلوكوز مقارنةً بنقطة الوقت عند 0 دقيقة.جلوكوز الدم الصائم ) والأنسولين ( تم قياسها عند نقطة الوقت 0 دقيقة.عينات لمجموعة المجتمع الأساسية (المشار إليها باللون الأبيض في B-K) وعينات لمجتمع القاعدةمجموعة scindens (المشار إليها باللون البرتقالي في B-K)؛-القيم تشير إلى الفروقات بين الفئران المستعمرة بالمجتمع الأساسي والمجتمع الأساسي + C. scindens التي تم تحليلها باستخدام اختبار مان-ويتني ذو الذيلين (E-H و J، K) أو تحليل التباين الثنائي مع اختبار المقارنات المتعددة Šidák (I). تشير أشرطة الخطأ إلى الخطأ المعياري للمتوسط (C، D و I). CA حمض الكوليك، DCA حمض الديوكسيكوليك، ßMCA حمض البيتا-موريتشوليك، TCA حمض التاوروكوليك، TDCA حمض التاوروديوكسيكوليك، Tحمض الموريكوليك ألفا-تاورو-إم سي إيه، Tحمض الموريتشوليك التوروي، 7-أوكسيCA حمض 7-أوكوشوليك.
طرق
سلالات البكتيريا وظروف الزراعة
تم زراعة سلالات المجتمع الاصطناعي في وسط ميغا المخفض مسبقًاأو على EGفيفي غرفة لاهوائية (مختبرات كوي، غراست ليك، ميشيغان، الولايات المتحدة الأمريكية) مع جو منالهيدروجين و تضمنت سلالات المجتمع الاصطناعي بكتيرويدس يونيورميس ATCC 8492، بكتيرويدس فولغاتوس ATCC 8482، بكتيرويدس ثيتايوتاوميكرون VPI-5482، بكتيرويدس كاكا ATCC 43185، بكتيرويدس أوفاتوس ATCC 8483، بارابكتيرويدس ديستاسونيس ATCC 8503، يوبكتيريوم ركتالي ATCC 33656، دوريا لونغيكاتينا DSM 13814، رومي نوكوكوس توركيس ATCC 27756، وكلوستريديوم سكندنس ATCC 35704. تم الحصول على جميع السلالات من مجموعة الثقافة الأمريكية (ATCC)، باستثناء D. لونغيكاتينا DSM 13814، التي تم الحصول عليها من معهد لايبنيز DSMZ (براونشفايغ، ألمانيا).
تجارب الحيوانات
تم الحفاظ على إناث فئران C57BI/6 خالية من الجراثيم في عوازل جنتوبيوتيك مرنة من البلاستيك تحت دورة ضوء صارمة مدتها 12 ساعة (الضوء من الساعة 7 صباحًا حتى 7 مساءً)،درجة الحرارة، رطوبة الهواءتحت ظروف خالية من الجراثيم القياسية، وتم تغذيتها بنظام غذائي مطبوخ بالبخار (طعام مختبر، سانت لويس، ميزوري؛ #5021) مكمل بنسبة 1% من CA (أنفيغو تيكلاد دايتس، ماديسون، ويسكونسن) بحرية. كانت العوازل الخالية من الجراثيم تُختبر بانتظام للتأكد من التعقيم من خلال زراعة وتحليل PCR للبراز الذي يعزز جين 16S rRNA.
تم استعمار الفئران الخالية من الجراثيم بعد صيام لمدة 4 ساعات عن طريق التغذية القسرية بمقدار 0.2 مل من المجتمعات المزروعة (المجتمع الأساسي أو المجتمع الأساسي بما في ذلك. scindens). قبل الاستعمار، تم وزن جميع الفئران وتوزيعها عشوائيًا إلى مجموعتين متساويتين في الوزن. تم إذابة مجتمعات البكتيريا وإعطاؤها للفئران الخالية من الجراثيم خلال ساعتين بعد الإذابة. تم تكرار الاستعمار بعد يومين. تم استعمار الفئران في عمر 17 أسبوعًا وتم الحفاظ عليها لمدة أسبوعين بعد الاستعمار. تم مراقبة تركيبة الميكروبات المعوية بعد الاستعمار من خلال التسلسل وPCR القطرات الرقمية (ddPCR) كما هو موضح أدناه.
تم إجراء اختبار تحمل الأنسولين عن طريق حقن الأنسولين كجم من وزن الجسم عن طريق الحقن داخل الصفاق بعد صيام لمدة 4 ساعات. تم جمع عينات دم من الذيل في ، وتم تحديد مستويات الجلوكوز في الدم بعد 120 دقيقة باستخدام جهاز قياس الجلوكوز (كونتور نكست، أسينسيا لرعاية السكري، ستوكهولم، السويد). تم تحديد تكوين الجسم باستخدام التصوير بالرنين المغناطيسي (EchoMRI، هيوستن، تكساس) بعد اختبار تحمل الأنسولين وتم جمع الدم من الوريد الأجوف السفلي تحت تخدير عميق باستخدام الإيزوفلوران. ثم تم قتل الفئران، وجمع المصل والأنسجة. تم تسجيل أوزان الجسم والكبد والقولون، وتم تجميد الأنسجة على الفور في النيتروجين السائل وتخزينها في حتى يتم التحليل بشكل أكبر.
في تجربة منفصلة، تم الحفاظ على إناث الفئران من سلالة C57BI/6 التي تم تربيتها تقليديًا تحت دورة ضوء صارمة مدتها 12 ساعة (الضوء من الساعة 7 صباحًا حتى 7 مساءً)،درجة الحرارة، رطوبة الهواءتحت ظروف خالية من مسببات الأمراض المحددة القياسية، وتم تغذيتها إما بنظام غذائي عادي (حمية المختبر، سانت لويس، ميزوري) أو نظام غذائي مكمل بنسبة 1% من CA (حمية إنفيغو تيكلاد، ماديسون، ويسكونسن) بحرية لمدة أسبوعين. ثم تم قتل الفئران، وجُمعت المصل والقولون للتحليلات. جميع الحيوانات تم إجراء التجارب في الطب الحيوي التجريبي، جامعة غوتنبرغ، باستخدام بروتوكولات معتمدة من لجنة أخلاقيات البحث على الحيوانات في غوتنبرغ (رقم الأخلاقيات 4805/23).
مجموعات بشرية
تم اختيار 200 فرد (100 مصاب بداء السكري من النوع الثاني غير المعالج و100 من ضوابط متطابقة في مؤشر كتلة الجسم مع تحمل جلوكوز طبيعي) من مجموعة ميكروبيوتا IGT (45 مصاب بداء السكري من النوع الثاني و45 تحمل جلوكوز طبيعي)، ومجموعة دراسة التصوير الحيوي القلبي الرئوي السويدية (SCAPIS) – مجموعة غوتنبرغ (55 مصاب بداء السكري من النوع الثاني و55 تحمل جلوكوز طبيعي). تم وصف تصميم المجموعتين البشريتين بالتفصيل في مكان آخر.تتكون المجموعتان من رجال ونساء تتراوح أعمارهم بين 50 و64 عامًا من منطقة غوتنبرغ في السويد، الذين تم اختيارهم عشوائيًا من سجل التعداد. كانت معايير الاستبعاد تشمل مرض السكري المعروف؛ الأمراض الالتهابية، مثل مرض كرون، والتهاب القولون التقرحي، والأمراض الروماتيزمية؛ العلاج بالستيرويدات أو الأدوية المناعية؛ السرطان (ما لم يكن خاليًا من الانتكاسات خلال السنوات الخمس السابقة)؛ ضعف الإدراك؛ والعلاج من الأمراض المعدية واستخدام المضادات الحيوية في الأشهر الثلاثة الماضية. وبناءً عليه، لم يكن أي من الضوابط المتطابقة في العمر والجنس ومؤشر كتلة الجسم يتناولون إما الميتفورمين أو أدوية تقليل الوزن مثل ناهضات GLP1. كما تم استبعاد الأفراد إذا لم يفهموا اللغة السويدية وإذا وُلِدوا خارج السويد. قدم جميع المشاركين موافقة مستنيرة، وتمت الموافقة على الدراسة من قبل لجنة مراجعة الأخلاقيات في غوتنبرغ (560-13) وفي أوميا 2010-228-31M (2012-285-32M و2014-33-32).
قمنا بجمع عينات البلازما والبراز من 200 فرد وتم تقييم المعايير الأيضية. تم تقييم أحماض الصفراء في البلازما والبراز.
استخراج الحمض النووي وتسلسل الثقافات البكتيرية وعينات الأعور من الفئران
تم استخراج الحمض النووي الكلي من محتويات أمعاء الفئران وكرات ثقافة البكتيريا باستخدام مجموعة عزل الحمض النووي PowerFecal (موبيو، كارلسباد، كاليفورنيا، الولايات المتحدة الأمريكية)، وتم قياسه باستخدام مجموعة اختبار Qubit dsDNA BR ومقياس الفلورة Qubit من إنفيتروجن (ثيرمو فيشر، وولثام، ماساتشوستس، الولايات المتحدة الأمريكية). تم إنشاء مكتبات التسلسل باستخدام مجموعة تحضير مكتبة الحمض النووي Nextera XT ومجموعة الفهرسة v2 (إيلومينا، هايوارد، كاليفورنيا، الولايات المتحدة الأمريكية) وتم تسلسلها على جهاز إيلومينا MiSeq مع مجموعة كواشف MiSeq v2 (300 دورة) (إيلومينا، هايوارد، كاليفورنيا، الولايات المتحدة الأمريكية). تم تعيين قراءات التسلسل إلى قاعدة بيانات مرجعية تم بناؤها باستخدام جينومات سلالات البكتيريا الموجودة في الدراسة فقط باستخدام Bowtie2 2.2.9. وتم قياس الوفرة النسبية لأعضاء المجتمع البكتيري باستخدام SAMtools 1.3.1 .
تقدير ddPCR
تم تصميم مجموعات البرايمر/البروبي IDT Taqman ddPCR لجين baiCD (الجين المستحث بواسطة الأحماض الصفراوية) لـ. scindens ATCC 35704، والجين الأساسي ذو النسخة الواحدة tilS (تيرنايلي-لايسيدين سينثاز) لـ B. caccae ATCC 43185 لضمان احتساب كل جينوم مرة واحدة فقط. باختصار، تفاعل يتضمنماءبيوراد ddPCR سوبرميكس للاختبارات (بدون dUTP) [رقم: 186-3023] NEB HindIII-HF، مجموعة بادئات/مجسات Taqman 20X في محلول IDTEتم استخدام الحمض النووي المدخل لكل عينة. تم توليد القطرات باستخدام QX200
الجدول 1 | خصائص المشاركين في الدراسة في مجموعتي NGT و T2D
خصائص المشاركين في الدراسة
NGT
داء السكري من النوع الثاني
-قيمة
جنس
44 (% إناث)
42 (% إناث)
نس
العمر (بالسنوات)
نس
الوزن (كجم)
نس
الطول (سم)
نس
مؤشر كتلة الجسم (BMI) )
نس
الخصر (سم)
0.01
الورك (سم)
نس
أين
ضغط الدم الانقباضي (مم زئبق)
نس
ضغط الدم الانبساطي (مم زئبق)
نس
الهيموغلوبين (غ/ل)
0.05
FBG (ملليمول/لتر)
OGTT 2 ساعة BG (مللي مول/لتر)
الأنسولين (بيكومول/لتر)
0.02
الهيموغلوبين A1c (%)
HOMA_IR
مؤشر TyG
TG (ملليمول/لتر)
إجمالي الكوليسترول (ملليمول/لتر)
نس
LDL (مللي مول/لتر)
نس
HDL (مللي مول/لتر)
CRP (ملغ/ل)
0.001
WBC ( )
0.04
نيوت ( )
0.02
مونو ( )
نس
EOS ( )
نس
اللمف )
نس
ستاتين
4 (% من الكل)
12 (% من الكل)
نس
مؤشر أسعار المنتجين
53 (% من الكل)
54 (% من الكل)
نس
تُعرض البيانات كمتوسطSD، الـلكل معلمة يتم الإشارة إليها داخل الأقواس، يتم تقديم الجنس كنسبة مئوية للإناث في المجموعة، ويتم تقديم الستاتينات ومثبطات مضخة البروتون كنسبة مئوية للمستخدمين في المجموعة.-القيم تشير إلى الفروقات بين مجموعة NGT ومجموعة T2D التي تم تحليلها باستخدام اختبار مان-ويتني ثنائي الاتجاه مع تعديل بنجاميني وهوشبرغ-القيم. كول كوليسترول، CRP بروتين سي التفاعلي، DBP ضغط الدم الانبساطي، إيوس الإيوزينيات، FBG سكر الدم الصائم، HbA1c الهيموغلوبين A1c، HDL البروتين الدهني عالي الكثافة، HOMA-IR نموذج التقييم المنزلي لمقاومة الأنسولين، LDL البروتين الدهني منخفض الكثافة، لمف اللمفاويات، مونو الوحيدات، NGT تحمل الجلوكوز الطبيعي، نيوت العدلات، OGTT سكر الدم بعد ساعتين من اختبار تحمل الجلوكوز الفموي، PPI مثبط مضخة البروتون، SBP ضغط الدم الانقباضي، T2D داء السكري من النوع 2، TG الدهون الثلاثية، TyG مؤشر الدهون الثلاثية والجلوكوز، WBC كريات الدم البيضاء، whr نسبة الخصر إلى الورك.
نظام PCR الرقمي القائم على القطرات (بايو راد) وتمت عملية التضخيم عن طريق الحضانة عند لمدة 5 دقائق تليها 40 دورة من لمدة 30 ثانية، لمدة دقيقة واحدة، ثم الثبات عندلمدة 10 دقائق لتعطيل الإنزيم. معدل الزيادة لـتم استخدامه في الثانية. تم قراءة التفاعلات باستخدام جهاز قراءة القطرات QX200 (بايو راد) وتحليلها باستخدام برنامج QuantaSoft الإصدار 1.6.
أيض أحماض الصفراء في الثقافة المشتركة
تم جمع الأنواع الأحادية من أعضاء المجتمع البكتيري التي نمت طوال الليل عن طريق الطرد المركزي اللاهوائي وإعادة تعليقها في وسط ميغا جديد مُخفض مسبقًا مع CA (سيغما-ألدريتش، سانت لويس، ميزوري، الولايات المتحدة الأمريكية) وسمح له بالت incubate في لمدة 48 ساعة لتحفيز استقلاب أحماض الصفراء. تم جمع هذه الثقافات عن طريق الطرد المركزي اللاهوائي، وتم غسل الكريات بالـ PBS المخفض مسبقًا وإعادة تعليقها بكثافة (نصف حجم الثقافة الأصلية) في محلول جديد مخفض مسبقًا.
ميغا ميديا. تم دمج ثقافات البكتيرويديتس والفيرميكوتيس، باستثناء C. scindens.حسب الحجم وتم تقسيمه إلى أجزاء. تم تخفيف C. scindens بالتسلسل بنسبة 1:10 إلى أجزاء من المجتمع الأساسي المشترك وتم إضافة CA إلى كل ثقافة مشتركة من محلول مخزون DMSO بتركيز 100 مللي مول لتر إلى تركيز نهائي منتم أخذ عينات من الثقافات لتحليل الأحماض الصفراوية والحمض النووي بعد 24 ساعة من ذروة CA. لتحليل الأحماض الصفراوية، تم أخذ 2 مل من الثقافة وتم تحميضها على الفور بـ6 ن من حمض الهيدروكلوريك لإيقاف جميع الأنشطة الأيضية وبروتنة أحماض الصفراء لجعلها أكثر قابلية للذوبان في المذيب العضوي. لتحليل الحمض النووي، تم أخذ عينة 1 مل من الثقافة، وتم ترسيبها، وتخزينها فيحتى الاستخراج والتسلسل.
تحليل أحماض الصفراء لثقافات البكتيريا بواسطة الكروماتوغرافيا الغازية-مطياف الكتلة (GC-MS)
تم تطبيق هذه الطريقة على التجارب المعروضة في الشكل 1A-D. تم استخراج الثقافات البكتيرية الحمضية مرتين بحجوم متساوية من الإيثيل أسيتات، وتم تجميع المستخلصات العضوية، وتم تمريرها عبر سدادة من، وتم التركيز حتى الجفاف. تم تنقية المتبقي باستخدام عمود بيبت سيليكا بارتفاع 1.5 سم. تم غسل العمود بـ 2 مل من الميثانول في ثنائي كلورو ميثان، تم إزالته باستخدام 6 مل منالميثانول في الأسيتات الإيثيلي، وتم تركيز المحلول إلى جفاف. قبل تحليل GC-MS، تم تحويل الأحماض الصفراوية المنقاة إلى مشتقات إستر الميثيل – إثير ثلاثي ميثيل سيلوكسي كما يلي. تم إعادة تعليق الأحماض الصفراوية فيتم إضافة الميثانول ومحلول 2 م من (تريميثيلسيلي)ديزوميثان في الإيثر ثنائي الإيثيل بالتنقيط حتى استمر اللون الأصفر. تم تركيز المحلول حتى الجفاف وتم إضافة محلول بنسبة 3:1 من N,O-bistrimethylsilyl) trifluoroacetamide (BSTFA) و chlorotrimethylsilane. تم إجراء تحليل GC-MS علىللمحلول الناتج. تم تحديد أحماض الصفراء بناءً إما على أوقات الاحتفاظ وأنماط تفتت الكتلة للمعايير أو على أنماط تفتت الكتلة المنشورة..
تحليل أحماض الصفراء لثقافات البكتيريا بواسطة الكروماتوغرافيا السائلة عالية الأداء (UPLC) – مطيافية الكتلة رباعية القطب بتقنية الزمن الطائر (Q-TOF-MS)
تم تعديل هذه الطريقة من طريقة تم تطويرها بواسطة جاستن ر. كروس وتم تطبيقه على التجارب المعروضة في الشكل 1E. باختصار، تم استخراج الثقافات البكتيرية الحمضية مرتين بحجوم متساوية من الإيثيل أسيتات، وتم تجميع المستخلصات العضوية، وتم تمريرها عبر سدادة من ، تم تركيزه حتى الجفاف وإعادة تكوينه فيالميثانول في الماء. تم فصل المركبات على جهاز Agilent 1290 Infinity II UPLC باستخدام عمود Kinetex C18، فينومينيكس، رقم الكاتالوج #00D-4475-AN) وتم الكشف عنه باستخدام جهاز Agilent 6530 Q-TOF المزود بمصدر ESI مزدوج الرذاذ في وضع التأين السالب، كانت درجة حرارة الغاز الغلافتم تحقيق الفصل الكروماتوغرافي بمعدل تدفق قدرهمعتدرج )، حيث كانت المرحلة المتنقلة A هي الماء مع حمض الفورميك ومرحلة التحرك ب كانت الأسيتون معحمض الفورميك.
تحليل أحماض الصفراء من عينات من الفئران والبشر بواسطة UPLC-MS/MS
تم تطبيق هذه الطريقة على التجارب المعروضة في الشكل 2C، D، الشكل 3A-G، الشكل التكميلي 1A، B والبيانات التكملية. تم تحليل أحماض الصفراء في مصل الدم والأمعاء الغليظة من الفئران، والبلازما والبراز من البشر، باستخدام UPLC-MS/MS وتم تقديرها باستخدام مزيج من المعايير غير الموسومة والمعايير الداخلية الموسومة بالديتيريوم.مصل ومن الأعور من الفئران، وبلازما وتم استخدام براز المشاركين البشريين في التحليلات.
تم استخراج عينات المصل والبلازما باستخدام 10 أضعاف من الميثانول المحتوي على d4-TCA و d4-GCA و d4-GCDCA و d4-GUDCA و d4-GLCA و d4-UDCA و d4-CDCA و d4-LCA (50 نانومول لكل منها). بعد 10 دقائق من دوامة و 10 دقائق من الطرد المركزي فيتم تبخير السائل العلوي باستخدام تيار من النيتروجين وإعادة تكوينه في (لسيروم الفأر) (للبلازما البشرية) ميثانول:ماء [1:1]. من العينات تم استخدامها لتحليل الأحماض الصفراوية.
تم استخراج عينات السائل المعوي والبراز باستخدام الميثانول، الذي يحتوي على نفس المعايير المعلمة بالديتيريوم كما هو مذكور أعلاه.لكل منها). تم تجانس العينات باستخدام كرات خزفية لمدة 10 دقائق باستخدام جهاز Qiagen Tissuelyser II. بعد 10 دقائق من الطرد المركزي عندتم تخفيف السائل العلوي بـميثانول:ماء 1:1.
كانت ظروف UPLC-MS/MS وفقًا للعمل السابقباختصار، بعد الحقن تم فصل أحماض الصفراء على عمود C18; كينيتكس، فينومينيكس، الولايات المتحدة) باستخدام الماء مع 7.5 مللي مولار من أسيتات الأمونيوم وحمض الفورميك (الطور المتحرك A)
الشكل 3 | زيادة مستويات DCA في البلازما من الأفراد المصابين بداء السكري من النوع الثاني. A-D مستويات البلازما من الأحماض الصفراوية الكلية (A)، DCA (B)، isoDCA (C) و 12-epiDCA (D) في الأفراد ذوي مستويات الجلوكوز الطبيعية (NGT) والأفراد المصابين بداء السكري من النوع الثاني (T2D). E، F مستويات الأحماض الصفراوية الكلية في البراز (E) و DCA (F) في ضوابط NGT والأفراد المصابين بداء السكري من النوع الثاني. G، H تحليل ارتباط سبيرمان بين مستويات DCA في البلازما والبراز. ) وبين DCA و LCA و isoUDCA و isoDCA و 12-epiDCA و 12oxoDCA في البراز أو البلازما والمعايير السريرية ( ). لــ ( )، يعني مع جميع النقاط مرسومة؛ عينات/مجموعة؛-القيم تشير إلى الفروقات بين مجموعة NGT (اللون الأزرق) ومجموعة T2D (اللون الوردي) التي تم تحليلها باستخدام اختبار مان-ويتني ذو الاتجاهين مع تعديل بنجاميني وهوشبرغ.-القيم. بالنسبة لـ H، اللون الأحمر يشير إلى ارتباط إيجابي واللون الأزرق يشير إلى ارتباط سلبي و مشار إليها باستخدام تعديل بنجاميني وهوشبرغ-القيم. الكوليسترول، بروتين سي التفاعلي، ضغط الدم الانبساطي، حمض الديوكسيكوليك، الإيوزينوفيلات، سكر الدم الصائم، الهيموغلوبين A1c، البروتين الدهني عالي الكثافة، نموذج التقييم المنزلي لمقاومة الأنسولين، حمض الإيزوديوكسيكوليك، حمض الإيزو-أورسوديوكسيكوليك، حمض الليثوكوليك، البروتين الدهني منخفض الكثافة، اللمفاويات، وحيدات النواة، تحمل الجلوكوز الطبيعي، العدلات، سكر الدم بعد ساعتين من اختبار تحمل الجلوكوز الفموي، ضغط الدم الانقباضي، السكري من النوع 2، الدهون الثلاثية، مؤشر الدهون الثلاثية والجلوكوز، كريات الدم البيضاء، حمض 12-إيبي-ديوكسيكوليك، حمض 12-أوكوديوكسيكوليك. وأسيتونيتريل معحمض الفورميك (الطور المتحرك ب). بدأت عملية الفصل الكروماتوغرافي بـفصل إيزوكراتي. ثم تم زيادة المرحلة ب إلى خلال 4 دقائق. خلال الدقائق العشر التالية، تم زيادة المرحلة ب إلىوأقيم فيلمدة 3.5 دقيقة قبل العودة إلىكان إجمالي وقت التشغيل 20 دقيقة. تم الكشف عن أحماض الصفراء باستخدام مراقبة التفاعل المتعدد في الوضع السالب على جهاز قياس الطيف الكتلي QTRAP 5500 (Sciex، كونكورد، كندا) وتمت عملية التقدير مقابل معايير داخلية مشبعة مناسبة مع تعديلات باستخدام منحنيات معايير فردية خارجية.
اختبارات وظائف الكبد
تم قياس إنزيمات الكبد، الألانين ترانس أميناز (ALT) والأسبارتات ترانس أميناز (AST) فيمن مصل الفئران. تم إجراء التحليلات في قسم الكيمياء السريرية، مستشفى ساهلجرينسكا الجامعي، باستخدام نظام Alinity ci 1303، إصدار البرنامج 3.4.0 (مختبرات أبوت).
تفاعل البوليميراز المتسلسل الكمي في الوقت الحقيقي لعينات الكبد
تم تجانس حوالي 30 ملغ من كبد الفئران المستعمرة باستخدام جهاز Tissuelyser II (Qiagen) وتم عزل RNA الكلي باستخدام مجموعة RNeasy mini (Qiagen، 74106). تم استخدام مجموعة تحويل النسخ العكسي cDNA عالية السعة (Applied Biosystems، 4368813) لتخليقتم الحصول على قوالب cDNA من 500 نانوجرام من RNA المنقى باستخدام بريميرات سداسية عشوائية، وتم تخفيف المنتجات بمعدل 7 مرات قبل استخدامها في التفاعلات اللاحقة.سايبرتم استخدام Green Supermix (Bio-Rad، 1708886) لعملية qRT-PCR بحجوم تفاعل نهائية منتم استخدام بادئات محددة للجينات في كل تفاعل وتم تطبيع جميع بيانات التعبير الجيني على تعبير جين بروتين الريبوسوم L32 (L32) (الأمامية: 5′-CCTCTGGTGA AGCCCAAGATC-3′; العكسية: 5′-TCTGGGTTTCCGCCAGTTT-3′). كانت الجينات والبادئات التي تم تحليلها هي الكوليسترول.-هيدروكسيلاز (Cyp7a1) (الأمام: -AGCAACTAAACAACCTGCCAGTACTA-3′; العكس: 5′-GTCCGGATATTCA AGGATGCA-3′) و 12α-هيدروكسيلاز الستيرول (Cyp8b1) (الأمام: 5′-GGCTGGCTTCCTGAGCTTATT-3′; العكس: 5′-ACTTCCTGAACAGCTCAT CGG-3′).
قياسات ثلاثي غليسريد الكبد
تم تجانس حوالي 50 ملغ من كبد الفئران المستعمرة في أنابيب بولي بروبيلين سعة 2 مل محملة بـ 6 كرات من أكسيد الزركونيوم (3 مم) ثم تم استخراجها باستخدام طريقة BUME.تم إجراء تحليل الدهون الثلاثية في الكبد على عينة من المستخلص الكلي باستخدام كروماتوغرافيا السائل عالية الأداء (HPLC) كما هو موصوف سابقًا.وتمت المعايرة مقابل منحنى المعايرة الخارجي.
تحليل الأحماض الدهنية قصيرة السلسلة لعينات الساكنة باستخدام GC-MS
تم قياس الأحماض الدهنية قصيرة السلسلة في القولون باستخدام تقنية الكروماتوغرافيا الغازية – مطياف الكتلة.تم خلط محتوى القولون من الفئران المستعمرة مع معايير داخلية، وأضيفت إلى قوارير زجاجية وتم تجفيفها بالتجميد. تم تحميض جميع العينات بحمض الهيدروكلوريك، وتم استخراج الأحماض الدهنية قصيرة السلسلة من خلال جولتين من استخراج الإيثر ثنائي الإيثيل. تم جمع الطور العضوي، وأضيف عامل التحويل N-tert-butyldimethylsilyl-Nmethyltrifluoroacetamide (Sigma-Aldrich)، وتم حضن العينات في درجة حرارة الغرفة طوال الليل. تمت كمية الأحماض باستخدام جهاز كروماتوغرافيا الغاز 7090A المتصل بجهاز مطياف الكتلة 5975C (أجيلة تكنولوجيز). تم الحصول على معايير مشعة من حمض البروبيونيك وأملاح الصوديوم من الأسيتات والبيوتيرات من سيغما-ألدريش (ستوكهولم، السويد). تم إجراء الكمية باستخدام معايرة نقطة واحدة مقابل المعيار الداخلي المشع المقابل. تم حقن العينات في جهاز كروماتوغرافيا الغاز 7890A من أجيلة تكنولوجيز المزود بعمود DB-5MS Ultra Inert.، عملت شركة أجيليت تكنولوجيز في وضع الانقسام (50:1). بدأت درجة الحرارة المتدرجة عندلمدة 3 دقائق، الانتقال إلىفي غضون 9.5 دقيقة تليها. زيادة تصل إلى والاحتفاظ لمدة 3.5 دقيقة، مما يعطي وقت تشغيل إجمالي قدره 15.7 دقيقة لكل عينة. تم الكشف عن المواد المحللة باستخدام مطياف الكتلة (Agilent Technologies 5975C) في وضع التأين الإلكتروني (EI). لكل مادة محللة، تم اختيار الأيون أو الشظية الأكثر كثافة للتقدير.
التحليلات الإحصائية
تم تحليل الفروقات بين الفئران المستعمرة مع المجتمع الأساسي والمجتمع الأساسي + C. scindens باستخدام اختبار مان-ويتني لتحليل المعلمات الفسيولوجية، واستخدام تحليل التباين ثنائي الاتجاه مع اختبار المقارنات المتعددة لشيداك لتحليل اختبار تحمل الأنسولين. تم استخدام اختبار كروسكال واليس مع اختبار المقارنات المتعددة لدن عندما تم مقارنة أكثر من مجموعتين. تم تحليل الفروقات بين مجموعة NGT ومجموعة T2D باستخدام مان-ويتني مع تعديل بنجاميني وهوشبرغ.-القيم. تحليل ارتباط سبيرمان مع تعديل بنجاميني وهوشبرغتم استخدام قيم -values لتحليل الارتباط. تم استخدام GraphPad Prism 9.5.0 لتحليل بيانات in vivo وتم استخدام R v4.3.2 لتحليل البيانات البشرية.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة ناتشر المرتبط بهذه المقالة.
توفر البيانات
جميع البيانات التي تدعم نتائج هذه الدراسة متاحة ضمن الورقة والمعلومات التكميلية لها. البيانات الخام لتحليل الأحماض الصفراوية من العينات البشرية متوفرة في ملف بيانات المصدر. يمكن طلب بيانات النمط الظاهري للمشاركين البشريين وفقًا للبروتوكول القياسي للوصول إلى البيانات من المؤلف المقابل. تم توفير بيانات المصدر مع هذه الورقة.
References
Fromentin, S. et al. Microbiome and metabolome features of the cardiometabolic disease spectrum. Nat. Med. 28, 303-314 (2022).
Krautkramer, K. A., Fan, J. & Backhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 19, 77-94 (2021).
Ferrell, J. M. & Chiang, J. Y. L. Understanding bile acid signaling in diabetes: from pathophysiology to therapeutic targets. Diab. Metab. J. 43, 257-272 (2019).
Sayin, S. I. et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 17, 225-235 (2013).
Wahlstrom, A., Sayin, S. I., Marschall, H. U. & Backhed, F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metab. 24, 41-50 (2016).
Chiang, J. Y. L. & Ferrell, J. M. Bile acid receptors FXR and TGR5 signaling in fatty liver diseases and therapy. Am. J. Physiol. Gastrointest. Liver Physiol. 318, G554-G573 (2020).
Ridlon, J. M., Kang, D. J. & Hylemon, P. B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 47, 241-259 (2006).
Funabashi, M. et al. A metabolic pathway for bile acid dehydroxylation by the gut microbiome. Nature 582, 566-570 (2020).
Wells, J. E., Berr, F., Thomas, L. A., Dowling, R. H. & Hylemon, P. B. Isolation and characterization of cholic acid 7alpha-dehydroxylating fecal bacteria from cholesterol gallstone patients. J. Hepatol. 32, 4-10 (2000).
Ridlon, J. M., Kang, D. J., Hylemon, P. B. & Bajaj, J. S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 30, 332-338 (2014).
Ridlon, J. M. et al. The ‘in vivo lifestyle’ of bile acid 7alphadehydroxylating bacteria: comparative genomics, metatranscriptomic, and bile acid metabolomics analysis of a defined microbial community in gnotobiotic mice. Gut Microbes 11, 381-404 (2020).
Devlin, A. S. & Fischbach, M. A. A biosynthetic pathway for a prominent class of microbiota-derived bile acids. Nat. Chem. Biol. 11, 685-690 (2015).
Hofmann, A. F. et al. A proposed nomenclature for bile acids. J. Lipid Res. 33, 599-604 (1992).
Doden, H. L. et al. Completion of the gut microbial epi-bile acid pathway. Gut Microbes 13, 1-20 (2021).
Wu, M. et al. Genetic determinants of in vivo fitness and diet responsiveness in multiple human gut Bacteroides. Science 350, aac5992 (2015).
Marion, S. et al. In vitro and in vivo characterization of Clostridium scindens bile acid transformations. Gut Microbes 10, 481-503 (2019).
Studer, N. et al. Functional intestinal bile acid 7alphadehydroxylation by clostridium scindens associated with protection from Clostridium difficile infection in a gnotobiotic mouse model. Front. Cell Infect. Microbiol. 6, 191 (2016).
Marion, S. et al. Biogeography of microbial bile acid transformations along the murine gut. J. Lipid Res. 61, 1450-1463 (2020).
Song, P., Zhang, Y. & Klaassen, C. D. Dose-response of five bile acids on serum and liver bile Acid concentrations and hepatotoxicty in mice. Toxicol. Sci. 123, 359-367 (2011).
Hylemon, P. B., Harris, S. C. & Ridlon, J. M. Metabolism of hydrogen gases and bile acids in the gut microbiome. FEBS Lett. 592, 2070-2082 (2018).
Narushima, S., Itoh, K., Takamine, F. & Uchida, K. Absence of cecal secondary bile acids in gnotobiotic mice associated with two human intestinal bacteria with the ability to dehydroxylate bile acids in vitro. Microbiol. Immunol. 43, 893-897 (1999).
Narushima, S. et al. Deoxycholic acid formation in gnotobiotic mice associated with human intestinal bacteria. Lipids 41, 835-843 (2006).
Ridlon, J. M., Kang, D.-J. & Hylemon, P. B. Isolation and characterization of a bile acid inducible -dehydroxylating operon in Clostridium hylemonae TN271. Anaerobe 16, 137-146 (2010).
Khan, M. T. et al. Synergy and oxygen adaptation for development of next-generation probiotics. Nature 620, 381-385 (2023).
Zaborska, K. E., Lee, S. A., Garribay, D., Cha, E. & Cummings, B. P. Deoxycholic acid supplementation impairs glucose homeostasis in mice. PLoS ONE 13, e0200908 (2018).
Yoshimoto, S. et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 499, 97-101 (2013).
Chavez-Talavera, O. et al. Bile acids associate with glucose metabolism, but do not predict conversion from impaired fasting glucose to diabetes. Metabolism 103, 154042 (2020).
Haeusler, R. A., Astiarraga, B., Camastra, S., Accili, D. & Ferrannini, E. Human insulin resistance is associated with increased plasma levels of 12alpha-hydroxylated bile acids. Diabetes 62, 4184-4191 (2013).
Atlas R. M. Handbook of Microbiological Media, 3rd edition (2004).
Wu, H. et al. The gut microbiota in prediabetes and diabetes: a population-based cross-sectional study. Cell Metab. 32, 379-90 e3 (2020).
Bergstrom, G. et al. The Swedish CArdioPulmonary Biolmage Study: objectives and design. J. Intern. Med. 278, 645-659 (2015).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357-359 (2012).
Danecek, P. et al. Twelve years of SAMtools and BCFtools. Gigascience 10, giab008 (2021).
Lawson, A. M., Setchell, K. D. R. The Bile Acids: Chemistry, Physiology, and Metabolism: Volume 4: Methods and Applications (eds Setchell, K. D. R., Kritchevsky, D. & Nair, P. P.) 167-267 (Springer US, 1988).
Buffie, C. G. et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 517, 205-208 (2015).
Tremaroli, V. et al. Roux-en-Y gastric bypass and vertical banded gastroplasty induce long-term changes on the human gut microbiome contributing to fat mass regulation. Cell Metab. 22, 228-238 (2015).
Lofgren, L. et al. The BUME method: a novel automated chloroformfree 96-well total lipid extraction method for blood plasma. J. Lipid Res. 53, 1690-1700 (2012).
Homan, R. & Anderson, M. K. Rapid separation and quantitation of combined neutral and polar lipid classes by high-performance liquid chromatography and evaporative light-scattering mass detection. J. Chromatogr. B Biomed. Sci. Appl. 708, 21-26 (1998).
شكر وتقدير
نشكر آنا هالن، كارينا أرفيدسون، ولويز هيلدين على المساعدة الفنية ومختبر آدم أركين على توفير Eubacterium rectale ATCC 33656. تم دعم هذا العمل من خلال منح من مؤسسة ليدوك (17CVD01)، ومؤسسة القلب والرئة السويدية (20210366)، ومؤسسة كنوت وأليس وولنبيرغ (2017.0026)، ومجلس البحث السويدي (2019-01599)، وتأمينات AFA (160337)، ومؤسسة نوفو نورديسك (NNF21OC0070298)، ومن الدولة السويدية بموجب الاتفاق بين الحكومة السويدية والمجالس المحلية، اتفاقية ALF (ALFGBG-718101). F.B. هو أستاذ تورستن سودربيرغ في الطب وعالم وولنبيرغ. كما تم دعم S.L.H. وM.A.F. جزئيًا من خلال منحة المعاهد الوطنية للصحة P01 HL147823.
تملك جامعة ستانفورد وChan Zuckerberg Biohub براءات اختراع قيد الانتظار لتقنيات الميكروبيوم التي يعتبر A.B. وM.A.F. من المخترعين المشاركين فيها. M.A.F. هو أحد المؤسسين ومدير شركة Federation Bio وKelonia، وأحد مؤسسي شركة Revolution Medicines، وعضو في اللجان الاستشارية العلمية لشركتي NGM Bio وZymergen. S.L.H. يذكر أنه تلقى مدفوعات حقوق ملكية عن اختراعات أو اكتشافات تتعلق بتشخيصات أو علاجات القلب والأوعية الدموية من Cleveland Heart Lab، وهي شركة تابعة مملوكة بالكامل لشركة Quest Diagnostics، وProcter & Gamble. كونه مستشارًا مدفوع الأجر لشركة Zehna Therapeutics، وقد تلقى أموال بحث من Procter & Gamble و Pfizer Inc. و Roche Diagnostics و Zehna Therapeutics. F.B. هو أحد مؤسسي ومالكي شركة Roxbiosens Inc و Implexion Pharma AB، ويتلقى تمويلًا بحثيًا من Biogaia AB، وهو عضو في المجلس الاستشاري العلمي لشركة Bactolife A/S. يعلن المؤلفون الآخرون عدم وجود مصالح متنافسة.
مختبر والينبرغ وقسم الطب الجزيئي والسريري، معهد الطب، أكاديمية ساهلجرينسكا، جامعة غوتنبرغ، غوتنبرغ، السويد.قسم الهندسة الحيوية، جامعة ستانفورد، ستانفورد، كاليفورنيا، الولايات المتحدة الأمريكية.قسم علم الأحياء الدقيقة والمناعة، كلية الطب بجامعة ستانفورد، جامعة ستانفورد، ستانفورد، كاليفورنيا، الولايات المتحدة الأمريكية.معهد ChEM-H، جامعة ستانفورد، ستانفورد، كاليفورنيا، الولايات المتحدة الأمريكية.مركز تشان زوكربيرغ للبيولوجيا، سان فرانسيسكو، كاليفورنيا، الولايات المتحدة الأمريكية.المختبر الوطني الرئيسي للهندسة الوراثية، معهد الفينوم البشري، مركز فودان للميكروبات، وقسم جراحة السمنة والتمثيل الغذائي، مستشفى هواشان، جامعة فودان، شنغهاي، الصين.قسم علوم القلب والأوعية الدموية والتمثيل الغذائي، معهد ليرنر للبحوث، كليفلاند، أوهايو، الولايات المتحدة الأمريكية.مركز الميكروبيوم وصحة الإنسان، عيادة كليفلاند، كليفلاند، أوهايو، الولايات المتحدة الأمريكية.قسم طب القلب والأوعية الدموية، معهد القلب والأوعية الدموية والصدر، عيادة كليفلاند، كليفلاند، أوهايو، الولايات المتحدة الأمريكية.منطقة فاسترا غوتالاند، مستشفى ساهلغرنسكا الجامعي، قسم الفيزيولوجيا السريرية، غوتنبرغ، السويد.مركز نوفو نورديسك للأبحاث الأساسية في الأيض، كلية العلوم الصحية، جامعة كوبنهاغن، كوبنهاغن، الدنمارك.الراحل: هانس-أولريش مارشال.البريد الإلكتروني: fischbach@fischbachgroup.org; Fredrik.Backhed@wlab.gu.se
Alterations in gut microbiota composition are suggested to contribute to cardiometabolic diseases, in part by producing bioactive molecules. Some of the metabolites are produced by very low abundant bacterial taxa, which largely have been neglected due to limits of detection. However, the concentration of microbially produced metabolites from these taxa can still reach high levels and have substantial impact on host physiology. To explore this concept, we focused on the generation of secondary bile acids by -dehydroxylating bacteria and demonstrated that addition of a very low abundant bacteria to a community can change the metabolic output dramatically. We show that Clostridium scindens converts cholic acid into the secondary bile acid deoxycholic acid (DCA) very efficiently even though the abundance of . scindens is low, but still detectable by digital droplet PCR. We also show that colonization of germ-free female mice with a community containing C. scindens induces DCA production and affects host metabolism. Finally, we show that DCA correlates with impaired glucose metabolism and a worsened lipid profile in individuals with type 2 diabetes, which implies that this metabolic pathway may contribute to the development of cardiometabolic disease.
Cardiometabolic diseases such as obesity and type 2 diabetes mellitus (T2D) are increasing globally and there is an urge for new treatments. Alterations in the gut microbiota are implicated in metabolic diseases , but how these alterations contribute to the disease is still not fully
understood. One key challenge in microbiota research is to understand how different bacteria function in consortia to produce bioactive molecules. Some microbial metabolites are produced by very low abundant bacterial taxa, yet the metabolites reach very high
concentrations in the gut and circulation where they can have substantial impact on host physiology . An example of such metabolites are secondary bile acids, which are among the most abundant microbiota-derived molecules, although only a few known bacterial species are able to produce them. Bile acids signal via host nuclear- and G-protein coupled receptors and can affect many different metabolic pathways . Different bile acids have varying affinities for their receptors; the gut microbiota can thus modulate signaling via the host receptors by changing the bile acid composition . The gut microbiota modifies the bile acid pool by metabolizing primary bile acids into secondary bile acids through dehydroxylation, dehydrogenation and epimerization . The two most prevalent secondary bile acids in humans are deoxycholic acid (DCA) and lithocholic acid (LCA), which are produced by -dehydroxylating bacteria from cholic acid (CA) and chenodeoxycholic acid (CDCA), respectively . Microbial dehydroxylation is a complex multistep process, which has just recently been fully unveiled and shown to require enzymes encoded by the bai-operon . The number of bacterial species that possess the whole bai-operon are limited and they have been identified in Eubacterium and Clostridium cluster XIVa and XI . Although, dehydroxylating bacteria are typically present at low abundance in the human gut, their metabolic potential is high and more than 95% of the bile acid pool in the large intestine consists of dehydroxylated bile acids . Furthermore, in addition to DCA and LCA there is a wide range of other microbially produced bile acid metabolites such as iso-, epi-, and oxo-bile acids, which might be biologically important but are less well studied .
In this study, we explore if addition of a low abundance bacterial species into a community could have significant impact on the metabolic output. We chose to focus on bile acids since they are involved in many physiological processes and secondary bile acids are among the most abundant microbial-derived metabolites. We investigated DCA production by a low abundant bacterial species, Clostridium scindens, both in vitro and in vivo, and demonstrated a link between DCA and impaired host metabolism.
Results
Generation of a simplified community with bile acidmetabolizing capacity
To investigate if we could produce a system to assess the function of low abundant bacteria, we first established an in vitro system in which a -dehydroxylating bacterial species was present at low abundance, to study the formation of DCA from CA. First, we constructed a ‘base’ synthetic community, based on previous work consisting of bacterial species that lack the bai genes. The nine selected bacteria were cultured individually in the presence of CA to assess their bile acidmetabolizing capacity. Three of them (Bacteroides caccae, Bacteroides thetaiotaomicron and Bacteroides ovatus) produced 7-oxoCA, while none of the base community members produced DCA (Fig. 1A). Similarly, the nine bacteria did not produce DCA in vitro when they were cultured together (Fig. 1B). In contrast, C. scindens, a known dehydroxylating bacterium , converted CA to DCA both when cultured alone (Fig. 1C) and together with the base community (Fig. 1D).
Finally, to mimic the situation in the human gut we performed serial dilutions to decrease the abundance of . scindens in the simplified community. To quantify the level of . scindens in our diluted communities we used shotgun metagenomic sequencing coupled with digital droplet PCR, with specific primer-probe sets for . scindens and B. caccae, a high abundance community member detected in the shotgun metagenomic data and used as a reference (Supplementary Table 1). We then assessed the capability of C. scindens to generate DCA in the different dilutions and we found that at dilutions that range from to . scindens could still convert of CA to DCA
in 24 h , establishing that in a controlled synthetic community setting, a low abundant organism can have a very high metabolic output (Fig. 1E).
DCA-producing C. scindens modulates physiology and glucose metabolism in colonized mice
We next investigated if the base community . scindens produced DCA in vivo by colonizing germ-free mice for two weeks. Since germfree mice have low levels of , the precursor of DCA, we fed the mice a diet containing . At termination, we analyzed microbiota composition in the inoculum and in caecum by performing shotgun metagenomic sequencing (Fig. 2A). We observed that the caecal microbiota was dominated by Bacteroides species and that the relative abundance of . scindens was below the limit of quantification. Hence, we performed digital droplet PCR and found that the ratio of . scindens genome copies to the total community was 1:14,397 (Fig. 2B). Importantly, C. scindens was not detected in any of the mice colonized with the base community, establishing that we could generate communities that differed in one low abundant species in vivo.
We then analyzed bile acids in serum from vena cava and caecal content from mice colonized with the base community and demonstrated that these mice could deconjugate tauro-beta-muricholic acid ( ) and tauro- CA (TCA) into and CA respectively, but they did not further metabolize them to secondary bile acids (Fig. 2C, D). In contrast, mice colonized with base community . scindens showed substantial DCA production despite low abundance of . scindens ( of all community members). DCA levels ranged from in serum and in caecum of mice colonized with base community + C. scindens, while DCA was undetectable in serum of mice colonized with base community and only traces were found in caecum (Fig. 2C, D). To compare the DCA levels in our colonized mice with physiological levels we also measured DCA in serum and caecum of conventionally raised mice on chow or 1% CAdiet. The conventionally raised mice on CA-diet had equally high DCA levels in serum as mice colonized with base community . scindens but higher DCA levels in caecum (Figure S1A). In concordance with previous findings, the CA-diet resulted in elevated serum levels of alanine transferase (ALT) and aspartate aminotransferase (AST), indicating a hepatotoxic effect of . However, there were no differences between mice colonized with base community and base community + C. scindens (Figure S1B). Of note, mice colonized with C. scindens had reduced hepatic gene expression of Cyp7a1 while there was no significant difference in Cyp8b1 expression between the two groups (Figure S1C).
Furthermore, mice colonized with base community . scindens had similar body weight as mice colonized with base community (Fig. 2E). However, mice colonized with base community + C. scindens had increased adiposity (Fig. 2F) and liver weight (Fig. 2G), but reduced caecum weight (Fig. 2H), compared to mice colonized with base community. In line with increased adiposity, liver triglycerides were also increased in mice colonized with base community + C. scindens and correlated with relative liver weight (Figure S1D). Analysis of caecal short chain fatty acids showed higher butyrate levels in mice colonized with base community + C. scindens, which could be due to increased carbohydrate fermentation and may in part explain the reduction in relative caecum weight (Figure S1E).
At the end of the experiment, we performed an intraperitoneal insulin tolerance test and observed that mice colonized with base community + C. scindens had impaired insulin tolerance, indicating that glucose metabolism may be altered (Fig. 2I) although there were no differences in fasting blood glucose or fasting insulin (Fig. 2J, K). Taken together, our in vitro and in vivo systems show that addition of a low abundance bacterial species into a community can have a large effect on the microbial metabolic output, which subsequently can alter host metabolism.
DCA is enriched in T2D individuals and correlates with worsened metabolic characteristics
To investigate if alterations in DCA levels could be implicated in human metabolic diseases, we selected 100 individuals with normal glucose tolerance (NGT) and 100 individuals with treatment naive T2D from the IGT Microbiota and SCAPIS cohorts and analyzed their bile acid profiles. The NGT and T2D groups were matched for sex, age, weight,
BMI, statins, and proton-pump inhibitor use and none in the T2D group were on diabetes medication (Table 1).
Bile acid analysis in plasma revealed that total bile acid levels were increased in individuals with T2D compared with NGT controls ( in the T2D group vs in the NGT group, ; Fig. 3A and Supplementary Data 1). Analysis of individual bile acids demonstrated that 12-hydroxylated secondary bile acids e.g.,
Fig. 1 | In vitro assessment of DCA formation by the bacterial species included in the simplified communities. A-D Assessment of DCA formation after 48 h culture in Mega Medium containing CA by the base community members Bacteroides caccae ATCC 43185, Bacteroides vulgatus ATCC 8482, Eubacterium rectale ATCC 33656, Bacteroides thetaiotaomicron VPI-5482, Bacteroides uniformis ATCC 8492, Parabacteroides distasonis ATCC 8503, Bacteroides ovatus ATCC 8483, Ruminococcus torques ATCC 27756 and Dorea longicatena DSM 13814 cultured individually (A), base community members cultured together (B), Clostridium
scindens ATCC 35704 cultured individually (C) and base community + Clostridium scindens cultured together (D). E Assessment of DCA production after 24 h in serial dilutions of Clostridium scindens in the base community spiked with CA from a 100 mM stock solution to a final concentration of . Abundance of CA, DCA and 7-oxoCA was measured by GC-MS in (A-D) and by UPLC-Q-TOF-MS in (E). CA, cholic acid; DCA, deoxycholic acid; 7-oxoCA, 7-oxocholic acid. Black line indicates bile acid abundance at 0 h and blue line indicates bile acid abundance at ; blue line indicates 7-oxoCA; black line indicates CA and red line indicates DCA (E).
DCA, isoDCA and 12 -epiDCA were the most significantly increased bile acids in individuals with T2D (Fig. 3B-D and Supplementary Fig. 2A) and 8 additional bile acids were also increased (Supplementary Fig. 2A and Supplementary Data 1).
Next, we analyzed bile acid composition in faeces but did not observe any significant differences in total bile acid levels (Fig. 3E) or in any of the analyzed bile acid species (Supplementary Fig. 2B and Supplementary Data 2). However, DCA was the most abundant bile acid and tended to be increased in faeces from individuals with T2D ( before adjustment for multiple testing) (Fig. 3F and Supplementary Data 2) and correlated with DCA levels in plasma (Fig. 3G).
To investigate associations between DCA and physiological parameters related to impaired glucose metabolism, we correlated clinical parameters (Table 1) with bile acids in faeces and plasma. Although we did not observe significant differences in faecal DCA between the NGT and T2D groups, we observed significant correlations between faecal DCA and parameters associated with impaired glucose metabolism such as HOMA-IR, fasting blood glucose (FBG), HbA1c, and insulin (Fig. 3H and Supplementary Data 2). In addition, DCA correlated positively with triglycerides (TG) and negatively with high-density lipoprotein (HDL), which is in line with a worsened lipid profile. The correlations were observed when all individuals were analyzed together and also when individuals with T2D were analyzed separately (Fig. 3H). In contrast, we did not observe any significant correlations when the NGT group was analyzed separately. DCA also correlated positively with C-reactive protein (CRP) and white blood cells (e.g., neutrophils) suggesting that it is associated with inflammation. Plasma DCA showed a similar correlation pattern as faecal DCA and the strongest correlation was found with HOMA-IR, both in the total cohort and when the T2D group was analyzed separately (Fig. 3H and Supplementary Data 1). In summary, DCA, in both faeces and plasma, is associated with impaired glucose metabolism and a worsened lipid profile.
Discussion
In this study we demonstrated that a low abundance bacterial species, which might be neglected due to detection limitations, can have a significant impact on the production of bioactive metabolites.
We showed that C. scindens, although present at low abundance, could produce large amounts of DCA and affect insulin tolerance in colonized mice, suggesting that DCA may contribute to impaired glucose metabolism. Furthermore, we demonstrated that DCA, in plasma and faeces, correlates with impaired glucose metabolism and a worsened lipid profile in individuals with T2D. Our findings highlight that low abundant bacterial species may have profound effects on metabolite levels in the gut and circulation and should receive more attention. One challenge of studying these organisms in metagenomes is that they may be close to the detection limit and thus robust analyses are lacking. Accordingly, this concept may be extended to other metabolites that are produced by low abundant taxa and emphasize the importance of studying gut microbial metabolites in the circulation of host organisms. Another challenge is that bile acid transformation is very complex and a variety of environmental factors, such as medications, diet, pH , gas atmosphere and presence of enzymatic cofactors can influence interactions between bile acids and bacteria in the gut . These parameters are dependent on the crosstalk between
different bacteria in a community and the function of a specific bacterial species can differ when it is cultured alone compared to when it is cultured in a consortium of other bacteria. There can also be discrepancies in bile acid transformation by bacteria in vitro and in vivo , which emphasize the importance of comprehensive studies on bile acid metabolizing bacteria.
Our results clearly show that very low abundance of dehydroxylating bacteria is sufficient to produce large amounts of DCA and this supports the idea that -dehydroxylation could be exclusive for very few bacteria in the human gut. However, DCA levels in caecum were lower than the levels in conventionally raised mice, which have a complete microbiota, and we cannot exclude that there are other unidentified bacterial species in the human gut that contribute to the formation of DCA, such as Clostridium hylemonae , and further studies are needed to provide an extended view of bile acid metabolizing bacteria. There could also be synergistic effects between C. scindens and other bacteria in the gut, comparable to what we previously have shown between Faecalibacterium prausnitzii and Desulfovibrio piger . In that study, co-colonization of mice with . piger increased the abundance and metabolic capacity of . prausnitzii and it will be important to elucidate similar mechanisms for . scindens and other DCA-producing bacteria.
Bile acids have generated significant interest as signaling molecules over the past years and here we demonstrate that insulin tolerance is affected in mice colonized with DCA-producing C. scindens. This is in line with a previous study where Zaborska et al showed that DCA supplementation in mice leads to impaired glucose homeostasis (both by insulin- and glucose tolerance tests) and increased hepatic ER stress and decreased hepatic insulin signaling . In another study, Yoshimoto and co-workers showed that DCA production was increased in mice with diet- or genetically induced obesity and that DCA contributed to cancer development in the obese mice . Contrary to Yoshimoto’s study, we used a chow diet supplemented with CA and our mice were not obese. Nevertheless, we found increased adiposity and liver triglycerides in our DCA-producing mice, supporting that there is a link between DCA and lipid accumulation although the mechanisms might be different.
Bile acid synthesis has also been implicated in metabolic changes and we observed decreased expression of Cyp7a1 in the livers of mice colonized with base community . scindens but no difference in the expression of Cyp8b1. However, the contribution of endogenously synthesized bile acids in our mouse model is probably negligible since the mice were administered a very high dose of exogenous CA through the diet.
We also showed that DCA levels are increased in plasma of individuals with T2D and that the altered bile acid profile is not a consequence of diabetes medication since all individuals with T2D were newly diagnosed and had not been treated for their diabetes. It is currently unclear why the faecal concentrations of DCA were not significantly different, but one possible explanation could be that we only obtained spot samples rather than samples, or that there are differences in bile acid uptake.
Previous studies have shown conflicting results regarding plasma bile acid profiles and glucose metabolism . Hausler et al showed that an increased ratio of non bile acids is associated with insulin resistance , while another study found no correlation between ratio and HOMA-IR or risk of new onset of diabetes . One potential reason for this discrepancy might be that there are differences in the microbial conversion within the bile acid species i.e., conversion of CA into DCA and/or iso-DCA and 12epiDCA, hence non ratio might be a crude measurement and needs to be further dissected. We found strong correlations
between DCA and parameters related to glucose and lipid metabolism and our clinical findings suggest that DCA may be a biomarker for inflammation and cardiometabolic diseases. However, further studies are needed to establish the causality between DCA and metabolic disease and prospective studies may reveal if DCA can be used to identify individuals at risk of developing T2D.
Fig. 2 | Colonization of germ-free mice with bacterial communities. A Relative abundance of bacteria in the inoculum and the caecal contents of mice colonized with base community or base community + C. scindens assessed by shotgun metagenomic sequencing. B Microbial genome copies per nanogram bacterial DNA in the caecal content of mice colonized with base community . scindens assessed by digital droplet PCR. The base community members were quantified using primer-probe sets for the gene tilS, and C. scindens was quantified using primer-probe sets for the baiCD gene (see Supplementary Table 1). C, D Bile acid levels in vena cava (C) and caecum (D) of mice colonized with base community or base community + C. scindens measured by UPLC-MS/MS. E-H Physiological parameters of the mice including body weight (E), adiposity (F), relative liver weight ( ) and relative caecum weight ( ) measured 2 weeks after colonization. I Insulin tolerance test performed by intraperitoneal injection of insulin. Glucose
was measured at after injection and data is presented as percentage of glucose compared to the 0 min time point. Fasting blood glucose ( ) and insulin ( ) were measured at the 0 min time point. samples for the base community group (indicated by white color in B-K) and samples for the base community . scindens group (indicated by orange color in B-K); -values indicate differences between mice colonized with base community and base community + C . scindens analyzed with two-tailed Mann-Whitney (E-H and J, K) or 2-way ANOVA with Šidák multiple comparisons test (I). Error bars indicate standard error of the mean (C,D and I). CA cholic acid, DCA deoxycholic acid, ßMCA beta-muricholic acid, TCA tauro-cholic acid, TDCA tauro-deoxycholic acid, T MCA tauro-alpha-muricholic acid, T MCA tauro-betamuricholic acid, 7-oxoCA 7-oxocholic acid.
Methods
Bacterial strains and culture conditions
Synthetic community strains were cultured in pre-reduced Mega Medium or on EG at in an anaerobic chamber (Coy Laboratories, Grass Lake, MI, USA) with an atmosphere of hydrogen, and . Synthetic community strains included Bacteroides uniformis ATCC 8492, Bacteroides vulgatus ATCC 8482, Bacteroides thetaiotaomicron VPI-5482, Bacteroides caccae ATCC 43185, Bacteroides ovatus ATCC 8483, Parabacteroides distasonis ATCC 8503, Eubacterium rectale ATCC 33656, Dorea longicatena DSM 13814, Ruminococcus torques ATCC 27756, and Clostridium scindens ATCC 35704. All strains were obtained from the American Type Culture Collection (ATCC), except D. longicatena DSM 13814, which was obtained from The Leibniz Institute DSMZ (Braunschweig, Germany).
Animal experiments
Germ-free female C57BI/6 mice were maintained in flexible plastic gnotobiotic isolators under a strict 12 h light cycle (light from 7 a.m. to 7 p.m.), temperature, air humidity of , under standard germ-free conditions, and fed an autoclaved chow diet (Lab diet, St Louis, MO; #5021) supplemented with 1% CA (Envigo Teklad Diets, Madison, WI) ad libitum. Germ-free isolators were routinely tested for sterility by culturing and PCR analysis of faeces amplifying the 16S rRNA gene.
Germ-free mice were colonized after 4 h fasting by gavage with 0.2 ml of the cultured communities (base community or base community including . scindens). Before colonization all the mice were weighed and randomized into two weight-matched groups. The communities of bacteria were thawed and gavaged into the germ-free mice within 2 h after thawing. The colonization was repeated after 2 days. Mice were colonized at 17 weeks of age and maintained for 2 weeks after colonization. Gut microbial composition following colonization was monitored by sequencing and digital droplet PCR (ddPCR) as detailed below.
An insulin tolerance test was performed by injecting insulin kg body weight intraperitoneally after 4 h fasting. Tail blood samples were collected at , and 120 min and blood glucose levels were determined using a glucose meter (Contour NEXT, Ascensia Diabetes Care, Stockholm, Sweden). Body composition was determined with magnetic resonance imaging (EchoMRI, Houston, TX) after the insulin tolerance test and blood was collected from the inferior vena cava under deep isoflurane-induced anesthesia. The mice were then killed, and serum and tissues were collected. Body, liver and caecum weights were recorded, and tissues were immediately frozen in liquid nitrogen and stored at until further analyzed.
In a separate experiment conventionally raised female C57BI/6 mice were maintained under a strict 12 h light cycle (light from 7 a.m. to 7 p.m.), temperature, air humidity of , under standard specific-pathogen-free conditions and fed either chow diet (Lab diet, St Louis, MO) or a diet supplemented with 1% CA (Envigo Teklad Diets, Madison, WI) ad libitum for two weeks. The mice were then killed, and serum and caecum were collected for analyses. All animal
experiments were performed at Experimental Biomedicine, University of Gothenburg, using protocols approved by the Gothenburg Animal Research Ethics Committee (Ethical number 4805/23).
Human cohorts
200 individuals (100 with treatment naive T2D and 100 BMI matched controls with normal glucose tolerance (NGT)) were selected from the IGT Microbiota cohort ( 45 T2D and 45 NGT), and the Swedish Cardiopulmonary Bioimage Study (SCAPIS)-Gothenburg cohort (55 T2D and 55 NGT). The design of the two human cohorts have been described in detail elsewhere . Both cohorts comprise men and women aged 50-64 years from the Gothenburg area, Sweden, who were recruited at random from the census register. Exclusion criteria were known diabetes; inflammatory diseases, such as Crohn’s disease, ulcerative colitis, rheumatic diseases; treatment with steroids or immunomodulatory drugs; cancer (unless relapse free for the preceding 5 years); cognitive dysfunction; and treatment for infectious diseases and with antibiotics in the past three months. Accordingly, none of the age, sex and BMI matched controls were taking either metformin or weight reduction medications like GLP1 agonists. Individuals were also excluded if they did not understand Swedish and if they were born outside Sweden. All participants gave informed consent, and the study was approved by the Ethics Review Board in Gothenburg (560-13) and in Umeå 2010-228-31M (2012-285-32M and 2014-33-32).
We collected plasma and faecal samples from the 200 individuals and metabolic parameters were evaluated. Bile acids were assessed in plasma and faeces.
DNA extraction and sequencing of bacterial cultures and mouse caecal samples
Total DNA from mouse caecal contents and bacterial culture pellets were extracted using the PowerFecal DNA Isolation kit (MoBio, Carlsbad, CA, USA), and quantified using the Qubit dsDNA BR Assay Kit and Invitrogen Qubit Fluorometer (Thermo Fischer, Waltham, MA, USA). Sequencing libraries were generated using the Nextera XT DNA Library Preparation kit and Index v2 kit (Illumina, Hayward, CA, USA) and sequenced on an Illumina MiSeq with MiSeq Reagent Kit v2 (300 cycles) (Illumina, Hayward, CA, USA). Sequencing reads were mapped to a reference database built with only the genomes of bacterial strains in the study using Bowtie2 2.2.9 and the relative abundance of bacterial community members was quantified using SAMtools 1.3.1 .
ddPCR quantification
IDT Taqman ddPCR primer/probe sets were designed to the baiCD gene (bile acid-inducible gene) for . scindens ATCC 35704, and the single copy essential gene tilS (tRNAIle-lysidine synthase) for B. caccae ATCC 43185 to ensure each genome was counted only once. In brief, a reaction comprising water, BioRad ddPCR Supermix for Probes (no dUTP) [cat: 186-3023], NEB HindIII-HF, Taqman Primer/Probe Set 20X in IDTE buffer, input DNA was used for each sample. Droplets were generated with the QX200
Table 1 | Characteristics of the study participants in the NGT and T2D groups
Characteristics of study participants
NGT
T2D
-value
Gender
44 (% females)
42 (% females)
ns
Age (years)
ns
Weight (kg)
ns
Height (cm)
ns
BMI ( )
ns
Waist (cm)
0.01
Hip (cm)
ns
whr
SBP (mmHg)
ns
DBP (mmHg)
ns
Hb (g/L)
0.05
FBG (mmol/L)
OGTT 2 h BG (mmol/L)
Insulin (pmol/L)
0.02
HbA1c (%)
HOMA_IR
TyG_index
TG (mmol/L)
Total Chol (mmol/L)
ns
LDL (mmol/L)
ns
HDL (mmol/L)
CRP (mg/L)
0.001
WBC ( )
0.04
Neut ( )
0.02
Mono ( )
ns
EOS ( )
ns
Lymph ( )
ns
Statin
4 (% av all)
12 (% of all)
ns
PPI
53 (% of all)
54 (% of all)
ns
Data are presented as mean SD, the for each parameter is indicated within the parenthesis, Gender is presented as the percentage of females in the group, statin and PPI are presented as the percentage of users in the group. -values indicate differences between the NGT and T2D group analyzed with two-tailed Mann-Whitney using Benjamini and Hochberg adjusted -values. Chol cholesterol, CRP C-reactive protein, DBP diastolic blood pressure, Eos eosinophils, FBG fasting blood glucose, HbA1c Hemoglobin A1c, HDL high-density lipoprotein, HOMA-IR Homeostatic Model Assessment for Insulin Resistance, LDL low-density lipoprotein, Lymph lymphocytes, Mono monocytes, NGT normal glucose tolerance, Neut neutrophils, OGTT 2 h BG blood glucose at 2 h after oral glucose tolerance test, PPI proton-pump inhibitor, SBP systolic blood pressure, T2D Type 2 diabetes, TG triglycerides, TyG triglyceride-glucose index, WBC white blood cells, whr waist-to-hip ratio.
Droplet Digital PCR System (Bio-Rad) and amplification was performed by incubating at for 5 min followed by 40 cycles of for 30 s , for 1 min , and then holding at for 10 min to deactivate the enzyme. A ramp rate of per second was used. Reactions were read using the QX200 Droplet reader (Bio-Rad) and analyzed using QuantaSoft software v1.6.
Bile acid metabolism in coculture
Overnight monocultures of bacterial community members were harvested by anaerobic centrifugation and resuspended in fresh prereduced Mega Medium with CA (Sigma-Aldrich, St. Louis, MO, USA) and allowed to incubate at for 48 h to induce bile acid metabolism. These cultures were harvested by anaerobic centrifugation, pellets were washed with pre-reduced PBS and resuspended densely (half the volume of the prime culture) in fresh pre-reduced
Mega Medium. Bacteroidetes and Firmicutes cultures, excluding C. scindens, were combined by volume and aliquoted. C. scindens was serially diluted 1:10 into aliquots of the combined base community and CA was spiked into each coculture from a 100 mM DMSO stock solution to a final concentration of . Cocultures were sampled for bile acid and DNA analyses 24 h post-CA spike. For bile acid analysis, 2 ml of culture was sampled and immediately acidified with of 6 N HCl to stop all metabolic activity and protonate bile acids to make them more soluble in organic solvent. For DNA analysis, 1 ml of culture was sampled, pelleted, and stored at until extraction and sequencing.
Bile acid profiling of bacterial cultures by gas chromatography-mass spectrometry (GC-MS)
This method was applied to experiments presented in Fig. 1A-D. Acidified bacterial cultures were extracted twice with equal volumes of ethyl acetate, and the organic extracts were pooled, passed through a plug of , and concentrated to dryness. The residue was purified using a 1.5 cm silica pipet column. The column was washed with 2 ml of methanol in dichloromethane, eluted with 6 ml of methanol in ethyl acetate, and the eluent was concentrated to dryness. Prior to GC-MS analysis, purified bile acids were converted to their methyl ester-trimethylsilyl ether derivatives as follows. Bile acids were resuspended in methanol and a 2 M solution of (trimethylsilyl)diazomethane in diethyl ether was added dropwise until a yellow color persisted. The solution was concentrated to dryness and of a 3:1 solution of N,O-bistrimethylsilyl) trifluoroacetamide (BSTFA) and chlorotrimethylsilane was added. GC-MS analysis was performed on of the resulting solution. Bile acids were identified based either on retention times and MS fragmentation patterns of standards or on published MS fragmentation patterns .
Bile acid profiling of bacterial cultures by ultra-performance liquid chromatography (UPLC)-quadrupole time of flight mass spectrometry (Q-TOF-MS)
This method was adapted from a method developed by Justin R. Cross and was applied to experiments presented in Fig. 1E. Briefly, acidified bacterial cultures were extracted twice with equal volumes of ethyl acetate, the organic extracts were pooled, passed through a plug of , concentrated to dryness and reconstituted in methanol in water. Compounds were separated on an Agilent 1290 Infinity II UPLC using a Kinetex C18 column ( , Phenomenex, Cat. #00D-4475-AN) and detected using an Agilent 6530 Q-TOF equipped with a dual-nebulizer ESI source in negative ionization mode, sheath gas temperature was . Chromatographic separation was achieved at a flow rate of with a gradient ), where Mobile Phase A was water with formic acid and Mobile Phase B was acetone with formic acid.
Bile acid profiling of samples from mice and humans by UPLC-MS/MS
This method was applied to experiments presented in Fig. 2C, D, Fig. 3A-G, Supplementary Fig. 1A, B and Supplementary Data. Bile acids in serum and caecum from mice, and plasma and faeces from the humans, were analyzed using UPLC-MS/MS and quantified using a combination of unlabeled standards and deuterium-labeled internal standards. serum and of caecum from the mice, and plasma and faeces from the human participants, were used for the analyses.
Serum and plasma samples were extracted with 10 volumes of methanol containing d4-TCA, d4-GCA, d4-GCDCA, d4-GUDCA, d4GLCA, d4-UDCA, d4-CDCA, d4-LCA (50 nM of each). After 10 min of
vortex and 10 min of centrifugation at , supernatants were evaporated using a stream of nitrogen and reconstituted in (for mouse serum) (for human plasma) methanol:water [1:1]. of the samples were used for the bile acid analysis.
Caecal and faecal samples were extracted with methanol, containing the same deuterium-labeled standards as above ( of each). The samples were homogenized with ceramic beads for 10 min
using a Qiagen Tissuelyser II. After 10 min of centrifugation at supernatant was diluted with methanol:water 1:1.
UPLC-MS/MS conditions were according to previous work . Briefly, after injection the bile acids were separated on a C18 column ( ; Kinetex, Phenomenex, USA) using water with 7.5 mM ammonium acetate and formic acid (mobile phase A)
Fig. 3 | Increased levels of DCA in plasma from T2D individuals. A-D Plasma levels of total bile acids (A), DCA (B), isoDCA (C) and 12-epiDCA (D) in NGT and T2D individuals. E, F Faecal levels of total bile acids (E) and DCA (F) in NGT controls and T2D individuals. G, H Spearman’s correlation analysis between DCA levels in plasma and faeces ( ) and between DCA, LCA, isoUDCA, isoDCA, 12-epiDCA and 12oxoDCA in faeces or plasma and clinical parameters ( ). For ( ), mean with all points are plotted; samples/group; -values indicate differences between the NGT (blue color) and T2D (pink color) groups analyzed with two-tailed Mann-Whitney using Benjamini and Hochberg adjusted -values. For H , red color indicates positive correlation and blue color indicate negative correlation and are indicated using Benjamini and Hochberg adjusted -values. Chol cholesterol, CRP C-reactive protein, DBP diastolic blood pressure, DCA deoxycholic acid, Eos eosinophils, FBG fasting blood glucose, HbA1c Hemoglobin A1c, HDL high-density lipoprotein, HOMA-IR Homeostatic Model Assessment for Insulin Resistance, isoDCA iso-deoxycholic acid, isoUDCA iso-ursodeoxycholic acid, LCA lithocholic acid, LDL low-density lipoprotein, Lymph lymphocytes, Mono monocytes, NGT normal glucose tolerance, Neut neutrophils, OGTT 2 h BG blood glucose at 2 h after oral glucose tolerance test, SBP systolic blood pressure, T2D Type 2 diabetes, TG triglycerides, TyG triglyceride-glucose index, WBC white blood cells, 12 -epiDCA 12 -epideoxycholic acid, 12 -oxoDCA 12 -oxodeoxycholic acid.
and acetonitrile with formic acid (mobile phase B). The chromatographic separation started with isocratic separation at . The B-phase was then increased to during 4 min . During the next 10 min the B-phase was increased to and it was held at for 3.5 min before returning to . The total runtime was 20 min . Bile acids were detected using multiple reaction monitoring in negative mode on a QTRAP 5500 mass spectrometer (Sciex, Concord, Canada) and quantification was made against appropriate deuterated internal standards with adjustments by using external individual standard curves.
Liver function tests
Liver transaminases, alanine transaminase (ALT) and aspartate transaminase (AST) were measured in of serum from the mice. Analyses were performed at Clinical Chemistry Department, Sahlgrenska University Hospital, using the system Alinity ci 1303, software version 3.4.0 (Abbott laboratories).
Quantitative real-time PCR of liver samples
Approximately 30 mg of livers from colonized mice were homogenized using Tissuelyser II (Qiagen) and total RNA was isolated using RNeasy mini kit (Qiagen, 74106). High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, 4368813) was used to synthesize cDNA templates from 500 ng purified RNA using random hexamer primers, and the products were diluted 7 x before use in subsequent reactions. SYBR Green Supermix (Bio-Rad, 1708886) was used for qRT-PCR at final reaction volumes of gene-specific primers were used in each reaction and all gene expression data was normalized to the ribosomal protein L32 (L32) gene expression (forward: 5′-CCTCTGGTGA AGCCCAAGATC-3′; reverse: 5′-TCTGGGTTTCCGCCAGTTT-3′). Analyzed genes and primers were cholesterol -hydroxylase (Cyp7a1) (forward: -AGCAACTAAACAACCTGCCAGTACTA-3′; reverse: 5′-GTCCGGATATTCA AGGATGCA-3′) and sterol 12a-hydroxylase (Cyp8b1) (forward: 5′-GGCTGGCTTCCTGAGCTTATT-3′; reverse: 5′-ACTTCCTGAACAGCTCAT CGG-3′).
Liver triglyceride measurements
Approximately 50 mg of livers from colonized mice were homogenized in 2 ml polypropylene tubes loaded with 6 zirconium oxide beads ( 3 mm ) and then extracted using the BUME method . Analysis of liver triglycerides was performed on an aliquot of the total extract using straight phase high-performance liquid chromatography (HPLC) as described previously and quantification was made against an external calibration curve.
Short-chain fatty acid analysis of caecal samples using GC-MS
Caecum short-chain fatty acids were measured using GC-MS. of caecal content from colonized mice were mixed with internal standards, added to glass vials and freeze-dried. All samples were acidified with HCl , and short-chain fatty acids were extracted with two rounds of diethyl ether extraction. The organic supernatant was collected, the derivatization agent N-tert-butyldimethylsilyl-Nmethyltrifluoroacetamide (Sigma-Aldrich) was added, and samples were incubated at room temperature overnight and short-chain fatty
acids were quantified with a 7090A gas chromatograph coupled to a 5975C mass spectrometer (Agilent Technologies). Isotopically labeled standards of propionic acid and sodium salts of acetate and butyrate were obtained from Sigma-Aldrich (Stockholm, Sweden). Quantification was made using a one-point calibration against the corresponding isotopically labeled internal standard. The samples were injected into an Agilent Technologies 7890A gas chromatograph equipped with a DB-5MS Ultra Inert column ( , Agilent Technologies) operated in split mode (50:1). Temperature gradient started at for 3 min , changing to within 9.5 min followed by a . increase up to and holding for 3.5 min , giving a total runtime of 15.7 min per sample. Analytes were detected using a mass spectrometer (Agilent Technologies 5975C) in electron ionization (EI) mode. For each analyte the most intense ion or fragment was selected for quantification.
Statistical analyses
Differences between mice colonized with base community and base community + C. scindens were analyzed with Mann-Whitney for analysis of physiological parameters, and 2-way ANOVA with Šidák multiple comparisons test for analysis of the insulin tolerance test. Kruskal Wallis with Dunn’s multiple comparison test was used when more than two groups were compared. Differences between the NGT and T2D group were analyzed with Mann-Whitney using Benjamini and Hochberg adjusted -values. Spearman’s correlation analysis with Benjamini and Hochberg adjusted -values was used for correlation analysis. GraphPad Prism 9.5.0 was used for analysis of the in vivo data and R v4.3.2 was used for the analyses of the human data.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data supporting the findings of this study are available within the paper and its Supplementary Information. Raw data for bile acid analysis of the human samples are provided in a Source data file. Phenotype data for the human participants can be requested following standard protocol for data access to the corresponding author. Source data are provided with this paper.
References
Fromentin, S. et al. Microbiome and metabolome features of the cardiometabolic disease spectrum. Nat. Med. 28, 303-314 (2022).
Krautkramer, K. A., Fan, J. & Backhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 19, 77-94 (2021).
Ferrell, J. M. & Chiang, J. Y. L. Understanding bile acid signaling in diabetes: from pathophysiology to therapeutic targets. Diab. Metab. J. 43, 257-272 (2019).
Sayin, S. I. et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 17, 225-235 (2013).
Wahlstrom, A., Sayin, S. I., Marschall, H. U. & Backhed, F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metab. 24, 41-50 (2016).
Chiang, J. Y. L. & Ferrell, J. M. Bile acid receptors FXR and TGR5 signaling in fatty liver diseases and therapy. Am. J. Physiol. Gastrointest. Liver Physiol. 318, G554-G573 (2020).
Ridlon, J. M., Kang, D. J. & Hylemon, P. B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 47, 241-259 (2006).
Funabashi, M. et al. A metabolic pathway for bile acid dehydroxylation by the gut microbiome. Nature 582, 566-570 (2020).
Wells, J. E., Berr, F., Thomas, L. A., Dowling, R. H. & Hylemon, P. B. Isolation and characterization of cholic acid 7alpha-dehydroxylating fecal bacteria from cholesterol gallstone patients. J. Hepatol. 32, 4-10 (2000).
Ridlon, J. M., Kang, D. J., Hylemon, P. B. & Bajaj, J. S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 30, 332-338 (2014).
Ridlon, J. M. et al. The ‘in vivo lifestyle’ of bile acid 7alphadehydroxylating bacteria: comparative genomics, metatranscriptomic, and bile acid metabolomics analysis of a defined microbial community in gnotobiotic mice. Gut Microbes 11, 381-404 (2020).
Devlin, A. S. & Fischbach, M. A. A biosynthetic pathway for a prominent class of microbiota-derived bile acids. Nat. Chem. Biol. 11, 685-690 (2015).
Hofmann, A. F. et al. A proposed nomenclature for bile acids. J. Lipid Res. 33, 599-604 (1992).
Doden, H. L. et al. Completion of the gut microbial epi-bile acid pathway. Gut Microbes 13, 1-20 (2021).
Wu, M. et al. Genetic determinants of in vivo fitness and diet responsiveness in multiple human gut Bacteroides. Science 350, aac5992 (2015).
Marion, S. et al. In vitro and in vivo characterization of Clostridium scindens bile acid transformations. Gut Microbes 10, 481-503 (2019).
Studer, N. et al. Functional intestinal bile acid 7alphadehydroxylation by clostridium scindens associated with protection from Clostridium difficile infection in a gnotobiotic mouse model. Front. Cell Infect. Microbiol. 6, 191 (2016).
Marion, S. et al. Biogeography of microbial bile acid transformations along the murine gut. J. Lipid Res. 61, 1450-1463 (2020).
Song, P., Zhang, Y. & Klaassen, C. D. Dose-response of five bile acids on serum and liver bile Acid concentrations and hepatotoxicty in mice. Toxicol. Sci. 123, 359-367 (2011).
Hylemon, P. B., Harris, S. C. & Ridlon, J. M. Metabolism of hydrogen gases and bile acids in the gut microbiome. FEBS Lett. 592, 2070-2082 (2018).
Narushima, S., Itoh, K., Takamine, F. & Uchida, K. Absence of cecal secondary bile acids in gnotobiotic mice associated with two human intestinal bacteria with the ability to dehydroxylate bile acids in vitro. Microbiol. Immunol. 43, 893-897 (1999).
Narushima, S. et al. Deoxycholic acid formation in gnotobiotic mice associated with human intestinal bacteria. Lipids 41, 835-843 (2006).
Ridlon, J. M., Kang, D.-J. & Hylemon, P. B. Isolation and characterization of a bile acid inducible -dehydroxylating operon in Clostridium hylemonae TN271. Anaerobe 16, 137-146 (2010).
Khan, M. T. et al. Synergy and oxygen adaptation for development of next-generation probiotics. Nature 620, 381-385 (2023).
Zaborska, K. E., Lee, S. A., Garribay, D., Cha, E. & Cummings, B. P. Deoxycholic acid supplementation impairs glucose homeostasis in mice. PLoS ONE 13, e0200908 (2018).
Yoshimoto, S. et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 499, 97-101 (2013).
Chavez-Talavera, O. et al. Bile acids associate with glucose metabolism, but do not predict conversion from impaired fasting glucose to diabetes. Metabolism 103, 154042 (2020).
Haeusler, R. A., Astiarraga, B., Camastra, S., Accili, D. & Ferrannini, E. Human insulin resistance is associated with increased plasma levels of 12alpha-hydroxylated bile acids. Diabetes 62, 4184-4191 (2013).
Atlas R. M. Handbook of Microbiological Media, 3rd edition (2004).
Wu, H. et al. The gut microbiota in prediabetes and diabetes: a population-based cross-sectional study. Cell Metab. 32, 379-90 e3 (2020).
Bergstrom, G. et al. The Swedish CArdioPulmonary Biolmage Study: objectives and design. J. Intern. Med. 278, 645-659 (2015).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357-359 (2012).
Danecek, P. et al. Twelve years of SAMtools and BCFtools. Gigascience 10, giab008 (2021).
Lawson, A. M., Setchell, K. D. R. The Bile Acids: Chemistry, Physiology, and Metabolism: Volume 4: Methods and Applications (eds Setchell, K. D. R., Kritchevsky, D. & Nair, P. P.) 167-267 (Springer US, 1988).
Buffie, C. G. et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 517, 205-208 (2015).
Tremaroli, V. et al. Roux-en-Y gastric bypass and vertical banded gastroplasty induce long-term changes on the human gut microbiome contributing to fat mass regulation. Cell Metab. 22, 228-238 (2015).
Lofgren, L. et al. The BUME method: a novel automated chloroformfree 96-well total lipid extraction method for blood plasma. J. Lipid Res. 53, 1690-1700 (2012).
Homan, R. & Anderson, M. K. Rapid separation and quantitation of combined neutral and polar lipid classes by high-performance liquid chromatography and evaporative light-scattering mass detection. J. Chromatogr. B Biomed. Sci. Appl. 708, 21-26 (1998).
Acknowledgements
We thank Anna Hallén, Carina Arvidsson, and Louise Helldén for technical assistance and the Adam Arkin Lab for providing Eubacterium rectale ATCC 33656. This work was supported by grants from the Leducq Foundation (17CVD01), the Swedish Heart Lung Foundation (20210366), the Knut and Alice Wallenberg Foundation (2017.0026), the Swedish Research Council (2019-01599), AFA insurances (160337), the Novo Nordisk Foundation (NNF21OC0070298), and from the Swedish state under the agreement between the Swedish government and the county councils, the ALF-agreement (ALFGBG-718101). F.B. is a Torsten Söderberg Professor in Medicine and Wallenberg Scholar. S.L.H. and M.A.F. were also partially supported by National Institutes of Health grant P01 HL147823.
Author contributions
Conceptualization: A.W., A.B., M.A.F., and F.B. Clinical investigation: G.B. Methodology: A.W., A.B., L.O., W.S., A.L., and M.H. Formal analysis: A.W., L.O., H.W., and W.S. Visualization: A.W., A.B., and L.O. Resources: M.A.F., S.L.H., and F.B. Writing-original draft: A.W. Writing-review & editing: A.W., L.O., K.M., W.S., S.L.H., H.-U.M., M.A.F., and F.B.
Funding
Open access funding provided by University of Gothenburg.
Competing interests
Stanford University and the Chan Zuckerberg Biohub have patents pending for microbiome technologies on which A.B. and M.A.F. are coinventors. M.A.F. is a co-founder and director of Federation Bio and Kelonia, a co-founder of Revolution Medicines, and a member of the scientific advisory boards of NGM Bio and Zymergen. S.L.H. reports having received royalty payments for inventions or discoveries related to cardiovascular diagnostics or therapeutics from Cleveland Heart Lab, a fully owned subsidiary of Quest Diagnostics, and Procter & Gamble,
being a paid consultant for Zehna Therapeutics, and having received research funds from Procter & Gamble, Pfizer Inc., Roche Diagnostics and Zehna Therapeutics. F.B. is co-founder and shareholder of Roxbiosens Inc and Implexion Pharma AB, receives research funding from Biogaia AB, and a member of the scientific advisory board of Bactolife A/ S . The remaining authors declare no competing interests.
Correspondence and requests for materials should be addressed to Michael A. Fischbach or Fredrik Bäckhed.
Peer review information Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Wallenberg Laboratory and Department of Molecular and Clinical Medicine, Institute of Medicine, Sahlgrenska Academy, University of Gothenburg, Gothenburg, Sweden. Department of Bioengineering, Stanford University, Stanford, CA, USA. Department of Microbiology and Immunology, Stanford University School of Medicine, Stanford University, Stanford, CA, USA. ChEM-H Institute, Stanford University, Stanford, CA, USA. Chan Zuckerberg Biohub, San Francisco, CA, USA. State Key Laboratory of Genetic Engineering, Human Phenome Institute, Fudan Microbiome Center, and Department of Bariatric and Metabolic Surgery, Huashan Hospital, Fudan University, Shanghai, China. Department of Cardiovascular & Metabolic Sciences, Lerner Research Institute, Cleveland, OH, USA. Center for Microbiome and Human Health, Cleveland Clinic, Cleveland, OH, USA. Department of Cardiovascular Medicine, Heart, Vascular and Thoracic Institute, Cleveland Clinic, Cleveland, OH, USA. Region Västra Götaland, Sahlgrenska University Hospital, Department of Clinical Physiology, Gothenburg, Sweden. Novo Nordisk Foundation Center for Basic Metabolic Research, Faculty of Health Sciences, University of Copenhagen, Copenhagen, Denmark. Deceased: Hanns-Ulrich Marschall. e-mail: fischbach@fischbachgroup.org; Fredrik.Backhed@wlab.gu.se