اختبار مصيدة المعادن وتحسين المخططات لتصميم تميؤ البروتينات الخلوية بعناصر مختلفة A metal-trap tests and refines blueprints to engineer cellular protein metalation with different elements
لقد كان من الصعب اختبار كيفية اكتساب البروتينات للمعادن المحددة في الخلايا. يُعتقد أن نوعية التمعدن تعتمد على تفضيلات البروتينات للمعادن المختلفة التي تتنافس على توافر المعادن داخل الخلايا. وهذا يعني أن التمعدن الخاطئ قد يحدث إذا أصبحت البروتينات غير متطابقة مع توافر المعادن في الخلايا غير المتجانسة. هنا نستخدم بكتيريا زرقاء…-كوبينكفخ معدني، لاختبار توقعات التمثيل المعدني. من خلال إعادة طي MncA في معادن متنافسة مخففة، يتم تحديد تفضيلات المعادن. ربط تفضيلات المعادن بتوافر المعادن المقدرة باستخدام حساسات المعادن الخلوية، يتنبأ بسوء التمثيل المعدني لـ MncA معفي الإشريكية القولونية. بعد التعبير في. القولونية، بشكل رئيسيتم عزل MncA المرتبط تجريبيًا. من المتوقع أنه في الخلايا القابلة للحياة المدعمة بالمعادن يجب أن يتغير نوع MncA المعدني. – – ، أو يتم استردادها من خلايا المعادن المضافة المعنية. تُستخدم الفروق بين التخصص المعدني لمركب MncA الملاحظ والمتوقع لتحسين تقديرات توافر المعادن. تُقدم القيم كخطط إرشادية لتوجيه هندسة تميؤ البروتينات البيولوجية بالمعادن.
هدف هذا البحث هو اختبار التفسيرات حول كيفية اكتساب البروتينات معادن مختلفة في الخلايا: تنوع المعادن في البيولوجيا (الشكل التوضيحي التكميلي 1). إن تحفيز الإنزيمات المعدنية غالبًا ما يكون محددًا للمعادن. ومع ذلك، فإن البروتينات المعدنية عادةً ما ترتبط بأحد المعادن الخاطئة أو أكثر تفضيلًا على المعادن المتوافقة.تكون مواقع المعادن الناشئة في البروتينات مرنة بحيث يمكن أن ترتبط المعادن غير المتوافقة بشكل غير محافظ من خلال استخدام مجموعة فرعية من الروابط الأصلية، من خلال استقطاب روابط إضافية و/أو من خلال اعتماد هندسات تنسيق غير متوافقة.مع وجود قيود محدودة مثل هذه، فإن ترتيب ارتباط المعادن يتبع عادة سلسلة إيرفينغ-ويليامز (الشكل 1أ).استثناء هو عندما يكون هناك تنظيم هيكلي سابق، على سبيل المثال من خلال التعاون في مواقع ثنائية المعادن.. هنا نقوم بتحديد تفضيلات الارتباط لبروتين (MncA) الذي يحبس المعادن بشكل حركي أثناء الطيثم نستخدم MncA لتحديد ما إذا كان يمكن التنبؤ بشكل صحيح بعملية ميتاليشن البروتين، ثم تعديلها بشكل متوقع، وأخيرًا لتحسين تقديرات توفر المعادن داخل الخلايا.
بالإضافة إلى تفضيلات ربط المعادن للبروتينات، فإن العامل الثاني الذي يحدد تنوع المعادن والبروتينات هو توفر المعادن.يمكن أن يحدث تقسيم المعادن مباشرة من برك المعادن القابلة للتبادل والمتاحة إلى البروتين المعني، أو إلى مسار تجميع لمساعد جزيئي صغير (مثلإلى الهيم أوإلى فيتامين )، أو إلى شاحنة معدنية توصل إلى مسار التجميع أو إلى البروتين المعني . هنا نستكشف خطوة التقسيم الأولية من خزانات المعادن داخل الخلايا المتاحة وغير المستقرة.
أهمية توفر المعادن داخل الخلايا لتحديد الأنواع المعدنية-البروتينية ظهرت على مدى عدة عقود. عندما يتم التعبير عنها في خلية زرقاء-خضراء – و -مستشعر معدني مرتبط بالحمض النووي المستجيب من المتفطرة السلية استجاب فقط إلى. تم نسب هذا التغيير في التخصص إلى كونه غير متوفر بشكل كافٍ داخل السيانوبكتيريا القابلة للحياة لتكوين معقد مع NmtR. وبالمثل، اكتسب DtxR المستجيب لفيل من Corynebacterium diphtheriae استجابة لـ
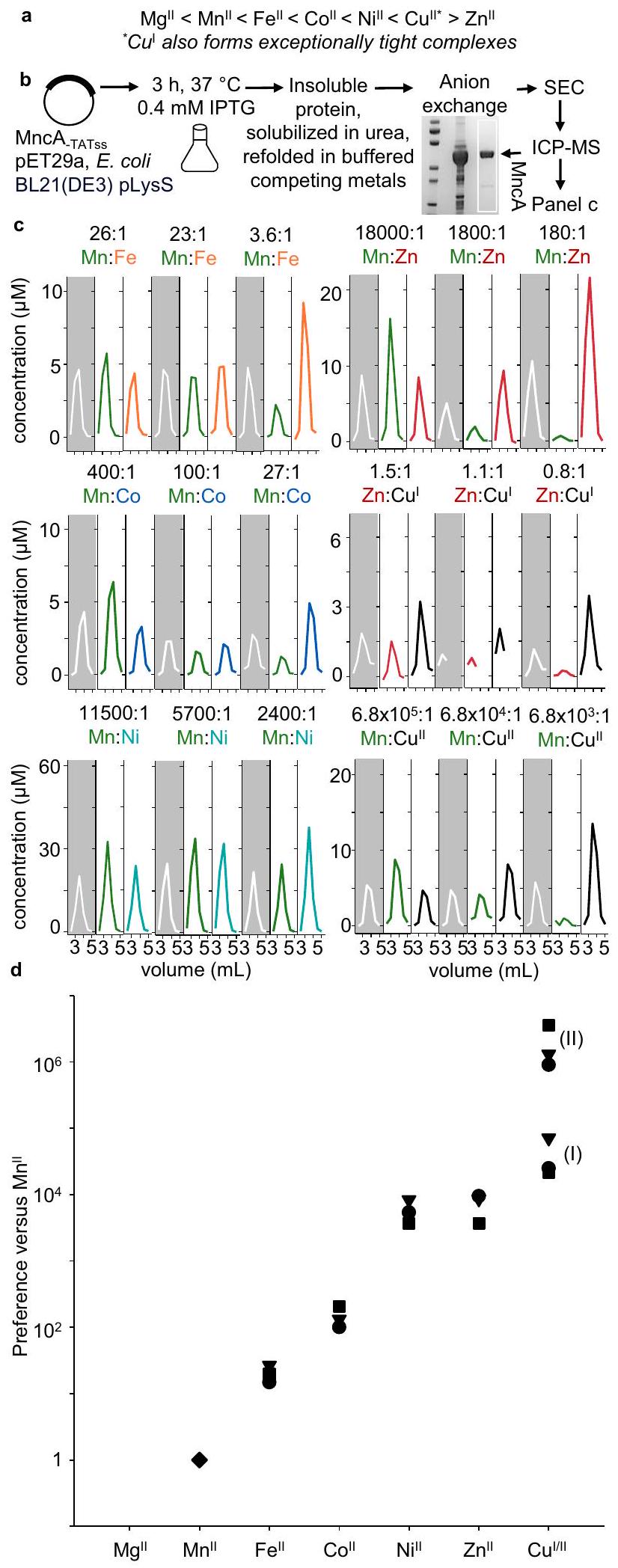
الشكل 1 | تفضيلات المعادن النسبية لـ MncA المطوية في المعادن المتنافسة المدعومة. أ ترتيب إيرفينغ-ويليامز لتكوين المعقدات مع المعادن، كحالات أيونية للمعادن القابلة للتبادل في السيتوسول. من الضعيف (يسار) إلى القوي، مع ملاحظة عكس السهم بعد النحاس. ب بروتوكول لاستعادة MncA غير المطوية من الأجسام الشاملة بعد التعبير بمستويات عالية في E. coli BL21(DE3) pLysS لمدة 3 ساعات باستخدام pET29a.في، تليها طي MncA المنكسر عن طريق التخفيف بالتنقيط في حجم كبير من العازلة الخالية من اليوريا التي تحتوي على أزواج من أيونات المعادن المتنافسة والمخزنة. تم تركيز MncA المطوي بواسطة تبادل الأنيون وتم إصداره باستخدام عازلة عالية الملح في خطوة واحدة. تم فصل MncA المرتبط بالمعادن عن المعادن غير المرتبطة بواسطة SEC (عمود PD-10)، وتم تحليل المعادن بواسطة ICP-MS. يظهر إدراج الجل، من اليسار، علامات الحجم (من الأعلى، )، تم تحميل MncA المذاب في اليوريا وMncA الذي تم إزالته من Q-Sepharose (الجل الكامل الشكل التكميلية 2a). ج محتويات المعادن في الفقرات التي تحتوي على MncA بعد الطي في فائض من المعادن المتنافسة المدعومة بنسب مختلفة من التوافرات (كما في اللوحة ، العوازل في الجدول التكميلي 1 التي تم إعدادها بمساعدة من البيانات التكميلية 1). نسب المعادن المتنافسة فوق كل لوحة. تركيز MncA يقدر منتقريب النسب الكيميائيةلكن تم حساب التمثيل المعدني النسبي من إجمالي المعدن كما هو موضح في الجدول 1. التجارب مع و تمت العملية في غرفة لاهوائية، وتم تحضير مخزونات المعادن حديثًا، واستخدمت هيدروكسيلامين (1 مللي مول) للحفاظ على النحاس المخفض. و تم التأكيد على أنهتم تقليلها من خلال التفاعل مع فائض من الفيروزين أو BCA على التوالي.تم عرض النسخ التجريبية المستقلة. د تم قياس تفضيلات MncA للمعادن الثنائية المختلفة، بالإضافة إلى النحاس الأحادي، بالنسبة لـ ( تظهر التجارب المستقلة، المثلثات، المربعات، والدوائر النسخ في (ج)، (I) و(II) تشير إلى أشكال النحاس)، التي تم تحديدها من ملفات الإلوت في (ج). يتبع ترتيب التفضيلات سلسلة إيرفينغ-ويليامز كما في (أ). يفضل MncA جميع المعادن غير المعروفة علىتُقدم بيانات المصدر كملف بيانات مصدر.
عند التعبير عنه في بكتيريا باسيلاس سوبتيليس، يُعزى مرة أخرى إلى اختلافات داخل الخلاياالتوافرات في الخلايا المختلفةاثنان من بروتينات ربط المعادن، و تم اكتشافها في الفضاء بين الغشاء الخلوي لسيانوبكتيريا (Synechocystis PCC 6803)، حيث ترتبط بشكل ملحوظ بالمعادن من نهايات مختلفة لسلسلة إيرفينغ-ويليامز، ومع ذلك تستغل طيات مشابهة وبقايا ربط المعادن المتطابقة.. عند الطي في المختبر بكميات مماثلة من النحاس المتنافس مقابل ، أو مقابل المعادن غير المتجانسة، النحاس أومربوط بـ. بينما يتم إفراز الكوبيرين النحاسي غير مطوي عبر نظام Sec، فإن -كوبين هو ركيزة تات تقوم بطي المعدن الأقل تنافسية واحتجازه حركيًا في السيتوسول قبل الإفراز. وبالتالي، فإن توفر المعدن في موقع طي البروتين يحدد نوع المعدن، ويجب أن تكون أكثر توفرًا من أو في السيتوسول البكتيري الأزرق. MncA هو ديكربوكسيلاز الأوكسالات المعتمد على (غير نشط مع النحاس أو )، و OxdC السيتوزولي المرتبط من بكتيريا Bacillus subtilis محاصر إما أو في الإشريكية القولونية اعتمادًا على إضافة المعادن إلى الوسطلذلك، تقدم MncA فرصًا مغرية لاستجواب الآليات والتنبؤات المتعلقة بالمعادن بشكل مباشر لأن (1) المعادن المحصورة حركيًا من غير المحتمل أن تتبادل أثناء التنقية، وبالتالي يجب أن تعكس MncA بدقة حالة المعادن في الجسم الحي، و(2) تشير السوابق إلى أنه قد يكون من الممكن تغيير نوع المعادن في MncA في E. coli واكتشاف ما إذا كانت حالات المعادن المختلفة قابلة للتنبؤ.
قمنا مؤخرًا بتطوير حاسبة للمعادن تأخذ في الاعتبار المنافسة بين المعادن داخل الخلايا.. تستخدم الحسابات تقديرات لتوافر المعادن داخل الخلايا مستمدة من استجابات الخلايا المنضبطة حرارياً لمُنظِّمات النسخ الحسية للمعادن المرتبطة بالحمض النووي (الشكل التوضيحي التكميلي 1) تُوصف التوافرات على أنها طاقات حرة متوافقة مع المعادن المرتبطة ولكن القابلة للتغيير والتي تتمتع بقدرة سريعة على تبادل الروابط مع البروتين المعني.من خلال استخدام تقديرات التوافر داخل الخلايا المعيارية عند نقاط منتصف نطاقات المستشعر (التي تمثل خلايا مثالية)، تم فك تشفير المعادن المتوافقة لأربعة بروتينات نموذجية بشكل صحيح.. تشجع هذه البيانات على وجهة نظر مفادها أن فهمنا لتنوع المعادن (بشكل عام) صحيح. ومع ذلك، على عكس الخلايا المثالية، ستكون أجهزة استشعار المعادن في مواقع مختلفة ضمن نطاقاتها في الخلايا الفعلية اعتمادًا على ظروف النمو. تم تقدير وفرة النسخ المترجمة التي تشفرها الجينات المنظمة بواسطة أجهزة استشعار المعادن بواسطة qPCR ثم تم معايرتها لتقدير توافر المعادن داخل إي. كولاي المزروعة في وسط غير مدعوم. حتى الآن، تم اختبار حسابات تنوع المعادن البروتينية تجريبيًا بشكل غير مباشر في بكتيريا الإشريكية القولونية المعدلة لتصنيع الفيتامين.. كوإنتاج يعتمد علىتم قياسه بالتالي كبديل لحالات المعدنة لـالمعدني الشابيرون كوب دبليو، وكوبنست. هذا العمل الآن تختبر التنبؤات مباشرة باستخدام MncA من السيانوبكتيريا لقراءة التمثيل المعدني في الكائنات الحية عند التعبير عنها فيخلايا الإشريكية القولونية تحت ظروف نمو مختلفة.
هنا نحدد ما إذا كانت حسابات التمثيل المعدني تتنبأ بشكل صحيح بالتمثيل المعدني الخاطئ الذي يُعتقد أنه يحدث في الخلايا المهندسة حيث تتعارض توافر المعادن مع تفضيلات المعادن للبروتينات المعبر عنها بشكل غير متجانس. في الخلايا المضافة إليها الكوبالت والنيكل والمنغنيز، تُستخدم الفروق المتبقية بين التمثيل المعدني المتوقع والملاحظ لـ MncA أيضًا لتحسين تقديرات توافر المعادن الأخرى، ونختبر ما إذا كانت التغيرات في ذرات المعادن في الخلايا…يتماشى مع هذه التحسينات. نستكشف الآلية التي يمكن من خلالها أن يؤثر التعرض لفلز واحد (مثل الكوبالت) على توفر فلز آخر (مثلتتضمن الآلات الحاسبة لتمكين استخدام MncA لاستكشاف توافر المعادن تحت ظروف نمو أخرى وفي أنواع خلايا أخرى. تُبذل جهود لتصميم بروتينات تتغلب على سلسلة إيرفينغ ويليامز.. تفرض هذه الأساليب قيودًا على، على سبيل المثال، المراكز الحفازة ثنائية المعدن. بالمقابل، هنا نوضح كيف يمكن استغلال البيولوجيا بطريقة يمكن التنبؤ بها للتغلب على التحدي الذي تقدمه سلسلة إيرفينغ-ويليامز، وبالتالي توسيع مجموعة التحفيز الحيوي المدفوع بالمعدن التي يمكن استخدامها بشكل يمكن التنبؤ به. تم توفير حاسبات لتوجيه تحسين تفاعل البروتين مع معادن مختلفة.
النتائج
Metal preferences at folding and trapping can also follow the Irving-Williams series
The first objective was to measure the metal-binding preferences of MncA in vitro. It is not feasible to measure affinities because is entrapped within the folded protein such that off-rates become negligible . Instead, relative preferences during folding were determined. Rapid, high-level, expression of MncA (minus secretion signal
peptide) in . coli produces MncA-containing inclusion bodies from which unfolded apo-protein can be recovered . MncA was thus expressed in E. coli BL21(DE3) pLysS, and urea-solubilised MncA, shown in Fig. 1b, was refolded by dilution into urea-free buffer. Refolding solutions contained pairs of competing metals buffered with NTA (or histidine for competitions) as in Supplementary Table 1, formulated via Supplementary Note 1 using the provided calculator (Supplementary Data 1). Competitions involving and were performed in an anaerobic chamber with -purged buffers and metal stocks confirmed > 95% reduced immediately prior to use. Metals were unbuffered in competitions between and . Refolded MncA (Fig. 1b), recovered by anion exchange chromatography, was resolved from unbound metal by size exclusion chromatography (SEC) with fractions ( 0.5 mL ) analysed for MncA by UV absorbance and metals by ICP-MS.
The proportion of each metal acquired by MncA was determined from the chromatograms in Fig. 1c. Challenges in generating NTAbuffered competitions initially led to competitions being performed without buffer before employing histidine-buffers. An extra replicate of histidine-buffered -competition was also performed (Supplementary Fig. 2, Supplementary Table 2). Competition between and bicinchoninic acid (BCA) buffered confirmed that MncA has -fold preference for , consistent with the determined -fold preference (Supplementary Fig. 2d, Table 1). Preferences of metal-binding to MncA at folding relative to were calculated from Table 1 to generate Fig. 1d. The order of binding follows the IrvingWilliams series (Fig. 1a). The exchangeable forms of metals in the cytosol are thought to be divalent except copper, which is monovalent. Fig. 1d illustrates the challenge to predict the metalation states of proteins in vivo and to decode cognate metals, since here seems least likely.
Table 1 | Preferences for metals trapped by MncA relative to
Metal1
Metal2
[Metal1] [Metal2]
Metal1 (%)
Metal2 (%)
Metal1/Metal2
Preference
Average preference
Fe”
56.7
43.3
1.31
Fe”
45.9
54.1
0.85
-50.8
Fe”
19.3
80.7
0.24
Co
20.1
79.9
0.25
Co
43.1
56.9
0.76
-55.6
Co
65.9
34.1
1.93
57.8
42.2
1.37
76.8
23.2
3.31
-65.3
Ni
39.3
60.7
0.65
51.3
48.7
1.06
Zn”
65.9
34.1
1.93
17.3
82.7
0.21
-65.5
3.1
96.9
0.03
Cu”
7.0
93
0.08
Cu
34.1
65.9
0.52
-79.1
Cu”
65.6
34.4
1.91
32.0
68.0
0.47
8.3
91.7
0.09
-69.5
29.5
70.5
0.42
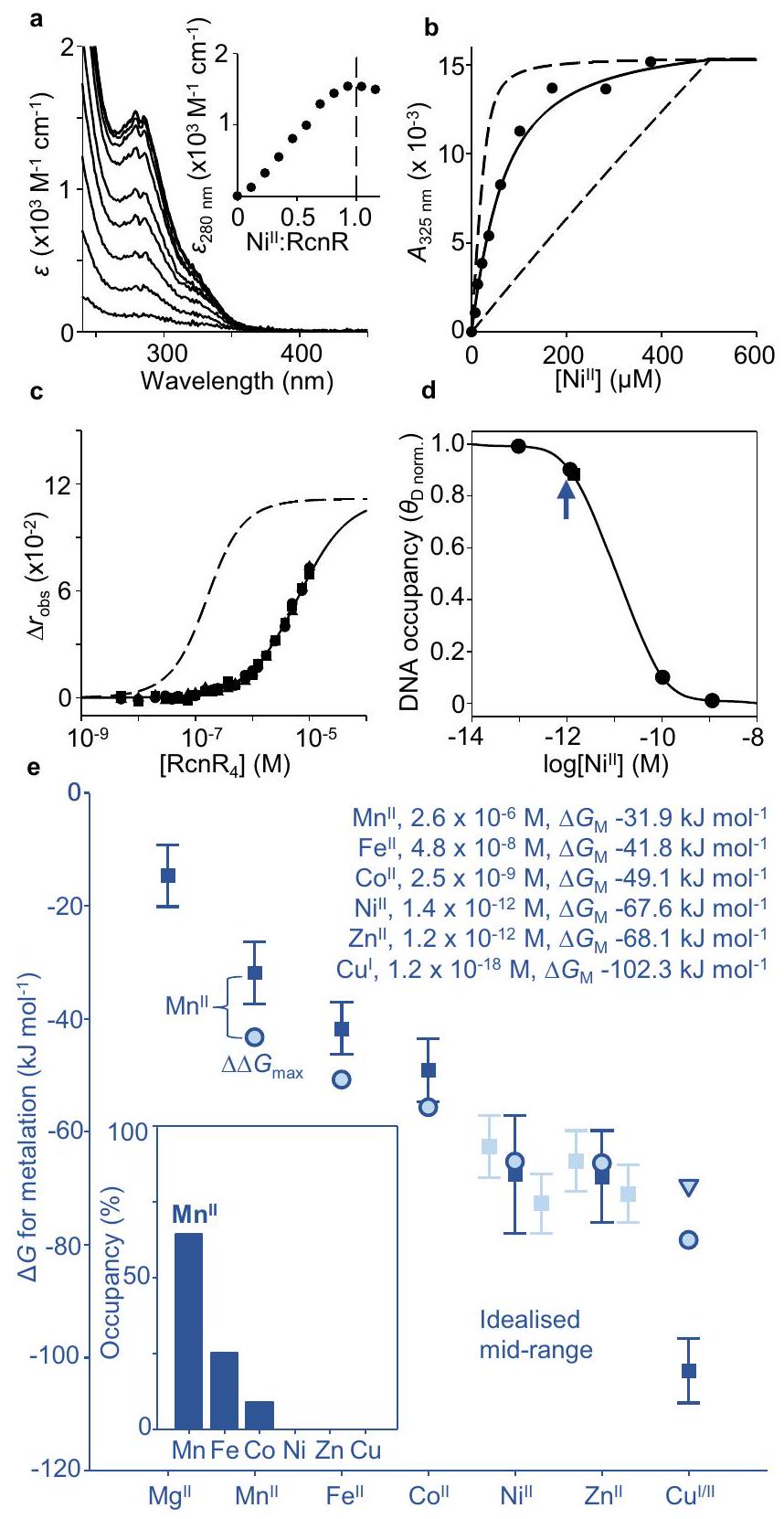
Fig. 2 | -RcnR refined mid-range metal availabilities decode as the cognate MncA metal. a Apo-subtracted difference spectra of RcnR ( RcnR monomer, calculated from total protein) titrated with , inset showing peak wavelength confirming 1:1 stoichiometry of to RcnR monomer, or 4:1 to RcnR4 ( ). RcnR ( monomer) also migrated with one equivalent of by SEC (Supplementary Fig. 3b). b Representative titration of RcnR ( monomer) in EGTA , solid line representing calculated for from simultaneously fitted RncR-EGTA competitions ( independent experiments) at varied EGTA concentrations (Supplementary Fig. 3c, fitting models in Supplementary Software). Dashed lines, simulations with affinities 10 -fold tighter and weaker than calculated . c RcnR binding to hexachlorofluorescein-labelled operator-promoter by fluorescence anisotropy. Solid line, best simultaneous fit to experimental replicates (circles, triangles, squares) for (fitting model in Supplementary Software), dashed line simulates apo-RcnR using published and maximum and DNA affinities determined above used with previously measured RcnR molecules cell to calculate (via Supplementary Data 2) relationship between intracellular availability and RcnR DNA occupancy , as for (circles show and 0.01 ). Combined mid-point of ranges for and also shown (blue arrow). e Metal availabilities (squares and inset text) as activities/concentrations and free energies, , at mid-points ( DNA occupancies, representing idealised cells) of metal sensors (bars are sensor ranges, to ), now including -RcnR. Pale bars show individual ranges where two cognate sensors. MncA metal preferences (pale blue circles, triangles, from Fig. 1d) as free energies ( ) from pseudoaffinities giving metalation at mid-range availability ( ) without competing metals (Supplementary Fig. 5 simulates alternative values for . Inset shows occupancies of MncA predicted from free energy differences between MncA and labile metal ( ). has the largest favourable gradient annotated . Supplementary Data 3 enables similar predictions of cognate metals for other proteins. Source data are provided as a Source Data file.
التوافرات المحددة بواسطة-تحسين توافر المعادن في النطاق المتوسط لـ RcnR
تقدم الشكل 1d تفضيلات المعادن لـ MncA بعد أخذ المنافسة من روابط أخرى (NTA أو الهيستيدين) في الاعتبار. يُعتقد أن تصنيف المعادن في البروتينات في السيتوسول المزدحم ناتج أيضًا عن المنافسة مع روابط متنوعة ترتبط بالمعادن القابلة للتبادل والمتاحة بمستويات مختلفة.تم تقدير توافر المعادن في السيتوسول لخلايا البكتيريا (السالمونيلا والإشريكية القولونية) كطاقات حرة ( ): الطاقات الحرة الرسمية لتكوين المركبات مع رافعة نصف معدنية افتراضية عند التوافر المعنيتكتشف حساسات المعادن المرتبطة بالحمض النووي التغيرات فيوقد تم ضبط استجاباتهم لهذه البكتيريا. ومع ذلك، الـاستجابات عاليةتم تجاهل RcnR الحساس سابقًا، ويتطلب معايرته تحديدالألفة زائد الألفة الحمض النووي-RcnR (الشكل التوضيحي 3a).
تغيرات امتصاص RcnR معويشبع طيف الفرق عند مكافئ واحد لكل مونومر، أو (الشكل 2أ). يتزامن RcnR مع واحد ذرة لكل مونومر بواسطة SEC (الشكل التكميلي 3b). EGTA يتنافس علىمع تمكين RcnR للتحديد عدم وجود ألفة RcnRمن ملاءمة متزامنة إلىالتعادل في تركيزات مختلفة من EGTA (الشكل التكميلي 3c). تؤكد مجموعة بيانات تمثيلية أن القيمة الملائمة تقع ضمن حدود الاختبار من الخطوط المتقطعة التي تحاكي القيم التي تكون أكثر تماسكًا وضعفًا بعشر مرات (الشكل 2b). ارتباط الأبو- وتمت مراقبة الحمض النووي سابقًا بواسطة الاستقطاب الفلوري باستخدام فلوريسئين سداسي الكلور.شظايا المشغل-المروج، لتحديد تقارب الحمض النووي. هنا تؤكد المعايرات المماثلة أن يضعف بشكل مشابه ارتباط الحمض النووي مع ملاءمة ثابتة قدرها 3.09 (الشكل 2ج). باستخدام هذه القيم، مع affinity DNA السابق لـ apo-RcnR من بالإضافة إلى جزيئات RcnR المعروفة في الخليةالعلاقة بين داخل الخليةالتوفر وتم حساب شغل المشغل المشغل كما هو لـ (البيانات التكميلية 2) تزداد نسخ rcnA مع انخفاض تواجد RcnR على الحمض النووي مع ارتفاع مستوى الداخل الخلويوفقًا للعلاقة في الشكل 2د. توافر المعادن عند إشغالات الحمض النوويو 0.01 موضحة. العلاقات المماثلة لمستشعرات المعادن الأخرى موجودة في الشكل التكميلي 4. تُظهر الأعمدة في الشكل 2e نطاقات التوفر (كـ ) التي تتوافق مع إشغالات . لالنطاق في الشكل 2e يجمع الآن بين تلك لـ-نيك آر و-RcnR. يتم عرض نقطة المنتصف للنطاق المدمج جنبًا إلى جنب مع نقاط المنتصف للمعادن الأخرى، مع توضيحها كتركيزات (M) وطاقة حرة (kJ mol-1) (الشكل 2e). سلسلة إيرفينغ-ويليامز غير واضحة بشأن ترتيب ضد لكن كلاهما أضعف من النحاس (الشكل 1أ). بما في ذلكRcnR، الشكل 2e يعكس ترتيب التوافر الداخلي للزنك. و الموضح في النسخ السابقةلكن لا تزال المستشعرات تحافظ على التوافرات لعكس سلسلة إيرفينغ-ويليامز.
-RcnR تم تحسين توفرات المعادن المتوسطة النطاق لفك تشفير التمعدن الصحيح
يجب أن يكون التفاعل المعدني داخل الخلية قابلاً للتنبؤ من تفضيلات الربط لـ MncA في الشكل 1d بالنسبة لمدى ارتباط المعادن الداخلية المتاحة (القابلة للتبادل) بإحكام، على سبيل المثال عند نقاط المنتصف لنطاقات المستشعر (الشكل 2e). لإجراء هذه المقارنة،تم تعيين انتماء زائف (إعطاء-معدنة
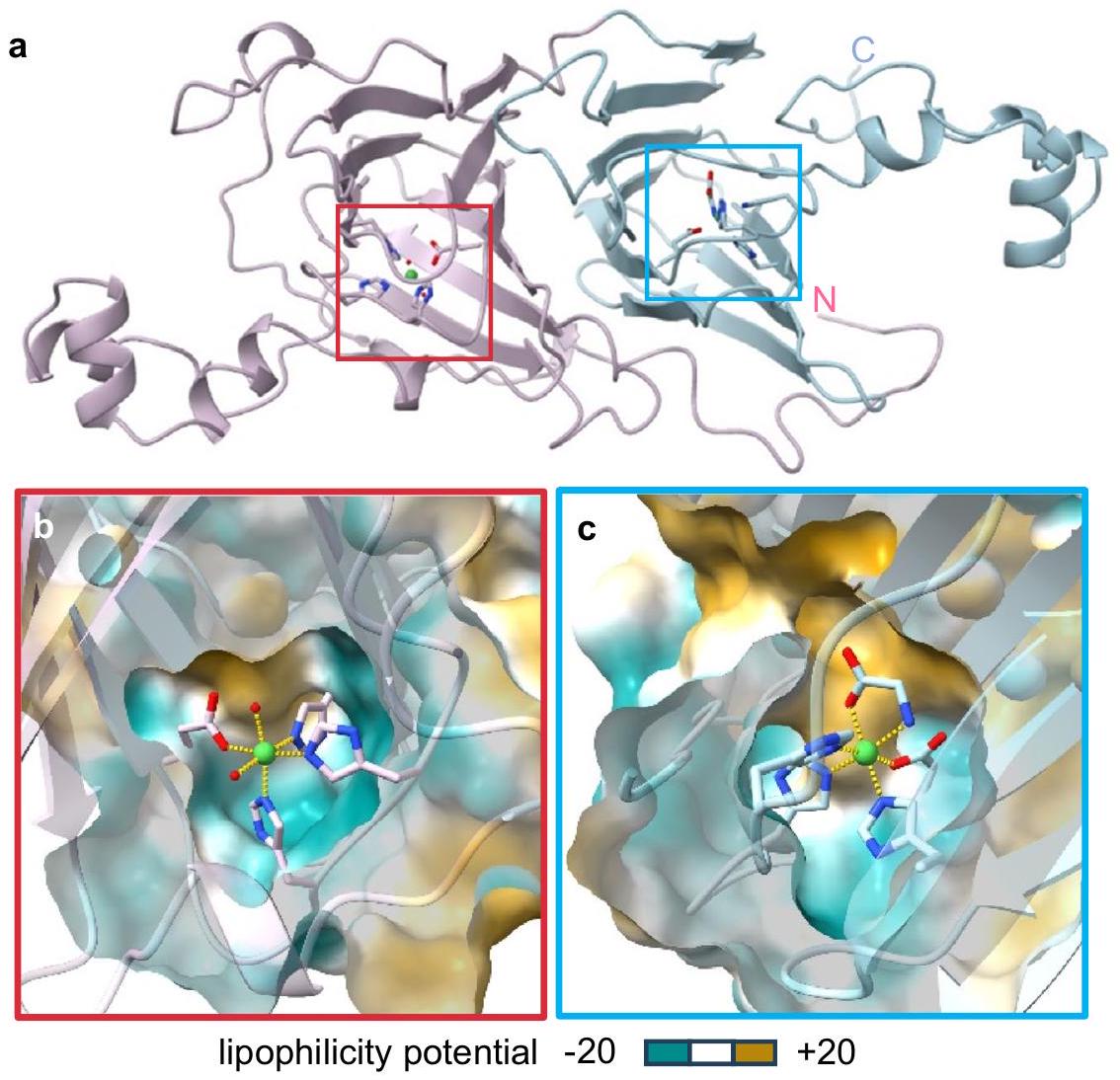
الشكل 3 |-تشير بنية MncA إلى أن المعدن غير المتوافق محاصر أيضًا حركيًا. تمثيل شريطي لبنية البلورة لـفيالقرار يظهر الطي الثنائي الكوبين المميز ومواقع المعادن (المحددة بإطار) تم نمذجتها من البقايا 39 من تسلسل البروتين الكامل بما في ذلك ببتيد الإشارة الذي لا يوجد في البروتين المعبر عنه (إحصائيات جمع البيانات والتنقيح موضحة في الجدول التكميلي 3). المجال الأميني (باللون الوردي)، المجال الكربوكسي (باللون الأزرق) من البقايا 238 (في التسلسل الكامل). ب، ج مقاطع عرضية لأسطح الوصول إلى المذيب (تم نمذجتها عندنصف قطر المذيب ليشمل الديناميات) المناطق المعدنية المحيطة لا تظهر قناة إلى الطرف الأمينيوقناة ضيقة محبة للدهون (كارهة للماء) إلى الطرف الكربوكسي.. يصبح محاصرًا في البروتين المطوي في هندسة قريبة من الأوكتاهيدرال (روابط صفراء؛ الشكل التوضيحي 6) مماثل لـيظهر نموذج MncA احتجازًا حركيًا للمعادن غير المتوافقة مما يشير إلى أن MncA قد يُستخدم للإبلاغ بدقة عن التمثيل المعدني داخل الخلايا. توضح الشكل التكميلية 7 أنه بعد الطيلم يتبادل معفي المختبر. فينقطة المنتصف لـ، وتم تحديد الطاقات الحرة لتكوين معقد المعدن-MncA. القيم ( ) للمعادن الأخرى تم حسابها بعد ذلك باستخدام الشكل 1d (الجدول 1). التدرجات من المواقع السيتوسولية القابلة للتبادل إلى تم حسابها وتم أخذ المنافسة بين المعادن في الاعتبار باستخدام آلة حاسبة Nill-RcnR المنقحة في البيانات التكميلية 3، للتنبؤ بالاحتلالات (الشكل 2e). المعدن المقابلتم فك تشفيره بشكل صحيح (الأكبر المواتي ). تم توليد تآلفات زائفة أكثر أو أقل بمعدل عشرة أضعاف بنفس النسب المئوية للاحتلال (الشكل التوضيحي 5a، b). وبالتالي، فإن تنوع المعادن هو وظيفة من تفضيلات ربط المعادن النسبية وتوافرها، ولكن إجمالي احتلال المعادن يختلف مع القيم المطلقة.قيم لـ. استخدام -RcnR- تم تعديل توافر النقاط الوسطى مع أربعة بروتينات نموذجية تم قياس تقاربهايفكك المعادن المماثلة لها على الرغم من تفضيلها جميعًا للنحاس (الشكل التوضيحي الإضافي 5c-f).-يمكن استخدام الآلة الحاسبة المعدلة RcnR في البيانات التكميلية 3 لفك تشفير التمثيل المعدني المتعلق.
المعدن غير المتجانس محاصر أيضًا حركيًا بواسطة MncA
الخصائص المتغيرة للمعادن المرتبطة بالبروتين تخلق تحديًا لتعريف حالات التمثيل المعدني للبروتينات المعدنية داخل الخلايا. يمكن فقدان المعادن أو اكتسابها أو تبادلها عند تحلل الخلايا و/أو أثناء التنقية والتحليل. يعتبر MncA جذابًا لأنيصبح محاصرًا في البروتين المطوي. ومع ذلك، قد يتبنى MncA طيات غير أصلية مع معادن غير متوافقة. قد لا تكون المعادن المنسقة بشكل غير طبيعي محاصر حركيًا.غالبًا ما يفضل الهندسات المسطحة ذات الأربعة تنسيقات، وتم اختيارها للتحليل الهيكلي. هيكل بلوري لـ ( القرار، الجدول التكميلي 3، الشكل التكميلي 6أ)، يظهر طية البي-كوبين لـمع ذرة معدنية في كل مجال كوبين (الشكل 3أ). كل الذرة مرتبطة بثلاثة بقايا هيستيدين وبقايا واحدة من الغلوتامات مع روابط غير بروتينية تكمل التنسيق السداسي.مجالات التنسيق.-MncA لديه بيئات تنسيق مشابهة بدون قناة إلى الطرف الأمينيوقناة ضيقة إلى الموقع الطرفي الكربوكسي مشابهة لـ (الشكل 3ب، ج) من المفترض أن القناة تسمح بالوصول إلى الركيزة للتفاعل التحفيزي. تم احتلال موقع الركيزة بشكل مجاني بواسطة الأسيتات في البنية ومع الجلايسين في الحالة المتبلورة-شكل (الشكل التوضيحي الإضافي 6، الشكل 3c). من غير المحتمل أن يسمح القناة الهيدروفوبية الضيقةالتبادل وبالتالي كلاهماتبدو الذرات محاصرة.
MncA لديهتفضيل مضاعف لـ فوق عند الطي (الشكل 1د). لاختبار ما إذا كان محاصر،-MncA تم حضنه لمدة 24 ساعة في فائض مولاري مزدوج من، تم فصل المعدن المرتبط والمعدن الحر بواسطة SEC تلاه ICP-MS (الشكل التوضيحي 7). ظل البروتين مرتبطًا حصريًا بـتأكيدمحاصر حركياً.
تعديل غير صحيح لمعدن MncA معالمتوقع والملاحظ في الإشريكية القولونية
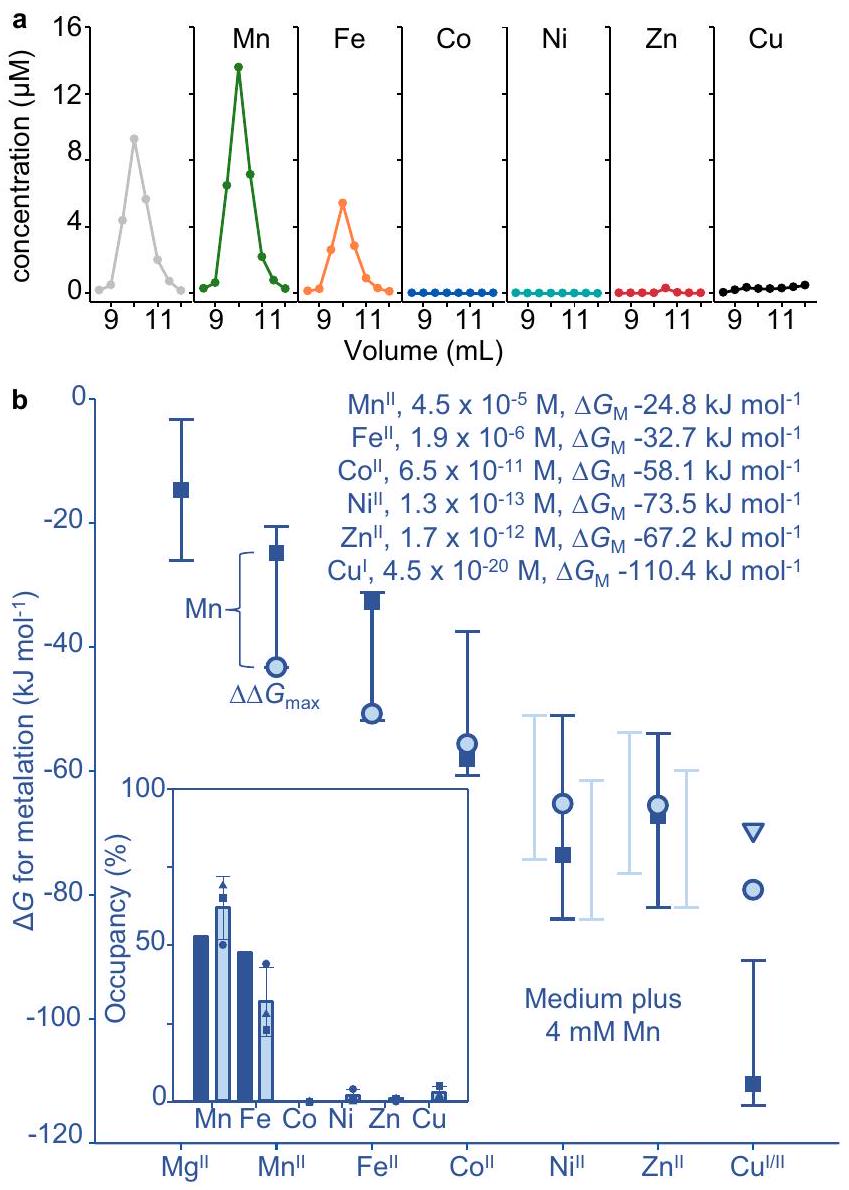
الشكل 4 (على عكس الشكل 2e) يظهر تقديرات توافر المعادن في الواقع. كولاي BW25113 (في مكان آخر. القولونية) تنمو هوائيًا، وتكوين المعدن تم قياس النسخ التي تنظمها حساسات المعادن سابقًا بواسطة qPCR، ثم تم ربطها بإشغال المحفزات وبالتالي توافر المعادن من خلال العلاقات في الشكل التوضيحي الإضافي.بشكل ملحوظ، داخل الخلاياالتوافر في الخلايا المزروعة هوائياً أقل من النطاق لـ – RcnR . أكبر تدرج إيجابي من المعدن القابل للتبادل المتاح إلى ، هو لـ غير متشابه (الشكل 4ج).
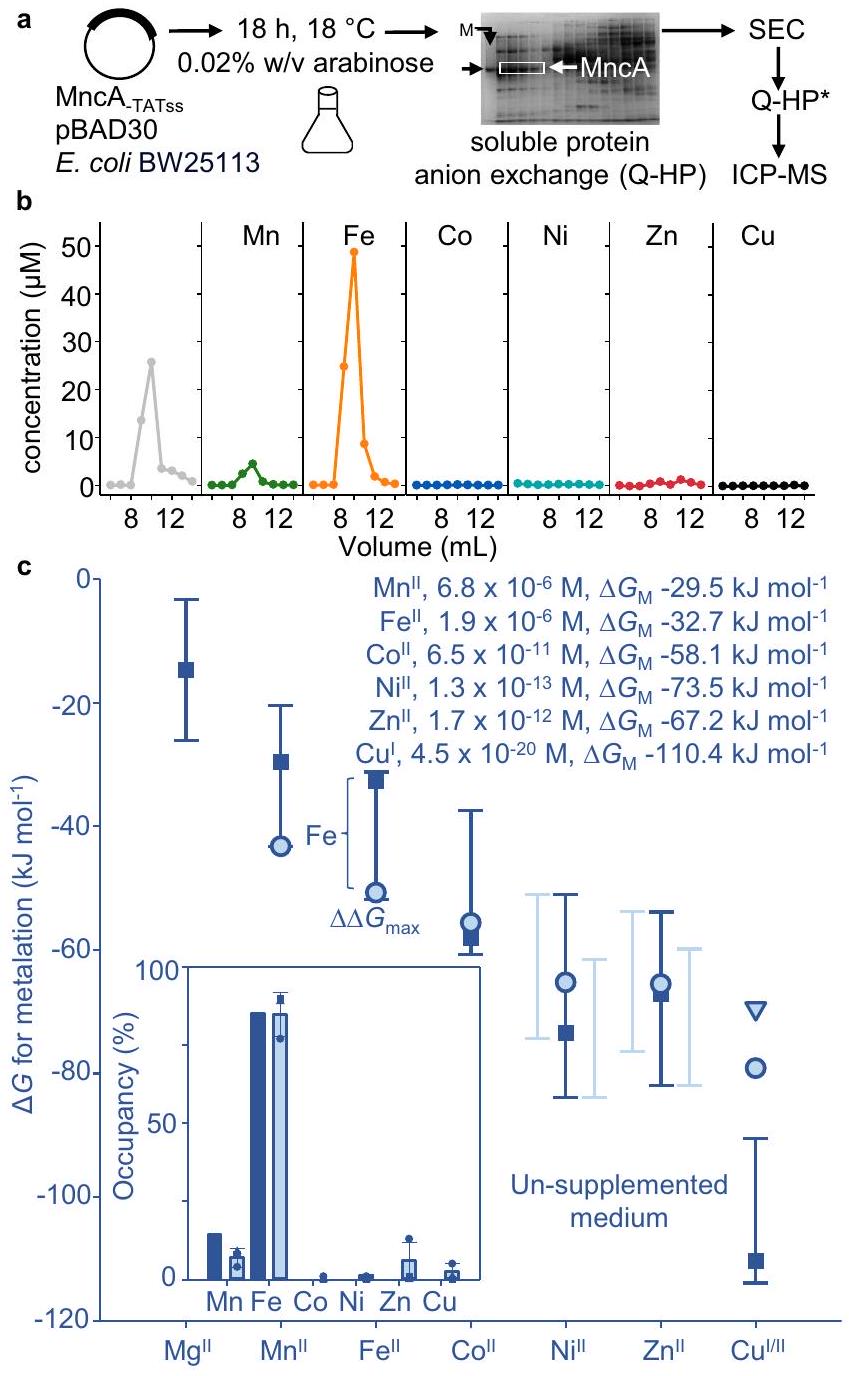
يمكن أن يؤدي التعبير عن البروتينات بمستويات عالية في خلايا العائل غير المتجانسة إلى استنفاد العوامل المساعدة، ومن المحتمل أن يسهم في تكوين أجسام الإدراج المحتوية على MncA (الشكل 1b). لمراقبة التمثيل المعدني داخل الخلايا، تم التعبير عن MncA بمستوى منخفض من محفز قابل للتعديل باستخدام محفز منخفض، ودرجة حرارة منخفضة، ونمو بطيء مطول طوال الليل في E. coli (الشكل 4a). تم إثراء MncA من مستخلص بروتين قابل للذوبان عبر تبادل الأنيونات وSEC تلاه ICP-MS. تم قياس التمثيل المعدني في ثلاث تجارب بيولوجية مستقلة (الشكل التكميلي 8، الجدول التكميلي 4a، b). تشير الامتصاصية عند 280 نانومتر إلى تركيز MncA بشكل كبير بسبب معامل الانقراض العالي له.، وهو واضح من النسبة الاستوكيومترية التي تقترب من 2:1، مما يشير إلى عدم وجود بروتين غير نشط (الجدول التكميلي 4أ، ب). قد تؤثر بعض المعادن على معامل الانقراض وقد تتسبب آثار البروتينات المتداخلة في إدخال تباين بين التجارب. وبالتالي، تم حساب النسب المئوية من إجمالي المعدن في واحدة أو أكثر من الفصائل المحتوية على MncA بدلاً من تركيز MncA، ولا سيما مع نتائج مشابهة عند التكرار (الجدول التكميلي 4ب). تم تكرار التجربة باستخدام خطوة تبادل أنيون واحدة وHPLC SEC التحليلية مع نتائج مشابهة (الجدول التكميلي 4). يتم عرض متوسط ± الانحراف المعياري لتخصص MncA (الإطار في الشكل 4ج). تم حساب التخصص الذي يأخذ في الاعتبار المنافسة بين المعادن باستخدام البيانات التكميلية 4. تم التنبؤ بـيتطابق التعديل الخاطئ للمعادن بشكل وثيق مع التعديل الخاطئ للمعادن الملحوظ في الإشريكية القولونية (الشكل 4ب، ج).
تم التنبؤ والملاحظة بوجود تفاعل معدنة موازٍ كبير لمركب MncAمنغنيز
كنا متحمسين لمعرفة ما إذا كانت داخل الخلايايمكن أن تزداد إلى توفر مقبول داخل قابل للحياة. القولونية، بما يكفي لتشكيل بشكل رئيسي الشكل 4c يوضح تفضيلات المعادن ( ) تتبع اتجاهًا مشابهًا للتوافرات ( )، متوقعًا أن التغييرات المتواضعة يمكن أن تحول نوعية التمثيل المعدني. باستخدام فقط مكملات المنغنيز، الحد الأقصى داخل الخلاياتم الكشف عن التوافر في الوسائط بتركيز 4 مللي مول من المنغنيزمن الجدير بالذكر أن المنغنيز العالي وبيروكسيد الهيدروجين حددا حدود المستشعر 0.99 (الشكل التكميلي 4)لأن استيراد المنغنيز يتم تنظيمه بواسطة. هنا، لم يمنع 4 مللي مول من المنغنيز كثافة الخلايا بعد زراعة طويلة الأمد (الشكل التوضيحي 9). تحول إلى السائد تمت ملاحظة التمثيل المعدني في ثلاث ثقافات مستقلة من E. coli تم تزويدها بـ 4 مللي مول من المنغنيز (الشكل 5أ، الشكل 5ب في الإطار، الجدول التكميلي 5أ، ب، الشكل التكميلي 10).
معايروفرة النسخ في 4 مللي مول من المنغنيز تعكس التوافر داخل الخلايا لـ (الشكل التوضيحي 4) . يتم عرض هذه القيمة في الشكل 5b بالإضافة إلى توافر المعادن الأخرى في الوسائط غير المدعمة. أكبر تدرج طاقة حرة مواتية من المعادن المتاحة القابلة للتبادل إلى MncA يصبح (الشكل 5ب). تم التنبؤ بالمعادن من خلال استبدال هذا الارتفاعالتوافر في البيانات التكميلية 4 (في الشكل 5ب). التحول إلى التمثيل المعدني السائد مع النظير بالإضافة إلى التعديل الجزئي غير الصحيح للمعادن مع وبالتالي يتم التنبؤ به، كما تم ملاحظته من خلال احتجاز MncA، في . القولونية المدعمة بـ 4 مللي مول من المنغنيز.
المعادن المحصورة في MncA تؤكد عدم وجود تأثير يذكر أو المعدنة في الخلايا المدعمة
تساءلنا أيضًا عما إذا كان يمكن أن يتم معدنته بشكل متوقع مع عناصر أخرى في بكتيريا الإشريكية القولونية القابلة للحياة المدعمة بالمعادن. توافرات و عند حدود المستشعر العليا (0.99) (مكمل)
الشكل 4 | التعديل الخاطئ بالمعادن معتم التنبؤ به وملاحظته في الإشريكية القولونية غير المتجانسة. تم تنقية MncA القابلة للذوبان والمطوية في الجسم الحي، بدون إشارة الإفراز، عن طريق كروماتوغرافيا تبادل الأنيون (5 مل Q-Sepharose، و1 مل Q-Sepharose) تم إزالتها باستخدام محلول ملحي عالي، مع SEC (Superdex 200 أو 75) بينهما. تم استرداد MncA من جزء البروتين القابل للذوبان من. كولاي بعد التعبير المنخفض المستوى طوال الليل فياستخدام pBAD30-مُستحث بـالعربينوز. ممثل لـتنقيات مستقلة بيولوجيًا. صورة الجل الكاملة في الشكل التكميلية 8a. ب تمثيلي (النسخ البيولوجية المستقلة) كروماتوغرام يظهر المعادن المكتسبة داخل الخلايا في الفractions المحتوية على MncA التي تم تحديدها بواسطة ICP-MS تكشف عن التمثيل الخاطئ للمعادن مع وآثار تم تأكيد الفractions المحتوية على MncA بواسطة SDS-PAGE (الشكل التوضيحي 8d) وتم قياسها بواسطة (استخدام ). تم استخدام محتويات المعادن في الفractions التي تحتوي على MncA (الجدول التكميلي 4a، b، الشكل التكميلي 8) لحساب النسب المئوية للاحتلال (%). يُظهر الجدول التكميلي 4 نتائج مشابهة من خلال بروتوكول معدل كما تم استخدامه في التجارب اللاحقة. ج توفر المعادن ( ، المربعات والنص المدخل) في السيتوسول من. القولونية التي نمت هوائيًا في LB، تم تقديرها كما في فוסטר وآخرون.عن طريق qPCR المعاير مع الجينات المنظمة بواسطة مستشعرات المعادن المتوافقة (الأشرطة هي نطاقات المستشعر منإلىتفضيلات المعادن MncA“دوائر زرقاء باهتة”، (مثلث). يظهر الإطار الداخلي إشغالات MncA المتوقعة من الفروق في الطاقة الحرة باستخدام البيانات التكميلية 4. يظهر الإطار الداخلي نتيجة التنبؤ بتلوث MncA بالمعادن. (الأعمدة الزرقاء الداكنة) في المضيف غير المتجانس بناءً على أكبر تدرج ملائم ( في الشكل الرئيسي). كما يظهر الشكل الفرعي المتوسط ( التعديل المعدني داخل الخلايا منالتكرارات البيولوجية (الأعمدة الزرقاء الفاتحة، المربعات، الدوائر، المثلثات، الجدول التكميلي 4ب)، تتطابق بشكل وثيق مع التوقعات. تم توفير بيانات المصدر كملف بيانات مصدر.
الشكل 5 | تم التنبؤ بتفاعل المعادن المماثلة الكبير وتأكيده في 4 مللي مول من المنغنيز. أ تمثيلي (النسخ البيولوجية المستقلة) كروماتوغرام يظهر المعادن المكتسبة داخل الخلايا في الفractions المحتوية على MncA التي تم تحديدها بواسطة ICP-MS، كما في الشكل 4a و b ولكن باستخدام خطوة تبادل أنيون واحدة (Q-Sepharose)، HPLC SEC (TSK SW3000) ويظهر زيادة في التمعدن المتعلق بـ“. تم تحديد MncA بواسطة SDS-PAGE في الشكل التوضيحي 10c. تم استخدام محتويات المعادن في الفقرات التي تحتوي على MncA (الجدول التوضيحي 5) من النسخ البيولوجية المستقلة (الشكل التوضيحي 10) لحساب النسب المئوية للاحتلال (%). ب توفر المعادن (، المربعات، والنص المدخل) في سيتوسول الإشريكية القولونية المزروعة هوائياً كما في الشكل 4c، باستثناءاستُبدل بتقديرات من qPCR المعايرة لهدف MntRفي الخلايا المزروعة في 4 مللي مول من المنغنيز (الشكل التوضيحي 4). تفضيلات المعادن لـ MncA كـدوائر زرقاء باهتة،، مثلث). القضبان هي نطاقات المستشعر منإلى. يظهر الشكل المرفق إشغالات MncA المتوقعة من تدرجات الطاقة الحرة باستخدام البيانات التكميلية 4 والاستبدالالتوفر بقيمة (كما في النص المدخل). يُظهر المدخل توقعات تميؤ ميتال MncA (الأعمدة الزرقاء الداكنة) مع استنادًا إلى أكبر تدرج ملائمفي الشكل الرئيسي)، بالإضافة إلى التمثيل الجزئي معيوضح الشكل الداخلي أيضًا المتوسط (± الانحراف المعياري) لتكوين المعادن داخل الخلايا منتكرارات بيولوجية مستقلة (مربعات، دوائر، مثلثات، أعمدة زرقاء باهتة، كما هو موضح في الجدول التكميلي 5)، تتطابق إلى حد كبير مع التوقعات. تم توفير بيانات المصدر كملف بيانات مصدر.
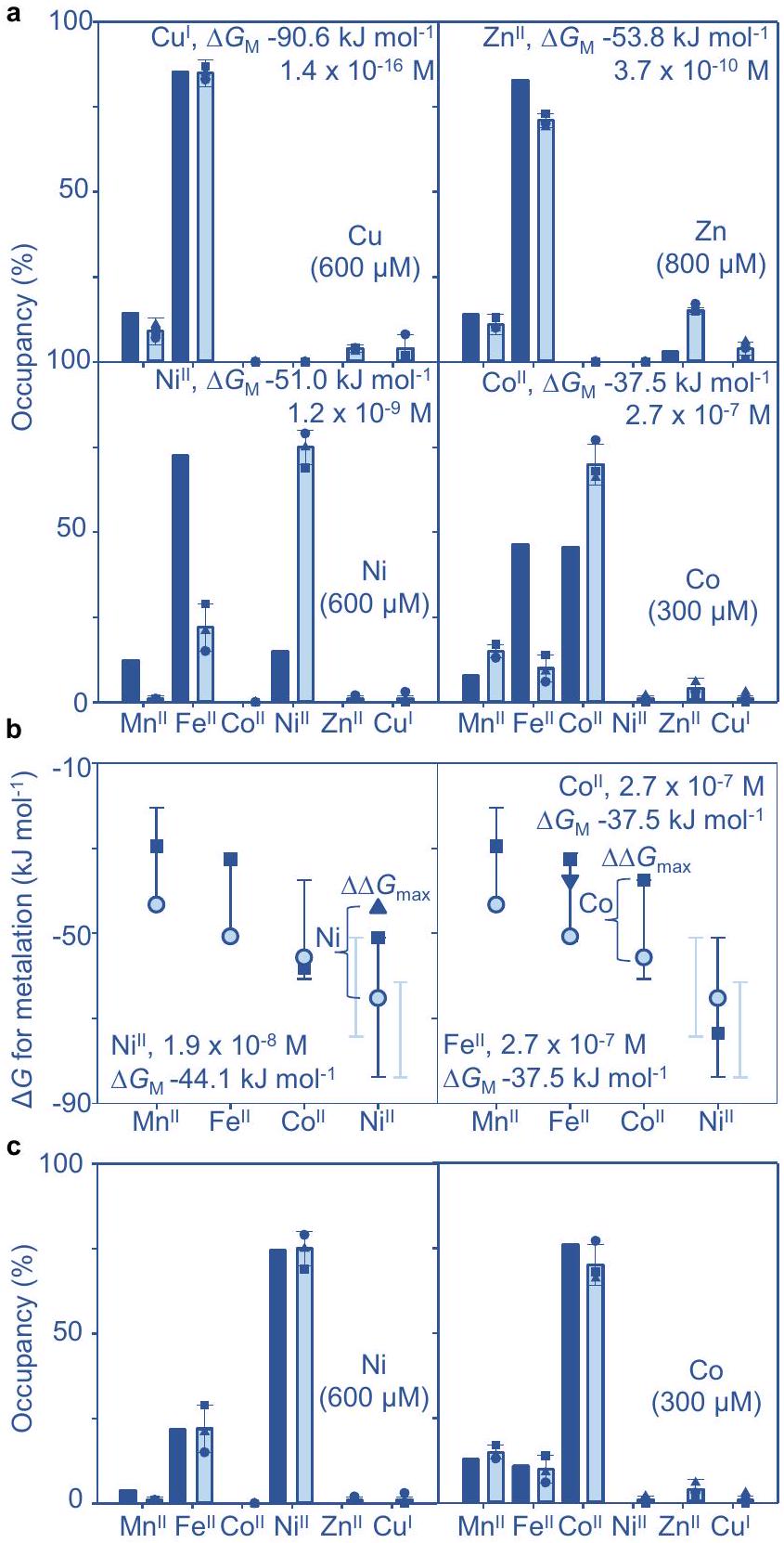
الشكل 4، كما هو موضح في الشكل 6أ، تم إدخالها في حاسبة التمثيل المعدني في البيانات التكميلية 4 كما في السابق. كانت نسبة الإشغال ضئيلة أو معدومة مع أي من المعدنين، ولكنتم التنبؤ بـ ‘الميس ميتالاسيون’ (الشكل 6أ).
توقعات بحدوث تفاعل غير صحيح مع المعدنغير ملحوظ ) لوحظ في الخلايا المزروعة في الزنك (الشكل 6أ). تم اختيار هذا العلاج لتقريب الحد الأقصى من الوفرة لـالسجلات المنظمة بواسطة، ومن ثم الحد الأقصىالتوافر مع تثبيط طفيف للنمو (الشكل التوضيحي التكميلي 9). الملفات الكروماتوغرافية لـ (على عكس و لا تتماشى بشكل غير كامل مع الامتصاص عند 280 نانومتر أو توزيع MncA على SDS-PAGE (الشكل التكميلي 11). لم نفعل تحديد الملوثتم تحديد البروتين من خلال تحليل المكونات الرئيسية وMncA-المعدنة من خلال كسر واحد يظهر أقل دليل على وجود بروتينات أخرى (الشكل التكميلي 11، الجدول التكميلي 6).
توقعات بحدوث تميؤ خاطئ مع (ليس ) لوحظ أيضًا في الخلايا المزروعة في النحاس (الشكل 6أ، الشكل التوضيحي 9). الملفات الخاصة بالنحاس (على عكس و ) مرة أخرى تتماشى بشكل غير كامل مع MncA ولكنها مرتبطة ببروتين من تطابق إنزيم الجليسرالديهيد-3 فوسفات ديهيدروجيناز (GAPDH) (الشكل التكميلي 12). من المعروف أن GAPDH يرتبط بالنحاس في الخلايا المعرضة للنحاس ويمكن إزالته باستخدام Blue Sepharose.. بما في ذلك هذه الخطوة احتفظت و تم القضاء على النحاس المتنقل مع MncA (الشكل التكميلي 13). تم قياس المعدنة من ثلاث ثقافات مستقلة بعد استخدام Blue Sepharose (الجدول التكميلي 7a، b). باختصار، توافرات داخل الخلايا لـ أو غير كافية لتعدين MncA في الخلايا المدعمة، مع تطابق التعدين مع التوقعات.
المعادن المحصورة في MncA في مستويات عالية و تنقيح التوافرات
محتجز MncA أو في الوسائط المدعمة بالمعادن المعنية (الشكل 6 أ، الشكل التكميلية 9، 14، 15، الجدول التكميلية 8، 9). تم التنبؤ بالمعالجة المعدنية كما هو الحال بالنسبة للتركيز العالي و لكن باستخدام قيم الحدود 0.01 لاحتلال RcnR-DNA (الشكل التكميلي 4). في وجود الكوبالت العالي، ارتبط MncA بشكل أكبرأقلوالمزيد، مما كان متوقعًا (الشكل 6أ). التفسير الأكثر اقتصادًا هو أنتراجع التوافر في المرتفعاتتم استخدام حاسبة بقايا MncA في البيانات التكميلية 5، التي تم صياغتها في الملاحظة التكميلية 2، لتقدير التراجع في التفضيلات المحددة لـ MncA.التوافر بالنسبة لـ (الشكل 6ب). الشكل 6ج يقارن بين الإشغالات الملحوظة والتوقعات باستخدام المنقحالتوافر. المتبقيات لـغير مستخدم في التنقيح، تم تقليله كما هو متوقع للحل الاقتصادي.
في العالي، ارتبط MncA أكثرأقل و ، مما كان متوقعًا (الشكل 6أ). هنا، التفسير الأكثر اقتصادًا هو أنمتاحة أكثر مما تم تقديره. قام حاسبة بقايا MncA بتقدير الزيادة الإضافية في داخل الخلاياالتوافر بالنسبة للحديد (الشكل 6ب). الشكل 6ج يقارن-الإشغالات المكررة مع تلك الملاحظة. المتبقيات لـتم تقليله مرة أخرى. باختصار، يمكن استخدام مقياس تواجد المعادن MncA بالإضافة إلى حاسبة بقايا MncA لاستكشاف وتحسين تقديرات توافر المعادن داخل الخلايا. يمكن أن تساعد هذه الأدوات في توقعات تواجد المعادن في البروتينات داخل خلايا أنواع أخرى وظروف نمو مختلفة. كما تشير التحسينات المستمدة من MncA إلىتراجع التوافر في. القولونية في كميات عالية من الكوبالت، وتم استكشاف ذلك لاحقًا.
سطحيتم استنفاد المخزون في الوسط غير المدعوم
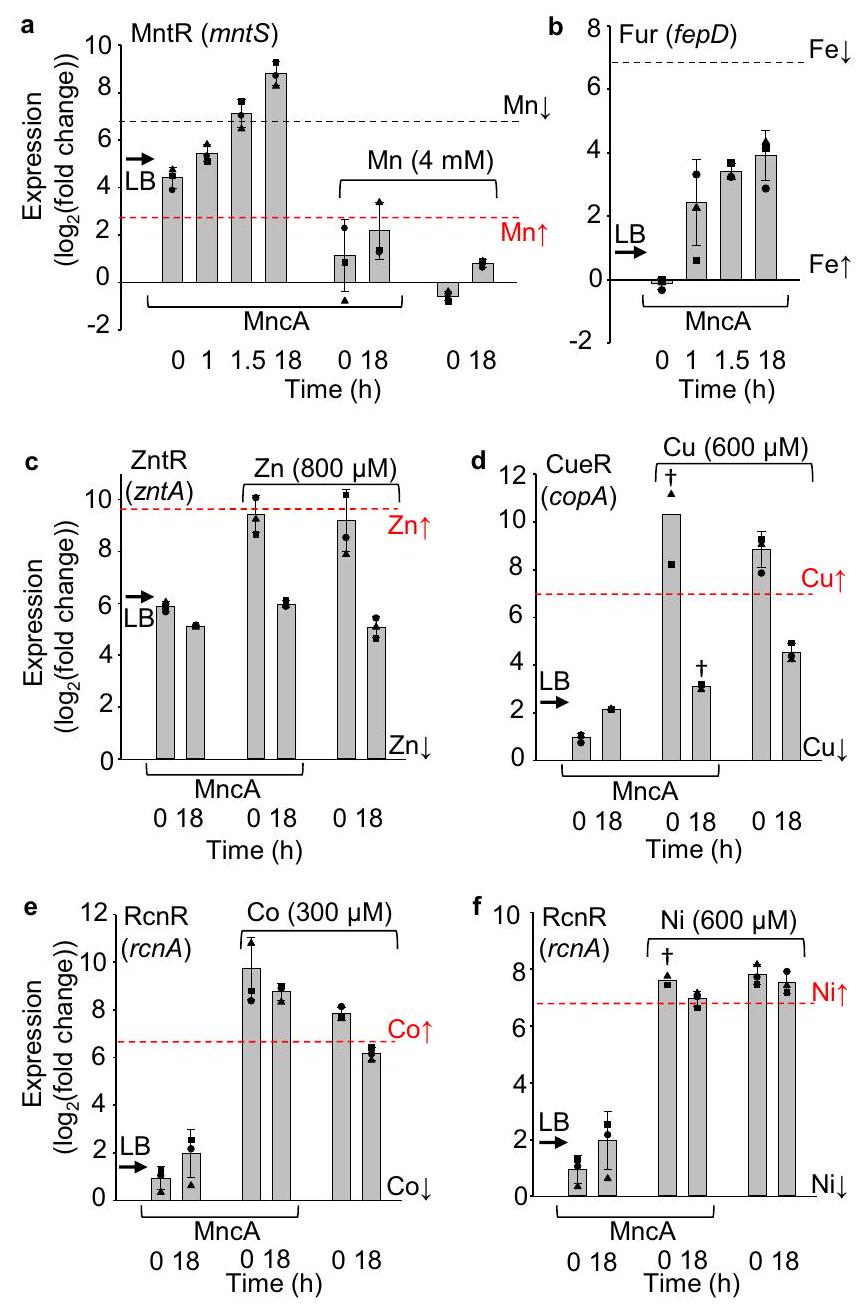
على الرغم منهو معدن متوفر بشكل كبير (الأقل سلبية، الشكل 4ج)، الـالمسبح في وسط غير مكمل ضحل، يحتوي فقط على بضع آلاف من الذرات لكل خليةتفاعل البوليميراز المتسلسل الكمي باستخدام بادئات لتنظيم MntRكشف عن زيادة في التعبير وبالتالي تقليل المتاح ( -تخفيف التثبيط المشترك لـ MntR) خلال التعبير المطول لـ MncA في وسط غير مكمل (الشكل 7أ). هذا يتماشى مع ملاحظة أقلمن المتوقع (الشكل 4c في الإطار). تشير النسخ المنظمة بواسطة Fur لـ fepD إلى انخفاض أصغر في المتاحبركة في وسط غير مكمل. تسلط هذه البيانات الضوء أيضًا على تحدٍ مع qPCR لتقدير توافر المعادن داخل الخلايا عند وفرة النسخ العليا، مما يتوافق مع توافرات أقل لمثبطات المعادن المعتمدة MntR و Fur. الطبيعة اللوغاريتمية لـ PCR كما تم تمثيلها بـالمقاييس في الشكل 7، تعني أن النصف العلوي من نطاق التعبير يتوافق مع دورة واحدة من تفاعل البوليميراز المتسلسل (PCR). وفرةتصل (وتتجاوز) السجلات الحد الأعلى، الأدنى المتاح، كما تم الإبلاغ عنه سابقًا بينما يبقى fepD 3 إلى 4 دورات أقل (الشكل 7 أ، ب). هذه البيانات تشير بالتالي إلى استنفاد داخل الخلاياحمام السباحة طوال الليل بواسطةلكن تغييرًا متواضعًا نسبيًا فيلذا فإن استخدام MncA لتحسين تقديرات التوافر المقدرة يكون ذا قيمة خاصة لتقديرات qPCR في
الشكل 6 | قراءات MncA لعملية التمثيل المعدني داخل الخلايا في مستويات عالية و تنقيح توافرات المعادن. أ عملية تميؤ المعادن داخل الخلية مع و ، ليس “ أو ، يتحول من في المكملات المعدنيةعلى التوالي كما هو موضح) وسائل الإعلام، تتطابق نوعياً مع التوقعات ولكنها أكبر كميًا و تفاعل المعادن أكثر مما تم التنبؤ به. تم التنبؤ بتفاعل المعادن (الأعمدة الزرقاء الداكنة) باستخدام البيانات التكميلية 4 من التوافرات في الوسط غير المضاف ولكن مع توافرات عالية معدلة بشكل منفصل. أو (كنص مدرج). تتوافق التوافرات العالية مع الحدود المعدنية العليا المعنية ( أو أو 0.01 ) الموضح في الشكل التوضيحي 4. متوسط ( ± الانحراف المعياري) قياسات التمثيل المعدني داخل الخلايا ( تكرارات بيولوجية مستقلة، مربع، دائرة، مثلث) في معدن عالي (أشرطة زرقاء باهتة) تم حسابها باستخدام البيانات في الجداول التكميليةاستنادًا إلى الأشكال التكميلية 11-15. ب توافر المعادن داخل الخلايا المختارة (المثلثات المنقحة باستخدام البيانات التكميلية 5 لـ (يسار) من الملاحظ في الخلية – و -احتلالات MncA في المرتفعات (بالنسبة لـ في وسط غير مكمل)، ولـ (يمين) من و الإشغالات في (بالنسبة إلى المرتفعكما هو موضح في النص المرفق). تفضيلات المعادن لـ MncA موضحة كـ (دوائر). القضبان هي نطاقات المستشعر من إلى. تم حساب التمثيل المعدني (الأعمدة الزرقاء الداكنة) باستخدام البيانات التكميلية 4 ولكن مع توافرات مصقولة كما في اللوحة (ب)، وتقليل المتبقيات لـ (الذي لم يتم تضمينه في عملية التنقيح) بالنسبة للملاحظة في تفاعل المعادن داخل الخلايا (الأعمدة الزرقاء الفاتحة مدرجة للمقارنة)، موضحة المتوسط ( ± الانحراف المعياري) (نسخ بيولوجية مستقلة، مربع، دائرة، مثلث). يمكن استخدام ميتاليشن MncA لتحسين توافر المعادن داخل الخلايا النسبي، وتم توفير البيانات التكميلية 5 لتمكين مثل هذه الحسابات. تم توفير بيانات المصدر كملف بيانات مصدر.
علوينطاقات التعبير. ينطبق هذا على بعض الخلايا المدعمة بالمعادن ولكن ليس على الخلايا غير المدعمة كما تم زراعتها سابقًا.عند مشاهدتهما معًا، تكشف الشكل 4c (الإدراج) والشكل 7a أن الفجوات بين التنبؤات والملاحظات لمعدنة MncA يمكن أن تشير إلى توافر المعادن داخل الخلايا في حالات غير مستقرة. ).
توفرات الحالة المستقرة المستدامة للارتفاع و ، ليس “ و
في بداية تعبير MncA، تصل الخلايا المعرضة للمعادن المرتفعة جميعها إلى أو تتجاوز وفرة النسخ المقدرة المحددة كحدود qPCR لتوافر المعادن داخل الخلايا بشكل مرتفع (الشكل 7).-MntR يثبط نسخ mntS، وهذا يعكس ارتفاع تواجد محفز mntS في الخلايا المعرضة لـ 4 مللي مول من المنغنيز وانخفاض وفرة النسخ. بالمقابل، هذا يعادل ارتفاع وفرة النسخ لـ و نصوص استجابة منظمة بواسطة المحفزات ZntR وCueR ومزيل الكبت المعتمد على المعادن RcnR. في نهاية إنتاج MncA (18 ساعة) في الخلايا المدعمة بالمعادن، و تظل التوافرات مرتفعة، لكنها تنخفض لـ و (الشكل 7 أ، ج-و). هذا صحيح أيضًا بالنسبة للثقافات التي لا تعبر عن MncA باستثناء الاحتمال الرسمي بأن دوران MncA المعدني حافظ على حالة مستقرة مرتفعة. و التوافرات ولكن ليس و عدم وجود حالة مستقرة مرتفعة و تشير إلى اختلاف في الميتالستاز لهذه المعادن مقارنة بـ و قد يكون من الأكثر تحديًا إدخال المعادن في البروتينات داخل خلايا قابلة للحياة مع أو ببساطة من خلال مكملات المعادن.
الفرو يسببذرات خلويةللانخفاض في
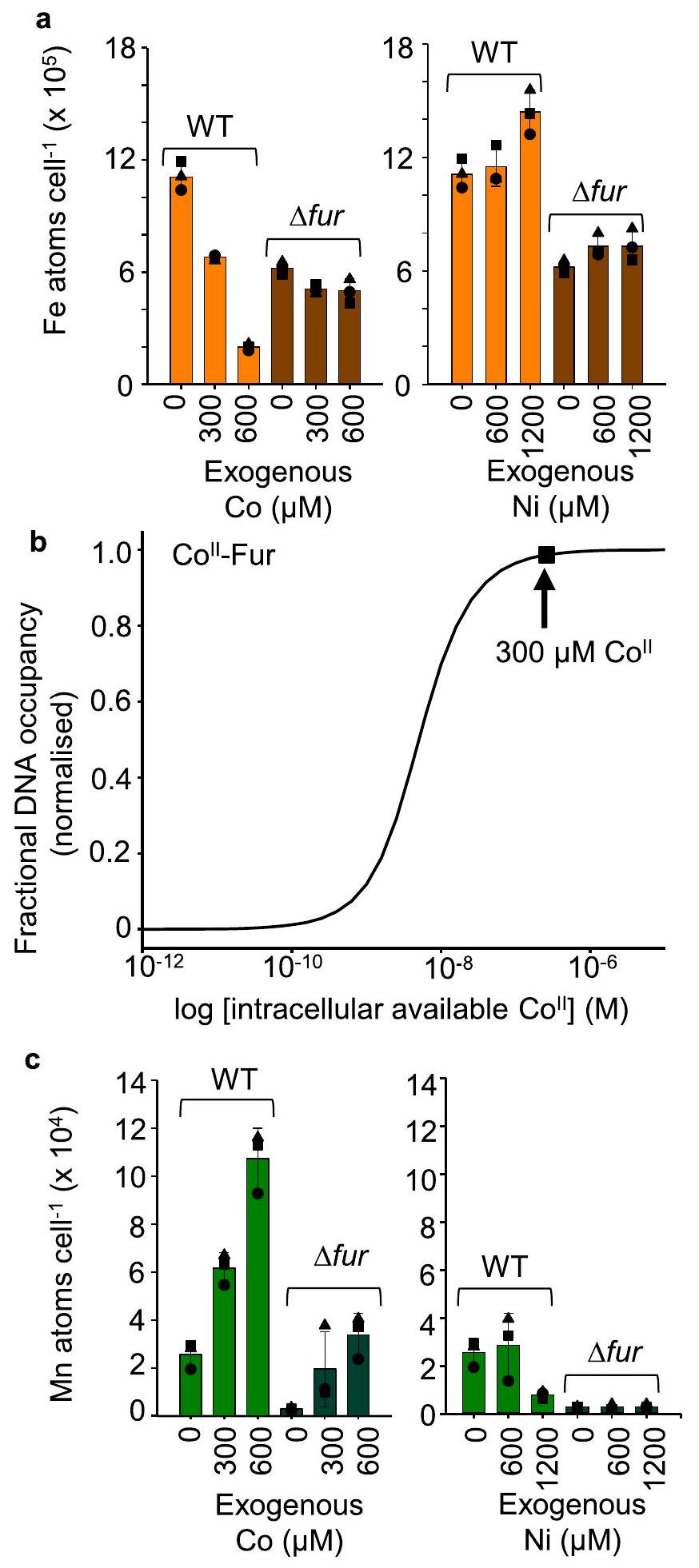
للتحقيق في الانخفاض المتوقع بواسطة MncA في داخل الخلاياالتوفر العالي (الشكل 6ب)، إجمالي ذرات الحديد في الخليةتم قياسها بواسطة ICP-MS (الشكل 8أ). انخفض محتوى الحديد فييتماشى ذلك مع التوقع (مع ملاحظة أن المعدن المتاح يمكن أن يتجه أحيانًا بشكل مختلف عن إجمالي المعدن). تحتوي الخلايا التي تفتقر إلى Fur على كمية أقل من الحديد في الوسائط القياسية كما تم شرحه سابقًا.من المهم أن تأثيرعلى الحديد غائب فيالفرو (الشكل 8أ). يرتبط الفرو و -الفور يربط الحمض النووي بكلا الألفة المعروفة بالفعلالشكل 8ب يحسب استجابة Fur داخل الخلايامحاكى كما لـاحتلال الحمض النووي فيتوافر داخل الخلايا المزروعة هنا في مستويات مرتفعة من الكوبالت يكشف عن استجابة متقاطعة شديدة لـ Fur. من المعروف أن المستشعرات غير المتجانسة تستجيب بشكل متقاطع لتوافرات في أعلى نطاقات المستشعرات المتجانسة.من الأهمية بمكان، التفاعل المتقاطع للمعادن مع الفور. يفسر الانخفاض في إجمالي الحديد في الشكل 8أ وفي الحديد المتاح داخل الخلايا تم قراءته بواسطة MncA (الشكل 6ب).
المعادن المحصورة بواسطة MncA تقوم بتحسين التقديرات المتاحة بشكل متكرر
تم استخدام الفروق المتبقية بين الإشغالات المرصودة والمتوقعة في الشكل 6c عبر حاسبة بقايا MncA (البيانات التكميلية 5) لتحسين توافر المعادن داخل الخلايا بشكل أكبر (الشكل التكميلية 16).يُستنتج أن التوافر ينخفض في المرتفعاتوزيادة في الارتفاعتمت عكس كلا الاتجاهين المتوقعين في خلية الذرات الكلية.، وآثار الـعلىلم تكن تعتمد على الفراء (الشكل 8c). تأثيرات الارتفاععلىلا تعتمد على-استشعار MntR (الشكل التوضيحي 17a). الانخفاض المستقل عن MntR في أقل حدة في النوع البري حيث يتحكم المستشعر في توازن المعادن مما يخفف من التغيرات. تشير المتبقيات في الخلايا المعرضة لـ 4 مللي مول من المنغنيز إلى تراجع في المتاح وهذا يتماشى مع انخفاض مستقل عن الفيريتين في إجمالي ذرات الحديد في الخلية (الشكل 5ب، الشكل التوضيحي 17ب). تم استخدام حاسبة بقايا MncA بشكل تكراري لمزيد من تحسين العالي و المخططات (الشكل التوضيحي التكميلي 16). التوافرات المنقحة والمعادن العالية المتعلقة بها
الشكل 7 | ضحلتم استنفاد المسبح في وسط غير مدعوم ولكن تم الحفاظ على توافر الحالة المستقرة في مستويات عالية و . qPCR لجين mntS (الذي يتم تنظيمه بواسطة-MntR)،فب د (حديد-فرو)، زنتا (-ZntR)، د ( -CueR)، e RcnR )، ، كـ التغيير بالنسبة لأدنى وفرة نسخ تم قياسها تم ملاحظتها في فاستر وآخرون.استخدام الخالب، المعدن و/أوالخلايا المدعمةالقيم في بيانات المصدر TXT للشكل 7، البيانات هي المتوسطانحراف معياري لـتكرارات بيولوجية مستقلة (مربع، دائرة، مثلث) باستثناء حيث يوجد خنجرتزداد نسخ جينات MntR و Fur في الظروف المنخفضة و (MntR و Fur هما مثبطان معتمدان على المعادن)، لكن نصوصًا أخرى تزداد في توافر المعادن المماثلة العالية (تُنظم بواسطة المنشطات أو مثبطات التحرير المعتمدة على المعادن). السابقتم عرض (تغيير الطي) qPCR عند حدود المعادن العالية (الخطوط الحمراء المتقطعة، 0 لـ Fur) وعند حدود المعادن المنخفضة (الخطوط السوداء المتقطعة لـ MntR و Fur، 0 للآخرين). تشير الأسهم إلى السابق (تغيير الطي) qPCR للنسخ المتجانسة المعزولة من الخلايا المزروعة في LBتظهر اللوحات (أ) و (ب) التعبير عندبعد إضافة الأرابينوز إلى الوسط غير المدعوم لتحفيز التعبير المنخفض لـ MncA عند. بعد 18 ساعة، تم تنظيم MntRالسجلات تتجاوز المستوى المنخفضحدود (شغل الحمض النووي) ) (اللوحة أ) تم تحديدها بشكل أكبر في الشكل التكميلية 18. المسبح ضحل ومُستنفد في LB. بالنسبة لـ fepD المنظم بواسطة Furتقرير المقياس عن نقص متواضع في المتاح داخل الخلايافي الساعة 18 في وسط غير مكمل (الشكل التوضيحي 18). العمودان الأولان (الألواح ) تظهر تغيرًا ضئيلًا في التوفر لـ أو بعد 18 ساعة من تعبير MncA في وسط غير مدعوم. الأعمدة الأربعة النهائية (الألواح ) تقرير (تغيير الطي) في وسائط مدعمة بالمعادن سواء مع أو بدون تعبير MncA كما هو موضح. حالة مستقرة من المنغنيزالتوفر الذي يقترب من الأعلىتم الحفاظ على الحدود حتى 18 ساعة في 4 مللي مول من المنغنيز (اللوحة أ). كما يتم الحفاظ على الحالة المستقرة التي تقترب من الحد العلوي للمعادن في مستويات عالية و ، لكن ليس و (الألواح تم توفير بيانات المصدر كملف بيانات المصدر.
تُقدم الآلات الحاسبة (البيانات التكميلية 6-8). يمكن استخدام MncA وآلة حساب بقايا MncA في البيانات التكميلية 5 لاستكشاف توافر المعادن النسبي في أنواع خلايا أخرى بما في ذلك (تصنيف) الخلايا التي تم هندستها لتحقيق التوازن المعدني لتحسين نوعية التمثيل المعدني للعناصر المختارة.
نقاش
هنا نكتشف أن تفضيلات ربط المعادن لبروتين يلتقط المعادن حركيًا أثناء الطي، MncA، تتبع جميعها سلسلة إيرفينغ-ويليامز (الشكل 1). قد يعكس هذا تفضيلات المواقع المرنة في مسار الطي قبل الحبس الحركي. من الممكن أيضًا أن المعادن المفضلة في السلسلة تشجع بشكل متزايد تقدم وسائط طي MncA. على الرغم من أن النحاس هو الأكثر تفضيلًا، إلا أن المعدن المقابل يتم فك شفرته بشكل صحيح ليكونمن خلال ربط هذه التفضيلات بتقديرات مثالية لتوافر المعادن داخل الخلايا، موحدة على النقاط الوسطى لنطاقات حساسات المعادن (الشكل 2). يقوم MncA بحبس المعادن بشكل حركي، بحيث أن تلك التي تتطابق في التنقية من المحتمل أن تعكس حالات التمثيل المعدني في الجسم الحي (الشكل 3، الشكل التكميلية 7). تقديرات توافر المعادن داخلتنبؤات بخصوص E. coli المزروعة هوائياً تشير إلى أن MncA (من السيانوبكتيريا) سيتم تميه المعادن بشكل خاطئ معفي. كولاي، وهذا ما تم إثباته من خلال تحليل ICP-MS لمركب MncA القابل للذوبان المنقى (الشكل 4). وبالتالي، يتم إظهار نوعية التمعدن بشكل مباشر على أنها تحددها تفضيلات الربط النسبية للمعادن المتنافسة عند التوافر الداخلي (الشكل التوضيحي 5a)، ويمكن التنبؤ بالتمعدن الخاطئ في العوائل غير المتجانسة. يمكن أن تؤدي التغيرات الطفيفة في توافر المعادن (أو التفضيل) إلى تغيير التمعدن لأن التوافر الداخلي يتبع عكس سلسلة إيرفينغ-ويليامز وبالتالي يتماشى مع التفضيلات (الأشكال 2c، 4c، 5b، 6a). يمكن استخدام تقديرات توافر المعادن الداخلية بالتالي للتنبؤ بنوعية التمعدن في الجسم الحي لإبلاغ هندسة البروتينات المعدنية الطبيعية والصناعية لتحسين التمعدن (باستخدام البيانات التكميلية 3، 4، 6-8).
توافر المعادن غير المكررة المستمدة من استجابات qPCR لمُنظِّمات النسخ الحساسة للمعادن توقعت بشكل صحيحتغيير المعادن في MncA في E. coli في وسائل غير مكملة (الشكل 4c). في وسائل مكملة بالمعادن، كانت إشغالات MncA مع، و تجاوزت (على الأقل إلى حد ما) التوقعات (الأشكال 5، 6a). تم استخدام هذه الإشغالات مع البيانات التكميلية 5 لتحسين تقديرات التوافر داخل الخلايا التي تتنبأ بانخفاض التوافر المتاح.في و انخفض المتاحفي، ولكن زادت المتاحةفي شركة (الشكل 6ب، الشكل التوضيحي 16). ومن المشجع أن هذا تزامن في كل حالة مع تغييرات مماثلة في الإجمالي أو ذرات خلوية (الشكل 8أ، ج، الشكل التكميلية 17أ، ب). الانخفاض في الخلايا في العاليلكن ليس بشكل عالٍ، كان يعتمد على الفراء وفسر السابق من خلال الاستجابات لـمن الفراء (الشكل 8 أ، ب، الشكل التوضيحي الإضافي 17 ب). مدى تداخل المعادن المتقاطع مع الفراءإن التغير غير المرغوب فيه في التمثيل المعدني لا يزال بحاجة إلى إثبات. معًا، تُظهر هذه البيانات قيمة MncA كأداة لاستكشاف توافر المعادن داخل الخلايا، بشكل موازٍ لمستشعرات المعادن الاصطناعية.وتكمل أجهزة استشعار المعادن الذاتية المعايرة، خاصةً حيث تكون تقنية qPCR أقل موثوقية و/أو من المحتمل حدوث تداخل..
تقرير إشغالات MncA تم تخفيضهالتوفر داخل الخلايا بعدالنمو في الوسط غير المدعوم (الشكل 4). هنا، السطح الضحلتُستنفد المسبح خلال تعبير MncA بحيث لا يتم الحفاظ على توفر الحالة المستقرة (الشكل 7 أ، الشكل التوضيحي 18). هذا مهم عندما يتم التعبير عن البروتينات المعدنية المؤتلفة بشكل مفرط في الإشريكية القولونية مما قد يؤدي إلى استنفاد المعادن: يمكن أن تعكس عملية التمعدن عمق أحواض المعادن المتاحة بدلاً من قوة المنافسة مع روابط أخرى. في الخلايا المدعمة بالمنغنيز والنيكل والكوبالت، تم الحفاظ على توفرات داخلية مرتفعة.لكن لم يكن هذا هو الحال في الزنك والنحاس المرتفعين (الشكل 7). آليات الميتالستاز لـ و يجب تعديل معدلات الاستيراد والتصدير و/أو الاستهلاك لاستعادة توافر الحالة الثابتة داخل الخلايا قبل التعرض، في حين أن الحالات الثابتة المرتفعة هي
الشكل 8 | الفراء يسبب ذرات الحديد في الخليةللانخفاض فيتطابق توافرات MncA المتوقعة. إجمالي ذرات الحديد في الخليةتحدد بواسطة ICP-MS لمستخلصات الخلايا، انخفاضات في الخلايا المزروعة في وسط مضاف إليه نسبة عالية من (اللوحة اليسرى). هذا يتماشى مع الانخفاض في داخل الخلاياالتوافر المقدر من المتبقيات في الشكل 6أ، المحسوبة باستخدام البيانات التكميلية 5 والم shown في الشكل 6ب. مرتفعلا يؤثر على إجمالي ذرات الحديد في الخلية (اللوحة اليمنى). إجمالي الحديد في الخليةأقل فيالفرو ولكن لا يتراجع أكثر في المستويات المرتفعةانخفاض في خلايا الحديداستجابةً لارتفاعيعتمد على الفراء. ب الفراء يرتبطلتعزيز ارتباط الحمض النووي: تم محاكاته باستخدام المعروفةألفة الفراء، ألفة الحمض النووي-الفور، بالإضافة إلى ارتباط الحمض النووي apo-Fur وجزيئات الفور في الخلية، كما تم محاكاته لـسيتم قمع مروجي الفراء بشكل شاذ فيالكوبالت (السهم الأزرق المربع) يسبب الانخفاض المتوقع في التوافر داخل الخلاياموضح في الشكل 6ب والانخفاض الملحوظ في إجمالي ذرات الحديد في الخليةفي اللوحة (أ). ج إجمالي ذرات المنغنيز في الخليةزيادة في الارتفاعمتسق مع زيادة طفيفة في التوافر داخل الخلاياقابل للتشكيل من المتبقيات المتبقية في العاليفي الشكل 6c، تم حسابه باستخدام البيانات التكميلية 5، الموضحة في الشكل التكميلية 16b. هذه التأثيرات للكوبالت علىمستقلة عن الفراء. إجمالي ذرات المنغنيز في الخليةانخفاض في الارتفاعهذا الاتجاه أكثر وضوحًا في (الشكل التوضيحي 17)، الاتجاه غير واضح في الفرو الذي يحتوي على نسبة منخفضة من المنغنيز (يمين). انخفاض مماثل في توفر الخلايا الداخليةيمكن حسابه (البيانات التكميلية 5) من المتبقيات المتبقية في العاليالشكل 6ب، موضح في الشكل التكميلي 16أ. استخدام MncA كأداة لقياس توافر المعادن داخل الخلايا بشكل نسبي يقوم بتحسين القيم الموضحة في الشكل التكميلي 16، المضمنة في البيانات التكميلية 6-8، والمقدمة كخطط للمساعدة في تعديل نوعية تفاعل المعادن مع البروتينات في الكائنات الحية. المتوسط ± الانحراف المعياري لـتكرارات بيولوجية مستقلة (مربعات، دوائر، مثلثات) في (أ) و(ج). يتم توفير بيانات المصدر كملف بيانات مصدر.
تم الحفاظ عليه للمعادن الأخرى (الشكل 7). ومع ذلك، يُتوقع أن يرتبط MncA بشكل ضئيل أو حتى لو تم الحفاظ على توافرات مستقرة مرتفعة. ومن الجدير بالذكر أن التدرجات الأكثر ملاءمة ( ) لـ و تستبعد إلى حد كبير (الشكل التوضيحي 19). أقل و في وسائل الإعلام، هندسة الخلايا لتقليل توافرها من و أو إضعاف ارتباط MncA بـ و يمكن أن يساعد في تراكم.
تكوين المعدن لـ MncA داخل. القولونية انتقلت بين، و كما تم التنبؤ به (الأشكال 4-6). يصبح هذا ممكنًا لأن توافر المعادن داخل الخلايا يتماشى مع تفضيلات ربط المعادن للبروتينات. قد يشير هذا إلى أن حدوث خطأ في تميؤ المعادن للبروتينات من المحتمل. في الجسم الحي. بدلاً من ذلك، يمكن أن تكون آليات توازن المعادن مضبوطة بدقة استثنائية لتتناسب مع البروتينات المعدنية الأصلية مما يقلل من حدوث التمثيل الخاطئ للمعادن. في كلتا الحالتين، يثير هذا احتمال حدوث تمثيل خاطئ للمعادن على نطاق واسع في هندسة البيولوجيا لأن توافر المعادن قد لا يكون مضبوطًا بشكل صحيح للبروتينات المعدنية غير الأصلية. ومن الجدير بالذكر أنه في الإشريكية القولونية، يُتوقع أن تكون الشابيرات المعدنية المعتمدة على GTP YeiR وYjiA معدنية بشكل أساسي مع بينما يصبح CobW غير المتجانس من Rhodobacter وCbiK من Salmonella، مثل MncA من السيانوبكتيريا، ميتالياً خاطئاً مع و على التوالي (الشكل 4، الشكل التكميلي 20). قياسالإنتاج في الهندسة. كولي قدمت دليلًا غير مباشر على التوقعات المتعلقة بسوء التمثيل المعدني لـ CobW، ومن المعروف أن CbiK يقوم بإدخال الحديدإلى السيروهم في السالمونيلا التي تفتقر إلى السيروهم كيلاتاز CysGمن الأهمية بمكان، أن MncA الآن يوفر قراءة مباشرة تؤكد التوقعات بشأن التمثيل الخاطئ للمعادن مع (الشكل 4).
يمكن أن توجه توافر المعادن داخل الخلايا المنقحة بواسطة MncA، جنبًا إلى جنب مع الآلات الحاسبة المقدمة هنا، تحسين عملية التمعدن من خلال استخدام مكملات المعادن أو العوامل الخالبة، أو التعديلات على التوازن الداخلي عن طريق هندسة سلالات المضيف (الهياكل الأساسية)، أو هندسة البروتينات لتتناسب مع التوافر. تعتمد حوالي نصف تفاعلات الحياة على كيمياء المعادن الصحيحة المرتبطة بالبروتينات المعدنية.يتم توجيه أبحاث كبيرة نحو إنتاج أنواع مختلفة من الإنزيمات المعدنية الاصطناعية.لا مفر من أن الجهود في الهندسة الأيضية، وعلم الأحياء الاصطناعي، والتطور الموجه ستعتمد غالبًا على الإنزيمات المعدنية.يمكن أن توفر المخططات الحياتية والآلات الحاسبة معلومات عن الإنزيمات المعدنية المهندسة لتطبيقات المعالجة الحيوية داخل الكائنات الحية. إن تحسين عملية التمعدن داخل الكائنات الحية يتيح فرصًا حيث يتم إعطاء الأولوية لهندسة الأنظمة البيولوجية من أجل التصنيع الحيوي (المستدام)..
طرق
تعبير وتنقية البروتين غير المطوي apo-MncA
لفحص تفضيلات الارتباط النسبية لـ MncA في المختبر، تم التعبير عن البروتين وتنقيته.. باختصار، تم التعبير عن إشارة إفراز MncA بدون TAT من pET29a-mncA في E. coli BL21(DE3) pLysS. إيزوبروبيلتم إضافة -D-1-thيوقالكوبيرانويد (IPTG، 1 مل 0.4 م) إلى ثقافة في مرحلة النمو المتوسطة (1 لتر في قنينة سعة 2 لتر)فيلتحفيز التعبير على مستوى عالٍ لمدة 3 ساعات قبل جمع الخلايا بالطرد المركزي ( ) والتجميد ( ). تم إعادة تعليق الكريات في 30 مل من محلول تريس بتركيز 100 مللي مولار ودرجة حموضة EDTA، 1 مللي مول من فلوريد الفينيل ميثيل سلفونيل (PMSF) ومُعالج بالموجات فوق الصوتيةعلى الثلج، بفواصل زمنية مدتها دقيقة واحدة). تم تصفية السائل الخلوي بواسطة الطرد المركزي، )، تم التخلص من السائل العلوي، وتم إعادة تعليق الرواسب في 100 مليمول من تريس ترايتون إكس-100مُعالج بالموجات فوق الصوتيةعلى الثلج، فترات زمنية مدتها دقيقة واحدة) وتم تكرار الطرد المركزي. تم غسل الرواسب في 30 مل من 100 مليمول تريسلإزالة الترايتون، ثم بالتتابع في 30 مل من 50 مليمول/لتر هيبز pH 7.5، 1 مولي يوريا تليها 15 مل من 50 مليمول/لتر هيبز pH 7.5، 2 مولي يوريا. خلال الغسل النهائي، تم تقسيم المستخلص إلى حصص 2 مل تليها الطرد المركزي.لاستعادة الأجسام الشاملة، المخزنة في.
تحضير مخزونات المعادن
أسهم المعادن و ) في الماء النقي للغاية تم تصفيته بشكل معقم وتم قياسه بواسطة مطيافية الكتلة مع اقتران البلازما المحفزة (ICP-MS). عند الحاجة للتجارب اللاهوائية،تم إعداد الأسهم فيتم تنقية المياه فائقة النقاء. ثم تم تأكيد تركيزات المنغنيز والحديد الكلية بواسطة ICP-MS.تم تأكيد المخزون على أنهتم تقليلها بواسطة التفاعل مع فائض (عشر مرات) من الفيروزين (Fz) باستخداملـمعقد. مخفضتم تحضير الأسهم كما هو موضح في تجارب محددة وتم التحقق من حالة النحاس الثنائي باستخدام حمض البيكينشونيك (BCA) و ICP-MS.
إنتاج المعادن المتنافسة المدعومة
كانت محاليل إعادة طي MncA في المختبر (في 50 مليمول MOPS pH 7.5) تحتوي على أزواج من المعادن والروابط بكميات متنوعة لتحقيق تركيزات مختلفة من المعادن المتاحة. تم إعداد المحاليل في قوارير مغسولة بحمض مع إضافة المكونات بالترتيب: عازل pH، ماء نقي للغاية، رابط (عادةً NTA، أو 1 مليمول L -هيستيدين للمنافسات مع ) تليها المعدنان. عادة ما كانت المحاليل العازلة تُحضّر بحجم 100 مل، وتُصفى عبر تصفية أي دليل على تشتت الضوء. يوضح الجدول التكميلي 1 الكميات الإجمالية لكل معدن و ligand (NTA، الهيستيدين أو لا شيء) بالإضافة إلى [المعادن المتاحة] المضافة في كل محلول إعادة الطي. تم استخدام البيانات التكميلية 1 (المشتقة في الملاحظة التكميلية 1) وتوفيرها هنا للمساعدة في إنتاج مثل هذه المحاليل المعدنية المزدوجة. لتحقيق فعالية العازل، يجب أن يتجاوز التركيز الإجمالي للمعادن بشكل كبير تركيز البروتين.-ضعف بشكل عام، 10 -ضعف للمسابقات معلتحقيق مخزن فعاليجب أن يتم معدنة عامل التخفيف.
تم إجراء تجربة أولية باستخدام BCA كعازلدون توقع للتخزين المؤقت. مع و ، متوقع مجاني هو الذي تم التنافس ضده. لأن < النحاس هاجر مع MncA، غير مخففتم التنافس ضد غير المخزنتركيزات نموذجية كانتكل منها تم التحقق منه بواسطة ICP-MS في محلول الطي. المنافسات بين و استخدم محلولًا مُعدًا حديثًا منوشملت تجارب إعادة الطي 1 مللي مول من الهيدروكسيلامين للحفاظ على النحاس في شكل مخفض. تم تحديد تركيزات كل معدن باستخدام برنامج HySS لتكوين معقدات تعتمد على المعادن بنسبة 2:1.. القيم لجميع التوازنات هي كما يلي: تفكك البروتون من الهيستيدين (HisH-9.08،تعقيدتكوين بشرته ( ، ) ، . لمجمعات BCA، اشتقاق التخفيف من ] في الملاحظة التكميلية 1.
Metal-binding preferences of MncA at folding
Inclusion bodies containing MncA were solubilized in HEPES pH 7.5 with 8 M urea, and the concentration calculated from using an experimentally determined extinction coefficient , typically in the range . Refolding was achieved by
dropwise dilution of urea-solubilised MncA in large volumes ( 100 mL ) of solutions containing pairs of competing buffered metals prepared as described earlier. Solutions were thoroughly mixed before adding unfolded MncA and gently mixed between additions. Dilute refolded MncA was recovered by binding to a 1 mL Q-Sepharose (Cytiva or GE Healthcare) anion exchange column pre-equilibrated with low-salt buffer Tris pH 7.5, 50 mM NaCl. The column was washed with 20 mL low-salt buffer before eluting MncA with high-salt buffer, 50 mM Tris pH . MncA was quantified by then resolved ( 0.5 mL at ) by SEC (PD-10, GE healthcare, previously washed with 0.5 mL 5 mM EDTA followed by ultrapure water and equilibrated with the low-salt buffer). Fractions ( 0.5 mL ) were analysed for protein via and metal by ICP-MS using corresponding matrix-matched calibration curves. To compete versus and , stocks were prepared as described earlier and refolding done in an anaerobic chamber. The concentrations of and metals (ICP-MS) were superimposed to identify metals co-eluting with MncA and estimate the metal:protein stoichiometry. The ratio of trapped metals was used to determine the relative binding preferences of the two metals at folding in buffer of known competing metal availabilities (Table 1, Supplementary Tables 1, 2).
البنية البلورية
أتم إضافة محلول MncA في 50 مللي مولار HEPES pH 7.5 و 8 مولار يوريا، المحضر من كتل الإدماج المترسبة، بالتنقيط مع التحريك إلى 100 مل من 50 مللي مولار MOPS، pH 7.5، ومرّ عبرتم تصفية وتحميل العينة على عمود Q-Sepharose بسعة 5 مل (Cytiva) متوازن مع 50 مللي مول من تريس، pH 7.5، و50 مللي مول من NaCl. تم غسل العمود بنفس المحلول. تم طيها وتركيزها.تم غسلها بمحلول يحتوي على 500 مللي مولار من NaCl. تم استبدال المحلول عبر عدة دورات من التخفيف والتركيز (Amicon Ultracel، 0.5 مل 10 كيلودالتون) للحصول علىفي 10 مللي مولار تريس، pH 7.5، 50 مللي مولار NaCl. تم تنمية البلورات في 100 مللي مولار أسيتات الصوديوم، pH 4.0 وPEGتم إعادة ذوبان جزء من البلورات في 50 مللي مول من تريس،تم تحليلها بواسطة ICP-MS وتم قياس تأكيد التفاعل المعدني الستيوكيومتري مع. بعد الحماية بالتبريد من خلال الإضافة التدريجية للجلسرين إلىتم تبريد البلورات بسرعة وتخزينها في النيتروجين السائل قبل جمع البيانات.
تم جمع البيانات في خط الشعاع I04، مصدر الضوء الماسي (الجدول التكميلي 3). تم جمع مجموعة بيانات ذات تكرار عالٍ من خلال الحصول علىلبيانات في أربع مسحات منفصلة على محور بلورة طويلة على شكل قضيب سداسي. تم معالجة البيانات باستخدام حزمة xia2 في دايموندالذي استخدم XDS للتكاملو AIMLESS لتوسيع ودمج البياناتتم الحصول على حل الهيكل من خلال استبدال الجزيئات مع الثلاثيالبنية (معرف PDB 2VQA) كنموذج بحث باستخدام فازرتم تنفيذها في فينيكساستمر تحسين النموذج في فينيكس بالتناوب مع النمذجة في كوتتم إعداد الأشكال باستخدام ChimeraX، وبرنامج PyMOL (Schrödinger). تم استخدام MOLE 2.5 لتحديد القنوات الموجودة في هيكل.
تعبير وتنقية RncR
تم وصف تنقية RcnR المفرط التعبير في E. coli BL21(DE3) من تسلسلات الترميز المدمجة في pET29a. بالمثل مع المنظمين الآخرين (MntR، Fur، NikR، ZntR، Zur، CueR)، كانت التسلسل من سلالة سالمونيلا إنتيريكا سيروفار تايفيموريوم SL1344 (المشار إليها بسالمونيلا)، وRcnR تشاركتطابق التسلسل مع E. coli RcnR. تم تحضير RcnR غير الهوائي، المخفف وapo-RcnR عن طريق تطبيق البروتين النقي المعالج بـ EDTA.عمود الهيبارين HiTrap، تم نقله إلى غرفة لاهوائية، وغسله بـأحجام الأعمدة المعالجة بـ Chelex،-مُطهَّر10 مللي مولار هيبس، pH 7.0، ثم تم إزالتها بـ10 مللي مولار هيبيس، pH 7.0. تم قياس RcnR بواسطةباستخدام معامل الانقراض المحدد تجريبيًا لـتم الحصول عليه عبر تحليل الأحماض الأمينية الكمي. تم تقييم محتوى الثيول المعدني والمخفض.وجميع عينات البروتين اللاهوائي (المحافظة في غرفة لاهوائية) كانتمخفضخالي من المعادن. تم إجراء جميع التجارب في المختبر تحت ظروف لاهوائية باستخدام مواد معالجة بـ Chelex و-المخازن المنظفة.
الستيوكيومترية والألفة لـ RcnR
تم إجراء جميع التجارب فيHEPES pH 7.5. لتحديد النسبة المولية، (كـ ) تم معايرته في بروتين منقى ( ) وطيف الامتصاص المسجل عند التوازن باستخدام طيف ضوء الأشعة فوق البنفسجية والمرئية (بيركن إلمر لعلوم الحياة). بالإضافة إلى ذلك، تم أخذ عينة منالمونومر) تم حضنه معوتم فصل المعدن المرتبط بواسطة SEC وتم إصداره معHEPES pH 7.5 (PD-10، جمع 0.5 مل من الفractions) وتم تحليله للبروتين بواسطة اختبار برادفورد المعاير مع تركيزات معروفة من RcnR والمعادن بواسطة ICP-MS. لتحديدتم إجراء المعايرات في وجود EGTA باستخدام أربعة تركيزات من أحادية RcnR؛ RcnR و EGTA RcnR و EGTA RcnR و EGTA RcnR و EGTA، مراقبةميزة معتمدة على-RcnR عند 326 نانومتر. تم إجراء ملاءمة متزامنة لجميع مجموعات البيانات باستخدام Dynafit (نماذج الملاءمة في البرنامج الإضافي)..
-تفاعل الحمض النووي RcnR بواسطة الاستقطاب الفلوري
مجسات الحمض النووي مزدوجة الشريط المعلّمة بالفلور (هيكسكلوروفلورسئين) التي تحتوي على موقع ارتباط RcnR المحدد upstream منتم تخليق وتلدين المحفز كما هو موضح. RcnR ( تمت معايرة رباعي RcnR) أو apo-RcnR في 10 نانومتر من الحمض النووي فيHEPES pH 7.5. تغييرات في الأنيسوتروبي ( تم قياسها باستخدام مطياف فلوريسcence Cary Eclipse المعدل (Agilent Technologies) المزود بمرشحات قطبية.متوسط الوقتيكرر، و )، مما يسمح للكوفية بالتوازن ( 3 دقائق ) قبل التسجيل. تم إجراء ملاءمة متزامنة لجميع مجموعات البيانات (نماذج الملاءمة في البرنامج الإضافي باستخدام الحد الأقصى ).
تعبير وتنقية MncA القابلة للذوبان لتحديد التمثيل المعدني في الجسم الحي
منطقة الترميز لـكما في pET29a-تم استنساخها فرعيًا لإنشاء pBAD30-mncA لتمكين التعبير المتناغم اعتمادًا على [الأرابينوز]. تم تلقيح E. coli BW25113 (المشار إليها فيما بعد بـ E. coli) في ثقافات ليلية (جلوكوز + كاربينيسيلين عند ) استخدم اليوم التالي لتطعيم قنينة سعة 2 لتر تحتوي على 1 لتر من وسط LB وكاربيسيلين (بدون جلوكوز) وتم حضنها في حتى وصلت OD إلى مرحلة منتصف اللوغتم نقل الثقافات إلى وتركيز منخفض ) تم إضافة L-arabinose لتحفيز تعبير الجينات بمستوى منخفض يتبعه زراعة طوال الليلإجمالي) قبل حصاد الخلايا عن طريق الطرد المركزي (تضمنت تنقية MncA المعدني القابل للذوبان في الجسم الحي بروتوكولات مشابهة للإجراءات المستخدمة لاستعادة MncA الأصلي من Synechocystis.تم إعادة تعليق الكتلة الخلوية بالكامل (من 1 لتر من الثقافة) في 30 مل من محلول التحلل (20 مللي مول من تريس pH 7.5، 1 مللي مول من EDTA، 1 مللي مول من PMSF) وتمت المعالجة بالموجات فوق الصوتية (4 دقائق نبض) ثم تم الطرد المركزي. ) 45 دقيقة لإزالة حطام الخلايا. تم تحميل السائل العلوي على عمود تبادل الأنيونات Q-Sepharose (5 مل، تم توازنها مسبقًا مع 20 مللي مولار تريس pH 7.5) ثم تم غسله بنفس العازلة. تم استرجاع MncA باستخدام تدرج في 30 مل، جمع 1 مل من الفractions. تم تحديد الفractions التي تحتوي على MncA بواسطة SDS-PAGE وتم تجميعها.تم استخدام بروتوكول تحليلي أسرع في التجارب اللاحقة بينما في التجارب السابقة تم تحميل MncA على أعمدة Superdex 75 أو Superdex 200 SEC (كما هو محدد) مع تحليل الفractions بواسطة SDS-PAGE. تم تخفيف الفractions التي تحتوي على MncA إلىإعادة تطبيقها على Q-Sepharoseغسل بعد ذلك تم غسلها بمحلول 300 مللي مولار من NaCl وتم تحليل الفractions بواسطة SDS-PAGE،“، و[المعدن] بواسطة ICP-MS. تم حساب النسب المئوية (%) لاحتلال MncA مع كل معدن أولاً من نسبة [المعدن]/[MncA] بافتراض وجود موقعين معدنيين لكل جزيء MncA، وتم تأكيد ذلك من خلال متوسط احتلال تجريبي قدره 99% (التكرارات 1-3 في الجدول التكميلي 4a، b). تم تبسيط البروتوكول إلى خطوتين حيث تم استرداد MncA أولاً عبر تبادل الأنيونات (عمود Q-Sepharose بسعة 5 مل، تدرج 0-300 مليمول من NaCl) تلاه فصل سريع على مقياس تحليلي باستخدام عمود SW3000 (TSK).تم تحليل الكسور مرة أخرى بواسطة SDS-PAGE، و ICP-MS. تم تنقية النسخة البيولوجية الرابعة المقارنة من MncA المستخرجة من خلايا نمت بدون إضافة المعادن عبر الطريقة المبسطة، مما أدى إلى الحصول على نسب معدنية مشابهة (الجدول التكميلي 4). بالإضافة إلى ذلك، تم حساب نسب مشابهة كنسبة من إجمالي محتوى المعادن في MncA لتجنب التباين في MncA.مع معادن مختلفة (الجدول التكميلي 4ب). في الثقافات المدعمة بالمعادن اللاحقة، تمت إضافة المعادن المعنية عند التلقيح 1-3 ساعات قبل إضافة L-arabinose. تم عزل MncA من خلايا مدعمة بالنحاس وتمت تنقيته مع بروتين نحاسي تم التعرف عليه بشكل مؤقت على أنه GAPDH. تم تمرير الفractions التي تحتوي على MncA عبر عمود Cibacron blue Sepharose (Blue Sepharose) بحجم 5 مل متوازن مع 50 مليمول من Tris pH 7.5. تم إزاحة MncA بسرعة بينما تم الاحتفاظ بالبروتين الملوث ثم تم إزاحته باستخدام 1 مليمول من NaCl في 50 مليمول من Tris pH 7.5. تم إخضاع محلول MncA المعالج بـ Blue Sepharose إلى SEC التحليلية كما هو موضح أعلاه.
تقدير وفرة النسخ في الإشريكية القولونية
مقتطفانتم جمعها من E. coli، بما في ذلك من خلايا تحتوي على pBAD30-mncA التي تعبر عن MncA مباشرة قبل إضافة الأربيانوز، وثانياً بعد النمو طوال الليل، وتم تثبيت RNA باستخدام مادة RNAProtect Bacteria Reagent، 2 مل (Qiagen). تم معالجة العينات كما هو موضحباختصار، تم استخراج RNA باستخدام مجموعة RNeasy Mini (Qiagen)، [تم تقدير RNA منثم تم معالجته بـ DNase I (فيرمنتاس). تم إنتاج cDNA باستخدام نظام ImProm-II للنسخ العكسي (بروماجا)، مع تفاعلات تحكم متوازية تستبعد النسخ العكسي. تم تحديد وفرة النسخ باستخدام بادئات لـ mntS و fepD و rcnA و nikA و znuA و zntA و copA و rpoD التي تضخملـ DNA مع تسلسلات مدرجة في البيانات التكميلية 9. تم تنفيذ تحليل تفاعل البوليميراز المتسلسل الكمي (qPCR) فيتفاعلات تحتوي على 5 نانوغرام من cDNA، و400 نانومتر من كل بادئ مكمل وPowerUP SYBR Green Master Mix (Thermo Fisher Scientific). تم تحليل ثلاث نسخ فنية من كل نسخة بيولوجية باستخدام جهاز Rotor-Gene Q 2plex (Qiagen؛ برنامج Rotor-Gene-Q Pure Detection) مع تفاعلات تحكم إضافية بدون قوالب cDNA (تم استخدام ماء بدرجة qPCR بدلاً من ذلك، مقدم من Thermo Fisher Scientific) تم تشغيلها لكل زوج من البادئات، بالإضافة إلى تفاعلات تحكم بدون إنزيم النسخ العكسي لزوج بادئات الجين المرجعي (rpoD).تم حساب القيم باستخدام LinRegPCR (الإصدار 2021.1) بعد تصحيح كفاءة الأمبليكون (بيانات المصدر التكميلية TXT، الموضحة في الشكل 7). تم حساب التغير في وفرة الجين، بالنسبة لحالة التحكم (المعرفة على أنها الحالة التي تم فيها ملاحظة الحد الأدنى من وفرة النسخ لكل جين مستهدف)، باستخدامطريقةباستخدام الجين المرجعي rpoD ومُقدّم كـ (تغيير الطي).
توفر المعادن داخل الخلايا وتوقعات التمثيل المعدني في الجسم الحي
استجابات حساسات المعادن (لتواجدات الحمض النووي لعوامل إزالة الكبت المعتمدة على المعادن وعوامل الكبت المشتركة،للمنشطات المعدنية) كدالة لتركيزات المعادن المدعومة المتاحة داخل الخلايا تم حسابها باستخدام تقارب المعادن في المستشعرات، وتقارب الحمض النووي، ووفرة البروتينات، وعدد مواقع ارتباط الحمض النووي لـأجهزة استشعار الإشريكية القولونية كما هو موصوف (البيانات التكميلية 2 لـ تمت مقارنة وفرة النسخ مع منحنيات الاستجابة لتمكين تقديرات توافر المعادن داخل الخلايا المعبر عنه كطاقة حرة لتكوين المركبات. ) في . القولونية تنمو هوائياً في LB كما هو موصوفباستخدام هذه التوافرات، تم التنبؤ بعملية المعادن للبروتينات في الجسم الحي، مع الأخذ في الاعتبار المنافسات المتعددة بين المعادن بما في ذلك المنافسة من العازل داخل الخلوي كما وصفه يونغ وزملاؤه.البيانات التكميلية 3 و 4 تقوم بإجراء هذه الحسابات للخلايا المثالية (أجهزة الاستشعار في النطاق المتوسط، بما في ذلك ) وتوافر المعادن داخل الخلايا في . القولونية التي نمت هوائيًا في LB على التوالي.
في الثقافات المدعمة بالمعادن، (تغير الطي) وفرة النسخ النسبية في هذا العمل تقارب القيم المبلغ عنها سابقًا مع تجاوز بعض الحدود (الشكل 7). تم استبدال القيم المبلغ عنها لتوافر المعادن المدعمة داخل الخلايا العالية في البيانات التكميلية 4، مع الاحتفاظ بالقيم الأصلية لجميع المعادن الأخرى، لإجراء التنبؤات الأولى لتأين المعادن في MncA عند مستويات عالية داخل الخلايا. أو . ل و تم استخدام الفروق المتبقية بين التنبؤات والملاحظات لعملية ميتاليشن MncA بشكل تكراري لتحسين توافر المعادن داخل الخلايا وفقًا للملاحظة التكميلية 2 واستخدام البيانات التكميلية 5. تحتوي البيانات التكميلية 6-8 على التحسينات ويمكن استخدامها للتنبؤ بعملية ميتاليشن بروتينات أخرى في E. coli في ظروف مرتفعة.، “ أو .
تحليلات العناصر بواسطة ICP-MS وخلايا ذرات المعادن
تم إجراء تحليل ICP-MS في منشأة Bio-ICP-MS بجامعة دورهام (نموذج ThermoFisher iCAP RQ) باستخدام منحنيات معيارية متطابقة مع المصفوفة ومعايير داخلية من الفضة. E. coliفراء وتم الحصول على السلالات من مجموعة كيوتم تلقيح مستعمرات فردية من E. coli أو الطفرات فياهتزاز عندلـمخفف في وسط جديدفي أنبوب طرد مركزي مخروطي سعة 50 مل مضاف إليه المعادن ) حيثما تم تحديده، إلى تم حضانة الخلايا مع التحريك فيبين عشية وضحاهامخفف 1:10 للقياسوتم استرداد الخلايا (من 9.9 مل المتبقية) عن طريق الطرد المركزي. تم غسل الرواسب أربع مرات عن طريق إعادة التعليق في 1 مل من محلول الغسيل.تريسسوربيتول، 0.2 مللي مول EDTA) تليها الطرد المركزي. أولترابورميركتمت إضافة إلى كل حبة وسمح لها بالت incubate لمدة لا تقل عن 16 ساعة حتى يتم هضمها بالكامل. ثم تم إعداد العينات لـ ICP-MS مع منحنيات معايرة متطابقة مع المصفوفة. من 1 يعادل عدد خلايا (عداد خلايا CASY) من خلايا استخدام هذا السلالة، مما يمكّن الذرات الخلوية ليتم حسابه من بيانات ICP-MS المعايرة.
الإحصائيات وإمكانية التكرار
تم اختيار أحجام العينات بناءً على الخبرة التجريبية السابقة، ولتقديم نتائج متسقة، وفقًا للعرف في الأدبيات للتحليلات المعادلة. تم إجراء التجارب المصممة لاشتقاق قيم كمية تستخدم للتنبؤ أو الاختبار وقياس التمثيل المعدني، أو لتحسين وتقييم تقديرات توافر المعادن داخل الخلايا، ثلاث مرات.أو أكثرلتمكين حساب الانحراف المعياري (المُدرج في الجداول أو النص أو المعروض كأشرطة خطأ في الأشكال) أو الخطأ القياسي لـ-ت affinities. لا تنتشر توقعات التمعدن SD من القيم المساهمة. تم الحصول على كروماتوغرامات مماثلة للبيانات التمثيلية في الأشكال 4b و 5a في مناسبتين إضافيتين ( البيانات في الأشكال 4أ و 1ب تمثل 18 و التنقيات المماثلة على التوالي. يتم عرض عدد التجارب المستقلة أو العينات البيولوجية المستقلة بطريقة أخرى في أساطير الأشكال أو الحواشي السفلية للجداول.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة ناتشر المرتبط بهذه المقالة.
توفر البيانات
جميع البيانات متاحة ضمن المقال، وملفات المعلومات التكميلية الخاصة به، بالإضافة إلى إدخال PDB 9GOF ومن المؤلفين المقابلين عند الطلب. يتم توفير بيانات المصدر مع هذه الورقة كملفات بيانات المصدر.
جداول بيانات إكسل (مع تعليمات) توفر آلة حاسبة لصياغة العوازل المعدنية المتنافسة، لحساب شغل الحمض النووي كدالة لـتوافر مثبط RcnR المعتمد على المعادن وتوفير آلة حاسبة لاستخدام معدلات المعادن المستعادة في الجسم الحي لـ MncA كأداة لتحسين تقديرات توافر المعادن داخل الخلايا، تم تقديمها كبيانات إضافية 1 و 2 و 5 على التوالي. جداول بيانات Excel التي تشكل آلات حاسبة للتعدين في-تم توفير خلايا مثالية مصقولة، E. coli نمت هوائيًا في LB، E. coli نمت هوائيًا في LB مضاف إليه المنغنيز والنيكل والكوبالت، كبيانات إضافية 3، 4، 6-8 على التوالي. تحتوي البيانات الإضافية 9 على تسلسلات أوليغونيوكليوتيد. الهياكل المنشورة المستخدمة هنا لـ MncA و MntR هي إدخالات PDB 2VQA و 9C4D على التوالي. تم توفير بيانات المصدر مع هذه الورقة.
توفر الشيفرة
اشتقاقات المعادلات موجودة في الملاحظات التكميلية 1 و 2 من المعلومات التكميلية. تم توفير نصوص Dynafit في البرنامج التكميلية.
References
Waldron, K. J., Rutherford, J. C., Ford, D. & Robinson, N. J. Metalloproteins and metal sensing. Nature 460, 823-830 (2009).
Irving, H. & Williams, R. J. P. Order of stability of metal complexes. Nature 162, 746-747 (1948).
Da Silva, J. F. & Williams, R. J. P. The biological chemistry of the elements: the inorganic chemistry of life. (Oxford University Press, 2001).
Robinson, N. J. & Glasfeld, A. Metalation: nature’s challenge in bioinorganic chemistry. J. Biol. Inorg. Chem. 25, 543-545 (2020).
Foster, A. W., Osman, D. & Robinson, N. J. Metal preferences and metallation. J. Biol. Chem. 289, 28095-28103 (2014).
Grāve, K., Griese, J. J., Berggren, G., Bennett, M. D. & Högbom, M. The Bacillus anthracis class Ib ribonucleotide reductase subunit NrdF intrinsically selects manganese over iron. J. Biol. Inorg. Chem. 25, 571-582 (2020).
Cotruvo, J. A. Jr & Stubbe, J. Metallation and mismetallation of iron and manganese proteins in vitro and in vivo: the class I ribonucleotide reductases as a case study. Metallomics 4, 1020-1036 (2012).
Tottey, S. et al. Protein-folding location can regulate manganesebinding versus copper- or zinc-binding. Nature 455, 1138-1142 (2008).
Raux, E., Thermes, C., Heathcote, P., Rambach, A. & Warren, M. J. A role for Salmonella typhimurium cbiK in cobalamin (vitamin B12) and siroheme biosynthesis. J. Bacteriol. 179, 3202-3212 (1997).
Schubert, H. L., Raux, E., Wilson, K. S. & Warren, M. J. Common chelatase design in the branched tetrapyrrole pathways of heme and anaerobic cobalamin synthesis. Biochemistry 38, 10660-10669 (1999).
Lacasse, M. J., Douglas, C. D. & Zamble, D. B. Mechanism of selective nickel transfer from HypB to HypA, Escherichia coli [NiFe]hydrogenase accessory proteins. Biochemistry 55, 6821-6831 (2016).
Watanabe, S. et al. Structural basis of a Ni acquisition cycle for [NiFe] hydrogenase by Ni-metallochaperone HypA and its enhancer. Proc. Natl. Acad. Sci. 112, 7701-7706 (2015).
Banci, L. et al. The Atx1-Ccc2 complex is a metal-mediated proteinprotein interaction. Nat. Chem. Biol. 2, 367-368 (2006).
Yang, X., Li, H., Lai, T.-P. & Sun, H. UreE-UreG complex facilitates nickel transfer and preactivates GTPase of UreG in Helicobacter pylori. J. Biol. Chem. 290, 12474-12485 (2015).
Crouzet, J. et al. Nucleotide sequence and genetic analysis of a 13.1-kilobase-pair Pseudomonas denitrificans DNA fragment containing five cob genes and identification of structural genes encoding Cob(I)alamin adenosyltransferase, cobyric acid synthase, and
bifunctional cobinamide kinase-cobinamide phosphate guanylyltransferase. J. Bacteriol. 173, 6074-6087 (1991).
Blaby-Haas, C. E., Flood, J. A., de Crécy-Lagard, V. & Zamble, D. B. YeiR: a metal-binding GTPase from Escherichia coli involved in metal homeostasis. Metallomics 4, 488-497 (2012).
Weiss, A. et al. Zn-regulated GTPase metalloprotein activator 1 modulates vertebrate zinc homeostasis. Cell 185, 2148-2163. e2127 (2022).
O’Halloran, T. V. & Culotta, V. C. Metallochaperones, an intracellular shuttle service for metal ions. J. Biol. Chem. 275, 25057-25060 (2000).
Cavet, J. S. et al. A nickel-cobalt-sensing ArsR-SmtB family repressor: contributions of cytosol and effector binding sites to metal selectivity. J. Biol. Chem. 277, 38441-38448 (2002).
Guedon, E. & Helmann, J. D. Origins of metal ion selectivity in the DtxR/MntR family of metalloregulators. Mol. Microbiol. 48, 495-506 (2003).
Moomaw, E. W. et al. Metal dependence of oxalate decarboxylase activity. Biochemistry 48, 6116-6125 (2009).
Young, T. R. et al. Calculating metalation in cells reveals CobW acquires Coll for vitamin B12 biosynthesis while related proteins prefer ZnII. Nat. Commun. 12, 1195 (2021).
Osman, D. et al. Bacterial sensors define intracellular free energies for correct enzyme metalation. Nat. Chem. Biol. 15, 241-249 (2019).
Grossoehme, N. E. & Giedroc, D. P. Energetics of allosteric negative coupling in the zinc sensor S. aureus CzrA. J. Am. Chem. Soc. 131, 17860-17870 (2009).
Foster, A. W. et al. A tight tunable range for sensing and buffering in cells. Nat. Chem. Biol. 13, 409-414 (2017).
Outten, C. E. & O’Halloran, T. V. Femtomolar sensitivity of metalloregulatory proteins controlling zinc homeostasis. Science 292, 2488-2492 (2001).
Foster, A. W. et al. Metalation calculators for E. coli strain JM109 (DE3): aerobic, anaerobic, and hydrogen peroxide exposed cells cultured in LB media. Metallomics 14, mfac058 (2022).
Young, T. R. et al. Two distinct thermodynamic gradients for cellular metalation of vitamin B12. JACS Au 3, 1472-1483 (2023).
Choi, T. S. & Tezcan, F. A. Overcoming universal restrictions on metal selectivity by protein design. Nature 603, 522-527 (2022).
Chivers, P. T., Basak, P. & Maroney, M. J. One His, two His… the emerging roles of histidine in cellular nickel trafficking. J. Inorg. Biochem. 259, 112668 (2024).
Krämer, U., Cotter-Howells, J. D., Charnock, J. M., Baker, A. J. M. & Smith, J. A. C. Free histidine as a metal chelator in plants that accumulate nickel. Nature 379, 635-638 (1996).
Ma, Z. et al. Bacillithiol is a major buffer of the labile zinc pool in Bacillus subtilis. Mol. Microbiol. 94, 756-770 (2014).
Murphy, J. T. et al. Histidine protects against zinc and nickel toxicity in Caenorhabditis elegans. PLoS Genet. 7, e1002013 (2011).
Nairn, B. L. et al. The response of Acinetobacter baumannii to zinc starvation. Cell Host Microbe 19, 826-836 (2016).
Morgan, M. T., Nguyen, L. A. H., Hancock, H. L. & Fahrni, C. J. Glutathione limits aquacopper(I) to sub-femtomolar concentrations through cooperative assembly of a tetranuclear cluster. J. Biol. Chem. 292, 21558-21567 (2017).
Brawley, H. N. & Lindahl, P. A. Low-molecular-mass labile metal pools in Escherichia coli: advances using chromatography and mass spectrometry. J. Biol. Inorg. Chem. 26, 479-494 (2021).
Stewart, L. J. et al. Role of glutathione in buffering excess intracellular copper in Streptococcus pyogenes. MBio 11, 02804-02820 (2020).
Hider, R. C. & Kong, X. L. Glutathione: a key component of the cytoplasmic labile iron pool. BioMetals 24, 1179-1187 (2011).
Osman, D. et al. Fine control of metal concentrations is necessary for cells to discern zinc from cobalt. Nat. Commun. 8, 1884 (2017).
Giedroc, D. P. & Arunkumar, A. I. Metal sensor proteins: nature’s metalloregulated allosteric switches. Dalton Trans. 29, 3107-3120 (2007).
Pennella, M. A., Arunkumar, A. I. & Giedroc, D. P. Individual metal ligands play distinct functional roles in the zinc sensor Staphylococcus aureus CzrA. J. Mol. Biol. 356, 1124-1136 (2006).
Aguirre, J. D. & Culotta, V. C. Battles with iron: manganese in oxidative stress protection. J. Biol. Chem. 287, 13541-13548 (2012).
Anjem, A., Varghese, S. & Imlay, J. A. Manganese import is a key element of the OxyR response to hydrogen peroxide in Escherichia coli. Mol. Microbiol. 72, 844-858 (2009).
Tarrant, E. et al. Copper stress in Staphylococcus aureus leads to adaptive changes in central carbon metabolism. Metallomics 11, 183-200 (2019).
Abdul-Tehrani, H. et al. Ferritin mutants of Escherichia coli are iron deficient and growth impaired, and fur mutants are iron deficient. J. Bacteriol. 181, 1415-1428 (1999).
Mills, S. A. & Marletta, M. A. Metal binding characteristics and role of iron oxidation in the ferric uptake regulator from Escherichia coli. Biochemistry 44, 13553-13559 (2005).
Carter, K. P., Young, A. M. & Palmer, A. E. Fluorescent sensors for measuring metal ions in living systems. Chem. Rev. 114, 4564-4601 (2014).
Morgan, M. T. et al. Ratiometric two-photon microscopy reveals attomolar copper buffering in normal and Menkes mutant cells. Proc. Nat. Acad. Sci. 116, 12167-12172 (2019).
Nolan, E. M. & Lippard, S. J. Small-molecule fluorescent sensors for investigating zinc metalloneurochemistry. Acc. Chem. Res. 42, 193-203 (2009).
Zou, Z., Higginson, B. & Ward, T. R. Creation and optimization of artificial metalloenzymes: Harnessing the power of directed evolution and beyond. Chem 10, 2373-2389 (2024).
Harnden, K. A., Wang, Y., Vo, L., Zhao, H. & Lu, Y. Engineering artificial metalloenzymes. Protein Eng., 177-205 (2021).
Helalat, S. H., Jers, C., Bebahani, M., Mohabatkar, H. & Mijakovic, I. Metabolic engineering of Deinococcus radiodurans for pinene production from glycerol. Microb. Cell Fact. 20, 1-14 (2021).
Shomar, H. & Bokinsky, G. Towards a synthetic biology toolset for metallocluster enzymes in biosynthetic pathways: what we know and what we need. Molecules 26, 6930 (2021).
Arnold, F. H. Directed evolution: bringing new chemistry to life. Angew. Chem. Int. Ed. 57, 4143 (2018).
United Kingdom Government, D. f. S. I. a. T. National Vision For Engineering Biology, https://www.gov.uk/government/ publications/national-vision-for-engineering-biology (2023).
Alderighi, L. et al. Hyperquad simulation and speciation (HySS): a utility program for the investigation of equilibria involving soluble and partially soluble species. Coord. Chem. Rev. 184, 311-318 (1999).
Smith, R. M. & Martell, A. E. Critical stability constants. 1 (Amino acids, 1974).
Winter, G. xia2: an expert system for macromolecular crystallography data reduction. J. Appl. Crystallogr. 43, 186-190 (2010).
Kabsch, W. xds. Acta Crystallogr. D. Biol. Crystallogr 66, 125-132 (2010).
Winn, M. D. et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D. Biol. Crystallogr 67, 235-242 (2011).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658-674 (2007).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D. Biol. Crystallogr 66, 213-221 (2010).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D. Biol. Crystallogr 66, 486-501 (2010).
Meng, E. C. et al. UCSF ChimeraX: Tools for structure building and analysis. Protein Sci. 32, e4792 (2023).
Pravda, L. et al. MOLEonline: a web-based tool for analyzing channels, tunnels and pores (2018 update). Nucleic Acids Res. 46, W368-W373 (2018).
Osman, D. et al. Generating a metal-responsive transcriptional regulator to test what confers metal sensing in cells. J. Biol. Chem. 290, 19806-19822 (2015).
Foster, A. W., Patterson, C. J., Pernil, R., Hess, C. R. & Robinson, N. J. Cytosolic Ni(II) sensor in cyanobacterium: nickel detection follows nickel affinity across four families of metal sensors. J. Biol. Chem. 287, 12142-12151 (2012).
Kuzmič, P. Program DYNAFIT for the analysis of enzyme kinetic data: application to HIV proteinase. Anal. Biochem. 237, 260-273 (1996).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the method. Methods 25, 402-408 (2001).
Baba, T. et al. Construction of Escherichia coli K-12 in-frame, singlegene knockout mutants: the Keio collection. Mol. Sys. Biol. 2, 2006 (2006).
شكر وتقدير
تم الاعتراف بمصدر ضوء الماس للوقت على خط الشعاع IO4 بموجب الاقتراح MX32736، البروفيسور تيموثي بلور، البروفيسور ليز موريس للمساعدة في جمع ومعالجة بيانات البلورات. تم دعم هذا العمل من قبل جوائز مجلس بحوث العلوم البيولوجية والتكنولوجيا BB/W015749/1 (N.J.R.)، فهم التغيرات غير الطبيعية في المعادن للبروتينات الأصلية مقابل البروتينات المعبر عنها بشكل غير متجانس، وBB/V006002/1 (N.J.R.)، آلة حاسبة لتحديد المعادن داخل الخلية، بالإضافة إلى BB/S009787/1 (N.J.R.) التي تدعم الشبكات في التكنولوجيا الحيوية الصناعية. يقر المؤلفون بمساهمات دينا مورتون (née عثمان) التي تم تكريس العمل لها.
مساهمات المؤلفين
ساهمت T.R.Y. و S.E.C. و A.G. و E.T. و N.J.R. في تجارب ربط المعادن MncA في الجسم الحي. ساهمت T.R.Y. و A.G. و S.E.C. و E.T. في تجارب إعادة الطي في المختبر، حيث قامت T.R.Y. بأعمال أساسية مع S.E.C. و A.G. التي أكملت الجزء الأكبر من هذه التجارب. استخلصت T.R.Y. حسابات لصياغة العوازل المعدنية المتنافسة واستخدام MncA كأداة لتحسين تقديرات توافر المعادن داخل الخلايا وصممت بروتوكولات تجريبية ذات صلة. أنشأت A.G. الهيكل البلوري لـ-MncA. تم تحليل P.T.C. و A.J.P.S.وخصائص ارتباط الحمض النووي لـ Nill-RcnR. A.G. و S.E.C. و E.T. حددوا محتويات المعادن في خلايا E. coli. قامت S.E.C. بإجراء وتحليل وفرة النسخ باستخدام qPCR. العلاقات بين إشغالات الحمض النووي لمستشعرات المعادن، وتوافر المعادن داخل الخلايا، وبيانات qPCR، استخدمت طرقًا وضعتها T.R.Y. و N.J.R. طورت T.R.Y. حسابات التمعدن بالشكل المستخدم هنا. كتب N.J.R. وS.E.C.، بمساعدة A.G. وE.T.، المسودة الأولى من المخطوطة وأنشأوا الرسوم البيانية بينما قام جميع المؤلفين بتحرير واعتماد النسخة النهائية. ساهم T.R.Y. ثم S.E.C. ثم A.G. بشكل متسلسل في الجزء الأكبر من العمل المختبري في مراحل مختلفة من البرنامج، حيث قدم T.R.Y. وS.E.C. مساهمات كبيرة بشكل متساوٍ. قام N.J.R. بدعم من جميع المؤلفين، وخاصة T.R.Y. وS.E.C. وA.G.، بتفسير أهمية البيانات. كان N.J.R. مسؤولاً بشكل عام عن تصميم المشروع وتمويله وإدارته طوال فترة المشروع.
يجب توجيه المراسلات والطلبات للحصول على المواد إلى نايجل ج. روبنسون.
معلومات مراجعة الأقران تشكر مجلة Nature Communications إريك يوكيل والمراجعين الآخرين المجهولين على مساهمتهم في مراجعة هذا العمل. يتوفر ملف مراجعة الأقران.
ملاحظة الناشر: تظل شركة سبرينغر ناتشر محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
الوصول المفتوح هذه المقالة مرخصة بموجب رخصة المشاع الإبداعي النسب 4.0 الدولية، التي تسمح بالاستخدام والمشاركة والتكيف والتوزيع وإعادة الإنتاج بأي وسيلة أو صيغة، طالما أنك تعطي الائتمان المناسب للمؤلفين الأصليين والمصدر، وتوفر رابطًا لرخصة المشاع الإبداعي، وتوضح إذا ما تم إجراء تغييرات. الصور أو المواد الأخرى من طرف ثالث في هذه المقالة مشمولة في رخصة المشاع الإبداعي الخاصة بالمقالة، ما لم يُشار إلى خلاف ذلك في سطر الائتمان للمواد. إذا لم تكن المادة مشمولة في رخصة المشاع الإبداعي الخاصة بالمقالة وكان استخدامك المقصود غير مسموح به بموجب اللوائح القانونية أو يتجاوز الاستخدام المسموح به، فسيتعين عليك الحصول على إذن مباشرة من صاحب حقوق الطبع والنشر. لعرض نسخة من هذه الرخصة، قم بزيارةhttp://creativecommons.org/رخصة/بواسطة/4.0/. (ج) المؤلف(ون) 2025
¹قسم علوم الحياة، جامعة دورهام، دورهام، المملكة المتحدة. ²قسم الكيمياء، جامعة دورهام، دورهام، المملكة المتحدة.ساهم هؤلاء المؤلفون بالتساوي: صوفي إي. كلوف، تيسا آر. يونغ. البريد الإلكتروني:nigel.robinson@durham.ac.uk
نسبة توافر المعادن المخففة وفقًا للجدول التكميلي 1 والبيانات التكميلية 1. نسبة كل معدن متنافس تم الحصول عليها بواسطة MncA كما هو محدد من الكروماتوغرامات في الشكل 1d. تُحسب تفضيل M 2 كنسبة التركيزات المخزنة لـ M 1 وعمودمقسومًا على نسب إشغالات المعدن 1 والمعدن 2عمودما لم يُذكر خلاف ذلك. محسوب لـ M2 (Cu’) بالنسبة إلى Mn” استنادًا إلى التفضيل الملحوظ لـبالنسبة إلى. بالنسبة لـ M2 بالنسبة لقيمة معينة منلـ (المحدد في الأسطورة الخاصة بالشكل 2e والمفصل في الأشكال التكميلية 5a و 5b). كثوابت زائفة للتفكك: ; الحديد الثنائي ; ; . المنافسة في التجربة الرابعةمعغير موضح في الشكل 1c والكروماتوغرامات في الشكل التكميلي 2b. هذه النتيجة تتعلق بالتفضيل المحسوب لـ (حيث التكرارات التجريبيةللتحديدات الأخرىSD).
A metal-trap tests and refines blueprints to engineer cellular protein metalation with different elements
Received: September 2024
Accepted: 10 January 2025
Published online: 18 January 2025
Sophie E. Clough , Tessa R. Young , Emma Tarrant , Andrew J. P. Scott , Peter T. Chivers , Arthur Glasfeld & Nigel J. Robinson
It has been challenging to test how proteins acquire specific metals in cells. The speciation of metalation is thought to depend on the preferences of proteins for different metals competing at intracellular metal-availabilities. This implies mis-metalation may occur if proteins become mis-matched to metalavailabilities in heterologous cells. Here we use a cyanobacterial -cupin as a metal trap, to test predictions of metalation. By re-folding MncA in buffered competing metals, metal-preferences are determined. Relating metal-preferences to metal-availabilities estimated using cellular metal sensors, predicts mis-metalation of MncA with in E. coli. After expression in . coli, predominantly -bound MncA is isolated experimentally. It is predicted that in metal-supplemented viable cells metal-MncA speciation should switch. – – , or are recovered from the respective metalsupplemented cells. Differences between observed and predicted metal-MncA speciation are used to refine estimated metal availabilities. Values are provided as blueprints to guide engineering biological protein metalation.
A purpose of this research is to test explanations of how proteins acquire different metals in cells: the speciation of metalation in biology (Supplementary Fig. 1). Metalloenzyme catalysis is mostly metalspecific. Yet metalloproteins typically bind one or more wrong metals in preference to the cognate metal(s) . Nascent metal sites in proteins are flexible such that non-cognate metals can bind non-conservatively by using a subset of the native ligands, by recruiting additional ligands and/or by adopting non-cognate coordination geometries . With such limited constraint the order of metal binding commonly follows the Irving-Williams series (Fig. 1a) . An exception is where there has been prior structural organisation for example via cooperativity in di-metal sites . Here we quantify the binding preferences of a protein (MncA) which kinetically traps metals during folding . We then use MncA to establish if protein metalation can be correctly predicted, then predictably adjusted, and finally to refine estimates of intracellular metal availability.
In combination with the metal-binding preferences of proteins, a second factor determining metal-protein speciation is metal availability . Metal partitioning can occur directly from pools of labile, exchangeable and hence available metals to the protein of interest, or to an assembly pathway for a small molecule cofactor (such as into heme or into vitamin ), or to a metallochaperone that delivers to an assembly pathway or to the protein of interest . Here we explore the initial partitioning step from labile, available intracellular metal pools.
The significance of intracellular metal availability to metal-protein speciation emerged over several decades. When expressed in a cyanobacterial cell a – and -responsive DNA-binding metal-sensor from Mycobacterium tuberculosis solely responded to . This change in specificity was attributed to being insufficiently available inside viable cyanobacteria to metalate NmtR. Similarly, Fell-responsive DtxR from Corynebacterium diphtheriae gained responsiveness to
Fig. 1 | Relative metal preferences of MncA folded in competing buffered metals. a The Irving-Williams order of formation of complexes with metals, as the ionic states of exchangeable metals in the cytosol. From weak (left) to tight, noting reversal of arrow after copper. b Protocol for recovery of unfolded MncA from inclusion bodies after high-level expression in E. coli BL21(DE3) pLysS for 3 h using pET29a at , followed by folding denatured MncA via dropwise dilution into a large volume of urea-free buffer containing pairs of buffered and competing metal ions. Folded MncA was concentrated by anion exchange and eluted with single-step high-salt buffer. Metal-bound MncA was separated from unbound metal by SEC (PD-10 column), and metal analysed by ICP-MS. Gel inset shows, from left, size markers (from top, ), overloaded urea-solubilised MncA and MncA eluted from Q-Sepharose (full gel Supplementary Fig. 2a). c Metal contents of MncA-containing fractions after folding in excesses of competing metals buffered to different ratios of availabilities (as in panel , buffers in Supplementary Table 1 prepared with assistance from Supplementary Data 1). Ratios of competing metals above each panel. The concentration of MncA approximated from . Stoichiometries approximate but proportional metalation was calculated from total metal as in Table 1. Experiments with and performed in an anaerobic chamber, metal stocks prepared freshly, hydroxylamine ( 1 mM ) used to sustain reduced copper. and confirmed to be reduced via reaction with excess ferrozine or BCA respectively. independent experimental replicates shown. d Quantified preferences of MncA for different divalent metals, plus monovalent copper, relative to ( independent experiments, triangles, squares, and circles depict replicates in (c), (I) and (II) denote forms of copper), determined from the elution profiles in (c). The order of preferences follows the Irving-Williams series as in (a). MncA prefers all noncognate metals over . Source data are provided as a Source Data file.
when expressed in Bacillus subtilis, again attributed to different intracellular availabilities in the different cells . Two metalbinding cupins, and , were discovered in the periplasm of a cyanobacterium (Synechocystis PCC 6803), notably binding metals from different ends of the Irving-Williams series yet exploiting similar folds and identical metal-binding residues . When folded in vitro in similar amounts of competing copper versus , or versus , the non-cognate metals, copper or , bound to . While the copper-cupin is secreted unfolded via the Sec-system, the -cupin is a Tat-substrate which folds and kinetically traps the less competitive metal in the cytosol before secretion. Thus, metal availability at the site of protein folding determines metal speciation, and must be more available than or in the cyanobacterial cytosol . MncA is an -dependent oxalate decarboxylase (inactive with copper or ), and the related cytosolic OxdC from Bacillus subtilis trapped either or in E. coli depending on media metal supplementation . MncA thus offers enticing opportunities to directly interrogate mechanisms and predictions of metalation because (1) kinetically trapped metals are unlikely to exchange during purification, and so MncA should faithfully report its in vivo metalation state, and (2) precedent suggests it could be possible to switch the speciation of MncA metalation in E. coli and discover if various metalation states are predictable.
We recently developed a metalation calculator which accounts for inter-metal competition within cells . The calculations use estimations of intracellular metal availabilities derived from thermodynamically calibrated responses of the cells’ DNA-binding metalsensing transcriptional regulators (Supplementary Fig. 1) . Availabilities are described as free energies consistent with bound but labile metals capable of rapid associative ligand exchange with the protein of interest . By using estimates of intracellular availabilities standardised to the mid-points of the sensor ranges (representing idealised cells) the cognate metals of four exemplar proteins have been correctly decoded . These data encourage a view that our understanding of the speciation of metalation is (broadly) correct. However, unlike in idealised cells, metal sensors will be at different positions in their ranges in actual cells depending on growth conditions. The abundance of transcripts encoded by metal-sensor regulated genes was estimated by qPCR and then calibrated to estimate metal availabilities inside . coli grown in un-supplemented medium . To date, calculations of protein metal speciation have only been experimentally tested indirectly in E. coli engineered to manufacture vitamin . Co -dependent production of was thus measured as a proxy for the metalation states of the metallochaperone CobW, and the chelatase CobNST. This work now
tests predictions directly by using cyanobacterial MncA to read out in vivo metalation when expressed in . coli cells under different growth conditions.
Here we determine whether metalation calculations correctly predict mis-metalation speculated to occur in engineered cells where metal availabilities are mismatched to the metal preferences of heterologously expressed proteins. In cells supplemented with cobalt, nickel and manganese, residual differences between predicted and observed MncA metalation are also used to refine estimated availabilities of other metals, and we test if changes in metal atoms cell align with these refinements. We explore the mechanism by which exposure to one metal (eg cobalt) can change availability of another (eg ). Calculators are included to enable use of MncA to probe metal availabilities under other growth conditions and in other cell types. Efforts are being made to engineer proteins that overcome the IrvingWilliams series . These approaches place constraints on, for example, bi-metallic catalytic centres. In contrast, here we show how biology can be exploited in a predictable way to overcome the challenge presented by the Irving-Williams series, thus expanding the repertoire of metaldriven biocatalysis that can be predictably utilised. Calculators are provided to guide the optimisation of protein metalation with different metals.
Results
Metal preferences at folding and trapping can also follow the Irving-Williams series
The first objective was to measure the metal-binding preferences of MncA in vitro. It is not feasible to measure affinities because is entrapped within the folded protein such that off-rates become negligible . Instead, relative preferences during folding were determined. Rapid, high-level, expression of MncA (minus secretion signal
peptide) in . coli produces MncA-containing inclusion bodies from which unfolded apo-protein can be recovered . MncA was thus expressed in E. coli BL21(DE3) pLysS, and urea-solubilised MncA, shown in Fig. 1b, was refolded by dilution into urea-free buffer. Refolding solutions contained pairs of competing metals buffered with NTA (or histidine for competitions) as in Supplementary Table 1, formulated via Supplementary Note 1 using the provided calculator (Supplementary Data 1). Competitions involving and were performed in an anaerobic chamber with -purged buffers and metal stocks confirmed > 95% reduced immediately prior to use. Metals were unbuffered in competitions between and . Refolded MncA (Fig. 1b), recovered by anion exchange chromatography, was resolved from unbound metal by size exclusion chromatography (SEC) with fractions ( 0.5 mL ) analysed for MncA by UV absorbance and metals by ICP-MS.
The proportion of each metal acquired by MncA was determined from the chromatograms in Fig. 1c. Challenges in generating NTAbuffered competitions initially led to competitions being performed without buffer before employing histidine-buffers. An extra replicate of histidine-buffered -competition was also performed (Supplementary Fig. 2, Supplementary Table 2). Competition between and bicinchoninic acid (BCA) buffered confirmed that MncA has -fold preference for , consistent with the determined -fold preference (Supplementary Fig. 2d, Table 1). Preferences of metal-binding to MncA at folding relative to were calculated from Table 1 to generate Fig. 1d. The order of binding follows the IrvingWilliams series (Fig. 1a). The exchangeable forms of metals in the cytosol are thought to be divalent except copper, which is monovalent. Fig. 1d illustrates the challenge to predict the metalation states of proteins in vivo and to decode cognate metals, since here seems least likely.
Table 1 | Preferences for metals trapped by MncA relative to
Metal1
Metal2
[Metal1] [Metal2]
Metal1 (%)
Metal2 (%)
Metal1/Metal2
Preference
Average preference
Fe”
56.7
43.3
1.31
Fe”
45.9
54.1
0.85
-50.8
Fe”
19.3
80.7
0.24
Co
20.1
79.9
0.25
Co
43.1
56.9
0.76
-55.6
Co
65.9
34.1
1.93
57.8
42.2
1.37
76.8
23.2
3.31
-65.3
Ni
39.3
60.7
0.65
51.3
48.7
1.06
Zn”
65.9
34.1
1.93
17.3
82.7
0.21
-65.5
3.1
96.9
0.03
Cu”
7.0
93
0.08
Cu
34.1
65.9
0.52
-79.1
Cu”
65.6
34.4
1.91
32.0
68.0
0.47
8.3
91.7
0.09
-69.5
29.5
70.5
0.42
Fig. 2 | -RcnR refined mid-range metal availabilities decode as the cognate MncA metal. a Apo-subtracted difference spectra of RcnR ( RcnR monomer, calculated from total protein) titrated with , inset showing peak wavelength confirming 1:1 stoichiometry of to RcnR monomer, or 4:1 to RcnR4 ( ). RcnR ( monomer) also migrated with one equivalent of by SEC (Supplementary Fig. 3b). b Representative titration of RcnR ( monomer) in EGTA , solid line representing calculated for from simultaneously fitted RncR-EGTA competitions ( independent experiments) at varied EGTA concentrations (Supplementary Fig. 3c, fitting models in Supplementary Software). Dashed lines, simulations with affinities 10 -fold tighter and weaker than calculated . c RcnR binding to hexachlorofluorescein-labelled operator-promoter by fluorescence anisotropy. Solid line, best simultaneous fit to experimental replicates (circles, triangles, squares) for (fitting model in Supplementary Software), dashed line simulates apo-RcnR using published and maximum and DNA affinities determined above used with previously measured RcnR molecules cell to calculate (via Supplementary Data 2) relationship between intracellular availability and RcnR DNA occupancy , as for (circles show and 0.01 ). Combined mid-point of ranges for and also shown (blue arrow). e Metal availabilities (squares and inset text) as activities/concentrations and free energies, , at mid-points ( DNA occupancies, representing idealised cells) of metal sensors (bars are sensor ranges, to ), now including -RcnR. Pale bars show individual ranges where two cognate sensors. MncA metal preferences (pale blue circles, triangles, from Fig. 1d) as free energies ( ) from pseudoaffinities giving metalation at mid-range availability ( ) without competing metals (Supplementary Fig. 5 simulates alternative values for . Inset shows occupancies of MncA predicted from free energy differences between MncA and labile metal ( ). has the largest favourable gradient annotated . Supplementary Data 3 enables similar predictions of cognate metals for other proteins. Source data are provided as a Source Data file.
availabilities defined by -RcnR refine mid-range metal availabilities
Figure 1d presents the metal preferences of MncA after accounting for competition from other ligands (NTA or histidine). The speciation of protein metalation in the crowded cytosol is similarly thought to result from competition with diverse ligands binding labile, exchangeable metals at different availabilities . Metal availabilities in the cytosol of bacterial cells (Salmonella and E. coli) have been estimated as free energies ( ): formally free energies for complex formation with a notional half-metalated ligand at the respective availability . DNAbinding metal sensors detect changes in and their responses have been calibrated for these bacteria . However, the responses of high -sensing RcnR were previously overlooked, and its calibration requires determination of affinity plus DNA affinity of -RcnR (Supplementary Fig. 3a) .
RcnR absorbance changes with and a difference spectrum saturates at one equivalent per monomer, or (Fig. 2a). RcnR co-elutes with one atom per monomer by SEC (Supplementary Fig. 3b). EGTA competes for with RcnR enabling determination
of RcnR Nill affinity from a simultaneous fit to titrations in different EGTA concentrations (Supplementary Fig. 3c). A representative data set confirms the fitted value is within the limits of the assay from dashed lines simulating values ten times tighter and weaker (Fig. 2b). Binding of apo- and of to DNA was previously monitored by fluorescence anisotropy using hexachlorofluorescein-labelled operator-promoter fragments, to determine DNA affinities . Here analogous titrations confirm that similarly weakens DNA-binding with fitted affinity of 3.09 (Fig. 2c). Using these values, with previous apo-RcnR DNA affinity of , plus known RcnR molecules cell , the relationship between intracellular availability and operatorpromoter occupancy was calculated as for (Supplementary Data 2) . rcnA transcripts increase as RcnR DNA occupancy decreases with elevated intracellular following the relationship in Fig. 2d. Metal availabilities at DNA occupancies of and 0.01 are indicated. Analogous relationships for other metal sensors are in Supplementary Fig. 4. The bars on Fig. 2e show ranges of availability (as ) corresponding to occupancies of . For the range in Fig. 2e now combines those for -NikR and -RcnR. The mid-point of the combined range is shown along with the mid-points for other metals, annotated as concentrations (M) and free energies (kJ mol-1) (Fig. 2e). The Irving-Williams series is ambiguous about the order of versus , but both are weaker than copper (Fig. 1a). Including RcnR, Fig. 2e reverses the order of intracellular availabilities of Zn and shown in previous iterations , but still the sensors maintain availabilities to the inverse of the Irving-Williams series .
-RcnR refined mid-range metal availabilities decode correct metalation
Metalation within a cell should be predictable from the binding preferences of MncA in Fig. 1d relative to how tightly available (exchangeable) intracellular metals are bound, for example at the midpoints of sensor ranges (Fig. 2e). To make this comparison, was assigned a pseudo-affinity ( ) giving -metalation
Fig. 3 | -MncA structure indicates non-cognate metal is also kinetically trapped. a Ribbon representation of crystal structure of at resolution showing characteristic bi-cupin fold and metal sites (boxed) modelled from residue 39 of the full protein sequence including signal peptide which is absent from the expressed protein (data collection and refinement statistics shown in Supplementary Table 3). Amino-terminal domain (pink), carboxy-terminal domain (blue) from residue 238 (in the full sequence). b, c Cross sections of solvent accessibility surfaces (modelled at solvent radius to encompass dynamics)
surrounding the metal sites show no channel to amino-terminal and narrow lipophilic (hydrophobic) channel to carboxy-terminal . becomes trapped in the folded protein in a near octahedral geometry (yellow bonds; Supplementary Fig. 6) analogous to cognate . The MncA model illustrates kinetic trapping of non-cognate metals suggesting MncA may be used to faithfully report in-cell metalation. Supplementary Fig. 7 shows that post-folding did not exchange with in vitro.
at the mid-point for , and free energies of metal-MncA complex formation were designated . Values ( ) for other metals were then calculated using Fig. 1d (Table 1). The gradients from exchangeable cytosolic sites to , were calculated and inter-metal competition accounted for using the Nill-RcnR-refined calculator in Supplementary Data 3, to predict occupancies (Fig. 2e). The cognate metal was correctly decoded (largest favourable ). Pseudo-affinities ten times tighter or weaker generated the same proportional occupancies (Supplementary Fig. 5a, b). Thus, the speciation of metalation is a function of relative metal-binding preferences and availabilities, but total metal occupancy does vary with absolute values for . Using the -RcnR-revised mid-point availabilities with four exemplar proteins whose affinities have been measured , decodes their cognate metals despite all preferring copper (Supplementary Fig. 5c-f). The -RcnR revised calculator in Supplementary Data 3 can be used to decode cognate metalation.
Non-cognate metal is also kinetically trapped by MncA
The labile character of protein-bound metals creates a challenge to define the in-cell metalation states of metalloproteins. Metals can be lost, gained, or exchanged at cell lysis and/or during purification and analysis. MncA is attractive because becomes trapped in the folded protein. However, MncA might adopt non-native folds with noncognate metals. Abnormally coordinated metals might not be
kinetically trapped. often prefers four-coordinate, planar geometries, and was chosen for structural analysis. A crystal structure of ( resolution, Supplementary Table 3, Supplementary Fig. 6a), shows the bi-cupin fold of with a metal atom in each cupin domain (Fig. 3a) . Each atom is coordinated to three histidine and one glutamate residue with non-protein ligands completing hexacoordinate coordination spheres. -MncA has similar coordination environments with no channel to the aminoterminal and a narrow channel to the carboxy-terminal site analogous to (Fig. 3b, c) . The channel is presumed to allow substrate access to catalytic . The substrate site was gratuitously occupied by acetate in the structure and with glycine in the crystalised -form (Supplementary Fig. 6, Fig. 3c). The narrow hydrophobic channel is unlikely to allow exchange and thus both atoms appear trapped.
MncA has -fold preference for over at folding (Fig. 1d). To test if is trapped, -MncA was incubated for 24 h in a two-fold molar excess of , bound and free metal separated by SEC followed by ICP-MS (Supplementary Fig. 7). The protein remained exclusively bound to confirming is kinetically trapped.
Mis-metalation of MncA with predicted and observed in E. coli
Figure 4 (unlike Fig. 2e) shows estimated metal availabilities in actual . coli BW25113 (elsewhere . coli) grown aerobically , and the metalation
of soluble MncA in E. coli. Transcripts regulated by metal-sensors were previously quantified by qPCR, then related to promoter occupancies and hence metal availabilities via the relationships in Supplementary Fig. . Notably, intracellular availability in aerobically grown cells is below the range for – RcnR . The largest favourable gradient from available exchangeable metal to , is for not cognate (Fig. 4c).
High-level protein expression in heterologous host cells can deplete cofactors, and likely contributed to formation of MncAcontaining inclusion bodies (Fig. 1b). To monitor in-cell metalation, MncA was thus expressed at a low level from a tuneable promoter using low inducer, low temperature, and prolonged slow growth overnight in E. coli (Fig. 4a). MncA was enriched from a soluble protein extract via anion exchange and SEC followed by ICP-MS. Metalation was quantified in three biologically independent experiments (Supplementary Fig. 8, Supplementary Table 4a, b). Absorbance at 280 nm largely reports MncA concentration due to its high extinction coefficient , evident from stoichiometry approximating 2:1, and suggesting no apo-protein (Supplementary Table 4a, b). Some metals might alter the extinction coefficient and traces of interfering proteins could introduce variation between experiments. Proportional occupancies have thus been calculated from total metal in one or more MncA-containing fraction(s) rather than MncA concentration, notably with similar outcomes when duplicated (Supplementary Table 4b). The experiment was repeated using a single anion exchange step and analytical HPLC SEC with similar outcome (Supplementary Table 4). The mean ± SD MncA-speciation is shown (inset Fig. 4c). Speciation accounting for inter-metal competition was calculated using Supplementary Data 4. Predicted mis-metalation closely matches observed mis-metalation in E. coli (Fig. 4b, c).
Substantial cognate MncA metalation predicted and observed in manganese
We were eager to know if intracellular could be increased to a tolerable availability inside viable . coli, sufficient to predominantly form . Fig. 4c shows metal preferences ( ) following a similar trend to availabilities ( ), predicting that modest changes could switch the speciation of metalation. Using only manganese supplementation maximum intracellular availability was detected in media plus 4 mM manganese . Notably, high manganese and hydrogen peroxide defined the 0.99 sensor boundary (Supplementary Fig. 4) , because manganese import is modulated by . Here, 4 mM manganese did not inhibit cell density after prolonged culturing (Supplementary Fig. 9). A switch to predominant metalation was observed in three independent E. coli cultures supplemented with 4 mM manganese (Fig. 5a, inset Fig. 5b, Supplementary Table 5a, b, Supplementary Fig. 10).
Calibrated transcript abundance in 4 mM manganese reads out intracellular availability of (Supplementary Fig. 4) . This value is shown in Fig. 5b plus availabilities for other metals in un-supplemented media. The largest favourable free energy gradient from exchangeable available metals to MncA becomes (Fig. 5b). Metalation was predicted by substituting this elevated availability into Supplementary Data 4 (inset Fig. 5b). A switch to predominant metalation with cognate plus partial mis-metalation with is thus predicted, as well as observed by MncA -trapping, in . coli supplemented with 4 mM manganese.
MncA-trapped metals confirm negligible or metalation in supplemented cells
We further wondered if MncA could be predictably metalated with other elements in metal-supplemented viable E. coli. Availabilities of and at the upper (0.99) sensor boundaries (Supplementary
Fig. 4 | Mis-metalation with predicted and observed in heterologous E. coli. a Purification of soluble MncA folded in vivo, minus secretion signal, by anion exchange chromatography ( 5 mL Q-Sepharose, and 1 mL Q-Sepharose) both eluted using high-salt buffer, with intervening SEC (Superdex 200 or 75). MncA was recovered from a soluble protein fraction of . coli after low-level expression overnight at using pBAD30- , induced with arabinose. Representative of biologically independent purifications. Full gel image in Supplementary Fig. 8a. b Representative ( independent biological replicates) chromatogram showing in-cell acquired metals in MncA-containing fractions determined by ICP-MS revealing mis-metalation with and traces of . MncAcontaining fractions were confirmed by SDS-PAGE (Supplementary Fig. 8d) and quantified by (using ). Metal contents of MncA-containing fractions (Supplementary Table 4a, b, Supplementary Fig. 8) were used to calculate fractional occupancies (%). Supplementary Table 4 shows similar outcomes via a modified protocol as used in subsequent experiments. c Metal availabilities ( , squares and text inset) in the cytosol of . coli grown aerobically in LB, estimated as in Foster et al. by calibrated qPCR with genes regulated by cognate metal sensors (bars are sensor ranges from to ). MncA metal-preferences ( , pale blue circles), (triangle). Inset shows occupancies of MncA predicted from the free energy differences using Supplementary Data 4. The inset shows the resulting predicted MncA mis-metalation with (dark blue columns) in the heterologous host based on the largest favourable gradient ( in main figure). Inset also shows mean ( SD) in-cell metalation from the biological replicates (pale blue columns, squares, circles, triangles, Supplementary Table 4b), closely matching predictions. Source data are provided as a Source Data file.
Fig. 5 | Substantial cognate metalation predicted and confirmed in 4 mM manganese. a Representative ( independent biological replicates) chromatogram showing in-cell acquired metals in MncA-containing fractions determined by ICP-MS, as in Fig. 4a, b but using a single anion exchange step (Q-Sepharose), HPLC SEC (TSK SW3000) and showing increased cognate metalation with . MncA was identified by SDS-PAGE in Supplementary Fig. 10c. Metal contents of MncAcontaining fractions (Supplementary Table 5) from independent biological replicates (Supplementary Fig. 10) were used to calculate fractional occupancies (%). b Metal availabilities ( , squares, and text inset) in E. coli cytosol grown aerobically as in Fig. 4c, except replaced with estimates from calibrated qPCR of MntR target in cells cultured in 4 mM manganese (Supplementary Fig. 4). MncA metal-preferences as (pale blue circles, , triangle). Bars are sensor ranges from to . Inset shows occupancies of MncA predicted from free energy gradients using Supplementary Data 4 and substituting availability with a value of (as in inset text). The inset shows predicted MncA metalation (dark blue columns) with , based on the largest favourable gradient ( in main figure), plus partial metalation with . Inset also shows mean ( ± SD) in-cell metalation from the independent biological replicates (square, circle, triangle, pale blue columns, as in Supplementary Table 5), largely matching predictions. Source data are provided as a Source Data file.
Fig. 4) , annotated in Fig. 6a, were entered into the metalationcalculator in Supplementary Data 4 as before. Negligible or no occupancy with either metal, but mis-metalation, was predicted (Fig. 6a).
Predicted mis-metalation with (negligible ) was observed in cells grown in zinc (Fig. 6a). This treatment was selected to approximate maximal abundance of transcripts regulated by , hence maximum availability with slight inhibition of growth (Supplementary Fig. 9). Chromatographic profiles for (unlike and ) imperfectly align with absorbance at 280 nm or the distribution of MncA on SDS-PAGE (Supplementary Fig. 11). We did not
identify the contaminating -protein by principal component analysis and MncA-metalation was determined from a single fraction showing least evidence of other proteins (Supplementary Fig. 11, Supplementary Table 6).
Predicted mis-metalation with (not ) was also observed in cells grown in copper (Fig. 6a, Supplementary Fig. 9). The profiles for copper (unlike and ) again imperfectly align with MncA but correlated with a protein of matching glyceraldehyde-3phosphate dehydrogenase (GAPDH) (Supplementary Fig. 12). GAPDH is known to bind copper in copper-exposed cells and is removable using Blue Sepharose . Including this step retained and MncA but eliminated co-migrating copper (Supplementary Fig. 13). Metalation was quantified from three independent cultures after using Blue Sepharose (Supplementary Table 7a, b). In summary, intracellular availabilities of or are insufficient to metalate MncA in supplemented cells, with metalation matching predictions.
MncA-trapped metals in high and refine availabilities
MncA trapped or in the respective metal supplemented media (Fig. 6a, Supplementary Fig. 9, 14, 15, Supplementary Table 8,9). Metalation was predicted as for high and but using 0.01 boundary values for RcnR-DNA occupancy (Supplementary Fig. 4). In high cobalt, MncA bound more , less and more , than predicted (Fig. 6a). The most parsimonious explanation is that availability declines in high . An MncA-residuals calculator in Supplementary Data 5, formulated in Supplementary Note 2, used the determined preferences of MncA to estimate the decrease in availability relative to (Fig. 6b). Figure 6c compares observed occupancies with predictions using the refined availability. Residuals for , not used in refinement, reduced as anticipated for the parsimonious solution.
In high , MncA bound more , less and , than predicted (Fig. 6a). Here, the most parsimonious explanation is that is more available than approximated. The MncA-residuals calculator estimated the further increase in intracellular availability relative to Fe (Fig. 6b). Figure. 6c compares -refined occupancies with those observed. Residuals for again reduced. In summary, MncA metaloccupancies plus the MncA-residuals calculator can be used to probe and refine estimates of intracellular metal availabilities. These tools can assist predictions of in-cell metalation of proteins in other cell types and growth conditions. MncA-derived refinements also suggest availability declines in . coli in high cobalt and this is subsequently explored.
Shallow pool is depleted in un-supplemented media
Although is a most available metal (least negative , Fig. 4c), the pool in un-supplemented media is shallow, containing only a few thousand atoms per cell . qPCR with primers to MntR-regulated revealed increased expression and hence reduced available ( -MntR co-repression being alleviated) during prolonged MncA expression in un-supplemented medium (Fig. 7a). This is consistent with observing less than predicted (Fig. 4c inset). Furregulated fepD transcripts indicate a smaller decline in the available pool in un-supplemented medium. These data also highlight a challenge with qPCR for estimating intracellular metal availabilities at upper transcript abundance, corresponding to lower availabilities for metal-dependent co-repressors MntR and Fur. The logarithmic nature of PCR as represented by the scales in Fig. 7, means the upper 50% of the expression range corresponds to a single PCR cycle. The abundance of transcripts reaches (and exceeds) the upper boundary, lowest available , reported previously whereas fepD remains 3 to 4 cycles below (Fig. 7a, b) . These data thus report depletion of the intracellular pool overnight by but a relatively modest change in . The use of MncA to refine estimated availabilities is thus especially valuable for qPCR-based estimates in the
Fig. 6 | MncA read-outs of in-cell metalation in high and refine metal availabilities. a In-cell metalation with and , not or , switches from in metal-supplemented respectively as shown) media, qualitatively matching predictions but quantitatively greater and metalation than predicted. Metalation (dark blue bars) predicted using Supplementary Data 4 from availabilities in un-supplemented medium but with separately altered high availabilities of or (as inset text). High availabilities correspond to the respective upper metal boundaries ( or or 0.01 ) shown in Supplementary Fig. 4. Mean ( ± SD) measurements of in-cell metalation ( independent biological replicates, square, circle, triangle) in high metal (pale blue bars) calculated using data in Supplementary Tables based on Supplementary Figs. 11-15. b Selected intracellular metal availabilities ( ) refined (triangles) using Supplementary Data 5 for (left) from observed in-cell – and -MncA occupancies in high (relative to in un-supplemented medium), and for (right) from and occupancies in (relative to elevated as show in inset text). Metal preferences of MncA shown as (circles). Bars are sensor ranges from to . c Calculated metalation (dark blue bars) using Supplementary Data 4 but with refined availabilities as in panel (b), reduced residuals for (which was not included in the refinement process) relative to observed in-cell metalation (pale blue bars included for comparison), showing mean ( ± SD) ( independent biological replicates, square, circle, triangle). MncA metalation can be used to refine relative intracellular metal availabilities and Supplementary Data 5 is provided to enable such calculations. Source data are provided as a Source Data file.
upper of expression ranges. This applies to some metalsupplemented cells but not un-supplemented cells as cultured previously . Viewed together, Fig. 4c (inset) and Fig. 7a reveal that disparities between predicted and observed MncA-metalation can report on non-steady-state intracellular metal availabilities ( ).
Steady-state availabilities sustained for high and , not and
At the outset of MncA expression, cells exposed to elevated metals all reach or exceed the estimated transcript abundances defined as the qPCR boundaries for high intracellular metal availabilities (Fig. 7). For -MntR co-repressed mntS transcripts, this reflects high occupancy of the mntS promoter in cells exposed to 4 mM manganese and low transcript abundance. In contrast this equates to high transcript abundance for and responsive transcripts regulated by activators ZntR, CueR and metal-dependent de-repressor RcnR. At the end of MncA production ( 18 h ) in metal-supplemented cells, and availabilities remain high, but they decline for and
(Fig. 7a, c-f). This is also true for cultures that are not expressing MncA excluding the formal possibility that turnover of metalated MncA sustained elevated steady-state and availabilities but not and . Lack of steady-state elevated and indicates a difference in metallostasis for these metals compared to and . It may be more challenging to metalate proteins in viable cells with or simply by metal supplementation.
Fur causes atoms cell to decline in
To investigate the MncA-predicted decline in intracellular availability in high (Fig. 6b), total atoms of iron cell were measured by ICP-MS (Fig. 8a). Iron content declined in consistent with the prediction (noting that available metal can sometimes trend differently to total metal). Cells lacking Fur contain less iron in standard media as explained previously . Importantly, the effect of on iron is absent in fur (Fig. 8a). Fur binds and -Fur binds DNA with both affinities already known . Fig. 8b calculates the Fur response to intracellular simulated as for . DNA occupancy at the availability inside cells grown here in elevated cobalt reveals an extreme cross-response of Fur. Non-cognate sensors are known to cross-respond to availabilities at the top of the cognate sensor ranges . Crucially, cross-metalation of Fur with explains the decline in total iron in Fig. 8a and in available intracellular read out by MncA (Fig. 6b).
The remaining residual differences between observed and predicted occupancies in Fig. 6c have been used via the MncA-residuals calculator (Supplementary Data 5), to further refine intracellular metal availabilities (Supplementary Fig. 16). availability is inferred to decrease in high and increase in high . Both predicted trends were reflected in the total atoms cell , and the effects of on were not Fur-dependent (Fig. 8c). The effects of high on are not dependent on -sensing MntR (Supplementary Fig. 17a). The MntRindependent decline in is less severe in wild-type where the sensor controls metallostasis dampening changes. The residuals in cells exposed to 4 mM manganese suggest a decline in available and this matched a Fur-independent decline in total iron atoms cell (Fig. 5b, Supplementary Fig. 17b). The MncA-residuals calculator was used iteratively to further refine high and blueprints (Supplementary Fig. 16). The refined availabilities and related high-metal
Fig. 7 | Shallow pool depleted in un-supplemented medium but steadystate availabilities sustained in high and . qPCR of a mntS (regulated by -MntR), fepD (Fe -Fur), zntA ( -ZntR), d ( -CueR), e RcnR ), , as change relative to respective lowest measured transcript abundance observed in Foster et al. using chelator, metal and/or supplemented cells values in Source Data TXT for Fig. 7, data are mean SD of independent biological replicates (square, circle, triangle) except where dagger . MntR and Fur target gene transcripts increase in low and (MntR and Fur are metal-dependent co-repressors), but other transcripts increase in high availabilities of cognate metals (regulated by activators or metal-dependent de-repressors). Previous (fold change) qPCR at high metalboundaries (red dashed lines, 0 for Fur) and at low metal boundaries (black dashed lines for MntR and Fur, 0 for others) are shown. Arrows show previous (fold change) qPCR for cognate transcripts isolated from cells grown in LB . Panels (a) and (b) show expression at after addition of arabinose to unsupplemented medium to induce low expression of MncA at . After 18 h MntR regulated transcripts pass the low boundary (DNA occupancy ) (panel a) further quantified in Supplementary Fig. 18. The pool is shallow and depleted in LB. For Fur-regulated fepD the scale reports modest depletion of intracellular available at 18 h in un-supplemented medium (Supplementary Fig. 18). The first two columns (panels ) show negligible change in availability of or after 18 h MncA expression in non-supplemented medium. Final four columns (panels ) report (fold change) in metal-supplemented media either with or without expression of MncA as indicated. Steady-state Mn availability approximating upper boundary sustained to 18 h in 4 mM manganese (panel a). Steady-state approximating the upper metal boundary is also sustained in high and , but not and (panels ). Source data are provided as a Source Data file.
calculators are provided (Supplementary Data 6-8). MncA and the MncA-residuals calculator in Supplementary Data 5, could be used to probe relative metal availabilities in other cell types including (rating) cells with metallostasis engineered to optimise the speciation of metalation for selected elements.
Discussion
Here we discover that the metal-binding preferences of a protein which kinetically traps metals at folding, MncA, all follow the Irving-Williams series (Fig. 1). This may reflect the preferences of flexible sites in the folding pathway prior to kinetic trapping. It is also possible that preferred metals in the series increasingly encourage progression of MncA folding intermediates. Although copper is most preferred, the cognate metal is correctly decoded to be by relating these preferences to idealised estimates of intracellular metal availabilities, standardised to the mid-points of the ranges of metal sensors (Fig. 2). MncA kinetically traps metals, such that those which co-purify are likely to reflect in vivo metalation states (Fig. 3, Supplementary Fig. 7). Estimates of metal availabilities inside . coli grown aerobically predict that (cyanobacterial) MncA will be mis-metalated with in . coli, and this is established by ICP-MS analysis of purified soluble MncA (Fig. 4). Thus, the speciation of metalation is directly shown to be determined by relative binding preferences for metals competing at intracellular availabilities (Supplementary Fig. 5a), and mis-metalation can be predicted in heterologous hosts. Subtle changes in a metal availability (or preference) can switch metalation because intracellular availabilities follow the inverse of the Irving-Williams series and hence trend with preferences (Figs. 2c, 4c, 5b, 6a). Estimates of intracellular metal-availabilities can thus be used to predict the speciation of in vivo metalation to inform the engineering of natural and synthetic metalloproteins to optimise metalation (using Supplementary Data 3, 4, 6-8).
Unrefined metal availabilities derived from qPCR-based responses of metal-sensing transcriptional regulators correctly predicted mismetalation of MncA in E. coli in un-supplemented media (Fig. 4c). In metal-supplemented media, MncA occupancies with , and exceeded (at least to some extent) predictions (Figs. 5, 6a). These occupancies were used with Supplementary Data 5 to refine estimates of intracellular availabilities predicting decreased available in and , decreased available in , but increased available in Co (Fig. 6b, Supplementary Fig. 16). Encouragingly, in every case this coincided with analogous changes in total or atoms cell (Fig. 8a, c, Supplementary Fig. 17a, b). The decline in cellular in high , but not in high , was Fur-dependent and the former explained by responses to of Fur (Fig. 8a, b, Supplementary Fig. 17b). The extent to which cross-metalation of Fur with is disadvantageous mis-metalation, remains to be established. Together these data show the value of MncA as a probe of intracellular metal-availabilities, orthogonal to synthetic metal sensors , and complementing calibrated endogenous metal sensors especially where qPCR is less reliable and/or crosstalk likely .
MncA occupancies report lowered availability inside cells after growth in un-supplemented medium (Fig. 4). Here, the shallow pool is depleted during MncA expression such that steady-state availability is not sustained (Fig. 7a, Supplementary Fig. 18). This is significant when recombinant metalloproteins are overexpressed in E. coli potentially depleting metals: metalation can then reflect depths of available metal pools rather than strengths of competition with other ligands. In manganese-, nickel- and cobaltsupplemented cells elevated intracellular availabilities were sustained for but this was not the case in elevated zinc and copper (Fig. 7). Mechanisms of metallostasis for and must adjust rates of import, export and/or consumption to restore pre-exposure steadystate intracellular availabilities, whereas elevated steady-states are
Fig. 8 | Fur causes iron atoms cell to decline in matching MncA-predicted availabilities. a Total iron atoms cell determined by ICP-MS of cell digests, declines in cells grown in medium supplemented with high (left panel). This is consistent with the decline in intracellular availability estimated from the residuals in Fig. 6a, calculated using Supplementary Data 5 and shown in Fig. 6b. Elevated does not affect total iron atoms cell (right panel). Total iron cell is less in fur but does not further decline in elevated . The decline in iron cell in response to elevated is Fur-dependent. b Fur binds to promote DNAbinding: simulated using known affinity of Fur, DNA affinity of -Fur, plus apo-Fur DNA affinity and Fur molecules cell , as simulated for . Fur promoters will be aberrantly repressed in cobalt (square and blue arrow) causing the predicted decline in intracellular available shown in Fig. 6b and observed decline in total iron atoms cell in panel (a). c Total manganese atoms cell increase in high consistent with slightly increased intracellular available , modellable from the remaining residuals in high in Fig. 6c, calculated using Supplementary Data 5, shown in Supplementary Fig. 16b. These effects of cobalt on are independent of Fur. Total manganese atoms cell decrease in high , this trend is more pronounced in (Supplementary Fig. 17), the trend is not evident in fur which contains low manganese (right). An analogous decrease in available intracellular calculable (Supplementary Data 5) from the remaining residuals in high , Fig. 6b, is shown in Supplementary Fig. 16a. Use of MncA as a probe of relative intracellular metal availabilities iteratively refines values shown in Supplementary Fig. 16, included in Supplementary Data 6-8, and provided as blueprints to assist manipulation of the speciation of in vivo protein metalation. Mean ± SD of independent biological replicates (squares, circles, triangles) in (a) and (c). Source data are provided as a Source Data file.
maintained for the other metals (Fig. 7). Nonetheless, it is predicted that MncA would bind negligible or even if elevated steadystate availabilities were sustained. Notably, more favourable gradients ( ) for and largely exclude (Supplementary Fig. 19). Less and in the media, engineering cells to have reduced availabilities of and , or weakening MncA binding to and , could assist accumulation of .
Metalation of MncA inside . coli switched between , and as predicted (Figs. 4-6). This becomes achievable because intracellular metal availabilities track with metal-binding preferences of proteins. This could indicate that protein mis-metalation is likely
in vivo. Alternatively, mechanisms of metal homoeostasis could be exceptionally finely tuned to native metallo-proteomes minimising mis-metalation. In either eventuality, this raises the prospect in engineering biology of widespread mis-metalation because metalavailabilities will not necessary be correctly tuned to non-native metalloproteins. Notably, in E. coli, GTP-dependent metallochaperones YeiR and YjiA are predicted to be predominantly metalated with presumed cognate , whereas heterologous CobW from Rhodobacter, CbiK from Salmonella, in common with cyanobacterial MncA become mis-metalated with and respectively (Fig. 4, Supplementary Fig. 20). Quantification of production in engineered . coli provided indirect evidence of the predicted mis-metalation of CobW, and CbiK is known to insert Fe into siroheme in Salmonella missing the siroheme chelatase CysG . Crucially, MncA now provides a direct read-out confirming predicted mis-metalation with (Fig.4).
MncA-refined intracellular metal availabilities along with the calculators provided here can guide optimisation of metalation via use of metal supplements or chelants, alterations to homeostasis by engineering host strains (chassis), or engineering proteins to match availabilities. Approximately half the reactions of life rely on the chemistries of the correct metals bound to metalloproteins . Considerable research is being directed to the generation of various types of artificial metalloenzymes . Inevitably, efforts in metabolic engineering, synthetic biology and directed evolution will often rely on metalloenzymes . The blueprints and calculators can inform metalloenzyme engineered for in vivo bioprocessing applications. The optimisation of in vivo metalation presents opportunities as the engineering of biological systems for (sustainable) bio-manufacturing is prioritised .
Methods
Expression and purification of unfolded apo-MncA
To examine the relative binding preferences of MncA in vitro, protein was expressed and purified . Briefly, MncA minus TAT secretion signal was expressed from pET29a-mncA in E. coli BL21(DE3) pLysS. Isopropyl -D-1-thiogalactopyranoside (IPTG, 1 mL 0.4 M ) was added to a mid-log phase culture ( 1 L in a 2 L flask) at , to induce high-level expression for 3 h before harvesting cells by centrifugation
( ) and freezing ( ). The pellet was resuspended in 30 mL 100 mM Tris pH EDTA, 1 mM phenylmethylsulphonyl fluoride (PMSF) and sonicated ( on ice, 1 min intervals). Lysate was cleared by centrifugation , ), supernatant discarded, and the pellet resuspended in 100 mM Tris Triton X-100 , sonicated ( on ice, 1 min intervals) and centrifugation repeated. The pellet was washed in 30 mL 100 mM Tris to remove the Triton, then sequentially in 30 mL 50 mM HEPES pH 7.5, 1 M urea followed by 15 mL 50 mM HEPES pH 7.5, 2 M urea. During the final wash the lysate was split into 2 mL aliquots followed by centrifugation to recover inclusion bodies, stored at .
Preparation of metal stocks
Metal stocks ( and ) in ultrapure water were sterile filtered and quantified by inductively coupled plasma mass spectrometry (ICP-MS). When required for anaerobic experiments, stocks were prepared in purged ultrapure water. Total Mn and Fe concentrations were then confirmed by ICP-MS. stock was confirmed to be reduced by reaction with excess (10-fold) ferrozine (Fz) using for the complex. Reduced stocks were prepared as described in specific experiments and cuprous state validated with bicinchoninic acid (BCA) and ICP-MS.
Production of buffered competing metals
MncA in vitro refolding buffers (in 50 mM MOPS pH 7.5) contained pairs of metals and ligand in varied amounts to achieve different buffered [available metals]. Buffers were prepared in acid-washed flasks with components added in the order, pH buffer, ultrapure water, ligand (typically NTA, or 1 mM L -histidine for competitions with ) followed by the two metals. Buffers were typically prepared in 100 mL volumes, filtered via filters if any evidence of light scatter. Supplementary Table 1 shows the total amounts of each metal and ligand (NTA, histidine or neither) plus the buffered [available metals] used in each refolding solution. Supplementary Data 1 (derivation in Supplementary Note 1) was used and is provided here to assist in the production of such paired metal buffers. For an effective buffer the total metal concentration should substantially exceed the protein concentration ( -fold generally, 10 -fold for competitions with ). To achieve an effective buffer of the buffering agent should be metalated.
A preliminary experiment was performed with BCA to buffer with no expected buffering of . With and , expected free is , which was competed against . Because < copper co-migrated with MncA , unbuffered was competed against unbuffered . Typical concentrations were each and were verified by ICP-MS on the folding solution. Competitions between and , used freshly prepared solution of and refolding experiments included 1 mM hydroxylamine to maintain copper in reduced form. For buffers forming 2:1 metal-dependent complexes, buffered concentrations of each metal were determined using HySS software for His . values for all equilibria are as follows: proton dissociation from histidine (HisH-9.08, complexation of , His complexation of ( , ), . For BCA complexes, derivation of buffering of [ ] in Supplementary Note 1.
Metal-binding preferences of MncA at folding
Inclusion bodies containing MncA were solubilized in HEPES pH 7.5 with 8 M urea, and the concentration calculated from using an experimentally determined extinction coefficient , typically in the range . Refolding was achieved by
dropwise dilution of urea-solubilised MncA in large volumes ( 100 mL ) of solutions containing pairs of competing buffered metals prepared as described earlier. Solutions were thoroughly mixed before adding unfolded MncA and gently mixed between additions. Dilute refolded MncA was recovered by binding to a 1 mL Q-Sepharose (Cytiva or GE Healthcare) anion exchange column pre-equilibrated with low-salt buffer Tris pH 7.5, 50 mM NaCl. The column was washed with 20 mL low-salt buffer before eluting MncA with high-salt buffer, 50 mM Tris pH . MncA was quantified by then resolved ( 0.5 mL at ) by SEC (PD-10, GE healthcare, previously washed with 0.5 mL 5 mM EDTA followed by ultrapure water and equilibrated with the low-salt buffer). Fractions ( 0.5 mL ) were analysed for protein via and metal by ICP-MS using corresponding matrix-matched calibration curves. To compete versus and , stocks were prepared as described earlier and refolding done in an anaerobic chamber. The concentrations of and metals (ICP-MS) were superimposed to identify metals co-eluting with MncA and estimate the metal:protein stoichiometry. The ratio of trapped metals was used to determine the relative binding preferences of the two metals at folding in buffer of known competing metal availabilities (Table 1, Supplementary Tables 1, 2).
crystal structure
A solution of MncA in 50 mM HEPES pH 7.5 and 8 M urea, prepared from pelleted inclusion bodies, was added dropwise, with stirring to 100 mL of 50 mM MOPS, pH 7.5 , and , passed through a filter and loaded on a 5 mL Q-Sepharose column (Cytiva) equilibrated with 50 mM Tris, pH 7.5, and 50 mM NaCl . The column was washed with the same buffer. Folded, concentrated was eluted with buffer containing 500 mM NaCl . Buffer was exchanged via several cycles of dilution and concentration (Amicon Ultracel, 0.5 mL 10 kDa ) to obtain in 10 mM Tris, pH 7.5, 50 mM NaCl . Crystals were grown in 100 mM sodium acetate, pH 4.0 and PEG . An aliquot of crystals redissolved in 50 mM Tris, were analysed by ICP-MS and measured confirming stoichiometric metalation with . Following cryo-protection by the stepwise addition of glycerol to , crystals were flash-cooled and stored in liquid nitrogen prior to data collection.
Data were collected at beamline I04, Diamond Light Source (Supplementary Table 3). A highly redundant data set was collected by obtaining of data in four separate scans along the axis of a long, hexagonal rod-shaped crystal. The data were processed using the xia2 package at Diamond , which employed XDS to integrate , and AIMLESS to scale and merge the data . The structure solution was obtained by molecular replacement with the trimeric structure (PDB ID 2VQA) as search model using Phaser implemented in Phenix . Refinement continued in Phenix alternating with modelling in Coot . Figures were prepared with ChimeraX , and PyMOL (Schrödinger) software. MOLE 2.5 was used to identify channels present in the structure .
Expression and purification of RncR
Purification of RcnR overexpressed in E. coli BL21(DE3) from coding sequences cloned in pET29a has been described . In common with the other regulators (MntR, Fur, NikR, ZntR, Zur, CueR), the sequence was from Salmonella enterica serovar Typhimurium strain SL1344 (referred to as Salmonella), and RcnR shares sequence identity to E. coli RcnR. Anaerobic, reduced and apo-RcnR was prepared by applying purified, EDTA treated, apo-protein to a HiTrap heparin column, transferred to an anaerobic chamber, washed with column volumes of Chelex-treated, -purged , 10 mM HEPES, pH 7.0, then eluted with , 10 mM HEPES, pH 7.0. RcnR was quantified by using experimentally determined extinction coefficient of obtained
via quantitative amino acid analysis. Reduced thiol and metal content were assayed , and all anaerobic protein samples (maintained in an anaerobic chamber) were reduced and metal-free. All in vitro experiments were carried out under anaerobic conditions using Chelex-treated and -purged buffers .
stoichiometry and affinity of RcnR
All experiments were conducted in HEPES pH 7.5. To determine stoichiometry, (as ) was titrated into purified protein ( ) and absorption spectra recorded at equilibrium using a UV-visible spectrophotometer (Perkin Elmer Life Sciences). Additionally, an aliquot of monomer) was incubated with and bound metal resolved by SEC eluted with HEPES pH 7.5 (PD-10, collecting 0.5 mL fractions) and analysed for protein by Bradford assay standardised with known concentrations of RcnR and metal by ICP-MS. To determine affinity, titrations were performed in the presence of EGTA using four RcnR monomer concentrations; RcnR and EGTA, RcnR and EGTA, RcnR and EGTA, RcnR and EGTA, monitoring a dependent feature of -RcnR at 326 nm . A simultaneous fit was made to all data sets using Dynafit (fitting models in Supplementary Software) .
-RcnR DNA-affinity by fluorescence anisotropy
Fluorescently-labelled (hexachlorofluorescein) double-stranded DNA probes containing the identified RcnR-binding site upstream of the promoter were synthesised and annealed as described . RcnR ( RcnR tetramer) or apo-RcnR were titrated into 10 nM DNA in HEPES pH 7.5. Changes in anisotropy ( ) were measured using a modified Cary Eclipse fluorescence spectrophotometer (Agilent Technologies) fitted with polarising filters ( , averaging time , replicates , and ), allowing the cuvette to equilibrate ( 3 min ) before recording. A simultaneous fit was made to all data sets (fitting models in Supplementary Software using maximum ).
Expression and purification of soluble MncA to determine in vivo metalation
The coding region of as in pET29a- was sub-cloned to create pBAD30-mncA to enable tuned expression dependent on [arabinose]. E. coli BW25113 (hereafter E. coli) containing pBAD30-mncA was inoculated into overnight cultures ( glucose + carbenicillin at ) used the following day to inoculate 2 L flask containing 1L LB medium and carbenicillin (no glucose) and incubated at , until OD reached mid-log phase . Cultures were transferred to and a low concentration ( ) L-arabinose added to induce low-level gene expression followed by culturing overnight total) before harvesting cells by centrifugation ( ). Purification of soluble in vivo metalated MncA involved protocols analogous to procedures used to recover native MncA from Synechocystis . The entire cell pellet (from 1L culture) was resuspended in 30 mL lysis buffer ( 20 mM Tris pH 7.5, 1 mM EDTA, 1 mM PMSF) sonicated ( 4 min pulsing) and centrifuged ( ) 45 min to remove cell debris. Supernatant was loaded onto a Q-Sepharose anion exchange column ( 5 mL , pre-equilibrated with 20 mM Tris pH 7.5) then washed with the same buffer. MncA was eluted using a gradient in 30 mL , collecting 1 mL fractions. SDS-PAGE identified MncAcontaining fractions which were pooled . Further purification and analysis used a more rapid analytical protocol in later experiments while in earlier experiments MncA was loaded onto Superdex 75 or Superdex 200 SEC columns (as specified) with fractions analysed by SDS-PAGE. MncA containing fractions were diluted to , reapplied to Q-Sepharose washed then
eluted with 300 mM NaCl and fractions analysed by SDS-PAGE, , and [metal] by ICP-MS. Proportional (%) occupancies of MncA with each metal was first calculated from the ratio of [metal]/[MncA] assuming 2 metal sites per MncA molecule, confirmed by mean experimental occupancy of 99% (replicates 1-3 in Supplementary Table 4a, b). The protocol was simplified to two steps with MncA first recovered via anion exchange ( 5 mL Q-Sepharose column, 0-300 mM NaCl gradient) followed by rapid analytical scale SEC using a SW3000 (TSK) column . Fractions were again analysed by SDS-PAGE, and ICP-MS. A comparative fourth biological replicate of MncA extracted from cells grown without metal supplementation was purified via the simplified approach obtaining similar metal occupancies (Supplementary Table 4). Additionally similar occupancies were calculated as a proportion of the total metal content of MncA avoiding variation in MncA with different metals (Supplementary Table 4b). In subsequent metal-supplemented cultures, respective metals were added at inoculation 1-3 h prior to L-arabinose addition. MncA isolated from cells supplemented with copper co-purified with a copper-protein tentatively identified as GAPDH. Fractions containing MncA were passed over a 5 mL Cibacron blue Sepharose (Blue Sepharose) column equilibrated with 50 mM Tris pH 7.5. MncA eluted rapidly while the contaminating protein was retained then eluted with 1 M NaCl in 50 mM Tris pH 7.5. Blue Sepharose-treated MncA-containing solution was subjected to analytical SEC as above.
Estimation of transcript abundance in E. coli
Two extracts were collected from E. coli, including from cells containing pBAD30-mncA expressing MncA immediately before arabinose addition, and secondly after overnight growth, and RNA stabilised using RNAProtect Bacteria Reagent, 2 mL (Qiagen). Samples were processed as described . Briefly, RNA was extracted using RNeasy Mini Kit (Qiagen), [RNA] estimated from then treated with DNase I (Fermentas). ImProm-II Reverse Transcriptase System (Promega) generated cDNA, with parallel control reactions excluding reverse transcriptase. Transcript abundance was determined using primers for mntS, fepD, rcnA, nikA, znuA, zntA, copA and rpoD that amplify of DNA with sequences listed in Supplementary Data 9. Quantitative polymerase chain reaction (qPCR) analysis was executed in reactions containing 5 ng of cDNA, 400 nM of each complementary primer and PowerUP SYBR Green Master Mix (Thermo Fisher Scientific). Three technical replicates of each biological replicate were analysed using a Rotor-Gene Q 2plex (Qiagen; Rotor-Gene-Q Pure Detection Software) with additional control reactions without cDNA templates (qPCR grade water used instead, supplied by Thermo Fisher Scientific) run for each primer pair, in addition to control reactions without reverse transcriptase for the reference gene primer pair (rpoD). values were calculated with LinRegPCR (version 2021.1) after correcting for amplicon efficiency (Supplementary Source Data TXT, shown in Fig. 7). Change in gene abundance, relative to the control condition (defined as the condition where the minimum transcript abundance was observed for each target gene), was calculated using the method using rpoD as the reference gene and presented as (fold change).
Intracellular metal availabilities and predictions of in vivo metalation
Responses of metal sensors ( for DNA occupancies of metaldependent de-repressors and co-repressors, for metalated activators) as a function of intracellular available buffered metal concentrations were calculated using sensor metal affinities, DNA affinities, protein abundances and numbers of DNA-binding sites for . coli sensors as described (Supplementary Data 2 for ). Transcript abundance was correlated with the response curves to enable estimations of intracellular metal availability expressed as a free energy for complex formation ( ) in . coli grown aerobically in LB
as described . Using these availabilities, metalation of proteins was predicted in vivo, accounting for multiple inter-metal competitions including competition from the intracellular buffer as described by Young and coworkers . Supplementary Data 3 and 4 perform these calculations for ideal cells (sensors at mid-range, including ) and at intracellular metal availabilities in . coli grown aerobically in LB respectively.
In metal-supplemented cultures, (fold change) relative transcript abundance in this work approximated values reported previously with some boundaries exceeded (Fig. 7). The reported values for high intracellular availabilities of supplemented metals were thus substituted into Supplementary Data 4, while retaining original values for all other metals, to make first predictions of MncA metalation at high intracellular or . For and , residual differences between predicted and observed MncA metalation was used to iteratively refine intracellular metal availabilities according to Supplementary Note 2 and using Supplementary Data 5 . Supplementary Data 6-8 contain the refinements and can be used to predict metalation of other proteins in E. coli in elevated , or .
Elemental analyses by ICP-MS and metal atoms cell
ICP-MS was performed at the Durham University Bio-ICP-MS Facility (ThermoFisher iCAP RQ model) with matrix matched standard curves and internal silver standards. E. coli fur and strains were obtained from the Keio collection . Individual colonies of E. coli or mutants were inoculated in shaking at for , diluted into fresh medium in a 50 mL conical centrifuge tube supplemented with metals ( ) where specified, to . Cells were incubated with shaking at , overnight, diluted 1:10 to measure , and cells recovered (from remaining 9.9 mL ) by centrifugation. Pellets were washed four times by resuspension in 1 mL wash buffer Tris sorbitol, 0.2 mM EDTA) followed by centrifugation. Ultrapure (Merck) was added to each pellet and allowed to incubate for a minimum of 16 h until fully digested. The samples were then prepared for ICP-MS with matrix-matched calibration curves. An of 1 equated to a cell count (CASY cell counter) of cells using this strain, enabling atoms cell to be calculated from calibrated ICP-MS data.
Statistics and reproducibility
Sample sizes were chosen based on prior experimental experience, and to give consistent results, following convention in the literature for equivalent analyses. Experiments designed to derive quantitative values used to predict or test and measure metalation, or to refine and evaluate estimates of intracellular metal availabilities, were performed in triplicate or more to enable calculation of SD (listed in tables or text or shown as error bars on figures) or SE for -RcnR affinities. Predictions of metalation do not propagate SD from contributing values. Analogous chromatograms to the representative data in Figs. 4b and 5a were obtained on two further occasions ( ), data in Figs. 4a and 1 b are representative of 18 and analogous purifications respectively. The number of independent experiments or biologically independent samples is otherwise shown in figure legends or footnotes of Tables.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data are available within the article, its Supplementary Information files, plus PDB entry 9GOF and from corresponding authors on request. Source data are provided with this paper as Source Data files.
Excel spreadsheets (with instructions) providing a calculator to formulate competing metal buffers, to calculate DNA occupancy as a function of availability for metal-dependent de-repressor RcnR and providing a calculator to use in vivo recovered metal occupancies of MncA as a probe to refine estimates of intracellular metal availabilities, are provided as Supplementary Data 1, 2 and 5 respectively. Excel spreadsheets constituting calculators of metalation in -RcnRrefined idealised cells, E. coli grown aerobically in LB, E. coli grown aerobically in LB supplemented with manganese, nickel, and cobalt, are provided as Supplementary Data 3, 4, 6-8 respectively. Supplementary Data 9 contains oligonucleotide sequences. Published structures used here for MncA and MntR are PDB entries 2VQA and 9C4D respectively. Source data are provided with this paper.
Code availability
Equation derivations are in Supplementary Notes 1 and 2 of the Supplementary Information. Dynafit scripts are provided in Supplementary Software.
References
Waldron, K. J., Rutherford, J. C., Ford, D. & Robinson, N. J. Metalloproteins and metal sensing. Nature 460, 823-830 (2009).
Irving, H. & Williams, R. J. P. Order of stability of metal complexes. Nature 162, 746-747 (1948).
Da Silva, J. F. & Williams, R. J. P. The biological chemistry of the elements: the inorganic chemistry of life. (Oxford University Press, 2001).
Robinson, N. J. & Glasfeld, A. Metalation: nature’s challenge in bioinorganic chemistry. J. Biol. Inorg. Chem. 25, 543-545 (2020).
Foster, A. W., Osman, D. & Robinson, N. J. Metal preferences and metallation. J. Biol. Chem. 289, 28095-28103 (2014).
Grāve, K., Griese, J. J., Berggren, G., Bennett, M. D. & Högbom, M. The Bacillus anthracis class Ib ribonucleotide reductase subunit NrdF intrinsically selects manganese over iron. J. Biol. Inorg. Chem. 25, 571-582 (2020).
Cotruvo, J. A. Jr & Stubbe, J. Metallation and mismetallation of iron and manganese proteins in vitro and in vivo: the class I ribonucleotide reductases as a case study. Metallomics 4, 1020-1036 (2012).
Tottey, S. et al. Protein-folding location can regulate manganesebinding versus copper- or zinc-binding. Nature 455, 1138-1142 (2008).
Raux, E., Thermes, C., Heathcote, P., Rambach, A. & Warren, M. J. A role for Salmonella typhimurium cbiK in cobalamin (vitamin B12) and siroheme biosynthesis. J. Bacteriol. 179, 3202-3212 (1997).
Schubert, H. L., Raux, E., Wilson, K. S. & Warren, M. J. Common chelatase design in the branched tetrapyrrole pathways of heme and anaerobic cobalamin synthesis. Biochemistry 38, 10660-10669 (1999).
Lacasse, M. J., Douglas, C. D. & Zamble, D. B. Mechanism of selective nickel transfer from HypB to HypA, Escherichia coli [NiFe]hydrogenase accessory proteins. Biochemistry 55, 6821-6831 (2016).
Watanabe, S. et al. Structural basis of a Ni acquisition cycle for [NiFe] hydrogenase by Ni-metallochaperone HypA and its enhancer. Proc. Natl. Acad. Sci. 112, 7701-7706 (2015).
Banci, L. et al. The Atx1-Ccc2 complex is a metal-mediated proteinprotein interaction. Nat. Chem. Biol. 2, 367-368 (2006).
Yang, X., Li, H., Lai, T.-P. & Sun, H. UreE-UreG complex facilitates nickel transfer and preactivates GTPase of UreG in Helicobacter pylori. J. Biol. Chem. 290, 12474-12485 (2015).
Crouzet, J. et al. Nucleotide sequence and genetic analysis of a 13.1-kilobase-pair Pseudomonas denitrificans DNA fragment containing five cob genes and identification of structural genes encoding Cob(I)alamin adenosyltransferase, cobyric acid synthase, and
bifunctional cobinamide kinase-cobinamide phosphate guanylyltransferase. J. Bacteriol. 173, 6074-6087 (1991).
Blaby-Haas, C. E., Flood, J. A., de Crécy-Lagard, V. & Zamble, D. B. YeiR: a metal-binding GTPase from Escherichia coli involved in metal homeostasis. Metallomics 4, 488-497 (2012).
Weiss, A. et al. Zn-regulated GTPase metalloprotein activator 1 modulates vertebrate zinc homeostasis. Cell 185, 2148-2163. e2127 (2022).
O’Halloran, T. V. & Culotta, V. C. Metallochaperones, an intracellular shuttle service for metal ions. J. Biol. Chem. 275, 25057-25060 (2000).
Cavet, J. S. et al. A nickel-cobalt-sensing ArsR-SmtB family repressor: contributions of cytosol and effector binding sites to metal selectivity. J. Biol. Chem. 277, 38441-38448 (2002).
Guedon, E. & Helmann, J. D. Origins of metal ion selectivity in the DtxR/MntR family of metalloregulators. Mol. Microbiol. 48, 495-506 (2003).
Moomaw, E. W. et al. Metal dependence of oxalate decarboxylase activity. Biochemistry 48, 6116-6125 (2009).
Young, T. R. et al. Calculating metalation in cells reveals CobW acquires Coll for vitamin B12 biosynthesis while related proteins prefer ZnII. Nat. Commun. 12, 1195 (2021).
Osman, D. et al. Bacterial sensors define intracellular free energies for correct enzyme metalation. Nat. Chem. Biol. 15, 241-249 (2019).
Grossoehme, N. E. & Giedroc, D. P. Energetics of allosteric negative coupling in the zinc sensor S. aureus CzrA. J. Am. Chem. Soc. 131, 17860-17870 (2009).
Foster, A. W. et al. A tight tunable range for sensing and buffering in cells. Nat. Chem. Biol. 13, 409-414 (2017).
Outten, C. E. & O’Halloran, T. V. Femtomolar sensitivity of metalloregulatory proteins controlling zinc homeostasis. Science 292, 2488-2492 (2001).
Foster, A. W. et al. Metalation calculators for E. coli strain JM109 (DE3): aerobic, anaerobic, and hydrogen peroxide exposed cells cultured in LB media. Metallomics 14, mfac058 (2022).
Young, T. R. et al. Two distinct thermodynamic gradients for cellular metalation of vitamin B12. JACS Au 3, 1472-1483 (2023).
Choi, T. S. & Tezcan, F. A. Overcoming universal restrictions on metal selectivity by protein design. Nature 603, 522-527 (2022).
Chivers, P. T., Basak, P. & Maroney, M. J. One His, two His… the emerging roles of histidine in cellular nickel trafficking. J. Inorg. Biochem. 259, 112668 (2024).
Krämer, U., Cotter-Howells, J. D., Charnock, J. M., Baker, A. J. M. & Smith, J. A. C. Free histidine as a metal chelator in plants that accumulate nickel. Nature 379, 635-638 (1996).
Ma, Z. et al. Bacillithiol is a major buffer of the labile zinc pool in Bacillus subtilis. Mol. Microbiol. 94, 756-770 (2014).
Murphy, J. T. et al. Histidine protects against zinc and nickel toxicity in Caenorhabditis elegans. PLoS Genet. 7, e1002013 (2011).
Nairn, B. L. et al. The response of Acinetobacter baumannii to zinc starvation. Cell Host Microbe 19, 826-836 (2016).
Morgan, M. T., Nguyen, L. A. H., Hancock, H. L. & Fahrni, C. J. Glutathione limits aquacopper(I) to sub-femtomolar concentrations through cooperative assembly of a tetranuclear cluster. J. Biol. Chem. 292, 21558-21567 (2017).
Brawley, H. N. & Lindahl, P. A. Low-molecular-mass labile metal pools in Escherichia coli: advances using chromatography and mass spectrometry. J. Biol. Inorg. Chem. 26, 479-494 (2021).
Stewart, L. J. et al. Role of glutathione in buffering excess intracellular copper in Streptococcus pyogenes. MBio 11, 02804-02820 (2020).
Hider, R. C. & Kong, X. L. Glutathione: a key component of the cytoplasmic labile iron pool. BioMetals 24, 1179-1187 (2011).
Osman, D. et al. Fine control of metal concentrations is necessary for cells to discern zinc from cobalt. Nat. Commun. 8, 1884 (2017).
Giedroc, D. P. & Arunkumar, A. I. Metal sensor proteins: nature’s metalloregulated allosteric switches. Dalton Trans. 29, 3107-3120 (2007).
Pennella, M. A., Arunkumar, A. I. & Giedroc, D. P. Individual metal ligands play distinct functional roles in the zinc sensor Staphylococcus aureus CzrA. J. Mol. Biol. 356, 1124-1136 (2006).
Aguirre, J. D. & Culotta, V. C. Battles with iron: manganese in oxidative stress protection. J. Biol. Chem. 287, 13541-13548 (2012).
Anjem, A., Varghese, S. & Imlay, J. A. Manganese import is a key element of the OxyR response to hydrogen peroxide in Escherichia coli. Mol. Microbiol. 72, 844-858 (2009).
Tarrant, E. et al. Copper stress in Staphylococcus aureus leads to adaptive changes in central carbon metabolism. Metallomics 11, 183-200 (2019).
Abdul-Tehrani, H. et al. Ferritin mutants of Escherichia coli are iron deficient and growth impaired, and fur mutants are iron deficient. J. Bacteriol. 181, 1415-1428 (1999).
Mills, S. A. & Marletta, M. A. Metal binding characteristics and role of iron oxidation in the ferric uptake regulator from Escherichia coli. Biochemistry 44, 13553-13559 (2005).
Carter, K. P., Young, A. M. & Palmer, A. E. Fluorescent sensors for measuring metal ions in living systems. Chem. Rev. 114, 4564-4601 (2014).
Morgan, M. T. et al. Ratiometric two-photon microscopy reveals attomolar copper buffering in normal and Menkes mutant cells. Proc. Nat. Acad. Sci. 116, 12167-12172 (2019).
Nolan, E. M. & Lippard, S. J. Small-molecule fluorescent sensors for investigating zinc metalloneurochemistry. Acc. Chem. Res. 42, 193-203 (2009).
Zou, Z., Higginson, B. & Ward, T. R. Creation and optimization of artificial metalloenzymes: Harnessing the power of directed evolution and beyond. Chem 10, 2373-2389 (2024).
Harnden, K. A., Wang, Y., Vo, L., Zhao, H. & Lu, Y. Engineering artificial metalloenzymes. Protein Eng., 177-205 (2021).
Helalat, S. H., Jers, C., Bebahani, M., Mohabatkar, H. & Mijakovic, I. Metabolic engineering of Deinococcus radiodurans for pinene production from glycerol. Microb. Cell Fact. 20, 1-14 (2021).
Shomar, H. & Bokinsky, G. Towards a synthetic biology toolset for metallocluster enzymes in biosynthetic pathways: what we know and what we need. Molecules 26, 6930 (2021).
Arnold, F. H. Directed evolution: bringing new chemistry to life. Angew. Chem. Int. Ed. 57, 4143 (2018).
United Kingdom Government, D. f. S. I. a. T. National Vision For Engineering Biology, https://www.gov.uk/government/ publications/national-vision-for-engineering-biology (2023).
Alderighi, L. et al. Hyperquad simulation and speciation (HySS): a utility program for the investigation of equilibria involving soluble and partially soluble species. Coord. Chem. Rev. 184, 311-318 (1999).
Smith, R. M. & Martell, A. E. Critical stability constants. 1 (Amino acids, 1974).
Winter, G. xia2: an expert system for macromolecular crystallography data reduction. J. Appl. Crystallogr. 43, 186-190 (2010).
Kabsch, W. xds. Acta Crystallogr. D. Biol. Crystallogr 66, 125-132 (2010).
Winn, M. D. et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D. Biol. Crystallogr 67, 235-242 (2011).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658-674 (2007).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D. Biol. Crystallogr 66, 213-221 (2010).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D. Biol. Crystallogr 66, 486-501 (2010).
Meng, E. C. et al. UCSF ChimeraX: Tools for structure building and analysis. Protein Sci. 32, e4792 (2023).
Pravda, L. et al. MOLEonline: a web-based tool for analyzing channels, tunnels and pores (2018 update). Nucleic Acids Res. 46, W368-W373 (2018).
Osman, D. et al. Generating a metal-responsive transcriptional regulator to test what confers metal sensing in cells. J. Biol. Chem. 290, 19806-19822 (2015).
Foster, A. W., Patterson, C. J., Pernil, R., Hess, C. R. & Robinson, N. J. Cytosolic Ni(II) sensor in cyanobacterium: nickel detection follows nickel affinity across four families of metal sensors. J. Biol. Chem. 287, 12142-12151 (2012).
Kuzmič, P. Program DYNAFIT for the analysis of enzyme kinetic data: application to HIV proteinase. Anal. Biochem. 237, 260-273 (1996).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the method. Methods 25, 402-408 (2001).
Baba, T. et al. Construction of Escherichia coli K-12 in-frame, singlegene knockout mutants: the Keio collection. Mol. Sys. Biol. 2, 2006 (2006).
Acknowledgements
The Diamond Light Source is acknowledged for time on beamline IO4 under proposal MX32736, Prof. Timothy Blower, Prof. Liz Morris for assistance in crystallographic data collection and processing. This work was supported by Biotechnology and Biological Sciences Research Council awards BB/W015749/1 (N.J.R.), Understanding mis-metalation of native versus heterologously expressed protein, and BB/V006002/1 (N.J.R.), A calculator for metalation inside a cell, along with BB/S009787/ 1 (N.J.R.) supporting networking in Industrial Biotechnology. The authors acknowledge the contributions of Deenah Morton (née Osman) to whom the work is dedicated.
Author contributions
T.R.Y., S.E.C., A.G., E.T., and N.J.R. contributed to in vivo MncA metalbinding experiments. T.R.Y., A.G., S.E.C., and E.T. contributed to in vitro refolding experiments, with T.R.Y. doing foundation work with S.E.C. and A.G. completing the bulk of these experiments. T.R.Y. derived calculations to formulate competing metal buffers and to use MncA as a probe to refine estimates of intracellular metal availabilities and designed related experimental protocols. A.G. generated the crystal structure of -MncA. P.T.C. and A.J.P.S. analysed and DNA binding properties of Nill-RcnR. A.G., S.E.C. and E.T. determined metal contents of E. coli cells. S.E.C. performed and analysed transcript abundance by qPCR. Relationships between DNA occupancies of metal sensors, intracellular metal availabilities, and qPCR data, used methods established by T.R.Y., N.J.R. T.R.Y. developed the metalation calculations in a form used here.
N.J.R., S.E.C., with input from A.G. and E.T., wrote a first draft of the manuscript and generated graphics while all authors edited and approved the final version. T.R.Y., then S.E.C., then A.G., sequentially contributed the bulk of laboratory work at different phases of the programme, with T.R.Y. and S.E.C. making equivalently large contributions. N.J.R. with support of all authors, and especially T.R.Y., S.E.C., and A.G., interpreted the significance of the data. N.J.R. had overall responsibility for the design, finance, and management of the project throughout.
Correspondence and requests for materials should be addressed to Nigel J. Robinson.
Peer review information Nature Communications thanks Erik Yukl and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/ licenses/by/4.0/.
(c) The Author(s) 2025
¹Department of Biosciences, University of Durham, Durham, UK. ²Department of Chemistry, University of Durham, Durham, UK. These authors contributed equally: Sophie E. Clough, Tessa R. Young. e-mail: nigel.robinson@durham.ac.uk
Ratio of metal availabilities buffered according to Supplementary Table 1 and Supplementary Data 1. The proportion of each competing metal acquired by MncA as determined from the chromatograms in Fig. 1d. Preference for M 2 is calculated as the ratio of buffered concentrations of M 1 and column divided by the ratios of occupancies of Metal1 and Metal2 column except where noted. Calculated for M2 (Cu’) relative to Mn” based on the observed preference of relative to . For M2 relative to an assigned value of for (defined in the legend to Fig. 2e and elaborated in Supplementary Figs. 5a, b). As pseudo-dissociation constants: ; FeII ; ; . fourth trial competing with is not shown in Fig. 1c and the chromatograms are in Supplemental Fig. 2b. This result is in the calculated preference for (where experimental replicates, for the other determinations SD).