يُفترض أن الإبداع ينشأ من حالة ذهنية توازن بين التفكير العفوي والتحكم المعرفي، مما يتوافق مع الاتصال الوظيفي بين شبكة الوضع الافتراضي في الدماغ (DMN) وشبكة التحكم التنفيذي (ECN). هنا، نقوم بإجراء دراسة واسعة النطاق ومتعددة المراكز لاختبار هذه الفرضية. باستخدام نهج علم الأعصاب الشبكي التحليلي، نقوم بتحليل بيانات التصوير بالرنين المغناطيسي الوظيفي في حالة الراحة وأداء المهام الإبداعية عبر 10 عينات مستقلة من النمسا وكندا والصين واليابان والولايات المتحدة.-تشكل أكبر وأشمل دراسة في علم الأعصاب للإبداع من حيث التنوع العرقي حتى الآن. باستخدام تحليل الشبكة المستند إلى الزمن، نحقق في العلاقة بين الإبداع (أي، القدرة على التفكير المتباين) والتبديل الديناميكي بين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN). نجد أن الإبداع، ولكن ليس الذكاء العام، يمكن التنبؤ به بشكل موثوق من خلال عدد تبديلات DMN-ECN. من المهم أن نحدد علاقة على شكل U مقلوب بين الإبداع ودرجة التوازن بين تبديل DMN-ECN، مما يشير إلى أن الأداء الإبداعي الأمثل يتطلب ديناميات متوازنة في الشبكات الدماغية. علاوة على ذلك، دراسة تحقق مستقلة باستخدام التصوير بالرنين المغناطيسي الوظيفي (fMRI) (تظهر نتائج الدراسة زيادة في التبديل بين الشبكتين الدماغيتين DMN-ECN أثناء توليد الأفكار الإبداعية (مقارنة بحالة التحكم) وتعيد تأكيد العلاقة المعكوسة على شكل حرف U. لذلك، نقدم أدلة قوية عبر مجموعات بيانات متعددة المراكز تشير إلى أن الإبداع مرتبط بالقدرة على التبديل الديناميكي بين الشبكات الدماغية التي تدعم التفكير العفوي والمتحكم فيه.
التفكير الإبداعي هو قدرة إنسانية حيوية، تمكن تقدم الحضارة من خلال الابتكار المستمر في مجالات علمية وفنية متنوعة. في العقود الأخيرة، أسفرت الدراسات السلوكية ودراسات التصوير العصبي عن فهم كبير للآليات العصبية والمعرفية التي تكمن وراء كيفية توليد الناس لأفكار جديدة ومفيدة (أي، إبداعية).تشير الأدلة المتزايدة إلى أن التفكير الإبداعي ينشأ من التفاعل بين الروابط العفوية وعمليات التحكم المعرفي المتعمدة، التي يقودها بشكل أساسي التفاعلات بين شبكة الوضع الافتراضي (DMN) ومجموعة متنوعة من الشبكات الأخرى.على وجه الخصوص، تم ربط التفكير الإبداعي الأكبر بترابط الشبكة الافتراضية (DMN) وشبكة التحكم التنفيذي (ECN).
يتكون الشبكة الافتراضية (DMN) من الخط الأوسط القشري (بما في ذلك القشرة الجبهية الإنسية والقشرة الحزامية الخلفية) والمناطق الجدارية السفلية الخلفية، والفص الصدغي الأوسط؛ يُعرف أن هذه الشبكة الدماغية تتضمن التفكير الموجه داخليًا أو التفكير الذاتي، مثل التنشيط العفوي للذكريات، والتجول الذهني، والتكامل الدلالي، والأفكار العاطلة المرتبطة، والمحاكاة الذهنية – العمليات المعرفية التي يُعتقد أنها تدعم توليد الأفكار الإبداعية.يتكون الشبكة التنفيذية المركزية من مناطق الفص الجبهي الجانبي والمناطق الجدارية السفلية الأمامية، وعندما تت coupled مع الشبكة الافتراضية، تدعم استرجاع المعاني بشكل منظم ومرونة معرفية، مما يفيد في اختيار وتقييم الأفكار الإبداعية.لقد وجدت العديد من دراسات تصوير الدماغ أن التفكير الإبداعي يستقطب مناطق داخل الشبكة الافتراضية (DMN) وبالإضافة إلى ذلك، يمكن التنبؤ بقدرة التفكير الإبداعي الفردية من خلال التفاعلات الوظيفية داخل وبين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) سواء في حالة الراحة أو أثناء أداء المهام الإبداعية باستخدام طرق الشبكات الثابتة.. على سبيل المثال، وظيفي
يمكن أن تتنبأ ميزات الاتصال المرتبطة بالنمط الافتراضي، والتحكم التنفيذي، وشبكات البروز بالأداء الإبداعي الفردي خلال مهام التفكير المتباين، ويمكن تعميم هذه الميزات عبر عدة عينات مستقلة من تصوير الرنين المغناطيسي الوظيفي في حالة الراحة.نظرًا لأدوار الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) في الإدراك العفوي والمتحكم فيه على التوالي، يُعتقد أن الاتصال بين DMN وECN يعكس تنسيقًا بين العمليات العفوية/التوليدية والعمليات المتحكم فيها/التقييمية.ومع ذلك، فإن الأساليب الثابتة للشبكات تفشل في التقاط التنسيق الديناميكي للحالات المعرفية المتعلقة بإعادة التهيئة والتفاعلات بين الشبكات.لذلك، هناك حاجة إلى تحليلات وظيفية قائمة على إعادة التكوين الديناميكي لتحديد أنماط التفاعل المحددة بين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) المتعلقة بالقدرة الإبداعية.
كيف يحقق الدماغ الإبداعي التنسيق الديناميكي بين العفوية والسيطرة؟ وفقًا للإطار الديناميكي للفكر العفويالإبداع هو حالة ذهنية تتطلب توازنًا مثاليًا بين توليد الأفكار العفوية والمعالجة الموجهة نحو الأهداف – مما يتوافق مع حالة متوازنة من التواصل بين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN). كما تؤكد نظريات العمليات المزدوجة للإبداع أن الإدراك الإبداعي ينطوي على مرحلتين مختلفتين – توليد الأفكار والتقييم – تهيمن عليهما التعاون بين الشبكات الوظيفية الكبيرة في الدماغ بما في ذلك الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN).لذلك، إحدى الاحتمالات هي أن الأشخاص الأكثر إبداعًا يمكنهم التبديل بمرونة بين الفصل والتكامل بين شبكة الوضع الافتراضي (DMN) وشبكة التحكم التنفيذية (ECN) – متأرجحين بين حالات تعمل فيها الشبكات الدماغية بشكل مستقل ومتداخل، على التوالي – مما قد يعكس توازنًا مثاليًا أو “نقطة حلاوة” بين الشبكات التي تستفيد من قدراتها المستقلة والجماعية. على الرغم من الجاذبية البديهية لهذه النظرية، إلا أن هناك القليل جدًا من الأدلة التجريبية لدعم هذا الادعاء. وقد أشارت عدة دراسات استندت إلى مراحل المعالجة الإبداعية إلى أن شبكة الوضع الافتراضي (DMN) تعمل بشكل أساسي في المرحلة التوليدية في المراحل المبكرة من الإنتاج الإبداعي، في حين أن زيادة الترابط بين شبكة الوضع الافتراضي (DMN) وشبكة التحكم التنفيذية (ECN) تكمن وراء المرحلة التقييمية في المرحلة اللاحقة.، مما يوحي بأن الشبكة العصبية التنفيذية قد تراقب وتوجه العمليات التوليدية/الارتباطية العفويةبالإضافة إلى ذلك، تدعم الترابط الزمني لشبكات الدماغ واسعة النطاق المستندة إلى بيانات الرنين المغناطيسي الوظيفي في حالة الراحة التكوين الوظيفي الديناميكي المرتبط بالإبداع المعرفي.ومع ذلك، فإن أنماط التفاعل الدقيقة والدور الميكانيكي لهذا الارتباط بين الشبكة الافتراضية الشبكية (DMN) والشبكة التنفيذية (ECN) في التفكير الإبداعي لا تزال غير واضحة.
يمكن تقسيم هذا إعادة تكوين ديناميكية لربط الشبكات الدماغية إلى نمطين متميزين أو “حالات” يحتلها الدماغ بشكل روتيني: الانفصال والتكامل. يتوافق النمط المنفصل مع المعالجة المستقلة داخل الشبكات، بينما يتوافق النمط المتكامل مع التعاون بين الشبكات.أفادت الأبحاث السابقة بأهمية فصل الشبكات الدماغية ودمجها لأداء مهام معرفية متنوعة.على الرغم من عدم وجود أعمال سابقة درست ديناميات الشبكات الدماغية المتعلقة بالإبداع. ومع ذلك، فقد لاحظت عدة دراسات وجود تباينات في التعاون الزمني بين الشبكات الوظيفية خلال الإدراك الإبداعي، مما يشير إلى وجود انتقال ديناميكي محتمل بين الانفصال الوظيفي والتكامل.من منظور الفروق الفردية، من المرجح أن يشارك الأفراد ذوو الإبداع العالي في التفكير الموجه داخليًا أثناء الراحة، والذي يتميز بالشبكات الوظيفية الذاتية وتفاعلاتها.نظرًا لأن الشبكات الوظيفية في حالة الراحة تشكل تنشيطات المهام المعرفيةالأفراد الذين يمتلكون قدرات تفكير إبداعي أعلى – والتي قد تسهم في دورة أكثر كفاءة من التوليد والتقييم – قد يظهرون أيضًا ديناميكيات متغيرة لشبكات الدماغ الكبيرة (وهي الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN)) في حالة الراحة.لقد أظهرت الدراسات باستمرار أن الديناميات الشبكية الداخلية أثناء الراحة تعكس الخصائص التنظيمية الأساسية التي تتنبأ بالاختلافات الفردية في القدرات المعرفية.يبدو أن نمط وتكرار التبديل بين الحالات المنفصلة والمندمجة للشبكة مهمان بشكل خاص، حيث يشير ذلك إلى قدرة الدماغ على تنسيق عمليات معرفية مختلفة بمرونة.. إن دراسة هذه الديناميات أثناء الراحة يسمح لنا بتوصيف الفروق الفردية المستقرة في تنظيم الشبكات عبر عينات كبيرة، مما يكمل ويوسع الرؤى المستخلصة من الدراسات المعتمدة على المهام في الإدراك الإبداعي. هذه المقاربة ذات قيمة خاصة نظرًا للأدلة التي تشير إلى أن
القدرة الإبداعية تتعلق بأنماط داخلية من تفاعل الشبكات الدماغية التي تستمر عبر حالات الراحة والمهام. يتماشى هذا الرأي مع الدراسات حول الاتصال الديناميكي التي أفادت بأن التغير الزمني في الاتصال الوظيفي في حالة الراحة بين المناطق داخل الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) كان مرتبطًا بشكل إيجابي بقدرة التفكير الإبداعي. علاوة على ذلك، قدمت دراسة حديثة فهمًا أكثر دقة للآليات العصبية المعرفية للدماغ الإبداعي، مشيرة إلى أن المراحل التوليدية والتقييمية قد تتضمن تركيبات مختلفة من العمليات الترابطية والمتحكم بها، وبالتالي تجند نسبًا مختلفة من نشاط DMN وECN.
بغض النظر عن التفكير الإبداعي، قد تعكس إعادة تكوين الشبكة الدماغية الأكثر مرونة (أي، التبديل المتكرر لحالات الدماغ) توازنًا وظيفيًا بين الحالات المنفصلة والمندمجة التي تساعد على تعزيز القدرات المعرفية مثل استرجاع الذاكرة. على سبيل المثال، أفادت تحقيقات حديثة أنه بينما يدعم الانفصال القوي للدماغ الذكاء المتبلور وسرعة المعالجة، فإن الاندماج يعزز القدرة المعرفية العامة بشكل أفضل، فإن التوازن بين الانفصال والاندماج مرتبط بذاكرة أفضل. بالمقابل، فإن الزيادة في الاتصال (أي، زيادة الاتصال الإيجابي) بين DMN وECN، أو الشبكات الأخرى، مرتبطة بسلوكيات وأفكار جامدة مرتبطة ببعض الاضطرابات النفسية (مثل اضطراب طيف التوحد)، بينما يرتبط انخفاض الاتصال (أي، انخفاض الاتصال الإيجابي) بضعف في الوظيفة التنفيذية والإدراك الاجتماعي. بالعودة إلى الدراسة الحالية، لم يتم إجراء أي بحث حتى الآن حول التفاعلات الديناميكية بين DMN وECN في التفكير الإبداعي فيما يتعلق بأنماط التبديل بين الحالات المنفصلة والمندمجة التي يمكن أن تفيد الإبداع الفردي.
في الدراسة الحالية، نحقق فيما إذا كانت الفروق الفردية في القدرة الإبداعية يمكن التنبؤ بها بشكل موثوق من خلال التفاعل الديناميكي بين DMN وECN. لهذا الغرض، نقوم بتحليل مجموعات بيانات كبيرة ومتعددة المراكز تشمل التصوير بالرنين المغناطيسي الهيكلي، والتصوير بالرنين المغناطيسي الوظيفي في حالة الراحة، وبيانات القدرة الإبداعية (المقاسة من خلال اختبارات التفكير المتباين التي تم إجراؤها خارج جهاز التصوير) من خمسة مراكز في النمسا وكندا والصين واليابان والولايات المتحدة ()-تشكل (حسب علمنا) أكبر وأشمل تحليل قائم على الدماغ للإبداع حتى الآن. قمنا بتطبيق نهج علم الأعصاب الشبكي التحليلي التلوي لدمج النتائج من خلال إجراء تحليل جديد باستخدام البيانات الخام من كل مجموعة بيانات. هنا، تم تحديد حالتين عصبيتين ثنائيتين، أنماط الاتصال الوظيفي المنفصل والمندمج بين DMN وECN، باستخدام تحليلات زمنية لحالة الراحة من بيانات التصوير بالرنين المغناطيسي الوظيفي. بناءً على الأبحاث السابقة, افترضنا أن تكرارًا أعلى للتبديل بين انفصال DMN-ECN والاندماج مع مرور الوقت سيتنبأ بأداء إبداعي أفضل. كما افترضنا أن الأداء الإبداعي الأعلى قد يستفيد من توازن وظيفي بين الانفصال والاندماج في هذه الشبكات الدماغية. أخيرًا، لتوفير تحقق أكثر مباشرة من ديناميات الاقتران العصبي، أجرينا تحليل تحقق خارجي باستخدام بيانات التصوير بالرنين المغناطيسي الوظيفي المعتمدة على المهام في عينة مستقلة من البالغين الصينيينواختبرنا ما إذا كانت ديناميات الشبكة الدماغية التي تم تقييمها أثناء الراحة في العينة الكاملة تمتد إلى الأداء النشط أثناء مهمة التفكير الإبداعي.
النتائج
تركيب العينة والنتائج السلوكية
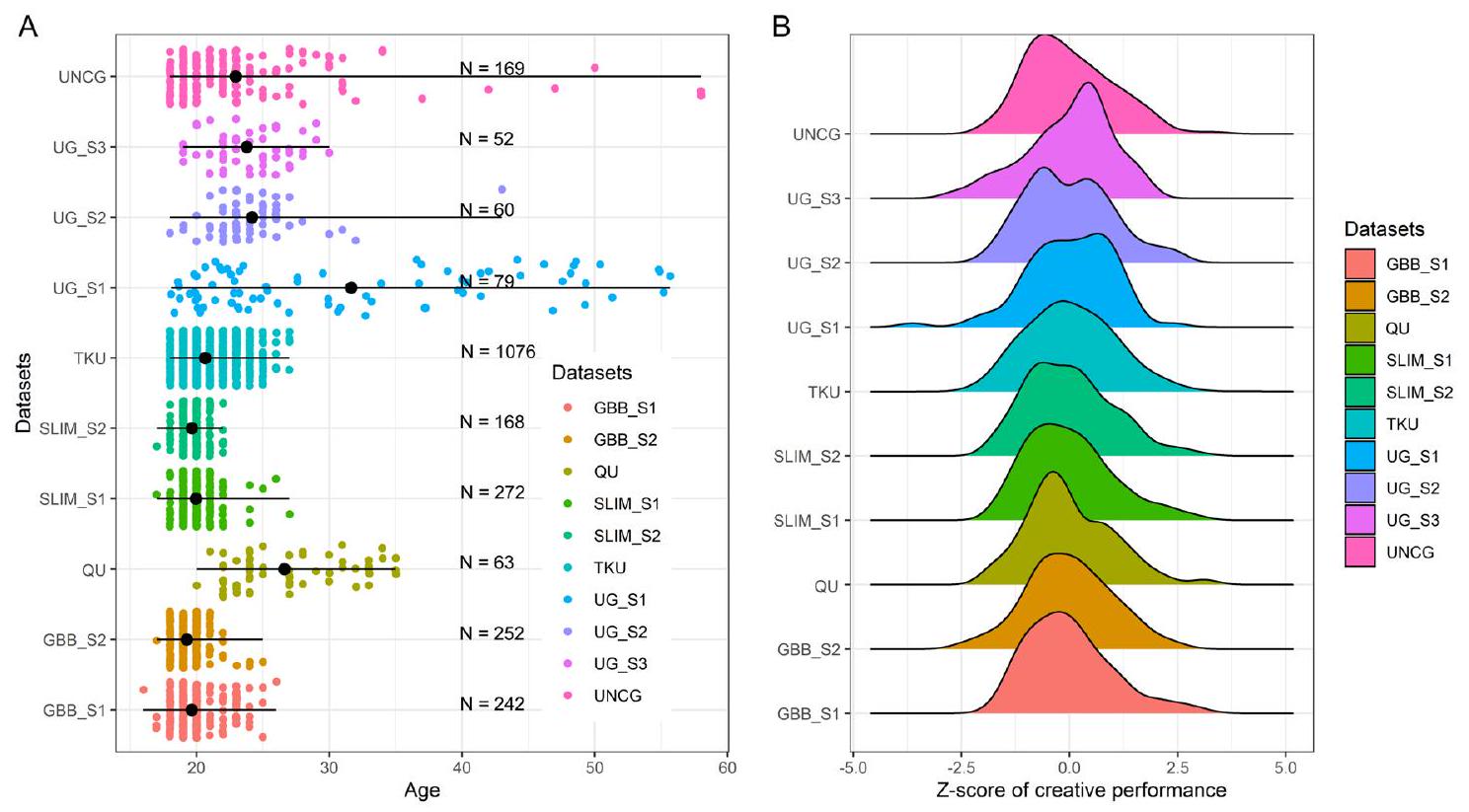
تم تحليل عشر مجموعات بيانات تغطي خمسة مراكز، بما في ذلك بيانات من 2433 فردًا صحيًا تتراوح أعمارهم بين 16 و58 عامًا. تلخص الشكل 1A أحجام العينات ونطاقات الأعمار لكل مجموعة بيانات (الجدول التكميلي 1) ويصف الشكل 1B توزيع بيانات الأداء الإبداعي لكل مجموعة بيانات. تم تقييم الأداء الإبداعي الفردي من خلال مهمة الاستخدامات البديلة المستخدمة على نطاق واسع, حيث يقوم المشاركون بتوليد استخدامات إبداعية للأشياء وتم تقييم ردودهم من قبل مقيمين مدربين (انظر معلومات مجموعة البيانات التكملية لمزيد من التفاصيل). أظهرت التحليلات السلوكية (الجدول التكميلي 2) عدم وجود اختلافات متسقة بين الجنسين عبر المجموعات العشر، وهو ما يتماشى مع الأدبيات السابقة. لم يتم العثور على ارتباط كبير بين درجات الإبداع والعمر باستثناء مجموعة بيانات واحدة (UG_S1). وبالتالي، فإن غياب النتائج المتسقة عبر مجموعات بيانات متنوعة يشير إلى أن اختلافات الجنس في درجات الإبداع والارتباطات بين الأداء الإبداعي والعمر تفتقر إلى القوة بسبب تباين العينة و
الشكل 1 | خصائص مجموعات البيانات. A العدد الإجمالي للمشاركين ونطاقات الأعمار لكل مجموعة بيانات. تشير موضع النقطة السوداء على كل خط إلى العمر الوسيط لتلك المجموعة. B توزيع الأداء الإبداعي بناءً على درجات الأصالة التي قيمها البشر في مهام التفكير المتباين في كل مجموعة بيانات.
توزيع إيجابي مائل في العمر. من بين ست مجموعات بيانات تحتوي على كل من درجات الذكاء والإبداع، أظهرت ثلاث مجموعات بيانات ارتباطًا كبيرًا بين البناءين: UNCG،CI; UG_S1،CI; UG_S3،CI; SLIM_S1،CI [-0.05 إلى 0.19]; GBB_S1،CI [-0.04 إلى 0.30]; وGBB_S2،إلى 0.12، بما يتماشى مع الأعمال السابقة التي تظهر ارتباطات إيجابية بين الذكاء والإبداع.
العلاقة بين تكرار تبديل الشبكة والأداء الإبداعي
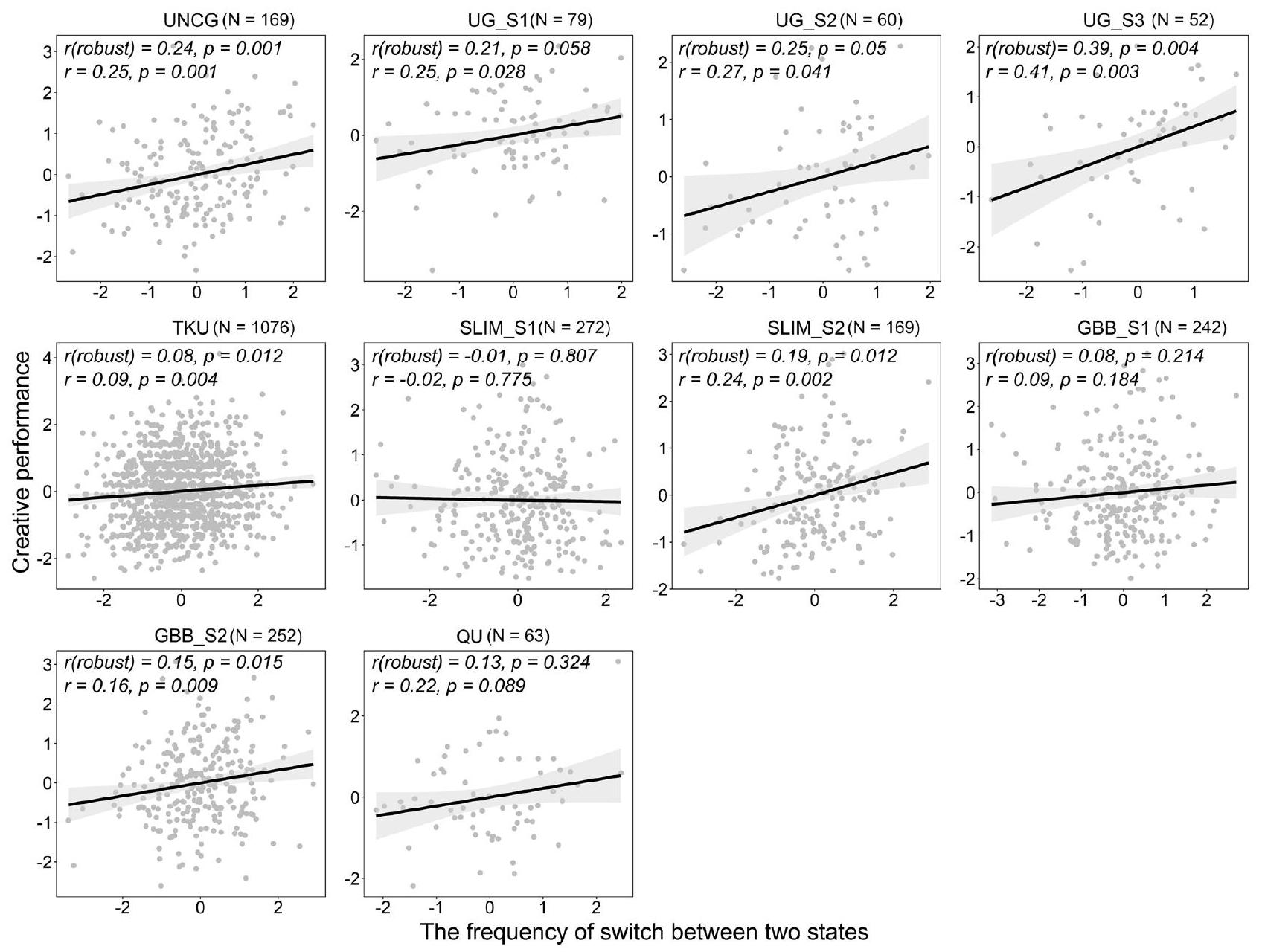
بعد ذلك، انتقلنا لاختبار فرضيتنا الرئيسية بشأن ما إذا كان التفكير الإبداعي يستفيد من الميل للتبديل ديناميكيًا بين الانفصال والاندماج لشبكة DMN وECN. تم حساب تكرار التبديل من خلال عدد الانتقالات بين الحالات المنفصلة والمندمجة عبر نوافذ الزمن (انظر “الطرق”). أكدت تحليلات الارتباط بيرسون فرضيتنا عبر 7 من 10 مجموعات بيانات (الشكل 2): كان تكرار التبديل بين انفصال DMN-ECN والاندماج مرتبطًا بشكل إيجابي وكبير بالأداء الإبداعي. بعد ذلك، قمنا بتطبيق نهج تحليلي تلوي لحساب حجم التأثير العام الموزون بين تكرار تبديل حالة DMN-ECN والقدرة الإبداعية عبر المجموعات العشر. كان حجم التأثير العام كبيرًا عند تطبيق تأثيرات عشوائية على مستوى مجموعة البيانات، (الشكل 3). كان اختبار Q للتباين كبيرًا،,,, مما يشير إلى ضرورة استكشاف المحاور المحتملة لهذه التأثيرات
. نظرًا لأن المتغيرات الثلاثة المعدلة – الموقع (المقابل لخمس مراكز)، نوع جهاز التصوير، وطرق التقييم – متداخلة تقريبًا، أجرينا تحليلًا معدلاً شمل جميع المتغيرات الثلاثة كمعدلات معًا. بالإضافة إلى ذلك، بعد التحليلات الفرعية، قمنا بحساب التأثير النقي لكل مجموعة بعد استبعاد المتغيرات الأخرى، مثل الموقع ونوع جهاز التصوير. كشفت التحليلات المعدلة التالية أن الموقع ونوع جهاز التصوير وطرق التقييم،, كانت بشكل كبير ومشترك معدلة للتأثير العام. بعد ذلك، قمنا بإجراء تحليلات فرعية ضمن المركز ونوع جهاز التصوير وطرق التقييم للإبداع حيث يكون عدد التأثيرات (k) على الأقل 3: مجموعات بيانات النمسا () لها تأثير إيجابي كبير ومتوسط،, ومجموعات بيانات الصين () لها تأثير إيجابي ضعيف نسبيًا وكبير
،. مجموعات البيانات التي تستخدم جهاز التصوير بالرنين المغناطيسي المغناطيسي () لها تأثير إيجابي كبير ومتوسط،، بينما مجموعات البيانات التي تستخدم جهاز التصوير بالرنين المغناطيسي Trio () لها تأثير إيجابي كبير وضعيف،CI. مجموعات البيانات التي تستخدم التقييم بناءً على الأصالة فقط () لها تأثير إيجابي كبير ومتوسط،، ومجموعات البيانات التي تستخدم التقييم بناءً على مجموع الأصالة والطلاقة () لها تأثير ضعيف ولكنه كبير،. بالإضافة إلى ذلك، حجم العينة،، العمر،، وقت المسح،، ونسبة الجنس في العينة،لم تؤثر بشكل كبير على التأثير العام، مما يشير إلى أن مصدر البيانات (بما في ذلك المواقع وبروتوكول التصوير بالرنين المغناطيسي والتقييم) ولكن ليس المتغيرات المتعلقة بالمشاركين هي التي أثرت على التأثير العام. وبالتالي، يميل الأفراد الأكثر إبداعًا إلى التبديل بشكل أكثر تكرارًا بين الفصل والتكامل لشبكة الوضع الافتراضي (DMN) والشبكة التنفيذية المركزية (ECN) أثناء الراحة.
علاوة على ذلك، ظلت النتائج ذات دلالة إحصائية عند استخدام طريقة الارتباط القوية، التي لا تعتمد على افتراضات الطبيعية أو الخطية ويمكنها التعامل مع القيم الشاذة بشكل أفضل من طرق الارتباط التقليدية. علاوة على ذلك، وُجد أن حجم التأثير الكلي كان ذا دلالة. سي آي [ ” ]، باستخدام نموذج التأثيرات العشوائية (الشكل التوضيحي التكميلي 1). لم يكن اختبار Q للتباين ذا دلالة هنا،مما يشير إلى تباين منخفض نسبيًا بين الدراسات.
العلاقة بين تردد التبديل والذكاء
نظرًا للعلاقة الشرطية بين الإبداع والذكاء، قمنا بتطبيق نهج تحليل ميتا لتحديد ما إذا كان الذكاء، الذي يُعتبر مهمة تحكم، يرتبط أيضًا بالفصل والتكامل بين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN)، كما توقعنا (ووجدنا) بالنسبة للإبداع. وبالتالي، تم إجراء نفس التحليلات بين 6 مجموعات بيانات.، انظر الجدول التكميلي 3) الذي شمل درجات الذكاء بالإضافة إلى درجات الإبداع. لم يكن حجم التأثير الكلي ذا دلالة إحصائية،، دون تباين كبير،، على النقيض من ذلك، كان حجم التأثير العام للعلاقة بين تردد التبديل والأداء الإبداعي كبيرًا بين نفس مجموعات البيانات الست.، فرق تقدير المعامل بين النموذجين (الذكاء مقابل الإبداع) ذو دلالة إحصائية.، مما يدل على
الشكل 2 | العلاقات بين الأداء الإبداعي وتكرار التحويل لفصل وتكامل الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN). (قوي) أشار إلى قيمة الارتباط القوي في كل مجموعة بيانات. يتم عرض خط الانحدار الملائم باللون الأسود، و الinterval الثقة للخط موضح بالتظليل الرمادي. كلا من (الأداء الإبداعي) و محاور (الأداء الإبداعي) هي قيم موحدة بعد التعديل وفقًا للجنس والعمر ومتوسط الإزاحة الإطارية ومتوسط الإشارة العالمية. أن تأثير تبديل DMN-ECN خاص بالقدرة الإبداعية ولا يمتد إلى الذكاء.
الأداء الإبداعي وتوازن فصل وتكامل شبكة الوضع الافتراضي والشبكة التنفيذية
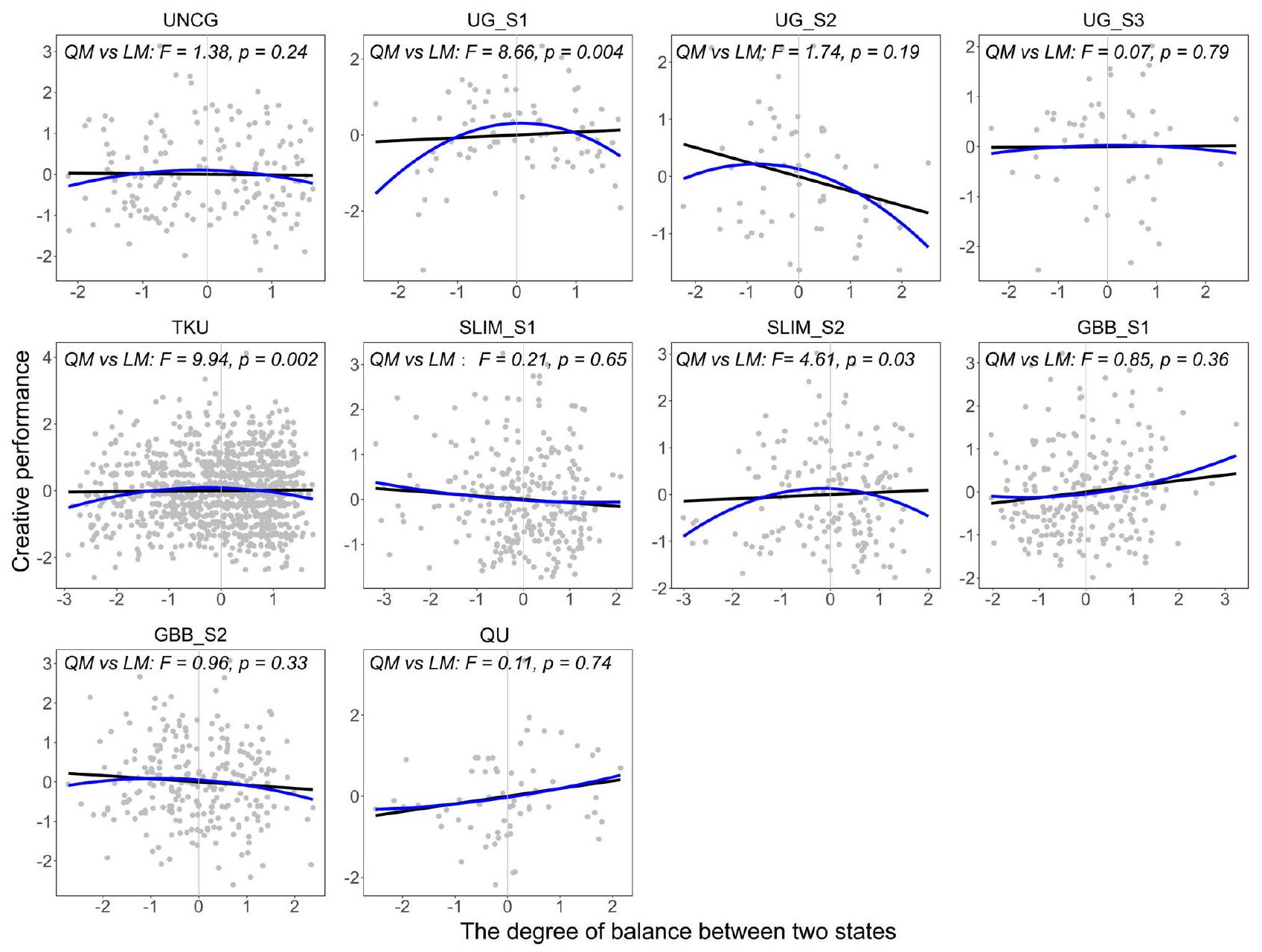
لقد أظهرت الأدلة التجريبية السابقة أن الأفراد الذين يتمتعون بدرجات مختلفة من التوازن الوظيفي بين تمايز الشبكة واندماجها يظهرون قدرات معرفية متفاوتة.علاوة على ذلك، فإن التبديل المتكرر بين الحالات المنفصلة والمندمجة يؤدي إلى توازن وظيفي، وقد يعكس التوازن الأمثل (أي عندما تكون قيمة التوازن مساوية للصفر) أقصى مرونة في الانتقالات الديناميكية بين الحالتين.. هنا، هدفنا إلى اختبار ما إذا كانت هناك علاقة غير خطية بين الأداء الإبداعي وتوازن فصل ودمج الشبكتين الدماغيتين DMN وECN؛ لذلك قمنا بإجراء اختبار ANOVA وقارنّا قيم AIC بين نموذج الانحدار الخطي ونموذج الانحدار التربيعي في كل مجموعة بيانات.
وجدنا أن نموذج الانحدار التربيعي كان ملائمًا بشكل أفضل بشكل ملحوظ من نموذج الانحدار الخطي (الشكل 4) في ثلاث مجموعات بيانات: UG_S1شكراً لك، و SLIM_S2 لم يكن هناك فرق كبير بين النموذجين في مجموعات البيانات السبع المتبقية (الجدول التكميلي 4).
لتلخيص أحجام التأثير لنموذج الانحدار التربيعي عبر جميع مجموعات البيانات، تم تحليل معاملات الارتباط الجزئي للحد التربيعي في كل نموذج انحدار باستخدام نموذج التأثيرات العشوائية مع أقصى احتمال مقيد. كان حجم التأثير الإجمالي صغيرًا ولكن
مجموعات البيانات
الموقع
وزن r -القيمة [95% فترة الثقة]
جامعة شمال كارولينا
الولايات المتحدة الأمريكية
فم
6.92%
0.26 [0.10، 0.41]
UG_S1
النمسا
5.25%
0.26 [0.03، 0.48]
UG_S2
النمسا
٤.٢٤٪
0.28 [0.02، 0.54]
UG_S3
النمسا
⟵ •
3.75%
0.44 [0.16، 0.72]
شكراً
اليابان
–
11.36%
0.09 [0.03، 0.15]
SLIM_S1
الصين
–
18.14%
-0.02 [-0.14, 0.10]
SLIM_S2
الصين
12.58%
0.24 [0.09، 0.40]
جي بي بي_س1
الصين
16.72%
0.09 [-0.04, 0.22]
جي بي بي_س2
الصين
17.21%
0.16 [0.04، 0.29]
كيو
كندا
3.81%
0.22 [-0.03, 0.48]
نموذج RE ذو المستويين لجميع مجموعات البيانات:
100.00%
0.17 [0.08، 0.27]
الشكل 3 | مخططات الغابة لعشر أحجام تأثير مدرجة في التحليل التلوي. مخطط الغابة لأحجام التأثير للارتباط بين الأداء الإبداعي وتكرار التبديل (محسوب باستخدام بيانات fMRI في حالة الراحة). على الجانب الأيسر، تعرض مخططات الغابة اسم وموقع كل مجموعة بيانات؛ على يسار الخط العمودي المتقطع (الذي يمثل عدم وجود تأثير)، توفر مخططات الغابة الوزن، التأثير الملحوظ، وفترة الثقة، حيث يكون حجم المربع متناسبًا مع الأوزان المحددة بواسطة عكس تباين مجموعات البيانات؛ الخط السفلي يظهر حجم التأثير الكلي (يمين) واختبار التباين (يسار).
مهم، مما يشير إلى أن التعاون المعتدل – لا أكثر ولا أقل – بين الشبكة الديناميكية للدماغ (DMN) والشبكة التنفيذية (ECN) قد يكون مفيدًا في توليد الأفكار الإبداعية. قد تشير هذه النتيجة إلى أن القدرة الإبداعية تستفيد من البيانات متعددة المراكز،
الشكل 4 | العلاقات بين الأداء الإبداعي وتوازن فصل الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) ودمجهما. العلاقة الخطية (الخط الأسود الملائم) والعلاقة التربيعية (الخط الأزرق الملائم) للأداء الإبداعي (-محور) وتوازن الفصل والتكامل بين DMN-ECN (-المحور)، ومع المقارنة بين النموذج الملائم الخطي ونموذج ملائم تربيعي لكل مجموعة بيانات (محسوب باستخدام بيانات fMRI في حالة الراحة). كلا من و المحاور هي قيم موحدة بعد التعديل وفقًا للجنس والعمر ومتوسط الإزاحة الإطارية ومتوسط الإشارة العالمية. التوازن الأمثل بين العمليات العفوية والمتحكم فيها، مدعومًا بالتبديل المعتدل بين فصل شبكة الوضع الافتراضي (DMN) وشبكة التحكم التنفيذية (ECN) ودمجهما. إحصائية اختبار Q،يشير إلى أن القليل من التباين موجود.
إمكانية التكرار
من المهم أننا قمنا بتقييم قابلية تكرار نتائجنا باستخدام التحليلات الضخمة، وهي نهج يجمع جميع البيانات الفردية المتاحة من مجموعات بيانات مختلفة. تتيح لنا هذه الطريقة استبعاد المتغيرات غير ذات الصلة، مثل العوامل الديموغرافية (مثل الجنس والعمر) وعوامل الموقع، والتركيز على التأثيرات المحددة ضمن نموذج واحد. تم تلخيص نتائج التحليل الضخم في الجدول التكميلية 5، مما يشير إلى أن الأفراد الذين لديهم تكرار أعلى لفصل ودمج الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) أظهروا أداءً إبداعيًا أعلى.، (الشكل 5A). من خلال مقارنة النماذج الخطية وغير الخطية باستخدام التحليل الضخم، أظهرت النتائج أن توازن التبديل الديناميكي بين فصل الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) والتكامل، كالعوامل التربيعية، يمكن أن يتنبأ بشكل كبير بالأداء الإبداعي، (الشكل 5B)، والنموذج غير الخطي (AIC = 6925.45، الجدول التكميلية 6) أفضل من النموذج الخطيالجدول التكميلي 7) الذي يتضمن فقط الحد الخطي (توازن حالات الدماغ الديناميكية) كعامل ثابت،. بشكل عام، أسفرت التحليل الضخم عن نتائج مشابهة لتلك التي حصلت عليها الطريقة التحليلية المجمعة. إعادة تحليل البيانات باستخدام أطلس دماغ شين، التي تتكون من 54 منطقة اهتمام داخل الشبكة الافتراضية للراحة (DMN) و
أكدت ECN أيضًا نتائجنا: كان حجم التأثير الكلي كبيرًا باستخدام نموذج التأثيرات العشوائية، (الشكل التوضيحي 2).
التحقق الخارجي باستخدام بيانات fMRI المستندة إلى المهام
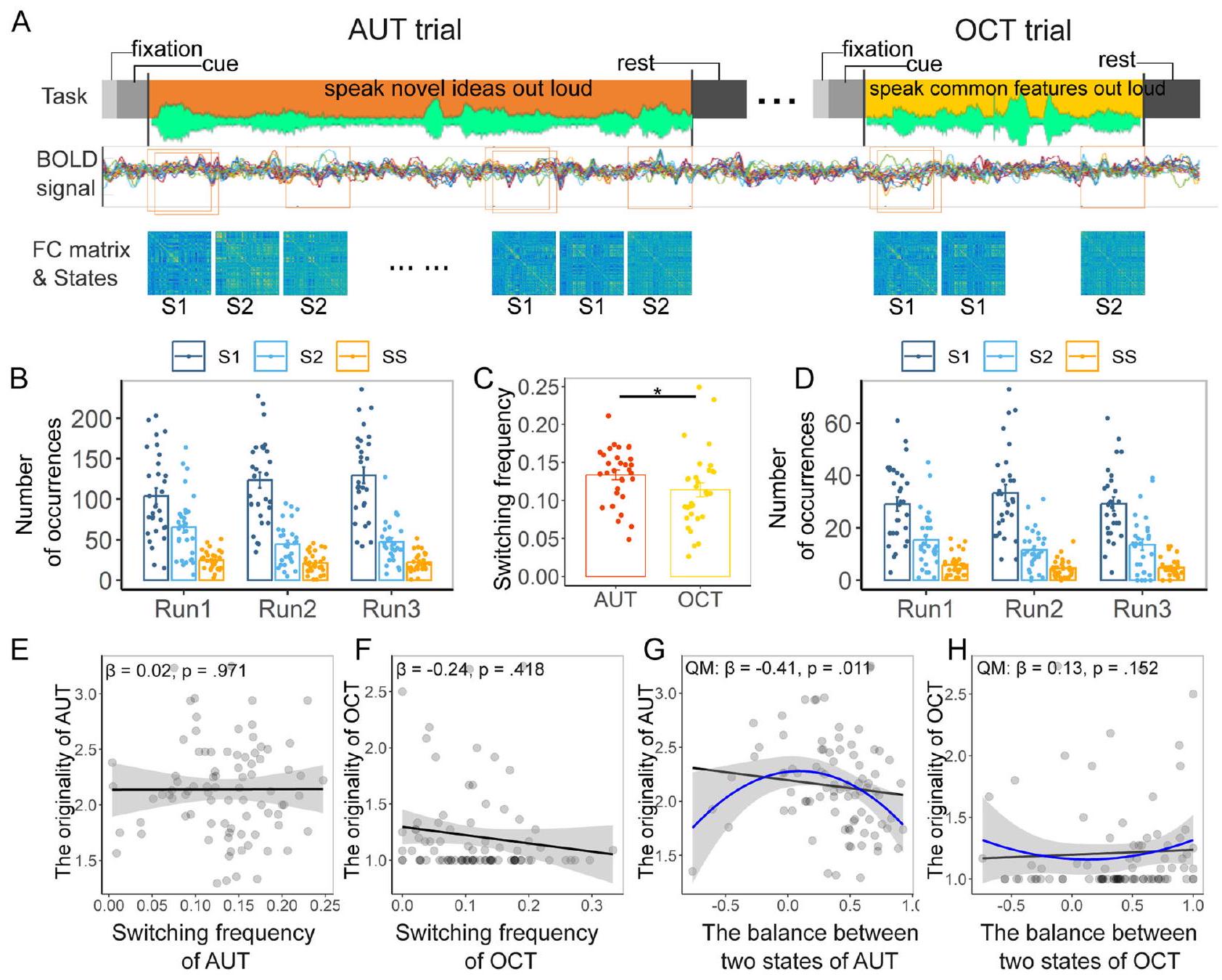
لقد حددنا أنماط تفاعلية ديناميكية بين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) المرتبطة بالأداء الإبداعي الفردي في مجموعة بيانات كبيرة متعددة المراكز من تصوير الرنين المغناطيسي الوظيفي في حالة الراحة. ومع ذلك، هل يمكن التحقق من صحة هذه الأنماط في سياق مهمة التفكير المتباين، التي تتضمن عمليات إدراكية إبداعية، على عكس مهمة التحكم التي تتطلب فقط إدراكات شائعة (مثل استرجاع الذاكرة)؟ لمعالجة ذلك، أجرينا تحليل تحقق خارجي باستخدام بيانات تصوير الرنين المغناطيسي الوظيفي أثناء المهمة، حيث أكمل واحد وثلاثون مشاركًا كل من مهمة الاستخدامات البديلة (AUT)، التي تتطلب إنتاج استخدامات إبداعية للأشياء، ومهمة خصائص الأشياء (OCT)، التي تتطلب إنتاج خصائص شائعة للأشياء، أثناء مسح الرنين المغناطيسي الوظيفي. تم اشتقاق تكرار التبديل ودرجة التوازن بين حالتين (أي، حالة DMN-ECN المنفصلة أو المتكاملة) من الأحداث ذات الصلة بالمهمة. على وجه التحديد، قمنا باستخراج هذين المؤشرين الديناميكيين من خلال تحديد تجارب AUT وOCT بدقة عندما بدأ المشاركون في التفكير، مما أدى إلى توليد ما يصل إلى أربعة أفكار (الشكل 6A). ثم تم حساب المؤشرين لكل مهمة لكل مشارك من خلال حساب تكرار التبديل وتوازن انفصال DMN-ECN ودمجه في كل جولة.
أظهرت النتائج أن AUT (; الشكل 6B) استقطبت تكرار تبديل أعلى بشكل ملحوظ من OCT (; الشكل 6D)، , كوهين’s (الشكل 6C). وهذا يشير إلى أن الإبداع
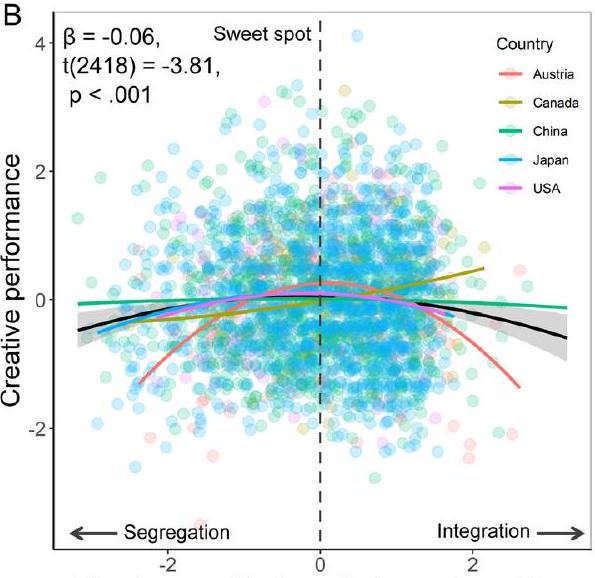
الشكل 5 | نماذج التأثير المختلط. أ العلاقة الخطية بين الأداء الإبداعي وتكرار التبديل بين حالات DMN-ECN المنفصلة والمتكاملة (محسوبة باستخدام بيانات تصوير الرنين المغناطيسي الوظيفي في حالة الراحة)؛ الخط الأسود يحدد الاتجاه العام، وinterval الثقة للخط موضح في الظل الرمادي. ب العلاقة التربيعية بين درجة الإبداع والتوازن الوظيفي بين حالات DMN-ECN المنفصلة والمتكاملة (محسوبة باستخدام بيانات تصوير الرنين المغناطيسي الوظيفي في حالة الراحة)؛ الخط الأسود يمثل الاتجاه العام، مع interval الثقة المشار إليه بالظل الرمادي، بينما تمثل الخطوط الملونة الاتجاهات لكل مركز. الجانب الأيسر من -المحور يشير إلى الانفصال الشديد، والجانب الأيمن يشير إلى الدمج الشديد، والخط المنقط في المنتصف يمثل التوازن الأمثل أو “نقطة الحلاوة”.
أ
درجة التوازن بين حالتين
قد يتم دعم التفكير من خلال التفاعلات بين DMN وECN مقارنة بالعمليات الإدراكية العامة، مثل استخراج المفاهيم من المعرفة الموجودة. لم يظهر نموذج التأثيرات المختلطة الخطية أن أداء AUT (درجة أصالة الأفكار) مرتبط بشكل كبير بتكرار تبديل انفصال DMN-ECN ودمجه أثناء التفكير الإبداعي، , كوهين’s (الشكل 6E)؛ وبالمثل، لم يكن أداء OCT مرتبطًا بتكرار التبديل، (الشكل 6F). ومع ذلك، أظهرت تحليلات نموذج التأثيرات المختلطة غير الخطية أن درجة التوازن بين انفصال DMN-ECN ودمجه مرتبطة بأداء AUT، , وأن النموذج غير الخطي AIC أفضل من النموذج الخطي AIC , الذي يتضمن فقط الحد الخطي (توازن حالات DMN-ECN الديناميكية) كعامل ثابت، , (الشكل 6G). لكن توازن تبديل DMN-ECN الديناميكي ليس مرتبطًا بأداء OCT في كل من النموذج غير الخطي، (54) , والنموذج الخطي، , (الشكل 6H).
لذا، فإن نتائج تصوير الرنين المغناطيسي الوظيفي المستندة إلى المهمة تحقق نتائج حالة الراحة: التفكير الإبداعي يستحث تبديل ديناميكي أكبر وتفاعلات أكثر توازنًا بين الشبكة الافتراضية والشبكات التنفيذية مقارنة بالإدراك غير الإبداعي، مما يدعم الأداء الإبداعي الفردي.
نقاش
كشفت تحليلاتنا الميتا-تحليلية أن الأداء الإبداعي يمكن التنبؤ به بشكل موثوق من تكرار التبديلات بين حالات DMN وECN المنفصلة والمتكاملة في حالة الراحة: الأشخاص الذين يتبدلون بين هذه الحالات بشكل متكرر ينتجون أفكارًا أكثر أصالة. علاوة على ذلك، كانت القدرة العالية على التفكير الإبداعي مرتبطة بتوازن بين الحالات المنفصلة والمتكاملة لأن الانتقالات المتكررة بين هذه الحالات وفرت وسيلة للحفاظ على هذا التوازن. إن التقارب بين هذه الأنماط في حالة الراحة عبر عينة كبيرة وظهورها الانتقائي أثناء أداء المهام الإبداعية (مقارنة بمهام التحكم) يشير إلى أن ديناميات الشبكة هذه تمثل علامة محددة على القدرة الإبداعية. هذه النتائج تعزز فهمنا للآليات العصبية للإبداع من خلال تحديد أنماط محددة من تفاعل الشبكة التي تميز كل من القدرة على والانخراط النشط في الفكر الإبداعي.
بينما تم إحراز تقدم كبير، لا يزال فهمنا للأسس العصبية للإبداع بعيدًا عن الاكتمال. الإبداع هو بناء معقد يستقطب كل من العمليات الإدراكية الخاصة بالإبداع والعمليات الإدراكية العادية مثل الذاكرة والانتباه والتحكم الإدراكي . إن نهج الشبكة الديناميكية مناسب بشكل خاص لمعالجة التفاعل المعقد للآليات العصبية التي تشغل الدماغ بأكمله أثناء التفكير الإبداعي . في هذا السياق، ركزت الدراسة الحالية على ECN بأكملها
وشبكة DMN في الإدراك الإبداعي، مع تجاهل الفروق بين المناطق المحددة والشبكات الفرعية داخل هذه الشبكات الدماغية الكبيرة. بالإضافة إلى ذلك، توقعنا العثور على أنماط عامة عبر المجالات من تواصل DMN-ECN التفاعلي المرتبط بالإدراك الإبداعي. في الواقع، في البحث الحالي، لم نجد أي ارتباط كبير بين الذكاء وتكرار تبديل DMN-ECN عبر ست مجموعات بيانات. بالإضافة إلى ذلك، لاحظنا تكرار تبديل DMN-ECN أقل بشكل ملحوظ تحت مهمة التحكم (استرجاع الذاكرة) مقارنة بمهمة التفكير المتباين. قد تشير هذه الملاحظة إلى أن هذه التفاعلات الديناميكية بين DMN وECN تميز الإدراك الإبداعي بشكل خاص، بدلاً من القدرات الإدراكية الأخرى.
أظهرت مجموعة من الدراسات أن التفكير الإبداعي يعتمد عمومًا على قدرة DMN على التزامن مع شبكات دماغية أخرى , مثل , شبكة البروز , شبكة الحركية الحسية , والشبكة البصرية . بينما تم تسليط الضوء على الارتباط بين DMN وECN باستمرار كعلامة على الإبداع، فقد تم إظهار مثل هذا الارتباط بشكل كبير في أبحاث الاتصال الوظيفي “الثابت” ودور ديناميكية الارتباط المحددة كمحرك للإبداع كان حتى الآن تخمينيًا إلى حد كبير من خلال تحليل الاتصال الوظيفي للدماغ بالكامل . ومع ذلك، ركزت دراسات قليلة على فحص التفاعلات داخل هذه الشبكات وكيف تؤثر على الإدراك الإبداعي . على الرغم من أن بحثنا يعتمد على طرق تتضمن ارتباطات بين نمط الدماغ والسلوك، من خلال دمج الأدلة من كل من الشبكات في حالة الراحة ذات الحجم الكبير والشبكات الوظيفية الإدراكية (أي، تصوير الرنين المغناطيسي الوظيفي المستند إلى المهمة)، تدعم نتائجنا فكرة أن الارتباط الوظيفي بين المناطق في DMN وECN قد يعكس آلية عامة عبر المجالات للإدراك الإبداعي. من المهم، تم ملاحظة نمط تفاعل ديناميكي جديد بين DMN وECN في هذه الدراسة، وهو أن الأفراد الأكثر إبداعًا يظهرون تعاونًا عصبيًا مثاليًا بين DMN وECN. على وجه التحديد، أظهرت درجة التوازن بين الحالات المنفصلة والمتكاملة علاقة على شكل U مقلوب مع الأداء الإبداعي، حيث قد يؤثر الكثير (أو القليل جدًا) من الارتباط بين DMN-ECN سلبًا على الأداء الإبداعي. تتماشى هذه النتيجة مع نظريات الإبداع السابقة، التي تقترح أن الفكرة الإبداعية تنشأ من التوازن الدقيق بين المرونة والثبات في الحالات الإدراكية . هذا التوازن ضروري للحفاظ على التركيبة المثلى من الجدة والفائدة طوال عملية الإدراك الإبداعي .
أشارت الأبحاث السابقة إلى أن DMN قد يكون متورطًا بشكل مركزي في استرجاع الذكريات النشطة والعمليات الترابطية التي تنتج أفكارًا متنوعة، بينما قد تعمل ECN على توجيه العمليات الترابطية من خلال المراقبة، والحد، وعمليات التحكم من أعلى إلى أسفل ذات الصلة . تشرح هذه الدراسات أدوار DMN وECN والشبكات الأخرى في الإدراك الإبداعي في مرحلتين: التوليد والتقييم . اقترحت نظريات الإبداع السابقة أن
الشكل 6 | تصميم مهمة fMRI، التحليل الرئيسي، والنتائج. أ. نموذج تجريبي لمهمة التفكير المتباين، حيث كان على المشاركين إكمال مهمة الاستخدامات البديلة (AUT) ومهمة خصائص الكائنات (OCT) أثناء مسح fMRI. كان نموذج المهمة يتكون من علامة تثبيت (1 ثانية)، وإشارة تشير إلى الحالة القادمة (“استخدامات جديدة” أو “خصائص شائعة”; 4 ثواني)، وفترة تفكير تعرض إشارة كائن في نص (60 ثانية لـ AUT؛ 30 ثانية لـ OCT). استجاب المشاركون بالضغط على مفتاح والتحدث بإجاباتهم بصوت عالٍ في ميكروفون متوافق مع MRI حتى يولدوا أربع أفكار أو تنفد منهم فترة التفكير. تلا ذلك فترة راحة لا تقل عن 6 ثوانٍ، أو الوقت المتبقي من مرحلة التفكير إذا كان ذلك مناسبًا. تم معالجة بيانات مهمة fMRI مسبقًا باستخدام fmriprep، ثم تم استخراج المسارات الزمنية من 106 مناطق اهتمام مرتبطة بالشبكات الافتراضية وشبكات التحكم التنفيذي المستمدة من خريطة شيفر؛ بعد ذلك، تم استخدام نهج النافذة المنزلقة لحساب مصفوفة الاتصال الوظيفي (FC)، وتم تقسيم كل مصفوفة إلى مناطق منفصلة (حالة ) ودمجت (الدولةتستخدم الدول تجميع k-means. يعرض اللوح الأيسر العدد المتوسط للدول المنفصلة، المدمجة، والدول المتغيرة (SS) خلال مهمة AUT عبر ثلاث جولات. خطأ تشير الأعمدة إلى الخطأ المعياري للمتوسط. ج متوسط تردد التبديل لمهمة AUT وOCT، وعينة مزدوجة-اختبارات مقارنة بين AUT و OCT (يمثل شريط الخطأ الأخطاء المعيارية). تظهر اللوحة اليمنى العدد المتوسط للدول المنفصلة، المدمجة، والتبديل خلال مهمة OCT عبر ثلاث جولات. تشير أشرطة الخطأ إلى الخطأ المعياري للمتوسط. E، F تُظهر الرسوم البيانية المبعثرة ارتباطات بيرسون بين درجة الأصالة (أي، الأداء الإبداعي في المهمة،-المحور) وتردد التبديل لفصل ودمج DMN-ECN (-المحور) في AUT وOCT، على التوالي. الخط الأسود يمثل الاتجاه العام، مع فترة الثقة المشار إليها بالتظليل الرمادي. ج، ح خطي (خط أسود ملائم، مع فترة الثقة المشار إليها بالتظليل الرمادي) والعلاقة التربيعية (أي، النموذج التربيعي، QM؛ الخط الأزرق الملائم، معفترة الثقة المشار إليها بالتظليل الرمادي) لدرجة الأصالة (-محور) وتوازن الفصل والتكامل بين شبكة الوضع الافتراضي (DMN) والشبكة التنفيذية (ECN) (-المحور)، ومع المقارنة بين النموذج الخطي والنموذج التربيعي الملائم لـ AUT و OCT، على التوالي. الإبداع هو استمراريةوتحدث المخرجات الإبداعية على طول مجموعة من الاستمراريات المعرفية، مثل استمرارية التفكير المتباين-المتجمع، واستمرارية التركيز-عدم التركيز، واستمرارية العفوية-التعمد، واستمرارية تشتت الذهن-الوعي الذهني، وغيرها.في الإطار العصبي المعرفي الحالي، يمكن أن تتوافق مثل هذه الاستمراريات المعرفية مع ارتباط الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN)، حيث تعمل الشبكتان معًا على تحقيق دورة التوليد والتقييم. على سبيل المثال، قد يعكس ارتباط DMN-ECN أشكالًا من استرجاع الذاكرة الموجهة نحو الهدف والمحاكاة العرضية في مرحلة توليد الأفكار التي تتطلب التحكم المعرفي.بينما تعمل أنماط الربط المماثلة على تقييد وتوجيه عمليات التقييم لإنتاج أفكار تكون جديدة ومناسبة في مرحلة تقييم الأفكار.. تشير بعض الأدلة إلى أن الاختلافات قد تت correspond نسب نشاط DMN و ECN ضمن أنماط الاقتران إلى مراحل توليد وتقييم التفكير الإبداعي.لذا، من المحتمل أن التحول الديناميكي بين التوليد والتقييم يمكن أن يوفر طريقة مثلى لإنتاج تدفق مستمر من المخرجات الإبداعية. ومع ذلك، يجب أن يكون هذا التحول الديناميكي بين التوليد والتقييم متوازنًا بعناية، حيث إن الإفراط في أحدهما دون الآخر من غير المرجح أن ينتج أفكارًا جديدة ومناسبة..
عملنا يعزز، وإن كان بشكل غير مباشر، العلاقة بين ديناميات تبديل الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) ونموذج الإبداع القائم على التوليد والتقييم. هناك حاجة إلى أدلة مباشرة تدعم هذه الفرضية، ويجب أن تهدف الأبحاث المستقبلية إلى توضيح الأساس العصبي والآليات الكامنة وراء هذا التوازن. باختصار، فإن نتائجنا من التحليل التلوي… تم التحقق من صحة نهج الشبكة التحليلية في علم الأعصاب باستخدام بيانات fMRI في حالة الراحة التي تم جمعها سابقًا من خلال دراسة fMRI قائمة على المهام. وبالتالي، تساهم نتائجنا في جهد معرفي وعلمي عصبي مركز لتوضيح الآليات المعرفية والعصبية التي تحقق التفكير الإبداعي..
القيود والاتجاهات المستقبلية
نحن نعترف بعدة قيود في دراستنا. أولاً، ندرك أن حجم التأثير “الحقيقي” صغير، مما يعني أن إعادة التكوين الديناميكي بين الشبكة الافتراضية القشرية (DMN) والشبكة التنفيذية (ECN) هي مجرد عامل واحد يساهم في القدرة الإبداعية. استنادًا إلى بيانات التصوير بالرنين المغناطيسي الوظيفي في حالة الراحة، تشير نتائجنا التحليلية التراكمية إلى أن التفاعلات الأكثر كفاءة بين DMN وECN تدعم قدرة إبداعية أعلى؛ بدلاً من ذلك، مع تطور الأفراد لمهاراتهم الإبداعية، قد يزيد ذلك من كفاءة التفاعلات بين DMN وECN، حتى في حالة الراحة (على سبيل المثال، المرجع 86). من المهم أن نلاحظ أن العديد من العوامل الأخرى بخلاف الأنماط الديناميكية لـ DMN-ECN الملاحظة هنا متورطة بالتأكيد في القدرة الإبداعية، مثل الشخصية، والانتباه، والإدراك، والفضول، والمعتقدات الذاتية الإبداعية. ومع ذلك، تم العثور على أنماط تفاعل مشابهة بين DMN وECN لأمور أخرى.أشكال التفكير العفوي (مثل، تشتت الذهن). لذلك، هناك حاجة إلى مزيد من البحث لاستكشاف الشبكات الدماغية الكبيرة وتفاعلاتها الديناميكية عبر سياقات المهام الإبداعية المختلفة، لتمييز بين استقطاب الأنماط العصبية العادية وتلك الخاصة بالإبداع.
القيود الثانية لدراستنا هي تطبيق نهج الشبكات العصبية التحليلية الميتا لجمع البيانات الخام من الدراسات السابقة. مؤخرًا، تم اقتراح أن أحجام العينات الكبيرة جدًا (في عشرات الآلاف) مطلوبة لإجراء دراسات الارتباط على مستوى الدماغ القابلة للتكرار.. ومع ذلك، جادل آخرون بأهمية حجم التأثير على حجم العينة في مثل هذه الدراسات المرتبطة بالدماغ.نهجنا يستخدم طرق تحليل ميتا قياسية ومقبولة، وأحجام العينات للدراسات المضمنة كافية لأبحاث علوم الأعصاب الشبكية.تقدم دراسة حديثة رؤى قيمة حول أحجام العينات وأحجام التأثير الحقيقية في ارتباطات النمط الظاهري السلوكي الدماغي، مشيرة إلى أن أحجام العينات النموذجية غالبًا ما تؤدي إلى تضخيم أحجام التأثير وفشل التكرار. ومع ذلك، مع زيادة أحجام العينات إلى الآلاف (على سبيل المثال،تتحسن معدلات التكرار، وتقلل تضخيم حجم التأثير، على الرغم من أن الروابط الحقيقية قد تكون أصغر.لذلك، فإن حجم التأثير الملحوظ في دراستنا يعكس على الأرجح العلاقة الحقيقية بين التفاعل الديناميكي بين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) وقدرة التفكير المتباين. ومع ذلك، سيكون حجم العينة الكبير بما فيه الكفاية ومجموعة بيانات ذات تجانس عالٍ في طرق جمع البيانات (بما في ذلك التصوير بالرنين المغناطيسي وقياسات الإبداع) أكثر فائدة للكشف عن هذه العلاقة الحقيقية. بالإضافة إلى ذلك، نؤكد على الرسالة التي قدمها غرافتون وآخرون.الذي يعترف بالحاجة إلى دراسات مركزة على عينات صغيرة مع دراسات قائمة على تجمعات كبيرة النطاق من أجل التقدم حقًا في فهمنا لكيفية تحقيق التفاعلات العصبية المعقدة للسلوك البشري.
حد آخر محتمل من قيود الدراسة الحالية هو أن دقة الفضاء للأطالس المختارة، وخاصة عدد المناطق المضمنة ضمن الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN)، قد تؤثر على التأثير العام لهذه الدراسة. وقد أشارت بعض الدراسات إلى أن اختيار أطلس الدماغ يؤثر بشكل كبير على قوة العلاقة بين الاتصال الوظيفي والقدرات المعرفية.لذلك، هناك حاجة إلى أبحاث مستقبلية لتكرار وتوسيع نتائجنا باستخدام أطلسات دماغية إضافية ذات دقة مكانية مختلفة.
الاستنتاجات
تكشف النتائج الحالية أن الأشخاص ذوي الإبداع العالي يتميزون بتفاعلات مرنة ومتوازنة بين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) (المقاسة في حالة الراحة). من المهم أن الأنماط العصبية المماثلة لوحظت أيضًا أثناء التفكير الإبداعي في تحقق خارجي مع بيانات fMRI المستندة إلى المهام. وبالتالي، يوفر دراستنا اتجاهًا واعدًا بشكل خاص لتوضيح العمليات العصبية والمعرفية الديناميكية التي تحقق الإدراك الإبداعي. باختصار، فإن دراسة الإدراك الإبداعي من إطار ديناميكي تحمل الكثير من الوعد لتحقيق فهم معزز للإبداع وتقديم رؤى جديدة. بشكل حاسم، تبرز دراستنا علامة عصبية عامة للقدرة الإبداعية، مما يساهم في إزالة الغموض عن تعقيد الإدراك الإبداعي.
طرق
مجموعات البيانات
تم تحليل مجموعات البيانات في الدراسة الحالية للتحليل التلوي من خمسة مراكز في النمسا.كنداالصيناليابان، والولايات المتحدة“، مما يشكل مجموعات بيانات متعددة المراكز تشمل التصوير بالرنين المغناطيسي الهيكلي، وتصوير الرنين المغناطيسي الوظيفي في حالة الراحة، وبيانات القدرة الإبداعية (المقيمة من خلال اختبارات التفكير المتباين). كان هناك 10 عينات من أربعة مراكز ومصدر مفتوح واحد، بما في ذلك 3533 مشاركًا صحيًا أكملوا عمليات مسح الرنين المغناطيسي و2772 مشاركًا أكملوا أيضًا تقييم الإبداع. كانت العينة النهائية الصالحة تتكون من 2433 مشاركًا (ذكور )، بمتوسط عمر 21.12 سنة (الانحراف المعياريسنوات)، بعد اجتياز مجموعة من معايير الاستبعاد بما في ذلك فشل تشغيل fMRIprep (بسبب ضعف التطبيع المكاني و/أو جودة الإشارة، وبيانات خام غير مكتملة)، حركة رأس مفرطة، وبيانات سلوكية غير مكتملة أو غير صالحة. بالنسبة لهذه البيانات العشر، تم نشر بيانات fMRI في حالة الراحة (بما في ذلك UNCG و GBB_S1 و GBB_S2 و SLIM_S1 و UG_S3 و TKU) في مكان آخر.تم نشر بيانات fMRI المتعلقة بالمهمة التي تم جمعها، ولكن بيانات fMRI في حالة الراحة التي تم جمعها خلال نفس الفترة لم يتم تحليلها في الأعمال السابقة وهي مدرجة في هذه الدراسة (على سبيل المثال، QU، UG_S1، UG_S2، UG_S3). من المهم أنه بالنسبة لجميع مجموعات البيانات التي تم تقييمها في البحث الحالي، لم تستخدم أي دراسة سابقة التحليلات التي تم إجراؤها في العمل الحالي أو فحصت أنماط التفاعل الديناميكي بين الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) باستخدام بيانات حالة الراحة فيما يتعلق بالأداء الإبداعي الفردي. لمزيد من التفاصيل حول كل عينة، انظر الجدول التكميلي 1. سمحت جميع مجالس المراجعة المؤسسية المحلية باستخدام البيانات التي تم إزالة تعريفها وت anonymized. قدم جميع المشاركين في الدراسة موافقة خطية مستنيرة في مؤسستهم المحلية قبل المشاركة. تم اتباع جميع اللوائح الأخلاقية المتعلقة بالمشاركين في أبحاث البشر.
التقييم السلوكي
في هذه الدراسة، تم تقييم القدرة الإبداعية الفردية من خلال مهام التفكير المتباين. على الرغم من استخدام مهام تفكير متباين مختلفة عبر عشرة مجموعات بيانات، كانت إرشادات ومتطلبات المهمة هي نفسها، بما في ذلك طلب من المشاركين تقديم أكبر عدد ممكن من الاستخدامات المثيرة والجديدة وغير الشائعة لبعض الأشياء في وقت محدود. يتماشى ذلك مع إجراءات التقييم التقليدية.قام 2-4 مقيمين بتقييم استجابات المشاركين في كل مهمة وحصلوا على موثوقية جيدة بين المقيمين، باستثناء عينة TKU التي استخدمت مقيمًا مدربًا واحدًا فقط. بالنسبة لكل نوع من التفكير المتباين المقاس في مجموعة بيانات واحدة، تم الحصول على متوسط الدرجات لكل مقيم، أو تم استخدام نمذجة المتغيرات الكامنة لنمذجة درجة الإبداع العامة. ومن الجدير بالذكر أن طرق التقييم الدقيقة للإبداع اختلفت عبر مراكز مختلفة، حيث شملت مقاييس الأصالة أو مجموع الأصالة والطلاقة (لمزيد من التفاصيل حول القياسات والتقييم، انظر معلومات مجموعة البيانات التكميلية).
اكتساب الصورة ومعالجتها
تم الحصول على البيانات الهيكلية والوظيفية محليًا، راجع معلمات المسح التفصيلية في معلومات مجموعة البيانات التكميلية. تم تحليل جميع الصور باستخدام fMRIPrep 1.4.1rc1 (RRID:SCR_016216).، الذي يعتمد على Nipype 1.2.0 (RRID:SCR_002502) يمكن العثور على وصف كامل لخط أنابيب المعالجة المسبقة في المعلومات التكميلية – “معالجة الصور المسبقة.” بعد المعالجة المسبقة، المشاركون الذين لديهم حركة رأس كبيرة (في الترجمة أوتم استبعاد (في التناوب) من التحليل الإضافي. تم حساب حركة الرأس بعد ICAAROMA حيث تم تحديد مكونات الضوضاء (بما في ذلك الحركة) واستبعادها خلال عمليات fmriprep، وبالتالي فإن القيمة هنا ليست حركة الرأس الخام. تم حساب العيوب الزمنية في كل مجموعة بيانات من خلال حساب الإزاحة الإطارية (FD) و FD.أو DVARSتم تحديدهاتم استبعاد المشاركين الذين تجاوزت نقاط وقت الراحة لديهم 10% من متوسط FD من التحليل الإضافي.
التحليل الوظيفي الديناميكي
بعد المعالجة المسبقة، قمنا أولاً بحساب نسبة الإشارة إلى الضوضاء (SNR) لكل منطقة اهتمام (ROI) من خلال متوسط إشارة الرنين المغناطيسي على مدى الزمن مقسومًا على الانحراف المعياري لسلسلة الزمن، وتلك المناطق التي تحتوي على إشارة غير كافية (أي، SNR الانحراف المعياري أعلى أو أدنى من متوسط المجموعة تم استبعادها. بعد ذلك، سلسلة الزمن المتوسطة تم استخراجها من مناطق الاهتمام (ROIs) المرتبطة بشبكتين من الاهتمام، شبكة الوضع الافتراضي وشبكة التحكم التنفيذي، والتي تم الحصول عليها في مساحة MNI القياسية من 300 قطعة قشرية مع 17 شبكة.. لكل مجموعة بيانات، تم تقدير الدقة الزمنية للاتصال الزمني في بيانات سلسلة الزمن BOLD لمناطق الاهتمام (ROIs) باستخدام نهج ضرب المشتقات الزمنية (MTD) ضمن نافذة زمنية منزلقة مكونة من 14 نقطة زمنية، مما أسفر عن مصفوفة تحتوي على الأوقاتعائدات الاستثمارتم حساب MTD من خلال حساب حاصل الضرب النقطي للمشتق الزمني لسلاسل الزمن الزوجية، والتي كانت موثوقة وقابلة للتكرار عبر مجموعات بيانات متعددة مقارنةً بارتباط بيرسون التقليدي باستخدام نافذة الانزلاق.. بعد ذلك، تم تقدير هيكل المجتمع الزمني المحسوب لكل نافذة زمنية باستخدام خوارزمية لوفيان للموارد من صندوق أدوات الاتصال الدماغيحيث تم تقييم مهمة المجتمع لكل منطقة 500 مرة، وتم تحديد تقسيم بالإجماع باستخدام خوارزمية ضبط دقيقة. استنادًا إلى مهام المجتمع المستندة إلى الزمن، قمنا بتقدير درجة z لقوة داخل الوحدة.لكل منطقة وقدرنا تنوع قوة بين الوحدات (أي، معامل المشاركة؛ ) لكل منطقة داخل كل نافذة زمنية .
باستخدام هذين المقياسين للاتصال (أي، و قمنا بحساب هيستوغرام مشترك لقياسات القوة داخل الوحدة والقوة بين الوحدات (عن طريق جمع حالات كل قيمة من القيمتين ضمن 100 حاوية محددة بالتساوي على طول كل محور) داخل كل نافذة زمنية كملف خرائطي. لاختبار ما إذا كان الملف الخرائطي يتقلب مع مرور الوقت، قمنا بعد ذلك بإجراء تحليل تجميع k-means لتصنيف كل ملف خرائطي على مر الزمن إلى واحدة من حالتين (مجموعتين) باستخدام معلمة مسبقة.التقسيم و100 إعادة تشغيل عشوائية)، والتي أظهرت الدراسات السابقة أنها مثاليةلتحديد الحالة المحددة لملف خرائطي واحد، قمنا بمقارنة المتوسطات الإقليمية. و الدرجات عبر الولايتين باستخدام عينات مستقلة-اختبار. هنا، دولة واحدة ذات مستوى أعلىالنتيجة وأقلتشير النتيجة إلى التعاون والتكامل بين الشبكة الافتراضية للراحة (DMN) والشبكة التنفيذية (ECN) (أي حالة متكاملة، )، والحالة الأخرى تعني الفصل بين الشبكتين (أي، حالة مفصولة، ؛ الشكل التوضيحي الإضافي 3B). بالإشارة إلى الدراسات السابقةيمكننا تحديد أي حالة (أي، حالة مفصولة أو حالة متكاملة) تهيمن على الروابط الوظيفية بين الشبكتين في كل نافذة زمنية. عبر النوافذ الزمنية، تم حساب تردد التبديل الذي يقيس عدد الانتقالات بين الحالات المفصولة والمتكاملة، والذي تم تعريفه على أنه: تردد التبديل; وتم حساب توازن الدولتين من خلال فرق زمن الإقامة في الدولتين مقسومًا على الزمن الكلي، والذي تم تعريفه على أنه: التوازن، حيث هو إجمالي وقت الإقامة لـ و أظهرت الدراسات السابقة أن التبديل المتكرر بين الحالات المنفصلة ديناميكيًا والمندمجة على مدى فترات زمنية طويلة يؤدي إلى التوازن. وبالتالي، يظهر تردد التبديل والتوازن علاقة على شكل حرف U مقلوب (الشكل التكميلي 3C)..
تحليلات داخل مجموعة البيانات
لكل مجموعة بيانات، قمنا أولاً بحساب الارتباط بين الأداء الإبداعي (أي، القدرة على التفكير المتباين) وتكرار التبديل (بين الفصل والتكامل لشبكة DMN-ECN) مع التحكم في الجنس والعمر ومتوسط FD والإشارة العالمية المتوسطة. بالإضافة إلى ذلك، قمنا بحساب هذه العلاقة في كل مجموعة بيانات باستخدام نهج ارتباط قوي.. كانت مسألتنا الثانية تتعلق بما إذا كانت العلاقة بين الأداء الإبداعي وتوازن حالتي الدماغ الديناميتين خطية أم تربيعية. بالنسبة لنماذج الانحدار الخطية والتربيعية، احتوى نموذج الانحدار المتعدد على مؤشر التوازن (أي، خطي) أو مؤشر التوازن وعبارته المربعة (أي، تربيعي) كمتغيرات مستقلة، ودرجة الإبداع الفردية كمتغير تابع، كما يلي:
بعد ذلك، تم استخدام تحليل التباين (ANOVA) ومعيار أكايكي للمعلومات (AIC) لتحديد أي نموذج يناسب البيانات بشكل أفضل إذا كانت النماذج الخطية والتربيعية ذات دلالة استنادًا إلى إحصائية F، حيث تشير قيمة AIC الأصغر إلى ملاءمة أفضل. فرق منفي AIC اعتُبر أنه يعكس فرقًا كبيرًا بين النماذج.
النهج التحليلي الشامل
قمنا بإجراء تحليلات ميتا-تحليل باستخدام الوظائف المتاحة في حزمة metafor (الإصدار 1.9-6)داخل بيئة البرمجيات مفتوحة المصدر R (الإصدار 3.2.0). في تحليلات داخل مجموعة البيانات، -تم حساب قيمة p بين تردد التبديل بين الحالات والأداء الإبداعي لكل مجموعة بيانات. بعد ذلك، تم تحويل قيم r بين المجموعات العشر إلى أحجام تأثير باستخدام دالة “escalc” مع خيار القياس المحدد على “ZCOR”. نظرًا لأن عدة مراكز مشمولة في هذا التحليل قدمت مجموعات بيانات متعددة كمشاريع مختلفة وقياسات للإبداع، استخدمنا أولاً نماذج التأثير العشوائي على مستوى الموقع مع “rma.mv” وظيفة مع خيار الطريقة المحدد إلى “REML” تم تنفيذها في حزمة metafor R. في الوقت نفسه، تم أيضًا ملاءمة نموذج التأثيرات العشوائية باستخدام وظيفة “rma”، التي قدمت أيضًا مخرجات تقليدية لاختبار Q لعدم التجانس. تم إجراء مزيد من التحليلات للوسطاء والمجموعات الفرعية لاستكشاف التأثير المحتمل للوسطاء، بما في ذلك المواقع، بروتوكول التصوير بالرنين المغناطيسي، طرق التقييم، حجم العينة، العمر، ونسبة الجنس.
إعادة الإنتاجية
بالإضافة إلى نهج علوم الأعصاب الشبكية التحليلية الميتا، قمنا أيضًا بإجراء تحليلات ضخمة من خلال تجميع جميع البيانات المتاحة على مستوى الأفراد من جميع مجموعات البيانات.. سمح لنا هذا النهج باستبعاد المتغيرات غير ذات الصلة، مثل العوامل الديموغرافية وعوامل الموقع، والتركيز على التأثيرات المحددة. لتمكين هذا التحليل، تم تحويل جميع أداء الإبداع، وتكرار التبديل، والتوازن الوظيفي بين حالات DMNECN المنفصلة والمندمجة إلى درجات Z ضمن كل مجموعة بيانات. في البداية، في التحليل الضخم، قمنا بفحص العلاقة بين أداء الإبداع وتكرار التبديل بين الانفصال والاندماج داخل DMN-ECN. في هذا التحليل، تم اعتبار تكرار التبديل عبر مجموعات البيانات كعامل عشوائي، بينما تم اعتبار تكرار التبديل والمتغيرات غير ذات الصلة كعوامل ثابتة. بعد ذلك، استكشفنا نموذجين: العلاقات الخطية وغير الخطية بين أداء الإبداع وتوازن الانفصال والاندماج في DMN-ECN. ثم أجرينا اختبار ANOVA وقارنّا قيم AIC بين النماذج الخطية والتربيعية ضمن التحليل الضخم. تم إجراء جميع التحليلات الضخمة باستخدام نموذج التأثيرات المختلطة الخطية (LME)، باستخدام حزمة ‘nlme’ في R (الإصدار 4.2.3). بالإضافة إلى ذلك، أعدنا تحليل البيانات باستخدام أطلس دماغ شين.التي تتكون من 54 منطقة اهتمام داخل الشبكة الافتراضية (DMN) والشبكة التنفيذية (ECN) لتأكيد نتائجنا.
التحقق الخارجي
لقد تم انتقاد استخدام أساليب التصوير بالرنين المغناطيسي الوظيفي في حالة الراحة لاشتقاق تفسيرات معرفية من قبل البعض.في هذا السياق، تقدم أساليب التصوير بالرنين المغناطيسي الوظيفي المرتبطة بالحدث، إلى حد ما، اختبارًا أكثر مباشرة للعمليات العقلية الإبداعية.. لذلك، قمنا بإجراء تحليل تحقق خارجي لتقييم ما إذا كان نمط التبديل الديناميكي بين الشبكتين DMN وECN يمكن التحقق منه في سياق مهمة التفكير المتباين التي تتضمن عمليات إدراكية إبداعية مقارنة بمهمة التحكم التي تتطلب إدراكات شائعة (مثل استرجاع الذاكرة). أكمل واحد وثلاثون مشاركًا كل من مهمة الاستخدامات البديلة (AUT) ومهمة خصائص الكائنات (OCT) أثناء تصوير الرنين المغناطيسي الوظيفي. في التحليل الإحصائي، استخدمنا في البداية عينة مرتبطة من -اختبار لفحص ما إذا كان هناك فرق كبير في تكرار التبديل بين مهام AUT وOCT. بعد ذلك، تم استخدام نموذج خطي مختلط أو غير خطي للتحقيق في العلاقة بين الأداء الإبداعي (أي، أداء AUT) وتكرار التبديل وتوازن حالتي الدماغ الديناميتين.
الإحصائيات وإمكانية التكرار
تفاصيل حجم العينة لكل مجموعة بيانات، بالإضافة إلى التحليلات الإحصائية لكل من بيانات السلوك وبيانات fMRI، موضحة في الأقسام المعنية من “النتائج” و”الطرق”.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
البيانات المعالجة التي تم توليدها في هذه الدراسة متاحة على إطار العلوم المفتوحة علىhttps://osf.io/83arw/، https://doi.org/10.17605/OSF.IO/83ARW. بعض البيانات الخام المستخدمة في الدراسة الحالية كانت متاحة للجمهور وتم تحليلها في دراسات سابقة، وجزء من البيانات غير العامة متاح من المؤلف المراسل عند الطلب المعقول.
تاريخ الاستلام: 11 يوليو 2024؛ تاريخ القبول: 6 يناير 2025؛ نُشر على الإنترنت: 15 يناير 2025
References
Beaty, R. E., Benedek, M., Silvia, P. J. & Schacter, D. L. Creative cognition and brain network dynamics. Trends Cogn. Sci. 20, 87-95 (2016).
Benedek, M., Beaty, R. E., Schacter, D. L. & Kenett, Y. N. The role of memory in creative ideation. Nat. Rev. Psychol. 2, 246-257 (2023).
Mayseless, N., Eran, A. & Shamay-Tsoory, S. G. Generating original ideas: the neural underpinning of originality. Neuroimage 116, 232-239 (2015).
Beaty, R. E. & Kenett, Y. N. Associative thinking at the core of creativity. Trends Cogn. Sci. 27, 671-683 (2023).
Lloyd-Cox, J., Chen, Q. & Beaty, R. E. The time course of creativity: multivariate classification of default and executive network contributions to creative cognition over time. Cortex 156, 90-105(2022).
Bendetowicz, D. et al. Two critical brain networks for generation and combination of remote associations. Brain 141, 217-233 (2018).
Marron, T. R. et al. Chain free association, creativity, and the default mode network. Neuropsychologia 118, 40-58 (2018).
Heinonen, J. et al. Default mode and executive networks areas: association with the serial order in divergent thinking. PLoS ONE 11, e0162234 (2016).
Andrews-Hanna, J. R., Smallwood, J. & Spreng, R. N. The default network and self-generated thought: component processes, dynamic control, and clinical relevance. Ann. N. Y. Acad. Sci. 1316, 29-52 (2014).
Andrews-Hanna, J. R., Reidler, J. S., Huang, C. & Buckner, R. L. Evidence for the default network’s role in spontaneous cognition. J. Neurophysiol. 104, 322-335 (2010).
Ovando-Tellez, M. et al. Brain connectivity-based prediction of reallife creativity is mediated by semantic memory structure. Sci. Adv. 8, eabl4294 (2022).
Marron, T. R., Berant, E., Axelrod, V. & Faust, M. Spontaneous cognition and its relationship to human creativity: a functional connectivity study involving a chain free association task. Neuroimage 220, 117064 (2020).
Shofty, B. et al. The default network is causally linked to creative thinking. Mol. Psychiatry 27, 1848-1854 (2022).
Raffaelli, Q. et al. Creative minds at rest: creative individuals are more associative and engaged with their idle thoughts. Creat. Res. J. 36, 396-412 (2024).
Shamay-Tsoory, S. G., Adler, N., Aharon-Peretz, J., Perry, D. & Mayseless, N. The origins of originality: the neural bases of creative thinking and originality. Neuropsychologia 49, 178-185 (2011).
Wang, M., Hao, N., Ku, Y., Grabner, R. H. & Fink, A. Neural correlates of serial order effect in verbal divergent thinking. Neuropsychologia 99, 92-100 (2017).
Kleinmintz, O. M. et al. Participation of the left inferior frontal gyrus in human originality. Brain Struct. Funct. 223, 329-341 (2018).
Chen, Q. et al. A meta-analysis of the effects of transcranial direct current stimulation on creative thinking. Psychol. Aesthet. Creat. Arts. https://doi.org/10.1037/aca0000704 (2024).
Herault, C. et al. Creative connections: the neural correlates of semantic relatedness are associated with creativity. Commun. Biol. 7, 810 (2024).
Ovando-Tellez, M. et al. An investigation of the cognitive and neural correlates of semantic memory search related to creative ability. Commun. Biol. 5, 604 (2022).
Ellamil, M., Dobson, C., Beeman, M. & Christoff, K. Evaluative and generative modes of thought during the creative process. Neuroimage 59, 1783-1794 (2012).
Wu, X. et al. A meta-analysis of neuroimaging studies on divergent thinking using activation likelihood estimation. Hum. Brain Mapp. 36, 2703-2718 (2015).
Benedek, M. et al. To create or to recall? Neural mechanisms underlying the generation of creative new ideas. Neuroimage 88, 125-133 (2014).
Benedek, M. et al. Creating metaphors: the neural basis of figurative language production. Neuroimage 90, 99-106 (2014).
Matheson, H. E., Kenett, Y. N., Gerver, C. & Beaty, R. E. Representing creative thought: a representational similarity analysis of creative idea generation and evaluation. Neuropsychologia 187, 108587 (2023).
Bartoli, E. et al. Default mode network electrophysiological dynamics and causal role in creative thinking. Brain 147, 3409-3425 (2024).
Beaty, R. E. et al. Robust prediction of individual creative ability from brain functional connectivity. Proc. Natl Acad. Sci. USA 115, 1087-1092 (2018).
Takeuchi, H. et al. The association between resting functional connectivity and creativity. Cereb. Cortex 22, 2921-2929 (2012).
Beaty, R. E. et al. Creativity and the default network: a functional connectivity analysis of the creative brain at rest. Neuropsychologia 64, 92-98 (2014).
Shi, L. et al. Large-scale brain network connectivity underlying creativity in resting-state and task fMRI: cooperation between default network and frontal-parietal network. Biol. Psychol. 135, 102-111 (2018).
Frith, E. et al. Intelligence and creativity share a common cognitive and neural basis. J. Exp. Psychol. Gen. 150, 609-632 (2021).
Patil, A. U. et al. Static and dynamic functional connectivity supports the configuration of brain networks associated with creative cognition. Sci. Rep. 11, 165 (2021).
Ren, Z. et al. Connectome-based predictive modeling of creativity anxiety. Neuroimage 225, 117469 (2021).
Beaty, R. E., Benedek, M., Kaufman, S. B. & Silvia, P. J. Default and executive network coupling supports creative idea production. Sci. Rep. 5, 10964 (2015).
Kleinmintz, O. M., Ivancovsky, T. & Shamay-Tsoory, S. G. The twofold model of creativity: the neural underpinnings of the generation and evaluation of creative ideas. Curr. Opin. Behav. Sci. 27, 131-138 (2019).
Fox, K. C. & Christoff, K. The Oxford Handbook of Spontaneous Thought: Mind-wandering, Creativity, and Dreaming (Oxford University Press, 2018).
Braun, U. et al. Dynamic reconfiguration of frontal brain networks during executive cognition in humans. Proc. Natl Acad. Sci. USA 112, 11678-11683 (2015).
Kenett, Y. N., Chrysikou, E. G., Bassett, D. S. & Thompson-Schill, S. L. Neural dynamics during the generation and evaluation of creative and non-creative ideas. Preprint at bioRxiv https://doi.org/10.1101/ 2024.04.15.589621 (2024).
Christoff, K., Irving, Z. C., Fox, K. C., Spreng, R. N. & AndrewsHanna, J. R. Mind-wandering as spontaneous thought: a dynamic framework. Nat. Rev. Neurosci. 17, 718-731 (2016).
Benedek, M. & Jauk, E. Spontaneous and controlled processes in creative cognition in The Oxford Handbook of Spontaneous Thought: MindWandering, Creativity, Dreaming and Clinical Conditions (eds Christoff, K. & Fox, K. C. R.) 285-298 (Oxford University Press, 2018).
Chrysikou, E. G. Creativity in and out of (cognitive) control. Curr. Opin. Behav. Sci. 27, 94-99 (2019).
Tromp, C. Integrated constraints in creativity: foundations for a unifying model. Rev. Gen. Psychol. 27, 41-61 (2023).
Sun, J. et al. Verbal creativity correlates with the temporal variability of brain networks during the resting state. Cereb. Cortex 29, 1047-1058 (2018).
Sporns, O. Network attributes for segregation and integration in the human brain. Curr. Opin. Neurobiol. 23, 162-171 (2013).
Shine, J. M. et al. The dynamics of functional brain networks: integrated network states during cognitive task performance. Neuron 92, 544-554 (2016).
Orwig, W. et al. Creativity at rest: exploring functional network connectivity of creative experts. Netw. Neurosci. 7, 1022-1033 (2023).
Spreng, R. N., Stevens, W. D., Chamberlain, J. P., Gilmore, A. W. & Schacter, D. L. Default network activity, coupled with the frontoparietal control network, supports goal-directed cognition. Neuroimage 53, 303-317 (2010).
Cole, M. W., Ito, T., Bassett, D. S. & Schultz, D. H. Activity flow over resting-state networks shapes cognitive task activations. Nat. Neurosci. 19, 1718-1726 (2016).
Tromp, C. Creativity from constraint exploration and exploitation. Psychol. Rep. 127, 1818-1843 (2024).
Girn, M., Mills, C., Roseman, L., Carhart-Harris, R. L. & Christoff, K. Updating the dynamic framework of thought: creativity and psychedelics. Neuroimage 213, 116726 (2020).
Beaty, R. E., Seli, P. & Schacter, D. L. Network neuroscience of creative cognition: mapping cognitive mechanisms and individual differences in the creative brain. Curr. Opin. Behav. Sci. 27, 22-30 (2019).
Cohen, J. R. The behavioral and cognitive relevance of time-varying, dynamic changes in functional connectivity. Neuroimage 180, 515-525 (2018).
Wang, R. et al. Segregation, integration, and balance of large-scale resting brain networks configure different cognitive abilities. Proc. Natl Acad. Sci. USA 118, e2022288118 (2021).
Kenett, Y. N., Betzel, R. F. & Beaty, R. E. Community structure of the creative brain at rest. Neuroimage 210, 116578 (2020).
Capouskova, K., Zamora-López, G., Kringelbach, M. L. & Deco, G. Integration and segregation manifolds in the brain ensure cognitive flexibility during tasks and rest. Hum. Brain Mapp. 44, 6349-6363 (2023).
Zhuang, K. et al. Diverse functional interaction driven by controldefault network hubs supports creative thinking. Cereb. Cortex 33, 11206-11224 (2023).
Li, J. et al. High transition frequencies of dynamic functional connectivity states in the creative brain. Sci. Rep. 7, 46072 (2017).
Cohen, J. R. & D’Esposito, M. The segregation and integration of distinct brain networks and their relationship to cognition. J. Neurosci. 36, 12083-12094 (2016).
Xu, J. et al. Anxious brain networks: a coordinate-based activation likelihood estimation meta-analysis of resting-state functional connectivity studies in anxiety. Neurosci. Biobehav. Rev. 96, 21-30 (2019).
Jahani, A. et al. Twinned neuroimaging analysis contributes to improving the classification of young people with autism spectrum disorder. Sci. Rep. 14, 20120 (2024).
Acar, S. & Runco, M. A. Divergent thinking: new methods, recent research, and extended theory. Psychol. Aesthet. Creat. Arts 13, 153-158 (2019).
Karwowski, M. et al. Is creativity without intelligence possible? A necessary condition analysis. Intelligence 57, 105-117 (2016).
Gerwig, A. et al. The relationship between intelligence and divergent thinking-a meta-analytic update. J. Intell. 9, 23 (2021).
Borenstein, M., Hedges, L. V., Higgins, J. P. & Rothstein, H. R. Introduction to Meta-analysis (Wiley, 2021).
Beaty, R. E., Merseal, H. M. & Zeitlen, D. C. in The Routledge International Handbook of Creative Cognition, 433-452 (Routledge, 2023).
Shen, X. et al. Using connectome-based predictive modeling to predict individual behavior from brain connectivity. Nat. Protoc. 12, 506-518 (2017).
Benedek, M. & Fink, A. Toward a neurocognitive framework of creative cognition: the role of memory, attention, and cognitive control. Curr. Opin. Behav. Sci. 27, 116-122 (2019).
Volle, E. Associative and Controlled Cognition in Divergent Thinking: Theoretical, Experimental, Neuroimaging Evidence, and New Directions, 333-362 (Cambridge University Press, 2018).
Beaty, R. E., Silvia, P. J. & Benedek, M. Brain networks underlying novel metaphor production. Brain Cogn. 111, 163-170 (2017).
Da Mota, P. A. et al. The dynamics of the improvising brain: a study of musical creativity using jazz improvisation. Preprint at bioRxiv https://doi.org/10.1101/2020.01.29.924415 (2020).
Limb, C. J. & Braun, A. R. Neural substrates of spontaneous musical performance: an FMRI study of jazz improvisation. PLoS ONE 3, e1679 (2008).
Liu, S. Y. et al. Brain activity and connectivity during poetry composition: toward a multidimensional model of the creative process. Hum. Brain Mapp. 36, 3351-3372 (2015).
Matheson, H. E. & Kenett, Y. N. The role of the motor system in generating creative thoughts. Neuroimage 213, 116697 (2020).
Chen, Q. et al. Brain hemispheric involvement in visuospatial and verbal divergent thinking. Neuroimage 202, 116065 (2019).
He, R. et al. The impact of knowledge on poetry composition: an fMRI investigation. Brain Lang. 235, 105202 (2022).
Bilder, R. M. & Knudsen, K. S. Creative cognition and systems biology on the edge of chaos. Front. Psychol. 5, 112333 (2014).
Simonton, D. K. Creative thought as blind-variation and selectiveretention: combinatorial models of exceptional creativity. Phys. Life Rev. 7, 156-179 (2010).
Boot, N., Baas, M., van Gaal, S., Cools, R. & De Dreu, C. K. W. Creative cognition and dopaminergic modulation of fronto-striatal networks: integrative review and research agenda. Neurosci. Biobehav. Rev. 78, 13-23 (2017).
Amabile, T. Componential Theory of Creativity (Harvard Business School, 2011).
Ding, K., He, R., Wang, X., Chen, Q. & Kenett, Y. N. Recognizing ideas generated in a creative task: the roles of the hippocampus and medial prefrontal cortex in facilitating self-generated learning. Cereb. Cortex 34, bhae219 (2024).
Wang, X. et al. Semantic associative abilities and executive control functions predict novelty and appropriateness of idea generation. Commun. Biol. 7, 703 (2024).
Kenett, Y. N. et al. Developing a neurally informed ontology of creativity measurement. Neuroimage 221, 117166 (2020).
Gerver, C. R., Griffin, J. W., Dennis, N. A. & Beaty, R. E. Memory and creativity: a meta-analytic examination of the relationship between memory systems and creative cognition. Psychon. Bull. Rev. 30, 2116-2154 (2023).
Green, A. E., Beaty, R. E., Kenett, Y. N. & Kaufman, J. C. The process definition of creativity. Creat. Res. J. 36, 544-572 (2024).
Japardi, K., Bookheimer, S., Knudsen, K., Ghahremani, D. G. & Bilder, R. M. Functional magnetic resonance imaging of divergent and convergent thinking in Big-C creativity. Neuropsychologia 118, 59-67 (2018).
Kam, J. W. et al. Default network and frontoparietal control network theta connectivity supports internal attention. Nat. Hum. Behav. 3, 1263-1270 (2019).
Zanesco, A. P., Denkova, E. & Jha, A. P. Self-reported mind wandering and response time variability differentiate prestimulus electroencephalogram microstate dynamics during a sustained attention task. J. Cogn. Neurosci. 33, 28-45 (2021).
Liu, S., Abdellaoui, A., Verweij, K. J. & van Wingen, G. A. Replicable brain-phenotype associations require large-scale neuroimaging data. Nat. Hum. Behav. 7, 1344-1356 (2023).
Marek, S. et al. Reproducible brain-wide association studies require thousands of individuals. Nature 603, 654-660 (2022).
Gratton, C., Nelson, S. M. & Gordon, E. M. Brain-behavior correlations: two paths toward reliability. Neuron 110, 1446-1449 (2022).
Kong, X.-Z., Zhang, C., Liu, Y. & Pu, Y. Scanning reproducible brainwide associations: sample size is all you need? Psychoradiology 2, 67-68 (2022).
Rosenberg, M. D. & Finn, E. S. How to establish robust brain-behavior relationships without thousands of individuals. Nat. Neurosci. 25, 835-837 (2022).
Spisak, T., Bingel, U. & Wager, T. D. Multivariate BWAS can be replicable with moderate sample sizes. Nature 615, E4-E7 (2023).
Helwegen, K., Libedinsky, I. & van den Heuvel, M. P. Statistical power in network neuroscience. Trends Cogn. Sci. 27, 282-301 (2023).
Bryce, N. V. et al. Brain parcellation selection: an overlooked decision point with meaningful effects on individual differences in resting-state functional connectivity. Neuroimage 243, 118487 (2021).
Messé, A. Parcellation influence on the connectivity-based structure-function relationship in the human brain. Hum. Brain Mapp. 41, 1167-1180 (2020).
Kong, R. Q. et al. A network correspondence toolbox for quantitative evaluation of novel neuroimaging results. Preprint at bioRxiv https:// doi.org/10.1101/2024.06.17.599426 (2024).
Beaty, R. E. et al. Personality and complex brain networks: the role of openness to experience in default network efficiency. Hum. Brain Mapp. 37, 773-779 (2016).
Jauk, E., Benedek, M., Dunst, B. & Neubauer, A. C. The relationship between intelligence and creativity: new support for the threshold hypothesis by means of empirical breakpoint detection. Intelligence 41, 212-221 (2013).
Jauk, E., Neubauer, A. C., Dunst, B., Fink, A. & Benedek, M. Gray matter correlates of creative potential: a latent variable voxel-based morphometry study. Neuroimage 111, 312-320 (2015).
Fink, A. et al. Modulation of resting-state network connectivity by verbal divergent thinking training. Brain Cogn. 128, 1-6 (2018).
Poppenk, J. Multimodal brain data and core dimensions of creativity. Data Brief 29, 105176 (2020).
Chen, Q. et al. Longitudinal alterations of frontoparietal and frontotemporal networks predict future creative cognitive ability. Cereb. Cortex 28, 103-115 (2018).
Liu, Z. et al. Neural and genetic determinants of creativity. Neuroimage 174, 164-176 (2018).
Takeuchi, H. et al. Originality of divergent thinking is associated with working memory-related brain activity: evidence from a large sample study. Neuroimage 216, 116825 (2020).
Orwig, W., Diez, I., Vannini, P., Beaty, R. & Sepulcre, J. Creative connections: computational semantic distance captures individual creativity and resting-state functional connectivity. J. Cogn. Neurosci. 33, 499-509 (2021).
Silvia, P. J. et al. Assessing creativity with divergent thinking tasks: exploring the reliability and validity of new subjective scoring methods. Psychol. Aesthet. Creat. Arts 2, 68-85 (2008).
Esteban, O. et al. fMRIPrep: a robust preprocessing pipeline for functional MRI. Nat. Methods 16, 111-116 (2019).
Gorgolewski, K. et al. Nipype: a flexible, lightweight and extensible neuroimaging data processing framework in Python. Front. Neuroinform. 5, 13 (2011).
Patel, A. X. et al. A wavelet method for modeling and despiking motion artifacts from resting-state fMRI time series. Neuroimage 95, 287-304 (2014).
Drysdale, A. T. et al. Resting-state connectivity biomarkers define neurophysiological subtypes of depression. Nat. Med. 23, 28-38 (2017).
Schaefer, A. et al. Local-global parcellation of the human cerebral cortex from intrinsic functional connectivity MRI. Cereb. Cortex 28, 3095-3114 (2018).
Rubinov, M. & Sporns, O. Complex network measures of brain connectivity: uses and interpretations. Neuroimage 52, 1059-1069 (2010).
Guimera, R. & Amaral, L. A. N. Functional cartography of complex metabolic networks. Nature 433, 895-900 (2005).
Pernet, C. R., Wilcox, R. & Rousselet, G. A. Robust correlation analyses: false positive and power validation using a new open source Matlab toolbox. Front. Psychol. 3, 606 (2013).
Viechtbauer, W. Conducting meta-analyses in with the metafor package. J. Stat. Softw. 36, 1-48 (2010).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation, 2013).
Costafreda, S. G. Pooling FMRI data: meta-analysis, mega-analysis and multi-center studies. Front. Neuroinform. 3, 33 (2009).
Davis, S. W., Stanley, M. L., Moscovitch, M. & Cabeza, R. Restingstate networks do not determine cognitive function networks: a commentary on Campbell and Schacter (2016). Lang. Cogn. Neurosci. 32, 669-673 (2017).
Finn, E. S. Is it time to put rest to rest? Trends Cogn. Sci. 25, 1021-1032 (2021).
شكر وتقدير
تم دعم Q.C. من قبل المؤسسة الوطنية للعلوم الطبيعية في الصين (NSFC31800919، NSFC32071070). تم دعم R.E.B. من خلال منح من المؤسسة الوطنية للعلوم الأمريكية [DRL-1920653؛ DUE-2155070]. تم دعم هذا العمل جزئيًا من قبل صندوق العلوم الثنائية الأمريكي-الإسرائيلي (BSF) منحة (رقم 2021040) لـ R.E.B. و Y.N.K.
مساهمات المؤلفين
قام Q.C. وY.N.K. وR.E.B. بتصميم الدراسة؛ ساهم H.T. وA.F. وM.B. وR.K. وJ.Q. وR.E.B. في جمع البيانات؛ قام Q.C. وY.N.K. وZ.C. وK.Z. وR.E.B. بإجراء التحليلات؛ كتب Q.C. وY.N.K. وR.E.B. المسودة الأصلية؛ قام Z.C. وM.B. وD.C.Z. وK.Z. وJ.L.C. وJ.Q. وR.E.B. بمراجعة وتحرير النص. قدم جميع المؤلفين المشاركين ملاحظات ووافقوا على المخطوطة.
كلية علم النفس، جامعة جنوب غرب، تشونغتشينغ، الصين. قسم علم النفس، جامعة ولاية بنسلفانيا، حديقة الجامعة، بنسلفانيا، الولايات المتحدة الأمريكية. كلية علوم البيانات وعلوم القرار، معهد التخنيون-معهد التكنولوجيا الإسرائيلي، حيفا، إسرائيل. المعهد الصيني لأبحاث الدماغ، بكين، الصين. قسم علوم الأعصاب المعرفية التنموية، معهد التنمية، الشيخوخة والسرطان، جامعة توهوكو، سنداي، اليابان. قسم علم النفس، جامعة غراتس، غراتس، النمسا. معهد العلوم والتكنولوجيا للذكاء المستوحى من الدماغ، جامعة فودان، شنغهاي، الصين. قسم علم النفس، غولدسميث، جامعة لندن، لندن، المملكة المتحدة. ساهم هؤلاء المؤلفون بالتساوي: كونلين تشين، يود ن. كينيت. □ البريد الإلكتروني: yoedk@technion.ac.il; qiuj318@swu.edu.cn
Dynamic switching between brain networks predicts creative ability
Qunlin Chen , Yoed N. Kenett , Zaixu Cui (), Hikaru Takeuchi , Andreas Fink , Mathias Benedek , Daniel C. Zeitlen , Kaixiang Zhuang , James Lloyd-Cox , Ryuta Kawashima , Jiang Qiu & Roger E. Beaty
Creativity is hypothesized to arise from a mental state which balances spontaneous thought and cognitive control, corresponding to functional connectivity between the brain’s Default Mode (DMN) and Executive Control (ECN) Networks. Here, we conduct a large-scale, multi-center examination of this hypothesis. Employing a meta-analytic network neuroscience approach, we analyze resting-state fMRI and creative task performance across 10 independent samples from Austria, Canada, China, Japan, and the United States -constituting the largest and most ethnically diverse creativity neuroscience study to date. Using time-resolved network analysis, we investigate the relationship between creativity (i.e., divergent thinking ability) and dynamic switching between DMN and ECN. We find that creativity, but not general intelligence, can be reliably predicted by the number of DMN-ECN switches. Importantly, we identify an inverted-U relationship between creativity and the degree of balance between DMN-ECN switching, suggesting that optimal creative performance requires balanced brain network dynamics. Furthermore, an independent task-fMRI validation study ( ) demonstrates higher DMN-ECN switching during creative idea generation (compared to a control condition) and replicates the inverted-U relationship. Therefore, we provide robust evidence across multi-center datasets that creativity is tied to the capacity to dynamically switch between brain networks supporting spontaneous and controlled cognition.
Creative thinking is a critical human capacity, enabling the progress of civilization through continuous innovation in diverse scientific and artistic domains. In recent decades, behavioral and neuroimaging studies have yielded a considerable understanding of the neural and cognitive mechanisms underlying how people generate novel and useful (i.e., creative) ideas . Growing evidence suggests that creative thought emerges from the interaction between spontaneous associations and deliberate, cognitive control processes, primarily driven by interactions between the Default Mode Network (DMN) and various other networks . In particular, greater creative thinking has been linked to the coupling of the DMN and the Executive Control Network (ECN).
The DMN consists of cortical midline (including the medial prefrontal cortex and posterior cingulate cortex), posterior inferior parietal regions,
and the medial temporal lobe; this brain network is known to involve internally directed or self-generated thought, such as the spontaneous activation of memories, mind wandering, semantic integration, associative idle thoughts, and mental simulation-cognitive processes thought to support creative idea generation . The ECN consists of lateral prefrontal and anterior inferior parietal regions, and when coupled with the DMN, supports controlled semantic retrieval and cognitive flexibility, which benefits the selection and evaluation of creative ideas . Numerous neuroimaging studies have consistently found that creative thinking recruits regions within the DMN and . In addition, an individual creative thinking ability can be predicted by functional interactions within and between the DMN and ECN both at rest and during creative task performance using static networks methods . For example, functional
connectivity features associated with the default mode, executive control, and salience networks can predict individual creative performance during divergent thinking tasks, and these features can be generalized across several independent resting-state fMRI samples . Given the roles of the DMN and ECN in spontaneous and controlled cognition, respectively, DMN-ECN connectivity is thought to reflect a coordination between spontaneous/ generative and controlled/evaluative processes . However, static network approaches fail to capture the dynamic coordination of cognitive states related to reconfiguration and interactions between networks . Therefore, dynamic reconfiguration-based functional analyses are needed to identify specific interaction patterns between the DMN and ECN related to creative ability.
How does the creative brain achieve dynamic coordination between spontaneity and control? According to the dynamic framework of spontaneous thought , creativity is a mental state that requires an optimal balance between spontaneous thought generation and goal-directed processing -corresponding to a balanced state of DMN and ECN communication. Dual-process theories of creativity also emphasize that creative cognition involves two different phases-idea generation and evaluation-dominated by cooperation between large-scale functional brain networks including the DMN and ECN . Therefore, one possibility is that higher creative people can flexibly switch between segregation and integration of DMN and ECN-oscillating between states where the brain networks operate independently and interdependently, respectively-which may reflect an optimal balance or “sweet spot” between the networks that leverage their independent and collective capacities. Despite the intuitive appeal of this theory, very little empirical evidence to support this claim exists. Several studies based on stages of creative processing have pointed out that the DMN predominantly acts on the generative phase in the early stages of creative production, whereas increased coupling between the DMN and ECN underlies the evaluative phase in the later stage , implying that the ECN may monitor and guide spontaneous associative/generative processes . Additionally, the temporal connectivity of large-scale brain networks based on resting-state fMRI data supports the dynamic functional configuration associated with creative cognition . However, the precise interaction patterns and the mechanistic role of this DMN-ECN coupling in creative thinking remain unclear.
This dynamic reconfiguration of brain network connectivity can be divided into two distinct patterns or “states” that the brain routinely occupies: segregation and integration. The segregated pattern corresponds to independent processing within networks, whereas the integrated pattern corresponds to cooperation between networks . Previous research has reported the importance of brain network segregation and integration for performance on diverse cognitive tasks , although no prior work has examined such brain network dynamics in relation to creativity. Several studies have, however, observed variations in temporal cooperation between functional networks during creative cognition, suggesting the existence of a possible dynamic transition between functional segregation and integration . From the perspective of individual differences, highly creative individuals are more likely to engage in internally directed cognition during rest, which is characterized by intrinsic functional networks and their interactions . Given that resting-state functional networks shape cognitive task activations , individuals with higher creative thinking abilities —potentially conducive to a more efficient cycle of generation and eva-luation-may also exhibit altered dynamics of large-scale brain networks (namely, the DMN and ECN) at rest . Studies have consistently shown that intrinsic network dynamics during rest reflect fundamental organizational properties that predict individual differences in cognitive abilities . The pattern and frequency of network switching between segregated and integrated states appear particularly important, as it indicates the brain’s capacity to flexibly coordinate different cognitive processes . Examining these dynamics during rest allows us to characterize stable individual differences in network organization across large samples, complementing and extending insights from task-based studies of creative cognition. This approach is especially valuable given evidence that
creative ability relates to intrinsic patterns of brain network interaction that persist across both rest and task states. Consistent with this view, dynamic connectivity studies have reported that the temporal variability of restingstate functional connectivity among regions within DMN and ECN was positively related to creative thinking ability . Further, a recent study provided a more nuanced understanding of the neurocognitive mechanisms of the creative brain, indicating that generative and evaluative stages may involve different combinations of associative and controlled processes, and thus recruit different proportions of DMN and ECN activity .
Independently from creative thinking, more flexible brain network reconfiguration (that is, highly frequent brain state switching) may reflect a functional balance between segregated and integrated states that helps to foster cognitive abilities such as memory retrieval . For example, a recent investigation reported that while stronger whole-brain segregation supports crystallized intelligence and processing speed and integration fosters better general cognitive ability, a balance between segregation and integration is associated with better memory . By contrast, hyperconnectivity (i.e., increased positive connectivity) between the DMN and the ECN, or other networks, is associated with rigid behaviors and thoughts linked to certain psychiatric disorders (e.g., autism spectrum disorder), while hypo-connectivity (i.e., decreased positive connectivity) is associated with impairments in executive function and social cognition . Returning to the present study, no research to date has investigated dynamic interactions between the DMN and ECN in creative thinking regarding which patterns of switching between segregated and integrated states can benefit individual creativity.
In the present study, we investigate whether individual differences in creative ability can be reliably predicted by the dynamic interaction between the DMN and ECN. To this end, we analyze large-scale, multi-center datasets including structural MRI, resting-state fMRI, and creative ability data (assessed by divergent thinking tests administered outside the scanner) from five centers in Austria, Canada, China, Japan, and the United States ( )-constituting (to our knowledge) the largest and most ethnically diverse brain-based analysis of creativity to date. We applied a meta-analytic network neuroscience approach to integrate the findings by conducting a new analysis using the raw data from each dataset. Here, two dichotomous neural states, segregated and integrated functional connectivity patterns between the DMN and ECN, were determined using time-resolved analyses of resting-state fMRI data. Building on previous research , we hypothesized that a higher frequency of switching between DMN-ECN segregation and integration over time will predict better creative performance. We also hypothesized that higher creative performance may benefit from a functional balance between segregation and integration in these brain networks. Finally, to provide a more direct validation of the neural coupling dynamics, we conducted an external validation analysis using task-based fMRI data in an independent sample of Chinese adults and tested whether the brain network dynamics assessed during rest in the full sample extend to active performance during a creative thinking task.
Results
Sample composition and behavioral findings
Ten datasets spanning five centers were analyzed, including data from 2433 healthy individuals aged from 16 to 58 . Figure 1 A summarizes the sample sizes and age ranges of each dataset (Supplementary Table 1) and Fig. 1B describes the data distribution of creative performance of each dataset. Individual creative performance was assessed through the widely used Alternate Uses Task , where participants generate creative uses to objects and their responses were scored by trained 2-4 raters (see Supplementary Dataset Information for more detail). Behavioral analyses (Supplementary Table 2) showed no consistent gender differences across the 10 datasets, which is in line with previous literature. No significant correlation was found between creativity scores and age except for one dataset (UG_S1). Thus, the absence of consistent findings across diverse datasets suggests that gender differences in creativity scores and the associations between creative performance and age lack robustness due to sample heterogeneity and the very
Fig. 1 | Datasets characteristics. A The total number of participants and the age ranges of each dataset. The position of the black dot on each line indicates the median age of that dataset. B The distribution of creative performance based on human-rated originality scores on divergent thinking tasks in each dataset.
positively skewed distribution in age. Among six datasets containing both intelligence and creativity scores, three datasets showed a significant correlation between the two constructs: UNCG, CI ; UG_S1, CI ; UG_S3, CI ; SLIM_S1, CI [-0.05 to 0.19]; GBB_S1, CI [ -0.04 to 0.30 ]; and GBB_S2, to 0.12, consistent with past work demonstrating positive associations between intelligence and creativity .
The relationship between network switching frequency and creative performance
Next, we turned to test our primary hypothesis regarding whether creative thinking benefits from a tendency to dynamically switch between segregation and integration of the DMN and ECN. The switching frequency was calculated by the number of transitions between segregated and integrated states across the time windows (see “Methods”). Pearson correlation analyses confirmed our hypothesis across 7 of the 10 datasets (Fig. 2): the switching frequency of DMN-ECN segregation and integration was significantly and positively associated with creative performance. Next, we applied a meta-analytic approach to calculate the overall weighted effect size between DMN-ECN state switching frequency and creative ability across the ten datasets. The overall effect size was significant applying random effects at the dataset level, (Fig. 3). The Q test for heterogeneity was significant, , , indicating the necessity for exploring potential moderators of these effects .
Considering that the three moderating variables-location (corresponding to five centers), scanner type, and scoring methods are almost confounded, we conducted a moderator analysis that included all three variables as moderators together. Additionally, following the subgroup analyses, we calculated the pure effect of each group after excluding other variables, such as location and scanner type. The following moderator analyses revealed location, scanner type, and scoring methods, , significantly and jointly moderated the overall effect. Subsequently, we performed subgroup analyses within center, scanner type, and scoring methods for creativity in which the number of effects ( k ) is at least 3: Austria datasets ( ) have a significant and medium positive effect, , and China datasets ( ) have a relatively weak and significant positive
effect, . Datasets with the magnetome MRI scanner ( ) have a significant and medium positive effect, , whereas datasets with the Trio MRI ( ) have a significant and weak positive effect, CI . Datasets with scoring using originality alone ( ) have a significant and medium positive effect, , and datasets with scoring using the sum of originality and fluency ( ) have a weak but significant effect, . In addition, sample size, , age, , scanning time, , and gender ratio of sample, , did not significantly moderate the overall effect, indicating that data origin (including the locations and MRI and scoring protocol) but not participant-related variables influenced the overall effect. Thus, more creative individuals tend to switch more frequently between segregation and integration of the DMN and ECN at rest.
Furthermore, the results remained statistically significant when using the robust correlation method, which does not rely on assumptions of normality or linearity and can handle outliers better than traditional correlation methods. Moreover, the overall effect size was found to be significant, CI [ ], , using random effects model (Supplementary Fig. 1). The Q test for heterogeneity was not significant here, , indicating relatively low variability among the studies.
The relationship between switching frequency and intelligence
Given the conditional relationship between creativity and intelligence, we applied a meta-analytic approach to determine whether intelligence, regarded as a control task, also relates to segregation and integration of DMN and ECN, as we expected (and found) for creativity. Thus, the same analyses were performed among 6 datasets ( , see Supplementary Table 3) which included intelligence scores in addition to creativity scores. The overall effect size was not significant, , with no significant heterogeneity, , . In contrast, the overall effect size for the relationship between switching frequency and creative performance was significant among the same 6 datasets, , . The parameter estimate difference between the two models (Intelligence v.s. Creativity) is significant, , indicating
Fig. 2 | Associations between creative performance and the switching frequency of DMN-ECN segregation and integration. The (robust) indicated the value of robust correlation in each dataset. The fitting regression line is shown in black, and
the confidence interval for the line is shown in gray shading. Both the (creative performance) and (creative performance) axes are standardized value after adjusting for gender, age, mean framewise displacement, and global mean signal.
that the DMN-ECN switching effect is specific to creative ability and does not extend to intelligence.
Creative performance and the balance of DMN-ECN segregation and integration
Previous empirical evidence has shown that individuals with various degrees of functional balance between network segregation and integration demonstrated differential cognitive ability . Furthermore, frequent switching between segregated and integrated states results in a functional balance, and optimal balance (i.e., when the magnitude of balance is equal to zero) might reflect maximum flexibility in dynamic transitions between the two states . Here, we aimed to test whether there is a nonlinear relationship between creative performance and DMN-ECN segregation and integration balance; we therefore performed an ANOVA test and compared AIC values between the linear and quadratic regression model in each dataset.
We found that the quadratic regression model was significantly better fitting than the linear regression model (Fig. 4) in three datasets: UG_S1 , TKU , and SLIM_S2 . There was no significant difference between the two models in the remaining seven datasets (Supplementary Table 4).
To summarize the effect sizes of the quadratic regression model across all datasets, the partial correlation coefficients of the quadratic term in each regression model were meta-analyzed using a random-effects model with restricted maximum likelihood. The overall effect size was small but
Datesets
Location
Weight r -value [95%CI]
UNCG
USA
口
6.92%
0.26 [ 0.10, 0.41]
UG_S1
Austria
5.25%
0.26 [ 0.03, 0.48]
UG_S2
Austria
4.24%
0.28 [ 0.02, 0.54]
UG_S3
Austria
⟵ •
3.75%
0.44 [ 0.16, 0.72]
TKU
Japan
–
11.36%
0.09 [ 0.03, 0.15]
SLIM_S1
China
–
18.14%
-0.02 [-0.14, 0.10]
SLIM_S2
China
12.58%
0.24 [ 0.09, 0.40]
GBB_S1
China
16.72%
0.09 [-0.04, 0.22]
GBB_S2
China
17.21%
0.16 [ 0.04, 0.29]
QU
Canada
3.81%
0.22 [-0.03, 0.48]
Two-level RE model for all datasets:
100.00%
0.17 [ 0.08, 0.27]
Fig. 3 | Forest plots of ten effect sizes included in the meta-analysis. Forest plot of effect sizes for the correlation between creative performance and switching frequency (calculated using resting-state fMRI data). On the left side, forest plots display the name and location of each dataset; on the left of dashed vertical line (representing no effect), the forest plots provide the weight, the observed effect, and the confidence interval, in which the size of a square is proportional to the weights determined by the inverse of datasets’ variance; the bottom line shows the overall effect size (right) and heterogeneity test (left).
significant, , indicating that moderate cooperation-neither more nor less-between the DMN and ECN may be conducive to generating creative ideas. This finding may suggest that across multi-center datasets, creative ability benefits from an
Fig. 4 | Associations between creative performance and the balance of DMN-ECN segregation and integration. Linear (black fitted line) and quadratic relationship (blue fitted line) of creative performance ( -axis) and DMN-ECN segregation and integration balance ( -axis), and with the comparison between linear fitted model
and quadratic fitted model for each dataset (calculated using resting-state fMRI data). Both the and axes are standardized value after adjusting for gender, age, mean framewise displacement, and global mean signal.
optimal balance between spontaneous and controlled processes, supported by moderate switching between DMN-ECN segregation and integration. The Q-test statistic, , suggests that little heterogeneity exists.
Reproducibility
Importantly, we evaluated the reproducibility of our findings using megaanalyses, an approach that pools all available individual-level data from various datasets. This method allows us to exclude non-relevant covariates, such as demographic (such as gender and age) and location factors, and focus on specific effects within a single model. The results of the megaanalysis are summarized in Supplementary Table 5, indicating that individuals with higher switching frequency of DMN-ECN segregation and integration exhibited higher creative performance, , (Fig. 5A). By contrasting the linear and nonlinear models using mega-analysis, the results showed that balance of dynamic switching between DMN-ECN segregation and integration, as the quadratic terms, can significantly predict the creative performance, (Fig. 5B), and the nonlinear model (AIC = 6925.45, Supplementary Table 6) is better than the linear model , Supplementary Table 7) that only includes the linear term (the balance of dynamic brain states) as a fixed factor, . Overall, the mega-analysis yielded results similar to those of the meta-analytic approach. Reanalyzing data using the Shen brain atlas , which consists of 54 regions of interest within DMN and
ECN also confirmed our results: the overall effect size was significant using the random-effects model, (Supplementary Fig. 2).
External validation using task-based fMRI data
We identified dynamic interactive patterns between the DMN and ECN associated with individual creative performance in a large-scale, multicenter, resting-state fMRI dataset. However, can these patterns be validated in the context of a divergent thinking task, which involves creative cognitive processes, as opposed to a control task that only requires common cognitions (e.g., memory retrieval)? To address this, we conducted an external validation analysis using task fMRI data, with thirty-one participants completing both the Alternate Uses Task (AUT), requiring production of creative object uses, and the Object Characteristics Task (OCT), requiring production of common object characteristics, during fMRI scanning. The switching frequency and the degree of balance between two states (i.e., DMN-ECN segregated or integrated state) were derived from task-relevant events. Specifically, we extracted these two dynamic indices by precisely pinpointing AUT and OCT trials when subjects initiated thinking, generating up to four ideations (Fig. 6A). The two indices for each task were then computed for each participant by calculating the switch frequency and balance of DMN-ECN segregation and integration in each run.
The results revealed that AUT ( ; Fig. 6B) recruited a significantly higher switching frequency than OCT ( ; Fig. 6D), , Cohen’s (Fig. 6C). This suggests that creative
Fig. 5 | Mixed effect models. A The linear relationship between creative performance and switching frequency between segregated and integrated DMN-ECN states (calculated using resting-state fMRI data); the black line marks the overall trend, and the confidence interval for the line is shown in gray shading. B The quadratic relationship between creativity score and functional balance between segregated and integrated DMN-ECN states (calculated using resting-state fMRI data); the black line represents the overall trend, with the confidence interval indicated by the gray shading, while the colored lines depict trends for each center. The left side of the -axis indicates extreme segregation, the right side indicates extreme integration, and the dotted line in the center represents the optimal balance or “sweet spot”.
A
The degree of balance between two states
thinking may be supported by interactions between the DMN and ECN compared to general cognitive processes, such as extracting concepts from existing knowledge. A linear mixed-effects model did not show that AUT performance (originality score of ideas) was significantly associated with the switching frequency of DMN-ECN segregation and integration during creative thinking, , Cohen’s (Fig. 6E); similarly, OCT performance was not related to the switching frequency, (Fig. 6F). However, a nonlinear mixed-effects model analyses showed that the degree of balance between DMN-ECN segregation and integration is related to AUT performance, , and the nonlinear model AIC is better than the linear model AIC , that only includes the linear term (the balance of dynamic DMN-ECN states) as a fixed factor, , (Fig. 6G). But the balance of dynamic DMN-ECN switching is not related to OCT performance in both the nonlinear model, (54) , and the linear model, , (Fig. 6H).
Thus, the task-based fMRI findings validate the resting-state results: creative thinking elicits greater dynamic switching and more balanced interactions between default mode and executive control networks compared to non-creative cognition, thereby supporting individual creative performance.
Discussion
Our meta-analytic analyses revealed that creative performance can be reliably predicted from the frequency of switches between the segregated and integrated states of the DMN and ECN at rest: people who switch between these states more often produce more original ideas. Furthermore, high creative thinking ability was associated with a balance between the segregated and integrated states because frequent transitions between these states provided a means to maintain this balance. The convergence between these resting-state patterns across a large sample and their selective emergence during creative task performance (compared to control tasks) suggests these network dynamics represent a specific marker of creative ability. These findings advance our understanding of the neural mechanisms of creativity by identifying specific patterns of network interaction that characterize both the capacity for and active engagement in creative thought.
While much progress has been made, our understanding of the neural bases of creativity remains far from complete. Creativity is a complex construct that recruits both creativity-specific and ordinary cognitive processes such as memory, attention, and cognitive control . A dynamic network-based approach is particularly well-suited to address the complex interplay of neural mechanisms that engage the whole brain during creative thinking . In this perspective, the present study focused on the entire ECN
and DMN in creative cognition, while disregarding differences between specific regions and subnetworks within these large brain networks. In addition, we expected to find domain-general patterns of interactive DMNECN connectivity related to creative cognition. Indeed, in the present research, we found no significant correlation between intelligence and DMN-ECN switching frequency across six datasets. Additionally, we observed a significantly lower DMN-ECN switching frequency under the control (memory retrieval) task compared to the divergent thinking task. This observation may suggest that these dynamic interactions between the DMN and ECN characterize creative cognition in particular, rather than other cognitive abilities.
A body of studies have shown that creative thinking is generally dependent on the capacity of the DMN to synchronize with other brain networks , such as the , salience network , sensorimotor network , and visual network . While coupling between the DMN and ECN has been consistently highlighted as a marker of creativity, such a coupling has been largely shown in “static” functional connectivity research and the role of a specific coupling dynamic as a driver of creativity has so far been largely speculative by analyzing the whole-brain functional connectivity . However, few studies have focused on examining the interactions within these networks and how they influence creative cognition . Although our research relies on methods involving brain-behavioral phenotype associations, by combining evidence from both resting-state networks with large sample size and cognitive functional networks (i.e., task-based fMRI), our findings further support the idea that functional coupling among regions in the DMN and ECN may reflect a domain-general mechanism of creative cognition. Importantly, a new dynamic interaction pattern between the DMN and ECN was observed in this study, namely that higher creative individuals exhibit optimal neural cooperation between the DMN and ECN. Specifically, the degree of balance between the segregated and integrated states demonstrated an inverted U-shaped relationship with creative performance, where too much (or too little) DMN-ECN coupling may impair creative performance. This finding aligns with previous creativity theories, proposing that creative ideation emerges from the delicate balance between flexibility and stability of cognitive states . This equilibrium is crucial for maintaining the optimal combination of novelty and utility throughout the process of creative cognition .
Previous research suggested that the DMN may be centrally involved in active recollection of memories and associative processes that yield diverse ideas, whereas the ECN may act to guide associative processes through monitoring, inhibition, and related top-down control processes . These studies explain the roles of the DMN, ECN, and other networks in creative cognition in two stages: generation and evaluation . Previous creativity theories have suggested that
Fig. 6 | fMRI task design, main analysis, and results. A Experimental paradigm of the divergent thinking task, in which participants had to complete an Alternate Uses Task (AUT) and an Object Characteristics Task (OCT) during fMRI scanning. The task paradigm consisted of a fixation cross ( 1 s ), a cue indicating the upcoming condition (“novel uses” or “common characteristics”; 4 s ), a thinking period presenting an object cue in text ( 60 s for AUT; 30 s for OCT). Participants responded by pressing a key and speaking their responses out loud into an MRI-compatible microphone until they either generated four ideas or ran out of thinking time. This was followed by a rest period of at least 6 s , or the remaining time from the thinking phase if applicable. Task fMRI data were preprocessed using fmriprep, and then time courses were extracted from 106 ROIs affiliated with the default and executive control networks derived from Schaefer Atlas; subsequently, a sliding-window approach was used to compute the functional connectivity (FC) matrix, and each matrix was partitioned into segregated (state ) and integrated (state ) states using k-means clustering. B The left panel shows the mean number of segregated, integrated, and switching states (SS) during the AUT task across three runs. Error
bars indicate the standard error of the mean. C The average switching frequency for the AUT and OCT task, and paired-sample -tests comparing AUT v.s. OCT (the error bar represents the standard errors) . The right panel shows the mean number of segregated, integrated states, and switching during the OCT task across three runs. Error bars indicate the standard error of the mean. E, F The scatterplots show the Pearson correlations between the originality score (i.e., creative performance in the task, -axis) and the switching frequency of DMN-ECN segregation and integration ( -axis) in AUT and OCT, respectively. The black line represents the overall trend, with the confidence interval indicated by the gray shading. G, H Linear (black fitted line, with the confidence interval indicated by the gray shading) and quadratic relationship (i.e., quadratic model, QM; blue fitted line, with the confidence interval indicated by the gray shading) of originality score ( -axis) and DMN-ECN segregation and integration balance ( -axis), and with the comparison between linear fitted model and quadratic fitted model for AUT and OCT, respectively.
creativity is a continuum and creative outputs occur along a set of cognitive continuums, such as the divergent-convergent thinking continuum, the defocused-focused continuum, the spontaneous-deliberate continuum, the mind-wandering-mindfulness continuum, etc. . In the present neurocognitive framework, such cognitive continuums can map onto the DMN-ECN coupling, in which the DMN and ECN together realize the cycle of generation and evaluation. For example, DMN-ECN coupling might reflect forms of goal-directed memory retrieval and episodic simulation in the stage of idea generation that requires cognitive control ; whereas similar coupling patterns act to constrain and direct evaluation processes to output ideas that are both novel and appropriate in the stage of idea evaluation . Some evidence suggests that different
proportions of DMN and ECN activity within coupling patterns might correspond to generative and evaluative stages of creative thinking . Thus, it is likely that dynamically shifting between generation and evaluation could provide an optimal way to produce a constant stream of creative outputs. Yet, such dynamic switching between generation and evaluation must be carefully balanced, as too much of one without the other is unlikely to produce both novel and appropriate ideas .
Our work further-albeit indirectly-establishes a connection between DMN-ECN switching dynamics and the theoretical generation-evaluation creativity model. Direct evidence supporting this hypothesis is needed, and future research should aim to elucidate the neural basis and underlying mechanisms driving this balance. In sum, our findings from the meta-
analytic network neuroscience approach using previously collected restingstate fMRI data were externally validated via a task-based fMRI study. Thus, our findings contribute to a concentrated cognitive and neuroscientific effort to elucidate the cognitive and neural mechanisms that realize creative thinking .
Limitations and future directions
We acknowledge several limitations in our study. First, we recognize that the “true” effect size is small, meaning that the dynamic reconfiguration between the DMN and ECN is only one factor contributing to creative ability. Based on resting-state fMRI data, our meta-analytic findings suggest that more efficient DMN-ECN interactions support higher creative ability; alternatively, as individuals develop their creative skills, that may increase the efficiency of DMN-ECN interactions, even at rest (e.g., Ref. 86). Importantly, many other factors beyond the dynamic DMN-ECN patterns observed here are certainly involved in creative ability, such as personality, attention, perception, curiosity, and creative self-beliefs. Yet, similar DMNECN interaction patterns have been found for other forms of spontaneous thinking (e.g., mind wandering). Therefore, future research is needed to further explore large-scale brain networks and their dynamic interactions across different creative task contexts, to distinguish between the recruitment of ordinary and creativity-specific neural patterns.
A second limitation of our study is the application of a meta-analytic network neuroscience approach to pool raw data from previous studies. Recently, it has been proposed that extremely large sample sizes (in the tens of thousands) are needed to conduct reproducible Brain-Wide Association studies . However, others have argued for the significance of effect size over sample size in such brain-wide association studies . Our approach employs standard and acceptable meta-analytic methods, and the sample sizes of the included studies are adequate for network neuroscience research . A recent study offers valuable insights into sample sizes and true effect sizes in brain-behavioral phenotype associations, suggesting that typical sample sizes often lead to inflated effect sizes and replication failures. However, as sample sizes increase into the thousands (e.g., ), replication rates improve, and effect size inflation decreases, even though the true associations may be smaller . Therefore, the observed effect size in our study likely reflects the true relationship between DMN-ECN dynamic interaction and divergent thinking ability. Nonetheless, a sufficiently large sample size and a dataset with high homogeneity in data collection methods (including MRI and creativity measures) would be more beneficial for detecting this true relationship. In addition, we echo the message advanced by Grafton et al. that recognizes the need for both small-sample focused studies with large-scale consortia-based studies to truly advance our understanding of how complex neural interactions realize human behavior.
Another potential limitation of the present study is that the spatial resolutions of the chosen atlases, particularly the number of regions included within the DMN and ECN, may affect the overall effect of this study. Some studies have indicated that the choice of brain atlas significantly influences the strength of the association between functional connectivity and cognitive abilities . Therefore, future research is needed to replicate and extend our findings using additional brain atlases with different spatial resolutions.
Conclusions
The present findings reveal that highly creative people are characterized by flexible and balanced interactions between the DMN and ECN (measured at rest). Importantly, similar neural patterns were also observed during creative thinking in an external validation with task-based fMRI data. Thus, our study provides a particularly promising direction for clarifying the dynamic neural and cognitive processes that realize creative cognition. In sum, studying creative cognition from a dynamic framework holds much promise for yielding an enhanced understanding of creativity and providing new insight. Critically, our study highlights a general neural marker of creative ability, thus further demystifying the complexity of creative cognition.
Methods
Datasets
The datasets analyzed in the current study for meta-analytic were from five centers in Austria , Canada , China , Japan , and the United States , constituting multi-center datasets including structural MRI, resting-state fMRI, and creative ability data (assessed by divergent thinking tests). There were 10 samples from four centers and one open-access source, including 3533 healthy participants who completed MRI scans and 2772 participants who also completed creativity assessment. The final valid sample consisted of 2433 participants (male ), with an average age of 21.12 years (SD years), after passing a set of exclusion criteria including failed fMRIprep running (due to poor spatial normalization and/or signal quality, and incomplete raw data), excessive head motion, and incomplete or invalid behavioral data. For these 10 datasets, the resting-state fMRI data (including UNCG, GBB_S1, GBB_S2, SLIM_S1, UG_S3, and TKU) have been published elsewhere ; the task-fMRI data collected has also been published, but the resting-fMRI data collected during the same period was not analyzed in prior work and is included in this study (e.g., QU, UG_S1, UG_S2, UG_S3). Importantly, for all the datasets assessed in the present research, no prior study has utilized the analyses conducted in the present work or examined the patterns of DMN-ECN dynamic interaction using resting-state data in relation to individual creative performance. For more details on each sample, see Supplementary Table 1. All local institutional review boards permitted the use of the de-identified and anonymized data. All study participants provided written informed consent at their local institution prior to participation. All ethical regulations relevant to human research participants were followed.
Behavioral assessment
In this study, individual creative ability was assessed by divergent thinking tasks. Despite different divergent thinking tasks employed across the ten datasets, the guidance and requirements of the task were the same, including participants being asked to come up with as many interesting, novel, and uncommon uses as possible for some objects in a limited time. Consistent with conventional scoring procedures , 2-4 raters scored participants’ responses in each task and obtained good interrater reliability, except for the TKU sample which employed one single trained rater. For each type of divergent thinking measured in one dataset, the average of the scores for each rater was obtained, or latent variable modeling was used to model the overall creativity score. Notably, the precise scoring methods for creativity varied across different centers, encompassing measures of originality or the sum of originality and fluency (for more details on measurements and scoring, see Supplementary Dataset Information).
Image acquisition and processing
Structural and functional data were acquired locally, see more detailed scanning parameters in the Supplementary Dataset Information. All images were analyzed using fMRIPrep 1.4.1rc1 (RRID:SCR_016216) , which is based on Nipype 1.2.0 (RRID:SCR_002502) . A full description of the preprocessing pipeline can be found in Supplementary Information -“Imaging preprocessing.” After preprocessing, participants with substantial head motion ( in translation or in rotation) were excluded from further analysis. Head motion was calculated after ICAAROMA in which noise components (including motion) were identified and excluded during fmriprep processes, thus the value here is not the raw head motion. Temporal artifacts were computed in each dataset by calculating framewise displacement (FD) and FD or DVARS were identified . Participants with greater than mean FD 10% of the resting time points exceeding these values were excluded from further analysis.
Dynamic functional analysis
Following preprocessing, we first computed the ROI-wise SNR via the mean magnetic resonance signal over time divided by the standard deviation of the time series, and those ROIs with inadequate signal (i.e., SNR SD above below the group mean) were excluded. Subsequently, the mean time series
was extracted from regions of interest (ROIs) affiliated with two networks of interest, default mode and executive control networks, which were obtained in MNI standard space from 300 cortical parcels with 17 networks . For each dataset, the temporal resolution of time-resolved connectivity in BOLD time series data of ROIs was estimated by the Multiplication of Temporal Derivatives (MTD) approach within a sliding temporal window of 14 time points, resulting in a matrix with times ROIs ROIs (Supplementary Fig. 3A). The MTD is computed by calculating the point-wise product of the temporal derivative of pairwise time series, which were reliable and replicable across multiple datasets compared to the conventional sliding-window Pearson’s correlation . Next, time-resolved community structure was estimated for each temporal window using the Louvain modularity algorithm from the Brain Connectivity Toolbox , in which the community assignment for each region was assessed 500 times, and a consensus partition was identified using a fine-tuning algorithm. Based on time-resolved community assignments, we estimated the within-module strength z-score for each region and estimated the diversity of between-module strength (i.e., participation coefficient; ) for each region within each temporal window .
Using these two connectivity measures (i.e., and ), we computed a joint histogram of within-module strength and between-module strength measures (by summing the instances of each value of the two measures within 100 equally defined bins along each axis) within each temporal window as a cartographic profile. To test whether the cartographic profile fluctuated over time, we subsequently performed k -means clustering analysis to classify each cartographic profile over time into one of two states (clusters) using a priori parameter (with partition and 100 random restarts), which has been shown to be optimal in previous studies . To determine the specific state for one cartographic profile, we compared the mean scores of regional and scores across the two states using an independent samples -test. Here, one state with a higher score and lower score suggests cooperation and integration between DMN and ECN (i.e., integrated state, ), and the other state means segregation between the two networks (i.e., segregated state, ; Supplementary Fig. 3B). Referring to previous studies , we can determine which state (i.e., segregated or integrated state) dominates the functional connections between the two networks in each time window. Across the time windows, the switching frequency that measures the transition number segregated and integrated states was calculated, which was defined as: Switching frequency ; and the balance of the two states was computed by the difference of dwell time in the two states divided by the whole time, which was defined as: Balance , where is the total dwell time of and . Previous studies have shown that frequent switching between dynamically segregated and integrated states over long time periods leads to balance. Consequently, switching frequency and balance exhibit an inverted U-shaped relationship (Supplementary Fig. 3C) .
Within-dataset analyses
For each dataset, we first computed the correlation between creative performance (i.e., divergent thinking ability) and switching frequency (between segregation and integration of DMN-ECN) controlling for gender, age, mean FD, and global mean signal. In addition, we calculated this relationship in each dataset using a robust correlation approach . Our second question concerned whether the relationship between creative performance and balance of the two dynamic brain states is linear or quadratic. For linear and quadratic regression models, the polynomial regression model contained the balance index (i.e., linear) or the balance index and its squared term (i.e., quadratic) as independent variables, and individual creativity score as the dependent variable, as follows:
Next, ANOVA and the Akaike information criterion (AIC) were used to determine which model better fit the data if both the linear and quadratic models were significant based on the F statistic, where a smaller AIC indicated a better fit. A difference of in the AIC was deemed to reflect a significant difference between models.
Meta-analytic approach
We performed meta-analytic analyses using functions available in the metafor package (version 1.9-6) within the R open-source software environment (version 3.2.0) . In within-dataset analyses, the -value between the state-switching frequency and creative performance was calculated for each dataset. Next, the r values among the ten datasets were transformed into effect sizes using “escalc” function with the measure option set to “ZCOR”. Considering that several centers included in this analysis provided multiple datasets as different projects and creativity measurements, we first used random-effects models at the location level with the “rma.mv” function with the method option set to “REML” implemented in metafor R package. Meanwhile, the random-effects model also fit using “rma” function, which also provided conventional output on the Q test for heterogeneity. Further moderator and subgroup analyses were performed to explore the potential impact of moderators, including locations, MRI protocol, scoring methods, sample size, age, and gender proportion.
Reproducibility
Beyond the meta-analytic network neuroscience approach, we also performed mega-analyses by pooling all available individual-level data from all datasets . This approach allowed us to exclude non-relevant covariates, such as demographic and location factors, and to focus on specific effects. To enable this analysis, all creative performance, switching frequency and functional balance between segregated and integrated DMNECN states were converted to Z-scores within each dataset. Initially, in the mega-analysis, we examined the relationship between creative performance and the frequency of switching between segregation and integration within the DMN-ECN. In this analysis, the switching frequency across datasets was treated as a random factor, while the frequency of switching and non-relevant covariates were considered as fixed factors. Subsequently, we explored two models: the linear and nonlinear relationships between creative performance and the balance of segregation and integration in the DMN-ECN. We then conducted an ANOVA test and compared the AIC values between the linear and quadratic models within the mega-analysis. All mega-analyses were conducted using a linear mixed-effects model (LME), employing the ‘nlme’ package in R (version 4.2.3). In addition, we reanalyzed the data using the Shen brain atlas , which consists of 54 regions of interest within DMN and ECN to confirm our results.
External validation
The use of resting-state fMRI approaches for deriving cognitive explanations has been criticized by some . In this context, eventrelated fMRI approaches offer, to some extent, a more direct test of creative mental operations . Therefore, we conducted an external validation analysis to assess whether the dynamic DMN-ECN switching pattern could be validated in the context of a divergent thinking task involving creative cognitive processes in contrast to a control task requiring common cognitions (e.g., memory retrieval). Thirty-one participants completed both the Alternate Uses Task (AUT) and the Object Characteristics Task (OCT) during fMRI scanning. In the statistical analysis, we initially employed a paired-sample -test to examine whether there was a significant difference in switching frequency between AUT and OCT tasks. Subsequently, a mixed linear or nonlinear model was used to investigate the relationship between creative performance (i.e., AUT performance) and switching frequency and balance of the two dynamic brain states.
Statistics and reproducibility
The sample size details for each dataset, as well as the statistical analyses for both behavioral data and fMRI data, are provided in the respective sections of the “Results” and “Methods.”
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The processed data generated in this study are available on the Open Science Framework at https://osf.io/83arw/, https://doi.org/10.17605/OSF.IO/ 83ARW. Some of the raw data used in the present study has been publicly available and analyzed in previous studies, and a part of the non-public data is available from the corresponding author upon reasonable request.
Received: 11 July 2024; Accepted: 6 January 2025;
Published online: 15 January 2025
References
Beaty, R. E., Benedek, M., Silvia, P. J. & Schacter, D. L. Creative cognition and brain network dynamics. Trends Cogn. Sci. 20, 87-95 (2016).
Benedek, M., Beaty, R. E., Schacter, D. L. & Kenett, Y. N. The role of memory in creative ideation. Nat. Rev. Psychol. 2, 246-257 (2023).
Mayseless, N., Eran, A. & Shamay-Tsoory, S. G. Generating original ideas: the neural underpinning of originality. Neuroimage 116, 232-239 (2015).
Beaty, R. E. & Kenett, Y. N. Associative thinking at the core of creativity. Trends Cogn. Sci. 27, 671-683 (2023).
Lloyd-Cox, J., Chen, Q. & Beaty, R. E. The time course of creativity: multivariate classification of default and executive network contributions to creative cognition over time. Cortex 156, 90-105(2022).
Bendetowicz, D. et al. Two critical brain networks for generation and combination of remote associations. Brain 141, 217-233 (2018).
Marron, T. R. et al. Chain free association, creativity, and the default mode network. Neuropsychologia 118, 40-58 (2018).
Heinonen, J. et al. Default mode and executive networks areas: association with the serial order in divergent thinking. PLoS ONE 11, e0162234 (2016).
Andrews-Hanna, J. R., Smallwood, J. & Spreng, R. N. The default network and self-generated thought: component processes, dynamic control, and clinical relevance. Ann. N. Y. Acad. Sci. 1316, 29-52 (2014).
Andrews-Hanna, J. R., Reidler, J. S., Huang, C. & Buckner, R. L. Evidence for the default network’s role in spontaneous cognition. J. Neurophysiol. 104, 322-335 (2010).
Ovando-Tellez, M. et al. Brain connectivity-based prediction of reallife creativity is mediated by semantic memory structure. Sci. Adv. 8, eabl4294 (2022).
Marron, T. R., Berant, E., Axelrod, V. & Faust, M. Spontaneous cognition and its relationship to human creativity: a functional connectivity study involving a chain free association task. Neuroimage 220, 117064 (2020).
Shofty, B. et al. The default network is causally linked to creative thinking. Mol. Psychiatry 27, 1848-1854 (2022).
Raffaelli, Q. et al. Creative minds at rest: creative individuals are more associative and engaged with their idle thoughts. Creat. Res. J. 36, 396-412 (2024).
Shamay-Tsoory, S. G., Adler, N., Aharon-Peretz, J., Perry, D. & Mayseless, N. The origins of originality: the neural bases of creative thinking and originality. Neuropsychologia 49, 178-185 (2011).
Wang, M., Hao, N., Ku, Y., Grabner, R. H. & Fink, A. Neural correlates of serial order effect in verbal divergent thinking. Neuropsychologia 99, 92-100 (2017).
Kleinmintz, O. M. et al. Participation of the left inferior frontal gyrus in human originality. Brain Struct. Funct. 223, 329-341 (2018).
Chen, Q. et al. A meta-analysis of the effects of transcranial direct current stimulation on creative thinking. Psychol. Aesthet. Creat. Arts. https://doi.org/10.1037/aca0000704 (2024).
Herault, C. et al. Creative connections: the neural correlates of semantic relatedness are associated with creativity. Commun. Biol. 7, 810 (2024).
Ovando-Tellez, M. et al. An investigation of the cognitive and neural correlates of semantic memory search related to creative ability. Commun. Biol. 5, 604 (2022).
Ellamil, M., Dobson, C., Beeman, M. & Christoff, K. Evaluative and generative modes of thought during the creative process. Neuroimage 59, 1783-1794 (2012).
Wu, X. et al. A meta-analysis of neuroimaging studies on divergent thinking using activation likelihood estimation. Hum. Brain Mapp. 36, 2703-2718 (2015).
Benedek, M. et al. To create or to recall? Neural mechanisms underlying the generation of creative new ideas. Neuroimage 88, 125-133 (2014).
Benedek, M. et al. Creating metaphors: the neural basis of figurative language production. Neuroimage 90, 99-106 (2014).
Matheson, H. E., Kenett, Y. N., Gerver, C. & Beaty, R. E. Representing creative thought: a representational similarity analysis of creative idea generation and evaluation. Neuropsychologia 187, 108587 (2023).
Bartoli, E. et al. Default mode network electrophysiological dynamics and causal role in creative thinking. Brain 147, 3409-3425 (2024).
Beaty, R. E. et al. Robust prediction of individual creative ability from brain functional connectivity. Proc. Natl Acad. Sci. USA 115, 1087-1092 (2018).
Takeuchi, H. et al. The association between resting functional connectivity and creativity. Cereb. Cortex 22, 2921-2929 (2012).
Beaty, R. E. et al. Creativity and the default network: a functional connectivity analysis of the creative brain at rest. Neuropsychologia 64, 92-98 (2014).
Shi, L. et al. Large-scale brain network connectivity underlying creativity in resting-state and task fMRI: cooperation between default network and frontal-parietal network. Biol. Psychol. 135, 102-111 (2018).
Frith, E. et al. Intelligence and creativity share a common cognitive and neural basis. J. Exp. Psychol. Gen. 150, 609-632 (2021).
Patil, A. U. et al. Static and dynamic functional connectivity supports the configuration of brain networks associated with creative cognition. Sci. Rep. 11, 165 (2021).
Ren, Z. et al. Connectome-based predictive modeling of creativity anxiety. Neuroimage 225, 117469 (2021).
Beaty, R. E., Benedek, M., Kaufman, S. B. & Silvia, P. J. Default and executive network coupling supports creative idea production. Sci. Rep. 5, 10964 (2015).
Kleinmintz, O. M., Ivancovsky, T. & Shamay-Tsoory, S. G. The twofold model of creativity: the neural underpinnings of the generation and evaluation of creative ideas. Curr. Opin. Behav. Sci. 27, 131-138 (2019).
Fox, K. C. & Christoff, K. The Oxford Handbook of Spontaneous Thought: Mind-wandering, Creativity, and Dreaming (Oxford University Press, 2018).
Braun, U. et al. Dynamic reconfiguration of frontal brain networks during executive cognition in humans. Proc. Natl Acad. Sci. USA 112, 11678-11683 (2015).
Kenett, Y. N., Chrysikou, E. G., Bassett, D. S. & Thompson-Schill, S. L. Neural dynamics during the generation and evaluation of creative and non-creative ideas. Preprint at bioRxiv https://doi.org/10.1101/ 2024.04.15.589621 (2024).
Christoff, K., Irving, Z. C., Fox, K. C., Spreng, R. N. & AndrewsHanna, J. R. Mind-wandering as spontaneous thought: a dynamic framework. Nat. Rev. Neurosci. 17, 718-731 (2016).
Benedek, M. & Jauk, E. Spontaneous and controlled processes in creative cognition in The Oxford Handbook of Spontaneous Thought: MindWandering, Creativity, Dreaming and Clinical Conditions (eds Christoff, K. & Fox, K. C. R.) 285-298 (Oxford University Press, 2018).
Chrysikou, E. G. Creativity in and out of (cognitive) control. Curr. Opin. Behav. Sci. 27, 94-99 (2019).
Tromp, C. Integrated constraints in creativity: foundations for a unifying model. Rev. Gen. Psychol. 27, 41-61 (2023).
Sun, J. et al. Verbal creativity correlates with the temporal variability of brain networks during the resting state. Cereb. Cortex 29, 1047-1058 (2018).
Sporns, O. Network attributes for segregation and integration in the human brain. Curr. Opin. Neurobiol. 23, 162-171 (2013).
Shine, J. M. et al. The dynamics of functional brain networks: integrated network states during cognitive task performance. Neuron 92, 544-554 (2016).
Orwig, W. et al. Creativity at rest: exploring functional network connectivity of creative experts. Netw. Neurosci. 7, 1022-1033 (2023).
Spreng, R. N., Stevens, W. D., Chamberlain, J. P., Gilmore, A. W. & Schacter, D. L. Default network activity, coupled with the frontoparietal control network, supports goal-directed cognition. Neuroimage 53, 303-317 (2010).
Cole, M. W., Ito, T., Bassett, D. S. & Schultz, D. H. Activity flow over resting-state networks shapes cognitive task activations. Nat. Neurosci. 19, 1718-1726 (2016).
Tromp, C. Creativity from constraint exploration and exploitation. Psychol. Rep. 127, 1818-1843 (2024).
Girn, M., Mills, C., Roseman, L., Carhart-Harris, R. L. & Christoff, K. Updating the dynamic framework of thought: creativity and psychedelics. Neuroimage 213, 116726 (2020).
Beaty, R. E., Seli, P. & Schacter, D. L. Network neuroscience of creative cognition: mapping cognitive mechanisms and individual differences in the creative brain. Curr. Opin. Behav. Sci. 27, 22-30 (2019).
Cohen, J. R. The behavioral and cognitive relevance of time-varying, dynamic changes in functional connectivity. Neuroimage 180, 515-525 (2018).
Wang, R. et al. Segregation, integration, and balance of large-scale resting brain networks configure different cognitive abilities. Proc. Natl Acad. Sci. USA 118, e2022288118 (2021).
Kenett, Y. N., Betzel, R. F. & Beaty, R. E. Community structure of the creative brain at rest. Neuroimage 210, 116578 (2020).
Capouskova, K., Zamora-López, G., Kringelbach, M. L. & Deco, G. Integration and segregation manifolds in the brain ensure cognitive flexibility during tasks and rest. Hum. Brain Mapp. 44, 6349-6363 (2023).
Zhuang, K. et al. Diverse functional interaction driven by controldefault network hubs supports creative thinking. Cereb. Cortex 33, 11206-11224 (2023).
Li, J. et al. High transition frequencies of dynamic functional connectivity states in the creative brain. Sci. Rep. 7, 46072 (2017).
Cohen, J. R. & D’Esposito, M. The segregation and integration of distinct brain networks and their relationship to cognition. J. Neurosci. 36, 12083-12094 (2016).
Xu, J. et al. Anxious brain networks: a coordinate-based activation likelihood estimation meta-analysis of resting-state functional connectivity studies in anxiety. Neurosci. Biobehav. Rev. 96, 21-30 (2019).
Jahani, A. et al. Twinned neuroimaging analysis contributes to improving the classification of young people with autism spectrum disorder. Sci. Rep. 14, 20120 (2024).
Acar, S. & Runco, M. A. Divergent thinking: new methods, recent research, and extended theory. Psychol. Aesthet. Creat. Arts 13, 153-158 (2019).
Karwowski, M. et al. Is creativity without intelligence possible? A necessary condition analysis. Intelligence 57, 105-117 (2016).
Gerwig, A. et al. The relationship between intelligence and divergent thinking-a meta-analytic update. J. Intell. 9, 23 (2021).
Borenstein, M., Hedges, L. V., Higgins, J. P. & Rothstein, H. R. Introduction to Meta-analysis (Wiley, 2021).
Beaty, R. E., Merseal, H. M. & Zeitlen, D. C. in The Routledge International Handbook of Creative Cognition, 433-452 (Routledge, 2023).
Shen, X. et al. Using connectome-based predictive modeling to predict individual behavior from brain connectivity. Nat. Protoc. 12, 506-518 (2017).
Benedek, M. & Fink, A. Toward a neurocognitive framework of creative cognition: the role of memory, attention, and cognitive control. Curr. Opin. Behav. Sci. 27, 116-122 (2019).
Volle, E. Associative and Controlled Cognition in Divergent Thinking: Theoretical, Experimental, Neuroimaging Evidence, and New Directions, 333-362 (Cambridge University Press, 2018).
Beaty, R. E., Silvia, P. J. & Benedek, M. Brain networks underlying novel metaphor production. Brain Cogn. 111, 163-170 (2017).
Da Mota, P. A. et al. The dynamics of the improvising brain: a study of musical creativity using jazz improvisation. Preprint at bioRxiv https://doi.org/10.1101/2020.01.29.924415 (2020).
Limb, C. J. & Braun, A. R. Neural substrates of spontaneous musical performance: an FMRI study of jazz improvisation. PLoS ONE 3, e1679 (2008).
Liu, S. Y. et al. Brain activity and connectivity during poetry composition: toward a multidimensional model of the creative process. Hum. Brain Mapp. 36, 3351-3372 (2015).
Matheson, H. E. & Kenett, Y. N. The role of the motor system in generating creative thoughts. Neuroimage 213, 116697 (2020).
Chen, Q. et al. Brain hemispheric involvement in visuospatial and verbal divergent thinking. Neuroimage 202, 116065 (2019).
He, R. et al. The impact of knowledge on poetry composition: an fMRI investigation. Brain Lang. 235, 105202 (2022).
Bilder, R. M. & Knudsen, K. S. Creative cognition and systems biology on the edge of chaos. Front. Psychol. 5, 112333 (2014).
Simonton, D. K. Creative thought as blind-variation and selectiveretention: combinatorial models of exceptional creativity. Phys. Life Rev. 7, 156-179 (2010).
Boot, N., Baas, M., van Gaal, S., Cools, R. & De Dreu, C. K. W. Creative cognition and dopaminergic modulation of fronto-striatal networks: integrative review and research agenda. Neurosci. Biobehav. Rev. 78, 13-23 (2017).
Amabile, T. Componential Theory of Creativity (Harvard Business School, 2011).
Ding, K., He, R., Wang, X., Chen, Q. & Kenett, Y. N. Recognizing ideas generated in a creative task: the roles of the hippocampus and medial prefrontal cortex in facilitating self-generated learning. Cereb. Cortex 34, bhae219 (2024).
Wang, X. et al. Semantic associative abilities and executive control functions predict novelty and appropriateness of idea generation. Commun. Biol. 7, 703 (2024).
Kenett, Y. N. et al. Developing a neurally informed ontology of creativity measurement. Neuroimage 221, 117166 (2020).
Gerver, C. R., Griffin, J. W., Dennis, N. A. & Beaty, R. E. Memory and creativity: a meta-analytic examination of the relationship between memory systems and creative cognition. Psychon. Bull. Rev. 30, 2116-2154 (2023).
Green, A. E., Beaty, R. E., Kenett, Y. N. & Kaufman, J. C. The process definition of creativity. Creat. Res. J. 36, 544-572 (2024).
Japardi, K., Bookheimer, S., Knudsen, K., Ghahremani, D. G. & Bilder, R. M. Functional magnetic resonance imaging of divergent and convergent thinking in Big-C creativity. Neuropsychologia 118, 59-67 (2018).
Kam, J. W. et al. Default network and frontoparietal control network theta connectivity supports internal attention. Nat. Hum. Behav. 3, 1263-1270 (2019).
Zanesco, A. P., Denkova, E. & Jha, A. P. Self-reported mind wandering and response time variability differentiate prestimulus electroencephalogram microstate dynamics during a sustained attention task. J. Cogn. Neurosci. 33, 28-45 (2021).
Liu, S., Abdellaoui, A., Verweij, K. J. & van Wingen, G. A. Replicable brain-phenotype associations require large-scale neuroimaging data. Nat. Hum. Behav. 7, 1344-1356 (2023).
Marek, S. et al. Reproducible brain-wide association studies require thousands of individuals. Nature 603, 654-660 (2022).
Gratton, C., Nelson, S. M. & Gordon, E. M. Brain-behavior correlations: two paths toward reliability. Neuron 110, 1446-1449 (2022).
Kong, X.-Z., Zhang, C., Liu, Y. & Pu, Y. Scanning reproducible brainwide associations: sample size is all you need? Psychoradiology 2, 67-68 (2022).
Rosenberg, M. D. & Finn, E. S. How to establish robust brain-behavior relationships without thousands of individuals. Nat. Neurosci. 25, 835-837 (2022).
Spisak, T., Bingel, U. & Wager, T. D. Multivariate BWAS can be replicable with moderate sample sizes. Nature 615, E4-E7 (2023).
Helwegen, K., Libedinsky, I. & van den Heuvel, M. P. Statistical power in network neuroscience. Trends Cogn. Sci. 27, 282-301 (2023).
Bryce, N. V. et al. Brain parcellation selection: an overlooked decision point with meaningful effects on individual differences in resting-state functional connectivity. Neuroimage 243, 118487 (2021).
Messé, A. Parcellation influence on the connectivity-based structure-function relationship in the human brain. Hum. Brain Mapp. 41, 1167-1180 (2020).
Kong, R. Q. et al. A network correspondence toolbox for quantitative evaluation of novel neuroimaging results. Preprint at bioRxiv https:// doi.org/10.1101/2024.06.17.599426 (2024).
Beaty, R. E. et al. Personality and complex brain networks: the role of openness to experience in default network efficiency. Hum. Brain Mapp. 37, 773-779 (2016).
Jauk, E., Benedek, M., Dunst, B. & Neubauer, A. C. The relationship between intelligence and creativity: new support for the threshold hypothesis by means of empirical breakpoint detection. Intelligence 41, 212-221 (2013).
Jauk, E., Neubauer, A. C., Dunst, B., Fink, A. & Benedek, M. Gray matter correlates of creative potential: a latent variable voxel-based morphometry study. Neuroimage 111, 312-320 (2015).
Fink, A. et al. Modulation of resting-state network connectivity by verbal divergent thinking training. Brain Cogn. 128, 1-6 (2018).
Poppenk, J. Multimodal brain data and core dimensions of creativity. Data Brief 29, 105176 (2020).
Chen, Q. et al. Longitudinal alterations of frontoparietal and frontotemporal networks predict future creative cognitive ability. Cereb. Cortex 28, 103-115 (2018).
Liu, Z. et al. Neural and genetic determinants of creativity. Neuroimage 174, 164-176 (2018).
Takeuchi, H. et al. Originality of divergent thinking is associated with working memory-related brain activity: evidence from a large sample study. Neuroimage 216, 116825 (2020).
Orwig, W., Diez, I., Vannini, P., Beaty, R. & Sepulcre, J. Creative connections: computational semantic distance captures individual creativity and resting-state functional connectivity. J. Cogn. Neurosci. 33, 499-509 (2021).
Silvia, P. J. et al. Assessing creativity with divergent thinking tasks: exploring the reliability and validity of new subjective scoring methods. Psychol. Aesthet. Creat. Arts 2, 68-85 (2008).
Esteban, O. et al. fMRIPrep: a robust preprocessing pipeline for functional MRI. Nat. Methods 16, 111-116 (2019).
Gorgolewski, K. et al. Nipype: a flexible, lightweight and extensible neuroimaging data processing framework in Python. Front. Neuroinform. 5, 13 (2011).
Patel, A. X. et al. A wavelet method for modeling and despiking motion artifacts from resting-state fMRI time series. Neuroimage 95, 287-304 (2014).
Drysdale, A. T. et al. Resting-state connectivity biomarkers define neurophysiological subtypes of depression. Nat. Med. 23, 28-38 (2017).
Schaefer, A. et al. Local-global parcellation of the human cerebral cortex from intrinsic functional connectivity MRI. Cereb. Cortex 28, 3095-3114 (2018).
Rubinov, M. & Sporns, O. Complex network measures of brain connectivity: uses and interpretations. Neuroimage 52, 1059-1069 (2010).
Guimera, R. & Amaral, L. A. N. Functional cartography of complex metabolic networks. Nature 433, 895-900 (2005).
Pernet, C. R., Wilcox, R. & Rousselet, G. A. Robust correlation analyses: false positive and power validation using a new open source Matlab toolbox. Front. Psychol. 3, 606 (2013).
Viechtbauer, W. Conducting meta-analyses in with the metafor package. J. Stat. Softw. 36, 1-48 (2010).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation, 2013).
Costafreda, S. G. Pooling FMRI data: meta-analysis, mega-analysis and multi-center studies. Front. Neuroinform. 3, 33 (2009).
Davis, S. W., Stanley, M. L., Moscovitch, M. & Cabeza, R. Restingstate networks do not determine cognitive function networks: a commentary on Campbell and Schacter (2016). Lang. Cogn. Neurosci. 32, 669-673 (2017).
Finn, E. S. Is it time to put rest to rest? Trends Cogn. Sci. 25, 1021-1032 (2021).
Acknowledgements
Q.C. was supported by the National Natural Science Foundation of China (NSFC31800919, NSFC32071070). R.E.B. was supported by grants from the US National Science Foundation [DRL-1920653; DUE-2155070]. This work was partially supported by the US-Israel Binational Science Fund (BSF) grant (number 2021040) to R.E.B. and Y.N.K.
Author contributions
Q.C., Y.N.K., and R.E.B. conceptualized the study; H.T., A.F., M.B., R.K., J.Q., and R.E.B. contributed the data; Q.C., Y.N.K., Z.C., K.Z., and R.E.B. performed the analyses; Q.C., Y.N.K., and R.E.B. wrote the original draft; Z.C., M.B., D.C.Z., K.Z., J.L.C., J.Q., and R.E.B. review and editing. All coauthors provided feedback and approved the manuscript.
Correspondence and requests for materials should be addressed to Yoed N. Kenett or Jiang Qiu.
Peer review information Communications Biology thanks Quentin Raffaelli and the other anonymous reviewer(s) for their contribution to the peer review of this work. Primary handling editor: Jasmine Pan. A peer review file is available.
Faculty of Psychology, Southwest University, Chongqing, China. Department of Psychology, Pennsylvania State University, University Park, Pennsylvania, USA. Faculty of Data and Decision Sciences, Technion-Israel Institute of Technology, Haifa, Israel. Chinese Institute for Brain Research, Beijing, China. Division of Developmental Cognitive Neuroscience, Institute of Development, Aging and Cancer, Tohoku University, Sendai, Japan. Department of Psychology, University of Graz, Graz, Austria. Ilnstitute of Science and Technology for Brain-Inspired Intelligence, Fudan University, Shanghai, China. Department of Psychology, Goldsmiths, University of London, London, UK. These authors contributed equally: Qunlin Chen, Yoed N. Kenett. □e-mail: yoedk@technion.ac.il; qiuj318@swu.edu.cn