تتحكم الخلايا الدبقية الصغيرة في انقراض سلوكيات القلق الناتجة عن الإجهاد الحاد في ذكور الفئران Microglia govern the extinction of acute stress-induced anxiety-like behaviors in male mice
تظهر الأعراض المرتبطة بالقلق بعد الإجهاد الحاد عادةً أنها تتلاشى تدريجياً خلال فترة من الزمن. ومع ذلك، فإن الآليات التي تكمن وراء كيفية تعامل الأفراد مع الإجهاد لتحقيق انقراض القلق ليست واضحة. هنا نوضح أن الإجهاد الناتج عن التقييد الحاد يسبب زيادة في نشاط الخلايا العصبية GABAergic في الـفي ذكور الفئران، مما يؤدي إلى سلوكيات شبيهة بالقلق خلال 12 ساعة؛ في حين أن مستوى GABA مرتفعإفراز CX3CL1 العصبي عبر MST4 (كيناز شبيه الستريل-20 من الثدييات 4) – NF-кB – إشارة CX3CL1 ينشط في النهاية الخلايا الدبقية الصغيرة في CeA. الخلايا الدبقية الصغيرة المنشطة بدورها تثبط GABA.النشاط العصبي من خلال ابتلاع أشواكها الشجرية، مما يؤدي في النهاية إلى انقراض سلوكيات القلق الناتجة عن ضغط القيود. تكشف هذه النتائج عن آلية جزيئية وخلوية ديناميكية حيث تدفع الخلايا الدبقية الصغيرة ردود فعل سلبية لتثبيط GABA.النشاط العصبي، مما يسهل الحفاظ على توازن الدماغ في استجابة للتوتر الحاد.
القلق الناتج عن الضغط الحاد من المحتمل أن يكون تكيفًا تطوريًا للحفاظ على اليقظة العالية والانتباه المستمر للمؤثرات التي تتطلب اهتمامًا وثيقًا أو لتجنب التعرض المتكرر لظروف خطرة.من المعروف أن القلق الحاد يمكن أن يتلاشى تدريجياً مع إزالة محفزات الضغط. ومع ذلك، فإن آليات كيفية تعامل الأفراد مع الضغط الحاد ودفع انقراض القلق لتجنب الآثار غير القابلة للعكس التي تسببها القلق المستمر لا تزال غير واضحة.
يمكن أن تتواصل الخلايا الدبقية الصغيرة والعصبونات في الدماغ بشكل ثنائي الاتجاه. عند تلقيها محفزات خارجية، تستجيب العصبونات أولاً وتقوم بوساطة سلسلة من الاستجابات في جميع أنحاء الدماغ، بما في ذلك تعديل نشاط خلايا المناعة العصبية، مثل الخلايا الدبقية الصغيرة، من خلال إفراز عوامل قابلة للذوبان (مثل الكيموكينات والسيتوكينات و ناقلات عصبيةبصفتها البلعميات المقيمة في الدماغ، تؤدي الخلايا الدبقية الصغيرة وظائف أساسية مطلوبة للحفاظ على توازن الدماغ، على سبيل المثال، إزالة الخلايا العصبية الميتة، تقليم المشابك غير الوظيفية، أو إنتاج الروابط التي تدعم بقاء الخلايا العصبية.من المعروف أيضًا أن تنشيط الخلايا الدبقية الصغيرة يشارك في تحفيز القلق في نماذج حيوانية من الإجهاد المزمن.على الرغم من عدة عقود من البحث المكثف الذي يركز على إعادة تشكيل الخلايا العصبية في الدماغ بعد الإجهاد الحاد أو المزمن، لا يزال فهمنا للآليات الديناميكية التي تنظم تفاعلات الخلايا العصبية والميكروغليا، ودورها في التعافي من القلق الناتج عن الإجهاد الحاد، غير مكتمل.
يمكن أن يؤدي التعرض الحاد أو المزمن للإجهاد إلى عدة تغييرات تكيفية طويلة الأمد في مناطق الدماغ الحساسة للإجهاد مثل اللوزة الدماغية، التي أظهرت أنها تخضع لتغييرات جذرية في
وظيفة وبنيةأظهرت بيانات التصوير العصبي أن النشاط العصبي مرتفع باستمرار في اللوزة الدماغية لدى المرضى الذين يعانون من اضطرابات المزاج، مثل الخوف والاكتئاب.بشكل خاص، يُعتبر النواة المركزية لللوزة الدماغية (CeA) النواة الرئيسية للإخراج لوظائف اللوزة الدماغية، وقد تم إثبات أن زيادة نشاطها تُعتبر محركًا قويًا للسلوكيات الشبيهة بالقلق في الفئران.على الرغم من أن نشاط الخلايا العصبية في CeA يبدو أنه يشارك في استجابات الإجهاد التي تؤدي إلى الاضطرابات النفسية العصبية، إلا أن الآليات الجزيئية والخلوية لهذه الاستجابات العصبية في CeA خلال انقراض القلق الناتج عن الإجهاد الحاد لا تزال غير واضحة.
في هذه الدراسة، نوضح كيف أن الضغط الحاد يحفز تنشيط الخلايا الدبقية الصغيرة عبر إشارة MST4 (كيناز شبيه بالستريل-20 من الثدييات 4)-NF-кB-CX3CL1 بطريقة تعزز لاحقًا انقراض سلوكيات القلق في الفئران. نجد أن انخفاض تعبير MST4 في الخلايا العصبية GABAergic في CeA (GABAيساهم في زيادة إفراز CX3CL1، مما يحفز تنشيط الخلايا الدبقية الصغيرة، والذي بدوره يزيد من ابتلاع GABA.الشوكات العصبية، مما يقلل من GABAالنشاط العصبي وتخفيف السلوكيات الشبيهة بالقلق خلال 12 ساعة بعد الإجهاد الناتج عن التقييد الحاد. تدعم هذه النتائج دور الميكروغليا التنظيمي الإيجابي في التنشيط المؤقت والقابل للعكس للاستجابة للإجهاد الحاد، وبالتالي، الحفاظ على توازن الدماغ في الفئران الذكور. توفر هذه النتائج أساسًا آليًا لزوال السلوكيات الشبيهة بالقلق وتقترح استراتيجية تدخل لعلاج آثار إعادة تشكيل الدماغ الناتجة عن الإجهاد.
النتائج
جاباتشارك النشاط العصبي في بداية وانقراض سلوكيات مشابهة للقلق في الفئران تحت الضغط الحاد
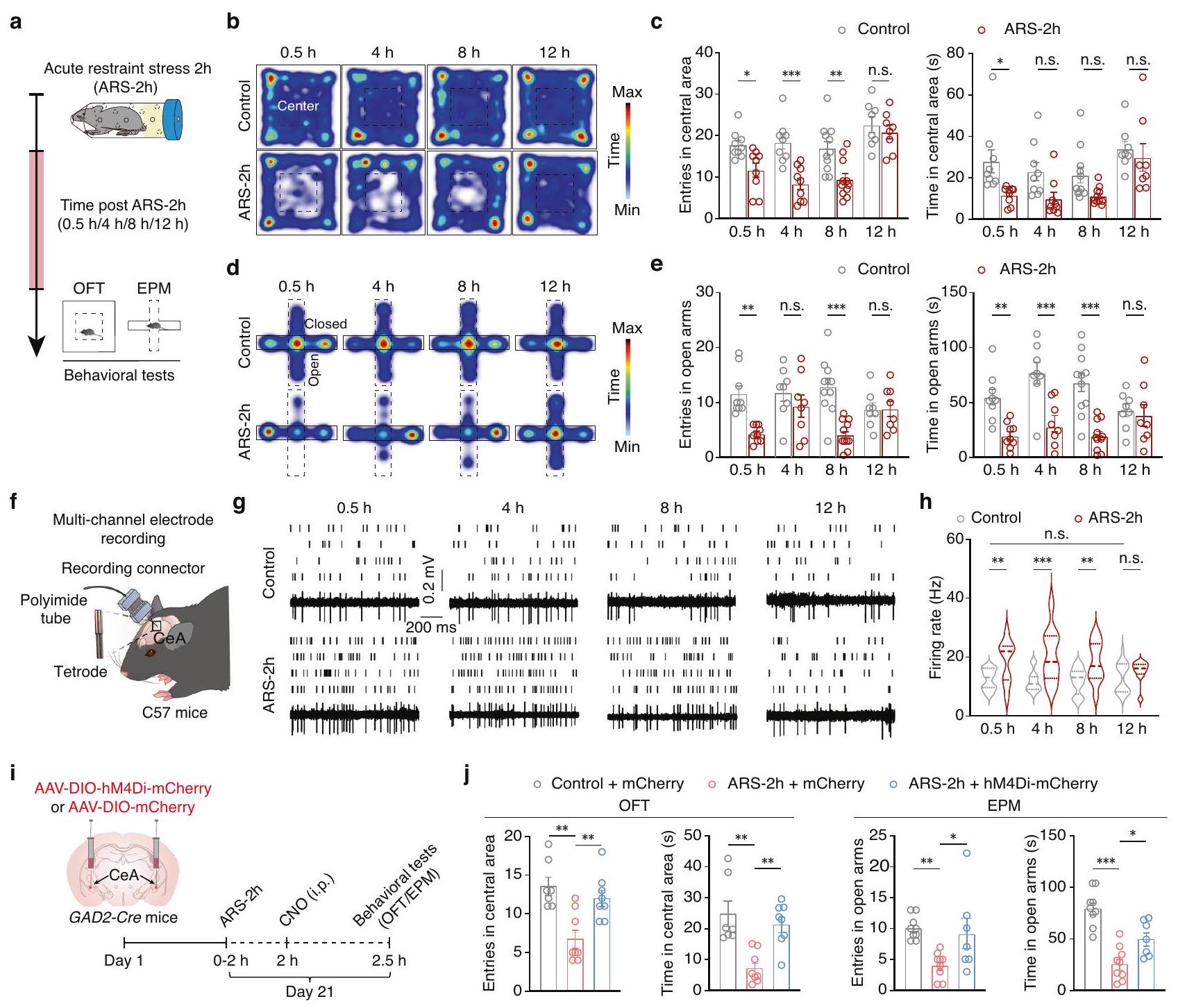
لتحقيق في الأساس العصبي لاستجابة الإجهاد الحاد، قمنا بتحفيز سلوكيات شبيهة بالقلق في ذكور الفئران من خلال إجهاد التقييد الحاد لمدة ساعتين (ARS-2h)، كما تم وصفه سابقًا.. ثم تم استخدام اختبار الحقل المفتوح (OFT) واختبار المتاهة المرتفعة (EPM) لتقييم سلوكيات الاستكشاف في الفئران في “، و 12 ساعة بعد ARS-2h وفي الضوابط غير المجهدة (الشكل 1a). تم ملاحظة تغييرات كبيرة في سلوكيات الاستكشاف، والتي ترتبط ارتباطًا وثيقًا بسلوكيات مشابهة للقلق، حيث تم الإشارة إلى قضاء وقت أقل في المنطقة المركزية من اختبار التفضيل المفتوح (OFT) وعدد أقل من الدخول إلى الأذرع المفتوحة من اختبار الأذرع المفتوحة (EPM).تم induction بعد الإجهاد في فئران ARS-2h مقارنة بالفئران الضابطة، بينما لم يُلاحظ أي فرق كبير بين الفئران الضابطة وفئران ARS-2h بعد 12 ساعة من التقييد (الشكل 1b-e). بالإضافة إلى ذلك، لا يوجد فرق كبير في المسافة المقطوعة في اختبار فتح المجال.بعد 2 ساعة من ARS-2h مقارنةً بالفئران الضابطة المقابلة (الشكل التوضيحي 1). أظهرت هذه البيانات أن التغيرات في سلوكيات الاستكشاف الناتجة عن الإجهاد الناتج عن القيود الحادة تختفي بعد 12 ساعة من بدء الإجهاد.
يلعب النواة المركزية (CeA) دورًا حاسمًا في توليد حالات القلق.. ثم قمنا بفحص GABAالنشاط العصبي في8 ساعات و 12 ساعة بعد ARS-2h في الفئران المتحركة بحرية بواسطة تسجيلات متعددة الأقطاب في الجسم الحي (الشكل 1f). خصائص موجات النبضات لـ GABAتم تحديد الخلايا العصبية باستخدام الوسم الضوئي في فئران GAD2-Cre (الشكل التوضيحي 2). أظهرت النتائج أن معدل إطلاق النبضات العفوي قد زاد في GABA.الخلايا العصبية لفئران ARS-2h مقارنة بالفئران الضابطة ولكنها انخفضت مع مرور الوقت حتى عادت إلى مستويات قابلة للمقارنة مع الضوابط غير المجهدة بعد 12 ساعة من القيود، مما يتماشى جيدًا مع نتائج الاختبارات السلوكية (الشكل 1g، h).
لمزيد من التحقيق في كيفية تأثير GABAالخلايا العصبية شاركت في السلوكيات الاستكشافية، GABAتم تثبيط النشاط العصبي عن طريق الحقن الموضعي لـ AAV-DIO-hM4Di-mCherry في CeA الثنائي الجانب لفئران GAD2-Cre (الشكل 1i والشكل التكميلي 3a). بعد ثلاثة أسابيع من تعبير الفيروس، كشفت تسجيلات توصيل الخلايا الكاملة في شرائح الدماغ الحادة أن جهد الغشاء الراحة ( ) تم تضخيمه في GABA الخلايا العصبية بعد التروية بـ Clozapine-N-oxide (CNO) ) (الشكل التكميلي 3ب، ج)، مما يدل على أن الفيروس تم التعبير عنه بنجاح في GABAالخلايا العصبية. بالإضافة إلى ذلك، في فئران ARS-2h، تم تثبيط GABA باستخدام الكيمياء الجينية.زاد عدد الخلايا العصبية من الوقت الذي قضاه والدخول إلى المنطقة المركزية من اختبار التفضيل المفتوح (OFT) والأذرع المفتوحة من اختبار القلق المفتوح (EPM) (الشكل 1j). علاوة على ذلك، كانت الفئران السليمة غير المجهدة مع تنشيط كيميائي جيني لـأظهرت الخلايا العصبية مستوى منخفضًا من السلوكيات الاستكشافية (الشكل التوضيحي 4). تشير هذه النتائج إلى أن الضغط الناتج عن القيود الحادة يزيد من مستوى GABA.النشاط العصبي ويحفز سلوكيات مشابهة للقلق في الفئران، بينما يرتبط انخفاض هذا النشاط العصبي بانقراض سلوكيات مشابهة للقلق.
تنشيط الخلايا الدبقية الصغيرة مرتبط بإخماد سلوكيات القلق بعد التعرض للإجهاد الحاد
تعمل الخلايا الدبقية الصغيرة كمنظمات حاسمة لنشاط الخلايا العصبية والاستجابات السلوكية المرتبطة بها في الفئران، بما في ذلك سلوكيات مشابهة للقلق.لفحص ما إذا كانت التغيرات في GABAالنشاط العصبي مع مرور الوقت مرتبط بتنشيط الخلايا الدبقية الصغيرة في فئران ARS-2h، قمنا بفحص تنشيط الخلايا الدبقية الصغيرة في CeA من خلال الكشف المناعي الفلوري عن علامة محددة للخلايا الدبقية الصغيرة وهي جزيء 1 المرتبط بالكالسيوم المؤين (lbal). عند 0.5 ساعة و 8 ساعات بعد ARS2 h، عددكانت خلايا الميكروغليا أعلى بشكل ملحوظ في الفئران النموذجية مقارنة بالفئران الضابطة المقابلة، ولكن هذا التغيير تم عكسه بعد 12 ساعة من ARS-2h (الشكل 2a، b). نظرًا لأن شكل الميكروغليا مرتبط بحالة تنشيطهاقمنا بتحليل شكل الخلايا الدبقية الصغيرة باستخدام قياسات مورفومترية ثلاثية الأبعاد شبه تلقائية، والتي كشفت عن عمليات أقصر بشكل ملحوظ ونقاط تفرع أقل بعد 0.5 ساعة و8 ساعات من ARS-2h مقارنة بالفئران الضابطة المقابلة (الشكل 2c، d). ومن الجدير بالذكر أن هذه الأنماط التنشيطية للخلايا الدبقية الصغيرة عادت إلى مستوياتها الطبيعية بعد 12 ساعة من ARS-2h في الفئران النموذجية (الشكل 2b-d).
من منظور جزيئي، قمنا أيضًا باختبار مستويات علامة الالتهاب MHCII ومجموعة من العوامل الالتهابية، بما في ذلك TNF-، إيل- ، و IL-6 في الخلايا الدبقية. أظهرت صبغة المناعة الفلورية أن تعبير MHCII زاد في فئران ARS-2h مقارنةً بالتحكمات بعد 0.5 ساعة من تحفيز الإجهاد المرتبط بتنشيط الخلايا الدبقية (الشكل التوضيحي التكميلي 5a، b). بالإضافة إلى ذلك، قمنا بعزل خلايا الدبقية من CeA باستخدام فرز الخلايا المعتمد على الفلورية باستخدام فئران Cx3cr1-GFP المعدلة وراثيًا، التي تعبر عن علامة GFP في خلايا الدبقية، ووجدنا أن mRNA لـ Tnf-تم التعبير عن IL-6 الذي تم تحليله بواسطة qPCR بمستويات أعلى بشكل ملحوظ في فئران ARS-2h مقارنةً بمجموعة التحكم غير المجهدة بعد 0.5 ساعة من علاج الإجهاد (الشكل التوضيحي 5c-e). للتحقق من ملاحظاتنا حول التغيرات في كثافة الخلايا الدبقية الصغيرة، قمنا بصبغ Ki67 (علامة لتكاثر الخلايا) ووجدنا تعبيرًا واضحًا عن Ki67 في الخلايا الدبقية الصغيرة في CeA لفئران ARS-2h بعد 0.5 ساعة و8 ساعات من العلاج مقارنةً بالمجموعة الضابطة، تلا ذلك انخفاض تدريجي على مر الزمن (الشكل التوضيحي 6a). علاوة على ذلك، أظهرت اختبارات TUNEL أن مستويات الموت الخلوي بلغت ذروتها بعد 8 ساعات من ARS-2h، ثم عادت إلى مستوياتها الأساسية بعد 12 ساعة من علاج ARS-2h (الشكل التوضيحي 6b). أدت هذه النتائج إلى افتراضنا أن تنشيط الخلايا الدبقية الصغيرة في CeA الناتج عن الإجهاد الحاد قد يكون له دور في انقراض سلوكيات القلق.
لفحص ما إذا كانت تنشيط الخلايا الدبقية الصغيرة في CeA مسؤولاً عن سلوكيات القلق الشبيهة الناتجة عن ARS، تم إجراء حقن دقيق داخل الجمجمة باستخدام المينوسكلين، الذي تم استخدامه على نطاق واسع لتثبيط نشاط الخلايا الدبقية الصغيرة، في CeA الثنائي بواسطة زراعة قنية (الشكل 2e). أظهرت الفئران النموذجية التي تم حقنها مسبقًا بالمينوسكلين انخفاضًا في الوقت الذي قضته والدخول إلى المنطقة المركزية من اختبار التفضيل المفتوح (OFT) والأذرع المفتوحة من اختبار الأذرع المفتوحة (EPM) بعد 12 ساعة من ARS-2h، بينما أظهرت الفئران المعالجة مسبقًا بعلاج ACSF التحكم مستويات عالية من السلوكيات الاستكشافية بعد 12 ساعة من تحفيز الإجهاد (الشكل 2f، g). ومن الجدير بالذكر أيضًا أن مستويات منخفضة من السلوكيات الاستكشافية كانت ملحوظة بعد 0.5 ساعة من ARS-2h في الفئران المعالجة مسبقًا إما بالمينوسكلين أو ACSF (الشكل 2f، g). وأكدت صبغة المناعة الفلورية لـ Iba1 أن تنشيط الخلايا الدبقية الصغيرة في CeA تم تثبيطه بشكل كبير في الفئران المعالجة مسبقًا بالمينوسكلين قبل تحفيز الإجهاد ARS-2h.
الشكل 1 | غاباتتوسط الخلايا العصبية سلوكيات القلق الناتجة عن الإجهاد الحاد. أ مخطط لتصميم التجربة. خرائط حرارية تمثيلية (ب) وبيانات ملخصة للدخول والوقت المستغرق في المنطقة المركزية (ج) من اختبار المجال المفتوح (OFT) من المجموعات المحددة.فئران لكل مجموعة؛فئران لكل مجموعة؛فئران لكل مجموعة؛الفئران لكل مجموعة). خرائط حرارية تمثيلية (د) وبيانات ملخصة للدخول والوقت المستغرق في الأذرع المفتوحة (هـ) من متاهة بلس المرتفعة (EPM) من المجموعات المحددة (فئران لكل مجموعة؛فئران لكل مجموعة؛فئران لكل مجموعة؛فئران لكل مجموعة؛ e اليسار،; صحيح، ). مخطط لتسجيلات الأقطاب الكهربائية متعددة القنوات. رسومات شبكية وآثار نموذجية (g) والبيانات الكمية ( ) من الإطلاقات العفوية لـ الخلايا العصبية من
i مخطط الإجراء التجريبي.بيانات ملخصة للمدخلات والوقت المستغرق في المنطقة المركزية والأذرع المفتوحة من OFT و EPM من المجموعات المحددة (OFT: التحكم + mCherry،فئران، ARS-8 فئران، ARSفئران؛ تركت،; صحيح، , ; EPM: التحكم + ميري، فئران، ARS-2مCherryفئران، ARS-2h; اليسار، ; صحيح، ، تم تقييم الأهمية باستخدام تحليل التباين المتكرر ذو الاتجاهين مع المقارنة بعد الاختبار بين المجموعات في (تحليل التباين الأحادي (ANOVA) مع مقارنة بعدية بين المجموعات في ( ). جميع البيانات مقدمة كمتوسط SEM., ; ن.س.، غير مهم. انظر أيضًا البيانات التكميلية 1. يتم تقديم بيانات المصدر كملف بيانات مصدر. مقارنة بالفئران التي تم معالجتها مسبقًا بـ ACSF (الشكل التوضيحي 7). لم يُلاحظ أي تفاعل واضح للخلايا الدبقية في CeA للفئران التي تم زرع قنية فيها (الشكل التوضيحي 8). بالإضافة إلى ذلك، أظهرت تسجيلات متعددة الأقطاب في vivo في فئران ARS-2h المتحركة بحرية مع حقن دقيق مسبق في CeA للمنوكسيلين أن النشاط العصبي التلقائي لـ GABAكان عدد الخلايا العصبية أعلى بشكل ملحوظ بعد 12 ساعة من ARS-2h مقارنةً بتلك الموجودة في الفئران التي تم إعطاؤها محلول ملحي قبل ARS-2h (الشكل 2h، i)، على الرغم من عدم وجود فرق في النشاط العصبي التلقائي لـ GABA.تم الكشف عن الخلايا العصبية بعد 0.5 ساعة بين الفئران ARS-2h المعالجة بالمنوocyكلين وتلك المعالجة بالمحلول الملحي (الشكل 2h، i). تشير هذه النتائج إلى أن تثبيط تنشيط الخلايا الدبقية يمنع استعادة GABA.فرط النشاط العصبي وانقراض السلوكيات الشبيهة بالقلق الناتجة عن الإجهاد الحاد.
ابتلاع الخلايا الدبقية الصغيرة لجاباتشير الشوكات العصبية إلى انقراض السلوكيات الشبيهة بالقلق الناتجة عن الإجهاد الحاد أظهرت التقارير السابقة أن الميكروغليا المنشطة يمكن أن تؤثر على النشاط العصبي من خلال الابتلاع المباشر للشوكات في الدماغ.للتحقيق في العلاقة المحتملة بين حساسية الخلايا الدبقية الصغيرة للإجهاد و GABAالنشاط المنخفض للخلايا العصبية خلال انقراض سلوكيات مشابهة للقلق، قمنا بفحص إعادة تشكيل GABAالشوكات العصبية بعد ARS-2h في الفئران النموذجية. لهذا الغرض، قمنا بإجراء وسم نادر محدد لنوع الخلية عن طريق حقن فيروس AAV-CSSP-YFP8E3 في CeA الثنائي الجانب لفئران GAD2-Cre لوسم الشوكات الشجرية لـ GABA بشكل محدد.الخلايا العصبية (الشكل 3أ). بالمقارنة مع الفئران الضابطة المقابلة، فإن كثافة GABAشجيرات عصبية
الشكل 2 | انقراض سلوكيات مشابهة للقلق يعتمد على الميكروغليا النشطة. أ صور تمثيلية لتلوين Ibal المناعي وإعادة بناء ثلاثي الأبعاد للميكروغليا في CeA من المجموعات المحددة. وحدات القياس، (نظرة عامة) و (الإدراج والرسم). تكميمأعداد الخلايا في CeA من المجموعات المحددة (فئران لكل مجموعة؛، ت quantification شبه تلقائي للمورفولوجيا الخلوية المعتمدة على IMARIS، بما في ذلك الطول الإجمالي للعملية (ج) وعدد نقاط التفرع (د) لـ Iba1الميكروغليا في CeA من المجموعات المحددةفئران لكل مجموعة؛ ). المخطط التجريبي لفئران ARS2 h المعالجة مسبقًا بـ ACSF أو مينو سيكلين (مينو). بيانات ملخصة للمدخلات والوقت المستغرق في المنطقة المركزية من OFT (0.5 ساعة: ARS-2 ساعة + ACSF،فئران، ARS-2h
مينو،فئران؛ 12 ساعة: ARS-2 ساعة + ACSF،فئران، ARS-2h + مينو،فئران؛ تركت،; صحيح، ). بيانات ملخصة عن المدخلات والوقت المستغرق في المنطقة المركزية من EPM (0.5 ساعة: ARS-ACSFفئران، ARS-2h + مينو،فئران؛ 12 ساعة: ARS-2 ساعة + ACSF،فئران، ARS-2h + مينو،فئران؛ تركت،; صحيح، ). مخططات النقطية والمسارات النموذجية ( ) والبيانات الكمية ( ) من الإطلاقات العفوية لـ GABA الخلايا العصبية من المجموعات المحددة (خلايا من ستة فئران لكل مجموعة؛ أناتم تقييم الأهمية باستخدام تحليل التباين المتكرر ذو الاتجاهين مع مقارنة بعدية بين المجموعات في (ب، ج، د، و، ز، ي). جميع البيانات مقدمة كمتوسطSEM.; ن.س.، غير مهم. انظر أيضًا البيانات التكميلية 1. يتم تقديم بيانات المصدر كملف بيانات المصدر. تم زيادة الشوكات عند 0.5 ساعة و 8 ساعات بعد ARS-2 ساعة في الفئران النموذجية ولكنها عادت إلى مستويات التحكم بحلول 12 ساعة بعد ARS-2 ساعة (الشكل 3ب، ج). ومع ذلك، فإن GABAلم يكن هناك فرق كبير في كثافة الشوكات الشجرية العصبية بين الفئران النموذجية التي تم علاجها مسبقًا بالمنوكسيلين (عن طريق الحقن داخل الصفاق) وتلك المعالجة بمحلول ملحي بعد 0.5 ساعة من ARS-2h، ولكن كانت الكثافة أعلى بشكل ملحوظ في مجموعة العلاج المسبق بالمنوكسيلين بعد 12 ساعة من تحفيز الإجهاد مقارنة بمجموعة العلاج المسبق بمحلول ملحي في الفئران النموذجية (الشكل 3d-f). تشير هذه النتائج إلى أن زيادة ابتلاع الخلايا الدبقية الصغيرة مطلوبة من أجل الانخفاض الملحوظ في كثافة الشوكات الشجرية العصبية لـ GABA.الخلايا العصبية خلال انقراض سلوكيات القلق الناتجة عن الضغط الناتج عن التقييد الحاد.
لتحقيق التفاعلات بين عمليات الخلايا الدبقية الصغيرة والشوكيات الشجرية لـ GABAفي الخلايا العصبية، قمنا بفحص وظيفة البلعمة للميكروغليا في فئران ARS-2h. أظهر تلوين المناعة الفلورية لعلامة البلعمة CD68 مستويات أعلى بشكل ملحوظ في فئران ARS-2h مقارنةً بالمجموعة الضابطة بعد 0.5 ساعة من تحفيز الإجهاد، ولكن لم يكن هناك فرق قابل للاكتشاف بين الفئران المجهدة والمجموعة الضابطة بعد 12 ساعة. بعد ARS-2h، بما يتماشى مع التغيرات الملحوظة في تعبير Ibal في الخلايا الدبقية الصغيرة (الشكل 3g). بالإضافة إلى ذلك، أظهرت اختبارات الابتلاع الكمية أن الخلايا الدبقية الصغيرة احتوت على المزيد منالشوكات الشجرية العصبية، مصحوبة بمناطق تماس أكبر بين الميكروغليا والشجرة، في فئران ARS-2h مقارنة بتلك الموجودة في الحيوانات الضابطة بعد 0.5 ساعة، ولكن ليس بعد 12 ساعة، من بدء الضغط (الشكل التوضيحي 9). علاوة على ذلك، أظهرت إعادة البناء ثلاثي الأبعاد زيادة ملحوظة في التداخل المكاني للنقاط المناعية GAD65/67، وهو علامة محددة للعصبونات GABAergic، والميكروغليا في CeA بعد 0.5 ساعة من ARS-2h، مقارنة بالفئران الضابطة، ولم يعد هذا الظاهرة تحدث بعد 12 ساعة من ARS-2h في الفئران النموذجية (الشكل 3h). بيانات التصوير المجهري التداخلي والتصوير السطحي ثلاثي الأبعاد أظهرت مزيدًا من التداخل بين النقاط المناعية التفاعلية لـ GAD65/67 و CD68 و Iba1.الميكروغليا في CeA لفئران ARS-2h بعد 0.5 ساعة، ولكن ليس بعد 12 ساعة من العلاج، ولا في الحيوانات الضابطة المقابلة (الشكل 3i). بعد المعالجة المسبقة بالمنوسكلين في فئران ARS2h، كانت بلعمة الميكروغليا لإشارات GAD65/67 في CeA أقل بكثير من تلك في مجموعة المعالجة المسبقة بالمحلول الملحي (الشكل 3j).
الشكل 3 | انقراض سلوكيات مشابهة للقلق من خلال ابتلاع الخلايا الدبقية
جاباالشوكات الشجرية. مخطط لعدوى فيروسية ثنائية الجانب. صور تمثيلية للتشعبات العصبية (ب) وقياس عدد الشوكات لكل ( فئران لكل مجموعة؛ ). قضبان القياس، “. د. مخطط تجريبي لحقن الفيروس. صور تمثيلية للتغصنات العصبية (هـ) وقياس عدد الشوكات ( ) من المجموعات المحددة ( فئران لكل مجموعة؛ ). قضبان القياس، . صور تمثيلية (يسار) وتحليلات كمية (يمين) للتلوين المناعي لـ CD68 (أحمر)، Iba1 (أخضر)، وDAPI (أزرق) في CeA من المجموعات المحددة.فئران لكل مجموعة؛ إيبال،شريط القياس،. صور تمثيلية ورسم ثلاثي الأبعاد (يسار) لميكروغليا Iba1+ (أخضر) تحتوي على نقاط GAD65/67+ (أحمر) وDAPI (أزرق) في CeA من المجموعات المحددة. الكمية (يمين) لـ GAD65/67نقاط في الميكروغليا في الفئرانخلايا من ستة فئران لكل مجموعة؛ ). قضبان القياس، (نظرة عامة) و (الإدراج والرسم). صور تمثيلية (يسار) وتحليلات كمية (يمين) للتلوين المناعي لـ CD68 (أحمر)، Iba1 (أخضر)، و GAD65نقاط (أرجوانية) في CeA من المجموعات المحددة (خلايا من ستة فئران لكل مجموعة؛ ). قضبان القياس، . صور تمثيلية (يسار) وقياس (يمين) لـ Iba1الميكروغليا (الخضراء) تحتوي على GAD65/نقاط (حمراء) في CeA من المجموعات المحددةخلايا من ستة فئران لكل مجموعة؛ ). قضبان القياس، (نظرة عامة) و (الإدراج والرسم). تم تقييم الأهمية باستخدام تحليل التباين المتكرر ذو الاتجاهين مع المقارنة بعد الاختبار بين المجموعات في ( اختبار الطالب ذو الذيلين غير المتزاوج-اختبار في (ج). جميع البيانات مقدمة كمتوسطSEM.; ن.س.، غير دال. انظر أيضًا البيانات التكميلية 1. يتم تقديم بيانات المصدر كملف بيانات المصدر.
تشير هذه النتائج مجتمعة إلى أن الميكروغليا المنشطة يمكن أن تثبط GABAالنشاط العصبي من خلال آلية تغذية راجعة سلبية ظاهرة بعد الإجهاد الناتج عن القيود الحادة لتعزيز التعافي من سلوكيات مشابهة للقلق في المرحلة اللاحقة.
فرط نشاط GABAتقوم الخلايا العصبية بتنشيط الميكروغليا في منطقة اللوزة الدماغية لدى الفئران المعرضة للإجهاد الحاد
لفهم كيفية عمل GABA بشكل أفضلتستجيب الخلايا العصبية وخلايا الميكروغليا لمحفزات الإجهاد الحاد، قمنا بفحص GABAالنشاط العصبي وشكل الخلايا الدبقية الصغيرة في الفئران التي تعرضت للإجهاد الحاد الناتج عن ARS- 5 دقائق و ARS- 30 دقيقة. أظهرت النتائج أن النشاط التلقائي معدل إطلاق GABAكان عدد الخلايا العصبية قد زاد بشكل ملحوظ بعد 5 دقائق من القيود، وأن هذا الزيادة ظهرت أيضًا بعد 30 دقيقة من ضغط القيود مقارنة بالفئران الضابطة (الشكل التوضيحي 10 أ، ب). علاوة على ذلك، على الرغم من أن عددلم تتغير الميكروغليا في كل من الفئران ARS-5min أو ARS-30min مقارنةً بالتحكم، وكان طول الفروع ونقاط الميكروغليا أقل بشكل ملحوظ في ARS-30min، ولم تكن هذه التغيرات ملحوظة في الفئران ARS-5min مقارنةً بالتحكم (الشكل التوضيحي 10c، d). تشير هذه النتائج مجتمعة إلى أن تنشيطتسبق الخلايا العصبية تنشيط الخلايا الدبقية الصغيرة في منطقة اللوزة الدماغية لدى الفئران المعالجة بالإجهاد الناتج عن التقييد الحاد.
بعد ذلك، قمنا بمزيد من التحقيق فيما إذا كان GABAالنشاط المفرط للعصبونات مرتبط بشكل سببي بتنشيط الخلايا الدبقية الصغيرة في الفئران المعرضة للإجهاد الناتج عن القيود الحادة. وجدنا أن التنشيط الكيميائي الجيني لـ GABAالخلايا العصبية في الفئران الساذجة أدت إلى تنشيط الخلايا الدبقية الصغيرة، مما يدل عليه زيادة عددالميكروغليا وطول الفروع والنقاط المتناقصة في هذه الخلايا (الشكل التكميلي 11 أ، ب). بالإضافة إلى ذلك، فإن تثبيط الجابا الكيميائي الجينيألغت الخلايا العصبية تنشيط الميكروغليا الملحوظ في فئران ARS-2h (الشكل التكميلي 11c، d). مجتمعة، اقترحت هذه النتائج أن تنشيط الميكروغليا ناتج عن زيادة مستوى GABA.النشاط العصبي الناتج عن الضغط الناتج عن القيود الحادة في الفئران.
إشارات CX3CL1-CX3CR1 تساهم في انقراض سلوكيات مشابهة للقلق بعد الإجهاد الحاد
لاستكشاف آليات جزيئية محتملة من خلال تفعيل الخلايا الدبقية الصغيرة الناتج عن فرط نشاط GABAبعد الإجهاد الحاد، استخدمنا تقنية qPCR لفحص تعبير عدد من الجزيئات التي تم الإبلاغ عنها سابقًا بأنها متورطة في الاستجابة للإجهاد ولتسهيل التفاعلات بين الخلايا العصبية والميكروغليا.وجدنا أن مستويات mRNA لـ، و كانت أعلى بشكل ملحوظ في عينات أنسجة CeA المجمعة بعد 0.5 ساعة من العلاج من ARS-2h مقارنة بتلك الموجودة في الفئران الضابطة (الشكل التكميلي 12). من المعروف جيدًا أن CX3CL1/فراتالكين، وهو كيموكين مُفرز يُعبر عنه بشكل خاص في الخلايا العصبية ويشارك على نطاق واسع في تقليم المشابك المعتمد على النشاط، يمكن أن يتم تقطيعه لإنتاج شكل قابل للذوبان يحفز تنشيط الخلايا الدبقية من خلال الارتباط بمستقبلها، CX3CR1.لذلك، ركزنا على إشارة CX3CL1-CX3CR1 في تنظيم التفاعلات بين الخلايا العصبية والميكروغليا. كما هو متوقع، أظهرت اختبارات Western blot أن مستوى بروتين CX3CL1 القابل للذوبان كان بالفعل أعلى بشكل ملحوظ عند 0.5 ساعة و8 ساعات في فئران ARS-2h مقارنة بالفئران الضابطة (الشكل 4a، b).
للتحقيق فيما إذا كان هذا الزيادة في CX3CL1 القابل للذوبان في CeA مرتبطًا بتنظيم تنشيط الخلايا الدبقية الصغيرة، قمنا بحقن مضاد محدد لـ CX3CR1 وهو JMS-17-2 أو التحكم في المركب في فئران ARS-2h. تم تثبيط تنشيط الخلايا الدبقية الصغيرة، الذي تم الإشارة إليه بتعبير Ibal، في CeA بشكل ملحوظ في الفئران المعرضة للإجهاد الحاد مع المعالجة المسبقة بـ JMS-17-2 مقارنة بتلك التي تم إعطاؤها المركب (الشكل 4c، d). بالإضافة إلى ذلك، أوضحت صبغة المناعة الفلورية أن بلع الخلايا الدبقية الصغيرة لنقاط GAD65/67 في CeA انخفض بشكل ملحوظ في فئران ARS-2h المعالجة مسبقًا بمضاد CX3CR1 مقارنة بتلك المعالجة مسبقًا بالمركب (الشكل 4e، f). بعد ذلك، قمنا بفحص ما إذا كان التغيير في GABAتم التأثير أيضًا على كثافة الشوكات الشجرية العصبية بواسطة مضاد CX3CR1 في فئران ARS-2h. GABAأظهر التعبير المحدد للخلايا العصبية لفيروس AAV-CSSP-YFP-8E3 في فئران GAD2-Cre عدم وجود فرق في كثافة الشوكات الشجرية لهذه الخلايا العصبية بعد 0.5 ساعة من ARS-2 ساعة بين فئران النموذج المعالجة بـ JMS-17-2 والفئران المعالجة بالعقار الوهمي (الشكل. )، بينما كانت كثافة الشوكات أعلى بشكل واضح في هذه الخلايا العصبية بعد 12 ساعة من ARS-2h بعد المعالجة المسبقة بـ JMS-172 مقارنة بمجموعة المركب بعد الإجهاد الناتج عن التقييد الحاد (الشكل. ).
لفحص ما إذا كان هذا الـ CX3CL1 القابل للذوبان المعزز في CeA مسؤولاً عن سلوكيات القلق الشبيهة بتلك الناتجة عن ARS، قمنا بحقن JMS-17-2، أو التحكم الوهمي، بشكل ثنائي في CeA للفئران ARS-2h. وجدنا أن الفئران التي تم معالجتها مسبقًا بـ JMS-17-2 أظهرت سلوكيات استكشافية أقل بشكل ملحوظ من الفئران المعالجة بالتحكم الوهمي بعد 12 ساعة من ARS-2h، كما يتضح من عدد المدخلات الأقل والوقت الأقل الذي قضوه في المنطقة المركزية من OFT والأذرع المفتوحة من EPM (الشكل 4i، j)، والتي لوحظت أيضًا في كل من مجموعتي JMS-17-2 والتحكم الوهمي بعد 0.5 ساعة من ARS-2h (الشكل 4i، j). علاوة على ذلك، أظهرت تسجيلات متعددة الأقطاب في الفئران ARS-2h المتحركة بحرية عدم وجود فرق في GABA.نشاط إطلاق النار العفوي العصبي بين الفئران المعالجة بـ JMS-17-2 والفئران المعالجة بالعقار الوهمي بعد 0.5 ساعة من ARS-2h، وهو ما يتماشى مع نتائج الاختبارات السلوكية؛ ومع ذلك،كانت نشاط إطلاق الخلايا العصبية أعلى بعد 12 ساعة من تحفيز الإجهاد في الفئران التي تم حقنها بمادة JMS-17-2 في منطقة CeA مقارنةً بمجموعة التحكم التي تلقت محلولاً ملهماً (الشكل. ).
لاستكشاف كيف زاد mRNA الخاص بـ Cx3cl1 بواسطة ARS-2h، استخدمنا qPCR لفحصتعبير mRNA، الذي يعد عامل نسخ لتوسط تعبير السيتوكينات، بما في ذلك CX3CL1. من المثير للاهتمام، وجدنا أن تم زيادة مستويات mRNA بشكل ملحوظ في CeA بعد 0.5 ساعة من ARS-2h مقارنة بالفئران غير المجهدة (الشكل 4م). بعد ذلك، قمنا بإعطاء Pyrrolidinedithiocarbamate ammonium (PDTC)، وهو مثبط انتقائي لـ NF-kB، إلى CeA. في هذه الفئران، تم منع زيادة الوقت والدخول في المنطقة المركزية من OFT والأذرع المفتوحة من EPM بعد 12 ساعة من ARS-2h بعد إعطاء PDTC (الشكل 4ن، o). بالإضافة إلى ذلك، لوحظ انخفاض كبير في تعبير mRNA لـ Cx3cl1 في الفئران النموذجية المعالجة بـ PDTC بعد 0.5 ساعة من ARS-2h مقارنة بتلك في الحيوانات الضابطة (الشكل 4ص). تشير هذه النتائج إلى أن مسار إشارة NF-kB من المحتمل أن يكون متورطًا في تنظيم تعبير بروتين CX3CL1 في حالات الإجهاد الحاد.
تشير هذه النتائج معًا إلى أنه، خلال 12 ساعة بعد تحفيز الإجهاد، يمكن أن يعزز بروتين CX3CL1 القابل للذوبان المرتفع ابتلاع الخلايا الدبقية الصغيرة لجابا.الشوكات العصبية، مما يقلل من GABAالنشاط العصبي ودفع التعافي من سلوكيات مشابهة للقلق في الفئران النموذجية.
جاباإشارات MST4-NF-кB-CX3CL1 العصبية تبدأ تنشيط الميكروغليا الناتج عن الإجهاد الحاد
في ضوء الأدلة المذكورة أعلاه التي تدعم دور زيادة إشارات NF-кB-CX3CL1 الناتجة عن ARS في CeA، قمنا بعد ذلك بفحص الآليات المحتملة المسؤولة عن بدء هذه العملية في ARS. وقد تم الإبلاغ سابقًا عن أن الكيناز MST4 يقوم بشكل مباشر بفوسفرة TRAF6 (عامل مرتبط بمستقبل TNF 6)، مما يمنع يوبيلتينه، وبالتالي يحد من الاستجابة الالتهابية.. وبالتالي، تم اعتباره بمثابة “فرامل” على الاستجابات الالتهابية المعتمدة على TLR-TRAF6-NF-кB. لاختبار ما إذا كان MST4 يشارك في تنشيط الخلايا الدبقية في ARS، قمنا بإجراء صبغ مناعي فلوري لـ MST4 في الدماغ ووجدنا أنه موزع على نطاق واسع في عدة مناطق، بما في ذلك النواة المتكئة، والذيل المخطط، ونواة ما تحت المهاد الجانبية، وCeA (الشكل التكميلي 13، والشكل 5a). كما وجدنا أن MST4 يتواجد بشكل رئيسي مع الأجسام المضادة الخاصة بـ GABA، ولكن ليس مع الخلايا الدبقية، في CeA (الشكل التكميلي 14). أظهرت اختبارات qRT-PCR وWestern blot أن تعبير MST4 انخفض بشكل كبير على كل من مستوى mRNA والبروتين في CeA بعد 0.5 ساعة من ARS-2h مقارنة بالفئران غير المجهدة (الشكل 5b، c)، وعاد إلى مستوى التحكم بعد 12 ساعة من ARS-2h (الشكل 5c).
لمزيد من استكشاف وظيفة MST4 في تنظيم GABAالنشاط العصبي والسلوكيات الاستكشافية في الفئران، قمنا بعد ذلك بإنشاء ناقل AAV يعتمد على Cre لزيادة التعبير عن MST4 في GABAالخلايا العصبية لفئران GAD2-Cre. بعد ثلاثة أسابيع من حقن CeA الثنائي مع rAAV-Ef1-DIO-MST4-P2A-mCherry-WPRE-pA (AAV-MST4) أو rAAV-Efl-DIO-mCherry-WPRE-pA (AAV-mCherry) الفيروسات في فئران GAD2-Cre (الشكل 5d)، أظهرت المجهرية الفلورية أن المCherryكان الإشارة متواجدة بشكل كبير مع الإشارة من الأجسام المضادة الخاصة بـ GABA في CeA للفئران السليمة (الشكل 5e). أظهرت التحاليل الغربية تعبير MST4 بشكل أكبر بشكل ملحوظ في GABAالخلايا العصبية للفئران الساذجة المصابة بـ AAV-MST4 كانت أكثر من تلك الموجودة في الفئران الضابطة المصابة بـ AAV-mCherry (الشكل 5f). ثم قمنا بفحص سلوكيات الاستكشاف في فئران GAD2Cre التي تم حقنها إما بفيروسات AAV-MST4 أو AAV-mCherry (الشكل 5g) ووجدنا أن GABAزيادة التعبير عن MST4 المحدد للعصبونات، ولكن ليس التحكم AAV-mCherry، يمكن أن تؤدي إلى زيادة الوقت والدخول إلى المنطقة المركزية من اختبار الانفتاح (OFT) والأذرع المفتوحة من اختبار الأذرع المفتوحة (EPM) في فئران ARS-2h (الشكل 5h، i). بالإضافة إلى ذلك، GABAتم استعادة النشاط العصبي في فئران ARS المصابة بـ AAV-MST4 (الشكل 5j، k)، مصحوبًا بانخفاض مستويات mRNA لـ و في CeA (الشكل 5l)، مقارنةً بفئران ARS المصابة بـ AAV-mCherry. علاوة على ذلك، تم عكس تنشيط الخلايا الدبقية بشكل ملحوظ في CeA لفئران ARS التي تم حقنها بـ AAVMST4، مع عدد أقل بشكل ملحوظ من Iba1.الخلايا وزيادة الطول الكلي للفروع الميكروغليالية والنقاط مقارنةً بفئران ARS-2h مع تعبير محدد في CeA عن ناقل AAV-mCherry (الشكل 5م، ن).
الشكل 4 | حجب إشارة CX3CL1-CX3CR1 في CeA يمنع انقراض سلوكيات القلق بعد الإجهاد الحاد. صور تمثيلية (أ) وتحليلات كمية (ب) لعملية Western blot لـ CX3CL1 في CeA (فئران لكل مجموعة). ج صور تمثيلية لتلوين Iba1 المناعي وإعادة بناء ثلاثي الأبعاد للخلايا الدبقية الصغيرة في CeA. مقياس الأشرطة، (نظرة عامة) و (الإدراج والرسم). د أظهرت الفئران المعالجة بـ ARS تثبيطًا في تنشيط الخلايا الدبقية الصغيرة بعد المعالجة بـ JMS-17-2 (فئران لكل مجموعة). صور تمثيلية (هـ) وكمية (و) منالميكروغليا (الخضراء) تحتوي على GAD65نقاط (حمراء) في CeA خلايا من خمسة فئران لكل مجموعة). قضبان القياس، (نظرة عامة) و (الإدراج والرسم). صور تمثيلية للتغصنات العصبية (g) وقياس عدد الشوكات ( ) ( فئران لكل مجموعة؛، ). قضبان القياس، بيانات ملخصة للمدخلات والوقت المستغرق في المنطقة المركزية والأذرع المفتوحة لـ OFT (i) و EPM (j) (تركت،; صحيح، يسار; صحيح، ، ). مخططات الشبكة وآثار نموذجية ( ) والبيانات الكمية ( ) من الإطلاقات العفوية لـ الخلايا العصبيةخلايا من ستة فئران لكل مجموعة؛ ل، ). من في CeA (الفئران لكل مجموعة). البيانات الملخصة للدخول والوقت المستغرق في المنطقة المركزية والأذرع المفتوحة من اختبار التفضيل المفتوح (OFT) واختبار الأذرع المفتوحة (EPM) (فئران لكل مجموعة).تفاعل البوليميراز المتسلسل الكمي في CeA (فئران لكل مجموعة؛تم تقييم الأهمية باستخدام تحليل التباين المتكرر ذو الاتجاهين مع المقارنة بعد الاختبار بين المجموعات في (ب،اختبار الطالب ذو الذيلين غير المتزاوج-اختبار في ( ). جميع البيانات مقدمة كمتوسط SEM.; ن.س.، غير مهم. انظر أيضًا البيانات التكميلية 1. يتم توفير بيانات المصدر كملف بيانات المصدر.
الشكل 5 | الإفراط في التعبير عن MST4 في GABAتخفف الخلايا العصبية سلوكيات القلق الناتجة عن الإجهاد الحاد. أ. صبغة المناعة الفلورية لـ MST4 (أخضر) وNeuN (أحمر) في CeA. مقياس الأشرطة، (يسار) و (يمين). ب. تفاعل البوليميراز المتسلسل الكمي (qPCR) لـ Mst4 في CeA (فئران لكل مجموعة). ج تحليل Western blot لـ MST4 في CeA (فئران لكل مجموعة؛فئران لكل مجموعة؛, ). د توضيح تخطيطي لاستراتيجية بناء فرط التعبير عن MST4. هـ صور تمثيلية تظهر المCherry مع وسم مشترك لمضاد محدد لـ GABA. تم تكرار التجربة ثلاث مرات. شريط القياس، (يسار) و (يمين). تحليل Western blot لـ MST4 في CeA (فئران لكل مجموعة؛ ). رسم تخطيطي للإجراء التجريبي. بيانات ملخصة للمدخلات والوقت المستغرق في المنطقة المركزية والأذرع المفتوحة لـ OFT (س) و EPM (ع). مخططات راستر وآثار نموذجية (ي) والبيانات الكمية (ك) لإطلاق النار التلقائي لـ الخلايا العصبيةخلايا من ستة فئران لكل مجموعة؛
. تحليل qPCR لـ و في CeA من المجموعات المحددة (فئران لكل مجموعة). م صور تمثيلية لتلوين المناعية لـ lbal وإعادة بناء ثلاثية الأبعاد للميكروغليا في CeA. مقياس الأشرطة، (نظرة عامة) و (إدراج ورسم). أظهرت الفئران المعالجة بـ ARS تثبيطًا في تنشيط الميكروغليا بعد التعبير المفرط عن MST4 في CeAفئران لكل مجموعة). صور تمثيلية (o) وقياس (p) لـالميكروغليا (الخضراء) تحتوي على GAD65نقاط (أرجوانية) في CeA خلايا من ستة فئران لكل مجموعة). قضبان القياس، (نظرة عامة) و (الإدراج والرسم). تم تقييم الأهمية باستخدام تحليل التباين المتكرر ذو الاتجاهين مع المقارنة بعد الاختبار بين المجموعات في (ج)، اختبار الطالب غير المتزاوج ذو الاتجاهين.-اختبار في (ب، ف، ح، ي، ك، ل، ن، ب). جميع البيانات مقدمة كمتوسطSEM.، . انظر أيضًا البيانات التكميلية 1. يتم توفير بيانات المصدر كملف بيانات مصدر.
من الجدير بالذكر أن ابتلاع الخلايا الدبقية الصغيرة لنقاط GAD65/67 قد انخفض بشكل ملحوظ في الفئران التي تعرضت للإجهاد الحاد الناتج عن التقييد مع GABA.الخلايا العصبية المصابة بـ AAV-MST4 مقارنة بتلك المصابة بـ AAV-mCherry (الشكل 50، ص). يجب أيضًا الإشارة إلى أن الفئران الساذجة التي تعبر عن MST4 بشكل مفرط في GABAلم تظهر الخلايا العصبية أي تغييرات في سلوكيات الاستكشاف ولم تظهر أي فرق كبير في GABAنشاط الخلايا العصبية مقارنة بالفئران التي تعبر عن فيروس التحكم (الشكل التكميلي 15).
لاستكشاف تأثيرات إشارات MST4 على تنشيط الخلايا الدبقية الصغيرة في CeA، قمنا بعد ذلك بإنشاء ناقل AAV يعتمد على Cre يعبر عن RNAs قصيرة الشعر (shRNAs) لـ MST4 لتقليل تعبير MST4. بعد ثلاثة أسابيع من حقن CeA بـ rAAV-CMV-DIO-mCherry-U6- shRNA (MST4) (AAV-shMST4) أو rAAV-CMV-DIO-mCherry-U6-shRNA (خليط) (AAV-mCherry) في فئران GAD2-Cre غير المدربة (الشكل 6a)، أظهر تلوين المناعة الفلورية أن المCherryتمت تسمية الخلايا بشكل متكرر باستخدام جسم مضاد محدد لـ GABA في CeA (الشكل 6ب). أظهرت اختبارات qPCR وWestern blot أن مستويات mRNA لـ Mst4 ومستويات بروتين MST4 قد انخفضت بشكل كبير، بينما و مستويات وتعبير بروتين CX3CL1 زاد في GABAالخلايا العصبية، في الفئران التي تم تقليل MST4 فيها مقارنة بالفئران المصابة بعقار AAVmCherry (الشكل 6c-e). علاوة على ذلك، تم تنشيط الخلايا الدبقية وزيادة كبيرة في ابتلاع الخلايا الدبقية لـ GAD65/67 في الفئران السليمة مع حقن CeA لعقار AAV-shMST4 مقارنة بمجموعة التحكم AAVmCherry (الشكل 6f-i). هذه النتائج تشير إلى أن
الشكل 6 | تقليل MST4 في GABAتقوم الخلايا العصبية بتنشيط الميكروغليا وزيادة ابتلاع GABAالشوكات العصبية. أ. توضيح تخطيطي لاستراتيجية البناء المستخدمة لتقليل MST4. ب. صور تمثيلية تظهر تعبير المCherry في الخلايا العصبية مع وسم مشترك للأجسام المضادة الخاصة بـ GABA. تم تكرار هذه التجارب ثلاث مرات. شريط القياس، (يسار) و (يمين). تحليل qPCR للتعبير عن mRNA لـ Mst4 و Nf-kb و Cx3cl1 في CeA للفئران السليمة المصابة بـ AAV-mCherry و AAV-shMST4 (mCherry، فئران لكل مجموعة، shMST4،فئران لكل مجموعة).تحليل Western blot لبروتين MST4 في CeA للفئران الساذجة المصابة بـ AAV-mCherry و AAV-shMST4فئران لكل مجموعة؛تحليل Western blot لبروتين CX3CL1 في CeA للفئران السليمة المصابة بـ AAV-mCherry و AAV-shMST4فئران لكل مجموعة).صور تمثيلية لتلوين Iba1 المناعي وثلاثي الأبعاد
إعادة بناء الميكروغليا في CeA للفئران الساذجة المصابة بـ AAV-mCherry و AAV-shMST4. مقياس الأشرطة، (نظرة عامة) و (إدراج ورسم). ج قياس أعداد الخلايا، الطول الإجمالي للعملية وعدد نقاط التفرع للميكروغليا في CeA من الفئران السليمة المصابة بـ AAV-mCherry و AAV-shMST4فئران لكل مجموعة). صور تمثيلية (هـ) وقياس (ط) لـالميكروغليا (الخضراء) تحتوي على GAD65/67نقاط (بنفسجي) في CeA من الفئران الساذجة المصابة بـ AAV-mCherry و AAV-shMST4 خلايا من ستة فئران لكل مجموعة). قضبان القياس، (نظرة عامة) و (الإدراج والرسم). تم تقييم الأهمية بواسطة اختبار الطالب غير المتزاوج ذو الطرفين.-اختبار في ( ). جميع البيانات مقدمة كمتوسط SEM.. انظر أيضًا البيانات التكميلية 1. يتم توفير بيانات المصدر كملف بيانات مصدر. يمكن أن يؤدي تقليل تنظيم MST4 مباشرة إلى تنشيط الخلايا الدبقية وزيادة ابتلاع GABAالشوكات العصبية.
تشير هذه النتائج التراكمية إلى أن انخفاض تعبير MST4 بعد الإجهاد الناتج عن القيود الحادة يؤدي إلى تنشيط إشارة NF-кВCX3CL1 في GABA.الخلايا العصبية، التي بدورها تعزز تنشيط الخلايا الدبقية الصغيرة وابتلاع GABAالشوكات العصبية، مما يؤدي إلى استعادة GABAفرط النشاط العصبي وانقراض السلوكيات الشبيهة بالقلق.
نقاش
تشير نتائج الدراسة الحالية إلى أن السلوكيات الشبيهة بالقلق التي تسببها ضغوط التقييد الحادة تختفي بعد 12 ساعة من بدء الضغط في الفئران الذكور وأن نشاط GABAتزداد الخلايا العصبية ولكنها تعود إلى مستويات غير متوترة. يتم تنظيم هذه العملية من خلال تنشيط الخلايا الدبقية الصغيرة في المنطقة المركزية من اللوزة وزيادة ابتلاع الخلايا الدبقية الصغيرة لـ GABA.الشوكات العصبية في الفئران المعرضة للإجهاد الحاد. تكشف نتائجنا أيضًا أن إشارة MST4-NF-кB-CX3CL1 في GABAتقوم الخلايا العصبية بوساطة تنشيط الخلايا الدبقية الصغيرة (الشكل التكميلي 16). تشير هذه النتائج التراكمية إلى أن آلية التغذية الراجعة السلبية المدفوعة بالخلايا الدبقية الصغيرة، التي تعمل بطريقة مشابهة للخلايا العصبية المثبطة، مهمة لحماية الدماغ من التنشيط المفرط للخلايا العصبية في حالات الصحة أو المرض.
يرتبط الإجهاد الحاد أو المزمن بزيادة خطر العديد من الاضطرابات النفسية، بما في ذلك اضطرابات القلق والاكتئاب.نظرًا لأن CeA هو النواة الرئيسية لإخراج المعلومات في اللوزة، فقد أفادت العديد من الدراسات أن فرط نشاط CeA بعد التعرض للإجهاد أو أحداث الضغط يساهم في مسببات القلق.؛ هذه النتائج تتماشى مع نتائجنا. بالمقابل، هنا، استكشفنا المزيد من العلاقة الزمنية بين GABAالنشاط العصبي وبداية وانقراض سلوكيات مشابهة للقلق بعد الإجهاد الناتج عن التقييد الحاد في الفئران. ومن الجدير بالذكر أنه بالإضافة إلى فرط نشاط GABAتمت ملاحظة تنشيط الخلايا الدبقية الصغيرة في النواة المركزية (CeA) أيضًا في الفئران خلال علاج الضغط الحاد الناتج عن التقييد، والذي كان يعتمد على تنشيط GABA.الخلايا العصبية بسبب تثبيط كيميائي وراثي لهذه الخلايا العصبية مما يعكس تنشيطها.
أظهرت دراسات متعددة أن تنشيط الخلايا الدبقية الصغيرة، وهي خلايا مراقبة المناعة في الدماغ، يشارك في الوساطة للاستجابات لمختلف الضغوط التي يمكن أن تكون محفزًا رئيسيًا للعديد من الاضطرابات النفسية العصبية.في الواقع، يمكن تنشيط الخلايا الدبقية الصغيرة بسرعة عندما يتعرض الجسم للإجهاد الحاد، كما يتضح من تغير الشكل، وزيادة القدرة البلعومية، وتغير الملف النسخي، للكشف عن التغيرات في النشاط العصبي في الوقت الحقيقي وتعزيز استعادة توازن الدماغ.ومع ذلك، غالبًا ما تتعرض وظيفة المناعة للخلايا الدبقية الصغيرة للخلل بسبب الإجهاد المزمن، مما يؤدي إلى الإفراط في إنتاج الوسائط الالتهابية وزيادة النشاط البلعومي، وهو ما يمكن أن يكون ضارًا لإعادة تشكيل الشبكة العصبية، مما يؤدي إلى تفاقم سلوكيات المرض.تظهر الدراسة الحالية أن الخلايا الدبقية الصغيرة في المنطقة المركزية من اللوزة (CeA) تكون مفعلة في الفئران التي تعاني من سلوكيات قلقية ناتجة عن ضغط الحبس الحاد، مصحوبة بزيادة في ابتلاع GABA.الشوكات الشجرية العصبية ومناطق الاتصال الأكبر مع هذه الخلايا العصبية. علاوة على ذلك، فإن تثبيط النشاط الميكروغليالي عن طريق الأدوية قبل الإجهاد الحاد لا يمنع ظهور سلوكيات مشابهة للقلق ولكنه يمنع تخفيف هذه الظاهرة في الفئران، مما يشير إلى أن تنشيط الميكروغليا في CeA هو المسؤول عن عملية تخفيف سلوكيات القلق الناتجة عن الإجهاد الحاد بدلاً من التوسط في حدوثها. وبالتالي، فإن الميكروغليا تستجيب إلىتنشيط الخلايا العصبية ثم دفع آلية التغذية الراجعة السلبية مشابهة لتلك الخاصة بالخلايا العصبية المثبطة، والتي تعتبر ضرورية لحماية الدماغ من التنشيط المفرط في الصحة والمرض.هناك بعض القيود المحتملة لفحص الانغماس المشبكي في دراستنا. على سبيل المثال، فشلنا في تقديم دليل مباشر لإثبات خصوصية جسم مضاد GAD65/67، على الرغم من أنه قد تم استخدامه على نطاق واسع في صبغ المناعية الفلورية. في العديد من الدراساتبالإضافة إلى ذلك، يمكن على الأرجح اكتشاف الفلورية الذاتية للليبوفوسين داخل الليزوزومات الميكروغليالية في دماغ الفأر البالغ بواسطة المجهر الضوئي.. لمعالجة هذه القضية العامة في هذا المجال، استخدمنا نفس الفئران المسنّة كضوابط مقابلة للكشف عن ابتلاع الخلايا الدبقية الصغيرة، والتي كانت قادرة على تقليل هذا التأثير قدر الإمكان.
بالإضافة إلى الخلايا الدبقية الصغيرة، تم الإشارة تاريخياً إلى أن الخلايا النجمية توفر الدعم الهيكلي والتمثيلي العصبي، وقد تم الإشارة أيضاً إلى أنها متورطة في تقليم المشابك. ومع ذلك، عادةً ما تبتلع الخلايا النجمية وتزيل المشابك خلال التطور وفي ظروف مثل مرض الزهايمر والصرع، ولا يوجد حالياً دليل يدعم دور الخلايا النجمية في تقليم المشابك استجابةً للمؤثرات المجهدة. تُعرف الخلايا الدبقية قليلة التغصن، وهي نوع آخر مهم من الخلايا الدبقية، بأنها مسؤولة بشكل أساسي عن تنظيم نمو المحاور، وإنتاج وصيانة الميالين. على الرغم من أن خلايا سلف الخلايا الدبقية قليلة التغصن تبتلع المشابك خلال إعادة تشكيل الدوائر في الفئران، إلا أن الخلايا الدبقية قليلة التغصن الناضجة ليست متورطة بشكل مباشر في تقليم المشابك. بناءً على الاستجابة السريعة للخلايا الدبقية الصغيرة للإجهاد الحاد وخصائصها في تقليم المشابك، من المحتمل أن تلعب الخلايا الدبقية الصغيرة دوراً مهيمناً في تنظيم تلاشي سلوكيات القلق الناتجة عن الإجهاد الحاد. تحتاج وظيفة الخلايا النجمية والخلايا الدبقية قليلة التغصن تحت ظروف الإجهاد الحاد إلى مزيد من التحقيق.
مسار التفاعل الكلاسيكي بين الخلايا العصبية والخلايا الدبقية الصغيرة هو تنظيم الفراكالكين المشتق من الخلايا العصبية (CX3CL1) لتنشيط الخلايا الدبقية الصغيرة بواسطة ارتباط CX3CR1 على الخلايا الدبقية الصغيرة. ومع ذلك، تظل المسارات الإشارية المسؤولة عن التحكم في إفراز CX3CL1 من الخلايا العصبية غير مفهومة جيداً. في التحقيق في الإشارات العليا التي تتوسط تعبير CX3CL1، حددنا كيناز عائلة MST4 من عائلة كيناز مركز الجنين III، وهو كيناز سيرين-ثريونين معبر عنه بشكل كبير في المشيمة، والغدة الصعترية، واللمفاويات، وكريات الدم البيضاء في الدم المحيطي. MST4 هو مكون من مجمع “الفوسفاتاز وكيناز المتفاعل مع النواة”، والذي يعمل في مجموعة واسعة من العمليات الأساسية، مثل نقل الإشارات، والتحكم في دورة الخلية، والموت الخلوي، ونقل الحويصلات، وهجرة الخلايا.
تقدم دراستنا دليلاً يظهر أن MST4 موزع على نطاق واسع في مناطق الدماغ المسؤولة عن العمليات العاطفية. بشكل أكثر تحديداً، ينخفض MST4 في CeA، ويمكن أن يخفف فرط نشاط GABA العصبي، مما يعكس سلوكيات القلق، في فئران ARS. وجدنا أيضاً أن عامل النسخ المؤيد للالتهاب NF-кВ ينظم نسخ Cx3cl1 في GABA الخلايا العصبية، مما يؤدي إلى تنشيط الخلايا الدبقية الصغيرة عبر الارتباط بـ CX3CR1. يمنع الإعطاء المحلي لمضاد مستقبل CX3CR1 JMS-17-2 الانقراض التدريجي لسلوكيات القلق، والذي يحدث خلال 12 ساعة بعد تحفيز الإجهاد في الفئران النموذجية، مما يؤدي في النهاية إلى استمرار نشاط GABA العصبي العالي. من الجدير بالذكر أن فرط تعبير MST4 يمكن أن يثبط مباشرة إفراز CX3CL1 من خلال مسار إشارة MST4-NF-кBCX3CL1 الذي يتوسط تفاعلات الخلايا العصبية والخلايا الدبقية الصغيرة في المرحلة المبكرة من الإجهاد الحاد لزيادة تنشيط الخلايا الدبقية الصغيرة وابتلاع الشوكات العصبية GABA، مما يؤدي بالتالي إلى تثبيط نشاط GABA العصبي في فئران ARS. توفر هذه الدراسة دليلاً أولياً يدعم دور العامل المضاد للالتهاب MST4، المعبر عنه بشكل خاص في GABA الخلايا العصبية، في تحفيز إفراز الفراكالكين لتنشيط خلايا الدبقية الصغيرة في CeA بعد الإجهاد الحاد.
تظهر هذه النتائج الجماعية الأساس العصبي على المستوى الجزيئي والخلوي الذي يكمن وراء انقراض سلوكيات القلق بعد الإجهاد الحاد. تتضمن هذه العملية الراجعة المثبطة إشارات مؤيدة للالتهاب الناتجة عن الإجهاد الحاد بواسطة GABA الخلايا العصبية لتنشيط الخلايا الدبقية الصغيرة، والتي بدورها تخفف من النشاط العصبي عبر ابتلاع الشوكات العصبية لتقليل سلوكيات القلق خلال 12 ساعة بعد تحفيز الإجهاد. تشير أبحاثنا بالتالي إلى أن تنشيط الخلايا الدبقية الصغيرة في المرحلة المبكرة من الإجهاد الحاد يمكن أن يلعب دوراً وقائياً في تنظيم النشاط العصبي، وهو مختلف عن الدور المؤيد للألم للإجهاد المزمن الموصوف في الدراسات السابقة.
طرق
الحيوانات
تمت الموافقة على جميع التجارب التي أجريت من قبل لجنة الرعاية في جامعة العلوم والتكنولوجيا في الصين (USTC) (USTCACUC25120123086). دون تعليمات خاصة، كانت الفئران الذكور من سلالة C57BL/6J، Ai14 (RCL-tdT)، Cx3cr1-GFP، وGAD2-Cre تتراوح أعمارها بين 8-10 أسابيع. تم الحصول على الحيوانات من تشارلز ريفر أو مختبر جاكسون وتم تربيتها في منشأة الحيوانات في USTC. كانت الفئران تعيش في مجموعات مع خمسة في كل قفص في مستعمرة في بيئة مستقرة درجة الحرارة المحيطة و الرطوبة) ما لم يتم زرع قنية أو مجموعة أقطاب كهربائية. تم الحفاظ عليها تحت دورة ضوء-ظلام مدتها 12 ساعة (الأضواء مشتعلة من 7:00 إلى 19:00) مع توفر الماء والطعام بحرية.
إجهاد التقييد الحاد
تم immobilizing الفئران في حقن بلاستيكية معدلة مرة واحدة لمدة ساعتين دون تعليمات خاصة. تم حفر ثقوب في نهايات الحقن للسماح للفئران بالتنفس. خلال فترة التقييد، تم السماح للفئران الضابطة بالتحرك بحرية في القفص دون توفير الماء أو الطعام. تم إجراء التقييد الحاد في الفئران بين الساعة 8:00 و10:00 صباحاً، وتم قياس التجارب السلوكية لاحقاً بعد 0.5 ساعة،، و12 ساعة بعد علاج الإجهاد.
التجارب السلوكية
للاختبار السلوكي، تم نقل الفئران إلى غرفة الاختبار قبل يوم واحد من الاختبار للتأقلم. تم استخدام دفعات مختلفة من الفئران لاختبار الحقل المفتوح (OFT) واختبارات المتاهة المرتفعة (EPM)، من أجل استبعاد تأثيرات الاختبار السابق على الاختبار الحالي. خلال جلسة الاختبار، تم تسجيل مسارات الحركة وتحليلها لاحقاً بشكل غير متصل باستخدام برنامج EthoVision XT (Noldus). تم حساب المدخلات والوقت الذي قضاه في مركز الحقل المفتوح أو الأذرع المفتوحة للمتاهة المرتفعة.
بالنسبة لـ OFT، تم استخدام جهاز حقل مفتوح يتكون من منطقة مربعة () ومنطقة هامشية () لاختبار القلق. تم وضع الفئران بسرعة في المنطقة المركزية من صندوق الاختبار وسمح لها بالتحرك بحرية. تم استخدام برنامج تحليل سلوك الحيوان لتسجيل نشاط الفئران في الصندوق، وعادةً ما كانت مدة التجربة 15 دقيقة. تم تنظيف الجدار الداخلي وقاع غرفة الاختبار باستخدام الإيثانول بين الاختبارات لإزالة تأثير الرائحة. تم تحليل عدد المرات التي دخلت فيها الفأر المنطقة المركزية وإجمالي الوقت الذي قضته في المنطقة المركزية باستخدام برنامج تحليل سلوك الحيوان.
بالنسبة لـ EPM، كان الجهاز يتكون من منصة مركزية () وذراعين مغلقين () وذراعين مفتوحين متقابلين. كانت المتاهة بارتفاع 100 سم عن الأرض. تم وضع الفئران في المنطقة المركزية من المتاهة، مع توجيه رؤوسها نحو الأذرع المفتوحة، وسمح لها بالاستكشاف بحرية. تم تحليل مسارات الحركة بشكل غير متصل، وتم حساب الوقت الذي قضته في الأذرع المفتوحة وعدد المدخلات إلى الأذرع المفتوحة بعد التسجيل. تم تنظيف الجهاز بين الاختبارات باستخدام 75% إيثانول.
تسجيلات الأقطاب الكهربائية متعددة القنوات في الجسم الحي
تم الاحتفاظ بالفئران التي سيتم زرع مجموعة أقطاب كهربائية فيها بمفردها. بعد ثلاثة أيام من تأقلم الفئران مع البيئة، تم زرع أربع مجموعات أقطاب كهربائية متحركة مصنوعة خصيصاً في CeA. تم استخدام محركات ميكروية مدفوعة بالبراغي محلية الصنع مع أربعة أقطاب كهربائية قابلة للزراعة لتسجيل الإشارات من عدة خلايا عصبية في وقت واحد. تم تسجيل الإشارات بعد أسبوعين على الأقل من التعافي من الجراحة، وتم تأقلم الفئران مع الكابلات المتصلة بالأقطاب الكهربائية على رؤوسها قبل التسجيل. تم وضع الفئران في صندوق أسطواني مغطى بشبكة نحاسية وسمح لها بالتحرك بحرية دون أي إزعاج، وتم تسجيل الإشارات الكهربائية متعددة القنوات طوال هذه الفترة. أخيراً، تم تدوير البرغي في حامل الأقطاب الكهربائية عكس اتجاه عقارب الساعة، وتم خفض الأقطاب الكهربائية تدريجياً بواسطة بعد كل اختبار تسجيل يومي. تم تضخيم الإشارات العصبية، وتم تصفيتها عند عرض النطاق، وتم تخزينه باستخدام برنامج NeuronStudio. ثم تم تصدير البيانات إلى Offline Sorter 4 (Plexon) و NeuroExplorer 4 (NexTechnologies) للتحليل غير المتصل. تم استبعاد الوحدات التي كانت نسبة الإشارة إلى الضوضاء أقل من 2 من التحليل. تم إجراء فرز النبضات باستخدام طريقة فرز تتضمن خوارزمية T-Dis E-M المدمجة في Offline Sorter 4. تم حساب معدلات إطلاق النار للوحدات المفروزة باستخدام Neuroexplorer 4.
لتحديد GABAالخلايا العصبية، نبضات الضوء الأزرق (تم توصيل (التردد 20 هرتز) في نهاية كل جلسة تسجيل بتردد عالٍ. فقط النبضات المستحثة بالليزر والنبضات العفوية ذات الأشكال الموجية المتشابهة للغاية (معامل الارتباط ) اعتُبرت ناتجة عن خلية عصبية واحدة. في التجارب اللاحقة، قمنا بتصنيف الوحدات المعزولة جيدًا وفقًا لنمط إطلاق النار النموذجي للخلايا العصبية GABAergic المستحثة بالضوء باستخدام خوارزمية تجميع غير خاضعة للإشراف تعتمد على -يعني الطريقةعلى وجه التحديد، تم تصنيف النبضات ذات عرض نصف النبضة ونصف الوادي الأقصر ومعدل إطلاق النار الأعلى على أنها خلايا عصبية محتملة من نوع GABAergic.
علم المناعة النسيجية، التصوير، وتحليل الصور
تم تخدير الفئران بشكل عميق باستخدام الإيزوفلوران (حوالي 15 ثانية) ثم تم غسلها بسرعة بمحلول ملحي لمدة 3 دقائق.محلول بارافورمالدهيد لمدة 4 دقائق. تم إزالة الدماغ ووضعه فيمحلول بارافورمالدهيد ثم تم تخزينه فيثلاجة للتثبيت طوال الليل. ثم استخدمنا و محلول السكروز ووضع الدماغ فيثلاجة لتجفيفها حتى تصبح متساوية التوتر. بعد تضمين نسيج الدماغ بوسيلة تضمين عندتم قطع نسيج الدماغ المدمج إلىشرائح كورونالية باستخدام نظام ميكروتوم كريوستات (CM1860، لايكا). تم غمر شرائح الدماغ في محلول مضاد للتجمد وتخزينها فيللتلوين، تم غسل شرائح الدماغ التي تحتوي على CeA بمحلول PBS لمدة 5 دقائق ثلاث مرات. تم إغلاق شرائح الدماغ في محلول PBS يحتوي على 5% BSA و5% Triton X-100 في درجة حرارة الغرفة لمدة ساعة واحدة. ثم تم تحضين شرائح الدماغ في الأجسام المضادة الأولية (البيانات التكميلية 2). بعد ذلك، تم تحضين شرائح الدماغ معترايتون X-100 و مصل الحصان في الأجسام المضادة الأولية (البيانات التكميلية 2) عندبينما كانت الشرائح مغمورة طوال الليل. بعد ذلك، تم غسل الأقسام ثلاث مرات بمحلول PBS لمدة 5 دقائق وتمت معالجتها بالأجسام المضادة الثانوية (البيانات التكميلية 2) لمدة 1.5 ساعة في درجة حرارة الغرفة في مكان مظلم. أخيرًا، تم صبغ جميع شرائح الدماغ بـ DAPI (رقم الكاتالوج D9542، سيغما-ألدريش) باستخدام محلول التخفيف (1:2000) لمدة 3 دقائق وغسلها مرتين. تم استخدام DAPI لتعليم النوى، وتم تثبيت جميع الأقسام بوسيط تثبيت مضاد للتلاشي (رقم الكاتالوج H-1000، فيكتور). تم مسح الشرائح وتصويرها باستخدام المجاهر (Zeiss LSM880، Zeiss LSM980، Olympus SpinSR، وOlympus FV3000) لرؤية إشارات الفلورسنت.
إعادة بناء ثلاثية الأبعاد
التم صبغ الشرائح بمضاد Ibal لمدة 24 ساعة، تلاها صبغ بمضاد ثانوي مرتبط بألكسا فلور 488 أو مضاد ثانوي مرتبط بألكسا فلور 643. تم إجراء التصوير باستخدام مجهر زيس LSM880 أو زيس LSM980 باستخدامكان الهدف من زيت NA، ومعلمات التصوير (قوة الليزر، الكسب، والانحراف) متسقة عبر جميع التجارب. تم إجراء تكديس Z بقيمة 1.0.خطوات في الاتجاه، و تم تحليل صور بدقة بكسل باستخدام برنامج IMARIS 9.6.2 (Bitplane). تم استخدام وظيفة IMARIS “Fliaments” لقياس طول العمليات المجهرية وعدد نقاط التفرع. تم استخدام كل فأر على حدة كعينات مستقلة.
تم تحليل ابتلاع الخلايا الدبقية الصغيرة لإنشاء عرض سطحي ثلاثي الأبعاد للخلايا الدبقية الصغيرة، مع تحديد عتبة لضمان إعادة بناء دقيقة لعمليات الخلايا الدبقية الصغيرة، والتي تم استخدامها بعد ذلك لإعادة البناء اللاحقة. تم استخدام وظيفة “النقاط” لإعادة بناء النقاط GAD65/67. أخيرًا، تم استخدام المكون الإضافي المستند إلى MATLAB من IMARIS (“تقسيم إلى كائنات سطحية”) لتقييم عدد النقاط GAD65/67 الموجودة بالكامل داخل سطح الخلايا الدبقية الصغيرة (المسافة).شريحتان تم اختيارهما عشوائيًا من تم تصوير كل فأر وقياسه لخمسة أو ستة فئران في كل مجموعة. تم استخدام النتيجة المتوسطة للتحليل الشكلي.
لتحليل عمليات الخلايا الدبقية الصغيرة واتصالات الشجيرات العصبية، تم إجراء التصوير باستخدام ميكروسكوب زيس LSM880. تم استخدام وظيفة “السطح” لإعادة بناء كل من lba1 بدقة.الميكروغليا وتم استخدام الشجيرات، وتم استخدام العتبات التي تم تحديدها في البداية لإعادة البناء التالية. أخيرًا، تم استخدام المكون الإضافي IMARIS المستند إلى MATLAB “مساحة الاتصال بين السطحين” لقياس حجم مناطق الاتصال بين عمليات الخلايا الدبقية الصغيرة والشجيرات العصبية.
تدفق الخلايا
تم تخدير فئران Cx3cr1-GFP بشكل عميق باستخدام بنتوباربيتال الصوديوم.، أي.ب.). بعد ذلك، تم ضخ الفئران داخل القلب بـ 20 مل من محلول هانك المتوازن للملح (HBSS)، تلاه جمع سريع للـ CeA الثنائي، الذي تم غسله بمحلول PBS بارد وقطعه إلى قطع صغيرة على الثلج. تم تجانس الأنسجة الصغيرة ميكانيكياً باستخدام إبرة 23G، مما أدى إلى إنتاج تعليقات خلوية تم تصفيتها من خلال منخل الخلايا. بعد ذلك، تم تصفية العينات وطردها مركزيًا عندتم إعادة تعليق الكريات فيمحلول بيركول ودوّرت في جهاز الطرد المركزي عند، 10 دقائق). بعد إزالة السائل العلوي بعناية، تم شطف الكريات باستخدام PBS. أخيرًا، تم قياس الخلايا باستخدام جهاز قياس التدفق CytoFLEX (بيكمان كولتر، الولايات المتحدة الأمريكية) وتم تحليل البيانات بواسطة FlowJo V10. GFPتم فرز الخلايا بواسطة BD FACSAria III (BD، الولايات المتحدة الأمريكية) لإجراء تجارب إضافية: التحقيق في تعبير الجينات لـ و عن طريق qPCR (أبلايد بايوسستمز، ثيرمو فيشر، الصين).
إدارة الأدوية
تم إذابة هيدروكلوريد المينوسكلين (مينو) (رقم الكاتالوج M9511، سيغما-ألدريتش) في الماء )، الذي تم إعطاؤه عن طريق الحقن داخل الصفاق مع للتسريب المجهري داخل الجمجمة، تم زرع القنية الموجهة (قطر داخلي 0.35 مم، RWD) عادةً 0.2 مم أعلى من النواة المستهدفة في CeA في الفئران قبل أسبوعين من اختبارات السلوك لضمان وقت كافٍ للتعافي. تم توصيل تركيز 40 مليمول من مينو مخفف في السائل الدماغي الشوكي الاصطناعي (ACSF) باستخدام حقنة واحدة عن طريق إدخال القنية الداخلية، التي كانت أعمق بمقدار 0.2 مم من القنية الموجهة، وتم تطبيق ACSF (250 نانولتر) محليًا كتحكم.
مضاد CX3CR1 JMS-17-2 (رقم الكاتالوج HY-123918، ميد كيم إكسبريس) تم إذابته في زيت الذرة )، الذي تم إعطاؤه عن طريق حقن داخل الصفاق مع تم تخفيف تركيز 10 مللي مولار من JMS-17-2 في زيت الذرة وتم حقنه ميكرويًا داخل الجمجمة في المنطقة الثنائية من CeA، وتم تطبيق محلول المركب (زيت الذرة، 150 نانولتر) محليًا كتحكم.
تم إذابة كلوزابين-N-أكسيد (CNO) (رقم الكاتالوج C0832، سيغما-ألدريتش) في DMSO ) كمحلول مخزون ومخفف في محلول ملحي لتجارب DREADDs، حيث تم حقنه (30 دقيقة قبل الاختبارات السلوكية.
تم إذابة ثيوكاربامات البيروليدين الأمونيوم (PDTC) (رقم الكاتالوج HY-18738، ميد كيم إكسبريس) في محلول فوسفات البفر. تم توصيل تركيز 50 مللي مولار من PDTC إلى المنطقة المركزية من اللوزة الثنائية، وتم تطبيق محلول المركب (ACSF، 150 نانولتر) محليًا كتحكم.
الجراحة الستيريوتاكسية
جميع إجراءات حقن الفيروسات اتبعت إرشادات السلامة الحيوية في المختبر المعتمدة من قبل USTC. قبل الجراحة، تم تثبيت الفئران في إطار ستيريوتاكتيكي (RWD) تحت حقن داخل الصفاق من البنتوباربيتال الصوديوم.تم الحفاظ على درجة حرارة جسم الفئران عندباستخدام وسادة تسخين. تم ملء ميكروإلكترود زجاجي مسحوب بالفيروس وتم توصيله بحقنة سعة 10 ميكرولتر. كان حجم الحقن للفيروسات المختلفة يتراوح من 100 إلى 300 نانولتر اعتمادًا على العيار الفيروسي وإمكانات التعبير، وكانت سرعة الحقن هيتم تضمين الإحداثيات كوسيط جانبي (ML) من الخط الأوسط (بالمليمتر)، أمامي-خلفي (AP) من البريغما، وعمودي-أفقي (DV) من السطح السحائي للدماغ.
للتلاعب الكيميائي الجيني، rAAV-Ef1-DIO-hM3D(Gq)-mCherry-WPRE-pA (AAV-DIO-hM3Dq-mCherry, AAV2/فيروس Cat# PT-0042، BrainVTA) أو rAAV-Ef1-DIO-hM4D(Gi)-mCherry-WPRE-pA (AAV-DIO-hM4Di-mCherry، AAV2/9،، رقم الكاتالوج PT-0043، BrainVTA) تم حقنه ثنائي الجانب في CeA لفئران GAD2-Cre. بعد ثلاثة أسابيع من حقن الفيروس، تم حقن CNO عن طريق البطن (Sigma)، وهو ربيطة اصطناعية لمستقبلات الماسكارين المهندسة البشرية التي ترتبط وتنشط hM3Dq و hM4Di من أجل التلاعب الكيميائي الوراثي بالنشاط العصبيتم إعطاؤه 30 دقيقة قبل الاختبارات السلوكية. الـ rAAV-Ef1-DIO-mCherry-WPRE-pA (AAV-DIO-mCherry، AAV2/9،تم استخدام فيروس Cat# PT0013، BrainVTA) كعنصر تحكم.
لتحديد أشواك الدندrites العصبية للخلايا العصبية GABAergic بشكل محدد، تم استخدام وسم نادر خاص بنوع الخلايا عن طريق حقن فيروس AAV-sparse-CSSP-YFP-8E3 (AAV-CSSP-YFP-8E3، Cat# BC-SL003، BrainCase) في CeA في فئران GAD2-Cre التي تبلغ من العمر 5 أسابيع. يتم إنتاج فيروس AAV-CSSP-YFP-8E3 عن طريق التعبئة المشتركة لبلازميد AAV-CMV-DIO-Flp وبلازميد AAV-FDIO-EYFP بنسبة 1:8E3 في خطوة إنتاج rAAV واحدة (إجمالي عيار الفيروس ).
لزيادة التعبير وانخفاض التعبير عن MST4 في GABAالخلايا العصبية، الرAAV-Efl-DIO-MST4-P2A-mCherry-WPRE-pA (AAV-MST4، AAV2/9،فيروسات و rAAV-CMV-DIO-mCherry-U6-shRNA (MST4) (AAV-shMST4، AAV2/9،تم استخدام الفيروسات للتقليل من التعبير الجيني والتعبير الزائد، على التوالي. فيروس AAV-DIOmCherry و rAAV-CMV-DIO-mCherry-U6-shRNA (خليط) (AAV-scramble، AAV2/9،، رقم القطعة PT-2788، تم استخدام فيروس BrainVTA كضوابط. تم تأكيد إحداثيات موقع الحقن في الفئران بعد حقن صبغة نيسل. كانت الإحداثيات النهائية بالنسبة للبريغما كما يلي: AP، -1.2 مم؛ ML،; DV، -4.00 مم .
تسجيلات الكهربية الفسيولوجية
لتحضير شرائح الدماغ، تم تخدير الفئران باستخدام الإيزوفلوران (حوالي 15 ثانية) تلاه البنتوباربيتال الصوديوم.، أي.ب.) وبعد ذلك تم ضخها داخل القلب بـ 20 مل من N-methyl-D-glucamine (NMDG) المبردة والمحتوية على الأكسجين وACSF التي تحتوي على 93 NMDG، -2-هيدروكسي إيثيل بيبيرازين-N-2 إيثان سلفونيك أسيد (HEPES)، 25 جلوكوز، 2 ثيو يوريا، 5 ناتريوم أسكوربات، 3 ناتريوم بايروفات،، و 3 جلاوتاثيون (GSH) (بالمليمول، الرقم الهيدروجيني: 7.3-7.4، الأسمولالية: 300-305 مليمول/كجم). بعد عملية التروية، شرائح الدماغ التي تحتوي على CeA تم قطعها بسرعة بواسطة مبردنموذج NMDG ACSF على ميكروتوم (VT1200s، لايكا، ألمانيا) يهتز عندالسرعة. تم تحضين شرائح الدماغ بسرعة في محلول NMDG ACSF المؤكسج (المشبع بـلتوفير درجة حموضة مستقرة وأكسجة مستمرةفيثم تم حضنه في محلول HEPES ACSF يحتوي علىهيبز25 جلوكوز، 2 ثيويوريا،5 نازكوربات، 3 ناز-بايروفات،، و 3 GSH (بالمليمول، الرقم الهيدروجيني: 7.3-7.4، الأسمولالية: ) لمدة ساعة واحدة على الأقل في . ثم تم نقل شرائح الدماغ إلى حجرة الشرائح (وارنر إنسترومنتس، هولستون، ميزوري، الولايات المتحدة الأمريكية) للتسجيل الكهربائي مع تدفق مستمر لمحلول ACSF القياسي الذي يحتوي علىجلوكوز، و (بالمليمول، الرقم الهيدروجيني: 7.3-7.4، الأسمولالية: تم نقل الشرائح إلى الحجرة التي كانت تتلقى تدفقًا مستمرًا من محلول ACSF القياسي المؤكسج الذي يحتوي على، و 10 جلوكوز (بالمليمول، الرقم الهيدروجيني: 7.3-7.4، الأسمولية: 300-305 مOsm/kg) للتسجيل في بواسطة سخان المحلول (TC-344B، وارنر إنسترومنتس، هولستون، ميزوري، الولايات المتحدة الأمريكية). كان المراقب معزولًا عن الأغراض التجريبية أثناء التسجيل والتحليلات.
لأغراض تسجيلات الخلايا الكاملة باستخدام تقنية الباتش كلومب، تم تصور الخلايا العصبية باستخدام ميكروسكوب تداخل الضوء التفاضلي بالأشعة تحت الحمراء (IR-DIC) (BX51WI، أوليمبوس، طوكيو، اليابان) مععدسة غمر الماء. تم إجراء تسجيلات توصيل الخلايا الكاملة باستخدام رقع ماصات ) التي تم سحبها من أنابيب زجاجية بوروسيليكات (VitalSense Scientific Instruments Co., Ltd.، ووهان، الصين) بقطر خارجي يبلغ 1.5 مم على جهاز سحب أفقي بأربع مراحل (P1000، Sutter Instruments، نوفاتو، كاليفورنيا، الولايات المتحدة الأمريكية). تم جمع الإشارات باستخدام مضخم MultiClamp 700B، وتم تصفيتها بتردد منخفض عند 2.8 كيلوهرتز، وتم تحويلها إلى بيانات رقمية عند 10 كيلوهرتز باستخدام نظام جمع البيانات Digidata 1440 A، وتم تحليلها باستخدام pClamp 10.7 (Molecular Devices، سوني فيل، كاليفورنيا، الولايات المتحدة الأمريكية). تم استبعاد الخلايا العصبية التي كانت مقاومتها السلسلية أكثر من أو التي تغيرت بأكثر منأثناء التسجيل.
لتسجيل الخصائص الغشائية الجوهرية، يتم ملء الميكروبيبتات بمحلول داخلي يحتوي علىكانت المحلول داخل الخلايا يحتوي على (بالمليمول):-غلوكونات، 2 ملغ ATP، 0.6 EGTA، و 0.3 Na-GTP (درجة الحموضة: 7.4، الأسمولية: 290-300 مOsM).
ما لم يُذكر خلاف ذلك، تم شراء جميع الأدوية المستخدمة في التسجيل الكهربائي من سيغما-ألدريتش، سانت لويس، ميزوري، الولايات المتحدة الأمريكية.
التحليل الغربي
تم الحصول على أنسجة CeA بسرعة من-شرائح سميكة مأخوذة على جهاز الاهتزاز. لاستخراج البروتين الكلي، تم تجانس الأنسجة في محلول RIPA البارد، الذي يحتوي على 50 مليمول من Tris-HCl (pH 7.6)، 1% Triton X-100،خليط مثبطات البروتياز، وديكسيكولات الصوديوم. ثم تم الحصول على البروتينات عن طريق الطرد المركزي عندفيلمدة 15 دقيقة، وتم قياس تركيزات البروتين بواسطة اختبار BCA. في كل حارة،تم فصل البروتين عن طريق هلام الكهربية SDS-PAGE بنسبة 10% ثم تم نقله إلى أغشية بولي فينيليدين فلوريد (PVDF) (بايو راد). بعد حجبها بحليب خالي من الدسم بنسبة 5%، تم تحضين الأغشية مع الأجسام المضادة الأولية، بما في ذلك MST4 (1:1000، أبكام، ab52491) وبيتا-أكتين (1:1000، أبسين، abs137975)، عندتمت المعالجة طوال الليل مع الأجسام المضادة الثانوية المعلمة بالبيروكسيداز (1:5000، جاكسون) في درجة حرارة الغرفة لمدة 90 دقيقة. تم تصور جميع شرائط البروتين باستخدام كاشف ECL عالي الحساسية (GE Healthcare) وتم تحليلها باستخدام برنامج ImageJ. تم توفير المسحات غير المعالجة للبلوط في ملف بيانات المصدر.
عزل RNA و
تم تخدير الفئران باستخدام الإيزوفلوران تلاه البنتوباربيتال صوديومتمت معالجة الحيوانات (i.p.) وتمت عملية الحقن القلبي بمحلول PBS البارد. تم قطع شرائح الدماغ كما هو الحال في التسجيلات الكهربية، وتم تشريح عينات CeA يدويًا باستخدام إبرة حقن على الثلج. تم عزل وتنقية RNA الكلي باستخدام مجموعة RNeasy (Qiagen)، تلاها النسخ العكسي باستخدام مجموعة النسخ العكسي SuperRT III (Biosharp، BL1013B). تم إجراء qPCR من 1 إلى 10 نانوغرام من قوالب cDNA باستخدام مجموعة SYBR qPCR Master Mix العالمية (Biosharp، BL697A) على نظام ABI StepOne (Applied Biosystems، وولثام، ميزوري، الولايات المتحدة الأمريكية). تم استخدام قياس mRNA لجين Gapdh كتحكم تحميل للتطبيع. تم حساب التغيرات النسبية لمستويات mRNA مقارنة بالتحكم باستخدام طريقة. تم إجراء كل تفاعل PCR في الوقت الحقيقي ثلاث مرات. تم توفير تسلسلات البرايمرات (Sangon Biotech) المستخدمة في PCR في البيانات التكميلية 3.
التكميم والتحليل الإحصائي
تم إجراء التحليلات الإحصائية والرسم البياني باستخدام GraphPad Prism 8.0 (GraphPad Software, Inc.، سان دييغو، كاليفورنيا، الولايات المتحدة الأمريكية). تم تعريف الدلالة الإحصائية على أنهاقمنا بإجراء مقارنات إحصائية بسيطة باستخدام اختبار ستودنت-اختبار، وتم استخدام ANOVA (أحادي الاتجاه وثنائي الاتجاه) لتحليل بيانات المجموعات التجريبية إحصائيًا مع مقارنات متعددة. اختبار Student غير المتزاوج أو المتزاوجتم استخدام اختبارات – لمقارنة المجموعتين. يتم الإشارة إلى مستويات الدلالة على أنها ; ; . ن.س. تمثل غير ذات دلالة. تم إجراء جميع التجارب السلوكية باستثناء قياس التغيرات في السلوكيات الاستكشافية (أي، OFT و EPM) مرتين على الأقل. جميع البيانات معبر عنها كمتوسطSEM.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
لا توجد قيود على توفر البيانات في المخطوطة. البيانات التي تم توليدها في هذه الدراسة متاحة في ملف البيانات المصدر. يتم توفير البيانات المصدر مع هذه الورقة.
References
McCall, J. G. et al. CRH engagement of the locus coeruleus noradrenergic system mediates stress-induced anxiety. Neuron 87, 605-620 (2015).
Zhang, G. W. et al. Medial preoptic area antagonistically mediates stress-induced anxiety and parental behavior. Nat Neurosci. 24, 516-528 (2021).
de Andrade, J. S. et al. Acute restraint differently alters defensive responses and fos immunoreactivity in the rat brain. Behav. Brain Res. 232, 20-29 (2012).
Wohleb, E. S. Neuron-microglia interactions in mental health disorders: “for better, and for worse. Front. Immunol. 7, 544 (2016).
Sugama, S., Takenouchi, T., Hashimoto, M., Ohata, H., Takenaka, Y. & Kakinuma, Y. Stress-induced microglial activation occurs through beta-adrenergic receptor: noradrenaline as a key neurotransmitter in microglial activation. J. Neuroinflamm. 16, 266 (2019).
Kettenmann, H., Hanisch, U. K., Noda, M. & Verkhratsky, A. Physiology of microglia. Physiol. Rev. 91, 461-553 (2011).
Werneburg, S., Feinberg, P. A., Johnson, K. M. & Schafer, D. P. A microglia-cytokine axis to modulate synaptic connectivity and function. Curr. Opin. Neurobiol. 47, 138-145 (2017).
Ransohoff, R. M. & Perry, V. H. Microglial physiology: unique stimuli, specialized responses. Annu. Rev. Immunol. 27, 119-145 (2009).
Rooney, S. et al. Neuroinflammatory alterations in trait anxiety: modulatory effects of minocycline. Transl. Psychiatry 10, 256 (2020).
Walker, F. R., Nilsson, M. & Jones, K. Acute and chronic stressinduced disturbances of microglial plasticity, phenotype and function. Curr. Drug Targets 14, 1262-1276 (2013).
Tabassum, S., Misrani, A., Huo, Q., Ahmed, A., Long, C. & Yang, L. Minocycline ameliorates chronic unpredictable mild stressinduced neuroinflammation and abnormal mPFC-HIPP oscillations in mice. Mol. Neurobiol. 59, 6874-6895 (2022).
Wang, Y. L. et al. Microglial activation mediates chronic mild stressinduced depressive- and anxiety-like behavior in adult rats. J. Neuroinflamm. 15, 21 (2018).
McEwen, B. S., Nasca, C. & Gray, J. D. Stress effects on neuronal structure: hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology 41, 3-23 (2016).
Liu, W. et al. The role of neural plasticity in depression: from hippocampus to prefrontal cortex. Neural Plast. 2017, 6871089 (2017).
Huang, S. H. et al. Association of increased amygdala activity with stress-induced anxiety but not social avoidance behavior in mice. Neurosci. Bull. 38, 16-28 (2022).
Bryant, R. A. et al. Enhanced amygdala and medial prefrontal activation during nonconscious processing of fear in posttraumatic stress disorder: an fMRI study. Hum. Brain Mapp. 29, 517-523 (2008).
Frodl, T., Moller, H. J. & Meisenzahl, E. Neuroimaging genetics: new perspectives in research on major depression? Acta Psychiatr. Scand. 118, 363-372 (2008).
Botta, P. et al. Regulating anxiety with extrasynaptic inhibition. Nat. Neurosci. 18, 1493-1500 (2015).
Ahrens, S. et al. A central extended amygdala circuit that modulates anxiety. J. Neurosci. 38, 5567-5583 (2018).
Pliota, P. et al. Stress peptides sensitize fear circuitry to promote passive coping. Mol. Psychiatry 25, 428-441 (2020).
Anthony, T. E., Dee, N., Bernard, A., Lerchner, W., Heintz, N. & Anderson, D. J. Control of stress-induced persistent anxiety by an extra-amygdala septohypothalamic circuit. Cell 156, 522-536 (2014).
Joffe, M. E. et al. Acute restraint stress redirects prefrontal cortex circuit function through receptor plasticity on somatostatinexpressing interneurons. Neuron 110, 1068-1083.e1065 (2022).
Pomrenze, M. B. et al. A corticotropin releasing factor network in the extended amygdala for anxiety. J. Neurosci. 39, 1030-1043 (2019).
Shackman, A. J. & Fox, A. S. Contributions of the central extended amygdala to fear and anxiety. J. Neurosci. 36, 8050-8063 (2016).
Wohleb, E. S., Powell, N. D., Godbout, J. P. & Sheridan, J. F. Stressinduced recruitment of bone marrow-derived monocytes to the brain promotes anxiety-like behavior. J. Neurosci. 33, 13820-13833 (2013).
Yasumoto, Y. et al. Ucp2-dependent microglia-neuronal coupling controls ventral hippocampal circuit function and anxiety-like behavior. Mol. Psychiatry 26, 2740-2752 (2021).
Paolicelli, R. C. & Ferretti, M. T. Function and dysfunction of microglia during brain development: consequences for synapses and neural circuits. Front. Synaptic Neurosci. 9, 9 (2017).
Nimmerjahn, A., Kirchhoff, F. & Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 308, 1314-1318 (2005).
Miyamoto, A. et al. Microglia contact induces synapse formation in developing somatosensory cortex. Nat. Commun. 7, 12540 (2016).
Wang, C. et al. Microglia mediate forgetting via complementdependent synaptic elimination. Science 367, 688-694 (2020).
Shi, Y. et al. Long-term diazepam treatment enhances microglial spine engulfment and impairs cognitive performance via the mitochondrial 18 kDa translocator protein (TSPO). Nat. Neurosci. 25, 317-329 (2022).
Cao, P. et al. Early-life inflammation promotes depressive symptoms in adolescence via microglial engulfment of dendritic spines. Neuron 109, 2573-2589.e2579 (2021).
Wohleb, E. S., Terwilliger, R., Duman, C. H. & Duman, R. S. Stressinduced neuronal colony-stimulating factor 1 provokes microgliamediated neuronal remodeling and depressive-like behavior. Biol. Psychiatry 83, 38-49 (2018).
Zhang, J. et al. IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis. Sci. Adv. 7, eabb9888 (2021).
Qing, H. et al. Origin and function of stress-induced IL-6 in murine models. Cell 182, 372-387.e314 (2020).
Taniguchi, S., Elhance, A., Van Duzer, A., Kumar, S., Leitenberger, J. J. & Oshimori, N. Tumor-initiating cells establish an IL-33-TGF-beta niche signaling loop to promote cancer progression. Science 369, eaay1813 (2020).
De Schepper, S. et al. Perivascular cells induce microglial phagocytic states and synaptic engulfment via SPP1 in mouse models of Alzheimer’s disease. Nat. Neurosci. 26, 406-415 (2023).
Morganti, J. M. et al. The soluble isoform of CX3CL1 is necessary for neuroprotection in a mouse model of Parkinson’s disease. J. Neurosci. 32, 14592-14601 (2012).
Geismann, C. et al. TRAIL/NF-kappaB/CX3CL1 mediated oncoimmuno crosstalk leading to TRAIL resistance of pancreatic cancer cell lines. Int. J. Mol. Sci. 19, 1661 (2018).
Jiao, S. et al. The kinase MST4 limits inflammatory responses through direct phosphorylation of the adaptor TRAF6. Nat. Immunol. 16, 246-257 (2015).
Wang, D. et al. Lateral hypothalamus orexinergic inputs to lateral habenula modulate maladaptation after social defeat stress. Neurobiol. Stress 14, 100298 (2021).
Shao, J. et al. Cav3.1-driven bursting firing in ventromedial hypothalamic neurons exerts dual control of anxiety-like behavior and energy expenditure. Mol. Psychiatry 27, 2901-2913 (2022).
Ciocchi, S. et al. Encoding of conditioned fear in central amygdala inhibitory circuits. Nature 468, 277-282 (2010).
Ma, J., du Hoffmann, J., Kindel, M., Beas, B. S., Chudasama, Y. & Penzo, M. A. Divergent projections of the paraventricular nucleus of the thalamus mediate the selection of passive and active defensive behaviors. Nat. Neurosci. 24, 1429-1440 (2021).
Ramirez, K., Fornaguera-Trias, J. & Sheridan, J. F. Stress-induced microglia activation and monocyte trafficking to the brain underlie the development of anxiety and depression. Curr. Top. Behav. Neurosci. 31, 155-172 (2017).
Crotti, A. & Ransohoff, R. M. Microglial physiology and pathophysiology: insights from genome-wide transcriptional profiling. Immunity 44, 505-515 (2016).
Salter, M. W. & Stevens, B. Microglia emerge as central players in brain disease. Nat. Med. 23, 1018-1027 (2017).
Vilalta, A. & Brown, G. C. Neurophagy, the phagocytosis of live neurons and synapses by glia, contributes to brain development and disease. FEBS J. 285, 3566-3575 (2018).
Badimon, A. et al. Negative feedback control of neuronal activity by microglia. Nature 586, 417-423 (2020).
Augustine, V. et al. Temporally and spatially distinct thirst satiation signals. Neuron 103, 242-249.e244 (2019).
Lee, S. et al. Chemosensory modulation of neural circuits for sodium appetite. Nature 568, 93-97 (2019).
Stillman, J. M., Mendes Lopes, F., Lin, J. P., Hu, K., Reich, D. S. & Schafer, D. P. Lipofuscin-like autofluorescence within microglia and its impact on studying microglial engulfment. Nat. Commun. 14, 7060 (2023).
Chung, W. S. et al. Astrocytes mediate synapse elimination through MEGF10 and MERTK pathways. Nature 504, 394-400 (2013).
Auguste, Y. S. S. et al. Oligodendrocyte precursor cells engulf synapses during circuit remodeling in mice. Nat. Neurosci. 25, 1273-1278 (2022).
Cardona, A. E. et al. Control of microglial neurotoxicity by the fractalkine receptor. Nat. Neurosci. 9, 917-924 (2006).
Qian, Z., Lin, C., Espinosa, R., LeBeau, M. & Rosner, M. R. Cloning and characterization of MST4, a novel Ste20-like kinase. J. Biol. Chem. 276, 22439-22445 (2001).
Lin, J. L., Chen, H. C., Fang, H. I., Robinson, D., Kung, H. J. & Shih, H. M. MST4, a new Ste20-related kinase that mediates cell growth and transformation via modulating ERK pathway. Oncogene 20, 6559-6569 (2001).
Kean, M. J. et al. Structure-function analysis of core STRIPAK Proteins: a signaling complex implicated in Golgi polarization. J. Biol. Chem. 286, 25065-25075 (2011).
Liu, L. et al. Cell type-differential modulation of prefrontal cortical GABAergic interneurons on low gamma rhythm and social interaction. Sci. Adv. 6, eaay4073 (2020).
Breton-Provencher, V. & Sur, M. Active control of arousal by a locus coeruleus GABAergic circuit. Nat. Neurosci. 22, 218-228 (2019).
Weston, M. et al. Olanzapine: a potent agonist at the hM4D(Gi) DREADD amenable to clinical translation of chemogenetics. Sci. Adv. 5, eaaw1567 (2019).
شكر وتقدير
تم دعم هذا العمل من قبل البرنامج الوطني الرئيسي للبحث والتطوير في الصين (STI2030-مشاريع كبرى 2021ZD0203100 إلى Z.Z.)، ومؤسسة العلوم الطبيعية الوطنية في الصين (المنح 32025017 إلى Z.Z.، 32121002 إلى Z.Z.، 82171218 إلى Y.J.، 82101300 إلى D.C.، U22A20305 إلى Y.J.)، وخطط المشاريع الكبرى للعلوم والتكنولوجيا في المقاطعات (202303a07020002 إلى Z.Z.)، ومشروع الأكاديمية الصينية للعلوم للعلماء الشباب في البحث الأساسي (YSBR-013 إلى Z.Z.)، والصين الوطنية
برنامج ما بعد الدكتوراه للمواهب المبتكرة (BX2O22O283 إلى P.C.)، جمعية تعزيز الابتكار للشباب CAS، برنامج الابتكار التعاوني لمركز هيفي للعلوم (2021HSC-CIP013 إلى Y.J.)، صناديق البحث الأساسية للجامعات المركزية (WK9100000030 إلى Y.J.)، صناديق البحث في USTC لمبادرة الفئة الأولى المزدوجة (YD9100002018 إلى Y.J.)، مؤسسة العلوم الطبيعية في مقاطعة آنهوي (2208085J30 إلى Y.J.)، فريق البحث المبتكر للجامعات المحلية عالية المستوى في شنغهاي (SHSMU-ZDCX2O211902 إلى Z.Z.)، معهد الصحة والطب (OYZD20220007 إلى Z.Z.)، مؤسسة العلوم ما بعد الدكتوراه في الصين (2023M733395 إلى P.C.)، ومنحة تانغ في USTC إلى Y.J. نشكر وحدة التصوير المجسم في مركز المرافق الأساسية لعلوم الحياة في جامعة العلوم والتكنولوجيا في الصين وX.S. على الخبرة الفنية القيمة والمساعدة.
مساهمات المؤلفين
P.C. وY.J. وZ.Z. وضعوا تصور الدراسة؛ D.C. وQ.L. وX.S. قاموا بمعظم التجارب؛ F.K. وA.L. وC.Z. وY.L. أجروا بعض التجارب السلوكية؛ D.C. وP.C. وY.J. وZ.Z. كانوا مسؤولين عن الكتابة؛ D.W. وS.Q. شاركوا في مراجعة المخطوطة النهائية؛ D.C. وP.C. وY.J. وZ.Z. أشرفوا على الدراسة.
قسم التخدير، المستشفى الأول التابع لجامعة USTC، مختبر هيفي الوطني للعلوم الفيزيائية على المقياس المجهري، قسم علوم الحياة والطب، جامعة العلوم والتكنولوجيا في الصين، هيفي 230026، الصين.قسم الفسيولوجيا، كلية العلوم الطبية الأساسية، جامعة آنهوي الطبية، هيفي 230022، الصين.قسم التخدير، المستشفى الأول التابع لكلية ونان الطبية، ووهوا 241002، الصين.مركز السكتة الدماغية وقسم الأعصاب، المستشفى الأول التابع لجامعة USTC، قسم علوم الحياة والطب، جامعة العلوم والتكنولوجيا في الصين، هيفي 230026، الصين.مركز العلوم المتقدمة متعددة التخصصات والطب الحيوي، معهد الصحة والطب، قسم علوم الحياة والطب، جامعة العلوم والتكنولوجيا في الصين، هيفي 230026، الصين.ساهم هؤلاء المؤلفون بالتساوي: Danyang Chen وQianqian Lou وXiang-Jie Song. البريد الإلكتروني: zhizhang@ustc.edu.cn; pengcao@ustc.edu.cn; jinyan@ustc.edu.cn
Anxiety-associated symptoms following acute stress usually become extinct gradually within a period of time. However, the mechanisms underlying how individuals cope with stress to achieve the extinction of anxiety are not clear. Here we show that acute restraint stress causes an increase in the activity of GABAergic neurons in the in male mice, resulting in anxiety-like behaviors within 12 hours; meanwhile, elevated GABA neuronal CX3CL1 secretion via MST4 (mammalian sterile-20-like kinase 4)-NF-кB-CX3CL1 signaling consequently activates microglia in the CeA. Activated microglia in turn inhibit GABA neuronal activity via the engulfment of their dendritic spines, ultimately leading to the extinction of anxiety-like behaviors induced by restraint stress. These findings reveal a dynamic molecular and cellular mechanism in which microglia drive a negative feedback to inhibit GABA neuronal activity, thus facilitating maintenance of brain homeostasis in response to acute stress.
Anxiety caused by acute stress is likely an evolutionary adaptation for maintaining heightened vigilance and sustained attention to stimuli that require close attention or for avoiding repeated exposure to dangerous conditions . It is well-known that the acute anxiety can gradually become extinct with the removal of stress stimuli. However, in this dynamic extinction process, the mechanisms of how individuals cope with acute stress and drive anxiety extinction to avoid the irreversible effects induced by persistent anxiety are still unclear.
Microglia and neurons in the brain can communicate bidirectionally. Upon receiving external stimuli, neurons respond first and mediate a cascade of responses throughout the brain, including modulating the activity of neural immune cells, such as microglia, via secretion of soluble factors (e.g., chemokines, cytokines, and
neurotransmitters) . As resident macrophages in the brain, microglia perform essential functions required for the maintenance of brain homeostasis, for instance, removing dying neurons, pruning nonfunctional synapses, or producing ligands that support neuronal survival . Microglial activation is also known to participate in triggering anxiety in animal models of chronic stress . Despite several decades of intensive research focusing on neuronal remodeling in the brain after acute or chronic stress, our understanding of the dynamic mechanisms regulating neuron-microglia interactions, and their role in recovery from acute stress-induced anxiety, remains incomplete.
Acute or chronic exposure to stress can lead to several longlasting adaptive changes in stress-sensitive brain regions such as the amygdala, which has been shown to undergo radical changes in
function and structure . Neuroimaging data have shown that neural activity is consistently elevated in the amygdala of patients with mood disorders, such as fear and depression . In particular, the central nucleus of the amygdala (CeA) serves as the primary output nucleus for amygdala functions, and its increased activity is well-established as a potent driver of anxiety-like behaviors in mice . Although CeA neuronal activity appears to participate in stress responses resulting in neuropsychiatric disorders, the molecular and cellular mechanisms of these CeA neuronal responses in the extinction of acute stress-induced anxiety remain unclear.
In this study, we demonstrate how acute stress induces microglial activation via MST4 (mammalian sterile-20-like kinase 4)-NF-кBCX3CL1 signaling in a manner that later promotes the extinction of anxiety-like behaviors in mice. We find that reduced MST4 expression in GABAergic neurons in the CeA (GABA ) contributes to the increased release of CX3CL1, inducing microglial activation, which in turn increases the engulfment of GABA neuronal spines, thereby reducing GABA neuronal activity and alleviating anxiety-like behaviors within 12 hours after acute restraint stress. These results support a positive regulatory role of microglia in the temporary and reversible activation of the response to acute stress and, thus, maintaining brain homeostasis in male mice. These findings provide a mechanistic basis for the extinction of anxiety-like behaviors and suggest an intervention strategy for treating the effects of stress-induced remodeling in the brain.
Results
GABA neuronal activity participates in the onset and extinction of anxiety-like behaviors in acute stress mice
To investigate the neural basis for an acute stress response, we induced anxiety-like behaviors in male mice through acute restraint stress for 2 hours (ARS-2h), as previously described . Then, the open field test (OFT) and elevated plus maze (EPM) test were used to assess exploratory behaviors in mice at , and 12 h post-ARS-2h and in non-stressed controls (Fig. 1a). Significant changes in exploratory behaviors, which are closely associated with anxiety-like behaviors, indicated by less time spent in the central area of the OFT and fewer entries into the open arms of the EPM, were observed at post-stress induction in ARS-2h mice compared with control mice, while no significant difference was observed between control mice and ARS-2h mice at 12 h after restraint (Fig. 1b-e). In addition, there is no significant difference in distance traveled in OFT at post ARS-2h from that of corresponding control mice (Supplementary Fig.1). These data showed that changes in exploratory behaviors caused by acute restraint stress are extinct by 12 h after stress induction.
The CeA plays a critical role in the generation of anxiety states . We then examined GABA neuronal activity at 8 h /and 12 h post-ARS-2h in freely moving mice by in vivo multitetrode recordings (Fig. 1f). The characteristics of spike waveforms of GABA neurons were identified by using optogenetic tagging in GAD2-Cre mice (Supplementary Fig. 2). The results showed that the spontaneous spike firing rate was augmented in GABA neurons of ARS-2h mice compared with control mice but decreased over time until it returned to levels comparable with the non-stressed controls at 12 h after restraint, aligning well with the results of behavioral tests (Fig. 1g, h).
To further investigate how GABA neurons participated in exploratory behaviors, GABA neuronal activity was inhibited by local injection of AAV-DIO-hM4Di-mCherry into the bilateral CeA of GAD2-Cre mice (Fig. 1i and Supplementary Fig. 3a). At three weeks after virus expression, whole-cell patch-clamp recordings in acute brain slices revealed that the resting membrane potential ( ) was hyperpolarized in GABA neurons after perfusion with Clozapine-N-oxide (CNO) ( ) (Supplementary Fig. 3b, c),
indicating that the virus was successfully expressed in GABA neurons. In addition, in ARS-2h mice, chemogenetic inhibition of GABA neurons increased the time spent and entries in the central area of the OFT and open arms of the EPM (Fig. 1j). Moreover, non-stressed naïve mice with chemogenetic activation of neurons displayed low level of exploratory behaviors (Supplementary Fig. 4). These results suggested that acute restraint stress increases GABA neuronal activity and induces anxiety-like behaviors in mice, while a decrease in this neuronal activity is accompanied by the extinction of anxiety-like behaviors.
Microglial activation is involved in the extinction of anxiety-like behaviors following acute stress
Microglia serve as critical modulators of neuronal activity and associated behavioral responses in mice, including anxiety-like behaviors . To examine whether changes in GABA neuronal activity over time are related to microglial activation in ARS-2h mice, we examined the activation of microglia in the CeA through immunofluorescence detection of the microglia-specific marker ionized calcium-binding adapter molecule 1 (lbal). At 0.5 h and 8 h post-ARS2 h , the number of microglia were significantly higher in model mice than in corresponding control mice, but this change was reversed at 12 h post-ARS-2h (Fig. 2a, b). Since microglial morphology is correlated with their activation status , we analyzed microglial morphology with semi-automatic quantitative morphometric three-dimensional (3D) measurements, which revealed significantly shorter processes and decreased branch points at 0.5 h and 8 h post-ARS-2h compared with corresponding control mice (Fig. 2c, d). Notably, these activation phenotypes of microglia returned to normal levels at 12 h post-ARS-2h in model mice (Fig. 2b-d).
From a molecular standpoint, we also tested the levels of inflammatory marker MHCII and a panel of inflammatory factors, including TNF- , IL- , and IL-6 in microglia. Immunofluorescence staining showed that the expression of MHCII increased in ARS-2h mice compared with controls at 0.5 h post-stress induction associated with microglial activation (Supplementary Fig. 5a, b). In addition, we isolated CeA microglia by fluorescence-activated cell sorting using Cx3cr1-GFP transgenic mice, which express a GFP label in microglial cells, and found that the mRNA of Tnf- and Il-6 analyzed by qPCR were expressed at significantly higher levels in ARS-2h mice than those of non-stressed controls at 0.5 h post-stress treatment (Supplementary Fig. 5c-e). To further validate our observations of changes in microglial density, we stained Ki67 (a marker for cell proliferation) and found that an obvious Ki 67 expression in CeA microglia of ARS-2h mice at 0.5 h and 8 h post treatment compared to that in controls, followed by a gradual decrease over time (Supplementary Fig. 6a). Moreover, TUNEL assays showed that apoptosis levels peaked at 8 h post-ARS-2h, then returned to baseline levels at 12 h post-ARS-2h treatment (Supplementary Fig. 6b). These results led us to hypothesize that the activation of microglia in the CeA produced by acute stress may be involved in the extinction of anxiety-like behaviors.
To examine whether microglial activation in the CeA is responsible for ARS-induced anxiety-like behaviors, intracranial microinfusion with minocycline, which has been widely used to inhibit microglial activity, were delivered in the bilateral CeA by cannula implantation (Fig. 2e). The model mice with pre-microinjection of minocycline still exhibited decreased the time spent and entries in the central area of the OFT and open arms of the EPM at 12 h post-ARS-2h, while ARS-2h mice pre-administered with an ACSF control treatment displayed high levels of exploratory behaviors at 12 h post-stress induction (Fig. 2f, g). It also deserves mention that low levels of exploratory behaviors were observable at 0.5 h post-ARS-2 h in mice pre-treated with either minocycline or ACSF (Fig. 2f, g). Immunofluorescence staining for Iba1 verified that microglial activation in the CeA was significantly inhibited in mice pre-treated with minocycline before ARS-2h stress induction
Fig. 1 | GABA neurons mediate acute restraint stress-induced anxiety-like behaviors. a Schematic of experimental design. Representative heatmaps (b) and summarized data of entries and the time spent in central area (c) of the open field test (OFT) from the indicated groups ( mice per group; mice per group; mice per group; mice per group). Representative heatmaps (d) and summarized data of entries and the time spent in the open arms (e) of the elevated plus maze (EPM) from the indicated groups ( mice per group; mice per group; mice per group; mice per group; e left, ; right, ). f Schematic of multi-channel electrode recordings. Raster plots and typical traces (g) and the quantitative data ( ) of the spontaneous firings of neurons from the
i Schematic of the experimental procedure. Summarized data of entries and the time spent in the central area and open arms of OFT and EPM from the indicated groups (OFT: Control + mCherry, mice, ARS- ,8 mice, ARS mice; left, ; right, , ; EPM: Control + mCherry, mice, ARS-2 mCherry, mice, ARS-2h ; left, ; right, , ). Significance was assessed by two-way repeated-measures ANOVA with post hoc comparison between groups in ( ), one-way ANOVA with post hoc comparison between groups in ( ). All data are presented as mean SEM. , ; n.s., not significant. See also Supplementary Data 1. Source data are provided as a Source Data file.
compared with mice pre-treated with ACSF (Supplementary Fig. 7). No apparent reactive gliosis was observed in the CeA of mice with cannula implantation (Supplementary Fig. 8). In addition, in vivo multitetrode recordings in freely moving ARS-2h mice with CeA pre-microinjection of minocycline showed that spontaneous neuronal activity of GABA neurons was significantly higher at 12 h post-ARS-2h compared with that in ARS-2h mice pre-administered saline (Fig. 2h, i), although no difference in spontaneous neuronal activity of GABA neurons was detected at 0.5 h between ARS-2h mice treated with minocycline and those treated with saline (Fig. 2h, i). These results indicated that inhibiting microglial activation blocks the recovery of GABA neuronal hyperactivity and the extinction of anxiety-like behaviors induced by acute stress.
Microglial engulfment of GABA neuronal spines contributes to the extinction of anxiety-like behaviors induced by acute stress
Previous reports have shown that activated microglia can influence neuronal activity through direct engulfment of spines in the brain . To investigate the potential relationship between microglial sensitivity to stress and GABA neuronal hypoactivity during the extinction of anxiety-like behaviors, we examined the remodeling of GABA neuronal spines following ARS-2h in model mice. To this end, we performed cell type-specific sparse labeling by injecting AAV-CSSP-YFP8E3 virus into the bilateral CeA of GAD2-Cre mice to specifically label the dendritic spines of GABA neurons (Fig. 3a). Compared with corresponding control mice, the density of GABA neuronal dendritic
Fig. 2 | Extinction of anxiety-like behaviors depends on activated microglia. a Representative images of Ibal immunostaining and three-dimensional reconstruction of microglia in the CeA from the indicated groups. Scale bars, (overview) and (inset and rendering). Quantification of cell numbers in the CeA from the indicated groups ( mice per group; , ). IMARIS-based semi-automatic quantification of cell morphometry, including the total process length (c) and number of branch points (d) of Iba1 microglia in the CeA from the indicated groups ( mice per group; ). e Experimental scheme of ARS2 h mice pre-treated with ACSF or minocycline (Mino). Summarized data of entries and the time spent in central area of OFT ( 0.5 h : ARS-2h + ACSF, mice, ARS-2h
Mino, mice; 12 h : ARS-2h + ACSF, mice, ARS-2h + Mino, mice; left, ; right, ). g Summarized data of entries and the time spent in central area of EPM ( 0.5 h : ARS- ACSF, mice, ARS-2h + Mino, mice; 12 h : ARS-2h + ACSF, mice, ARS-2h + Mino, mice; left, ; right, ). Raster plots and typical traces ( ) and the quantitative data ( ) of the spontaneous firings of GABA neurons from the indicated groups ( cells from six mice per group; i ). Significance was assessed by two-way repeated-measures ANOVA with post hoc comparison between groups in (b, c, d, f, g, i). All data are presented as mean SEM. ; n.s., not significant. See also Supplementary Data 1. Source data are provided as a Source Data file.
spines was increased at 0.5 h and 8 h post-ARS-2 h in model mice but recovered to control levels by 12 h post-ARS-2h (Fig. 3b, c). However, GABA neuronal dendritic spine density was not significantly different between model mice pre-treated with minocycline (i.p.) and those treated with saline at 0.5 h post-ARS-2h, but was markedly higher in the minocycline pre-treatment group at 12 h post-stress induction compared with the saline pre-treatment group in model mice (Fig. 3d-f). These results suggested that enhanced microglial engulfment is required for the observed reduction in dendritic spine density of GABA neurons during the extinction of anxiety-like behaviors induced by acute restraint stress.
To investigate the interactions between microglial processes and dendritic spines of GABA neurons, we examined the phagocytic function of microglia in ARS-2h mice. Immunofluorescence staining for the macrophage marker CD68 showed significantly higher levels in ARS-2h mice compared with controls at 0.5 h post-stress induction, but no detectable difference between stressed and control mice at 12 h
post-ARS-2h, consistent with observed changes in microglial Ibal expression (Fig. 3g). In addition, quantitative engulfment assays showed that microglia contained more neuronal dendritic spines, accompanied by greater microglia-dendrite contact areas, in ARS-2h mice than those in control animals at 0.5 h , but not at 12 h , post-stress induction (Supplementary Fig. 9). Moreover, 3D reconstruction showed a significant increase in the colocalization of GAD65/67 immunoreactive puncta, a GABAergic neuron-specific marker, and microglia in the CeA at 0.5 h post-ARS-2h, compared with control mice, and this phenomenon no longer occurred at 12 h post-ARS-2h in model mice (Fig. 3h). Confocal imaging data and 3D surface rendering further depicted abundant colocalization between immunoreactive puncta of GAD65/67, CD68, and Iba1 microglia in the CeA of ARS-2h mice at 0.5 h , but not at 12 h post-treatment, nor in the corresponding control animals (Fig. 3i). Following minocycline pre-treatment in ARS2h mice, microglial phagocytosis of GAD65/67 signals in the CeA was significantly lower than that in the saline pre-treatment group (Fig. 3j).
Fig. 3 | Extinction of anxiety-like behaviors via microglial engulfment of
GABA dendritic spines. a Schematic of bilateral virus infection. Representative images of neuronal dendrites (b) and quantification of spine numbers per ( mice per group; ). Scale bars, . d Experimental scheme for virus injection. Representative images of neuronal dendrites (e) and quantification of spine numbers ( ) from the indicated groups ( mice per group; ). Scale bars, . Representative images (left) and quantitative analyses (right) of immunostaining for CD68 (red), Iba1 (green), and DAPI (blue) in the CeA from the indicated groups ( mice per group; Ibal, . Scale bars, .
h Representative images and three-dimensional surface rendering (left) of Iba1+ microglia (green) containing GAD65/67+ puncta (red) and DAPI (blue) in the CeA from the indicated groups. Quantification (right) of GAD65/67 puncta in microglia
in mice ( cells from six mice per group; ). Scale bars, (overview) and (inset and rendering). i Representative images (left) and quantitative analyses (right) of immunostaining for CD68 (red), Iba1 (green), and GAD65 puncta (purple) in the CeA from the indicated groups ( cells from six mice per group; ). Scale bars, . Representative images (left) and quantification (right) of Iba1 microglia (green) containing GAD65/ puncta (red) in the CeA from the indicated groups ( cells from six mice per group; ). Scale bars, (overview) and (inset and rendering). Significance was assessed by two-way repeated-measures ANOVA with post hoc comparison between groups in ( ), two-tailed unpaired Student’s -test in (j). All data are presented as mean SEM. ; n.s., not significant. See also Supplementary Data 1. Source data are provided as a Source Data file.
These results together suggested that activated microglia can inhibit GABA neuronal activity through an apparent negative feedback mechanism following acute restraint stress to promote the recovery from anxiety-like behaviors in the later stage.
Hyperactivity of GABA neurons activates microglia in the CeA of acute stress mice
To better understand how GABA neurons and microglial cells respond to acute stress stimuli, we examined GABA neuronal activity and microglial morphology in mice with acute stress induced by ARS- 5 min and ARS- 30 min . The results showed that the spontaneous
firing rate of GABA neurons was significantly increased after 5 min of restraint and that this increase also appeared after 30 min of restraint stress compared with control mice (Supplementary Fig. 10a, b). Furthermore, although the number of microglia did not change in either ARS-5min or ARS-30min mice compared that with controls, the branch length and points of microglia were significantly lower at ARS30 min , and these changes were not observable in ARS-5min mice compared with controls (Supplementary Fig. 10c, d). These collective findings suggested that activation of neurons precedes microglial activation in the CeA of mice treated with acute restraint stress.
Next, we further investigated whether GABA neuronal hyperactivity is causally related to microglial activation in acute restraint stress mice. We found that chemogenetic activation of GABA neurons in naïve mice led to microglial activation, indicated by an increased number of microglia and decreased branch length and points in these cells (Supplementary Fig. 11a, b). In addition, chemogenetic inhibition of GABA neurons abolished the observed activation of microglia in ARS-2h mice (Supplementary Fig. 11c, d). Taken together, these results suggested that microglial activation results from elevated GABA neuronal activity induced by acute restraint stress in mice.
CX3CL1-CX3CR1 signaling contributes to the extinction of anxiety-like behaviors following acute stress
To further explore potential molecular mechanisms through which microglial activation induced by hyperactivity of GABA neurons following acute stress, we used qPCR to screen for expression of a number of molecules previously reported to be involved in the response to stress and to mediate neuronal-microglia interactions . We found that the mRNA levels of , and were significantly higher in samples of the CeA tissues collected at 0.5 h post treatment from ARS-2h than those in control mice (Supplementary Fig. 12). It is well known that CX3CL1/fractalkine, which is a secreted chemokine specifically expressed in neurons and widely involved in activity-dependent synaptic pruning, can be cleaved to yield a soluble isoform that triggers microglial activation through binding to its receptor, CX3CR1 . We, therefore, focused on the CX3CL1-CX3CR1 signaling in the regulation of the interactions between neurons and microglia. As expected, Western blot assays showed that the level of soluble CX3CL1 protein was indeed significantly higher at 0.5 h and 8 h in ARS-2h mice than that in control mice (Fig. 4a, b).
To investigate whether this enhanced soluble CX3CL1 in the CeA was involved in regulating microglial activation, we injected the CX3CR1-specific antagonist JMS-17-2 or the vehicle control in ARS-2h mice. Microglial activation, indicated by Ibal expression, in the CeA was significantly inhibited in JMS-17-2 pre-treated acute restraint stress mice compared with those given the vehicle (Fig. 4c, d). In addition, immunofluorescence staining further illustrated that microglial phagocytosis of GAD65/67 puncta in the CeA decreased significantly in ARS-2h mice pretreated with the CX3CR1 antagonist compared with those pre-treated with vehicle (Fig. 4e, f). Next, we examined whether the change in GABA neuronal dendritic spine density was also affected by the CX3CR1 antagonist in ARS-2h mice. GABA neuronspecific expression of AAV-CSSP-YFP-8E3 virus in GAD2-Cre mice showed no difference in the dendritic spine density of these neurons at 0.5 h post-ARS- 2 h between JMS-17-2- and vehicle-pretreated model mice (Fig. ), while the spine density was obviously greater in these neurons at 12 h post-ARS-2h following pre-treatment with JMS-172 compared with the vehicle group after acute restraint stress (Fig. ).
To examine whether this enhanced soluble CX3CL1 in the CeA is responsible for ARS-induced anxiety-like behaviors, we bilaterally injected the JMS-17-2, or the vehicle control, into the CeA of ARS-2h mice. We found that mice pre-treated with JMS-17-2 showed significantly lower exploratory behaviors than vehicle-treated mice at 12 h post-ARS-2h, indicated by fewer entries and less time spent in the central area of the OFT and open arms of the EPM (Fig. 4i, j), which were still observed in both the JMS-17-2 and vehicle groups at 0.5 h post-ARS-2h (Fig. 4i, j). Moreover, in vivo multitetrode recordings in freely moving ARS-2h mice showed no difference in GABA neuronal spontaneous firing activity between the JMS-17-2 and vehicle-treated mice at 0.5 h post-ARS-2h, which was consistent with the results of behavioral tests; however, neuronal firing activity was higher at 12 h post-stress induction in mice with the CeA pre-injection of JMS-17-2 than in vehicle controls (Fig. ).
To explore how Cx3cl1 mRNA increased by ARS-2h, we used qPCR to examine mRNA expression, which is a transcription factor for mediating the expression of cytokines, including CX3CL1 . Interestingly, we found that mRNA levels were significantly increased in the CeA at 0.5 h post-ARS-2h compared with non-stressed control mice (Fig. 4m). Next, we administrated Pyrrolidinedithiocarbamate ammonium (PDTC), a selective inhibitor of NF-kB, into the CeA. In these mice, the increased time and entries in the central area of the OFT and open arms of the EPM at 12 h post-ARS-2h was prevented following PDTC administration (Fig. 4n, o). In addition, a significant decrease in Cx3cl1 mRNA expression was observed in PDTC-treated model mice at 0.5 h post-ARS-2h compared to that in vehicle control animals (Fig. 4p). These findings indicated that the NF-kB signaling pathway is likely involved in regulating CX3CL1 protein expression in acute stress states.
Together, these findings suggest that, within 12 h after stress induction, elevated soluble CX3CL1 protein can promote microglial phagocytosis of GABA neuronal spines, thereby reducing GABA neuronal activity and driving the recovery from anxiety-like behaviors in model mice.
In light of the above mentioned evidence supporting the role of ARSinduced NF-кB-CX3CL1 signaling upregulation in the CeA, we next examined the mechanisms potentially responsible for initiating this process in ARS. The kinase MST4 has been previously reported to directly phosphorylate TRAF6 (TNF receptor-associated factor 6), inhibiting its ubiquitination, and subsequently limiting inflammatory response . It thus has been considered as a “brake” on TLR-TRAF6-NF-кB-mediated inflammatory responses. To test whether MST4 participates in microglial activation in ARS, we conducted immunofluorescence staining of MST4 in the brain and found that it was widely distributed in several regions, including the accumbens nucleus, caudate putamen, paraventricular hypothalamic nucleus, and CeA (Supplementary Fig. 13, and Fig. 5a). We also found that MST4 predominantly colocalized with GABA-specific antibody, but not microglia, in the CeA (Supplementary Fig. 14). qRT-PCR and Western blot assays showed that MST4 expression was significantly decreased at both the mRNA and protein levels in the CeA at 0.5 h post-ARS-2h compared with non-stressed control mice (Fig. 5b, c), and returned to that of controls at 12 h post-ARS-2h (Fig. 5c).
To further explore MST4 function in the regulation of GABA neuronal activity and exploratory behaviors in mice, we next generated a Cre-dependent AAV vector to overexpress MST4 in GABA neurons of GAD2-Cre mice. Three weeks after bilateral CeA injection with rAAV-Ef1 -DIO-MST4-P2A-mCherry-WPRE-pA (AAV-MST4) or rAAV-Efl -DIO-mCherry-WPRE-pA (AAV-mCherry) viruses in GAD2-Cre mice (Fig. 5d), fluorescence microscopy showed that the mCherry signal was largely co-localized with the signal from GABA-specific antibody in the CeA of naïve mice (Fig. 5e). Western blots showed significantly greater MST4 expression in GABA neurons of naïve mice infected with AAV-MST4 than in control mice infected with AAVmCherry (Fig. 5f). We then examined exploratory behaviors in GAD2Cre mice injected with either AAV-MST4 or AAV-mCherry viruses (Fig. 5g) and found that GABA neuron-specific MST4 overexpression, but not the AAV-mCherry control, could lead to increasing time and entries in the central area of the OFT and open arms of the EPM in ARS-2h mice (Fig. 5h, i). In addition, GABA neuronal activity was restored in AAV-MST4-infected ARS mice (Fig. 5j, k), accompanied by lower mRNA levels of and in the CeA (Fig. 5l), compared with AAV-mCherry-infected ARS mice. Moreover, microglial activation was significantly reversed in the CeA of ARS mice injected with AAVMST4, with markedly fewer Iba1 cells and increased total microglial branch length and points compared with ARS-2h mice with CeA-specific expression of the AAV-mCherry vector (Fig. 5m, n).
Fig. 4 | Blocking CX3CL1-CX3CR1 signaling in the CeA prevents the extinction of anxiety-like behaviors following acute stress. Representative images (a) and quantitative analyses (b) for Western blot of CX3CL1 in the CeA ( mice per group). c Representative images of Iba1 immunostaining and three-dimensional reconstruction of microglia in the CeA. Scale bars, (overview) and (inset and rendering). d The ARS-treated mice showed inhibited microglial activation after treated with JMS-17-2 ( mice per group). Representative images (e) and quantification (f) of microglia (green) containing GAD65 puncta (red) in the CeA ( cells from five mice per group). Scale bars, (overview) and (inset and rendering). Representative images of neuronal dendrites (g) and quantification of spine numbers ( ) ( mice per group; , ). Scale bars, . Summarized data of entries and the time spent in
the central area and open arms of OFT (i) and EPM (j) (i left, ; right, left, ; right, , ). Raster plots and typical traces ( ) and the quantitative data ( ) of the spontaneous firings of neurons ( cells from six mice per group; l, ). of in the CeA ( mice per group). Summarized data of entries and the time spent in the central area and open arms of OFT (n) and EPM (o) ( mice per group). qPCR of in the CeA ( mice per group; ). Significance was assessed by two-way repeatedmeasures ANOVA with post hoc comparison between groups in (b, ), two-tailed unpaired Student’s -test in ( ). All data are presented as mean SEM. ; n.s., not significant. See also Supplementary Data 1. Source data are provided as a Source Data file.
Fig. 5 | Overexpression of MST4 in GABA neurons alleviates anxiety-like behaviors induced by acute stress. a Immunofluorescence staining of MST4 (green) and NeuN (red) in the CeA. Scale bars, (left) and (right). b qPCR of Mst4 in the CeA ( mice per group). c Western blot analysis of MST4 in the CeA ( mice per group; mice per group; , ). d Schematic illustration of the MST4 overexpression construction strategy. e Representative images showing mCherry with co-labeling of GABAspecific antibody. The experiment was repeated three times. Scale bars, (left) and (right). f Western blot analysis of MST4 in the CeA ( mice per group; ). g Schematic of the experimental procedure. Summarized data of entries and the time spent in the central area and open arms of OFT (h) and EPM (i). Raster plots and typical traces (j) and the quantitative data (k) of the spontaneous firings of neurons ( cells from six mice per group;
. I qPCR of and in the CeA from the indicated groups ( mice per group). m Representative images of lbal immunostaining and three-dimensional reconstruction of microglia in the CeA. Scale bars, (overview) and (inset and rendering). The ARS-treated mice showed inhibited activation of microglia after overexpression of MST4 in the CeA ( mice per group). Representative images (o) and quantification (p) of microglia (green) containing GAD65 puncta (purple) in the CeA ( cells from six mice per group). Scale bars, (overview) and (inset and rendering). Significance was assessed by two-way repeated-measures ANOVA with post hoc comparison between groups in (c), two-tailed unpaired Student’s -test in (b, f, h, i, k, l, n, p). All data are presented as mean SEM. , . See also Supplementary Data 1. Source data are provided as a Source Data file.
Notably, microglial engulfment of GAD65/67 puncta was also significantly decreased in acute restraint stress mice with GABA neurons infected with AAV-MST4 compared with those with AAV-mCherry infection (Fig. 50, p). It should also be mentioned that naïve mice overexpressing MST4 in GABA neurons exhibited no changes in exploratory behaviors and showed no significant difference in GABA neuronal activity compared with mice expressing control virus (Supplementary Fig. 15).
To explore the effects of MST4 signaling on microglial activation in the CeA, we next generated a Cre-dependent AAV vector expressing MST4 short-hairpin RNAs (shRNAs) to knock down MST4 expression. At three weeks after CeA injection with rAAV-CMV-DIO-mCherry-U6-
shRNA (MST4) (AAV-shMST4) or rAAV-CMV-DIO-mCherry-U6-shRNA (scramble) (AAV-mCherry) in naïve GAD2-Cre mice (Fig. 6a), immunofluorescence staining showed that mCherry cells were invariably colabeled with GABA-specific antibody in the CeA (Fig. 6b). qPCR and Western blot assays showed that Mst4 mRNA levels and MST4 protein levels significantly decreased, while and levels and CX3CL1 protein expression increased in GABA neurons, in naïve MST4 knockdown mice compared with mice infected with the AAVmCherry control (Fig. 6c-e). Furthermore, microglia were activated and microglial engulfment of GAD65/67 was significantly increased in naïve mice with CeA injection of AAV-shMST4 compared with the AAVmCherry control group (Fig. 6f-i). These results suggested that the
Fig. 6 | MST4 knockdown in GABA neurons activates microglia and increases the engulfment of GABA neuronal spines. a Schematic illustration of the construction strategy used for MST4 knockdown. b Representative images showing mCherry expression in neurons with co-labeling of GABA-specific antibody. These experiments have been repeated three times. Scale bars, (left) and (right). c qPCR of Mst4, Nf-kb, and Cx3cl1 mRNA expression in the CeA of naïve mice infected with AAV-mCherry and AAV-shMST4 (mCherry, mice per group, shMST4, mice per group). Western blot analysis of MST4 protein in the CeA of naïve mice infected with AAV-mCherry and AAV-shMST4 ( mice per group; ). e Western blot analysis of CX3CL1 protein in the CeA of naïve mice infected with AAV-mCherry and AAV-shMST4 ( mice per group). Representative images of Iba1 immunostaining and three-dimensional
reconstruction of microglia in the CeA of naïve mice infected with AAV-mCherry and AAV-shMST4. Scale bars, (overview) and (inset and rendering). g Quantification of cell numbers, total process length and number of branch points of microglia in the CeA from naïve mice infected with AAV-mCherry and AAV-shMST4 ( mice per group). Representative images (h) and quantification (i) of microglia (green) containing GAD65/67 puncta (purple) in the CeA from naïve mice infected with AAV-mCherry and AAV-shMST4 ( cells from six mice per group). Scale bars, (overview) and (inset and rendering). Significance was assessed by two-tailed unpaired Student’s -test in ( ). All data are presented as mean SEM. . See also Supplementary Data 1. Source data are provided as a Source Data file.
downregulation of MST4 could directly activate microglia and increase the engulfment of GABA neuronal spines.
These cumulative findings suggest that a decrease in MST4 expression after acute restraint stress results in the activation of NF-кВCX3CL1 signaling in GABA neurons, which in turn promotes microglial activation and engulfment of GABA neuronal spines, leading to the recovery of GABA neuronal hyperactivity and the extinction of anxiety-like behaviors.
Discussion
The findings of the current study suggest that the anxiety-like behaviors induced by acute restraint stress are extinct 12 h after stress induction in male mice and that the activity of GABA neurons is increased but returns to non-stress levels. This process is mediated by microglial activation in the CeA and enhanced microglial engulfment of GABA neuronal spines in acute restraint stress mice. Our results further reveal that MST4-NF-кB-CX3CL1 signaling in GABA neurons mediates microglial activation (Supplementary Fig. 16). These cumulative findings suggest that a microglia-driven negative feedback mechanism, which operates similarly to inhibitory neurons, is important for protecting the brain from excessive activation of neurons in health or disease states.
Acute or chronic stress is associated with a higher risk of many psychological disorders, including anxiety and depressive disorders . As the CeA is the main information output nucleus in the amygdala, many studies have reported that CeA hyperactivity following exposure to stress or stressor events contributes to the pathogenesis of anxiety ; these findings are consistent with our results. By contrast, here, we further explored a time-dependent correlation between GABA neuronal activity and the onset and extinction of anxiety-like behaviors after acute restraint stress in mice. Notably, in addition to the hyperactivity of GABA neurons, microglial activation in the CeA was also observed in mice during acute restraint stress treatment, which was dependent on the activation of GABA neurons due to chemogenetic inhibition of these neurons reversing their activation.
Multiple studies have suggested that activation of microglia, immune monitoring cells in the brain, is involved in mediating responses to various stresses that can serve as a major trigger for many neuropsychiatric disorders . In fact, microglia could be activated rapidly when the body suffers acute stress, indicated by changed morphology, enhanced phagocytic capacity, and an altered transcriptional profile, to detect changes in neuronal activity in real time and to promote the restoration of the brain homeostasis . However, microglial immune function often becomes impaired by chronic stress, resulting in overproduction of inflammatory mediators and exacerbated phagocytic activity, which can be detrimental to the remodeling of neural network, leading to further exacerbation of disease behaviors . The present study shows that microglia in the CeA are activated in mice with acute restraint stress-induced anxietylike behaviors, accompanied by increased engulfment of GABA neuronal dendritic spines and larger contact areas with these neurons. Moreover, pharmacological inhibition of microglial activity prior to acute stress does not prevent the onset of the anxiety-like behaviors but does block the relief of this phenotype in mice, suggesting that microglial activation in the CeA is responsible for the process of relieving acute stress-induced anxiety-like behaviors rather than mediating their occurrence. It follows that microglia respond to neuronal activation and then drive a negative feedback mechanism similar to that of inhibitory neurons, which is essential for protecting the brain from excessive activation in health and disease . There are some potential limitations to examine synaptic engulfment in our study. For example, we failed to provide a direct evidence for proving the specificity of GAD65/67 antibody, although it has been widely used in immunofluorescence staining
in many studies . In addition, the autofluorescence of lipofuscin can be likely detected within microglial lysosomes in the adult mouse brain by light microscopy . To address this general issue in the field, we used the same aged mice as corresponding controls to detect microglial engulfment, which was able to minimize this effect as possible.
In addition to microglia, astrocytes, historically thought to provide structural and metabolic support for neurons, have also been implicated in synaptic pruning. However, astrocytes usually engulf and eliminate synapses during development and in conditions such as Alzheimer’s disease and epilepsy , and there is currently no evidence supporting a role of astrocytes in synaptic pruning in response to stressful stimuli. Oligodendrocytes, another important type of glial cell, are known to be primarily responsible for the regulation of axon growth, and the production and maintenance of the myelination. Although oligodendrocyte precursor cells engulf synapses during circuit remodeling in mice, mature oligodendrocytes are not directly involved in synaptic pruning . Based on the rapid response of microglia to acute stress and its characteristics of synaptic pruning, microglia probably play a dominant role in regulation of the fading of acute stress-induced anxiety-like behaviors. The function of astrocytes and oligodendrocytes under acute stress conditions needs to be further investigated.
The classic neuron-microglia interaction pathway is neuronderived fractalkine (CX3CL1) regulation of microglial activation mediated by binding CX3CR1 on microglia . However, the signaling pathways responsible for controlling neuronal secretion of CX3CL1 remain poorly understood. In the investigation of the upstream signals that mediate CX3CL1 expression, we identified the germinal center kinase subfamily III family kinase MST4, a serine-threonine kinase highly expressed in the placenta, thymus, lymphocytes, and peripheral blood leukocytes . MST4 is a component of the “striatalinteracting phosphatase and kinase” complex, which functions in a wide range of fundamental processes, such as signal transduction, cell cycle control, apoptosis, vesicle trafficking, and cell migration .
Our study provides evidence showing that MST4 is widely distributed in brain regions responsible for emotional processes. More specifically, MST4 is decreased in the CeA, and its overexpression can alleviate GABA neuronal hyperactivity, reversing the anxiety-like behaviors, in ARS mice. We also found that the pro-inflammatory transcription factor NF-кВ regulates Cx3cl1 transcription in GABA neurons, leading to microglial activation via binding with CX3CR1. Local administration of the CX3CR1 receptor antagonist JMS-17-2 blocks the gradual extinction of anxiety-like behaviors, which occurs within 12 h post-stress induction in model mice, ultimately resulting in sustained high GABA neuronal activity. Notably, MST4 overexpression can directly inhibit CX3CL1 release through a MST4-NF-кBCX3CL1 signaling pathway that mediates neuron-microglia interactions in the early stage of acute stress to increase microglial activation and engulfment of GABA neuronal spines, consequently inhibiting GABA neuronal activity in ARS mice. This work provides initial evidence supporting the role of the anti-inflammatory factor MST4, specifically expressed in GABA neurons, in stimulating fractalkine secretion to activate CeA microglia after acute stress.
These collective findings depict the molecular- and cellular-level neural basis underlying the extinction of anxiety-like behaviors following acute stress. This inhibitory feedback process involves acute stress-induced pro-inflammatory signaling by GABA neurons to activate microglia, which, in turn, attenuate neuronal activity via neuronal spine engulfment to suppress anxiety-like behaviors within 12 h after stress induction. Our research thus indicates that microglial activation in the early stage of acute stress can play a protective role in regulating neuronal activity, which is different from the pro-nociceptive role of chronic stress described in previous studies.
Methods
Animals
All the conducted experiments were approved by the Care Committee of the University of Science and Technology of China (USTC) (USTCACUC25120123086). Without special instruction, C57BL/6J, Ai14 (RCL-tdT), Cx3cr1-GFP, and GAD2-Cre male mice were aged 8-10 weeks. Animals were obtained from Charles River or The Jackson Laboratory and bred at the animal facility at USTC. The mice were group-housed with five per cage in a colony in a stable environment ambient temperature and humidity) unless a cannula or tetrode array was implanted. They were maintained under a 12 -h light-dark cycle (lights on from 7:00 to 19:00) with water and food available ad libitum.
Acute restraint stress
Mice were immobilized in modified plastic syringes once for 2 h without special instruction. Holes were drilled in the ends of the syringes to allow the mice to breathe. During the restraint period, the control mice were allowed to freely move in the cage without water or food provided. Acute restraint in mice was conducted between 8:0010:00 am, and behavioral assays were subsequently measured at 0.5 h , , and 12 h post-stress treatment.
Behavioral experiments
For the behavioral test, the mice were transported into the testing room 1 day prior to testing for habitation. Different batches of mice were used for open field test (OFT) and elevated plus maze (EPM) tests, in order to exclude the effects of previous test on the current test. During the testing session, the movement trajectories were recorded and subsequently analyzed offline using EthoVision XT software (Noldus). The entries into and time spent in the center of the open field or the open arms of the elevated plus maze were counted.
For OFT, an open field apparatus consisting of a square area ( ) and a marginal area ( ) was used to assay the anxiety. The mice were quickly placed in the central area of the test box and allowed to move freely. The animal behavior analysis software was used to record the activity of mice in the box, and the experiment time was usually 15 min . The inner wall and bottom of the test chamber were cleaned with ethanol between tests to eliminate the odor effect. The number of times the mouse entered the central area and the total time spent in the central area were analyzed using animal behavior analysis software.
For EPM, the apparatus consisted of a central platform ( ), two closed arms ( ), and two opposing open arms . The maze was 100 cm high from the ground. The mice were placed in the central area of the maze, with their heads facing open arms, and allowed to explore freely. The movement trajectories were analyzed offline, and the time spent in the open arms and the number of entries into the open arms were calculated after recording. The apparatus was cleaned between tests using 75% ethanol.
In vivo multi-channel electrode recordings
The mice to be implanted with a tetrode array were kept alone. Three days after the mice adapted to the environment, four custom-made movable tetrode arrays were implanted into CeA . Homemade screwdriven microdrives with four implantable tetrodes were used to record simultaneously from multiple neurons. Signals were recorded after at least two weeks of recovery from the surgery, and mice were habituated to the cables connected to the electrode on their heads before recording. Subject mice were put in a cylindrical box wrapped with a copper mesh and allowed to freely move without any disturbance, and multichannel electrical signals were recorded throughout this period. Finally, the screw in the electrode holder was turned counterclockwise, and the tetrodes were gradually lowered by after each daily recording test. Neuronal signals were amplified, filtered at a bandwidth, and stored using NeuronStudio software. Then, data were exported to Offline Sorter 4 (Plexon) and NeuroExplorer 4 (NexTechnologies) for offline analysis. Units with a signal-tonoise ratio smaller than 2 were excluded from the analysis. Spike sorting was performed with a sorting method involving the T-Dis E-M algorithm built in Offline Sorter 4. The firing rates of sorted units were calculated using Neuroexplorer 4.
To identify GABA neurons, blue-light pulses ( duration, 20 Hz ) were delivered at the end of each recording session at high frequency. Only laser-evoked and spontaneous spikes with highly similar waveforms (correlation coefficient ) were considered as originating from a single neuron. In subsequent experiments, we classified well-isolated units according to the typical firing pattern of light-evoked GABAergic neurons using an unsupervised clustering algorithm based on a -means method . Specifically, spikes with a shorter half-spike width and half-valley width and higher firing rate were classified as putative GABAergic neurons.
Immunohistochemistry, imaging, and image analysis
Mice were deeply anesthetized with isoflurane (about 15 s ) and then quickly perfused with saline for 3 min and paraformaldehyde solution for 4 min . The brain was removed and placed in paraformaldehyde solution and then stored in a refrigerator for overnight fixation. We then used and sucrose solution and placed the brain in a refrigerator to dehydrate until it became isotonic. After embedding brain tissue with an embedding agent at , the embedded brain tissue was cut into coronal sections with a cryostat microtome system (CM1860, Leica). Brain slices were immersed in antifreeze solution and stored at . For staining, the brain slices containing CeA were washed with PBS for 5 min three times. The brain slices were sealed in PBS containing 5% BSA and 5% Triton X-100 at room temperature for 1 h . The brain slices were then incubated in the primary antibody (Supplementary Data 2). Then the brain slices were incubated with Triton X-100 and donkey serum in the primary antibody (Supplementary Data 2) at overnight. Next, sections were washed three times with PBS for 5 min and incubated with secondary antibody (Supplementary Data 2) for 1.5 h at room temperature in a dark place. Finally, all brain slices were stained with DAPI (Cat. No. D9542, Sigma-Aldrich) dilution buffer (1:2000) for 3 min and washed twice. DAPI was used to label the nuclei, and all the sections were mounted with antifade mounting medium (Cat. No. H-1000, Vector). The slices were scanned and imaged using microscopes (Zeiss LSM880, Zeiss LSM980, Olympus SpinSR, and Olympus FV3000) to visualize the fluorescence signals.
Three-dimensional reconstruction
The slices were stained with anti-Ibal for 24 h , followed by Alexa Fluor 488-conjugated secondary antibody or Alexa Fluor 643 -conjugated secondary antibody staining. Imaging was performed on a Zeiss LSM880 or Zeiss LSM980 microscope using a NA oil objective, and imaging parameters (laser power, gain, and offset) were consistent across all experiments. Z stacking was performed with 1.0 steps in the direction, and -pixel resolution images were analyzed using IMARIS 9.6.2 software (Bitplane). The IMARIS function “Fliaments” was used to quantify the microglial process length and number of branch points. Individual mice were used as independent samples.
Microglial engulfment was analyzed to create a 3D surface rendering of the microglia, with a threshold established to ensure accurate reconstruction of microglial processes, which was then used for subsequent reconstructions. The function of “Spots” was used to reconstruct the GAD65/67 puncta. Finally, the IMARIS MATLAB-based (MathWorks, Natick, MA, USA) plugin “Split into Surface Objects” was used to assess the number of GAD65/67 puncta entirely within the microglial surface (distance ). Two slices randomly picked from
each mouse were imaged and quantified for five or six mice in each group. The mean result was used for morphological analysis.
For analysis of microglial processes and neuronal dendrites contacts, imaging was performed on a Zeiss LSM880 microscope. The “Surface” function was used to accurately reconstruct both lba1 microglia and dendrites, and initially established thresholds were then used for the following reconstructions. Finally, the MATLABbased IMARIS plugin “Surface-Surface Contact Area” was used to measure the size of contact areas between microglial processes and neuronal dendrites.
Flow cytometry
Cx3cr1-GFP mice were deeply anesthetized with pentobarbital sodium , i.p.). Subsequently, mice were perfused intracardially with 20 ml of cold Hank’s Balanced Salt Solution (HBSS), followed by a rapid collection of the bilateral CeA, which was washed with cold PBS and chopped into small pieces on ice. Small tissue was mechanically homogenized using 23G needle, producing cell suspensions which were filtered through a cell strainer. Subsequently, the samples were strained and centrifuged at . The pellets were resuspended in percoll solution and centrifuged at , 10 min ). After careful removal of the supernatant, the pellets were rinsed with PBS. Finally, cells were measured using CytoFLEX (Beckman Coulter, USA) flow cytometer and data were analyzed by FlowJo V10. GFP cells were sorted by BD FACSAria III (BD, USA) for further experiments: investigation the gene expression of and by qPCR (Applied Biosystems, ThermoFisher, China).
Drug administration
Minocycline hydrochloride (Mino) (Cat# M9511, Sigma-Aldrich) was dissolved in water ( ), which was administered via i.p. injection with . For intracranial microinfusion, the guide cannula (internal diameter 0.35 mm , RWD) was generally implanted 0.2 mm higher than the target nucleus of the CeA in mice two weeks before behavioral tests to ensure sufficient recovery time. A concentration of 40 mM Mino diluted in artificial cerebrospinal fluid (ACSF) was delivered using a single injection by insertion of the internal cannula, which was 0.2 mm deeper than the guide cannula, and ACSF ( 250 nl ) was locally applied as a control.
CX3CR1 antagonist JMS-17-2 (Cat# HY-123918, MedChemExpress) was dissolved in corn oil ( ), which was administered via i.p. injection with . A concentration of 10 mM JMS-17-2 diluted in corn oil intracranial microinfused into the bilateral CeA , and the vehicle solution (corn oil, 150 nl ) was locally applied as a control.
Clozapine-N-oxide (CNO) (Cat# C0832, Sigma-Aldrich) was dissolved in DMSO ( ) as a stock solution and diluted in saline for DREADDs experiments, where it was injected ( , i.p.) 30 min before behavioral tests.
Pyrrolidinedithiocarbamate ammonium (PDTC) (Cat# HY-18738, MedChemExpress) was dissolved in PBS. A concentration of 50 mM PDTC was delivered into the bilateral CeA , and the vehicle solution (ACSF, 150 nl ) was locally applied as a control.
Stereotaxic surgery
All virus injection procedures followed the Laboratory Biosafety Guidelines approved by the USTC. Before surgery, the mice were fixed in a stereotactic frame (RWD) under an i.p. injection of pentobarbital sodium ( ). The core temperature of the mice was maintained at using a heating pad. A pulled glass microelectrode was backfilled with virus and connected to a 10 -microliter syringe. The injection volume of different viruses varied from 100 to 300 nl depending on the viral titer and expression potential, and the infusion rate was min. The coordinates were included as medio-lateral (ML) from the midline (in mm ), anterior-posterior (AP) from bregma, and dorsoventral (DV) from the pial surface of the brain.
For chemogenetic manipulation, the rAAV-Ef1 -DIO-hM3D(Gq)-mCherry-WPRE-pA (AAV-DIO-hM3Dq-mCherry, AAV2/ , Cat# PT-0042, BrainVTA) virus or rAAV-Ef1 -DIO-hM4D(Gi)-mCherry-WPRE-pA (AAV-DIO-hM4Di-mCherry, AAV2/9, , Cat# PT-0043, BrainVTA) was bilaterally injected into the CeA of GAD2-Cre mice. Three weeks after viral injection, an intraperitoneal injection of CNO ( , Sigma), which is a synthetic ligand for human muscarinic engineered receptors that binds and activates hM3Dq and hM4Di for the chemogenetic manipulation of neuronal activity , was given 30 min before the behavioral tests. The rAAV-Ef1 -DIO-mCherry-WPRE-pA (AAV-DIO-mCherry, AAV2/9, , Cat# PT0013, BrainVTA) virus was used as the control.
To specifically label neuronal dendritic spines of GABAergic neurons, the cell type-specific sparse labeling by injection of AAV-sparse-CSSP-YFP-8E3 (AAV-CSSP-YFP-8E3, Cat# BC-SL003, BrainCase) virus into CeA in 5-week GAD2-Cre mice. AAV-CSSP-YFP-8E3 virus is produced by co-packaging the AAV-CMV-DIO-Flp plasmid and AAV-FDIO-EYFP plasmid with the ratio of 1:8E3 in a single rAAV production step (total virus titer ).
For overexpression and knockdown of MST4 in GABA neurons, the rAAV-Efl -DIO-MST4-P2A-mCherry-WPRE-pA (AAV-MST4, AAV2/9, ) viruses and rAAV-CMV-DIO-mCherry-U6-shRNA (MST4) (AAV-shMST4, AAV2/9, ) viruses were used for knockdown and overexpression experiments, respectively. AAV-DIOmCherry virus and rAAV-CMV-DIO-mCherry-U6-shRNA (scramble) (AAV-scramble, AAV2/9, , Cat# PT-2788, BrainVTA) virus were used as the controls. The coordinates of the injection site were confirmed in mice after Nissl dye injection. The final coordinates relative to bregma were as follows: AP, -1.2 mm ; ML, ; DV, -4.00 mm .
Electrophysiological recordings
For brain slices preparation, the mice were anesthetized with isoflurane (around 15 s ) followed by pentobarbital sodium ( , i.p.) and afterward intra-cardinally perfused with 20 mL oxygenated icecold N-methyl-D-glucamine (NMDG) and ACSF containing 93 NMDG, -2-hydroxyethyl piperazine-N-2ethane sulfonic acid (HEPES), 25 glucoses, 2 thiourea, 5 Na -ascorbate, 3 Na-pyruvate, , and 3 glutathione (GSH) (in mM, pH: 7.3-7.4, osmolality: 300-305 mOsm). After perfusion, the brain slices containing CeA were quickly sectioned by chilled NMDG ACSF on a microtome (VT1200s, Leica, Germany) vibrating at velocity. The brain slices were quickly incubated in oxygenated NMDG ACSF (saturated with to provide stable pH and continuous oxygenation) for at and then incubated in HEPES ACSF containing HEPES, , 25 glucose, 2 thiourea, , 5 Naascorbate, 3 Na-pyruvate, , and 3 GSH (in mM, pH: 7.3-7.4, the osmolality: ) for at least 1 h at . Then the brain slices were transferred to a slice chamber (Warner Instruments, Holliston, MO, USA) for electrophysiological recording with continuous perfusion with standard ACSF that contained glucose, , and (in mM, pH: 7.3-7.4, osmolality: ). The slices were moved to the chamber that was constantly perfused with oxygenated standard ACSF containing , and 10 glucose (in mM, pH: 7.3-7.4 osmolarity: 300-305 mOsm/kg) for recording at by solution heater (TC-344B, Warner Instruments, Holliston, MO, USA). The observer was blinded to the experimental purposes during recording and analyses.
For whole-cell patch-clamp recordings, neurons were visualized using an infrared (IR)-differential interference contrast (DIC) microscope (BX51WI, Olympus, Tokyo, Japan) with a water-immersion objective. Whole-cell patch-clamp recordings were performed using
patch pipettes ( ) that were pulled from borosilicate glass capillaries (VitalSense Scientific Instruments Co., Ltd., Wuhan, China) with an outer diameter of 1.5 mm on a four-stage horizontal puller (P1000, Sutter Instruments, Novato, CA, USA). The signals were collected using a MultiClamp 700B amplifier, lowpass filtered at 2.8 kHz , digitized at 10 kHz with a Digidata 1440 A Data Acquisition System, and analyzed using pClamp 10.7 (Molecular Devices, Sunnyvale, CA, USA). Neurons were excluded with a series resistance of more than or that changed by more than during the recording.
To record the intrinsic membrane properties, micropipettes are filled with an internal solution containing . The intracellular solution contained (in mM): -gluconate, 2 Mg ATP, 0.6 EGTA, and 0.3 Na-GTP (pH: 7.4, osmolarity: 290-300 mOsm).
Unless otherwise stated, all drugs used for electrophysiological recording were purchased from Sigma-Aldrich, St. Louis, MO, USA.
Western blotting
The CeA tissues were quickly obtained from -thick slices taken on the vibratome. To extract total protein, the tissues were homogenized in ice-cold RIPA buffer, which contained 50 mM Tris- HCl (pH 7.6), 1% Triton X-100, , a protease inhibitor cocktail, and sodium deoxycholate. Then, the proteins were obtained by centrifuging at at for 15 min , and the protein concentrations were measured by BCA assay. In each lane, protein was separated via 10% SDS-PAGE gel electrophoresis and then transferred to polyvinylidene difluoride (PVDF) membranes (Bio-Rad). After blocking with 5% skim milk, the membranes were incubated with primary antibodies, including MST4 (1:1000, Abcam, ab52491) and beta-actin (1:1000, Absin, abs137975), at overnight and with peroxidase-labeled secondary antibody (1:5000, Jackson) at room temperature for 90 min . All the protein bands were visualized using high-sensitivity ECL reagent (GE Healthcare) and analyzed using ImageJ software. Unprocessed scans of the blots are provided in the Source Data file.
RNA isolation and
Mice were anesthetized with isoflurane followed by pentobarbital sodium ( , i.p.) and intracardially perfused with ice-cold PBS. Brain slices were sectioned as for electrophysiological recordings, and CeA samples were manually dissected using a syringe needle on ice. Total RNA was isolated and purified using the RNeasy kit (Qiagen), followed by reverse transcription using a SuperRT III Reverse Transcription kit (Biosharp, BL1013B). qPCR from 1 to 10 ng of cDNA templates was performed using a universal SYBR qPCR Master Mix (Biosharp, BL697A) on an ABI StepOne system (Applied Biosystems, Waltham, MO, USA). Gapdh mRNA quantification was used as a loading control for normalization. Fold changes of mRNA levels over controls were calculated using the method. Each real-time PCR reaction was performed in triplicate. Sequences of the primers (Sangon Biotech) used in PCR are provided in Supplementary Data 3.
Quantification and statistical analysis
The statistical analyses and graphing were performed using GraphPad Prism 8.0 (GraphPad Software, Inc., San Diego, CA, USA). Statistical significance was defined as . We performed simple statistical comparisons using Student’s -test, and ANOVA (one-way and twoway) was used to statistically analyze the experimental groups’ data with multiple comparisons. Unpaired or paired Student’s -tests were used to compare the two groups. Significance levels are indicated as ; ; . n.s. represents not significant. All behavioral experiments except for measuring the changes in exploratory behaviors (i.e., OFT and EPM) were performed at least twice. All data are expressed as the mean SEM.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
There are no restrictions on data availability in the manuscript. The data generated in this study are provided in the Source Data file. Source data are provided with this paper.
References
McCall, J. G. et al. CRH engagement of the locus coeruleus noradrenergic system mediates stress-induced anxiety. Neuron 87, 605-620 (2015).
Zhang, G. W. et al. Medial preoptic area antagonistically mediates stress-induced anxiety and parental behavior. Nat Neurosci. 24, 516-528 (2021).
de Andrade, J. S. et al. Acute restraint differently alters defensive responses and fos immunoreactivity in the rat brain. Behav. Brain Res. 232, 20-29 (2012).
Wohleb, E. S. Neuron-microglia interactions in mental health disorders: “for better, and for worse. Front. Immunol. 7, 544 (2016).
Sugama, S., Takenouchi, T., Hashimoto, M., Ohata, H., Takenaka, Y. & Kakinuma, Y. Stress-induced microglial activation occurs through beta-adrenergic receptor: noradrenaline as a key neurotransmitter in microglial activation. J. Neuroinflamm. 16, 266 (2019).
Kettenmann, H., Hanisch, U. K., Noda, M. & Verkhratsky, A. Physiology of microglia. Physiol. Rev. 91, 461-553 (2011).
Werneburg, S., Feinberg, P. A., Johnson, K. M. & Schafer, D. P. A microglia-cytokine axis to modulate synaptic connectivity and function. Curr. Opin. Neurobiol. 47, 138-145 (2017).
Ransohoff, R. M. & Perry, V. H. Microglial physiology: unique stimuli, specialized responses. Annu. Rev. Immunol. 27, 119-145 (2009).
Rooney, S. et al. Neuroinflammatory alterations in trait anxiety: modulatory effects of minocycline. Transl. Psychiatry 10, 256 (2020).
Walker, F. R., Nilsson, M. & Jones, K. Acute and chronic stressinduced disturbances of microglial plasticity, phenotype and function. Curr. Drug Targets 14, 1262-1276 (2013).
Tabassum, S., Misrani, A., Huo, Q., Ahmed, A., Long, C. & Yang, L. Minocycline ameliorates chronic unpredictable mild stressinduced neuroinflammation and abnormal mPFC-HIPP oscillations in mice. Mol. Neurobiol. 59, 6874-6895 (2022).
Wang, Y. L. et al. Microglial activation mediates chronic mild stressinduced depressive- and anxiety-like behavior in adult rats. J. Neuroinflamm. 15, 21 (2018).
McEwen, B. S., Nasca, C. & Gray, J. D. Stress effects on neuronal structure: hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology 41, 3-23 (2016).
Liu, W. et al. The role of neural plasticity in depression: from hippocampus to prefrontal cortex. Neural Plast. 2017, 6871089 (2017).
Huang, S. H. et al. Association of increased amygdala activity with stress-induced anxiety but not social avoidance behavior in mice. Neurosci. Bull. 38, 16-28 (2022).
Bryant, R. A. et al. Enhanced amygdala and medial prefrontal activation during nonconscious processing of fear in posttraumatic stress disorder: an fMRI study. Hum. Brain Mapp. 29, 517-523 (2008).
Frodl, T., Moller, H. J. & Meisenzahl, E. Neuroimaging genetics: new perspectives in research on major depression? Acta Psychiatr. Scand. 118, 363-372 (2008).
Botta, P. et al. Regulating anxiety with extrasynaptic inhibition. Nat. Neurosci. 18, 1493-1500 (2015).
Ahrens, S. et al. A central extended amygdala circuit that modulates anxiety. J. Neurosci. 38, 5567-5583 (2018).
Pliota, P. et al. Stress peptides sensitize fear circuitry to promote passive coping. Mol. Psychiatry 25, 428-441 (2020).
Anthony, T. E., Dee, N., Bernard, A., Lerchner, W., Heintz, N. & Anderson, D. J. Control of stress-induced persistent anxiety by an extra-amygdala septohypothalamic circuit. Cell 156, 522-536 (2014).
Joffe, M. E. et al. Acute restraint stress redirects prefrontal cortex circuit function through receptor plasticity on somatostatinexpressing interneurons. Neuron 110, 1068-1083.e1065 (2022).
Pomrenze, M. B. et al. A corticotropin releasing factor network in the extended amygdala for anxiety. J. Neurosci. 39, 1030-1043 (2019).
Shackman, A. J. & Fox, A. S. Contributions of the central extended amygdala to fear and anxiety. J. Neurosci. 36, 8050-8063 (2016).
Wohleb, E. S., Powell, N. D., Godbout, J. P. & Sheridan, J. F. Stressinduced recruitment of bone marrow-derived monocytes to the brain promotes anxiety-like behavior. J. Neurosci. 33, 13820-13833 (2013).
Yasumoto, Y. et al. Ucp2-dependent microglia-neuronal coupling controls ventral hippocampal circuit function and anxiety-like behavior. Mol. Psychiatry 26, 2740-2752 (2021).
Paolicelli, R. C. & Ferretti, M. T. Function and dysfunction of microglia during brain development: consequences for synapses and neural circuits. Front. Synaptic Neurosci. 9, 9 (2017).
Nimmerjahn, A., Kirchhoff, F. & Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 308, 1314-1318 (2005).
Miyamoto, A. et al. Microglia contact induces synapse formation in developing somatosensory cortex. Nat. Commun. 7, 12540 (2016).
Wang, C. et al. Microglia mediate forgetting via complementdependent synaptic elimination. Science 367, 688-694 (2020).
Shi, Y. et al. Long-term diazepam treatment enhances microglial spine engulfment and impairs cognitive performance via the mitochondrial 18 kDa translocator protein (TSPO). Nat. Neurosci. 25, 317-329 (2022).
Cao, P. et al. Early-life inflammation promotes depressive symptoms in adolescence via microglial engulfment of dendritic spines. Neuron 109, 2573-2589.e2579 (2021).
Wohleb, E. S., Terwilliger, R., Duman, C. H. & Duman, R. S. Stressinduced neuronal colony-stimulating factor 1 provokes microgliamediated neuronal remodeling and depressive-like behavior. Biol. Psychiatry 83, 38-49 (2018).
Zhang, J. et al. IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis. Sci. Adv. 7, eabb9888 (2021).
Qing, H. et al. Origin and function of stress-induced IL-6 in murine models. Cell 182, 372-387.e314 (2020).
Taniguchi, S., Elhance, A., Van Duzer, A., Kumar, S., Leitenberger, J. J. & Oshimori, N. Tumor-initiating cells establish an IL-33-TGF-beta niche signaling loop to promote cancer progression. Science 369, eaay1813 (2020).
De Schepper, S. et al. Perivascular cells induce microglial phagocytic states and synaptic engulfment via SPP1 in mouse models of Alzheimer’s disease. Nat. Neurosci. 26, 406-415 (2023).
Morganti, J. M. et al. The soluble isoform of CX3CL1 is necessary for neuroprotection in a mouse model of Parkinson’s disease. J. Neurosci. 32, 14592-14601 (2012).
Geismann, C. et al. TRAIL/NF-kappaB/CX3CL1 mediated oncoimmuno crosstalk leading to TRAIL resistance of pancreatic cancer cell lines. Int. J. Mol. Sci. 19, 1661 (2018).
Jiao, S. et al. The kinase MST4 limits inflammatory responses through direct phosphorylation of the adaptor TRAF6. Nat. Immunol. 16, 246-257 (2015).
Wang, D. et al. Lateral hypothalamus orexinergic inputs to lateral habenula modulate maladaptation after social defeat stress. Neurobiol. Stress 14, 100298 (2021).
Shao, J. et al. Cav3.1-driven bursting firing in ventromedial hypothalamic neurons exerts dual control of anxiety-like behavior and energy expenditure. Mol. Psychiatry 27, 2901-2913 (2022).
Ciocchi, S. et al. Encoding of conditioned fear in central amygdala inhibitory circuits. Nature 468, 277-282 (2010).
Ma, J., du Hoffmann, J., Kindel, M., Beas, B. S., Chudasama, Y. & Penzo, M. A. Divergent projections of the paraventricular nucleus of the thalamus mediate the selection of passive and active defensive behaviors. Nat. Neurosci. 24, 1429-1440 (2021).
Ramirez, K., Fornaguera-Trias, J. & Sheridan, J. F. Stress-induced microglia activation and monocyte trafficking to the brain underlie the development of anxiety and depression. Curr. Top. Behav. Neurosci. 31, 155-172 (2017).
Crotti, A. & Ransohoff, R. M. Microglial physiology and pathophysiology: insights from genome-wide transcriptional profiling. Immunity 44, 505-515 (2016).
Salter, M. W. & Stevens, B. Microglia emerge as central players in brain disease. Nat. Med. 23, 1018-1027 (2017).
Vilalta, A. & Brown, G. C. Neurophagy, the phagocytosis of live neurons and synapses by glia, contributes to brain development and disease. FEBS J. 285, 3566-3575 (2018).
Badimon, A. et al. Negative feedback control of neuronal activity by microglia. Nature 586, 417-423 (2020).
Augustine, V. et al. Temporally and spatially distinct thirst satiation signals. Neuron 103, 242-249.e244 (2019).
Lee, S. et al. Chemosensory modulation of neural circuits for sodium appetite. Nature 568, 93-97 (2019).
Stillman, J. M., Mendes Lopes, F., Lin, J. P., Hu, K., Reich, D. S. & Schafer, D. P. Lipofuscin-like autofluorescence within microglia and its impact on studying microglial engulfment. Nat. Commun. 14, 7060 (2023).
Chung, W. S. et al. Astrocytes mediate synapse elimination through MEGF10 and MERTK pathways. Nature 504, 394-400 (2013).
Auguste, Y. S. S. et al. Oligodendrocyte precursor cells engulf synapses during circuit remodeling in mice. Nat. Neurosci. 25, 1273-1278 (2022).
Cardona, A. E. et al. Control of microglial neurotoxicity by the fractalkine receptor. Nat. Neurosci. 9, 917-924 (2006).
Qian, Z., Lin, C., Espinosa, R., LeBeau, M. & Rosner, M. R. Cloning and characterization of MST4, a novel Ste20-like kinase. J. Biol. Chem. 276, 22439-22445 (2001).
Lin, J. L., Chen, H. C., Fang, H. I., Robinson, D., Kung, H. J. & Shih, H. M. MST4, a new Ste20-related kinase that mediates cell growth and transformation via modulating ERK pathway. Oncogene 20, 6559-6569 (2001).
Kean, M. J. et al. Structure-function analysis of core STRIPAK Proteins: a signaling complex implicated in Golgi polarization. J. Biol. Chem. 286, 25065-25075 (2011).
Liu, L. et al. Cell type-differential modulation of prefrontal cortical GABAergic interneurons on low gamma rhythm and social interaction. Sci. Adv. 6, eaay4073 (2020).
Breton-Provencher, V. & Sur, M. Active control of arousal by a locus coeruleus GABAergic circuit. Nat. Neurosci. 22, 218-228 (2019).
Weston, M. et al. Olanzapine: a potent agonist at the hM4D(Gi) DREADD amenable to clinical translation of chemogenetics. Sci. Adv. 5, eaaw1567 (2019).
Acknowledgements
This work was supported by the National Key Research and Development Program of China (STI2030-Major Projects 2021ZD0203100 to Z.Z.), the National Natural Science Foundation of China (grants 32025017 to Z.Z., 32121002 to Z.Z., 82171218 to Y.J., 82101300 to D.C., U22A20305 to Y.J.), the Plans for Major Provincial Science & Technology Projects (202303a07020002 to Z.Z.), the CAS Project for Young Scientists in Basic Research (YSBR-013 to Z.Z.), the China National
Postdoctoral Program for Innovative Talents (BX2O22O283 to P.C.), Youth Innovation Promotion Association CAS, CAS Collaborative Innovation Program of Hefei Science Center (2021HSC-CIP013 to Y.J.), the Fundamental Research Funds for the Central Universities (WK9100000030 to Y.J.), USTC Research Funds of the Double FirstClass Initiative (YD9100002018 to Y.J.), the Natural Science Foundation of Anhui Province (2208085J30 to Y.J.), the Innovative Research Team of High-level Local Universities in Shanghai (SHSMU-ZDCX2O211902 to Z.Z.), the Institute of Health and Medicine (OYZD20220007 to Z.Z.), the China Postdoctoral Science Foundation (2023M733395 to P.C.), and USTC Tang Scholar to Y.J. We thank the Confocal Imaging Unit at the Core Facility Centre for Life Science of the University of Science and Technology of China and X.S. for valuable technical expertise and assistance.
Author contributions
P.C., Y.J., and Z.Z. conceptualized the study; D.C., Q.L., and X.S. performed most of the experiments; F.K., A.L., C.Z., and Y.L. conducted some of the behavioral experiments; D.C., P.C., Y.J., and Z.Z. were responsible for writing; D.W. and S.Q. were involved in the revision of the final manuscript; D.C., P.C., Y.J., and Z.Z. supervised the study.
Correspondence and requests for materials should be addressed to Zhi Zhang, Peng Cao or Yan Jin.
Peer review information Nature Communications thanks the anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Department of Anesthesiology, the First Affiliated Hospital of USTC, Hefei National Laboratory for Physical Sciences at the Microscale, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei 230026, China. Department of Physiology, School of Basic Medical Sciences, Anhui Medical University, Hefei 230022, China. Department of Anesthesiology, the First Affiliated Hospital of Wannan Medical College, Wuhu 241002, China. Stroke Center and Department of Neurology, The First Affiliated Hospital of USTC, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei 230026, China. The Center for Advanced Interdisciplinary Science and Biomedicine, Institute of Health and Medicine, Division of Life Sciences and Medicine, University of Science and Technology of China, Hefei 230026, China. These authors contributed equally: Danyang Chen, Qianqian Lou, Xiang-Jie Song. e-mail: zhizhang@ustc.edu.cn; pengcao@ustc.edu.cn; jinyan@ustc.edu.cn