تعديل غير جراحي لللمفاويات السحائية يحسن من الشيخوخة ومرض الزهايمر المرتبط بالمرض والإدراك في الفئران Non-invasive modulation of meningeal lymphatics ameliorates ageing and Alzheimer’s disease-associated pathology and cognition in mice
لقد أظهرت الأوعية اللمفاوية السحائية (mLVs) أنها تشارك في إزالة الأميلويد بيتا ، والتي تعتبر هدفًا علاجيًا محتملاً لمرض الزهايمر (AD). في هذه الدراسة، استنادًا إلى التوزيع المكاني السطحي لـ mLVs، تم استخدام ضوء قريب من الأشعة تحت الحمراء لتعديل تصريف اللمف، مما يحسن بشكل كبير الإدراك لكل من الفئران المسنّة ومرض الزهايمر (5xFAD و APP/PS1)، ويخفف من علم الأمراض المرتبطة بمرض الزهايمر عن طريق تقليل ترسب A ، الالتهاب العصبي والأضرار العصبية. علاوة على ذلك، تشير بيانات تصوير المجهر الإلكتروني الناقل وتسلسل RNA إلى تحسين في التمثيل الغذائي للميتوكوندريا والاتصال الخلوي لخلايا بطانة اللمفاويات السحائية (mLECs) بواسطة تعديل الضوء. تشير هذه الدراسات مجتمعة إلى أن علاج الضوء القريب من الأشعة تحت الحمراء يمكن أن يحسن الوظيفة الإدراكية من خلال تعزيز قدرة التنظيف لـ mLVs من خلال استعادة وظيفة mLEC. في الختام، فإن تعزيز تصريف اللمف بواسطة الضوء يعزز الشفاء المرضي وتحسين الإدراك في نماذج الفئران المسنّة ومرض الزهايمر، مما يقدم استراتيجية تحسين محتملة للأمراض التنكسية العصبية.
لفترة طويلة، اعتُبر الدماغ محميًا من المناعة بسبب عدم وجود نظام تصريف لمفاوي. في عام 2015، تم اكتشاف الأوعية اللمفاوية السحائية (mLVs) الموجودة في الأم الجافية، والتي تشكل شبكة تصريف لمفاوي واسعة لإزالة النفايات الجزيئية الكبيرة والوسائط الالتهابية، ونقل خلايا المناعة، وتنسيق الاستجابات المناعية في الجهاز العصبي المركزي (CNS). كشفت الدراسات الحديثة أن نظام mLVs مرتبط بتقدم الشيخوخة، ومرض الزهايمر (AD)، ومرض باركنسون (PD)، وإصابة الدماغ الرضحية (TBI)، ونزيف تحت العنكبوتية (SAH)، والعدوى الفيروسية في CNS وأمراض الجهاز العصبي الأخرى، وأن التغيرات في قدرة نقل mLVs يمكن أن تؤثر بشكل كبير على تطور المرض. تشير هذه الدراسات إلى أن
تعديل تصريف mLVs يمكن أن يكون استراتيجية فعالة للأمراض العصبية.
مرض الزهايمر هو مرض تنكسي عصبي مرتبط بالعمر مع معدل وفيات مرتفع. الميزات المرضية الرئيسية لمرض الزهايمر هي-تجمع غير طبيعي للأميلويد (A ) وتشابكات عصبية في الدماغ تساهم في خلل وظيفي عصبي وتدهور إدراكي. تم تأكيد أن mLVs قد تدهورت وظيفيًا مع تقدم العمر أو مرض الزهايمر، مما قد يكون عاملاً أساسيًا في تفاقم الخلل الوظيفي الإدراكي والضرر العصبي. يمكن أن يعزز العلاج بواسطة عامل نمو بطانة الأوعية الدموية C (VEGF-C) عبر حقن داخل القناة الشوكية (i.c.m.) وظيفة mLVs بشكل فعال، ثم يعزز تصريف اللمف لإزالة الجزيئات السامة في CNS، وكذلك تحسين
القدرة على التعلم والذاكرة. لذلك، قد تكون mLVs هدفًا علاجيًا للعيوب الإدراكية المرتبطة بالعمر. ومع ذلك، بالنسبة للأمراض التنكسية العصبية المزمنة والمتقدمة، مثل مرض الزهايمر، فإن نماذج العلاج الجراحي ليست قابلة للتطبيق. بشكل عام، فإن تطوير نمط علاج غير جراحي ضروري لعلاج مرض الزهايمر.
توزع شبكة mLVs بشكل سطحي في الأم الجافية، مما يوفر استراتيجية واعدة للعلاجات العصبية عبر الجمجمة لتخفيف أمراض CNS من خلال تعديل تصريف mLVs. في هذا العمل، استنادًا إلى النافذة البصرية لاختراق الأنسجة، نوضح أن الضوء القريب من الأشعة تحت الحمراء يمكن أن يعدل وظيفة خلايا بطانة اللمفاويات السحائية (mLECs)، والتي بدورها يمكن أن تساهم في تصريف mLVs من أجل تخفيف المرض وتعزيز الوظيفة الإدراكية للفئران المسنّة ومرض الزهايمر. تكشف نتائجنا أن تعزيز التوازن الأيضي الميتوكوندري لـ mLECs بواسطة الضوء يحدد تعزيز التصاق الخلايا والنمو، مما قد يعزز وظائف الخلايا والاتصالات بين mLECs لتعزيز نقل اللمفاويات السحائية في CNS من أجل تحسين المرض.
النتائج
الضوء يعدل تصريف mLVs ويخفف من التدهور الإدراكي في الفئران المسنّة
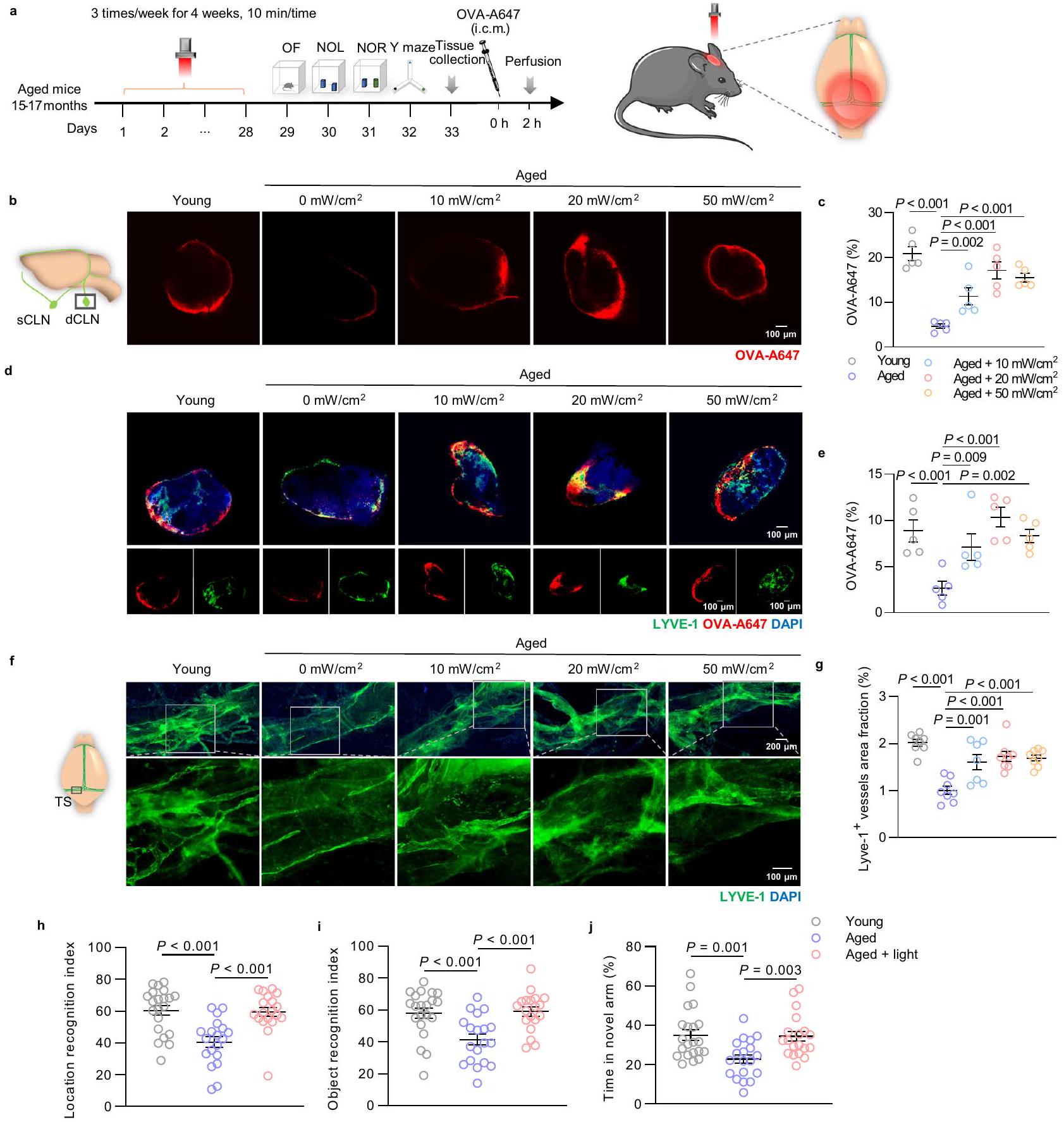
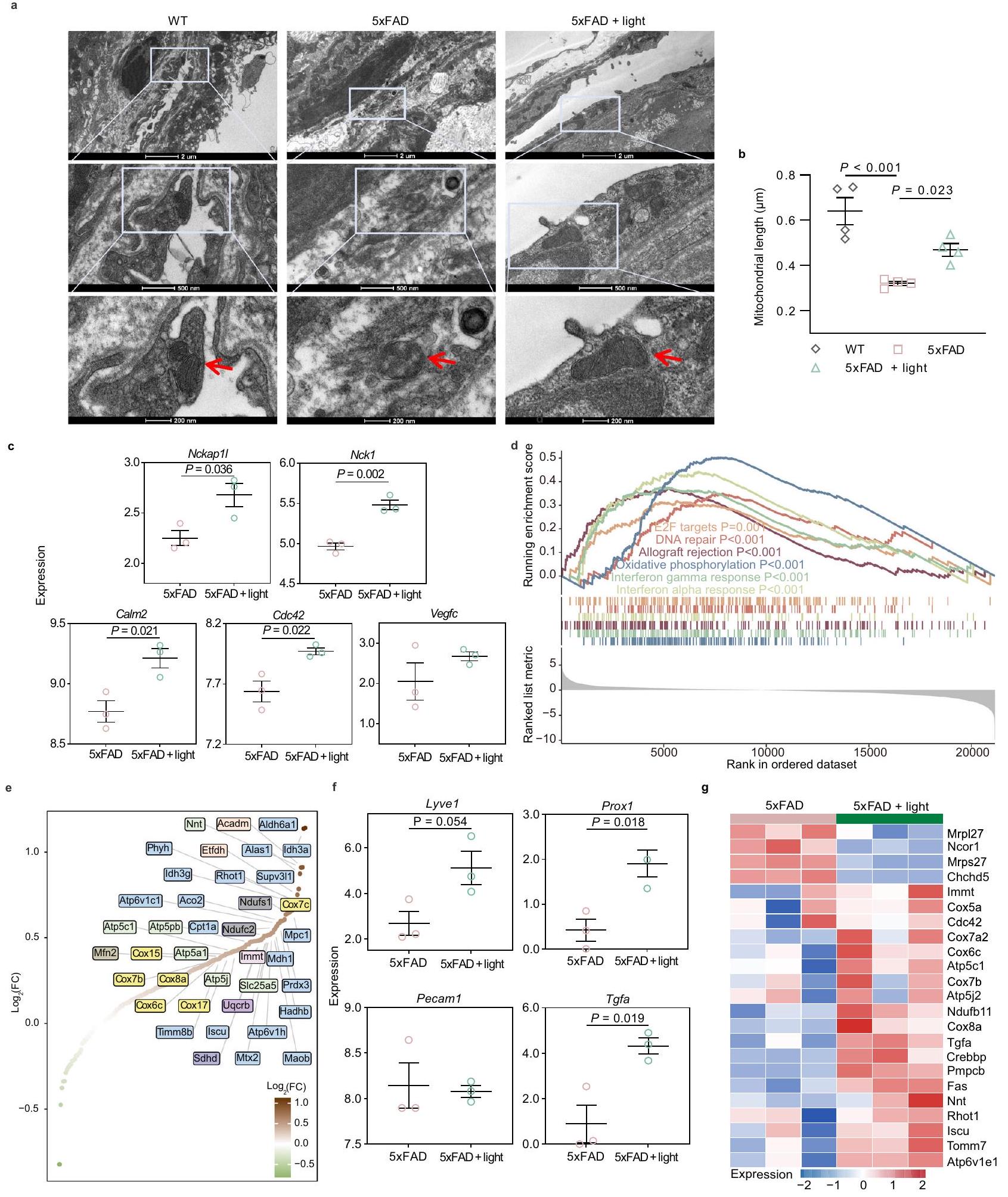
لتحديد الفرضية القائلة بأن الضوء يمكن أن يعدل اللمفاويات السحائية، تم إجراء علاج ضوئي غير جراحي عبر الجمجمة باستخدام ليزر بطول موجي 808 نانومتر على فئران C57BL/6J المسنّة (15-17 شهرًا، ذكور وإناث) لمدة 4 أسابيع (3 مرات في الأسبوع، 10 دقائق في المرة). تم تخدير الفئران، وتم حلق وتنظيف فروة الرأس الخلفية، ثم خضعت للإضاءة عبر الجمجمة بواسطة ضوء 808 نانومتر (فئران معالجة بالضوء) أو إضاءة داخلية (فئران معالجة كتحكم) لمدة 10 دقائق. بعد العلاج، تم تقييم وظيفة التصريف وتغيرات الشكل لـ mLVs، وحركة الفئران ووظيفتها الإدراكية (الشكل 1a). من أجل الحصول على جرعة فعالة لعلاج الضوء، تم اختيار كثافات طاقة مختلفة (10، 20، و ) للمقارنة، استنادًا إلى الجرعة التقليدية للضوء القريب من الأشعة تحت الحمراء (NIR) المستخدمة لفوائد الصحة ( )، ومعدل اختراق ضوء 808 نانومتر على جمجمة الفأر مع فروة رأس خالية من الشعر (حوالي ، بغض النظر عن تغييرات الطاقة الناتجة) (الشكل التكميلي 1a). بالإضافة إلى ذلك، تم تسجيل درجة حرارة سطح فروة الرأس أثناء إشعاع الضوء. لم يكن هناك تقلبات ملحوظة في درجة الحرارة، مما استبعد تأثير تغييرات درجة الحرارة على وظيفة اللمفاويات السحائية (الشكل التكميلي 1b، c).
أولاً، تم تقييم قدرة تصريف نظام اللمفاويات السحائية من خلال الكشف عن كثافة الفلورسنت لمؤشر CSF OVA-A647 (i.c.m.) في العقد اللمفاوية العنقية العميقة (dCLNs). كانت منطقة توزيع OVA-A647 التي تم تصريفها إلى dCLNs أعلى بكثير في الفئران المعالجة بالضوء مقارنة بتلك غير المعالجة (المسنّة مقابل المسنّة: ; المسنّة : مقابل المسنّة: ; المسنّة + مقابل المسنّة: ) (الشكل 1b-e). ومع ذلك، لم يكن هناك تمييز كبير في تدفق الدم الدماغي للفئران المسنّة بعد علاج الضوء (الشكل التكميلي 1d، e). تم تقييم هيكل الأوعية اللمفاوية بواسطة الكيمياء المناعية مع صبغة مستقبلات الهيالورونان 1 (LYVE-1) (علامة كلاسيكية لخلايا بطانة اللمفاويات (LECs)). تم استعادة فقدان مساحة الأوعية اللمفاوية السحائية في الفئران المسنّة إلى الوضع الطبيعي بواسطة علاج الضوء (المسنّة : مقابل المسنّة: ; المسنّة + مقابل المسنّة: ; المسنّة مقابل المسنّة: , ) (الشكل 1f، g). أظهرت النتائج أن الضوء يمكن أن يحسن تصريف وتوزيع mLVs في الفئران المسنّة، وأن الضوء عند أظهر تأثيرات أكثر استقرارًا وفعالية.
لتقييم حركة ووظائف الإدراك للفئران المعالجة، تم إجراء اختبارات الحقل المفتوح (OF)، وتحديد موقع الجسم الجديد (NOL)، والتعرف على الجسم الجديد (NOR) واختبارات المتاهة Y على الفئران المعالجة بالضوء . أظهرت نتائج اختبار OF أنه لم يكن هناك فرق كبير في المسافة الإجمالية، أو السرعة المتوسطة، أو الوقت الذي قضاه في المركز بين المجموعات، مما يشير إلى أن الضوء لم يؤثر على الحركة أو يزيد من سلوك القلق لدى الفئران المسنّة (الشكل التكميلي 1f-i). من المثير للاهتمام، أن الفئران المسنّة التي خضعت لعلاج الضوء قضت وقتًا ملحوظًا أكثر مع الجسم في الموقع الجديد في اختبار NOL أو الجسم الجديد في اختبار NOR مقارنة بالجسم المألوف (المسنّة + ضوء: مقابل المسنّة: في اختبار NOR)، مشابهًا لأداء الفئران الشابة (1.5 شهرًا، ذكور وإناث) في نفس الاختبار. على العكس، فضلت الفئران المسنّة غير المعالجة الجسم المألوف في اختبار NOL وNOR (الشكل 1h، i). في الوقت نفسه، أظهرت الفئران المسنّة التي تلقت علاج الضوء مؤشر تمييز أعلى بشكل ملحوظ (DI، DIفي اختبارات NOL وNOR، كانت الفئران غير المعالجة أقل أداءً (الشكل التوضيحي 1j-o). في اختبار المتاهة Y، استكشفت الفئران المسنّة أقل في الذراع الجديدة مقارنة بالفئران الشابة، مما يدل على وجود عجز في الذاكرة. ومع ذلك، بعد العلاج بالضوء، زادت الفئران المسنّة بشكل كبير من الوقت الذي قضته في الذراع الجديدة. ومن المثير للاهتمام أن الوقت الذي قضته الفئران المسنّة المعالجة بالضوء في الذراع الجديدة كان مشابهًا لأداء الفئران الشابة (الشكل 1j). كشفت النتائج أعلاه أن الضوء على الممرات اللمفاوية الصغيرة يمكن أن يحسن من التعرف والذاكرة المكانية في الفئران المسنّة. لذلك، يمكن أن تكون العلاج الضوئي المستهدف للممرات اللمفاوية الصغيرة فعالًا في استعادة توزيع ووظيفة تصريف الممرات اللمفاوية الصغيرة، فضلاً عن تحسين الإدراك في الفئران.
تعديل الضوء يخفف من التدهور المعرفي والأضرار المرضية في الفئران المصابة بمرض الزهايمر
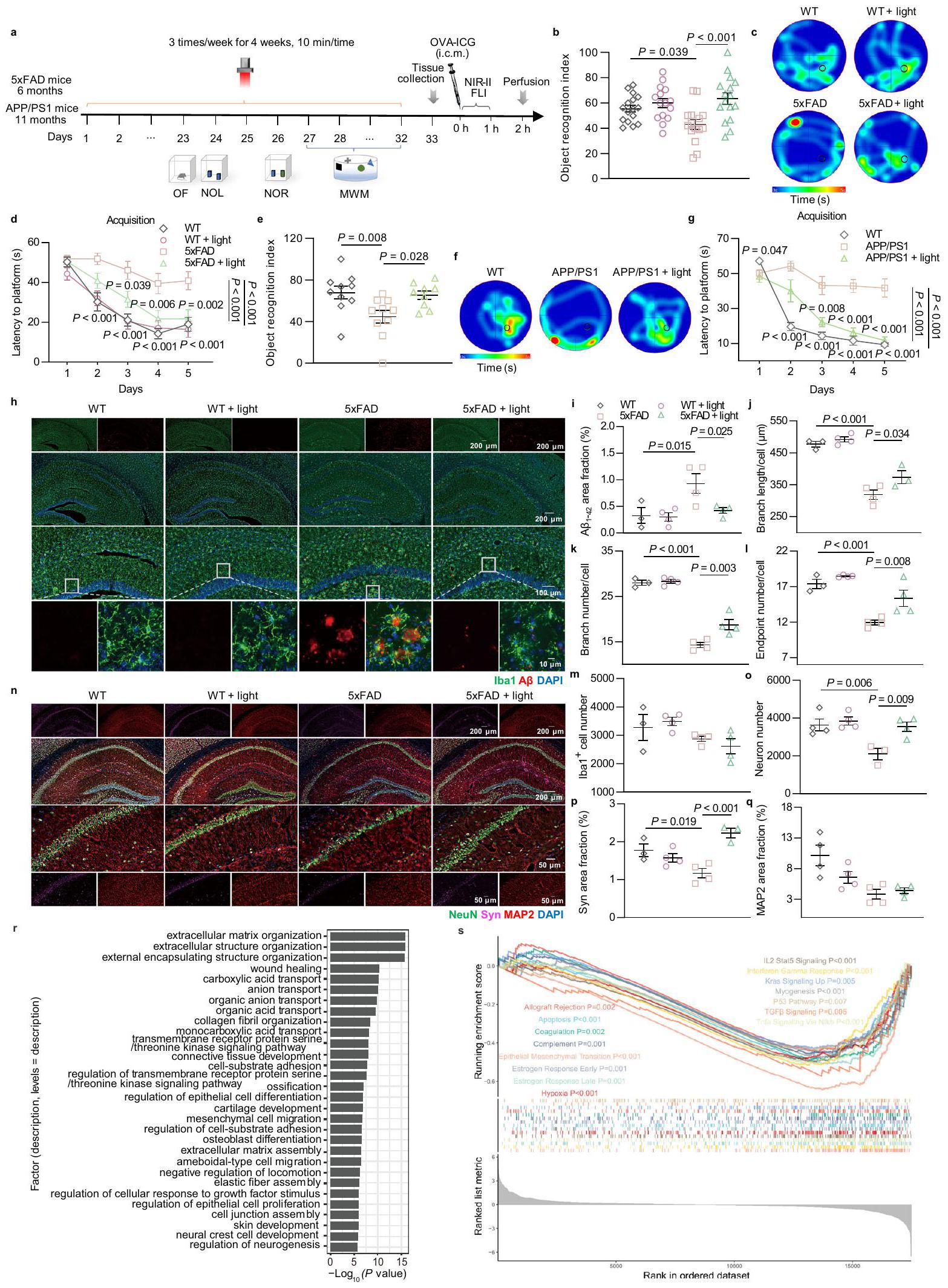
لاستكشاف تأثير تعديل mLVs بواسطة الضوء على قدرة التعلم والذاكرة لدى الفئران المصابة بمرض الزهايمر، قمنا أيضًا بإجراء علاج بالضوء وأجرينا اختبارات OF و NOL و NOR على فئران 5xFAD (6 أشهر، ذكور) و APPswe/PS1.فئران (APP/PS1) (11 شهرًا، ذكر) (الشكل 2أ). في اختبار OF، لم يكن هناك فرق ملحوظ في المسافة الإجمالية المقطوعة، أو السرعة المتوسطة، أو الوقت في المركز بين المجموعات، مما يشير إلى أن الفئران لم تعاني من أي عجز حركي بعد العلاج بالضوء (الشكل التوضيحي 2أ-د والشكل التوضيحي 3أ-د). كما هو متوقع، أظهرت فئران AD المعالجة بالضوء استعادة في الإدراك في الاختبارات السلوكية (5xFAD + ضوء: مقابل 5xFAD: في اختبار NOR؛ APP/PS1 + ضوء: مقابل APP/PS1: في اختبار NOR). ومع ذلك، لم يكن هناك زيادة ملحوظة في الأداء المعرفي في الفئران من النوع البري (WT) المعالجة بالضوء (WT + ضوء: ضد WT: في اختبار NOR) (الشكل 2ب، هـ، الشكل التكميلية 2هـ-ك والشكل التكميلية 3هـ-ز). بشكل عام، أدى الضوء أيضًا إلى تعديل مميز في التعلم والذاكرة المكانية في فئران 5xFAD وAPP/PS1، بينما لم يحدث ذلك في الفئران WT. للتحقيق بشكل أكبر في فائدة الضوء على التعلم والذاكرة على المدى الطويل لفئران AD، تم إجراء اختبار متاهة مورس المائية (MWM). مقارنةً بفئران AD غير المعالجة، قلل العلاج بالضوء من زمن الانتظار للعثور على المنصة في تجارب الاكتساب (5xFAD + ضوء مقابل 5xFAD،; APP/PS1 + ضوء مقابل APP/PS1، ) وزيادة الزيارات إلى موقع المنصة والزيارات إلى الربع المستهدف في اختبار الاستكشاف، دون أي تأثير على سرعة السباحة. ومع ذلك، لم يؤدِ الضوء إلى أي تعديلات في مؤشرات السلوك في الفئران من النوع البري المتطابقة في العمر (WT + ضوء مقابل WT، ) (الشكل 2ج، د، و، ز، الشكل التوضيحي 2ل-ع والشكل التوضيحيأظهرت هذه النتائج أن الضوء يمكن أن يحسن العيوب في التعلم المكاني والذاكرة لدى الفئران المصابة بمرض الزهايمر، ولكن ليس لدى الفئران السليمة. تعتبر التراكم، الالتهاب العصبي، والأضرار العصبية من الميزات المرضية لمرض الزهايمر.“تم تقييم ترسبات الالتهاب في الحُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُوبروتين Iba1 (بروتين ربط الكالسيوم المؤين 1، علامة الخلايا الدبقية). أظهرت النتائج أن تفاقم الدماغ Aحدث العبء في HPC و PFC من فئران 5xFAD، والذي تم تخفيفه بشكل ملحوظ في فئران 5xFAD المعالجة بالضوء إلى مستوى مشابه لأقران WT. بالإضافة إلى ذلك، تم ملاحظة تجمع بارز للخلايا الدبقية الصغيرة إلىوتفعيل مع تقليل
الشكل 1 | تأثيرات الضوء على الإدراك وتصريف الميكروفيسكلات الكبيرة في الفئران المسنّة. أ جدول العلاجات واختبارات السلوك للفئران المسنّة.صور تمثيلية للعقد اللمفاوية العنقية العميقة (dCLNs) المتراكمة مع OVA-A647 بعد ساعتين من الحقن (i.c.m.) (من نسختين). مقياس الرسمتقدير توزيع الفلورسنت لـ OVA-A647 في dCLNs.فئران في كل مجموعة. د صور مقاطع تمثيلية من العقد اللمفية الدهنية المتراكمة OVA-A647 ملونة بـ LYVE-1 و DAPI (من 2 تكرارات). مقياس الرسمتحديد توزيع الفلورسنت لـ OVA-A647 في مقاطع dCLN.فئران في كل مجموعة.صور تمثيلية للأغشية السحائية ملونة بـ LYVE-1 و DAPI (من 2 تكرارات).
شريط القياس أو تحديد نسبة المساحة لـ LYVE-1الأوعية اللمفاوية.فئران في الشيخوخةمجموعة،الفئران في مجموعات أخرى.مؤشر التعرف على الموقع لاختبار موقع الكائن الجديد (NOL). i مؤشر التعرف على الكائن لاختبار التعرف على الكائن الجديد (NOR).نسبة الوقت المستغرق في الذراع الجديدة من اختبار متاهة Y.فئران في مجموعة الشبابالفئران في مجموعات العمر والعمر + الضوء في البيانات. بيانات في تُعرض كمتوسطتم تحليل SEM بواسطة ANOVA أحادي الاتجاه مع اختبار المقارنات المتعددة لسيداك لمقارنات المجموعات المتعددة. يتم توفير بيانات المصدر كملف بيانات مصدر. تم اكتشاف طول الفروع، والعدد، ونقاط النهاية في فئران 5xFAD. وقد قلل الضوء من تنشيط الميكروغليا الشاذ في HPC كما يتضح من زيادة طول الفروع، وعدد الفروع، وعدد نقاط النهاية، مع ميل مشابه في PFC. ومع ذلك، Iba1لم يتغير عدد الخلايا بشكل كبير ولكنه أظهر اتجاهًا تنازليًا بعد العلاج بالضوء (الشكل 2h-m والشكل التوضيحي 4a-f). بشكل عام، خفف الضوء أعراض مرضية للدماغترسب والتهاب عصبي في فئران الزهايمر.
لإجراء مزيد من الفحص لحماية الأعصاب الناتجة عن تعديل mLVs بواسطة الضوء في الفئران المصابة بمرض الزهايمر، تم إجراء تحليلات مناعية نسيجية باستخدام صبغة NeuN (علامة عصبية) و synaptophysin (Syn) وبروتين الأنابيب الدقيقة المرتبط بالبروتين 2 (MAP2). يعتبر synaptophysin جزءًا أساسيًا بروتين غشائي موضعي في الحويصلات ما قبل المشبك يشارك في نقل الناقلات العصبية، وMAP2 هو المنظم الهيكلي الخلوي السائد داخل الشجيرات العصبية.مقارنةً بأقرانهم من النوع البري،تم تقليل خلايا العصبونات وSyn في HPC وPFC لفئران 5xFAD بشكل كبير واستعادتها بواسطة العلاج بالضوء إلى مستويات مشابهة لتلك الموجودة في الأقران WT، مما يشير إلى استعادة كبيرة للعصبونات. الخلايا. في الوقت نفسه، MAP2أظهر تعبير الشجيرات زيادة طفيفة في مجموعة العلاج بالضوء. ومن المثير للاهتمام أن شكل الشجيرات أظهر توزيعًا غير منظم في قشرة الحُصين (HPC) والقشرة الجبهية (PFC) لفئران 5xFAD، وقد عزز الضوء إعادة ترتيب أكثر انتظامًا (الشكل 2n-q والشكل التكميلية). ). وبالمثل، فإن العلاج بالضوء قلل أيضًا تراكم وتنشيط الخلايا الدبقية الصغيرة، بالإضافة إلى تلف الخلايا العصبية و
الشكل 2 | تأثيرات تحسين الضوء على الإدراك والمرض في الفئران المصابة بمرض الزهايمر. أ جدول العلاجات واختبارات السلوك للفئران المصابة بمرض الزهايمر. ب-د مؤشر التعرف على الكائنات في اختبار NOR (ب)، وخرائط حرارة الإشغال التمثيلية (ج) والزمن المستغرق للوصول إلى المنصة (د) في اختبار MWM.فئران FAD.الفئران في WT ومجموعات الضوء،الفئران في مجموعات WT + الضوء و 5xFAD. e-g مؤشر التعرف على الكائنات في اختبار NOR (e)، خرائط الحرارة التمثيلية للاحتلال (f) وزمن الانتظار للمنصة (g) في اختبار MWM لفئران APP/PS1.الفئران في مجموعات WT و APP/PS1،الفئران في مجموعة APP/PS1 + الضوء في البياناتالفئران في كل مجموعة في البيانات“صور تمثيلية لمقاطع الدماغ في الحُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُ“، Iba1 و DAPI. مقياس الرسم أو . قياس نسبة المساحة من في الحوسبة عالية الأداء.تحديد طول الفروع لكل خلية )، رقم الفرع لكل خلية ( رقم نقطة النهاية ( ) ورقم الهاتف ( ) من الخلايا في HPC. الفئران في مجموعة WT،الفئران في مجموعات أخرى.صور تمثيلية لمقاطع الدماغ في HPC ملونة بـ NeuN و synaptophysin و MAP2 و DAPI. مقياس الرسم أو تحديد عدد الخلايا العصبية )، ونسبة المساحة لـ Syn (p) و MAP2 (q) في HPC. r مصطلحات GO عند معالجة الضوء في فئران 5xFAD.فئران في كل مجموعة. رسومات إثراء GSEA.فئران في كل مجموعة. البيانات فيتُعرض كمتوسطتم تحليل SEM بواسطة ANOVA أحادي الاتجاه (e) أو ANOVA ثنائي الاتجاه (b، d، g، i-m، o-q) مع اختبار المقارنة المتعددة لسيداك لمقارنات المجموعات المتعددة.القيم في و تم حسابها باستخدام clusterProfiler باستخدام اختبار هايبرجومتري واختبار التبديل القائم على النمط الظاهري التجريبي، على التوالي، دون أي تعديلات على المقارنات المتعددة. شملت الاختبارات الإحصائية تحليلًا ثنائي الجانب. تم توفير بيانات المصدر كملف بيانات مصدر. فقدان المشابك في HPC وPFC لفئران APP/PS1 (الشكل التوضيحي 5a-t). وبناءً عليه، يمكن أن يؤدي الضوء إلى تأثير وقائي عصبي من خلال تحسينتراكم، التهاب عصبي، وفقدان الخلايا العصبية والمشابك، مما قد يحول بيئة الجهاز العصبي المركزي نحو مرحلة مرضية مخففة من مرض الزهايمر.
“لتقييم تأثير الضوء على التغيرات المرتبطة بمرض الزهايمر، تم إجراء تحليل تسلسل RNA في الحُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُبالإضافة إلى ذلك، أظهرت مجموعات الجينات المميزة من خلال تحليل إثراء مجموعات الجينات (GSEA) إثراءً مميزًا لمجموعات الجينات المنخفضة التنظيم المرتبطة بتطور مرض الزهايمر مثل الموت الخلوي المبرمج، والتخثر، ونقص الأكسجة، ومسار p53، بالإضافة إلى استجابة الالتهاب بما في ذلك المكمل، وإشارات IL2 Stat5، وإشارات TNF-alpha عبر NF-кB (الشكل 2s). تشير هذه النتائج إلى أن العلاج بالضوء قد يساهم في تنظيم بقاء الخلايا العصبية، والالتهاب العصبي، وغيرها من الأمراض ذات الصلة لتحسين مرض الزهايمر.
تعديل على هيكلبواسطة الضوء يعززتصريف
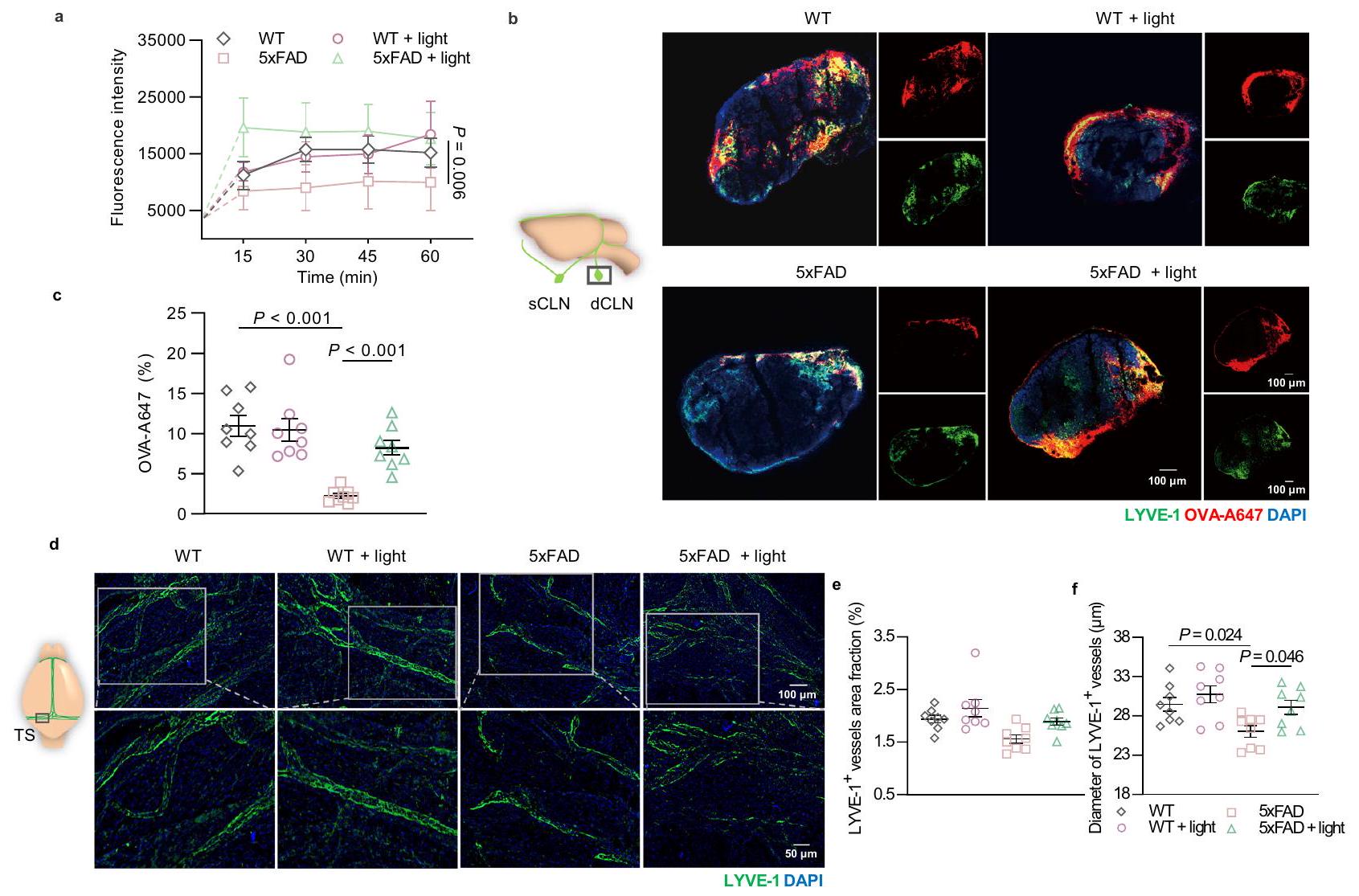
وبالمثل، قمنا بالتحقق من قدرة تصريف اللمف في الفئران المصابة بمرض الزهايمر المعالجة بالضوء من خلال التصوير الضوئي بالأشعة تحت الحمراء NIR-II في العقد اللمفاوية القريبة (CLNs) وصبغة dCLNs المناعية. تدفق OVA-ICG أو OVA-A647 إلى العقد اللمفاوية القريبة (5xFAD + ضوء: 8.225 مقابل 5xFAD: ) أو حول المLVs في السحايا كان أعلى في المعالجة بالضوء فئران 5xFAD غير المعالجة (الشكل 3أ-ج والشكل التكميلي 7أ-د). كما هو متوقع، لم يكن هناك تغيير ملحوظ في تدفق الدم الدماغي لفئران AD بعد العلاج بالضوء (الشكل التكميلي 7هـ، و و). للتحقق مما إذا كان تحسين نقل السائل اللمفاوي مرتبطًا بالتغيرات الهيكلية في الأوعية اللمفاوية الصغيرة، تم تقييم التوسع اللمفاوي في السحايا من خلال صبغة المناعية باستخدام LYVE-1. أشارت النتائج إلى أن تغطية LYVE-1لم تتناقص الأوعية في فئران 5xFAD، على الرغم من أنه تم ملاحظة ضعف تصريف اللمف بالفعل. ومع ذلك، بعد العلاج بالضوء، زاد قطر LYVE-1.الأوعية في السحايا حدثت في الفئران المصابة بمرض الزهايمرضوء مقابل 5xFAD: لذلك، أدى الضوء إلى توسع هيكلي ولكن ليس تغييرات في توزيع الممرات اللمفية الصغيرة في فئران 5xFAD (الشكل 3d-f). بعد ذلك، في فئران APP/PS1، تم ملاحظة ضعف في تصريف الممرات اللمفية الصغيرة وانحلال هيكلي لها من خلال انخفاض إشارات الفلورسنت OVA-ICG في العقد اللمفية العنقية والـ LYVE.نسبة مساحة الأوعية في السحايا، تكشف عن تدهور وظيفة وتوزيع اللمف السحائي. بعد الضوء تم عرض تحسين تصريف اللمف في الفئران المصابة بمرض الزهايمر من خلال تعزيز فلورية OVA-ICG في العقد اللمفية المركزية، بالإضافة إلى تكوين الأوعية اللمفية من خلال زيادة LYVE-1.منطقة في السحايا (الشكل التكميلي 8a-d). ومع ذلك، لم يكن هناك أي تأثيرات على توزيع الأوعية الدموية في السحايا (الشكل التكميلي 8e). ونتيجة لذلك، يمكن أن يحسن الضوء من قدرة التصريف والتوسع الوظيفي للأوعية اللمفاوية الصغيرة في كل من فئران 5xFAD وAPP/PS1.
تعزيز تصريف المLVs مطلوب لتحسين الإدراك بواسطة الضوء
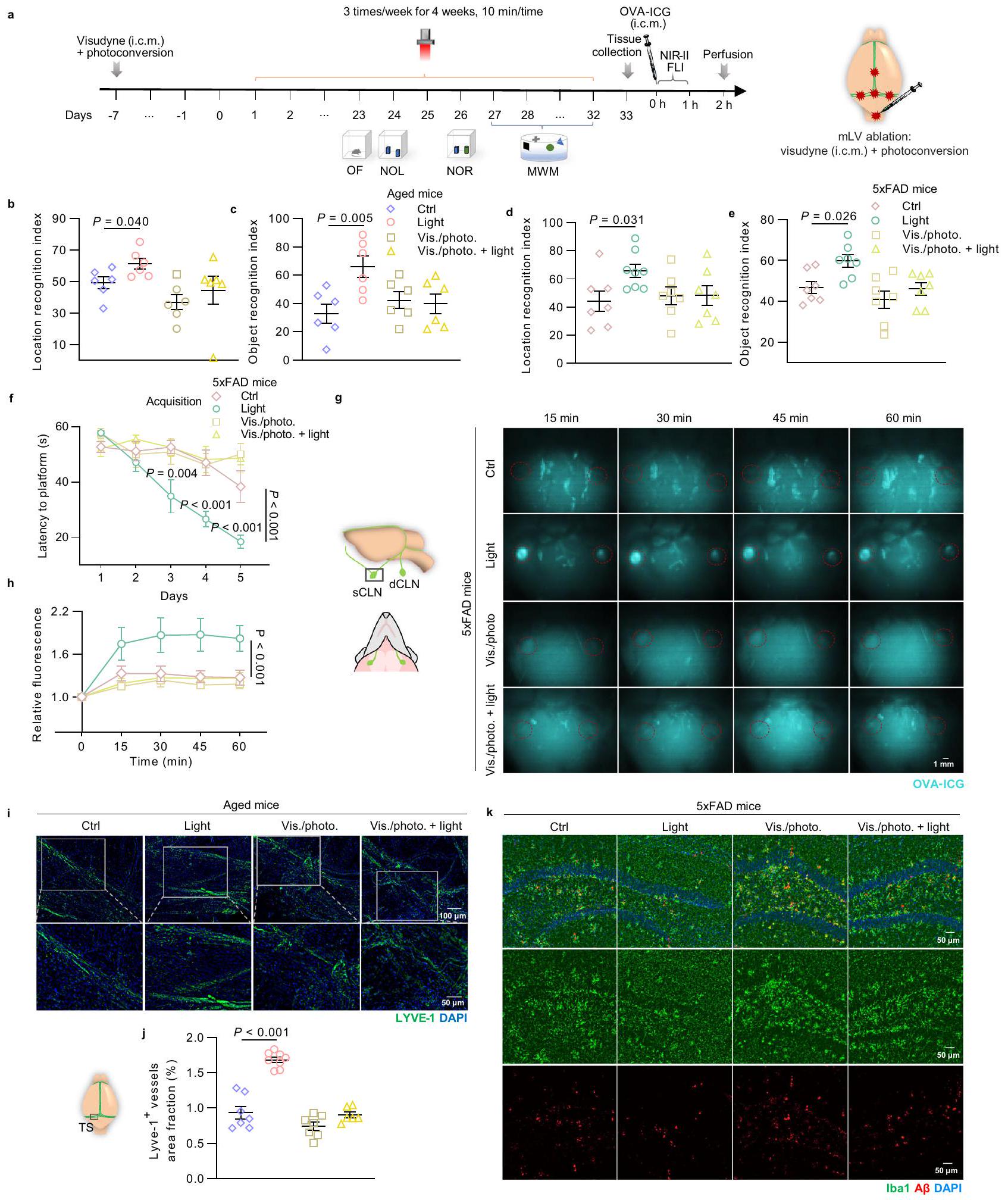
لتحديد ما إذا كان التأثير المفيد للضوء على الوظيفة الإدراكية ومرض الزهايمر المرتبط بالمرض يعتمد على زيادة تصريف الأوعية اللمفية الصغيرة، قمنا بإجراء استئصال للأوعية اللمفية الصغيرة على الفئران المسنّة (15-17 شهرًا، ذكور وإناث) وفئران 5xFAD (6 أشهر، ذكور) وفقًا للإجراء الموصوف في الدراسات السابقة.. ثم قمنا بإجراء علاج بالضوء ( ) على الفئران المسنّة التي تم استئصال mLVs و تمت معالجة الفئران FAD لمدة 4 أسابيع، ثم تم تقييم القدرة الحركية، والقلق، والتعلم والذاكرة لدى الفئران من خلال اختبارات سلوكية (الشكل 4a). لم يكن للضوء أي تأثير على المسافة الإجمالية، أو السرعة المتوسطة، أو الوقت في المركز في اختبار OF (الشكل التكميلي 9a-d والشكل التكميلي 10a-d)، مما يشير إلى عدم وجود تغيير في الحركة أو سلوك القلق. مشابهًا للنتائج المذكورة أعلاه، حسّن علاج الضوء وظيفة التعلم والذاكرة لدى الفئران المسنّة وفئران AD، بينما لم تكن هناك تأثيرات في الفئران التي تم استئصال mLVs منها (مؤشر التعرف على الكائن: Vis./photo. +light للفئران المسنّة:مقابل الصور/الفوتوغرافية للفئران المسنّة:، في اختبار NOR؛ رؤية/صورة + ضوء من فئران 5xFAD: مقابل صورة/فوتوغرافية لـ فئران، في اختبار NOR) (الشكل 4ب-و، الشكل التوضيحي 9هـ-ك والشكل التوضيحي 10هـ-ن). أظهرت هذه النتائج التأثير الإيجابي لعلاج الضوء في تحسين الإدراك لدى الفئران المسنّة وفئران الزهايمر، والذي تم إلغاؤه بعد إزالة الممرات اللمفية المتوسطة، مما يكشف أن الممرات اللمفية المتوسطة لعبت دورًا مهمًا في تعزيز الإدراك من خلال علاج الضوء.
لتقدير ما إذا كان إلغاء فوائد العلاج بالضوء في الفئران المسنّة والمصابة بمرض الزهايمر (AD) المعالجة بإزالة الأوعية اللمفية الصغيرة (mLVs) ناتجًا عن عدم قدرة تعديل وظيفة الأوعية اللمفية الصغيرة، تم تقييم ديناميات التصريف وشكل الأوعية اللمفية الصغيرة. تم الكشف عن توزيع حيوي حركي في الجسم لعقار OVA-ICG في العقد اللمفية السطحية العنقية (sCLNs) (حتى ساعة واحدة بعد الحقن)، وتم تسجيل شدة الفلورسنت لعقار OVA-ICG المتراكم في العقد اللمفية العميقة (dCLNs) والدماغ (بعد ساعتين من الحقن) بواسطة تصوير الفلورسنت بالأشعة تحت الحمراء NIR-II. ارتفعت شدة الفلورسنت في العقد اللمفية السطحية بسرعة إلى ذروتها خلال عدة دقائق في الفئران المسنّة والمصابة بمرض الزهايمر المعالجة بالضوء، ثم انخفضت تدريجياً لتستقر عند مستوى أعلى من الفئران غير المعالجة. ومع ذلك، ظلت الإشارات الفلورية في العقد اللمفية السطحية لكل من الفئران المعالجة بإزالة الأوعية اللمفية الصغيرة مع العلاج بالضوء وبدونه ثابتة عند مستويات منخفضة خلال 60 دقيقة (الشكل التوضيحي التكميلي 9l، m والشكل 4g، h). أخيرًا، تم تصريف المزيد من OVA-ICG إلى العقد اللمفية العميقة والدماغ في فئران 5xFAD التي خضعت للعلاج بالضوء مقارنة بالفئران غير المعالجة بعد ساعتين من الحقن، بينما لم يكن هناك فرق بين الفئران المعالجة بإزالة الأوعية اللمفية الصغيرة التي خضعت للعلاج بالضوء والفئران غير المعالجة (شدة الفلورسنت للعقد اللمفية العميقة:ضوء مقابل : ; رؤية/صورة. +ضوء: مقابل فيس./صورة: أظهرت النتائج زيادة في شدة الفلورية للعلامة في العقد اللمفية العنقية
الشكل 3 | تأثيرات تحسين الضوء على وظيفة التصريف والتغيرات الهيكلية في المLVs في فئران 5xFAD. أ قياس شدة الفلورسنت لـ OVA-ICG في sCLNs بعد الحقن (i.c.m.).فئران في كل مجموعة.صور تمثيلية لقسم من dCLNs المتراكمة بـ OVA-A647 والمصبوغة بـ LYVE-1 و DAPI (من 2 تكرارات). شريط القياستحديد توزيع الفلورسنت لـ OVA-A647 في مقاطع dCLN.فئران في كل مجموعة.ممثل
صور للأغشية السحائية ملونة بـ LYVE-1 و DAPI (من أكثر من 3 تكرارات). شريط القياس أو تحديد كمية نسبة المساحة (e) والقطر (f) لـ LYVE-الأوعية اللمفاوية في السحايا.الفئران في كل مجموعة. البيانات في أ، ج، هـ، و تُعرض كمتوسطتم استخدام SEM، وتم تحليلها بواسطة ANOVA ثنائية الاتجاه مع اختبار المقارنة المتعددة لسيداك لمقارنات المجموعات المتعددة. تم توفير بيانات المصدر كملف بيانات مصدر. وأدمغة الفئران المعالجة بالضوء، مما يشير إلى أن كل من تصريف السحايا وتصريف الغليمف كانا معززين. عندما تم استئصال الأوعية اللمفاوية السحائية، بالإضافة إلى الحظر المتوقع لتصريف السحايا، تم أيضًا إلغاء التأثيرات المفيدة للضوء على وظيفة تصريف الغليمف (الشكل التوضيحي 10o-r). تشير هذه النتائج إلى أن أنظمة اللمف السحائي والغليمف تشكل تدفق السائل الدماغي الشوكي، وقد يؤثر تصريف اللمف السحائي المعزز بالضوء بطريقة ما على النظام الدوري الكامل للسائل الدماغي الشوكي لإزالة الجزيئات الكبيرة. بالإضافة إلى ذلك، فقدان شديد لـ LYVE-“تمت ملاحظة منطقة الأوعية في الفئران المسنّة التي خضعت لاستئصال الممرات اللمفية المتوسطة، ولم تكن هناك تغييرات في الممرات اللمفية المتوسطة تحت العلاج بالضوء (الشكل 4i، j). وبالمثل، تم إلغاء النتائج المفيدة للعلاج بالضوء فيما يتعلق بتحسين تنشيط الخلايا الدبقية الصغيرة وحمل الأميلويد في منطقة الحُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُ
تتم إدارة التوازن الميتوكوندري والوظائف الخلوية لخلايا بطانة اللمف السحائي (mLECs) بواسطة الضوء
للمزيد من التحقيق فيما إذا كان التأثير المفيد للضوء على تصريف mLVs يعتمد على التعديل الوظيفي لـ mLECs، تم دراسة الهياكل الدقيقة داخل mLECs للذكورتمت رؤية الفئران بواسطة المجهر الإلكتروني الناقل (TEM). كما هو موضح في الشكل 5a، بالمقارنة مع الفئران WT المتطابقة في العمر، كانت خلايا الميكروفاج (mLECs) داخل الأوعية اللمفاوية (mLVs) في فئران AD مرتبة بشكل أكثر تشتتًا، مع ميتوكوندريا ذات شكل دائري، بالإضافة إلى كريستا ميتوكوندريا مكسورة وضبابية. تعتبر الميتوكوندريا مواقع لتمثيل الطاقة الخلوية، وهي مرتبطة ارتباطًا وثيقًا بالوظائف الخلوية. تعتبر غشاء الكريستا والمصفوفة في الميتوكوندريا مناطق وظيفية رئيسية للتمثيل الغذائي، وبالتالي فإن الشكل غير الطبيعي للميتوكوندريا أو تقليل وضبابية هيكل الكريستا تعكس بشكل مباشر تلف التمثيل الغذائي.بشكل مفاجئ، في الفئران المصابة بمرض الزهايمر المعالجة بالضوء، كانت خلايا اللمف اللمفاوية المتوسطة (mLECs) أكثر انتظامًا وكثافة، تقريبًا مثل الأقارب من النوع البري، مما يشير إلى أن الضوء قد ينقذ من اضطراب الوصلات في mLEC. في الوقت نفسه، أظهرت الفئران المصابة بمرض الزهايمر المعالجة بالضوء ميتوكوندريا ذات شكل بيضاوي أو على شكل قضيب مع كريستا أكثر انتظامًا ووضوحًا في mLECs، بالإضافة إلى زيادة في طول الميتوكوندريا، مما يكشف عن استعادة وظيفة الميتوكوندريا بواسطة الضوء (الشكل 5 أ، ب). لتقييم توازن الميتوكوندريا بشكل أكبر في mLECs للفئران المعالجة بالضوء، قمنا بقياس إنتاج السوبر أكسيد الميتوكوندري وكذلك جهد الغشاء الميتوكوندري باستخدام مؤشر السوبر أكسيد الميتوكوندري (MitoSOX) ومسبار التترا ميثيل رودامين (TMRE)، على التوالي. أظهرت هذه النتائج زيادة في إنتاج السوبر أكسيد الميتوكوندري وانخفاض في جهد الغشاء الميتوكوندري في mLECs للفئران المصابة بمرض الزهايمر والفئران المسنّة، مما يشير إلى خلل في وظيفة الميتوكوندريا في mLECs. أنقذت معالجة الضوء الشذوذات الميتوكوندرية في mLECs مع تقليل إنتاج السوبر أكسيد الميتوكوندري وزيادة في جهد الغشاء الميتوكوندري (الشكل التكميلية 11 أ-و). بشكل عام، يعزز الضوء استعادة الأيض الميتوكوندري في mLECs للحفاظ على وظيفة الخلايا، مع الحفاظ على الهيكل الكامل والترتيب المضغوط لـ mLV.
علاوة على ذلك، تم التحقق من وظيفة الخلايا لـ mLVs التي تم تعديلها بواسطة الضوء باستخدام تحليل RNA-seq. تم استخراج RNA الكلي من أنسجة السحايا وتم تسلسلها. أظهر مخطط البركان وخريطة الحرارة فرقًا كبيرًا في تعبير الجينات في السحايا لـالفئران المعالجة بالضوء أو بدون ضوء (الشكل التكميلي 11g، h). أظهر إثراء GO لجينات التوقيع زيادة في تنظيم تكاثر الخلايا، والالتصاق، والتمايز، والتطور، والتوازن، بالإضافة إلى النقل في السحايا لدى الفئران المصابة بمرض الزهايمر المعالجة بالضوء.
الشكل 4 | تأثيرات الضوء على الإدراك وتصريف اللمف في المLVs-
فئران مُزالة. جدول العلاجات واختبارات السلوك لفئران مسنّة مُزالة mLVs وفئران 5xFAD. ب، ج مؤشر التعرف على الموقع في اختبار NOL (ب) ومؤشر التعرف على الكائن في اختبار NOR (ج) لفئران مسنّة مُزالة mLVs.الفئران في كل مجموعة. د-و مؤشر التعرف على الموقع في اختبار NOL (د)، مؤشر التعرف على الكائن في اختبار NOR (هـ) والمدة الزمنية للعثور على المنصة في اكتساب اختبار MWM (و) لفئران mLVsablated 5xFAD.الفئران في كل مجموعة في البياناتالفئران في مجموعة التحكم،الفئران في مجموعة الضوء،الفئران في الرؤية/الصورة. ومجموعات الضوء في البيانات. صور تمثيلية للعقد اللمفية السطحية من الأوعية اللمفية المعالجةفئران FAD مع تراكم OVAICG (مُعلمة بدوائر حمراء) في نقاط زمنية مختلفة بعد الحقن (i.c.m.) (من نسختين مكررتين). شريط القياستQuantification of fluorescence شدة OVA-ICG في العقد اللمفية السطحية.فئران في Ctrl، الرؤية/الصورة، والرؤية/الصورة + مجموعات الضوء،الفئران في مجموعة الضوء.صور تمثيلية للأغشية السحائية للفئران المسنّة التي تم استئصالها والمصبوغة بـ LYVE-1 و DAPI (من نسختين مكررتين). مقياس الرسم = أو . تحديد نسبة مساحة LYVE-1الأوعية اللمفاوية.الفئران في مجموعات التحكم والصورة.الفئران في مجموعة الضوء،فئران في التصوير/الصورة. + مجموعة الضوء. ك صور تمثيلية لمقاطع الدماغ من mLVsablatedفئران FAD في HPC مصبوغة بـ Iba1،و DAPI (من نسختين متماثلتين).فئران في كل مجموعة. شريط القياس. بيانات في تُعرض كمتوسطتم استخدام SEM، وتم تحليلها بواسطة ANOVA ثنائية الاتجاه مع اختبار المقارنة المتعددة لسيداك لمقارنات المجموعات المتعددة. تم توفير بيانات المصدر كملف بيانات مصدر.
الشكل 5 | تأثيرات تحسين الضوء على البنية المجهرية والتعبير الجيني لـ mLVs في فئران 5xFAD. أ صور تمثيلية لميكروسكوب الإلكترون الناقل (TEM) لترتيب mLEC وشكل الميتوكوندريا (رؤوس الأسهم الحمراء: ميتوكوندريا mLECs) (من 2 تكرارات). مقياس الرسمأو 200 نانومتر. ب قياس طول الميتوكوندريا في خلايا المصل اللمفية.الفئران في كل مجموعة. ج تعبير Nckap1l وNck1 وCalm2 وCdc42 وVegfc في السحايا بين مجموعة AD المعالجة بالضوء ومجموعة AD. د مخطط إثراء لمجموعات الجينات الرئيسية بواسطة GSEA للسحايا، يظهر ملف درجة الإثراء الجارية ومواقع أعضاء مجموعة الجينات في القائمة المرتبة. هـ مخطط انتشار للجينات المثرية في الفسفرة التأكسدية مرتبة وفقًا لـ (FC: تغيير الطي).
تعبير Lyve1 و Prox1 و Pecam1 و Tgfa في خلايا المصل اللمفية المتوسطة (mLECs) بين مجموعة مرض الزهايمر المعالجة بالضوء ومجموعة مرض الزهايمر. خريطة حرارية تظهر مستويات التعبير النسبية للجينات المعبر عنها بشكل مختلف (DEGs) في خلايا المصل اللمفية المتوسطة (mLECs) المعنية بعملية الأيض الميتوكوندري.الفئران في كل مجموعة في البيانات ج-ط. البيانات فيتُعرض كمتوسطتم تحليل SEM باستخدام تحليل التباين الأحادي الاتجاه مع اختبار المقارنات المتعددة لسيداك للمقارنات بين مجموعات متعددة (ب)، أو اختبار t لطلاب غير المقترنين ذو الذيلين للمقارنات بين مجموعتين (ج، ف).القيم فيتم حسابه باستخدام clusterProfiler باستخدام اختبار التبديل القائم على الظاهرة التجريبية، دون أي تعديلات على المقارنات المتعددة. كانت الاختبارات الإحصائية تتضمن تحليلًا ثنائي الجانب. تم توفير بيانات المصدر كملف بيانات مصدر.
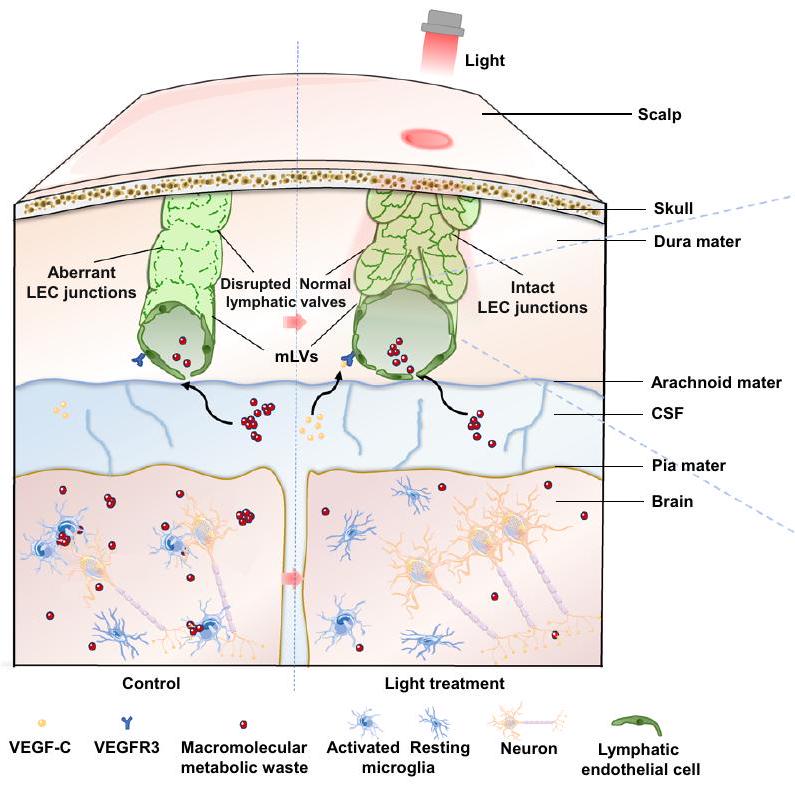
الشكل 6 | توضيح تخطيطي للآلية التي تستهدف بها العلاج الضوئي باستخدام mLVs لتحسين علم الأمراض المرتبط بمرض الزهايمر. يعزز الضوء القريب من الأشعة تحت الحمراء التنفس الميتوكوندري في خلايا الأوعية اللمفاوية عبر مستقبل الضوء CcO، مما يصلح خلايا اللمف.
التقاطعات والترتيبات لاستعادة تصريف المLVs، وتعزيز إزالة الجزيئات الكبيرة من النفاياتوفي النهاية، تخفيف العمليات التنكسية العصبية في الفئران.
(الشكل التوضيحي 11i). بالإضافة إلى ذلك، لم يكن هناك تغيير واضح في تعبير Vegfc (المتعلق بتكوين الأوعية اللمفاوية) في السحايا بعد العلاج بالضوء، بينما كانت الجينات المشاركة في إشارات VEGF بما في ذلك Nckap1l.نكا1كالْم2 و Cdc42تم تنظيمها بشكل ملحوظ (الشكل 5c). علاوة على ذلك، كما هو موضح في الجدول التكميلي 1، تم تنظيم البروتينات الرئيسية لتجميع الوصلات الضيقة بين الخلايا والالتصاق الخلوي، مما يشير إلى أن الوصلات الضيقة بين الخلايا كانت متورطة في التنظيم الإيجابي للأوعية اللمفاوية السحائية بواسطة الضوء. أظهرت النتيجة إمكانية تعزيز تكاثر الخلايا والتصاق الأوعية اللمفاوية السحائية بعد العلاج بالضوء، والتي كانت الأساس لترتيب الأوعية اللمفاوية السحائية بشكل مضغوط. المصطلحات في KEGG التي تم إثراؤها بواسطة الجينات المتغيرة بين المجموعات، سلطت الضوء على التغيرات النسخية في مسار الإشارات لتطور الخلايا، والتواصل الخلوي ومرض الزهايمر. أدى العلاج بالضوء إلى تنظيم مرتفع للجينات المرتبطة بدورة الخلية، وإصلاح الأخطاء، وانقباض العضلات الملساء الوعائية، وتنظيم منخفض للجينات المتعلقة بمرض الزهايمر ومسار التنكس العصبي في السحايا، مما قد يشير إلى تعزيز نشاط الخلايا ووظيفة تصريف النظام التصريف السحائي، فضلاً عن تحسين مرض الزهايمر (الشكل التكميلي 11j-1).
لاستكشاف آلية تعديل mLVs بشكل أعمق، تم إجراء تحليل GSEA باستخدام قاعدة بيانات هالمارك للتعبير الجيني. أظهرت النتائج غنىً كبيرًا في 6 مجموعات جينية مرتفعة التعبير مرتبطة بالفوسفوريلation التأكسدي، والتحكم في دورة الخلية، وإصلاح الحمض النووي، وتنظيم المناعة. ساهم تعبير مجموعة من الجينات المرتبطة بالفوسفوريلation التأكسدي، مثل Cox7c وCox17 وAtp5pb وMfn2 وImmt وIscu كجينات رئيسية مرتفعة التعبير، في نشاط وتركيب سيتوكروم ج (CcO) والنقل الإلكتروني التنفسي، وتخليق أدينوسين ثلاثي الفوسفات (ATP)، واندماج الميتوكوندريا، وتشكيل الكريستا، واستتباب الأيض (الشكل 5d، e). الفوسفوريلation التأكسدي هو استجابة لنقل الطاقة في الميتوكوندريا، والتي ترتبط بنقل الإلكترونات وتنتج ATP لتوفير الطاقة للتخليق الخلوي والأيض. في هذه العملية، تلعب CcO دورًا رئيسيًا كعنصر نهائي في سلسلة التنفس الموجودة في الغشاء الداخلي للميتوكوندريا، والتي تعمل أيضًا ككاشف ضوئي يخضع لتغيير في الشكل الجزيئي ويزيد من نشاط الإنزيم بعد تحفيز الضوء، مما يعزز نقل الإلكترونات ويزيد من إنتاج ATP، وبالتالي يعزز نشاط الخلية ووظيفتها.. لذلك، من المحتمل أن الضوء قد حفز توسع الأوعية اللمفاوية وأدى إلى ترتيب خلايا المصل اللمفاوي المجمعة والمنظمة بواسطة
تعديل CcO من أجل الحفاظ على توازن الميتوكوندريا واستعادة وظيفة الخلايا، مما يساهم في استعادة ديناميات التصريف.
بعد ذلك، من أجل التحقق بشكل أكبر من آلية تعديل mLVs، تم عزل mLEC من السحايا لتحليل RNA-seq (الشكل التكميلي 12a-e). أدى تعديل الضوء إلى زيادة تعبير Lyve1 وProx1 (التي تشفر علامات كلاسيكية لـ LECs)، بالإضافة إلى Tgfa (عامل النمو المحول)، ولكنه لم يغير مستوى تعبير Pecam1 (الذي يشفر علامة الأوعية الدموية) (الشكل 5f)، مما يظهر اللمفاوية الناتجة عن العلاج بالضوء.الفئران. بالإضافة إلى ذلك، كانت الجينات التي تم تنظيمها لأعلى مرتبطة أيضًا بالفوسفوريلات المؤكسدة وعمليات الأيض الميتوكوندرية، وهو ما يتماشى مع تحليل تسلسل RNA السحائي المعالج بالضوء.الفئران (الشكل 5 ج). عرضت مصطلحات KEGG المسارات الغنية بالجينات المنخفضة التنظيم في خلايا الميكروفاج اللمفاوية (mLECs) المعنية بإصلاح القواعد، وإصلاح الأخطاء، والتمثيل الغذائي الحيوي في فئران 5xFAD مقارنة بفئران WT، والتي كانت مرتفعة التنظيم في فئران 5xFAD مع تعديل الضوء (الشكل التكميلي 12f، g). كشفت النتائج أعلاه أن الاستعادة، والتمثيل الحيوي، والتمثيل الغذائي قد تكون وظائف أساسية لخلايا الميكروفاج اللمفاوية (mLEC) لتطوير ونقل اللمف، والتي تأثرت في فئران 5xFAD (6 أشهر)، وربما تم إنقاذها من خلال استعادة وظائف الميتوكوندريا لخلايا الميكروفاج اللمفاوية (mLECs) عبر تعديل الضوء، مما يعيد أيضًا شبكة الأوعية اللمفاوية الصغيرة (mLVs) وقدرة التصريف للحفاظ على توازن الجهاز العصبي المركزي.
نقاش
في هذه الدراسة، أظهرنا أن العلاج الضوئي غير الجراحي عبر الجمجمة يمكن أن يحسن بشكل كبير من الإدراك لدى الفئران المسنّة وفئران الزهايمر من خلال تعزيز استهداف تصريف اللمف السحائي. كما أظهرنا أن الضوء يعزز التوازن الأيضي للميتوكوندريا لتحسين وظيفة خلايا اللمف السحائية، مما أنقذ الوصلات الضعيفة لخلايا اللمف السحائية، مما أدى إلى زيادة تصريف اللمف من أجل إزالةترسب والتهاب عصبي، مما يؤدي إلى تخفيف المرض وتحسين الإدراك في فئران AD (الشكل 6).
تم تأكيد ضعف وظيفة اللمف في السحايا في مجموعة متنوعة من الأمراض العصبية، بما في ذلك الأمراض التنكسية العصبية، الشيخوخة، إصابة الدماغ الرضحية، والعدوى الفيروسية العصبية.. هنا، وجدنا أن الوظيفة الإدراكية وقدرة تصريف اللمف متضررت في الفئران (6 أشهر)، على الرغم من أن تغطية mLVs أظهرت فقط اتجاهًا هبوطيًا غير ذي دلالة، متسقة مع نتائج الدراسات السابقة. (الشكل 3د-و). تشير النتيجة إلى أن حدث تلف في وظيفة تصريف المفاوز اللمفية قبل تدهور التوزيع في الفئران المصابة بمرض الزهايمر.
توزيع الأوعية الدموية السطحية في الأم الجافية يوفر اتجاهًا لاستراتيجية تعديل الأعصاب عبر الجمجمة لتخفيف الأمراض العصبية من خلال تنظيم التصريف. في خلايا الحيوانات، تم اعتبار السيتوكروم ج oxidase (CcO) (المجمع الرابع) كأول مستقبل ضوئي في نافذة بصرية شفافة من الضوء الأحمر (630 نانومتر) إلى الضوء القريب من تحت الأحمر (900 نانومتر). يمكن تعزيز CcO بواسطة الفوتونات لتعزيز كفاءة سلسلة التنفس وتخليق ATP لتحسين نشاط الخلايا وإصلاح الوظائف.. بشكل خاص،لقد وُجد أن له أعمق اختراق مع امتصاص محدود إما عن طريق الماء أو الدم في الأنسجة، مما يجعله أكثر ملاءمة للتعديل عبر الجمجمة من أجل تحسينات مرضية طويلة الأمد.
أكدت الدراسات السابقة أن العلاج بالضوء عبر الجمجمة عند 1267 نانومتر يمكن أن يعزز من إزالة المادة المحقونة.ببتيد من الدماغ، كما هو موضح بواسطة التراكمفي الدماغ ومؤخراً، تم إجراء علاج الضوء عبر الجمجمة عند 1267 نانومتر على ذكور حديثي الولادة والجرذان البالغة لعلاج النزيف داخل البطينات من خلال تعزيز إزالة كريات الدم الحمراء عبر اللمفاويات الدماغية.. ومع ذلك، لا يزال غير واضح كيف ينظم ضوء 1267 نانومتر نقل اللمف. من الضروري تحديد الآلية التنظيمية لتصريف اللمف السحائي بواسطة الضوء وكيف تساهم التغيرات في التصريف في تحسين الإدراك. من خلال مراقبة الهيكل الدقيق لخلايا اللمف السحائية (mLECs) باستخدام تصوير TEM، وجدنا تغييرات في شكل الميتوكوندريا، حيث كانت معظمها بيضاوية أو على شكل قضيب وتوزيع الكريستا منتظم في خلايا mLECs للفئران المصابة بمرض الزهايمر والمعالجة بالضوء، بالإضافة إلى زيادة في طول الميتوكوندريا، مما يشير إلى استعادة وظيفة الميتوكوندريا لتعزيز وظيفة الخلايا (الشكل 5 أ، ب). علاوة على ذلك، أظهر تحليل RNA-seq للسحايا تنظيمًا إيجابيًا للفوسفوريل المؤكسد والتمثيل الغذائي للحفاظ على توازن الخلايا، مما قد يعزز بشكل أكبر و تعبير عن هجرة الخلايا واستعادة الوصلات الالتصاقية البطانية فيالفئران المعرضة للضوء (الشكل التكميلي 11c، الشكل 5c، d والجدول التكميلي 1). بالإضافة إلى ذلك، ساهمت الجينات التي تم تنظيمها لأعلى في الفسفرة التأكسدية المرتبطة بنشاط CcO، ونقل الإلكترونات التنفسية، وتخليق الأدينوزين ثلاثي الفوسفات (ATP) وتوازن الأيض الميتوكوندري (الشكل 5e). علاوة على ذلك، أظهر تحليل RNAseq لـ mLEC التعبير المرتفع للجينات المرتبطة بالفسفرة التأكسدية، والأيض، وتكوين الأوعية اللمفاوية وأكد استعادة الأيض الميتوكوندري وتطور الخلايا لـ mLECs في الفئران المعالجة بالضوء.الفئران (الشكل 5f و g والشكل التكميلي 12f و g). تشير هذه النتائج إلى أن التحفيز الضوئي على CcO قادر على تعزيز الفسفرة التأكسدية للميتوكوندريا، مما يعزز توفر الطاقة وانتقال الإشارات للحفاظ على وظيفة خلايا المصل اللمفية الصغيرة (mLECs)، مما يسهم بشكل أكبر في إصلاح الوصلات البطانية. إن تعديل وظيفة mLEC بواسطة الضوء يعزز قدرة النقل اللمفاوي على إزالة الجزيئات الكبيرة المهدرة، مما يخفف بشكل أكبر من الإصابة المرضية المرتبطة بمرض الزهايمر من خلال تقليلترسب، التهاب عصبي وتلف عصبي، مما يساهم في النهاية في تحسين الإدراك.
من أجل النتائج المفيدة لعلاج الضوء NIR، من الممكن أن تتواجد آليات تنظيمية أخرى في الوقت نفسه. قد يزيد الضوء من نشاط خلايا المناعة الناقلة في الغشاء السحائي، مما ينظم النظام المناعي السحائي لتحقيق التوازن الدماغي. بالإضافة إلى ذلك، قد يكون التنظيم المباشر لنشاط الخلايا العصبية في القشرة الدماغية آلية أخرى محتملة من خلالها ينظم الضوء وظيفة الدماغ لتحسين الإدراك. ومن المثير للاهتمام أن الضوء كان غير فعال في تنظيم الإدراك في الفئران WT، ربما بسبب تأثير السقف للقدرة الحالية على التصريف. (الشكل 3 أ-ج والشكل التكميلية 7 أ-د). قد تلعب الضوء دورًا في الوقاية أو التخفيف من العجز المعرفي اعتمادًا على وقت العلاج. في المستقبل، نخطط للتحقيق فيما إذا كان الحفاظ على وظيفة تصريف اللمف من خلال العلاج الضوئي الدوري في المراحل المبكرة من المرض في نماذج الحيوانات المصابة بمرض الزهايمر يمكن أن يخفف أو يؤخر تقدم المرض.
في دراستنا، أكدنا أن العلاج الضوئي المستهدف لـ mLVs عزز الإدراك في الفئران المسنّة وفئران الزهايمر. علاوة على ذلك، كشفنا أن الضوء يمكن أن يستعيد الأيض الميتوكوندري والوظيفة الخلوية لـ mLECs، مما يساهم في تحسين ترتيب الخلايا لـ mLVs لتحسين ديناميات التصريف لتخفيف الأمراض في فئران الزهايمر. بشكل عام، نقدم علاجًا محتملاً للأمراض العصبية من خلال تعديل نقل mLVs لتنظيف النفايات في الجهاز العصبي المركزي، مما قد يخفف من ضعف الإدراك والأمراض المرتبطة.
طرق
الحيوانات
كانت جميع الفئران من سلالة C57BL/6J التي تم شراؤها من شركة جينزهيه للتكنولوجيا الحيوية المحدودة (يانغتشو، الصين). تم استخدام ذكور وإناث الفئران البرية (WT) التي تتراوح أعمارها بين 15 إلى 17 شهرًا كنموذج لكبار السن، بينما كانت الفئران التي تبلغ من العمر 1.5 شهرًا تمثل مجموعة الشباب المقابلة. تم استخدام نموذجين ذكور من مرض الزهايمر في هذه الدراسة: 5xFAD بعمر 6 أشهر وAPPswe/PS1 بعمر 11 شهرًا.9 من فئران APP/PS1، مع أقرانهم من نفس السلالة غير الحاملة للجين كجموعات مقارنة. يتم رؤية ترسب الأميلويد مصحوبًا بالتهاب الدبق في فئران 5xFAD عند عمر شهرين. يحدث العجز المعرفي في حوالي 5 أشهر من العمر ويبدأ فقدان الخلايا العصبية في مناطق الدماغ التي تحتوي على الأميلويد عند عمر 6 أشهر.في فئران APP/PS1، تتكون ترسبات الأميلويد في الدماغ بين 4 إلى 6 أشهر ويحدث العجز المعرفي بين 6 إلى 10 أشهر.تم تربية الفئران من جميع السلالات تحتدورات متناوبة من الضوء والظلام مع درجة حرارة مضبوطة والرطوبة ( )، وتم السماح لهم بالوصول الحر إلى النظام الغذائي القياسي والماء. تم إجراء علاجات الفئران وفقًا لدليل رعاية واستخدام الحيوانات المخبرية. تم مراجعة جميع الدراسات الحيوانية والموافقة عليها من قبل لجنة رعاية واستخدام الحيوانات المؤسسية في جامعة هاينان (رقم الموافقة: HNUAUCC-2021-00025).
حقن داخل الحجرة الجافية الكبرى (i.c.m.)
تم تخدير الفئران بـتم استخدام الإيزوفلوران مع إضافة الدكسميديتوميدين. تم حلاقة وتنظيف السطح الظهري للرقبة. بعد إجراء شق بطول 1.5 سم في جلد الرقبة في المنتصف، تم سحب طبقات العضلات باستخدام الملقط لفتح الحجرة السحائية الكبرى. تم حقن المحلول المطلوب في حجرة السحائية الكبرى.لتقييم تصريف اللمف،من مركب أليكسافلور 647 مع البيض الزلال (عندأو OVA-A647، ثيرمو فيشر ساينتيفيك، الولايات المتحدة الأمريكية) أو OVA-ICG (عندتم إعطاء الحقنة باستخدام حقنة هاملتون. بعد الحقن، تم تثبيت الإبرة في مكانها لمدة إضافية قدرها دقيقتان لتجنب تدفق السائل الدماغي الشوكي إلى الوراء. أخيرًا، تم تنظيف جلد ورقبة الفأر وخياطته، وتم وضع الفئران على وسادة تدفئة للتعافي من التخدير حتى استعادة الوعي، ثم تم نقلها مرة أخرى إلى قفصها.
استئصال الأوعية اللمفاوية السحائية (mLVs)
تم إجراء علاج فيسودين وفقًا للبروتوكولات المنشورة.تم تخدير الفئران بـالإيزوفلورين المدعوم بالدكسمديتوميدين. ثمفيسودين، APExBIO، الولايات المتحدة الأمريكية) تم حقنه (داخل العضلة) بمعدلبعد 15 دقيقة، تم تحويل الفيزودين ضوئيًا باستخدام ضوء غير حراري بطول موجي 689 نانومتر (LSR689CP-3.6W-FC، LASEVER، الصين) على خمسة مواقع مختلفة على الجمجمة (واحد في موقع الحقن، واحد عند التقاء الجيوب، اثنان عند الجيوب المستعرضة الثنائية وواحد عند الجيب السهمي العلوي)، معإشعاعتم خياطة جلد فروة الرأس وتم وضع الفئران على وسادة تدفئة للتعافي من التخدير حتى استعادوا وعيهم، ثم تم نقلهم مرة أخرى إلى قفصهم المنزلي.
قياس اختراق الضوء
تم euthanizing الفئران عن طريق خلع العنق وتم حلاقة فروة الرأس وتنظيفها باستخدام شفرة الحلاقة وكريم إزالة الشعر. ثم تم عزل الرأس وإزالة الفك السفلي ونسج الدماغ، وتم جمع الجمجمة مع فروة الرأس الخالية من الشعر. تم تثبيت الجمجمة تحت مصدر ضوء 808 نانومتر. تم قياس اختراقات الجمجمة مع فروة الرأس الخالية من الشعر عند الطول الموجي. تم تسجيل النقطة تحت الضوء عند قوى مختلفة (القوى المكتشفة بواسطة مقياس الطاقة والقدرة البصرية المحمولة الرقمية (PM100D، ثورلابس إنك، نيوتن، نيو جيرسي، الولايات المتحدة الأمريكية)). تم حساب النفاذية كتيار الإشعاع/القدرة الساقطة.
علاج الضوء
تم إدخال الفئران إلى غرفة التجارب قبل 30 دقيقة على الأقل من علاج الضوء للتكيف. تم تخدير الفئران بـتم استخدام الإيزوفلورين، وتم حلاقة وتنظيف فروة الرأس الظهرية. ثم تم تطبيق ضوء مستمر بطول موجي 808 نانومتر (LSR808H-5W-FC، LASEVER، الصين) لإضاءة الجمجمة مع فروة رأس خالية من الشعر على مساحة من، في لمدة 10 دقائق، والتي انتهت بجرعات منتم قياس درجة حرارة سطح فروة الرأس خلال 10 دقائق من التعرض للإشعاع الضوئي باستخدام كاميرا حرارية قريبة من الأشعة تحت الحمراء (NIR) (226S، FOTRIC، الصين) وتم تحليلها باستخدام AnalyzlR (الإصدار 4.3.2.71). خضعت الفئران لعلاج ضوئي ثلاث مرات في الأسبوع لمدة أربعة أسابيع متتالية. بالنسبة للفئران في مجموعة التحكم، تم تنفيذ نفس الإجراء التجريبي. تم تخدير الفئران باستخدام الإيزوفلورين، وتم حلاقة وتنظيف فروة الرأس الظهرية، ثم خضعت للإضاءة عبر الجمجمة بواسطة إضاءة داخلية لمدة 10 دقائق. بعد العلاجات، تم وضع الفئران على وسادة تدفئة للتعافي من التخدير حتى تستعيد وعيها، ثم تم نقلها مرة أخرى إلى قفصها المنزلي.
تصوير العقد اللمفية العنقية (CLNs)
تم تخدير الفئران باستخدام 1% إيزوفلوران مع إضافة دكسمديتوميدين، وتم استئصال جلد الرقبة لكشف العقد اللمفاوية العنقية. تم تنظيف السطح الظهري للرقبة وشقه. تم سحب طبقات العضلات إلى الجانبين لكشف الحويصلة الكبيرة.من OVA-ICG (عندتم إعطاؤه (i.c.m.) للفئران فيباستخدام حقنة هاملتون. بعد الحقن، تم تثبيت الإبرة لمدة إضافية قدرها دقيقتان لتجنب تدفق السائل الدماغي الشوكي إلى الوراء. أخيرًا، تم تنظيف جلد ورقبة الفأر وعضلاته وخياطتها، وتم السماح للفئران بالتعافي على وسادة تدفئة معصيانة الإيزوفلورين. بعد خمس عشرة دقيقة من بدء الحقن، تم وضع الفئران في وضع الاستلقاء تحت نظام التصوير الحي MARS (أرتيميس إنتليجنت إيميجينغ، الصين) للتصوير. تم جمع صور العقد اللمفاوية العنقية (CLNs) بعد 15 دقيقة إلى ساعة واحدة من بدء الحقن، مع فواصل زمنية قدرها 15 دقيقة. تم تسجيل شدة الفلورسنت المعتمدة على الزمن للمتتبعات المتراكمة في العقد اللمفاوية العنقية. بعد ساعتين من بدء الحقن، تم ضخ الفئران عبر القلب بمحلول PBS وبارافورمالدهيد (PFA). تم قطع العضلات العنقية وسحبها، وتم جمع العقد اللمفاوية السطحية والعميقة (sCLNs و dCLNs) لتصويرها بواسطة نظام التصوير. تم إجراء تحليل كمي لشدة الفلورسنت باستخدام برنامج فيجي (الإصدار 1.53 q) وLightField (الإصدار 6.14) لتوصيف وظيفة تصريف اللمف.
تصوير تباين النقاط الليزرية (LSCI)
بعد علاج استمر لمدة شهر، تم تخدير الفئران باستخدام 1% إيزوفلوران مضاف إليه دكسمديتوميدين، ثم تم استئصال فروة الرأس واللفافة لكشف الجمجمة. بعد ذلك، تم تثبيت رأس الفئران باستخدام إطار تثبيت الرأس ووضعها مستلقية تحت ميكروسكوب LSCI (سيموبتو، الصين) للكشف عن تدفق الدم في الدماغ. تم تضخيم الإشارات المثارة بواسطة ليزر 785 نانومتر ورقمنتها لتصوير تدفق الدم. تم تسجيل تدفق الدم النسبي (%) للفئران لمدة 3 دقائق متواصلة وتم حساب مؤشر تدفق الدم (وحدات التروية، PU) بواسطة برنامج SIM BFI (الإصدار 3.1.45، سيموبتو).
اختبار الحقل المفتوح (OF)
تم إجراء اختبار OF في ساحة مكعبة غير شفافة (بجانب 40 سم). قبل الاختبار، تم تعويد الفئران على غرفة الاختبار لمدة 30 دقيقة. ثم تم وضع الفئران في الساحة وسمح لها بالاستكشاف لمدة 10 دقائق. تم تسجيل وتحليل المسافة الإجمالية (سم)، السرعة (سم/ث) والوقت المستغرق في المركز (%) باستخدام نظام تحليل سلوك DigBehv (الإصدار 4.1، شركة جيليان للبرمجيات والتكنولوجيا المحدودة).
اختبار التعرف على الكائنات الجديدة/اختبار موقع الكائنات الجديدة (NOR/NOL)
تم إجراء اختبار NOR أو NOL في نفس الساحة المكعبة المستخدمة في اختبار OF. قبل 24 ساعة من الاختبار، تم وضع الفئران في الساحة لاستكشافها لمدة 10 دقائق كعملية تأقلم. ثم تم وضع جسمين متطابقين في زوايا مختلفة، على بعد حوالي 6 سم من الجدران. خلال التدريب، تم السماح للفئران باستكشاف الأجسام في الساحة لمدة 10 دقائق ثم العودة إلى أقفاصها المنزلية. بعد 1 أو 24 ساعة، تم اختبار الذاكرة لموقع الجسم أو التعرف عليه. في اختبار NOL، تم السماح للفئران باستكشاف الأجسام لمدة 10 دقائق في نفس الساحة مع نقل جسم واحد إلى موقع جديد. في اختبار NOR، تم السماح للفئران بالاستكشاف لمدة 10 دقائق في نفس الساحة مع استبدال جسم واحد بجسم جديد في الموقع الأصلي. تم تسجيل استكشاف الأجسام باستخدام برنامج Visutrack (الإصدار 3.0، شركة Xinruan لتكنولوجيا المعلومات المحدودة) وتم حساب مؤشر التعرف (RI) ومؤشر التمييز (DI) وتفضيل الجسم (نسبة الوقت مع الجسم).تفضيل الكائن أو .
اختبار متاهة Y
تم إجراء اختبار المتاهة على شكل حرف Y في متاهة على شكل حرف Y تحتوي على ثلاثة أذرع متطابقة، كل منها بطول 35 سم، وعرض 6 سم، وارتفاع 15 سم. تم وضع أشكال هندسية مختلفة داخل كل ذراع كمعالم مكانية. تم تعيين الأذرع بشكل عشوائي كذراع جديدة، وذراع البداية، وذراع أخرى. خلال التدريب، تم استخدام فاصل لحجب الذراع الجديدة. تم وضع الفئران في ذراع البداية من المتاهة، متجهة بعيدًا عن مركز المتاهة، من أجل ““استكشاف. في تجربة الاختبار، تم وضع الفئران في متاهة Y بعد ساعة واحدة لاستكشافها لمدة 5 دقائق مع فتح جميع الأذرع للوصول الحر. تم تسجيل الوقت الذي قضاه كل فأر في كل ذراع باستخدام نظام تحليل السلوك DigBehv (الإصدار 4.1، شركة جيليانغ للبرمجيات والتكنولوجيا المحدودة) وتم حساب نسبة الوقت الذي قضاه في الذراع الجديدة (%)”.
اختبار متاهة مورس المائية (MWM)
تم إجراء اختبار MWM في بركة دائرية (قطرها 120 سم) مملوءة بالماء غير الشفاف عند حواليإلى عمق 30 سم. تم وضع أشكال هندسية مختلفة حول المسبح كمعالم مرجعية مكانية. خلال خمسة أيام تدريب متتالية، تم وضع الفئران في الخزان للعثور على منصة ثابتة مخفية (بقطر 10 سم، تقع على عمق 1 سم تحت سطح الماء) خلال 60 ثانية. الفئران التي فشلت في العثور على المنصة خلال 60 ثانية تم توجيهها بلطف لتسلق المنصة والراحة لمدة 15 ثانية. تم تسجيل زمن الوصول إلى المنصة باستخدام برنامج Visutrack (الإصدار 3.0، شركة Xinruan لتكنولوجيا المعلومات المحدودة) وتم حساب متوسط زمن الوصول إلى المنصة من التجارب الأربعة كل يوم. في اليوم السادس (تجربة الاستكشاف)، تمت إزالة المنصة وأجرت الفئران استكشافًا لمدة 60 ثانية. تم حساب نسبة الوقت المستغرق في كل ربع (%)، وعدد العبور لمنطقة المنصة وسرعة السباحة (تم تسجيلها وتحليلها.
جمع الأنسجة
تم تخدير الفئران ثم تم ضخها عبر القلب بمحلول PBS و4% PFA. تم قطع وإبعاد جلد ورقبة الفئران، وتم جمع العقد اللمفاوية العنقية وتثبيتها فييرجى العثور على المرفق لمدة 24 ساعة فيتمت إزالة الجلد والعضلات من الرؤوس، ثم تم جمع الأدمغة والجمجمة وتثبيتها فييرجى العثور على المرفق لمدة 24 ساعة في. ثم تم إزالة السحايا بالكامل بعناية من الجماجم ثم تم تخزينها في PBS عندحتى الاستخدام لاحقًا. تم غسل الأدمغة الثابتة وCLNs بمحلول PBS، وتجفيفها بـ، و السكروز لمدة ثلاثة أيام ومجمد في مركب Tissue-Tek OCT (ساكورا، الولايات المتحدة الأمريكية) عند تم تقطيع الأدمغةالأقسام) وتم الاحتفاظ بها في مادة حماية التجميد عندللاستخدام لاحقًا.
علم المناعة النسيجية والتصوير
لإعادة استرجاع المستضد، تم تسخين مقاطع الدماغ أو مقاطع العقد اللمفاوية الجانبية باستخدام محلول سترات لمدة 30 دقيقة. بالنسبة لصبغة المناعة الفلورية، تم غسل مقاطع الدماغ ومقاطع العقد اللمفاوية الجانبية والأغشية السحائية بمحلول PBS و تم حجبها بمحلول PBS يحتوي على 0.3% من Triton X-100 (سيغما، الولايات المتحدة) و5% من ألبومين مصل البقر (BSA، سيغما) لمدة ساعة واحدة في درجة حرارة الغرفة، تلاها حضانة مع تخفيفات الأجسام المضادة الأولية طوال الليل في : فأر مضاد-LYVE-1 (رقم الكات# 14-0443-82، رقم الدفعة# 2494966، النسخة# ALY7، 1:200، إنفيتروجين، الولايات المتحدة الأمريكية)، فأر مضاد للأميلويد- (رقم الكات 805501، رقم الدفعة B326654، رقم النسخة 12F4، 1:1000، Biolegend، الولايات المتحدة الأمريكية)، الأجسام المضادة للأرنب ضد Iba1 (رقم الكات 019-19741، رقم الدفعة LEN4341، 1:500، Abcam، المملكة المتحدة)، الأجسام المضادة للفأر ضد NeuN (رقم الكات ab279297، رقم الدفعة GR3400636-6، رقم النسخة EPR12763، 1:1000، Abcam، المملكة المتحدة)، الأجسام المضادة للأرنب ضد Syn (رقم الكات MA5-14532، رقم الدفعة WI3369463، رقم النسخة SP11، 1:100، Invitrogen)، الأجسام المضادة للفأر ضد MAP2 (رقم الكات MA5-12826، رقم الدفعة XF3603732A، رقم النسخة AP18، 1:200، Invitrogen)، الأجسام المضادة للأرنب ضد CD31 (رقم الكات SAB4502167100UG، رقم الدفعة 210547، 1:200، Sigma). ثم تم غسل الأنسجة ثلاث مرات بمحلول PBS، تلا ذلك حضانة مع الأجسام المضادة الثانوية Alexa Fluor 488/555/633 ضد الفأر/الأرنب/الفأر IgG (H+L) في PBS يحتوي على 0.3% Triton X-100 لمدة ساعة واحدة في الظلام عند درجة حرارة الغرفة: الأجسام المضادة الثانوية Alexa Fluor 488 ضد الفأر IgG (H + L) (رقم الكات A-11006، رقم الدفعة 2247986، 1:500، Invitrogen)، الأجسام المضادة الثانوية Alexa Fluor 488 ضد الأرنب IgG (الأجسام المضادة الثانوية (رقم الكات# A-32731، رقم الدفعة# VG302077، 1:500، إنفيتروجن)، أليكسا فلور 555 ماعز مضاد للفأر IgG (H + L) الجسم المضاد الثانوي (رقم الكات# A21434، رقم الدفعة# 2184321، 1:500، إنفيتروجن)، أليكسا فلور 555 ماعز مضاد للأرنب IgG (H + L) الجسم المضاد الثانوي (رقم الكات# A-32732، رقم الدفعة# YA361054، 1:500، إنفيتروجن)، وأليكسا فلور 633 ماعز مضاد للفأر IgG (H + L) الجسم المضاد الثانوي (رقم الكات# A-21052، رقم الدفعة# 2418505، 1:500، إنفيتروجن). ثم تم غسل الأنسجة ثلاث مرات وتمت معالجتها مع DAPI لمدة 5 دقائق في درجة حرارة الغرفة. أخيرًا، تم تثبيت الأنسجة باستخدام SlowFade Diamond Antifade Mountant (إنفيتروجن) وتم تصويرها بواسطة مجهر المسح بالليزر (FV3000، أوليمبوس، اليابان) أو ماسح الشرائح (VS200، أوليمبوس). تم تصدير الصور وتحليلها الكمي باستخدام برنامج OLYMPUS OlyVIA (الإصدار 4.1) وبرنامج Fiji (الإصدار 1.53q).
قياس السوبر أكسيد الميتوكوندريا وإمكانات غشاء الميتوكوندريا
تم استخدام MitoSOX Red (Invitrogen) ومؤشر التترا ميثيل رودامين (TMRE، Invitrogen) لتقييم مستويات السوبر أكسيد الميتوكوندريا وإمكانات غشاء الميتوكوندريا في خلايا بطانة الأوعية اللمفاوية السحائية (mLECs). تم جمع السحايا الكاملة الطازجة واحتضانها في محلول MitoSOX لمدة 10 دقائق أو في محلول TMRE لمدة 30 دقيقة في، تليها فترة تحضين بتخفيف من الأجسام المضادة الفأرية Alexa Fluor 488 ضد LYVE-1 (رقم الكاتالوج# 53-0443-80، رقم الدفعة# 2547846، النسخة# ALY7، 1:250، Invitrogen) لمدة 30 دقيقة فيتم تصوير MLECs بواسطة ميكروسكوب ضوئي متماسك (FV3000، أوليمبوس).
المجهر الإلكتروني الناقل (TEM)
تم euthanizing الفئران عن طريق خلع الرقبة، ثم تم تشريح قشور الجمجمة وتم إزالة السحايا بالكامل. تم تقليم مناطق MLVs من السحايا إلىتم قطع الأنسجة بشفرة وغسلها مرتين بمحلول PBS. ثم تم تثبيت الأنسجة بـمحلول عازل الجلوتارالدهيد معبارافورمالدهيد لمدة 30 دقيقة في درجة حرارة الغرفة وتم تخزينه فيحتى يتم معالجتها كعينات TEM. ثم تم شطف الأنسجة بمحلول PBS، وثم تم تثبيتها بـ رباعي أكسيد الأوزميوم (OsO4)، تم شطفه بماء مزدوج التقطير، وتجفيفه بالإيثانول، وتخلله الأسيتون، وتم تضمينه في راتنج الإيبوكسي. تم قطع العينات إلى أقسام بسمك 70 نانومتر، وتم صبغها بأسيتات اليورانيوم وسترات الرصاص، ثم تم تصويرها باستخدام مجهر إلكتروني نافذ (Tecnai G2 Spirit، FEI، الولايات المتحدة الأمريكية). تم إجراء تحليل كمي لطول الميتوكوندريا في خلايا بطانة اللمف السحائي (mLECs) باستخدام برنامج Fiji. تم الحصول على صور خلايا LECs في السحايا باستخدام المجهر الإلكتروني النافذ وتم حساب متوسط قياسات طول الميتوكوندريا لعشر قياسات فردية لكل عينة في مجموعات مختلفة. ).
فرز خلايا الميكروفاج اللمفاوية
للحصول على تعليق من خلايا الميكروغليا من فئران 5xFAD باستخدام فرز الخلايا المعتمد على الفلورية (FACS)، تم تخدير الفئران ثم تم غسلها عبر القلب بمحلول PBS. تم تشريح أغطية الجمجمة وتم إزالة السحايا بالكامل في وسط دوبلكو المعدل من إيجل. (DMEM، جيبكو، الولايات المتحدة الأمريكية)، مزود بـFBS (جيبكو)، بنسلين (مل ، جيبكو) وستربتوميسين (، سيغما). ثم تم حضن السحايا في DMEM يحتوي علىمن الكولاجيناز الثامن (سيغما) ومن DNase I (سيغما) لمدة 30 دقيقة عندتم تصفية السحايا المهضومة من خلالشبكة نايلون للخلايا المفردة. ثم تم طرد الخلايا (280 جرام) عندلمدة 10 دقائق ثم أعيد تعليقها في PBS. بعد ذلك، تم حضن الخلايا مع تخفيفات الأجسام المضادة في لمدة 30 دقيقة: فأر مضاد CD45-PE (رقم الكاتالوج 12-0451-82، رقم الدفعة 2356222، النسخة 30-F11، 1:150، eBioscience)، فأر مضاد CD31-FITC (رقم الكاتالوج 11-0311-85، رقم الدفعة 2373735، النسخة 390، 1:100، eBioscience)، هامستر مضاد بودوبلانين-PE-Cy7 (رقم الكاتالوج 127412، رقم الدفعة B355692، النسخة 8.1.1، 1:100، Biolegend) وفأر مضاد CD11b-PerCP (رقم الكاتالوج 101230، رقم الدفعة B352939، النسخة M1/70، 1:80، BioLegend)، وأخيرًا تم الحضانة مع DAPI لمدة 5 دقائق. ثم تم إعادة تعليق الخلايا في محلول FACS. تم تحديد الخلايا الفردية باستخدام عرض النبضة من التشتت الجانبي والتشتت الأمامي. تم التعرف على الخلايا السلبية لـ DAPI واختيارها كخلايا قابلة للحياة. الخلية الفردية تم تصنيف الخلايا وفرزها باستخدام تقنية تحليل تدفق الخلايا (MoFlo Astrios، بيكمان كولتر، الولايات المتحدة الأمريكية). تم توزيع خلايا العينة في القطرات بحجموتم إسقاطها في أنابيب PCR التي تحتوي على 4 ميكرولترات من محلول التحلل. تم تخزين العينات مؤقتًا فيللمزيد من التحليل.
استخراج RNA وتسلسلها
تمت معالجة جميع عينات RNA (بما في ذلك توليد مكتبة cDNA) وتسلسل RNA (RNA-seq) بواسطة شركة E-GENE Technology Co., Ltd (شنتشن، الصين). تم إعداد مكتبات RNA-seq للعينات باستخدام بروتوكول Smart-seq2.“مع تعديلات طفيفة. بالنسبة لعينات الأنسجة، تم استخراج RNA الكلي من كل عينة من نسيج الحُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُُ ) أو نسيج واحد من السحايا ( ) عبر مادة استخلاص RNA الكلي TRIzol (Thermo Fisher Scientific). ثم تم قياس كمية RNA للعينة باستخدام جهاز Qubit Fluorometer وتم الكشف عن سلامة RNA بواسطة الرحلان الكهربائي في هلام الأجاروز. بالنسبة لعينات LEC، تم فرز خلايا LEC (التي تم فرزها مسبقًا بواسطة FACS من غشاء واحد فردي،تم تحللها بـمنترايتونمحلول يحتوي علىمثبط RNase. تم خلط RNA الكلي المستخرج أو تحلل الخلايا معمنبادئة Oligo-dT لتغنيت RNA، وتم إخضاعها لعملية النسخ العكسي في نظام عازل الشريط الأول Super Script II بتركيز 1X مع 200 وحدة من إنزيم النسخ العكسي Super Script II.قالب تبديل أوليغو (TSO،20 وحدة مثبط RNase،ديثيوثريتال (DTT، 100 مللي مولار)،من البيتين (5 م) ومنتم تخليق cDNA الكامل وتضخيمه من خلال 10 إلى 15 دورة من تضخيم PCR. ثم تم تنقية المنتج واستخدامه في بناء المكتبة. بعد قياسه بواسطة جهاز Agilent 2100 Bioanalyzer (Agilent Technologies، الولايات المتحدة الأمريكية)، تم تسلسل المكتبة باستخدام منصة Illumina Novaseq.
تم تصفية القراءات الخام بواسطة fastp (الإصدار 0.18.0) لإزالة المحولات أو القواعد ذات الجودة المنخفضة للتركيب والتحليل التالي. تم حساب قيمة الشظية لكل كيلوباز من النسخة لكل مليون قراءة موجهة (FPKM) لقياس وفرة التعبير وت variations النسخ باستخدام RSEMالبرمجيات. تم إجراء جميع التحليلات لتصور بيانات RNA-seq باستخدام البرنامج الإحصائي R (الإصدار 4.3.1). تم إجراء تحليل التعبير التفاضلي وتصور النتائج باستخدام حزمة R DESeq. (الإصدار 1.40.2) و ggplot2 (الإصدار 3.4.2). الجينات التي لديها قيمة تغيير مضاعف أكبر من 2.0 أو أقل من 0.5 ومعدل الاكتشاف الخاطئ (FDR) المصححتم اعتبارها جينات معبرة بشكل مختلف بشكل كبير (DEGs). قدم تحليل إثراء مصطلحات علم الجينات (GO) ومسارات موسوعة كيوتو للجينات والجنوم (KEGG) مصطلحات GO و KEGG التي كانت غنية بشكل كبير في DEGs مقارنة بخلفية الجينوم، وقام بتصفية DEGs التي تت correspond إلى الوظائف البيولوجية أو المسارات. تم إجراء تحليل إثراء مصطلحات GO و KEGG باستخدام برنامج clusterProfiler (الإصدار 4.8.3). تم تنفيذ تشابه مصطلحات GO بناءً على مصفوفات التشابه للمصطلحات الوظيفية باستخدام حزم R simplifyEnrichment (الإصدار 1.10.0). من أجل الحصول على فهم أفضل للآلية، تم إجراء تحليل إثراء مجموعة الجينات. تم إجراء تحليل مجموعة الجينات (GSEA) من خلال ترتيب جميع الجينات بناءً على مستوى اختلافها باستخدام حزمة R clusterProfiler (الإصدار 4.8.3) وتمت visualized باستخدامحزمة enrichplot (الإصدار 1.20.3). تم الحصول على مجموعات الجينات المميزة التي عملت كمجموعات مرجعية لتحليل المسار المعتمد على التعبير الجيني (GSEA) من قاعدة بيانات التوقيعات الجزيئية.https://www.gsea-msigdb.org/gsea/تم تصور خريطة الحرارة باستخدام ggplot2 (الإصدار 3.4.2) و pheatmap (الإصدار 1.0.12). لم نقم بإجراء أي تعديلات على البيانات، وتم تسلسل البيانات على نفس المنصة ومعالجتها بنفس الطريقة.
التحليل الإحصائي
لضمان العشوائية، تم تقسيم الحيوانات من أقفاص مختلفة في نفس المجموعة التجريبية. كان الباحثون غير مدركين لتوزيع المجموعات للفئران في جميع التجارب خلال جمع البيانات وتحليلها. تم إجراء الدلالة الإحصائية باستخدام اختبار Student غير المقترن ذو الطرفين.اختبار لمقارنات المجموعتين، وتحليل التباين الأحادي أو تحليل التباين الثنائي مع اختبار المقارنات المتعددة لسيداك لمقارنات المجموعات المتعددة.تم اعتبار القيمة الأقل من 0.05 ذات دلالة إحصائية. تُعرض البيانات كمتوسطتم تحليل البيانات الإحصائية باستخدام برنامج GraphPad Prism (الإصدار 8.4.2، شركة GraphPad Software، Inc.).
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة نيتشر المرتبط بهذه المقالة.
توفر البيانات
البيانات التي تدعم نتائج هذه الدراسة متاحة ضمن المقال، وملفات المعلومات التكميلية وملفات بيانات المصدر. يمكن العثور على بيانات تسلسل RNA التي تم إنشاؤها لهذه الدراسة في مستودع GEO تحت رقم الوصول GSE245658. تم الحصول على مجموعات الجينات المميزة التي خدمت كمجموعات مرجعية لتحليل GSEA في تحليل تسلسل RNA من قاعدة بيانات التوقيعات الجزيئية.https://www.gseamsigdb.org/gsea/msigdb/تم توفير بيانات المصدر مع هذه الورقة.
توفر الشيفرة
لم يتم استخدام رموز أو خوارزميات مخصصة لتوليد النتائج في الدراسة الحالية.
References
Hasek, M., Chutna, J., Sládecek, M. & Lodin, Z. Immunological tolerance and tumor allografts in the brain. Nature 268, 68-69 (1977).
Louveau, A. et al. Structural and functional features of central nervous system lymphatic vessels. Nature 523, 337-341 (2015).
Aspelund, A. et al. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 212, 991-999 (2015).
Da Mesquita, S. et al. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature 560, 185-191 (2018).
Da Mesquita, S. et al. Meningeal lymphatics affect microglia responses and anti-Aß immunotherapy. Nature 593, 255-260 (2021).
Ding, X. et al. Impaired meningeal lymphatic drainage in patients with idiopathic Parkinson’s disease. Nat. Med. 27, 411-418 (2021).
Bolte, A. C. et al. Meningeal lymphatic dysfunction exacerbates traumatic brain injury pathogenesis. Nat. Commun. 11, 4524 (2020).
Chen, J. et al. Meningeal lymphatics clear erythrocytes that arise from subarachnoid hemorrhage. Nat. Commun. 11, 3159 (2020).
Li, X. et al. Meningeal lymphatic vessels mediate neurotropic viral drainage from the central nervous system. Nat. Neurosci. 25, 577-587 (2022).
Wu, K. M. et al. The role of the immune system in Alzheimer’s disease. Ageing Res. Rev. 70, 101409 (2021).
Wiedenmann, B., Franke, W. W., Kuhn, C., Moll, R. & Gould, V. E. Synaptophysin: a marker protein for neuroendocrine cells and neoplasms. Proc. Natl. Acad. Sci. USA. 83, 3500-3504 (1986).
DeGiosio, R. A. et al. More than a marker: potential pathogenic functions of MAP2. Front. Mol. Neurosci. 15, 974890 (2022).
Liu, Q. et al. Apelin alleviated neuroinflammation and promoted endogenous neural stem cell proliferation and differentiation after spinal cord injury in rats. J. Neuroinflammation 19, 160 (2022).
Takahashi, J. et al. Oxytocin reverses A -induced impairment of hippocampal synaptic plasticity in mice. Biochem. Biophys. Res. Commun. 528, 174-178 (2020).
Leikin-Frenkel, A., Schnaider Beeri, M. & Cooper, I. How alpha linolenic acid may sustain blood-brain barrier integrity and boost brain resilience against Alzheimer’s disease. Nutrients 14, 5091 (2022).
Spinelli, J. B. & Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nat. Cell Biol. 20, 745-754 (2018).
Schlame, M. Mitochondrial cristae as insulated transformers of metabolic energy. EMBO J. 38, e103472 (2019).
Kondadi, A. K., Anand, R. & Reichert, A. S. Cristae membrane dynamics – a paradigm change. Trends Cell Biol. 30, 923-936 (2020).
Castro, C. N. et al. NCKAP1L defects lead to a novel syndrome combining immunodeficiency, lymphoproliferation, and hyperinflammation. J. Exp. Med. 217, e20192275 (2020).
Alfaidi, M., Acosta, C. H., Wang, D., Traylor, J. G. & Orr, A. W. Selective role of Nck1 in atherogenic inflammation and plaque formation. J. Clin. Invest. 130, 4331-4347 (2020).
Dormond, O., Foletti, A., Paroz, C. & Rüegg, C. NSAIDs inhibit integrin-mediated and Cdc42/Rac-dependent endothelial-cell spreading, migration and angiogenesis. Nat. Med. 7, 1041-1047 (2001).
Broman, M. T., Mehta, D. & Malik, A. B. Cdc42 regulates the restoration of endothelial adherens junctions and permeability. Trends Cardiovasc. Med. 17, 151-156 (2007).
Yao, C. et al. Mitochondrial fusion supports increased oxidative phosphorylation during cell proliferation. eLife 8, e41351 (2019).
Karu, T. I. Mitochondrial signaling in mammalian cells activated by red and near-IR radiation. Photochem. Photobiol. 84, 1091-1099 (2008).
Salehpour, F., Khademi, M., Bragin, D. E. & DiDuro, J. O. Photobiomodulation therapy and the glymphatic system: promising applications for augmenting the brain lymphatic drainage system. Int. J. Mol. Sci. 23, 2975 (2022).
Wu, S. & Xing, D. Intracellular signaling cascades following light irradiation. Laser Photonics Rev. 8, 115-130 (2014).
Semyachkina-Glushkovskaya, O. et al. Transcranial photobiomodulation of clearance of beta-amyloid from the mouse brain: effects on the meningeal lymphatic drainage and blood oxygen saturation of the brain. Adv. Exp. Med. Biol. 1269, 57-61 (2021).
Semyachkina-Glushkovskaya, O. et al. Night photostimulation of clearance of beta-amyloid from mouse brain: new strategies in preventing Alzheimer’s disease. Cells 10, 3289 (2021).
Li, D. et al. Photostimulation of brain lymphatics in male newborn and adult rodents for therapy of intraventricular hemorrhage. Nat. Commun. 14, 6104 (2023).
Richard, B. C. et al. Gene dosage dependent aggravation of the neurological phenotype in the 5xFAD mouse model of Alzheimer’s disease. J. Alzheimer Dis. 45, 1223-1236 (2015).
Minkeviciene, R. et al. Age-related decrease in stimulated glutamate release and vesicular glutamate transporters in APP/PS1 transgenic and wild-type mice. J. Neurochem. 105, 584-594 (2008).
Jackson, H. M., Soto, I., Graham, L. C., Carter, G. W. & Howell, G. R. Clustering of transcriptional profiles identifies changes to insulin signaling as an early event in a mouse model of Alzheimer’s disease. BMC Genomics 14, 831 (2013).
Picelli, S. et al. Full-length RNA-seq from single cells using Smartseq2. Nat. Protoc. 9, 171-181 (2014).
Chen, S., Zhou, Y., Chen, Y. & Gu, J. Fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884-i890 (2018).
Li, B. & Dewey, C. N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 12, 323 (2011).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
شكر وتقدير
نشكر جميع أعضاء مختبر زو على المناقشات والمساعدة الفنية خلال تنفيذ هذا المشروع. تم دعم هذه الدراسة من قبل مشاريع STI2030 الكبرى (رقم 2022ZDO212200 إلى F.Z.) وبرنامج البحث والتطوير في المنطقة الرئيسية بمقاطعة هاينان (KJRC2O23C30 إلى F.Z.).
مساهمات المؤلفين
ف. زو، ق. لو، ق. ليو، و س. وو تصوروا وصمموا التجارب. م. وانغ، ج. يان، وإكس.ليأجرى التجارب وحلل البيانات. م. وانغ،إكس.ليوشارك T.Yang في تجارب الحيوانات. قام T.Yang وM.Wang بإجراء تصوير الفلورسنت NIR-II. كتب F.Zhou وM.Wang وS.Wu المخطوطة. قرأ جميع المؤلفين ووافقوا على المخطوطة.
يجب توجيه المراسلات والطلبات للحصول على المواد إلى تشينغمينغ ليو أو فيفان تشو.
معلومات مراجعة الأقران تشكر مجلة Nature Communications أكسل مونتاني، والمراجع (المراجعين) الآخرين المجهولين على مساهمتهم في مراجعة هذا العمل. يتوفر ملف مراجعة الأقران.
معلومات إعادة الطباعة والتصاريح متاحة على http://www.nature.com/reprints ملاحظة الناشر: تظل شركة سبرينجر ناتشر محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
المختبر الوطني الرئيسي للهندسة الطبية الرقمية، كلية الهندسة الطبية الحيوية، جامعة هاينان، هايكو 570100، الصين. المختبر الرئيسي لتقييم وتدخل الصحة العقلية الذكي، وزارة التعليم، كلية التكنولوجيا الطبية، معهد بكين للتكنولوجيا، 100081 بكين، الصين. – البريد الإلكتروني: qluo@hainanu.edu.cn; zhouff@hainanu.edu.cn
Meningeal lymphatic vessels (mLVs) have been shown to be involved in amyloid beta clearance, which is considered as a potential therapeutic target for Alzheimer’s disease (AD). In this study, based on the superficial spatial distribution of mLVs, a near-infrared light is employed to modulate lymphatic drainage, significantly improving cognition of both aged and AD (5xFAD and APP/PS1) mice, and alleviating AD-associated pathology by reducing A deposition, neuroinflammation and neuronal damage. Furthermore, transmission electron microscopy imaging and RNA sequencing data indicate amelioration of mitochondrial metabolism and cellular junction of meningeal lymphatic endothelial cells (mLECs) by light modulation. These studies collectively suggest that near-infrared light treatment can improve cognitive function by strengthening scavenging ability of mLV s through restoring mLEC function. In conclusion, lymphatic drainage potentiation by light promotes pathological remission and cognitive enhancement in aging and AD mouse models, which offers a potential amelioration strategy for neurodegenerative diseases.
For a long time, the brain has been considered immune privileged owing to the lack of lymphatic drainage system . In 2015, meningeal lymphatic vessels (mLVs) located in the dura were discovered, which constitute an extensive lymphatic drainage network to remove macromolecular waste and inflammatory mediators, direct immune cell transport, and coordinate immune responses in central nervous system (CNS) . Recent studies have revealed that mLVs system is associated with the progression of ageing, Alzheimer’s disease (AD), Parkinson’s disease (PD), traumatic brain injury (TBI), subarachnoid hemorrhage (SAH), CNS viral infection and other nervous system diseases, and the changes of mLVs transport capability can significantly affect the disease development . These studies indicate that
modulating mLVs drainage can be an effective strategy for neurologic diseases.
AD is an age-associated neurodegenerative disease with high mortality. The main pathological features of AD are -amyloid (A ) abnormal aggregation and neuronal tangles in brain that contribute to neuronal dysfunction and cognitive decline . MLVs have been confirmed to be functionally degenerated with ageing or AD progression, which might be an underlying factor for exacerbated cognitive dysfunction and neural impairment. Viral-mediated vascular endothelial growth factor C (VEGF-C) treatment via intracisterna magna (i.c.m.) injection could efficiently boost function of mLVs , and then enhance lymphatic drainage to clear toxic molecules in CNS, as well as improve
learning and memory ability . Therefore, mLVs might be a therapeutical target for age-associated cognitive deficits. However, for chronic and progressive neurodegenerative disease, like AD, invasive therapy models are not feasible. Altogether, the development of noninvasive treatment modality is necessary for AD treatment.
The mLVs network is superficially distributed in the dura mater, which provides a promising strategy for transcranial neuromodulation therapies to alleviate CNS diseases through mLVs drainage modulation. In this work, based on the optical window for tissue penetration, we demonstrate that near-infrared light could modulate the function of meningeal lymphatic endothelial cells (mLECs), which in turn could contribute to mLVs drainage for the remission of pathology and the enhancement of cognitive function of aged and AD mice. Our findings reveal that the enhancement on mitochondrial metabolic homeostasis of mLECs by light is determined to promote cell adhesion and growth, which possibly potentiates cell functions and mLECs junctions to further boost meningeal lymphatic transport in the CNS for pathology amelioration.
Results
Light modulates mLVs drainage and attenuates cognitive decline in aged mice
To determine the hypothesis that light could modulate meningeal lymphatics, a non-invasive transcranial light treatment with a laser wavelength of 808 nm was performed on aged C57BL/6J mice (15-17 months, male and female) for 4 weeks ( 3 times per week, 10 min per time). Mice were anesthetized, had their dorsal scalp shaved and cleaned, and then underwent transcranial illumination by 808 nm light (light-treated mice) or indoor lighting (control-treated mice) for 10 min . After the treatment, the drainage function and morphology changes of mLVs, and the mobility and cognitive function of mice were evaluated (Fig. 1a). In order to obtain an efficacious dose for light treatment, different power densities ( 10,20 , and ) were chosen for comparison, based on the conventional dose of nearinfrared (NIR) light used for health benefits ( ), and the penetration rate of 808 nm light on the mouse skull with hairless scalp (about , independent of output power changes) (Supplementary Fig. 1a). Additionally, the surface temperature of scalp was recorded during light irradiation. There was insignificant fluctuation of the temperature, which excluded the effect of temperature changes on meningeal lymphatic function (Supplementary Fig. 1b, c).
Firstly, drainage capacity of meningeal lymphatic system was assessed by fluorescence intensity detection of CSF tracer OVA-A647 (i.c.m.) in deep cervical lymph nodes (dCLNs). The distribution area of OVA-A647 drained into dCLNs was distinctly higher in light-treated mice than that in untreated mice (Aged versus Aged: ; Aged : versus Aged: ; Aged + versus Aged: ) (Fig. 1b-e). However, there was no significant distinction of cerebral blood flow of aged mice after light treatment (Supplementary Fig. 1d, e). Structure of lymphatic vessel was evaluated by immunohistochemistry with lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1) staining (a classical marker of lymphatic endothelial cells (LECs)). The loss of area fraction of meningeal lymphatic vessels in aged mice was restored to normal by light treatment (Aged : versus Aged: ; Aged + versus Aged: ; Aged versus Aged: , ) (Fig. 1f, g). The results demonstrated that light could improve drainage and distribution of mLVs in aged mice, and light at showed more stable and efficient effects.
To evaluate the mobility and cognitive functions of treated mice, open field (OF), novel object localization (NOL), novel object recognition (NOR) and Y-maze tests were performed on light-treated mice . The results from OF test demonstrated that there was no significant difference in total distance, average velocity, or time spent in center between groups, indicating that light did not influence mobility or increase anxiety-like behavior of aged mice (Supplementary Fig. 1f-i). Interestingly, the aged mice underwent light treatment spent noticeably more time with the object at the novel location in NOL test or the novel object in NOR test than the familiar one (Aged + light: versus Aged: in NOR test), similar to the performance of young mice (1.5 months, male and female) in the same test. On the contrary, untreated aged mice preferred the familiar object in NOL and NOR test (Fig. 1h, i). Concurrently, aged mice with light treatment displayed a significantly higher discrimination index (DI, DI in the NOL and NOR tests than untreated mice (Supplementary Fig. 1j-o). In Y-maze test, aged mice explored less in novel arm than young mice, showing deficits in memory. However, following light treatment, aged mice significantly increased the time spent in the novel arm. Interestingly, the time spent in the novel arm of light treated aged mice was similar to the performance of young mice (Fig. 1j). The above results revealed that light on mLVs could improve recognition and spatial memory in aged mice. Therefore, mLVs targeted phototherapy could effectively restore the distribution and drainage function of mLVs , as well as improve cognition in mice.
Light modulation alleviates cognitive decline and pathological damage in AD mice
To explore the effect of mLVs modulation by light on the learning and memory ability of AD mice, we similarly conducted light treatment and performed OF, NOL, and NOR tests on 5xFAD mice (6 months, male) and APPswe/PS1 (APP/PS1) mice ( 11 months, male) (Fig. 2a). In the OF test, there was no noticeable difference in total distance traveled, average speed, or time in center between groups, suggesting that mice had no motor impairments after light treatment (Supplementary Fig. 2a-d and Supplementary Fig. 3a-d). As expected, light-treated AD mice exhibited cognition restoration in the behavioral tests (5xFAD + light: versus 5xFAD: in NOR test; APP/PS1 + light: versus APP/PS1: in NOR test). However, there was no significant increase in cognitive performance in wild-type (WT) mice treated with light (WT + light: versus WT: in NOR test) (Fig. 2b, e, Supplementary Fig. 2e-k and Supplementary Fig. 3e-g). Altogether, light also induced distinct learning and spatial memory modifications in 5xFAD and APP/ PS1 mice, while not in WT mice. To further investigate the benefit of light on long-term learning and memory of AD mice, the Morris water maze (MWM) test was performed. Compared with untreated AD mice, light treatment decreased latency to find the platform in acquisition trials (5xFAD + light versus 5xFAD, ; APP/PS1 + light versus APP/PS1, ) and increased visits to platform location and visits to the target quadrant in probe test, without any effect on swimming velocity. However, light did not induce any behavioral indices modifications in age-matched WT mice (WT + light versus WT, ) (Fig. 2c, d, f, g, Supplementary Fig. 2l-o and Supplementary Fig. ). These results demonstrated that light could ameliorate impairments in spatial learning and memory of AD mice, but not WT mice. accumulation, neuroinflammation and neurological damage are considered as AD pathological features. deposition and inflammation in hippocampus (HPC) and prefrontal cortex (PFC) of AD mice were evaluated by immunostaining with and Iba1 (Ionized calcium-binding adapter protein 1 , microglial marker). The results demonstrated that aggravated brain A burden occurred in HPC and PFC of 5xFAD mice, which were noticeably alleviated in light-treated 5xFAD mice to the similar level in WT littermates. Additionally, prominent microglia aggregation to and activation with reduced
Fig. 1 | Effects of light on cognition and mLVs drainage in aged mice. a Schedule of treatments and behavior tests of aged mice. Representative images of deep cervical lymph nodes (dCLNs) accumulated with OVA-A647 at 2 h after injection (i.c.m.) (from 2 replicates). Scale bar . c Quantification of fluorescence distribution of OVA-A647 in dCLNs. mice in each group. d Representative section images of OVA-A647-accumulated dCLNs stained with LYVE-1 and DAPI (from 2 replicates). Scale bar . e Quantification of fluorescence distribution of OVA-A647 in dCLN sections. mice in each group. Representative images of meninges stained with LYVE-1 and DAPI (from 2 replicates).
Scale bar or Quantification of area fraction of LYVE-1 lymphatic vessels. mice in Aged group, mice in other groups. Location recognition index of novel object location (NOL) test. i Object recognition index of novel object recognition (NOR) test. Percentage of time spent in novel arm of Y-maze test. mice in Young group, mice in Aged and Aged + light groups in data . Data in are presented as mean SEM, and analyzed by one-way ANOVA with Sidak’s multiple comparison test for comparisons of multiple groups. Source data are provided as a Source data file.
branch length, number and endpoints were detected in 5xFAD mice. Light diminished aberrant microglia activation in HPC displayed by increased branch length, branch number and endpoint number, with a similar tendency in the PFC. However, Iba1 cell number did not change significantly but presented a decreasing trend after light treatment (Fig. 2h-m and Supplementary Fig. 4a-f). Overall, light attenuated
pathological symptoms of brain deposition and neuroinflammation in AD mice.
To further examine neuroprotection of mLVs modulation by light in AD mice, immunohistochemical analyses were performed with NeuN (neuronal marker), synaptophysin (Syn) and microtubule associated protein 2 (MAP2) staining. Synaptophysin is an integral
membrane protein localized to presynaptic vesicles participating in neurotransmitter transport, and MAP2 is the predominant cytoskeletal regulator within neuronal dendrites . Compared to WT littermates, neuron cells and Syn in HPC and PFC of 5xFAD mice were significantly reduced and restored by light treatment to the similar levels as in WT littermates, indicating a significant restoration of neuronal
cells. Concurrently, MAP2 dendrites expression showed a slight increase in the light treatment group. Interestingly, dendrite morphology exhibited disordered distribution in HPC and PFC of 5xFAD mice, and light promoted more regular rearrangement (Fig. 2n-q and Supplementary Fig. ). Similarly, light treatment also reduced accumulation and microglia activation, as well as neuron damage and
Fig. 2 | Ameliorating effects of light on cognition and pathology in AD mice. a Schedule of treatments and behavior tests of AD mice. b-d Object recognition index of NOR test (b), and representative occupancy heatmaps (c) and latency to platform (d) of MWM test of FAD mice. mice in WT and light groups, mice in WT + light and 5xFAD groups. e-g Object recognition index of NOR test (e), representative occupancy heatmaps (f) and latency to platform (g) of MWM test of APP/PS1 mice. mice in WT and APP/PS1 groups, mice in APP/PS1 + light group in data mice in each group in data . h Representative images of brain sections in hippocampus (HPC) stained with , Iba1 and DAPI. Scale bar or . i Quantification of area fraction of in HPC. Quantification of branch length per cell ( ), branch number per cell ( ), endpoint number ( ) and cell number ( ) of cells in HPC. mice in WT group, mice in other groups. Representative images of brain sections in HPC stained with NeuN, synaptophysin, MAP2 and DAPI. Scale bar or Quantification of neuron number ( ), and area fraction of Syn (p) and MAP2 (q) in HPC. r GO terms upon light treatment in 5xFAD mice. mice in each group. s Enrichment plots of GSEA. mice in each group. Data in are presented as mean SEM, and analyzed by one-way ANOVA (e) or two-way ANOVA (b, d, g, i-m, o-q) with Sidak’s multiple comparison test for comparisons of multiple groups. values in and were calculated with clusterProfiler using the hypergeometric test and empirical phenotype-based permutation test, respectively, without any adjustments for multiple comparisons. The statistical tests involved two-sided analysis. Source data are provided as a Source data file.
synapse loss in HPC and PFC of APP/PS1 mice (Supplementary Fig. 5a-t). Accordingly, light could elicit a neuroprotective effect via ameliorating accumulation, neuroinflammation, and neuron and synapse loss, possibly shifting CNS environment toward a mitigatory pathological stage of AD.
To further evaluate the effect of light on AD-associated pathology changes, hippocampal RNA-seq analysis of AD mice was performed. Total RNA was extracted from HPC tissue and sequenced. Volcano plot and heatmap showed expression levels of differential genes in HPC between 5xFAD mice treated with and without light (Supplementary Fig. 6a, b). Gene ontology (GO) functional enrichment analysis showed changes in gene sets involved in ion transport, cell migration, neurogenesis and development (Fig. 2r and Supplementary Fig. 6c). Kyoto Encyclopedia of Genes and Genomes (KEGG) terms enrichment also revealed transcriptional alterations related to PI3K-AKT signaling pathway, apelin signaling pathway, TGF-beta signaling pathway, oxytocin signaling pathway, and alpha-linolenic acid metabolism, which participate in neuronal survival and differentiation as well as AD progression (Supplementary Fig. 6d, e) . In addition, Hallmark gene sets by gene set enrichment analysis (GSEA) further demonstrated distinct enrichment of down-regulated gene sets associated with AD development such as apoptosis, coagulation, hypoxia, and p53 pathway, as well as inflammation response including complement, IL2 Stat5 signaling, TNF-alpha signaling via NF-кB (Fig. 2s). These results indicated that light treatment might contribute to regulation of neuronal survival, neuroinflammation and other related pathology for AD amelioration.
Modulation on the structure of by light enhances drainage
Similarly, we validated lymphatic drainage capacity of light-treated AD mice by in vivo CLNs NIR-II FLI and dCLNs immunostaining. The outflow of OVA-ICG or OVA-A647 into CLNs (5xFAD + light: 8.225 versus 5xFAD: ) or around the mLVs in meninges was higher in light-treated mice than that in untreated 5xFAD mice (Fig. 3a-c and Supplementary Fig. 7a-d). As expected, there was unnoticeable change of cerebral blood flow of AD mice after light treatment (Supplementary Fig. 7e, f). To investigate whether improvement of lymphatic fluid transport was associated with the structural changes of mLVs, lymphatic expansion in meninges was assessed by immunofluorescence staining with LYVE-1. The results indicated that coverage of LYVE-1 vessels did not diminish in 5xFAD mice, though impaired lymphatic drainage has already been observed. However, following light treatment, increased diameter of LYVE-1 vessels in meninges occurred in AD mice ( light: versus 5xFAD: ). Therefore, light induced structural expansion but not distribution changes of mLVs in 5xFAD mice (Fig. 3d-f). Subsequently, in APP/PS1 mice, drainage impairment and structural degeneration of mLVs were observed by diminished OVA-ICG fluorescent signals in CLNs and LYVE vessels area fraction in meninges, revealing degeneration of meningeal lymphatic function and distribution. Following light
operation, enhancement of lymphatic drainage in AD mice was exhibited by strengthened OVA-ICG fluorescence in CLNs, as well as lymphangiogenesis by increased LYVE-1 area in meninges (Supplementary Fig. 8a-d). However, there was no effects on blood vessel distribution in meninges (Supplementary Fig. 8e). As a result, light could improve drainage capacity and functional expansion of mLVs in both 5xFAD and APP/PS1 mice.
Boosting mLVs drainage is required for cognition improvement by light
To determine whether the beneficial impact of light on cognitive function and AD-associated pathology relied on the increase of mLVs drainage, we performed mLVs ablation on aged mice (15-17 months, male and female) and 5xFAD mice ( 6 months, male) following the procedure described in previous studies . Then, we conducted light treatment ( ) on mLVs -ablated aged mice and FAD mice for 4 weeks, and subsequently evaluated the motor ability, anxiety, learning and memory of mice by behavioral tests (Fig. 4a). Light had no effect on total distance, average velocity, or center time in the OF test (Supplementary Fig. 9a-d and Supplementary Fig. 10a-d), indicating no alteration in mobility or anxiety-like behavior. Similar to the results above, light treatment improved learning and memory function of aged mice and AD mice, while there were no effects in mLVs-ablated mice (Object recognition index: Vis./photo. +light of aged mice: versus Vis./photo. of aged mice: , in NOR test; Vis./photo. +light of 5xFAD mice: versus Vis./photo. of mice: , in NOR test) (Fig. 4b-f, Supplementary Fig. 9e-k and Supplementary Fig. 10e-n). These results showed the beneficial impact of light treatment in cognitive improvement of aged and AD mice, which was abrogated after mLVs ablation, revealing that mLVs played an important role in cognition enhancement by light treatment.
To further estimate whether abrogation of light treatment benefits in mLVs-ablated aged and AD mice is due to inability of mLVs functional modulation, drainage kinetics and mLVs morphology were evaluated. In vivo kinetic biodistribution of OVA-ICG in superficial cervical lymph nodes (sCLNs) (up to 1 h after injection), and fluorescence intensity of OVA-ICG-accumulated dCLNs and brains (at 2 h after injection) were detected and recorded by NIR-II FLI. The fluorescence intensity of sCLNs rapidly rose to peak within several minutes in lighttreated aged and AD mice, then gradually declined to flatten out and maintained at a higher intensity than untreated mice. However, fluorescent signals in sCLNs of both mLVs-ablated mice with and without light treatment remained steady at lower levels within 60 min (Supplementary Fig. 9l, m and Fig. 4g, h). Finally, more OVA-ICG drained into dCLNs and brains of 5xFAD mice underwent light than untreated mice at 2 h after injection, while no difference between mLVs -ablated mice with light operation versus untreated mice (fluorescence intensity of dCLNs: light: versus : ; Vis./photo. +light: versus Vis./photo.: ). The results showed higher fluorescence intensity of tracer in cervical lymph nodes
Fig. 3 | Improvement effects of light on drainage function and structural changes of mLVs in 5xFAD mice. a Quantification of fluorescence intensity of OVA-ICG in sCLNs after injection (i.c.m.). mice in each group. Representative section images of OVA-A647-accumulated dCLNs stained with LYVE-1 and DAPI (from 2 replicates). Scale bar Quantification of fluorescence distribution of OVA-A647 in dCLN sections. mice in each group. Representative
images of meninges stained with LYVE-1 and DAPI (from >3 replicates). Scale bar or . e-f Quantification of area fraction (e) and diameter (f) of LYVE- lymphatic vessels in meninges. mice in each group. Data in a, c, e, f are presented as mean SEM, and analyzed by two-way ANOVA with Sidak’s multiple comparison test for comparisons of multiple groups. Source data are provided as a Source data file.
and brains of light-treated mice, suggesting that both meningeal and glymphatic drainages were enhanced. When mLVs were ablated, in addition to the expected blockage of meningeal drainage, the beneficial impacts of light on glymphatic drainage function were also abrogated (Supplementary Fig. 10o-r). These results indicated that meningeal lymphatic and glymphatic systems constructs the flow of CSF, and light-facilitated meningeal lymphatic drainage may somehow affect the entire CSF circulatory system for macromolecule clearance. Additionally, severe loss of LYVE- vessel area was observed in aged mice that underwent mLVs ablation, and there were no mLVs changes under light treatment (Fig. 4i, j). Similarly, the beneficial outcomes by light in regard to amelioration of microglial activation and amyloid load in HPC were also abrogated in mLVs-ablated AD mice (Fig. 4k).
Mitochondrial homeostasis and cellular functions of meningeal lymphatic endothelial cells (mLECs) are regulated by light
To further investigate whether the beneficial effect of light on mLVs drainage relied on the functional modulation of mLECs, fine structures within mLECs of male mice were visualized by transmission electron microscopy (TEM). As shown in Fig. 5a, compared to agematched WT mice, in AD mice, mLECs within mLVs were arranged more loosely, with round-shape mitochondria, as well as fractured and fuzzy mitochondrial cristae. Mitochondria are the sites of cellular energy metabolism, closely related to cellular functions. Cristae membrane and matrix of mitochondria are major functional regions of metabolism, and thus abnormal morphology of mitochondria or reduction and blurring of cristae structure directly reflect metabolism damage . Surprisingly,
in light-treated AD mice, mLECs were noticeably more orderly and compact, almost similar to WT littermates, indicating that light might rescue mLEC junction disruption. Simultaneously, light-treated AD mice showed mostly oval or rod-shaped mitochondria with much regular and clear cristae in mLECs, as well as increased mitochondrial length, revealing restored mitochondrial function by light (Fig. 5a, b). To further evaluate mitochondrial homeostasis in mLECs of light-treated mice, we measured the mitochondrial superoxide generation as well as mitochondrial membrane potential using mitochondrial superoxide indicator (MitoSOX) and tetramethylrhodamine (TMRE) probe, respectively. These results demonstrated that there was an increase in mitochondrial superoxide production and a decrease trend in mitochondrial membrane potential in mLECs of AD mice and aged mice, indicating mitochondrial dysfunction of mLECs. Light treatment rescued mitochondrial abnormalities in mLECs with less production of mitochondrial superoxide and an increased trend of mitochondrial membrane potential (Supplementary Fig. 11a-f). All in all, light promotes mitochondrial metabolism recovery of mLECs for cellular function preservation, maintaining the complete structure and compact arrangement of mLV .
Furthermore, the cellular function of mLVs modulated by light was verified using RNA-seq analysis. Total RNA was extracted from meninge tissues and sequenced. Volcano plot and heatmap demonstrated significant difference in gene expression in meninges of mice treated with or without light (Supplementary Fig. 11g, h). GO enrichment of signature genes further exhibited up-regulation of cell proliferation, adhesion, differentiation, development and homeostasis, as well as transport in meninges of light-treated AD mice
Fig. 4 | Effects of light on cognition and lymphatic drainage in mLVs-
ablated mice. a Schedule of treatments and behavior tests of mLVs-ablated aged mice and 5xFAD mice. b, c Location recognition index of NOL test (b) and object recognition index of NOR test (c) of mLVs-ablated aged mice. mice in each group. d-f Location recognition index of NOL test (d), object recognition index of NOR test (e) and latency to find platform in the acquisition of MWM test (f) of mLVsablated 5xFAD mice. mice in each group in data mice in Ctrl group, mice in Light group, mice in Vis./photo. and Vis./photo. + light groups in data . Representative images of sCLNs of mLVs -ablated FAD mice with OVAICG accumulation (marked with red circles) at different time points after injection (i.c.m.) (from 2 replicates). Scale bar Quantification of fluorescence
intensity of OVA-ICG in sCLNs. mice in Ctrl, Vis./photo. and Vis./photo. + light groups, mice in Light group. Representative images of meninges of mLVs ablated aged mice stained with LYVE-1 and DAPI (from 2 replicates). Scale bar = or . Quantification of area fraction of LYVE-1 lymphatic vessels. mice in Ctrl and Vis./photo. groups, mice in Light group, mice in Vis./photo. + light group. k Representative images of brain sections of mLVsablated FAD mice in HPC stained with Iba1, and DAPI (from 2 replicates). mice in each group. Scale bar . Data in are presented as mean SEM, and analyzed by two-way ANOVA with Sidak’s multiple comparison test for comparisons of multiple groups. Source data are provided as a Source data file.
Fig. 5 | Improvement effects of light on microstructure and gene expression of mLVs in 5xFAD mice. a Representative transmission electron microscopy (TEM) images of mLEC arrangement and mitochondrial morphology (red arrowheads: mitochondria of mLECs) (from 2 replicates). Scale bar or 200 nm . b Quantification of mitochondrial length of mLECs. mice in each group. c Expression of Nckap1l, Nck1, Calm2, Cdc42 and Vegfc in meninges between lighttreated AD group and AD group. d Enrichment plot of Hallmark gene sets by GSEA of meninges, showing the profile of the running enrichment score and positions of gene set members on the rank-ordered list. e Scatter plot of genes enriched in oxidative phosphorylation and ranked according to (FC: fold change).
f Expression of Lyve1, Prox1, Pecam1 and Tgfa in mLECs between light-treated AD group and AD group. g Heatmap showing relative expression levels of DEGs in mLECs involved in mitochondrial metabolism. mice in each group in data c-g. Data in are presented as mean SEM, and analyzed by one-way ANOVA with Sidak’s multiple comparison test for comparisons of multiple groups (b), or twotailed unpaired Student’s t-test for two-group comparisons (c, f). values in was calculated with clusterProfiler using the empirical phenotype-based permutation test, without any adjustments for multiple comparisons. The statistical tests involved two-sided analysis. Source data are provided as a Source data file.
Fig. 6 | Schematic illustration of the mechanism by which mLVs-targeting phototherapy to improve AD pathology. Near-infrared light enhances mitochondrial respiratory in mLECs via photoreceptor CcO , which repairs lymphatic cell
junctions and arrangement for restoration of mLVs drainage, promoting clearance of waste macromolecules , ultimately alleviating neurodegenerative processes in mice.
(Supplementary Fig. 11i). In addition, there was no distinct change of Vegfc expression (associated with lymphangiogenesis) in meninge after light treatment, while the genes involved in signaling of VEGF including Nckap1l , Nck1 , Calm2 and Cdc42 significantly upregulated (Fig. 5c). Furthermore, as shown in Supplementary Table 1, the key proteins of bicellular tight junction assembly and cell adhesion were up-regulated, suggesting that tight junction between cells was involved in the positive regulation of meningeal lymphatic vessels by light. The result demonstrated possible enhancement in cell proliferation and adhesion of mLVs after light treatment, which were the fundament for compact arrangement of mLVs. KEGG terms enriched by altered genes between groups, highlighted transcriptional changes in signaling pathway of cell development, cell communication and Alzheimer’s disease. Light treatment led to up-regulation of genes involved in cell cycle, mismatch repair and vascular smooth muscle contraction, and down-regulation of genes related to Alzheimer’s disease and pathway of neurodegeneration in meninges, which might indicate enhanced cell activity and drainage function of the meningeal drainage system, as well as AD amelioration (Supplementary Fig. 11j-1).
To further explore the mechanism of mLVs modulation, GSEA analysis was performed using the Hallmark database of gene expression. The results showed significant enrichment of 6 up-regulated gene sets associated with oxidative phosphorylation, cell cycle control, DNA repair, and immune regulation. The expression of the subset of genes contributed to oxidative phosphorylation, like Cox7c, Cox17, Atp5pb, Mfn2, Immt and Iscu as key up-regulated genes were associated with cytochrome c oxidase ( CcO ) activity and assembly, respiratory electron transport, adenosine triphosphate (ATP) synthesis, mitochondrial fusion, cristae formation and metabolic homeostasis (Fig. 5d, e). Oxidative phosphorylation is an energy transfer response in mitochondria, which couples with electron transport and produces ATP to provide energy for cellular synthesis and metabolism. In the process, CcO plays a key role as the terminal component of respiratory chain located in mitochondrial inner membrane, which also acts as a photoreceptor that undergoes molecular conformational change and heightens enzyme activity after light stimulation, promoting electron transfer and increasing ATP production, thus enhancing cell activity and function . Therefore, light probably induced lymphatic expansion and led to the clustered and structured mLEC arrangement by
CcO modulation for mitochondrial homeostasis and cellular function restoration, contributing to drainage dynamics recovery.
Subsequently, in order to further verify the mechanism of mLVs modulation, mLEC were isolated from meninges for RNA-seq analysis (Supplementary Fig. 12a-e). Light modulation led to up-regulated expression of Lyve1 and Prox1 (encoding classical markers of LECs), as well as Tgfa (transforming growth factor), but did not alter expression level of Pecam1 (encoding blood vessel marker) (Fig. 5f), showing lymphangiogenesis of light-treated mice. In addition, upregulated genes were also associated with oxidative phosphorylation and mitochondrial metabolism, which was consistent with meningeal RNA-seq analysis of light-treated mice (Fig. 5 g ). KEGG terms presented the pathways enriched by down-regulated genes in mLECs involved in base excision repair, mismatch repair and biosynthetic metabolism in 5xFAD mice compared with WT mice, which were upregulated in 5xFAD mice with light modulation (Supplementary Fig. 12f, g). The results above revealed that restoration, biosynthesis and metabolism might be essential functions of mLEC for lymphatic development and transport, which were impaired in 5xFAD mice (6 months), and possibly rescued by restoration of mitochondria functions of mLECs via light modulation, further restoring the mLVs network and drainage capability for CNS homeostasis.
Discussion
In this study, we showed that non-invasive transcranial light treatment could significantly improve cognition of aged and AD mice by targeted enhancement of meningeal lymphatic drainage. We also demonstrated that light promoted mitochondrial metabolic homeostasis to improve mLEC functionality, rescuing loose junction of mLECs, which led to increase of lymphatic drainage for clearance of deposition and neuroinflammation, resulting in pathology alleviation and cognition improvement in AD mice (Fig. 6).
Meningeal lymphatic function impairment has been confirmed in various neurologic diseases, including neurodegenerative diseases, ageing, traumatic brain injury and neurotropic viral infection . Here, we found that cognitive function and the capacity of lymphatic drainage impaired in mice ( 6 months), though mLVs coverage merely presented an insignificant downward trend, consistent with the finding of previous studies (Fig. 3d-f). The result suggested that
damage of mLVs drainage function occurred before distributional degradation in AD mice.
Superficial-distributing mLVs in the dura mater provide a direction for transcranial neuromodulation strategy to attenuate neurologic diseases by drainage regulation. In animal cells, cytochrome c oxidase (CcO) (complex IV) was considered as the primary photoreceptor in a transparent optical window from red light ( 630 nm ) to near-infrared light ( 900 nm ). CcO can be boosted by photons to promote efficiency of respiratory chain and ATP synthesis for cell activity improvement and function repairation . Particularly, has been found to have the deepest penetration with limited absorption either by water or blood in tissues, which is more suitable for transcranial modulation for longterm pathological improvement.
Previous studies confirmed that transcranial light treatment at 1267 nm could enhance the clearance of injected peptide from the brain, shown by accumulated in the brain and . Recently, transcranial light treatment at 1267 nm was performed in male newborn and adult rodents for treatment of intraventricular hemorrhage by enhancing the removal of red blood cells through brain lymphatics . Nevertheless, how 1267 nm light regulates lymphatic transport remains unclear. It is critical to determine the regulatory mechanism of meningeal lymphatic drainage by light and how drainage changes participate in cognitive improvement. By further observing the fine structure of mLECs with TEM imaging, we found changes in mitochondrial morphology with mostly oval or rod shape and regular cristae distribution in mLECs of light-treated AD mice, as well as increased mitochondrial length, implying mitochondrial function recovery for cellular function enhancement (Fig. 5a, b). Moreover, meningeal RNA-seq analysis showed positive regulation of oxidative phosphorylation and metabolic homeostasis for enhancement of cell growth, probably further promoting and expression for cell migration and restoration of endothelial adherens junctions in mice with light (Supplementary Fig. 11c, Fig. 5c, d and Supplementary Table 1). Additionally, the up-regulated genes contributed to oxidative phosphorylation associated with CcO activity, respiratory electron transport, adenosine triphosphate (ATP) synthesis and mitochondrial metabolic homeostasis (Fig. 5e). Furthermore, mLEC RNAseq analysis demonstrated the up-regulated expression of oxidative phosphorylation-, metabolism- and lymphangiogenesis-associated genes and confirmed restoration of mitochondrial metabolism and cellular development of mLECs in light-treated mice (Fig. 5f, g and Supplementary Fig. 12f, g). These results indicated that light stimulation on CcO is able to enhance oxidative phosphorylation of mitochondria, promoting energy availability and signal transduction for cellular function maintenance of mLECs, further mediating repair of endothelial junctions. The modulation of mLEC function by light enhances lymphatic transport capacity for clearance of waste macromolecules, further alleviating AD-associated pathological injury by reducing deposition, neuroinflammation and neuronal damage, finally contributing to cognitive improvement.
For the beneficial outcomes by NIR light treatment, it is possible that other regulatory mechanisms may coexist simultaneously. Light may increase the activity of the transporting immune cells in dura, thereby regulating the meningeal immune system for cerebral homeostasis. In addition, direct regulation of neuronal activity on the cerebral cortex may be another possible mechanism by which light regulates brain function for cognition improvement. Interestingly, light was ineffective in regulation of cognition in WT mice, probably owing to the ceiling effect of existing draining ability (Fig. 3a-c and Supplementary Fig. 7a-d). Light may play a role in the prevention or amelioration of cognitive deficits depending on the time of treatment. In the future, we plan to investigate whether maintaining lymphatic drainage function by periodic light treatment at the early disease stages in AD animal models could alleviate or delay disease progression.
In our study, we confirmed that mLVs -targeting phototherapy enhanced cognition in aged and AD mice. Furthermore, we revealed that light could restore mitochondrial metabolism and cellular function of mLECs, which contribute to cell arrangement amelioration of mLVs to improve drainage dynamics for pathology relief in AD mice. Overall, we provide a potential treatment for neurologic diseases by mLVs transport modulation for CNS waste clearance, which could alleviate cognitive impairment and related pathology.
Methods
Animals
All the mice were in C57BL/6J background purchased from Jinzhihe Biotechnology Co., Ltd (Yangzhou, China). Male and female WT mice aged between 15 to 17 months served as the elderly model, and 1.5-month-old ones served as the counterpart young group. Two male AD models were used in this study: 6-month-old 5xFAD and 11-month-old APPswe/PS1 9 (APP/PS1) mice, with their transgene non-carrier littermates as counterpart groups. Amyloid deposition accompanied by gliosis is seen in 5xFAD mice at 2 months of age. Cognitive deficit occurs at about 5 months old and neuron loss in the brain regions with amyloidosis begins at 6 months of age . In APP/PS1 mice, amyloid deposits in brain between 4 to 6 months and cognitive deficit occurs between 6 to 10 months . Mice of all strains were bred under light and darkness alternating cycles with controlled temperature and humidity ( ), and were allowed free access to standard diet and water. The mice treatments were performed in compliance with the Guide for the Care and Use of Laboratory Animals. All animal studies were reviewed and approved by the Institutional Animal Care and Use Committee of Hainan University (approval number: HNUAUCC-2021-00025).
Intra-cisterna magna (i.c.m.) injections
Mice were anesthetized with isoflurane supplemented with dexmedetomidine. The dorsal surface of neck was shaved and cleaned. After making a 1.5 cm incision of the neck skin at midline, the muscle layers were retracted with forceps for the cisterna magna exposure. The desired solution was injected into the cisterna magna compartment . For evaluation of lymphatic drainage, of Alexa Fluor 647 conjugate ovalbumin (at , OVA-A647, Thermo Fisher Scientific, USA) or OVA-ICG (at ) was administered using a Hamilton syringe. After the injection, the needle was held in place for an extra 2 min to avoid CSF backflow. Finally, the neck skin and muscle were cleaned and sutured, and the mice were placed on a warming pad for recovery from anesthesia until conscious, then transported back to home cage.
Meningeal lymphatic vessels (mLVs) ablation
Visudyne treatment was performed as published protocols . Mice were anesthetized with isoflurane supplemented with dexmedetomidine. Then visudyne (at , APExBIO, USA) was injected (i.c.m.) at a rate of . After 15 min , visudyne was photoconverted with a non-thermal 689 nm light (LSR689CP-3.6W-FC, LASEVER, China) on five different spots on skull (one at the injection site, one at the confluence of the sinuses, two at the bilateral transverse sinuses and one at the superior sagittal sinus), with irradiation ( , 83 s each spot). The scalp skin was sutured and the mice were placed on a warming pad for recovery from anesthesia until conscious, then transported back to home cage.
Light penetration measurement
Mice were euthanized by cervical dislocation and the scalps were shaved and cleaned with razor and depilatory cream. Then the head was isolated, the lower jaw and brain tissue was removed, and the skull with hairless scalp was harvested. The skull was fixed under the 808 nm light source. The penetrations of the skull with hairless scalp at lambda
point under the light at different powers were recorded (powers detected by a Digital Handheld Optical Power and Energy Meter (PM100D, Thorlabs Inc, Newton, NJ, USA)). The transmittances were calculated as radiation flux/incident power.
Light treatment
Mice were brought into experimental room at least 30 min before light treatment for habituation. Mice were anesthetized with isoflurane, and dorsal scalp was shaved and cleaned. Then a continuous 808 nm light (LSR808H-5W-FC, LASEVER, China) was applied to illuminate the skull with intact hairless scalp over an area of , at for 10 min , which ended up with doses of . The surface temperature of scalp was measured along the 10 min of exposure to the light irradiation using a near-infrared (NIR) thermal camera (226S, FOTRIC, China) and analyzed using AnalyzlR (version 4.3.2.71). Mice underwent light treatment three times a week lasting for continuous four weeks. For the mice in control group, the same experimental procedure was performed. Mice were anesthetized with isoflurane, and dorsal scalp was shaved and cleaned, then underwent transcranial illumination by indoor lighting for 10 min . After treatments, the mice were placed on a warming pad for recovery from anesthesia until conscious, then transported back to home cage.
Cervical lymph nodes (CLNs) imaging
Mice were anesthetized with 1% isoflurane supplemented with dexmedetomidine, and neck skin was resected to expose CLNs. The dorsal surface of neck was cleaned and incised. The muscle layers were retracted to both sides to expose the cisterna magna. Then of OVA-ICG (at ) was administered (i.c.m.) to mice at using a Hamilton syringe. After the injection, the needle was held for additional 2 min to avoid CSF backflow. Finally, the neck skin and muscle were cleaned and sutured, and the mice were allowed to recover on a warming pad with isoflurane maintenance. Fifteen minutes post-injection start, the mice were placed supine under the MARS in vivo imaging system (Artemis Intelligent Imaging, China) for imaging. Images of CLNs were collected 15 min to 1 h after injection start, with intervals of 15 min . The timedependent fluorescence intensity of accumulated tracers in CLNs was recorded. Two hours after injection start, the mice were transcardially perfused with PBS and paraformaldehyde (PFA). Cervical muscles were cut and retracted, and sCLNs and dCLNs were collected to be imaged by the imaging system. Quantitative analysis of fluorescence intensity was performed using Fiji software (version 1.53 q ) and LightField (version 6.14) to characterize the lymphatic drainage function.
Laser speckle contrast imaging (LSCI)
After one-month treatment, mice were anesthetized with 1% isoflurane supplemented with dexmedetomidine, followed by the scalp and the fascia resected to expose the skull. Then the mice were head-fixed using a head-fixation frame and placed prostrate under the LSCI microscope (Simopto, China) for detection of cerebral blood flow. The signals excited by 785 nm laser were amplified and digitized for blood flow imaging. The relative blood flow (%) of mice was record for continuous 3 min and the blood flow index (perfusion units, PU) was calculated by SIM BFI software (version 3.1.45, Simopto).
Open field (OF) test
OF test was performed in an opaque cubic arena ( 40 cm edge). Before the test, mice were habituated to the test room for 30 min . Mice were then placed into the arena and allowed to explore for 10 min . Total distance (cm), velocity (cm/s) and time spent in the center (%) were recorded and analyzed using DigBehv behavior analysis system (version 4.1, Jiliang Software Science&Technology Co., Ltd.).
Novel object recognition/novel object location (NOR/NOL) test
NOR or NOL task was conducted in the same cubic arena used in the OF test. 24 h before the test, the mice were placed in the arena to explore for 10 min as habituation. Then two identical objects were placed in different corners, around 6 cm away from the walls. During training, mice were allowed to explore the objects in the arena for 10 min and then return to the home cages. 1 or 24 h later, memory of the location or object recognition was tested. For NOL test, mice were allowed to explore the objects for 10 min in the same arena with one object moved to a novel location. For NOR test, mice were allowed to explore for 10 min in the same arena with one object replaced with a novel one in the original position. Object exploration was recorded using Visutrack software (version 3.0, Xinruan Information Technology Co., Ltd.) and recognition index (RI), discrimination index (DI) and object preference (percentage of time with object) were calculated. , object preference or .
Y-maze test
Y-maze test was conducted in a Y-shaped maze with three identical arms, each 35 cm length, 6 cm width and 15 cm height. Different geometrical figures were placed inside each arm as spatial landmarks. The arms were randomly designated as the novel, start and other arm. During training, a divider was used to block the novel arm. The mice were placed into the start arm of the maze, heading away from the maze center, for a exploration. In test trial, the mice were placed into the Y -maze 1 h later to explore for 5 min with all arms open to free access. The time spent in each arm was recorded using DigBehv behavior analysis system (version 4.1, Jiliang Software Science&Technology Co., Ltd.) and percentage of time spent in the novel arm (%) was calculated.
Morris Water Maze (MWM) test
MWM test was conducted in a circular pool ( 120 cm diameter) filled with opaque water at around to a 30 cm depth. Different geometrical figures were placed surrounding the pool as spatial reference landmarks. During five continuous training days, mice were placed into the tank to find a hidden fixed platform ( 10 cm diameter, located at 1 cm below the water surface) within 60 s . The mice failing to find the platform during 60 s were gently guided to climb on the platform and rest for 15 s . The latency to platform was recorded using Visutrack software (version 3.0, Xinruan Information Technology Co., Ltd.) and the mean latency to platform of the four trials each day was calculated. On the sixth day (probe trial), the platform was removed and mice performed a 60-s exploration. The percentage of time spent in each quadrant (%), the number of crossings of the platform area and swimming velocity ( ) were recorded and analyzed.
Tissue collection
Mice were anesthetized and then transcardially perfused with PBS and 4% PFA. Cervical skin and muscles were cut and retracted, and CLNs were collected and fixed in PFA for 24 h at . The skin and muscle of the heads were removed from the bones, and then brains and skulls were collected and fixed in PFA for 24 h at . Then the wholemount meninges were carefully stripped from skulls and then stored in PBS at until further use. The fixed brains and CLNs were washed with PBS, dehydrated with , and sucrose for three days and frozen in Tissue-Tek OCT compound (SAKURA, USA) at . The brains were sliced ( sections) and kept in cryoprotectant at for further use.
Immunohistochemistry and imaging
For antigen retrieval, the frozen brain or CLN sections were heated with citrate buffer for 30 min . For immunofluorescence staining, brain sections, CLN sections and meninges were washed with PBS and
blocked with PBS containing 0.3% Triton X-100 (Sigma, USA) and 5% bovine serum albumin (BSA, Sigma) for 1 h at room temperature (RT), followed by incubation with of primary antibodies dilutions overnight at : rat anti-LYVE-1 (cat# 14-0443-82, lot# 2494966, clone# ALY7, 1:200, Invitrogen, USA), mouse anti-amyloid- (cat# 805501, lot# B326654, clone# 12F4, 1:1000, Biolegend, USA), rabbit anti-Iba1 (cat# 019-19741, lot# LEN4341, 1:500, Abcam, UK), rat anti-NeuN (cat# ab279297, lot# GR3400636-6, clone# EPR12763, 1:1000, Abcam, UK), rabbit anti-Syn (cat# MA5-14532, lot# WI3369463, clone# SP11, 1:100, Invitrogen), mouse anti-MAP2 (cat# MA5-12826, lot# XF3603732A, clone# AP18, 1:200, Invitrogen), rabbit anti-CD31 (cat# SAB4502167100UG, lot# 210547, 1:200, Sigma). Tissues were then washed three times with PBS, followed by incubation with Alexa Fluor 488/555/633 goat anti-rat/rabbit/mouse IgG (H+L) secondary antibodies in PBS containing 0.3% Triton X-100 for 1 h in dark at RT: Alexa Fluor 488 goat anti-rat IgG (H + L) secondary antibody (cat# A-11006, lot# 2247986, 1:500, Invitrogen), Alexa Fluor 488 goat anti-rabbit IgG ( ) secondary antibody (cat# A-32731, lot# VG302077, 1:500, Invitrogen), Alexa Fluor 555 goat anti-rat IgG (H + L) secondary antibody (cat# A21434, lot# 2184321, 1:500, Invitrogen), Alexa Fluor 555 goat anti-rabbit IgG (H + L) secondary antibody (cat# A-32732, lot# YA361054, 1:500, Invitrogen), and Alexa Fluor 633 goat anti-mouse IgG (H + L) secondary antibody (cat# A-21052, lot# 2418505, 1:500, Invitrogen). Then tissues were washed three times and incubated with DAPI for 5 min at RT. Finally, tissues were mounted with SlowFade Diamond Antifade Mountant (Invitrogen) and imaged by confocal laser scanning microscopy (FV3000, Olympus, Japan) or Slide Scanner (VS200, Olympus). Image export and quantitative analysis were performed using OLYMPUS OlyVIA (version 4.1) and Fiji software (version 1.53q).
Mitochondrial superoxide and mitochondrial membrane potential measurement
MitoSOX Red (Invitrogen) and tetramethylrhodamine (TMRE, Invitrogen) probe were used to assess the mitochondrial superoxide and mitochondrial membrane potential levels in meningeal lymphatic endothelial cells (mLECs). The fresh whole-mount meninges were collected and incubated in MitoSOX reagent for 10 min or in TMRE reagent for 30 min at , followed by Alexa Fluor 488 rat anti-LYVE-1 (cat# 53-0443-80, lot# 2547846, clone# ALY7, 1:250, Invitrogen) dilution incubation for 30 min at . MLECs were imaged by a confocal microscope (FV3000, Olympus).
Transmission electron microscopy (TEM)
Mice were euthanized by cervical dislocation, then the skullcaps were dissected and the whole-mount meninges were stripped. MLVs regions of meninges were trimmed to tissues with a blade and rinsed twice with PBS. Then tissues were fixed with glutaraldehyde buffer with paraformaldehyde for 30 min at RT and stored at until processed as TEM samples. Then the tissues were rinsed with PBS, secondly fixed with osmium tetroxide (OsO4), rinsed with doubledistilled water, dehydrated with ethanol, infiltrated with acetone, and embedded in epoxy resin. The samples were cut as 70 nm sections, stained with uranyl acetate and lead citrate, and then imaged using a transmission electron microscope (Tecnai G2 Spirit, FEI, USA). Quantitative analysis of the mitochondrial length of meningeal lymphatic endothelial cells (mLECs) was performed using Fiji. The images of LECs in the meninges were acquired using a transmission electron microscopy and the mean of 10 individual mitochondrial length measurements was calculated for each sample in different groups ( ).
Sorting of mLECs
To obtain a suspension of mLECs of 5xFAD mice using fluorescenceactivated cell sorting (FACS), mice were anesthetized and then transcardially perfused with PBS. Skullcaps were dissected and wholemount meninges were stripped in Dulbecco’s Modified Eagle Medium
(DMEM, Gibco, USA), supplied with FBS (Gibco), penicillin ( ml , Gibco) and streptomycin ( , Sigma). Then meninges were incubated in DMEM containing of collagenase VIII (Sigma) and of DNase I (Sigma) for 30 min at . The digested meninges were filtrated through nylon cell meshed for single-cell suspensions. Cells were then centrifuged ( 280 g ) at for 10 min and resuspended in PBS. Then cells were incubated with antibodies dilutions at for 30 min : rat anti-CD45-PE (cat# 12-0451-82, lot# 2356222, clone# 30-F11, 1:150, eBioscience), rat anti-CD31-FITC (cat# 11-0311-85, lot# 2373735, clone# 390, 1:100, eBioscience), hamster anti-podoplanin-PE-Cy7 (cat# 127412, lot# B355692, clone# 8.1.1, 1:100, Biolegend) and rat anti-CD11b-PerCP (cat# 101230, lot# B352939, clone# M1/70, 1:80, BioLegend), and finally incubated with DAPI for 5 min . Cells were then resuspended in FACS buffer. The singlets were gated using the pulse width of the side scatter and forward scatter. Cells negative for DAPI were identified and selected as viable cells. The single cells were gated and sorted using flow cytometry (MoFlo Astrios, Beckman Coulter, USA). The sample cells were dispensed in the droplets with a volume of and dropped into the PCR tubes containing 4 ul of lysis buffer. The samples were temporarily stored at for further analysis.
RNA extraction and sequencing
All RNA sample processing (including cDNA library generation) and RNA sequencing (RNA-seq) were performed by E-GENE Technology Co., Ltd (Shenzhen, China). The RNA-seq libraries for samples were prepared using the Smart-seq2 protocol with minor modifications. For tissue samples, the total RNA of each simple was extracted from one individual hippocampus (HPC) tissue of brain ( ) or one individual meninge tissue ( ) via TRIzol total RNA extraction reagent (Thermo Fisher Scientific). Then RNA of sample was quantified by Qubit Fluorometer and RNA integrity was detected by agarose gel electrophoresis. For LEC samples, the LECs (previously sorted by FACS from one individual meninge, ) were lysed with of Triton solution containing of RNase inhibitor. The extracted total RNA or the cell lysis was mixed with of Oligo-dT primer for RNA enrichment, and was subjected to reverse transcription in 1X Super Script II first-strand buffer system with 200 U of Super Script II reverse transcriptase, of Template switch oligo (TSO, ), 20 U RNase Inhibitor, of Dithiothreitol (DTT, 100 mM ), of Betaine ( 5 M ) and of . The full-length cDNA was synthesized and amplified with 10 to15 cycles of PCR amplification. Then the product was purified and used for library construction. After quantified by Agilent 2100 Bioanalyzer (Agilent Technologies, USA), the library was sequenced with Illumina Novaseq platform.
The raw reads were filtered by fastp (version 0.18.0) to remove adapters or low-quality bases for following assembly and analysis. Fragment per kilobase of transcript per million mapped reads (FPKM) value was calculated to quantify expression abundance and variations of transcription using RSEM software. All analyses of visualizing the RNA-seq data were performed with the statistical software R (version 4.3.1). Differential expression analysis was conducted and visualized using the R package DESeq (version 1.40.2) and ggplot2 (version 3.4.2). The genes with a fold change value greater than 2.0 or lower than 0.5 and false discovery rate (FDR)-corrected were considered significantly differentially expressed genes (DEGs). Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis provided GO and KEGG terms that significantly enriched in DEGs comparing to the genome background, and filtered the DEGs that correspond to biological functions or pathways. GO and KEGG term enrichment analysis were carried out with the clusterProfiler software (version 4.8.3). GO terms similarity based on similarity matrices of functional terms was performed using R packages simplifyEnrichment (version 1.10.0). In order to gain a better understanding of the mechanism, a Gene Set Enrichment Analysis
(GSEA) was performed by ranking all genes based on their level of difference using R package clusterProfiler (version 4.8.3) and visualized using package enrichplot (version 1.20.3). The hallmark gene sets served as reference sets for GSEA were obtained from the Molecular Signatures Database (https://www.gsea-msigdb.org/gsea/ msigdb). Heatmap were visualized using ggplot2 (version 3.4.2) and pheatmap (version 1.0.12). We didn’t make any adjustments on data, and the data were sequenced on the same platform and processed in the same way.
Statistical analysis
To ensure randomization, animals from different cages were divided in the same experimental group. Experimenters were blind to the group allocation of mice in all the experiments during data collection and analysis. Statistical significance was conducted using two-tailed unpaired Student’s test for two-group comparisons, and one-way ANOVA or two-way ANOVA with Sidak’s multiple comparison test for multiple-group comparisons. A value less than 0.05 was considered statistically significant. Data are presented as mean SEM. Statistical data were analyzed using GraphPad Prism (version 8.4.2, GraphPad Software, Inc.).
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The data that support the findings of this study are available within the article, the Supplementary Information files and the Source Data files. The RNA sequencing data generated for this study can be found in the GEO repository under accession number GSE245658. The hallmark gene sets served as reference sets for GSEA in RNA-seq analysis were obtained from the Molecular Signatures Database (https://www.gseamsigdb.org/gsea/msigdb/). Source data are provided with this paper.
Code availability
Customized code or algorithms were not used to generate results in the present study.
References
Hasek, M., Chutna, J., Sládecek, M. & Lodin, Z. Immunological tolerance and tumor allografts in the brain. Nature 268, 68-69 (1977).
Louveau, A. et al. Structural and functional features of central nervous system lymphatic vessels. Nature 523, 337-341 (2015).
Aspelund, A. et al. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 212, 991-999 (2015).
Da Mesquita, S. et al. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature 560, 185-191 (2018).
Da Mesquita, S. et al. Meningeal lymphatics affect microglia responses and anti-Aß immunotherapy. Nature 593, 255-260 (2021).
Ding, X. et al. Impaired meningeal lymphatic drainage in patients with idiopathic Parkinson’s disease. Nat. Med. 27, 411-418 (2021).
Bolte, A. C. et al. Meningeal lymphatic dysfunction exacerbates traumatic brain injury pathogenesis. Nat. Commun. 11, 4524 (2020).
Chen, J. et al. Meningeal lymphatics clear erythrocytes that arise from subarachnoid hemorrhage. Nat. Commun. 11, 3159 (2020).
Li, X. et al. Meningeal lymphatic vessels mediate neurotropic viral drainage from the central nervous system. Nat. Neurosci. 25, 577-587 (2022).
Wu, K. M. et al. The role of the immune system in Alzheimer’s disease. Ageing Res. Rev. 70, 101409 (2021).
Wiedenmann, B., Franke, W. W., Kuhn, C., Moll, R. & Gould, V. E. Synaptophysin: a marker protein for neuroendocrine cells and neoplasms. Proc. Natl. Acad. Sci. USA. 83, 3500-3504 (1986).
DeGiosio, R. A. et al. More than a marker: potential pathogenic functions of MAP2. Front. Mol. Neurosci. 15, 974890 (2022).
Liu, Q. et al. Apelin alleviated neuroinflammation and promoted endogenous neural stem cell proliferation and differentiation after spinal cord injury in rats. J. Neuroinflammation 19, 160 (2022).
Takahashi, J. et al. Oxytocin reverses A -induced impairment of hippocampal synaptic plasticity in mice. Biochem. Biophys. Res. Commun. 528, 174-178 (2020).
Leikin-Frenkel, A., Schnaider Beeri, M. & Cooper, I. How alpha linolenic acid may sustain blood-brain barrier integrity and boost brain resilience against Alzheimer’s disease. Nutrients 14, 5091 (2022).
Spinelli, J. B. & Haigis, M. C. The multifaceted contributions of mitochondria to cellular metabolism. Nat. Cell Biol. 20, 745-754 (2018).
Schlame, M. Mitochondrial cristae as insulated transformers of metabolic energy. EMBO J. 38, e103472 (2019).
Kondadi, A. K., Anand, R. & Reichert, A. S. Cristae membrane dynamics – a paradigm change. Trends Cell Biol. 30, 923-936 (2020).
Castro, C. N. et al. NCKAP1L defects lead to a novel syndrome combining immunodeficiency, lymphoproliferation, and hyperinflammation. J. Exp. Med. 217, e20192275 (2020).
Alfaidi, M., Acosta, C. H., Wang, D., Traylor, J. G. & Orr, A. W. Selective role of Nck1 in atherogenic inflammation and plaque formation. J. Clin. Invest. 130, 4331-4347 (2020).
Dormond, O., Foletti, A., Paroz, C. & Rüegg, C. NSAIDs inhibit integrin-mediated and Cdc42/Rac-dependent endothelial-cell spreading, migration and angiogenesis. Nat. Med. 7, 1041-1047 (2001).
Broman, M. T., Mehta, D. & Malik, A. B. Cdc42 regulates the restoration of endothelial adherens junctions and permeability. Trends Cardiovasc. Med. 17, 151-156 (2007).
Yao, C. et al. Mitochondrial fusion supports increased oxidative phosphorylation during cell proliferation. eLife 8, e41351 (2019).
Karu, T. I. Mitochondrial signaling in mammalian cells activated by red and near-IR radiation. Photochem. Photobiol. 84, 1091-1099 (2008).
Salehpour, F., Khademi, M., Bragin, D. E. & DiDuro, J. O. Photobiomodulation therapy and the glymphatic system: promising applications for augmenting the brain lymphatic drainage system. Int. J. Mol. Sci. 23, 2975 (2022).
Wu, S. & Xing, D. Intracellular signaling cascades following light irradiation. Laser Photonics Rev. 8, 115-130 (2014).
Semyachkina-Glushkovskaya, O. et al. Transcranial photobiomodulation of clearance of beta-amyloid from the mouse brain: effects on the meningeal lymphatic drainage and blood oxygen saturation of the brain. Adv. Exp. Med. Biol. 1269, 57-61 (2021).
Semyachkina-Glushkovskaya, O. et al. Night photostimulation of clearance of beta-amyloid from mouse brain: new strategies in preventing Alzheimer’s disease. Cells 10, 3289 (2021).
Li, D. et al. Photostimulation of brain lymphatics in male newborn and adult rodents for therapy of intraventricular hemorrhage. Nat. Commun. 14, 6104 (2023).
Richard, B. C. et al. Gene dosage dependent aggravation of the neurological phenotype in the 5xFAD mouse model of Alzheimer’s disease. J. Alzheimer Dis. 45, 1223-1236 (2015).
Minkeviciene, R. et al. Age-related decrease in stimulated glutamate release and vesicular glutamate transporters in APP/PS1 transgenic and wild-type mice. J. Neurochem. 105, 584-594 (2008).
Jackson, H. M., Soto, I., Graham, L. C., Carter, G. W. & Howell, G. R. Clustering of transcriptional profiles identifies changes to insulin signaling as an early event in a mouse model of Alzheimer’s disease. BMC Genomics 14, 831 (2013).
Picelli, S. et al. Full-length RNA-seq from single cells using Smartseq2. Nat. Protoc. 9, 171-181 (2014).
Chen, S., Zhou, Y., Chen, Y. & Gu, J. Fastp: an ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884-i890 (2018).
Li, B. & Dewey, C. N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 12, 323 (2011).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Acknowledgements
We thank all members of the Zhou lab for discussion and technical assistance during the execution of this project. This study was supported by STI2030-Major Projects (No. 2022ZDO212200 to F.Z.) and Hainan Province Key Area R&D Program (KJRC2O23C30 to F.Z.).
Author contributions
F.Zhou, Q.Luo, Q.Liu, and S.Wu conceived and designed the experiments. M.Wang, C.Yan, and X.Li performed the experiments and analyzed the data. M.Wang, X.Li, and T.Yang participated in animal experiments. T.Yang and M.Wang performed NIR-II fluorescence imaging. F.Zhou, M.Wang, and S.Wu wrote the manuscript. All the authors read and approved the manuscript.
Correspondence and requests for materials should be addressed to Qingming Luo or Feifan Zhou.
Peer review information Nature Communications thanks Axel Montagne, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Reprints and permissions information is available at http://www.nature.com/reprints
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
State Key Laboratory of Digital Medical Engineering, School of Biomedical Engineering, Hainan University, Haikou 570100, China. Key Laboratory of Brain Health Intelligent Evaluation and Intervention, Ministry of Education, School of Medical Technology, Beijing Institute of Technology, 100081 Beijing, China. – e-mail: qluo@hainanu.edu.cn; zhouff@hainanu.edu.cn