دوبيزيلا نيويوركensis تعدل التسامح المناعي في التهاب القولون عبر مسار L-لايسين المنشط AhR-IDO1-Kyn Dubosiella newyorkensis modulates immune tolerance in colitis via the L-lysine-activated AhR-IDO1-Kyn pathway
تنتج البكتيريا المعوية المتعايشة مجموعة متنوعة للغاية من المستقلبات النشطة للحفاظ على توازن الأمعاء، ومع ذلك فإن دورها الأساسي في إنشاء بيئة ميكروية مناعية متسامحة في القناة المعوية لا يزال غامضًا. هنا، نوضح أن بكتيريا متعايشة في الفئران لم يتم دراستها بشكل كافٍ، وهي Dubosiella newyorkensis، ونظيرتها البشرية Clostridium innocuum، لها تأثير بروبيوتيك مناعي على التهاب القولون الناتج عن كبريتات الدكستران الصوديوم باستخدام نماذج الفئران التقليدية، المعالجة بالمضادات الحيوية، والخالية من الجراثيم. نحدد دورًا مهمًا لـ D. newyorkensis في إعادة توازن استجابات Treg/Th17 وتحسين إصابة الحاجز المخاطي من خلال إنتاج الأحماض الدهنية قصيرة السلسلة، وخاصة البروبيونات والليسين (Lys). نوضح أيضًا أن Lys يعزز قدرة الخلايا التغصنية (DCs) على التحمل المناعي من خلال تعزيز استقلاب التربتوفان نحو مسار الكينورينين (Kyn) من خلال تنشيط إنزيم الاستقلاب إندولامين-2،3-ديوكسيجيناز 1 (IDO1) بطريقة تعتمد على مستقبل الهيدروكربون العطري (AhR). تحدد هذه الدراسة تواصلًا استقلابيًا غير معترف به سابقًا حيث تمارس البكتيريا المتعايشة المنتجة لـ Lys قدرتها على تنظيم المناعة لإنشاء بيئة مثبطة مناعية متوسطة بواسطة Treg من خلال تنشيط دائرة الاستقلاب AhR-IDO1-Kyn في DCs. تمثل هذه الدائرة الاستقلابية هدفًا علاجيًا محتملاً لعلاج أمراض الأمعاء الالتهابية.
يحتوي الجهاز الهضمي البشري على مجموعة هائلة ومتنوعة وحرجة من الكائنات الدقيقة (المعروفة باسم ‘الميكروبيوتا’) التي تتفاعل بشكل وثيق مع مضيفها البشري لتشكيل نظام بيئي متبادل الاعتماد ومقيد بشكل متبادل، حيث تؤدي وظائف أيضية مثل تخمير الكربوهيدرات وتخليق الفيتامينات.الأمعاء تنتج الميكروبيوتا مجموعة متنوعة بشكل هائل من المستقلبات الصغيرة النشطة حيوياً التي يمكن أن تشارك في الإشارات، وصيانة الحاجز المخاطي، أو تعديل نظام المناعة.نظرًا لمثل هذه المجموعة الواسعة من التأثيرات على فسيولوجيا المضيف وعلم المناعة، لا يزال من غير الواضح كيف تتفاعل الميكروبات المحددة (والجزيئات الصغيرة التي تنتجها) مع
تسبب، تحافظ، تخفف، أو تتنبأ بالأمراض المتعلقة بالأمعاء مثل مرض الأمعاء الالتهابي (IBD)، التهاب القولون التقرحي (UC) ومرض كرون (CD).
تنشيط مفرط لـ Th1/Th17 وضعف في Foxp3تم وصف خلايا T التنظيمية (Treg) في مسببات التهاب الأمعاء (IBD)تثبيط المناعة بواسطة Treg من خلال إفراز السيتوكينات المثبطة مثل TGF-وIL-10 له أهمية كبيرة في تثبيط التهاب الأمعاء. كما أنها تحافظ على فسيولوجيا الأمعاء من خلال تعزيز وظائف الحاجز الظهاري وإصلاح الأنسجة. على الرغم من أنه تم التأكيد على أن ميكروبات الأمعاء المحددة والمواد الأيضية المرتبطة بها تشارك في تحفيز التسامح المناعي المعتمد على خلايا Treg.آلياتها الجزيئية الأساسية غير معروفة إلى حد كبير.
تتأثر تطوير خلايا Tregs المعوية بعدة مستقلبات مشتقة من النظام الغذائي. الأحماض الدهنية قصيرة السلسلة (SCFAs)، مثل الأسيتات، البروبيونات، والبيوتيرات هي المنتجات الرئيسية لتخمر الكربوهيدرات غير المهضومة في الأمعاء، وخاصة في القولون. تحفز الأحماض الدهنية قصيرة السلسلة تراكم خلايا Tregs القولونية (cTregs) من خلال آليات متعددة.يمكنهم تحفيز تكاثر Treg مباشرة من خلال تنشيط GPR43 (المعروف أيضًا باسم FFAR2)والتمييز بين الساذجالخلايا إلى خلايا Treg من خلال أسيتيل هيسون H3 لـ Foxp3 عن طريق تثبيط هيستون ديأسيتيلز (HDAC).
التريبتوفان (Trp) هو حمض أميني أساسي وعطري يجب على البشر الحصول عليه من النظام الغذائي. يمكن استقلاب التريبتوفان الغذائي إلى كينورينين (Kyn) بواسطة نشاط إنزيم إندولامين-2،3-ديوكسيجيناز 1 (IDO1) في خلايا الظهارة المعوية (IECs) والخلايا الشجيرية (DCs).ويساهم في تمايز الخلايا الساذجة خلايا إلى تريغسمن خلال تنشيط مستقبل الهيدروكربون العطريعامل نسخ مُفعل بواسطة الليغاند ومستشعر مناعيالذي تم الإشارة إليه في مسببات التهاب الأمعاءعلى الرغم من أنه تم إثبات أن الميكروبيوم المعوي مهم لتمثيل التريبتوفان وتنشيط مستقبلات أريل الهيدروكربون خلال التهاب الأمعاء.لم يتم استكشاف الأنواع المتعايشة المحددة والمواد الأيضية الميكروبية التي تشارك في تنشيط الدائرة الأيضية AhR-IDO1Kyn وتأثيرها في إنشاء بيئة ميكروبية مخاطية متسامحة بوساطة Treg بشكل كامل.
في هذه الدراسة، حددنا بكتيريا متعايشة قوية تنتج الأحماض الدهنية القصيرة السلسلة، وهي Dubosiella newyorkensis، التي يمكن أن تحسن التهاب القولون الناتج عن كبريتات الدكستران (DSS) في الفئران من خلال إعادة توازن استجابات Treg/Th17 وتحسين سلامة الحاجز المخاطي. كشفنا أن D. newyorkensis ونظيرتها البشرية Clostridium innocuum (C. innocuum) تنتج L-Lys لتعديل تثبيط المناعة الذي تسببه Treg عن طريق تنشيط دوائر AhR-IDO1-Kyn في خلايا DCs للفئران والبشر. بشكل جماعي، تشير هذه النتائج إلى استراتيجية متعددة العوامل تستخدمها D. newyorkensis وC. innocuum للحفاظ على توازن المناعة في أمعاء الفئران والبشر، على التوالي.
النتائج
تلاعب بميكروبات الأمعاء عن طريق علاج بمضاد حيوي واحد يقيّد التهاب القولون الناتج عن DSS
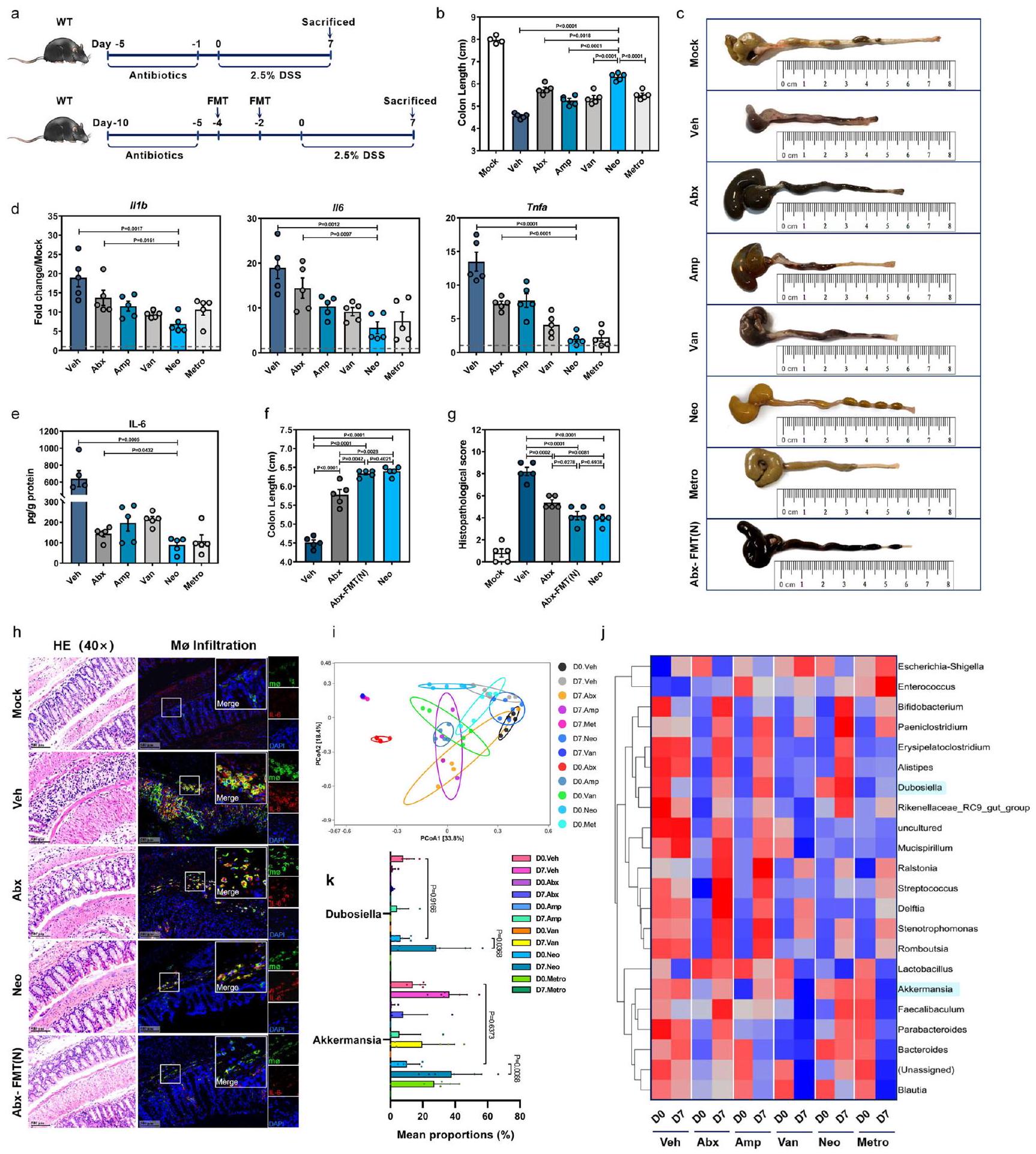
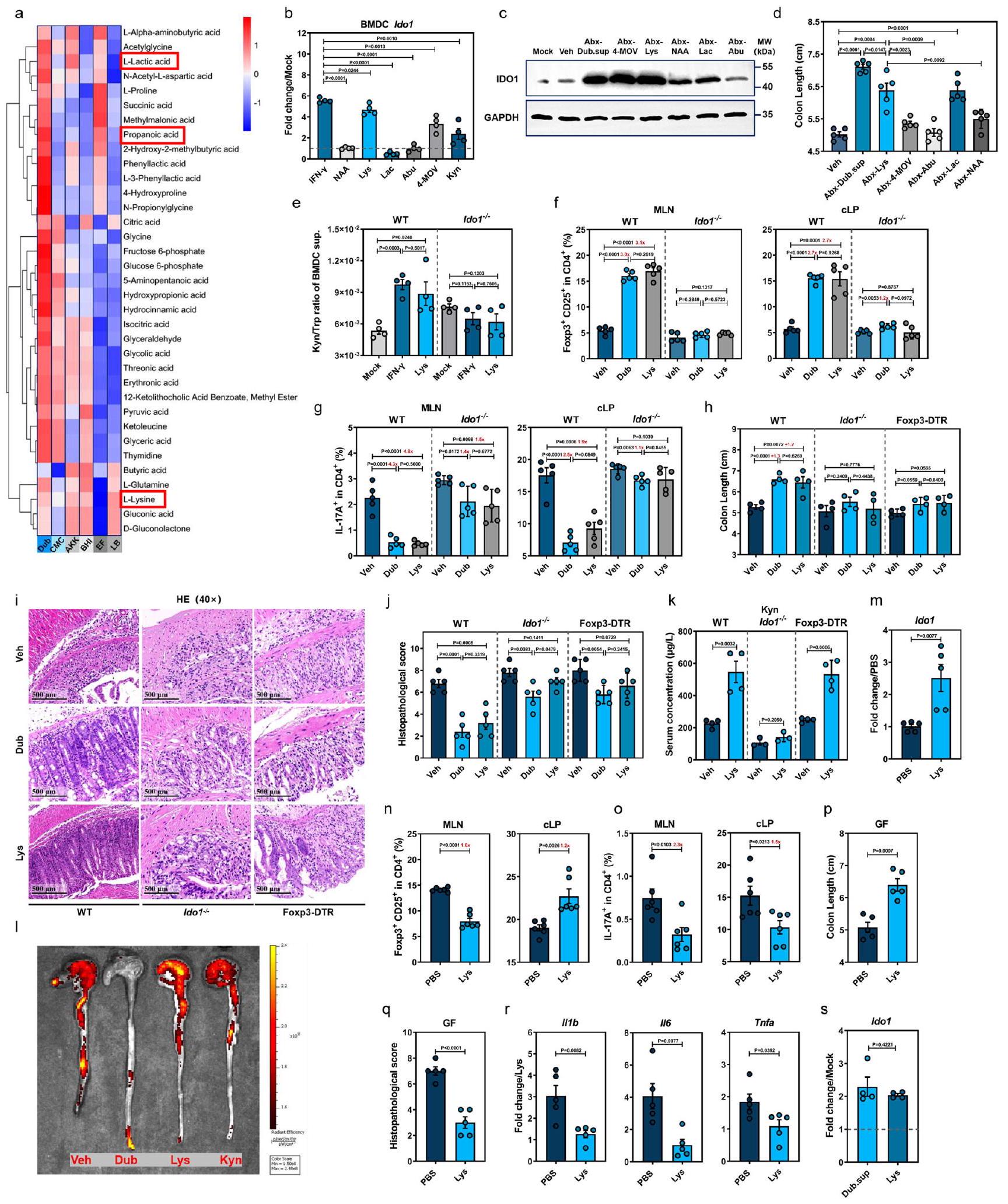
تم إعطاء فئران C57BL/6J من النوع البري (WT) الأمبيسيلين (Amp) والفانكومايسين (Van) والنيوميسين (Neo) أو المترونيدازول (Metro) إما بشكل فردي أو ككوكتيل يحتوي على الأربعة (Abx) عن طريق التغذية الفموية، ثم تعرضت لـ DSS (الشكل 1a). تم تأكيد اضطراب ميكروبيوم الأمعاء بواسطة كل من Abx وجميع علاجات المضادات الحيوية الأربعة الفردية كخفض كبير في نسخ 16 S rDNA (الشكل التوضيحي 1a). كان لعلاج Neo أفضل تأثير في تخفيف التهاب القولون الناتج عن DSS، حيث تمثل ذلك في أقل فقدان للوزن (الشكل التوضيحي 1b)، ووجود قولون أطول (الشكل 1b، c) بالإضافة إلى انخفاض مستويات mRNA في القولون للسيتوكينات المؤيدة للالتهابات (الشكل 1d) وبروتين IL-6 (الشكل 1e).
بعد ذلك، قمنا بإعداد عينات براز مجمعة من فئران نيو وأجرينا زراعة ميكروبات البراز (FMT) في فئران أبكس التي تم استنفاد ميكروباتها. (الشكل 1أ). كما هو متوقع، استعاد نقل البراز (FMT) بشكل كبير العدد الإجمالي لنسخ 16S rDNA في البراز قبل علاج DSS، ولكن ليس إلى نفس مستوى التحكم في المركبة (Veh) (الشكل التكميلي 1ج). ومن المثير للاهتمام، فقدان الوزن (الشكل التكميلي 1د)، القولون تم تخفيف تقصير (الشكل 1ج، و) والآفات النسجية المرضية (الشكل 1هـ، و) وتسلل البلعميات المفرزة لـ IL-6 (الشكل 1هـ) بشكل كبير فيتشير هذه البيانات إلى أن الميكروبات المعوية في فئران نيو قد تحتوي بالفعل على ميكروبات معوية محددة يمكن أن توفر الحماية ضد التهاب القولون الناتج عن DSS.
قمنا بإجراء تسلسل وتحليل جين 16S rRNA على عينات البراز المجمعة من الفئران المعالجة بالمركب الوهمي (Veh) والمضادات الحيوية (Abx) والفئران المعالجة بمضاد حيوي واحد في اليوم 0 (D0) واليوم 7 (D7) بعد إعطاء DSS ووجدنا اختلافات ملحوظة في تركيب الميكروبيوم بناءً على العلاج بالمضادات الحيوية، مع أو بدون تحفيز DSS (الشكل 1i). كان هناك زيادة ملحوظة في الوفرة النسبية لبكتيريا Bifidobacterium وBacteroides وAkkermansia وDubosiella في اليوم 7، وكانت أعلى في الفئران المعالجة بـ Neo مقارنة بالفئران المعالجة بمضاد حيوي واحد الأخرى والفئران المعالجة بالمضادات الحيوية (الشكل 1j، k).
تخفف الاستعمار بواسطة D. newyorkensis التهاب القولون الناتج عن DSS من خلال استعادة وظيفة الحاجز المخاطي وتثبيط الاستجابات الالتهابية
من بين الأنواع البكتيرية التي تم تعزيزها بواسطة علاج نيو، تم الإبلاغ عن أن بيفيدوباكتيريوم، باكتيرويديس، وأكرمانسيا تلعب أدوارًا واقية خلال التهاب الأمعاء.، وبالتالي ركزنا على جنس دوبوسيلا (الموجود ضمن عائلة إريسبيلوترشاسي) للدراسة الحالية. اخترنا D. newyorkensis (المختصرة هنا باسم دوب) المعزولة من فأر مختبريكعينة ممثلة لتحديد دور جنس دوبوسيلا في التهاب القولون الناتج عن DSS. أكدت اختبارات حساسية المضادات الحيوية لدوب مقاومتها للنيوميسين والميترونيدازول ولكنها كانت حساسة للأمبيسيلين والفانكومايسين (الشكل التوضيحي 2أ).
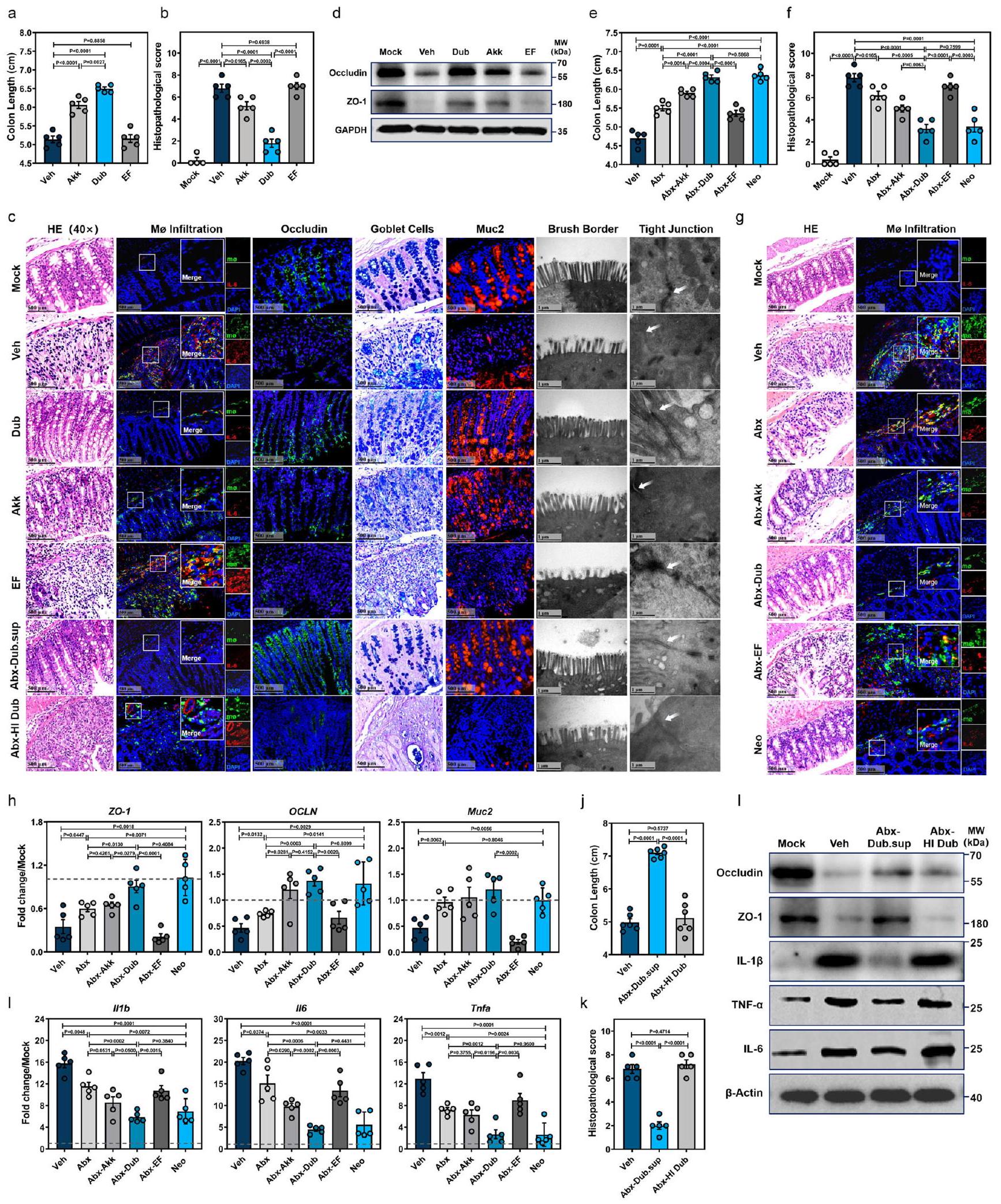
تم إعطاء مجموعات من فئران WT دواء Dub مرتين ثم تم إعطاؤها DSS لتحفيز التهاب القولون (الشكل التوضيحي 2b). نظرًا لأن Akkermansia muciniphila (المختصرة بـ Akk فيما بعد) كانت أيضًا غنية بعد علاج Neo (الشكل 1j، k) وقد تم توثيق أنها تحمي خلال التهاب الأمعاء الالتهابي.تم اختياره كتحكم بروبيوتيك في الدراسة. كانت Enterococcus faecalis (المختصرة بـ EF فيما بعد)، والتي تعد سببًا رئيسيًا للعدوى المكتسبة في المستشفيات، بمثابة تحكم بكتيري غير مرتبط من نوع جرام إيجابي. في الواقع، كان لاستعمار Dub تأثير وقائي أكبر ضد التهاب القولون الناتج عن DSS مقارنة بـ Akk، مع فقدان وزن جسم أقل (الشكل التوضيحي 2c)، وطول قولون أطول (الشكل 2a والشكل التوضيحي 2d)، ودرجة نسيجية أقل (الأشكال 2b، 2c) وتسلل أقل لمكروفاجات القولون (الشكل 2c). بالمقابل، أدى استعمار EF إلى نمط ظاهري التهابي مشابه لمجموعة التحكم Veh (الأشكال 2a-d والشكل التوضيحي 2c، d).
علاوة على ذلك، أدى الاستعمار باستخدام Dub إلى حماية أفضل لحاجز الغشاء المخاطي القولوني، كما يتضح من انخفاض apoptosis (الشكل التوضيحي 2e)، وزيادة مساحة الخلايا الإيجابية للأوكلاودين (الشكل التوضيحي 2f)، وزيادة عدد خلايا الجوبلت المفرزة للميوسين في الظهارة القولونية وأقل ت disruption للهيكل الدقيق لحدود الفرشاة والوصلات الضيقة (الشكل 2c). كما أكدنا الفروق في تعبير الأوكلاودين وZO-1 وMuc2 في خلايا الظهارة المعوية القولونية (cIECs) (الشكل 2c، d والشكل التوضيحي 2g)، بينما أظهرنا أيضًا أن تعبير السيتوكينات المؤيدة للالتهابات IL-1IL-6 وعامل نخر الورم- (TNF- كان ( ) أقل بكثير في WT-Dub مقارنة بـ WT-EF أو WT-Veh، سواء في الغشاء المخاطي القولوني (cLP) (الشكل التوضيحي 2h) أو في الدم المحيطي (الشكل التوضيحي 2i).
بالإضافة إلى ذلك، أظهرت الفئران WT التي تم إعطاؤها Dub عن طريق الفم لمدة 5 أيام متتالية بدءًا من اليوم الثالث بعد إعطاء DSS (الشكل التوضيحي 2j) طول قولون أطول (الشكل التوضيحي 2k، l) وتحسن في علم الأمراض النسيجية (الشكل التوضيحي 2m، n) مقارنة بالفئران التي تلقت EF أو الشواهد غير المعالجة، مما يشير إلى أن Dub له أيضًا تأثيرات علاجية على التهاب القولون الناتج عن DSS.
لتقييم تأثير البروبيوتيك لدوب على التهاب القولون بشكل أكبر، استخدمنا نموذج الفئران لالتهاب القولون الناتج عن حمض 2,4,6-ثلاثي نيتروبنزين سلفونيك (TNBS) (الشكل التكميلي 3a). في هذا النموذج، خفف استعمار دوب من إصابة الأمعاء كما يتضح من طول القولون (الشكل التكميلي 3b، c) وتقليل الآفات النسجية المرضية.
الشكل 1 | تأثير تعديل ميكروبيوتا الأمعاء على قابلية الفئران للإصابة بالتهاب القولون الناتج عن DSS. أ مخطط أنظمة العلاج. ب-هـ فئران C57BL/6J من النوع البري (WT، ) تم استلام المركبة (Veh) أو المضادات الحيوية عن طريق الفم (جرعة واحدة من أمبيسيلين، فانيكوميسين، نيومايسين، أو ميترو نيدازول أو بالاشتراك [Abx]، ) يوميًا لمدة 5 أيام وتمت معالجته بـ تم استخدام DSS في مياه الشرب لمدة 7 أيام. في اليوم السابع (D7) بعد علاج DSS، تم تحديد طول القولون (ب، ج)، التعبير النسبي لـ mRNA للسيتوكينات المؤيدة للالتهابات IIIb و II6 و Tnfa بواسطة qRTPCR (د) ومستوى بروتين IL-6 بواسطة ELISA (هـ) في كل مجموعة.تم إعطاء فئران Abx زراعة ميكروبيوتا البراز (FMT) من فئران معالجة بـ Neo مرتين [Abx-FMT(N)] أو Veh بفاصل 48 ساعة ثم تم علاجها بـ DSS لمدة 7 أيام.تم استخدام فئران WT B6 المعالجة بـ Neo (Neo) كتحكم. تم قياس طول القولون (f) وتم تسجيل التغيرات النسيجية المرضية بواسطة صبغة HE. ) و تم تحديد تسرب البلعميات (Mø) التي تفرز IL-6 بواسطة IFA (ساعة).تحليل المكونات الرئيسية (PCoA) (i) وخريطة الحرارة للوفرة النسبية (j) للبكتيريا في عينات البراز من الفئران المعالجة بالمضادات الحيوية أو بمضاد حيوي واحد في اليوم 0 واليوم 7 بعد إعطاء DSS. k الوفرة النسبية لدوبوسيلا وأكمانسيا في اليوم 0 واليوم 7 بين المجموعات المعالجة بمضادات حيوية مختلفة (اليوم 7. أمب، اليوم 7. فان،; D0. في, D0. أبكس, D0. مترو, D0. أمب, D0. فان, ; D0. نيو، D7. فيه، D7. نيو، D7. أبكس، D7. مترو، ). تشير الخطوط المتقطعة عند 1 إلى أن العلاجات لها قيمة متساوية كضوابط موحدة. النتائج تمثل بيانات تم توليدها في تجربتين مستقلتين على الأقل وتُعبر عنها كمتوسط SEM، وذو جانبين-تم فحص القيم بواسطة اختبار الطالب-اختبار. يتم توفير بيانات المصدر كملف بيانات المصدر. (الشكل التكميلي 3d، e). زادت استعمار الدوب من تعبير OCLN و ZO-1 و Muc2 في خلايا الظهارة المعوية (الشكل التكميلي 3f)، وانخفض تعبير Illb و Il6 و Tnfa في الطبقة تحت المخاطية المعوية (الشكل التكميلي 3g)، بما يتماشى مع دور الدوب في حماية الحاجز المخاطي والوقاية من الالتهاب المعوي.
لإزالة تأثير الميكروبات الخلفية وربط تأثير البروبيوتيك مباشرةً باستعمار دوب، تم إعطاء الفئران المعالجة بالمضادات الحيوية دوب (Abx-Dub) أو أك (Abx-Akk) أو EF (Abx-EF) عن طريق التغذية القسرية ومن ثم تمت الإدارة باستخدام DSS (الشكل التكميلي 3h). كانت جميع العزلات البكتيرية الثلاثة قادرة على إعادة توطين ميكروبيوتا الأمعاء في الفئران المعالجة بالمضادات الحيوية (الشكل التكميلي 3i). لاحظنا فقدان وزن جسم أقل (الشكل التكميلي 3k)، وطول قولون أطول (الشكل 2e والشكل التكميلي 3l)، ودرجة نسيجية أقل (الشكل 2f، g) وتسلل أقل للخلايا البالعة (الشكل 2g) في قولون مجموعة Abx-Dub مقارنة بمجموعات Abx-Akk وAbx-EF أو مجموعة Abx. كانت هذه المعايير متساوية بين مجموعتي Abx-Dub وNeo (الشكل 2e-g والشكل التكميلي 3l). مشابه لمجموعة Neo
الشكل 2 | D. newyorkensis يحمي الفئران من التهاب القولون الناتج عن DSS ويخفف الالتهاب المخاطي والأضرار في الحاجز الناتجة. أ-د تم استعمار الفئران التقليدية من النوع البري C57BL/6J بـCFU من D. newyorkensis (Dub)،. ميوسينيفيلا (Akk) ، E. فايساليس (EF) أو المركبة (Veh) مرتين مع فاصل مدته يومين بينهما، ثم تم العلاج بـ DSS كما هو موصوف سابقًا ( ). في اليوم السابع بعد إدارة DSS، تم جمع عينات من القولون لتحديد طول القولون (أ)، درجة الهستوباثولوجيا (ب) بواسطة صبغة HE، تسلل البلعميات المفرزة لـ IL-6، تعبير Muc2 وOccludin بواسطة IFA، عدد خلايا الجوبلت بواسطة صبغة ألكيان الأزرق والميكروهيكل للظهارة القولونية بواسطة TEM (ج). تم تحديد تعبير Occludin وZO-1 في كل مجموعة بواسطة التحليل الغربي (WB) (د). e-i تم استعمار الفئران WT المعالجة بـ Abx بـ CFU دوب، أك، EF، أو Veh وتعرض لـ DSS . في اليوم السابع بعد علاج DSS، طول القولون (e)، درجة الفحص النسيجي بواسطة صبغة HE (تسلل البلعميات المفرزة لـ IL-6تم فحصها.
في هذه الأثناء، تعبير ZO-1 وOCLN وMuc2 في خلايا الظهارة المعوية القولونية المعزولة )، وتم الكشف عن تعبير السيتوكينات المؤيدة للالتهابات Illb و Il6 و Tnfa في الغشاء المخاطي القولوني (cLP) (i) بواسطة qRT-PCR.-فئران WT التقليدية (تم إعطاء Dub المعطل حرارياً (HI) أو السائل الفائق من Dub (Dub.sup) عن طريق الفم وتم تعريضهم لـ DSS. في اليوم السابع بعد إعطاء DSS، تم قياس طول القولون (j) وتم تسجيل التغيرات النسيجية المرضية بواسطة صبغة HE.مستوى بروتينات الوصل الضيق أوكلودين و ZO-1، وتعبير السيتوكينات المؤيدة للالتهابات IL-1IL-6 و TNF-تم تحديدها بواسطة WB (I). الخطوط المتقطعة عند 1 تشير إلى أن العلاجات لها قيمة متساوية كضوابط موحدة. النتائج تمثل بيانات تم توليدها في تجربتين مستقلتين على الأقل وتُعبر عنها كمتوسط.SEM، وذو جانبينتم فحص القيم بواسطة اختبار الطالب-اختبار. يتم توفير بيانات المصدر كملف بيانات المصدر. بالنسبة للمجموعات الضابطة، كان هناك زيادة ملحوظة في تعبير ZO-1 وOCLN وMuc2 في خلايا الظهارة المعوية (clECs) (الشكل 2h) وانخفاض كبير في Illb وIl6 وTnfa في الأنسجة المعوية (cLP) (الشكل 2i) مقارنة بمجموعات Abx-Akk وAbx-EF وAbx أو Veh.
لتحديد المكون الوقائي لدوب بشكل أفضل، تم إعطاء فئران Abx محلول دوب المصفى (Dub.sup) أو دوب المعطل حرارياً (HI) عن طريق الفم، ثم تم تعريضها لعلاج DSS (الشكل التكميلي 3h). من المثير للاهتمام أن المعالجة المسبقة بـ Dub.sup، وليس HI Dub، حسنت فقدان الوزن الناتج عن DSS (الشكل التكميلي 3j) والظواهر المرضية (الشكل 2j، k والشكل التكميلي 3l)، واستعادت وظائف الحاجز المخاطي (الشكل 2c، 1)، وأعاقت تعبير السيتوكينات الالتهابية في القولون (الشكل 21)، على الرغم من أنه تم الكشف عن خلايا دوب المعقمة في براز فئران Abx قبل إعطاء DSS (الشكل التكميلي 3i). مجتمعة، تظهر هذه النتائج دورًا وظيفيًا محتملاً لمستقلبات دوب المستمدة بدلاً من النشاط المباشر لمكونات البكتيريا في تخفيف التهاب القولون الناتج عن DSS.
يمنع دوب الأمراض المناعية الناتجة عن DSS من خلال إعادة التوازنفوكس بي 3تريغ و IL17CD4استجابات خلايا T في بيئة الأمعاء الدقيقة
باستخدام تحليلات RNA-Seq، اكتشفنا انخفاض تنظيم المسارات المؤيدة للالتهابات، بما في ذلك تلك المرتبطة بتسرب الكريات البيضاء، وتنشيط NF-kB، ومسارات إشارات IL-17، في القولون من فئران Dub مقارنةً بمجموعة التحكم Veh في اليوم السابع بعد علاج DSS (الشكل 3a والشكل التوضيحي 4a). وبشكل متسق، تم انخفاض تنظيم II17 وIfng المؤيدين للالتهابات وزيادة تنظيم II10 المضاد للالتهابات وتمت ملاحظة خلايا المناعة cLP بكميات كبيرة في مجموعة دوب أو أك، ولكن ليس في مجموعة EF (الشكل التوضيحي 4b).
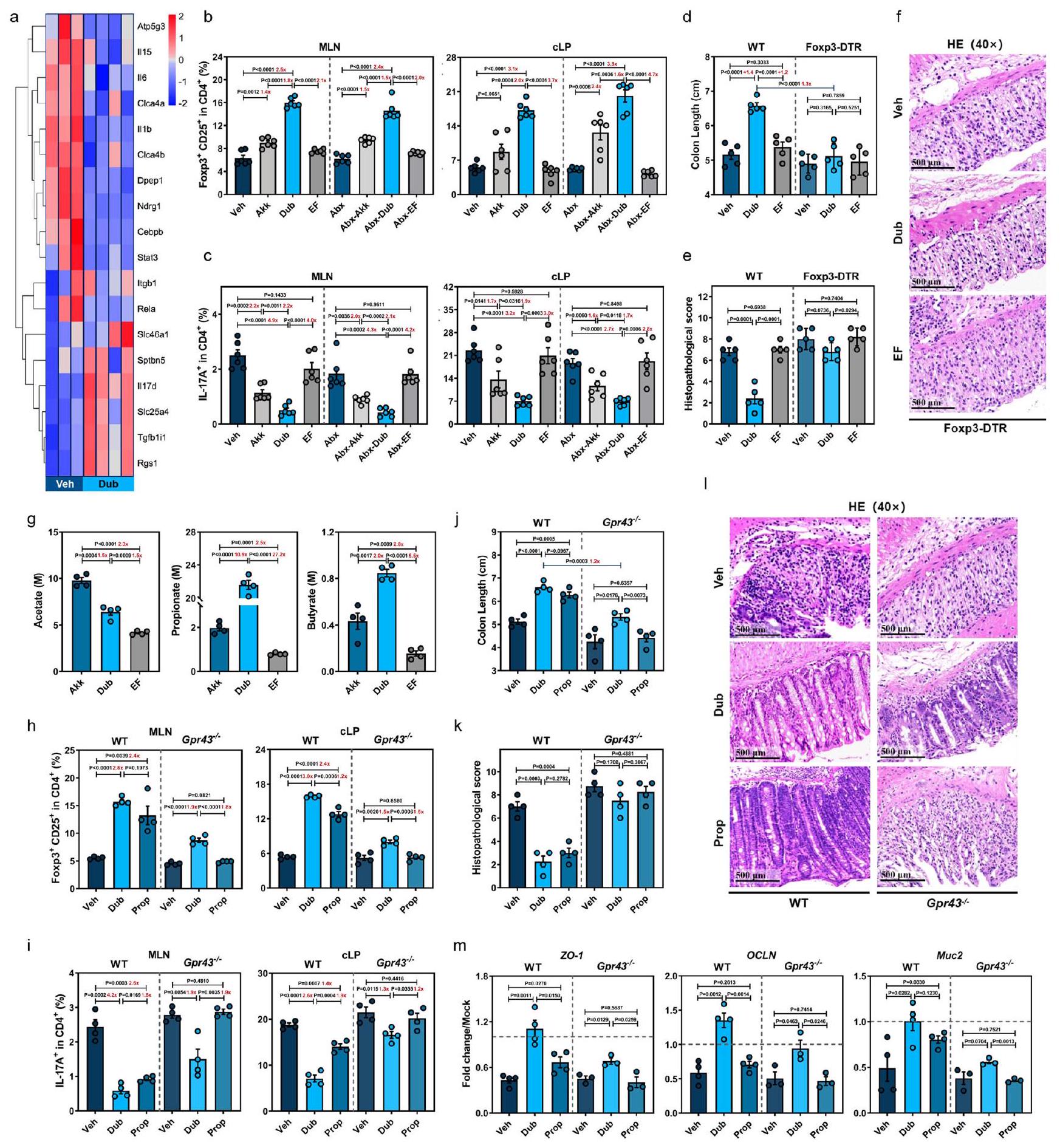
نظرًا لأن الخلايا التائية التنظيمية تلعب دورًا مركزيًا في الحفاظ على البيئة المناعية المثبطة في الأمعاء عبر TGF-وأن إفراز IL-10 وعدم التوازن بين استجابات Treg و Th1/Th17 مرتبطان بتطور مرض التهاب الأمعاء (IBD)افترضنا أن استعمار دوب يثبط الالتهاب الناتج عن DSS من خلال تحفيز Foxp3تريغز وكبح إنترفيرونالخلايا و IL17الخلايا. لاختبار ذلك، قمنا بتعقيم مجموعات من الفئران المعالجة بالمضادات الحيوية (Abx) باستخدام Dub، وأعطينا DSS، وقمنا بقياس مجموعات فرعية محددة من خلايا T في الطحال، والعقد اللمفاوية المساريقية (MLNs)، و cLP باستخدام تحليل التدفق الخلوي في اليوم السابع (الرسوم التوضيحية التكميلية 2b و3h). استراتيجيات التصفية المعنية موضحة في الشكل التوضيحي التكميلي 5. نظرًا لأن Akk يمكن أن يحفز كل من Tregs القولونية والمحيطية في التهاب القولونقمنا بتضمين Akk كعنصر تحكم إيجابي وEF كعنصر تحكم سلبي. زادت استعمار Dub أو Akk بشكل كبير من CD25.فوكس بي 3تراجس وانخفاض ملحوظ في IL17CD4خلايا T في العقد اللمفاوية المتوسطة و cLP (الشكل 3ب، ج)، ولكن تم تقليل IFN بشكل طفيف فقطالخلايا في cLP (الشكل التوضيحي 4c) مقارنةً بالتحكمات EF أو Veh، مع أو بدون ميكروبات الأمعاء في اليوم السابع. لوحظ نمط مشابه في الطحال، حيثفوكس بي 3كانت خلايا Tregs أعلى بشكل ملحوظ و IL17CD4كانت خلايا T أقل بشكل ملحوظ في مجموعات Dub أو Abx-Dub مقارنةً بالتحكم غير المستعمر في اليوم السابع (الشكل التوضيحي 4d)، مما يشير إلى أن Dub أعاد توازن استجابة Treg/Th17 ليس فقط في القولون، ولكن أيضًا في الدم المحيطي في التهاب القولون الناتج عن DSS. ومن الجدير بالذكر،
كان دوب متفوقًا على أك في إعادة توازن استجابة Treg/Th17 سواء بوجود ميكروبيوتا الأمعاء intact أو بدونها (الشكل 3ب، ج والشكل التوضيحي 4ب-د).
بالإضافة إلى ذلك، قمنا بإعطاء الفئران التقليدية أو فئران Abx دواء Dub في غياب DSS (الشكل التوضيحي 4e) ووجدنا أن CD25فوكس بي 3تم زيادة Tregs أيضًا في الطحال و cLP، بينما IL17CD4تم تقليل خلايا T في العقد اللمفاوية المتوسطة و cLP دون تحفيز DSS (الشكل التكميلي 4f، g)، مما يوحي بأن Dub يساهم في توازن استجابة Treg/Th17 حتى في ظل التوازن الداخلي للأمعاء.
أخيرًا، تم تغذية فئران Foxp3-DTR المعالجة بـ WT أو سم الدفتيريا (DT) بدوب أو EF قبل إدارة DSS (الشكل التوضيحي 4h). كما هو متوقع، فإن معالجة DT قد أضعفت بشكل كبير CD25.فوكس بي 3تراجي في الطحال، العقد اللمفاوية المتوسطة و cLP في فئران Foxp3-DTR (الشكل التوضيحي 4i). أدى استنفاد تراجي في فئران Foxp3-DTR إلى عكس الفروقات الملحوظة بين الفئران المعالجة بـ Veh (WT-Veh) والفئران المستعمرة Dub (WT-Dub) في أنماط المرض (الشكل 3d والشكل التوضيحي 4j)، التقييم النسيجي (الشكل 3e، f) وتعبير السيتوكينات المؤيدة للالتهابات (الشكل التوضيحي 4k)، مما يشير إلى أن Foxp3تعتبر خلايا Tregs ضرورية لتأثير الحماية الذي يوفره Dub ضد التهاب القولون الناتج عن DSS. ومن المثير للاهتمام أن هناك اختلافات صغيرة لا تزال موجودة في تعبير ZO-1 وOCLN وMuc2 بين الفئران المعالجة بـ Veh والفئران المستعمرة بـ Dub من نوع Foxp3-DTR (الشكل التكميلي 41)، مما يشير إلى أن Dub قد يكون قادرًا على تعزيز الشفاء المخاطي بشكل مستقل عن تحفيز تثبيط المناعة المعتمد على Treg.
تمنع الاستعمار باستخدام دوب التهاب القولون الناتج عن DSS من خلال تنظيمفوكس بتريغس جزئيًا من خلال محور الإشارة SCFA-GPR43
نظرًا لأن دوب هو قريب جيني من فاكيليباكولوم رودنتيوموهو منتج فعال للأحماض الدهنية قصيرة السلسلة وقد تم الإبلاغ عن قدرته على الحماية من نمو الأورام المعويةافترضنا أن النمط الظاهري الوقائي لدوب قد يكون بسبب إنتاج الأحماض الدهنية قصيرة السلسلة بشكل قوي. في الواقع، كانت هناك تركيزات أعلى بشكل ملحوظ من الأسيتات، البروبيونات (Prop)، والبيوتيرات في عينات البراز من فئران دوب مقارنة بمجموعات EF أو مجموعة التحكم Veh، وكانت تركيزات البروبيونات والبيوتيرات متقاربة بين فئران دوب وفئران أكك المستعمرة (الشكل التوضيحي 6a). متسقة مع هذه البيانات، كانت تركيزات البروبيونات والبيوتيرات في دوب.sup أعلى بـ 10.9 مرة و 2 مرة على التوالي مقارنة بـ Akk.sup. ومع ذلك، كان تركيز الأسيتات أعلى بـ 1.5 مرة في Akk.sup مقارنة بـ Dub.sup (الشكل 3g).
نظرًا لأن البروبيونات لديها أعلى تقارب لمستقبل GPR43الذي يُعتبر ضروريًا لتوسع ووظيفة تثبيط الخلايا التائية التنظيمية القولونية (cTregs) في الإصابة الناتجة عن DSS، كنا نتوقع أن الاختلافات فيفوكس بي 3قد يتم تكرار تحفيز Treg بين فئران WT-Dub و WT-Veh من خلال إعطاء Prop، وقد تفقد كلا النمطين في فئران Gpr43-/-. لذلك، قمنا بإعطاء فئران WT و Gpr43-/- Dub عن طريق الفم مرتين، أو أضفنا Prop إلى مياه الشرب لمدة 3 أسابيع، ثم تم إعطاؤهم DSS وتم تقييم ترددات CD25.فوكس بي 3تريغز و IL17CD4خلايا T في الطحال، العقد اللمفاوية المتوسطة و cLP (الشكل التوضيحي 6b). في الواقع، أدى إعطاء Prop إلى
الشكل 3 | D. newyorkensis يمنع التهاب القولون الناتج عن DSS من خلال تنظيم فوكس بي 3تعمل خلايا Tregs جزئيًا من خلال محور الإشارة SCFA-GPR43. خريطة حرارية تظهر تعبير mRNA الذي تم تحديده بواسطة تسلسل RNA الكمي في عينات القولون من المعالجة بالسيارة (Veh) ( ) أو D. newyorkensis (Dub) – مستعمرات من فئران C57BL/6J البرية من النوع البري ( ) في D7 بعد علاج DSS. ب، ج CD25+Foxp3+Tregs (ب) و ILالخلايا (ج) في العقد اللمفاوية المساريقية (MLN) والطبقة الغشائية القولونية (cLP) للفئران البرية التقليدية أو المعالجة بالمضادات الحيوية التي استعمرت بـ Dub أو A. muciniphila (Akk) أو E. faecalis (EF)تم فحصها بواسطة تحليل تدفق الخلايا في اليوم السابع بعد علاج DSS. د-ف الفئران التقليدية WT أو Foxp3-DTR ( ) تم استعمارها بـ تمت معالجة CFU من Dub أو Akk أو EF بعلاج DSS. في اليوم السابع بعد إدارة DSS، تم قياس طول القولون (د) وإجراء التقييم النسيجي (هـ، و). ج تم تحديد تركيز SCFA في السوبرناتانت الثقافي لـ Dub أو Akk أو EF بواسطة GC/MS.. فئران WT التقليدية و Gpr43-/- ) كانوا
مستعمرة مع دوب ( CFU) مرتين أو تم إعطاء البروبيونات (Prop) في مياه الشرب لمدة 3 أسابيع قبل التعرض لـ DSS. في اليوم السابع بعد إدارة DSS، CD25فوكس بي 3تريغس ) و IL-17 خلايا T (i) في العقد اللمفاوية المتوسطة و cLP للفئران العادية WT أو Gpr43-/تم قياس ( ) بواسطة قياس التدفق الخلوي. في الوقت نفسه، تم قياس طول القولون (ج)، والدرجة النسيجية المرضية بواسطة صبغة HE (ك، ل) ( )، وتعبير ZO1 وOCLN وMuc2 في خلايا الظهارة المعوية القولونية المعزولة (cIECs) (WT، ; Gpr43-/, ) ( كان محددًا. تغيير في ترددات CD25فوكس بي 3تريغس أو IL-17CD4خلايا Tطول القولون ( تم حساب تركيزات الأحماض الدهنية قصيرة السلسلة (SCFA) (غ) بين المجموعات وعرضها بأرقام باللون الأحمر. تشير الخطوط المتقطعة عند 1 إلى أن العلاجات لها قيمة متساوية كضوابط موحدة. النتائج تمثل بيانات تم توليدها في تجربتين مستقلتين على الأقل وتُعبر عنها كمتوسط.SEM، وذو جانبينتم فحص القيم بواسطة اختبار الطالب-اختبار. يتم توفير بيانات المصدر كملف بيانات المصدر. تركيز مرتفع بشكل ملحوظ في البراز (الشكل التوضيحي 6c) وأظهر تأثيرات مماثلة في زيادة CD25فوكس بي 3تريغس وتقليل IL17CD4خلايا T في الأنسجة الثلاثة مقارنةً بتكوين مستعمرات Dub في الفئران WT. بشكل غير متوقع، ظلت هناك اختلافات صغيرة في ترددات مجموعتي خلايا T بين مجموعتي Gpr43-/-Dub وGpr43-/-Veh، في حين لم تُلاحظ أي اختلافات بين Gpr43-/-Prop أو Gpr43.-مجموعات Veh (الشكل 3h و i والشكل التوضيحي 6d)، مما يشير إلى مشاركة كاملة لـ GPR43 في إعادة التوازن بين Treg/Th17 التي تتوسطها البروبيونات في سياق التهاب القولون الناتج عن DSS.
متسقًا مع النتائج الملاحظة في تنظيم Treg/Th17، تم ممارسة تأثير وقائي مشابه بواسطة WT-Prop مقارنةً بفئران WTDub فيما يتعلق بظاهرة المرض (الشكل 3j والشكل التوضيحي 6e، f)، التغيرات النسيجية (الشكل 3k، I والشكل التوضيحي 6g)، سلامة الحاجز المخاطي (الشكل 3m والشكل التوضيحي ) والالتهاب (الشكل التوضيحي 6j)، في حين كانت الظاهرة الواقية معطلة في Gpr43الفئران. ومع ذلك، على الرغم من تضييق النمط الظاهري، كانت هناك اختلافات لا تزال قائمة بين مجموعتي Gpr43-/-Dub وGpr43-/-Veh، مما يشير إلى أنه بالإضافة إلى الأحماض الدهنية قصيرة السلسلة (وخاصة البروبيونات)، قد تكون هناك أيضًأ مستخلصات أخرى مشتقة من Dub أو مسارات مستقلة عن GPR43 قد تكون متورطة في التحفيز المدفوع من Dub للخلايا التائية التنظيمية (cTregs) وتحسين التهاب القولون الناتج عن DSS.
يحفز دوب خلايا Tregs الإيجابية لـ CD25 و Foxp3 من خلال تعزيز استقلاب التربتوفان بواسطة IDO1 في خلايا الدندريت في القولون
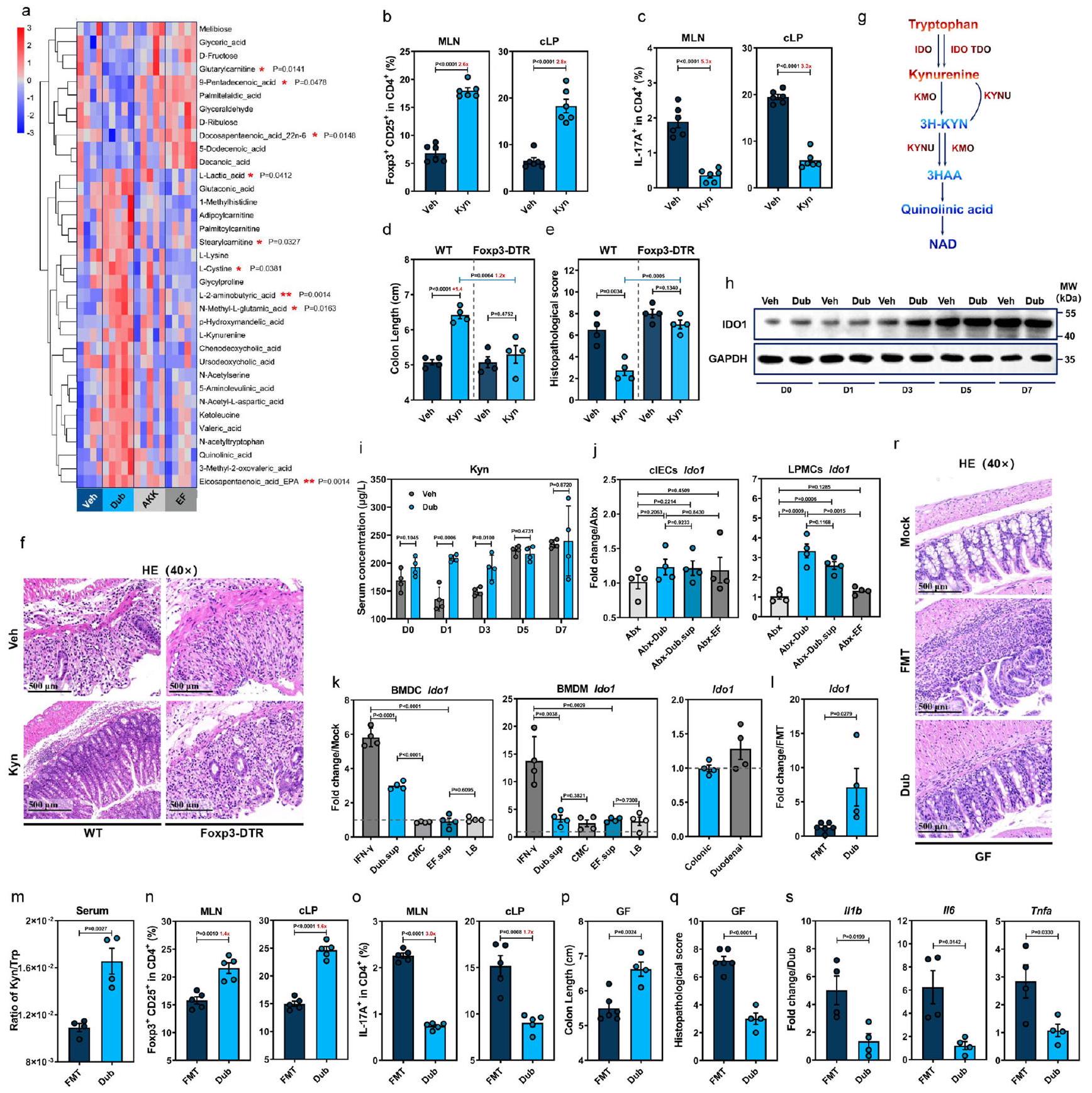
للبحث عن مستقلبات أخرى مشتقة من دوب قد تعزز CD25فوكس بي 3لتحفيز Treg، قمنا بإجراء تحليل استقلاب غير مستهدف على عينات القولون المجمعة من الفئران البرية التقليدية المستعمرة بـ Dub وAkk وEF أو ضوابط Veh في اليوم السابع، وتم الكشف عن إجمالي 253 مركبًا. أظهر تحليل التمييز باستخدام المربعات الجزئية أن هناك اختلافات ملحوظة في الملفات الاستقلابية لـ Dub مقارنةً بضوابط Veh (الشكل التوضيحي التكميلي 7a). من بين أعلى 35 من المستقلبات المختلفة الأكثر وفرة المدرجة في خريطة الحرارة، كان هناك 9 منها تختلف بشكل ملحوظ إحصائيًا بين الفئران المستعمرة بـ Dub والحيوانات الضابطة غير المستعمرة (الشكل 4a). أشارت تحليلات KEGG إلى أن استقلاب التربتوفان كان أقل بشكل ملحوظ في المجموعة المستحثة بـ DSS في اليوم السابع (الشكل التوضيحي التكميلي 7b)، بينما أدت الاستعمار بـ Dub إلى زيادة في L-Kyn وحمض الكينولين مقارنةً بفئران Akk وEF أو Veh على الرغم من أن الفرق لم يكن ذا دلالة إحصائية (الشكل 4a).
لتأكيد الدور المحتمل لـ Kyn في إعادة توازن استجابات Treg/Th17 في سياقات الالتهاب المعوي، تم إخضاع الفئران التقليدية WT و Foxp3-DTR لعلاج Kyn قبل وأثناء إدارة DSS (الشكل التوضيحي التكميلي 7c). يتماشى ذلك مع منشور سابقأدى علاج Kyn إلى مستوى أعلى بشكل ملحوظ من CD25فوكس بي 3تريغس ومستوى أقل بشكل كبير منCD4خلايا T في الطحال، العقد اللمفاوية المتوسطة و cLP في اليوم السابع في الفئران WT (الشكل 4ب، ج والمكمل الشكل 7د). علاوة على ذلك، كانت أنماط المرض (الشكل 4د والمكمل الشكل 7هـ)، التغيرات النسيجية المرضية (الشكل 4هـ، و) أو الاستجابات الالتهابية (المكمل الشكل 7و) هي نفسها في فئران Foxp3-DTR، على الرغم من الكشف عن تركيزات كينورينين في المصل بشكل كبير في كلا خطي الفئران في اليوم السابع (المكمل الشكل 7ز). لذلك، فإن استعمار دوب يعزز إنتاج الكاتابوليت تريبتوفان كينورينين، الذي يحفز CD25.فوكس بي 3تعمل خلايا Tregs على exert تأثير وقائي على التهاب القولون الناتج عن DSS.
يتم استقلاب غالبية التربتوفان المشتق من الطعام عبر مسار الكينورينين من خلال إنزيم IDO1 في الخلايا المخاطية والمناعية أو IDO2 وإنزيم تريبتوفان 2,3-ديوكسيجيناز (TDO) في الكبد. (الشكل 4g). وبالتالي، قمنا بقياس حركية تعبير IDO1 في القولون من WT-Dub أو WT-Veh في أوقات مختلفة بعد علاج DSS (الشكل التوضيحي 7c) ووجدنا أن IDO1 كان مرتفعًا بشكل ملحوظ في D1 وD3 وD5، ولكن ليس في D7 في WT-Dub مقارنةً بمجموعات التحكم WT-Veh (الشكل 4h والشكل التوضيحي 7h، i). وبما يتماشى مع هذه النتائج، أدى استعمار Dub إلى زيادة كبيرة في تركيز كينورين في المصل (الشكل 4i) في المراحل المبكرة من تقدم التهاب القولون (D0-D3).
لتحديد المصادر الخلوية لزيادة تعبير IDO1 استجابةً لمستقلبات مشتقة من Dub، قمنا باختبار تعبير Ido1 في مجموعات Abx-Dub و Abx-Dub.sup (الشكل التوضيحي 7c) وأظهرنا زيادة ملحوظة في تعبير Ido1 في خلايا أحادية النواة في الغشاء المخاطي القولوني (cLPMCs)، ولكن ليس في خلايا الظهارة القولونية (clECs) قبل وبعد 3 أيام من إعطاء DSS (الشكل 4j). علاوة على ذلك، أدى إضافة Dub.sup إلى خلايا دندريتية مشتقة من نخاع العظام من النوع البري (BMDCs) إلى زيادة ملحوظة في تعبير Ido1 مقارنةً بمجموعة التحكم في وسط الثقافة، بينما لم يكن لـ EF.sup أي تأثير معزز مماثل (الشكل 4k)؛ كان هناك اتجاه مشابه لخلايا البلعمة المشتقة من نخاع العظام من النوع البري (BMDMs) تحت علاج Dub.sup على الرغم من أنه لم يكن ذا دلالة إحصائية (الشكل 4k). على العكس من ذلك، لم يتغير تعبير Ido1 في الأعضاء القولونية وارتفع قليلاً في الأعضاء الاثني عشرية في وجود Dub.sup (الشكل 4k). تشير هذه النتائج إلى أن مستقلبات مشتقة من Dub تعزز بشكل تفضيلي تعبير IDO1 في البلعميات أحادية النواة المعوية.
لاستبعاد إمكانية أن تكون الأنماط الظاهرية قد تأثرت بالبكتيريا المعوية المقاومة للمضادات الحيوية المتبقية في مجموعة المضادات الحيوية لدينا، تم استعماري الفئران الخالية من الجراثيم (GF) مع Dub (GF-Dub) أو تلقت زراعة ميكروبية من فئران WT التقليدية (GF-FMT) قبل علاج DSS (الشكل التوضيحي 7j). بشكل متسق، كان لدى GF-Dub أيضًا مستويات أعلى بشكل ملحوظ من Ido1 في cLPMCs (الشكل 4I) ونسبة Kyn/Trp في المصل (الشكل 4m) مقارنةً بـ GF-FMT. وبالمثل، أظهر GF-Dub مستويات أعلى بشكل ملحوظ من CD25.فوكس بي 3تريغس ومستويات منخفضة بشكل ملحوظ من IL17CD4تظهر خلايا T في الطحال، والعقد اللمفاوية المتوسطة، والقولون الجانبي أكثر من GF-FMT (الشكل 4n، o والشكل التكميلية 7k). أيضًا، أظهر GF-Dub ظاهرة مرضية مخففة (الشكل 4p والشكل التكميلية 7l)، وآفات نسيجية (الشكل 4q، r) واستجابات التهابية (الشكل 4s) مقارنة بـ GF-FMT. من المثير للاهتمام أن الاستعمار الأحادي لـ Dub أدى إلى زيادة في ZO-1، ومستويات OCLN قابلة للمقارنة، وحتى مستويات Muc2 أقل من تلك الموجودة في فئران GF-FMT (الشكل التكميلية 7m). بشكل عام، أظهرنا أن استقلاب التربتوفان في DCs تم تعزيزه بواسطة Dub، وأن تعزيز Kyn الناتج عن زيادة تعبير IDO1 أعاد توازن استجابات Treg/Th17 وأدى في النهاية إلى تأثير وقائي على التهاب القولون الناتج عن DSS.
المستقلب المشتق من دوب L-Lys ينظم تعبير IDO1 المعتمد على AhR في خلايا DCs القولونية
للتحقيق في المستقلبات البكتيرية التي تتوسط زيادة تنظيم IDO1، قمنا بإجراء تحليلات غير مستهدفة للميتابولوم على Dub.sup بعد زراعته في وسط كربوهيدرات اللحم المفروم (CMC) (الشكل 5a). أسفرت تحليلات الكروماتوغرافيا السائلة-الطيف الكتلي المت tandem (LC-MS/MS) عن 5 مستقلبات مشتقة من Dub، N-acetyl-L-Asp (NAA)، L-Lysine (Lys)، L-Lactic acid (Lac)، L-حمض -أمينوبوتيريك (أبو) وكيتوليوسين (حمض 4-ميثيل-2-أوكوبنتانويك، 4-MOV) بتركيزات أعلى نسبيًا في كل من كروماتوغرافيا تبادل الأيونات الموجبة والسالبة (الشكل التوضيحي 8a). كانت هذه المستقلبات أيضًا مرتفعة في القولون لدى الفئران المستعمرة بدوب (الشكل 4a).
بعد معالجة WT mBMDCs بهذه المركبات الخمسة، اكتشفنا أن Lys و4-MOV قد أديا إلى زيادة كبيرة في Ido1 (الشكل 5b). IFN-تم استخدام محفز IDO1 المدروس جيدًا في خلايا DCs كتحكم إيجابيوفقًا لما ذكره غارغارو وآخرون.كما حددنا أيضًا أن معالجة Kyn الخارجي لـ mBMDCs زادت من تعبير Ido1 (الشكل 5b). وبشكل متسق، زادت إدارة Lys أو 4-MOV للفئران WT (الشكل التوضيحي 8b) بشكل كبير من تعبير IDO1 في القولون في اليوم الثالث (الشكل 5c). من بين 5 مستقلبات، حسنت Lys وLac بشكل ملحوظ من تقصير القولون (الشكل 5d)، وتلف الوصلات الضيقة والالتهاب المعوي (الشكل التوضيحي 8c)، على الرغم من أن النمط الظاهري الوقائي لم يكن قويًا بشكل عام مقارنةً بمعالجة Dub.sup.
نظرًا لأن 4-MOV لم يظهر أي تأثير وقائي ضد التهاب القولون الناتج عن DSS، اخترنا التركيز على الدور المحتمل لللايسين في تعديل استقلاب التربتوفان المدفوع بـ IDO1. في الواقع، أدى المعالجة المسبقة باللايسين إلى زيادة ملحوظة في تعبير Ido1 في خلايا WT mBMDCs بطريقة تعتمد على الجرعة والوقت (الشكل التوضيحي التكميلي 8d، e). علاوة على ذلك، أدى إضافة كل من اللايسين والتربتوفان معًا إلى خلايا WT mBMDCs إلى انخفاض كبير.
الشكل 4 | D. نيويوركنسيس يعزز CD25فوكس بي 3تريغس من خلال تعزيز استقلاب التربتوفان بواسطة IDO1 في الخلايا التغصنية. خريطة حرارية تظهر المستقلبات المختلفة في القولون في اليوم السابع بعد علاج DSS من فئران C57BL/6J البرية (WT) المعالجة بالسيارة (Veh، ) أو مستعمرة مع D. newyorkensis (Dub) أو A. muciniphila (Akk) أو E. faecalis (EF) ( ). تمثل التعليقات الإحصائية المقابلة للمواد الأيضية المختلفة الفروق بين الفئران المستعمرة بدوب وغير المستعمرة. ب، ج مجموعات خلايا T في العقد اللمفاوية المساريقية (MLN) والطبقة الظهارية القولونية (cLP) من المعالجة بـ Veh أو Kyn ( تم فحص فئران WT التقليدية بواسطة التدفق. فئران Foxp3-DTR المعالجة بـ WT أو DTتم علاجهم بـ Kyn وتعرضوا لإدارة DSS لمدة 7 أيام، وتم فحص طول القولون (د) وعلم الأمراض النسيجي (هـ، و). ج رسم تخطيطي لمسار تحلل Kyn في استقلاب التربتوفان. ح، ط حركية تعبير IDO1 في القولون (ح) وتركيز Kyn في المصل (ط) في الفئران التقليدية WT المستعمرة بـ Dub أو Veh. ) في الأوقات المحددة بعد إدارة DSS. تعبير IDO1 في خلايا الظهارة المعوية القولونية (cIECs) أو خلايا أحادية النواة في الطبقة الأساسية (cLPMCs) المستخرجة
من فئران Abx-Dub و Abx-EF و Abx-Veh أو Abx-Dub.sup ( ) في إدارة ما بعد DSS في D3. تعبير Ido1 في البلعميات المشتقة من نخاع العظام الفأري (mBMDMs) والخلايا التغصنية المشتقة من نخاع العظام (mBMDCs) أو الأعضاء المعوية المعالجة بمستخلصات ثقافة Dub أو EF لمدة 18 ساعة. الفئران الخالية من الجراثيم (GF) التي تتلقى زراعة ميكروبيوتا البراز (FMT، ) من فئران WT التقليدية أو مستعمرة مع Dub ( ) تعرضت لعلاج DSS لمدة 7 أيام. تعبير Ido1 في cLPMCs ( نسبة تركيز المصل منمجموعات الخلايا ( ) ( طول القولون في MLN و cLPالتغيرات المرضية ) (FMT, ; دوب، ) والسيتوكينات المؤيدة للالتهابات في cLPMCs ( ) ( تم تحديدها. تم تغيير ترددات مجموعات خلايا T ( ) أو طول العمود (d) بين المجموعات تم حسابه وعرضه بأرقام باللون الأحمر. الخطوط المتقطعة عند 1 تشير إلى أن العلاجات لها قيمة متساوية كضوابط موحدة. النتائج تمثل بيانات تم توليدها في تجربتين مستقلتين على الأقل وتُعبر عنها كمتوسط SEM، وذو جانبينتم فحص القيم بواسطة اختبار الطالب-اختبار. يتم توفير بيانات المصدر كملف بيانات المصدر. من تريبتوفان وزيادة ملحوظة في كينورينين في السائل الخلوي الفائق (الشكل التكميلي 8f). كانت هذه النتيجة مرتبطة بتعبير أكثر قوة عن Ido1 (الشكل التكميلي 8g) كما يتضح من زيادة نسبة كينورينين/تريبتوفان، والتي فقدت في خلايا الماكرومونوسيت المشتقة من نخاع العظام المعالجة بشكل مشابه والمجمعة من Ido1الفئران (الشكل 5e)، مما يشير إلى أن مسار Kyn تم تعزيزه بواسطة Lys بطريقة تعتمد على IDO1.
يتم نقل الليسين خارج الخلية عبر SLC7A1 و SLC7A2 و SLC7A3بينما يتم نقل التربتوفان بواسطة النظام L، وهو هيتيروديمر يتكون من سلسلة ثقيلة (مشفرة بواسطة Slc3a2) وسلسلة تحفيزية L LAT1 (مشفرة بواسطة Slc7a5)تمت ملاحظة زيادة في تعبير Slc7a1 وSlc7a2 وSlc7a5 وSlc3a2 في خلايا mBMDCs المجمعة من كل من الفئران التقليدية وGF والمعالجة بـ Lys، مقارنة بالخلايا غير المعالجة (الشكل التوضيحي 8h، i)، بينما لم يتم الكشف عن Slc7a3 (البيانات غير معروضة). وبالتالي، قد تؤدي خلايا mBMDCs المعالجة بـ Lys إلى زيادة تعبير SLC7A5 وSLC3A2 لزيادة امتصاص Trp، مما يغذي مسار Kyn في تحلل Trp.
الشكل 5 | يعزز L-Lys استقلاب التربتوفان في الخلايا الشجرية لتحفيز تثبيط المناعة المعتمد على Treg من خلال تعزيز تعبير IDO1. أ خريطة حرارية تظهر المستقلبات المعبر عنها بشكل مختلف من D. newyorkensis (Dub) و A. muciniphila (Akk) و E. faecalis (EF) والمستخلصات المزروعة المقابلة (مرق الكربوهيدرات من اللحم المفروم، CMC؛ وسط تغذية القلب والدماغ، BHI؛ وسط لوريا-بيرتاني، LB) بعد 48 ساعة. ب تعبير Ido1 في خلايا دهنية مشتقة من نخاع العظام في الفئران (BMDCs) المعالجة بـ N-acetyl-L-Asp (NAA؛ 1 مليمول)، L-Lys (8 مليمول)، حمض اللبنيك L- (Lac؛ 100 مليمول)، L-حمض -أمينوبوتيريك (أبو؛ 0.1 مللي مول) ، كيتوليوسين ( ) و Kyn ( 0.2 مليمول ) ( ). تم قياس تعبير IDO1 وطول القولون في فئران Abx المعالجة بالمواد الأيضية في اليوم الثالث أو السابع بعد إعطاء DSS، على التوالي. ). نشاط IDO1 في خلايا BMDCs المعالجة بـ Lys أو IFN تم تحديده بواسطة نسبة كينين/تريبتوفان في سوائل زراعة الخلاياتم استعمار الفئران التقليدية WT، والفئران المعالجة بـ DT Foxp3DTR، أو الفئران Ido1-/- بدوب أو تم علاجها بـ ليس. ) وتعرضت لإدارة DSS لمدة 7 أيام. مجموعات خلايا T ( ) في العقد اللمفاوية المساريقية (MLN) وطبقة الغشاء المخاطي القولوني ( طول القولون (cLP) ) ( ) و علم الأمراض النسيجيةكانوا مصممين.تركيز سيروم كين في الفئران التقليدية WT المعالجة، وفئران Foxp3-DTR المعالجة بـ DT أو Ido1-/- قبل التعرض لـ DSS (WT، Foxp3-DTR،; إيدو1 1 التصوير الحي للقولون من الفئران المعدلة وراثيًا IL-17-EGFP المعالجة بمركب السيارة (Veh) أو Dub أو Lys أو Kyn. تم تعريض الفئران الخالية من الجراثيم المعالجة بـ Lys أو المركب Veh لعلاج DSS لمدة 7 أيام وتعبير Ido1 في القولون.مجموعات خلايا Tفي MLN و cLPطول القولونعلم الأمراض النسيجية (q) والسيتوكينات المؤيدة للالتهابات ( ) ( تم تقييم تعبير Ido1 في خلايا الدندريت المشتقة من نخاع العظام (BMDCs) المعالجة بـ Lys- (8 مليمول) أو السائل الفائق لـ Dub (Dub.sup) المستخرجة من الفئران الخالية من الجراثيم (GF) بعد 18 ساعة من المعالجة. ). طي التغير في ترددات مجموعات خلايا T ( ) أو طول النقطتين ( ) تم حسابها وعرضها بأرقام باللون الأحمر. الخطوط المتقطعة عند 1 تشير إلى أن العلاجات لها قيمة متساوية كضوابط موحدة. النتائج تمثل بيانات تم توليدها في ثلاث تجارب مستقلة وتُعبر عنها كمتوسط SEM، وثنائي الجانب-تم فحص القيم بواسطة اختبار الطالب-اختبار. يتم توفير بيانات المصدر كملف بيانات المصدر.
لاختبار الأهمية الفسيولوجية لتعزيز IDO1 الذي يتم بوساطة Lys في قمع الالتهاب من خلال تحفيز Treg في الجسم الحي، قمنا بإعطاء فئران WT التقليدية، Ido1-/- أو Foxp3-DTR Dub أو Lys قبل التعرض لـ DSS (الشكل التوضيحي التكميلي 8j). وُجد أن علاج Lys زاد منفوكس بي 3تريغس وانخفاضخلايا T إلى حد مشابه لما فعلته استعمار دوب في الطحال، والعقد اللمفية المتوسطة، والقولون السفلي مقارنة بالفئران WT المعالجة بـ Veh، لكن Lys فشل في التعديلفوكس بي 3تريغس في جميع الأنسجة الثلاثة أوالخلايا في cLP من Ido1الفئران (الشكل 5f و g والشكل التوضيحي 8k).
علاوة على ذلك، كان لليس تأثير مفيد واضح على أنماط الأمراض (الشكل 5h والشكل التكميلي 81)، والتغيرات النسيجية المرضية (الشكل 5i، j)، ووظيفة الحاجز المخاطي (الشكل التكميلي 8m)، والاستجابات الالتهابية (الشكل التكميلي 8n) في الفئران البرية التقليدية، في حين تم إلغاء التأثير الوقائي فيأو فئران Foxp3DTR. يتماشى مع بيانات التدفق، ظل تأثير وقائي خفيف في فئران Ido1-/-Dub و Foxp3-DTR-Dub كما يتضح من التقييم النسيجي (الشكل 5i، j). علاوة على ذلك، حددنا أن فئران WT-Lys، ولكن ليس فئران Ido1-/-Lys، أظهرت زيادة في تركيز كينورين في المصل (الشكل 5k). على الرغم من أن التأثير كان أصغر من استعمار Dub، فإن إدارة Lys و Kyn لفئران IL-17-EGFP المعالجة بـ DSS أدت إلى انخفاض مماثل في تعبير IL-17 القولوني، كما تم قياسه بواسطة التصوير الحي (الشكل 51).
بعد ذلك، حددنا ما إذا كانت الظاهرة الملحوظة في الفئران البرية التقليدية ذات الفلورا المعوية الطبيعية صحيحة أيضًا في البيئات الخالية من الجراثيم. في الواقع، أظهرت الفئران الخالية من الجراثيم (GF-Lys) زيادة في تركيز الليسين في المصل قبل التعرض لـ DSS، وتم تأكيد زيادة تركيز الليسين في البراز في مجموعة GF-Dub (الشكل التوضيحي 9b). كما هو متوقع، أدى إعطاء الليسين للفئران الخالية من الجراثيم (الشكل التوضيحي 9a) إلى زيادة تعبير Ido1 في cLP (الشكل 5m)، وزيادة تركيز كينورين في المصل (الشكل التوضيحي 9b)، وزيادة تنظيم…فوكس بي 3تريغس وانخفاض التنظيمالخلايا في الطحال و cLP (الشكل 5n، o والشكل التوضيحي 9c)، ولكن تم تقليل CD25 بشكل غير متوقعفوكس بي 3تريغس في MLN (الشكل 5n). كان لليس تأثير مريح على نمط المرض (الشكل 5p والشكل التكميلية 9d)، وإصابة الأنسجة (الشكل 5q والشكل التكميلية 9e)، وتلف الحاجز المخاطي بالإضافة إلى الالتهاب المحلي والمحيطي (الشكل 5r والشكل التكميلية 9f). علاوة على ذلك، قمنا بمعالجة خلايا GF mBMDCs مسبقًا بالليس ووجدنا زيادة ملحوظة في تعبير Ido1 (الشكل 5s)، مما يوحي بأن تحفيز IDO1 في DCs بواسطة اليس كان على الأرجح نتيجة لتغيرات داخل الخلايا لا تعتمد على إشارات مشتقة من الميكروبات.
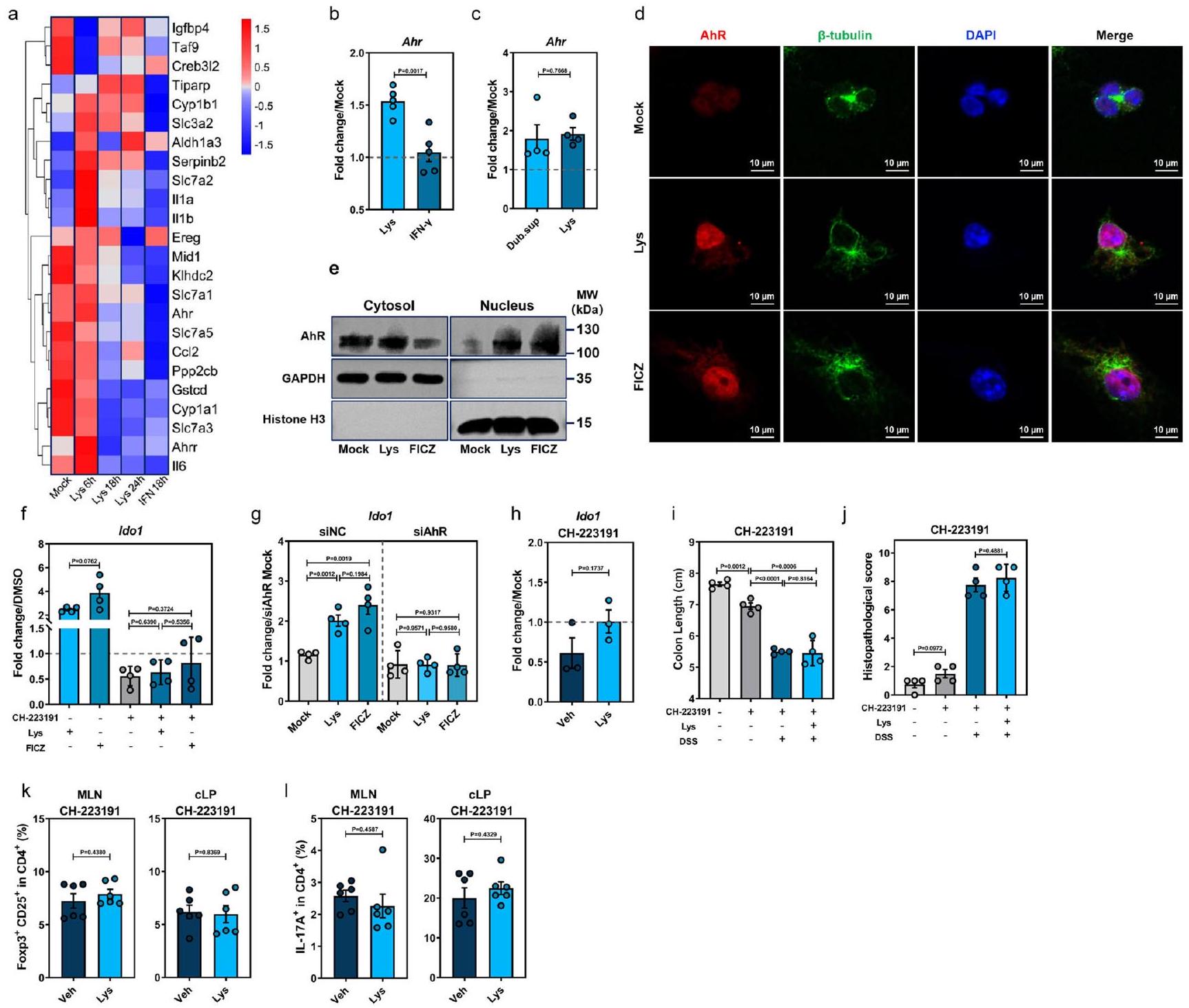
لفهم أفضل لكيفية تعزيز Lys تعبير Ido1 في DCs، قمنا بإجراء تسلسل RNA على BMDCs WT المعالجة بـ Lys، مما كشف عن تحفيز واسع لجينات استجابة AhR (الشكل 6a). أكدنا زيادة التعبير عن Ahr (الشكل 6b، c) وجينات الهدف AhR Aldh1a3 وCyp1a1 وCyp1b1 وTiparp وIl1b وIl6 في كل من BMDCs WT التقليدية وGF عند 6 ساعات بعد تحفيز Lys (الشكل التكميلية 10a، b). علاوة على ذلك، أدى Lys إلى انتقال AhR إلى النواة بعد 6 ساعات من العلاج، مما يشير إلى تأثير فوري لـ Lys على AhR (الشكل 6d). وفقًا لذلك، أظهر التحليل الغربي لـ mBMDCs المعالجة بـ Lys انخفضت التوطين السيتوبلازمي بالتوازي مع زيادة تراكم AhR النووي، مقارنةً بتلك التي يسببها المنبه AhR 6-for-mylindolo[3,2-b]carbazole (FICZ) (الشكل 6e). تم تثبيط تنشيط AhR وزيادة تعبير Ido1 استجابةً لـ Lys بواسطة مضاد AhR CH-223191 (الشكل 6f) أو تقليل Ahr (الشكل 6g والشكل التكميلية 10c).
لتقييم تأثيرات ليسين على إشارات AhR في الجسم الحي، تم معالجة مجموعات من الفئران البرية قبل إعطاء DSS إما مع ليسين بالاشتراك مع، أو وحيدًا (الشكل التوضيحي 10d). يتماشى مع بيانات المختبر، لم تعزز إدارة الليزين تعبير Ido1 في cLPMCs في وجود CH-223191 في اليوم الثالث بعد علاج DSS (الشكل 6h). يتفق ذلك مع التأثيرات الوقائية المبلغ عنها لـ AhR في التهاب القولون الناتج عن DSS.، أظهرت مجموعة WT-CH-223191 تعبيرًا متضررًا بوضوح لمستقبل Ahr في القولون (الشكل التوضيحي 10e) وساءت ظاهرة المرض (الشكل 6i، j). الاختلافات فيفوكس بتريغس أو IL17تم فقدان الخلايا والسيتوكينات المؤيدة للالتهابات في cLPMCs بين WT-Lys و WT-Veh في وجود CH-223191 (الشكل 6k، I والشكل التكميلي 10f، g). بشكل جماعي، تشير هذه النتائج إلى أن Lys يعزز تعبير IDO1 لتوفير الحماية ضد التهاب القولون الناتج عن DSS من خلال تنشيط AhR.
المتماثل البشري لدوب، C. innocuum، ينتج L-لايسين ويعززفوكس بي 3تريغس لتخفيف الإصابة الناتجة عن DSS من خلال زيادة تعبير IDO1 في خلايا DCs
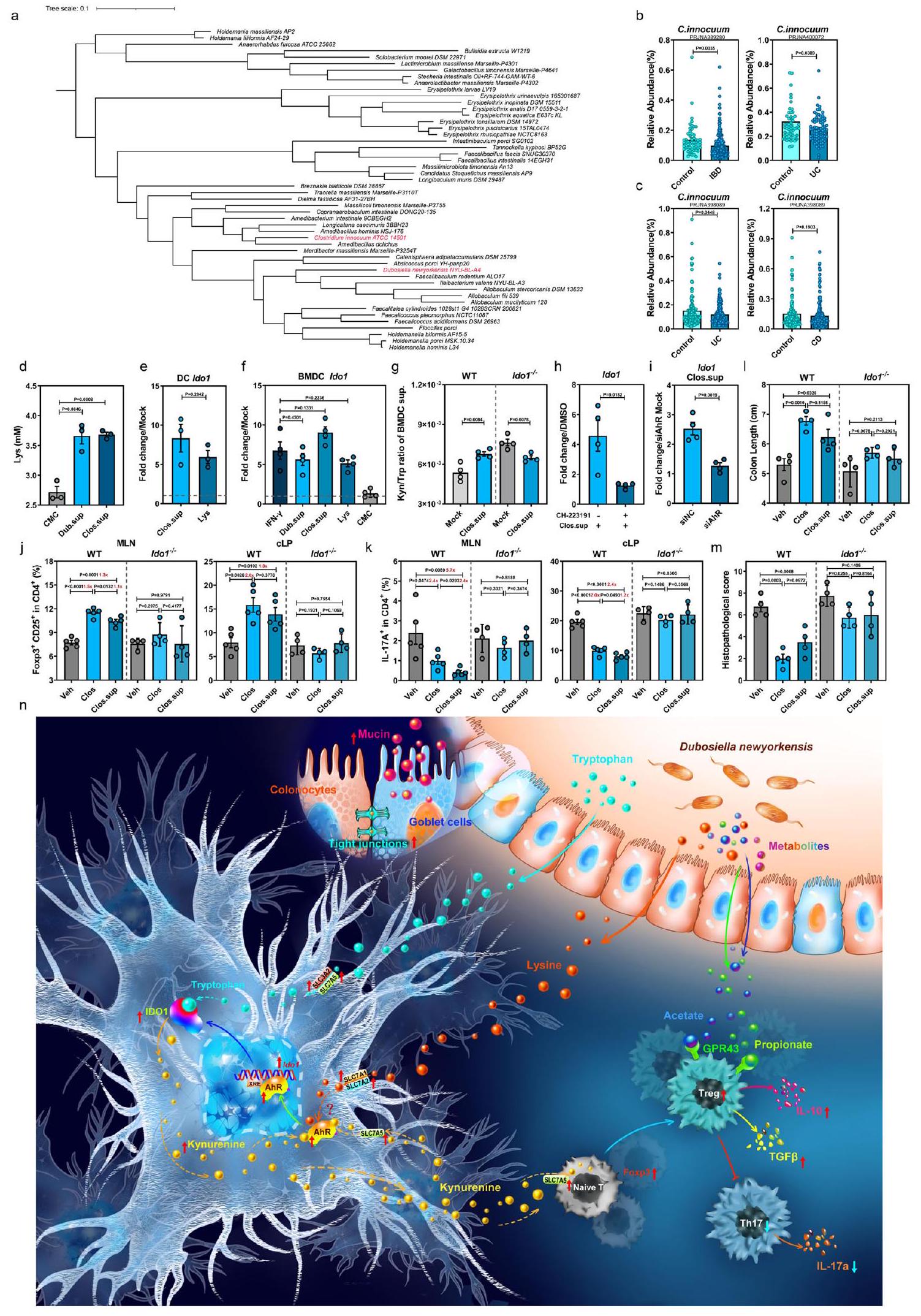
مؤخراً، تم توثيق أن الزيادات في عائلة Erysipelotrichaceae المعوية كانت مرتبطة بشكل سببي بانخفاض خطر الإصابة بمرض التهاب الأمعاء (IBD).. وبالتالي، قمنا بإجراء تحليل تطوري لعائلة Erysipelotrichaceae وأظهرنا أن نوعًا بشريًا متعايشًا غير ممثل بشكل كافٍ، C. innocuum (المختصر هنا بـ Clos)، هو قريب جدًا من Dub (الشكل 7a، المسافة التطورية ). قمنا بمقارنة مجموعات البيانات الميتاجينية المعوية لمجموعات مرض التهاب الأمعاء المتاحة للجمهور مع الضوابط الصحية المتطابقة.اكتشاف أن مستوى Clos كان منخفضًا بشكل ملحوظ في المرضى الذين يعانون من التهاب الأمعاء (IBD)، وخاصة أولئك الذين يعانون من التهاب القولون التقرحي (UC)، مقارنة بالأفراد الأصحاء (الشكل 7ب). بالإضافة إلى ذلك، أظهرت تحليل قائم على مجموعة من الولايات المتحدة انخفاضًا في وفرة Clos في المرضى الذين يعانون من UC. أو قرص مضغوط بالمقارنة مع الضوابط الصحية (الشكل 7c).
مماثل لدوب، أنتج كلوس ليس في السائل الثقافي (الشكل 7د). علاوة على ذلك، عزز كل من كلوس.سوب وليس تعبير إيدو1 في خلايا DC الأولية المستخرجة من PBMCs البشرية (hPBMCs) (الشكل 7هـ) و WT mBMDCs (الشكل 7و)، مع زيادة نسبة كين/ترب في السائل الثقافي لـ mBMDC (الشكل 7ز). على النقيض من ذلك، لم يتم ملاحظة هذه الظاهرة في mBMDCs Ido1-/- (الشكل 7ز)، وتم إلغاؤها بواسطة CH-223191 (الشكل 7ح) أو RNA صغير متداخل محدد لـ AhR (siAhR) (الشكل 7ط). في mBMDCs GF المعالجة بـ Lys، لوحظ زيادة في Ahr وجيناته المستهدفة، بالإضافة إلى ناقلات Lys وTrp بعد 6 ساعات من العلاج (الشكل التكميلي 11أ، ب). وبالمثل، تم عرض تعبير أعلى لـ Ido1 بعد 18 ساعة من العلاج (الشكل التكميلي 11ج). معًا، تشير هذه النتائج إلى أن كلوس، النظير البشري لدوب، قد
الشكل 6 | L-Lys ينشط محور AhR-IDO1 في خلايا DCs لتحفيز المناعة المثبطة استجابات Treg. تحليل النسخ الجيني لعينة وهمية، Lys، IFN--معالجة خلايا BMDCs من الفئران في الأوقات المحددة بعد المعالجة (وهمية،; ليس IFN 18 ساعة. ب تعبير في ليسين- أو إنترفيرون-BMDCs المعالجة المستخرجة من الفئران العادية WT. مع التعبير في خلايا BMDCs المعالجة بـ Lys- أو Dub.sup المستخرجة من الفئران GF ( نقل AhR (باللون الأحمر) إلى النواة (DAPI، باللون الأزرق) من السيتوبلازم (-توبولين، أخضر) من خلايا BMDCs الفأرية بعد 6 ساعات من المعالجة بـ Lys أو المنبه AhR 6-formylindolo[3,2-b]carbazole (FICZ). تحليل بروتين AhR في الكسور النووية والسيتوبلازمية من خلايا BMDCs الفأرية المعالجة بـ mock أو Lys أو FICZ بعد 6 ساعات من المعالجة بواسطة تقنية الويسترن بلوت.تعبير Ido1 في خلايا BMDCs الفأرية المعالجة بـ Lys أو FICZ أو مضاد AhR CH-223191، أو مجموعة من CH223191 و Lys/FICZ ). تعبير Ido1 في خلايا BMDCs الفأرية التي تم نقلها بواسطة التحكم السلبي (siNC) أو RNA صغير متداخل محدد لـ AhR (siAhR) وتم علاجها بـ Lys أو FICZ بعد 18 ساعة من العلاج ( ” ). للتجارب الموضحة في ( )
فيما سبق، تم استخدام ما يلي: ليسين (8 مللي مول)، FICZ (300 نانومول)، و CH-223191 ( ). للتجارب الموضحة في ( ) أدناه، فئران WT التقليدية ( ) تم علاجها بـ Lys ( )، CH-223191 ( )، أو ليس مع CH-223191 وتعرض لإدارة DSS.تعبير Ido1 في cLPMCs من الفئران WT المعالجة بـ Veh أو Lys في اليوم الثالث بعد إعطاء DSS في وجود CH-223191طول القولون (i)، التغيرات النسيجية المرضية بواسطة صبغة HECD25فوكس بي 3تريغس ) و IL-17 CD4خلايا T ) ( تم فحص (MLN و cLP من الفئران التقليدية WT المعالجة بـ Veh أو Lys في وجود CH-223191 في اليوم السابع بعد علاج DSS. تشير الخطوط المتقطعة عند 1 إلى أن العلاجات لها قيمة متساوية كضوابط موحدة. النتائج تمثل بيانات تم توليدها في ثلاث تجارب مستقلة وتعبر عنها كمتوسط.SEM، وثنائي الجانب-تم فحص القيم بواسطة اختبار الطالب-اختبار. يتم توفير بيانات المصدر كملف بيانات المصدر. كما أن لها تأثيرًا معززًا على استقلاب التريبتوفان من خلال مسار الكينورينين عن طريق تعزيز تعبير IDO1 في كل من خلايا المناعة البشرية والفأرية عبر تنشيط AhR.
بعد ذلك، قمنا باختبار Clos أو Clos.sup في الفئران البرية WT والفئران Ido1-/- (الشكل التكميلي 11d)، ولاحظنا أن كل من استعمار Clos (الشكل التكميلي 11e) وإدارة Clos.sup زادت بشكل كبير من CD25.فوكس بي 3تريغس وانخفاض IL17CD4خلايا T في الطحال، والعقد اللمفاوية المتوسطة، والأنسجة اللمفاوية المعوية في الفئران الطبيعية، ولكن ليس في Ido1الفئران (الشكل 7j و k والشكل التكميلي 11f). علاوة على ذلك، فإن استعمار Clos أو العلاج بـ Clos.sup في الفئران WT خفف من تقصير القولون (الشكل 71 و
الشكل التكميلي 11g)، إصابات الأنسجة القولونية (الشكل 7m والشكل التكميلي 11h) والاستجابات الالتهابية (الشكل التكميلي 11i)، وهو تأثير لم يُلاحظ فيلذا، يُقترح أن Clos قد يوفر الحماية ضد التهاب القولون في البشر من خلال تحسين استجابات Treg/Th17 غير المتوازنة من خلال تعزيز تعبير IDO1.
نقاش
تشير مجموعة متزايدة من الأدبيات إلى أن الميكروبيوم المعوي يلعب دورًا أساسيًا في الحفاظ على توازن الأمعاء. من خلال تعزيز وظيفة الحاجز الظهاري والتحمل المناعيظهر الأيض كلاعب جديد في وظائف المناعة المخاطية والالتهاب مع ظهور مجال المناعة الأيضية الجديد.على الرغم من أن دور المستقلبات المشتقة من الميكروبات المحددة وآلية عملها لا تزال غامضة إلى حد كبير. هنا كشفنا أن البكتيريا المتعايشة المقاومة للنيوميسين، دوب، تلعب دورًا دور محوري في تعزيز سلامة الحاجز المخاطي وإعادة توازن استجابات Treg/Th17 للحفاظ على التوازن المعوي من خلال إنتاج الأحماض الدهنية قصيرة السلسلة (خصوصًا البروبيونات) والليسين. لقد أظهرنا دورًا غير معروف سابقًا لليسين المشتق من الميكروبات المعوية في تعزيز تعبير IDO1 في خلايا DCs القولونية. من خلال إعطاء دوب أو ليسين، تمكنا من تحريف استقلاب التربتوفان الداخلي.
الشكل 7 | النظير البشري لـ D. newyorkensis، Clostridium innocuum، ينتج L-Lys ويحمي من التهاب القولون الناتج عن DSS من خلال تنشيط تعبير IDO1 في خلايا DCs. أ تحليل تطوري لعائلة Erysipelotrichaceae وعزل Clostridium innocuum. ب، ج وفرة نسبية لـ C. innocuum (Clos) في عينات البراز من الأفراد الأصحاء والمرضى الذين يعانون من مرض الأمعاء الالتهابي (IBD) (ب)، وخاصة التهاب القولون التقرحي (UC) ومرض كرون (CD) (ج). مجموعة PRJNA389280 (صحي، ; التهاب الأمعاء، )، مجموعة PRJNA400072 (صحي، ; التهاب الأمعاء، )، مجموعة PRJNA398089 (صحي، ; UC، ; سي دي، ). د مستويات ليس في سوائل الثقافة من دوب (Dub.sup) أو كلوس (Clos.sup) ( ). هـ qPCR من mRNA Ido1 في DCs البشرية المعالجة بـ Lys- أو Clos.sup المستخرجة من خلايا الدم المحيطية الوحيدة النواة بعد 18 ساعة من العلاج ( ). qPCR من mRNA Ido1 بعد 18 ساعة من العلاج في خلايا DC المشتقة من نخاع العظام من الفئران C57BL/6J من النوع البري (WT) المعالجة بـ 8 مللي مولار ليس، IFN ، Dub.sup، Clos.sup أو مرق الكربوهيدرات من اللحم المفروم (CMC) ( ). ز نسبة كينورينين/تريبتوفان في خلايا BMDCs المعالجة بـ Clos.sup، أو غير المعالجة المستخرجة من الفئران WT أو Ido1-/- بعد 18 ساعة من العلاج . ح تعبير Ido1 في خلايا BMDCs الفأرية المعالجة بـ Clos.sup،
CH-223191، أو مجموعة من CH-223191 و Clos.sup ( ). ط تعبير Ido1 في خلايا BMDCs الفأرية التي تم نقلها بـ siNC (تحكم سلبي) أو siAhR (RNA صغير متداخل محدد لـ AhR) وتمت معالجتها بـ Clos.sup بعد 18 ساعة من النقل ( ). ج-م تم استعمار الفئران WT أو Ido1 بـ CFU من Clos أو تمت معالجتها بـ Clos.sup وتعرضت لإدارة DSS. في اليوم السابع بعد إدارة DSS، تم فحص CD25 Foxp3 Tregs ( ) و IL-17 CD4 T cells ( ) في العقد اللمفاوية المساريقية (MLN) وطبقة الغشاء المخاطي القولوني (cLP) (WT، ؛ Ido1 )، طول القولون (1) والتغيرات النسيجية المرضية بواسطة صبغة HE (م) تم فحصها في اليوم السابع بعد علاج DSS . (ن) توضيح تخطيطي لمستقلب D. newyorkensis ليس الذي يحمي من التهاب القولون الناتج عن DSS من خلال إعادة توازن استجابة Treg/Th17 من خلال تنشيط دائرة AhR-IDO1-Kyn. الخطوط المتقطعة عند 1 تشير إلى أن العلاجات لها قيمة متساوية كضوابط طبيعية. النتائج تمثل بيانات تم توليدها في تجربتين مستقلتين على الأقل ويتم التعبير عنها كمتوسط SEM، وتم فحص القيم ذات الجانبين – بواسطة اختبار -Student. تم توفير بيانات المصدر كملف بيانات مصدر.
نحو مسار كينورينين وإعادة توازن استجابات Treg/Th17 لحماية الفئران من التهاب القولون.
أظهرت دراسات متعددة أن العلاج بالمضادات الحيوية يمكن أن يحسن أعراض التهاب القولون، سواء في التجارب البشرية ونماذج الفئران المستحثة بـ DSS ، من خلال تشكيل تركيبة الميكروبيوتا. تم ربط التأثير المفيد على الأعراض السريرية وكبت الالتهاب بزيادة البكتيريا البروبيوتيك مثل Bifidobacteria، . prausnitzii أو Enterobacter ludwigii (E. ludwigii) . في الدراسة الحالية، وجدنا أن علاج Neo يمكن أن يمنع التهاب القولون الناتج عن DSS بشكل أكثر فعالية بين جميع المضادات الحيوية واسعة الطيف الأربعة (Amp، Van، Neo وMetro) (الشكل 1). لا تشبه هذه النتيجة نتائج دراسة حديثة جدًا أجراها Li وآخرون، والتي استخدمت أيضًا علاجات متعددة بمضادات حيوية فردية في نموذج الفأر المستحث بـ DSS، حيث تم الإبلاغ عن أن Metro، ولكن ليس Neo، تسبب في تخفيف التهاب القولون الناتج عن DSS المنسوب إلى توسع E. ludwigii . يمكن تفسير هذه التناقضات من خلال الاختلافات في أنظمة المضادات الحيوية بين الدراستين (جرعة عالية عن طريق الفم في دراستنا مقابل توزيع جرعة منخفضة في الماء في Li وآخرون)، حيث أظهرنا أن هذين النظامين العلاجيين يمكن أن يؤديان إلى اختلاف كبير في القابلية لالتهاب البنكرياس الحاد الناتج عن caerulein بسبب اختلافات كبيرة في تركيبات الميكروبيوتا .
تم الإبلاغ عن أن وفرة Dub المعوية مرتبطة بالسمنة ومرض الزهايمر في نماذج الفئران، ومع ذلك لم يتم إجراء دراسات وظيفية لتأكيد أهميتها البيولوجية في vivo. للمرة الأولى، قمنا بتحليل شامل لدورها المحتمل كبروبيوتيك في كل من نماذج الفئران التقليدية ونماذج الفئران المستنفدة من الميكروبيوتا. يمتلك Dub قدرة بروبيوتيك تفوق البكتيريا المعوية المعروفة Akk في تحسين تلف الحاجز المخاطي القولوني والالتهاب المعوي الناتج عن DSS وTNBS (الشكل 2). تتماشى نتائجنا التي تفيد بأن Dub يخفف الالتهاب المخاطي والالتهاب خارج الأمعاء من خلال إعادة توازن استجابات Treg/Th17 تمامًا مع دراستين بارزتين تظهران أن الأسيتات والبروبيونات تنظم كبت المناعة المعتمد على GPR43 بواسطة cTregs تحت ظروف DSS أو نقل خلايا T .
على الرغم من أن الدور الرئيسي للميكروبيوتا المعوية في تحفيز نشاط IDO1 قد تم إثباته في كل من نماذج الفئران GF وAbx المعالجة ، إلا أن هناك القليل من التركيز على التأثيرات التعديلية على استقلاب تريبتوفان المضيف من خلال مسار كينورينين الذي تمارسه بكتيريا معوية محددة ومستقلبات ميكروبية في IBD. هنا، كشفنا عن دور جديد لـ Dub في تعزيز استقلاب تريبتوفان المضيف من خلال مسار كينورينين لتنظيم استجابات Treg/Th17 تحت ظروف الحالة الثابتة أو الالتهابية من خلال تعزيز تعبير IDO1 في cLPMCs (الشكل 4). علاوة على ذلك، أظهرنا أن ليس المستمد من Dub يميل وظيفة DCs نحو وظيفة تنظيمية وتحمل لتعزيز Tregs من خلال زيادة Kyn، من خلال نشاط IDO1 المعتمد على AhR (الشكل 6). وفقًا لـ Gargaro وآخرين، يحتوي كل من المنطقة العليا والمنطقة غير المشفرة الداخلية من Ido1
تحتوي على عناصر استجابة للمواد الغريبة الكلاسيكية (XREs)، وهي مواقع ربط AhR . بالإضافة إلى ذلك، أظهرنا أن ليس يزيد من SLC7A5 في DCs (الشكل S8h، i)، وهو ناقل أساسي لـ Kyn المدفوع بـ IDO1 في إنشاء دائرة الأيض AhR-IDO1-Kyn في DCs عبر الإشارات الذاتية والإشارات الجانبية . يجب ملاحظة أن هناك اختلافات طفيفة لا تزال موجودة في Treg وTh17 بين مجموعات Ido1 -Dub وIdo1-/-Veh/Lys (الشكل 5f، g)، والتي يمكن تفسيرها بشكل معقول من خلال حقيقة أن Dub قادر أيضًا على إعادة توازن استجابات Treg/Th17 من خلال محور SCFA-GPR43 (الشكل 3h، i). وبالتالي، نحن نحاول إنتاج فئران Gpr43-/-Ido1 ثنائية الطفرة لتأكيد أن هذين العاملين المناعيين ينظمان بشكل تآزري استجابات Treg/Th17 في التهاب القولون الناتج عن DSS.
لم تظهر أي دراسات تقريبًا أن ليس يؤثر على نشاط AhR، سواء بشكل مباشر أو غير مباشر، ولا يزال الرابط الجزيئي بين استقلاب ليس وتنشيط AhR غير مؤكد، على الرغم من أنه من الممكن أن تراكم كروتونيل-CoA الناتج عن استقلاب ليس يعيد برمجة إشارات IFN ، مما قد ينشط بدوره محور AhR-IDO1 . على الرغم من أنه تم مؤخرًا إظهار أن مجموعات DC المختلفة تكتسب مسار IDO1 التحمل وتستجيب لمستقلبات تريبتوفان النشطة مناعياً بشكل مختلف ، هناك حاجة إلى مزيد من الدراسات لاستكشاف الدور التنظيمي لـ ليس في دائرة AhR-IDO1-Kyn في مجموعات DC المحددة. هناك حاجة إلى تجارب مع حذف شرطي محدد لنوع الخلية لـ Ahr أو Ido1 لمعالجة هذه المسألة. أخيرًا، نبذل جهودًا كبيرة لإنتاج سلالة متحورة من Dub مع (ترميز الإيبيمراز الأساسي للديامينوبيميلات من أجل تخليق ليس) نقص، مما سيمكننا من تفكيك الدور المحدد لـ ليس الذي تم تصنيعه بواسطة Dub في تخفيف التهاب القولون الناتج عن DSS.
باختصار، توضح تحليلاتنا الشاملة دورًا غير مكشوف سابقًا لـ Dub في تنشيط دائرة AhR-IDO1-Kyn في DCs، مما حسن استجابة Treg/Th17 غير المتوازنة لتخفيف كل من الالتهاب المخاطي والنظامي في التهاب القولون الناتج عن DSS. قد يوفر الاكتشاف بأن النظير البشري Clos ومستقلبه المرتبط ليس يمكن أن يخفف الالتهاب المخاطي ويحسن الشفاء المخاطي في نموذج الفأر أساسًا لتطوير أساليب علاجية قائمة على الميكروبيوتا لعلاج IBD السريري لدى البشر.
طرق
بيان الأخلاقيات
تم إجراء جميع تجارب الحيوانات بدقة وفقًا للبروتوكول رقم 117113 المعتمد من قبل لجنة رعاية واستخدام الحيوانات المؤسسية (IACUC) في جامعة تشجيانغ. تم جمع الدم من متبرعين أصحاء بموافقة اللجنة الأخلاقية للمستشفى الأول التابع لجامعة غوانغدونغ للصيدلة (رقم التصريح: 20210221). قدم جميع المشاركين موافقة خطية مستنيرة لجمع العينات والتحليلات اللاحقة.
سلالات بكتيرية
تم الحصول على A. muciniphila (Akk) من مجموعة الثقافة الأمريكية (ATCC، رقم BAA835) وزرعها في وسط مغذي من مغذيات القلب والدماغ (BHI؛ Oxoid) عند تحت ظروف لاهوائية. تم شراء D. newyorkensis (Dab) وC. innocuum من مجموعة الميكروبات الألمانية (DSMZ، DSM103457 وDSM1286-1213001) وزرعها في مرق الكربوهيدرات من اللحم المفروم (CMC، Hopebio) عند تحت ظروف لاهوائية. تم شراء E. faecalis (EF) من مركز الصين لمجموعة الثقافة النمطية (CCTCC، AB 2018154) وزرعها في وسط مرق ليوجين (Oxoid) فيتم تقدير تركيز كل نوع من البكتيريا بناءً على الكثافة الضوئية عند.
سلالات الفئران
تم استخدام فئران تتراوح أعمارها بين ستة وثمانية أسابيع ومتطابقة في الجنس من خلفية C57BL/6J في هذه الدراسة. تم تربية وإيواء فئران C57BL/6J خالية من الجراثيم (GF) في شركة Shenzhen Gnotobio Biotechnology Co., Ltd. وتم تأكيد حالة GF من خلال تحليل 16 S qPCR قبل استخدامها في التجارب النسبية، التي أجريت أيضًا في شركة Shenzhen Gnotobio Biotechnology Co., Ltd. تم شراء فئران C57BL/6J من النوع البري (WT) من مركز أبحاث الحيوانات النموذجية بجامعة نانجينغ (نانجينغ، الصين). كانت فئران Foxp3-DTR هدية من الدكتور بين لي من قسم المناعة والميكروبيولوجيا، معهد شنغهاي للمناعة، كلية الطب بجامعة شنغهاي جياو تونغ. تم إعطاء فئران Foxp3-DTR حقنًا داخل الصفاق (i.p.) بجرعة 1 ملغ من DTx (50 نانوغرام DT/كغ من وزن الجسم) مرة واحدة في اليوم لمدة 7 أيام متتالية لضمان استنفاد Tregs بنجاح. تم تحليل خلايا الطحال للتحقق من كفاءة الإزالة لـ Tregs باستخدام تقنية تدفق الخلايا. تم استخدام فئران knockout لإنزيم إندولامين-2،3-دي أوكسيجيناز 1 (Ido1-“) تم توفيرها بلطف من قبل الدكتور ياجينغ وانغ من المختبر الوطني الرئيسي للأدوية الطبيعية، قسم الفسيولوجيا، جامعة الصين للصيدلة. تم شراء فئران knockout لمستقبلات البروتين G المرتبطة (Gpr43–) وفئران ترانسجين IL-17-EGFP من شركة Cyagen Biosciences Inc (سوتشو، الصين). تم الحفاظ على الفئران بشكل روتيني في منشأة خالية من مسببات الأمراض المحددة مع بيئة تتحكم في درجة الحرارة والرطوبة (، الرطوبة)، وتحت دورة ضوء/ظلام ثابتة لمدة 12 ساعة، وتمت إتاحة الوصول الحر إلى نظام غذائي عادي (Gat# P1101F-25، شنغهاي SLACOM) والماء طوال فترة الدراسة في جامعة تشجيانغ. تم تنفيذ جميع الإجراءات وفقًا لبروتوكول معتمد من لجنة رعاية واستخدام الحيوانات في جامعة تشجيانغ، الصين.
علاج المضادات الحيوية، زراعة ميكروبات البراز (FMT)، واستعمار البكتيريا
تم إعطاء الفئران الأمبيسيلين (Amp 10 ملغ)، النيومايسين (Neo 10 ملغ)، الميترونيدازول (Metro 10 ملغ)، أو الفانكومايسين (Van 10 ملغ)، بشكل فردي أو مجتمعة (يشار إليها باسم Abx) يوميًا لمدة 5 أيام عن طريق التغذية الفموية. تم جمع عينات البراز من الفئران التي تم استنفاد الميكروبيوتا في اليوم الخامس بعد العلاج، وتم تجانسها، وزرعها على أجار BHI معدم الأغنام، وزُرع تحت ظروف لاهوائية فيلمدة يومين تليها حضانة تحت ظروف هوائية عندلمدة يوم واحد لتأكيد استنفاد الميكروبات بكفاءة.
للتجارب المتعلقة بزراعة البراز، تم تجانس 200 ملغ من كريات البراز المجمعة من الفئران البرية باستخدام كرات سيليكا معقمة في 1.5 مل من محلول فوسفات البفر (PBS) بسرعة 45 هرتز لمدة دقيقة واحدة وتم تصفيتها باستخدامالمصفاة. تم إجراء التغذية القسرية للفئران البرية المعالجة بالمضادات الحيوية أو الفئران الخالية من الجراثيم.مستخلصات البراز المصفاة (تجارب FMT) أوعدد وحدات تشكيل المستعمرات للبكتيريا المزروعة (تجارب الاستعمار) مرتين بفاصل 48 ساعة. بعد 48 ساعة من زراعة البكتيريا أو زراعة البكتيريا، تم جمع عينات البراز لتحديد كفاءة الاستعمار قبل إعطاء DSS لتحفيز نموذج التهاب القولون.
نموذج الفأر لالتهاب القولون الناتج عن DSS
ما لم يُذكر خلاف ذلك، تم إعطاء الفئران البرية أو بعض الفئران المعدلة وراثيًا 2.5% DSS (رقم الكاتالوج: 60316ES76؛ ياسين، شنغهاي، الصين) في مياه الشرب لمدة 7 أيام لإنشاء نموذج التهاب القولون الناتج عن DSS.
لدراسة تأثير المضادات الحيوية على قابلية الفئران للإصابة بالتهاب القولون الناتج عن DSS، تم معالجة الفئران البرية بمضاد حيوي واحد لمدة 5 أيام، وتم إيقاف إعطاء المضادات الحيوية في اليوم السادس، ثم تعرضت لعلاج DSS. تم وزن الحيوانات المعالجة يوميًا وجُمعت عينات البراز في الأوقات المحددة بعد علاج DSS.
لتقييم تأثير سلالات البكتيريا على التهاب القولون الناتج عن DSS، تم إعطاء الفئران البرية التقليدية عن طريق الفمCFU من D. newyorkensis (Dub) و A. muciniphila (Akk) أو E. faecalis (EF) مرتين بفاصل 48 ساعة، ثم تم تعريضها لعلاج DSS. لتقييم التأثير العلاجي للسلالات الفردية على التهاب القولون التجريبي دون معالجة مسبقة، تم علاج الفئران البرية التقليدية.تم استخدام DSS في مياه الشرب لمدة 3 أيام، ثم تم إعطاؤه عن طريق الفم من اليوم الثالث إلى اليوم الثامن مع CFU من داب، أك، أو EF يوميًا، مع الحفاظ على DSS في مياه الشرب.
لدراسة التأثير الرئيسي لسلالات البكتيريا الفردية على التهاب القولون الناتج عن DSS في سياق استنفاد الميكروبيوتا، تم معالجة الفئران البرية بمضادات حيوية لمدة 5 أيام و…يوم تم تغذيته عن طريق أنبوبCFU من دوب، أك، أو EF مرتين مع فاصل 48 ساعة بين العلاجات قبل إعطاء DSS.
للتجارب المتعلقة بالبسترة (HI)، تم إعطاء الفئران البرية المعالجة بالمضادات الحيوية عن طريق الفم لمدة 5 أيام بـCFU من داب تم تعطيله بواسطة البسترة لمدة 30 دقيقة عندمخفف فيتم إعطاء PBS، وتم إدارة DSS في اليوم السادس.
تمت طرد المحاليل فوق الطافية من ثقافات دوب أو C. innocuum (كلوس) في جهاز الطرد المركزي عندلمدة 10 دقائق عندثم مرت عبر فلاتر بولي إيثر-سولفون (; ميرك ميلبورو) لإزالة الخلايا البكتيرية المتبقية. تم تغذية الفئران البرية المعالجة أو غير المعالجة بالمضادات الحيوية بمستخلصات البكتيريا المناسبة (لكل فأر) لمدة 5 أيام وتم إعطاء DSS في اليوم السادس.
نموذج الفأر لالتهاب القولون الناتج عن TNBS
للت sensibilization، تم حلاقة ظهر الفئران أسفل الرقبة مباشرة وتم دهنها بمحلول 1% TNBS (Meilunbio MB5523) مخلوط في محلول من الأسيتون وزيت الزيتون. ثم تم تحفيز التهاب القولون بـTNBS فيالإيثانول عن طريق الحقن الشرجى بعد 7 أيام من التحسس المسبق كما هو موصوف سابقًا.
دراسة حماية الحيوانات مع المستقلبات
في دراسة حماية SCFA، تم إعطاء الفئران WT و Gpr43-/ 200 مللي مول من البروبيونات في مياه الشرب لمدة 3 أسابيع ثم تم إعطاؤهم DSS. لاختبار تأثير المستقلبات المشتقة من دوب، تم إعطاء الفئران حمض اللاكتيك L (Lac؛ )، كيتوليوسين ( ; ن-أسيتيل-ل-أسب (NAA؛ل-لايس ), L- حمض الأمينوبوتيريك (أبو؛ “ ) يوميًا لمدة 5 أيام قبل علاج DSS. بالنسبة لدراسة حماية Kyn، تم حقن الفئران WT أو Foxp3-DTR عن طريق الحقن داخل الصفاق بـكين كل يومين، 7 جرعات إجمالية مناليوم السابق لإدارة DSS إلى اليوم السادس بعد إدارة DSS. لدراسة حماية Lys، تم استخدام فئران WT، Foxp3-DTR، أو Ido1.تم إعطاء الفئران L-Lys عبر التغذية الفموية مرة واحدة يوميًا لمدة 5 أيام قبل إدارة DSS. بالنسبة لتجارب مضادات AhR، تم علاج الفئران البرية عن طريق الحقن داخل الصفاق بـ CH-223191 (، MedChemExpress) يوميًا لمدة 10 أيام متتالية (من اليوم -7 إلى اليوم 2). خلال هذه الفترة، تم علاج الفئران بـتم إعطاء ليس عن طريق التغذية الفموية لمدة 5 أيام (من اليوم -5 إلى اليوم 0) ثم تم إعطاء DSS.
تقدير بكتيريا البراز
تم قياس بكتيريا البراز باستخدام تقنية qPCR (تم إدراج البرايمرات في جدول الموارد الأساسية). تم عزل الحمض النووي البكتيري من البراز باستخدام مجموعة TIANamp Stool DNA Kit (TIANGEN)، وتم إجراء qPCR باستخدام SYBR Green Real-time PCR Master Mix (TOYOBO).
استخراج الحمض النووي، تسلسل الأمبليكون 16S rDNA، وتحليل البيانات
عينات برازتم إعادة تعليقها في محلول ASL من كياجن وتم تجانسها لمدة دقيقتين. تم استخراج الحمض النووي البرازي الكلي من الطور العلوي الناتج باستخدام مجموعة QIAamp DNA Stool Mini Kit (كياجن). تم قياس تركيز و نقاء الحمض النووي باستخدام جهاز Qubit (Thermo Fisher Scientific). ثم تم تضخيم الحمض النووي من البراز باستخدام Phusion High-Fidelity PCR Master Mix (New England Biolabs) من خلال تفاعل البوليميراز المتسلسل (PCR) الذي يستهدف المناطق المتغيرة 3 و 4 (V3-V4) من 16S rDNA. تم إجراء تسلسل متعدد للمنتجات باستخدام رموز شريطية خاصة بكل عينة باستخدام منصة MiSeq Illumina (Guangdong Magigene Biotechnology Co., Ltd.). تم دمج القراءات المزدوجة الأطراف في تسلسلات طويلة باستخدام FLASH v.1.2.7، وهي أداة تحليل سريعة ودقيقة مصممة لدمج القراءات المزدوجة الأطراف عندما تكون هناك تداخلات بين القراءات1 و القراءات2. ثم تم تحليل التسلسلات المدمجة باستخدام حزمة البرمجيات QIIME v.1.9.1.
تفاعل البوليميراز المتسلسل العكسي الكمي
تم استخراج RNA الكلي من عينات الأنسجة الممزوجة بالخرز أو زراعة الخلايا باستخدام كاشف TRIzol (Invitrogen) وفقًا لتعليمات الشركة المصنعة. تم إجراء تفاعلات PCR باستخدام مجموعة HiScript II One Step qRT-PCR SYBR Green Kit (Vazyme) على نظام Gentier 96R RealTime PCR (TIANLONG، شيآن، الصين). تم تطبيع مستويات النسخ للجينات المحددة إلى التحكم الداخلي GAPDH لكل عينة فردية باستخدام البرايمرات المدرجة في جدول الموارد الرئيسية وتم قياسها باستخدام دورة العتبة الحرجة المقارنة.طريقة.
تحليلات تعبير السيتوكينات
IL-6 و TNF-تم قياس مستويات البروتين في عينات المصل وهوموجينات القولون باستخدام مجموعات الفحص المناعي المرتبط بالإنزيم (ELISA) المقابلة (70-EK201B/3-96، 70-EK206/3-96، 70-EK282/4-96 MultiSciences) وفقًا لتعليمات الشركة المصنعة.
تحليل البقعة الغربية
تم تحلل الأنسجة والخلايا المعالجة كما هو موضح باستخدام محلول التحلل RIPA (بيوتايم) وتم إخضاعها لـتم إجراء الرحلان الكهربائي للهلام SDS-البولي أكريلاميد ثم تم نقله إلى أغشية بولي فينيليدين فلوريد (Millipore). تم تحضين البروتينات بعد ذلك مع الأجسام المضادة الأولية المحددة ثم مع الأجسام المضادة الثانوية المرتبطة بالبيروكسيداز من الفجل. تم تصور شرائط البروتين باستخدام مجموعة كيمياء مضيئة معززة (Vazyme) مع نظام تصوير هلام ChemiDoc Touch (Bio-Rad).
علم الأنسجة النسيجية، التلوين المناعي، والتصوير الحي
لفائف سويسرية منقوعة فيتم تجفيف محلول البارافورمالدهيد، وتضمينه في الشمع، وقطعه إلىأقسام سميكة، وتم صبغها بالهيماتوكسيلين والإيوزين (HE) باستخدام الإجراءات القياسية. تم تقييم الشرائح بواسطة طبيب أمراض ذو خبرة بطريقة عمياء، وتم تقييم الدرجات النسيجية بناءً على المعايير التالية وفقًا للأبحاث السابقة: الالتهاب، عيوب الظهارة، ضمور الغدد، التغيرات التنسجية/الأورام، والمساحة المتأثرة بالتغيرات التنسجية.لتلوين خلايا الكأس، تم تلوين مقاطع القولون أيضًا بصبغة الألكيان الأزرق لـومجفف فيالكحول والزيلين.
لتحليل المناعة الفلورية (IFA)، تم حجب مقاطع القولون المنزوع البارافين بـ مصل الماعز العادي لمدة 30 دقيقة في درجة حرارة الغرفة (RT). ثم تم تحضين الشرائح مع أجسام مضادة أولية محددة في لمدة 12 ساعة. الأجسام المضادة المستخدمة: مضاد IL-6 (1:50، 21865-1-AP بروتين تك)، مضاد F4/80 (1:50، 28463-1-AP بروتين تك)، مضاد أوكلودين (1:100، بروتين تك 13409-1-AP) ومضاد ميوك2 (1:100، 27675-1-AP بروتين تك). ثم تم تحضين الشرائح مع أجسام مضادة ثانوية موسومة بالفلور (جاكسون إيمونوريسيرش). تم صبغ النوى باستخدام DAPI (روش، سويسرا). تم إجراء صبغة TUNEL باستخدام مجموعة الكشف عن موت الخلايا في الموقع POD (روش داياغنوستكس) على جهاز Discovery XT وفقًا لبروتوكول الشركة المصنعة. تم إجراء جميع التحليلات باستخدام برنامج ImageJ.
تم استعماره الفئران المعدلة وراثيًا IL-17-EGFP بـ CFU داب، المعالج بـكين أوليس وخضع لـنظام دعم القرار تمت الإدارية كما هو موصوف أعلاه. في اليوم السابع بعد إعطاء DSS، تم تشريح القولون من فئران IL-17-EGFP المتحولة جينياً في كل مجموعة. تم إجراء التصوير الحي باستخدام PerkinElmer (CLS136341/ F) وتم التقاط صور تمثيلية.
تحليل المجهر الإلكتروني الناقل
أتم غسل قطعة من أنسجة القولون الطازجة بمحلول PBS وثُبّتت فيجلوتارالدهيد فيلمدة 4 ساعات. بعد الشطف في PBS، تم تثبيت الأنسجة بشكل إضافي في PBS يحتوي على 1% من رباعي أكسيد الأوزميوم لمدة ساعتين في درجة حرارة الغرفة، ثم تم شطفها في PBS وتجفيفها. بعد ذلك، تم تضمين الأنسجة في Epon 812 طوال الليل، ثم تم معالجتها في فرن عندلمدة 48 ساعة. تم قطع مقاطع بسمك 80 نانومتر باستخدام ميكروتوم فائق (RMC MTX) بواسطة سكين ماسية. تم إيداع المقاطع على شبكات ذات ثقب واحد مغطاة بالفورمفار والكربون وتم صبغها مرتين في محاليل مائية منأسيتات اليورانيوم لمدة 25 دقيقة عندورصاصات الرصاص لمدة 3 دقائق في درجة حرارة الغرفة. تم التقاط الصور باستخدام مجهر إلكتروني نافذ H-7650 (هيتاشي، إباراكي، اليابان) عند جهد تسريع 80 كف. تم تصوير مناطق الاهتمام في الأغشية المتغيرة بواسطة كاميرا CCD Gatan 830 (جاتان، كاليفورنيا، الولايات المتحدة الأمريكية).
عزل خلايا الظهارة المعوية القولونية وخلايا أحادية النواة في الطبقة الخاصة
تم الحصول على خلايا الأمعاء الظهارية الفأرية وخلايا المناعة الموجودة في الغشاء المخاطي من القولون كما تم وصفه سابقًاباختصار، تم فتح القولون طوليًا وقطعه إلى قطع. بعد الحضانة مع EDTA (5.5 مليمول) وDTT (1 مليمول) في محلول ملح هانك المتوازن (HBSS)، تم الخلط باستخدام جهاز الخلط وتمريره عبرمنخل الخلايا، تم غسل تعليق IECs مرتين بواسطة الطرد المركزي فيلمدة دقيقتين وتم جمعها للتجارب المستقبلية. تم تحضين نسيج الغشاء المخاطي المتبقي مع محلول الهضم الذي يحتوي على الكولاجيناز ( ) و DNase تم تعريض تعليق خلايا LPMC الناتج لفصل باستخدام تدرج بيركول وجمعه لإجراء تجارب إضافية.
تدفق الخلايا
لتلوين سطح الخلايا، تم تحضين تعليقات الخلايا المفردة على الثلج لمدة 30 دقيقة مع الأجسام المضادة التالية: مضاد CD3 المترافق مع FITC (11-0032-82، eBioscience، 1:150)، مضاد CD4 المترافق مع eFluor 450 (48-0041-82، eBioscience، 1:150)، ومضاد CD25 المترافق مع APC (17-025182، eBioscience، 1:150).
لتحليل خلايا Treg بعد صبغ الخلايا باستخدام الأجسام المضادة CD3 وCD4 وCD25، تم تثبيت وتعقيم تعليق اللمفاويات باستخدام مجموعات عازلة لعوامل النسخ (562574، BD Pharmingen) وفقًا لتعليمات الشركة المصنعة وتم صبغها باستخدام anti-Foxp3-PE (12-5773-82، eBioscience، 1:75). لتحليل خلايا Th1 وTh17، تم تحفيز اللمفاويات النسيجية المعزولة لمدة 5 ساعات باستخدام خليط تحفيز الخلايا بالإضافة إلى مثبطات نقل البروتين (00-4975-93، eBioscience). بعد الحضانة لمدة 5 ساعات، تم غسل الخلايا في PBS وصبغها لاكتشاف موت الخلايا باستخدام صبغة حيوية قابلة للتثبيت 570 (564995، BD Horizon، 1:100)، FITC-anti-CD3، وeFluor 450-anti-CD4. تم تثبيت الخلايا المصبوغة في عازلة التثبيت (00-8222-49، eBioscience)، وتم تعقيمها باستخدام عازلة غسل التثبيت الداخلي (00-8333-56، eBioscience)، وصبغها باستخدام anti-IL-17A المرتبط بـ PerCPCyanine5.5 (45-7177-82، eBioscience، 1:75) وphycoerythrin المرتبط بـ anti-IFN- (505808، بايوليجند، 1:75).
عزل خلايا بون مار من الفئران BMDMs و BMDCs
تم عزل البلعميات المشتقة من نخاع العظام من الفئران (mBMDMs) من نخاع العظام المستخرج من عظام الفخذ والساق للفئران كما هو موصوف سابقًا.تم زراعة الخلايا في و في DMEM المضاف إليه FBS و السائل العلوي لخلايا L929 المصفاة،البنسلينستربتوميسين في و لمدة 7 أيام قبل الإجراء التجريبي. تم الحصول على خلايا دندريتية مشتقة من نخاع العظم الفأري (mBMDCs) من خلايا نخاع العظم وتم تحفيزها بـ GM-CSF (سيرفيس بيو) وIL-4 (PeproTech) لمدة 6 أيام.من mBMDMs أو mBMDCs كانت بعد ذلك مزروعة في لوحة 24 بئرًا ومعالجة بحمض اللاكتيك L (100 مللي مول) وKyn (0.2 مللي مول)نAA (1 مليمول)، L-Lys (8 مليمول)، أبو (0.1 مليمول) لاكتشاف Ido1 بعد 18 ساعة من العلاج بواسطة qRT-PCR.
فرز خلايا الدندريت البشرية
تم تجنيد الأفراد الأصحاء الذين تم إدخالهم إلى المستشفى لإجراء فحص صحي كمتبرعين أصحاء في الدراسة. تم جمع عينات من الدم المحيطي من سبعة أفراد أصحاء (3 ذكور و4 إناث) تتراوح أعمارهم بين 22 و41 عامًا وتم استخراج خلايا الدم البيضاء أحادية النواة (PBMCs) بواسطة الطرد المركزي بتدرج الكثافة باستخدام مجموعات عزل PBMC البشرية (LTS10771 TBD) وفقًا لبروتوكول الشركة المصنعة. بعد العزل، تم إثراء الخلايا الشجرية (DCs) من PBMCs باستخدام EasySep.مجموعة إثراء خلايا دندريتية المايلويد البشرية (19061 ستيمسيل)، وتم صبغ خلايا دندريتية المثرية باستخدام BV421 مضاد CD11c البشري (301628 بايو ليجند، لكل عينة). تم استخدام خلايا DCs المعززة للاختبارات اللاحقة. تم مراجعة البروتوكول والموافقة عليه من قبل لجنة المراجعة الأخلاقية في المستشفى الأول التابع لجامعة قوانغدونغ للصيدلة، وتم الحصول على موافقة خطية مستنيرة من كل مشارك.
زراعة الأورغانويد
تم زراعة الأعضاء العضوية المعوية والقولونية الفأرية كما هو موصوف سابقًاباختصار، تم عدّ تجاويف الاثني عشر وتضمينها في من مادة ماتريجيل (كورنينج) بتركيز 10,000 كريبت/مل وزُرعت في مجموعة الأعضاء العضوية القولونية للفئران (K2204-MC، بايوجينوس). تم قياس مساحة سطح الأعضاء العضوية باستخدام برنامج إيميج جي (المعاهد الوطنية للصحة). تم عزل الكريبتات القولونية وزراعتها كما هو موضح سابقًا.تم عد الكروم في القولون وتضمينها في ماتريجيل وزراعتها في مجموعة الأعضاء المعوية للفئران (K2001-MI، bioGenous). في اليوم الرابع (للاثني عشر) أو اليوم الثالث (للقولون)، تم تحفيز الأعضاء العضوية بـ L-Lys لمدة 18 ساعة قبل استخراج RNA واكتشاف qRT-PCR.
علاج الأعضاء العضوية، خلايا الدم البيضاء المستمدة من نخاع العظم، وخلايا البلعمة المستمدة من نخاع العظم باستخدام سوائل الثقافة البكتيرية
تمت طرد المحاليل الفائقة البكتيرية في جهاز الطرد المركزي عندلمدة 10 دقائق عندثم مرت عبر فلاتر بولي إيثر-سولفون (; ميرك ميلبورو). تم تحضين الأعضاء العضوية، وخلايا الدم البيضاء المستمدة من نخاع العظم، وخلايا البلعمة المستمدة من نخاع العظم مع سوائل الثقافة البكتيرية (8%) المخففة في وسط زراعة الخلايا المناسب لمدة 18 ساعة في .
تحديد نشاط IDO1
تم قياس نشاط IDO1 في المختبر كقدرته على تحويل التربتوفان إلى L-Kyn.تم إضافة L-Trp إلى BMDCs بالاشتراك مع 8 مللي مول من L-Lys أوإنترفيرون- وتم حضنه في تم جمع مستخلصات زراعة الخلايا بعد 18 ساعة وتم قياس تركيزات L-Kyn و L-Trp باستخدام LC-MS/MS.
إسكات وتثبيط AhR في خلايا الدم الجذعية المستمدة من نخاع العظم
تم إضافة ليبوفكتامين 2000 (إنفيتروجين 11668019) إلىتم خلط Opti-MEM (Gibco 31985070) بلطف، وتركه لمدة 5 دقائق. مجموعة siRNA محددة لـتم خلطه وتركه لمدة 20 دقيقة. تم زراعة خلايا BMDCs في أطباق 24 بئر وتم تحضينها مع الخليط لمدة 10 دقائق عند درجة حرارة الغرفة.تم إضافة من DMEM الكامل (Pricella PM150210). بعد 24 ساعة، تم استخدام خلايا BMDCs المعدلة وراثيًا للاختبارات اللاحقة. بالنسبة لتجارب التثبيط، تم معالجة خلايا BMDCs مسبقًا بـCH-223191 (مضاد AhR) لمدة 24 ساعة واستخدمت للاختبارات اللاحقة.
كشف انتقال AhR
تم تعريض خلايا mBMDCs لـ 8 مللي مولار من ليسين أو 300 نانومولار من FICZ لمدة 6 ساعات. تم تثبيت الخلايا باستخدام 4% من البارافورمالدهيد في PBS لمدة 15 دقيقة في درجة حرارة الغرفة، تلاها خطوة اختراق باستخدام PBS يحتوي على 0.5% من Triton X-100 لمدة 20 دقيقة في درجة حرارة الغرفة. بعد ذلك، تم إجراء تفاعل Click عن طريق إضافةمن خليط التفاعل (Tublin 1:50، AF2835؛ AhR 1:100، AF6165 بييوتيم) وتم حضنه لمدة 30 دقيقة في درجة حرارة الغرفة. تم صبغ النوى بـ
DAPI. تم إجراء التحليل باستخدام مجهر المسح بالليزر المجهري (LSM880) مع عدسة تكبير 60x مغمورة بالزيت. تم الكشف عن التوبولين باستخدام الليزر، تم الكشف عن AhR باستخدامالليزر، وتم الكشف عن DAPI باستخدامليزر. تم جمع الصور باستخدام برنامج زين.
تم عزل البروتين من فئتين خلويتين مختلفتين باستخدام مجموعة استخراج السيتوبلازم والنواة (R0050 Solarbio)، وفقًا لتعليمات الشركة المصنعة. بعد ذلك، تم إخضاع العينات لتحليل الويسترن بلوت كما هو موضح أعلاه. تم استخدام التوبولين الخاص بالسيتوبلازم والهستون الخاص بالنواة كضوابط للتفريق المناسب. الأجسام المضادة المستخدمة: مضاد التوبولين (1:50، AF2835 Beyotime)، مضاد الهستون (1:50، AF0009 Beyotime) ومضاد AHR (1:100، AF6165 Beyotime).
تحليل النسخ الجيني
تم معالجة خلايا BMDCs مسبقًا بـ 8 مللي مول من Lys لمدة 6 أو 18 أو 24 ساعةإنترفيرون-لمدة 18 ساعة، ثم تم جمع العينات لاستخراج RNA الكلي باستخدام كاشف TRIzol (Invitrogen). تم تقييم العينات في نفس الوقت على نظام Agilent 4200 (Agilent Technologies)، Qubit 3.0 (Thermo Fisher Scientific)، وNanodrop One (Thermo Fisher Scientific). تم إنشاء مكتبات RNA-Seq وتسلسلها بواسطة Guangdong Magigene Biotechnology. تم بناء عينات ثلاثية من جميع الاختبارات في مكتبة مستقلة، وتم تنفيذ التسلسل والتحليل التالي: تم إنشاء مكتبات RNA-Seq الكاملة باستخدام مجموعة تحضير مكتبة RNA غير الاتجاهية Next Ultra لـ Illumina (New England Biolabs) وفقًا لتوصيات الشركة المصنعة. تم تنفيذ تجميع العينات المشفرة بالفهرس على نظام توليد التجمع cBot. بعد توليد التجمع، تم تسلسل المكتبات على منصة Illumina NovaSeq 6000، وتم إنتاج قراءات مزدوجة بطول 150 قاعدة. تم معالجة البيانات الخام بتنسيق fastq بواسطة Trimmomatic (الإصدار 0.36) للحصول على قراءات نظيفة، والتي تم تعيينها إلى قواعد بيانات NCBI Rfam لإزالة تسلسلات rDNA باستخدام Bowtie2 (الإصدار 2.33). تم تعيين تسلسلات mRNA المتبقية إلى الجينوم المرجعي بواسطة Hisat2 (2.1.0). تم استخدام HTSeq-count (الإصدار 0.9.1) للحصول على عدد القراءات ومعلومات الوظيفة لكل جين وفقًا لنتائج التعيين. تم تحديد الجينات المعبر عنها بشكل مختلف في حالتين/مجموعتين باستخدام edgeR (الإصدار 3.16.5)، وتم تنفيذ تحليل GO باستخدام clusterProfiler (الإصدار 3.4.4)، حيث تم تصحيح انحياز طول الجين.
الميتabolوميات شبه المستهدفة
تم استخراج المستقلبات من مستحلبات القولون أو عينات SUP البكتيرية وإرسالها إلى BGI (شنتشن، الصين) لإجراء تحليلات الميتابولوميات المستهدفة. تم استخدام LC-MS/MS عالي الأداء لإجراء قياس دقيق وعالي الحساسية وذو تغطية واسعة وذو إنتاجية عالية لـ 350 مستقلبًا مستهدفًا في هذه العينات. بالتفصيل، تم إعادة تعليق كميات مناسبة من كريات الخلايا التجريبية أو كريات التحكم في الجودة (QC) فيمنمحلول الماء/الميثانول، تم تحليله، وتم الطرد المركزي، وتم نقل السائل العلوي إلى أنبوب جديد. تم إعداد منحنى قياسي عن طريق التخفيف المتسلسل لمعيار مختلط HM350. تم إخضاع العينة التجريبية، وعينة مراقبة الجودة، والمعيار لتفاعل تحويل، وتم تخفيف المركبات الناتجة في مخفف HM350 وتم الطرد المركزي عندفيلمدة 10 دقائق. تم تطبيق السائل العلوي على تحليل LC-MS/MS باستخدام جهاز LC-MS QTRAP. (SCIEX). تم إجراء الكروماتوغرافيا على عمود BEH C18 (تم إجراء قياس الطيف الكتلي باستخدام مصدر ESI+/ESI-؛ وتمت إجراء تحليلات إحصائية وحيوية إضافية بواسطة BGI باستخدام إجراءات روتينية.
تحديد تركيزات التربتوفان والكينيورين باستخدام تقنية LC-MS/MS
مصل ) أو تم إعادة تعليق سوائل الثقافة BMDC في من الأسيتونيتريل/الميثانول (8:2) وتم الطرد المركزي عندلمدة 20 دقيقة. ثم تم تجفيف السائل العلوي باستخدام منفاخ نيتروجين. تم إعادة تكوين الرواسب فيمن الماء/أسيتونيتريل (8:2) يحتوي علىحمض الفورميك عن طريق الخلط الجيد والدوران. أخيرًا، الـ SUP (تم حقنه في نظام LC-MS/MS لـ تم استخدام نظام UHPLC-MS/MS (ExionLC AD UHPLC-QTRAP 6500+، شركة AB SCIEX) لتحديد كمية التربتوفان (Trp) والكينيورينين (Kyn) في نوفوجين. تم استخدام LCMS/MS للكشف عن تركيز سلسلة من المحاليل القياسية. تم استخدام تركيز المحلول القياسي كالمتغير المستقل، وتم استخدام نسبة مساحة قمة المعيار الداخلي كالمتغير التابع للتحقق من خطية المحلول القياسي.
تحليل GC-MS/MS لتركيزات الأحماض الدهنية قصيرة السلسلة
تم وضع 20 ملغ من كل عينة براز في أنبوب EP سعة 2 مل، وتم إضافة 1 مل من حمض الفوسفوريك ) تم إضافة محلول وكرة فولاذية صغيرة. تم طحن العينات بشكل موحد، ثم تم خلطها في جهاز دوار لمدة 10 دقائق وتم استخدام الموجات فوق الصوتية لـتم جمع SUP بعد أن تم طرد الخليط مركزياً عندلمدة 10 دقائق عندتم إضافة محلول MTBE (الذي يحتوي على معيار داخلي)، وتم خلط المزيج باستخدام جهاز الخلط الدوار لمدة 3 دقائق وتم استخدام الموجات فوق الصوتية لمدة 5 دقائق. بعد ذلك، تم طرد المزيج باستخدام جهاز الطرد المركزي عندلمدة 10 دقائق عندثم تم جمع الـ SUP واستخدامه لتحليل GC-MS/MS.
لتحليل الأحماض الدهنية قصيرة السلسلة باستخدام GC-MS/MS، تم استخدام جهاز كروماتوغرافيا الغاز Agilent 7890B متصل بمطياف الكتلة 7000D مع عمود DB-FFAP (طول 30 م)هويةتم استخدام سمك الفيلم، J&W Scientific، الولايات المتحدة الأمريكية). تم استخدام الهيليوم كغاز حامل، بمعدل تدفق قدرهتمت الحقن في وضع الانقسام بحجم حقن قدرهتم الحفاظ على درجة حرارة الفرن عندلمدة دقيقة واحدة، ثم ارتفع إلىبمعدلثم ارتفع إلىبمعدل، تم الاحتفاظ به لمدة 36 ثانية، تم رفعه إلىبمعدلدقيقة، تم الاحتفاظ بها لمدة 0.5 دقيقة وتشغيلها لمدة 3 دقائق. تم تحليل جميع العينات في وضع المراقبة المتعددة التفاعلات؛ كانت درجات حرارة مدخل الحقن وخط النقل و على التوالي. تم الكشف عن محتوى الأحماض الدهنية قصيرة السلسلة بواسطة MetWare (http://www.metware.cn/) استنادًا إلى منصة GC-MS/MS Agilent 7890B-7000D.
الإحصائيات وإمكانية التكرار
تم إجراء التحليلات الإحصائية باستخدام برنامج Prism GraphPad الإصدار 8.0. تمثل أشرطة الخطأ الخطأ المعياري للمتوسطات في جميع الأشكال، و-تم تحديد القيم بواسطة اختبار الطالب ذو الاتجاهين-اختبارات أو تحليل التباين (ANOVA).تم تقديرها لتحليل الارتباط بين متغيرين مستمرين. تم استخدام اختبار ذو جانبين-قيمةاعتُبر ذا دلالة إحصائية. تم تكرار جميع التجارب، مع ذكر عدد النسخ في أساطير الأشكال. كانت الصور التمثيلية لبلوتات ويسترن من على الأقل 3 تحضيرات عينات مستقلة.
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقرير Nature Portfolio المرتبط بهذه المقالة.
توفر البيانات
بيانات تسلسل جين 16 S rRNA متاحة في أرشيف قراءة التسلسل (SRA) تحت رقم مشروع BioProject PRJNA1013230. بيانات RNA-seq متاحة في SRA تحت رقم مشروع BioProject PRJNA1012836. تم إيداع بيانات الميتابولوم لمستخلصات القولون الفأرية ووسط زراعة الخلايا البكتيرية المبلغ عنها في هذه الدراسة في قاعدة بيانات NGDC OMIX (معرف OMIX: OMIX005711؛ OMIX005712). جميع البيانات الأخرى التي تدعم استنتاجات هذه الدراسة متاحة في الورقة والمواد التكميلية. تم توفير بيانات المصدر مع هذه الورقة.
References
Wang, G. et al. Bridging intestinal immunity and gut microbiota by metabolites. Cell. Mol. Life Sci. 76, 3917-3937 (2019).
Nicolas, G. R. & Chang, P. V. Deciphering the chemical lexicon of host-gut microbiota interactions. Trends Pharmacol. Sci. 40, 430-445 (2019).
Skelly, A. N., Sato, Y., Kearney, S. & Honda, K. Mining the microbiota for microbial and metabolite-based immunotherapies. Nat. Rev. Immunol. 19, 305-323 (2019).
Rooks, M. G. & Garrett, W. S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 16, 341-352 (2016).
Subramanian, B. C. Inflammatory bowel disease: DCs sense LTB(4) to drive and differentiation. Cell. Mol. Immunol. 17, 307-309 (2020).
Tanoue, T., Atarashi, K. & Honda, K. Development and maintenance of intestinal regulatory T cells. Nat. Rev. Immunol. 16, 295-309 (2016).
Ramanan, D. et al. Regulatory T cells in the face of the intestinal microbiota. Nat. Rev. Immunol. 23, 749-762 (2023).
Smith, P. M. et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 341, 569-573 (2013).
Furusawa, Y. et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446-450 (2013).
Sharma, M. D. et al. Inhibition of the BTK-IDO-mTOR axis promotes differentiation of monocyte-lineage dendritic cells and enhances anti-tumor T cell immunity. Immunity 54, 2354-2371.e8 (2021).
Puccetti, P. & Grohmann, U. IDO and regulatory T cells: a role for reverse signalling and non-canonical NF-kappaB activation. Nat. Rev. Immunol. 7, 817-823 (2007).
Fallarino, F. et al. The combined effects of tryptophan starvation and tryptophan catabolites down-regulate T cell receptor zetachain and induce a regulatory phenotype in naive T cells. J. Immunol. 176, 6752-6761 (2006).
Mezrich, J. D. et al. An interaction between kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J. Immunol. 185, 3190-3198 (2010).
Di Meglio, P. et al. Activation of the aryl hydrocarbon receptor dampens the severity of inflammatory skin conditions. Immunity 40, 989-1001 (2014).
Monteleone, I. et al. Aryl hydrocarbon receptor-induced signals upregulate IL-22 production and inhibit inflammation in the gastrointestinal tract. Gastroenterology 141, 237-248 (2011).
Ala, M. Tryptophan metabolites modulate inflammatory bowel disease and colorectal cancer by affecting immune system. Int. Rev. Immunol. 41, 326-345 (2022).
Schirmer, M., Garner, A., Vlamakis, H. & Xavier, R. J. Microbial genes and pathways in inflammatory bowel disease. Nat. Rev. Microbiol. 17, 497-511 (2019).
Engevik, M. A. et al. Bifidobacterium dentium-derived y-glutamylcysteine suppresses ER-mediated goblet cell stress and reduces TNBS-driven colonic inflammation. Gut Microbes 13, 1-21 (2021).
Liu, Y. et al. TLR4 regulates RORgammat(+) regulatory T-cell responses and susceptibility to colon inflammation through interaction with Akkermansia muciniphila. Microbiome 10, 98 (2022).
Cox, L. M. et al. Corrigendum: description of two novel members of the family Erysipelotrichaceae: Ileibacterium valens gen. nov., sp. nov. and Dubosiella newyorkensis, gen. nov., sp. nov., from the murine intestine, and emendation to the description of Faecalibacterium rodentium. Int. J. Syst. Evol. Microbiol. 67, 4289 (2017).
Rodrigues, V. F. et al. Akkermansia muciniphila and gut immune system: a good friendship that attenuates inflammatory bowel disease, obesity, and diabetes. Front. Immunol. 13, 934695 (2022).
Kuczma, M. P. et al. Commensal epitopes drive differentiation of colonic T(regs). Sci. Adv. 6, eaaz3186 (2020).
Zagato, E. et al. Endogenous murine microbiota member Faecalibaculum rodentium and its human homologue protect from intestinal tumour growth. Nat. Microbiol. 5, 511-524 (2020).
Le Poul, E. et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 278, 25481-25489 (2003).
Maslowski, K. M. et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 461, 1282-1286 (2009).
Cervenka, I., Agudelo, L. Z. & Ruas, J. L. Kynurenines: tryptophan’s metabolites in exercise, inflammation, and mental health. Science 357, eaaf9794 (2017).
Mellor, A. L. & Munn, D. H. IDO expression by dendritic cells: tolerance and tryptophan catabolism. Nat. Rev. Immunol. 4, 762-774 (2004).
Gargaro, M. et al. Indoleamine 2,3-dioxygenase 1 activation in mature cDC1 promotes tolerogenic education of inflammatory cDC2 via metabolic communication. Immunity 55, 1032-1050.e14 (2022).
Yuan, H. et al. Lysine catabolism reprograms tumour immunity through histone crotonylation. Nature 617, 818-826 (2023).
Yang, L. et al. Amino acid metabolism in immune cells: essential regulators of the effector functions, and promising opportunities to enhance cancer immunotherapy. J. Hematol. Oncol. 16, 59 (2023).
Liu, C. et al. Baicalein restores the balance of Th17/Treg cells via aryl hydrocarbon receptor to attenuate colitis. Mediat. Inflamm. 2020, 5918587 (2020).
Zhuang, Z. et al. GWAS-associated bacteria and their metabolites appear to be causally related to the development of inflammatory bowel disease. Eur. J. Clin. Nutr. 76, 1024-1030 (2022).
Franzosa, E. A. et al. Gut microbiome structure and metabolic activity in inflammatory bowel disease. Nat. Microbiol. 4, 293-305 (2019).
Schirmer, M. et al. Dynamics of metatranscription in the inflammatory bowel disease gut microbiome. Nat. Microbiol. 3, 337-346 (2018).
Zhang, Y. et al. Discovery of bioactive microbial gene products in inflammatory bowel disease. Nature 606, 754-760 (2022).
Michaudel, C. & Sokol, H. The gut microbiota at the service of immunometabolism. Cell Metab. 32, 514-523 (2020).
Maccaferri, S. et al. Rifaximin modulates the colonic microbiota of patients with Crohn’s disease: an in vitro approach using a continuous culture colonic model system. J. Antimicrob. Chemother. 65, 2556-2565 (2010).
Abraham, B. & Quigley, E. M. M. Antibiotics and probiotics in inflammatory bowel disease: when to use them? Frontline Gastroenterol. 11, 62-69 (2020).
Li, Q. et al. Enterobacter ludwigii protects DSS-induced colitis through choline-mediated immune tolerance. Cell Rep. 40, 111308 (2022).
Rath, H. C. et al. Different subsets of enteric bacteria induce and perpetuate experimental colitis in rats and mice. Infect. Immun. 69, 2277-2285 (2001).
Li, H. et al. Bifidobacterium spp. and their metabolite lactate protect against acute pancreatitis via inhibition of pancreatic and systemic inflammatory responses. Gut Microbes 14, 2127456 (2022).
Rodriguez-Daza, M. C. et al. Berry polyphenols and fibers modulate distinct microbial metabolic functions and gut microbiota enterotype-like clustering in obese mice. Front. Microbiol. 11, 2032 (2020).
Sun, P. et al. Comparative metagenomics and metabolomes reveals abnormal metabolism activity is associated with gut microbiota in Alzheimer’s disease mice. Int. J. Mol. Sci. 23, 11560 (2022).
Agus, A., Planchais, J. & Sokol, H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe 23, 716-724 (2018).
Bacsi, S. G. & Hankinson, O. Functional characterization of DNAbinding domains of the subunits of the heterodimeric aryl hydrocarbon receptor complex imputing novel and canonical basic helix-loop-helix protein-DNA interactions. J. Biol. Chem. 271, 8843-8850 (1996).
Sinclair, L. V., Neyens, D., Ramsay, G., Taylor, P. M. & Cantrell, D. A. Single cell analysis of kynurenine and system L amino acid transport in T cells. Nat. Commun. 9, 1981 (2018).
Rothhammer, V. et al. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat. Med. 22, 586-597 (2016).
Sanmarco, L. M. et al. Identification of environmental factors that promote intestinal inflammation. Nature 611, 801-809 (2022).
Allen, I. C. et al. NLRP12 suppresses colon inflammation and tumorigenesis through the negative regulation of noncanonical NFkappaB signaling. Immunity 36, 742-754 (2012).
Weigmann, B. et al. Isolation and subsequent analysis of murine lamina propria mononuclear cells from colonic tissue. Nat. Protoc. 2, 2307-2311 (2007).
Steed, A. L. et al. The microbial metabolite desaminotyrosine protects from influenza through type I interferon. Science 357, 498-502 (2017).
Matsuzawa-Ishimoto, Y. et al. Autophagy protein ATG16L1 prevents necroptosis in the intestinal epithelium. J. Exp. Med. 214, 3687-3705 (2017).
Yui, S. et al. Functional engraftment of colon epithelium expanded in vitro from a single adult Lgr5(+) stem cell. Nat. Med. 18, 618-623 (2012).
الشكر والتقدير
نحن نقدر الدكتور ياجينغ وانغ من جامعة الصين للصيدلة والدكتور بين لي من كلية الطب بجامعة شنجهاي جياو تونغ لتقديمهم قيمة وأيضًا فئران Foxp3-DTR. نشكر الدكتور لينغدونغ شو من مركز الحيوانات المخبرية بجامعة تشجيانغ على الدعم الفني الأساسي في إجراء المجهر الضوئي التداخلي. تم استخدام خدمة التحرير المهنية NB Revisions للإعداد الفني للنص قبل التقديم. هذا العمل مدعوم بمنح من المؤسسة الوطنية للعلوم الطبيعية في الصين (رقم 32172864 ورقم U21A2O261) وخطة البحث والتطوير الوطنية الرئيسية في الصين (2022YFD1800804) إلى S.J.Z.
مساهمات المؤلفين
صمم S.J.Z. وY.Z. التجارب. قام Y.Z. وS.T. وX.J. وJ.W. وJ.G. وX.S. وS.S. وG.W. وJ.Q. وZB.Z. وH.C. وZY.Z. بتنفيذ التجارب. أجرى Y.Z. وJ.M. تحليل المعلومات الحيوية. جمع S.S. وX.S. العينات والبيانات. نسق S.C. المشروع. علق L.Z. على مسودات المخطوطة وراجعها. كتب S.J.Z. وY.Z. وS.T. الورقة. أشرف S.J.Z. على البحث والتنسيق والاستراتيجية.
قسم الطب البيطري، كلية علوم الحيوان، جامعة تشجيانغ، هانغتشو، تشجيانغ 310058، جمهورية الصين الشعبية.قسم أمراض القلب، المستشفى الأول التابع لجامعة غوانغدونغ للصيدلة، غوانغتشو، غوانغدونغ 510080، جمهورية الصين الشعبية.قسم طب الرعاية الحرجة، مستشفى سير ران ران شاو، كلية الطب بجامعة تشجيانغ، هانغتشو، تشجيانغ 310016، جمهورية الصين الشعبية.كلية الطب البيطري، جامعة تشينغداو الزراعية، تشينغداو، شاندونغ 266109، جمهورية الصين الشعبية.مستشفى الشعب في شاوكسينغ، مستشفى جامعة تشجيانغ شاوكسينغ، شاوكسينغ، تشجيانغ 312000، جمهورية الصين الشعبية.كلية الطب البيطري، مركز أبحاث الهندسة الإقليمي في جيلين للبروبيوتيك الحيوانية، المختبر الرئيسي الإقليمي لجيلين للميكروبيولوجيا الحيوانية والتربية الصحية، جامعة جيلين الزراعية، تشانغتشون 130118، جمهورية الصين الشعبية.البريد الإلكتروني: shuzhu@zju.edu.cn
Commensal bacteria generate immensely diverse active metabolites to maintain gut homeostasis, however their fundamental role in establishing an immunotolerogenic microenvironment in the intestinal tract remains obscure. Here, we demonstrate that an understudied murine commensal bacterium, Dubosiella newyorkensis, and its human homologue Clostridium innocuum, have a probiotic immunomodulatory effect on dextran sulfate sodium-induced colitis using conventional, antibiotic-treated and germ-free mouse models. We identify an important role for the D. newyorkensis in rebalancing Treg/Th17 responses and ameliorating mucosal barrier injury by producing short-chain fatty acids, especially propionate and L-Lysine (Lys). We further show that Lys induces the immune tolerance ability of dendritic cells (DCs) by enhancing Trp catabolism towards the kynurenine (Kyn) pathway through activation of the metabolic enzyme indoleamine-2,3-dioxygenase 1 (IDO1) in an aryl hydrocarbon receptor (AhR)-dependent manner. This study identifies a previously unrecognized metabolic communication by which Lysproducing commensal bacteria exert their immunoregulatory capacity to establish a Treg-mediated immunosuppressive microenvironment by activating AhR-IDO1-Kyn metabolic circuitry in DCs. This metabolic circuit represents a potential therapeutic target for the treatment of inflammatory bowel diseases.
The human gastrointestinal tract harbors an enormous, diverse, and critical population of microorganisms (termed ‘microbiota’) that interacts intimately with its human host to form an interdependent and mutually restrained ecosystem, performing metabolic functions such as carbohydrate fermentation and vitamin biosynthesis . The gut
microbiota generates immensely diverse bioactive small-molecule metabolites that can be involved in signaling, mucosal barrier maintenance, or immune system modulation . Given such a broad range of effects on host physiology and immunology, it remains unclear how specific microbes (and the small molecules they produce) interact to
cause, sustain, mitigate, or predict gut-related diseases such as inflammatory bowel disease (IBD), ulcerative colitis (UC) and Crohn’s disease (CD).
Excessive activation of Th1/Th17 and impairment of Foxp3 regulatory T (Treg) cells have been described in the pathogenesis of IBD . Treg-mediated immunosuppression via secretion of inhibitory cytokines of TGF- and IL-10 is of great importance for inhibition of IBD . They also preserve intestinal physiology by promoting epithelial barrier functions and tissue repair. Although specific intestinal microbes and related metabolites have been confirmed to be involved in the induction of Treg-mediated immune tolerance , the underlying molecular mechanisms are largely unknown.
The development of intestinal Tregs is affected by several metabolites derived from diet. Short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate are the main products of the fermentation of undigested carbohydrate in the gut, particularly in the colon. SCFAs induce accumulation of colonic Tregs (cTregs) through multiple mechanisms . They can directly stimulate Treg proliferation through activation of GPR43 (also known as FFAR2) and the differentiation of naïve cells into Treg cells through histone H3 acetylation of Foxp3 by histone deacetylase (HDAC) inhibition .
Tryptophan (Trp) is an essential, aromatic amino acid that humans must acquire from diet. Dietary Trp can be metabolized to kynurenine (Kyn) by indoleamine-2,3-dioxygenase 1 (IDO1) activity in intestinal epithelial cells (IECs) and dendritic cells (DCs) and contributes to the differentiation of naïve cells into Tregs through activation of the aryl hydrocarbon receptor , a ligandactivated transcription factor and immune sensor that has been implicated in IBD pathogenesis . Although it has been demonstrated that the intestinal microbiome is important for Trp catabolism and AhR activation during IBD , the specific commensal species and the microbial metabolites that participate in activation of the AhR-IDO1Kyn metabolic circuitry and their impact in establishing a Tregmediated tolerogenic mucosal microenvironment has not been fully explored.
In this study, we identified an understudied, robust SCFAproducing commensal bacterium, Dubosiella newyorkensis, that can ameliorate dextran sulfate sodium (DSS)-induced colitis in mice by rebalancing Treg/Th17 responses and improving mucosal barrier integrity. We revealed that D. newyorkensis and its human homolog Clostridium innocuum (C. innocuum) generate L-Lys to modulate Tregmediated immunosuppression by activating AhR-IDO1-Kyn circuits in mouse and human DCs. Collectively, these results point to a multifactorial strategy used by D. newyorkensis and C. innocuum to maintain mouse and human gut immune homeostasis, respectively.
Results
Manipulation of the gut microbiota by single-antibiotic treatment restricts DSS-induced colitis
Wild-type C57BL/6J mice (WT) were given ampicillin (Amp), vancomycin (Van), neomycin (Neo), or metronidazole (Metro) either individually or as a cocktail containing all 4 (Abx) via oral gavage and subsequently exposed to DSS (Fig. 1a). Perturbation of the gut microbiota by both Abx and all 4 single-antibiotic treatments was confirmed as a significant reduction in 16 S rDNA copies (Supplementary Fig. 1a). Neo treatment exerted the best effect in attenuating DSSinduced colitis, manifested by the least body weight loss (Supplementary Fig. 1b), longer colon (Fig. 1b, c) as well as lower colonic mRNA levels of proinflammatory cytokines (Fig. 1d) and IL-6 protein (Fig. 1e).
Next, we prepared fecal samples pooled from Neo mice and performed fecal microbiota transplantation (FMT) in microbiota-depleted Abx mice (Fig. 1a). As expected, FMT significantly restored the total fecal 16S rDNA copy number prior to DSS treatment, but not to the same level of vehicle (Veh) control (Supplementary Fig. 1c). Intriguingly, weight loss (Supplementary Fig. 1d), colon
shortening (Fig. 1c, f), histopathological lesions (Fig. 1g, h) and infiltration of IL-6-secreting macrophages (Fig. 1h) were greatly alleviated in the , with results comparable to Neo group. These data suggest that the fecal microbiota of Neo mice may indeed contain specific intestinal microbes that can confer protection against DSSinduced colitis.
We carried out 16S rRNA gene sequencing and analysis on fecal samples collected from Veh, Abx, and single-antibiotic-treated mice at Day 0 (D0) and 7 (D7) post-DSS administration and found remarkable differences in microbiome composition based on antibiotic treatment, with or without DSS induction (Fig. 1i). There was a significantly increased relative abundance of Bifidobacterium, Bacteroides, Akkermansia and Dubosiella at D7, higher in the Neo-treated than other single-antibiotic-treated, and Abx mice (Fig. 1j, k).
Colonization with D. newyorkensis mitigates DSS-induced colitis by restoring mucosal barrier function and inhibiting inflammatory responses
Among the bacterial taxa enriched by Neo treatment, Bifidobacterium, Bacteroides, and Akkermansia have already been reported to play protective roles during IBD , thus we focused on the Dubosiella genus (located within the family Erysipelotrichaceae) for the present study. We chose D. newyorkensis (abbreviated as Dub hereafter) isolated from a laboratory mouse as the representative isolate to determine the role of genus Dubosiella in DSS-induced colitis. Antibiotic susceptibility testing of Dub confirmed its resistant to Neo and Metro but susceptibility to Amp and Van (Supplementary Fig. 2a).
Groups of WT mice were gavaged with Dub twice and then administrated DSS for colitis induction (Supplementary Fig. 2b). Since Akkermansia muciniphila (abbreviated as Akk hereafter) was also enriched after Neo treatment (Fig. 1j, k) and has been documented to be protective during IBD , it was chosen as a probiotic control in the study. Enterococcus faecalis (abbreviated as EF hereafter), which is a major cause of nosocomial infection, served as an unrelated Grampositive commensal bacterial control. Indeed, Dub colonization exerted an even greater protective effect against DSS-induced colitis than Akk, with lower body weight loss (Supplementary Fig. 2c), longer colon length (Fig. 2a and Supplementary Fig. 2d), lower histopathological score (Fig. 2b, c) and less colonic macrophage infiltration (Fig. 2c). By contrast, EF colonization resulted in a colitic phenotype comparable to Veh controls (Fig. 2a-d and Supplementary Fig. 2c, d).
Furthermore, colonization with Dub led to better protection of the colonic mucosal barrier, as reflected by reduced apoptosis (Supplementary Fig. 2e), greater area of occludin-positive cells (Supplementary Fig. 2f), greater number of mucin-secreting goblet cells in the colonic epithelia and less disruption to the fine structure of the brush border and tight junctions (Fig. 2c). We also confirmed the differences of occludin, ZO-1 and Muc2 expression in the colonic intestinal epithelial cells (cIECs) (Fig. 2c, d and Supplementary Fig. 2g), while also showing that the expression of proinflammatory cytokines IL-1 , IL-6 and tumor necrosis factor- (TNF- ) was significantly lower in the WT-Dub vs the WT-EF or WT-Veh, both in the colonic lamina propria (cLP) (Supplementary Fig. 2h) and peripheral blood (Supplementary Fig. 2i).
Additionally, WT mice gavaged with Dub for 5 consecutive days starting from day 3 post-DSS administration (Supplementary Fig. 2j) exhibited longer colon length (Supplementary Fig. 2k, l) and improved histopathology (Supplementary Fig. 2m, n) compared with mice receiving EF or untreated controls, suggesting that Dub also has therapeutic effects on DSS-induced colitis.
To further evaluate the probiotic effect of Dub on colitis, we used the mouse model of 2,4,6-trinitrobenzene sulfonic acid (TNBS)-induced colitis (Supplementary Fig. 3a). In this model, Dub colonization alleviated intestinal injury as indicated by longer colon (Supplementary Fig. 3b, c) and reduced histopathological lesions
Fig. 1 | Manipulation of gut microbiota affects mouse susceptibility to DSSinduced colitis. a Outline of treatment regimens. b-e Wild-type C57BL/6J mice (WT, ) received vehicle (Veh) or oral antibiotics (single Amp, Van, Neo, or Metro or in combination [Abx], ) daily for 5 days and treated with DSS in drinking water for 7 days. At day 7 (D7) post-DSS treatment, colon length (b, c), relative mRNA expression of proinflammatory cytokines IIIb, II6, and Tnfa by qRTPCR (d) and IL-6 protein level by ELISA (e) were determined in each group. Abx mice were given fecal microbiota transplantation (FMT) from Neo-treated mice twice [Abx-FMT(N)] or Veh with a 48 h interval and then treated with DSS for 7 days . WT B6 mice treated with Neo (Neo) were served as control. Colon length (f) was measured, histopathological changes were scored by HE staining ( ) and
infiltration of IL-6-secreting macrophages (Mø) was determined by IFA (h). Principal coordinate analysis (PCoA) (i) and heatmap of the relative abundance (j) of bacteria in fecal samples from mice treated with Abx or single antibiotics at D0 and D7 post-DSS administration. k Relative abundance of Dubosiella and Akkemansia at D0 and D7 among groups treated with different antibiotics (D7. Amp, D7. Van, ; D0. Veh, D0. Abx, D0. Metro, D0. Amp. D0. Van, ; D0. Neo, D7. Veh, D7. Neo, D7. Abx, D7. Metro, ). Dashed lines at 1 indicate that the treatments have equal value as normalized controls. Results are representative of data generated in at least two independent experiments and are expressed as mean SEM, and 2 -sided -values were examined by the Student’s -test. Source data are provided as a Source Data file.
(Supplementary Fig. 3d, e). Dub colonization also increased the expression of OCLN, ZO-1, and Muc2 in the clECs (Supplementary Fig. 3f), and decreased the expression of Illb, Il6, and Tnfa in the cLP (Supplementary Fig. 3g), in agreement with a role for Dub in protection of the mucosal barrier and prevention of intestinal inflammation.
To remove the effect of background microbes and link the probiotic effect directly to Dub colonization, Abx mice were gavaged with Dub (Abx-Dub), Akk (Abx-Akk) or EF (Abx-EF) and subsequently
administrated with DSS (Supplementary Fig. 3h). All 3 bacterial isolates were able to repopulate the gut microbiota in Abx mice (Supplementary Fig. 3i). We observed less body weight loss (Supplementary Fig. 3k), longer colon length (Fig. 2e and Supplementary Fig. 3l), lower histopathological score (Fig. 2f, g) and less macrophage infiltration (Fig. 2g) in the colon of Abx-Dub compared with Abx-Akk, Abx-EF or Abx groups. These parameters were equivalent between Abx-Dub and Neo groups (Fig. 2e-g and Supplementary Fig. 3l). Similar to the Neo
Fig. 2 | D. newyorkensis protects mice from DSS-induced colitis and attenuates resulting mucosal inflammation and barrier damages. a-d Conventional wildtype C57BL/6J mice were colonized with CFU of D. newyorkensis (Dub), . muciniphila (Akk), E. faecalis (EF) or vehicle (Veh) twice with 2-day break in between, then treated with DSS as previously described ( ). At D7 post-DSS administration, colon samples were collected to determine colon length (a), histopathological score (b) by HE staining, IL-6-secreting macrophage infiltration, Muc2 and Occludin expression by IFA, number of goblet cells by Alcian blue staining and microstructure of colonic epithelia by TEM (c). The expression of Occludin and ZO-1 in each group was determined by western blot (WB) (d). e-i Abxtreated WT mice were colonized with CFU Dub, Akk, EF, or Veh and exposed to DSS . At D7 post-DSS treatment, colon length (e), histopathological score by HE staining ( ), IL-6-secreting macrophage infiltration ( ) were examined.
Meanwhile, the expression of ZO-1,OCLN, and Muc2 in isolated colonic intestinal epithelial cells ( ), and the expression of proinflammatory cytokines Illb, Il6, and Tnfa in colonic lamina propria (cLP) (i) was detected by qRT-PCR. -I Conventional WT mice ( ) were orally administered heat-inactivated ( HI ) Dub, or Dub supernatant (Dub.sup) and exposed to DSS. At D7 post-DSS administration, colon length (j) was measured, histopathological changes were scored by HE staining ( ), level of tight junction proteins Occludin and ZO-1, and expression of proinflammatory cytokines IL-1 , IL-6, and TNF- were determined by WB (I). Dashed lines at 1 indicate that the treatments have equal value as normalized controls. Results are representative of data generated in at least two independent experiments and are expressed as the mean SEM, and 2 -sided -values were examined by the Student’s -test. Source data are provided as a Source Data file.
controls, Abx-Dub had markedly increased expression of ZO-1, OCLN and Muc2 in the clECs (Fig. 2h) and significantly reduced of Illb, Il6, and Tnfa in the cLP (Fig. 2i) compared with Abx-Akk, Abx-EF, Abx or Veh groups.
In order to better define the protective component of Dub, Abx mice were gavaged with filtered Dub supernatant (Dub.sup) or heatinactivated (HI) Dub and then subjected to DSS treatment (Supplementary Fig. 3h). Interestingly, pretreatment with Dub.sup, but not HI Dub, ameliorated DSS-induced weight loss (Supplementary Fig. 3j) and disease phenotypes (Fig. 2j, k and Supplementary Fig. 3l), restored mucosal barrier functions (Fig. 2c, 1), and inhibited inflammatory cytokine expression in the colon (Fig. 21), even though the pasteurized Dub cells were detected in the feces of Abx mice before DSS administration (Supplementary Fig. 3i). Taken together, these results demonstrate a likely functional role for Dub-derived metabolites rather than direct activity of bacterial components in the attenuation of DSS-induced colitis.
Dub prevents DSS-induced immunopathology by rebalancing Foxp3 Treg and IL17 CD4 T cell responses in the gut microenvironment
Using RNA-Seq analyses, we detected the downregulation of proinflammatory pathways, including those associated with leukocyte extravasation, NF-kB activation, and IL-17 signaling pathways, in colons from Dub mice compared with Veh controls at D7 post-DSS treatment (Fig. 3a and Supplementary Fig. 4a). Consistently, downregulation of proinflammatory II17 and Ifng and upregulation of anti-inflammatory II10 and in bulk cLP immune cells were observed in Dub or Akk, but not in EF groups (Supplementary Fig. 4b).
Given that Tregs play a central role in maintaining the immunosuppressive microenvironment in the gut via TGF- and IL-10 secretion and an imbalance of Treg and Th1/Th17 responses is involved in the pathogenesis of IBD , we hypothesized that Dub colonization inhibits DSS-induced inflammation by inducing Foxp3 Tregs and suppressing IFN cells and IL17 cells. To test this, we colonized groups of Abx mice with Dub, administered DSS, and measured specific T cell subpopulations in the spleen, mesenteric lymph nodes (MLNs), and cLP by flow cytometry at D7 (Supplementary Figs. 2b and 3h). Respective gating strategies are presented in Supplementary Fig. 5. Since Akk can induce both colonic and peripheral Tregs in colitis , we included Akk as a positive control and EF as a negative control. Dub or Akk colonization significantly increased CD25 Foxp3 Tregs and markedly decreased IL17 CD4 T cells in the MLNs and cLP (Fig. 3b, c), but only mildly reduced IFN cells in the cLP (Supplementary Fig. 4c) compared to EF or Veh controls, with or without gut microbiota at D7. A similar pattern was observed in the spleen, where Foxp3 Tregs were significantly higher and IL17 CD4 T cells were significantly lower in Dub or Abx-Dub groups compared with noncolonized controls at D7 (Supplementary Fig. 4d), indicating that Dub rebalanced Treg/Th17 response not only in the colon, but also in the peripheral blood in DSS-induced colitis. Notably,
Dub was superior to Akk in rebalancing Treg/Th17 response with or without an intact gut microbiota (Fig. 3b, c and Supplementary Fig. 4b-d).
Additionally, we gavaged conventional or Abx mice with Dub in the absence of DSS (Supplementary Fig. 4e) and found that CD25 Foxp3 Tregs were also increased in the spleen and cLP, while IL17 CD4 T cells were decreased in the MLNs and cLP without DSS induction (Supplementary Fig. 4f, g), implying that Dub contributes to the balance of Treg/Th17 response even under intestinal homeostasis.
Finally, WT or diphtheria toxin (DT)-treated Foxp3-DTR mice were gavaged with Dub or EF before DSS administration (Supplementary Fig. 4h). As expected, DT treatment severely compromised CD25 Foxp3 Tregs in the spleen, MLNs and cLP in Foxp3-DTR mice (Supplementary Fig. 4i). Treg depletion in Foxp3-DTR mice reversed the differences observed between Veh-treated (WT-Veh) and Dubcolonized WT mice (WT-Dub) in disease phenotypes (Fig. 3d and Supplementary Fig. 4j), histological evaluation (Fig. 3e, f) and proinflammatory cytokine expression (Supplementary Fig. 4k), indicating that Foxp3 Tregs are essential for the Dub protective effect against DSS-induced colitis. Interestingly, small differences still remained in the expression of ZO-1,OCLN, and Muc2 between Veh-treated and Dubcolonized Foxp3-DTR mice (Supplementary Fig. 41), suggesting that Dub might be able to promote mucosal healing independent of inducing Treg-mediated immunosuppression.
Colonization with Dub prevents DSS-induced colitis by regulating Foxp Tregs in part through the SCFA-GPR43 signaling axis
Given that Dub is a close genetic relative of Faecalibaculum rodentium , which is an efficient SCFA-producer that has been reported to be capable of protecting from intestinal tumor growth , we hypothesized that the protective phenotype of Dub might be due to robust SCFA production. Indeed, there were significantly higher concentrations of acetate, propionate (Prop), and butyrate in the fecal samples of Dub mice compared with EF groups or the Veh controls and the concentrations of Prop and butyrate were comparable between Dub- and Akk-colonized mice (Supplementary Fig. 6a). Consistent with this data, propionate and butyrate concentrations in Dub.sup were 10.9-fold and 2-fold higher than in Akk.sup, respectively. However, the concentration of acetate was 1.5-fold higher in Akk.sup than in Dub.sup (Fig. 3g).
Since propionate has the highest affinity for GPR43 , which is required for the expansion and suppressive function of colonic Tregs (cTregs) in the DSS-induced injury , we expected that the differences in Foxp3 Treg induction between WT-Dub and WT-Veh mice might be reproduced by Prop administration and both phenotypes might be lost in Gpr43-/- mice. Thus, we gavaged WT and Gpr43-/- mice with Dub twice, or supplemented with Prop in drinking water for 3 weeks, then administrated with DSS and assessed the frequencies of CD25 Foxp3 Tregs and IL17 CD4 T cells in the spleen, MLNs and cLP (Supplementary Fig. 6b). Indeed, Prop administration led to its
Fig. 3 | D. newyorkensis prevents DSS-induced colitis by regulating Foxp3 Tregs partially through the SCFA-GPR43 signaling axis. a Heat map showing mRNA expression determined by bulk RNA-Seq in colon samples from vehicle (Veh)-treated ( ) or D. newyorkensis (Dub)-colonized wild-type C57BL/6J mice ( ) at D7 post-DSS treatment. b, c CD25+Foxp3+Tregs (b) and IL cells (c) in mesenteric lymph nodes (MLN) and colonic lamina propria (cLP) of conventional or Abx-treated WT mice colonized with Dub, A. muciniphila (Akk), or E. faecalis (EF) ( ) were examined by flow cytometry at D7 post-DSS treatment. d-f Conventional WT or Foxp3-DTR mice ( ) were colonized with CFU of Dub, Akk, or EF and subjected to DSS treatment. At D7 post-DSS administration, colon length measurement (d) and histopathological evaluation (e, f) were performed. g SCFA concentration in Dub, Akk, or EF culture supernatant determined by GC/MS . h-m Conventional WT and Gpr43-/- mice ( ) were
colonized with Dub ( CFU) twice or administrated propionate (Prop) in drinking water for 3 weeks before DSS exposure. At D7 post-DSS administration, CD25 Foxp3 Tregs ( ) and IL-17 T cells (i) in MLNs and cLP of conventional WT or Gpr43-/ mice ( ) were quantified by flow cytometry. Meanwhile, colon length (j), histopathological score by HE staining (k, l) ( ), and expression of ZO1, OCLN, and Muc2 in isolated colonic intestinal epithelial cells (cIECs) (WT, ; Gpr43-/, ) ( ) was determined. Fold of change in frequencies of CD25 Foxp3 Tregs or IL-17 CD4 T cells ( ), colon length ( ), or SCFA concentrations (g) between groups was calculated and presented with numbers in red. Dashed lines at 1 indicate that the treatments have equal value as normalized controls. Results are representative of data generated in at least two independent experiments and are expressed as mean SEM, and 2-sided -values were examined by the Student’s -test. Source data are provided as a Source Data file.
significantly elevated concentration in the feces (Supplementary Fig. 6c) and exerted comparable effects in upregulating CD25 Foxp3 Tregs and downregulating IL17 CD4 T cells in all 3 tissues compared with Dub colonization in WT mice. Unexpectedly, small differences in the frequencies of the 2 T cell subsets remained between Gpr43-/-Dub and Gpr43-/-Veh groups, whereas no differences were observed between Gpr43-/-Prop or Gpr43 -Veh groups (Fig. 3h, i and Supplementary Fig. 6d), indicating a full involvement for GPR43 in the propionate-mediated Treg/Th17 rebalancing in the context of DSSinduced colitis.
Consistent with the results observed in Treg/Th17 regulation, a similar protective effect was exerted by WT-Prop compared with WTDub mice with respect to disease phenotype (Fig. 3j and Supplementary Fig. 6e, f), histological changes (Fig. 3k, I and Supplementary Fig. 6g), mucosal barrier integrity (Fig. 3m and Supplementary Fig. ) and inflammation (Supplementary Fig. 6j), while the protective phenotype was impaired in Gpr43 mice. However, despite a narrowing of the phenotype, there were still differences between the Gpr43-/-Dub and Gpr43-/-Veh groups, suggesting that in addition to SCFAs (especially propionate), other Dub-derived metabolites or GPR43-independent pathways might be involved in the Dub-driven induction of cTregs and amelioration of DSS-induced colitis.
Dub induces CD25+Foxp3+ Tregs via enhancement of IDO1mediated Trp metabolism in colonic dendritic cells
To search for other Dub-derived metabolites that might promote CD25 Foxp3 Treg induction, we performed untargeted metabolomic analysis on colon samples collected from conventional WT mice colonized with Dub, Akk, EF or Veh controls at D7, and a total of 253 compounds were detected. Partial least-squares discriminant analysis showed remarkably different metabolic profiles for Dub compared with the Veh controls (Supplementary Fig. 7a). Among the top 35 most abundant differential metabolites listed in the heatmap, 9 of them were statistically significantly differentiated between Dub-colonized mice and noncolonized control animals (Fig. 4a). KEGG analyses suggested that Trp metabolism was significantly lower in the DSS-induced group at D7 (Supplementary Fig. 7b), while colonization with Dub resulted in greater L-Kyn and quinolinic acid compared with the Akk, EF or Veh mice although the difference was not statistically significant (Fig. 4a).
To confirm the potential role of Kyn in rebalancing Treg/Th17 responses in intestinal inflammation settings, conventional WT and Foxp3-DTR mice were subjected to Kyn treatment prior to and throughout DSS administration (Supplementary Fig. 7c). Consistent with a previous publication , Kyn treatment resulted in a significantly higher level of CD25 Foxp3 Tregs and a drastically lower level of CD4 T cells in the spleen, MLNs and cLP at D7 in WT mice (Fig. 4b, c and Supplementary Fig. 7d). Moreover, disease phenotypes (Fig. 4d and Supplementary Fig. 7e), histopathological changes (Fig. 4e, f) or inflammatory responses (Supplementary Fig. 7f) were the same in Foxp3-DTR mice, despite significant serum Kyn concentrations detected in both mouse lines at D7 (Supplementary Fig. 7g). Therefore, Dub colonization promotes the production of the Trp catabolite Kyn, which induces CD25 Foxp3 Tregs to exert a protective effect on DSSinduced colitis.
The majority of food-derived Trp is metabolized via the Kyn pathway through IDO1 in mucosal and immune cells or IDO2 and Trp 2,3-dioxygenase (TDO) in the liver (Fig. 4g). Thus, we measured the kinetics of IDO1 expression in the colon of WT-Dub or WT-Veh at different times post-DSS treatment (Supplementary Fig. 7c) and determined that IDO1 was markedly elevated at D1, D3, and D5, but not on D7 in WT-Dub compared with WT-Veh controls (Fig. 4h and Supplementary Fig. 7h, i). In agreement with these results, Dub colonization led to a significantly increased serum Kyn concentration (Fig. 4i) at early phases of colitis progression (D0-D3).
To identify the cellular sources of IDO1 upregulation responding to Dub-derived metabolites, we tested Ido1 expression in Abx-Dub and Abx-Dub.sup groups (Supplementary Fig. 7c) and demonstrated significantly increased Ido1 expression in colonic lamina propria mononuclear cells (cLPMCs), but not in clECs prior to and at D3 post-DSS administration (Fig. 4j). Moreover, adding Dub.sup to WT bone marrow-derived dendritic cells (BMDCs) resulted in significantly augmented Ido1 expression compared to culture medium control, whereas EF.sup had no such enhancing effect (Fig. 4k); there was a similar trend for WT bone marrow-derived macrophages (BMDMs) under the treatment of Dub.sup although it was not statistically significant (Fig. 4k). On the contrary, Ido1 expression was not altered in colonic organoids and slightly elevated in duodenal organoid at the presence of Dub.sup (Fig. 4k). These findings suggest that Dub-derived metabolites preferentially promote IDO1 expression in intestinal mononuclear phagocytes.
To exclude the possibility that the phenotypes were influenced by residual Abx-resistant commensal bacteria in our Abx group, germfree (GF) mice were colonized with Dub (GF-Dub) or received FMT from conventional WT mice (GF-FMT) before DSS treatment (Supplementary Fig. 7j). Consistently, GF-Dub also had markedly higher Ido1 levels in cLPMCs (Fig. 4I) and serum Kyn/Trp ratio (Fig. 4m) compared with GF-FMT. Correspondingly, GF-Dub displayed markedly higher CD25 Foxp3 Tregs and remarkably lower levels of IL17 CD4 T cells in the spleen, MLNs and cLP than GF-FMT (Fig. 4n, o and Supplementary Fig. 7k). Also, GF-Dub exhibited mitigated disease phenotype (Fig. 4p and Supplementary Fig. 7l), histological lesions (Fig. 4q, r) and inflammatory responses (Fig. 4s) compared with GF-FMT. Interestingly, Dub monocolonization resulted in higher ZO-1, comparable OCLN, and even lower Muc2 level than in GF-FMT mice (Supplementary Fig. 7m). Altogether, we demonstrated that Trp metabolism in DCs was promoted by Dub, and the resulting Kyn enhancement from strengthened IDO1 expression rebalanced Treg/Th17 responses and ultimately exerted a protective effect on DSS-induced colitis.
The Dub-derived metabolite L-Lys upregulates AhR-dependent IDO1 expression in colonic DCs
To investigate the bacterial metabolites mediating IDO1 upregulation, we performed untargeted metabolome analyses on Dub.sup after culture in chopped meat carbohydrate medium (CMC) (Fig. 5a). Liquid chromatography-tandem mass spectrometry (LC-MS/MS) analysis yielded 5 Dub-derived metabolites, N-acetyl-L-Asp (NAA), L-Lysine (Lys), L-Lactic acid (Lac), L- -aminobutyric acid (Abu) and ketoleucine (4-methyl-2-oxopentanoic acid, 4-MOV) with relatively higher concentrations in both positive and negative ion-exchange chromatography (Supplementary Fig. 8a). These metabolites were also upregulated in the colon of Dub-colonized mice (Fig. 4a).
After pretreatment of WT mBMDCs with these 5 metabolites, we discovered that Lys and 4-MOV induced significantly higher Ido1 (Fig. 5b). IFN- , a well-studied IDO1 stimulator in DCs, was used as a positive control . In agreement with Gargaro et al. , we also determined that exogenous Kyn treatment of mBMDCs increased Ido1 expression (Fig. 5b). Consistently, administration of Lys or 4-MOV to the WT mice (Supplementary Fig. 8b) significantly increased IDO1 expression in the colon at D3 (Fig. 5c). Among the 5 metabolites, Lys and Lac remarkably ameliorated colon shortening (Fig. 5d), tight junction damage and intestinal inflammation (Supplementary Fig. 8c), although the protective phenotype was not as robust overall compared with Dub.sup treatment.
Because 4-MOV showed no protective effect against DSS-induced colitis, we chose to focus on the potential role of Lys in modulating IDO1-driven Trp metabolism. Indeed, Lys pretreatment markedly elevated Ido1 expression in WT mBMDCs in a dose- and time-dependent manner (Supplementary Fig. 8d, e). Furthermore, the addition of both Lys and Trp together to WT mBMDCs resulted in a significant decrease
Fig. 4 | D. newyorkensis promotes CD25 Foxp3 Tregs through enhancement of IDO1-mediated Trp metabolism in dendritic cells. a Heat map showing differential metabolites in colon at D7 post-DSS treatment from wild-type C57BL/6J mice (WT) treated with vehicle (Veh, ) or colonized with D. newyorkensis (Dub), A. muciniphila (Akk), or E. faecalis (EF) ( ). Corresponding statistical annotations of differential metabolites represent the distinctions between Dub-colonized and noncolonized mice. b, c T cell subsets in mesenteric lymph nodes (MLN) and colonic lamina propria (cLP) from Veh- or Kyn-treated ( ) conventional WT mice were examined by flow . d-f WT or DT-treated Foxp3-DTR mice were treated with Kyn and subjected to DSS administration for 7 days, colon length (d) and histopathology (e,f) were examined. g Schematic of the Kyn catabolic pathway of Trp metabolism. h, i Kinetics of colonic IDO1 expression (h) and Kyn serum concentration (i) in Dub- or Veh-colonized conventional WT mice ( ) at indicated times post-DSS administration. j IDO1 expression in colonic intestinal epithelial cells (cIECs) or lamina propria mononuclear cells (cLPMCs) extracted
from Abx-Dub, Abx-EF, Abx-Veh, or Abx-Dub.sup mice ( ) at D3 post-DSS administration. Ido1 expression in murine bone marrow-derived macrophages (mBMDMs) and bone marrow-derived dendritic cells (mBMDCs) or intestinal organoids treated with Dub or EF culture supernatants for 18 h . 1-s Germ-free (GF) mice receiving fecal microbiota transplantation (FMT, ) from conventional WT mice or colonized with Dub ( ) were exposed to DSS treatment for 7 days. Ido1 expression in cLPMCs ( ), serum concentration ratio of cell subsets ( ) ( ) in MLN and cLP, colon length ( ), pathological changes ( ) (FMT, ; Dub, ) and proinflammatory cytokines in cLPMCs ( ) ( ) were determined. Fold of change in frequencies of T cell subsets ( ) or colon length (d) between groups was calculated and presented with numbers in red. Dashed lines at 1 indicate that the treatments have equal value as normalized controls. Results are representative of data generated in at least two independent experiments and are expressed as mean SEM, and 2 -sided -values were examined by the Student’s -test. Source data are provided as a Source Data file.
of Trp and remarkable increase of Kyn in the cellular supernatant (Supplementary Fig. 8f). This result was correlated with a more robust Ido1 expression (Supplementary Fig. 8g) as shown by an increased Kyn/ Trp ratio, which was lost in similarly treated mBMDCs collected from Ido1 mice (Fig. 5e), indicating that the Kyn pathway was enhanced by Lys in an IDO1-dependent manner.
Extracellular Lys is transported through SLC7A1, SLC7A2, and SLC7A3 , while Trp is transported by system L, a heterodimer
composed of a heavy chain (encoded by Slc3a2) and a catalytic L chain LAT1 (encoded by Slc7a5) . Increased expression of Slc7a1, Slc7a2, Slc7a5, and Slc3a2 was observed in mBMDCs collected from both conventional and GF mice and treated with Lys, compared with untreated cells (Supplementary Fig. 8h, i), while Slc7a3 was not detected (data not shown). Thus, Lys-treated mBMDCs may upregulate SLC7A5 and SLC3A2 to increase Trp uptake, which fuels the Kyn pathway of Trp catabolism.
Fig. 5 | L-Lys enhances Trp metabolism in dendritic cells to induce Tregdependent immunosuppression by promoting IDO1 expression. a Heat map showing differentially expressed metabolites of D. newyorkensis (Dub), A. muciniphila (Akk), E. faecalis (EF) and respective cultured supernatants (chopped meat carbohydrate broth, CMC; brain heart infusion medium, BHI; Luria-Bertani medium, LB) at 48 h. b Ido1 expression in mouse bone marrow-derived dendritic cells (BMDCs) treated with N-acetyl-L-Asp (NAA; 1 mM ), L-Lys ( 8 mM ), L-lactic acid (Lac; 100 mM ), L- -aminobutyric acid (Abu; 0.1 mM ), ketoleucine ( ) and Kyn ( 0.2 mM ) ( ). IDO1 expression and colon length of Abx mice treated with metabolites were measured at D3 or D7 post-DSS administration, respectively ( ). e IDO1 activity in BMDCs treated with Lys or IFN was determined by Kyn/ Trp ratio of cell culture supernatant Conventional WT, DT-treated Foxp3DTR, or Ido1-/- mice were colonized with Dub or treated with Lys ( ) and subjected to DSS administration for 7 days. T cell subsets ( ) in mesenteric lymph nodes (MLN) and colonic lamina propria ( ) (cLP) colon length ( ) ( ) and
histopathology were determined. Serum Kyn concentration of Lystreated conventional WT, DT-treated Foxp3-DTR or Ido1-/- mice before DSS exposure (WT, Foxp3-DTR, ; Ido1 ). 1 In vivo imaging of colon from vehicle (Veh)-, Dub-, Lys- or Kyn-treated transgenic IL-17-EGFP mice. m-r Lys- or Vehtreated germ-free mice were exposed to DSS treatment for 7 days and colonic Ido1 expression , T cell subsets in MLN and cLP , colon length , histopathology (q) and proinflammatory cytokines ( ) ( ) were evaluated. s Ido1 expression in Lys- ( 8 mM ) or Dub supernatant (Dub.sup)-treated bone marrowderived dendritic cells (BMDCs) extracted from GF mice at 18 h post-treatment ( ). Fold of change in frequencies of T cell subsets ( ) or colon length ( ) was calculated and presented with numbers in red. Dashed lines at 1 indicate that the treatments have equal value as normalized controls. Results are representative of data generated in three independent experiments and are expressed as mean SEM, and 2 -sided -values were examined by the Student’s -test. Source data are provided as a Source Data file.
To test the physiological relevance of the Lys-mediated IDO1 enhancement in suppressing inflammation via Treg induction in vivo, we gavaged conventional WT, Ido1-/- or Foxp3-DTR mice with Dub or Lys before DSS exposure (Supplementary Fig. 8j). It was found that Lys treatment augmented Foxp3 Tregs and diminished T cells to a similar extent that Dub colonization did in the spleen, MLNs, and cLP compared with Veh-treated WT mice, but Lys failed to modify Foxp3 Tregs in all 3 tissues or cells in cLP of Ido1 mice (Fig. 5f, g and Supplementary Fig. 8k).
Moreover, Lys conferred clear beneficial effect on disease phenotypes (Fig. 5h and Supplementary Fig. 81), histopathological changes (Fig. 5i, j), mucosal barrier function (Supplementary Fig. 8m), and inflammatory responses (Supplementary Fig. 8n) in conventional WT mice, whereas the protective effect was abolished in or Foxp3DTR mice. Consistent with the flow data, a mild protective effect remained in Ido1-/-Dub and Foxp3-DTR-Dub mice as manifested by histopathological assessment (Fig. 5i, j). Furthermore, we determined that WT-Lys, but not Ido1-/-Lys mice exhibited elevated serum Kyn concentration (Fig. 5k). Despite a smaller effect than Dub colonization, Lys and Kyn administration to DSS-treated IL-17-EGFP mice resulted in a similar downregulation in colonic IL-17 expression, as measured by in vivo imaging (Fig. 51).
Subsequently, we determined whether the phenotype observed in conventional WT mice with normal gut flora was also true in GF settings. Indeed, GF-Lys displayed an increased serum Lys concentration prior to DSS exposure and an elevated fecal Lys concentration was confirmed in GF-Dub group (Supplementary Fig. 9b). As expected, Lys administration to GF mice (Supplementary Fig. 9a) caused higher Ido1 expression in cLP (Fig. 5m), elevated serum Kyn concentration (Supplementary Fig. 9b), upregulated Foxp3 Tregs and downregulated cells in the spleen and cLP (Fig. 5n, o and Supplementary Fig. 9c), but unexpectedly downregulated CD25 Foxp3 Tregs in the MLN (Fig. 5n). Lys also had a relieving effect on disease phenotype (Fig. 5p and Supplementary Fig. 9d), tissue injury (Fig. 5q and Supplementary Fig. 9e), mucosal barrier damage as well as local and peripheral inflammation (Fig. 5r and Supplementary Fig. 9f). Moreover, we pretreated GF mBMDCs with Lys and found significantly increased Ido1 expression (Fig. 5s), implying that the stimulation of IDO1 in DCs by Lys was more likely due to cell-intrinsic changes not dependent on microbiota-derived signals.
For a better understanding of how Lys enhances Ido1 expression in DCs, we performed RNA-Seq on Lys-treated WT BMDCs, revealing broad induction of AhR response genes (Fig. 6a). We confirmed the upregulation of Ahr (Fig. 6b, c) and the AhR target genes Aldh1a3, Cyp1a1, Cyp1b1, Tiparp, Il1b, and Il6 in both conventional WT and GF mBMDCs at 6 h post-Lys induction (Supplementary Fig. 10a, b). Moreover, Lys led to translocation of AhR into the nucleus at 6 h posttreatment, implying an immediate effect of Lys on AhR (Fig. 6d). In accordance with this, western blot of Lys-treated mBMDCs showed
decreased cytoplasmic localization in parallel to increased nuclear AhR accumulation, comparable to that induced by the AhR agonist 6-for-mylindolo[3,2-b]carbazole (FICZ) (Fig. 6e). Activation of AhR and the resulting upregulation of Ido1 expression in response to Lys were both inhibited by the AhR antagonist CH-223191 (Fig. 6f) or knockdown of Ahr (Fig. 6g and Supplementary Fig. 10c).
To evaluate the effects of Lys on AhR signaling in vivo, groups of WT mice were treated prior to DSS administration either with Lys in combination with , or alone (Supplementary Fig. 10d). Consistent with the in vitro data, Lys administration did not boost Ido1 expression in cLPMCs in the presence of CH-223191 at D3 post-DSS treatment (Fig. 6h). In agreement with the reported protective effects of AhR in DSS-induced colitis , WT-CH-223191 group displayed clearly impaired colonic Ahr expression (Supplementary Fig. 10e) and worsened disease phenotype (Fig. 6i, j). The differences in Foxp Tregs or IL17 cells and proinflammatory cytokines in cLPMCs between WT-Lys and WT-Veh were lost in the presence of CH-223191 (Fig. 6k, I and Supplementary Fig. 10f, g). Collectively, these results suggest that Lys enhances IDO1 expression to confer protection against DSS-induced colitis via AhR activation.
The human homolog of Dub, C. innocuum, produces L-Lys and promotes Foxp3 Tregs to alleviate DSS-induced injury by upregulating IDO1 expression in DCs
Recently, it has been documented that increases in the intestinal Erysipelotrichaceae family were causally related to a lower risk of IBD . Thus, we performed a phylogenetic analysis of the Erysipelotrichaceae family and showed that an underrepresented human commensal species, C. innocuum (abbreviated as Clos hereafter), is a close relative to Dub (Fig. 7a, phylogenetic distance ). We compared the gut metagenomic datasets of the publicly available IBD cohorts and healthy matched controls , discovering that Clos was significantly decreased in patients with IBD, particularly those with UC, compared with healthy individuals (Fig. 7b). In addition, a USA cohort-based analysis displayed a lower abundance of Clos in patients with UC or CD in comparison with the healthy controls (Fig. 7c).
Similar to Dub, Clos generated Lys in the culture supernatant (Fig. 7d). Further, the Clos.sup and Lys both promoted Ido1 expression in primary DCs extracted from human PBMCs (hPBMCs) (Fig. 7e) and WT mBMDCs (Fig. 7f), with increased Kyn/Trp ratio in mBMDC culture supernatant (Fig. 7g). In contrast, this phenotype was not observed in Ido1-/- mBMDCs (Fig. 7g), and was abrogated by CH-223191 (Fig. 7h) or AhR-specific small interfering RNA (siAhR) (Fig. 7i). In Lystreated GF mBMDCs, upregulation of Ahr and its target genes, as well as Lys and Trp transporters were observed at 6 h post-treatment (Supplementary Fig. 11a, b). Similarly, higher Ido1 expression was exhibited at 18 h post-treatment (Supplementary Fig. 11c). Together, these results indicate that Clos, the human homolog of Dub, may
Fig. 6 | L-Lys activates the AhR-IDO1 axis in DCs to induce immunosuppressive
Treg responses. a Transcriptomic analysis of mock-, Lys-, IFN- -treated mouse BMDCs at indicated times post-treatment (mock, ; Lys , IFN 18 h , . b expression in Lys- or IFN- -treated BMDCs extracted from conventional WT mice . с expression in Lys- or Dub.sup-treated BMDCs extracted from GF mice ( ). d Translocation of AhR (red) into the nucleus (DAPI, blue) from cytoplasm ( -tubulin, green) of mouse BMDCs after 6 h treatment with Lys or the AhR agonist 6-formylindolo[3,2-b]carbazole (FICZ). e AhR protein analysis in nuclear and cytoplasmic fractions of mock-, Lys-, or FICZ-treated mouse BMDCs at 6 h post-treatment by western blot. Expression of Ido1 in mouse BMDCs treated with Lys, FICZ, or the AhR antagonist CH-223191, or a combination of CH223191 and Lys/FICZ ( ). g Expression of Ido1 in mouse BMDCs transfected with negative control (siNC) or AhR-specific small interfering RNA (siAhR) and treated with Lys or FICZ at 18 h post-treatment ( ). For the experiments shown in ( )
above, the following were used: Lys ( 8 mM ), FICZ ( 300 nM ), and CH-223191 ( ). For experiments shown in ( ) below, conventional WT mice ( ) were treated with Lys ( ), CH-223191 ( ), or Lys plus CH-223191 and subjected to DSS administration. Expression of Ido1 in the cLPMCs from Veh- or Lys-treated WT mice at D3 post-DSS administration in the presence of CH-223191 . Colon length (i), histopathological changes by HE staining , CD25 Foxp3 Tregs ( ) and IL-17 CD4 T cells ( ) ( ) in MLN and cLP of Veh- or Lys-treated conventional WT mice in the presence of CH-223191 were examined at D7 post-DSS treatment. Dashed lines at 1 indicate that the treatments have equal value as normalized controls. Results are representative of data generated in three independent experiments and are expressed as mean SEM, and 2 -sided -values were examined by the Student’s -test. Source data are provided as a Source Data file.
also have an enhancing effect on Trp metabolism through the Kyn pathway by promoting IDO1 expression in both human and mouse DCs via AhR activation.
Next, we tested Clos or Clos.sup in WT mice and Ido1-/- mice (Supplementary Fig. 11d), observing that both Clos colonization (Supplementary Fig. 11e) and Clos.sup administration significantly increased CD25 Foxp3 Tregs and decreased IL17 CD4 T cells in the spleen, MLNs and cLP in WT mice, but not in Ido1 mice (Fig. 7j, k and Supplementary Fig. 11f). Further, colonization of Clos or treatment with Clos.sup in WT mice mitigated colon shortening (Fig. 71 and
Supplementary Fig. 11g), colonic tissue injuries (Fig. 7m and Supplementary Fig. 11h) and inflammatory responses (Supplementary Fig. 11i), an effect which was not observed in mice. Thus, it is suggested that Clos may confer protection against colitis in humans by ameliorating unbalanced Treg/Th17 responses through enhancement of IDO1 expression.
Discussion
A growing body of literature suggests that the intestinal microbiome plays a fundamental role in the maintenance of gut homeostasis
through enhancing epithelial barrier function and immune tolerance . Metabolism has emerged as a new player in mucosal immune functions and inflammation with the advent of the new field of immunometabolism , though the role of specific microbiota-derived metabolites and their mode of action remain mostly elusive. Here we revealed that the Neo-resistant commensal bacterium, Dub, plays a
pivotal role in enhancing mucosal barrier integrity and rebalancing Treg/Th17 responses to maintain intestinal homeostasis through production of SCFAs (particularly propionate) and Lys (Fig. 7n). We have demonstrated a previously unknown role for intestinal microbederived Lys in promoting IDO1 expression in colonic DCs. By administrating Dub or Lys, we were able to skew endogenous Trp metabolism
Fig. 7 | The human homolog of D. newyorkensis, Clostridium innocuum, generates L-Lys and protects against DSS-induced colitis by activating IDO1 expression in DCs. a Phylogenic analysis of the Erysipelotrichaceae family and the Clostridium innocuum isolate. b, c Relative abundance of C. innocuum (Clos) in fecal samples from healthy individuals and patients with inflammatory bowel disease (IBD) (b), and specifically ulcerative colitis (UC) and Crohn’s disease (CD) (c). Cohort PRJNA389280 (healthy, ; IBD, ), cohort PRJNA400072 (healthy, ; IBD, ), cohort PRJNA398089 (healthy, ; UC, ; CD, ). d Levels of Lys in the culture supernatants of Dub (Dub.sup) or Clos (Clos.sup) ( ). e qPCR of Ido1 mRNA in the Lys- or Clos.sup-treated human DCs extracted from peripheral blood mononuclear cells at 18 h post-treatment ( ). qPCR of Ido1 mRNA at 18 h post-treatment in wild-type C57BL/6J mouse (WT) bone marrowderived dendritic cells (BMDCs) treated with 8 mM Lys, IFN , Dub.sup, Clos.sup or chopped meat carbohydrate broth (CMC) ( ). g Kyn/Trp ratio of Clos.sup-treated, or nontreated BMDCs extracted from WT or Ido1-/- mice at 18 h post-treatment . h Expression of Ido1 in mouse BMDCs treated with Clos.sup,
CH-223191, or a combination of CH-223191 and Clos.sup ( ). i Expression of Ido1 in mouse BMDCs transfected with negative control (siNC) or AhR-specific small interfering RNA (siAhR) and treated with Clos.sup at 18 h post-transfection ( ). j-m Conventional WT or Ido1 mice were colonized with CFU of Clos or treated with Clos.sup and subjected to DSS administration. At D7 post-DSS administration, CD25 Foxp3 Tregs ( ) and IL-17 CD4 T cells ( ) in mesenteric lymph nodes (MLN) and colonic lamina propria (cLP) (WT, ; Ido1 ), colon length (1) and histopathological changes by HE staining (m) were examined at D7 post-DSS treatment . (n) Schematic illustration of the D. newyorkensis metabolite Lys protecting against DSS-induced colitis by rebalancing Treg/Th17 response through the activation of the AhR-IDO1-Kyn circuitry. Dashed lines at 1 indicate that the treatments have equal value as normalized controls. Results are representative of data generated in at least two independent experiments and are expressed as mean SEM, and 2 -sided -values were examined by the Student’s -test. Source data are provided as a Source Data file.
toward the Kyn pathway and rebalance Treg/Th17 responses to protect mice from colitis.
Multiple studies have demonstrated that antibiotic treatment can improve the symptoms of colitis, both in human trials and DSSinduced mouse models , by shaping the composition of microbiota. Beneficial effect to clinical symptoms and inflammation suppression was linked to increased probiotic bacteria such as Bifidobacteria, . prausnitzii or Enterobacter ludwigii (E. ludwigii) . In the present study, we found that Neo treatment could prevent DSS-induced colonic inflammation most effectively among all 4 broad-spectrum antibiotics (Amp, Van, Neo and Metro) (Fig. 1). This outcome does not resemble the results of a very recent study conducted by Li et al., also using multiple single-antibiotic treatments in a DSS-induced mouse model, in which Metro, but not Neo, was reported to induce DSS-colitis remission attributed to the expansion of E. ludwigii . These discrepancies may be explained by differences in antibiotic regimens between the 2 studies (high-dose oral gavage in our study vs low-dose water distribution in Li et al.), as we have shown that these 2 treatment regimens could result in remarkably disparate susceptibility to caerulein-induced acute pancreatitis due to greatly distinct microbiota compositions .
The intestinal abundance of Dub has been reported to be associated with obesity and Alzheimer’s disease in mouse models, however no functional studies have been performed to confirm its biological relevance in vivo. For the very first time, we comprehensively analyzed its potential role as a probiotic in both conventional and microbiota-depleted mouse models. Dub possesses a probiotic capacity superior to the well-known probiotic commensal bacterium Akk in improving colonic mucosal barrier damage and intestinal inflammation caused by DSS and TNBS (Fig. 2). Our findings that Dub relieves host mucosal and extraintestinal inflammation by rebalancing Treg/Th17 responses are perfectly in line with 2 landmark studies that demonstrate acetate and propionate regulate GPR43-dependent immunosuppression by cTregs under DSS or T cell-transfer conditions .
Although the key role of the gut microbiota in stimulating IDO1 activity has been demonstrated in both GF and Abx-treated mouse models , there has been very little focus on the modulatory influences on host Trp catabolism through the Kyn pathway exerted by specific commensal bacteria and microbial metabolites in IBD. Herein, we unraveled a novel role for Dub in enhancing host Trp metabolism through the Kyn pathway to regulate Treg/Th17 responses under steady-state or inflammatory conditions by promoting IDO1 expression in cLPMCs (Fig. 4). Further, we showed that Dub-derived Lys biased the function of DCs toward a regulatory and tolerogenic function to promote Tregs by upregulation of Kyn, through AhRdependent IDO1 activity (Fig. 6). According to Gargaro et al. and others, both the upstream region and internal noncoding region of Ido1
contains canonical xenobiotic responsive elements (XREs), which are AhR binding sites . Additionally, we demonstrated that Lys upregulates SLC7A5 in DCs (Fig. S8h, i), which is a fundamental carrier for the IDO1-driven Kyn in establishing the AhR-IDO1-Kyn metabolic circuit in DCs via autocrine and paracrine signaling . It needs to be noted that mild discrepancies still existed in Treg and Th17 between Ido1 -Dub and Ido1-/-Veh/Lys groups (Fig. 5f, g), which could be plausibly explained by the fact that Dub is also able to rebalance Treg/ Th17 responses through the SCFA-GPR43 axis (Fig. 3h, i). Thus, we are trying to generate Gpr43-/-Ido1 double knockout mice to confirm that these two immunomodulatory factors synergistically regulate Treg/Th17 responses in DSS-induced colitis.
Hardly any studies have shown that Lys impacts AhR activity, either directly or indirectly, and the molecular link between Lys catabolism and AhR activation remains uncertain, although it is possible that accumulation of crotonyl-CoA resulting from Lys catabolism reprograms IFN signaling , which may in turn activate the AhR-IDO1 axis . Although it has been recently shown that different DC subsets acquire the IDO1 tolerogenic pathway and respond to immune-active Trp metabolites differently , further studies are needed to explore the regulatory role of Lys in the AhR-IDO1-Kyn circuitry in particular DC subsets. Experiments with cell type-specific conditional deletion of Ahr or Ido1 are needed to address this concern. Lastly, we are making great efforts to generate a mutant Dub strain with (encodes essential diaminopimelate epimerase for Lys biosynthesis) depletion, which will enable us to mechanistically dissect the specific role of Dub-synthesized Lys for mitigating DSS-induced colitis.
In summary, our comprehensive analysis demonstrates a previously unrevealed role of Dub in activating the AhR-IDO1-Kyn circuit in DCs, which improved the unbalanced Treg/Th17 response to ameliorate both mucosal and systemic inflammation in DSS-induced colitis. The finding that the human homolog Clos and its associated metabolite Lys can mitigate mucosal inflammation and improve mucosal healing in a mouse model may provide a basis for the development of microbiota-based therapeutic approaches for clinical IBD in humans.
Methods
Ethics statement
All animal experiments were strictly carried out in accordance with protocol no. 117113 approved by the Institutional Animal Care and Use Committee (IACUC) of Zhejiang University. Blood collection from healthy human donors was performed with the approval of the Ethical Committee of The First Affiliated Hospital of Guangdong Pharmaceutical University (Permit Number: 20210221). All participants provided written informed consent for sample collection and subsequent analyses.
Bacterial strains
A. muciniphila (Akk) was obtained from American Type Culture Collection (ATCC, No. BAA835) and cultured in brain heart infusion medium (BHI; Oxoid) at under anaerobic conditions. D. newyorkensis (Dab) and C. innocuum were purchased from the German collection of microorganisms (DSMZ, DSM103457 and DSM1286-1213001) and cultured in chopped meat carbohydrate broth (CMC, Hopebio) at under anaerobic conditions. E. faecalis (EF) was purchased from the China Center for Type Culture Collection (CCTCC, AB 2018154 ) and cultured in Lysogeny broth medium (Oxoid) at . The concentration of each bacterial species was estimated based on the optical density at .
Mouse strains
Six- to eight-week-old and sex-matched mice on C57BL/6J background were used in this study. Germ-free (GF) C57BL/6J mice were bred and housed at the Shenzhen Gnotobio Biotechnology Co., Ltd. GF status was confirmed through 16 S qPCR analysis before used for relative experiments, which were also carried out at Shenzhen Gnotobio Biotechnology Co., Ltd. Wild-type C57BL/6J (WT) mice were purchased from the Model Animal Research Center of Nanjing University (Nanjing, China). Foxp3-DTR mice were a gift from Dr. Bin Li from Department of Immunology and Microbiology, Shanghai Institute of Immunology, Shanghai Jiao Tong University School of Medicine. Foxp3-DTR mice were given intraperitoneal (i.p.) injections of 1 mg DTx ( 50 ng DT/g body weight) once per day for 7 consecutive days to guarantee successful depletion of Tregs. Splenocytes were analyzed to verify the elimination efficiency of Tregs by flow cytometry. Indolea-mine-2,3-dioxygenase 1 knockout mice (Ido1- ) were kindly provided by Dr. Yajing Wang from State Key Laboratory of Natural Medicines, Department of Physiology, China Pharmaceutical University. G-protein coupled receptor 43 knockout mice (Gpr43–) and IL-17-EGFP transgenic mice were purchased from Cyagen Biosciences Inc (Suzhou, China). The mice were routinely maintained in a specific-pathogen-free facility with a temperature- and humidity-controlled environment ( , humidity), and under a constant 12 h light/dark cycle, and were given free access to a regular chow diet (Gat# P1101F-25, Shanghai SLACOM) and water throughout study at Zhejiang University. All procedures were conducted in compliance with a protocol approved by the IACUC at Zhejiang University, China.
Antibiotic treatment, fecal microbiota transplantation (FMT), and bacterial colonization
Mice were given ampicillin (Amp 10 mg ), neomycin ( Neo 10 mg ), metronidazole (Metro 10 mg ), or vancomycin (Van 10 mg ), individually or in combination (referred to as Abx) daily for 5 days via oral gavage. Fecal samples collected from microbiota-depleted mice at the 5th day post-treatment were homogenized, plated on BHI agar with sheep blood, and cultured under anaerobic conditions at for 2 days followed by incubation under aerobic conditions at for 1 day to confirm efficient microbial depletion.
For FMT experiments, 200 mg of pooled fecal pellets from WT mice were homogenized with sterile silica beads in 1.5 mL PBS at 45 Hz for 1 min and filtered with strainers. Abx-treated WT mice or GF mice were subjected to gavage with filtered stool homogenates (FMT experiments) or CFU of cultured bacteria (colonization experiments) twice with a 48 – h interval. At 48 h after FMT or bacterial colonization, stool samples were collected to determine the efficiency of colonization prior to DSS administration for colitis model induction.
DSS-induced colitis mouse model
Unless otherwise specified, WT or certain knockout mice were administered 2.5% DSS (Cat No: 60316ES76; Yeasen, shanghai, China) in drinking water for 7 days to establish the DSS-induced colitis mouse model.
To study the effect of antibiotics on mouse susceptibility to DSSinduced colitis, WT mice were pretreated with single antibiotics for 5 days, antibiotics administration was discontinued on day 6, and exposed to DSS treatment. Treated animals were weighed daily and fecal samples were collected at indicated times post-DSS treatment.
To evaluate the effect of bacterial strains on DSS-induced colitis, conventional WT mice were gavaged with CFU of D. newyorkensis (Dub), A. muciniphila (Akk), or E.faecalis (EF) twice with a 48 -h interval, then subjected to DSS treatment. To evaluate the therapeutic effect of individual strains on experimental colitis without pretreatment, conventional WT mice were treated with DSS in drinking water for 3 days, then gavaged from the 3rd day to the 8th day with CFU of Dub, Akk, or EF daily, maintaining DSS in the drinking water.
To study the major effect of individual bacterial strains on DSSinduced colitis in the context of microbiota depletion, WT mice were pretreated with Abx for 5 days and on the day gavaged with CFU of Dub, Akk, or EF twice with a 48-h break between treatments prior to DSS administration.
For pasteurization (HI) experiments, Abx-treated WT mice were orally gavaged for 5 days with CFU of Dub inactivated by pasteurization for 30 min at , diluted in PBS, and administered DSS on the 6th day.
Supernatants from cultured Dub or C. innocuum (Clos) were centrifuged at for 10 min at and then passed through polyether-sulfone filters ( ; Merck Millipore) to remove residual bacterial cells. Conventional or Abx-treated WT mice were gavaged with respective bacterial supernatants ( per mouse) for 5 days and given DSS on the 6th day.
TNBS-induced colitis mouse model
For presensitization, mice were shaved on the back just below the neck and painted with 1% TNBS (Meilunbio MB5523) mixed in an acetone/ olive oil solution. Colitis was subsequently induced with TNBS in ethanol by rectal injection 7 days after presensitization as described previously .
Animal protection study with metabolites
For the SCFA protection study, WT and Gpr43-/ mice were given 200 mM propionate in drinking water for 3 weeks and then administered DSS. To test the effect of Dub-derived metabolites, mice were gavaged with L-lactic acid (Lac; ), ketoleucine ( ; ), N-acetyl-L-Asp (NAA; ), L-Lys ( ), L- aminobutyric acid (Abu; ) daily for 5 days before DSS treatment. For the Kyn protection study, WT or Foxp3-DTR mice were i.p. injected with Kyn every other day, 7 total doses from the day previous to DSS to the 6th day post-DSS administration. For the Lys protection study, WT mice, Foxp3-DTR, or Ido1 mice were given L-Lys ( ) via oral gavage once per day for 5 days before DSS administration. For AhR antagonist experiments, WT mice were treated i.p. with CH-223191 ( , MedChemExpress) daily for 10 consecutive days (from day -7 to day 2 ). During this period, mice were treated with Lys via oral gavage for 5 days (from day -5 to day 0 ) and then administered DSS.
Fecal bacteria quantification
Fecal bacteria were quantified by qPCR (primers are listed in the table of Key Resources). Fecal bacterial DNA was isolated using a TIANamp Stool DNA Kit (TIANGEN), and qPCR was performed using SYBR Green Real-time PCR Master Mix (TOYOBO).
DNA extraction, 16S rDNA amplicon sequencing, and data analyses
Fecal samples ( ) were resuspended in Qiagen’s ASL buffer and homogenized for 2 min . Total fecal DNA was extracted from the resulting supernatant using a QIAamp DNA Stool Mini Kit (Qiagen),
and DNA concentration and purity were measured by Qubit (Thermo Fisher Scientific). The stool DNA was then amplified using Phusion High-Fidelity PCR Master Mix (New England Biolabs) by PCR targeting the variable regions 3 and 4 (V3-V4) of the 16S rDNA. Multiplex sequencing of amplicons with sample-specific barcodes was performed using the MiSeq Illumina platform (Guangdong Magigene Biotechnology Co., Ltd.). Paired-end reads were merged into long sequences using FLASH v.1.2.7, a very fast and accurate analysis tool designed to merge paired-end reads when there are overlaps between reads1 and reads2. The merged sequences were then analyzed using the QIIME v.1.9.1 software package.
Quantitative RT-PCR
Total RNA from bead-homogenized tissue samples or cell culture was extracted using TRIzol reagent (Invitrogen) following the manufacturer’s instructions. PCR reactions were performed with HiScript II One Step qRT-PCR SYBR Green Kit (Vazyme) on a Gentier 96R RealTime PCR System (TIANLONG, Xi’an, China). Transcript levels of the indicated genes were normalized to endogenous control GAPDH for each individual sample using the primers listed in the Key Resources table and quantified using the comparative critical threshold cycle method.
Cytokine expression analyses
, IL-6, and TNF- protein levels in serum samples and colon homogenates were measured by the corresponding enzyme-linked immunosorbent assay (ELISA) kits (70-EK201B/3-96, 70-EK206/3-96, 70-EK282/4-96 MultiSciences) following the manufacturer’s instructions.
Western blot analysis
Tissues and cells treated as indicated were lysed with RIPA lysis buffer (Beyotime) and subjected to SDS-polyacrylamide gel electrophoresis and then transferred to polyvinylidene difluoride membranes (Millipore). Proteins were further incubated with the indicated primary antibodies and then horseradish peroxidase-conjugated secondary antibodies. Protein bands were visualized using an enhanced chemiluminescence kit (Vazyme) with a ChemiDoc Touch Gel Imaging System (Bio-Rad).
Tissue histology, immunostaining, and in vivo imaging
Colon ‘Swiss rolls’ soaked in paraformaldehyde solution were dehydrated, embedded in paraffin, cut into thick sections, and stained with haematoxylin and eosin (HE) using standard procedures. Slices were evaluated by an experienced pathologist in a blinded manner, and histological scores were assessed based on the following parameters according to previous research: inflammation, epithelial defects, crypt atrophy, dysplasia/neoplasia, and the area affected by dysplasia . For the staining of goblet cells, colon sections were also stained in Alcian blue for and dehydrated in alcohol and xylene.
For immunofluorescence (IFA) analysis, deparaffinized colon sections were blocked with normal goat serum for 30 min at room temperature (RT). The slides were then incubated with specific primary antibodies at for 12 h . Antibodies used: anti-IL-6 (1:50, 21865-1-AP Proteintech), anti-F4/80 (1:50, 28463-1-AP Proteintech), antioccludin (1:100, Proteintech 13409-1-AP) and anti-Muc2 (1:100, 27675-1-AP Proteintech). Slides were then incubated with fluorescently labeled secondary antibodies (Jackson ImmunoResearch). Nuclei were stained with DAPI (Roche, Switzerland). TUNEL staining was performed using the In situ Cell Death Detection POD kit (Roche Diagnostics) on the Discovery XT according to the manufacturer’s protocol. All analyses were performed using ImageJ software.
IL-17-EGFP transgenic mice were colonized with CFU Dub, treated with Kyn or Lys and subjected to DSS
administration as described above. At day 7 post-DSS administration, the colon was dissected from IL-17-EGFP transgenic mice in each group. In vivo imaging was performed using PerkinElmer (CLS136341/ F) and representative images were taken.
Transmission electron microscopy analysis
A segment of fresh colon tissues was flushed with PBS and fixed in glutaraldehyde at for 4 h . After being rinsed in PBS, the tissue was further fixed in PBS containing 1% osmium tetroxide for 2 h at RT, rinsed in PBS, and dehydrated. The tissues were then embedded in Epon 812 overnight, then cured in an oven at for 48 h . Sections of 80 nm thickness were cut on an ultramicrotome (RMC MTX) using a diamond knife. The sections were deposited on single-hole grids coated with Formvar and carbon and double-stained in aqueous solutions of uranyl acetate for 25 min at and lead citrate for 3 min at RT. Images were acquired on an H-7650 TEM (Hitachi, Ibaraki, Japan) at 80 kV of accelerating voltage. Regions of interest in the altered membranes were photographed by a Gatan 830 CCD camera (Gatan, CA, USA).
Isolation of colonic intestinal epithelial cells and lamina propria mononuclear cells
Murine IECs and LPMCs were obtained from the colon as previously described . Briefly, the colon was opened longitudinally and cut into pieces. After incubation with EDTA ( 5.5 mM ) and dithiothreitol (DTT) ( 1 mM ) in Hank’s balanced salt solution (HBSS), vortexing and passing through a cell strainer, the suspension of IECs was washed twice by centrifugation at for 2 min and collected for future experiments. The remaining lamina propria tissue was incubated with digestion solution containing collagenase ( ) and DNase . The resulting LPMC cell suspension was subjected to Percoll-gradient separation and harvested for further experiments.
Flow cytometry
For cell surface staining, single-cell suspensions were incubated on ice for 30 min with the following antibodies: FITC-conjugated anti-CD3 (11-0032-82, eBioscience, 1:150), eFluor 450-conjugated anti-CD4 (48-0041-82, eBioscience, 1:150), and APC-conjugated anti-CD25 (17-025182, eBioscience, 1:150).
For Treg cell analysis after staining of cells with CD3, CD4, and CD25 antibodies, lymphocyte suspensions were fixed and permeabilized using transcription factor buffer sets (562574, BD Pharmingen) according to the manufacturer’s instructions and stained with anti-Foxp3-PE (12-5773-82, eBioscience, 1:75). For analysis of Th1 and Th17 cells, isolated tissue lymphocytes were stimulated for 5 h with cell stimulation cocktail plus protein transport inhibitors (00-4975-93, eBioscience). After incubation for 5 h , cells were washed in PBS and stained for cell death using fixable viability stain 570 (564995, BD Horizon, 1:100), FITC-anti-CD3, and eFluor 450-anti-CD4. Stained cells were fixed in fixation buffer (00-8222-49, eBioscience), permeabilized with intracellular staining permeabilization wash buffer (00-8333-56, eBioscience), and stained with anti-IL-17A conjugated to PerCPCyanine5.5 (45-7177-82, eBioscience, 1:75) and phycoerythrinconjugated anti-IFN- (505808, Biolegend, 1:75).
Isolation of murine BMDMs and BMDCs
Mouse bone marrow-derived macrophages (mBMDMs) were isolated from bone marrow extracted from mouse femurs and tibiae as previously described . Cells were cultured at and in DMEM supplemented with FBS and supernatant of filtered L929 cells, penicillin, streptomycin at and for 7 days prior to the experimental procedure. Murine bone marrowderived dendritic cells (mBMDCs) were derived from bone marrow cells and stimulated with GM-CSF (Servicebio) and IL- 4 (PeproTech) for 6 days. of mBMDMs or mBMDCs were then
seeded into a 24-well plate and treated with L-lactic acid ( 100 mM ), Kyn ( 0.2 mM ), , NAA ( 1 mM ), L-Lys ( 8 mM ), Abu ( 0.1 mM ) for Ido1 detection at 18 h post-treatment by qRT-PCR.
Sorting of human dendritic cells
Healthy individuals admitted to the hospital for healthy examination were recruited as healthy donors in the study. Peripheral blood specimens were collected from seven healthy individuals ( 3 males and 4 females) aged from 22 to 41 and peripheral blood mononuclear cells (PBMCs) were extracted by density gradient centrifugation using human PBMC isolation kits (LTS10771 TBD) followed the manufacturer’s protocol. After isolation, dendritic cells (DCs) were enriched from PBMCs using the EasySep Human Myeloid DC Enrichment Kit (19061 Stemcell), and enriched DCs were stained with BV421 antihuman CD11c ( 301628 BioLegend, per sample). The enriched DCs were used for downstream assays. The protocol was reviewed and approved by the Committee for Ethical Review of The First Affiliated Hospital of Guangdong Pharmaceutical University, and written informed consent was obtained from each participant.
Organoid culture
Murine duodenal and colonic organoids were cultured as previously described . Briefly, crypts of duodenum were counted and embedded in of Matrigel (Corning) at 10,000 crypts/mL and cultured in the mouse colonic organoid kit (K2204-MC, bioGenous). Surface area of organoids was quantified with ImageJ (National Institutes of Health). Colonic crypts were isolated and cultured as described previously . Crypts of colon were counted and embedded in Matrigel and cultured in the mouse intestinal organoid kit (K2001-MI, bioGenous). At day 4 (duodenum) or day 3 (colon), organoids were stimulated with L-Lys for 18 h prior to RNA extraction and qRT-PCR detection.
Organoid, BMDC, and BMDM treatment with bacterial culture supernatants
Bacterial supernatants were centrifuged at for 10 min at and then passed through polyether-sulfone filters ( ; Merck Millipore). Organoids, BMDCs, and BMDMs were incubated with bacterial culture supernatants (8%) diluted in respective cell culture medium for 18 h at .
IDO1 activity determination
IDO1 activity was measured in vitro as the ability to convert Trp into L-Kyn. Thus, L-Trp was added to BMDCs in combination with 8 mM L-Lys or IFN- and incubated at for 18 h . Cell culture SUPs were collected and concentrations of L-Kyn and L-Trp were measured by LC-MS/MS.
AhR silencing and antagonization in BMDCs
Lipofectamine 2000 (Invitrogen 11668019) was added to Opti-MEM (Gibco 31985070), gently mixed, and allowed to stand for 5 min . An siRNA pool specific for was mixed and allowed to stand for 20 min . BMDCs seeded in 24 -well plates were incubated with the mixture for 10 min at RT and of complete DMEM was added (Pricella PM150210). After 24 h , transfected mBMDCs were used for downstream assays. For inhibition experiments, BMDCs were pretreated with CH-223191 (AhR antagonist) for 24 h and used for downstream assays.
Detection of AhR translocation
mBMDCs were exposed to 8 mM Lys or 300 nM FICZ for 6 h . Cells were fixed with 4% paraformaldehyde in PBS for 15 min at RT, followed by a permeabilization step with PBS containing 0.5% Triton X-100 for 20 min at RT. Next, the Click reaction was performed by adding of the reaction cocktail (Tublin 1:50, AF2835; AhR 1:100, AF6165 Beyotime) and incubated for 30 min at RT. Nuclei were stained with
DAPI. Analysis was conducted using a confocal laser scanning microscopy (LSM880) with a 60 x oil immersion objective. Tubulin was detected using the laser, AhR was detected using the laser, and DAPI was detected using the laser. Images were collected using Zen software.
Protein was isolated from the 2 different cellular fractions using a Cytoplasmic and Nuclear Extraction Kit (R0050 Solarbio), following the manufacturer’s instructions. Subsequently, samples were subjected to western blot analysis as described above. The cytoplasm-specific tubulin and nucleus-specific histone served as controls for appropriate fractionation. Antibodies used: anti-tubulin (1:50, AF2835 Beyotime), antihistone (1:50, AF0009 Beyotime) and anti-AHR (1:100, AF6165 Beyotime).
Transcriptomics analysis
BMDCs were pretreated with 8 mM Lys for 6,18 , or 24 h or IFN- for 18 h , then harvested for total RNA extraction with TRIzol reagent (Invitrogen). Samples were simultaneously assessed on an Agilent 4200 system (Agilent Technologies), Qubit 3.0 (Thermo Fisher Scientific), and Nanodrop One (Thermo Fisher Scientific). RNA-Seq libraries were generated and sequenced by Guangdong Magigene Biotechnology. Triplicate samples of all assays were constructed in an independent library, and the following sequencing and analysis were performed: Whole messenger RNA-Seq libraries were generated using a Next Ultra Nondirectional RNA Library Prep Kit for Illumina (New England Biolabs) following the manufacturer’s recommendations. Clustering of the index-coded samples was performed on a cBot Cluster Generation System. After cluster generation, libraries were sequenced on an Illumina NovaSeq 6000 platform, and 150-bp pairedend reads were generated. Raw data in fastq format were processed by Trimmomatic (v.0.36) to acquire clean reads, which were mapped to NCBI Rfam databases to remove the rDNA sequences using Bowtie2 (v.2.33). Remaining mRNA sequences were mapped to the reference genome by Hisat2 (2.1.0). HTSeq-count (v.0.9.1) was used to obtain the read count and function information of each gene according to mapping results. Differentially expressed genes of 2 conditions/groups were determined using edgeR (v.3.16.5), and GO analysis was implemented using clusterProfiler (v.3.4.4), in which gene-length bias was corrected.
Quasi-targeted metabolomics
Metabolites were extracted from the colon homogenates or bacterial SUP samples and sent to BGI (Shenzhen, China) for targeted metabolomics analyses. High-performance LC-MS/MS was used to perform high-sensitivity, wide-coverage, and high-throughput HM350-targeted quantification of 350 metabolites in these samples. In detail, appropriate amounts of experimental or quality control (QC) cell pellets were resuspended in of a water/methanol solution, lysed, and centrifuged, and the SUP was transferred to a new tube. A standard curve was prepared by serial dilution of an HM350 mixed standard. The experimental sample, QC sample, and standard were subjected to a derivatization reaction, and the resulting compounds were diluted in HM350 diluent and centrifuged at at for 10 min . The supernatant was applied to LC-MS/MS analysis on an LC-MS QTRAP (SCIEX). Chromatography was performed on a BEH C18 column ( , Waters). Mass spectrometry was performed with an ESI+/ESI- source; further statistical and bioinformatics analyses were performed by BGI using routine procedures.
LC-MS/MS determination of Trp and Kyn concentrations
Serum ( ) or BMDC culture supernatants were resuspended in of acetonitrile/methanol (8:2) and centrifuged at for 20 min . The supernatant was then dried using a nitrogen blower. The precipitates were reconstituted in of water/acetonitrile (8:2) containing formic acid by thorough vortexing and centrifugation. Finally, the SUP ( ) was injected into the LC-MS/MS system for
analysis. A UHPLC-MS/MS system (ExionLC AD UHPLC-QTRAP 6500+, AB SCIEX Corp.) was used to quantitate Trp and Kyn at Novogene. LCMS/MS was used to detect the concentration of a series of the standard solution. The concentration of the standard was used as the abscissa, and the ratio of the internal standard peak area was used as the ordinate to investigate the linearity of the standard solution.
GC-MS/MS analysis of SCFA concentrations
20 mg of each fecal sample was placed in a 2 mL EP tube, and 1 mL of phosphoric acid ( ) solution and a small steel ball were added. Samples were ground uniformly, then vortexed for 10 min and ultrasonicated for of SUP was collected after the mixture was centrifuged at for 10 min at MTBE (containing internal standard) solution was added, and the mixture was vortexed for 3 min and ultrasonicated for 5 min . After that, the mixture was centrifuged at for 10 min at , then the SUP was collected and used for GC-MS/MS analysis.
For GC-MS/MS analysis of SCFAs, an Agilent 7890B gas chromatograph coupled to a 7000D mass spectrometer with a DB-FFAP column ( 30 m length i.d. film thickness, J&W Scientific, USA) was employed. Helium was used as carrier gas, at a flow rate of . Injection was done in the split mode with an injection volume of . The oven temperature was held at for 1 min , then raised to at a rate of , then raised to at a rate of , held for 36 s , raised to at a rate of min , held for 0.5 min and run for 3 min . All samples were analyzed in the multiple reaction monitoring mode; the injector inlet and transfer line temperatures were and , respectively. SCFA content was detected by MetWare (http://www.metware.cn/) based on the Agilent 7890B-7000D GC-MS/MS platform.
Statistics and reproducibility
Statistical analyses were performed with Prism GraphPad software v.8.0. Error bars represent standard error of the means in all figures, and -values were determined by 2 -tailed Student’s -tests or ANOVA. was estimated for the correlation analysis of 2 continuous variables. A 2 -sided -value was considered statistically significant. All experiments were repeated, with the number of replicates stated in the figure legends. Representative images for western blots were from at least 3 independent sample preparations.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
16 S rRNA gene sequence data are available in the Sequence Read Archive (SRA) under BioProject accession PRJNA1013230. RNA-seq data are available in the SRA under BioProject accession PRJNA1012836. The mouse colon homogenates and bacterial cell culture supernatants metabolome data reported in this study have been deposited in the NGDC OMIX database (OMIX ID: OMIX005711; OMIX005712). All other data supporting the conclusions of this study are available in the paper and supplemental materials. Source data are provided with this paper.
References
Wang, G. et al. Bridging intestinal immunity and gut microbiota by metabolites. Cell. Mol. Life Sci. 76, 3917-3937 (2019).
Nicolas, G. R. & Chang, P. V. Deciphering the chemical lexicon of host-gut microbiota interactions. Trends Pharmacol. Sci. 40, 430-445 (2019).
Skelly, A. N., Sato, Y., Kearney, S. & Honda, K. Mining the microbiota for microbial and metabolite-based immunotherapies. Nat. Rev. Immunol. 19, 305-323 (2019).
Rooks, M. G. & Garrett, W. S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 16, 341-352 (2016).
Subramanian, B. C. Inflammatory bowel disease: DCs sense LTB(4) to drive and differentiation. Cell. Mol. Immunol. 17, 307-309 (2020).
Tanoue, T., Atarashi, K. & Honda, K. Development and maintenance of intestinal regulatory T cells. Nat. Rev. Immunol. 16, 295-309 (2016).
Ramanan, D. et al. Regulatory T cells in the face of the intestinal microbiota. Nat. Rev. Immunol. 23, 749-762 (2023).
Smith, P. M. et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 341, 569-573 (2013).
Furusawa, Y. et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504, 446-450 (2013).
Sharma, M. D. et al. Inhibition of the BTK-IDO-mTOR axis promotes differentiation of monocyte-lineage dendritic cells and enhances anti-tumor T cell immunity. Immunity 54, 2354-2371.e8 (2021).
Puccetti, P. & Grohmann, U. IDO and regulatory T cells: a role for reverse signalling and non-canonical NF-kappaB activation. Nat. Rev. Immunol. 7, 817-823 (2007).
Fallarino, F. et al. The combined effects of tryptophan starvation and tryptophan catabolites down-regulate T cell receptor zetachain and induce a regulatory phenotype in naive T cells. J. Immunol. 176, 6752-6761 (2006).
Mezrich, J. D. et al. An interaction between kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J. Immunol. 185, 3190-3198 (2010).
Di Meglio, P. et al. Activation of the aryl hydrocarbon receptor dampens the severity of inflammatory skin conditions. Immunity 40, 989-1001 (2014).
Monteleone, I. et al. Aryl hydrocarbon receptor-induced signals upregulate IL-22 production and inhibit inflammation in the gastrointestinal tract. Gastroenterology 141, 237-248 (2011).
Ala, M. Tryptophan metabolites modulate inflammatory bowel disease and colorectal cancer by affecting immune system. Int. Rev. Immunol. 41, 326-345 (2022).
Schirmer, M., Garner, A., Vlamakis, H. & Xavier, R. J. Microbial genes and pathways in inflammatory bowel disease. Nat. Rev. Microbiol. 17, 497-511 (2019).
Engevik, M. A. et al. Bifidobacterium dentium-derived y-glutamylcysteine suppresses ER-mediated goblet cell stress and reduces TNBS-driven colonic inflammation. Gut Microbes 13, 1-21 (2021).
Liu, Y. et al. TLR4 regulates RORgammat(+) regulatory T-cell responses and susceptibility to colon inflammation through interaction with Akkermansia muciniphila. Microbiome 10, 98 (2022).
Cox, L. M. et al. Corrigendum: description of two novel members of the family Erysipelotrichaceae: Ileibacterium valens gen. nov., sp. nov. and Dubosiella newyorkensis, gen. nov., sp. nov., from the murine intestine, and emendation to the description of Faecalibacterium rodentium. Int. J. Syst. Evol. Microbiol. 67, 4289 (2017).
Rodrigues, V. F. et al. Akkermansia muciniphila and gut immune system: a good friendship that attenuates inflammatory bowel disease, obesity, and diabetes. Front. Immunol. 13, 934695 (2022).
Kuczma, M. P. et al. Commensal epitopes drive differentiation of colonic T(regs). Sci. Adv. 6, eaaz3186 (2020).
Zagato, E. et al. Endogenous murine microbiota member Faecalibaculum rodentium and its human homologue protect from intestinal tumour growth. Nat. Microbiol. 5, 511-524 (2020).
Le Poul, E. et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 278, 25481-25489 (2003).
Maslowski, K. M. et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 461, 1282-1286 (2009).
Cervenka, I., Agudelo, L. Z. & Ruas, J. L. Kynurenines: tryptophan’s metabolites in exercise, inflammation, and mental health. Science 357, eaaf9794 (2017).
Mellor, A. L. & Munn, D. H. IDO expression by dendritic cells: tolerance and tryptophan catabolism. Nat. Rev. Immunol. 4, 762-774 (2004).
Gargaro, M. et al. Indoleamine 2,3-dioxygenase 1 activation in mature cDC1 promotes tolerogenic education of inflammatory cDC2 via metabolic communication. Immunity 55, 1032-1050.e14 (2022).
Yuan, H. et al. Lysine catabolism reprograms tumour immunity through histone crotonylation. Nature 617, 818-826 (2023).
Yang, L. et al. Amino acid metabolism in immune cells: essential regulators of the effector functions, and promising opportunities to enhance cancer immunotherapy. J. Hematol. Oncol. 16, 59 (2023).
Liu, C. et al. Baicalein restores the balance of Th17/Treg cells via aryl hydrocarbon receptor to attenuate colitis. Mediat. Inflamm. 2020, 5918587 (2020).
Zhuang, Z. et al. GWAS-associated bacteria and their metabolites appear to be causally related to the development of inflammatory bowel disease. Eur. J. Clin. Nutr. 76, 1024-1030 (2022).
Franzosa, E. A. et al. Gut microbiome structure and metabolic activity in inflammatory bowel disease. Nat. Microbiol. 4, 293-305 (2019).
Schirmer, M. et al. Dynamics of metatranscription in the inflammatory bowel disease gut microbiome. Nat. Microbiol. 3, 337-346 (2018).
Zhang, Y. et al. Discovery of bioactive microbial gene products in inflammatory bowel disease. Nature 606, 754-760 (2022).
Michaudel, C. & Sokol, H. The gut microbiota at the service of immunometabolism. Cell Metab. 32, 514-523 (2020).
Maccaferri, S. et al. Rifaximin modulates the colonic microbiota of patients with Crohn’s disease: an in vitro approach using a continuous culture colonic model system. J. Antimicrob. Chemother. 65, 2556-2565 (2010).
Abraham, B. & Quigley, E. M. M. Antibiotics and probiotics in inflammatory bowel disease: when to use them? Frontline Gastroenterol. 11, 62-69 (2020).
Li, Q. et al. Enterobacter ludwigii protects DSS-induced colitis through choline-mediated immune tolerance. Cell Rep. 40, 111308 (2022).
Rath, H. C. et al. Different subsets of enteric bacteria induce and perpetuate experimental colitis in rats and mice. Infect. Immun. 69, 2277-2285 (2001).
Li, H. et al. Bifidobacterium spp. and their metabolite lactate protect against acute pancreatitis via inhibition of pancreatic and systemic inflammatory responses. Gut Microbes 14, 2127456 (2022).
Rodriguez-Daza, M. C. et al. Berry polyphenols and fibers modulate distinct microbial metabolic functions and gut microbiota enterotype-like clustering in obese mice. Front. Microbiol. 11, 2032 (2020).
Sun, P. et al. Comparative metagenomics and metabolomes reveals abnormal metabolism activity is associated with gut microbiota in Alzheimer’s disease mice. Int. J. Mol. Sci. 23, 11560 (2022).
Agus, A., Planchais, J. & Sokol, H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe 23, 716-724 (2018).
Bacsi, S. G. & Hankinson, O. Functional characterization of DNAbinding domains of the subunits of the heterodimeric aryl hydrocarbon receptor complex imputing novel and canonical basic helix-loop-helix protein-DNA interactions. J. Biol. Chem. 271, 8843-8850 (1996).
Sinclair, L. V., Neyens, D., Ramsay, G., Taylor, P. M. & Cantrell, D. A. Single cell analysis of kynurenine and system L amino acid transport in T cells. Nat. Commun. 9, 1981 (2018).
Rothhammer, V. et al. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat. Med. 22, 586-597 (2016).
Sanmarco, L. M. et al. Identification of environmental factors that promote intestinal inflammation. Nature 611, 801-809 (2022).
Allen, I. C. et al. NLRP12 suppresses colon inflammation and tumorigenesis through the negative regulation of noncanonical NFkappaB signaling. Immunity 36, 742-754 (2012).
Weigmann, B. et al. Isolation and subsequent analysis of murine lamina propria mononuclear cells from colonic tissue. Nat. Protoc. 2, 2307-2311 (2007).
Steed, A. L. et al. The microbial metabolite desaminotyrosine protects from influenza through type I interferon. Science 357, 498-502 (2017).
Matsuzawa-Ishimoto, Y. et al. Autophagy protein ATG16L1 prevents necroptosis in the intestinal epithelium. J. Exp. Med. 214, 3687-3705 (2017).
Yui, S. et al. Functional engraftment of colon epithelium expanded in vitro from a single adult Lgr5(+) stem cell. Nat. Med. 18, 618-623 (2012).
Acknowledgements
We appreciate Dr. Yajing Wang of China Pharmaceutical University and Dr. Bin Li of Shanghai Jiao Tong University School of Medicine for providing valuable and Foxp3-DTR mice. We thank Dr. Lingdong Xu of Laboratory Animal Center, Zhejiang University for fundamental technical support with performing confocal microscopy. The professional editing service NB Revisions was used for technical preparation of the text prior to submission. This work is supported by grants from the National Natural Science Foundation of China (No. 32172864 and No. U21A2O261) and the National Key Research and Development Plan of China (2022YFD1800804) to S.J.Z.
Author contributions
S.J.Z. and Y.Z. designed the experiments. Y.Z., S.T., X.J., J.W., J.G., X.S., S.S., G.W., J.Q. ZB.Z., H.C. and ZY.Z. performed the experiments. Y.Z. and J.M. conducted bioinformatics analysis. S. S. and X.S. collected samples and data. S. C coordinated the project. L.Z. commented on and revised drafts of the manuscript. S.J.Z., Y.Z. and S.T. wrote the paper. S.J.Z. supervised research, coordination, and strategy.
Correspondence and requests for materials should be addressed to Shu Jeffrey Zhu.
Peer review information Nature Communications thanks Bahtiyar Yilmaz and the other, anonymous, reviewers for their contribution to the peer review of this work. A peer review file is available.
Department of Veterinary Medicine, College of Animal Sciences, Zhejiang University, Hangzhou, Zhejiang 310058, PR China. Department of Cardiology, The First Affiliated Hospital of Guangdong Pharmaceutical University, Guangzhou, Guangdong 510080, PR China. Department of Critical Care Medicine, Sir Run Run Shaw Hospital, Zhejiang University School of Medicine, Hangzhou, Zhejiang 310016, PR China. College of Veterinary Medicine, Qingdao Agricultural University, Qingdao, Shandong 266109, PR China. Shaoxing People’s Hospital, Zhejiang University Shaoxing Hospital, Shaoxing, Zhejiang 312000, PR China. College of Veterinary Medicine, Jilin Provincial Engineering Research Center of Animal Probiotics, Jilin Provincial Key Laboratory of Animal Microecology and Healthy Breeding, Jilin Agricultural University, Changchun 130118, PR China. e-mail: shuzhu@zju.edu.cn