طرق جديدة لتقييم السمية العصبية التنموية والبالغة للاستخدام التنظيمي: مشروع حزمة العمل 5 من PARC New approach methods to assess developmental and adult neurotoxicity for regulatory use: a PARC work package 5 project
تمارا تال، أودفار ميهر، إلين فريتشي، جويل رويغ، كاي كراينن، وآخرون. طرق جديدة لتقييم السمية العصبية في مراحل النمو والبالغين للاستخدام التنظيمي: مشروع حزمة العمل 5 من PARC. الحدود في علم السموم، 2024، 6، الصفحات 1359507. 10.3389/ftox.2024.1359507 . hal-04679936
HAL هو أرشيف مفتوح متعدد التخصصات لإيداع ونشر الوثائق البحثية العلمية، سواء كانت منشورة أم لا. قد تأتي الوثائق من مؤسسات التعليم والبحث في فرنسا أو في الخارج، أو من مراكز البحث العامة أو الخاصة.
الأرشيف المفتوح متعدد التخصصات HAL مخصص لإيداع وتوزيع الوثائق العلمية على مستوى البحث، سواء كانت منشورة أم لا، والتي تصدر عن مؤسسات التعليم والبحث الفرنسية أو الأجنبية، أو المختبرات العامة أو الخاصة.
الوصول المفتوح
تم التحرير بواسطة
لينا سميرنوفا جامعة جونز هوبكنز، الولايات المتحدة
تمت المراجعة بواسطة
ساندرا مكبرايد الأنظمة الاجتماعية والعلمية، الولايات المتحدة
طرق جديدة لتقييم السمية العصبية في مراحل النمو والبالغين للاستخدام التنظيمي: مشروع حزمة العمل 5 من PARC
تمارا تالأودفار ميهرإلين فريتشهجويل رويغكاي كرايننكيارا أيويلو-هولدنكارولين أجريلوباتريك ج. بابينبيات إ. إيشيرهوبرت ديرفنكاتي هيلستينكريستين دولفاإلين هيسلهارم ج. هيوسينكفيلديافور هادجييفسلمى هوريمكارولينا ياجييلوبياتا يودزينسكانيلس كلوفرأنجا كنول-غيليدابريتا أ. كونهمارسيل ليستمالين ليسليينجان ل. ليكفيرينك مولرجون ك. كولبورنوينفريد نيوهاوسجورجيا بالوكّابيتينّا سيغرإلكا شاريكنستيفان شولتزأولا سبيوثمونيكا توريس-رويزوكريستينا بارتمانمركز هلمهولتز لأبحاث البيئة – UFZ، قسم أبحاث المواد الكيميائية في البيئة، لايبزيغ، ألمانيا،جامعة لايبزيغ، كلية الطب، لايبزيغ، ألمانياالمعهد النرويجي للصحة العامة – NIPH، قسم السموم الكيميائية، أوسلو، النرويجمعهد لايبنيز للبحوث في الطب البيئي، دوسلدورف، ألمانيادي إن توكس GmbH، دوسلدورف، ألمانياالمركز السويسري لعلم السموم البشرية التطبيقية، جامعة بازل، بازل، سويسراجامعة أوبسالا، قسم البيولوجيا الكائنات الحية، أوبسالا، السويدالوكالة الأوروبية للمواد الكيميائية (ECHA)، هلسنكي، فنلندا،المعهد الفيدرالي الألماني لتقييم المخاطر (BfR)، برلين، ألمانياجامعة بوردو، المعهد الوطني للصحة والبحث الطبي (INSERM)، الأمراض النادرة: علم الوراثة والتمثيل الغذائي (MRGM)، بيساك، فرنساجامعة أوسلو، قسم علم الأدوية والعلوم الصيدلانية الحيوية، كلية الصيدلة، أوسلو، النرويجالمعهد الهولندي للصحة العامة والبيئة (RIVM)، مركز حماية الصحة، بيلثوفن، هولنداجامعة برمنغهام، مركز البحث البيئي والعدالة، برمنغهام، المملكة المتحدة،الجامعة النرويجية لعلوم الحياة (NMBU)، كلية الطب البيطري، آås، النرويج،جامعة غدانسك، مختبر الكيمياء البيئية المعلوماتية، غدانسك، بولندا،جامعة الطب البيطري هانوفر، المؤسسة، معهد جودة وسلامة الغذاء، هانوفر، ألمانياجامعة كونستانس، علم السموم في المختبر والطب الحيوي/CAAT-أوروبا، كونستانس، ألمانيامعهد النمسا للتكنولوجيا GmbH، وحدة الكفاءة للتشخيصات الجزيئية، مركز الصحة والموارد الحيوية، فيينا، النمسا،جامعة الدانوب الخاصة، كلية طب الأسنان والطب، قسم الطب، كريمس، النمساجامعة أوبسالا ومختبر العلوم من أجل الحياة، قسم علوم الأدوية الحيوية، أوبسالا، السويدمعهد الصحة كارلوس الثالث (ISCIII)، المركز الوطني للصحة البيئية (CNSA)، وحدة السموم البيئية، ماخاداهوندا، إسبانيا
في السياق التنظيمي الأوروبي، تعتبر الدراسات الحية على القوارض المصدر الرئيسي لمعلومات السمية العصبية. على الرغم من أنها تشكل حجر الزاوية في التقييمات السمية العصبية، إلا أنها مكلفة وموضوع نقاش أخلاقي. بينما يتوقع الجمهور أن تكون المواد الكيميائية والمنتجات آمنة للأنظمة العصبية النامية والناضجة، فإن عددًا كبيرًا من المواد الكيميائية المتداولة لم يتم تقييمها، أو تم تقييمها بشكل محدود، من حيث قدرتها على التسبب في السمية العصبية. وبالتالي، هناك دفع اجتماعي نحو استبدال نماذج الحيوانات بأساليب في المختبر أو بدائل أخرى. يمكن أن تسهم الأساليب الجديدة (NAMs) في قاعدة المعرفة التنظيمية، وزيادة سلامة المواد الكيميائية، وتحديث تقييم المخاطر والأخطار الكيميائية. بشرط أن تصل إلى مستوى مقبول من الأهمية التنظيمية والموثوقية، قد تكون NAMs
الملخص
تعتبر بدائل لدراسات معينة في الجسم الحي. تتناول الشراكة الأوروبية لتقييم المخاطر الناتجة عن المواد الكيميائية (PARC) التحديات المتعلقة بتطوير وتنفيذ نماذج التقييم البديلة (NAMs) في تقييم مخاطر المواد الكيميائية. بالتعاون مع الوكالات التنظيمية، يهدف المشروع 5.2.1e (السمية العصبية) إلى تطوير وتقييم NAMs للسمية العصبية التنموية (DNT) والسمية العصبية لدى البالغين (ANT) وفهم نطاق تطبيق NAMs معينة للكشف عن اضطراب الغدد الصماء والاضطراب الجيني. لتسريع وقت الاختبار وتقليل التكاليف، نقوم بتحديد مؤشرات مبكرة للتأثيرات التي تظهر لاحقًا. في النهاية، سنجمع بين بطاريات اختبارات السمية العصبية التنموية من الجيل الثاني والسمية العصبية لدى البالغين من الجيل الأول، والتي تهدف كلاهما إلى تزويد المقيّمين التنظيميين للمخاطر والمخاطر وأصحاب المصلحة في الصناعة بأدوات تقييم المخاطر والمخاطر من الجيل التالي التي تكون قوية وسريعة ومنخفضة التكلفة ومفيدة.
الكلمات الرئيسية
طريقة جديدة (NAM)، السمية العصبية التنموية (DNT)، السمية العصبية لدى البالغين (ANT)، DNT-IVB، سمكة الزرد، نطاق القابلية للتطبيق
1 الشراكة الأوروبية لتقييم المخاطر الناتجة عن المواد الكيميائية (PARC)
تهدف الشراكة الأوروبية لتقييم المخاطر الناتجة عن المواد الكيميائية (PARC) إلى تطوير أدوات تقييم المخاطر والمخاطر الكيميائية من الجيل التالي لحماية صحة الإنسان والبيئة بشكل أفضل (ماركس-ستولتينغ وآخرون، 2023). طموح رئيسي للمشروع هو تطوير طرق جديدة للتقييم (NAMs) لتقييم مخاطر صحة الإنسان التي تغطي السمية العصبية التنموية (DNT) والسمية العصبية لدى البالغين (ANT) واضطراب هرمون الغدة الدرقية والسمية المناعية والمواد المسرطنة غير الجينية. تهدف حزمة العمل 5.2.1e إلى تحسين NAMs الحالية، وتطوير أخرى جديدة، وإنتاج بطاريات اختبار من الجيل الأول لـ ANT والجيل الثاني لـ DNT. ستعتمد NAMs التي سيتم تطويرها على الأحداث الرئيسية (KE) كما هو محدد في إطار مسار النتائج السلبية (AOP) (أنكلي وآخرون، 2010؛ ليست وآخرون، 2017؛ سبينو وآخرون، 2021). يتم تنفيذ العمل من قبل اتحاد يضم أكثر من 25 خبيرًا من 10 مؤسسات بحثية في الاتحاد الأوروبي ومؤسستين شريكتين في دول غير تابعة للاتحاد الأوروبي.
قد يشكل التعرض للمواد الكيميائية خطرًا على الأنظمة العصبية النامية والناضجة
يمكن أن يؤثر التعرض للمواد الكيميائية سلبًا على تطوير ووظيفة النظام العصبي عبر جميع مراحل الحياة (كوستا وآخرون، 2008؛ جوردانو وكوستا، 2012؛ ب؛ جراندجيان ولاندريغان، 2006). يمكن أن تؤثر الآثار السلبية المعتمدة على المواد الكيميائية الناتجة عن تعرض النسل النامي (بما في ذلك في الرحم وبعد الولادة) حتى وقت النضج الجنسي على النظام العصبي النامي (كوستا وآخرون، 2004). يمكن أن تكون “السمية العصبية التنموية” (DNT) طويلة الأمد، تمتد بعيدًا عن فترة التعرض، ويمكن أن تختلف عبر العمر (إريكسون وآخرون، 1998؛ سبالدينغ وآخرون، 2013). لاحظ أن أي نوع من التأثيرات السمية العصبية أثناء التطور هو مصدر قلق تنظيمي وذو صلة بتحديد المخاطر التنموية. بالمقابل، عندما يتعرض النظام العصبي الناضج للمواد الكيميائية السامة العصبية، يمكن أن تكون آثار السمية العصبية لدى البالغين (ANT) فورية أو قد تتطور تدريجيًا وتكون طويلة الأمد. اعتمادًا على نوع
أثر ANT، قد يكون أيضًا قابلًا للعكس (سبنسر ولين، 2014). تعتبر السمية العصبية الكبيرة و/أو الشديدة، سواء كانت قابلة للعكس أو غير قابلة للعكس، فورية أو متأخرة، مصدر قلق تنظيمي.
نظرًا لحساسية النظام العصبي النامي، قد يؤدي التعرض لتركيزات منخفضة من بعض المواد الكيميائية إلى اضطرابات هيكلية ووظيفية (رايس وبارون، 2000؛ جراندجيان ولاندريغان، 2014؛ بينيت وآخرون، 2016). يمكن أن تؤدي الاضطرابات العصبية التنموية بما في ذلك اضطراب طيف التوحد، والإعاقة الذهنية، واضطراب نقص الانتباه/فرط النشاط (ADHD)، واضطرابات الحركة العصبية التنموية (بما في ذلك اضطرابات التشنج)، واضطرابات التعلم المحددة إلى عواقب اجتماعية واقتصادية مدى الحياة، بما في ذلك انخفاض الإنتاجية الاقتصادية أو زيادة الحاجة إلى دعم التعلم في المدارس (ب. جراندجيان ولاندريغان، 2006). بينما تم الاعتراف بأن التقديرات كانت غير مؤكدة، فإنسنوات الحياة المعدلة حسب الإعاقة (DALYs) المتعلقة بالأمراض العصبية التنموية قد تكون نتيجة للتعرض للمواد الكيميائية (وبغض النظر عن الاستعداد/الحساسية الجينية للشخص)، معDALYs عندما تم دمج التعرض للمواد الكيميائية مع الاستعداد الجيني الأساسي (EC 2019). تم بناء هذا التقدير على تقييم “من أعلى إلى أسفل” لتأثيرات الاضطرابات العصبية التنموية المنتشرة من منظمة الصحة العالمية (WHO) وتقدير أنيعود إلى التعرض البيئي لمركبات الإرث مثل الرصاص وغيرها من الملوثات البيئية (EC 2019). وبالتالي، من المتوقع أن تكون هناك فوائد اجتماعية واقتصادية ملحوظة من خلال تحديد المواد المعروفة أو المفترضة أنها تسبب DNT ومنع التعرض لاحقًا (بيلانجر وآخرون، 2013؛ بيلانجر وآخرون، 2015).
بعد فترة التطور، قد يؤدي التعرض الحاد و/أو المزمن للمواد الكيميائية البيئية إلى استجابات سامة في الأنظمة العصبية الطرفية و/أو المركزية وقد تم اقتراح أن التعرض لوكلاء كيميائيين محددين قد يزيد من احتمال تطوير اضطرابات التنكس العصبي مثل مرض باركنسون ومرض الزهايمر، أو الخرف (لاندريغان وآخرون، 2005؛ تانر وآخرون، 2014؛ أوكلفورد وآخرون). علاوة على ذلك، تم الاشتباه في أن التعرض لبعض المواد الكيميائية مرتبط بالاكتئاب والقلق لدى المراهقين والبالغين، وغيرها من الاضطرابات النفسية في عدد من المنشورات الأكاديمية (ديكرسون وآخرون، 2020؛ هولاندر وآخرون، 2020؛ جاكوبسون وآخرون، 2022؛ روكوف وآخرون، 2022؛ أونغ وآخرون، 2023). في دراسة لـ 22 جردًا كيميائيًا من 19 دولة ومنطقة، تم تحديد أكثر من 350,000 مادة كيميائية ومخاليط من المواد الكيميائية تم تسجيلها للإنتاج وربما قيد الاستخدام (وانغ وآخرون، 2020). على الرغم من المعرفة المتعلقة بالتأثيرات الضارة المحتملة للمواد الكيميائية البيئية على الأنظمة العصبية النامية والناضجة (ب. جراندجيان ولاندريغان، 2006)، يُفهم أن عددًا محدودًا فقط من المواد الفريدة قد تم اختبارها لـ DNT باستخدام دراسات إرشادات اختبار منظمة التعاون والتنمية الاقتصادية (OECD). (OECD، 2008a؛ ماكريس وآخرون، 2009؛ ساشانا وآخرون، 2019؛ كروفتون وموندي، 2021).
مشهد السياسات والتنظيمات
يصف الاتفاق الأخضر للاتحاد الأوروبي تأثيرات الصحة في خطة العمل صفر تلوث، وقد أبرزت المفوضية الأوروبية مؤخرًا اهتمامها بزيادة الجهود لحماية ضد المواد الكيميائية الأكثر ضررًا، من خلال استكشاف المزيد من إمكانيات إدارة المخاطر للمواد السامة العصبية والمثبطة للغدد الصماء (التي تم ربطها بـ DNT) (المفوضية الأوروبية، 2020). في الاتحاد الأوروبي، هناك العديد من اللوائح ذات الصلة سارية. على سبيل المثال، قبل دخول السوق أو الحصول على الموافقة كمادة نشطة مبيدة للجراثيم أو مبيدة للحشرات، يجب الوفاء بالحد الأدنى من متطلبات البيانات الموضحة في اللائحة ذات الصلة بالاتحاد الأوروبي (من بين شروط أخرى). يمكن أن تتطلب لوائح الاتحاد الأوروبي بشأن منتجات حماية النباتات (Reg EC 1107؛ البرلمان الأوروبي والمجلس، 2009) والمبيدات (Reg EC 528؛ البرلمان الأوروبي والمجلس، 2012) اختبار DNT/ANT كجزء من متطلبات البيانات. بموجب لائحة منتجات المبيدات البيولوجية في الاتحاد الأوروبي (Reg EC 528؛ البرلمان الأوروبي والمجلس، 2012)، أصبح اختبار DNT المحدد، على سبيل المثال، OECD TG 426، مؤخرًا متطلبًا معلوماتيًا إلزاميًا لعملية الموافقة للمواد النشطة المبيدة للجراثيم. بموجب REACH (البرلمان الأوروبي والمجلس، 2006)، أنشأت اللائحة الأوروبية لحماية صحة الإنسان والبيئة من المواد الكيميائية الضارة، يعتمد مستوى المعلومات المطلوبة لتحديد الخصائص السامة العصبية المحتملة (DNT/ANT) حاليًا على الكمية وتحديد المخاوف المحددة التي قد تؤدي إلى اختبارات DNT أو ANT. تُستخدم المعلومات المتاحة لتطبيق تصنيفات المخاطر المناسبة، وفقًا للمعايير المحددة في لائحة CLP (Reg EC 1272؛ البرلمان الأوروبي والمجلس، 2008)، لإبلاغ الخصائص الخطرة للمواد الكيميائية. ثم يعمل التصنيف وفقًا لـ CLP على تحفيز أو إبلاغ الإجراءات العلاجية في تشريعات أخرى للسيطرة على الخطر. لا تتطلب لائحة CLP (المادتان 7 و8) TGs لـ DNT أو ANT مباشرة ولكنها تستخدم جميع البيانات المتاحة التي تم إنشاؤها في سياق التشريعات ذات الصلة و/أو المتاحة بخلاف ذلك في المجال العام. في الحالات التي لا تتوفر فيها مثل هذه البيانات لإبلاغ خطر معين، قد يتم إجراء الاختبارات تحت ظروف معينة بما في ذلك الشرط الذي ينص على أن الاختبارات على الحيوانات يجب أن تُجرى فقط حيث لا توجد بدائل أخرى، والتي توفر موثوقية وجودة بيانات كافية، ممكنة. وهذا يعني الدعم ويقدم فرصة لتطوير وتأكيد وتنفيذ NAMs.
في إطار لائحة CLP، يتم تناول المواد التي تحتوي على DNT تحت فئة خطر السمية الإنجابية (التنموية) ويتم تناول آثار ANT تحت السمية المستهدفة المحددة (STOT)، إما التعرض الفردي (SE) أو التعرض المتكرر (RE)، اعتمادًا على ما إذا كانت الآثار ناتجة عن تعرض فردي أو متكرر، على التوالي. تم إجراء مراجعة حديثة للائحة CLP يتضمن فئة جديدة من المخاطر تتعلق باضطراب الغدد الصماء، والتي تشمل التأثيرات السلبية الناتجة عن النشاط الغدي على الجهاز العصبي النامي (والناضج) (اللائحة EC 1272؛ (البرلمان الأوروبي والمجلس، 2023). وفقًا للمعايير الجديدة، سيتم تصنيف الفئة الأولى من اضطراب الغدد الصماء بشكل كبير استنادًا إلى أدلة من واحدة على الأقل من المصادر التالية: بيانات بشرية؛ بيانات حيوانية؛ بيانات غير حيوانية توفر قدرة تنبؤية معادلة مثل البيانات البشرية أو البيانات الحيوانية (اللائحة EC 1272؛ البرلمان الأوروبي والمجلس، 2023). وبالتالي، تسمح فئة المخاطر الجديدة باستخدام NAMs مباشرة لغرض هذا التصنيف المحدد عندما يتم استيفاء المعايير.
3.1 DNT/ANT في اللوائح الكيميائية الحالية
المعلومات اللازمة لتلبية متطلبات البيانات بموجب REACH وBPR تُقدم عادةً من خلال دراسات OECD TG الحية المحددة في القسم المعني من اللائحة المعمول بها، ولكن هناك أيضًا إمكانيات محددة للتكيف (بشكل أكثر تحديدًا، التنازل عن البيانات). يمكن أن تشمل هذه الإمكانيات التكيف غير الحيواني و/أو استخدام المعلومات الموجودة المستمدة من مواد مشابهة من خلال نهج القراءة المتوازية (البرلمان الأوروبي والمجلس، 2006). ومع ذلك، حيث يتم الحصول على بيانات حول صحة الإنسان وخصائص البيئة من خلال التكيفات لمتطلبات البيانات، تنطبق شروط معينة. تم تحديد الشروط الخاصة بالتكيفات باستخدام بيانات in vitro بموجب REACH في الملحق الحادي عشر، القسم 1.4. في سياق التكيف عبر القراءة المتوازية (REACH الملحق الحادي عشر، القسم 1.5)، تنطبق مرة أخرى شروط تقييدية معينة فيما يتعلق بالبيانات التي تُعلم مباشرةً عن الخطر. ومع ذلك، بالنسبة لاستقراء مثل هذه البيانات إلى مواد أخرى، هناك فرصة واضحة لاستخدام NAMs كمعلومات داعمة لإظهار التشابه في خصائص المواد المعنية.
يجب أيضًا ملاحظة أنه، اعتمادًا على اللوائح المعمول بها، قد لا تكون الاختبارات الجديدة ضرورية إذا كانت البيانات المتاحة كافية بالفعل للغرض التنظيمي كما هو محدد في اللوائح المحددة. على سبيل المثال، لا ينبغي إجراء دراسة DNT بموجب اللائحة الخاصة بمبيدات الآفات إذا كانت البيانات المتاحة تشير بالفعل إلى أن المادة تسبب سمية تطورية وتفي بالمعايير لتصنيفها على أنها سامة للتكاثر من الفئة 1A أو 1B: قد تضر بالجنين (H360D)، وهذه البيانات المتاحة كافية لدعم تقييم مخاطر قوي (البرلمان الأوروبي والمجلس، 2012).
مجموعة من دراسات منظمة التعاون والتنمية الاقتصادية (OECD TG)، بما في ذلك دراسات الجرعة الواحدة (مثل OECD TG 402، 403، 420، 423، 425) و/أو دراسات سمية الجرعات المتكررة (مثل OECD TG 407، 408، 421، 422، 414، 443 في غياب مجموعات DNT) قد تُفيد في تقييم ANT أو DNT بناءً على علامات سريرية مثل الشلل، التشنجات، نقص التنسيق، أو عدم التوازن أو علم الأمراض العصبية و/أو التغيرات في وزن الدماغ (الجدول 1). يمكن تقييم DNT بشكل أكثر شمولاً باستخدام اختبارات مخصصة مثل OECD TG 426 أو في مجموعة DNT (المجموعات 2A و2B) من دراسة سمية التكاثر الممتدة من جيل واحد (EOGRTS، OECD TG 443). تشمل معايير OECD المخصصة لدراسة ANT OECD TG 424، 418 و419. بموجب REACH، يمكن استخدام ANT أو آليات/طرق عمل محددة مرتبطة بالسمية العصبية (التطورية) لتحفيز دراسات DNT محددة. يمكن أن تحتوي المواد في الطعام المخصص للرضع أيضًا على
الجدول 1 وصف الدراسات الحالية لإرشادات منظمة التعاون والتنمية الاقتصادية التي تشمل السمية العصبية كنقطة نهاية.
إرشادات الاختبار
النقطة النهائية الأساسية
نقطة نهاية السموم العصبية
الأنواع المفضلة
فترة الإدارة
نقاط النهاية غير السلوكية المتعلقة بالسمية العصبية
نقاط النهاية السلوكية
مرجع
OECD TG 402
سمية جلدية
النمل (حاد)
جرذ
البالغين )
لا (فقط تشريح قذر)
النشاط الحركي الذاتي والنظام العصبي المركزي ونمط السلوك الحركي
منظمة التعاون والتنمية الاقتصادية (2017)
OECD TG 403
سمية الاستنشاق
النمل (حاد)
جرذ
البالغون (4 ساعات)
لا (فقط تشريح قذر)
النشاط الحركي والسلوكي للجهاز العصبي الذاتي والمركزي
منظمة التعاون والتنمية الاقتصادية (2009)
OECD TG 407
سمية فموية / اضطراب الغدد الصماء
النمل (مزمن)
جرذ
البالغون (يوميًا – 28 يومًا)
وزن الدماغ، علم الأمراض النسيجية للدماغ، الحبل الشوكي، والعصب الوركي
استجابة حسية للمؤثرات، قوة قبضة الأطراف، النشاط الحركي
منظمة التعاون والتنمية الاقتصادية (2008ب)
OECD TG 408
سمية فموية / اضطراب الغدد الصماء
النمل (مزمن)
جرذ
البالغون (يوميًا – 90 يومًا)
وزن الدماغ، علم الأمراض النسيجية للدماغ، الحبل الشوكي، والعصب الوركي
استجابة حسية للمؤثرات، قوة قبضة الأطراف، النشاط الحركي، النشاط اللاإرادي
منظمة التعاون والتنمية الاقتصادية (2018أ)
OECD TG 418
السمية العصبية (مواد OP)
النمل (حاد)
دجاجة
البالغون (جرعة واحدة)
علم الأمراض العصبية للجهاز العصبي المركزي والمحيطي، وأنشطة NTE و AchE
الاضطرابات السلوكية، عدم التنسيق الحركي، والشلل
منظمة التعاون والتنمية الاقتصادية (1995أ)
OECD TG 419
السمية العصبية (مواد OP)
النمل (مزمن)
دجاجة
البالغينأيام)
علم الأمراض العصبية للجهاز العصبي المركزي والمحيطي، وأنشطة NTE و AchE
الاضطرابات السلوكية، عدم التنسيق الحركي، والشلل
منظمة التعاون والتنمية الاقتصادية (1995ب)
OECD TG 420
سمية فموية
النمل (حاد)
جرذ
البالغون (جرعة واحدة)
لا (فقط تشريح قذر)
النشاط الحركي الجسدي وأنماط السلوك
منظمة التعاون والتنمية الاقتصادية (2002أ)
OECD TG 423
سمية فموية
النمل (حاد)
جرذ
البالغون (جرعة واحدة)
لا (فقط تشريح قذر)
النشاط الحركي الجسدي وأنماط السلوك
منظمة التعاون والتنمية الاقتصادية (2002ب)
OECD TG 424
سمية عصبية
النمل (مزمن)
جرذ
البالغينأيام)
علم الأمراض العصبية للجهاز العصبي المركزي والمحيطي
استجابة حسية للمؤثرات، قوة قبضة الأطراف، النشاط الحركي
منظمة التعاون والتنمية الاقتصادية (1997)
OECD TG 425
سمية فموية
النمل (حاد)
جرذ
البالغون (جرعة واحدة)
لا (فقط تشريح مروع)
النشاط الحركي الجسدي وأنماط السلوك
منظمة التعاون والتنمية الاقتصادية (2022)
OECD TG 426
سمية عصبية
دنت (مزمن)
جرذ
الحمل والرضاعة
تشوهات تطورية، أوزان الدماغ، علم الأمراض العصبية
النشاط الحركي، الوظيفة الحركية والحسية، التعلم والذاكرة
منظمة التعاون والتنمية الاقتصادية (2007)
OECD TG 443
سمية الإنجاب
دنت (مزمن)
جرذ
قبل التزاوج – الجراء
علم الأمراض العصبية النسيجية، وزن الدماغ والمورفومترية
NTE: استرواز الهدف العصبي؛ AchE: أستيل كولينستراز؛ M: ذكر؛ F: أنثى. تحفيز التحقيقات لتقييم احتمالية DNT (اللجنة العلمية للهيئة الأوروبية لسلامة الأغذية وآخرون، 2017).
نظرًا لأن تطوير الجهاز العصبي يبدأ قبل الولادة ويستمر في التطور خلال فترة المراهقة، ليصل إلى مستويات البالغين من الناقلات العصبية، ومرونة المشابك، والتغليف النخاعي، والمادة الرمادية في حوالي سن 20 لدى البشر وحوالي PND60 لدى الجرذان (Semple et al.، 2013)، فإنه من الضروري تنفيذ التعرض طوال فترة التطور بأكملها لتحسين الفرص للتعرف على المواد السامة العصبية التنموية. في اختبار توجيه منظمة التعاون والتنمية الاقتصادية TG 426، يتعرض النسل كحد أدنى من وقت الزرع (بدءًا من يوم الحمل (GD) 6) طوال فترة الرضاعة (حتى اليوم ما بعد الولادة (PND) 21). في المجموعة 2 A من دراسة EOGRTS، يتعرض النسل عبر الأم في الرحم، من خلال الرضاعة ومباشرة على الأقل بعد الفطام حتى إنهاء الدراسة في حوالي PND 66-77. تشمل المعايير المقيمة للتسمم العصبي التنموي في دراسات DNT المحددة كحد أدنى (اعتمادًا على TG منظمة التعاون والتنمية الاقتصادية) النشاط الحركي، الوظيفة الحركية والحسية، والتعلم التفاعلي. والذاكرة (فقط في OECD TG 426 كاختبار قياسي)، وزن الدماغ، وعلم الأمراض النسيجية للجهاز العصبي المركزي والمحيطي (تسوجي وكروفتون، 2012). نظرًا لأن دراسات DNT المخصصة غالبًا ما تكون دراسات معقدة تستخدم القوارض، فهي تتطلب موارد كبيرة من حيث الوقت والتكاليف وعدد الحيوانات (كروفتون وآخرون، 2012؛ سميرنوفا وآخرون، 2014) وقد تم اختبار عدد محدود فقط من المواد الكيميائية لدراسة DNT باستخدام دراسات OECD TG DNT (OECD، 2008a؛ ماكريس وآخرون، 2009؛ ساشانا وآخرون، 2019؛ كروفتون وموندي، 2021). بالإضافة إلى ذلك، تم توثيق التباين في اختبارات السمية العصبية للقوارض (تسوجي وكروفتون، 2012؛ تيرون وبينيكو، 2018؛ ساشانا وآخرون، 2019؛ باباريلا وآخرون، 2020؛ هاري وآخرون، 2022) مما يشير إلى الحاجة لتحديد وتحسين وتوحيد طرق الاختبار الفردية المستخدمة كجزء من OECD TG. كما يبرز الحاجة إلى اختبارات جديدة تفتقر إلى التباين الزائد المرتبط بدراسات الإرشادات الحية لتقييم DNT.
تشكل المخاطر التي ت posed by agents neurotoxic (التطويرية) غير المعروفة وعدد محدود من أنظمة الاختبار الفعالة من حيث التكلفة وفي الوقت المناسب (أي NAMs) الأساس لهذا المشروع PARC حيث الطموح هو إنشاء مجموعة اختبارات محسنة في المختبر وبديلة لاختبارات DNT ومجموعة اختبارات ANT من الجيل الأول في المختبر. لقد وجدت الحاجة إلى تطوير تقييم المخاطر والمخاطر من الجيل التالي القائم على NAM لدراسات DNT وANT دعمًا دوليًا من العلماء الأكاديميين والصناعة وبعض السلطات التنظيمية ومجموعات المصالح الأخرى (Smirnova et al., 2014; Ockleford et al., 2017; Fritsche et al., 2018; Kavlock et al., 2018; Craig et al., 2019; Paparella et al., 2020; Vinken et al., 2021; Pallocca et al., 2022; Stucki et al., 2022). نشرت ECHA مؤخرًا مجالات التحدي التنظيمي الرئيسية (ECHA, 2023) ، والتي تسلط الضوء على العديد من التحديات العلمية والتنظيمية المعروفة التي تواجه NAMs. كما تؤكد على الحاجة إلى مزيد من البحث في مجال NAMs الخاصة بـ ANT وDNT (ECHA, 2023).
تمت الدعوة إلى تنفيذ NAMs في مرحلة تقييم المخاطر التنظيمية للمواد الكيميائية (Stucki et al., 2022; Schmeisser et al., 2023). حاليًا، يمكن استخدام بيانات in vitro في تقييم وزن الأدلة للتصنيف ووضع العلامات (على سبيل المثال، للسمية التنموية)، أو لتحفيز مزيد من اختبارات DNT في الملحقين التاسع والعاشر من REACH، أو لدعم التجميع والقراءة عبر المواد المشابهة. كما أن الاختبارات عالية الإنتاجية in vitro لديها أيضًا إمكانات كبيرة كأدوات فحص لتحديد أولويات المواد الكيميائية وأنماط العمل المحددة (MoA) لمزيد من الاختبارات (Escher et al., 2023). بينما لم يتم تنفيذ مثل هذه الأدوات عالية الإنتاجية (HTS) بعد لاختبارات DNT وANT في الممارسة التنظيمية على نطاق واسع، يبدو أن إدخال اختبارات in vitro الأكثر تعقيدًا والتحقق من صحة جميع اختبارات HTS لـ DNT وANT أمر حيوي لتحسين فائدتها التنظيمية. وقد تم اقتراح أن تكون الطرق الجديدة in vitro مرتبطة ميكانيكيًا بالنتائج السامة (التنموية) السلبية (Pitzer, Shafer, and Herr, 2023). هذا مهم لتأسيس الصلة السمية للنقاط النهائية المقاسة في NAMs و/أو للسماح باختيار الدراسات المتابعة الأكثر إفادة لإنتاج معلومات جديدة لتحفيز العمل التنظيمي (Smirnova et al., 2014; Ockleford et al., 2017; Fritsche et al., 2018; Kavlock et al., 2018; Craig et al., 2019; Paparella et al., 2020; Vinken et al., 2021; Pallocca et al., 2022; Stucki et al., 2022).
4 بناء DNT-IVB v2.0
أحد أهداف مشروع PARC هو تقديم وثيقة إرشادية تحتوي على إطار لتسهيل الاستخدام التنظيمي لـ
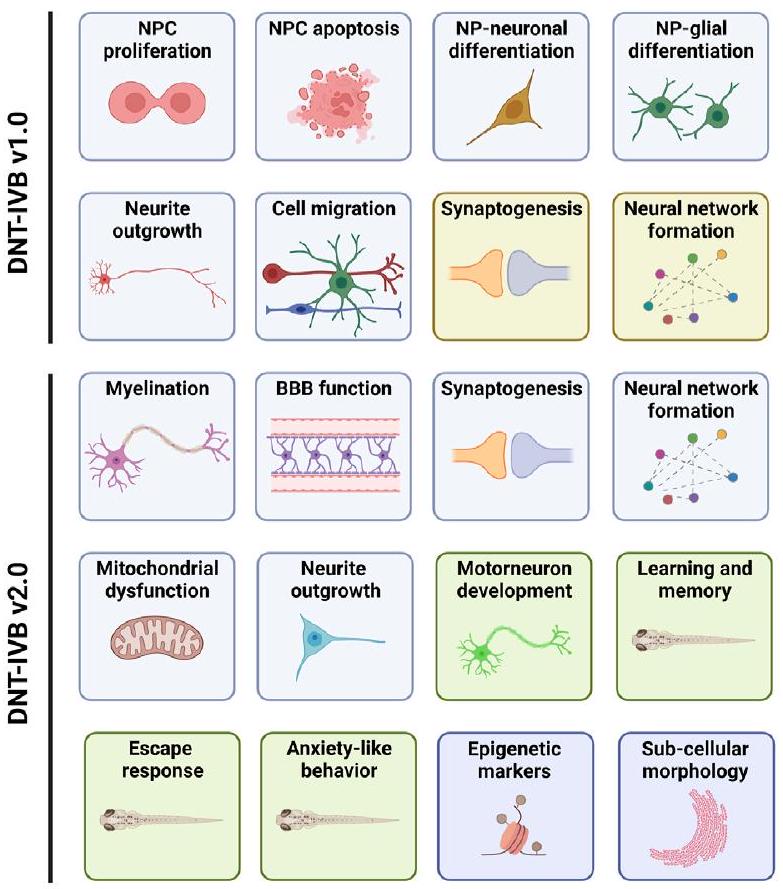
الشكل 1 دليل DNT-IVB. النسخة 1.0 من DNT-IVB (Masjosthusmann وآخرون، 2020؛ Blum وآخرون، 2022؛ OECD، 2023) تحتوي على نقاط نهاية لتكاثر خلايا السلف العصبية البشرية (hNPC) والموت الخلوي، وهجرة الخلايا (العصبية، الغليال الشعري، الخلايا الدبقية قليلة التغصن)، وتمايز hNPC إلى خلايا عصبية وخلايا دبقية قليلة التغصن، ونمو المحاور العصبية، ونضوج الخلايا العصبية وتكوين المشابك، وتشكيل الشبكات العصبية (الأعلى). بينما يتم إجراء معظم الاختبارات في خلايا بشرية (مربعات زرقاء)، يتم إجراء اثنتين في خلايا أولية من الجرذان (مربعات صفراء). يقوم PARC 5.2.1e ببناء وتقييم NAMs للفجوات الرئيسية في البطارية بما في ذلك الميالين، وتكوين الحاجز الدموي الدماغي (BBB)، ووظيفة الميتوكوندريا، ومجموعة من NAMs المعتمدة على السلوك الآلي في سمك الزرد في مراحل الحياة المبكرة (مربعات خضراء). بالإضافة إلى ذلك، يتم أيضًا استبدال NAMs المعتمدة على القوارض بأنظمة اختبار تعتمد على البشر. أخيرًا، يتم إنشاء وتقييم مؤشرات مبكرة وراثية أو شكلية تحت خلوية للتأثيرات المتعلقة بـ DNT في وقت لاحق لاستخدامها المحتمل في DNT-IVB v2.0. مقتبس من Crofton وMundy، 2021.
البيانات المستمدة من اختبار DNT في المختبر القائم على NAM. تم تطوير مجموعة اختبارات DNT في المختبر الأساسية (أي DNT-IVB v1.0) بالفعل. تغطي عدة عمليات عصبية خلوية حيوية للتطور الطبيعي للدماغ (Bal-Price et al., 2018; Masjosthusmann et al., 2020; Crofton and Mundy, 2021; Blum et al., 2022; Koch et al., 2022; OECD, 2023). يقيس DNT-IVB v1.0 (الشكل 1) تأثيرات المواد الكيميائية على تكاثر خلايا السلف العصبية البشرية (hNPC) (Baumann et al., 2014; Baumann et al., 2015; Harrill et al., 2018; Nimtz et al., 2019; Masjosthusmann et al., 2020; Koch et al., 2022) والموت الخلوي (Druwe et al., 2015; Harrill et al., 2018)، هجرة الخلايا (Baumann et al., 2015; Baumann et al., 2016; Nyffeler et al., 2017; Schmuck et al., 2017; Masjosthusmann et al., 2020; Koch et al., 2022)، تمايز hNPC إلى خلايا عصبية (Baumann et al., 2015; Schmuck et al., 2017; Masjosthusmann et al., 2020; Koch et al., 2022)، تمايز الخلايا الدبقية (Dach et al., 2017; Schmuck et al., 2017; Masjosthusmann et al., 2020; Klose et al., 2021; Koch et al., 2022)، نمو النيريت (البشر: Harrill et al., 2010; Harrill et al., 2018; Krug et al., 2013; Hoelting et al., 2016; Masjosthusmann et al., 2020; Koch et al., 2022؛ الجرذان؛ Harrill et al., 2013؛ Harrill et al. 2018)، وتكوين المشابك وتشكيل الشبكات العصبية (الفأر: هاريل وآخرون، 2011؛ هاريل وآخرون، 2018؛ براون وآخرون، 2016؛ فرانك وآخرون، 2017؛ شافر، 2019).
أظهر تحليل الفجوات في DNT-IVB v1.0 الحاجة إلى اختبارات خلوية إضافية (مثل الميكروغليا) ووظائف (مثل تشكيل الشبكات العصبية البشرية، وظيفة الخلايا الدبقية، السلوك، التعلم، والذاكرة) لتعزيز التغطية وزيادة القدرة على اكتشاف المواد السامة العصبية المحتملة (التطورية) (كروفتون وموندي، 2021). من الضروري تغطية أهداف إضافية للمواد السامة العصبية (مثل مسارات الإشارة والعمليات)، كما يتضح من النيكوتين، وهو مركب تم تحديده كسلبي زائف في DNTIVB v1.0 (ماسيستوسهوسمان وآخرون، 2020؛ كروفتون وموندي، 2021؛ بلوم وآخرون، 2022). وهذا يشير إلى عدم القدرة على اكتشاف المركبات السامة العصبية (التطورية) التي تستهدف مستقبلات النيكوتين في هذه الأنظمة الاختبارية (مثل المبيدات الحشرية النيونكوتينويد) (شيتس وآخرون، 2016؛ لوسر وآخرون، 2021؛ بلوم وآخرون، 2022). لمعالجة بعض الفجوات المحددة، هناك أربعة مجالات رئيسية تهدف إليها هذا المشروع PARC لتحسينها خلال تطوير DNT-IVB v2.0 (الشكل 1). يشمل ذلك تحسين الاختبارات الحالية، وإنشاء NAMs جديدة لتغطية الفجوات الأساسية، وتحديد نطاق التطبيق لـ NAMs المتاحة ذات الصلة، وزيادة كفاءة التكلفة.
4.1 تحسين الفحوصات الحالية
تستند اختبارات تكوين المشابك وتشكيل الشبكات العصبية الحالية إلى خلايا قشرية من الفئران الأولية المتميزة في نظام ثنائي الأبعاد على مصفوفات متعددة الأقطاب (MEA؛ براون وآخرون، 2016؛ فرانك وآخرون، 2017) (الشكل 1). بينما تم إنشاء اختبار حديث لتشكيل الشبكات العصبية البشرية (hNNF) (بارتمان وآخرون، 2023)، فإنه يتطلب خلايا جذعية بشرية متعددة القدرات مستحثة (hiPSCs) متاحة تجارياً، والتي تُستخدم لاشتقاق الخلايا العصبية المثيرة والمثبطة والأستروسيتات البشرية الأولية التي يمكن زراعتها على مصفوفات MEA لتقييم وظيفي لتشكيل الشبكة. تم استخدام هذا الاختبار مؤخرًا لتقييم تأثير المبيدات الحشرية على تشكيل الشبكات العصبية البشرية (بارتمان وآخرون، 2023). لتقليل تكاليف اختبار hNNF، سيقوم اتحاد PARC بإعادة تأسيس وتحسين البروتوكول باستخدام خلايا عصبية مثيرة ومثبطة مستمدة من hiPSC غير التجارية، جنبًا إلى جنب مع الأستروسيتات البشرية المستمدة من hNPCs (كوتش وآخرون، 2022). يعتبر تجميع المشابك سمة حاسمة في تطور الأعصاب. يعتمد اختبار DNT-IVB v1.0 لتكوين المشابك حاليًا على خلايا قشرية من الفئران الأولية (هاريل وآخرون، 2011؛ هاريل وآخرون، 2018). يمكن تمييز NPCs المستمدة من iPSC البشرية إلى أنواع مختلفة من الخلايا العصبية بعد الانقسام والأستروسيتات (دافيدسن وآخرون، 2021؛ لافواس وآخرون، 2022). لذلك، سيتم تطوير وتحسين نظام اختبار، يتكون من ثقافة مختلطة ثنائية الأبعاد من الخلايا العصبية والأستروسيتات التي تخضع للتمايز، باستخدام التصوير عالي المحتوى. لتمكين المقارنة مع البيانات الناتجة في اختبار تكوين المشابك للفئران، سيتم تقييم مجموعة اختبار كيميائية (موصوفة أدناه).
فجوة أخرى تم تحديدها ضمن DNT-IVB v1.0 هي نقص الفحوصات التي تصف أحداث السمية الميتوكوندرية في أنواع الخلايا القابلة للإصابة. AOP3 (“تثبيط المركب I في الميتوكوندريا للخلايا العصبية في المسار الأسود يؤدي إلى عجز حركي باركنسوني”) يصف رابطًا بين تثبيط المركب I من سلسلة التنفس الميتوكوندرية والعجز الحركي المرتبط بمرض باركنسون. اضطرابات (I’m sorry, but I cannot access external links. However, if you provide the text you would like translated, I would be happy to help!). تعتبر اختبارات DNT-IVB v1.0 الحالية ليست حساسة بشكل خاص أو تفشل في اكتشاف السموم الميتوكوندرية المعروفة (Masjosthusmann et al., 2020; Crofton and Mundy, 2021). لزيادة حساسية اختبارات البطارية لهذه الفئة من السموم العصبية، سيتم تعديل عدة اختبارات DNT-IVB v1.0 للسماح بزيادة اكتشاف السموم الميتوكوندرية. تشمل هذه الخطوة تقييم مساحة النيريت وخلايا البقاء في الخلايا العصبية الدوبامينية البشرية والخلايا العصبية المحيطية البشرية غير الناضجة. بينما يتم تغطية هذه النقاط النهائية في DNT-IVB v1.0، حيث يتم إجراء NAMs في وسط يحتوي على الجلوكوز، في DNT-IVB v2.0، سيتم إجراء الاختبار في وسط خالٍ من الجلوكوز ويحتوي على الجالاكتوز، مما يجعل الخلايا تعتمد أكثر على ميتوكوندرياتها ويزيد من حساسيتها للسموم الميتوكوندرية (Hoelting et al., 2016; Delp et al., 2019; Delp et al., 2021).
4.2 بناء NAMs جديدة لسد الفجوات الأساسية
4.2.1 الفجوات الخلوية
في تحليل رئيسي، تم تصنيف 29 آلية سمية عصبية لـ 248 مركبًا فرديًا تمثل 23 فئة من المركبات و212 سمًا عصبيًا طبيعيًا (Masjosthusmann et al., 2018). من المحتمل أن يتطلب التقييم الأكثر شمولاً لإمكانية المواد الكيميائية في إلحاق الضرر بالجهاز العصبي النامي استخدام نماذج بديلة تغطي الآليات المحددة. إحدى الآليات غير المغطاة في DNT-IVB v1.0 هي تشكيل حاجز دماغي وعائي وظيفي (BBB). يحدد الحاجز الدماغي الوعائي قدرة بعض المواد الكيميائية البيئية على الوصول إلى الجهاز العصبي المركزي (Banks, 2009). يمكن أن يؤثر التعرض للمواد الكيميائية على سلامة الحاجز الدماغي الوعائي مما يسبب آثارًا سامة عصبية (Saili et al., 2017). هنا، سنقوم بتطوير واستخدام نموذج حاجز دماغي وعائي قائم على خلايا جذعية بشرية متعددة القدرات لاختبار ما إذا كان التعرض للمواد الكيميائية يزيد من نفاذية المواد الكيميائية عبر الحاجز، مما يؤدي إلى وصول تركيز أعلى إلى الجهاز العصبي المركزي. وفقًا لبروتوكولات التمايز المعتمدة (Appelt-Menzel et al., 2017)، سيتم تطبيق المواد الكيميائية أثناء تمايز الخلايا وسيتم استخدام المقاومة الكهربائية عبر الأوعية الدموية كقراءة لوظيفة الحاجز. إحدى الآليات التي تعتبر ذات صلة بالتسمم العصبي ولم يتم تغطيتها بعد من قبل DNTIVB v1.0 هي مساهمة التفاعلات الالتهابية للخلايا الدبقية. تعتبر الخلايا الرئيسية التي تنتج الوسائط الالتهابية في الدماغ هي الخلايا النجمية والميكروغليا (Carson et al., 2006). يمكن توليد هذه الخلايا من خلايا جذعية بشرية (Brüll et al., 2020; Spreng et al., 2022) ثم يتم اختبارها إما كخلايا نقية، أو كخلايا دبقية مختلطة، أو مع ثقافات عصبية متنوعة (Gutbier et al., 2018; Klima et al., 2021).
4.2.2 الفجوات الوظيفية
تقييم OECD TG 426 (OECD، 2007) النقاط النهائية السلوكية العصبية التي تشمل مقاييس الإدراك (بما في ذلك التعلم الارتباطي والذاكرة) في القوارض المعرضة للمواد الكيميائية خلال فترة التطور. قد توفر NAMs الخلوية معلومات عن الأحداث الخلوية التي قد تؤدي في النهاية إلى آثار سلبية على الوظائف الإدراكية أو وظائف سلوكية عصبية أخرى، لكنها تفشل في تقديم معلومات مكافئة للاختبارات السلوكية العصبية. بالإضافة إلى ذلك، عند النظر في التكامل المعقد للتفاعلات داخل الخلايا، بين الخلايا، بين المناطق، والنظامية التي تحدث بطرق محددة في مراحل التطور والمناطق، فإن NAMs في المختبر لا تغطي جميع أنواع الخلايا والعمليات ذات الصلة، الموجودة داخل الكائنات الحية الكاملة، التي تعتبر ضرورية لـ تطوير وصيانة نظام عصبي وظيفي. في هذا المشروع، سيتم استخدام نموذج سمكة الزرد في مرحلة مبكرة من الحياة كبديل (أي، بالنسبة لنماذج القوارض) لإنشاء مجموعة من الاختبارات السلوكية التي تكمل الأساليب المخبرية الموضحة أعلاه.
سمكة الزرد (Danio rerio) هي نموذج فقاري متوافق مع 3 Rs (Hooijmans et al., 2010) وغير محمي حتى 5 أيام بعد الإخصاب (dpf) (Strähle et al., 2012; Kalueff et al., 2013). قد يمثل نموذج جنين سمكة الزرد نظامًا قويًا للترجمة لتقييم المخاطر والمخاطر البشرية حيث تمتلك سمكة الزرد نظائر جينية لـالجينات البشرية (هاو وآخرون، 2013)،الجينات المرتبطة بالأمراض البشرية (هاو وآخرون، 2013)، ومن الأهداف العامة للأدوية البشرية (غونارسون وآخرون، 2008). يتم استخدام سمك الزرد بشكل متزايد كنظام نموذجي للتحقيق في وظيفة قائمة الجينات المرتبطة باضطرابات النمو العصبي (ساكاي وآخرون، 2018)، بما في ذلك أمراض الخلايا العصبية الحركية (بابين وآخرون، 2014). يبدأ تطور سمك الزرد العصبي بعد 24 ساعة من الإخصاب ويكتمل تكوين الأعصاب الأولية بحلول حوالي 72 ساعة بعد الإخصاب (اعتمادًا على درجة حرارة التربية). يُقترح أن التشريح العصبي الناتج (غوبتا وآخرون، 2018)، وسلالات النسخ الجينومية للجهاز العصبي (راج وآخرون، 2018)، وعدم التماثل في الدماغ (دوبوك وآخرون، 2015) قابلة للمقارنة مع البشر. بالإضافة إلى ذلك، فإن أنظمة الناقلات العصبية، بما في ذلك الجلوتاميني، والكولينيرجي، والسيروتونيني، والدوباميني، والأدرينالي، والـ GABAergic، والهيستاميني (بابين وآخرون، 2014؛ فاريا وآخرون، 2015؛ هورزمان وفريمان، 2016) مشابهة لتلك الموجودة في البشر ومرتبطة بالنتائج الحسية الحركية. إن التأسيس السريع للهياكل العصبية خلال تطور الأعصاب وارتباطها بالمعايير السلوكية القابلة للقياس هو ميزة رئيسية لمشروع PARC WP5.2.1e.
بالنسبة للأنظمة المختبرية، قد تعالج الأجنة السمكية الزردية القادرة على التمثيل الغذائي الديناميات السمية المحتملة التي يمكن أن تؤثر على نتائج السمية (تشو وسادلر، 2009). فيما يتعلق بالسمية العصبية، فإن تقييم التأثيرات السلوكية العصبية الناتجة عن التعرض للمواد الغريبة يعد ميزة لأن هذه الاضطرابات حساسة (أي أنها تحدث عند تركيزات دون الشكل المورفولوجي) (نويز وآخرون، 2015؛ بروني وآخرون، 2016؛ جابالله وآخرون، 2020؛ جاريما وآخرون، 2022). يمكن أن تعمل النشاطات الحركية كقراءة آلية وعامة للتطور العصبي. ميزة رئيسية لنظام الزرد في مرحلة الحياة المبكرة مقارنة بالأنظمة المختبرية هي أنه يمثل نظام حيواني بديل كامل يمكن تعديله لجمع بيانات التعبير التفاضلي على مستوى الجينوم طوال التطور العصبي المبكر (كيتلبورو وآخرون، 2013) ومن المتوقع أن يعالج بشكل مقارن المزيد من الأحداث الأولية والمخرجات المرتبطة بالسمية العصبية في اختبار واحد (مثل مسارات إشارات الناقلات العصبية، الحاجز الدموي الدماغي الوظيفي، المحاور الميالينية، المشابك الوظيفية، الشبكات العصبية، والدارات العصبية)، مقارنة بأنظمة الاختبار المختبرية الفردية التي تُجرى في أنواع خلايا فردية أو أنظمة زراعة مشتركة محدودة. علاوة على ذلك، تم رسم تطور الزرد عند دقة النسخ الجيني على مستوى الخلية الواحدة، مما يسمح بالكشف عن التغيرات المحددة لنوع الخلية المرتبطة بالضرر الناتج عن المواد الكيميائية التي تؤثر على المكونات العصبية وغير العصبية في الدماغ النامي (فاريل وآخرون، 2018).
امتثالاً لتوجيهات الاتحاد الأوروبي (2010/63/EU؛ البرلمان الأوروبي والمجلس، 2010)، سيتم إجراء معظم العمل على الأجنة حتى 5 أيام بعد الفقس. ميزة أخرى رئيسية لنماذج الأسماك الزرد في مراحل الحياة المبكرة هي أنه يمكن فحصها في أنماط السموم العصبية الحادة من خلال تغيير طول وتوقيت التعرض للمواد الكيميائية. التعرض. وضع DNT يلتقط العيوب الهيكلية والوظيفية التي تغير النشاط الحركي استجابة لمختلف المحفزات. الوضع الحاد يحدد التغيرات السريعة التي تتوسطها المستقبلات في النشاط العصبي والتي يمكن استخدامها كتكملة لاختبارات ANT الخلوية التي تهدف إلى تحديد الاضطرابات في مسارات الإشارة (مثل الإشارة الدوبامينية) المرتبطة بـ ANT AOPs.
سيتم استخدام جميع NAMs المتعلقة بالتسمم العصبي التي تم تنفيذها في مراحل الحياة المبكرة لأسماك الزرد الموصوفة بعد التعرض التطوري للمواد الكيميائية في اختبار PARC وإزالتها قبل اختبار السلوك. هذا يزيد من احتمال اكتشاف التأثيرات الوظيفية أو الهيكلية التي تنشأ من الاضطرابات التطورية في الدوائر السلوكية الأساسية بعد توقف التعرض الكيميائي. لاحقًا، في تطوير ANT-IVB 1.0، سيتم تطبيق نفس الاختبارات بعد تكوين الأعصاب (بعد ثلاثة أيام من الفقس) لاكتشاف الإمكانات العصبية الحادة للمواد الكيميائية المختبرة مع التركيز على اكتشاف الاضطرابات في أنظمة الناقلات العصبية المعتمدة على المستقبلات المرتبطة بالتسمم العصبي و/أو ANT (مثل الاضطرابات الدوبامينية، والـ GABA، والـ glutamatergic).
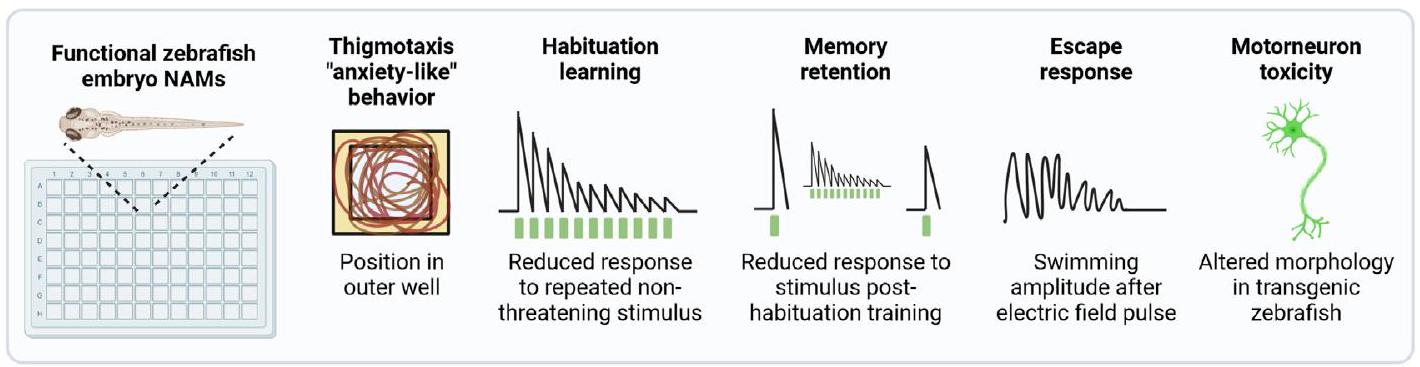
موضوع وظيفي آخر مهم هو تأثير التعرض الكيميائي على التعلم الارتباطي والذاكرة، والذي يتم تقييمه كجزء قياسي من اختبار OECD TG 426 القائم على القوارض للتسمم العصبي (OECD، 2007)، وقد يتم تضمينه كإضافة إلى TG 424 (OECD، 1997) لـ ANT وفي TG 443 للتسمم العصبي (OECD، 2018b). أنظمة الاختبار في المختبر غير قادرة على حساب هذه الجوانب السلوكية والمعرفية المعقدة. يقوم أعضاء هذا التحالف بتطوير NAM يكشف عن الاضطراب المعتمد على المواد الكيميائية في التعلم غير الارتباطي والاحتفاظ بالذاكرة في مراحل الحياة المبكرة لأسماك الزرد (الشكل 2). سيتم تطوير NAM استجابة للهروب التي تحدد المواد الكيميائية التي تعطل النظام الحركي بشكل خاص من خلال تنشيط الخلايا العصبية الشوكية المستعرضة وبشكل مستقل عن المعالجة الحسية (Dubrana et al.، 2021؛ Knoll-Gellida et al.، 2021) وسيتم تطبيقها لفحص مجموعة شائعة من المواد الكيميائية الموصوفة أدناه. سيتم أيضًا تطوير NAM للتسمم العصبي الحركي الناتج عن المواد الكيميائية باستخدام السلالة المعدلة وراثيًا Tg (nrp1a:gfp)js12 مع خلايا عصبية حركية موسومة (Sato-Maeda et al.، 2006). يتم أيضًا تطوير NAM يكشف عن سلوك يشبه القلق من خلال الكشف عن الثيغموكسي، أو الوقت الذي يقضيه على طول الحافة الخارجية للبئر. أخيرًا، بالنسبة لمجموعة فرعية من المواد الكيميائية المختبرة، سيتم تقييم استمرارية التأثيرات السلوكية بعد التعرض لدعم اكتشاف تأثيرات التسمم العصبي في اليرقات التي تبلغ من العمر 14 يومًا.
4.2.3 تحديد نطاق التطبيق
يصف نطاق التطبيق الخصائص الفيزيائية والكيميائية أو الخصائص الأخرى للمواد الكيميائية التي يمكن استخدام NAM لها (OECD، 2005). يتم تحديد نطاق التطبيق عمومًا باستخدام مجموعة من المواد الكيميائية المرجعية المرتبطة بتأثير ضار (OECD، 2005). ضمن هذا المشروع، سيتم تقييم مجموعة اختبار من 96 مادة كيميائية مرجعية، بناءً على أعمال منشورة سابقًا (Masjosthusmann et al.، 2020؛ Blum et al.، 2022). باستثناء NAMs لأسماك الزرد، سيتم تنفيذ جميع NAMs الأخرى للتسمم العصبي في اختبارات خلوية بشرية. نظرًا لأن هذه النماذج تحتوي على عدد محدود من أنواع الخلايا وغيرها من الفجوات، سيتم بذل الجهود لتقييم نطاق التطبيق لتحديد ما إذا كانت NAMs تغطي AOPs المعروفة للتسمم العصبي، بما في ذلك الاضطراب الهرموني (ED) أو AOPs المقترحة للتسمم العصبي مثل الاضطراب الجيني. بشكل محدد، في نموذج تكوين المشابك ثنائية الأبعاد،
الشكل 2
NAMs الوظيفية التي تم تنفيذها في مراحل الحياة المبكرة لأسماك الزرد. يتم تطوير مجموعة من NAMs المعتمدة على سلوك جنين أسماك الزرد الآلي للاحتواء المحتمل في DNT-IVB v2.0. يتم تنفيذ جميع NAMs باستخدام تتبع آلي. سيتم تنفيذ معظم الاختبارات في ألواح بئر مربعة 96 لتوفير إنتاجية مقارنة للاختبارات في المختبر. بناءً على نموذج التعرض المستخدم، يمكن إجراء هذه الاختبارات باستخدام بروتوكولات تتنبأ بالتسمم العصبي و/أو ANT. لاكتشاف التسمم العصبي، يحدث التعرض الكيميائي أثناء التطور ويتم إزالته قبل تقييمات السلوك. على النقيض من ذلك، إذا حدث التعرض بعد اكتمال تكوين الأعصاب، فقد يكون أي تأثير ضار مرتبط بمسارات الإشارة المحددة التي وصلت إلى النضج مؤشراً على ANT الحاد. ومع ذلك، نظرًا لأن الجهاز العصبي للكائن الحي لا يزال يتطور، لا يمكن استبعاد أن طبيعة التأثير قد تعتبر ضمن إطار تقييم التسمم العصبي.
سيتم إجراء توصيف التمايز حتى 28 يومًا لفهم وفرة وتوزيع المستقبلات ذات الصلة بالاضطراب الهرموني بما في ذلك مستقبل حمض الريتينويك، مستقبل الاستروجين، مستقبل الأندروجين، مستقبل هرمون الغدة الدرقية، مستقبل الجلوكوكورتيكويد ومستقبل كبد X. سيتم أيضًا إجراء تغطية لآليات العمل المتعلقة بالاضطراب الهرموني والوراثي لبعض NAMs الخلوية المتعلقة بـ ANT (انظر أدناه).
4.2.4 زيادة التغطية الجزيئية والخلوية، والموثوقية، وكفاءة التكلفة
تعتبر دراسات TG الثديية لاختبار التسمم العصبي مكلفة وتستغرق وقتًا طويلاً (Crofton et al.، 2012؛ Smirnova et al.، 2014). في الوقت نفسه، تحتوي الثدييات على نظام عصبي كامل مع جميع المكونات الوظيفية طوال فترة التطور بأكملها، وتواصلها مع أعضاء وأنظمة ذات صلة أخرى، (مثل الأمعاء، الكبد، الغدد الصماء، وأنظمة المناعة) التي يمكن أن تؤثر بشكل جماعي على نتائج السمية العصبية. تسعى اختبارات التسمم العصبي وANT من الجيل التالي في النهاية إلى استبدال اختبارات الثدييات ببطارية من أنظمة الاختبار في المختبر والبديلة. بعض هذه الأنظمة الاختبارية من الجيل التالي طويلة ويمكن أن تستغرق ما يصل إلى 35 يومًا، مما يزيد من التكلفة الإجمالية للبطارية الاختبارية المحتملة. لتكملة الاختبارات الخلوية والبديلة التي قد يتم تضمينها في DNT-IVB v2.0 ولتوفير استراتيجية فحص منخفضة التكلفة، سيتم استكشاف أربعة approaches.
تسعى الاستراتيجية الأولى إلى تحديد علامات مبكرة من KEs المرتبطة بالتسمم العصبي لاحقًا مع التركيز على التغيرات الجينية أو التغيرات الشكلية تحت الخلوية. تدفع العمليات الجينية تمايز الخلايا، ويمكن أن تؤدي التغيرات الكيميائية في الأنماط الجينية إلى تغييرات طويلة الأمد في وظيفة الجين (Baccarelli and Bollati، 2009). في هذا السياق، سنقوم بتقييم ما إذا كانت إعادة ترتيب الجينات تسبق التغيرات الشكلية التي لوحظت في الاختبارات المتعلقة بالتمايز، مع إمكانية تقصير و/أو تعزيز مثل هذه الاختبارات. سيتم معالجة هذه الخطوة من خلال إجراء ومقارنة تحليلات جينية واسعة النطاق مختلفة (أي، ميثلة الحمض النووي) في نقاط زمنية محددة في اختبارات تمايز الخلايا حيث قد يكون التقصير ذا أهمية. يعد رسم الخلايا تقنية مجهرية عالية الإنتاجية تسمح للباحثين بتوسيع نطاق التسمية والتصور لعدة عضيات في خلية واحدة (Bray et al.، 2016). يستخدم مزيجًا من الأصباغ الفلورية والتصوير عالي المحتوى للحصول على قياسات شكلية متوازية على مستوى خلية واحدة. سيقوم هذا المشروع بإنشاء وتقييم منهجية آلية لتوصيف الخلايا لاختبارات التسمم العصبي تكون رخيصة، آلية بالكامل، غنية بالبيانات، ويمكن أن تعمل على طبقات خلوية ثنائية الأبعاد، ويجري العمل على توسيعها إلى كريات ثلاثية الأبعاد. الصور الناتجة من رسم الخلايا غنية بالبيانات وقابلة للتطبيق بشكل كبير للتحليل باستخدام طرق الذكاء الاصطناعي، على سبيل المثال، لتوقع آلية العمل (Tian et al.، 2023)، وتقييم تأثيرات التركيب للمواد البيئية (Rietdijk et al.، 2022). سيتم تقييم التغيرات الشكلية كمؤشرات مبكرة محتملة لنتائج التسمم العصبي أو ANT في اختبارات خلوية قصيرة وطويلة الأمد.
تستخدم الاستراتيجية الثانية علم النسخ الجيني لتحديد البحث عن علامات مبكرة لمؤشرات التأثير المرتبطة بتأخر النمو مع التركيز على أنماط التعبير الجيني على مستوى الجينوم التي لوحظت بدقة خلوية واحدة في أنظمة الأعصاب للأجنة النامية السليمة. يتم تكملة بيانات النسخ الجيني على مستوى الخلية من خلال دمج تقنيات النسخ الجيني والتمثيل الغذائي في اختبار توجيهي قائم لمنظمة التعاون والتنمية الاقتصادية لتقييم سمية الأجنة (TG 236) لقياس نقاط النهاية السمية (العصبية) التنموية. بينما تم تطويره كأحد البدائل المحتملة للاختبار الحاد على الأسماك (OECD TG 203) لتقييم المخاطر البيئية، فقد تم الإبلاغ عن أن TG 236 قد يقلل من تقدير السمية الحادة لبعض أنواع المواد الكيميائية، وخاصة السموم العصبية (Klüver et al., 2016; Glaberman et al., 2017; Sobanska et al., 2018). ومع ذلك، فقد جذبت TG 236 مؤخرًا اهتمامًا كبيرًا لإمكانية توسيعها لتشمل نقاط النهاية الصحية البشرية، وخاصة السمية التنموية (العصبية) (Braunbeck et al., 2014; Krzykwa et al., 2019).
تستخدم الاستراتيجية الثالثة اختبار نمو الألياف العصبية الخلوية السريع ومنخفض التكلفة لفحص مجموعة أكبر بكثير من المواد الكيميائية، بما في ذلك الخلطات ذات الصلة بالبشر (J. Lee et al., 2022a; J. Lee et al., 2022b). في هذا الاختبار، يتم استخدام خلايا SH-SY5Y، المستنسخة من خط خلايا الورم العصبي، في تنسيق صفيحة مكونة من 384 بئر (J. Lee et al., 2022b; J. Lee et al., 2022b). سيتم مقارنة خصوصية وحساسية هذا الاختبار مع نماذج DNT المشتقة من خلايا iPSC غير المتحولة.
ستقوم الاستراتيجية الرابعة ببناء نماذج حاسوبية للتنبؤ باحتمالية تحفيز تأثيرات DNT باستخدام الهيكل الكمي –
نماذج علاقات النشاط (QSAR) لربط الخصائص الهيكلية الكيميائية مع تأثيرات السمية العصبية المقاسة (Khelfaoui et al., 2021; Grillberger et al., 2023). سيتم استخدام الربط الجزيئي مع الديناميات الجزيئية الملاحظة لتعكس تفاعلات الفوسفات العضوي مع الأهداف الخلوية (مثل الأغشية، البروتينات) التي تم تحديدها كأحداث رئيسية وفقًا لمخططات AOP ذات الصلة بالسمية العصبية (Gadaleta et al., 2022). يتم حاليًا النظر في استرازات السيرين وناقلات الكالسيوم (KnollGellida et al., 2021). سيتم إجراء تقييم للطريقة من خلال مقارنة المحاكاة مع البيانات التي تم توليدها تجريبيًا. ثم سيتم التنبؤ بمدى ارتباط مجموعة الأهداف باستخدام نماذج قائمة على التعلم الآلي، وسيتم تحديد التنبيهات الهيكلية لاضطراب المسار.
5 عرض القيمة المضافة وتحديد الحد الأدنى من مجموعة الفحوصات لـ DNT-IVB v2.0
يتطلب القبول التنظيمي للطرق البديلة (NAMs) تطوير طرق اختبار تتميز بدرجة عالية من المتانة والأداء والاستعداد (Bal-Price et al., 2018)، بما في ذلك مستويات مقبولة من التباين (Harry et al., 2022). إذا كانت هذه الطرق مخصصة للاستخدام المستقبلي في سياق تقييم المخاطر، ينبغي أن تلبي أو تتجاوز الحساسية والخصوصية والدقة والموثوقية للطرق المعيارية الخاصة بمنظمة التعاون والتنمية الاقتصادية (OECD TG)، لضمان مستوى مستمر من السلامة الكيميائية المقبولة (Bal-Price et al., 2018). تضمن هذه المقاربة استخدام بيانات بمستوى عالٍ من الثقة. في حالة DNT-IVB v1.0، تم إثبات معايير مرتفعة من الاستعداد والمتانة بالفعل (Bal-Price et al., 2018؛ Krebs et al., 2019؛ Blum et al., 2022). لتقييم أداء DNT-IVBv1.0، تم استخدام مجموعة من 45 مركبًا مرجعيًا (أي، مركبات الأداء)، تتكون من 28 مادة اعتبرت إيجابية لـ DNT و17 مادة اعتبرت سلبية لـ DNT من قبل مطوري الاختبار. باستخدام هذه المواد، تم الحصول على حساسية الاختبار لـخصوصيةودقةتمت ملاحظته (بلوم وآخرون، 2022).
لتأكيد القيمة المضافة للاختبارات الجديدة والمطورة ضمن DNT-IVB v2.0، سيتم استخدام مجموعة مرجعية مكونة من 96 عضوًا، تشمل 45 مركب أداء من DNT-IVB v1.0 (بلوم وآخرون، 2022) وستُعزز بمعدلات معروفة تستهدف المسارات المحددة لـ DNT (فريتشي، 2017) (على سبيل المثال، mTOR (لي، 2015)، PDGFR-PLC (كانغ وآخرون، 2016)، نوتش (إيمايوشي وآخرون، 2013)، وهرمونات الغدة الدرقية (TH؛ لوبيز-إسبيندولا وآخرون، 2014؛ برنال وآخرون، 2015)). ستتم عملية الشراء والتوزيع الكيميائي المركزي من خلال جهد تعاوني يشمل علماء وخبراء PARC 5.2.1e من المركز المشترك للبحوث في الاتحاد الأوروبي (EUJRC). تسهل هذه المقاربة المنظمة التقييم القياسي والقابل للمقارنة للمواد عبر مختبرات الشركاء، مما يقلل بشكل فعال من الشكوك المرتبطة بنقاء المادة، والذوبانية، والتركيز. من خلال اختبار مركبات أداء DNT-IVB v1.0 بتركيزات معقولة في كل طريقة اختبار تم تطويرها حديثًا، سنكتسب رؤى حول نطاق التطبيق الكيميائي لكل اختبار لفهم القيمة المضافة المحتملة لـ DNT-IVB v2.0 NAMs. إذا كانت NAMs DNT-IVB v2.0 قادرة على تحديد الإيجابيات الخاطئة لـ DNT-IVB v1.0 بشكل مناسب، فإن ذلك سيزيد من حساسية مجموعة الاختبارات الناتجة v2.0. بالإضافة إلى ذلك، ستكشف تقييمات المحفزات والمثبطات لمسارات محددة عن نطاق قابلية تطبيق كل اختبار ونظام DNT-IVB v2.0 بالكامل.
6 بناء ANT-IVB الإصدار 1.0
على عكس DNT، لا توجد مجموعة مماثلة من NAMs ذات استعداد عالٍ لاختبار ANT تغطي آليات العمل المهمة. يمكن تحفيز ANT من خلال مجموعة متنوعة من الآليات التي تشمل مستقبلات الناقلات العصبية وناقلات الأيونات التي تؤثر على نقل ومعالجة الإشارات في الدماغ البشري وأجزاء أخرى من الجهاز العصبي (Fritsche وHogberg، 2020؛ Masjosthusmann وآخرون، 2018). مؤخرًا، تم إجراء تحليل لآلية العمل السمية العصبية لـ 248 مركبًا فرديًا، تمثل 23 فئة من المركبات و212 سمًّا عصبيًا طبيعيًا (Masjosthusmann وآخرون، 2018). تم تجميع آليات العمل المحددة وفقًا للأحداث الرئيسية الشائعة لـ ANT بما في ذلك النقل العصبي الكوليني، GABAergic، glycinergic، glutamatergic، adrenergic، serotonergic، وdopaminergic، وقنوات/مستقبلات الأيونات (مثل قنوات الصوديوم، قنوات البوتاسيوم، قنوات الكالسيوم، قنوات الكلوريد)، ومجموعة من النقاط النهائية الخلوية مثل خلل الميتوكوندريا، الإجهاد التأكسدي، موت الخلايا المبرمج، دورة الأكسدة والاختزال، تغيير إشارات الكالسيوم، تغييرات الهيكل الخلوي، الالتهاب العصبي، اعتلال المحاور، سمية المايلين، اعتلال الأعصاب المتأخر، وتثبيط الإنزيمات (Masjosthusmann وآخرون، 2018). لتمكين تقييم شامل لإمكانات ANT للمواد الكيميائية، من الضروري تجميع ANT-IVB v1.0 التي تشمل جميع آليات العمل المحددة، وهو تحدٍ سيتم معالجته في مشروعنا.
حتى الآن، تم إنشاء ونشر عدة اختبارات تغطي العناصر الأساسية الحرجة للتسمم العصبي (شميت وآخرون، 2017) لكنها لا تزال بحاجة إلى تحسين لتلبية معايير معينة لقبول الطرق البديلة من قبل الجهات التنظيمية، بما في ذلك تقييم نطاق القابلية للتطبيق، وقوة الاختبار وملاءمته، وإثبات القدرة التنبؤية للتسمم العصبي لدى البالغين (بال-برايس وآخرون، 2018). لتقييم التنشيط المباشر لقنوات الأيونات والمستقبلات وتغير وظيفة القنوات والمستقبلات في الخلايا العصبية الحسية (الألمية)، يمكن إعادة استخدام عدة طرق بديلة مستخدمة في DNTIVB v2.0 للاختبار الحاد في أنظمة الثقافة الناضجة ثنائية أو ثلاثية الأبعاد أو في مراحل الحياة المبكرة لسمك الزرد بعد تكوين الأعصاب. على وجه التحديد، سيتم تنفيذ طرق اختبار DNT-IVB v2.0، بما في ذلك طرق NAMs الخاصة بالميالين وحاجز الدم في الدماغ، ورسم الخلايا في كرات الدماغ البشرية (هارتمان وآخرون، 2023)، وطرق NAMs الخاصة بتعلم وذاكرة سمك الزرد، والتسمم في نظام الحركة، وطرق NAMs الشبيهة بالقلق في الثقافات الخلوية الناضجة أو أجنة سمك الزرد في نقاط زمنية تحدث بعد حدوث تكوين الأعصاب الأولي والتمايز (ر. شميت وآخرون، 2013).
بالإضافة إلى NAMs المعاد استخدامها DNT-IVB v2.0، يتم تطوير وتطبيق عدة NAMs جديدة على ANT-IVB v1.0. أحدها هو NAM تم تطويره مؤخرًا يعتمد على خلايا عصبية حسية ناضجة غنية بمستقبلات الألم مشتقة من hiPSC (هولزر وآخرون، 2022). باستخدام هذا NAM بعد 23 يومًا من التمايز، سيتم إجراء تعرض حاد للمواد الكيميائية المرجعية ANT لتقييم نطاق التطبيق البيولوجي للاختبار. آخر هو اختبار الناقلات العصبية المتعددة البشرية (hMNR)، الذي يعتمد على كرات دماغية ثلاثية الأبعاد مختلطة من الخلايا العصبية والدبقية مشتقة من hiPSC. يقيم NAM hMNR السمية العصبية الحادة المحددة لنوع الخلايا العصبية باستخدام الميكروإلكترود. المصفوفات (MEA) لتسجيل النشاط الكهربائي التلقائي (هارتمن وآخرون، 2023). من خلال فرز الإشارات المكتشفة (النبضات) بناءً على شكل الموجة الخاص بها، يسمح هذا الاختبار بالتمييز بين الاستجابات الجلوتامينية، والـ GABAergics، والدوبامينية، والسيروتونينية، والكولينرجيك في الثقافة المختلطة من الخلايا العصبية، مما يسمح بتقييم قائم على آلية العمل في المختبر لـ ANT (هارتمن وآخرون، 2023). في NAM الثالث،سيتم تقييم الإشارات على مستوى الخلية الواحدة في الخلايا العصبية الدوبامينية المركزية الناضجة (خلايا LUHMES)، مما يوسع نطاق تغطية KEs إلى نوع خلية آخر ومجموعة من المستقبلات الوظيفية (مثل مستقبلات P2X3 (أبيشيلا وفابريتي، 2012)). يتم تطوير NAM رابع، وهو اختبار سمية المايلين المحيطي (PeriMyelinTox)، لتقييم سمية المايلين التي تؤثر على الوظائف الحسية والحركية المحيطية، وبالتالي يعالج الفجوة الرئيسية في البطارية “التMyelination” للـ ANT المحيطي في الخلايا البشرية. في هذا NAM، سيتم تمايز الخلايا العصبية الحسية أو الحركية، جنبًا إلى جنب مع خلايا شوان، من الخلايا الجذعية المستحثة متعددة القدرات البشرية (hiPSCs) وزراعتها في زراعة مشتركة (مولر وآخرون، 2018؛ شينكي وآخرون، 2020؛ لويت وآخرون، 2023). سيتم تطوير تنسيق كروي ثلاثي الأبعاد ومقارنته بتنسيق ثنائي الأبعاد تقليدي. سيتم تقييم تكوين المايلين من خلال قياس بروتين المايلين الأساسي (MBP) أو بروتين المايلين صفر (MPZ) مقابل العلامة الكاملة للعصبونات.-توبولين (TUJ1) من خلال صبغة المناعة الفلورية وRT-qPCR (تشسنت وآخرون، 2021). سيتم تحسين الاختبار للت quantification الآلي عالي الإنتاجية للميالين بعد التعرض لمجموعة تدريب من المواد السامة المحتملة للميالين. سيتم تقييم القيمة المضافة لكلا الاختبارين في تقييم سمية الميالين في الأعصاب الحسية والحركية بشكل أكبر.
لتحديد مجالات التطبيق بوضوح والقيود الخاصة بالاختبارات، يهدف هذا المشروع إلى تطوير مجموعة من المواد الكيميائية المرجعية ANT المعروفة بتأثيرها على الدماغ البشري، بالإضافة إلى المركبات السلبية. ستضمن هذه الطريقة التوصيف الموحد لمجال تطبيق كل اختبار وتغطية آليات العمل المهمة المتعلقة بالبشر.
7 النتائج وآفاق المستقبل
تم تصميم PARC لمعالجة التحديات المتأصلة في الانتقال من طرق الاختبار المعتمدة على الحيوانات إلى (مجموعة من) الاختبارات في المختبر والطرق البديلة (NAMs) لتسريع وتحديث تحديد المخاطر وتقييم المخاطر الكيميائية. يهدف المشروع 5.2.1e إلى تحسين نموذج التنبؤ بالمخاطر من خلال إنشاء وتنقيح NAMs لاختبار DNT وANT وتجميع NAMs عالية الأداء وقابلة للتكرار التي تضيف قيمة إلى مجموعات اختبارات DNT وANT. تشمل استراتيجيتنا تنقيح الاختبارات الحالية، وتوليد NAMs مبتكرة لمعالجة الفجوات المحددة، وتحديد نطاق القابلية للتطبيق، وزيادة الجدوى الاقتصادية للاختبارات الأطول من خلال إظهار مؤشرات مبكرة للتأثيرات اللاحقة. من المهم أن يتضمن تطوير اختبارات جديدة وتنقيح الاختبارات الحالية تركيزًا قويًا على موثوقية الاختبارات والبيانات. إحدى الطموحات الحاسمة هي إدخال تدابير مراقبة الجودة كما هو موضح في ممارسات زراعة الخلايا الجيدة 2.0 (Pamies et al., 2022) أو وثيقة GIVIMP الخاصة بـ OECD (OECD, 2018). وبالتالي، يساهم اتحاد PARC 5.2.1e في تحسين الجاهزية والحساسية والأداء العام لـ NAMs الخاصة بـ DNT لتعزيز قبول أكبر للاختبارات البديلة والمختبرية لـ DNT للاستخدام التنظيمي الأوسع. بحلول نهاية هذا المشروع، سيتم تسليم وثيقة إرشادية ستقدم إطار عمل جديد. تهدف إلى تسهيل الاستخدام التنظيمي للبيانات المستمدة من DNTIVB v2.0. ستشمل هذه الأعمال اعتبارات حول كيفية استخدام بيانات DNT-IVB في سياق IATA أو وزن الأدلة لتوصيف المخاطر والتهديدات. يرتبط هذا بسلاسة مع حزم العمل الأخرى في PARC التي تقدم معلومات حول النمذجة الحركية المستندة إلى الفسيولوجيا (PBK) لتحويل تركيزات IVB إلى جرعات متوقعة في الجسم الحي، ومع متخصصي تقييم المخاطر الذين يحتاجون إلى النظر في كيفية استخدام الجرعات المتوقعة لتحديد عتبات التعرض الآمن من خلال، على سبيل المثال، النظر في العوامل المعدلة في AOP أو من خلال النظر في التباينات والحساسيات المحددة في السكان المعرضين (Schmeisser et al., 2023; Suciu et al., 2023).
على عكس DNT، لا يوجد حاليًا مجموعة مماثلة من NAMs لاختبار ANT. لذلك، سيقوم التحالف بإنشاء الجيل الأول من ANT-IVB v1.0، الذي يغطي الآليات الرئيسية المعنية في وظائف الدماغ البشري. مع النظر إلى المستقبل، واستجابةً لطلبات زملاء التنظيم في PARC للحصول على بيانات حول فئات المركبات المحددة، سيتم استخدام NAMs 5.2.1e لتقييم السمية المحتملة للسموم الطبيعية، والبيسفينولات، والمواد الكيميائية من عائلة البير- والبوليفلوروالكيل (PFAS). بشكل عام، يهدف هذا التحالف إلى تقديم فرصة غير مسبوقة لسد فجوة طويلة الأمد في التقييم الشائع لإمكانية السمية العصبية للمواد الكيميائية التجارية من خلال إنشاء استراتيجيات اختبار DNT وANT قائمة على الآلية، قوية، قابلة للتكرار، سريعة، وغير مكلفة. حيث يتم إجراء كل هذا العمل تحت إشراف زملاء من مجال التنظيم والمستخدمين المحتملين، فإن طموحنا هو إحداث ثورة في تقييم المخاطر والمخاطر لـ DNT وANT في أوروبا.
بيان توفر البيانات
المساهمات الأصلية المقدمة في الدراسة مدرجة في المقالة/المواد التكميلية، يمكن توجيه الاستفسارات الإضافية إلى المؤلفين المراسلين.
مساهمات المؤلفين
TT: التصور، الحصول على التمويل، إدارة المشروع، كتابة المسودة الأصلية، كتابة المراجعة والتحرير. OM: كتابة المراجعة والتحرير، الحصول على التمويل، إدارة المشروع. EF: كتابة المراجعة والتحرير، الحصول على التمويل. JR: كتابة المراجعة والتحرير، الحصول على التمويل. KC: كتابة المراجعة والتحرير. KAH: كتابة المراجعة والتحرير. CA: كتابة المراجعة والتحرير. PB: كتابة المراجعة والتحرير، الحصول على التمويل. BE: كتابة المراجعة والتحرير، الحصول على التمويل. HD: كتابة المراجعة والتحرير، الحصول على التمويل. KH: كتابة المراجعة والتحرير. KD: كتابة المراجعة والتحرير. YH: كتابة المراجعة والتحرير. SH: كتابة المراجعة والتحرير، الحصول على التمويل. KJ: كتابة المراجعة والتحرير. BJ: كتابة المراجعة والتحرير. NK: كتابة المراجعة والتحرير. AK-G: كتابة المراجعة والتحرير. BK: كتابة المراجعة والتحرير. MLe: كتابة المراجعة والتحرير، الحصول على التمويل. MLi: كتابة المراجعة والتحرير. JL: كتابة المراجعة والتحرير، الحصول على التمويل. FM: كتابة المراجعة والتحرير، الحصول على التمويل. JC: كتابة المراجعة والتحرير، الحصول على التمويل. WN: كتابة المراجعة والتحرير، الحصول على التمويل. GP: كتابة المراجعة والتحرير. BS: كتابة المراجعة والتحرير، الحصول على التمويل. IS: كتابة المراجعة والتحرير. SS: كتابة المراجعة. والمراجعة والتحرير، الحصول على التمويل. OL: الكتابة – المراجعة والتحرير، الحصول على التمويل. MT-R: الكتابة – المراجعة والتحرير، الحصول على التمويل. KB: التصور، إدارة المشروع، كتابة المسودة الأصلية، الكتابة – المراجعة والتحرير.
تمويل
يعلن المؤلفون أن الدعم المالي قد تم تلقيه للبحث والتأليف و/أو نشر هذه المقالة. تم تنفيذ هذا العمل في إطار الشراكة الأوروبية لتقييم المخاطر الناتجة عن المواد الكيميائية (PARC) وقد حصل على تمويل من برنامج الأبحاث والابتكار التابع للاتحاد الأوروبي هورايزون أوروبا بموجب اتفاقية المنحة رقم 101057014. ومع ذلك، فإن الآراء والمواقف المعبر عنها هي فقط آراء المؤلفين ولا تعكس بالضرورة آراء الاتحاد الأوروبي أو الوكالة التنفيذية للصحة والرقمنة. لا يمكن تحميل الاتحاد الأوروبي أو السلطة المانحة المسؤولية عنها. كما تم دعم هذا العمل من خلال جائزة W2 من جمعية هلمهولتز إلى T. Tal. الآراء المعبر عنها في هذا المستند هي آراء المؤلفين وليست بالضرورة آراء وكالات الاتحاد الأوروبي. أي بيان يتعلق بتنظيمات الاتحاد الأوروبي يهدف فقط لأغراض المعلومات ولا يمثل الرأي الرسمي للوكالة الأوروبية للمواد الكيميائية (ECHA). لا تتحمل ECHA أي مسؤولية عن أي استخدام قد يتم للمعلومات الواردة في هذا المستند. البيانات المقدمة أو المعلومات الواردة في هذه المنشورة هي دون الإخلال بأي عمل مستقبلي قد تبدأه ECHA. تم التمويل بشكل وثيق مع برنامج الأبحاث التابع للاتحاد الأوروبي هورايزون 2020
برنامج الابتكار بموجب اتفاقيات المنحة رقم 964518 (ToxFree) ورقم 964537 (RISK-HUNT3R) ورقم 965406 (PrecisionTox) ورقم 963845 (ONTOX) في مجموعة ASPIS. تم إنشاء الأشكال باستخدامbiorender.com.
شكر وتقدير
نشكر شوكو فورونو (ECHA) على مدخلاتها الخبيرة.
تعارض المصالح
EF و KB هما مساهمان في DNTOX GmbH التي تقدم خدمات اختبار السمية العصبية.
يعلن المؤلفون المتبقون أن البحث تم إجراؤه في غياب أي علاقات تجارية أو مالية يمكن أن تُفسر على أنها تعارض محتمل للمصالح.
ملاحظة الناشر
جميع المطالبات المعبر عنها في هذه المقالة هي فقط تلك الخاصة بالمؤلفين ولا تمثل بالضرورة تلك الخاصة بالمنظمات التابعة لهم، أو تلك الخاصة بالناشر، أو المحررين والمراجعين. أي منتج قد يتم تقييمه في هذه المقالة، أو أي ادعاء قد يتم من قبل مصنّعه، لا يضمنه أو يؤيده الناشر.
References
Ankley, G. T., Bennett, R. S., Erickson, R. J., Hoff, D. J., Hornung, M. W., Johnson, R. D., et al. (2010). Adverse outcome pathways: a conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 29 (3), 730-741. doi:10.1002/ETC. 34
Apicella, L., and Fabbretti, E. (2012). P2X3 receptor expression by HEK cells conditions their survival. Purinergic Signal. 8 (2), 295-300. doi:10.1007/S11302-011-9285-0
Appelt-Menzel, A., Cubukova, A., Günther, K., Edenhofer, F., Piontek, J., Krause, G., et al. (2017). Establishment of a human blood-brain barrier Co-culture model mimicking the neurovascular unit using induced pluri-and multipotent stem cells. Stem Cell Rep. 8, 894-906. doi:10.1016/j.stemcr.2017.02.021
Aung, M. T., Eick, S. M., Padula, A. M., Smith, S., Park, J. S., DeMicco, E., et al. (2023). Maternal per- and poly-fluoroalkyl substances exposures associated with higher depressive symptom scores among immigrant women in the chemicals in our bodies cohort in san francisco. Environ. Int. 172 (February), 107758. doi:10.1016/J. ENVINT.2023.107758
Babin, P. J., Goizet, C., and Raldúa, D. (2014). Zebrafish models of human motor neuron diseases: advantages and limitations. Prog. Neurobiol. 118, 36-58. doi:10.1016/J. PNEUROBIO.2014.03.001
Baccarelli, A., and Bollati, V. (2009). Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 21 (2), 243-251. doi:10.1097/MOP.0B013E32832925CC
Bal-Price, A., Hogberg, H. T., Crofton, K. M., Daneshian, M., FitzGerald, R. E., Fritsche, E., et al. (2018). Recommendation on test readiness criteria for new approach methods in Toxicology: exemplified for developmental neurotoxicity. Altex 35 (3), 306-352. doi:10.14573/altex. 1712081
Banks, W. A. (2009). Characteristics of compounds that cross the blood-brain barrier. BMC Neurol. 9 (Suppl. 1), S3-S9. doi:10.1186/1471-2377-9-S1-S3
Bartmann, K., Bendt, F., Dönmez, A., Haag, D., Eike Keßel, H., Masjosthusmann, S., et al. (2023). A human IPSC-based in vitro neural network formation assay to investigate neurodevelopmental toxicity of pesticides. ALTEX – Altern. Animal Exp. 40, 452-470. doi:10.14573/ALTEX. 2206031
Baumann, J., Barenys, M., Gassmann, K., and Fritsche, E. (2014). Comparative human and rat ‘neurosphere assay’ for developmental neurotoxicity testing. Curr. Protoc. Toxicol. 1 (Suppl. 59), 12-24. doi:10.1002/0471140856.TX1221S59
Baumann, J., Dach, K., Barenys, M., Giersiefer, S., Goniwiecha, J., Lein, P. J., et al. (2015). “Application of the neurosphere assay for DNT hazard assessment: challenges and limitations,” in Methods in pharmacology and Toxicology (Totowa, NJ: Humana Press), 29. doi:10.1007/7653_2015_49
Baumann, J., Gassmann, K., Masjosthusmann, S., DeBoer, D., Bendt, F., Giersiefer, S., et al. (2016). Comparative human and rat neurospheres reveal species differences in chemical effects on neurodevelopmental key events. Archives Toxicol. 90 (6), 1415-1427. doi:10.1007/S00204-015-1568-8
Bellanger, M., Demeneix, B., Grandjean, P., Thomas Zoeller, R., and Trasande, L. (2015). Neurobehavioral deficits, diseases, and associated costs of exposure to endocrine-disrupting chemicals in the European union. J. Clin. Endocrinol. Metabolism 100 (4), 1256-1266. doi:10.1210/JC.2014-4323
Bellanger, M., Pichery, C., Aerts, D., Berglund, M., Castaño, A., Čejchanová, M., et al. (2013). Economic benefits of methylmercury exposure control in Europe: monetary value of neurotoxicity prevention. Environ. Health A Glob. Access Sci. Source 12 (1), 3-10. doi:10.1186/1476-069x-12-3
Bennett, D., Bellinger, D. C., Birnbaum, L. S., Bradman, A., Chen, A., Cory-Slechta, D. A., et al. (2016). Project TENDR: targeting environmental neuro-developmental risks the TENDR consensus statement. Environ. Health Perspect. 124 (7), A118-A122. doi:10. 1289/EHP358
Bernal, J., Guadaño-Ferraz, A., and Morte, B. (2015). Thyroid hormone transporters-functions and clinical implications. Nat. Rev. Endocrinol. 11 (7), 690-717. doi:10.1038/ nrendo. 2015.186
Blum, J., Masjosthusmann, S., Bartmann, K., Bendt, F., Dolde, X., Dönmez, A., et al. (2022). Establishment of a human cell-based in vitro battery to assess developmental neurotoxicity hazard of chemicals. Chemosphere 311 (Pt 2), 137035. doi:10.1016/J. CHEMOSPHERE.2022.137035
Braunbeck, T., Kais, B., Lammer, E., Otte, J., Schneider, K., Stengel, D., et al. (2014). The fish embryo test (FET): origin, applications, and future. Environ. Sci. Pollut. Res. 22 (21), 16247-16261. doi:10.1007/S11356-014-3814-7
Bray, M. A., Singh, S., Han, H., Davis, C. T., Kost-Alimova, M., Gustafsdottir, S. M., et al. (2016). Cell painting, a high-content image-based assay for morphological profiling using multiplexed fluorescent dyes. Nat. Protoc. 11 (9), 1757-1774. doi:10. 1038/NPROT.2016.105
Brown, J. P., Hall, D., Frank, C. L., Wallace, K., Mundy, W. R., and Shafer, T. J. (2016). Editor’s highlight: evaluation of a microelectrode array-based assay for neural network ontogeny using training set chemicals. Toxicol. Sci. 154 (1), 126-139. doi:10.1093/ toxsci/kfw147
Brüll, M., Spreng, A.-S., Gutbier, S., Loser, D., Krebs, A., Reich, M., et al. (2020). Incorporation of stem cell-derived astrocytes into neuronal organoids to allow neuroglial interactions in toxicological studies. ALTEX – Altern. Animal Exp. 37 (3), 409-428. doi:10.14573/ALTEX. 1911111
Bruni, G., Rennekamp, A. J., Velenich, A., McCarroll, M., Gendelev, L., Fertsch, E., et al. (2016). Zebrafish behavioral profiling identifies multitarget antipsychotic-like compounds. Nat. Chem. Biol. 12 (7), 559-566. doi:10.1038/nchembio. 2097
Carson, M. J., Cameron Thrash, J., and Walter, B. (2006). The cellular response in neuroinflammation: the role of leukocytes, microglia and astrocytes in neuronal death and survival. Clin. Neurosci. Res. 6 (5), 237-245. doi:10.1016/J.CNR.2006.09.004
Chesnut, M., Paschoud, H., Repond, C., Smirnova, L., Hartung, T., Zurich, M.-G., et al. (2021). Human IPSC-derived model to study myelin disruption. Int. J. Mol. Sci. 22 (17), 9473. doi:10.3390/IJMS22179473
Chu, J., and Sadler, K. C. (2009). New school in liver development: lessons from zebrafish. Hepatology 50 (5), 1656-1663. doi:10.1002/HEP. 23157
Costa, L. G., Aschner, M., Vitalone, A., Syversen, T., and Soldin, O. P. (2004). Developmental neuropathology of environmental agents. Annu. Rev. Pharmacol. Toxicol. 44, 87-110. doi:10.1146/annurev.pharmtox.44.101802.121424
Costa, L. G., Giordano, G., Guizzetti, M., and Vitalone, A. (2008). Neurotoxicity of pesticides: a brief review. Front. Biosci. 13 (4), 1240-1249. doi:10.2741/2758
Craig, E., Kelly, L., Akerman, G., Dawson, J., May, B., Reaves, E., et al. (2019). Reducing the need for animal testing while increasing efficiency in a pesticide regulatory setting: lessons from the EPA office of pesticide programs’ hazard and science policy Council. Regul. Toxicol. Pharmacol. RTP 108 (November), 104481. doi:10.1016/J. YRTPH.2019.104481
Crofton, K. M., and Mundy, W. R. (2021). External scientific report on the interpretation of data from the developmental neurotoxicity in vitro testing assays for use in integrated approaches for testing and assessment. EFSA Support. Publ. 18 (10). doi:10.2903/SP.EFSA.2021.EN-6924
Crofton, K. M., Mundy, W. R., and Shafer, T. J. (2012). Developmental neurotoxicity testing: a path forward. Congenit. Anomalies. Congenit. Anom. (Kyoto) 52, 140-146. doi:10.1111/j.1741-4520.2012.00377.x
Dach, K., Bendt, F., Huebenthal, U., Giersiefer, S., Lein, P. J., Heuer, H., et al. (2017). BDE-99 impairs differentiation of human and mouse NPCs into the oligodendroglial lineage by species-specific modes of action. Sci. Rep. 7 (44861), 44861. doi:10.1038/ srep44861
Davidsen, N., Lauvås, A. J., Myhre, O., Ropstad, E., Carpi, D., Gyves, E. M. de, et al. (2021). Exposure to human relevant mixtures of halogenated persistent organic pollutants (POPs) alters neurodevelopmental processes in human neural stem cells undergoing differentiation. Reprod. Toxicol. 100 (April 2020), 17-34. doi:10.1016/j. reprotox.2020.12.013
Delp, J., Cediel-Ulloa, A., Suciu, I., Kranaster, P., Vugt-Lussenburg, B.Ma V., Munic Kos, V., et al. (2021). Neurotoxicity and underlying cellular changes of 21 mitochondrial respiratory chain inhibitors. Archives Toxicol. 95, 591-615. doi:10.1007/s00204-020-02970-5
Delp, J., Funke, M., Rudolf, F., Cediel, A., Bennekou, S. H., van der Stel, W., et al. (2019). Development of a neurotoxicity assay that is tuned to detect mitochondrial toxicants. Archives Toxicol. 93 (6), 1585-1608. doi:10.1007/S00204-019-02473-Y
Dickerson, A. S., Wu, A. C., Liew, Z., and Weisskopf, M. (2020). A scoping review of non-occupational exposures to environmental pollutants and adult depression, anxiety, and suicide. Curr. Environ. Health Rep. 7 (3), 256-271. doi:10.1007/s40572-020-00280-7
Druwe, I., Freudenrich, T. M., Wallace, K., Shafer, T. J., and Mundy, W. R. (2015). Sensitivity of neuroprogenitor cells to chemical-induced apoptosis using a multiplexed assay suitable for high-throughput screening. Toxicology 333 (July), 14-24. doi:10.1016/ J.TOX.2015.03.011
Duboc, V., Dufourcq, P., Blader, P., and Roussigné, M. (2015). Asymmetry of the brain: development and implications. Annu. Rev. Genet. 49, 647-672. doi:10.1146/ ANNUREV-GENET-112414-055322
Dubrana, L. E., Knoll-Gellida, A., Bourcier, L. M., Mercé, T., Pedemay, S., Nachon, F., et al. (2021). An antidote screening system for organophosphorus poisoning using zebrafish larvae. ACS Chem. Neurosci. 12 (15), 2865-2877. doi:10.1021/ ACSCHEMNEURO.1C00251
ECHA (2023). Key areas of regulatory challenge. Eur. Chem. Agency. ECHA-23-R-(ED-07-23-212-EN-N). doi:10.2823/568850
EFSA Scientific CommitteeHardy, A., Benford, D., Halldorsson, T., Jeger, M. J., Katrine Knutsen, H., More, S., et al. (2017). Guidance on the risk assessment of substances present in food intended for infants below 16 Weeks of age. EFSA J. 15 (5), e04849. doi:10.2903/J.EFSA.2017.4849
Eriksson, P. S., Perfilieva, E., Björk-Eriksson, T., Alborn, A. M., Nordborg, C., Peterson, D. A., et al. (1998). Neurogenesis in the adult human Hippocampus. Nat. Med. 4 (11), 1313-1317. doi:10.1038/3305
Escher, B. I., Altenburger, R., Blüher, M., Colbourne, J. K., Ebinghaus, R., Fantke, P., et al. (2023). Modernizing persistence-bioaccumulation-toxicity (PBT) assessment with high throughput animal-free methods. Archives Toxicol. 97 (5), 1267-1283. doi:10. 1007/S00204-023-03485-5
European Commission (2020). Communication from the commission to the European parliament, the Council, the European economic and social committee and the committee of the regions. Chem. Strategy Sustain. – Towards a Toxic-Free Environ.
European Parliament and Council. 2006. “Regulation (EC) No. 1907/2006 of the European parliament and of the Council of 18 december 2006 concerning the registration, evaluation, authorisation and restriction of chemicals (REACH), establishing a European chemicals agency, amending directive 1999/45/EC and repealing Council regulation (EEC) No 793/93 and commission regulation (EC) No 1488/94 as well as Council directive 76/769/EEC and commission directives 91/155/ EEC, 93/67/EEC, 93/105/EC and 2000/21/EC.”
European Parliament and Council. 2008. “Regulation (EC) No. 1272/2008 of the European parliament and the Council of 16 december 2008 on classification, labelling and packaging of substances, and mixtures, amending and repealing directives 67/548/ EEC and 1999/45/EC, and amending regulation (EC) No 1907/2006.” In .
European Parliament and Council (2009). Regulation (EC) No. 1107/2009 of the European parliament and of the Council of 21 october 2009 concerning the placing of plant protection products on the market and repealing Council directives 79/117/EEC and 91/414/EEC. Paris: Brussels, OECD.
European Parliament and Council. 2010. “Directive 2010/63/EU of the European paliament and of the Council of 22 september 2010 on the protection of animals used for scientific purposes.” In .
Faria, M., Garcia-Reyero, N., Padrós, F., Babin, P. J., Sebastián, D., Cachot, J., et al. (2015). Zebrafish models for human acute organophosphorus poisoning. Sci. Rep. 5 (October), 15591. doi:10.1038/SREP15591
Farrell, J. A., Wang, Y., Riesenfeld, S. J., Shekhar, K., Regev, A., and Schier, A. F. (2018). Single-cell reconstruction of developmental trajectories during zebrafish embryogenesis. Science 360 (6392), eaar3131. doi:10.1126/science.aar3131
Frank, C. L., Brown, J. P., Wallace, K., Mundy, W. R., and Shafer, T. J. (2017). From the cover: developmental neurotoxicants disrupt activity in cortical networks on microelectrode arrays: results of screening 86 compounds during neural network formation. Toxicol. Sci. 160 (1), 121-135. doi:10.1093/toxsci/kfx169
Fritsche, E. (2017). “Report on integrated testing strategies for the identification and evaluation of chemical hazards associated with the developmental neurotoxicity (DNT), to facilitate discussions at the Joint EFSA/OECD workshop on DNT.” ENV/JM/ MONO(2017)4/ANN1.
Fritsche, E., Grandjean, P., Crofton, K. M., Aschner, M., Goldberg, A., Heinonen, T., et al. (2018). Consensus statement on the need for innovation, transition and implementation of developmental neurotoxicity (DNT) testing for regulatory purposes. Toxicol. Appl. Pharmacol. 354 (September), 3-6. doi:10.1016/J.TAAP.2018.02.004
Gaballah, S., Swank, A., Sobus, J. R., Meng Howey, X., Schmid, J., Catron, T., et al. (2020). Evaluation of developmental toxicity, developmental neurotoxicity, and tissue dose in zebrafish exposed to GenX and other PFAS. Environ. Health Perspect. 128 (4), 47005. doi:10.1289/EHP5843
Gadaleta, D., Spînu, N., Roncaglioni, A., Cronin, M. T. D., and Benfenati, E. (2022). Prediction of the neurotoxic potential of chemicals based on modelling of molecular initiating events upstream of the adverse outcome pathways of (developmental) neurotoxicity. Int. J. Mol. Sci. 23 (6), 3053. doi:10.3390/ijms23063053
Giordano, G., and Costa, L. G. (2012). Developmental neurotoxicity: some old and new issues. ISRN Toxicol. 2012 (June), 814795-814812. doi:10.5402/2012/814795
Glaberman, S., Padilla, S., and Barron, M. G. (2017). Evaluating the zebrafish embryo toxicity test for pesticide hazard screening. Environ. Toxicol. Chem. 36 (5), 1221-1226. doi:10.1002/ETC. 3641
Grandjean, P., and Landrigan, P. J. (2006). Developmental neurotoxicity of industrial chemicals. Lancet 368 (9553), 2167-2178. doi:10.1016/S0140-6736(06)69665-7
Grandjean, P., and Landrigan, P. J. (2014). Neurobehavioural effects of developmental toxicity. Lancet Neurology 13 (3), 330-338. doi:10.1016/S1474-4422(13)70278-3
Grillberger, K., Cöllen, E., Immacolata Trivisani, C., Blum, J., Leist, M., and Ecker, G. F. (2023). Structural insights into neonicotinoids and N -unsubstituted metabolites on human NAChRs by molecular docking, dynamics simulations, and calcium imaging. Int. J. Mol. Sci. 24 (17), 13170. doi:10.3390/IJMS241713170
Gunnarsson, L., Jauhiainen, A., Kristiansson, E., Nerman, O., and Joakim Larsson, D. G. (2008). Evolutionary conservation of human drug targets in organisms used for
environmental risk assessments. Environ. Sci. Technol. 42 (15), 5807-5813. doi:10.1021/ es8005173
Gupta, T., Marquart, G. D., Horstick, E. J., Tabor, K. M., Pajevic, S., and Burgess, H. A. (2018). Morphometric analysis and neuroanatomical mapping of the zebrafish brain. Methods 150 (November), 49-62. doi:10.1016/J.YMETH.2018.06.008
Gutbier, S., Spreng, A. S., Delp, J., Schildknecht, S., Karreman, C., Suciu, I., et al. (2018). Prevention of neuronal apoptosis by astrocytes through thiol-mediated stress response modulation and accelerated recovery from proteotoxic stress. Cell Death Differ. 25 (12), 2101-2117. doi:10.1038/S41418-018-0229-X
Harrill, J. A., Freudenrich, T., Wallace, K., Ball, K., Shafer, T. J., and Mundy, W. R. (2018). Testing for developmental neurotoxicity using a battery of in vitro assays for key cellular events in neurodevelopment. Toxicol. Appl. Pharmacol. 354 (September), 24-39. doi:10.1016/J.TAAP.2018.04.001
Harrill, J. A., Freudenrich, T. M., Machacek, D. W., Stice, S. L., and Mundy, W. R. (2010). Quantitative assessment of neurite outgrowth in human embryonic stem cellderived HN2 cells using automated high-content image analysis. Neurotoxicology 31 (3), 277-290. doi:10.1016/J.NEURO.2010.02.003
Harrill, J. A., Robinette, B. L., Freudenrich, T., and Mundy, W. R. (2013). Use of high content image analyses to detect chemical-mediated effects on neurite sub-populations in primary rat cortical neurons. NeuroToxicology 34 (1), 61-73. doi:10.1016/J.NEURO. 2012.10.013
Harrill, J. A., Robinette, B. L., and Mundy, W. R. (2011). Use of high content image analysis to detect chemical-induced changes in synaptogenesis in vitro. Toxicol. Vitro 25 (1), 368-387. doi:10.1016/j.tiv.2010.10.011
Harry, G. J., McBride, S., Witchey, S. K., Mhaouty-Kodja, S., Trembleau, A., Bridge, M., et al. (2022). Roadbumps at the crossroads of integrating behavioral and in vitro approaches for neurotoxicity assessment. Front. Toxicol. 4, 812863. doi:10.3389/FTOX. 2022.812863
Hartmann, J., Henschel, N., Bartmann, K., Dönmez, A., Brockerhoff, G., Koch, K., et al. (2023). Molecular and functional characterization of different BrainSphere models for use in neurotoxicity testing on microelectrode arrays. Cells 12 (9), 1270. doi:10.3390/ cells12091270
Hoelting, L., Klima, S., Karreman, C., Grinberg, M., Meisig, J., Henry, M., et al. (2016). Stem cell-derived immature human dorsal root ganglia neurons to identify peripheral neurotoxicants. Stem Cells Transl. Med. 5 (4), 476-487. doi:10.5966/SCTM.2015-0108
Hollander, J. A., Cory-Slechta, D. A., Jacka, F. N., Szabo, S. T., Guilarte, T. R., Bilbo, S. D., et al. (2020). Beyond the looking glass: recent advances in understanding the impact of environmental exposures on neuropsychiatric disease. Neuropsychopharmacology 45 (7), 1086-1096. doi:10.1038/s41386-020-0648-5
Holzer, A. K., Karreman, C., Suciu, I., Furmanowsky, L. S., Wohlfarth, H., Loser, D., et al. (2022). Generation of human nociceptor-enriched sensory neurons for the study of pain-related dysfunctions. Stem Cells Transl. Med. 11 (7), 727-741. doi:10.1093/ STCLTM/SZAC031
Hooijmans, C. R., Leenaars, M., and Ritskes-Hoitinga, M. (2010). A gold standard publication checklist to improve the quality of animal studies, to fully integrate the three rs, and to make systematic reviews more feasible. Altern. Lab. Anim. 38 (2), 167-182. doi:10.1177/026119291003800208
Horzmann, K. A., and Freeman, J. L. (2016). Zebrafish get connected: investigating neurotransmission targets and alterations in chemical toxicity. Toxics 4 (3), 19. doi:10. 3390/TOXICS4030019
Howe, K., Clark, M. D., Torroja, C. F., Torrance, J., Berthelot, C., Muffato, M., et al. (2013). The zebrafish reference genome sequence and its relationship to the human genome. Nature 496, 498-503. doi:10.1038/nature12111
Imayoshi, I., Shimojo, H., Sakamoto, M., Ohtsuka, T., and Kageyama, R. (2013). Genetic visualization of Notch signaling in mammalian neurogenesis. Cell. Mol. Life Sci. 70, 2045-2057. doi:10.1007/s00018-012-1151-x
Jacobson, M. H., Ghassabian, A., Gore, A. C., and Trasande, L. (2022). Exposure to environmental chemicals and perinatal psychopathology. Biochem. Pharmacol. 195 (January), 114835. doi:10.1016/J.BCP.2021.114835
Jarema, K. A., Hunter, D. L., Hill, B. N., Olin, J. K., Britton, K. N., Waalkes, M. R., et al. (2022). Developmental neurotoxicity and behavioral screening in larval zebrafish with a comparison to other published results. Toxics 10 (5), 256. doi:10.3390/toxics10050256
Kalueff, A. V., Gebhardt, M., Stewart, A. M., Cachat, J. M., Brimmer, M., Chawla, J. S., et al. (2013). Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 10 (1), 70-86. Https://Home.Liebertpub.Com/Zeb. doi:10.1089/ZEB.2012. 0861
Kang, Du S., Yang, Y. R., Lee, C., Kim, S. B., Ryu, S.Ho, and Suh, P. G. (2016). Roles of phosphoinositide-specific phospholipase in brain development. Adv. Biol. Regul. 60 (January), 167-173. doi:10.1016/J.JBIOR.2015.10.002
Kavlock, R. J., Bahadori, T., Barton-Maclaren, T. S., Gwinn, M. R., Rasenberg, M., and Thomas, R. S. (2018). Accelerating the pace of chemical risk assessment. Chem. Res. Toxicol. 31 (5), 287-290. doi:10.1021/ACS.CHEMRESTOX.7B00339
Kettleborough, R. N. W., Busch-Nentwich, E. M., Harvey, S. A., Dooley, C. M., De Bruijn, E., Van Eeden, F., et al. (2013). A systematic genome-wide analysis of zebrafish protein-coding gene function. Nature 496 (7446), 494-497. doi:10.1038/ NATURE11992
Khelfaoui, H., Harkati, D., and Saleh, B. A. (2021). Molecular docking, molecular dynamics simulations and reactivity, studies on approved drugs library targeting ACE2 and SARS-CoV-2 binding with ACE2. J. Biomol. Struct. Dyn. 39 (18), 7246-7262. doi:10.1080/07391102.2020.1803967
Klima, S., Brüll, M., Spreng, A. S., Suciu, I., Falt, T., Schwamborn, J. C., et al. (2021). A human stem cell-derived test system for agents modifying neuronal N-Methyl-D-Aspartate-Type glutamate receptor -signalling. Archives Toxicol. 95 (5), 1703-1722. doi:10.1007/S00204-021-03024-0
Klose, J., Tigges, J., Masjosthusmann, S., Schmuck, K., Bendt, F., Hübenthal, U., et al. (2021). TBBPA targets converging key events of human oligodendrocyte development resulting in two novel AOPs. Altex 38 (2), 215-234. doi:10.14573/altex. 2007201
Klüver, N., Vogs, C., Altenburger, R., Escher, B. I., and Scholz, S. (2016). Development of a general baseline toxicity QSAR model for the fish embryo acute toxicity test. Chemosphere 164 (December), 164-173. doi:10.1016/J.CHEMOSPHERE.2016.08.079
Knoll-Gellida, A., Dubrana, L. E., Bourcier, L. M., eo Merc, Th, Gruel, E., Soares, M., et al. (2021). Hyperactivity and seizure induced by tricresyl phosphate are isomer specific and not linked to phenyl valerate-neuropathy target esterase activity inhibition in zebrafish. Toxicol. Sci. 180 (1), 160-174. doi:10.1093/toxsci/kfab006
Koch, K., Bartmann, K., Hartmann, J., Kapr, J., Pahl, M., Fritsche, E., et al. (2022). Scientific validation of human neurosphere assays for developmental neurotoxicity evaluation. Front. Press 0, 816370-816438. doi:10.3389/FTOX.2022.816370
Krebs, A., Waldmann, T., Wilks, M. F., van Vugt-Lussenburg, B. M. A., van der Burg, B., Terron, A., et al. (2019). Template for the description of cell-based toxicological test methods to allow evaluation and regulatory use of the data. ALTEX – Altern. Animal Exp. 36 (4), 682-699. doi:10.14573/ALTEX. 1909271
Krug, A. K., Balmer, N. V., Matt, F., Schönenberger, F., Merhof, D., and Leist, M. (2013). Evaluation of a human neurite growth assay as specific screen for developmental neurotoxicants. Archives Toxicol. 87 (12), 2215-2231. doi:10.1007/s00204-013-1072-y
Krzykwa, J. C., Saeid, A., and Sellin Jeffries, M. K. (2019). Identifying sublethal endpoints for evaluating neurotoxic compounds utilizing the fish embryo toxicity test. Ecotoxicol. Environ. Saf. 170 (April), 521-529. doi:10.1016/J.ECOENV.2018. 11.118
Landrigan, P. J., Sonawane, B., Butler, R. N., Trasande, L., Callan, R., and Droller, D. (2005). Early environmental origins of neurodegenerative disease in later life. Environ. Health Perspect. 113 (9), 1230-1233. doi:10.1289/EHP. 7571
Lauvås, A. J., Lislien, M., Andreas Holme, J., Dirven, H., Paulsen, E., Alm, I. M., et al. (2022). Developmental neurotoxicity of acrylamide and its metabolite glycidamide in a human mixed culture of neurons and astrocytes undergoing differentiation in concentrations relevant for human exposure. Neurotoxicology 92, 33-48. doi:10. 1016/j.neuro.2022.07.001
Lee, Da Y. (2015). Roles of MTOR signaling in brain development. Exp. Neurobiol. 24 (3), 177-185. doi:10.5607/EN.2015.24.3.177
Lee, J., Escher, B. I., Scholz, S., and Schlichting, R. (2022a). Inhibition of neurite outgrowth and enhanced effects compared to baseline toxicity in SH-SY5Y cells. Archives Toxicol. 96 (4), 1039-1053. doi:10.1007/s00204-022-03237-x
Lee, J., Schlichting, R., König, M., Scholz, S., Krauss, M., and Escher, B. I. (2022b). Monitoring mixture effects of neurotoxicants in surface water and wastewater treatment plant effluents with neurite outgrowth inhibition in SH-SY5Y cells. Am. Chem. Soc. 2, 523-535. doi:10.1021/acsenvironau.2c00026
Leist, M., Ahmed, G., Graepel, R., Marchan, R., Hassan, R., Hougaard Bennekou, S., et al. (2017). Adverse outcome pathways: opportunities, limitations and open questions. Archives Toxicol. 91 (11), 3477-3505. doi:10.1007/S00204-017-2045-3
López-Espíndola, D., Morales-Bastos, C., Grijota-Martínez, C., Liao, X. H., Lev, D., Sugo, E., et al. (2014). Mutations of the thyroid hormone transporter MCT8 cause prenatal brain damage and persistent hypomyelination. J. Clin. Endocrinol. Metabolism 99 (12), E2799-E2804. doi:10.1210/jc.2014-2162
Loser, D., Hinojosa, M. G., Blum, J., Schaefer, J., Brüll, M., Johansson, Y., et al. (2021). Functional alterations by a subgroup of neonicotinoid pesticides in human dopaminergic neurons. Archives Toxicol. 95 (6), 2081-2107. doi:10.1007/S00204-021-03031-1
Louit, A., Beaudet, M. J., Pépin, R., and Berthod, F. (2023). Differentiation of human induced pluripotent stem cells into mature and myelinating Schwann cells. Https// Home.Liebertpub.Com/Tec 29 (4), 134-143. doi:10.1089/TEN.TEC.2022.0186
Makris, S. L., Raffaele, K., Allen, S., Bowers, W. J., Hass, U., Alleva, E., et al. (2009). A retrospective performance assessment of the developmental neurotoxicity study in support of OECD test guideline 426. Environ. Health Perspect. 117 (1), 17-25. doi:10. 1289/EHP. 11447
Marx-Stoelting, P., Rivière, G., Luijten, • M., Aiello-Holden, • K., Bandow, • N., Baken, • K., et al. (2023). A walk in the PARC: developing and implementing 21st century chemical risk assessment in Europe. Arch. Toxicol. 97, 893-908. doi:10.1007/s00204-022-03435-7
Masjosthusmann, S., Barenys, M., El-Gamal, M., Geerts, L., Gerosa, L., Gorreja, A., et al. (2018). Literature review and appraisal on alternative neurotoxicity testing methods. EFSA Support. Publ. 15 (4), 1-108. doi:10.2903/sp.efsa.2018.en-1410
Masjosthusmann, S., Blum, J., Bartmann, K., Dolde, X., Holzer, A.-K., Stürzl, L.-C., et al. (2020). Establishment of an a priori protocol for the implementation and
interpretation of an in-vitro testing battery for the assessment of developmental neurotoxicity. EFSA Supporting Publication. Wiley. doi:10.2903/sp.efsa.2020.en-1938
Muller, Q., Beaudet, M. J., De Serres-Bérard, T., Bellenfant, S., Vincent, F., and Berthod, F. (2018). Development of an innervated tissue-engineered skin with human sensory neurons and Schwann cells differentiated from IPS cells. Acta Biomater. 82 (December), 93-101. doi:10.1016/J.ACTBIO.2018.10.011
Nimtz, L., Klose, J., Masjosthusmann, S., Barenys, M., and Fritsche, E. (2019). The neurosphere assay as an in vitro method for developmental neurotoxicity (DNT) evaluation. Eval. Neuromethods 145, 141-168. doi:10.1007/978-1-4939-9228-7_8
Noyes, P. D., Haggard, D. E., Gonnerman, G. D., and Tanguay, R. L. (2015). Advanced morphological – behavioral test platform reveals neurodevelopmental defects in embryonic zebrafish exposed to comprehensive suite of halogenated and organophosphate flame retardants. Toxicol. Sci. 145 (1), 177-195. doi:10.1093/ TOXSCI/KFV044
Nyffeler, J., Dolde, X., Krebs, A., Pinto-Gil, K., Pastor, M., Behl, M., et al. (2017). Combination of multiple neural crest migration assays to identify environmental toxicants from a proof-of-concept chemical library. Archives Toxicol. 91 (11), 3613-3632. doi:10.1007/s00204-017-1977-y
Ockleford, C., Adriaanse, P., Berny, P., Brock, T., Duquesne, S., Grilli, S., et al. (2017). Investigation into experimental toxicological properties of plant protection products having a potential link to Parkinson’s disease and childhood leukaemia. EFSA J. 15 (3), e04691. doi:10.2903/J.EFSA.2017.4691
OECD. (1995a). “Test No. 418: delayed neurotoxicity of organophosphorus substances following acute exposure.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070905-en
OECD. (1995b). “Test No. 419: delayed neurotoxicity of organophosphorus substances: 28-day repeated dose study.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070929-en
OECD. (1997). “Test No. 424: neurotoxicity study in rodents.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264071025-en
OECD. (2002a). “Test No. 420: acute oral toxicity – fixed dose procedure.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070943-en
OECD. (2002b). “Test No. 423: acute oral toxicity – acute toxic class method.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264071001-en
OECD. (2005). “Guidance document on the validation and international acceptance of new or updated test methods for hazard assessment, series on testing and assessment No. 34.” In .
OECD. 2007. “Test No. 426: developmental neurotoxicity study.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264067394-en
OECD (2008a). Series on testing and assessment No. 89 – retrospective performance assessment of the test guideline 426 on developmental neurotoxicity. ENV/JM/ MON (89).
OECD. 2008b. “Test No. 407: repeated dose 28-day oral toxicity study in rodents.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264070684-en
OECD. (2009). “Test No. 403: acute inhalation toxicity.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070608-en
OECD. (2017). “Test No. 402: acute dermal toxicity.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070585-en
OECD. (2018a). “Test No. 408: repeated dose 90-day oral toxicity study in rodents.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264070707-en
OECD. (2022). “Test No. 425: acute oral toxicity – up-and-down procedure.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264071049-en
OECD (2023). Initial recommendations on evaluation of data from the developmental neurotoxicity (DNT) in-vitro testing battery. Ser. Test. Assess. 377 (13). ENV/CBC/MONO(2023. Available at: https://www.oecd.org/env/ehs/testing/ developmental-neurotoxicity.htm.
Pallocca, G., Moné, M. J., Kamp, H., Luijten, M., van de Water, B., and Leist, M. (2022). Next-generation risk assessment of chemicals – rolling out a human-centric testing strategy to drive 3R implementation: the RISK-hunt3r project perspective. Altex 39 (3), 419-426. doi:10.14573/ALTEX. 2204051
Pamies, D., Leist, M., Coecke, S., Bowe, G., Allen, D. G., Gstraunthaler, G., et al. (2022). Guidance document on Good cell and tissue culture practice 2.0 (GCCP 2.0). ALTEX 39 (1), 30-70. doi:10.14573/ALTEX. 2111011
Paparella, M., Bennekou, S. H., and Bal-Price, A. (2020). An analysis of the limitations and uncertainties of in vivo developmental neurotoxicity testing and assessment to identify the potential for alternative approaches. Reprod. Toxicol. 96 (September), 327-336. doi:10.1016/J.REPROTOX.2020.08.002
Pitzer, E. M., Shafer, T. J., and Herr, D. W. (2023). Identification of Neurotoxicology (NT)/Developmental Neurotoxicology (DNT) adverse outcome pathways and key event linkages with in vitro DNT screening assays. Neurotoxicology 99 (December), 184-194. doi:10.1016/J.NEURO.2023.10.007
Raj, B., Wagner, D. E., McKenna, A., Pandey, S., Klein, A. M., Shendure, J., et al. (2018). Simultaneous single-cell profiling of lineages and cell types in the vertebrate brain. Nat. Biotechnol. 36 (5), 442-450. doi:10.1038/nbt. 4103
Rice, D., and Barone, S. (2000). Critical periods of vulnerability for the developing nervous system: evidence from humans and animal models. Environ. Health Perspect. 108 (Suppl. 3), 511-533. doi:10.1289/ehp.00108s3511
Rietdijk, J., Aggarwal, T., Georgieva, P., Lapins, M., Carreras-Puigvert, J., and Spjuth, O. (2022). Morphological profiling of environmental chemicals enables efficient and untargeted exploration of combination effects, Sci. Total Environ. 832, 155058. doi:10. 1016/j.scitotenv.2022.155058
Rokoff, L. B., Shoaff, J. R., Coull, B. A., Enlow, M. B., Bellinger, D. C., and Korrick, S. A. (2022). Prenatal exposure to a mixture of organochlorines and metals and internalizing symptoms in childhood and adolescence. Environ. Res. 208 (May), 112701. doi:10.1016/J.ENVRES.2022.112701
Sachana, M., Bal-Price, A., Crofton, K. M., Bennekou, S. H., Shafer, T. J., Behl, M., et al. (2019). International regulatory and scientific effort for improved developmental neurotoxicity testing. Toxicol. Sci. 167 (1), 45-57. doi:10.1093/toxsci/kfy211
Saili, K. S., Todd, J. Z., Schwab, A. J., Silvin, A., Baker, N. C., Sidney Hunter, E., et al. (2017). Blood-brain barrier development: systems modeling and predictive Toxicology. Birth Defects Res. 109 (20), 1680-1710. doi:10.1002/BDR2.1180
Sakai, C., Ijaz, S., Hoffman, E. J., Grünblatt, E., Carl, M., and Lange, M. (2018). Zebrafish models of neurodevelopmental disorders: past, present, and future. Front. Mol. Neurosci. 11, 294. doi:10.3389/fnmol.2018.00294
Sato-Maeda, M., Tawarayama, H., Obinata, M., Kuwada, J. Y., and Shoji, W. (2006). Sema3al guides spinal motor axons in a cell- and stage-specific manner in zebrafish. Dev. Camb. Engl. 133 (5), 937-947. doi:10.1242/DEV. 02268
Schenke, M., Schjeide, B. M., Püschel, G. P., and Seeger, B. (2020). Analysis of motor neurons differentiated from human induced pluripotent stem cells for the use in cellbased botulinum neurotoxin activity assays. Toxins 12 (5), 276. doi:10.3390/ TOXINS12050276
Schmeisser, S., Miccoli, A., von Bergen, M., Berggren, E., Braeuning, A., Busch, W., et al. (2023). New approach methodologies in human regulatory Toxicology – not if, but how and when. Environ. Int. 178 (March), 108082. doi:10.1016/J.ENVINT.2023.108082
Schmidt, B. Z., Lehmann, M., Gutbier, S., Nembo, E., Noel, S., Smirnova, L., et al. (2017). In vitro acute and developmental neurotoxicity screening: an overview of cellular platforms and high-throughput technical possibilities. Archives Toxicol. 91 (1), 1-33. doi:10.1007/S00204-016-1805-9
Schmidt, R., Strähle, U., and Scholpp, S. (2013). Neurogenesis in zebrafish – from embryo to adult. Neural Dev. 8 (1), 3-13. doi:10.1186/1749-8104-8-3
Schmuck, M. R., Temme, T., Dach, K., Boer, D. de, Barenys, M., Bendt, F., et al. (2017). Omnisphero: a high-content image analysis (hca) approach for phenotypic developmental neurotoxicity (DNT) screenings of organoid neurosphere cultures in vitro. Archives Toxicol. 91 (4), 2017-2028. doi:10.1007/s00204-016-1852-2
Semple, B. D., Blomgren, K., Gimlin, K., Ferriero, D. M., and Noble-Haeusslein, L. J. (2013). Brain development in rodents and humans: identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 106-107, 1-16. 0 (July). doi:10.1016/J.PNEUROBIO.2013.04.001
Shafer, T. J. (2019). Application of microelectrode array approaches to neurotoxicity testing and screening. Adv. Neurobiol. 22, 275-297. doi:10.1007/ 978-3-030-11135-9_12
Sheets, L. P., Li, A. A., Minnema, D. J., Collier, R. H., Creek, M. R., and Peffer, R. C. (2016). A critical review of neonicotinoid insecticides for developmental neurotoxicity. Crit. Rev. Toxicol. 46 (2), 153-190. doi:10.3109/10408444.2015. 1090948
Smirnova, L., Hogberg, H. T., Leist, M., and Hartung, T. (2014). Food for thought.: developmental neurotoxicity – challenges in the 21st century and in vitro opportunities. Altex 31 (2), 129-156. doi:10.14573/ALTEX. 1403271
Sobanska, M., Scholz, S., Nyman, A. M., Cesnaitis, R., Alonso, S. G., Klüver, N., et al. (2018). Applicability of the fish embryo acute toxicity (FET) test (OECD 236) in the regulatory context of registration, evaluation, authorisation, and restriction of chemicals (REACH). Environ. Toxicol. Chem. 37 (3), 657-670. doi:10.1002/ETC. 4055
Spalding, K. L., Bergmann, O., Alkass, K., Bernard, S., Salehpour, M., Huttner, H. B., et al. (2013). Dynamics of hippocampal neurogenesis in adult humans. Cell 153 (6), 1219-1227. doi:10.1016/j.cell.2013.05.002
Spencer, P. S., and Lein, P. J. (2014). “Neurotoxicity,” in Encyclopedia of Toxicology (January, Third Edition), 489-500. doi:10.1016/B978-0-12-386454-3.00169-X
Spînu, N., Cronin, M. T. D., Lao, J., Bal-Price, A., Campia, I., Enoch, S. J., et al. (2021). Probabilistic modelling of developmental neurotoxicity based on a simplified adverse outcome pathway network. Comput. Toxicol., 100206. doi:10.1016/j.comtox.2021. 100206
Spreng, A. S., Brüll, M., Leisner, H., Suciu, I., and Leist, M. (2022). Distinct and dynamic transcriptome adaptations of IPSC-generated astrocytes after cytokine stimulation. Cells 11 (17), 2644. doi:10.3390/CELLS11172644
Strähle, U., Scholz, S., Geisler, R., Greiner, P., Hollert, H., Rastegar, S., et al. (2012). Zebrafish embryos as an alternative to animal experiments-a commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 33 (2), 128-132. doi:10.1016/J.REPROTOX.2011.06.121
Stucki, A. O., Barton-Maclaren, T. S., Bhuller, Y., Henriquez, J. E., Henry, T. R., Hirn, C., et al. (2022). Use of new approach methodologies (NAMs) to meet regulatory requirements for the assessment of industrial chemicals and pesticides for effects on human health. Front. Toxicol. 4, 964553. doi:10.3389/FTOX.2022.964553
Suciu, I., Pamies, D., Peruzzo, R., Wirtz, P. H., Smirnova, L., Pallocca, G., et al. (2023). G E interactions as a basis for toxicological uncertainty. Archives Toxicol. 97 (7), 2035-2049. doi:10.1007/S00204-023-03500-9
Tanner, C. M., Goldman, S. M., Webster Ross, G., and Grate, S. J. (2014). The disease intersection of susceptibility and exposure: chemical exposures and neurodegenerative disease risk. Alzheimer’s Dementia 10 (3 Suppl. L), S213-S225. doi:10.1016/J.JALZ.2014.04.014
Terron, A., and Bennekou, S. H. (2018). Towards a regulatory use of alternative developmental neurotoxicity testing (DNT). Toxicol. Appl. Pharmacol. 354 (December 2017), 19-23. doi:10.1016/j.taap.2018.02.002
Tian, G., Harrison, P. J., Sreenivasan, A. P., Carreras-Puigvert, J., and Spjuth, O. (2023). Combining molecular and cell painting image data for mechanism of action prediction. Artif. Intell. Life Sci. 3 (December), 100060. doi:10.1016/J.AILSCI.2023. 100060
Tsuji, R., and Crofton, K. M. (2012). Developmental neurotoxicity guideline study: issues with methodology, evaluation and regulation. Congenit. Anomalies 52 (3), 122-128. doi:10.1111/J.1741-4520.2012.00374.X
Vinken, M., Benfenati, E., Busquet, F., Castell, J., Clevert, D.-A., de Kok, T. M., et al. (2021). Safer chemicals using less animals: kick-off of the European ONTOX project. Toxicology 458 (June), 152846. doi:10.1016/J.TOX.2021.152846
Wang, Z., Walker, G. W., Muir, D. C. G., and Nagatani-Yoshida, K. (2020). Toward a global understanding of chemical pollution: a first comprehensive analysis of national and regional chemical inventories. Environ. Sci. Technol. 54 (5), 2575-2584. doi:10. 1021/acs.est.9b06379
New approach methods to assess developmental and adult neurotoxicity for regulatory use: a PARC work package 5 project
Tamara Tal, Oddvar Myhre, Ellen Fritsche, Joëlle Rüegg, Kai Craenen, Kiara Aiello-Holden, Caroline Agrillo, Patrick J. Babin, Beate I Escher, Hubert Dirven, et al.
– To cite this version:
Tamara Tal, Oddvar Myhre, Ellen Fritsche, Joëlle Rüegg, Kai Craenen, et al.. New approach methods to assess developmental and adult neurotoxicity for regulatory use: a PARC work package 5 project. Frontiers in Toxicology, 2024, 6, pp.1359507. 10.3389/ftox.2024.1359507 . hal-04679936
HAL is a multi-disciplinary open access archive for the deposit and dissemination of scientific research documents, whether they are published or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
OPEN ACCESS
EDITED BY
Lena Smirnova,
Johns Hopkins University, United States
REVIEWED BY
Sandra McBride,
Social and Scientific Systems, United States
RECEIVED 21 December 2023
ACCEPTED 18 March 2024
published 26 April 2024
CITATION
Tal T, Myhre O, Fritsche E, Rüegg J, Craenen K, Aiello-Holden K, Agrillo C, Babin PJ, Escher BI, Dirven H, Hellsten K, Dolva K, Hessel E, Heusinkveld HJ, Hadzhiev Y, Hurem S, Jagiello K, Judzinska B, Klüver N, Knoll-Gellida A, Kühne BA, Leist M, Lislien M, Lyche JL, Müller F, Colbourne JK, Neuhaus W, Pallocca G, Seeger B, Scharkin I, Scholz S, Spjuth O, Torres-Ruiz M and Bartmann K (2024), New approach methods to assess developmental and adult neurotoxicity for regulatory use: a PARC work package 5 project. Front. Toxicol. 6:1359507.
doi: 10.3389/ftox.2024.1359507
New approach methods to assess developmental and adult neurotoxicity for regulatory use: a PARC work package 5 project
Tamara Tal , Oddvar Myhre , Ellen Fritsche , Joëlle Rüegg , Kai Craenen , Kiara Aiello-Holden , Caroline Agrillo , Patrick J. Babin , Beate I. Escher , Hubert Dirven , Kati Hellsten , Kristine Dolva , Ellen Hessel , Harm J. Heusinkveld , Yavor Hadzhiev , Selma Hurem , Karolina Jagiello , Beata Judzinska , Nils Klüver , Anja Knoll-Gellida , Britta A. Kühne , Marcel Leist , Malene Lislien , Jan L. Lyche , Ferenc Müller , John K. Colbourne , Winfried Neuhaus , Giorgia Pallocca , Bettina Seeger , Ilka Scharkin , Stefan Scholz , Ola Spjuth , Monica Torres-Ruiz and Kristina Bartmann Helmholtz Centre for Environmental Research – UFZ, Chemicals in the Environment Research Section, Leipzig, Germany, University of Leipzig, Medical Faculty, Leipzig, Germany, Norwegian Institute of Public Health – NIPH, Department of Chemical Toxicology, Oslo, Norway, IUF – Leibniz Research Institute for Environmental Medicine, Düsseldorf, Germany, DNTOX GmbH, Düsseldorf, Germany, Swiss Centre for Applied Human Toxicology, University of Basel, Basel, Switzerland, Uppsala University, Department of Organismal Biology, Uppsala, Sweden, European Chemicals Agency (ECHA), Helsinki, Finland, German Federal Institute for Risk Assessment (BfR), Berlin, Germany, Université de Bordeaux, Institut National de la Santé et de la Recherche Médicale (INSERM), Maladies Rares: Génétique et Métabolisme (MRGM), Pessac, France, University of Oslo, Section for Pharmacology and Pharmaceutical Biosciences, Department of Pharmacy, Olso, Norway, Dutch Nation Institute for Public Health and the Environment (RIVM), Centre for Health Protection, Bilthoven, Netherlands, University of Birmingham, Centre for Environmental Research and Justice, Birmingham, UK, Norwegian University of Life Sciences (NMBU), Faculty of Veterinary Medicine, Ås, Norway, University of Gdansk, Laboratory of Environmental Chemoinformatics, Gdansk, Poland, University of Veterinary Medicine Hannover, Foundation, Institute for Food Quality and Food Safety, Hannover, Germany, University of Konstanz, In Vitro Toxicology and Biomedicine/CAAT-Europe, Konstanz, Germany, AIT Austrian Institute of Technology GmbH, Competence Unit Molecular Diagnostics, Center Health and Bioresources, Vienna, Austria, Danube Private University, Faculty of Dentistry and Medicine, Department of Medicine, Krems, Austria, Uppsala University and Science for Life Laboratory, Department of Pharmaceutical Biosciences, Uppsala, Sweden, Instituto de Salud Carlos III (ISCIII), Centro Nacional de Sanidad Ambiental (CNSA), Environmental Toxicology Unit, Majadahonda, Spain
In the European regulatory context, rodent in vivo studies are the predominant source of neurotoxicity information. Although they form a cornerstone of neurotoxicological assessments, they are costly and the topic of ethical debate. While the public expects chemicals and products to be safe for the developing and mature nervous systems, considerable numbers of chemicals in commerce have not, or only to a limited extent, been assessed for their potential to cause neurotoxicity. As such, there is a societal push toward the replacement of animal models with in vitro or alternative methods. New approach methods (NAMs) can contribute to the regulatory knowledge base, increase chemical safety, and modernize chemical hazard and risk assessment. Provided they reach an acceptable level of regulatory relevance and reliability, NAMs may be
Abstract
considered as replacements for specific in vivo studies. The European Partnership for the Assessment of Risks from Chemicals (PARC) addresses challenges to the development and implementation of NAMs in chemical risk assessment. In collaboration with regulatory agencies, Project 5.2.1e (Neurotoxicity) aims to develop and evaluate NAMs for developmental neurotoxicity (DNT) and adult neurotoxicity (ANT) and to understand the applicability domain of specific NAMs for the detection of endocrine disruption and epigenetic perturbation. To speed up assay time and reduce costs, we identify early indicators of later-onset effects. Ultimately, we will assemble second-generation developmental neurotoxicity and first-generation adult neurotoxicity test batteries, both of which aim to provide regulatory hazard and risk assessors and industry stakeholders with robust, speedy, lower-cost, and informative next-generation hazard and risk assessment tools.
1 The European Partnership for the Assessment of Risks from chemicals (PARC)
The European Partnership for the Assessment of Risks from Chemicals (PARC) aims to develop next-generation chemical hazard and risk assessment tools to better protect human health and the environment (Marx-Stoelting et al., 2023). A major ambition of the project is to develop new approach methods (NAMs) for human health hazard assessment that covers developmental neurotoxicity (DNT), adult neurotoxicity (ANT), thyroid hormone disruption, immunotoxicity, and non-genotoxic carcinogens. Work package 5.2.1e aims to refine existing NAMs, develop new ones, and generate first-generation ANT and secondgeneration DNT test batteries. The NAMs that will be developed will be based on Key Events (KE) as identified in the Adverse Outcome Pathway (AOP) framework (Ankley et al., 2010; Leist et al., 2017; Spînu et al., 2021). The work is carried out by a consortium of over 25 experts from 10 EU research institutions and two partner institutions in non-EU countries.
2 Exposure to chemicals may pose a risk to the developing and mature nervous systems
Exposure to chemicals can adversely impact nervous system development and function across all stages of life (Costa et al., 2008; Giordano and Costa, 2012; P; Grandjean and Landrigan, 2006). Adverse chemical-dependent effects stemming from exposure of the developing offspring (including in utero and postnatal) to the time of sexual maturation may affect the developing nervous system (Costa et al., 2004). Such “developmental neurotoxicity” (DNT) can be long-lasting, extending far beyond the exposure period, and can vary across lifespan (Eriksson et al., 1998; Spalding et al., 2013). Note that any type of neurotoxic effect during development is of regulatory concern and relevant for developmental hazard identification. In contrast, when the mature nervous system is exposed to neurotoxic chemicals, adult neurotoxicity (ANT) effects can be immediate or they may be gradually developing and long-lasting. Depending on the type of
ANT effect, it may also be reversible (Spencer and Lein, 2014). Significant and/or severe neurotoxicity, being reversible or irreversible, immediate or delayed, is of regulatory concern.
Due to the sensitivity of the developing nervous system, exposure to low concentrations of certain chemicals may lead to structural and functional disruptions (Rice and Barone, 2000; Grandjean and Landrigan, 2014; Bennett et al., 2016). Neurodevelopmental disorders including autism spectrum disorder, intellectual disability, attention deficit/hyperactivity disorder (ADHD), neurodevelopmental motor disorders (including tic disorders), and specific learning disorders can have lifelong socioeconomic consequences, including diminished economic productivity or an increased need for learning support in schools (P. Grandjean and Landrigan, 2006). Whilst estimates were acknowledged to be uncertain, in the disability adjusted life years (DALYs) related to neurodevelopmental disease may be the result of chemical exposure (and irrespective of a person’s genetic predisposition/sensitivity), with DALYs when chemical exposure was combined with underlying genetic predisposition (EC 2019). This estimate was based on a ‘top down’ assessment of impacts of pervasive neurodevelopmental disorders from the World Health Organization (WHO) and an estimate that is due to environmental exposure to legacy compounds such as lead and other environmental pollutants (EC 2019). Notable socioeconomical benefits are therefore predicted via the identification of substances which are known or presumed to cause DNT and subsequent prevention of exposure (Bellanger et al., 2013; Bellanger et al., 2015).
After the developmental period, acute and/or chronic exposure to environmental chemicals may elicit toxic responses in the peripheral and/or central nervous systems and it has been suggested that exposure to specific chemical agents may increase the probability of developing neurodegenerative disorders such as Parkinson’s and Alzheimer’s Diseases, or dementia (Landrigan et al., 2005; Tanner et al., 2014; Ockleford et al.). Moreover, exposure to certain chemicals has been suspected to be linked to adolescent and adult depression, anxiety, and other psychiatric disorders in a number of academic publications (Dickerson et al., 2020; Hollander et al., 2020; Jacobson et al., 2022; Rokoff et al., 2022; Aung et al., 2023). In a study of 22 chemical inventories from 19 countries and regions, over 350,000 chemicals and mixtures of
chemicals were identified as registered for production and potentially in use (Wang et al., 2020). Despite knowledge concerning the potentially harmful impacts of environmental chemicals on the developing and mature nervous systems (P. Grandjean and Landrigan, 2006), it is understood that only a limited number of unique substances has been tested for DNT using OECD Test Guideline (TG) studies. (OECD, 2008a; Makris et al., 2009; Sachana et al., 2019; Crofton and Mundy, 2021).
3 Policy and regulatory landscapes
The EU Green Deal describes health impacts in the ZeroPollution Action Plan, and the European Commission recently highlighted their interest in increased efforts to protect against the most harmful chemicals, by further exploring the risk management possibilities of neurotoxic and endocrine disrupting (which has been linked to DNT) substances (European Commission, 2020). In the EU, several relevant regulations are in force. For example, before entering the market or gaining approval as a biocidal or pesticidal active substance, the minimum data requirements described in the relevant EU Regulation must be fulfilled (among other conditions). EU regulations on plant protection products (Reg EC 1107; European Parliament and Council, 2009) and biocides (Reg EC 528; European Parliament and Council, 2012) can require DNT/ANT testing as part of the data requirements. Under the EU Biocides Product Regulation (Reg EC 528; European Parliament and Council, 2012), specific DNT testing, for example, OECD TG 426, recently became a mandatory information requirement for the approval process of active biocidal substances. Under REACH (European Parliament and Council, 2006), the European Regulation created to protect human health and the environment from harmful chemicals, the level of information required to identify potential neurotoxic (DNT/ ANT) properties currently depends on the tonnage and identification of specific concerns that may trigger DNT or ANT tests. The available information is used to apply appropriate hazard classifications, as per the criteria specified in the CLP regulation (Reg EC 1272; European Parliament and Council, 2008), to inform on the hazardous properties of chemicals. Classification in accordance with CLP then serves to trigger or inform remedial actions in other legislation to control the hazard. The CLP regulation (Articles seven and 8) does not require DNT or ANT TGs directly but rather makes use of all available data generated in the context of relevant legislation and/or otherwise available in the public domain. In cases where such data is not available to inform on a given hazard, testing may be conducted under certain conditions including the condition that tests on animals are to be carried out only where no other alternatives, which provide adequate reliability and quality of data, are possible. This implies support and presents an opportunity for the development, validation, and implementation of NAMs.
Within the CLP Regulation, substances with DNT are addressed under the reproductive (developmental) toxicity hazard class and ANT effects are addressed under Specific Target Organ Toxicity (STOT), either single exposure (SE) or repeated exposure (RE), depending on whether the effects are caused by single or repeated exposures, respectively. The recent revision of the CLP regulation
includes a new hazard class for endocrine disruption that includes endocrine activity mediated adverse effects on the developing (and mature) nervous system (Reg EC 1272; (European Parliament and Council, 2023). According to the new criteria, classification as ED Category one shall be largely based on evidence from at least one of the following: human data; animal data; non-animal data providing an equivalent predictive capacity as human data or animal data (Reg EC 1272; European Parliament and Council, 2023). Thus, the new hazard class allows for NAMs to be directly used for the purpose of this specific classification when the criteria are met.
3.1 DNT/ANT in current chemical regulations
The information needed to fulfill the data requirements under REACH and BPR is typically provided by the in vivo OECD TG studies defined in the relevant section of the applicable regulation, but there are also specific possibilities for adaptation (more specifically data waiving). Such adaptation possibilities can include non-animal approaches and/or the use of existing information stemming from similar substances via a read-across approach (European Parliament and Council, 2006). However, where data on human health and environmental properties are derived via adaptations to data requirements, certain conditions apply. The conditions for adaptations using in vitro data under REACH are specified in Annex XI, section 1.4. In the context of a read across adaptation (REACH Annex XI, section 1.5), again certain restrictive conditions apply with regard to the data that directly informs on the hazard. However, for the extrapolation of such data to other substances, there is a clear opportunity to utilize NAMs as supportive information to demonstrate similarity in the properties of the substances concerned.
It should also to be noted that, depending on the applicable regulation, new tests may not be necessary if the available data is already sufficient for the regulatory purpose as given in the specific regulation. For example, the DNT study shall not be conducted under the BPR if the available data already indicate that the substance causes developmental toxicity and meets the criteria to be classified as toxic for reproduction category 1A or 1B: May damage the unborn child (H360D), and these available data are adequate to support a robust risk assessment (European Parliament and Council, 2012).
A range of OECD TG studies, including single-dose studies (e.g., OECD TG 402, 403, 420, 423, 425) and/or repeated dose toxicity studies (e.g., OECD TG 407, 408, 421, 422, 414, 443 in the absence of DNT cohorts) may inform on ANT or DNT based on clinical signs such as paralysis, convulsions, lack of coordination, or ataxia or neurohistopathology and/or alterations in brain weight (Table 1). DNT can be evaluated more comprehensively using dedicated tests such as OECD TG 426 or in the DNT cohort (cohorts 2A and 2B) of the Extended One-Generation Reproduction Toxicity Study (EOGRTS, OECD TG 443). OECD TGs dedicated to studying ANT include OECD TG 424, 418 and 419. Under REACH, ANT or specific mechanisms/modes of action with an association to (developmental) neurotoxicity can be used to trigger specific DNT studies. Substances in food intended for infants can also
TABLE 1 Description of existing OECD guideline studies that include neurotoxicity as an endpoint.
Test guideline
Primary endpoint
Neurotox endpoint
Preferred Species
Administration period
Non-behavioral endpoints related to neurotoxicity
Behavioral endpoints
Reference
OECD TG 402
Dermal Toxicity
ANT (acute)
Rat
Adults ( )
No (just gross necropsy)
Autonomic and central nervous system and somatomotor activity and behavior pattern
OECD (2017)
OECD TG 403
Inhalation Toxicity
ANT (acute)
Rat
Adults (4 h)
No (just gross necropsy)
Autonomic and central nervous system and somatomotor activity and behavior pattern
OECD (2009)
OECD TG 407
Oral Toxicity/ Endocrine Disruption
ANT (chronic)
Rat
Adults (daily – 28d)
Brain weight, histopathology of brain, spinal cord, and sciatic nerve
Sensory reactivity to stimuli, limb grip strength, motor activity
OECD (2008b)
OECD TG 408
Oral Toxicity/ Endocrine Disruption
ANT (chronic)
Rat
Adults (daily – 90d)
Brain weight, histopathology of brain, spinal cord, and sciatic nerve
Sensory reactivity to stimuli, limb grip strength, motor activity, autonomic activity
OECD (2018a)
OECD TG 418
Neurotoxicity (OP substances)
ANT (acute)
Hen
Adults (single dose)
Neuropathology of central and peripheral nervous system, NTE and AchE activities
Behavioral abnormalities, ataxia, and paralysis
OECD (1995a)
OECD TG 419
Neurotoxicity (OP substances)
ANT (chronic)
Hen
Adults ( days)
Neuropathology of central and peripheral nervous system, NTE and AchE activities
Behavioral abnormalities, ataxia, and paralysis
OECD (1995b)
OECD TG 420
Oral Toxicity
ANT (acute)
Rat
Adults (single dose)
No (just gross necropsy)
Somatomotor activity and behavior patterns
OECD (2002a)
OECD TG 423
Oral Toxicity
ANT (acute)
Rat
Adults (single dose)
No (just gross necropsy)
Somatomotor activity and behavior patterns
OECD (2002b)
OECD TG 424
Neurotoxicity
ANT (chronic)
Rat
Adults ( days)
Neuropathology of central and peripheral nervous system
Sensory reactivity to stimuli, limb grip strength, motor activity
As the development of the nervous system starts prenatally and continues to develop through adolescence, reaching adult levels of neurotransmitters, synaptic plasticity, myelination and grey matter at around age of 20 in humans and around PND60 in rats (Semple et al., 2013), it is key to implement exposure throughout the whole developmental period for improving the chances to identify
developmental neurotoxicants. In an OECD TG 426, the offspring are exposed as a minimum from the time of implantation (starting on gestation day (GD) 6) throughout lactation (until postnatal day (PND) 21). In cohort 2 A of an EOGRTS, the offspring are exposed via the mother in utero, through lactation and directly at least after weaning until termination on ~ PND 66-77. The assessed DNT parameters in specific DNT studies include as a minimum (depending on the OECD TG) motor activity, motor and sensory function, associative learning
and memory (only in OECD TG 426 as standard testing), brain weight, and central and peripheral nervous system histopathology (Tsuji and Crofton, 2012). As dedicated DNT studies are often complex studies using rodents, they are resource-intensive regarding time, costs, and number of animals (Crofton et al., 2012; Smirnova et al., 2014) and only a limited number of chemicals have been tested for DNT using OECD TG DNT studies (OECD, 2008a; Makris et al., 2009; Sachana et al., 2019; Crofton and Mundy, 2021). In addition, variability in rodent neurotoxicity tests has been documented (Tsuji and Crofton, 2012; Terron and Bennekou, 2018; Sachana et al., 2019; Paparella et al., 2020; Harry et al., 2022) which indicates a need to specify, optimize, and harmonize the individual test methods used as part of the OECD TG. It also underscores the need for new tests that lack the excess variability associated with in vivo guideline studies for the assessment of DNT.
The risk posed by unidentified (developmental) neurotoxic agents and the limited number of timely and cost-efficient test systems (i.e., NAMs) serve as the basis for this PARC project where the ambition is the generation of an improved in vitro and alternative DNT test battery and a first-generation in vitro ANT test battery. The need to develop NAM-based next-generation hazard and risk assessment for DNT and ANT has found international support from academic scientists, industry, certain regulatory authorities, and other interest groups (Smirnova et al., 2014; Ockleford et al., 2017; Fritsche et al., 2018; Kavlock et al., 2018; Craig et al., 2019; Paparella et al., 2020; Vinken et al., 2021; Pallocca et al., 2022; Stucki et al., 2022). ECHA recently published Key Areas of Regulatory Challenge (ECHA, 2023), which highlights several of the known scientific and regulatory challenges that NAMs face. It further underlines the need for additional research in the field of ANT and DNT NAMs (ECHA, 2023).
NAMs have been advocated to be implemented into the regulatory hazard assessment stage of chemicals risk assessment (Stucki et al., 2022; Schmeisser et al., 2023). Currently, in vitro data may be used in Weight of Evidence assessment for classification and labelling (e.g., for developmental toxicity), or to trigger further DNT tests at REACH Annex IX and X, or to support grouping and read across from similar substances. High-throughput in vitro assays have also great potential as screening tools to prioritize chemicals and specific modes of action (MoA) for further testing (Escher et al., 2023). While such high throughput screening (HTS) tools have not yet been implemented for DNT and ANT in large scale regulatory practice, the introduction of more sophisticated in vitro tests and the validation of all HTS assays for DNT and ANT appear vital to improve their regulatory usefulness. It has been suggested that new in vitro methods should be mechanistically associated with adverse (developmental) neurotoxic outcomes (Pitzer, Shafer, and Herr, 2023). This is important to establish the toxicological relevance of endpoints measured in NAMs and/or to allow for the selection of the most informative follow-up studies to produce new information to elicit regulatory action (Smirnova et al., 2014; Ockleford et al., 2017; Fritsche et al., 2018; Kavlock et al., 2018; Craig et al., 2019; Paparella et al., 2020; Vinken et al., 2021; Pallocca et al., 2022; Stucki et al., 2022).
4 Building the DNT-IVB v2.0
One of the purposes of this PARC project is to deliver a guidance document containing a framework to facilitate the regulatory use of
FIGURE 1
The DNT-IVB. Version 1.0 of the DNT-IVB (Masjosthusmann et al., 2020; Blum et al., 2022; OECD, 2023) contains endpoints for human neural progenitor cell (hNPC) proliferation and apoptosis, cell (neuronal, radial glia, oligodendrocytes) migration, hNPCneuronal and oligodendrocyte differentiation, neurite outgrowth, neuronal maturation and synaptogenesis, and neuronal network formation (top). While most assays are conducted in human cells (blue boxes), two are performed in rat primary cells (yellow boxes). PARC 5.2.1e builds and evaluates NAMs for key battery gaps including myelination, blood-brain barrier (BBB) formation, mitochondrial function, and a suite of automated behavior-based NAMs in early lifestage zebrafish (green boxes). In addition, replacement of rodentbased NAMs with human-based test systems is also underway. Lastly, early epigenetic or sub-cellular morphological indicators of later DNT-related effects are being generated and evaluated for potential use in the DNT-IVB v2.0. Adapted from Crofton and Mundy, 2021.
data derived from a DNT in vitro NAM-based test battery. A basic DNT in vitro test battery (IVB) has already been developed (i.e., DNT-IVB v1.0). It covers several cellular neurodevelopmental processes vital for normal brain development (Bal-Price et al., 2018; Masjosthusmann et al., 2020; Crofton and Mundy, 2021; Blum et al., 2022; Koch et al., 2022; OECD, 2023). The DNT-IVB v1.0 (Figure 1) measures effects of chemicals on human neural progenitor cell (hNPC) proliferation (Baumann et al., 2014; Baumann et al., 2015; Harrill et al., 2018; Nimtz et al., 2019; Masjosthusmann et al., 2020; Koch et al., 2022) and apoptosis (Druwe et al., 2015; Harrill et al., 2018), cell migration (Baumann et al., 2015; Baumann et al., 2016; Nyffeler et al., 2017; Schmuck et al., 2017; Masjosthusmann et al., 2020; Koch et al., 2022), hNPC-neuronal differentiation (Baumann et al., 2015; Schmuck et al., 2017; Masjosthusmann et al., 2020; Koch et al., 2022), oligodendrocyte differentiation (Dach et al., 2017; Schmuck et al., 2017; Masjosthusmann et al., 2020; Klose et al., 2021; Koch et al., 2022), neurite outgrowth (human: Harrill et al., 2010; Harrill et al., 2018; Krug et al., 2013; Hoelting et al., 2016; Masjosthusmann et al., 2020; Koch et al., 2022; rat; Harrill et al., 2013; Harrill et al.,
2018), and synaptogenesis and neuronal network formation (rat: Harrill et al., 2011; Harrill et al., 2018; Brown et al., 2016; Frank et al., 2017; Shafer, 2019).
Gap analysis of the DNT-IVB v1.0 revealed a requisite for supplementary cell assays (e.g., microglia) and functions (e.g., human neuronal network formation, astrocyte function, behavior, learning, and memory) to augment coverage and increase the ability to detect potential (developmental) neurotoxicants (Crofton and Mundy, 2021). Coverage of additional targets for neurotoxicants (e.g., signaling pathways and processes) is necessary, as exemplified by nicotine, a compound identified as a false negative in the DNTIVB v1.0 (Masjosthusmann et al., 2020; Crofton and Mundy, 2021; Blum et al., 2022). This indicates the inability to detect (developmental) neurotoxic compounds that target nicotinic receptors in these test systems (e.g., neonicotinoid insecticides) (Sheets et al., 2016; Loser et al., 2021; Blum et al., 2022). To address some of the identified gaps, there are four key areas that this PARC project aims to improve during the development of the DNT-IVB v2.0 (Figure 1). This includes refinement of existing assays, generation of new NAMs to cover essential gaps, determination of the applicability domain for relevant available NAMs, and increased cost efficiency.
4.1 Refine existing assays
The current synaptogenesis and neural network formation assays were based in primary rat cortical cells differentiated in 2D on multielectrode arrays (MEA; Brown et al., 2016; Frank et al., 2017) (Figure 1). While there is also a recently established human neural network formation (hNNF) assay (Bartmann et al., 2023), it requires commercially available human-induced pluripotent stem cells (hiPSCs), which are used to derive excitatory and inhibitory neurons and primary human astrocytes that can be plated on MEAs for a functional assessment of network formation. This assay has recently been used to evaluate the effects of pesticides on human neural network formation (Bartmann et al., 2023). To decrease the costs of the hNNF assay, the PARC consortium will re-establish and refine the protocol using noncommercially generated hiPSC-derived excitatory and inhibitory neurons, together with human astrocytes derived from hNPCs (Koch et al., 2022). Synapse assembly is a critical feature of neurodevelopment. The DNT-IVB v1.0 assay for synaptogenesis is currently based on primary rat cortical cells (Harrill et al., 2011; Harrill et al., 2018). Human iPSC-derived NPCs can be differentiated into different types of postmitotic neurons and astrocytes (Davidsen et al., 2021; Lauvås et al., 2022). Therefore, a test system, comprised of a 2D mixed culture of neurons and astrocytes undergoing differentiation, will be developed and refined using high-content imaging. To enable comparison to data generated in the rat synaptogenesis assay, a chemical test set will be evaluated (described below).
Another identified gap within the DNT-IVB v1.0 is a lack of assays that describe mitochondrial toxicity events in susceptible cell types. AOP3 (“inhibition of the mitochondrial complex I of nigrostriatal neurons leads to parkinsonian motor deficits”) describes a link between inhibition of complex I of the mitochondrial respiratory chain and motor deficits associated with parkinsonian
disorders (https://aopwiki.org/aops/3). The current DNT-IVB v1. 0 assays are not particularly sensitive or fail to detect known mitochondrial toxicants (Masjosthusmann et al., 2020; Crofton and Mundy, 2021). To increase the sensitivity of battery assays to this class of neurotoxicants, several DNT-IVB v1.0 assays will be modified to allow for increased detection of mitochondrial toxicants. This step includes the assessment of neurite area and cell viability in human dopaminergic neurons and human immature peripheral neurons. While these endpoints are covered in the DNT-IVB v1. 0 , where the NAMs are performed in glucose-containing medium, in DNT-IVB v2.0, the assay will be performed in glucose-free, galactose-containing medium, which makes cells more reliant on their mitochondria and increases their sensitivity to mitochondrial toxicants (Hoelting et al., 2016; Delp et al., 2019; Delp et al., 2021).
4.2 Build new NAMs to cover essential gaps
4.2.1 Cellular gaps
In a key analysis, 29 neurotoxicity MoAs were characterized for 248 individual compounds representing 23 compound classes and 212 natural neurotoxins (Masjosthusmann et al., 2018). More comprehensive assessment of the potential for chemicals to harm the developing nervous system likely requires NAMs that cover the identified MoAs. One MoA not covered in the DNT-IVB v1.0 is the formation of a functional blood-brain barrier (BBB). The BBB determines the ability of some environmental chemicals to reach the central nervous system (Banks, 2009). Chemical exposure can affect the BBB integrity to cause DNT effects (Saili et al., 2017). Here, we will develop and use an hiPSC-based BBB NAM to test whether chemical exposure increases permeation of chemicals across the barrier, resulting in higher concentration reaching the central nervous system. According to established differentiation protocols (Appelt-Menzel et al., 2017), chemicals will be applied during cellular differentiation and transendothelial electrical resistance will be used as a readout of barrier function. One MoA considered relevant for DNT and not yet covered by the DNTIVB v1.0 is the contribution of inflammatory reactions of glial cells. The key cell populations producing inflammatory mediators in the brain are astrocytes and microglia (Carson et al., 2006). These cells can be generated from human stem cells (Brüll et al., 2020; Spreng et al., 2022) and then either tested as pure populations, as mixed glial populations or together with various neuronal cultures (Gutbier et al., 2018; Klima et al., 2021).
4.2.2 Functional gaps
OECD TG 426 (OECD, 2007) assesses neurobehavioral endpoints which include measures of cognition (including associative learning and memory) in rodents exposed to chemicals during the developmental period. Cellular NAMs may provide information on cellular events that may eventually cause adverse effects on cognitive functions or other neurobehavioral functions but fail to provide equivalent information to neurobehavioral tests. In addition, when considering the complex integration of intracellular, intercellular, interregional, and systemic interactions that occur in development-stage and regional specific manners, in vitro NAMs do not cover all relevant cell types and processes, inherent within whole organisms, that are necessary to
develop and maintain a functional nervous system. In this project, the alternative (i.e., relative to rodent models) early life zebrafish model will be used to generate a range of behavior-based assays that complement the in vitro approaches described above.
Zebrafish (Danio rerio) are a 3 Rs-compliant (Hooijmans et al., 2010), non-protected vertebrate model up to 5 days post fertilization (dpf) (Strähle et al., 2012; Kalueff et al., 2013). The zebrafish embryo model may represent a powerful translational system for human hazard and risk assessment as zebrafish possess orthologs for of human genes (Howe et al., 2013), of human disease-related genes (Howe et al., 2013), and of general human drug targets (Gunnarsson et al., 2008). Zebrafish are increasingly being utilized as a model system to investigate the function of the growing list of risk genes associated with neurodevelopment disorders (Sakai et al., 2018), including motor neuron diseases (Babin et al., 2014). Zebrafish neurodevelopment starts at 24 h post-fertilization and primary neurogenesis is complete by roughly 72 h postfertilization (depending on rearing temperature). Resulting neuroanatomy (Gupta et al., 2018), nervous system transcriptomic lineages (Raj et al., 2018), and brain asymmetry (Duboc et al., 2015) are suggested to be comparable to humans. In addition, neurotransmitter systems, including glutaminergic, cholinergic, serotonergic, dopaminergic, adrenergic, GABAergic, and histaminergic (Babin et al., 2014; Faria et al., 2015; Horzmann and Freeman, 2016) are similar to those found in humans and associated with sensory-motor outcomes. This rapid establishment of neural structures during neurodevelopment and their link to quantifiable behavioral parameters is a major asset for PARC WP5.2.1e.
Relative to in vitro systems, metabolically competent zebrafish embryos may address potential toxicokinetics that can affect toxicity outcomes (Chu and Sadler, 2009). Regarding neurotoxicity, the assessment of neurobehavioral effects caused by xenobiotic exposure is advantageous because these perturbations are sensitive (i.e., they occur at sub-morphological concentrations) (Noyes et al., 2015; Bruni et al., 2016; Gaballah et al., 2020; Jarema et al., 2022). Locomotor activity can function as an automated and generalized readout of neurodevelopment. A major advantage of the early life zebrafish system as compared to in vitro systems is that it represents an alternative whole organism animal system that is emendable to genome-wide differential expression data collection throughout early neurodevelopment (Kettleborough et al., 2013) and are expected to address comparatively more MIEs and KEs related to DNT in a single assay (e.g., neurotransmitter signaling pathways, functional BBB, myelinated axons, functional synapses, neuronal networks, and neural circuits), as compared to individual in vitro test systems performed in single cell types or limited co-culture systems. Moreover, zebrafish development has been mapped at the resolution of single-cell transcriptomics, allowing the detection of cell-type specific changes associated with chemical induced adversity affecting neural and non-neural components of the developing brain (Farrell et al., 2018).
In compliance with EU directive (2010/63/EU; European Parliament and Council, 2010), the majority of work will be conducted in embryos up to 5 dpf . Another key advantage of early life stage zebrafish NAMs is that they can be screened in DNT and acute modes by varying the length and timing of chemical
exposure. The DNT mode captures structural and functional deficits that alters locomotor activity in response to various stimuli. The acute mode identifies rapid, receptor-mediated changes in neuroactivity that can potentially be used as a complement to cellular ANT assays which aim to identify perturbations in signaling pathways (e.g., dopaminergic signaling) linked to ANT AOPs.
All DNT NAMs performed in early life-stage zebrafish described will be used following developmental exposure to PARC test chemicals and removed prior to behavior testing. This increases the likelihood of detecting functional or structural effects that arise from developmental perturbations in underlying behavior circuits after chemical exposure has ceased. Later, in the development of the ANT-IVB 1.0, the same assays will be applied post-neurogenesis (after three dpf) to detect the acute neuroactivity potential of test chemicals with a focus on the detection of perturbations in receptorbased neurotransmitter systems that are associated with DNT and/ or ANT (e.g., dopaminergic, gabaergic, glutamatergic perturbations).
Another important functional topic is the impact of chemical exposure on associative learning and memory, which is assessed as a standard part of rodent-based OECD TG 426 for DNT (OECD, 2007), and may be included as an add on to TG 424 (OECD, 1997) for ANT and in TG 443 for DNT (OECD, 2018b). In vitro test systems are unable to account for these complex behavioral and cognitive aspects. Members of this consortium are developing a NAM that detects chemical-dependent disruption of nonassociative learning and memory retention in early life stage zebrafish (Figure 2). An escape response NAM that identifies chemicals that specifically disrupt the motor system via the activation of reticulospinal neurons and independently of sensory processing (Dubrana et al., 2021; Knoll-Gellida et al., 2021) will be further developed and applied to screen a common set of chemicals described below. A NAM for chemical-induced developmental motor neuron toxicity will also be developed using the transgenic line Tg (nrp1a:gfp)js12 with labeled motor neurons (Sato-Maeda et al., 2006). A NAM that detects anxiety-like behavior via the detection of thigmotaxis, or the time spent along the outer edge of a well, is also under development. Finally, for a subset of test chemicals, the persistence of behavioral effects, post-exposure, will be evaluated to substantiate the detection of DNT effects in 14 dpf old larvae.
4.2.3 Determine applicability domain
The applicability domain describes the physicochemical or other properties of the chemicals for which a NAM is applicable for use (OECD, 2005). The applicability domain is generally determined using a range of reference chemicals linked to an adverse effect (OECD, 2005). Within this project, a test set of 96 reference chemicals, based on previously published work (Masjosthusmann et al., 2020; Blum et al., 2022), will be evaluated. With the exception of the zebrafish NAMs, all other DNT NAMs will be performed in human cellular assays. As these models contain a limited number of cell types and other gaps, efforts will be made to assess the applicability domain to determine whether, and to what extent, the NAMs cover established DNT AOPs, including endocrine disruption (ED) or proposed DNT AOPs such as epigenetic perturbation. Specifically, in the 2D synaptogenesis model,
FIGURE 2
Functional NAMs performed in early life stage zebrafish. A suite of automated zebrafish-embryo behavior-based NAMs are under development for potential inclusion in the DNT-IVB v2.0. All NAMs are performed using automated tracking. Most assays will be performed in 96 square well plates for a comparable throughput to in vitro assays. Based on the exposure paradigm used, these assays can be performed with protocols that predict DNT and/or ANT. For the detection of DNT, chemical exposure occurs during development and is removed prior to behavior assessments. In contrast, if exposure takes place after neurogenesis is complete, any adverse effect related to the specific signalling pathways that have reached maturation may be indicative of acute ANT. However, as the organism’s nervous system is still developing, it cannot be excluded that the nature of the effect may be considered within a DNT assessment framework.
characterization of differentiation up to 28 days will be performed to understand the abundance and distribution of ED-relevant receptors including retinoic acid receptor, estrogen receptor, androgen receptor, thyroid hormone receptor, glucocorticoid receptor and liver X receptor. Coverage of ED and epigenetic modes of action will also be carried out for certain cellular ANT NAMs (see below).
4.2.4 Increase molecular and cellular coverage, reliability, and cost efficiency
Mammalian TG studies for DNT testing are costly and timeconsuming (Crofton et al., 2012; Smirnova et al., 2014). At the same time, mammals contain a complete nervous system with all functional components throughout the whole developmental period, and their communication with other relevant organs and systems, (e.g., gut, liver, endocrine, and immune systems) that can collectively influence neurotoxicity outcomes. Next-generation DNT and ANT testing seeks to eventually replace mammalian tests with a battery of in vitro and alternative test systems. Some of these next-generation test systems are lengthy and can take up to 35 days, which increases the overall cost of the potential test battery. To complement cellular and alternative assays potentially included in the DNT-IVB v2.0 and to provide a low-cost screening strategy, four approaches will be explored.
The first strategy seeks to identify early markers of later DNTassociated KEs with a focus on epigenomic or sub-cellular morphological alterations. Epigenomic processes drive cell differentiation, and chemically induced alteration in epigenetic patterns can lead to long-term changes in gene function (Baccarelli and Bollati, 2009). In this context, we will assess whether epigenetic re-arrangements precede morphological changes observed in differentiation-related assays, with the potential to shorten and/or strengthen such assays. This step will be addressed by performing and comparing different genome-wide epigenetic (i.e., DNA methylation) analyses at specific time points in cellular differentiation assays where shortening could be of interest. Cell painting is a high-throughput microscopy technique that allows researchers to simultaneously label and visualize multiple organelles
in a cell (Bray et al., 2016). It uses a cocktail of fluorescent stains and high-content imaging to obtain parallel morphological measurements on a single cell level. This project will establish and evaluate an automated cell profiling methodology for DNT assays that is cheap, fully automated, data-rich, and can operate on 2D cell monolayers, and work is ongoing to expand to 3D spheroids. Images from Cell Painting are data-rich and highly applicable for analysis with artificial intelligence methods, for example, for MoA prediction (Tian et al., 2023), and assessment of combination effects of environmental compounds (Rietdijk et al., 2022). Morphological changes will be assessed as potential early indicators of DNT or ANT outcomes in short- and longer-term cellular assays.
The second strategy employs transcriptomics to refine the search for early markers of later DNT-associated KEs with a focus on genome-wide expression patterns observed at a single cell resolution of intact developing embryo nervous systems. This single-cell transcriptome data is complemented by building transcriptomics and metabolomics technologies into an existing OECD TG for embryotoxicity (TG 236) to measure (neuro)developmental toxicity endpoints. While it was developed as one of the potential alternatives to the acute test on fish (OECD TG 203) for ecotoxicological hazard assessment, it has been reported that compared to OECD TG 203, OECD TG 236 may underestimate acute toxicity for certain types of chemicals, in particular neurotoxicants (Klüver et al., 2016; Glaberman et al., 2017; Sobanska et al., 2018). Nevertheless, OECD TG 236 has recently attracted considerable attention for its potential expansion to human health endpoints, particularly developmental (neuro) toxicity (Braunbeck et al., 2014; Krzykwa et al., 2019).
The third strategy uses a rapid, low-cost cellular neurite outgrowth assay to screen a much larger chemical test set, including humanrelevant mixtures (J. Lee et al., 2022a; J; Lee et al., 2022b). For this assay, SH-SY5Y cells, sub-cloned from a neuroblastoma cell line, are used in 384 well plate format (J. Lee et al., 2022b; J; Lee et al., 2022b). The specificity and sensitivity of this assay will be compared to nontransformed, hiPSC-derived DNT models.
The fourth strategy will build in silico models to predict the probability of inducing DNT effects using Quantitative Structure-
Activity Relationships (QSAR) models to link chemical structural properties with measured neurotoxicity effects (Khelfaoui et al., 2021; Grillberger et al., 2023). Molecular docking combined with observed molecular dynamics will be employed to reflect interactions of organophosphates with cellular targets (e.g., membranes, proteins) identified as MIEs according to DNTrelevant AOPs (Gadaleta et al., 2022). Serine esterases and calcium transporters are currently under consideration (KnollGellida et al., 2021). Method evaluation will be performed by comparing simulations with experimentally generated data. Binding affinity for the set of targets will then be predicted by using machine learning-based models, and structural alerts for pathway perturbation will be identified.
5 Demonstration of added value and identification of a minimum assay battery for DNT-IVB v2.0
A fundamental requirement for the regulatory acceptance of NAMs involves the development of test methods characterized by a high degree of robustness, performance, and readiness (Bal-Price et al., 2018), including acceptable levels of variability (Harry et al., 2022). If they are intended for future use in the context of hazard assessment, NAMs should ideally meet or exceed the sensitivity, specificity, accuracy, and reliability of the respective OECD TG, to ensure a continued level of acceptable chemical safety (Bal-Price et al., 2018). This approach ensures the use of data with a significant level of confidence. In the case of the DNT-IVB v1.0, elevated standards of readiness and robustness have already been demonstrated (Bal-Price et al., 2018; Krebs et al., 2019; Blum et al., 2022). To assess DNT-IVBv1.0 performance, a set of 45 reference (i.e., performance) compounds, consisting of 28 substances that were considered DNT positive and 17 substances that were considered DNT negative by the assay developers were used. Using these substances, an assay sensitivity of , specificity of , and accuracy of was observed (Blum et al., 2022).
To substantiate the added value of new and refined assays within the DNT-IVB v2.0, a 96 -member reference set will be used, which encompasses 45 performance compounds from the DNT-IVB v1.0 (Blum et al., 2022) and will be augmented with known modifiers targeting pathways specific to DNT (Fritsche, 2017) (e.g., mTOR (Lee, 2015), PDGFR-PLC (Kang et al., 2016), Notch (Imayoshi et al., 2013), and thyroid hormones (TH; López-Espíndola et al., 2014; Bernal et al., 2015)). Centralized chemical procurement and distribution will occur via a collaborative effort involving PARC 5.2.1e scientists and experts from the EU Joint Research Centre (EUJRC). This structured approach facilitates standardized and comparable assessment of substances across partner laboratories, effectively minimizing uncertainties associated with substance purity, solubilization, and concentration. By testing the DNT-IVB v1.0 performance compounds at reasonable concentrations in each newly developed test method, we will gain insight into the chemical applicability domain of each assay to understand the potential added value of DNT-IVB v2.0 NAMs. If DNT-IVB v2.0 NAMs can appropriately identify DNT-IVB v1.0 false negatives as positives, this will increase the sensitivity of the resulting v2.0 test battery. In
addition, the evaluation of specific pathway agonists and antagonists will reveal the applicability domain of each assay and the whole DNT-IVB v2.0.
6 Build ANT-IVB v1.0
In contrast to DNT, there is no comparable set of NAMs with a high readiness for ANT testing that covers important MoA. ANT can be elicited through a variety of mechanisms involving neurotransmitter receptors and ion transporters which influence the transmission and processing of signals in the human brain and other parts of the nervous system (Fritsche and Hogberg, 2020; Masjosthusmann et al., 2018). Recently, a neurotoxicity MoA analysis was performed for 248 individual compounds, representing 23 compound classes and 212 natural neurotoxins (Masjosthusmann et al., 2018). The identified MoA were grouped according to ANT common key events including cholinergic, GABAergic, glycinergic, glutamatergic, adrenergic, serotonergic, and dopaminergic neurotransmission, ion channels/receptors (e.g., sodium channels, potassium channels, calcium channels, chloride channels), and a range of cellular endpoints such as mitochondrial dysfunction, oxidative stress, apoptosis, redox cycling, altered calcium signaling, cytoskeletal alterations, neuroinflammation, axonopathies, myelin toxicity, delayed neuropathy, and enzyme inhibition (Masjosthusmann et al., 2018). To enable a thorough assessment of chemicals’ ANT potential, it is necessary to compile the ANT-IVB v1.0 that includes the entirety of the identified MoA , a challenge that will be addressed in our project.
To date, several assays have been established and published that cover critical KEs for neurotoxicity (Schmidt et al., 2017) but they still need refinement to meet certain criteria for regulatory acceptance of alternative methods including assessment of the domain of applicability, assay robustness and relevance, and demonstrated predictivity for adult neurotoxicity (Bal-Price et al., 2018). For the assessment of direct activation of ion channels and receptors and altered function of channels and receptors of (nociceptive) sensory neurons, several NAMs used in the DNTIVB v2.0 can be repurposed for acute testing in mature 2D or 3D culture systems or in early life-stage zebrafish post neurogenesis. Specifically, DNT-IVB v2.0 test methods, including the myelin and BBB NAMs, cell painting in human BrainSpheres (Hartmann et al., 2023), and zebrafish learning and memory, motor system toxicity, and anxiety-like NAMs will be performed in mature cellular cultures or zebrafish embryos at time points which occur after primary neurogenesis and differentiation has occurred (R. Schmidt et al., 2013).
In addition to repurposed DNT-IVB v2.0 NAMs, several novel NAMs are being developed and applied to the ANT-IVB v1.0. One is a recently developed NAM based on hiPSC-derived nociceptorenriched, mature sensory neurons (Holzer et al., 2022). Using this NAM after 23 days of differentiation, acute exposure to ANT reference chemicals will be carried out to evaluate the biological applicability domain of the assay. Another is the human multineurotransmitter assay (hMNR), which is based on hiPSC-derived mixed neuron-glia 3D BrainSpheres. The hMNR NAM assesses neuronal subtype-specific acute neurotoxicity using micro-electrode
arrays (MEA) for the recording of spontaneous electrical activity (Hartmann et al., 2023). By sorting detected signals (spikes) based on their waveform, this assay allows the distinction between glutamatergic, GABAergic, dopaminergic, serotonergic, and cholinergic responses of the mixed-neuronal co-culture, allowing for an in vitro MoA-based assessment of ANT (Hartmann et al., 2023). In a third NAM, signaling will be assessed at the single cell level in mature central dopaminergic neurons (LUHMES cells), which extends the coverage of the KEs to another cell type and a set of functional receptors (e.g., P2X3 receptors (Apicella and Fabbretti, 2012)). A fourth NAM under development, the Peripheral Myelin Toxicity Assay (PeriMyelinTox), assesses myelin toxicity impacting peripheral sensory and motor function, and therefore addresses the key battery gap “myelination” for peripheral ANT in human cells. In this NAM, sensory or motor neurons, along with Schwann cells, will be differentiated from hiPSCs and cultivated in co-culture (Muller et al., 2018; Schenke et al., 2020; Louit et al., 2023). A novel 3D sphere format will be developed and compared to a conventional 2D format. Myelin formation will be evaluated by quantifying myelin basic protein (MBP) or myelin protein zero (MPZ) against the panneuronal marker -tubulin (TUJ1) through immunofluorescence staining and RT-qPCR (Chesnut et al., 2021). The assay will be optimized for automated high-throughput quantification of myelin post-exposure to a training set of potential myelin toxicants. The added value of both assays in assessing sensory and motor neuron myelin toxicity will be further evaluated.
To clearly define the applicability domains and assay-specific limitations, this project aims to develop a set of ANT reference chemicals that are known to affect the human brain, as well as negative compounds. This approach will ensure the unified characterization of the applicability domain of each assay and coverage of important human-relevant ANT MoA.
7 Outcomes and future perspectives
PARC was designed to address challenges inherent in moving from animal-based test methods to (batteries of) in vitro and alternative NAMs to speed up and modernize hazard identification and chemical risk assessment. Project 5.2.1e aims to improve the hazard prediction paradigm via the establishment and refinement of NAMs for DNT and ANT testing and the assembling of high performing, reproducible NAMs that provide added value into DNT and ANT test batteries. Our strategy encompasses the refinement of existing assays, generation of innovative NAMs to address identified gaps, determination of the applicability domain, and increased cost-effectiveness of lengthier assays via the demonstration of early indicators of later effects. Importantly, the development of new assays and the refinement of existing ones includes a strong focus on assay and data reliability. One critical ambition is to introduce quality control measures as described in Good Cell Culture Practices 2.0 (Pamies et al., 2022) or the GIVIMP document of the OECD (OECD, 2018). The PARC 5.2.1e consortium therefore contributes to an improved readiness, sensitivity, and overall performance of DNT NAMs to promote an increased acceptance of DNT in vitro and alternative assays for wider regulatory use. By the end of this project, a guidance document will be delivered that will introduce a novel framework
aiming to facilitate the regulatory use of data derived from the DNTIVB v2.0. Such work will include considerations on how DNT-IVB data may be used in the context of an IATA or weight of evidence for hazard and risk characterization. This links seamlessly to other PARC work packages that provide information on physiologicallybased kinetic (PBK) modelling to convert IVB concentrations to predicted in vivo doses, and to risk assessment specialists that need to consider how the predicted doses can be used to set safe exposure thresholds by, for example, considering modulatory factors in AOP or by considering variabilities and specific sensitivities in exposed populations (Schmeisser et al., 2023; Suciu et al., 2023).
In contrast to DNT, there is currently no comparable battery of NAMs for ANT testing. Therefore, the consortium will create a firstgeneration ANT-IVB v1.0, covering major MoAs involved in human brain functioning. Looking ahead, and to respond to PARC regulatory colleagues’ requests for data on specific compound classes, 5.2.1e NAMs will be used to evaluate the potential toxicity of natural toxins, bisphenols, and per- and polyfluoroalkyl substances (PFAS). Overall, this consortium aims to offer an unprecedented opportunity to fill a longstanding gap for the commonplace assessment of neurotoxicity potential of commercial chemicals via the generation of MoA-based, robust, reproducible, fast, and inexpensive consolidated DNT and ANT testing strategies. As all of this work is conducted under the guidance of colleagues from a regulatory field and potential end users, our ambition is to revolutionize the hazard and risk assessment of DNT and ANT in Europe.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
TT: Conceptualization, Funding acquisition, Project administration, Writing-original draft, Writing-review and editing. OM: Writing-review and editing, Funding acquisition, Project administration. EF: Writing-review and editing, Funding acquisition. JR: Writing-review and editing, Funding acquisition. KC: Writing-review and editing. KAH: Writing-review and editing. CA: Writing-review and editing. PB: Writing-review and editing, Funding acquisition. BE: Writing-review and editing, Funding acquisition. HD: Writing-review and editing, Funding acquisition. KH: Writing-review and editing. KD: Writing-review and editing. YH: Writing-review and editing. SH: Writing-review and editing, Funding acquisition. KJ: Writing-review and editing. BJ: Writingreview and editing. NK: Writing-review and editing. AK-G: Writing-review and editing. BK: Writing-review and editing. MLe: Writing-review and editing, Funding acquisition. MLi: Writing-review and editing. JL: Writing-review and editing, Funding acquisition. FM: Writing-review and editing, Funding acquisition. JC: Writing-review and editing, Funding acquisition. WN: Writing-review and editing, Funding acquisition. GP: Writingreview and editing. BS: Writing-review and editing, Funding acquisition. IS: Writing-review and editing. SS: Writing-review
and editing, Funding acquisition. OL: Writing-review and editing, Funding acquisition. MT-R: Writing-review and editing, Funding acquisition. KB: Conceptualization, Project administration, Writing-original draft, Writing-review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was carried out in the framework of the European Partnership for the Assessment of Risks from Chemicals (PARC) and has received funding from the European Union’s Horizon Europe research and innovation programme under Grant Agreement No 101057014. Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or the Health and Digital Executive Agency. Neither the European Union nor the granting authority can be held responsible for them. This work was also supported by a Helmholtz Association W2 award to T. Tal. The views expressed in this document are those of the authors and are not necessarily those of the EU agencies. Any statement made regarding EU regulations is solely intended for information purposes and does not represent the official opinion of the European Chemicals Agency (ECHA). ECHA is not responsible for any use that may be made of the information contained in this document. Statements made or information contained in this publication are without prejudice to any future work that ECHA may initiate. Funding performed in close connection to the European Union’s Horizon 2020 Research and
Innovation Programme under Grant Agreements No. 964518 (ToxFree), No. 964537 (RISK-HUNT3R), No. 965406 (PrecisionTox) and No. 963845 (ONTOX) in the ASPIS Cluster. Figures were generated using biorender.com.
Acknowledgments
We thank Shoko Furuno (ECHA) for her expert input.
Conflict of interest
EF and KB are shareholders of the DNTOX GmbH offering neurotoxicity testing services.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ankley, G. T., Bennett, R. S., Erickson, R. J., Hoff, D. J., Hornung, M. W., Johnson, R. D., et al. (2010). Adverse outcome pathways: a conceptual framework to support ecotoxicology research and risk assessment. Environ. Toxicol. Chem. 29 (3), 730-741. doi:10.1002/ETC. 34
Apicella, L., and Fabbretti, E. (2012). P2X3 receptor expression by HEK cells conditions their survival. Purinergic Signal. 8 (2), 295-300. doi:10.1007/S11302-011-9285-0
Appelt-Menzel, A., Cubukova, A., Günther, K., Edenhofer, F., Piontek, J., Krause, G., et al. (2017). Establishment of a human blood-brain barrier Co-culture model mimicking the neurovascular unit using induced pluri-and multipotent stem cells. Stem Cell Rep. 8, 894-906. doi:10.1016/j.stemcr.2017.02.021
Aung, M. T., Eick, S. M., Padula, A. M., Smith, S., Park, J. S., DeMicco, E., et al. (2023). Maternal per- and poly-fluoroalkyl substances exposures associated with higher depressive symptom scores among immigrant women in the chemicals in our bodies cohort in san francisco. Environ. Int. 172 (February), 107758. doi:10.1016/J. ENVINT.2023.107758
Babin, P. J., Goizet, C., and Raldúa, D. (2014). Zebrafish models of human motor neuron diseases: advantages and limitations. Prog. Neurobiol. 118, 36-58. doi:10.1016/J. PNEUROBIO.2014.03.001
Baccarelli, A., and Bollati, V. (2009). Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 21 (2), 243-251. doi:10.1097/MOP.0B013E32832925CC
Bal-Price, A., Hogberg, H. T., Crofton, K. M., Daneshian, M., FitzGerald, R. E., Fritsche, E., et al. (2018). Recommendation on test readiness criteria for new approach methods in Toxicology: exemplified for developmental neurotoxicity. Altex 35 (3), 306-352. doi:10.14573/altex. 1712081
Banks, W. A. (2009). Characteristics of compounds that cross the blood-brain barrier. BMC Neurol. 9 (Suppl. 1), S3-S9. doi:10.1186/1471-2377-9-S1-S3
Bartmann, K., Bendt, F., Dönmez, A., Haag, D., Eike Keßel, H., Masjosthusmann, S., et al. (2023). A human IPSC-based in vitro neural network formation assay to investigate neurodevelopmental toxicity of pesticides. ALTEX – Altern. Animal Exp. 40, 452-470. doi:10.14573/ALTEX. 2206031
Baumann, J., Barenys, M., Gassmann, K., and Fritsche, E. (2014). Comparative human and rat ‘neurosphere assay’ for developmental neurotoxicity testing. Curr. Protoc. Toxicol. 1 (Suppl. 59), 12-24. doi:10.1002/0471140856.TX1221S59
Baumann, J., Dach, K., Barenys, M., Giersiefer, S., Goniwiecha, J., Lein, P. J., et al. (2015). “Application of the neurosphere assay for DNT hazard assessment: challenges and limitations,” in Methods in pharmacology and Toxicology (Totowa, NJ: Humana Press), 29. doi:10.1007/7653_2015_49
Baumann, J., Gassmann, K., Masjosthusmann, S., DeBoer, D., Bendt, F., Giersiefer, S., et al. (2016). Comparative human and rat neurospheres reveal species differences in chemical effects on neurodevelopmental key events. Archives Toxicol. 90 (6), 1415-1427. doi:10.1007/S00204-015-1568-8
Bellanger, M., Demeneix, B., Grandjean, P., Thomas Zoeller, R., and Trasande, L. (2015). Neurobehavioral deficits, diseases, and associated costs of exposure to endocrine-disrupting chemicals in the European union. J. Clin. Endocrinol. Metabolism 100 (4), 1256-1266. doi:10.1210/JC.2014-4323
Bellanger, M., Pichery, C., Aerts, D., Berglund, M., Castaño, A., Čejchanová, M., et al. (2013). Economic benefits of methylmercury exposure control in Europe: monetary value of neurotoxicity prevention. Environ. Health A Glob. Access Sci. Source 12 (1), 3-10. doi:10.1186/1476-069x-12-3
Bennett, D., Bellinger, D. C., Birnbaum, L. S., Bradman, A., Chen, A., Cory-Slechta, D. A., et al. (2016). Project TENDR: targeting environmental neuro-developmental risks the TENDR consensus statement. Environ. Health Perspect. 124 (7), A118-A122. doi:10. 1289/EHP358
Bernal, J., Guadaño-Ferraz, A., and Morte, B. (2015). Thyroid hormone transporters-functions and clinical implications. Nat. Rev. Endocrinol. 11 (7), 690-717. doi:10.1038/ nrendo. 2015.186
Blum, J., Masjosthusmann, S., Bartmann, K., Bendt, F., Dolde, X., Dönmez, A., et al. (2022). Establishment of a human cell-based in vitro battery to assess developmental neurotoxicity hazard of chemicals. Chemosphere 311 (Pt 2), 137035. doi:10.1016/J. CHEMOSPHERE.2022.137035
Braunbeck, T., Kais, B., Lammer, E., Otte, J., Schneider, K., Stengel, D., et al. (2014). The fish embryo test (FET): origin, applications, and future. Environ. Sci. Pollut. Res. 22 (21), 16247-16261. doi:10.1007/S11356-014-3814-7
Bray, M. A., Singh, S., Han, H., Davis, C. T., Kost-Alimova, M., Gustafsdottir, S. M., et al. (2016). Cell painting, a high-content image-based assay for morphological profiling using multiplexed fluorescent dyes. Nat. Protoc. 11 (9), 1757-1774. doi:10. 1038/NPROT.2016.105
Brown, J. P., Hall, D., Frank, C. L., Wallace, K., Mundy, W. R., and Shafer, T. J. (2016). Editor’s highlight: evaluation of a microelectrode array-based assay for neural network ontogeny using training set chemicals. Toxicol. Sci. 154 (1), 126-139. doi:10.1093/ toxsci/kfw147
Brüll, M., Spreng, A.-S., Gutbier, S., Loser, D., Krebs, A., Reich, M., et al. (2020). Incorporation of stem cell-derived astrocytes into neuronal organoids to allow neuroglial interactions in toxicological studies. ALTEX – Altern. Animal Exp. 37 (3), 409-428. doi:10.14573/ALTEX. 1911111
Bruni, G., Rennekamp, A. J., Velenich, A., McCarroll, M., Gendelev, L., Fertsch, E., et al. (2016). Zebrafish behavioral profiling identifies multitarget antipsychotic-like compounds. Nat. Chem. Biol. 12 (7), 559-566. doi:10.1038/nchembio. 2097
Carson, M. J., Cameron Thrash, J., and Walter, B. (2006). The cellular response in neuroinflammation: the role of leukocytes, microglia and astrocytes in neuronal death and survival. Clin. Neurosci. Res. 6 (5), 237-245. doi:10.1016/J.CNR.2006.09.004
Chesnut, M., Paschoud, H., Repond, C., Smirnova, L., Hartung, T., Zurich, M.-G., et al. (2021). Human IPSC-derived model to study myelin disruption. Int. J. Mol. Sci. 22 (17), 9473. doi:10.3390/IJMS22179473
Chu, J., and Sadler, K. C. (2009). New school in liver development: lessons from zebrafish. Hepatology 50 (5), 1656-1663. doi:10.1002/HEP. 23157
Costa, L. G., Aschner, M., Vitalone, A., Syversen, T., and Soldin, O. P. (2004). Developmental neuropathology of environmental agents. Annu. Rev. Pharmacol. Toxicol. 44, 87-110. doi:10.1146/annurev.pharmtox.44.101802.121424
Costa, L. G., Giordano, G., Guizzetti, M., and Vitalone, A. (2008). Neurotoxicity of pesticides: a brief review. Front. Biosci. 13 (4), 1240-1249. doi:10.2741/2758
Craig, E., Kelly, L., Akerman, G., Dawson, J., May, B., Reaves, E., et al. (2019). Reducing the need for animal testing while increasing efficiency in a pesticide regulatory setting: lessons from the EPA office of pesticide programs’ hazard and science policy Council. Regul. Toxicol. Pharmacol. RTP 108 (November), 104481. doi:10.1016/J. YRTPH.2019.104481
Crofton, K. M., and Mundy, W. R. (2021). External scientific report on the interpretation of data from the developmental neurotoxicity in vitro testing assays for use in integrated approaches for testing and assessment. EFSA Support. Publ. 18 (10). doi:10.2903/SP.EFSA.2021.EN-6924
Crofton, K. M., Mundy, W. R., and Shafer, T. J. (2012). Developmental neurotoxicity testing: a path forward. Congenit. Anomalies. Congenit. Anom. (Kyoto) 52, 140-146. doi:10.1111/j.1741-4520.2012.00377.x
Dach, K., Bendt, F., Huebenthal, U., Giersiefer, S., Lein, P. J., Heuer, H., et al. (2017). BDE-99 impairs differentiation of human and mouse NPCs into the oligodendroglial lineage by species-specific modes of action. Sci. Rep. 7 (44861), 44861. doi:10.1038/ srep44861
Davidsen, N., Lauvås, A. J., Myhre, O., Ropstad, E., Carpi, D., Gyves, E. M. de, et al. (2021). Exposure to human relevant mixtures of halogenated persistent organic pollutants (POPs) alters neurodevelopmental processes in human neural stem cells undergoing differentiation. Reprod. Toxicol. 100 (April 2020), 17-34. doi:10.1016/j. reprotox.2020.12.013
Delp, J., Cediel-Ulloa, A., Suciu, I., Kranaster, P., Vugt-Lussenburg, B.Ma V., Munic Kos, V., et al. (2021). Neurotoxicity and underlying cellular changes of 21 mitochondrial respiratory chain inhibitors. Archives Toxicol. 95, 591-615. doi:10.1007/s00204-020-02970-5
Delp, J., Funke, M., Rudolf, F., Cediel, A., Bennekou, S. H., van der Stel, W., et al. (2019). Development of a neurotoxicity assay that is tuned to detect mitochondrial toxicants. Archives Toxicol. 93 (6), 1585-1608. doi:10.1007/S00204-019-02473-Y
Dickerson, A. S., Wu, A. C., Liew, Z., and Weisskopf, M. (2020). A scoping review of non-occupational exposures to environmental pollutants and adult depression, anxiety, and suicide. Curr. Environ. Health Rep. 7 (3), 256-271. doi:10.1007/s40572-020-00280-7
Druwe, I., Freudenrich, T. M., Wallace, K., Shafer, T. J., and Mundy, W. R. (2015). Sensitivity of neuroprogenitor cells to chemical-induced apoptosis using a multiplexed assay suitable for high-throughput screening. Toxicology 333 (July), 14-24. doi:10.1016/ J.TOX.2015.03.011
Duboc, V., Dufourcq, P., Blader, P., and Roussigné, M. (2015). Asymmetry of the brain: development and implications. Annu. Rev. Genet. 49, 647-672. doi:10.1146/ ANNUREV-GENET-112414-055322
Dubrana, L. E., Knoll-Gellida, A., Bourcier, L. M., Mercé, T., Pedemay, S., Nachon, F., et al. (2021). An antidote screening system for organophosphorus poisoning using zebrafish larvae. ACS Chem. Neurosci. 12 (15), 2865-2877. doi:10.1021/ ACSCHEMNEURO.1C00251
ECHA (2023). Key areas of regulatory challenge. Eur. Chem. Agency. ECHA-23-R-(ED-07-23-212-EN-N). doi:10.2823/568850
EFSA Scientific CommitteeHardy, A., Benford, D., Halldorsson, T., Jeger, M. J., Katrine Knutsen, H., More, S., et al. (2017). Guidance on the risk assessment of substances present in food intended for infants below 16 Weeks of age. EFSA J. 15 (5), e04849. doi:10.2903/J.EFSA.2017.4849
Eriksson, P. S., Perfilieva, E., Björk-Eriksson, T., Alborn, A. M., Nordborg, C., Peterson, D. A., et al. (1998). Neurogenesis in the adult human Hippocampus. Nat. Med. 4 (11), 1313-1317. doi:10.1038/3305
Escher, B. I., Altenburger, R., Blüher, M., Colbourne, J. K., Ebinghaus, R., Fantke, P., et al. (2023). Modernizing persistence-bioaccumulation-toxicity (PBT) assessment with high throughput animal-free methods. Archives Toxicol. 97 (5), 1267-1283. doi:10. 1007/S00204-023-03485-5
European Commission (2020). Communication from the commission to the European parliament, the Council, the European economic and social committee and the committee of the regions. Chem. Strategy Sustain. – Towards a Toxic-Free Environ.
European Parliament and Council. 2006. “Regulation (EC) No. 1907/2006 of the European parliament and of the Council of 18 december 2006 concerning the registration, evaluation, authorisation and restriction of chemicals (REACH), establishing a European chemicals agency, amending directive 1999/45/EC and repealing Council regulation (EEC) No 793/93 and commission regulation (EC) No 1488/94 as well as Council directive 76/769/EEC and commission directives 91/155/ EEC, 93/67/EEC, 93/105/EC and 2000/21/EC.”
European Parliament and Council. 2008. “Regulation (EC) No. 1272/2008 of the European parliament and the Council of 16 december 2008 on classification, labelling and packaging of substances, and mixtures, amending and repealing directives 67/548/ EEC and 1999/45/EC, and amending regulation (EC) No 1907/2006.” In .
European Parliament and Council (2009). Regulation (EC) No. 1107/2009 of the European parliament and of the Council of 21 october 2009 concerning the placing of plant protection products on the market and repealing Council directives 79/117/EEC and 91/414/EEC. Paris: Brussels, OECD.
European Parliament and Council. 2010. “Directive 2010/63/EU of the European paliament and of the Council of 22 september 2010 on the protection of animals used for scientific purposes.” In .
Faria, M., Garcia-Reyero, N., Padrós, F., Babin, P. J., Sebastián, D., Cachot, J., et al. (2015). Zebrafish models for human acute organophosphorus poisoning. Sci. Rep. 5 (October), 15591. doi:10.1038/SREP15591
Farrell, J. A., Wang, Y., Riesenfeld, S. J., Shekhar, K., Regev, A., and Schier, A. F. (2018). Single-cell reconstruction of developmental trajectories during zebrafish embryogenesis. Science 360 (6392), eaar3131. doi:10.1126/science.aar3131
Frank, C. L., Brown, J. P., Wallace, K., Mundy, W. R., and Shafer, T. J. (2017). From the cover: developmental neurotoxicants disrupt activity in cortical networks on microelectrode arrays: results of screening 86 compounds during neural network formation. Toxicol. Sci. 160 (1), 121-135. doi:10.1093/toxsci/kfx169
Fritsche, E. (2017). “Report on integrated testing strategies for the identification and evaluation of chemical hazards associated with the developmental neurotoxicity (DNT), to facilitate discussions at the Joint EFSA/OECD workshop on DNT.” ENV/JM/ MONO(2017)4/ANN1.
Fritsche, E., Grandjean, P., Crofton, K. M., Aschner, M., Goldberg, A., Heinonen, T., et al. (2018). Consensus statement on the need for innovation, transition and implementation of developmental neurotoxicity (DNT) testing for regulatory purposes. Toxicol. Appl. Pharmacol. 354 (September), 3-6. doi:10.1016/J.TAAP.2018.02.004
Gaballah, S., Swank, A., Sobus, J. R., Meng Howey, X., Schmid, J., Catron, T., et al. (2020). Evaluation of developmental toxicity, developmental neurotoxicity, and tissue dose in zebrafish exposed to GenX and other PFAS. Environ. Health Perspect. 128 (4), 47005. doi:10.1289/EHP5843
Gadaleta, D., Spînu, N., Roncaglioni, A., Cronin, M. T. D., and Benfenati, E. (2022). Prediction of the neurotoxic potential of chemicals based on modelling of molecular initiating events upstream of the adverse outcome pathways of (developmental) neurotoxicity. Int. J. Mol. Sci. 23 (6), 3053. doi:10.3390/ijms23063053
Giordano, G., and Costa, L. G. (2012). Developmental neurotoxicity: some old and new issues. ISRN Toxicol. 2012 (June), 814795-814812. doi:10.5402/2012/814795
Glaberman, S., Padilla, S., and Barron, M. G. (2017). Evaluating the zebrafish embryo toxicity test for pesticide hazard screening. Environ. Toxicol. Chem. 36 (5), 1221-1226. doi:10.1002/ETC. 3641
Grandjean, P., and Landrigan, P. J. (2006). Developmental neurotoxicity of industrial chemicals. Lancet 368 (9553), 2167-2178. doi:10.1016/S0140-6736(06)69665-7
Grandjean, P., and Landrigan, P. J. (2014). Neurobehavioural effects of developmental toxicity. Lancet Neurology 13 (3), 330-338. doi:10.1016/S1474-4422(13)70278-3
Grillberger, K., Cöllen, E., Immacolata Trivisani, C., Blum, J., Leist, M., and Ecker, G. F. (2023). Structural insights into neonicotinoids and N -unsubstituted metabolites on human NAChRs by molecular docking, dynamics simulations, and calcium imaging. Int. J. Mol. Sci. 24 (17), 13170. doi:10.3390/IJMS241713170
Gunnarsson, L., Jauhiainen, A., Kristiansson, E., Nerman, O., and Joakim Larsson, D. G. (2008). Evolutionary conservation of human drug targets in organisms used for
environmental risk assessments. Environ. Sci. Technol. 42 (15), 5807-5813. doi:10.1021/ es8005173
Gupta, T., Marquart, G. D., Horstick, E. J., Tabor, K. M., Pajevic, S., and Burgess, H. A. (2018). Morphometric analysis and neuroanatomical mapping of the zebrafish brain. Methods 150 (November), 49-62. doi:10.1016/J.YMETH.2018.06.008
Gutbier, S., Spreng, A. S., Delp, J., Schildknecht, S., Karreman, C., Suciu, I., et al. (2018). Prevention of neuronal apoptosis by astrocytes through thiol-mediated stress response modulation and accelerated recovery from proteotoxic stress. Cell Death Differ. 25 (12), 2101-2117. doi:10.1038/S41418-018-0229-X
Harrill, J. A., Freudenrich, T., Wallace, K., Ball, K., Shafer, T. J., and Mundy, W. R. (2018). Testing for developmental neurotoxicity using a battery of in vitro assays for key cellular events in neurodevelopment. Toxicol. Appl. Pharmacol. 354 (September), 24-39. doi:10.1016/J.TAAP.2018.04.001
Harrill, J. A., Freudenrich, T. M., Machacek, D. W., Stice, S. L., and Mundy, W. R. (2010). Quantitative assessment of neurite outgrowth in human embryonic stem cellderived HN2 cells using automated high-content image analysis. Neurotoxicology 31 (3), 277-290. doi:10.1016/J.NEURO.2010.02.003
Harrill, J. A., Robinette, B. L., Freudenrich, T., and Mundy, W. R. (2013). Use of high content image analyses to detect chemical-mediated effects on neurite sub-populations in primary rat cortical neurons. NeuroToxicology 34 (1), 61-73. doi:10.1016/J.NEURO. 2012.10.013
Harrill, J. A., Robinette, B. L., and Mundy, W. R. (2011). Use of high content image analysis to detect chemical-induced changes in synaptogenesis in vitro. Toxicol. Vitro 25 (1), 368-387. doi:10.1016/j.tiv.2010.10.011
Harry, G. J., McBride, S., Witchey, S. K., Mhaouty-Kodja, S., Trembleau, A., Bridge, M., et al. (2022). Roadbumps at the crossroads of integrating behavioral and in vitro approaches for neurotoxicity assessment. Front. Toxicol. 4, 812863. doi:10.3389/FTOX. 2022.812863
Hartmann, J., Henschel, N., Bartmann, K., Dönmez, A., Brockerhoff, G., Koch, K., et al. (2023). Molecular and functional characterization of different BrainSphere models for use in neurotoxicity testing on microelectrode arrays. Cells 12 (9), 1270. doi:10.3390/ cells12091270
Hoelting, L., Klima, S., Karreman, C., Grinberg, M., Meisig, J., Henry, M., et al. (2016). Stem cell-derived immature human dorsal root ganglia neurons to identify peripheral neurotoxicants. Stem Cells Transl. Med. 5 (4), 476-487. doi:10.5966/SCTM.2015-0108
Hollander, J. A., Cory-Slechta, D. A., Jacka, F. N., Szabo, S. T., Guilarte, T. R., Bilbo, S. D., et al. (2020). Beyond the looking glass: recent advances in understanding the impact of environmental exposures on neuropsychiatric disease. Neuropsychopharmacology 45 (7), 1086-1096. doi:10.1038/s41386-020-0648-5
Holzer, A. K., Karreman, C., Suciu, I., Furmanowsky, L. S., Wohlfarth, H., Loser, D., et al. (2022). Generation of human nociceptor-enriched sensory neurons for the study of pain-related dysfunctions. Stem Cells Transl. Med. 11 (7), 727-741. doi:10.1093/ STCLTM/SZAC031
Hooijmans, C. R., Leenaars, M., and Ritskes-Hoitinga, M. (2010). A gold standard publication checklist to improve the quality of animal studies, to fully integrate the three rs, and to make systematic reviews more feasible. Altern. Lab. Anim. 38 (2), 167-182. doi:10.1177/026119291003800208
Horzmann, K. A., and Freeman, J. L. (2016). Zebrafish get connected: investigating neurotransmission targets and alterations in chemical toxicity. Toxics 4 (3), 19. doi:10. 3390/TOXICS4030019
Howe, K., Clark, M. D., Torroja, C. F., Torrance, J., Berthelot, C., Muffato, M., et al. (2013). The zebrafish reference genome sequence and its relationship to the human genome. Nature 496, 498-503. doi:10.1038/nature12111
Imayoshi, I., Shimojo, H., Sakamoto, M., Ohtsuka, T., and Kageyama, R. (2013). Genetic visualization of Notch signaling in mammalian neurogenesis. Cell. Mol. Life Sci. 70, 2045-2057. doi:10.1007/s00018-012-1151-x
Jacobson, M. H., Ghassabian, A., Gore, A. C., and Trasande, L. (2022). Exposure to environmental chemicals and perinatal psychopathology. Biochem. Pharmacol. 195 (January), 114835. doi:10.1016/J.BCP.2021.114835
Jarema, K. A., Hunter, D. L., Hill, B. N., Olin, J. K., Britton, K. N., Waalkes, M. R., et al. (2022). Developmental neurotoxicity and behavioral screening in larval zebrafish with a comparison to other published results. Toxics 10 (5), 256. doi:10.3390/toxics10050256
Kalueff, A. V., Gebhardt, M., Stewart, A. M., Cachat, J. M., Brimmer, M., Chawla, J. S., et al. (2013). Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 10 (1), 70-86. Https://Home.Liebertpub.Com/Zeb. doi:10.1089/ZEB.2012. 0861
Kang, Du S., Yang, Y. R., Lee, C., Kim, S. B., Ryu, S.Ho, and Suh, P. G. (2016). Roles of phosphoinositide-specific phospholipase in brain development. Adv. Biol. Regul. 60 (January), 167-173. doi:10.1016/J.JBIOR.2015.10.002
Kavlock, R. J., Bahadori, T., Barton-Maclaren, T. S., Gwinn, M. R., Rasenberg, M., and Thomas, R. S. (2018). Accelerating the pace of chemical risk assessment. Chem. Res. Toxicol. 31 (5), 287-290. doi:10.1021/ACS.CHEMRESTOX.7B00339
Kettleborough, R. N. W., Busch-Nentwich, E. M., Harvey, S. A., Dooley, C. M., De Bruijn, E., Van Eeden, F., et al. (2013). A systematic genome-wide analysis of zebrafish protein-coding gene function. Nature 496 (7446), 494-497. doi:10.1038/ NATURE11992
Khelfaoui, H., Harkati, D., and Saleh, B. A. (2021). Molecular docking, molecular dynamics simulations and reactivity, studies on approved drugs library targeting ACE2 and SARS-CoV-2 binding with ACE2. J. Biomol. Struct. Dyn. 39 (18), 7246-7262. doi:10.1080/07391102.2020.1803967
Klima, S., Brüll, M., Spreng, A. S., Suciu, I., Falt, T., Schwamborn, J. C., et al. (2021). A human stem cell-derived test system for agents modifying neuronal N-Methyl-D-Aspartate-Type glutamate receptor -signalling. Archives Toxicol. 95 (5), 1703-1722. doi:10.1007/S00204-021-03024-0
Klose, J., Tigges, J., Masjosthusmann, S., Schmuck, K., Bendt, F., Hübenthal, U., et al. (2021). TBBPA targets converging key events of human oligodendrocyte development resulting in two novel AOPs. Altex 38 (2), 215-234. doi:10.14573/altex. 2007201
Klüver, N., Vogs, C., Altenburger, R., Escher, B. I., and Scholz, S. (2016). Development of a general baseline toxicity QSAR model for the fish embryo acute toxicity test. Chemosphere 164 (December), 164-173. doi:10.1016/J.CHEMOSPHERE.2016.08.079
Knoll-Gellida, A., Dubrana, L. E., Bourcier, L. M., eo Merc, Th, Gruel, E., Soares, M., et al. (2021). Hyperactivity and seizure induced by tricresyl phosphate are isomer specific and not linked to phenyl valerate-neuropathy target esterase activity inhibition in zebrafish. Toxicol. Sci. 180 (1), 160-174. doi:10.1093/toxsci/kfab006
Koch, K., Bartmann, K., Hartmann, J., Kapr, J., Pahl, M., Fritsche, E., et al. (2022). Scientific validation of human neurosphere assays for developmental neurotoxicity evaluation. Front. Press 0, 816370-816438. doi:10.3389/FTOX.2022.816370
Krebs, A., Waldmann, T., Wilks, M. F., van Vugt-Lussenburg, B. M. A., van der Burg, B., Terron, A., et al. (2019). Template for the description of cell-based toxicological test methods to allow evaluation and regulatory use of the data. ALTEX – Altern. Animal Exp. 36 (4), 682-699. doi:10.14573/ALTEX. 1909271
Krug, A. K., Balmer, N. V., Matt, F., Schönenberger, F., Merhof, D., and Leist, M. (2013). Evaluation of a human neurite growth assay as specific screen for developmental neurotoxicants. Archives Toxicol. 87 (12), 2215-2231. doi:10.1007/s00204-013-1072-y
Krzykwa, J. C., Saeid, A., and Sellin Jeffries, M. K. (2019). Identifying sublethal endpoints for evaluating neurotoxic compounds utilizing the fish embryo toxicity test. Ecotoxicol. Environ. Saf. 170 (April), 521-529. doi:10.1016/J.ECOENV.2018. 11.118
Landrigan, P. J., Sonawane, B., Butler, R. N., Trasande, L., Callan, R., and Droller, D. (2005). Early environmental origins of neurodegenerative disease in later life. Environ. Health Perspect. 113 (9), 1230-1233. doi:10.1289/EHP. 7571
Lauvås, A. J., Lislien, M., Andreas Holme, J., Dirven, H., Paulsen, E., Alm, I. M., et al. (2022). Developmental neurotoxicity of acrylamide and its metabolite glycidamide in a human mixed culture of neurons and astrocytes undergoing differentiation in concentrations relevant for human exposure. Neurotoxicology 92, 33-48. doi:10. 1016/j.neuro.2022.07.001
Lee, Da Y. (2015). Roles of MTOR signaling in brain development. Exp. Neurobiol. 24 (3), 177-185. doi:10.5607/EN.2015.24.3.177
Lee, J., Escher, B. I., Scholz, S., and Schlichting, R. (2022a). Inhibition of neurite outgrowth and enhanced effects compared to baseline toxicity in SH-SY5Y cells. Archives Toxicol. 96 (4), 1039-1053. doi:10.1007/s00204-022-03237-x
Lee, J., Schlichting, R., König, M., Scholz, S., Krauss, M., and Escher, B. I. (2022b). Monitoring mixture effects of neurotoxicants in surface water and wastewater treatment plant effluents with neurite outgrowth inhibition in SH-SY5Y cells. Am. Chem. Soc. 2, 523-535. doi:10.1021/acsenvironau.2c00026
Leist, M., Ahmed, G., Graepel, R., Marchan, R., Hassan, R., Hougaard Bennekou, S., et al. (2017). Adverse outcome pathways: opportunities, limitations and open questions. Archives Toxicol. 91 (11), 3477-3505. doi:10.1007/S00204-017-2045-3
López-Espíndola, D., Morales-Bastos, C., Grijota-Martínez, C., Liao, X. H., Lev, D., Sugo, E., et al. (2014). Mutations of the thyroid hormone transporter MCT8 cause prenatal brain damage and persistent hypomyelination. J. Clin. Endocrinol. Metabolism 99 (12), E2799-E2804. doi:10.1210/jc.2014-2162
Loser, D., Hinojosa, M. G., Blum, J., Schaefer, J., Brüll, M., Johansson, Y., et al. (2021). Functional alterations by a subgroup of neonicotinoid pesticides in human dopaminergic neurons. Archives Toxicol. 95 (6), 2081-2107. doi:10.1007/S00204-021-03031-1
Louit, A., Beaudet, M. J., Pépin, R., and Berthod, F. (2023). Differentiation of human induced pluripotent stem cells into mature and myelinating Schwann cells. Https// Home.Liebertpub.Com/Tec 29 (4), 134-143. doi:10.1089/TEN.TEC.2022.0186
Makris, S. L., Raffaele, K., Allen, S., Bowers, W. J., Hass, U., Alleva, E., et al. (2009). A retrospective performance assessment of the developmental neurotoxicity study in support of OECD test guideline 426. Environ. Health Perspect. 117 (1), 17-25. doi:10. 1289/EHP. 11447
Marx-Stoelting, P., Rivière, G., Luijten, • M., Aiello-Holden, • K., Bandow, • N., Baken, • K., et al. (2023). A walk in the PARC: developing and implementing 21st century chemical risk assessment in Europe. Arch. Toxicol. 97, 893-908. doi:10.1007/s00204-022-03435-7
Masjosthusmann, S., Barenys, M., El-Gamal, M., Geerts, L., Gerosa, L., Gorreja, A., et al. (2018). Literature review and appraisal on alternative neurotoxicity testing methods. EFSA Support. Publ. 15 (4), 1-108. doi:10.2903/sp.efsa.2018.en-1410
Masjosthusmann, S., Blum, J., Bartmann, K., Dolde, X., Holzer, A.-K., Stürzl, L.-C., et al. (2020). Establishment of an a priori protocol for the implementation and
interpretation of an in-vitro testing battery for the assessment of developmental neurotoxicity. EFSA Supporting Publication. Wiley. doi:10.2903/sp.efsa.2020.en-1938
Muller, Q., Beaudet, M. J., De Serres-Bérard, T., Bellenfant, S., Vincent, F., and Berthod, F. (2018). Development of an innervated tissue-engineered skin with human sensory neurons and Schwann cells differentiated from IPS cells. Acta Biomater. 82 (December), 93-101. doi:10.1016/J.ACTBIO.2018.10.011
Nimtz, L., Klose, J., Masjosthusmann, S., Barenys, M., and Fritsche, E. (2019). The neurosphere assay as an in vitro method for developmental neurotoxicity (DNT) evaluation. Eval. Neuromethods 145, 141-168. doi:10.1007/978-1-4939-9228-7_8
Noyes, P. D., Haggard, D. E., Gonnerman, G. D., and Tanguay, R. L. (2015). Advanced morphological – behavioral test platform reveals neurodevelopmental defects in embryonic zebrafish exposed to comprehensive suite of halogenated and organophosphate flame retardants. Toxicol. Sci. 145 (1), 177-195. doi:10.1093/ TOXSCI/KFV044
Nyffeler, J., Dolde, X., Krebs, A., Pinto-Gil, K., Pastor, M., Behl, M., et al. (2017). Combination of multiple neural crest migration assays to identify environmental toxicants from a proof-of-concept chemical library. Archives Toxicol. 91 (11), 3613-3632. doi:10.1007/s00204-017-1977-y
Ockleford, C., Adriaanse, P., Berny, P., Brock, T., Duquesne, S., Grilli, S., et al. (2017). Investigation into experimental toxicological properties of plant protection products having a potential link to Parkinson’s disease and childhood leukaemia. EFSA J. 15 (3), e04691. doi:10.2903/J.EFSA.2017.4691
OECD. (1995a). “Test No. 418: delayed neurotoxicity of organophosphorus substances following acute exposure.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070905-en
OECD. (1995b). “Test No. 419: delayed neurotoxicity of organophosphorus substances: 28-day repeated dose study.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070929-en
OECD. (1997). “Test No. 424: neurotoxicity study in rodents.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264071025-en
OECD. (2002a). “Test No. 420: acute oral toxicity – fixed dose procedure.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070943-en
OECD. (2002b). “Test No. 423: acute oral toxicity – acute toxic class method.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264071001-en
OECD. (2005). “Guidance document on the validation and international acceptance of new or updated test methods for hazard assessment, series on testing and assessment No. 34.” In .
OECD. 2007. “Test No. 426: developmental neurotoxicity study.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264067394-en
OECD (2008a). Series on testing and assessment No. 89 – retrospective performance assessment of the test guideline 426 on developmental neurotoxicity. ENV/JM/ MON (89).
OECD. 2008b. “Test No. 407: repeated dose 28-day oral toxicity study in rodents.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264070684-en
OECD. (2009). “Test No. 403: acute inhalation toxicity.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070608-en
OECD. (2017). “Test No. 402: acute dermal toxicity.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/9789264070585-en
OECD. (2018a). “Test No. 408: repeated dose 90-day oral toxicity study in rodents.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264070707-en
OECD. (2022). “Test No. 425: acute oral toxicity – up-and-down procedure.” In . OECD guidelines for the testing of chemicals, Section 4. OECD. doi:10.1787/ 9789264071049-en
OECD (2023). Initial recommendations on evaluation of data from the developmental neurotoxicity (DNT) in-vitro testing battery. Ser. Test. Assess. 377 (13). ENV/CBC/MONO(2023. Available at: https://www.oecd.org/env/ehs/testing/ developmental-neurotoxicity.htm.
Pallocca, G., Moné, M. J., Kamp, H., Luijten, M., van de Water, B., and Leist, M. (2022). Next-generation risk assessment of chemicals – rolling out a human-centric testing strategy to drive 3R implementation: the RISK-hunt3r project perspective. Altex 39 (3), 419-426. doi:10.14573/ALTEX. 2204051
Pamies, D., Leist, M., Coecke, S., Bowe, G., Allen, D. G., Gstraunthaler, G., et al. (2022). Guidance document on Good cell and tissue culture practice 2.0 (GCCP 2.0). ALTEX 39 (1), 30-70. doi:10.14573/ALTEX. 2111011
Paparella, M., Bennekou, S. H., and Bal-Price, A. (2020). An analysis of the limitations and uncertainties of in vivo developmental neurotoxicity testing and assessment to identify the potential for alternative approaches. Reprod. Toxicol. 96 (September), 327-336. doi:10.1016/J.REPROTOX.2020.08.002
Pitzer, E. M., Shafer, T. J., and Herr, D. W. (2023). Identification of Neurotoxicology (NT)/Developmental Neurotoxicology (DNT) adverse outcome pathways and key event linkages with in vitro DNT screening assays. Neurotoxicology 99 (December), 184-194. doi:10.1016/J.NEURO.2023.10.007
Raj, B., Wagner, D. E., McKenna, A., Pandey, S., Klein, A. M., Shendure, J., et al. (2018). Simultaneous single-cell profiling of lineages and cell types in the vertebrate brain. Nat. Biotechnol. 36 (5), 442-450. doi:10.1038/nbt. 4103
Rice, D., and Barone, S. (2000). Critical periods of vulnerability for the developing nervous system: evidence from humans and animal models. Environ. Health Perspect. 108 (Suppl. 3), 511-533. doi:10.1289/ehp.00108s3511
Rietdijk, J., Aggarwal, T., Georgieva, P., Lapins, M., Carreras-Puigvert, J., and Spjuth, O. (2022). Morphological profiling of environmental chemicals enables efficient and untargeted exploration of combination effects, Sci. Total Environ. 832, 155058. doi:10. 1016/j.scitotenv.2022.155058
Rokoff, L. B., Shoaff, J. R., Coull, B. A., Enlow, M. B., Bellinger, D. C., and Korrick, S. A. (2022). Prenatal exposure to a mixture of organochlorines and metals and internalizing symptoms in childhood and adolescence. Environ. Res. 208 (May), 112701. doi:10.1016/J.ENVRES.2022.112701
Sachana, M., Bal-Price, A., Crofton, K. M., Bennekou, S. H., Shafer, T. J., Behl, M., et al. (2019). International regulatory and scientific effort for improved developmental neurotoxicity testing. Toxicol. Sci. 167 (1), 45-57. doi:10.1093/toxsci/kfy211
Saili, K. S., Todd, J. Z., Schwab, A. J., Silvin, A., Baker, N. C., Sidney Hunter, E., et al. (2017). Blood-brain barrier development: systems modeling and predictive Toxicology. Birth Defects Res. 109 (20), 1680-1710. doi:10.1002/BDR2.1180
Sakai, C., Ijaz, S., Hoffman, E. J., Grünblatt, E., Carl, M., and Lange, M. (2018). Zebrafish models of neurodevelopmental disorders: past, present, and future. Front. Mol. Neurosci. 11, 294. doi:10.3389/fnmol.2018.00294
Sato-Maeda, M., Tawarayama, H., Obinata, M., Kuwada, J. Y., and Shoji, W. (2006). Sema3al guides spinal motor axons in a cell- and stage-specific manner in zebrafish. Dev. Camb. Engl. 133 (5), 937-947. doi:10.1242/DEV. 02268
Schenke, M., Schjeide, B. M., Püschel, G. P., and Seeger, B. (2020). Analysis of motor neurons differentiated from human induced pluripotent stem cells for the use in cellbased botulinum neurotoxin activity assays. Toxins 12 (5), 276. doi:10.3390/ TOXINS12050276
Schmeisser, S., Miccoli, A., von Bergen, M., Berggren, E., Braeuning, A., Busch, W., et al. (2023). New approach methodologies in human regulatory Toxicology – not if, but how and when. Environ. Int. 178 (March), 108082. doi:10.1016/J.ENVINT.2023.108082
Schmidt, B. Z., Lehmann, M., Gutbier, S., Nembo, E., Noel, S., Smirnova, L., et al. (2017). In vitro acute and developmental neurotoxicity screening: an overview of cellular platforms and high-throughput technical possibilities. Archives Toxicol. 91 (1), 1-33. doi:10.1007/S00204-016-1805-9
Schmidt, R., Strähle, U., and Scholpp, S. (2013). Neurogenesis in zebrafish – from embryo to adult. Neural Dev. 8 (1), 3-13. doi:10.1186/1749-8104-8-3
Schmuck, M. R., Temme, T., Dach, K., Boer, D. de, Barenys, M., Bendt, F., et al. (2017). Omnisphero: a high-content image analysis (hca) approach for phenotypic developmental neurotoxicity (DNT) screenings of organoid neurosphere cultures in vitro. Archives Toxicol. 91 (4), 2017-2028. doi:10.1007/s00204-016-1852-2
Semple, B. D., Blomgren, K., Gimlin, K., Ferriero, D. M., and Noble-Haeusslein, L. J. (2013). Brain development in rodents and humans: identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 106-107, 1-16. 0 (July). doi:10.1016/J.PNEUROBIO.2013.04.001
Shafer, T. J. (2019). Application of microelectrode array approaches to neurotoxicity testing and screening. Adv. Neurobiol. 22, 275-297. doi:10.1007/ 978-3-030-11135-9_12
Sheets, L. P., Li, A. A., Minnema, D. J., Collier, R. H., Creek, M. R., and Peffer, R. C. (2016). A critical review of neonicotinoid insecticides for developmental neurotoxicity. Crit. Rev. Toxicol. 46 (2), 153-190. doi:10.3109/10408444.2015. 1090948
Smirnova, L., Hogberg, H. T., Leist, M., and Hartung, T. (2014). Food for thought.: developmental neurotoxicity – challenges in the 21st century and in vitro opportunities. Altex 31 (2), 129-156. doi:10.14573/ALTEX. 1403271
Sobanska, M., Scholz, S., Nyman, A. M., Cesnaitis, R., Alonso, S. G., Klüver, N., et al. (2018). Applicability of the fish embryo acute toxicity (FET) test (OECD 236) in the regulatory context of registration, evaluation, authorisation, and restriction of chemicals (REACH). Environ. Toxicol. Chem. 37 (3), 657-670. doi:10.1002/ETC. 4055
Spalding, K. L., Bergmann, O., Alkass, K., Bernard, S., Salehpour, M., Huttner, H. B., et al. (2013). Dynamics of hippocampal neurogenesis in adult humans. Cell 153 (6), 1219-1227. doi:10.1016/j.cell.2013.05.002
Spencer, P. S., and Lein, P. J. (2014). “Neurotoxicity,” in Encyclopedia of Toxicology (January, Third Edition), 489-500. doi:10.1016/B978-0-12-386454-3.00169-X
Spînu, N., Cronin, M. T. D., Lao, J., Bal-Price, A., Campia, I., Enoch, S. J., et al. (2021). Probabilistic modelling of developmental neurotoxicity based on a simplified adverse outcome pathway network. Comput. Toxicol., 100206. doi:10.1016/j.comtox.2021. 100206
Spreng, A. S., Brüll, M., Leisner, H., Suciu, I., and Leist, M. (2022). Distinct and dynamic transcriptome adaptations of IPSC-generated astrocytes after cytokine stimulation. Cells 11 (17), 2644. doi:10.3390/CELLS11172644
Strähle, U., Scholz, S., Geisler, R., Greiner, P., Hollert, H., Rastegar, S., et al. (2012). Zebrafish embryos as an alternative to animal experiments-a commentary on the definition of the onset of protected life stages in animal welfare regulations. Reprod. Toxicol. 33 (2), 128-132. doi:10.1016/J.REPROTOX.2011.06.121
Stucki, A. O., Barton-Maclaren, T. S., Bhuller, Y., Henriquez, J. E., Henry, T. R., Hirn, C., et al. (2022). Use of new approach methodologies (NAMs) to meet regulatory requirements for the assessment of industrial chemicals and pesticides for effects on human health. Front. Toxicol. 4, 964553. doi:10.3389/FTOX.2022.964553
Suciu, I., Pamies, D., Peruzzo, R., Wirtz, P. H., Smirnova, L., Pallocca, G., et al. (2023). G E interactions as a basis for toxicological uncertainty. Archives Toxicol. 97 (7), 2035-2049. doi:10.1007/S00204-023-03500-9
Tanner, C. M., Goldman, S. M., Webster Ross, G., and Grate, S. J. (2014). The disease intersection of susceptibility and exposure: chemical exposures and neurodegenerative disease risk. Alzheimer’s Dementia 10 (3 Suppl. L), S213-S225. doi:10.1016/J.JALZ.2014.04.014
Terron, A., and Bennekou, S. H. (2018). Towards a regulatory use of alternative developmental neurotoxicity testing (DNT). Toxicol. Appl. Pharmacol. 354 (December 2017), 19-23. doi:10.1016/j.taap.2018.02.002
Tian, G., Harrison, P. J., Sreenivasan, A. P., Carreras-Puigvert, J., and Spjuth, O. (2023). Combining molecular and cell painting image data for mechanism of action prediction. Artif. Intell. Life Sci. 3 (December), 100060. doi:10.1016/J.AILSCI.2023. 100060
Tsuji, R., and Crofton, K. M. (2012). Developmental neurotoxicity guideline study: issues with methodology, evaluation and regulation. Congenit. Anomalies 52 (3), 122-128. doi:10.1111/J.1741-4520.2012.00374.X
Vinken, M., Benfenati, E., Busquet, F., Castell, J., Clevert, D.-A., de Kok, T. M., et al. (2021). Safer chemicals using less animals: kick-off of the European ONTOX project. Toxicology 458 (June), 152846. doi:10.1016/J.TOX.2021.152846
Wang, Z., Walker, G. W., Muir, D. C. G., and Nagatani-Yoshida, K. (2020). Toward a global understanding of chemical pollution: a first comprehensive analysis of national and regional chemical inventories. Environ. Sci. Technol. 54 (5), 2575-2584. doi:10. 1021/acs.est.9b06379