العين، وهي امتداد تشريحي للجهاز العصبي المركزي، تظهر العديد من التوازي الجزيئي والخلوية مع الدماغ. تظهر الأبحاث الناشئة أن التغيرات في الدماغ غالبًا ما تنعكس في العين، لا سيما في الشبكية.ومع ذلك، لا تزال إمكانية وجود رابط مناعي بين الجزء الخلفي من العين وبقية أنسجة الجهاز العصبي المركزي غير مستكشفة. هنا، من خلال دراسة الاستجابات المناعية لفيروس الهربس البسيط في الدماغ، لاحظنا أن التحصين داخل الجسم الزجاجي يحمي الفئران من التحدي الفيروسي داخل الجمجمة. امتد هذا الحماية لتشمل البكتيريا وحتى الأورام، مما سمح باستجابات مناعية علاجية ضد الورم الدبقي من خلال التحصين داخل الجسم الزجاجي. نوضح أيضًا أن الأقسام الأمامية والخلفية من العين لديها أنظمة تصريف لمفاوي متميزة، حيث تصرف الأخيرة إلى العقد اللمفاوية العنقية العميقة عبر الأوعية اللمفاوية في غمد العصب البصري. يمكن تعديل هذا التصريف اللمفاوي الخلفي، مثل تصريف اللمفاويات السحائية، بواسطة المحفز اللمفاوي VEGFC. وعلى العكس، نوضح أن تثبيط الإشارات اللمفاوية على العصب البصري يمكن أن يتغلب على قيد رئيسي في العلاج الجيني من خلال تقليل الاستجابة المناعية لفيروس الأدينو المرتبط وضمان استمرار الفعالية بعد عدة جرعات. تكشف هذه النتائج عن دائرة لمفاوية مشتركة قادرة على تقديم استجابة مناعية موحدة بين الجزء الخلفي من العين والدماغ، مما يبرز ميزة مناعية غير مدروسة للعين ويفتح المجال لاستراتيجيات علاجية جديدة في الأمراض العينية وأمراض الجهاز العصبي المركزي.
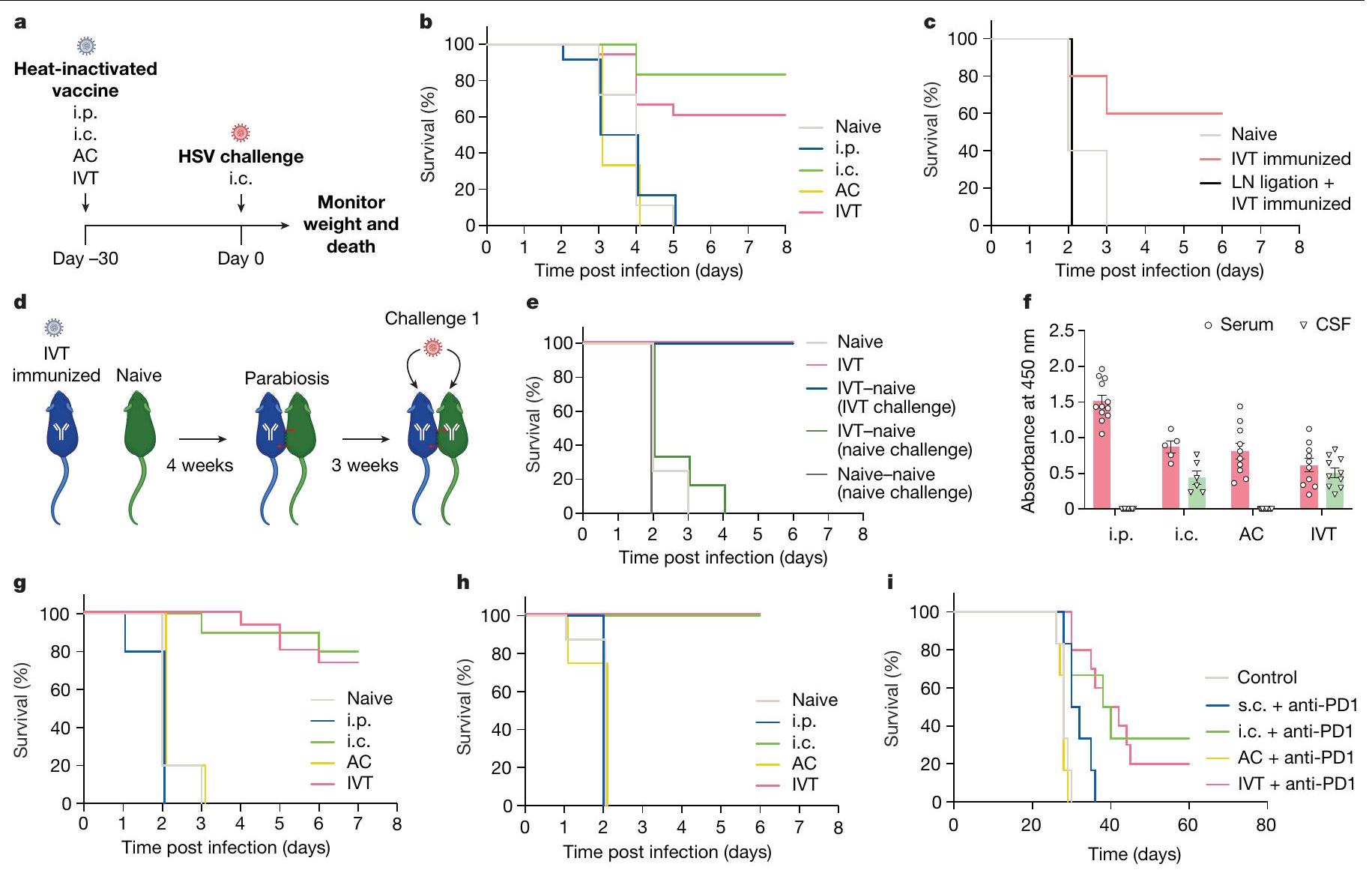
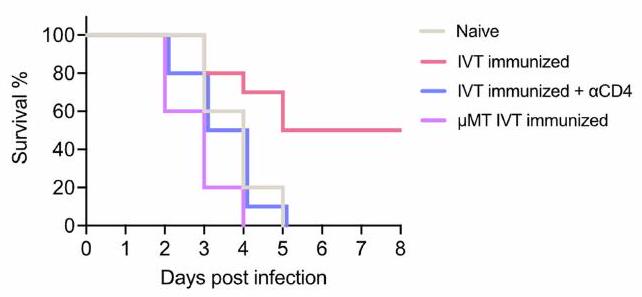
فيروس الهربس البسيط (HSV) لديه القدرة على التأثير على العديد من الأنسجة بما في ذلك الأنسجة العصبية المتعددة مثل العقد الجذرية الظهرية، والعينين، والدماغ – وهو السبب الأكثر شيوعًا لالتهاب الدماغ المميت المتقطع في جميع أنحاء العالم.لفهم المناعة الوقائية ضد فيروس الهربس البسيط (HSV) في الدماغ، قمنا بتطعيم الفئران بفيروس HSV-2 المعطل حرارياً من خلال أربع طرق إعطاء – داخل الصفاق (i.p.) لتحفيز المناعة الجهازية، داخل الجمجمة (i.c.) لتحفيز المناعة المحلية في الدماغ، داخل الغرفة الأمامية (AC)، وداخل الجسم الزجاجي (IVT) – حيث أن الطريقتين الأخيرتين هما مقصورات عينية لها اتصالات تشريحية فريدة مع الجهاز العصبي (الشكل 1a). بعد تحدي مميت بفيروس HSV-2 داخل الجمجمة، توفيت جميع الفئران التي خضعت للتطعيم داخل الصفاق بسبب العدوى، بينما حمت المناعة داخل الجمجمة حواليالفئران (الشكل 1ب)، مما يظهر أن المناعة الجهازية ليست غير كافٍ في توفير الحماية ضد عدوى الجهاز العصبي المركزي. من المRemarkably، أن التطعيم عن طريق الوريد (IVT) حمى أيضًا الفئران، لكن جميع الفئران الملقحة بواسطة AC، مثل الفئران الملقحة عن طريق الحقن داخل الصفاق، لم تنجُ من التحدي (الشكل 1ب)، مما يشير إلى أن التطعيم عن طريق الوريد يحفز استجابة مناعية واقية في الدماغ.
كما هو الحال مع الأعضاء الأخرى، استنتجنا أن الاستجابة المناعية في محور العين-الدماغ يجب أن تكون متوسطة بواسطة العقد اللمفاوية المحلية. للتحقيق في الآلية الخلوية التي تسهل حماية الدماغ بعد التطعيم عن طريق الحقن داخل الجسم الزجاجي، قمنا أولاً بربط الأوعية اللمفاوية للعقد اللمفاوية العنقية العميقة، التي تعتبر حاسمة لاستثارة الاستجابات المناعية في الدماغ. وقد أدى ذلك إلى إلغاء الحماية التي توفرها التطعيم عن طريق الحقن داخل الجسم الزجاجي، مما يدل على أهمية دائرة محلية بين الجهاز العصبي المركزي والعقد اللمفاوية التي تصرف المستضد إلى العقد اللمفاوية العنقية العميقة. بالإضافة إلى ذلك،
الشكل 1: المستضدات في الجزء الخلفي من العين تحفز استجابات مناعية في الدماغ. a، مخطط جدول الإجراءات للتجارب الموصوفة أدناه. ب، تم تحصين فئران C57BL/6J من النوع البري باستخدام حقن HSV-2 المعطل حرارياً من خلال الإدارة عن طريق البطن، داخل الجمجمة، AC و IVT. تم مراقبة البقاء على قيد الحياة بعد التحدي داخل الجمجمة بجرعة قاتلة من HSV-2 بعد 30 يومًا (غير محصن، ; i.p., ; أي.س.، ; مكيف هواء، ; IVT، ). تم ربط dCLNs للفئران باستخدام جهاز الكي. بعد سبعة أيام، تم حقن الفئران عبر الطريق الوريدي مع فيروس HSV-2 المعطل حرارياً. تم مراقبة بقائها بعد التحدي داخل الجمجمة بجرعة مميتة من HSV-2 بعد 30 يومًا (غير محصنة، ; تم تحصين IVT، ; ربط LN، ). د، مخطط نموذج الفأر المتصل والعلاجات المخطط لها. هـ، تم حقن الفئران عبر الطريق الوريدي مع فيروس الهربس البشري من النوع الثاني المعطل حرارياً. بعد أربعة أسابيع، تم ضم الفئران المحصنة إلى الفئران الساذجة. تم تحدي الفئران المحصنة أو الفئران الساذجة عبر الطريق داخل القحف بجرعة قاتلة من فيروس الهربس البشري من النوع الثاني بعد 3 أسابيع، وتم مراقبة بقائها (الساذجة، ; IVT، ; غير المعالج بـ IVT (تحدي IVT)، ; غير المعالج بـ IVT (تحدي غير المعالج)، ; ساذج-ساذج
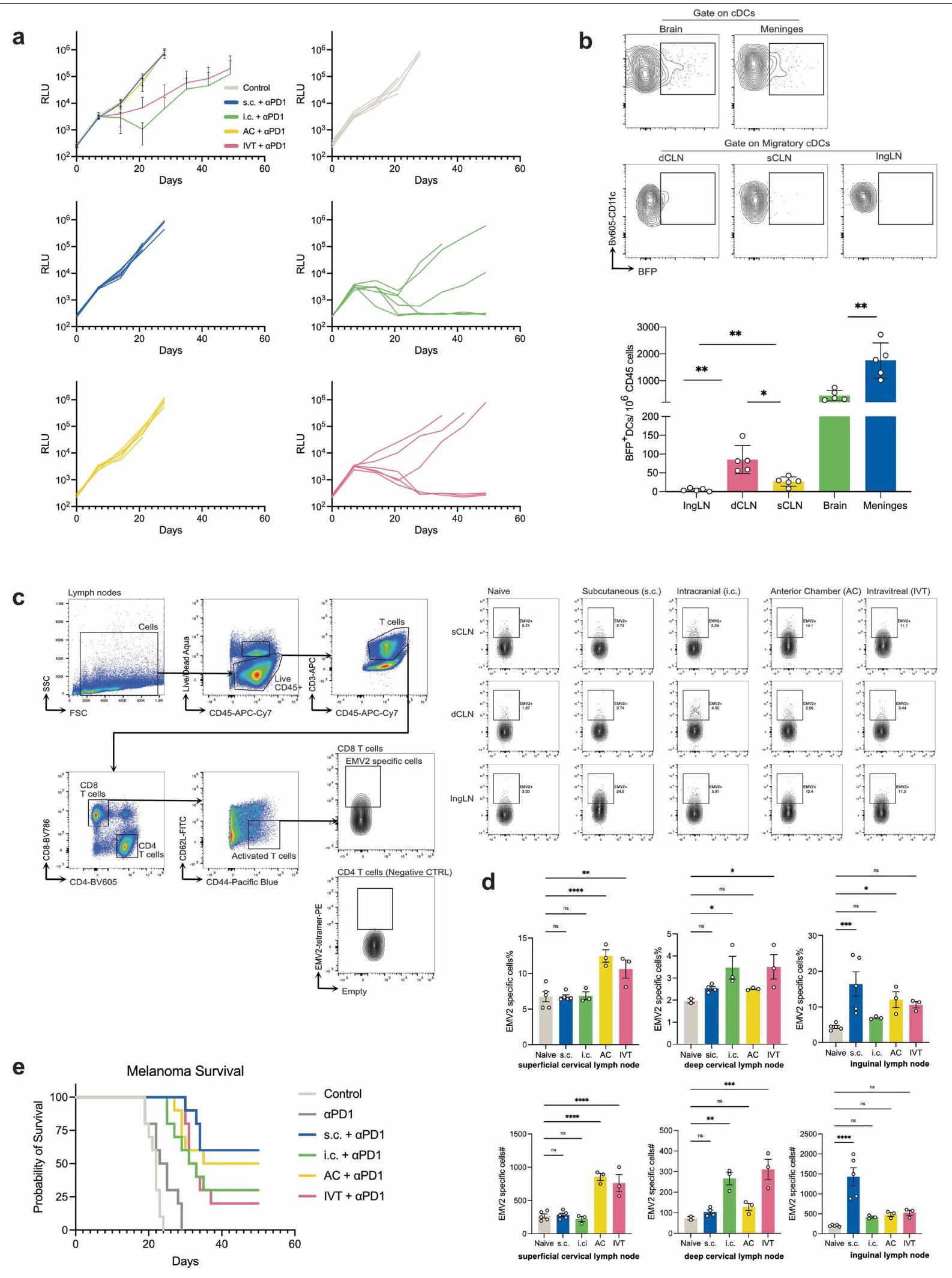
(تحدي ساذج)، ). تم قياس الأجسام المضادة المحددة ضد فيروس الهربس البسيط من النوع 2 بواسطة اختبار الامتزاز المناعي المرتبط بالإنزيم بعد طرق مختلفة من التطعيم ضد فيروس الهربس البسيط 2 (عن طريق الحقن داخل الصفاق،; أي.س.، ; ; IVT، ). تُعرض البيانات كمتوسط تم حقن فئران C57BL/6J من النوع البري بفيروس HSV-1 المعطل حرارياً من خلال الإدارة عن طريق البطن، أو داخل الجمجمة، أو AC أو IVT. وتمت مراقبة بقائهم بعد التحدي داخل الجمجمة بجرعة مميتة من HSV-1 بعد 30 يومًا (غير المعالجة،; i.p.,; أي.س., ; مكيف هواء، ; IVT، ). هـ، كما في أ، ولكن تم استخدام سلالة S. pneumoniae TIGR4 (بريء، ; i.p., ; أي. سي.,; مكيف هواء، ; IVT، تم تلقيح الفئران عن طريق الطريق داخل القحف معخلايا ورم الدماغ التي تعبر عن اللوسيفيراز (GL261-Luc)، تم علاجها بخلايا GL261-Luc المعرضة للإشعاع من خلال الإدارة تحت الجلد، داخل الجمجمة، AC أو IVT (اليوم 7) مع الأجسام المضادة المضادة لـ PD1 (RMP1-14) (الأيام 7 و9 و11) وتمت مراقبتها من أجل البقاء (غير المعالجة،; س.م.، ; أي. سي., ). البيانات تمثل تجربتين مستقلتين. الرسوم البيانية في a و d تم إنشاؤها باستخدام بايو ريندر.كوم. بينما قمنا بربط الأوعية اللمفاوية بعد التحضير ولكن قبل إعادة التحدي، أشارت نتائجنا إلى أن الأوعية اللمفاوية في العقد اللمفاوية الدهنية كانت مطلوبة لإطلاق استجابة تذكارية خلال التحدي (الشكل 1b من البيانات الموسعة). يُعتقد أن الوظائف الفعالة بعد تطعيم فيروس الهربس البسيط تُدار بواسطة الأجسام المضادة المضادة للفيروسات وخلاياوفقًا لهذه التقارير، أظهرت ملاحظاتنا أن CD4خلايا T المنقوصة أو خلايا B الناقصةلم تعد الفئران محمية بعد التطعيم عن طريق IVT (الشكل 1a من البيانات الموسعة). حيث أن التمايز الفعال لخلايا T المساعدة الجريبية وخلايا B في المركز الجرمي مطلوب لإنتاج خلايا بلازما طويلة العمر وذاكرة.الخلايا التي تحفز استجابة مناعية هومورالية متفوقة بعد إعادة التعرض لمستضدقمنا بتحديد تمايز خلايا B المركزية الجرثومية المحددة للمستضد من خلال النقل التبني. خلايا بعد تحصين الفئران بـ NP-OVA من خلال طرق AC أو IVT. على الرغم من أن كلا طريقتي التحصين يمكن أن تحفز بشكل فعال تمايز خلايا B في المركز الجرثومي في العقد اللمفية السطحية (sCLNs)، إلا أن التحصين عن طريق IVT فقط هو الذي يحفز تمايز خلايا B في المركز الجرثومي في العقد اللمفية العميقة (dCLNs) (الشكل البياني الممتد 1d-f). وبالمثل، فإن التحصين عن طريق IVT يزيد بشكل كبير من مستوى CD4 المحدد بمستضد.
تكاثر خلايا T في العقد اللمفاوية الدهنية المقارنة مع التحصين باللقاح AC (الشكل البياني الموسع) ). تشير هذه البيانات إلى أن إنتاج خلايا CD4 المحددة بمستضد محلي تستجيب خلايا T و B في العقد اللمفاوية الدرقية (dCLNs) من خلال التطعيم عن طريق الوريد (IVT) لحماية الجهاز العصبي المركزي المضاد للفيروسات في المضيف.
لمعالجة ما إذا كانت الحماية الناتجة عن التطعيم بواسطة IVT قد تم تطويرها محليًا في الدماغ، قمنا بإنشاء أزواج من الفئران المترافقة بحالات تطعيم مختلفة (الشكل 1d). عندما تم إقران الفئران الملقحة بواسطة IVT مع فئران naive، كانت الحماية موجودة فقط في الفئران التي تم تلقيحها في البداية (يسار) وليس في شركائها المترافقين (يمين) ضد تحدي داخل الجمجمة (الشكل 1e). وبالمثل، كانت مستويات الأجسام المضادة ضد HSV-2 قابلة للاكتشاف فقط في الفئران الملقحة بواسطة IVT، وليس في شركائها المترافقين naive، في السائل الدماغي الشوكي (CSF)، مما يدل على أن التطعيم المباشر ضروري للحصول على الأجسام المضادة ضد HSV-2 في CSF (الشكل 1c من البيانات الموسعة). على النقيض من ذلك، بسبب الدورة الدموية المشتركة، كان لدى الفئران naive التي تم إقرانها مع الفئران الملقحة بواسطة IVT كمية مماثلة من الأجسام المضادة ضد HSV-2 في مصلها (الشكل 1c من البيانات الموسعة). وهذا يشير إلى أن الأجسام المضادة في المصل لا تتوسط الحماية المناعية في الجهاز العصبي المركزي؛ بل تحدث الحماية المناعية.
مقالة
محليًا من خلال السائل الدماغي الشوكي. علاوة على ذلك، بعد فحص توطين الأجسام المضادة المضادة لفيروس الهربس البسيط بعد طرق التطعيم المختلفة، وجدنا أنه على الرغم من أن جميع طرق التطعيم أدت إلى وجود أجسام مضادة مضادة لفيروس الهربس البسيط من النوع 2 في المصل، إلا أن الفئران التي تم تطعيمها عن طريق الحقن داخل الدماغ والحقن داخل السائل الدماغي الشوكي كانت لديها مستويات قابلة للاكتشاف من الأجسام المضادة المضادة لفيروس الهربس البسيط من النوع 2 في السائل الدماغي الشوكي (الشكل 1f). معًا، تُظهر هذه البيانات أن التطعيم عن طريق الحقن داخل السائل الدماغي الشوكي يحقق حماية فريدة للجهاز العصبي المركزي من خلال استجابة محلية تعتمد على الأجسام المضادة.
أخيرًا، لتحديد ما إذا كانت حماية الجهاز العصبي المركزي التي تتوسطها محور العين-الدماغ تمتد إلى ما هو أبعد من فيروس الهربس البسيط من النوع 2، قمنا بفحص فعالية استراتيجية تطعيم مماثلة ضد فيروس الهربس البسيط من النوع 1 والمكورات الرئوية، العامل المسبب الرئيسي لالتهاب السحايا البكتيري.تم تحصين الفئران باستخدام HSV-1 المعطل حرارياً أو S. pneumoniae من خلال طرق الحقن داخل الصفاق (i.p.)، داخل الجمجمة (i.c.)، عبر الأنبوب الهوائي (AC) أو الحقن داخل الوريد (IVT) وتم إعادة تحديها بجرعة قاتلة من نفس الممرض بعد 30 يومًا. مشابهًا للنتائج في نموذج HSV-2، أظهرت البيانات للفئران المحصنة عبر IVT وi.c. أنها تتمتع بحماية شبه كاملة. بالمقابل، جميع الفئران السليمة والمحصة عبر i.p. وAC لم تنجُ من التحدي (الشكل 1g، h). حتى في بيئة علاجية حيث تم إنشاء ورم داخل الجمجمة قبل إعطاء لقاح خلايا السرطان، أدى التحصين عبر طرق IVT أو i.c. إلى حماية متفوقة مقارنة بالتحصين تحت الجلد (s.c.) وAC، الذي أثار فقط مناعة جهازية (الشكل 1i والشكل الإضافي 2a). كانت هذه البيانات متسقة مع النتائج الموصوفة سابقًا التي أظهرت أن التحصين عبر IVT وi.c. أدى إلى نسب وأعداد أكبر بشكل ملحوظ من CD8 المحددة بمستضد.خلايا T مقارنة بتلك التي تتبع التطعيم عن طريق الحقن العضلي والتطعيم تحت الجلد في العقد اللمفاوية الدرقية، ولكن ليس في العقد اللمفاوية السطحية أو العقد اللمفاوية الإربية. (IngLNs؛ الشكل البياني الموسع 2b-d). بالمقابل، في نموذج الميلانوما الجلدية، أظهرت التحصين تحت الجلد حماية أفضل من التحصين داخل الوريد أو التحصين داخل الجمجمة (الشكل البياني الموسع 2e)، مما يشير إلى أن التحصين داخل الوريد والتحصين داخل الجمجمة يحفزان استجابة مناعية موضعية ولكن لا يوفران حماية ضد الأورام المحيطية كما تفعل طريقة التحصين المحلية. معًا، تُظهر هذه البيانات أن الاتصال المناعي بين العين والدماغ ذو صلة محددة وقابل للتطبيق على نطاق واسع للعديد من أمراض الجهاز العصبي المركزي.
للعين نظام تصريف مقسم
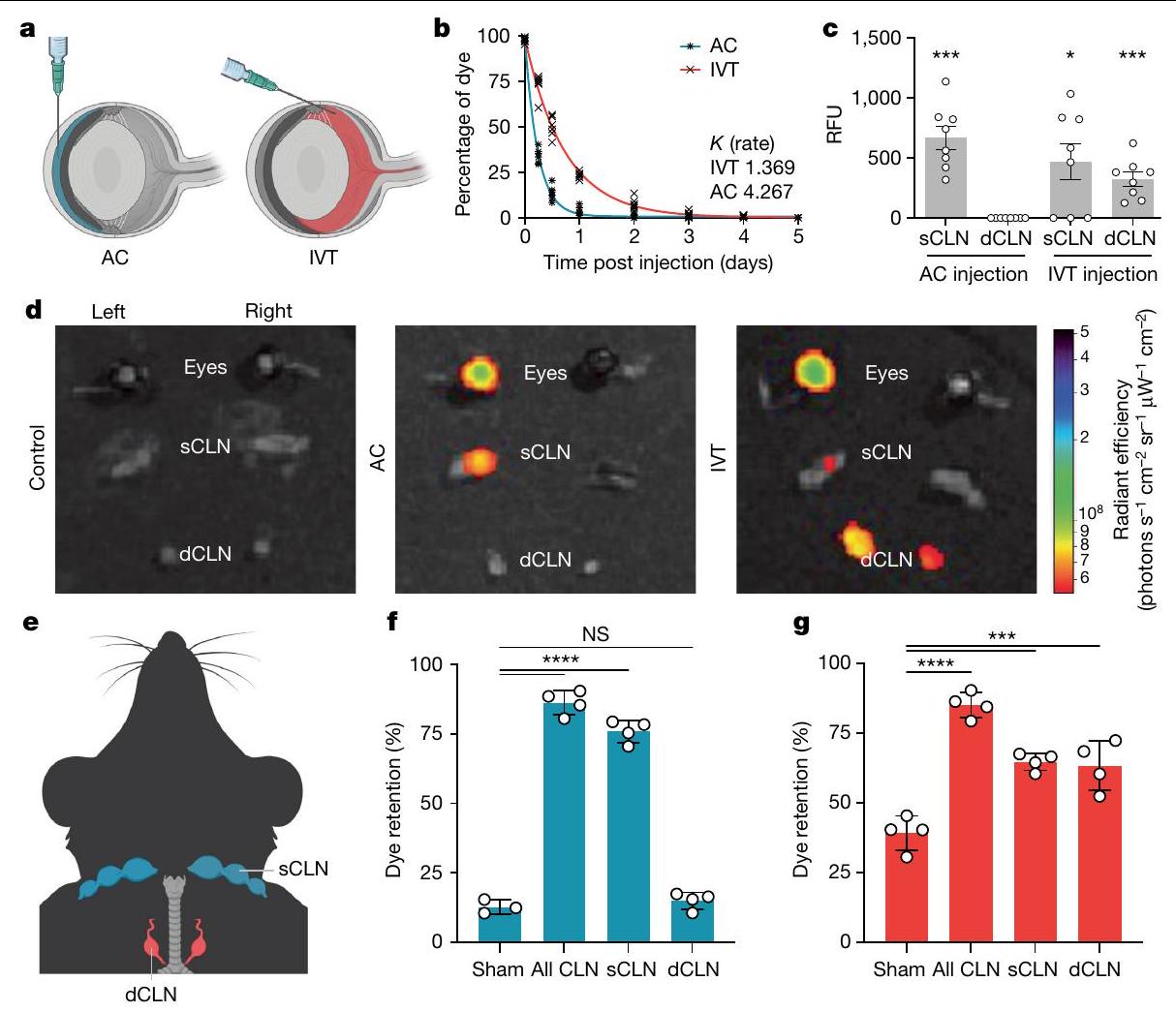
كشفت دراساتنا حول التطعيم عن استجابة مناعية متباينة بين الغرفة الأمامية (AC) والزجاجية في العين. يُعتقد أن سائل العين (الخلط المائي) يتدفق من الغرفة الأمامية عبر مسار التدفق التقليدي الذي يتضمن قناة شليم والشبكة التربيقية.بالإضافة إلى ذلك، تشمل طريقة غير تقليدية تم تقديرها مؤخرًا التدفق من خلال العضلة الهدبية والمساحات فوق الهدبية وفوق المشيمية.على النقيض من ذلك، فإن تصريف السائل من الحجرة العينية الخلفية، وخاصة الزجاجية، غير واضح نظرًا للأوعية اللمفاوية في العين ولا يزال قيد البحث..
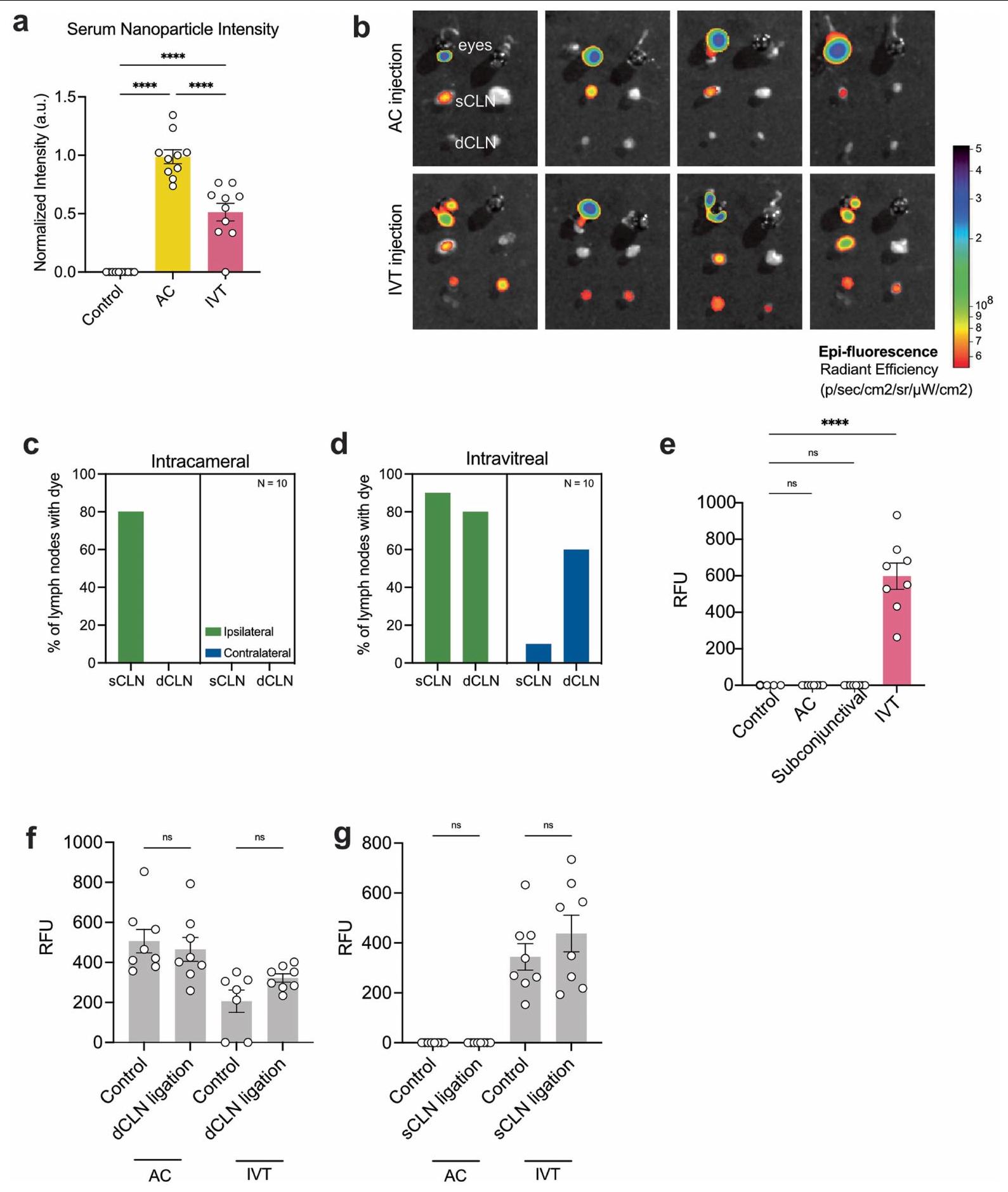
لتحقيق في أنظمة الصرف المختلفة للأقسام الأمامية والخلفية من العين، قمنا بحقن الدكسترانات المعلمة بالفلوريسنت عبر طرق AC أو IVT وقمنا بت quantifying حركيات احتفاظ الدكستران (الشكل 2a). كانت معدل إزالة الدكستران من العين بعد الإدارة عبر IVT (كان أبطأ بشكل ملحوظ من ذلك بعد إدارة AC“; الشكل 2ب). تم اختبار الدكسترانات بأطوال موجية مختلفة من الفلورية وأوزان جزيئية مختلفة للتأكد من أن هذه الظاهرة ليست محددة بصبغة معينة. افترضنا أن هذه الملاحظة يمكن تفسيرها بإمكانية واحدة من اثنتين؛ إما أن تكون للأقسام طرق تصريف فريدة، أو أن الخطوة المحددة لمعدل تصريف كل قسم مختلفة (أي أن الانتشار عبر الزجاجي قد يكون أبطأ خطوة). افترضنا أن هذه التفسيرات المحتملة يمكن توضيحها من خلال مراقبة توطين الصبغة في الجسم الحي. باستخدام طريقة تم وصفها سابقًاأكدنا أنه يمكن الكشف عن الصبغة التي تم حقنها في الحجرة الأمامية (AC) وحقنها داخل الجسم الزجاجي (IVT) في المصل. ومع ذلك، تم الكشف عن تركيزات أعلى بكثير من الصبغة في الدم بعد حقن AC مقارنةً بعد حقن IVT (الشكل 3a من البيانات الموسعة). بينما كانت الصبغة التي تم حقنها في AC محصورة في العقد اللمفاوية السطحية الجانبية المقابلة، كما تم الإبلاغ عنه سابقًا.صبغة تم حقنها عن طريق IVT تم تحديدها في العقد اللمفاوية الدرقية الثنائية بالإضافة إلى العقد اللمفاوية السفلية من نفس الجانب
الشكل البياني الممتد 3b-d). كما لوحظ بعد حقن السائل الأمينوسي، لا يتم تصريف الصبغة إلى العقد اللمفاوية الدهنية بعد الحقن تحت الملتحمة (الشكل البياني الممتد 3e)، مما يشير إلى أن الاختراق إلى الجزء الخلفي من العين أو الزجاجي مطلوب للتصريف إلى العقد اللمفاوية الدهنية. لاختبار متطلبات هذه الهياكل اللمفاوية المحلية لتصريف السائل المائي والزجاجي، قمنا بربط العقد اللمفاوية السطحية، العقد اللمفاوية الدهنية أو كليهما جراحيًا. أولاً، أكدنا أن ربط العقد اللمفاوية الدهنية لم يؤدي إلى زيادة التدفق إلى العقد اللمفاوية السطحية والعكس صحيح (الشكل البياني الممتد 3f,g). أدى ربط العقد اللمفاوية السطحية، ولكن ليس العقد اللمفاوية الدهنية، إلى زيادة كبيرة في احتباس الصبغة في العين بعد حقن السائل الأمينوسي (الشكل 2f)، مما يتماشى مع الملاحظة السابقة. بالنسبة لحقن الزجاجي، أدى ربط العقد اللمفاوية السطحية أو العقد اللمفاوية الدهنية بمفردها إلى زيادة احتباس الصبغة في العين (الشكل 2g). معًا، تُظهر هذه النتائج نظام تصريف مقسم في العين بين السائل الأمينوسي والسائل الزجاجي مع مسار محدد من السائل الزجاجي إلى العقد اللمفاوية الدهنية.
غمد العصب البصري يحتوي على لمفاويات وظيفية
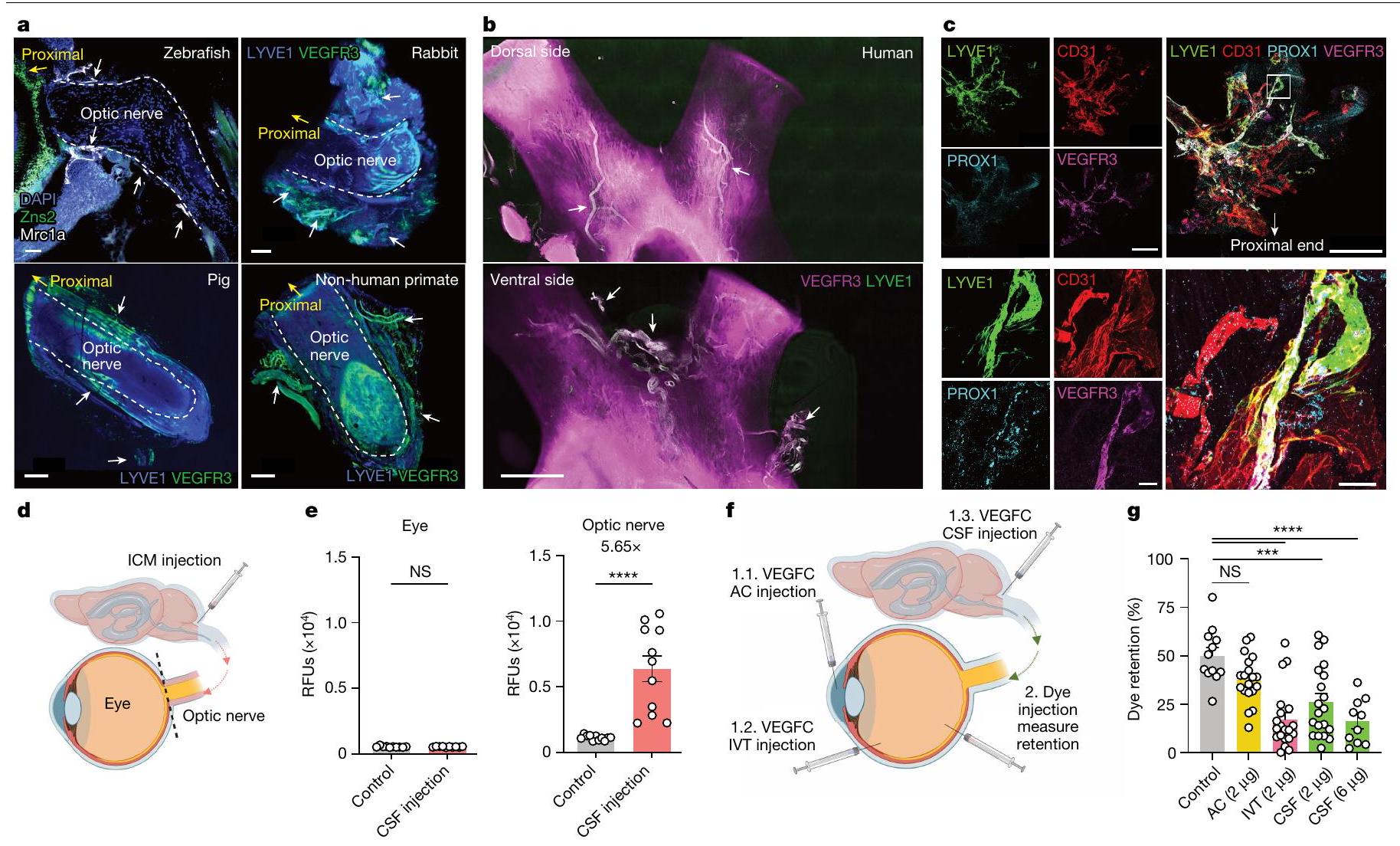
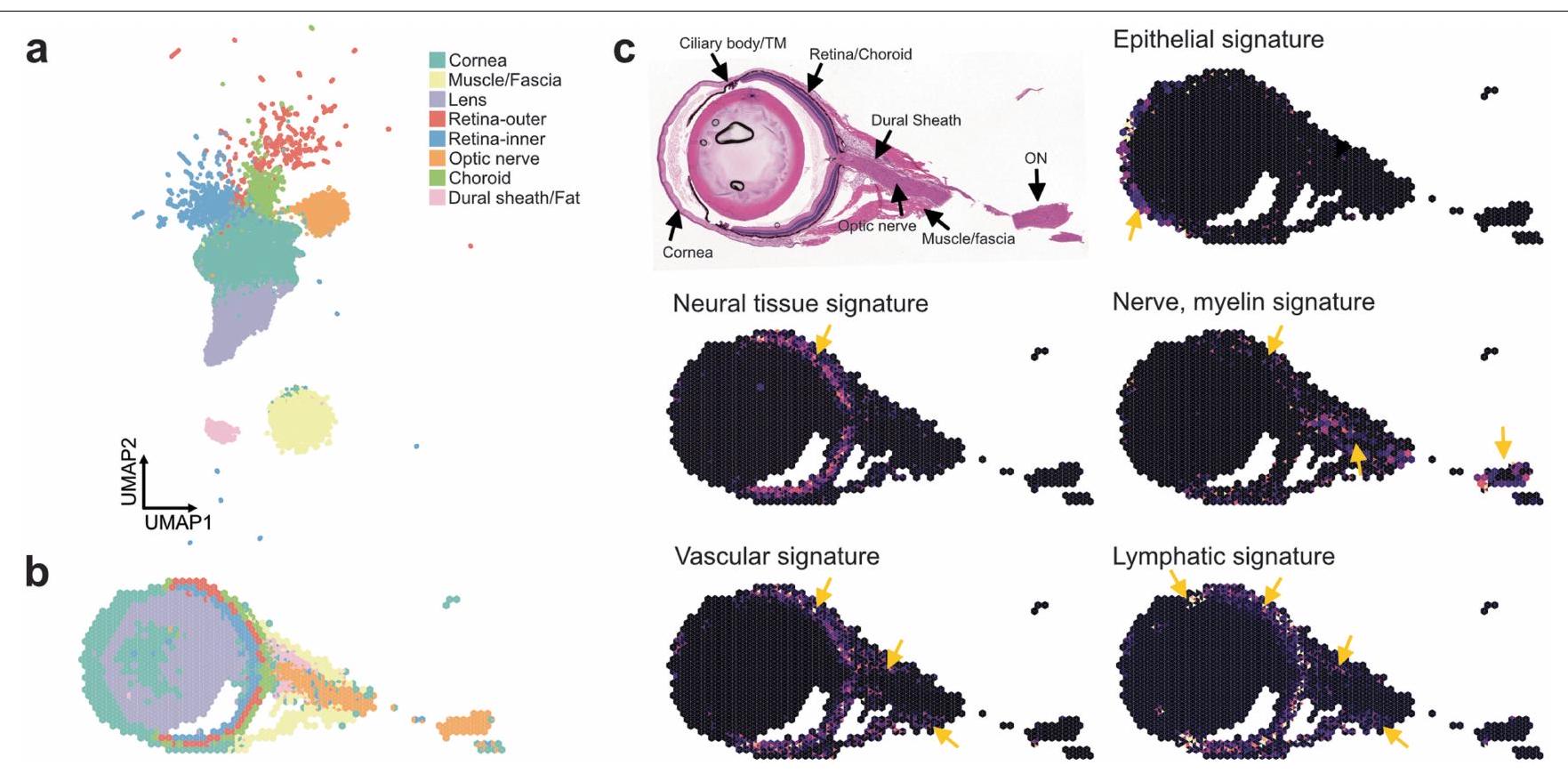
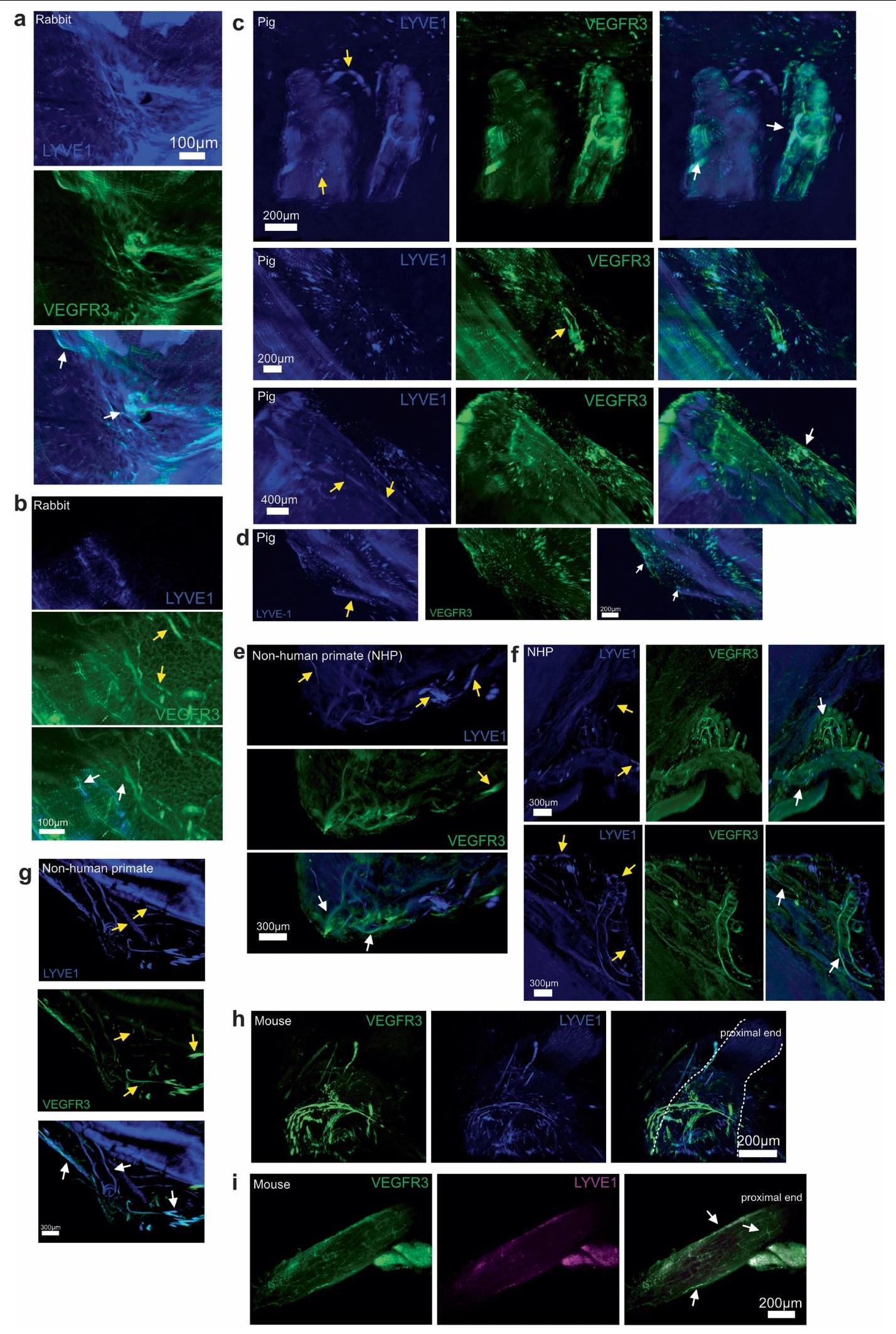
تشير موضع الجزيئات الكبيرة في العقد اللمفاوية المتميزة إلى أن شبكة الأوعية اللمفاوية ربما تكون وسيلة لتصريف كل من هذه الأقسام. على الرغم من أن دراسة حديثة وصفت وجود نظام غليمفاتي عيني في الفئران، مشابه لذلك الموجود في الدماغ.لم يتم حل شبكة وظيفية من الأوعية اللمفاوية التي تتوسط تصريف الزجاجي. لذلك، استخدمنا نهجًا ترنسكريبتوميًا جنبًا إلى جنب مع التصوير ثلاثي الأبعاد المدعوم بتوسيم المناعية للأعضاء المنظفة بالمذيب (iDISCO).للحصول على رؤية شاملة لشبكة الأوعية اللمفاوية المرتبطة بالعصب البصري في أنواع الفقاريات المختلفة. ساعدت تقنيات النسخ الجيني المكاني في جمع رؤية على مستوى العضو لتوقيعات جينات الأوعية اللمفاوية الشبيهة في العين والعصب البصري. بالإضافة إلى الشبكة التربيقية.والشبكية، التي وُجد أنها تحتوي على البلعميات التي تعبر عن بعض توقيعات الجينات الشبيهة باللمفاويةوجدنا أن غمد العصب البصري يحتوي أيضًا على توقيعات جينية تشبه الأوعية اللمفاوية (الشكل التمديدي 4). بالتوازي، كشفت تقنية iDISCO عن وجود أوعية لمفاوية تغطي أعصاب البصر في الأسماك الزعنفية، والفئران، والأرانب، والخنازير، والرئيسيات غير البشرية، والبشر (الشكل 3 أ، ب والشكل التمديدي 5)، مما يدل على سمة محفوظة تطوريًا للجهاز اللمفاوي. في جميع الثدييات، رأينا هياكل وعائية مع تلوين متزامن لـ LYVE1 ومستقبل عامل نمو بطانة الأوعية الدموية 3 (VEGFR3؛ الأسهم البيضاء في الشكل التمديدي 5). كما رأينا أوعية دموية ملونة لأحدهما وليس الآخر، مما يشير إلى احتمال وجود أوعية لمفاوية جمع سلبية لـ LYVE1.أو البلعميات التي تعبر عن LYVE1 (الأسهم الصفراء في الشكل 5 من البيانات الموسعة). ومن الجدير بالذكر أن الأوعية اللمفاوية كانت محصورة في الأغشية المحيطة بالعصب، وليس في العصب نفسه، بما يتماشى مع ما يُلاحظ في الدماغ والأغشية السحائية الجافية..
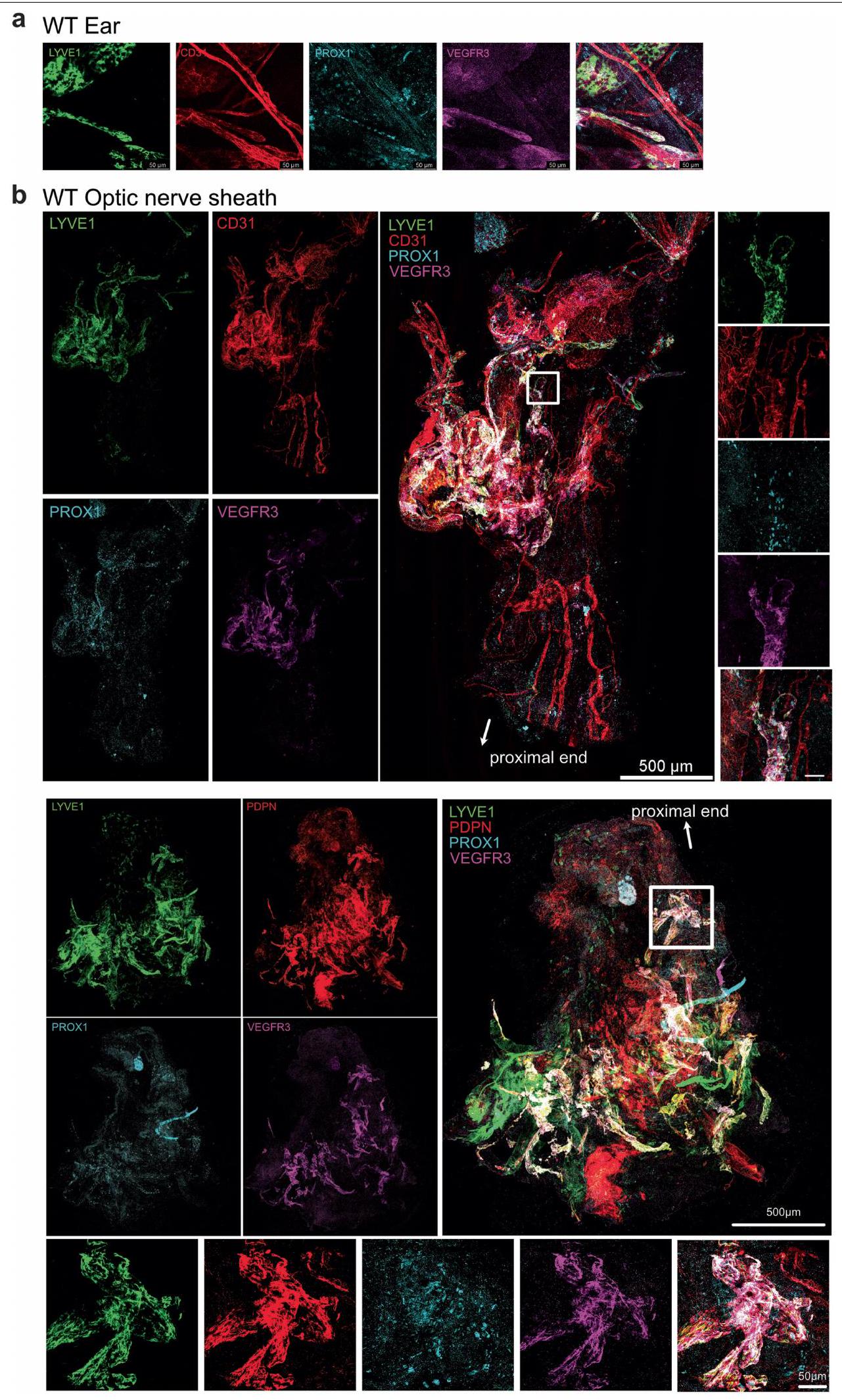
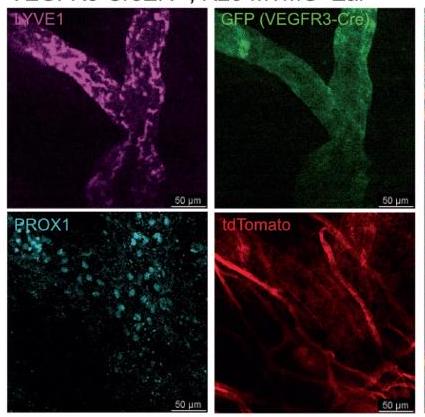
لتحسين حساسية الصبغات المطبقة، قمنا بتشريح غلاف العصب البصري بالكامل من العصب البصري. دون تثبيت التروية في الفئران، انكمش غلاف العصب البصري بعد الاستئصال، مما أدى إلى تجمع الأوعية اللمفاوية نحو الطرف القريب من العصب (الشكل التمديدي 5h). لمعالجة ذلك، قمنا بإجراء تثبيت التروية قبل الاستئصال، مما حافظ على تشريح الشبكة اللمفاوية على طول العصب (الشكل التمديدي 5i). باستخدام نظام الأوعية اللمفاوية في الأذن المعروف جيدًا كتحكم (الشكل التمديدي 6a)، قمنا بالتحقق من وجود الأوعية اللمفاوية في غلاف العصب البصري باستخدام علامات إضافية (VEGFR3، PROX1، LYVE1، CD31 وpodoplanin؛ الشكل 3c والشكل التمديدي 6b). لتوصيف هذه الأوعية بشكل أكبر باستخدام نهج وراثي، استخدمنا نوعين مختلفين من الفئران المبلغة مما يتيح صبغًا محددًا للغاية للأوعية اللمفاوية: VEGFR3-CreER.;فئران R26-mTmG، و PROX1-CreER;CDH5-Dre;R26-STOP-mCherry الفئران (الشكل البياني الموسع 7). معًا، أكدت هذه الطرق المتوازية أن الأوعية اللمفاوية موجودة بالفعل في غمد العصب البصري.
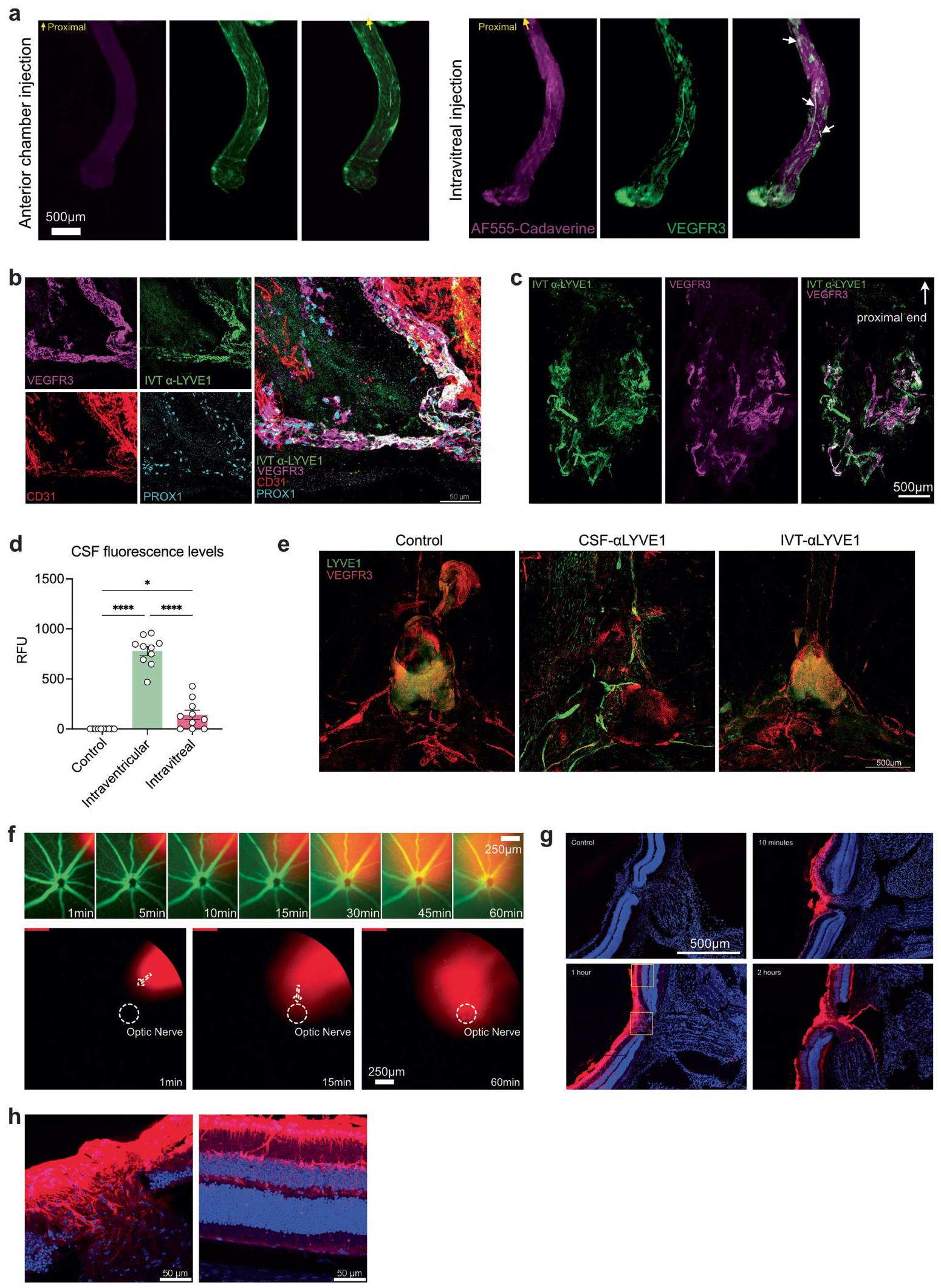
ثم سعينا لتقييم وظيفة هذه الأوعية اللمفاوية. بعد حقن الوريد، ولكن ليس حقن الغرفة الأمامية، لمؤشر فلوري، تم تصور المؤشر في VEGFR3.الأوعية الدموية على طول العصب البصري
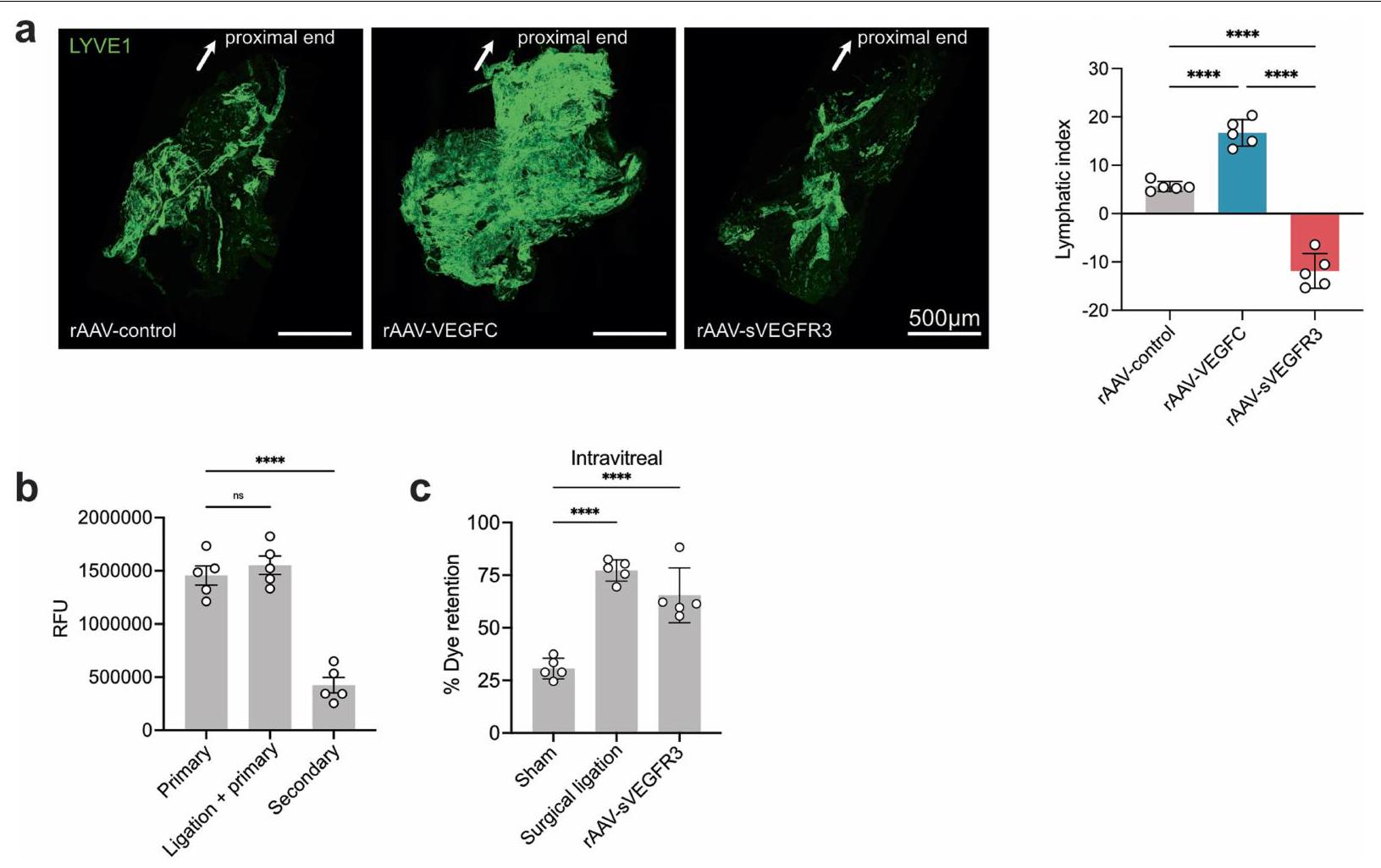
الشكل 2 | تمتلك العينان نظام تصريف لمفاوي مقسم. أ. مخطط حقن الصبغة عبر الغرفة الأمامية (AC) والطريق داخل الزجاجي (IVT). ب. تم حقن فئران C57BL/6J بالصبغة عبر الطريق AC أو IVT. تم تحليل نسبة احتفاظ الصبغة في العين من 6 ساعات إلى اليوم الخامس بعد الحقن (AC،; IVT، ). تم حقن الصبغة في العين من خلال الطريق الأمامي للغرفة (AC) أو الطريق الزجاجي (IVT). تم جمع قياسات العقد اللمفاوية السطحية والعميقة (SCLNs و dCLNs) باستخدام جهاز قراءة الفلورسنت بعد ساعة من الحقن. RFU، وحدة الفلورسنت النسبية (AC، ; IVT، ). تُعرض البيانات كمتوسط س.م., AC sCLN; , IVT sCLN; تم حقن الصبغة في العين اليسرى من خلال الطريق الأمامي للغرفة (AC) أو الطريق داخل الزجاجي (IVT)، وتم جمع العينين والعقد اللمفاوية السطحية والعميقة (sCLNs و dCLNs) للتصوير بالإضاءة الفلورية باستخدام نظام IVIS.
تظهر خرائط الحرارة الممثلة بعد طرح الخلفية للصبغة في العين، والعقد اللمفاوية السطحية (sCLNs) والعقد اللمفاوية العميقة (dCLNs) بعد ساعة من الحقن. e، مخطط للمواقع التشريحية للعقد اللمفاوية السطحية والعميقة. f، g، تم ربط العقد اللمفاوية السطحية، أو العميقة، أو كلاهما جراحيًا. بعد يومين، تم حقن الصبغة في العين عبر الطريق الأمامي (f) أو الطريق داخل الزجاجي (g). تم قياس نسبة احتفاظ الصبغة في العين بعد 12 ساعة. تُعرض البيانات كمتوسط.س.م. في و (شام AC، في جميع الظروف الأخرى)., IVT sCLN; IVT dCLN؛ NS، غير مهم.تم حساب القيم باستخدام تحليل التباين أحادي الاتجاه (ANOVA) مع اختبار المقارنات المتعددة (Dunnett). تم إنشاء الرسوم البيانية في a و e باستخدامبايو ريندر.كوم. (الشكل البياني الممتد 8a). بشكل متسق، يمكن للأجسام المضادة LYVE1 التي تم حقنها عن طريق IVT أن تميز بشكل فعال الأوعية اللمفاوية في غمد العصب البصري (الشكل البياني الممتد 8b، c)، مما يشير إلى أن الجزيئات الكبيرة مثل البروتينات يمكن أن تخترق بحرية هذه الأوعية اللمفاوية من الزجاجية. كانت فرضيتنا الرئيسية حول كيفية انتقال الجزيئات إلى هذه الأوعية اللمفاوية مبنية على فهم الأوعية اللمفاوية السحائية في الدماغ.لقد استنتجنا أن الجزيئات من العين تمر عبر السائل الدماغي الشوكي إلى اللمفاويات المحيطة بالعصب البصري، مشابهة لكيفية انتقال الجزيئات من الدماغ إلى اللمفاويات السحائية. لاختبار ذلك، قمنا بفحص توزيع الصبغة والأجسام المضادة بعد الحقن داخل الزجاجية. لقد اكتشفنا كمية ضئيلة من الصبغة في السائل الدماغي الشوكي بعد الحقن داخل الزجاجية مقارنة بعد الحقن داخل البطين الدماغي (الشكل التمديدي 8d). علاوة على ذلك، لم تتمكن الأجسام المضادة التي تم حقنها عن طريق الحقن داخل الزجاجية من وسم اللمفاويات السحائية بشكل كافٍ. تشير هذه البيانات إلى أن الصبغة المحقونة داخل الزجاجية ربما دخلت الفضاء المحلي للسائل الدماغي الشوكي حول العصب البصري ولم تصرف إلى العقد اللمفاوية الدرقية من خلال السحايا الدماغية التقليدية التي تم وصفها سابقًا (الشكل التمديدي 8e). لتحديد كيفية وصول الجزيئات الكبيرة إلى الأوعية اللمفاوية حول غمد العصب البصري، تتبعنا كيفية مغادرة الجزيئات المحقونة في الجسم الزجاجي للعين في الجسم الحي. بعد حقن AF647-OVA داخل الزجاجية، لاحظنا أن هناك تدفقًا اتجاهيًا نحو رأس العصب البصري (الشكل التمديدي 8f)، وهو ما يتماشى مع النتائج السابقة التي تشير إلى أن فرق الضغط بين الفضاءات داخل العين والفضاء داخل القحف يدفع تدفق السوائل العينية عبر العصب البصري.أظهر الفحص بدقة أعلى بعد قطع العين أن المؤشر كان مركزًا بشكل كبير في رأس العصب البصري (الشكل 8g من البيانات الموسعة)، مع بعض الانتشار عبر الشبكيات. أظهر الانتشار عبر الشبكية نمطًا مشابهًا لأنماط الغليمفاتيك الموصوفة سابقًا في الشبكيات، وهو شريط عمودي بين الخلايا عبر الشبكية. (الشكل 8h من البيانات الموسعة).
لإثبات قدرة هذه الأوعية اللمفاوية على تصريف الزجاجية، قمنا بإيصال VEGFC، وهو محفز لمفاوي.من خلال ثلاثة مسارات مقسمة مختلفة: AC (يستهدف اللمفاويات في الجزء الأمامي من العين)، IVT (يستهدف الجزء الخلفي من العين، العصب البصري وربما الجزء الأمامي من العين) أو الحقن داخل الفضاء الجافية (ICM؛ يستهدف العصب البصري). أولاً، باستخدام علامات فلورية، أكدنا أن العلامات التي تم حقنها في ICM كانت لها وصول فقط إلى العصب البصري وليس إلى العين. (الشكل 3د، هـ). بعد يومين من توصيل VEGFC، تم حقن الدكسترانات الفلورية عن طريق الطريق الوريدي داخل الزجاجي، وتم تحليل الاحتفاظ بعد 12 ساعة (الشكل 3و). لم تؤثر إدارة VEGFC عن طريق الحجرة الأمامية على تصريف الدكستران من الزجاجي، مما يشير إلى أن تحفيز اللمفاويات الأمامية غير كافٍ لدفع تصريف الزجاجي. على العكس من ذلك، زادت إدارة VEGFC عن طريق الوريد داخل الزجاجي وICM من تصريف الدكستران من الزجاجي، والأخيرة كانت بطريقة تعتمد على الجرعة (الشكل 3ز)، مما يدعم الملاحظة بأن الحجرة الخلفية للعين تصرف إلى العقد اللمفاوية القريبة من العمود الفقري من خلال الأوعية اللمفاوية على غمد العصب البصري. لتأكيد دور VEGFC في تعديل الأوعية اللمفاوية لغمد العصب البصري، قمنا أيضًا بتوصيل فيروس مرتبط بالأدينوفيروس (rAAV)-VEGFC أو VEGFR3 القابل للذوبان (sVEGFR3) عبر الطريق الوريدي داخل الزجاجي ووجدنا أن التعبير عن VEGFC أو sVEGFR3 أدى إلى زيادات أو انخفاضات كبيرة في
الشكل 3|تصريف الأوعية اللمفاوية لغمد العصب البصري في الجزء الخلفي من العين. a,b, تلوين المناعية لمقاطع من العصب البصري من سمك الزرد مع الأوعية اللمفاوية المعلّمة لـ Mrc1a (الأسهم البيضاء؛ أ، أعلى اليسار) وتلوين المناعية iDISCO للأوعية اللمفاوية السحائية للأرنب (أ، أعلى اليمين)، والخنزير (أ، أسفل اليسار)، والقرود غير البشرية (أ، أسفل اليمين) والإنسان (ب) العصب البصري والتقاطع، مع الأوعية اللمفاوية التي تظهر التوضع المشترك لـ LYVE1 و VEGFR3 (الأسهم البيضاء). DAPI، 4’،6-diamidino-2-phenylindole. ج، أغلفة العصب البصري من الفئران البرية من النوع البري الملوّنة لـ LYVE1 و CD31 و PROX1 و VEGFR3. الصور في الأسفل تظهر عرضًا أكبر تكبيرًا للمنطقة المميزة في الصورة المدمجة في أعلى اليمين. قضبان القياس، (أ، أعلى اليسار؛ ج، أسفل مدمج)، (أ، أعلى اليمين؛ ج، أعلى مدمج)، ، أسفل اليسار واليمين) و. رسم تخطيطي لحقن ICM وتشريح العين والعصب البصري. تم حقن الصبغة ICM، وتم قياس شدة إشارة الفلورسنت بعد ساعة واحدة في العين والعصب البصري (تحكم،حقن السائل الدماغي الشوكي ). **** .f، مخطط لطرق الحقن لـتم حقن VEGFC من خلال طريق الإدارة AC أو IVT أو ICM. بعد يومين، تم حقن صبغة عن طريق IVT في العين، وتم قياس نسبة احتفاظ الصبغة بعد 12 ساعة من حقن الصبغة (التحكم،; مكيف هواء، ; IVT، ; السائل الدماغي الشوكي ( ) ، ; السائل الدماغي الشوكي ( ) ، ) تُعرض البيانات كمتوسطس.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع اختبار المقارنات المتعددة (دونيت) أو اختبار الطالب غير المقترن ذو الجانبين.-اختبار. تم إنشاء الرسومات في د، ف باستخدامبايو ريندر.كوم. تظهر هذه البيانات وجود شبكة من الأوعية اللمفاوية الوظيفية على غمد العصب البصري التي تربط الجزء الخلفي من العين بمجموعة من العقد اللمفاوية الفريدة من نوعها عن بقية العين ولكنها مشتركة مع بقية الجهاز العصبي المركزي.
تعزيز العلاج الجيني من خلال اللمفاويات العينية
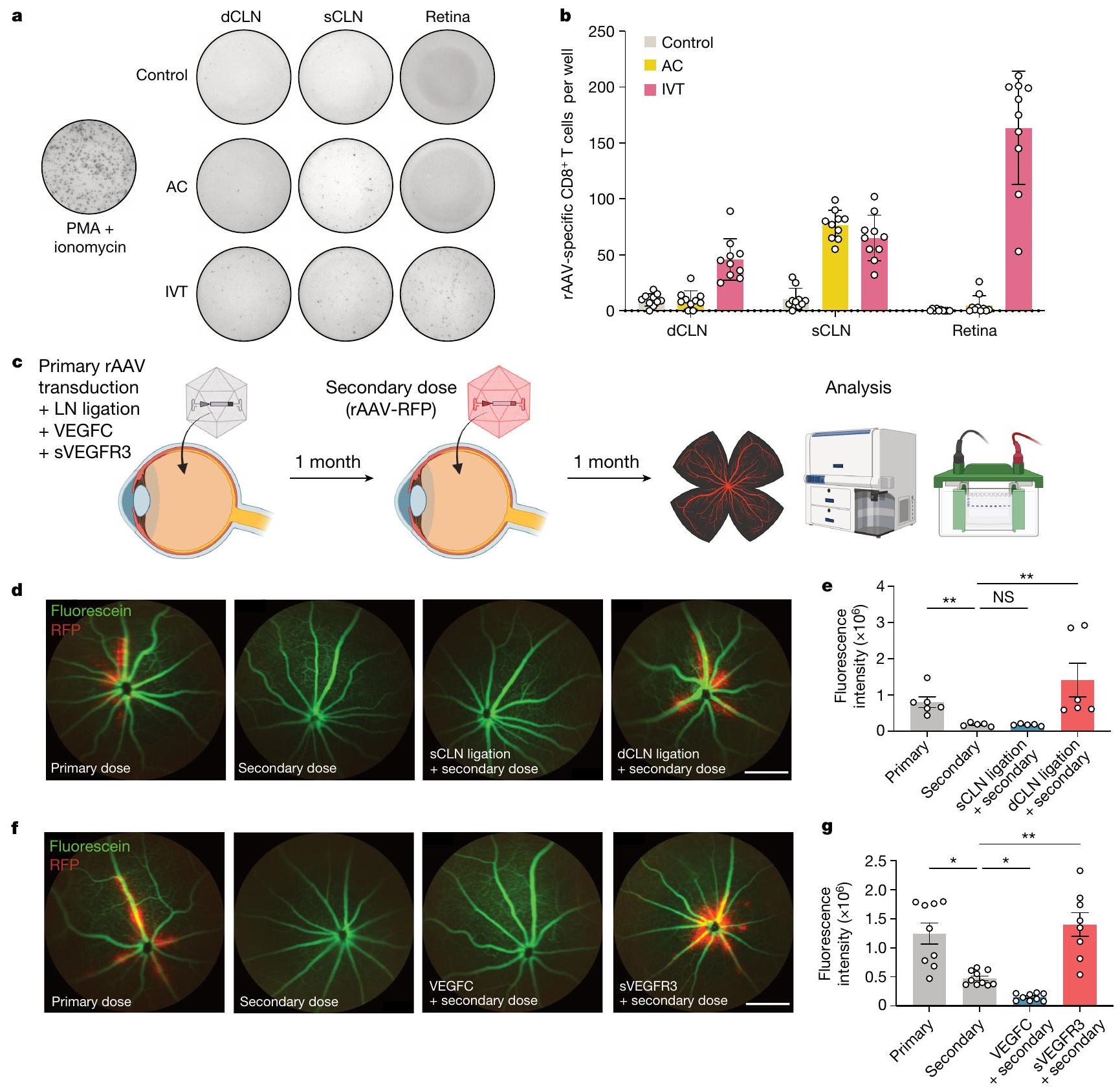
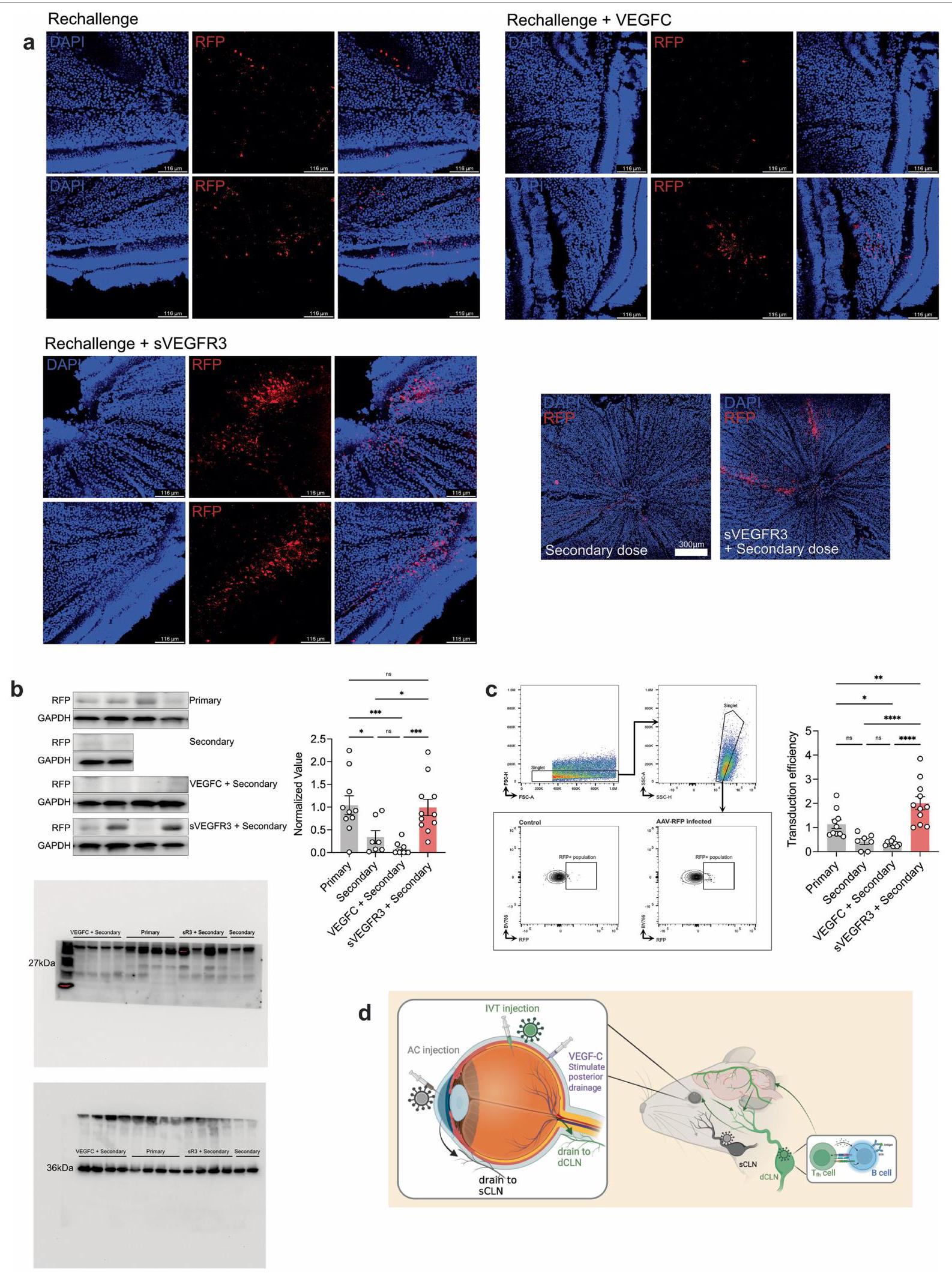
بعد تأكيد الأهمية المناعية لهذا التصريف العيني الخلفي، قمنا بعد ذلك بالتحقيق في التطبيقات العلاجية المحتملة التي تستفيد من هذه النتيجة. تعتبر الفيروسات المعاد هندستها (rAAVs) الأكثر شيوعًا كوسائط توصيل الجينات في الجسم بسبب قدرتها على النقل بشكل آمن وفعال.. بالإضافة إلى ذلك، كان استخدام AAV2 كناقل لعلاج المرضى الذين يعانون من ضمور الشبكية الوراثي المرتبط بـ RPE65 هو أول علاج جيني تم الموافقة عليه من قبل إدارة الغذاء والدواء الأمريكية. ومع ذلك، على الرغم من خصائصه المناعية المعدلة، فإن المناعة المكتسبة ضد rAAV تحد من فعالية إدارته المتكررة. قمنا أولاً بتوصيف CD8 المحدد لـ rAAVتحفيز خلايا T في العقد اللمفاوية والشبكية بعد إعطاء الرAAV عن طريق الحقن داخل الجسم الزجاجي والحقن تحت الجلد باستخدام اختبار ELISpot. على الرغم من أن كل من الحقن تحت الجلد والحقن داخل الجسم الزجاجي أدى إلى تحفيز كمية مماثلة من خلايا CD8 الخاصة بالرAAV.الخلايا التائية في العقد اللمفاوية السطحية، حقن IVT أدى بشكل خاص إلى تحفيز المزيد من خلايا CD8 المحددة لـ rAAVالخلايا التائية في العقد اللمفاوية الدرقية والشبكية، التي لم تُلاحظ بعد حقن AC (الشكل 4 أ، ب). وقد استنتجنا أن تصريف اللمف على يساهم العصب البصري في الاستجابة المناعية لـ rAAV، خاصةً بالنسبة للنهج العلاجي القائم على الشبكية، مما يحد من فائدته. قمنا بدراسة ما إذا كان استئصال الأوعية اللمفاوية العينية يمكن أن يمكّن من فعالية علاجية مطولة بعد حقن rAAV المتكرر في العين. لاختبار ذلك، قمنا أولاً بإعطاء الجرعة الأولية من rAAV، وبعد شهر، أعطينا جرعة ثانية من rAAV المعبر عن RFP (rAAV-RFP). كتحكم أساسي، تم إعطاء الفئران السليمة rAAV-RFP في وقت الجرعة الثانية (الشكل 4c). مشابهًا للنتائج التي توصلت إليها التقارير السابقة ومتسقًا مع ملاحظتنا بشأن المناعة المحلية في الجهاز العصبي المركزي، كان مستوى تعبير RFP بعد العدوى الثانوية أقل بكثير من ذلك بعد العدوى الأولية (الشكل 4d، e). تم تأكيد أهمية الأوعية اللمفاوية العينية من خلال إجراء تجربة مماثلة بعد ربط sCLN أو dCLN، والتي أظهرت أن ربط dCLN فقط كان كافيًا في تخفيف الاستجابة المناعية لـ rAAV واستعادة كفاءة نقل rAAV-RFP الثانوي (الشكل 4d، e). بالإضافة إلى ذلك، أكدنا أن إجراء الربط لم يؤثر على التحدي الأولي (الشكل 9b من البيانات الموسعة). نظرًا لأن ربط العقد اللمفاوية جراحيًا لن يكون خيارًا علاجيًا قابلاً للتطبيق، استخدمنا طريقة بديلة لتثبيط النشاط الجزيئي للأوعية اللمفاوية: الإدارة الوريدية لـ sVEGFR3، والتي وجدنا أنها تثبط تصريف اللمف بنفس القدر تقريبًا مثل الربط الجراحي (الشكل 9c من البيانات الموسعة). رأينا أن الاستجابة المناعية لـ rAAV تفاقمت أكثر بوجود VEGFC، والذي هو
الشكل 4 | تثبيط اللمف يمكن من إعطاء rAAV المتكرر. أ، ب، تم حقن الفئران بـ rAAV-RFP عبر الطريق الوريدي أو الطريق البصري. تم جمع العقد اللمفاوية الدهنية والعقد اللمفاوية السطحية والشبكية بعد 10 أيام، وتم قياس الاستجابات المناعية المحددة لـ rAAV باستخدام اختبار ELISpot. من أجلتم حقن فئران C57BL/6J بـ rAAV عن طريق الوريد، وبعد شهر، تم إعادة تحديها بـ rAAV-RFP. تم تحليل كفاءة نقل rAAV-RFP من خلال التصوير بعد شهر. ج، مخطط لخطط التجارب. د، تصوير قاع العين الفلوري في الجسم الحي لتصور الأوعية (باللون الأخضر) ونقل RFP (باللون الأحمر) في ظروف ربط العقد اللمفاوية المختلفة. مقياس الرسم،. e، قياس شدة RFP من د (أساسي، ; ثانوي، ; ربط sCLN، ; ربط dCLN، ).
معروف بزيادة الاستجابات المناعية من خلال تحفيز تصريف اللمف. على العكس من ذلك، فإن تثبيط إشارات VEGFC خلال النقل الأولي لـ rAAV باستخدام sVEGFR3 سمح للجرعة الثانية بأن تكون فعالة تمامًا مثل العدوى الأولية (الشكل 4f، g والشكل الإضافي 10a-c). تؤكد هذه التجارب على دور اللمفاويات العينية الخلفية في تعديل فعالية العلاجات الجينية المعتمدة على rAAV وتقترح منصة لتجاوز الحواجز الحالية التي تحد من تكرار العلاج الجيني. ، أولية مقابل ثانوية؛ ربط الثانوي مقابل ربط dCLN. التصوير الفلوري في vivo لشبكية العين لتصور الأوعية (باللون الأخضر) وانتقال RFP (باللون الأحمر) مع إضافة VEGFC أو sVEGFR3. شريط القياس،. تحديد شدة RFP من (أساسي، ; ثانوي، ; VEGFC، ; sVEGFR ). ، الأولية مقابل الثانوية؛، الثانوي مقابل VEGFC؛ ، الثانوي مقابل sVEGFR3.تم حساب القيم باستخدام تحليل التباين الأحادي مع اختبار المقارنات المتعددة (دونيت). تُعرض البيانات كمتوسطس.م في ب، هـ وج. الرسومات في ج تم إنشاؤها بـبايو ريندر.كوم.
نقاش
لقد أظهرنا أن الجزء الخلفي من العين يحتوي على نظام تصريف لمفاوي فريد يتفاعل تشريحياً مع شبكة اللمف السحائي للجهاز العصبي المركزي عند العقد اللمفاوية العميقة. وقد وصفت الدراسات السابقة وجود شعيرات لمفاوية نادرة على أعصاب البصر لدى البشر والرئيسيات غير البشرية.؛ باستخدام النسخ الجيني المكاني، وتقنيات التصوير المتعددة، والتلاعب الجيني في الجسم الحي وفي تجارب حقن المتعقب، قمنا برسم شبكة الأوعية اللمفاوية الأكبر في غمد العصب البصري ووصفنا وظيفتها في تصريف الزجاجية إلى العقد اللمفاوية العميقة. هذه النتائج تتماشى مع العلاقة المعروفة بين العين والجهاز العصبي المركزي.وتوسيع هذا المفهوم من خلال إظهار آلية مراقبة مناعية مشتركة بين الموقعين (الشكل البياني الممتد 10d).
في السابق، كان يُعتبر كل من العين والدماغ، اللذان يفتقران إلى الأوعية اللمفاوية التقليدية، أنسجة محمية من المناعة لأن زراعة الأنسجة الأجنبية يمكن أن تبقى على قيد الحياة بعد الزرع.. علاوة على ذلك، تم الإبلاغ عن أن المستضدات التي يتم توصيلها إلى كل من الغرفة الأمامية والغرفة الخلفية للعين تحفز التسامح المناعي المحيطي. ومع ذلك، كشفت دراستنا أن المستضدات الممرضة التي يتم توصيلها إلى الجزء الخلفي من العين من خلال التطعيم عن طريق الحقن داخل الجسم الزجاجي يمكن أن تصرف إلى العقد اللمفاوية القريبة من الدماغ وتبدأ استجابة مناعية محلية واقية في الدماغ تعتمد على الخلايا، خلايا B والأجسام المضادة المحلية في حجرة الجهاز العصبي المركزي. على الرغم من أن استخدام نماذج مرضية مختلفة وتدخلات علاجية في دراساتنا يظهر عالمية نتائجنا، فمن المحتمل أن تؤدي مسببات الأمراض أو الأمراض المختلفة إلى انحراف نحو أذرع معينة من الاستجابة المناعية التكيفية. من الجدير بالذكر، في تجاربنا وتجارب الآخرين، ليس أن العقد اللمفاوية السطحية تفتقر إلى المستضدات من الجهاز العصبي المركزي؛ بل إن التصريف إلى العقد اللمفاوية السطحية غير كافٍ لإطلاق استجابة مناعية في الجهاز العصبي المركزي، وقد تكشف الدراسات المستقبلية التي تكتشف الفرق الأساسي بين هذين النوعين من العقد اللمفاوية عن رؤى أفضل حول الملاحظات الموصوفة هنا. إحدى الاحتمالات هي أن العقد اللمفاوية السطحية والعقد اللمفاوية العميقة قد تختلف في قدرتها على إنتاج خلايا T فعالة تتجه نحو الدماغ أو خلايا T مقيمة في الدماغ، والتي يُبلغ عنها بأنها توفر الحماية ضد إعادة العدوى بالمسببات في الدماغ.بالإضافة إلى الدور الوقائي للأجسام المضادة المحلية في تطعيم IVT، سيكون من المهم التحقيق فيما إذا كانت خلايا T المقيمة في الدماغ تلعب دورًا أثناء التحدي.
بالإضافة إلى ذلك، لا تزال الآليات التي توجه هذه الاستجابات المناعية نحو الدماغ بحاجة إلى مزيد من التحقيق. تشير النتائج الحالية إلى أن التسامح المناعي تجاه المستضدات الأجنبية في الجزء الخلفي من العين والدماغ مدفوع بقمع المناعة، سواء كان سلبياً أو إيجابياً، وليس بسبب نقص في الاستجابة المناعية. وبالتالي، قد يلعب نظام التصريف الخلفي للعين دوراً محورياً في تعديل تنشيط المناعة وقمعها لمختلف الأمراض التي تشمل الجهاز العصبي المركزي. الطرق التقليدية الحالية لعلاج أو تشخيص أمراض الجهاز العصبي المركزي إما أنها تدخلية أو تفشل في بدء المناعة المحلية. في الحالات التي يُشتبه فيها بزيادة تأثير الكتلة في الجهاز العصبي المركزي بسبب العدوى أو الأورام، قد يكون من الخطير غالباً الوصول إلى السائل الدماغي الشوكي من خلال طرق مثل البزل القطني. على الرغم من أنها بعيدة عن التطبيق السريري، تقترح دراساتنا الزجاجية كمسار غير تدخلي نسبياً وأكثر وصولاً لاستثارة مناعة الجهاز العصبي المركزي.
نستخدم هذه المفاهيم لتوضيح أن تعطيل الأوعية اللمفاوية الخلفية يمكن أن يلغي الاستجابة المناعية لـ rAAV المقدم إلى الجزء الخلفي من العين وبالتالي يستعيد الفائدة العلاجية. على عكس فكرة الامتياز المناعي، تواجه علاج rAAV للأمراض العينية عقبات كبيرة بسبب الاستجابة المناعية الخلوية والمصلية التي يبدأها.تؤكد هذه النتائج بشكل أكبر أن نظام تصريف العصب البصري الذي تم تحديده في دراستنا هو المسؤول عن النتيجة المناعية استجابةً للمستضدات في الجسم الزجاجي. كما تشير النتائج إلى أن نظام تصريف اللمف الخلفي للعين هو هدف متعدد الاستخدامات وسهل الوصول له للتلاعب بالاستجابات المناعية في العديد من السياقات المختلفة.
بعيدًا عن الآثار المناعية لنظام التصريف الخلفي، فإن كيفية استغلال ميكانيكا مسار التصريف الخلفي لعلاج الأمراض العينية تستحق مزيدًا من الدراسة. على وجه التحديد، فإن تحديد نظام تصريف لمفاوي خلفي للعين يوفر مسار تصريف فعال محتمل لتعديل ضغط العين الداخلي أو إزالة السوائل غير المرغوب فيها في حالات المرض مثل الزرق أو الوذمة البقعية، على التوالي. أيضًا، مع وجود نظام الغليمفاتي الذي تم وصفه مؤخرًا في العين.يتطلب الأمر المزيد من العمل لتوضيح كيفية تفاعل هذه المسارات مع بعضها البعض لتوفير التوازن في العين، كما هو الحال في الدماغ. معًا، تثبت نتائج الدراسة الحالية أن العلاقة بين العين الخلفية والدماغ ليست فقط علاقة عصبية، بل هي أيضًا دائرة مناعية مشتركة تنسق المناعة من المحيط إلى الجهاز العصبي المركزي.
لماذا تطور هذا النظام المعقد من تصريف اللمف في العين يطرح سؤالًا مثيرًا. إحدى الاحتمالات هي أن مركزية الجهاز العصبي المركزي وظهور الشبكات اللمفاوية في الفقاريات قد دفعا إلى تمييز نظامي تصريف اللمف في العين. قد تكون هناك إشارة إلى ذلك في التوجه الجانبي الذي نلاحظه أثناء التصريف إلى العقد اللمفاوية السطحية ولكن ليس إلى العقد اللمفاوية العميقة، وهو ما يعد على الأرجح نتيجة للفصل التشريحي بين الجلد والعضلات والأنسجة المحيطة بالمدار. على العكس من ذلك، يحدث التصريف الثنائي إلى العقد اللمفاوية العميقة من الجزء الخلفي للعين من خلال شبكة من الأوعية اللمفاوية التي تصرف السائل الدماغي الشوكي. تعمل الأوعية اللمفاوية الأمامية بالتزامن مع القرنية والملتحمة، وهي سطح مخاطي عام يتعرض باستمرار للغزاة الخارجيين الذين قد لا يصلون أبدًا إلى الدماغ. الأوعية اللمفاوية للعصب البصري التي نصفها هنا تربط الجزء الخلفي للعين (الشبكية) بالدماغ، مما يسمح باستجابات مناعية منسقة قد تهيئ الدماغ للتهديدات الوشيكة. قد يسبق فصل استجابات المناعة في العين إلى الشبكات اللمفاوية الأمامية والخلفية الثدييات، وقد تساعد الدراسات المستقبلية في الفقاريات الأخرى في توضيح متى ولماذا ظهرت هذه الابتكار لأول مرة.
المحتوى عبر الإنترنت
أي طرق، مراجع إضافية، ملخصات تقارير Nature Portfolio، بيانات المصدر، بيانات موسعة، معلومات تكميلية، شكر وتقدير، معلومات مراجعة الأقران؛ تفاصيل مساهمات المؤلفين والمصالح المتنافسة؛ وبيانات توفر البيانات والرموز متاحة علىhttps://doi.org/10.1038/s41586-024-07130-8.
لندن، أ.، بنهار، إ. وشوارز، م. الشبكية كنافذة إلى الدماغ – من أبحاث العين إلى اضطرابات الجهاز العصبي المركزي. نات. ريف. نيورول. 9، 44-53 (2013).
ماركوتشي، م. إ. وآخرون. فيروس الهربس البسيط-1 في الدماغ: الجانب المظلم لعدوى خفية. اتجاهات الميكروبيولوجيا. 28، 808-820 (2020).
إيجيمَا، ن. وإيوَاسَاكِي، أ. وصول الأجسام المضادة الفيروسية الواقية إلى الأنسجة العصبية يتطلب مساعدة خلايا T من نوع CD4. ناتشر 533، 552-556 (2016).
فيكتورا، ج. د. ونوسنزاويغ، م. س. المراكز الجرثومية. مراجعة سنوية للمناعة. 40، 413-442 (2022).
سيستر، ج. ج. وآلن، س. د. استجابات خلايا ب: ديناميات تفاعل الخلايا والقرارات. خلية 177، 524-540 (2019).
شih، T.-A. Y.، رويدرر، M. و نوسنزاويغ، M. C. دور تقارب مستقبلات المستضد في استجابات الأجسام المضادة المستقلة عن الخلايا التائية في الجسم الحي. نات. إيمونول. 3، 399-406 (2002).
كويدل، يو.، شيلد، و. م. وبيفستر، هـ.-و. مسببات المرض وعلم وظائف الأعضاء المرضية لالتهاب السحايا بالمكورات الرئوية. لانسيت للأمراض المعدية 2، 721-736 (2002).
إيوينو، ف.، سينيين، ج.، هنريكس-نورمارك، ب. وفان ديل، ج. م. كيف يغزو المكورات الرئوية الدماغ؟ اتجاهات الميكروبيولوجيا. 24، 307-315 (2016).
أغنية، إ. وآخرون. تصريف اللمف المدفوع بـ VEGF-C يمكّن المراقبة المناعية للأورام الدماغية. الطبيعة 577، 689-694 (2020).
تومسون، ب. ر. وآخرون. عيب لمفاوي يسبب ارتفاع ضغط العين والزرق في الفئران. ج. التحقيقات السريرية 124، 4320-4324 (2014).
جونستون، م.، مكلاين، ج. و أوفربي، د. ر. تدفق سائل العين غير التقليدي: مراجعة. أبحاث العين التجريبية 158، 94-111 (2017).
وانغ، إكس. وآخرون. نظام تصريف غليمفاتيكي عيني يزيل بيتا-أميلويد من عين القوارض. علوم. ترانسلات. ميد. 12، eaaw3210 (2020).
كاسي، أ.، ليو، ج.، فائق، م. أ. و تشان، ك. س. التصوير الغليمفاتي وتعديل العصب البصري. أبحاث تجديد الأعصاب 17، 937-947 (2022).
تيتجن، ج. ت.، ديريتو، ج.، بوبير، ج. س. وسالتزمان، و. م. قياسات كمية تعتمد على المجهر لتركيز الجسيمات النانوية المتداولة باستخدام أحجام دم ميكرولتر. النانوميديسين 13، 1863-1867 (2017).
كاميلو، س.، كيزيتش، ج.، شانلي، أ.، ريجبي، ب. ومكمانامين، ب. ج. المستضد من الغرفة الأمامية للعين ينتقل في شكل قابل للذوبان إلى الأعضاء اللمفاوية الثانوية عبر الطرق اللمفاوية والوعائية. تحقيقات في طب العيون وعلم الرؤية 47، 1039-1046 (2006).
تام، أ. ل. س.، غوبتا، ن.، زانغ، ز. و يوجيل، ي. هـ. النقاط الكمومية تتبع تصريف اللمف من عين الفأر. تكنولوجيا النانو 22، 425101 (2011).
لي، ج. ي. وآخرون. التأكيد الهيكلي لتدفق اللمف من الفقاعات تحت الملتحمة في البشر الأحياء. علوم العيون 1، 100080 (2021).
يوسل، ي. هـ. وآخرون. تصريف لمفاوي نشط من العين تم قياسه بواسطة تصوير صوتي ضوئي غير جراحي لجزيئات نانوية قريبة من الأشعة تحت الحمراء. تحقيقات في طب العيون وعلوم الرؤية 59، 2699-2707 (2018).
رينير، ن. وآخرون. iDISCO: طريقة بسيطة وسريعة لوضع علامات مناعية على عينات الأنسجة الكبيرة للتصوير الحجمي. خلية 159، 896-910 (2014).
شو، هـ.، تشين، م.، ريد، د. م. وفورستر، ج. ف. توجد البلعميات الإيجابية لـ LYVE-1 في عيون الفئران الطبيعية. تحقيقات في طب العيون وعلوم الرؤية 48، 2162-2171 (2007).
ماكينن، ت. وآخرون. موقع تفاعل PDZ في الإيفرين B2 مطلوب لإعادة تشكيل الأوعية اللمفاوية. جينات التطور. 19، 397-410 (2005).
لوفيو، أ. وآخرون. الميزات الهيكلية والوظيفية للأوعية اللمفاوية في الجهاز العصبي المركزي. ناتشر 523، 337-341 (2015).
أسبيلوند، أ. وآخرون. نظام وعائي لمفاوي دورالي يصرف السائل بين الخلايا الدماغية والجزيئات الكبيرة. ج. التجارب الطبية. 212، 991-999 (2015).
بريسلاين، ج. و. وآخرون. عامل نمو البطانية الوعائية-C يحفز مضخة اللمف من خلال آلية تعتمد على مستقبل VEGF-3. المجلة الأمريكية لعلم وظائف الأعضاء. قلب. علم وظائف الأعضاء. 293، H709-H718 (2007).
جيلتش، م. وآخرون. تضخم الأوعية اللمفاوية في الفئران المعدلة وراثياً VEGF-C. ساينس 276، 1423-1425 (1997).
ماثيو، إ. وآخرون. دليل على دخول السائل الدماغي الشوكي إلى العصب البصري عبر مسار غليمفاتي. تحقيقات في طب العيون ورؤية العلوم 58، 4784-4791 (2017).
راسل، س. وآخرون. فعالية وسلامة فورتجين نيبارفوك (AAV2-hRPE65v2) في المرضى الذين يعانون من اعتلال الشبكية الوراثي الناتج عن RPE65: تجربة عشوائية، محكومة، مفتوحة، من المرحلة 3. لانسيت 390، 849-860 (2017).
كوليلا، ب.، رونزيتّي، ج. ومينغوزي، ف. قضايا ناشئة في العلاج الجيني داخل الجسم المعتمد على AAV. طرق العلاج الجزيئي. طرق التطوير السريرية 8، 87-104 (2018).
مينغوزي، ف. وهاي، ك. أ. الاستجابات المناعية تجاه ناقلات AAV: التغلب على الحواجز لتحقيق علاج جيني ناجح. بلود 122، 23-36 (2013).
شيرمان، د. د. وآخرون. تحديد الأوعية اللمفاوية المدارية: دراسات مجهرية ضوئية وإنزيمية ومجهرية إلكترونية. جراحة تجميلية وإصلاحية للعيون. 9، 153-169 (1993).
سترايلين، ج. و. امتياز المناعة العينية: فرص علاجية من تجربة طبيعية. نات. ريف. إيمونول. 3، 879-889 (2003).
مدوار، ب. ب. المناعة تجاه الجلد المزروع المتجانس؛ مصير زراعة الجلد المتجانس المزروعة في الدماغ، وفي الأنسجة تحت الجلد، وفي الغرفة الأمامية للعين. المجلة البريطانية لعلم الأمراض التجريبي 29، 58-69 (1948).
كاميلو، س.، شانلي، أ.، فوان، أ. س. ب. & مكمانامين، ب. ج. توزيع المستضد في الأنسجة اللمفاوية بعد حقنه في الغرفة الأمامية لعيني الجرذ. ج. المناعة. 172، 5388 (2004).
بونمان، ز. ف. هـ. م. وآخرون. مضاد الورم داخل العين يتدفق بشكل محدد إلى العقد اللمفاوية تحت الفك السفلي، مما يؤدي إلى رد فعل غير مكتمل للخلايا التائية السامة. ج. المناعة. 172، 1567 (2004).
سونودا، ك.-هـ. وآخرون. تحليل التحمل الجهازي الناتج عن حقن المستضد في تجويف الزجاجية: الانحراف المناعي المرتبط بتجويف الزجاجية. المناعة 116، 390-399 (2005).
أياصوفي، ك. وآخرون. خلايا الذاكرة التائية المقيمة في الدماغ تتوسع بسرعة وتبدأ استجابات التهاب الأعصاب بعد الإصابة الفيروسية في الجهاز العصبي المركزي. سلوك الدماغ. المناعة. 112، 51-76 (2023).
شتاينباخ، ك. وآخرون. تمثل خلايا الذاكرة التائية المقيمة في الدماغ حاجزًا ذاتيًا مضادًا للفيروسات. ج. تجريب. ميد. 213، 1571-1587 (2016).
ملاحظة الناشر: تظل شركة سبرينجر ناتشر محايدة فيما يتعلق بالمطالبات القضائية في الخرائط المنشورة والانتماءات المؤسسية.
الوصول المفتوح هذه المقالة مرخصة بموجب رخصة المشاع الإبداعي النسب 4.0 الدولية، التي تسمح بالاستخدام والمشاركة والتكيف والتوزيع وإعادة الإنتاج بأي وسيلة أو صيغة، طالما أنك تعطي الائتمان المناسب للمؤلفين الأصليين والمصدر، وتوفر رابطًا لرخصة المشاع الإبداعي، وتوضح إذا ما تم إجراء تغييرات. الصور أو المواد الأخرى من طرف ثالث في هذه المقالة مشمولة في رخصة المشاع الإبداعي الخاصة بالمقالة، ما لم يُشار إلى خلاف ذلك في سطر الائتمان للمواد. إذا لم تكن المادة مشمولة في رخصة المشاع الإبداعي الخاصة بالمقالة وكان استخدامك المقصود غير مسموح به بموجب اللوائح القانونية أو يتجاوز الاستخدام المسموح به، فستحتاج إلى الحصول على إذن مباشرة من صاحب حقوق الطبع والنشر. لعرض نسخة من هذه الرخصة، قم بزيارةhttp://creativecommons.org/licenses/by/4.0/. (ج) المؤلف(ون) 2024
طرق
فئران
خلايا مختلطة من الذكور والإناث تتراوح أعمارها بين ستة إلى عشرة أسابيع من سلالة C57BL/6J، B6.Cg-Tg(TcraTcrb)425Cbn/J (OT-II)، B6.129P2(C)-Ightm2Cgn/J (B1-8) و B6.129S2-IghtmICgn/J (تم شراء الفئران من مختبر جاكسون وشارلز ريفر، وتم تربيتها وإيواؤها لاحقًا في جامعة ييل. PROX1-CreER; CDH5-Dre; R26-STOP-mCherry و VEGFR3-CreERكانت فئران R26-mTmG هدايا من مختبر J.-L.T. جميع الإجراءات المستخدمة في هذه الدراسة تتوافق مع الإرشادات الفيدرالية وسياسات المؤسسة الخاصة بلجنة رعاية واستخدام الحيوانات في كلية الطب بجامعة ييل. تم تعيين الحيوانات المتطابقة في العمر والجنس عشوائيًا إلى مجموعات التحكم أو العلاج في كل تجربة. لم تُستخدم طرق إحصائية لتحديد أحجام العينات مسبقًا. تم تحديد أحجام العينات تجريبيًا بناءً على دراسات منشورة سابقًا ولضمان قوة إحصائية كافية. لم يكن الباحثون معزولين عن المجموعات التجريبية، حيث لم تكن القياسات ذات طابع ذاتي.
الخلايا
تم الحصول على خلايا GL261-Luc كهدية من J. Zhou (جراحة الأعصاب في جامعة ييل) وتم زراعتها في RPMI مضاف إليه 10% FBS و1% بنسلين-ستربتوميسين و1% صوديوم بايروفات. تم الحصول على خلايا CT2A-BFP كهدية من T. Mathivet (مركز الأبحاث القلبية الوعائية في باريس). تم الحصول على خلايا B16 كهدية من N. Palm (علم المناعة في جامعة ييل). كانت الخلايا سلبية لتلوث الميكوبلازما.
البكتيريا والفيروسات
سلالة HSV-1 KOS وسلالات HSV-2 186synتي كيهو 186synكانت هدايا من د. كنيب (كلية الطب بجامعة هارفارد). تم تكاثر هذه الفيروسات وتقدير قوتها على خلايا فيرو (ATCC CCL-81) كما تم وصفه سابقًا.كانت خلايا فيرو سلبية لتلوث الميكوبلازما. تم زراعة S. pneumoniae (ATCC 6303) على أطباق أجار دم الأغنام بنسبة 10% (BD Biosciences) طوال الليل فيثم تم نقل هذه المستعمرات إلى وسط تود هيويت ونمت طوال الليل. تم عد عدد البكتيريا باستخدام الكثافة الضوئية عند 600 نانومتر.
أنسجة العصب البصري والتألق المناعي الخلوي
Tg(mrc1a:eGFP)ذكور وإناث سمك زيبرا بالغين بعمر سنة واحدةتم توفيرها من قبل ب. و. وينشتاين في المعاهد الوطنية للصحة الأمريكية (بيثيسدا، ماريلاند). تم قتل أسماك الزرد بجرعة زائدة من MS-222 وتم استئصال الرؤوس بالكامل وثبتت في محلول 60 مللي مول من HEPES مع 4% PFA (درجة الحموضة 7.4) طوال الليل في درجة حرارة الغرفة. ثم تم إزالة الكالسيوم من العينات فيمحلول EDTA (رقم الهيدروجين 7.4) لمدة 5 أيام في درجة حرارة الغرفة. ثم تم تجميد الرؤوس بسرعة في OCT وتم الحصول على مقاطع سميكة بالتبريد. تم صبغ المقاطع التي تحتوي على العصب البصري بالأجسام المضادة الأولية الفأرية المضادة لزنبق الماء Zns2 (ZIRC، ZDB-ATB-081002-34؛ 1:50) والدجاج المضاد لـ GFP (Aves، رقم الكتالوج GFP-1010؛ 1:500) تليها الأجسام المضادة الثانوية المترافقة مع AF488 المربوطة بجلد الماعز المضاد للفأر IgG (Jackson Immuno 115-545146) و Cy5 المربوطة بدونكي المضاد للدجاج IgY (Jackson Immuno 703-175-155) مع DAPI. ) للتلوين النووي. تم الحصول على الصور باستخدام جهاز Zeiss LSM.
تم جمع عيون من الأرانب والخنازير بعد الوفاة من حيوانات تم euthanized بسبب حالات صحية غير مرتبطة. تم إزالة العيون خلال ساعة من الوفاة وثُبتت في محلول.فورمالين معزول محايد. تم جمع عيون سليمة من اثنتين من قردة الريسوس الإناث (Macaca mulatta)، تتراوح أعمارهن بين 25-28 عامًا وتم euthanized بسبب حالات صحية غير مرتبطة بعد الوفاة. تم الحصول على الأعصاب البصرية البشرية والتقاطعات من خلال خدمات أنسجة علم الأمراض في جامعة ييل من خلال خدمة الحصول على الأنسجة والتوزيع. تم كتابة وإقرار إجراء تشغيل قياسي مصغر لهذه العينات. تمت إزالة العيون خلال ساعة من الوفاة وثُبتت في فورمالين معزول محايد بنسبة 10%. تم قطع الأعصاب البصرية مع الأغشية السحائية ومعالجتها من خلال بروتوكول iDISCO أدناه.
تم تشريح أغشية العصب البصري للفأر بعد تخدير الفئران وإجراء غسيل قلبي باستخدام محلول PBS البارد وتم استخدام PFA (Sigma-Aldrich) بشكل متسلسل. تم قطع رأس الفئران أولاً، وتم كشف الجمجمة عن طريق قطع الجلد وفروة الرأس. تم إجراء قطع في منتصف الجمجمة وقطع عرضية على كلا جانبي الجمجمة. تم استخدام ملاقط لخلع وإزالة نصفي الجمجمة لكشف الدماغ. تم رفع الدماغ من الجزء الخلفي، وتم قطع الأوعية الدموية التي تربط الدماغ بأجزاء أخرى من الرأس حتى يمكن رفع الدماغ بما يكفي لكشف المسار البصري. تم إزالة نسيج الدماغ بعد القطع مباشرة أمام تقاطع العصب البصري. تم إزالة سقف القناة البصرية خارج الجمجمة عن طريق إزالة أجزاء من الجمجمة فوق العينين لكشف الأجزاء داخل القناة والقناة من العصب البصري. تم تشريح النسيج المحيط بالعصب البصري بعناية لتحرير العصب، وتم قطع العين عند رأس العصب البصري. بعد إزالة العصب البصري، تم قطع غلاف العصب البصري على طول طول العصب وإزالته للتلوين والتصوير الكامل.
PROX1-CreER;CDH5-Dre;R26-STOP-mCherry و VEGFR3CreERتم حقن فئران R26-mTmG عبر الطريق البطنيتاموكسيفين; سيغما-ألدريتش، T5648) لمدة 5 أيام متتالية وتم جمع أغشية العصب البصري بعد يومين كما هو موضح أعلاه. ثم تم تثبيت أغشية العصب البصري في 1% PFA لمدة ساعة واحدة ومعالجتها على الفور في محلول حجب (10% مصل حيواني طبيعي، 1% ألبومين مصل بقري، 0.3% PBS-Triton X-100) لمدة ساعة واحدة في درجة حرارة الغرفة. للكشف عن الأوعية اللمفاوية، تم حضانة العينات مع الأجسام المضادة الأولية طوال الليل في ، ثم تم غسلها خمس مرات في درجة حرارة الغرفة في PBS مع 0.5% Triton X-100، قبل الحضانة مع الأجسام المضادة الثانوية المترافقة مع الفلورسنت المخففة في PBS مع 5% مصل حيوانات دونكي العادي. تم الحصول على صور الأوعية اللمفاوية باستخدام ميكروسكوب ليكا المجهري (Stellaris 8). تم استخدام الأجسام المضادة التالية: جسم مضاد من الماعز ضد VEGFR3 الفأري (R&D، رقم AF743، 1:400)، جسم مضاد من الجرذ ضد LYVE1 الفأري (R&D، رقم MAB2125، 1:400)، جسم مضاد من هامستر سوري ضد بودوبلانين الفأري (BioLegend، 127402، 1:500)، جسم مضاد من الأرنب ضد Prox1 (Angiobio، 11-002 P، 1:200)، جسم مضاد من هامستر أرميني ضد CD31 الفأري (Gene Tex، 2H8، 1:1,000). تم الكشف عن الأجسام المضادة الأولية باستخدام الأجسام المضادة الثانوية المناسبة المترافقة مع AF405 وAF488 وAF555 وAF647 (Thermo Fisher، 1:500) بعد ساعتين من الحضانة في درجة حرارة الغرفة. تم استخدام ProLong Gold Antifade Mountant (Invitrogen، P36930) لتثبيت الشرائح.
بالنسبة لأنسجة الدماغ، تم جمع الأنسجة وحضانتها في خليط هضم يحتوي علىكولاجيناز D (روش) وDNase I (سيغما-ألدريتش) في RPMI عندلمدة 45 دقيقة. تم استخدام الماصة لتفتيت الأنسجة وتم تصفيتها من خلالفلتر. ثم، تم خلط الخلايا في 3 مل منتم تحضير محلول بيركول (سيغما-ألدريتش) وتم الطرد المركزي عند 580 جرام لمدة 15 دقيقة دون فرامل. تمت إزالة طبقة بيركول، وتم معالجة كريات الخلايا بـ 0.5 مل من محلول ACK وتم الطرد المركزي لمدة 5 دقائق عند 500 جرام. ثم، تم إعادة تعليق كريات الخلايا في محلول FACS (PBS + 2% FBS + 1 مليمول EDTA) للتلوين.
عند تحليل اللمفاويات، تم وضع عقدة لمفاوية أو طحال فيطبق بتري بقطر 15 مم يحتوي على 2 مل من محلول FACS وتم طحنه بين شريحتين ميكروسكوب مزججتين. عند تحليل خلايا DCs، تم هضم عقدة لمفاوية أو طحال كما هو موضح أعلاه. تم تصفية تعليقات الخلايا من خلالتمت تصفية العينة ودورانها لمدة 5 دقائق عند 500 جرام. ثم، تم إعادة تعليق كريات الخلايا في محلول FACS للتلوين.
تدفق الخلايا
تم وصف إعداد تعليقات الخلايا المفردة من الطحال والعقد اللمفاوية والدماغ أعلاه. تم حجب الارتباط غير المحدد باستخدام محلول حجب مستقبلات Fc (TruStain FcX، BioLegend، 101320، 1:200) لمدة 10 دقائق فيقبل التلوين المناعي. بعد ذلك، تم تلوين الخلايا بالأجسام المضادة المناسبة لمدة 30 دقيقة فيثم تم غسل الخلايا لإزالة الأجسام المضادة الزائدة وإعادة تعليقها في محلول FACS. تم تشغيل العينات على جهاز قياس التدفق Attune NxT ثم تم تحليلها باستخدام برنامج FlowJo (10.8.1، Tree Star).
اختبار الامتصاص المناعي المرتبط بالإنزيم
تم جمع السائل الدماغي الشوكي والمصل من الفئران كما هو موصوف سابقًا. تم تخفيف المصل والسائل الدماغي الشوكي بمحلول PBS يحتوي علىبكالوريوس فينسبة. تم طلاء الألواح (96 بئر) بـمن HSV-2 المنقى المعطل بالحرارة أو المعطل بواسطة PFA (إلىوحدات تشكيل اللويحات المعادلة لكلقياس الأجسام المضادة الخاصة بالفيروس أو الأجسام المضادة الموجهة ضد الأجسام المضادة للفأر من الماعز (SouthernBiotech، 1010-01، 1:1,000) ثم تم الحضانة طوال الليل في. ثم تم غسل هذه الأطباق بمحلول PBS-Tween 20 وتم حجبها لمدة ساعتين باستخدام 5% FBS في PBS. بعد ذلك، تم وضع العينات في الآبار وتم حضنها لمدة لا تقل عن 4 ساعات في درجة حرارة الغرفة. بعد غسلها بمحلول PBS-Tween 20، تم إضافة الأجسام المضادة الموجهة ضد الغلوبولين المناعي من نوع الفأر المرتبطة بـ HRP (SouthernBiotech، 1010-05، 1:5,000) في الآبار لمدة ساعة، تلتها عملية الغسل وإضافة محلول TMB (eBioscience). تم إيقاف التفاعلات باستخدام 1 Nتم قياس الامتصاص عند 450 نانومتر. تم تعريف عناوين الأجسام المضادة الكلية باستخدام معيار المناعية (لوحة المناعية للأجسام المضادة لفأر C57BL/6؛ SouthernBiotech).
البلوت الغربي
تم هضم الشبكيات المصابة بـ rAAV-RFP أو الشبكيات الضابطة باستخدام خليط يحتوي علىكولاجيناز D (روش) وDNase I (سيغما-ألدريتش) في RPMI عندلمدة 45 دقيقة. تم استخدام الماصة لتفتيت الأنسجة وتم تصفيتها من خلالتم تصفية الخلايا. ثم تم تحلل الخلايا في محلول RIPA وغليها لمدة 5 دقائق مع محلول العينة. تم إجراء التحليل الغربي بطريقة مشابهة لتلك التي تم الإبلاغ عنها سابقًا.باختصار،تم استخدام الجل وتشغيله عند 10 مللي أمبير لكل جل لمدة 30 دقيقة و40 مللي أمبير لكل جل حتى فصل السلم. تم إجراء النقل الرطب عند 120 مللي أمبير لكل جل لمدة 90 دقيقة على الثلج. تم استخدام الأجسام المضادة متعددة النسائل ضد RFP-Tag من الأرانب (تقنيات أوريجين، رقم الكتالوج AP09229PU-N) بتركيز 1:1000 وتم حضنها طوال الليل في غرفة باردة. بعد الغسل، تم استخدام الأجسام المضادة الثانوية المرتبطة بـ HRP ضد IgG من الأرانب (ثيرمو فيشر، G-21234) بتركيز 1:5000 في درجة حرارة الغرفة لمدة ساعتين وتم تصويرها باستخدام نظام تصوير ChemiDoc MP (بايو راد).
اختبار ELISpot
تم حقن الفئران بـ rAAV-RFP عبر الطريق الوريدي أو الطريق البطني. تم جمع العقد اللمفاوية الدرقية والعقد اللمفاوية السطحية والشبكية بعد 10 أيام. تم إعداد تعليقات خلوية مفردة وزرعها مع خلايا الطحال بنسبة 1:5 مع وجود ببتيدات فيروس rAAV-RFP (SNYNKSVNV و NGRDSLVNPGPAMAS). تم قياس الاستجابات المناعية المحددة لـ rAAV باستخدام اختبار ELISpot (إنترفيرون الفأر).مجموعة ELISpot؛ R&D، رقم الكتالوج EL485)، وفقًا لتعليمات الشركة المصنعة للاختبار.
حقن وعلاج ورم الجنب
تم تخدير الفئران باستخدام مزيج من الكيتامين ( ) وزيلزين ( )، وتم حلاقة الجنب وتعقيمه. أتم استخدام حقنة بإبرة 30-G لتوصيل500,000 خلية B16 تحت الجلد. ثم تم علاج الفئران بخلايا B16 المعرضة للإشعاع (250,000 خلية) من خلال طرق الإدارة تحت الجلد، داخل الجمجمة، عبر القناة الهوائية أو عن طريق الوريد (اليوم 7) مع أجسام مضادة مضادة لـ PD1 (RMP1-14) (الأيام 7 و9 و11) عبر الطريق البطني، وتم مراقبة بقائهم.
نقل التبني
لتحليل الاستجابة المناعية (خلايا T و B المحددة لمستضد) بشكل مباشر في العقد اللمفية الدهنية العميقة والعقد اللمفية السطحية بعد حقن IVT أو AC، قمنا بنقل خلايا OT-II و B1-8 وحللنا استجابتها بعد طرق التطعيم المختلفة.
لتحليل استجابة خلايا B المحددة بمستضد، اتبعنا طريقة تم الإبلاغ عنها سابقًاباختصار، تم تحفيز فئران المتلقين CD45.2 C57BL/6 (من عمر 6-8 أسابيع) عن طريق التطعيم داخل الصفاق بجرعة 50 ملغ من OVA (سيغما، A5503) المترسبة في الألوم عند نسبة 2:1 في PBS. بعد أسبوعين، كانت الفئران في حالة راحةتم عزل الخلايا من فئران B1-8 CD45.1.2 باستخدام مجموعة عزل خلايا B من الفئران (Stemcell، 19854). ثم تم وسم خلايا B باستخدام مجموعة وسم تكاثر الخلايا CellTrace Violet (Thermo Fisher، C34557). تم نقل ما مجموعه 5 ملايين خلية عن طريق الوريد إلى الفئران المستقبلة. بعد ثماني ساعات، تم تحصين الفئران بـNP20-OVA (بايو سيرش تك، ) من خلال الحقن الوريدي أو الحقن العضلي. تم تحليل تكاثر الخلايا وتكوين المراكز الجرثومية في العقد اللمفاوية الدرقية (dCLNs) والعقد اللمفاوية السطحية (sCLNs) في اليوم السابع.
للاستجابة المناعية الخاصة بالخلايا التائية المستجيبة للمستضد، خلايا CD4 الخاصة بـ OVAتم عزل خلايا T من فئران CD45.1 OT-II باستخدام CD4 الفأريمجموعة عزل خلايا T (ستيمسيل، 19852). ثم، CD4تم وسم خلايا T باستخدام مجموعة انتشار الخلايا CFSE (ثيرمو فيشر، C34554). تم نقل ما مجموعه 5 ملايين خلية عن طريق الوريد إلى فئران متلقية من نوع CD45.2. بعد ثمانية عشر ساعة، تم تحصين الفئران بـأوفا (سيغما، A5503) زائد بولي (I:C) (إنفيوجن، tlrl-picw) من خلال حقن IVT أو AC. A150 تم حقن كمية من FTY720 (Sigma، SML0700) عن طريق الحقن داخل الصفاق لتثبيط دوران خلايا T المهيأة بعد 24 ساعة من التحصين.CD4 المحدد لـ OVAتم تحليل تكاثر خلايا T في العقد اللمفاوية الجانبية الداخلية (ingLNs) والعقد اللمفاوية الجانبية العميقة (dCLNs) والعقد اللمفاوية الجانبية السطحية (sCLNs) بعد 72 ساعة من التحصين.
حقن IVT و AC و i.c. و ICM
تم تخدير الفئران التي تتراوح أعمارها بين 6-10 أسابيع من خلال حقن داخل الصفاق لمزيج من الكيتامين ( ) وزيلزين ( ). بالنسبة لحقن IVT أو AC، تم توسيع بؤبؤ العين باستخدام محلول تروبيكاميد العيني. لحقن IVT، تم استخدام إبرة 30-G لثقب فتحة على بعد 1 مم خلف نقطة التقاء القرنية والصلبة. تم استخدام حقنة هاملتون ذات النهاية غير الحادة مع من صبغة، HSV-1، HSV-2، S. pneumoniae أو خلايا ورمية مشعة تم إدخالها في الجسم الزجاجي حواليعميق ويتم إعطاؤه بمعدل. بالنسبة لحقن الغرفة الأمامية، تم ثقب الثقب بالقرب من تقاطع القرنية والصلبة وتم إدخال حقنة هاملتون ذات النهاية غير الحادة في الغرفة الأمامية بعمق حوالي 1 إلى 2 مم. بعد حقن داخل الزجاجية أو حقن الغرفة الأمامية، تم تطبيق مرهم بتروليوم العيني على العين لمنع تكوّن الساد. كانت طريقة الحقن داخل القحف مشابهة لتلك المستخدمة في تلقيح الورم، ولكن تم إعطاء l من HSV-1 و HSV-2 و S. pneumoniae أو خلايا الورم المعرضة للإشعاع. تم إجراء حقن ICM كما هو موصوف سابقًا.تم الاحتفاظ بالفئران على وسادات تدفئة ومراقبتها باستمرار حتى التعافي بعد الحقن.
تحصين الفيروسات والبكتيريا والتحدي
تم تخدير الفئران بمزيج من الكيتامين ( ) وزيلزين( ). للتطعيم ضد فيروس الهربس البسيط، تم حقن وحدات تشكيل اللويحات من فيروس الهربس البشري 1 أو 2 المعطل حرارياً عن طريق الحقن تحت الجلد، الوريدي، داخل الصفاق أو داخل الجمجمة. بعد ثلاثين يوماً، تم تحدي هذه الفئران عبر الطريق داخل الجمجمة معتمت مراقبة بقاء فيروس HSV-1 أو HSV-2. لبعض التجارب، تم استخدام CD4أجسام مضادة تستنفد خلايا T (BioXCell، GK1.5، رقم BE0003-1،تم حقن (لمدة 3 أيام) قبل إعادة التحدي. تم إجراء تجارب مماثلة مع S. pneumoniae؛ كانت جرعة التحصين هيمن البكتيريا المعطلة حرارياً وكانت جرعة التحدي هيالبكتيريا الحية.
تصوير وقياس نقل المتتبع
لتحليل شدة الفلورية في العين أو العقد اللمفاوية، تم حقن الدكستران المرتبط إما بـ FITC (40 كيلودالتون و70 كيلودالتون) أو بالتترا ميثيل رودامين (40 كيلودالتون و70 كيلودالتون) في الغرفة الأمامية أو الجسم الزجاجي. من خلال حقن AC أو IVT، على التوالي. لتحليل حركية تصريف الصبغة من العين، تم جمع العين في نقاط زمنية متسلسلة وتم تجانسها فيتم استخدام PBS بطريقة خلط الخرز (Matrix D، 116913500، MP Biomedicals). ثم تم طرد الأنسجة المهروسة في جهاز الطرد المركزي عند لمدة 10 دقائق و تم جمع السائل العلوي في لوحة 96 بئر وتم قراءة شدة الفلورسنت باستخدام أطوال موجية للإشعاع والتحفيز تبلغ 494 نانومتر و514 نانومتر أو 555 نانومتر و585 نانومتر. لقياس تصريف الصبغة إلىتم جمع dCLNs بعد 12 ساعة من حقن الصبغة. تم قياس شدة الفلورسنت كما هو موضح أعلاه. لقياس تصريف الجسيمات النانوية في المصل، استخدمنا بروتوكولًا تم نشره سابقًاباختصار، تم عزل مصل من الفئران ووضعه على شرائح ميكروسكوبية للسماح بالكشف عن نقل المؤشرات في الدم بحساسية عالية وبحجم صغير. للتصوير باستخدام IVIS لنقل المؤشر، تم جمع العين، والعقد اللمفاوية السطحية (sCLNs) والعقد اللمفاوية العميقة (dCLNs) بعد حقن الصبغة من خلال إما طريق إدارة AC أو IVT. تم تصويرها باستخدام نظام التصوير الحي IVIS Spectrum (PerkinElmer). لنقل متتبع IVT في الجسم الحي،تم حقن (Thermo Fisher، O34784) عن طريق IVT في الجسم الزجاجي.حجم الفلورسئين الصوديوم (تم حقن (سانتا كروز، sc-206026) في الدم لتحديد الأوعية الدموية. تم تتبع حركية تصريف AF647-OVA باستخدام ميكروسكوب التصوير Phoenix MICRON IV. لتحليل التوضع المشترك لليمفاويات في غمد العصب البصري مع متتبع IVT،تم حقن جسم مضاد مضاد لـ LYVE1 (R&D، MAB2125) عن طريق الوريد في الجسم الزجاجي. تم جمع أغشية العصب البصري بعد ساعتين وتم صبغها كما هو موضح أعلاه. لتتبع حركية تصريف الصبغة في العين،AF647-OVA (AF647-OVA (“; تم حقن Thermo Fisher، O34784) عن طريق IVT في الجسم الزجاجي. تم استئصال العينين في النقاط الزمنية المحددة ومعالجتهما بطريقة مشابهة لتلك التي تم الإبلاغ عنها سابقًا.باختصار، تم تثبيت العيون في محلول تثبيت هارتمن، وتم إنشاء ثلاث نوافذ على العين باستخدام تقنية النوافذ الموصوفة سابقًا.. مسلسل تم الحصول على المقاطع باستخدام جهاز التجميد (لايكا CM190) بعد تجفيف الأنسجة في تدرج السكروز حتى السكروز. تم تثبيت الأقسام باستخدام مادة تثبيت مضادة للتلاشي ProLong Gold مع DAPI (Invitrogen، P36931) وتم تصويرها باستخدام ميكروسكوب ليكا الفلوري (Stellaris 8).
جمع السائل الدماغي الشوكي
لجمع السائل الدماغي الشوكي، تم تخدير الفئران من خلال حقن داخل الصفاق لمزيج من الكيتامين ( ) وزيلزين ( تم حلاقة وتعقيم الجزء العلوي من الرقبة. تم إجراء شق بطول 1 سم عند قاعدة الجمجمة، وتم فصل عضلات الرقبة العلوية باستخدام الملقط لكشف الحجرة الكبيرة. تم استخدام ميكروبيبتيد مخصص (0.75/11brl GF؛ ستولتينغ) لاختراق السحايا لجمع السائل الدماغي الشوكي.
تلقيح ورم الدماغ وتصوير IVIS
تم إجراء تلقيح الورم كما هو موصوف سابقًا مع بعض التعديلات الطفيفة.تم تخدير الفئران عن طريق الحقن داخل الصفاق باستخدام مزيج من الكيتامين ) وزيلزين ( تم حلق رؤوس الفئران وتعقيم فروة الرأس. تم إجراء شق في منتصف فروة الرأس وحُفرت ثقب باستخدام مثقاب 2 مم على الجانب الجانبي من الدرز السهمي و0.5 مم خلف البريغما باستخدام إبرة 25-G. ثم تم وضع الفئران في إطار ستيريوتاكسي.حقنة هاملتون محملة بـخلايا GL261-Lucتم إدخال (الخلايا) في ثقب الحفر بعمق 4 مم من سطح الدماغ وترك لتوازن لمدة دقيقة واحدة قبل الحقن. تم استخدام مضخة حقن دقيقة (World Precision) للحقن عندتم ترك الحقنة في مكانها لمدة دقيقة أخرى بعد انتهاء التسريب. تم تثبيت الجلد وتنظيفه. بعد الإدارة العضلية لمسكّن (ميلوكسيكام وبوبرينورفين،تم وضع الفئران في قفص مدفأ حتى الشفاء التام. قمنا بتتبع حجم الورم أسبوعياً من خلال تصوير IVIS. تم تخدير الفئران باستخدام الإيزوفلوران وتم حقنها عبر الطريق البطني بمادة د-لوكسيفيرين بوتاسيوم الملحية المتلألئة (PerkinElmer، بعد 10 دقائق، تم تصوير الفئران باستخدام نظام التصوير الحي IVIS Spectrum (بيركين إلمر).
تحضير الأنسجة لتحليل تصريف مستضد ورم CT2A-BFP
تم تحليل أورام i.c. CT2A-BFP بعد 14 يومًا من الحقن. تم جمع الأورام، السحايا، العقد اللمفاوية الداخلية، العقد اللمفاوية العميقة والعقد اللمفاوية السطحية. تم عزل الخلايا الوحيدة النواة وصبغها.
بارابيوز
تم إجراء البارابيوزيس كما هو موصوف سابقًا مع بعض التعديلات الطفيفة.تم تخدير فئران C57BL/6 غير المناعية أو الممنعة بواسطة IVT من نفس العمر والوزن بمزيج من الكيتامين ( ) وزيلزين ( بعد حلاقة الجوانب الجانبية المقابلة لكل فأر، تم تنظيف الجلد وتعقيمه باستخدام وسادة تحضير بالكحول وفرك جراحي بيتادين. تم إجراء شقوق جلدية متطابقة من فوق الركبة مباشرة إلى الكوع، وتم خياطة فأرين معًا باستخدام خيوط فيكرل القابلة للامتصاص من نوع إيثيكون 5-0. ثم، تم تثبيت الجلد باستخدام دبابيس وتم تطبيق مرهم نيوسبورين + مسكن للألم على الشقوق. خلال الجراحة، تم إبقاء الفئران على وسائد تدفئة ومراقبتها باستمرار حتى التعافي.
ربط العقد اللمفاوية السطحية أو العقد اللمفاوية العميقة
لربط العقد اللمفاوية، تم تخدير الفئران بمزيج من الكيتامين ( ) وزيلزين ( تم حلاقة وتعقيم الرقبة الأمامية. تم إجراء شق بطول 2 سم، وتم تعريض العقد اللمفاوية السطحية والعميقة بشكل متتابع باستخدام الملقط. تم كوي الأوعية اللمفاوية الواردة أو الإبقاء عليها سليمة بناءً على ظروف التجربة. ثم تم إغلاق الشق باستخدام خياطة فيكرل 5-0، وتم إخضاع الفئران لنفس الإجراءات بعد العملية كما هو مذكور أعلاه.
نقل rAAV والتصوير
تم حقن الفئران البرية بواسطة IVT مع rAAVالجينومات الفيروسية) مع PBS، VEGFC ( ) أو sVEGFR3 ( ). ثم، بعد شهر واحد، تم إعادة تحدي هذه الفئران بـ rAAV-RFP ( الجينومات الفيروسية). تم تصوير العيون إما على جهاز Phoenix Micron IV أو تم جمعها بعد شهر من نقل rAAV-RFP وثُبتت بـيرجى العثور على المرفق في الليل فيتم تشريح صورة الشبكية بالكامل بعناية وتصويرها باستخدام مجهر ليكا الفلوري (Stellaris 8).
تصوير ثلاثي الأبعاد للأعضاء المزالة بالمذيبات
تم تنفيذ iDISCO كما تم وصفه سابقًا (http://www.idisco. معلومات) تم استخدام الأجسام المضادة التالية: جسم مضاد من الماعز ضد VEGFR3 الفأري (R&D، رقم AF743، 1:400)، جسم مضاد من الجرذان ضد LYVE1 الفأري (R&D، MAB2125، 1:400)، جسم مضاد من الأرانب ضد LYVE1 الفأري (AngioBio، رقم 11-034، 1:200)، جسم مضاد من الفئران ضد VEGFR3 البشري (Santa Cruz Biotechnology، SC-28297، 1:200)، جسم مضاد من الأرانب ضد LYVE1 البشري (Angio-Proteomie، 102-PA50S، 1:200)، جسم مضاد من الماعز ضد IgG-AF647 الفأري (Invitrogen، A21235، 1:500)، جسم مضاد من الحمار ضد IgG-AF647 الماعزي (Invitrogen، A21447، 1:500)، جسم مضاد من الماعز ضد IgG-AF555 الأرانبي (Invitrogen، A21428، 1:500). بعد ذلك، تم تصوير الأعصاب البصرية الشفافة مع أغشية الأعصاب البصرية باستخدام مجهر ليكا الفلوري (Stellaris 8). تم الانتهاء من العرض ثلاثي الأبعاد باستخدام برنامج Imaris 8 (Oxford Instruments).
الترانسكريبتوميات المكانية
تم استخدام تعبير الجينات المكاني 10X Visium لشريحة FFPE (PN-1000185) والبروتوكولات المرتبطة بها (CG000483) من 10X Genomics. تم تثبيت عيون الفئران.معالج ومقسمكما تم وصفه سابقًا. مقاطع عرضية منتم قطع سمك كرة العين باستخدام ميكروتوم (RM2255، لايكا بيوسيستمز) ووضعها بعناية داخل الإطار المرجعي على شريحة فيزيم، ثم تم تجفيف الشرائح في الهواء في درجة حرارة الغرفة طوال الليل وتخزينها في حاوية مجففة قبل تجربة النسخ الجزيئي المكاني. تم خبز الشرائح المحفوظة في الفورمالين، وصبغها بصبغة الهيماتوكسيلين-إيوزين، ثم تم تصويرها باستخدام ميكروسكوب فلووروسينس شامل من كيينس bz-x800. ثم، تمت عملية اختراق الخلايا وإعداد المكتبة وفقًا لدليل مستخدم تعبير الجينات المكاني Visium FFPE باستخدام المواد الكيميائية الموردة (10X Genomics). تم تسلسل المكتبات الناتجة وتحليلها باستخدام Space Ranger (الإصدار 2.1.0)، وتم تحليل البيانات باستخدام Seurat 4.9.9.9040.
معالجة الصور وتحليلها
تم إجراء التحليل الكمي للخلايا المصابة بـ rAAV باستخدام برنامج معالجة الصور FIJI أو ImageJ (NIH أو بيثيسدا).
التحليل الإحصائي
تم إجراء جميع التحليلات الإحصائية باستخدام برنامج GraphPad Prism. تم تحليل البيانات باستخدام اختبار Student غير المقترن ذو الذيلين.-اختبار أو تحليل التباين الأحادي مع اختبارات المقارنات المتعددة (دونيت) باستخدام برنامج بريزم. يتم تعريف الدلالة الإحصائية على أنها، و .
ملخص التقرير
معلومات إضافية حول تصميم البحث متاحة في ملخص تقارير مجموعة ناتشر المرتبط بهذه المقالة.
توفر البيانات
مجموعة بيانات النسخ الجيني المكاني لعين الفأر والعصب البصري متاحة للجمهور في المركز الوطني لمعلومات التكنولوجيا الحيوية تحت رقم الوصول PRJNA1046563. جميع مجموعات البيانات التي تم إنشاؤها و/أو تحليلها خلال الدراسة الحالية متاحة في المقالة. تم توفير بيانات المصدر مع هذه الورقة.
توفر الشيفرة
جميع الشيفرات المستخدمة في التحليل موصوفة في الطرق. ستتوفر الملفات التفصيلية عند الطلب. 43. إيجيما، ن. وآخرون. خلايا الشجرة وخلايا B تعظم استجابة الذاكرة Th1 المخاطية لفيروس الهربس البسيط. ج. تجريبي. طب. 205، 3041-3052 (2008). 44. يونغ، هـ. م. وآخرون. تطوير النظام اللمفاوي اليرقي في سمك الزرد. التنمية 144، 2070-2081 (2017). 45. جتلين، أ. د.، شلمان، ز. ونوسنزاويغ، م. ج. الاختيار النسلي في المركز الجرثومي من خلال التكاثر المنظم والطفرة المفرطة. الطبيعة 509، 637-640 (2014). 46. Ciabattini، أ. وآخرون. توزيع خلايا T المهيأة وخلايا تقديم المستضد المحملة بالمستضد بعد التحصين عن طريق الأنف في الفئران. PLoS ONE 6، e19346 (2011). 47. بانغ، ج. وآخرون. التحضير خطوة بخطوة لشرائح عيون الفئران للتشريح النسيجي الروتيني، المناعة الفلورية، وتعدد الهجين في الموقع للـ RNA. بروتوكولات ستار 2، 100879 (2021). 48. بانغ، ج. وآخرون. نهج استبدال التجميد مع مثبت جلايكزال القائم على المذيبات لمنع تشوه الهياكل العينية. ج. تاريخ التكنولوجيا. 45، 172-181 (2022).
الشكر والتقدير نشكر ن. براون، ل. ديل بريفور، م. لاينهان، د. كيسليش ود. رالستون، الذين لعبوا دورًا حاسمًا في إنشاء البنى التحتية اللازمة لإكمال المخطوطة، على الرغم من أنهم لم يكونوا متورطين بشكل مباشر في إجراء التجارب؛ وجميع أعضاء المجتمع العلمي في جامعة ييل على ملاحظاتهم خلال هذا المشروع. كانت الدراسة مدعومة من تمويل عميد جامعة ييل (إلى إ.س.). أ.ي. هو باحث في معهد هوارد هيوز الطبي. تم دعم ج.-ل.ت. من خلال منحة المعاهد الوطنية للصحة R01NS1210130؛ وتم دعم و.م.س. من خلال منحة المعاهد الوطنية للصحة UH3HL147352. يتم دعم ل.س. من خلال زمالة باركر ب. فرانسيس. تم إنشاء الرسوم البيانية في الأشكال 1أ، د، 2أ، هـ، 3د، ف و4ج والأشكال الممتدة 1د و10د باستخدامبايو ريندر.كوم.
مساهمات المؤلفين: X.Y. و S.Z. و J.H.L. هم المؤلفون المشاركون الأوائل في هذا العمل. قام X.Y. و S.Z. بتنفيذ وتصميم التجارب في هذه المخطوطة. كتب X.Y. و S.Z. و J.H.L. المخطوطة. قام J.H.L. و H.D. و G.M. و G.T. و A.K. و L.H.G. و M.P. و S.F. و T.Z. و F.S.M. بتنفيذ التجارب التي ساهمت في هذا المشروع. ساهم J.Z. و Y.W. و S.M. و N.R. و L.S. و I.S. و A.E. و J.-L.T. و W.M.S. و A.H. و C.Z. و A.R. و A.I. في المواد والموارد التي سمحت بإكمال هذا المشروع. قام E.S. بتصور العمل والحصول على التمويل والإشراف على العمل. ساهم جميع المؤلفين في تحرير النصوص والرسوم التوضيحية لهذه المخطوطة.
المصالح المتنافسة: إ.س.، أ.ر. و أ.ي. هم مؤسسو شركة رهو بايو. أ.ي. هو عضو في مجلس إدارة شركة روش هولدينغ المحدودة. إ.س.، ج.-ل.ت. و أ.ي. هم مخترعون مشتركين في طلب براءة الاختراع رقم 62/929,527، “التلاعب بالأوعية اللمفاوية السحائية لعلاج أورام الدماغ والجهاز العصبي المركزي”. و.م.س. هو مؤسس مشارك لشركات B3 Therapeutics و Stradefy و Xanadu Bio. و.م.س. هو مستشار لشركة Xanadu Bio و Stradefy Biosciences و Johnson & Johnson و Celanese و Cranius و CMC Pharma.
معلومات إضافية
معلومات إضافية النسخة الإلكترونية تحتوي على مواد إضافية متاحة فيhttps://doi.org/10.1038/s41586-024-07130-8. يجب توجيه المراسلات والطلبات للحصول على المواد إلى إريك سونغ. تُعرب مجلة Nature عن شكرها للمراجعين المجهولين على مساهمتهم في مراجعة هذا العمل. معلومات إعادة الطبع والتصاريح متاحة علىhttp://www.nature.com/reprints.
مقالة
أ
\نهاية{شكل}
ح
الشكل 1 من البيانات الموسعة | انظر الصفحة التالية للتعليق.
الشكل البياني الموسع 1| يمكن أن يؤدي التطعيم من خلال الحجرة الخلفية للعين إلى تحفيز حماية مناعية في الدماغ. أ، مشابه للشكل 1أ، ولكن الفئران التي تفتقر إلى خلايا B (تم حقن IVT بـ HSV-2 المعطل حرارياً، وتم علاج مجموعة من الفئران WT IVT بمضاد استنفاد CD4 قبل التحدي. ب، مشابه للشكل 1c، تم ربط dCLNs بعد تحفيز الفئران ولكن قبل إعادة التحدي داخل الجمجمة. تم الانتهاء من التطعيم أولاً. ثم، قبل 7 أيام من إعادة التحدي، تم ربط dCLNs، وتم مراقبة بقاء الفئران بعد التحدي داخل الجمجمة بجرعة قاتلة من HSV-2. (نايف،; تم تحصين IVT، ; ربط العقدة اللمفاوية، ” ). ج، توسيع النتائج من الشكل 1د، تم جمع مصل الدم والسائل الدماغي الشوكي من كل من الفئران الملقحة عن طريق IVT والفئران الساذجة بعد 3 أسابيع من التزاوج. تم قياس الأجسام المضادة المحددة ضد فيروس الهربس البسيط بواسطة ELISA ( ). د، مخطط الخطط التجريبية. تم تحضير فئران المتلقين CD45.2 من سلالة C57BL/6J عن طريق التطعيم. بعد أسبوعين، تم استخدام CD45.1.2 B1-8خلايا B الموسومة بـ CellTraceتم نقل مجموعة تكاثر الخلايا الأرجوانية عن طريق الوريد إلى الفئران المستقبلة. بعد 18 ساعة، تم تحصين الفئران بـNP-OVA على الرغم من حقن IVT أو AC.سلسلة خفيفة إيجابية B1-تم تحليل تكاثر الخلايا وتكوين المركز الجرثومي (GC) في العقد اللمفاوية الدهنية (dCLNs) والعقد اللمفاوية السطحية (sCLNs) في اليوم السابع.استراتيجية تحديد البوابة في تحليل تدفق الخلايا لب1- خلايا B GC. ف، قياس خلايا B1-8 GC في dCLNs و sCLNs ( ). ج ، مخطط الخطط التجريبية. CD45.1OVA محدد CD4 تم نقل خلايا T المعلّمة بـ CFSE عن طريق الوريد إلى فئران متلقية من نوع CD45.2. بعد 18 ساعة، تم تحصين الفئران بـأوفا بلس بولي (I:C) من خلال IVT أو حقن AC. تم حقن FTY720 عن طريق البطن لتثبيط دوران خلايا T المهيأة بعد 24 ساعة من التحصين. خلايا CD4 الخاصة بـ OVAتم تحليل تكاثر خلايا T في العقد اللمفاوية الدهنية (dCLNs) والعقد اللمفاوية السطحية (sCLNs) بعد 72 ساعة من التحصين. h، استراتيجية تصنيف تدفق الخلايا لتحليل CD4 المحدد بواسطة OVAانتشار الأسلحة النووية ). أنا، قياس CD4 المحدد لـ OVA السلبي CFSE خلايا T في العقد اللمفاوية الدهنية، والعقد اللمفاوية السطحية، والعقد اللمفاوية الإربية. البيانات معروضة كمتوسطس.م.تم حساب القيم باستخدام اختبار t لطلاب غير المقترنين ذو الذيلين. تم إنشاء الرسوم البيانية في (د) باستخدامبايو ريندر.كوم.
الشكل البياني الممتد 2 | انظر الصفحة التالية للتعليق.
الشكل البياني الممتد 2 | يمكن أن يوفر التطعيم من خلال الحجرة الخلفية للعين علاجًا علاجيًا لأورام الدماغ. أ، تم حقن الفئران عن طريق القحف بـ 50,000 خلية GL261-Luc وتم علاج الفئران بخلايا GL261-Luc المعرضة للإشعاع من خلال طرق الإدارة تحت الجلد (s.c.)، داخل القحف (i.c.)، عبر الحجرة الأمامية (AC)، أو عبر الحقن داخل الجسم الزجاجي (IVT) (اليوم 7) جنبًا إلى جنب مع الأجسام المضادة المضادة لـ PD-1 (الأيام 7 و9 و11) وتمت مراقبة نمو الورم. RLU، وحدات اللمعان النسبية. (s.c.،، ;IVT، ). ب، تم حقن الفئران عن طريق الحقن داخل القحف بـ 50,000 خلية CT2A-BFP. نسبة تم تحليل خلايا DCs في الورم، والعقد اللمفاوية الإقليمية، والسحايا، والعقد اللمفاوية العميقة، والعقد اللمفاوية السطحية بعد 14 يومًا من حقن الورم. ). ج، د، تجربة مشابهة لـ أ. نسبة CD8 المحدد بـ EMV-2تم تحليل خلايا T في dCLN و sCLN و ingLN في اليوم 14.c، استراتيجية تصنيف تدفق السيتومتر لخلايا EMV-2 المحددةخلايا T. د، قياس CD8 المحدد لـ EMV-2خلايا T في العقد اللمفاوية الدهنية، والعقد اللمفاوية السطحية، والعقد اللمفاوية الإربية. (ناجيف; س.م. ; أي. سي., IVT، تم حقن الفئران تحت الجلد بـ 50,000 خلية B16، وتم علاجها بخلايا B16 المعرضة للإشعاع من خلال طرق الحقن تحت الجلد، داخل الجمجمة، داخل التجويف البطني، وإدارة IVT (في اليوم السابع) مع الأجسام المضادة المضادة لـ PD-1 (في الأيام 7 و9 و11) وتمت مراقبتها من أجل البقاء على قيد الحياة.لكل المجموعات). البيانات معروضة كمتوسطس.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع اختبار المقارنات المتعددة (دونيت).
الشكل البياني الموسع 3 | تمتلك العينان تصريفًا لمفاويًا مقسمًا تم حقن الجسيمات النانوية عن طريق الحقن داخل الجسم الزجاجي أو الحقن الوريدي في العين. تم جمع المصل، ووضعه على شريحة، وتصويره باستخدام ميكروسكوب فلوري بعد ساعة من الحقن. ). تم حقن الصبغة عن طريق الحقن داخل الغرفة الأمامية (AC) أو الحقن داخل الزجاجية (IVT) في العين اليسرى. تم جمع العينين والعقد اللمفاوية السطحية (sCLNs) والعقد اللمفاوية العميقة (dCLNs) لتصوير الإضاءة الفلورية باستخدام نظام IVIS. خرائط حرارية تمثل الصبغة بعد طرح الخلفية في العين وsCLNs وdCLNs بعد ساعة واحدة من الحقن. تم حقن الصبغة عن طريق AC (c) أو IVT (d) في العين وتم قياس وجود الصبغة باستخدام قارئ الألواح الفلورية. تم قياس نسبة العقد اللمفاوية المتماثلة والعقد اللمفاوية المتقابلة التي تحتوي على الصبغة بعد 12 ساعة.فئران).
تم حقن الصبغة عن طريق الحقن الوريدي أو تحت الملتحمة في العين اليسرى. تم جمع العقد اللمفاوية الدرقية وقياسها باستخدام جهاز قراءة اللوحات الفلورية بعد ساعة واحدة من الحقن. ). “، dCLNs ( ) أو sCLNs ( ) تم ربطها جراحيًا. بعد يومين، تم حقن صبغة عن طريق AC أو IVT في العين. تم جمع sCLNs (f) أو dCLNs (g) وقياسها باستخدام جهاز قراءة اللوحات الفلورية بعد ساعة واحدة من الحقن. RFU، وحدة الفلورية النسبية. (AC، ;IVT، ). تم حساب القيم باستخدام تحليل التباين الأحادي مع اختبار المقارنات المتعددة (دونيت). البيانات معروضة كمتوسطس.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع اختبار المقارنات المتعددة (دونيت) أو اختبار t لعينتين غير متطابقتين ذو طرفين.
الشكل البياني الممتد 4| تسلسل مكاني بتقنية 10x VISIUM لعينات كرة العين والعصب البصري في الفأر. تم معالجة عيون الفأر باستخدام إعداد علم الأنسجة التقليدي القائم على FFPE. تم معالجة العينات باستخدام منصة 10x VISIUM وتحليلها باستخدام Seurat. أ، UMAP لكتل التسلسل المكاني المختلفة مع تصنيف إلى هيكل مستوى الأنسجة العام. ب، إسقاط مجموعات نوع الأنسجة على الكرة الأرضية والهياكل المحيطة. ج، صورة لصبغة H&E وتوقيع نوع الخلية المعني تم إسقاطه على بيانات التسلسل المكاني (الأسهم الصفراء تشير إلى المكان الذي يظهر فيه توقيع الجين للتوقيع المعطى).
مقالة
الشكل البياني الموسعالتوصيف التشريحي للأوعية اللمفاوية السحائية التي تغطي العصب البصري. صور إضافية لتلوين VEGFR3 و LYVE1 على الأعصاب البصرية للأرانب (أ، ب)، الخنازير (ج، د)، الرئيسيات غير البشرية (هـ، و، ز)، و فئران (تشير الأسهم الصفراء إلى الهياكل الوعائية التي تم صبغها بشكل فردي إما لـ LYVE1 أو VEGFR3. تشير الأسهم البيضاء إلى الهياكل الوعائية الملوّنة بشكل مشترك.
الشكل 6 من البيانات الموسعة | تحليل مفصل لليمفاويات في غمد العصب البصري. شريحة أذن بطنية كاملة من الفئران ملونة بـ LYVE1 و CD31 و PROX1 و تم تلوين PROX1 و VEGFR3 معًا. الأسفل: تم تلوين LYVE1 و PDPN و PROX1 و VEGFR3 معًا. VEGFR3.b، عينة كاملة من أغشية العصب البصري لفئران WT. الأعلى: LYVE1، CD31،
الشكل 7 من البيانات الموسعة | تحليل مفصل لليمفاويات في غمد العصب البصري باستخدام فئران تقارير لمفاوية. أ، ب، شريحة أذن سفلية كاملة من الفأر وغمد العصب البصري لفئران VEGFR3-CreER“;فئران R26-mTmG التي تم صبغها بـ LYVE1 و PROX1 (أ، مقياس:; ب، مقياس: ). ج، د، التثبيت الكامل
C PROX1-CreER;CDH5-Dre;R26-STOP-mCherry أذن
PROX1-CreER;CDH5-Dre;R26-STOP-mCherry غمد العصب البصري ورقة الأذن البطنية الفأرية وغمد العصب البصري لـ PROX1-CreER“;CDH5-Dre; فئران R26-STOP-mCherry التي تم صبغها بـ LYVE1 و CD31 و VEGFR3 (ج، المقياس:; د، مقياس: ).
الشكل البياني الموسع | انظر الصفحة التالية للتعليق.
مقالة
الشكل التمديدي 8|تقوم الأوعية اللمفاوية في غمد العصب البصري بتصريف الجزء الخلفي من العين. أ، صبغة VEGFR3 في العصب البصري الفأري الكامل و إشارة المتتبع بعد ساعة من حقن AC (يسار) أو IVT (يمين). تشير الأسهم البيضاء إلى التوطن المشترك لـ VEGFR3 والمتتبع. ب، ج، تم حقن جسم مضاد مضاد لـ LYVE1 عن طريق IVT في الجسم الزجاجي. تم جمع أغلفة العصب البصري بعد ساعتين وتم صبغها مع CD31 و PROX1 و VEGFR3. د، تم حقن صبغة عن طريق IVT في العين أو داخل البطين في الدماغ. تم جمع السائل الدماغي الشوكي من خلال cisterna magna وتم قياسه باستخدام قارئ لوحات فلورية بعد ساعة من الحقن. هـ، تم حقن جسم مضاد مضاد لـ LYVE1 عن طريق IVT في العين أو ICM في السائل الدماغي الشوكي. dura تم جمع العينة بعد ساعتين من الحقن وتم تحليل ارتباط الأجسام المضادة LYVE1 على الأوعية اللمفاوية الإيجابية VEGFR3 بواسطة التألق المناعي. تم استخدام الفئران التي لم تتلق حقن الأجسام المضادة anti-LYVE1 كتحكم. تم حقن AF647-OVA عن طريق الوريد في الجسم الزجاجي. تم حقن الفلورسئين عن طريق الحقن تحت الجلد لتوسيم الأوعية الدموية. تم تتبع حركية تصريف AF647-OVA باستخدام Phoenix MICRON.ميكروسكوب تصوير IV. تم حقن AF647-OVA عن طريق IVT في الجسم الزجاجي. تم جمع العيون التي تحتوي على العصب البصري، وتم تحليل موقع AF647-OVA في النقاط الزمنية المحددة.صور مكبرة لـ.
الشكل البياني الموسع 9 | تثبيط اللمف يمكن من إعطاء rAAV المتكرر. أ، تم حقن الفئران عن طريق الوريد بـ rAAV-VEGFC، rAAV-sVEGFR3 أو rAAV-control. بعد شهرين، تم جمع أغشية العصب البصري الخاصة بهم وصبغها بـ LYVE1. ب، مشابه للشكل 4ج، تم ربط العقد اللمفية الدرقية قبل حقن rAAV-RFP الأساسي. تم قياس شدة RFP الخاصة بهم. ج، النسبة المئوية تم قياس احتفاظ الصبغة في العين للفئران الضابطة، والفئران المربوطة بـ dCLN، أو الفئران التي تم حقنها بـ rAAV-sVEGFR3. البيانات معروضة كمتوسطس.م.تم حساب القيم باستخدام تحليل التباين الأحادي مع اختبار المقارنات المتعددة (دونيت).
مقالة
الشكل 10 من البيانات الموسعة | تثبيط اللمف يسمح بتكرار rAAV التحويل. صور مجهرية متداخلة كاملة من إعادة تحدي rAAV-RFP في شبكية العين الفأرية بعد العلاج بـ VEGFC أو sVEGFR3 خلال العدوى الأولية. تم جمع شبكية العين الفأرية، وتم التعبير عن RFP كما هو مشابه للشكل 4c تم قياسها باستخدام تقنية Western blot. ج، مشابهة للشكل 4ج، تم جمع شبكية العين الفأرية، وتم قياس تعبير RFP باستخدام تقنية تحليل تدفق الخلايا. د، ملخص تخطيطي للنتائج. الرسوم البيانية فيتم إنشاؤها بـبايو ريندر.كوم.
محفظة الطبيعة
المؤلف (المؤلفون) المراسلون: إريك سونغ آخر تحديث من المؤلف(ين): ديسمبر 132023
ملخص التقرير
تسعى Nature Portfolio إلى تحسين إمكانية تكرار العمل الذي ننشره. يوفر هذا النموذج هيكلًا للاتساق والشفافية في التقرير. لمزيد من المعلومات حول سياسات Nature Portfolio، يرجى الاطلاع على سياسات التحرير وقائمة مراجعة سياسة التحرير.
الإحصائيات
لجميع التحليلات الإحصائية، تأكد من أن العناصر التالية موجودة في أسطورة الشكل، أسطورة الجدول، النص الرئيسي، أو قسم الطرق.
مؤكد
حجم العينة بالضبط ( لكل مجموعة/شرط تجريبي، معطاة كرقم منفصل ووحدة قياس
بيان حول ما إذا كانت القياسات قد أُخذت من عينات متميزة أو ما إذا كانت نفس العينة قد تم قياسها عدة مرات
اختبار(ات) الإحصاء المستخدمة وما إذا كانت أحادية الجانب أو ثنائية الجانب يجب أن تُوصف الاختبارات الشائعة فقط بالاسم؛ واصفًا التقنيات الأكثر تعقيدًا في قسم الطرق.
وصف لجميع المتغيرات المشتركة التي تم اختبارها
وصف لأي افتراضات أو تصحيحات، مثل اختبارات الطبيعية والتعديل للمقارنات المتعددة
وصف كامل للمعلمات الإحصائية بما في ذلك الاتجاه المركزي (مثل المتوسطات) أو تقديرات أساسية أخرى (مثل معامل الانحدار) و التباين (مثل الانحراف المعياري) أو تقديرات مرتبطة بعدم اليقين (مثل فترات الثقة)
لاختبار الفرضية الصفرية، فإن إحصائية الاختبار (على سبيل المثال، ) مع فترات الثقة، أحجام التأثير، درجات الحرية وقيمة ملحوظة أعطِالقيم كقيم دقيقة كلما كان ذلك مناسبًا. لتحليل بايزي، معلومات حول اختيار القيم الأولية وإعدادات سلسلة ماركوف مونت كارلو للتصاميم الهرمية والمعقدة، تحديد المستوى المناسب للاختبارات والتقارير الكاملة عن النتائج تقديرات أحجام التأثير (مثل حجم تأثير كوهين)بيرسون )، مما يشير إلى كيفية حسابها تحتوي مجموعتنا على الإنترنت حول الإحصائيات لعلماء الأحياء على مقالات تتناول العديد من النقاط المذكورة أعلاه.
البرمجيات والشيفرة
معلومات السياسة حول توفر كود الكمبيوتر
جمع البيانات لم يتم استخدام أي برنامج تحليل البيانات
بالنسبة للمخطوطات التي تستخدم خوارزميات أو برامج مخصصة تكون مركزية في البحث ولكن لم يتم وصفها بعد في الأدبيات المنشورة، يجب أن تكون البرمجيات متاحة للمحررين والمراجعين. نحن نشجع بشدة على إيداع الشيفرة في مستودع مجتمعي (مثل GitHub). راجع إرشادات مجموعة Nature لتقديم الشيفرة والبرمجيات لمزيد من المعلومات.
بيانات
معلومات السياسة حول توفر البيانات
يجب أن تتضمن جميع المخطوطات بيانًا حول توفر البيانات. يجب أن يتضمن هذا البيان المعلومات التالية، حيثما ينطبق:
رموز الانضمام، معرفات فريدة، أو روابط ويب لمجموعات البيانات المتاحة للجمهور
وصف لأي قيود على توفر البيانات
بالنسبة لمجموعات البيانات السريرية أو بيانات الطرف الثالث، يرجى التأكد من أن البيان يتماشى مع سياستنا
البحث الذي يتضمن مشاركين بشريين، بياناتهم، أو مواد بيولوجية
معلومات السياسة حول الدراسات التي تشمل مشاركين بشريين أو بيانات بشرية. انظر أيضًا معلومات السياسة حول الجنس، الهوية/التقديم الجنسي، والتوجه الجنسي والعرق، والعرقية والعنصرية.
التقارير عن الجنس والنوع الاجتماعي
لم تتضمن الدراسة مشاركين بشريين
التقارير عن العرق أو الإثنية أو غيرها من التجمعات الاجتماعية ذات الصلة
لم تتضمن الدراسة مشاركين بشريين
خصائص السكان
لم تتضمن الدراسة مشاركين بشريين
التوظيف
لم تتضمن الدراسة مشاركين بشريين
رقابة الأخلاقيات
لم تتضمن الدراسة مشاركين بشريين
يرجى ملاحظة أنه يجب أيضًا تقديم معلومات كاملة حول الموافقة على بروتوكول الدراسة في المخطوطة.
التقارير الخاصة بالمجال
يرجى اختيار الخيار أدناه الذي يناسب بحثك بشكل أفضل. إذا لم تكن متأكدًا، اقرأ الأقسام المناسبة قبل اتخاذ قرارك. علوم الحياة X العلوم السلوكية والاجتماعية العلوم البيئية والتطورية والبيئية لنسخة مرجعية من الوثيقة مع جميع الأقسام، انظرnature.com/documents/nr-reporting-summary-flat.pdf
تصميم دراسة العلوم الحياتية
يجب على جميع الدراسات الإفصاح عن هذه النقاط حتى عندما يكون الإفصاح سلبياً.
حجم العينة
تم الإبلاغ عن أحجام العينات لجميع التجارب. قمنا برسم ملخص للسمات الرئيسية التي لاحظناها خلال عدة تجارب. مع عرض هذين التجربين
معدلات الاستجابة، فإن حساب حجم العينة المطلوب لمستوى ألفا قدره 0.05 هو (قوة ) و (قوة ).
استثناءات البيانات
تم استبعاد البيانات فقط عندما فشلت آلة جمع التجارب ولم يتم جمع أي نقاط بيانات، حيث لم نتمكن من إنتاج ملفات نتائج قابلة للقراءة.
التكرار
تم تكرار جميع التجارب كتنويع (تغييرات في نقاط الزمن) أو في حجم أصغرالأرقام، وأظهرت اتجاهات مماثلة واستنتاجات تجريبية. بالنسبة لمعظم التجارب الموصوفة، فهي أمثلة متسلسلة لجميع التجارب التي تم إجراؤها (ليس صحيحًا بالنسبة للتجارب التي تكون فيها التباينات بين الأيام المختلفة أعلى، أي قياسات MFI). كانت جميع التجارب قابلة للتكرار. تم تكرار جميع التجارب على الأقل مرتين وتم الإبلاغ عنها وفقًا لذلك في الأساطير.
العشوائية
تم توزيع الفئران عشوائيًا على كل مجموعة وتم التحكم في التجارب مع وجود ضوابط في كل يوم حتى للتجارب المكررة.
عمى
تم تعمية محقق واحد على الأقل لكل دراسة للسماح بتقارير غير متحيزة للبيانات
التقارير عن مواد وأنظمة وطرق محددة
نحتاج إلى معلومات من المؤلفين حول بعض أنواع المواد والأنظمة التجريبية والأساليب المستخدمة في العديد من الدراسات. هنا، يرجى الإشارة إلى ما إذا كانت كل مادة أو نظام أو طريقة مدرجة ذات صلة بدراستك. إذا لم تكن متأكدًا مما إذا كان عنصر القائمة ينطبق على بحثك، يرجى قراءة القسم المناسب قبل اختيار رد.
تم شراء الأجسام المضادة من الماعز ضد VEGFR3 الفأري (#AF743) والأجسام المضادة من الجرذ ضد LYVE1 الفأري (# MAB2125) من R&D. تم شراء الأجسام المضادة من الفأر ضد VEGFR3 البشري (SC-28297) من Santa Cruz Biotechnology. تم شراء الأجسام المضادة من الأرنب ضد Prox1 (11-002P) والأجسام المضادة من الأرنب ضد LYVE-1 البشري (102-PA50S) من Angio-proteomie. تم شراء Podoplanin (127402) من biolegend. تم شراء الأجسام المضادة من هامستر أرميني ضد CD31 الفأري (2H8) من Gene Tex. تم شراء الأجسام المضادة من الماعز ضد IgG-AF647 الفأري (A21235)، والأجسام المضادة من الحمار ضد IgG- AF647 للماعز (A21447)، والأجسام المضادة من الماعز ضد IgG-AF555 للأرنب (A21428). تم شراء الأجسام المضادة من الماعز ضد IgG- AF-488 لهامستر أرميني (# A78963) والأجسام المضادة من الماعز ضد IgG- AF-488 لهامستر سوري (# A78958) من Invitrogen.
تم شراء جسم مضاد متعدد النسائل ضد علامة RFP-Tag (#AP09229PU-N) من شركة أوريجين تكنولوجيز.
تم شراء الأجسام المضادة للفأر ضد زيبرا فيش zns-2 (ZDB-ATB-081002-34) من ZIRC. تم شراء الأجسام المضادة للدجاج ضد GFP (Cat# GFP-1010) من Aves. تم شراء الأجسام المضادة المربوطة بـ AF488 من الماعز ضد IgG للفأر AF488 (115-545-146) والأجسام المضادة المربوطة بـ Cy5 من الحمار ضد IgY للدجاج (703-175-155) من Jackson Immuno Research.
تم شراء الأجسام المضادة Goat anti-mouse immunoglobulin (1010-01) والأجسام المضادة HRP-conjugated anti-mouse Ig (1010-05) من SouthernBiotech.
تم شراء جسم مضاد CD4 من الفئران المضادة للفئران (#BE0003-1، GK1.5) من BioXCell.
التحقق
جميع الأجسام المضادة المستخدمة هي أجسام مضادة شائعة الاستخدام ومُعتمدة وفقًا لتعليمات الشركة المصنعة. يرجى الرجوع إلى كل شركة مصنعة كما هو مذكور أعلاه. لم يتم استخدام أي أجسام مضادة تم توليدها حديثًا أو أجسام مضادة غير تقليدية.
خطوط خلايا حقيقية النواة
معلومات السياسة حول خطوط الخلايا والجنس والنوع في البحث
مصدر(s) خط الخلايا
تم الحصول على خلايا GL261-Luc كهدية من J. Zhou (جراحة الأعصاب في جامعة ييل) وتم زراعتها في RPMI معززة بـFBS،البنسلين/الستربتوميسين وبيوروفات الصوديوم. كانت خلايا CT-2A-BFP هدية من T. Mathivet (مركز الأبحاث القلبية الوعائية في باريس). كانت خلايا B16 هدية من N. Palm (علم المناعة في ييل).
المصادقة
لم يتم توثيق الخلايا بشكل منفصل، ولكن تم التحقق من الأنماط الظاهرية باستخدام خطوط خلايا موثقة مماثلة من المعهد الوطني للصحة.
تلوث الميكوبلازما
جميع الخلايا أظهرت نتائج سلبية لوجود تلوث بالميكوبلازما
الخطوط التي يتم التعرف عليها بشكل خاطئ بشكل شائع (انظر سجل ICLAC)
لم تستخدم الدراسة الخطوط التي يتم التعرف عليها بشكل خاطئ بشكل شائع
الحيوانات وغيرها من الكائنات البحثية
معلومات السياسة حول الدراسات التي تشمل الحيوانات؛ توجيهات ARRIVE الموصى بها للإبلاغ عن أبحاث الحيوانات، والجنس والنوع في البحث
الحيوانات المخبرية
فئران مختلطة الجنس من سلالة C57BL/6 تتراوح أعمارها بين ستة إلى عشرة أسابيع، B6.Cg-Tg(TcraTcrb)425Cbn/J (OT-II)، B6.129P2(C)-Ightm2Cgn/J (B1-8) و B6.129S2IghtmICgn/J (تم شراء الفئران من مختبر جاكسون وشارلز ريفر، وتم تربيتها وإيواؤها لاحقًا في جامعة ييل. كانت فئران PROX1CreERT2;CDH5 Dre;R26-STOP-mCherry وVEGFR3-CreERT2; R26-MTMG هدايا من مختبر توماس. جميع الإجراءات المستخدمة في هذه الدراسة (مطابقة من حيث الجنس والعمر) تتوافق مع الإرشادات الفيدرالية وسياسات المؤسسة الخاصة بلجنة رعاية واستخدام الحيوانات في كلية الطب بجامعة ييل.
الحيوانات البرية
لم تتضمن الدراسة حيوانات برية
التقارير عن الجنس
تم استخدام فئران مختلطة الجنس في دراساتنا ولم نلاحظ أي أنماط ظاهرة متحيزة للجنس.
عينات تم جمعها من الميدان
لم تتضمن الدراسة عينات تم جمعها من الميدان
رقابة الأخلاقيات
تمت الموافقة على جميع الأعمال المتعلقة بالحيوانات من قبل لجنة رعاية واستخدام الحيوانات في جامعة ييل.
يرجى ملاحظة أنه يجب أيضًا تقديم معلومات كاملة حول الموافقة على بروتوكول الدراسة في المخطوطة.
مخزونات البذور
لم تتضمن الدراسة النباتات
أنماط جينية نباتية جديدة
لم تتضمن الدراسة النباتات
المصادقة
لم تتضمن الدراسة النباتات
تدفق الخلايا
مؤامرات
أكد أن:
X تسرد تسميات المحاور العلامة والفوروشروم المستخدم (مثل CD4-FITC).
X محاور المقياس مرئية بوضوح. قم بتضمين الأرقام على المحاور فقط للرسم البياني في أسفل اليسار من المجموعة (المجموعة هي تحليل للعلامات المتطابقة).
جميع الرسوم البيانية هي رسوم بيانية متساوية الارتفاع مع نقاط شاذة أو رسوم بيانية بالألوان الزائفة.
X يتم توفير قيمة عددية لعدد الخلايا أو النسبة المئوية (مع الإحصائيات).
المنهجية
تحضير العينة
بالنسبة لأنسجة الدماغ، تم جمع الأنسجة وحضنها في خليط هضم يحتوي علىكولاجيناز D (روش) وDNase I (سيغما-ألدريتش) في RPMI عندلمدة 45 دقيقة. تم استخدام الماصة لتفتيت الأنسجة وتم تصفيتها من خلالفلتر. ثم، تم خلط الخلايا في 3 مل منتم تحضير محلول بيركول (سيغما-ألدريتش) وتم الطرد المركزي عند 580 جرام لمدة 15 دقيقة دون فرامل. تمت إزالة طبقة بيركول، وتم معالجة كريات الخلايا بـ 0.5 مل من محلول ACK وتم الطرد المركزي لمدة 5 دقائق عند 500 جرام. ثم تم إعادة تعليق كريات الخلايا في محلول FACS (PBS).FBS +1 مليمول EDTA) للتلوين.
آلة
تم وصف إعداد تعليقات الخلايا المفردة من الطحال والعقد اللمفاوية والدماغ أعلاه. تم حجب الارتباط غير المحدد باستخدام محلول حجب مستقبلات Fc (TruStain FcX).101320، BioLegend) لمدة 10 دقائق فيقبل التلوين المناعي. بعد ذلك، تم تلوين الخلايا بالأجسام المضادة المناسبة لمدة 30 دقيقة فيثم تم غسل الخلايا لإزالة الأجسام المضادة الزائدة وإعادة تعليقها في محلول FACS. تم تشغيل العينات على جهاز قياس التدفق Attune NxT ثم تم تحليلها باستخدام برنامج FlowJo (10.8.1، Tree Star).
برمجيات
برنامج FlowJo (10.8.1، شجرة النجمة)
وفرة تجمع الخلايا
لم يتم إجراء أي فرز
استراتيجية البوابة
يرجى الرجوع إلى الأشكال والأساليب التكميلية لاستراتيجيات التصفية التفصيلية
قم بتحديد هذا المربع لتأكيد أنه تم تقديم رقم يوضح استراتيجية البوابة في المعلومات التكميلية.
Xiangyun Yin , Sophia Zhang , Ju Hyun Lee , Huiping Dong , George Mourgkos , Gordon Terwilliger , Aurora Kraus , Luiz Henrique Geraldo , Mathilde Poulet , Suzanne Fischer , Ting Zhou , Farrah Shalima Mohammed , Jiangbing Zhou , Yongfu Wang , Seth Malloy , Nicolas Rohner , Lokesh Sharma , Irene Salinas , Anne Eichmann , Jean-Leon Thomas , W. Mark Saltzman , Anita Huttner , Caroline Zeiss , Aaron Ring , Akiko Iwasaki & Eric Song
Abstract
The eye, an anatomical extension of the central nervous system (CNS), exhibits many molecular and cellular parallels to the brain. Emerging research demonstrates that changes in the brain are often reflected in the eye, particularly in the retina . Still, the possibility of an immunological nexus between the posterior eye and the rest of the CNS tissues remains unexplored. Here, studying immune responses to herpes simplex virus in the brain, we observed that intravitreal immunization protects mice against intracranial viral challenge. This protection extended to bacteria and even tumours, allowing therapeutic immune responses against glioblastoma through intravitreal immunization. We further show that the anterior and posterior compartments of the eye have distinct lymphatic drainage systems, with the latter draining to the deep cervical lymph nodes through lymphatic vasculature in the optic nerve sheath. This posterior lymphatic drainage, like that of meningeal lymphatics, could be modulated by the lymphatic stimulator VEGFC. Conversely, we show that inhibition of lymphatic signalling on the optic nerve could overcome a major limitation in gene therapy by diminishing the immune response to adeno-associated virus and ensuring continued efficacy after multiple doses. These results reveal a shared lymphatic circuit able to mount a unified immune response between the posterior eye and the brain, highlighting an understudied immunological feature of the eye and opening up the potential for new therapeutic strategies in ocular and CNS diseases.
Herpes simplex virus (HSV) has the propensity to affect many tissuesincluding multiple neural tissues such as the dorsal root ganglia, the eyes and the brain-and is the most common cause of sporadic fatal encephalitis worldwide . To understand protective immunity against HSV in the brain, we vaccinated mice with heat-inactivated HSV-2 through four administration routes-intraperitoneal (i.p.) to induce systemic immunity, intracranial (i.c.) to induce local brain immunity, intracameral (anterior chamber(AC)), and intravitreal (posterior chamber (IVT))-with the last two being ocular compartments that have unique anatomical connections to the nervous system (Fig. 1a). After a lethal i.c. HSV-2 challenge, all mice that underwent i.p. immunization succumbed to the infection, whereas i.c. immunization protected about of the mice (Fig. 1b), showing that systemic immunity is not
sufficient in providing protection against CNS infection. Remarkably, IVT immunization also protected the mice, but all AC-immunized mice, like i.p.-immunized mice, succumbed to the challenge (Fig. 1b), suggesting that IVT immunization drives a protective immune response in the brain.
As is the case for other organs, we reasoned that the immune response in the eye-brain axis must be mediated by local lymph nodes (LNs). To investigate the cellular mechanism facilitating the protection of the brain after IVT immunization, we first surgically ligated lymphatic vessels of the deep cervical LNs (dCLNs), which are critical to eliciting immune responses in the brain. This completely abrogated the protection provided by IVT immunization, indicating the importance of a local CNS-LN circuit that drains antigen to dCLNs (Fig. 1c). Additionally,
Fig. 1 Antigens in the posterior eye elicit immune responses in the brain.
a, Schematic of the schedule of procedures for the experiments described below.b, Wild-type C57BL/6J mice were immunized using heat-inactivated HSV-2 injection through i.p., i.c., AC and IVT administration. Survival was monitored after i.c. challenge with a lethal dose of HSV-230 days later (naive, ; i.p., ; i.c., ; AC, ; IVT, ). c, dCLNs of mice were ligated using a cauterizer. Seven days later, mice were injected through the IVT route with heat-inactivated HSV-2. Their survival was monitored after i.c. challenge with a lethal dose of HSV-230 days later (naive, ; IVT immunized, ; LN ligation, ). d, Schematic of the parabiosis mouse model and treatment plans. e, Mice were injected through the IVT route with heat-inactivated HSV-2. Four weeks later, the immunized mice were joined to naive mice. The immunized mice or naive mice were challenged through the i.c. route with a lethal dose of HSV-2 after 3 weeks, and their survival was monitored (naive, ; IVT, ; IVT-naive (IVT challenge), ; IVT-naive (naive challenge), ; naive-naive
(naive challenge), ). , Anti-HSV-specific antibody was measured by enzyme-linked immunosorbent assay after different routes of HSV-2 immunization (i.p., ; i.c., ; ; IVT, ). Data are shown as mean s.e.m.g, Wild-type C57BL/6J mice were injected with heat-inactivated HSV-1 through i.p., i.c., AC or IVT administration. Their survival was monitored after i.c. challenge with a lethal dose of HSV-130 days later (naive, ; i.p., ; i.c., ; AC, ; IVT, ). h, As in a, but S. pneumoniae strain TIGR4 was used (naive, ; i.p., ; i.c., ; AC, ; IVT, ). i, Mice were inoculated through the i.c. route with luciferase-expressing (GL261-Luc) brain tumour cells, treated with irradiated GL261-Luc cells through s.c., i.c., AC or IVT administration (day 7) along with anti-PD1 (RMP1-14) antibodies (days 7, 9 and 11) and monitored for survival (naive, ; s.c., ; i.c., ). Data are representative of two independent experiments. The graphics in a,d were created with BioRender.com.
as we ligated the lymphatic vasculature after priming but before the rechallenge, our results indicated that the lymphatic vessels of dCLNs were required to mount a recall response during the challenge (Extended Data Fig. 1b). The effector functions after HSV immunization are thought to be mediated by antiviral antibodies and cells . Consistent with these reports, our observations showed that CD4 T cell-depleted or B cell-deficient ( ) mice were no longer protected after IVT immunization (Extended Data Fig. 1a). As efficient differentiation of follicular helper T cells and germinal centre B cells is required for the production of long-lived plasma cells and memory cells that drive a superior humoral immune response following re-exposure to an antigen , we quantified antigen-specific germinal centre B cell differentiation by adoptive transfer of cells after immunizing mice with NP-OVA through AC or IVT routes. Although both immunization routes can efficiently induce germinal centre B cell differentiation in superficial CLNs (sCLNs), only IVT immunization induces germinal centre B cell differentiation in dCLNs (Extended Data Fig. 1d-f). Similarly, IVT immunization significantly increases the level of antigen-specific CD4
T cell proliferation in dCLNs compared to AC immunization (Extended Data Fig. ). These data indicate that the generation of localized antigen-specific CD4 T and B cell responses in dCLNs by IVT immunization is required for host antiviral CNS protection.
To further address whether the IVT immunization-mediated protection was developed locally in the brain, we generated parabiotic mouse pairs with various immunization states (Fig.1d). When IVT-immunized mice were paired with naive mice, only the mice that were initially immunized (left) and not their parabiotic partners (right) were protected against an i.c. challenge (Fig. 1e). Similarly, only the IVT-immunized mice, but not their naive parabiotic partners, had detectable levels of anti-HSV-2 antibodies in cerebrospinal fluid (CSF), which indicated that direct immunization is necessary to acquire anti-HSV-2 antibodies in CSF (Extended Data Fig. 1c). By contrast, because of the shared blood circulation, naive mice that were paired with IVT-immunized mice had a similar amount of anti-HSV-2 antibodies in their serum (Extended Data Fig. 1c). This suggested that serum antibodies do not mediate immune protection in the CNS; rather, immune protection occurs
Article
locally through the CSF. Furthermore, following examination of the localization of anti-HSV antibodies after different immunization routes, we found that, although all routes of immunization led to the presence of anti-HSV-2 antibodies in serum, only the i.c.- and IVT-immunized mice had detectable levels of anti-HSV-2 antibodies in CSF (Fig. 1f). Together, these data demonstrate that IVT immunization uniquely mediates CNS protection through a local antibody-dependent response.
Finally, to determine whether this ocular-brain axis-mediated CNS protection extends beyond HSV-2, we examined the efficacy of a similar vaccination strategy against HSV-1 and Streptococcus pneumoniae, the main causative agent of bacterial meningitis . Mice were immunized with heat-inactivated HSV-1 or S. pneumoniae through an i.p., i.c., AC or IVT route and rechallenged with a lethal dose of the same pathogen 30 days later. Similar to the results in the HSV-2 model, the data for both IVT- and i.c.-immunized mice showed them to have almost complete protection. By contrast, all of the naive, i.p.- and AC-immunized mice succumbed to the challenge (Fig. 1g,h). Even in a therapeutic setting in which an i.c. tumour was established before the administration of a cancer cell vaccine, immunization through the IVT or i.c. route resulted in superior protection compared to subcutaneous (s.c.) and AC immunization, which evoked only systemic immunity (Fig. 1i and Extended Data Fig. 2a). These data were consistent with previously described findings that IVT and i.c. immunization resulted in significantly greater proportions and numbers of antigen-specific CD8 T cells compared to those following AC and s.c. immunization in dCLNs, but not in sCLNs or inguinal LNs (IngLNs; Extended Data Fig. 2b-d). By contrast, in a model of cutaneous melanoma, s.c. immunization showed better protection than IVT or i.c. immunization (Extended Data Fig. 2e), indicating that IVT and i.c. immunization induce a localized immune response but do not provide protection against peripheral tumours as well as a local immunization method does. Together, these data demonstrate that the ocular-brain immunological connection is specifically relevant and broadly applicable to many CNS diseases.
The eye has a compartmentalized drainage system
Our immunization studies uncovered a divergent immune response between the AC and vitreous of the eye. It is thought that aqueous humour flows out from the AC through the conventional outflow pathway that includes the Schlemm’s canal and trabecular meshwork . Additionally, a more recently appreciated unconventional route includes flow through the ciliary muscle and supraciliary and suprachoroidal spaces . By contrast, the drainage from the posterior ocular compartment, particularly the vitreous, is unclear given the lymphatic vasculature in the eye and is still being explored .
To investigate the different drainage systems of the anterior and posterior compartments of the eye, we injected fluorescently labelled dextrans through AC or IVT routes and quantified the kinetics of dextran retention (Fig. 2a). The rate of dextran clearance in the eye after IVT administration ( ) was significantly slower than that after AC administration ( ; Fig. 2b). Dextrans with two different wavelengths of fluorescence and molecular weights were tested to ensure that this phenomenon was not dye specific. We reasoned that this observation could be explained by two possibilities; either the compartments have unique routes of drainage, or the rate-limiting step in the drainage of each compartment is different (that is, diffusion through the vitreous may be the slowest step). We reasoned that these potential explanations could be parsed out by observing dye localization in vivo. Using a previously described method , we confirmed that both AC- and IVT-injected dye could be detected in the serum. However, significantly higher concentrations of dye were detected in the blood after AC injection compared to after IVT injection (Extended Data Fig. 3a). Whereas dye injected into the AC localized to the ipsilateral sCLNs, as previously reported , IVT-injected dye localized to bilateral dCLNs in addition to the ipsilateral sCLNs (Fig. 2c-e and
Extended Data Fig. 3b-d). As was observed after AC injection, dye does not drain to the dCLNs after subconjunctival injection (Extended Data Fig. 3e), indicating that penetration into the posterior eye or vitreous is required for drainage to the dCLNs. To test the requirement of these local lymphatic structures for drainage of the aqueous and vitreous humour, we surgically ligated the sCLNs, dCLNs or both. We first confirmed that ligation of dCLNs did not result in increased efflux into the sCLNs and vice versa (Extended Data Fig. 3f,g). Ligation of sCLNs, but not dCLNs, resulted in significantly increased dye retention in the eye following AC injection (Fig. 2f), consistent with the above observation. For IVT injection, either sCLN or dCLN ligation alone led to increased dye retention in the eye (Fig. 2g). Together, these results demonstrate a compartmentalized drainage system in the eye between the AC and the vitreous humour with a specific route from the vitreous humour to the dCLNs.
Optic nerve sheath harbours functional lymphatics
Macromolecule localization into distinct LNs indicates that a lymphatic vessel network is probably mediating drainage from each of these compartments. Although a recent study described the presence of an ocular glymphatic system in mice, similar to that of the brain , a functional network of lymphatic vessels mediating vitreous drainage has not been resolved. Therefore, we used a transcriptomic approach along with immunolabelling-enabled three-dimensional imaging of solvent-cleared organs (iDISCO) to obtain a global view of the lymphatic vascular network associated with the optic nerve in different vertebrate species. Spatial transcriptomics helped gather an organ-level view of lymphatic-like vasculature gene signatures in the eye and the optic nerve. In addition to the trabecular meshwork and retina, which have been found to harbour macrophages expressing some lymphatic-like gene signatures , we found that the optic nerve sheath also contained lymphatic-like gene signatures (Extended Data Fig.4). In parallel, iDISCO revealed lymph vessels covering zebrafish, mouse, rabbit, pig, non-human primate and human optic nerves (Fig. 3a,b and Extended Data Fig. 5), demonstrating an evolutionarily conserved feature of the lymphatic system. In all mammals, we saw vascular structures with co-localized staining of LYVE1 and vascular endothelial growth factor receptor 3 (VEGFR3; white arrows in Extended Data Fig. 5). We also saw vasculature stained for one but not the other, indicating the likely presence of LYVE1-negative collecting lymphatic vasculature or LYVE1-expressing macrophages (yellow arrows in Extended Data Fig.5). Notably, the lymphatic vasculature was localized to membranes surrounding the nerve, not the nerve itself, in parallel to what is observed in the brain and the dural meninges .
To improve the sensitivity of the stains applied, we dissected out the optic nerve sheath completely from the optic nerve. Without perfusion fixation of mice, the optic nerve sheath retracted following enucleation, resulting in bunching of the lymphatic vasculature towards the proximal end of the nerve (Extended Data Fig. 5h). To address this, we carried out perfusion fixation before enucleation, preserving the anatomy of the lymphatic network along the nerve (Extended Data Fig. 5i). Using the well-characterized ear lymphatic system as a control (Extended Data Fig. 6a), we validated the presence of optic nerve sheath lymphatics using additional markers (VEGFR3, PROX1, LYVE1, CD31 and podoplanin; Fig. 3c and Extended Data Fig. 6b). To further characterize these vasculatures using a genetic approach, we used two different reporter mice enabling highly specific staining of lym-phatics:VEGFR3-CreER ;R26-mTmG mice, and PROX1-CreER ;CDH5-Dre;R26-STOP-mCherry mice (Extended Data Fig. 7). Together, these parallel methods confirmed that lymphatics indeed exist in the optic nerve sheath.
We then sought to evaluate the functionality of these lymphatics. Following IVT injection, but not AC injection, of a fluorescent tracer, the tracer was visualized in VEGFR3 vasculature along the optic nerve
Fig. 2 | Eyes have a compartmentalized lymphatic drainage system.
a, Schematic of AC and IVT injection of dye.b, C57BL/6J mice were injected with dye through the AC or IVT route. The percentage of dye retention in the eye was analysed from 6 h to day 5 post injection (AC, ; IVT, ). c, Dye was injected into the eye through the AC or IVT route. SCLNs and dCLNs were collected and measured using a fluorescence plate reader 1 h after injection. RFU, relative fluorescence unit (AC, ; IVT, ). Data are shown as mean s.e.m. , AC sCLN; , IVT sCLN; , IVT dCLN. d, Dye was injected into the left eye through the AC or IVT route, and eyes, sCLNs and dCLNs were collected for IVIS epifluorescence imaging.
Representative background-subtracted heat maps of dye in the eye, sCLNs and dCLNs 1 h after injection are shown. e, Schematic of the anatomical locations of the sCLNs and dCLNs.f,g, sCLNs, dCLNs or both CLNs were surgically ligated. Two days later, dye was injected into the eye through the AC (f) or IVT (g) route. The percentage of dye retention in the eye was measured 12 h later. Data are shown as mean s.e.m. in and (sham AC, in all other conditions). , IVT sCLN; , IVT dCLN; NS, not significant. values were calculated using a one-way analysis of variance (ANOVA) with multiple comparisons testing (Dunnett). The graphics in a,e were created with BioRender.com.
(Extended Data Fig. 8a). Consistently, IVT-injected LYVE1 antibodies could effectively label optic nerve sheath lymphatics (Extended Data Fig. 8b,c), suggesting that macromolecules such as proteins can freely penetrate into these lymphatics from the vitreous. Our leading hypothesis of how molecules travel into these lymphatics was based on an understanding of the dural lymphatics of the brain . We reasoned that molecules from the eye pass through the CSF into the optic nerve sheath lymphatics, similar to how molecules move from the brain to the dural lymphatics. To test this, we examined the distribution of dye and antibodies after IVT injection. We detected a trace amount of dye in the CSF after IVT injection compared to after brain intraventricular injection (Extended Data Fig. 8d). Furthermore, antibodies injected by the IVT route were not able to sufficiently label the dural lymphatics. These data indicated that IVT-injected dye probably entered the local CSF space around the optic nerve and did not drain to the dCLNs through previously characterized conventional brain dural meninges (Extended Data Fig. 8e). To parse out how macromolecules reach the lymphatic vasculature around the optic nerve sheath, we tracked how molecules injected into the vitreous left the eye in vivo. After IVT AF647-OVA injection, we observed that there was directional flow towards the optic nerve head (Extended Data Fig. 8f), consistent with previous findings that the pressure gradient between intraocular and i.c. spaces drives efflux of ocular fluid through the optic nerve . Examination at a higher resolution after sectioning the eye revealed that the tracer was largely concentrated
at the optic nerve head (Extended Data Fig. 8g), with some diffusion through the retinas. The diffusion through the retina showed a pattern similar to the previously described glymphatic patterns of the retinas, a paracellular perpendicular streak across the retina (Extended Data Fig. 8h).
To validate the ability of these lymphatics to drain the vitreous, we delivered VEGFC, a lymphatic stimulator , through three different compartmentalized routes: AC (targeting lymphatics of the anterior eye), IVT (targeting the posterior eye, optic nerve and possibly the anterior eye) or intracisterna magna (ICM; targeting optic nerve). First, using fluorescent tracers, we confirmed that ICM-injected tracers had access only to the optic nerve and not to the eye (Fig.3d,e). Two days after VEGFC delivery, fluorescent dextrans were injected by the IVT route, and retention was analysed 12 h later (Fig. 3f). AC administration of VEGFC did not change dextran drainage from the vitreous, indicating that the anterior lymphatic stimulation is not sufficient in driving vitreous drainage. By contrast, IVT and ICM administration of VEGFC increased dextran drainage from the vitreous, the latter in a dose-dependent manner (Fig. 3g), supporting the observation that the posterior compartment of the eye drains to the dCLNs through lymphatic vasculature on the optic nerve sheath. To further confirm the role of VEGFC in modulating optic nerve sheath lymphatics, we also delivered recombinant adeno-associated virus (rAAV)-VEGFC or soluble VEGFR3 (sVEGFR3) through the IVT route and found that expression of VEGFC or sVEGFR3 resulted in significant increases or decreases in
Fig. 3|Optic nerve sheath lymphatics drain the posterior eye.
a,b, Immunostaining of sections of optic nerve from zebrafish with lymphatics labelled for Mrc1a (white arrows; a, top left) and iDISCO immunolabelling of meningeal lymphatic vessels of rabbit (a, top right), pig (a, bottom left), non-human primate (a, bottom right) and human (b) optic nerve and chiasma, with lymphatics showing colocalization of LYVE1 and VEGFR3 (white arrows). DAPI, 4′,6-diamidino-2-phenylindole.c, Whole-mount wild-type mouse optic nerve sheaths stained for LYVE1, CD31, PROX1 and VEGFR3. The images at the bottom show a higher-magnification view of the area highlighted in the merged image at the top right. Scale bars, (a, top left; c, bottom merged), (a, top right; c, top merged), , bottom left and right) and .
d, Schematic of ICM injection and the eye and optic nerve dissection for e.e, Dye was ICM injected, and fluorescence signal intensity was measured 1 h later in the eye and optic nerve (control, ; CSF injection, ). **** . f, Schematic of injection methods for , VEGFC was injected through the AC, IVT or ICM administration route. Two days later, dye was IVT injected into the eye, and the percentage of dye retention was measured 12 h after dye injection (control, ; AC, ; IVT, ; CSF ( ), ; CSF ( ), ) . Data are shown as mean s.e.m. values were calculated using a one-way ANOVA with multiple comparisons testing (Dunnett) or two-tailed unpaired Student’s -test. The graphics in d,f were created with BioRender.com.
optic nerve sheath lymphatic vasculature, respectively (Extended Data Fig. 9a). In summary, these data highlight the existence of a functional lymphatic vessel network on the optic nerve sheath that connects the posterior eye to a set of LNs unique from the rest of the eye but shared with the rest of the CNS.
Enhancing gene therapy via ocular lymphatics
Having confirmed the immunological importance of this posterior ocular drainage, we next investigated potential therapeutic applications that harness this finding. rAAVs are the most common in vivo gene delivery vectors owing to their safe and effective transduction . Additionally, using AAV2 as a vector to treat patients with RPE65-mediated inherited retinal dystrophy was the first gene therapy approved by the US Food and Drug Administration . However, despite its modified immunological properties, acquired immunity to rAAV limits the efficacy of its repeated administration . We first characterized the rAAV-specific CD8 T cell induction in LNs and retinas after AC and IVT administration of rAAV using an ELISpot assay. Although both AC and IVT injection induced a similar amount of rAAV-specific CD8 T cells in the sCLNs, IVT injection specifically induced more rAAV-specific CD8 T cells in the dCLNs and the retinas, which were not observed after the AC injection (Fig. 4a,b). We reasoned that lymphatic drainage on
the optic nerve contributes to the immunological response to rAAV, especially for retina-based therapeutic approaches, thereby limiting its utility. We examined whether ablation of ocular lymphatic vessels could enable prolonged therapeutic efficacy after repeated rAAV injection into the eye. To test this, we first administered the primary dose of rAAV and, 1 month later, administered a second dose of RFP-expressing rAAV (rAAV-RFP). As a baseline control, naive mice were given rAAV-RFP at the time of the second dose (Fig. 4c). Similar to the findings of previous reports and consistent with our observation regarding local CNS immunity, the level of RFP expression after the secondary infection was significantly lower than that after the primary infection (Fig. 4d,e). The importance of ocular lymphatic vessels was confirmed by carrying out a similar experiment following either sCLN or dCLN ligation, which demonstrated that only dCLN ligation was sufficient in mitigating the immune response to rAAV and recovering secondary rAAV-RFP transduction efficiency (Fig. 4d,e). Additionally, we confirmed that the ligation procedure did not affect the primary challenge (Extended Data Fig. 9b). As surgically ligating the LNs would not be a therapeutically viable option for treatment, we used an alternative method to inhibit the molecular activity of lymphatics: IVT administration of sVEGFR3, which we found to inhibit lymphatic drainage to a similar extent as surgical ligation (Extended Data Fig. 9c). We saw that immune response to rAAV was further exacerbated by the presence of VEGFC, which is
Fig. 4 | Lymphatic inhibition enables repeat rAAV administration. a,b, Mice were injected with rAAV-RFP through the IVT or AC route. Their dCLNs, sCLNs and retinas were collected 10 days later, and rAAV-specific immune responses were quantified using an ELISpot assay. For , C57BL/6J mice were IVT injected with rAAV, and, 1 month later, were rechallenged with rAAV-RFP. The efficiency of rAAV-RFP transduction was analysed by imaging 1 month later. c, Schematic of experimental plans. d, In vivo fluorescence fundus imaging to visualize vessels (green) and RFP transduction (red) in different LN ligation conditions. Scale bar, . e, Quantification of RFP intensity from d (primary, ; secondary, ; sCLN ligation, ; dCLN ligation, ).
known to increase immune responses by stimulating lymphatic drainage . Conversely, inhibiting VEGFC signalling during the primary rAAV transduction using sVEGFR3 allowed for the second dose to be just as effective as the primary infection (Fig. 4f,g and Extended Data Fig. 10a-c). These experiments establish the role of posterior ocular lymphatics in modulating the efficacy of rAAV-based gene therapies and suggest a platform to overcome current barriers limiting repeated gene therapy. , primary versus secondary; , secondary versus dCLN ligation.f, In vivo fluorescence fundus imaging to visualize vessels (green) and RFP transduction (red) with addition of VEGFC or sVEGFR3. Scale bar, . , Quantification of RFP intensity from (primary, ; secondary, ; VEGFC, ; sVEGFR ). , primary versus secondary; , secondary versus VEGFC; , secondary versus sVEGFR3. values were calculated using a one-way ANOVA with multiple comparisons testing (Dunnett). Data are shown as mean s.e.m in b, e and g. The graphics in c were created with BioRender.com.
Discussion
We have demonstrated that the posterior compartment of the eye has a unique lymphatic drainage system that anatomically engages with the CNS meningeal lymphatic network at the dCLNs. Previous studies have described the existence of sparse lymphatic capillaries on human and non-human primate optic nerves ; using spatial transcriptomics, several imaging techniques, genetic manipulation in vivo
and tracer-injection experiments, we mapped the larger lymphatic vasculature network in the optic nerve sheath and described its functionality in draining the vitreous to the dCLNs. These results are in accordance with the known nexus between the eye and CNS and further extend this concept by demonstrating a shared immune surveillance mechanism between the two sites (Extended Data Fig. 10d).
Previously, both the eye and brain, which are devoid of traditional lymphatic vessels, were considered immune-privileged tissues because foreign tissue grafts can survive after implantation . Furthermore, antigens delivered to both the anterior and posterior chamber of the eye have been reported to induce peripheral immune tolerance . However, our study revealed that pathogenic antigens delivered to the posterior eye through IVT vaccination can drain to dCLNs and initiate a local protective immune response in the brain that is dependent on cells, B cells and local antibodies in the CNS compartment. Although the use of different disease models and therapeutic interventions in our studies shows the universality of our findings, it is likely that different pathogens or diseases may skew towards specific arms of the adaptive immune response. Of note, in our experiments and those of others, it is not that the sCLNs lack antigens from the CNS; rather drainage into the sCLNs is not sufficient to mount an immune response in the CNS, and future studies uncovering the fundamental difference between these two LNs may give us better insight into the observations described here. One possibility is that sCLNs and dCLNs may differ in their ability to generate effective brain-homing T cells or brain-resident T cells, which are reported to provide protection against pathogen reinfection in the brain . Besides the protective role of local antibodies in IVT vaccination, it will be important to investigate whether brain-resident T cells play a role during challenge.
In addition, the mechanisms that guide these immune responses to the brain still need to be further investigated. The present findings imply that immune tolerance to foreign antigens in the posterior eye and brain is driven by immune suppression, both passive and active, and not due to a lack of an immune response. Thus, the posterior drainage system of the eye may play a pivotal role in modulating immune activation and suppression for various diseases involving the CNS. Current conventional methods to treat or diagnose diseases of the CNS are either invasive or fail to initiate local immunity. In cases with a suspected increased mass effect in the CNS due to infection or malignancy, it can often be dangerous to access the CSF through methods such as a lumbar puncture. Although far from clinical applicability, our studies propose the vitreous as a relatively non-invasive and more accessible route to elicit CNS immunity.
We use these concepts to illustrate that disruption of the posterior lymphatic vessels can negate immune response to rAAV delivered to the posterior eye and thus restore therapeutic benefit. Contrary to the notion of immune privilege, rAAV therapy for ocular diseases faces substantial hurdles due to the cellular and humoral immune response that it initiates . These findings further confirm that this optic nerve drainage system identified in our study is responsible for the immunological outcome in response to antigens in the vitreous humour. The results also signify that the posterior lymphatic drainage system of the eye is a versatile and accessible target for manipulating immune responses in many different contexts.
Beyond the immunological implications of the posterior drainage system, how the mechanics of the posterior drainage pathway can be harnessed to treat ocular diseases merits further study. Specifically, the identification of a posterior eye lymphatic drainage system provides a potential efficient drainage route to modulate intraocular pressure or remove unwanted fluid in disease states such as glaucoma or macular oedema, respectively. Also, with the recently described presence of the glymphatic system in the eye , more work is required to elucidate how these pathways interact with each other to provide homoeostasis in the eye, similar to in the brain. Together, the
findings of the present study establish that the connection between the posterior eye and the brain is not only a neuronal one but also a shared immune circuit that coordinates immunity from the periphery to the CNS.
Why this compartmentalized lymphatic drainage system in the eye has evolved poses an interesting question. One possibility is that the centralization of the CNS and the emergence of lymphatic networks in vertebrates drove the polarization of the two lymphatic drainage systems in the eye. A clue to this may be in the lateralization that we observe during drainage to the sCLNs but not dCLNs, which is probably a consequence of the anatomical separation of skin, muscle and tissue surrounding the orbit. Contrarily, bilateral drainage to the dCLNs from the posterior eye occurs through a network of CSF-draining lymphatics. The anterior lymphatics act in synchrony with the cornea and conjunctiva, a generalized mucosal surface with constant exposure to outside invaders that may never have access to the brain. The optic nerve lymphatics we describe here connect the posterior eye (retina) with the brain, allowing coordinated immune responses that may prepare the brain for imminent threats. The segregation of eye immune responses into anterior and posterior lymphatic networks may predate mammals, and future studies in other vertebrates may help illuminate when and why this innovation first emerged.
Online content
Any methods, additional references, Nature Portfolio reporting summaries, source data, extended data, supplementary information, acknowledgements, peer review information; details of author contributions and competing interests; and statements of data and code availability are available at https://doi.org/10.1038/s41586-024-07130-8.
London, A., Benhar, I. & Schwartz, M. The retina as a window to the brain-from eye research to CNS disorders. Nat. Rev. Neurol. 9, 44-53 (2013).
Marcocci, M. E. et al. Herpes simplex virus-1 in the brain: the dark side of a sneaky infection. Trends Microbiol. 28, 808-820 (2020).
Iijima, N. & Iwasaki, A. Access of protective antiviral antibody to neuronal tissues requires CD4 T-cell help. Nature 533, 552-556 (2016).
Victora, G. D. & Nussenzweig, M. C. Germinal centers. Annu. Rev. Immunol. 40, 413-442 (2022).
Cyster, J. G. & Allen, C. D. C. B cell responses: cell interaction dynamics and decisions. Cell 177, 524-540 (2019).
Shih, T.-A. Y., Roederer, M. & Nussenzweig, M. C. Role of antigen receptor affinity in T cellindependent antibody responses in vivo. Nat. Immunol. 3, 399-406 (2002).
Koedel, U., Scheld, W. M. & Pfister, H.-W. Pathogenesis and pathophysiology of pneumococcal meningitis. Lancet Infect. Dis. 2, 721-736 (2002).
Iovino, F., Seinen, J., Henriques-Normark, B. & van Dijl, J. M. How does Streptococcus pneumoniae invade the brain? Trends Microbiol. 24, 307-315 (2016).
Song, E. et al. VEGF-C-driven lymphatic drainage enables immunosurveillance of brain tumours. Nature 577, 689-694 (2020).
Thomson, B. R. et al. A lymphatic defect causes ocular hypertension and glaucoma in mice. J. Clin. Investig. 124, 4320-4324 (2014).
Park, D.-Y. et al. Lymphatic regulator PROX1 determines Schlemm’s canal integrity and identity. J. Clin. Investig. 124, 3960-3974 (2014).
Aspelund, A. et al. The Schlemm’s canal is a VEGF-C/VEGFR-3-responsive lymphatic-like vessel. J. Clin. Investig. 124, 3975-3986 (2014).
Goel, M., Picciani, R. G., Lee, R. K. & Bhattacharya, S. K. Aqueous humor dynamics: a review. Open Ophthalmol. J. 4, 52-59 (2010).
Johnson, M., McLaren, J. W. & Overby, D. R. Unconventional aqueous humor outflow: a review. Exp. Eye Res. 158, 94-111 (2017).
Wang, X. et al. An ocular glymphatic clearance system removes beta-amyloid from the rodent eye. Sci. Transl. Med. 12, eaaw3210 (2020).
Kasi, A., Liu, C., Faiq, M. A. & Chan, K. C. Glymphatic imaging and modulation of the optic nerve. Neural Regen. Res. 17, 937-947 (2022).
Tietjen, G. T., DiRito, J., Pober, J. S. & Saltzman, W. M. Quantitative microscopy-based measurements of circulating nanoparticle concentration using microliter blood volumes. Nanomedicine 13, 1863-1867 (2017).
Camelo, S., Kezic, J., Shanley, A., Rigby, P. & McMenamin, P. G. Antigen from the anterior chamber of the eye travels in a soluble form to secondary lymphoid organs via lymphatic and vascular routes. Investig. Ophthalmol. Vis. Sci. 47, 1039-1046 (2006).
Tam, A. L. C., Gupta, N., Zhang, Z. & Yücel, Y. H. Quantum dots trace lymphatic drainage from the mouse eye. Nanotechnology 22, 425101 (2011).
Lee, J. Y. et al. Structural confirmation of lymphatic outflow from subconjunctival blebs of live humans. Ophthalmol. Sci. 1, 100080 (2021).
Yücel, Y. H. et al. Active lymphatic drainage from the eye measured by noninvasive photoacoustic imaging of near-infrared nanoparticles. Investig. Ophthalmol. Vis. Sci. 59, 2699-2707 (2018).
Renier, N. et al. iDISCO: a simple, rapid method to immunolabel large tissue samples for volume imaging. Cell 159, 896-910 (2014).
Xu, H., Chen, M., Reid, D. M. & Forrester, J. V. LYVE-1-positive macrophages are present in normal murine eyes. Investig. Ophthalmol. Vis. Sci. 48, 2162-2171 (2007).
Makinen, T. et al. PDZ interaction site in ephrinB2 is required for the remodeling of lymphatic vasculature. Genes Dev. 19, 397-410 (2005).
Louveau, A. et al. Structural and functional features of central nervous system lymphatic vessels. Nature 523, 337-341 (2015).
Aspelund, A. et al. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 212, 991-999 (2015).
Breslin, J. W. et al. Vascular endothelial growth factor-C stimulates the lymphatic pump by a VEGF receptor-3-dependent mechanism. Am. J. Physiol. Heart Circ. Physiol. 293, H709-H718 (2007).
Jeltsch, M. et al. Hyperplasia of lymphatic vessels in VEGF-C transgenic mice. Science 276, 1423-1425 (1997).
Mathieu, E. et al. Evidence for cerebrospinal fluid entry into the optic nerve via a glymphatic pathway. Investig. Ophthalmol. Vis. Sci. 58, 4784-4791 (2017).
Wang, D., Tai, P. W. L. & Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat. Rev. Drug Discov. 18, 358-378 (2019).
Russell, S. et al. Efficacy and safety of voretigene neparvovec (AAV2-hRPE65v2) in patients with RPE65-mediated inherited retinal dystrophy: a randomised, controlled, open-label, phase 3 trial. Lancet 390, 849-860 (2017).
Colella, P., Ronzitti, G. & Mingozzi, F. Emerging issues in AAV-mediated in vivo gene therapy. Mol. Ther. Methods Clin. Dev. 8, 87-104 (2018).
Mingozzi, F. & High, K. A. Immune responses to AAV vectors: overcoming barriers to successful gene therapy. Blood 122, 23-36 (2013).
Killer, H. E., Laeng, H. R. & Groscurth, P. Lymphatic capillaries in the meninges of the human optic nerve. J. Neuroophthalmol. 19, 222-228 (1999).
Sherman, D. D. et al. Identification of orbital lymphatics: enzyme histochemical light microscopic and electron microscopic studies. Ophthalmic Plast. Reconstr. Surg. 9, 153-169 (1993).
Streilein, J. W. Ocular immune privilege: therapeutic opportunities from an experiment of nature. Nat. Rev. Immunol. 3, 879-889 (2003).
Medawar, P. B. Immunity to homologous grafted skin; the fate of skin homografts transplanted to the brain, to subcutaneous tissue, and to the anterior chamber of the eye. Br. J. Exp. Pathol. 29, 58-69 (1948).
Camelo, S., Shanley, A., Voon, A. S. P. & McMenamin, P. G. The distribution of antigen in lymphoid tissues following its injection into the anterior chamber of the rat eye. J. Immunol. 172, 5388 (2004).
Boonman, Z. F. H. M. et al. Intraocular tumor antigen drains specifically to submandibular lymph nodes, resulting in an abortive cytotoxic T cell reaction. J. Immunol. 172, 1567 (2004).
Sonoda, K.-H. et al. The analysis of systemic tolerance elicited by antigen inoculation into the vitreous cavity: vitreous cavity-associated immune deviation. Immunology 116, 390-399 (2005).
Ayasoufi, K. et al. Brain resident memory T cells rapidly expand and initiate neuroinflammatory responses following CNS viral infection. Brain Behav. Immun. 112, 51-76 (2023).
Steinbach, K. et al. Brain-resident memory T cells represent an autonomous cytotoxic barrier to viral infection. J. Exp. Med. 213, 1571-1587 (2016).
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
(c) The Author(s) 2024
Methods
Mice
Six- to ten-week-old mixed-sex C57BL/6J,B6.Cg-Tg(TcraTcrb)425Cbn/J (OT-II), B6.129P2(C)-Ightm2Cgn/J (B1-8) and B6.129S2-IghtmICgn/J ( ) mice were purchased from Jackson Laboratory and Charles River and subsequently bred and housed at Yale University. PROX1-CreER ; CDH5-Dre;R26-STOP-mCherry and VEGFR3-CreER ;R26-mTmG mice were gifts from the laboratory of J.-L.T. All procedures used in this study complied with federal guidelines and the institutional policies of the Yale School of Medicine Animal Care and Use Committee. Age- and sex-matched animals were randomly assigned to control or treatment groups in each experiment. No statistical methods were used to predetermine sample sizes. Sample sizes were empirically determined based on previously published studies and to ensure sufficient statistical power. Investigators were not blinded to experimental groups, as measurements were not subjective.
Cells
GL261-Luc cells were a gift from J. Zhou (Yale Neurosurgery) and were cultured in RPMI supplemented with 10% FBS, 1% penicillinstreptomycin and 1% sodium pyruvate. CT2A-BFP cells were a gift from T. Mathivet (Paris Centre de Recherche Cardiovasculaire). B16 cells were a gift from N. Palm (Yale Immunobiology). Cells were negative for mycoplasma contamination.
Bacteria and viruses
The HSV-1 KOS strain and HSV-2 strains 186syn TK and 186syn were gifts from D. Knipe (Harvard Medical School). These viruses were propagated and titrated on Vero cells (ATCCCCL-81) as previously described . Vero cells were negative for mycoplasma contamination.
S. pneumoniae (ATCC 6303) was grown on 10% sheep blood agar plates (BD Biosciences) overnight at . These colonies were then transferred to Todd Hewitt broth and grown overnight. The number of bacteria was enumerated using the optical density at 600 nm .
Optic nerve tissues and fluorescence immunocytochemistry
Tg(mrc1a:eGFP) adult male and female 1-year-old zebrafish ( ) were provided by B.W. Weinstein at the US National Institutes of Health (Bethesda, MD). Zebrafish were euthanized by an overdose of MS-222 and whole heads were excised and fixed in 60 mM HEPES-buffered 4% PFA ( pH 7.4 ) overnight at room temperature. Samples were then decalcified in EDTA solution ( pH 7.4 ) for 5 days at room temperature. Heads were then snap frozen in OCT and -thick cryosections were obtained. Cryosections containing the optic nerve were stained with the primary antibodies mouse anti-zebrafish Zns2 (ZIRC, ZDB-ATB-081002-34; 1:50) and chicken anti-GFP (Aves, catalogue no. GFP-1010; 1:500) followed by the secondary antibodies AF488-conjugated goat anti-mouse IgG (Jackson Immuno 115-545146) and Cy5-conjugated donkey anti-chicken IgY (Jackson Immuno 703-175-155) with DAPI ( ) for nuclear staining. Images were acquired in a Zeiss LSM.
Eyes from rabbits and pigs were collected post-mortem from animals euthanized owing to unrelated health conditions. Eyes were removed within an hour of death and immersion fixed in neutral-buffered formalin. Intact eyes from two female rhesus macaques (Macaca mulatta), between 25-28 years of age and euthanized owing to unrelated health conditions were collected post-mortem. Human optic nerves and chiasmas were obtained through the Yale Pathology Tissue Services through the Tissue Procurement and Distribution service. A mini standard operating procedure was written and approved for these samples. Eyes were removed within an hour of death and immersion fixed in 10% neutral-buffered formalin. Optic nerves with meningeal sheaths were cut and processed through the iDISCO protocol below.
Mouse optic nerve sheaths were dissected after mice were anaesthetized and transcardially perfused with cold PBS and PFA (Sigma-Aldrich) sequentially. Mice were first decapitated, and the skull was exposed by cutting off the skin and scalp. One midline cut down the skull and two transverse cuts on both sides of the skull were made. Tweezers were used to peel and remove both halves of the skull to expose the brain. The brain was lifted from the posterior, and vasculature connecting the brain to other parts of the head was cut until the brain could be lifted enough to expose the optic tract. Brain tissue was removed after cutting just anteriorly to the optic chiasma. The extracranial optic canal was unroofed by removing parts of the skull above the eyes to expose the intracanalicular and canal segments of the optic nerve. Tissue surrounding the optic nerve was carefully dissected to free the nerve, and the eye was cut off at the optic nerve head. After removing the optic nerve, the optic nerve sheath was cut along the length of the nerve and removed for staining and whole-mount imaging.
The PROX1-CreER ;CDH5-Dre;R26-STOP-mCherry and VEGFR3CreER ; R26-mTmG mice were injected through the i.p. route with tamoxifen ( ; Sigma-Aldrich, T5648) for 5 consecutive days and optic nerve sheaths were collected 2 days later as above. Then optic nerve sheaths were fixed in 1% PFA for 1 h and immediately processed in a blocking solution (10% normal donkey serum,1% bovine serum albumin, 0.3% PBS-Triton X-100) for 1 h at room temperature. For detection of lymphatic vessels, samples were incubated with primary antibodies overnight at , and then washed five times at room temperature in PBS with 0.5% Triton X-100, before incubation with fluorescence-conjugated secondary antibodies diluted in PBS with 5% normal donkey serum. Lymphatic vessel images were acquired using a Leica confocal microscope (Stellaris 8). The following antibodies were used: goat anti-mouse VEGFR3 (R&D, No. AF743, 1:400), rat anti-mouse LYVE1 (R&D, No. MAB2125, 1:400), Syrian hamster anti-mouse podoplanin (BioLegend,127402,1:500), rabbit anti-Prox1 (Angiobio, 11-002 P, 1:200), Armenian hamster anti-mouse CD31 (Gene Tex, 2H8, 1:1,000). The primary antibodies were detected with appropriate AF405-, AF488-, AF555- and AF647-conjugated secondary antibodies (Thermo Fisher, 1:500) after 2 h of incubation at room temperature. ProLong Gold Antifade Mountant (Invitrogen, P36930) was used for mounting the sections.
For brain tissues, tissues were collected and incubated in a digestion cocktail containing collagenase D (Roche) and DNase I (Sigma-Aldrich) in RPMI at for 45 min . Tissues were pipetted to break tissue down and filtered through a filter. Then, cells were mixed in 3 ml of Percoll (Sigma-Aldrich) solution and centrifuged at 580 g for 15 min without brake. The Percoll layer was removed, and cell pellets were treated with 0.5 ml ACK buffer and spun for 5 min at 500 g . Then, the cell pellets were resuspended in FACS buffer (PBS + 2% FBS + 1 mM EDTA) for staining.
When analysing lymphocytes, an LN or a spleen was put in a 15 mm Petri dish containing 2 ml FACS buffer and was ground between two frosted microscope slides. When analysing DCs, an LN or a spleen was digested as above. Cell suspensions were filtered through a filter and spun for 5 min at 500 g . Then, the cell pellets were resuspended in FACS buffer for staining.
Flow cytometry
Preparation of single-cell suspensions from spleens, LNs and brains is described above. Nonspecific binding was blocked using an Fc receptor-blocking solution (TruStain FcX, BioLegend, 101320, 1:200) for 10 min at before immunostaining. Subsequently, the cells were stained with corresponding antibodies for 30 min at . Then, cells were washed to remove excess antibodies and resuspended in FACS buffer. Samples were run on an Attune NxT flow cytometer and then analysed using FlowJo software (10.8.1, Tree Star).
Enzyme-linked immunosorbent assay
CSF and serum were collected from mice as previously described . The serum and CSF were then diluted with PBS containing BSA in a ratio. Plates (96-well) were coated with of heat-inactivated or PFA-inactivated purified HSV-2 ( to plaque-forming units equivalent per ) for virus-specific immunoglobulin measurement or goat anti-mouse immunoglobulin (SouthernBiotech, 1010-01, 1:1,000) and then incubated overnight at . These plates were then washed with PBS-Tween 20 and blocked for 2 h with 5% FBS in PBS. Samples were then plated in the wells and incubated for at least 4 h at room temperature. After being washed in PBS-Tween 20, HRP-conjugated anti-mouse immunoglobulin antibodies (SouthernBiotech, 1010-05, 1:5,000) were added in the wells for 1 h , followed by washing and addition of TMB solution (eBioscience). Reactions were stopped with 1 N , and absorbance was measured at 450 nm . The total antibody titres were defined by using an immunoglobulin standard (C57BL/6 mouse immunoglobulin panel; SouthernBiotech).
Western blot
rAAV-RFP-infected retinas or control retinas were digested with cocktail containing collagenase D (Roche) and DNase I (Sigma-Aldrich) in RPMI at for 45 min . Tissues were pipetted to break tissue down and filtered through a filter. Then, cells were lysed in RIPA buffer and boiled for 5 min with sample buffer. Western blotting was carried out in a similar manner to that previously reported . In brief, gels were used and run at 10 mA per gel for 30 min and 40 mA per gel until ladder separation. Wet transfer was carried out at 120 mA pergel for 90 min on ice. RFP-Tag rabbit polyclonal antibodies (OriGene Technologies, catalogue no. AP09229PU-N) were used at a concentration of 1:1,000 and incubated overnight in a cold room. After being washed, HRP-conjugated anti-rabbit IgG secondary antibodies (Thermo Fisher, G-21234) were used at a concentration of 1:5,000 at room temperature for 2 h and imaged using the ChemiDoc MP imaging system (Bio-Rad).
ELISpot assay
Mice were injected with rAAV-RFP through the IVT or AC route. Their dCLNs, sCLNs and retinas were collected 10 days later. Single-cell suspensions were prepared and co-cultured with splenocytes at a ratio of 1:5 with the presence of rAAV-RFP virus peptides (SNYNKSVNV and NGRDSLVNPGPAMAS).rAAV-specific immune responses were quantified using an ELISpot assay (mouse IFN ELISpot Kit; R&D, catalogue no. EL485), following the manufacturer’s instructions for the assay.
Flank tumour inoculation and treatment
Mice were anaesthetized using a mixture of ketamine ( ) and xylazine ( ), and the flank was shaved and disinfected. A syringe with a 30-G needle was used to deliver of 500,000 B16 cells subcutaneously. Then, mice were treated with irradiated B16 cells
(250,000 cells) through s.c., i.c., AC or IVT administration routes (day 7) along with anti-PD1 (RMP1-14) antibodies (days 7, 9 and 11) through the i.p. route, and their survival was monitored.
Adoptive transfer
To directly analyse the immune response (antigen-specific T and B cells) in dCLNs and sCLNs after IVT or AC injection, we transferred OT-II and B1-8 cells and analysed their response after different immunization routes.
To analyse antigen-specific B cell response, we followed a previously reported method . In brief, CD45.2 C57BL/6 recipient mice (6-8 weeks of age) were primed by i.p. immunization with 50 mg of OVA (Sigma, A5503) precipitated in alum at a 2:1 ratio in PBS. Two weeks later, resting cells were isolated from CD45.1.2 B1-8 mice with a mouse B cell isolation kit (Stemcell, 19854). Then, the B cells were labelled with CellTrace Violet Cell Proliferation Kit (Thermo Fisher, C34557). A total of 5 million cells were transferred intravenously into recipient mice. Eight hours later, the mice were immunized with NP20-OVA (Biosearchtech, ) through IVT or AC injection. cell proliferation and germinal centre formation in dCLNs and sCLNs were analysed at day 7.
For antigen-specific T cell response, OVA-specific CD4 T cells were isolated from CD45.1 OT-II mice with mouse CD4 T Cell Isolation Kit (Stemcell, 19852). Then, the CD4 T cells were labelled using the CFSE Cell Proliferation Kit (Thermo Fisher, C34554). A total of 5 million cells were transferred intravenously into CD45.2 recipient mice. Eighteen hours later, the mice were immunized with OVA (Sigma, A5503) plus poly(I:C) (Invivogen, tlrl-picw) through IVT or AC injection. A150 quantity of FTY720 (Sigma, SML0700) was i.p. injected to inhibit the circulation of primed T cells 24 h after immunization . OVA-specific CD4 T cell proliferation in ingLNs, dCLNs and sCLNs was analysed 72 h after immunization.
IVT, AC, i.c. and ICM injection
Mice aged 6-10 weeks old were anaesthetized through i.p. injection of a mixture of ketamine ( ) and xylazine ( ). For IVT or AC injection, the eye was dilated with tropicamide ophthalmic solution. For IVT injection, a 30-G needle was used to puncture a hole at 1 mm posterior to the corneoscleral junction. A blunt-ended Hamilton syringe with of dye, HSV-1, HSV-2,S. pneumoniae or irradiated tumour cells was inserted into the vitreous humour about deep and administered at a rate of . For AC injection, the hole was punctured close to the corneoscleral junction and a blunt-ended Hamilton syringe was inserted into the AC about 1 to 2 mm deep. After IVT or AC injection, petrolatum ophthalmic ointment was applied on the eye to prevent cataract formation. The method for i.c. injection was similar to that for tumour inoculation, but l of HSV-1, HSV-2,S. pneumoniae or irradiated tumour cells was administered. ICM injection was carried out as previously described . The mice were kept on heating pads and continuously monitored until recovery after the injection.
Virus and bacteria immunization and challenge
Mice were anaesthetized with a mixture of ketamine ( ) and xylazine( ). For HSV immunization, plaque-forming units of heat-inactivated HSV-1 or HSV-2 were AC, IVT, i.p. or i.c. injected. Thirty days later, these mice were challenged through the i.c. route with HSV-1 or HSV-2 and their survival was monitored. For some experiments, CD4 T cell-depleting antibodies (BioXCell, GK1.5, No. BE0003-1, for 3 days) were injected before rechallenge. Similar experiments were carried out with S. pneumoniae; the immunization dose was of heat-inactivated bacteria and the challenge dose was of live bacteria.
Imaging and quantification of tracer transport
For quantification of fluorescence intensity in the eye or LNs, dextran conjugated to either FITC ( 40 kDa and 70 kDa ) or tetramethylrhodamine ( 40 kDa and 70 kDa ) was injected into the AC or vitreous humour
through AC or IVT injection, respectively. To analyse the kinetics of dye drainage from the eye, the eye was collected at serial time points and was homogenized in PBS using the bead beating method (Lysing Matrix D, 116913500, MP Biomedicals). Then, homogenized tissue was centrifuged at for 10 min and supernatant was collect into a 96-well plate and fluorescence intensity was read with emission and excitation wavelengths of 494 nm and 514 nm or 555 nm and 585 nm . To measure the dye drainage into and dCLNs were collected 12 h after dye injection. The fluorescence intensity was measured as above.
For measuring nanoparticle draining in the serum, we used a previously published protocol . In brief, serum from mice was isolated and placed on microscope slides to allow small-volume high-sensitivity detection of tracer transport in the blood.
For IVIS imaging of tracer transport, the eye, sCLNs and dCLNs were collected after dye injection through either an AC or IVT administration route. They were imaged using the IVIS Spectrum In Vivo Imaging System (PerkinElmer).
For IVT tracer transport in vivo, , Thermo Fisher, O34784) was IVT injected into the vitreous humour. A volume of sodium fluorescein ( , Santa Cruz, sc-206026) was injected into blood to label blood vessels. The kinetics of AF647-OVA drainage was tracked with the Phoenix MICRON IV imaging microscope.
For analysing the co-localization of optic nerve sheath lymphatics with IVT tracer, anti-mouse LYVE1 antibody (R&D, MAB2125) was IVT injected into the vitreous humour. Optic nerve sheaths were collected 2 h later and stained as above.
For tracking the kinetics of dye draining in the eye, AF647-OVA (AF647-OVA ( ; Thermo Fisher, O34784) was IVT injected into the vitreous humour. The eyes were enucleated at the indicated time points and processed in a manner similar to that previously reported . In brief, eyes were fixed in Hartman’s fixation buffer, and three windows on the eye were created using the previously described window technique . Serial sections were obtained using a cryostat (Leica CM190) following dehydration of the tissue in a sucrose gradient up to sucrose. The sections were mounted with ProLong Gold Antifade Mountant with DAPI (Invitrogen, P36931) and imaged with a Leica confocal microscope (Stellaris 8).
CSF collection
For CSF collection, mice were anaesthetized through i.p. injection of a mixture of ketamine ( ) and xylazine ( ). The dorsal neck was shaved and sterilized. A1-cm incision was made at the base of the skull, and the dorsal neck muscles were separated using forceps to expose the cisterna magna. A custom-pulled micropipette (0.75/11brl GF; Stoelting) was used to penetrate the dura to collect CSF.
Brain tumour inoculation and IVIS imaging
Tumour inoculation was carried out as previously described with slight modifications . Mice were anaesthetized through i.p. injection using a mixture of ketamine ( ) and xylazine ( ). Mouse heads were shaved and scalps were sterilized. A midline scalp incision was made and a burr hole was drilled 2 mm lateral to the sagittal suture and 0.5 mm posterior to the bregma with a 25-G needle. Then, mice were placed in a stereotaxic frame. A10- l Hamilton syringe loaded with GL261-Luc cells ( cells) was inserted into the burr hole at a depth of 4 mm from the surface of the brain and left to equilibrate for 1 min before infusion. A micro-infusion pump (World Precision) was used to infuse at . The syringe was left in place for another minute after the infusion was finished. The skin was stapled and cleaned. Following intramuscular administration of an analgesic (meloxicam and buprenorphine, ), mice were placed in a heated cage until full recovery. We tracked tumour size weekly thorough IVIS imaging. Mice were anaesthetized using isoflurane and injected through the i.p. route with D-luciferin potassium salt bioluminescent substrate (PerkinElmer, . After 10 min , mice were imaged using the IVIS Spectrum In Vivo Imaging System (PerkinElmer).
Preparation of tissue for analysis of CT2A-BFP tumour antigen drainage
i.c. CT2A-BFP tumours were analysed 14 days after injection. Tumours, meninges, IngLNs, dCLNs and sCLNs were collected. Mononuclear cells were isolated and stained.
Parabiosis
Parabiosis was carried out as previously described with slight modifications . Naive or IVT-immunized C57BL/6 mice of similar age and weight were anaesthetized with a mixture of ketamine ( ) and xylazine ( ). After shaving the corresponding lateral aspects of each mouse, the skin was cleaned and sterilized with an alcohol prep pad and Betadine surgical scrub. Matching skin incisions were made from just above the knee upwards to the olecranon, and two mice were sutured together with Ethicon 5-0 coated Vicryl absorbable sutures. Then, the skin was stapled and Neosporin + Pain Relief Ointment was applied on the incisions. During the surgery, mice were kept on heating pads and continuously monitored until recovery.
Ligation of sCLNs or dCLNs
For ligation of LNs, mice were anaesthetized with a mixture of ketamine ( ) and xylazine ( ) and the rostral neck was shaved and sterilized. A 2 -cm incision was made, and the sCLNs and dCLNs were sequentially exposed using forceps. Their afferent lymph vessels were cauterized or kept intact on the basis of the experiment conditions. Then, the incision was closed with a 5-0 Vicryl suture, and mice were subjected to the same postoperative procedures as above.
rAAV transduction and imaging
Wild-type mice were IVT injected with rAAV ( viral genomes) with PBS, VEGFC ( ) or sVEGFR3 ( ). Then, 1 month later, these mice were rechallenged with rAAV-RFP ( viral genomes). Eyes were either imaged on the Phoenix Micron IV or collected 1 month after rAAV-RFP transduction and fixed with PFA overnight at . The retina whole mount was carefully dissected and imaged with a Leica confocal microscope (Stellaris 8).
Three-dimensional imaging of solvent-cleared organs
iDISCO was carried out as previously described (http://www.idisco. info) . The following antibodies were used: goat anti-mouse VEGFR3 (R&D, No. AF743,1:400), rat anti-mouse LYVE1 (R&D, MAB2125,1:400), rabbit anti-mouse LYVE1 (AngioBio, No. 11-034,1:200), mouse anti-human VEGFR3 (Santa Cruz Biotechnology, SC-28297, 1:200), rabbit anti-human LYVE1 (Angio-Proteomie, 102-PA50S, 1:200), goat anti-mouse IgG-AF647 (Invitrogen, A21235, 1:500), donkey anti-goat IgG-AF647 (Invitrogen, A21447, 1:500), goat anti-rabbit IgG-AF555 (Invitrogen, A21428, 1:500). Subsequently, the transparent optic nerves with optic nerve sheaths were imaged using a Leica confocal microscope (Stellaris 8). Three-dimensional rendering was completed using Imaris 8 software (Oxford Instruments).
Spatial transcriptomics
The 10X Visium Spatial Gene Expression for FFPE slide (PN-1000185) and associated protocols (CG000483) from 10X Genomics were used. Mouse eyeballs were fixed , processed and sectioned as previously described. Transverse sections of in thickness of eyeball were cut using a microtome (RM2255, Leica Biosystems) and carefully placed within the fiducial frame on the Visium slide, and then sections were air dried at room temperature overnight and stored in a desiccant container before spatial transcriptomics experiment. The FFPE sections were baked, stained with haematoxylin-eosin and then imaged using a Keyence bz-x800 all-in-one fluorescence microscope. Then,
cell permeabilization and library preparation was carried out following the Visium Spatial Gene Expression FFPE User Guide using the supplied reagents (10X Genomics). The generated libraries were sequenced and analysed using Space Ranger (version 2.1.0), and data were analysed using Seurat 4.9.9.9040.
Image processing and analysis
Quantitative analysis of rAAV-infected cells was carried out using either FIJI or ImageJ image-processing software (NIH or Bethesda).
Statistical analysis
All statistical analyses were carried out using GraphPad Prism software. Data were analysed with a two-tailed unpaired Student’s -test or a one-way ANOVA with multiple comparisons testing (Dunnett) with Prism software. Statistical significance is defined as , and .
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The spatial transcriptomics dataset of the mouse eye and optic nerve is publicly available at the National Center for Biotechnology Information Gene Expression Omnibus under accession number PRJNA1046563. All datasets generated and/or analysed during the current study are available in the article. Source data are provided with this paper.
Code availability
All of the code used for analysis is described in the Methods. Detailed files will be made available upon on request.
43. Iijima, N. et al. Dendritic cells and B cells maximize mucosal Th1 memory response to herpes simplex virus. J. Exp. Med. 205, 3041-3052 (2008).
44. Jung, H. M. et al. Development of the larval lymphatic system in zebrafish. Development 144, 2070-2081 (2017).
45. Gitlin, A. D., Shulman, Z. & Nussenzweig, M. C. Clonal selection in the germinal centre by regulated proliferation and hypermutation. Nature 509, 637-640 (2014).
46. Ciabattini, A. et al. Distribution of primed T cells and antigen-loaded antigen presenting cells following intranasal immunization in mice. PLoS ONE 6, e19346 (2011).
47. Pang, J. et al. Step-by-step preparation of mouse eye sections for routine histology, immunofluorescence, and RNA in situ hybridization multiplexing. STAR Protocols 2, 100879 (2021).
48. Pang, J. et al. A freeze-substitution approach with solvent-based glyoxal fixative to prevent distortion of ocular structures. J. Histotechnol. 45, 172-181 (2022).
Acknowledgements We thank N. Brown, L. Del Priore, M. Linehan, D. Kieslich and D. Ralston, who played a crucial role in establishing the infrastructures necessary for completing the manuscript, even though they were not directly involved in conducting the experiments; and all members of the Yale scientific community for their feedback through this project. The study was supported by Yale Dean’s funding (to E.S.). A.I. is an investigator of the Howard Hughes Medical Institute. J.-L.T. was supported by NIH grant R01NS1210130; W.M.S. was supported by NIH grant UH3HL147352. L.S. is supported by the Parker B. Francis Fellowship. The graphics in Figs. 1a,d, 2a,e, 3d,f and 4c and Extended Data Figs. 1d and 10d were created with BioRender.com.
Author contributions X.Y., S.Z. and J.H.L. are co-first authors of this work. X.Y. and S.Z. carried out and designed experiments in this manuscript. X.Y., S.Z. and J.H.L. wrote the manuscript. J.H.L., H.D., G.M., G.T., A.K., L.H.G., M.P., S.F., T.Z. and F.S.M. carried out experiments that contributed to this project. J.Z., Y.W., S.M., N.R., L.S., I.S., A.E., J.-L.T., W.M.S., A.H., C.Z., A.R. and A.I. contributed to materials and resources that allowed this project to be completed. E.S. conceptualized, obtained funding and supervised the work. All authors contributed to the editing of the text and figures of this manuscript.
Competing interests E.S., A.R. and A.I. are co-founders of Rho Bio. A.I. is a member of the Board of Directors of Roche Holding Ltd. E.S., J.-L.T. and A.I. are co-inventors on patent application No. 62/929,527, “Manipulation of meningeal lymphatic vasculature for brain and CNS tumour therapy”. W.M.S. is a co-founder of B3 Therapeutics, Stradefy and Xanadu Bio. W.M.S. is a consultant to Xanadu Bio, Stradefy Biosciences, Johnson & Johnson, Celanese, Cranius and CMC Pharma.
Additional information
Supplementary information The online version contains supplementary material available at https://doi.org/10.1038/s41586-024-07130-8.
Correspondence and requests for materials should be addressed to Eric Song.
Peer review information Nature thanks the anonymous reviewers for their contribution to the peer review of this work.
Reprints and permissions information is available at http://www.nature.com/reprints.
Article
a
end{figure}
h
Extended Data Fig. 1| See next page for caption.
Extended Data Fig. 1| Immunization through the posterior compartment of the eye can induce an immunological protection in the brain. a, Similar to Fig. 1a, but B cell-deficient mice ( ) were IVT injected with heat-inactivated HSV-2 and one group of WT IVT immunized mice were treated with CD4 depletion antibody before challenge. b, Similar to Fig. 1c, dCLNs were ligated after priming of the mice but before the intracranial rechallenge. Vaccination was completed first. Then, 7 days before the rechallenge, dCLNs were ligated, and mice survival was monitored after i.c. challenge with a lethal dose of HSV-2. (Naïve, ; IVT immunized, ; LN ligation, ). c, Extending findings from Fig. 1d, the serum and CSF of both IVT immunized mice and naive mice were collected 3 weeks after parabiosis. Anti-HSV specific antibody was measured by ELISA ( ). d, Schematic of experimental plans. CD45.2 C57BL/6J recipient mice were primed by immunization. 2 weeks later, CD45.1.2 B1-8 B cells labelled with CellTrace Violet Cell Proliferation Kit were transferred intravenously into recipient mice. 18 h later, the mice were immunized with NP-OVA
though IVT or AC injection. light chain positive B1- cell proliferation and germinal center (GC) formation in dCLNs and sCLNs were analyzed at day 7. , Flow cytometry gating strategy for B1- GC B cells. f, Quantification of B1-8 GC B cells in dCLNs and sCLNs ( ). g , Schematic of experimental plans. CD45.1OVA specific CD4 T cells labelled with CFSE were transferred intravenously into CD45.2 recipient mice. 18 h later, the mice were immunized with OVA plus Poly(I:C) though IVT or AC injection. FTY720 was i.p. injected to inhibit the circulation of primed T cells 24 h after immunization. The OVA-specific CD4 T cell proliferation in dCLNs and sCLNs was analyzed 72 h after immunization. h, Flow cytometry gating strategy for analyzing OVA-specific CD4 T proliferation ( ). i, Quantification of CFSE-negative OVA-specific CD4 T cells in dCLNs, sCLNs, and ingLNs. Data shown as mean s.e.m. -values were calculated using a two-tailed unpaired Student’s t-test. The graphics in d were created with BioRender.com.
Extended Data Fig. 2 | See next page for caption.
Extended Data Fig. 2 | Immunization through the posterior compartment of the eye can provide therapeutic treatment of brain tumours.a, Mice were i.c. inoculated with 50,000 GL261-Luc cells and mice were treated with irradiated GL261-Luc through the routes of s.c., i.c., AC, or IVT administration (day 7) along with anti-PD-1 antibodies (days 7, 9 and 11) and monitored for tumour growth. RLU, relative luminescence units. (s.c., , ;IVT, ). b, Mice were i.c. inoculated with 50,000 CT2A-BFP cells. The percentage of DCs in the tumour, ingLNs, meninges, dCLNs and sCLNs were analyzed 14 days after tumour injection ( ). c, d, Similar experiment as
a. The percentage of EMV-2-specific CD8 T cells were analyzed in dCLN, sCLN and ingLN at day 14.c , Flow cytometry gating strategy for EMV-2-specific T cells. d, Quantification of EMV-2-specific CD8 T cells in dCLNs, sCLNs and ingLNs. (Naïve ; s.c., ; i.c., IVT, ) e, Mice were s.c. inoculated with 50,000 B16 cells, treated with irradiated B16 cells through the routes of s.c., i.c., AC, IVT administration (day 7) along with anti-PD-1 antibodies (days 7, 9 and 11) and monitored for survival. ( for all groups). Data shown as mean s.e.m. -values were calculated using a one-way ANOVA with multiple comparisons testing (Dunnett).
Extended Data Fig. 3 | Eyes have a compartmentalized lymphatic drainage
system.a, Nanoparticles were AC or IVT injected into the eye. Serum was collected, placed on a slide and imaged on a fluorescent microscope 1 h later after injection ( ). , Dye was AC or IVT injected into the left eye. Eyes, sCLNs and dCLNs were collected for IVIS epifluorescence imaging. Representative background-subtracted heat maps of dye in the eye, sCLNs and dCLNs 1 h later after injection. c, d, Dye was AC (c) or IVT(d) injected into the eye and dye presence was measured using a fluorescent plate reader. The percentage of ipsilateral and contralateral LNs with dye was measured 12 h later ( mice).
e, Dye was AC, IVT, or subconjunctivally injected into the left eye. dCLNs were collected and measured using a fluorescent plate reader 1 h later after injection ( ). , dCLNs ( ) or sCLNs ( ) were surgically ligated. 2 days later, dye was AC or IVT injected into the eye. sCLNs (f) or dCLNs (g) were collected and measured using a fluorescent plate reader 1 h later after injection. RFU, relative fluorescence unit. (AC, ;IVT, ). -values were calculated using a one-way ANOVA with multiple comparisons testing (Dunnett). Data shown as mean s.e.m. -values were calculated using a one-way ANOVA with multiple comparisons testing (Dunnett) or two-tailed unpaired Student’s t-test.
Extended Data Fig. 4|10x VISIUM Spatial sequencing of the mouse globe and optic nerve. Mouse eyes were processed using traditional FFPE-based histology preparation. Samples were processed using the 10x VISIUM platform and analyzed using Seurat.a, UMAP of different spatial sequencing blocks with
categorization into overall tissue level structure.b, Projection of tissue-type clusters onto the globe and surrounding structures. c, Image of H&E staining and respective cell type gene signature projected onto spatial sequencing data (yellow arrows denote where the gene signature appears for given signature).
Article
Extended Data Fig. Anatomical characterization of meningeal lymphatic vessels covering optic nerve. Additional images of VEGFR3 and LYVE1 staining on optic nerves of rabbits (a,b), pigs (c,d), non-human primates (e,f,g), and
mice ( ). Yellow arrows indicate vascular structures with single staining for either LYVE1 or VEGFR3. White arrows indicate co-stained vascular structures.
Extended Data Fig. 6| Detailed analysis of optic nerve sheath lymphatics.
a, Whole-mount murine ventral ear sheet stained with LYVE1, CD31, PROX1 and
PROX1 and VEGFR3 were co-stained. Bottom: LYVE1, PDPN, PROX1 and VEGFR3 were co-stained.
VEGFR3.b, Whole-mount of optic nerve sheaths of WT mice. Top: LYVE1, CD31,
Article
a VEGFR3-CreER ; R26-mTmG Ear
b VEGFR3-CreER ;R26-mTmG Optic nerve sheath
Extended Data Fig. 7|Detailed analysis of optic nerve sheath lymphatics with lymphatic reporter mice. a, b, Whole-mount murine ventral ear sheet and optic nerve sheaths of VEGFR3-CreER ;R26-mTmG mice that were stained with LYVE1 and PROX1 (a, scale: ; b, scale: ). c, d, Whole-mount
C PROX1-CreER ;CDH5-Dre;R26-STOP-mCherry Ear
d
PROX1-CreER ;CDH5-Dre;R26-STOP-mCherry Optic nerve sheath
murine ventral ear sheet and optic nerve sheaths of PROX1-CreER ;CDH5-Dre; R26-STOP-mCherry mice that were stained with LYVE1, CD31 and VEGFR3 (c, scale: ; d, scale: ).
Extended Data Fig. | See next page for caption.
Article
Extended Data Fig. 8|Optic nerve sheath lymphatics drain the posterior eye. a, Whole-mount murine optic nerve VEGFR3 staining and tracer signal on optic nerve 1 h later after AC (left) or IVT (right) injection. White arrowheads point to co-localization of VEGFR3 and tracer.b, c, Anti-LYVE1 antibody was IVT injected into the vitreous humor. Optic nerve sheaths were harvested 2 h later and co-stained with CD31, PROX1 and VEGFR3. d, Dye was IVT injected into the eye or intraventricularly into the brain. CSF was collected through the cisterna magna and measured using a fluorescent plate reader 1 h later after injection. e, Anti-LYVE1 antibody was injected IVT into the eye or ICM into the CSF. Dura
was collected 2 h after injection and LYVE1 antibody binding on VEGFR3 positive lymphatic vessels was analyzed by immunofluorescence. Mice without anti-LYVE1 antibody injection were used as the control.f, AF647-OVA was IVT injected into the vitreous humor. Fluorescein was i.p. injected to label blood vessels. The kinetics of AF647-OVA drainage was tracked with Phoenix MICRON IV imaging microscope. g, AF647-OVA was IVT injected into the vitreous humor. The eyes with optic nerve were harvested, and the location of AF647OVA was analyzed at the indicated time points. , Zoomed in images of .
Extended Data Fig. 9 | Lymphatic inhibition enables repeated rAAV administrations. a, Mice were IVT injected with rAAV-VEGFC, rAAV-sVEGFR3 or rAAV-control. Two months later, their optic nerve sheaths were harvested and stained with LYVE1.b, Similar to Fig. 4c, dCLNs were ligated before the primary rAAV-RFP injection. Their RFP intensity were quantified. c, Percentage
of dye retention in the eye was measured for control, dCLN-ligated, or rAAV-sVEGFR3-injected mice. Data shown as mean s.e.m. -values were calculated using a one-way ANOVA with multiple comparisons testing (Dunnett).
Article
Extended Data Fig. 10 | Lymphatic inhibition enables repeated rAAV
transduction.a, Whole mount confocal images of rAAV-RFP rechallenge in
murine retinas after treatment with VEGFC or sVEGFR3 during primary infection.
b, Similar to Fig. 4c, murine retinas were collected, and RFP expression was
quantified using western blot. c, Similar to Fig. 4c, murine retinas were collected, and RFP expression was quantified using flow cytometry. d, Schematic summary of findings. The graphics in were created with BioRender.com.
natureportfolio
Corresponding author(s): Eric Song
Last updated by author(s): December 132023
Reporting Summary
Nature Portfolio wishes to improve the reproducibility of the work that we publish. This form provides structure for consistency and transparency in reporting. For further information on Nature Portfolio policies, see our Editorial Policies and the Editorial Policy Checklist.
Statistics
For all statistical analyses, confirm that the following items are present in the figure legend, table legend, main text, or Methods section.
Confirmed
The exact sample size ( ) for each experimental group/condition, given as a discrete number and unit of measurement
A statement on whether measurements were taken from distinct samples or whether the same sample was measured repeatedly
The statistical test(s) used AND whether they are one- or two-sided
Only common tests should be described solely by name; describe more complex techniques in the Methods section.
A description of all covariates tested
A description of any assumptions or corrections, such as tests of normality and adjustment for multiple comparisons
A full description of the statistical parameters including central tendency (e.g. means) or other basic estimates (e.g. regression coefficient) AND variation (e.g. standard deviation) or associated estimates of uncertainty (e.g. confidence intervals)
For null hypothesis testing, the test statistic (e.g. ) with confidence intervals, effect sizes, degrees of freedom and value noted Give values as exact values whenever suitable.
For Bayesian analysis, information on the choice of priors and Markov chain Monte Carlo settings
For hierarchical and complex designs, identification of the appropriate level for tests and full reporting of outcomes
Estimates of effect sizes (e.g. Cohen’s , Pearson’s ), indicating how they were calculated
Our web collection on statistics for biologists contains articles on many of the points above.
Software and code
Policy information about availability of computer code
Data collection
No software was used
Data analysis
For manuscripts utilizing custom algorithms or software that are central to the research but not yet described in published literature, software must be made available to editors and reviewers. We strongly encourage code deposition in a community repository (e.g. GitHub). See the Nature Portfolio guidelines for submitting code & software for further information.
Data
Policy information about availability of data
All manuscripts must include a data availability statement. This statement should provide the following information, where applicable:
Accession codes, unique identifiers, or web links for publicly available datasets
A description of any restrictions on data availability
For clinical datasets or third party data, please ensure that the statement adheres to our policy
Research involving human participants, their data, or biological material
Policy information about studies with human participants or human data. See also policy information about sex, gender (identity/presentation), and sexual orientation and race, ethnicity and racism.
Reporting on sex and gender
The study did not involve human participants
Reporting on race, ethnicity, or other socially relevant groupings
The study did not involve human participants
Population characteristics
The study did not involve human participants
Recruitment
The study did not involve human participants
Ethics oversight
The study did not involve human participants
Note that full information on the approval of the study protocol must also be provided in the manuscript.
Field-specific reporting
Please select the one below that is the best fit for your research. If you are not sure, read the appropriate sections before making your selection.
X Life sciences Behavioural & social sciences Ecological, evolutionary & environmental sciences
For a reference copy of the document with all sections, see nature.com/documents/nr-reporting-summary-flat.pdf
Life sciences study design
All studies must disclose on these points even when the disclosure is negative.
Sample size
Sample sizes of all experiments are reported,. We graphed a summary of key phenotypes we observed throughout multiple experiments. With these two experiments showing
response rates, the sample size calculation required for an alpha of 0.05 is (power ) and (power ).
Data exclusions
Data was only excluded when experiment collection machine failed and no data points were collected. as were not able to produce readable result files
Replication
All experiments were repeated as a variation (changes in time points) or in smaller numbers, and showed similar trends and experimental conclusions. For most experiments depicted, they are concatenated examples of all experiments performed (not true for experiments where variation between different days are higher, ie. MFI measurements). All experiments were reproducible. All experiments were replicated at least twice and reported accordingly in the legends
Randomization
Mice were randomized to each group and experiments were controlled with controls in each day even for replication experiments
Blinding
At least one investigator was blinded for each study to allow for non-biased reporting of data
Reporting for specific materials, systems and methods
We require information from authors about some types of materials, experimental systems and methods used in many studies. Here, indicate whether each material, system or method listed is relevant to your study. If you are not sure if a list item applies to your research, read the appropriate section before selecting a response.
Goat anti-mouse VEGFR3 (#AF743) and Rat-anti-mouse LYVE1 (# MAB2125) were purchased from R&D. Mouse anti-human VEGFR3 (SC-28297) were purchased from Santa Cruz Biotechnology. Rabbit anti-Prox1 (11-002P) and Rabbit anti Human LYVE-1 (102-PA50S) were purchased from Angio-proteomie. Podoplanin (127402) were purchased from biolegend. Armenian Hamster anti- mouse CD31 (2H8) were purchased from Gene Tex. Goat anti-mouse IgG-AF647 (A21235), Donkey anti-goat IgG- AF647 (A21447), Goat anti-rabbit IgG-AF555 (A21428). Goat anti-Armenian Hamster IgG- AF-488 (# A78963), Goat anti-Syrian Hamster IgG-AF-488 (# A78958) were purchased from Invitrogen.
RFP-Tag rabbit polyclonal antibody (#AP09229PU-N) were purchased from OriGene Technologies.
Mouse anti- Zebrafish zns-2 (ZDB-ATB-081002-34) were purchased from ZIRC. Chicken anti-GFP (Cat# GFP-1010) were purchased from Aves. AF488 conjugated goat anti-mouse IgG AF488 (115-545-146) and Cy5-conjugated donkey anti-chicken IgY (703-175-155) were purchased from Jackson Immuno Research.
Goat anti-mouse immunoglobulin (1010-01) and HRP-conjugated anti-mouse Ig antibodies (1010-05) were purchased from SouthernBiotech.
Rat anti-mouse CD4 antibody (#BE0003-1, GK1.5) were purchased from BioXCell.
Validation
All the antibodies are commonly used and validated antibodies per manufacturer instructions. Please refer to each manufacturer as listed above. No antibodies used were newly generated or unconventionally used antibodies.
Eukaryotic cell lines
Policy information about cell lines and Sex and Gender in Research
Cell line source(s)
GL261-Luc cells were a gift from J. Zhou (Yale Neurosurgery) and were cultured in RPMI supplemented with FBS, penicillin/streptomycin and sodium pyruvate. CT-2A-BFP cells were a gift from T. Mathivet (Paris Centre de Recherche Cardiovasculaire). B16 cells were a gift from N. Palm (Yale Immunobiology).
Authentication
Cells were not authenticated separately, but phenotypes were validated with similar authenticated cell lines from the NIH.
Mycoplasma contamination
All Cells tested negative for mycoplasma contamination
Commonly misidentified lines (See ICLAC register)
The study did not use commonly misidentified lines
Animals and other research organisms
Policy information about studies involving animals; ARRIVE guidelines recommended for reporting animal research, and Sex and Gender in Research
Laboratory animals
Six-to-ten-week-old mixed sex C57BL/6 mice, B6.Cg-Tg(TcraTcrb)425Cbn/J (OT-II), B6.129P2(C)-Ightm2Cgn/J (B1-8) and B6.129S2IghtmICgn/J ( ) mice were purchased from Jackson Laboratory and Charles River and subsequently bred and housed at Yale University. PROX1CreERT2;CDH5 Dre;R26-STOP-mCherry and VEGFR3-CreERT2; R26-MTMG mice were gifts from the Thomas lab. All procedures used in this study (sex-matched, age-matched) complied with federal guidelines and the institutional policies of the Yale School of Medicine Animal Care and Use Committee
Wild animals
The study did not involve wild animals
Reporting on sex
Mixed sex mice were used for our studies ans we did not see any sex-biased phenotypes
Field-collected samples
The study did not involve samples collected from the field
Ethics oversight
All animal work was approved by Yale Institutional Animal Care & Use Committee
Note that full information on the approval of the study protocol must also be provided in the manuscript.
Seed stocks
The study did not involve plants
Novel plant genotypes
The study did not involve plants
Authentication
The study did not involve plants
Flow Cytometry
Plots
Confirm that:
X The axis labels state the marker and fluorochrome used (e.g. CD4-FITC).
X The axis scales are clearly visible. Include numbers along axes only for bottom left plot of group (a ‘group’ is an analysis of identical markers).
X All plots are contour plots with outliers or pseudocolor plots.
X A numerical value for number of cells or percentage (with statistics) is provided.
Methodology
Sample preparation
For brain tissues, tissues were collected and incubated in a digestion cocktail containing collagenase D (Roche) and DNase I (Sigma-Aldrich) in RPMI at for 45 min . Tissues were pipetted to break tissue down and filtered through a filter. Then, cells were mixed in 3 ml of Percoll (Sigma-Aldrich) solution and centrifuged at 580 g for 15 min without brake. The Percoll layer was removed, and cell pellets were treated with 0.5 ml ACK buffer and spun for 5 minutes at 500 g . Then the cell pellets were resuspended in FACS buffer (PBS FBS +1 mM EDTA) for staining.
Instrument
Preparation of single-cell suspensions from spleen, LNs and brains are described above. Nonspecific binding was blocked using a Fc receptor-blocking solution (TruStain FcX , 101320, BioLegend) for 10 minutes at prior to immunostaining. Subsequently, the cells were stained with corresponding antibodies for 30 min at . Then, cells were washed to remove excess antibodies and resuspended in FACS buffer. Samples were run on an Attune NxT flow cytometer and then analyzed using FlowJo software (10.8.1, Tree Star).
Software
FlowJo software (10.8.1, Tree Star)
Cell population abundance
No sorts were performed
Gating strategy
Please refer to supplemental figures and methods for detailed gating strategies
Tick this box to confirm that a figure exemplifying the gating strategy is provided in the Supplementary Information.